Изобретение относится к медицинской биотехнологии и может найти применение при выполнении задач, связанных с распознаванием антигена PRAME человека, в том числе при иммуногистохимическом анализе экспрессии белка PRAME в опухолях человека и для разработки лекарственного средства для таргетной терапии PRAME- экспрессирующих заболеваний. Изобретение относится также к обнаружению последовательности эпитопа антигена PRAME человека, который может быть использован для получения высокоспецифичных в отношении PRAME антител человека. Полученные антитела или их фрагменты образуют комплексы с белком PRAME в иммуногистохимческих препаратах опухолевой ткани, либо в вестернблотинге, либо в проточной цитометрии, и могут быть окрашены вторичными антителами. На основе антител\фрагментов также можно проводить выявление наличия белка PRAME в клеточных лизатах методом иммуноблоттинга и иммуноферментного анализа. Полученные антитела могут применяться также для выявления PRAME-экспрессирующих опухолевых клеток в образцах тканей человека или животного. Применение антител в иммунохимической диагностике позволит их использовать в качестве диагностического и прогностического маркера при онкологическом заболевании.
Белок PRAME (preferentially expressed antigen in melanoma, также известен как СИЗО, МАРЕ, OIP-4, OIP4) человека представляет собой раково-тестикулярный белок, активный в эмбриональных тканях и клетках гамет, и не экспрессирующийся в соматических тканях. Белок PRAME экспрессируется при онкологических заболеваниях любого гистологического происхождения и является существенным прогностическим маркером. При меланоме и других солидных опухолях экспрессия PRAME в основном связана с метастазированием и лекарственной резистентностью, а также с неблагоприятным исходом заболевания. Чем больший уровень экспрессии белка наблюдается, тем более выражено неблагоприятное влияние на прогноз заболевания PRAME не экспрессируется в здоровых соматических клетках взрослого человека. При онкологических заболеваниях активность PRAME наблюдается приблизительно в половине случаев, и вызвана сбоями регуляции экспрессии генов. Практически при всех онкологических заболеваниях наличие белка PRAME в опухолевой клетке связано с сокращением времени выживаемости, увеличением риска метастазирования и рецидивов [Мисюрин В.А. Клиническое значение экспрессии гена PRAME при онкогематологических заболеваниях. Клиническая онкогематология. Фундаментальные исследования и клиническая практика. 2018;11(1):26-33.] [Мисюрин В.А. Прогностическое значение экспрессии гена PRAME при солидных опухолях. Иммунология. 2018; 39(1): 67- 73. DOI: http://dx.doi.Org/10.18821/0206-4952-2018-39-l-67-73.l. С другой стороны, поскольку PRAME представлен только в опухолевой клетке, и не экспрессируется в здоровой, методы, направленные на устранение PRAME-положительной клетки, будут полезны в терапии опухолей. В настоящее время экспрессия PRAME обнаружена при многочисленных онкологических заболеваниях и локализациях: гемангиома печени, рак прямой кишки, меланома кожи, увеальная меланомы, опухоли матки, опухоли простаты, рак гортани, рак желудка, рак кожи, рак кости, рак легких, рак мозга, опухоли гипофиза, рак мочевого пузыря, рак печени, рак пищевода рак, рак почки, киста яичника, рак простаты, саркома молочной железы, хондросаркома, рак слюнной железы, рак шейки матки, рак щитовидной железы, рак яичка, рак яичника, острый миелоидный лейкоз, острый лимфоидный лейкоз, хронический В-клеточный лимфоидный лейкоз, хронический Т-клеточный лимфоидный лейкоз, волосатоклеточный лейкоз, множественная миелома, хронический миелоидный лейкоз, лимфома Бёркитта, фолликулярная лимфома, диффузная В-крупноклеточная лимфома, лимфома из клеток маргинальной зоны селезёнки, лимфома из клеток зоны мантии, Т-клеточные лимфомы кожи, миелодиспластический синдром и другие. Для любого вида опухоли и опухолевой ткани может быть проведен анализ наличия в ней мРНК или белка PRAME.
Количество белка PRAME в опухолевой клетке хорошо коррелирует с уровнем экспрессии мРНК гена PRAME, благодаря чему возможно проводить диагностику как с помощью метода ПЦР с обратной транскрипцией, так и при помощи иммунологических методов. Поскольку экспрессия PRAME является онко-специфичной, её выявление может однозначно свидетельствовать о поражении ткани опухолевыми клетками, что позволяет сделать выводы о степени распространённости опухолевого роста. Кроме этого, выявление белка PRAME может быть полезно при оценке минимальной остаточной болезни после проведения терапии.
Экспрессия белка PRAME опухолевой клеткой может также использоваться для таргетной терапии. PRAME-специфическое антитело может связаться с PRAME-позитивной опухолевой клеткой и инициировать реакцию антитело-зависимой клеточно-опосредованной цитотоксичности либо антитело-зависимой комплемент-опосредованной цитотоксичности. Поскольку это может приводить к гибели опухолевой клетки, данные антитела могут быть полезны в противоопухолевой терапии.
Краткое описание чертежей.
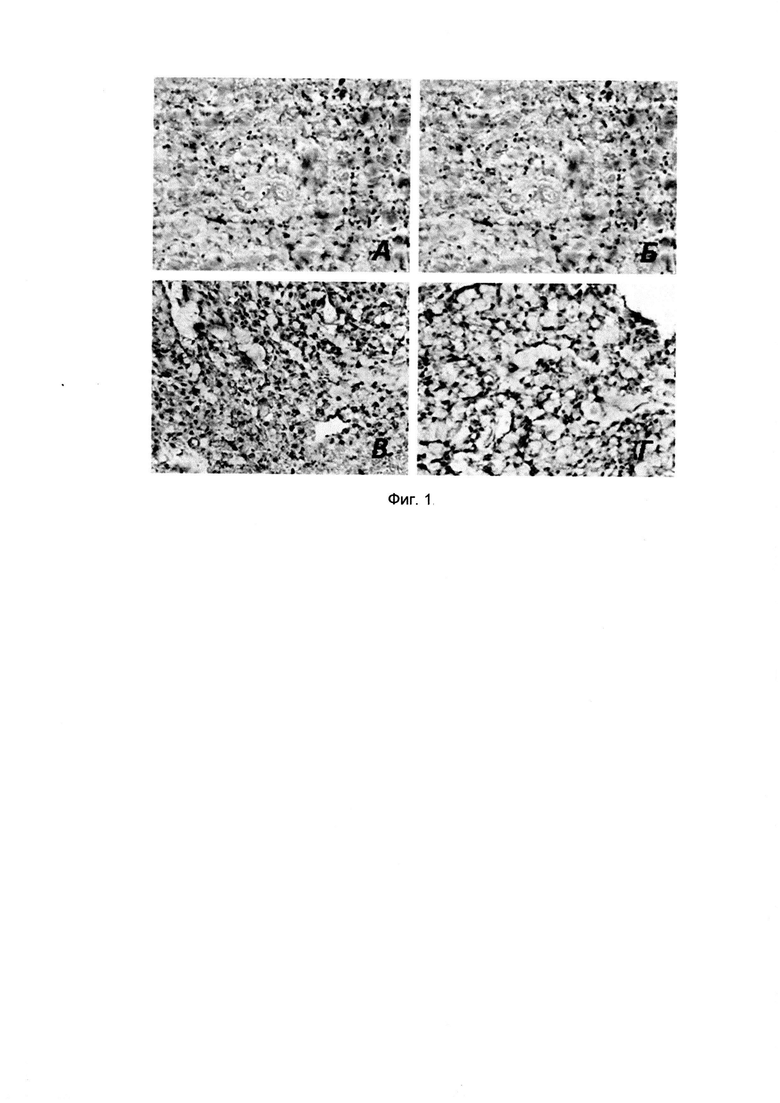
Фиг. 1. Иммуногистохимическое окрашивание антителами ксенографтов, полученных из линии рака лёгкого А549 при помощи мышиных и химерных антител в разведении 1:50: А - 5D3, Б - 6Н8, В - 5D3chim, Г - 6H8chim.
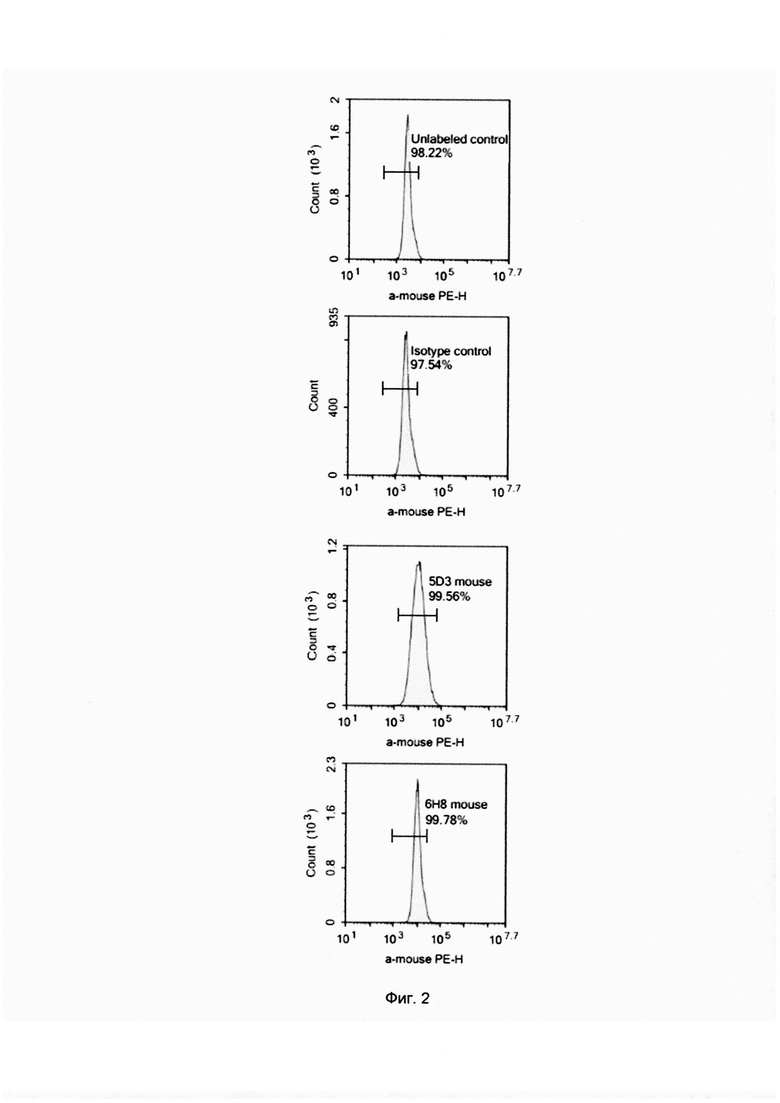
Фиг. 2. Окрашивание клеток линии ТНР1 при помощи антител 5D3 и 6Н8.
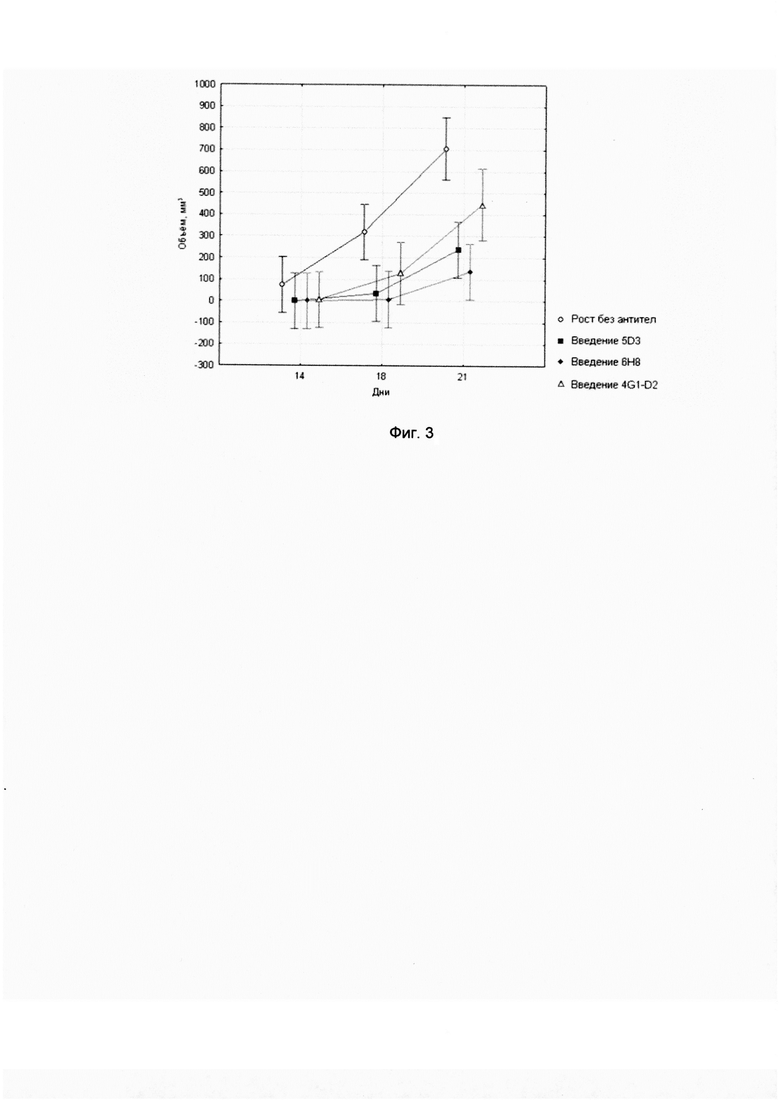
Фиг. 3. Скорость роста клеток линии B16F10-pCEP4-PRAME in vivo в условиях введения различных антител.
Предшествующий уровень техники
В настоящее время известно 9 коммерчески доступных моноклональных антитела, связывающих белок PRAME. Четыре антитела - кроличьи, причём два из них поставляются фирмой Abeam в виде одного реагента аЬ244107 для выполнения ELISA. Ещё одно кроличье антитело аЬ219650 от фирмы Abeam предназначено для выполнения вестернблоттинга, иммуногистохимии, иммуноферментного анализа, проточной цитометрии и иммунопреципитации. Наконец, антитело E7I1B от Creative biolabs предназначено для выполнения вестернблоттинга и иммунопреципитации. Пять мышиных антител представлены клонами Н-10 и D-12 от Santa Cruz biotechnology, которые могут быть использованы в протоколах вестернблоттинга, иммуногистохимии, иммуноферментного анализа, проточной цитометрии и иммунопреципитации, клоном 2В4 от Creative biolabs, которые были разработаны для проведения ELISA и вестернблоттинга, и клонами CL5148 и CL5146 от Thermofisher, разработанным для проведения иммуногистохимии и вестернблоттинга. Доступна информация о 282 поликлональных антител, распознающих человеческий белок PRAME, и пригодных использованию при определении наличия белка PRAME различными методами.
Перечисленные антитела имеют ряд ограничений в использовании. Поликлональные антитела могут быть использованы в лабораторной практике, но не могут быть быстро преобразованы в антитела других типов, например, гуманизированных или химерных, без предварительного клонирования. Известные анти-PRAME моноклональные антитела недостаточно специфично распознают целевой белок. Согласно опубликованным данным вестернблоттинга, из девяти клонов только четыре распознают белок с массой, соответствующей массе PRAME около 59 кДа - это клоны ab219650, Н-10, CL5148 и CL5146. Клон D-12 распознаёт белок другой массы, равной приблизительно 47 кДа. Данные о массе белков, распознаваемых при помощи продукта аЬ244107 и клонов E7I1B и 2В4 в настоящий момент недоступны. В настоящее время в уровне техники нет информации об эпитопах белка PRAME, которые должны присутствовать в антигене при создании PRAME-направленного антитела. Проблема разработки специфичного антитела против белка PRAME человека осложняется тем, что PRAME имеет значительную степень гомологии с белками группы PRAME-F, а также белком ингибитором рибонуклеаз и белками То11-подобных рецепторов. Клон аЬ219650 разрабатывался против полипептида, имеющего структуру белка PRAME в положении 410-509 аминокислотных остатков. Клоны, включенные в состав продукта аЬ244107 были разработаны путём иммунизации полноразмерным белком PRAME человека. Клоны Н-10 и D-12 разрабатывались против полипептида, имеющего структуру белка PRAME в положении 126-205 аминокислотных остатков. Клон 2В4 разрабатывался против полипептида, имеющего структуру белка PRAME в положении 1-151 аминокислотных остатков. Наконец, клоны CL5148 и CL5146 разрабатывались против полипептида, имеющего структуру белка PRAME в положении 265-385 аминокислотных остатков. Все представленные полипептидные фрагменты слишком велики, чтобы не иметь значительной гомологии с белками, которыми гомологичен белок PRAME. Таким образом, специфичность перечисленных клонов вызывает сомнения.
Задачей данного изобретения являлась разработка специфических антител, пригодных для проведения иммуногистохимического анализа экспрессии белка PRAME и элиминации PRAME-экспрессирующих клеток из организма млекопитающего, определение CDR данных антител, что позволит создавать усовершенствованные антитела на их основе. Определение эпитопа PRAME, к которому направлены данные антитела, позволит создать на основании данного эпитопа синтезировать синтетический антиген, при использовании которого могут быть получены высокоспецифичные PRAME-направленные антитела. Разработанные антитела 5D3 и 6Н8 показали эффективное окрашивание белка PRAME в иммуногистохимических реакциях, что позволяет их использовать в онкологической диагностике. В примерах изобретения продемонстрировано использование 5D3 и 6Н8 в иммуногистохимическом анализе экспрессии белка PRAME в клеточных линиях меланомы человека, а также в клеточных линиях меланомы мыши, модифицированных для экспрессии белка PRAME человека, однако данные примеры не ограничивают область применения антител, и они могут применяться для обнаружения экспрессии PRAME в любых видах PRAME- экспрессирующих опухолевых клеток и, соответственно, для выявления любых видов опухолевых заболеваний. Эксперименты по изучению эффективности и специфичности 5D3 и 6Н8 проводились на образцах тканей, в которых активность PRAME подтверждена методом ПЦР в реальном времени. PRAME -негативные образцы не окрашиваются полученными антителами, что демонстрирует их высокую специфичность. Образцы, экспрессирующие мРНК PRAME, окрашивались антителами. Наблюдалась прямая корреляция интенсивности экспрессии гена PRAME со степенью окрашивания белка PRAME в иммуногистохимии. Так, при проведении вестернблоттинга лизатов клеток здорового человека и лизатов опухолевых клеток окрашивается белок с массой 59 кДа, что соответствует массе белка PRAME. В то же время отсутствует окрашивание белков другой массы. Полученные антитела распознают небольшие фрагменты белка PRAME. Данные фрагменты встречаются только в белке PRAME человека, и не представлены в других белках, кодируемых в нормальных или опухолевых клетках человека. Таким образом, связывание антитела с эпитопом, расположенным внутри области 160-200 аминокислотных оснований белка PRAME человека, при условии отсутствия его связывания с белком PRAME за пределами данной области, определяет получение технического результата изобретения - специфическое распознавание белка PRAME человека полученными антителами и специфичное выявление PRAME-экспрессирующих клеток в образцах тканей млекопитающих. Таким образом, если антиген будет содержать последовательность, идентичную последовательности, расположенной в области 160-200 аминокислотных оснований белка PRAME или ее части, и не будет содержать последовательность, расположенную за пределами этой области, с его помощью могут быть получены антитела со специфичностью, аналогичной специфичности антител 6Н8 и 5D3. Из уровня техники известно, что длина эпитопа в составе антигена должна составлять более восьми аминокислотных оснований. Таким образом, предпочтительно, чтобы длина последовательности PRAME в составе антигена включала в себя не менее 9 аминокислотных оснований.
Связывание антител с клеточной мембраной запускает реакцию клеточно- и комплемент-опосредованной цитотоксичности, что приводит к гибели клетки. Именно этот механизм лежит в основе противоопухолевого эффекта, достигаемого антителами. Поскольку PRAME присутствует на поверхности только опухолевой клетки, здоровые клетки не подвергнутся воздействию антител, и не будут уничтожены. Таким образом, антитела, распознающие белок PRAME, могут быть не только эффективным противоопухолевым препаратом, но также отличаются безопасностью.
Список сокращений:
V - вариабельный домен
VL - вариабельный домен легкой цепи
VH - вариабельный домен тяжелой цепи
CDR - определяющий комплементарность участок, гипервариабельный участок вариабельной цепи, образующий антигенсвязывающий центр молекулы антитела.
FR - каркасный регион
Fc-фрагмент - кристаллизующийся фрагмент иммуноглобулина (Fc, fragment crystallizable region, Fc region).
BLAST - онлайн сервис для сравнения, поиска гомологии и определения структурных элементов в нуклеотидных и аминокислотных последовательностях человека и животных.
Антитело - молекула иммуноглобулина, для которого существует антиген.
Аффинность - термодинамическая характеристика, количественно описывающая силу взаимодействия антигена и антитела; определяется по закону действующих масс как отношение концентрации комплекса антиген - антитело к произведению концентраций компонентов.
Авидность - характеристика общей стабильности комплекса антигена и антитела; определяется аффинностью антитела к антигену, количеством антигенсвязывающих центров в молекуле антитела и особенностями пространственной структуры антигена, создающими стерические препятствия для создания комплекса. В отличие от аффинности, которая является термодинамическим параметром, количественно описывающим силу единственного взаимодействия антигена и антитела, авидность описывает силу кооперативных аффинных взаимодействий.
Антигенсвязывающий участок антитела - часть антитела, распознающая эпитоп. Имеет несколько названий: активный или антигенсвязывающий центр антител, антидетерминанта или паратоп.
Комплементарность антитела - способность антитела к специфическому взаимодействию с антигеном. За редким исключением, антитело взаимодействуют только с тем антигеном, который индуцировал их образование и подходит к ним по пространственной структуре.
Опухолевые антигены - антигены, продуцируемые раковыми клетками и способные вызвать иммунный ответ организма. Являются результатом проявления измененного генома раковой клетки.
Специфичность - способность антитела проявлять различную аффинность по отношению к различным антигенам.
Эпитоп, или антигенная детерминанта - часть макромолекулы антигена, небольшой участок молекулы антигена, который взаимодействует со специфичным к нему антителом. Этот участок распознаётся клетками или молекулами иммунной системы (антителами, В-лимфоцитами, Т-лимфоцитами).
PRAME-экспрессирующая клетка - клетка млекопитающего, в которой экспрессируется белок PRAME
PRAME-экспрессирующее заболевание - заболевание, при котором опухолевые клетки экспрессируют белок PRAME
Тяжёлые и лёгкие цепи антител 5D3 и 6Н8 были получены в результате иммунизации мышей рекомбинантным белком PRAME.
Антитела для распознавания антигена PRAME состоят из тяжёлой и лёгкой вариабельных цепей, причём тяжёлая цепь включает области CDRH1, CDRH2, FR3 и CDRH3, которые могут иметь последовательности SEQ ID N0:1, SEQ ID N0:2, SEQ ID N0:3 и SEQ ID N0:4 или SEQ ID N0:5, SEQ ID N0:6, SEQ ID N0:7 и SEQ ID N0:8, и лёгкая цепь включает области CDRL1, CDRL2 и CDRL3, которые имеют последовательности SEQ ID N0:9, SEQ ID N0:10 и SEQ ID N0:11 или SEQ ID N0:12, SEQ ID N0:13 и SEQ ID N0:14.
Антитело или фрагмент антитела для связывания PRAME по изобретению содержит вариабельный домен тяжелой цепи, имеющий аминокислотные последовательности SEQ ID N0:1 и вариабельный домен лёгкой цепи, имеющий аминокислотные последовательности SEQ ID N0:2, либо вариабельный домен тяжелой цепи, имеющий аминокислотные последовательности SEQ ID N0:3 и вариабельный домен лёгкой цепи, имеющий аминокислотные последовательности SEQ ID N0:4.
Антитело или фрагмент антитела по изобретению способны связываться с эпитопами PRAME, имеющими следующие последовательности: KVDGLSTEAEQPFIPVEVLVD (эпитоп, к которому направлено антитело 5D3) DLFLKEGACDELFSYLIEKVK (эпитоп, к которому направлено антитело 6Н8). Эпитоп с последовательностью KVDGLSTEAEQPFIPVEVLVD имеет координаты 160-180 в последовательности природного белка PRAME. Для распознавания данного эпитопа антитело должно в тяжёлой цепи содержать последовательности SEQ ID N0:1, SEQ ID N0:2, SEQ ID N0:3 и SEQ ID N0:4 и в лёгкой цепи последовательности SEQ ID N0:9, SEQ ID N0:10 и SEQ ID N0:11. Данный эпитоп имеет 5 из 21 совпадающих аминокислотных остатков с белками семейства PRAME, 1 из 21 совпадающих аминокислотных остатков с гомологичным участком последовательности белка ингибитора рибонуклеаз и 3 из 21 совпадающего с последовательностью белков семейства TLR. Эпитоп с последовательностью SEQ ID N0:19 имеет координаты 180-200 в последовательности природного белка PRAME. Для распознавания данного эпитопа антитело должно в тяжёлой цепи содержать последовательности SEQ ID N0:5, SEQ ID N0:6, SEQ ID N0:7 и SEQ ID N0:8 и в лёгкой цепи последовательности SEQ ID N0:12, SEQ ID N0:13 и SEQ ID N0:14. Данный эпитоп имеет 5 из 21 совпадающих аминокислотных остатков с белками семейства PRAME, 1 из 21 совпадающих аминокислотных остатков с гомологичным участком последовательности белка ингибитора рибонуклеаз и 1 из 21 совпадающего с последовательностью белков семейства TLR. Всё это показывает высокую специфичность разработанных антител.
Антитело или его фрагмент по изобретению способны образовывать комплекс с белком PRAME человека, окрашивающийся вторичными антителами, что обеспечивает получение ряда технических результатов - возможность выявления экспрессии белка PRAME в образце опухолевых клеток методами иммуногистохимического окрашивания, возможность выявления белка PRAME в лизатах клеток методами иммуноферментного анализа и возможность белка PRAME в лизатах клеток методами вестерн-блоттинга. Представленные в настоящем изобретении фрагменты антител имеют установленную аминокислотную последовательность, на основании которой можно сконструировать антитела различных вариантов, в том числе Fv, scFv, Fab, F(ab') фрагменты, линейные антитела которые могут быть скомбинированы с другими аминокислотными последовательностями, для ориентированной посадки на сорбент, например, с полилизиновым тагом, различные комбинации фрагментов, их димеры, тримеры (диатела, триатела, тетратела, Bis-scFv, минитела, Fab2, Fab3) и так далее. Эти фрагменты антиген-связывающего участка любого антитела определяют специфичность его связывания с антигеном. Моновалентные F(ab) фрагменты имеют один антиген- связывающий сайт, тогда как двухвалентные F(ab')2 -фрагменты имеют две антигенсвязывающие области, которые связаны дисульфидными связями. Уменьшение F(ab')2 фрагментов дает 2 моновалентных Fab' фрагмента, которые имеют свободную сульфгидрильную группу, которая используется для конъюгации с другими молекулами. Fv фрагмент - это наименьший фрагмент, получаемый в результате ферментативного расщепления антител класса IgG и IgM. Fv фрагмент имеет антиген-связывающий сайт, состоящий из областей VH и ВК, но отсутствуют СН1 и CL области. Цепи VH и VL удерживаются вместе в Fv фрагменте нековалентными взаимодействиями.
Методы генной инженерии позволяют использовать нуклеотидные последовательности, экспрессирующие созданные нами вариабельные домены антител, распознающих PRAME, при конструировании антител различных вариантов. Так, можно создать вариабельные одноцепочечные фрагменты (ScFv), которые представляют собой фрагменты типа Fv и включают домены VH и VL, соединенные гибким пептидом. Когда линкер имеет длину по меньшей мере 12 остатков, фрагменты ScFv в основном мономерные. Манипуляция ориентации V-доменов и длины линкера создает различные формы молекул Fv. Линкеры, которые в длину 3-11 остатков, дают ScFv молекулы, которые не могут фолдировать в функциональный домен Fv. Эти молекулы связываются со второй молекулой ScFv, создавая двухвалентное диатело - димер фрагмента антитела формата scFv, "diabody". Диатело может быть и биспецифическое (Бис-scFv), а может и моноспецифическое дивалентное. Если длина линкера составляет менее трех остатков, молекулы scFv объединяться в тритела или тетратела. Мультивалентные scFv обладают большей функциональной аффинностью связывания со своими антигенами-мишенями, чем их аналоги одновалентные из-за связывания с двумя антигенами-мишенями, что уменьшает скорость диссоциации фрагмента антитела. Минитела представляют собой scFv-СНЗ слитые белки,- которые собираются в двухвалентные димеры. Миниатюрные фрагменты scFv могут быть сгенерированы с содержанием двух различных вариабельных доменов, что позволяет этим Бис-scFv молекулам одновременно связываться с двумя различными эпитопами. Генетические методы также используются для создания биспецифических (Fab-димеров) и Fab2 триспецифических Fab тримеров (Fab3). Подобные структуры могут создаваться также путём химического конъюгирования Fab-фрагментов. Эти фрагменты антител способны связываться с 2 (Fab2) или 3 (Fab3) различными антигенами сразу. В дополнение, на основании данных о включении данных фрагментов в состав антитела, могут быть созданы камелидные антитела (нанотела, nanobody), которые содержат набор специфических Heavy Chain Antibodies (hcAb), исключительно состоящих из гомодимера тяжелых цепей и не имеют легких цепей. Fab часть этих антител называется VHH (вариабельный домен тяжелой цепи антитела), является наименьшей антиген-связывающей областью, найденной в природе. Нанотела представляют собой рекомбинантные домены из VHH, способные связывать антигены. Они очень стабильны и могут быть получены в огромном количестве с помощью традиционных систем, таких как бактерии и, таким образом, представляет собой перспективный инструмент для диагностических и терапевтических целей. Нанотела также могут быть экспрессированы непосредственно in vivo. BiTE антитела, составленные из двух одноцепочечных Fv- фрагментов, одним из которых может быть антитело против PRAME, а другим - ScFv фрагмент любого другого антитела, соединённых гибкими линкерами, содержащими повторы глицина-серина. Короткий линкер предотвращает неправильное спаривание внутри цепочки, но не между цепочками. Длинный гибкий линкер позволяет антигенсвязывающим участкам свободно вращаться. Возможно создание наночастиц, в которых один или несколько Fv-фрагментов конъюгированы с биоразлагаемыми разрешенными для внутривенного введения полимерными гранулами PLGA. Формат Trand позволяет создавать антитела, в которых в одну цепь соединены две пары VL и VH доменов, при этом формируется антитело с четырьмя валентностями. Формат кроссмаб предусматривает перестановку константного домена тяжелой цепи СН1 и константного домен CL легкой цепи для правильной сборки биспецифического антитела. Возможна конъюгация Fv-фрагментов с любыми белками неиммуноглобулиновой природы.
Фрагменты имеют преимущества по сравнению с полноразмерными антителами для некоторых целей - например, они достаточно малы, чтобы проникать в ткани, в которые полноразмерные антитела не могут проникать. Это преимущество используется в терапевтических и иммуногистохимических процедурах. Фрагменты антител, как правило, не имеют гликозилирования, что позволяет их производить в прокариотических системах экспрессии. Отсутствие домена Fc является существенным преимуществом для первичных антител, используемых в иммуногистохимических и других целях, так как они значительно уменьшают неспецифическое связывание с рецептором Fc. Фрагменты антител не имеющие Fc-области, имеют редуцированное неспецифическое связывание. Нанотела могут быть использованы также для коиммунопреципитации или в сочетании с флуоресцентными белками для отслеживания в реальном времени внутриклеточных мишеней в живых клетках.
Для снижения иммуногенности полученных антител для человека могут быть получены различные виды химерных и гуманизированных антител. Одним из таких примеров является способ получения химерного антитела, в котором вариабельный участок (V-участок) антитела получен из исходного антитела, а его константный участок (С- участок) получен из подходящего человеческого антитела. Так как полученное таким образом химерное антитело содержит вариабельный участок исходного мышиного антитела в интактной форме, ожидается, что оно будет связываться с антигеном со специфичностью, идентичной специфичности исходного мышиного антитела. Кроме того, в химерном антителе количество аминокислотных последовательностей, полученных из организма, отличного от человеческого, существенно снижено, и поэтому ожидается, что такое антитело будет обладать пониженной иммуногенностью по сравнению с исходным мышиным антителом. Химерное антитело может связываться с антигеном таким же образом, как и исходное мышиное моноклональное антитело, и может включать иммунологические реакции против вариабельного участка мышиного антитела, несмотря на то, что его иммуногенность понижена (LoBuglio A.F. et al., Proc. Natl. Acad. Sci. USA, 86, 4220-4224, 1989). Второй способ снижения иммуногенности мышиного антитела, хотя и гораздо более сложный, позволяет еще более снизить потенциальную иммуногенность мышиного антитела. В этом способе лишь участок, определяющий комплементарность (CDR) вариабельного участка мышиного антитела трансплантируют в вариабельный участок человеческого антитела, получая вариабельный участок "реконструированного" человеческого антитела. Однако для того чтобы сделать структуру вариабельного участка реконструированного человеческого антитела как можно более близкой к структуре исходного мышиного антитела, необходимо, чтобы часть аминокислотной последовательности каркасного участка (FR), который поддерживает CDR, можно было бы трансплантировать из вариабельного участка мышиного антитела в вариабельный участок человеческого антитела. Затем этот V-участок гуманизированного реконструированного человеческого антитела связывают с константным участком человеческого антитела. Часть, которая получена из отличной от человеческой аминокислотной последовательности в окончательно реконструированном гуманизированном антителе является CDR, и составляет лишь часть FR. CDR состоит из гипервариабельной аминокислотной последовательности, которая не содержит видоспецифических последовательностей. Таким образом, гуманизированное антитело, которое также называют перестроенным, содержащее мышиный CDR, не должно быть более иммуногенным, нежели природное человеческое антитело, содержащее CDR человеческого антитела. Общая технология рекомбинантной ДНК для получения таких антител также известна (см. European Patent Application ЕР 125023 и International Patent Application WO 92-19759).
Все использованные штаммы бактерий, плазмиды, праймеры полимеразной цепной реакции (ПЦР), рекомбинантные белки, а также клеточная линия, продуцирующая антитела, коммерчески доступны.
Изобретение иллюстрируется следующими примерами.
Пример 1. Получение гибридом для продукции мышиных антител против PRAME человека.
Для получения антител мышей линии BALB/c (самки массой 16 - 18 г.) иммунизировали дважды с интервалом в 2 недели в подушечки задних лап раствором иммуногена в полном (для первой иммунизации) или неполном (для второй иммунизации) адъюванте Фрейнда. В качестве иммуногена использовали рекомбинантный бело к PRAME, имеющий в своём составе 509 аминокислотных остатков, соответствующих природному белку PRAME, и б остатков гистидина, введённых в структуру белка для удобства очистки. Во всех случаях на одну иммунизацию брали 25 мкг белка, белок вводили в виде эмульсии, полученной из раствора белка в фосфатно- солевом буфере и равного объёма полного (1-я иммунизация) или неполного (2-я иммунизация) адъюванта Фрейнда.
Тестирование сывороток и культуральных супернатантов проводили при помощи непрямого иммуноферментного анализа (Engvall Е, Perlmann P. Enzyme-linked immunosorbent assay (ELISA). Quantitative assay of immunoglobulin G., Immunochemistry. 1971, 8(9): P.871-4). Для этого рекомбинантный белок PRAME в концентрации 1 мкг/мл сорбировали в течение ночи при 4°С в фосфатно-солевом буфере и затем вносили сыворотки в серийных разведениях или культуральные супернатанты на 1 ч при 37°С. Связавшиеся с иммобилизованным антигеном МкАт выявляли с помощью вторичных антител козы, разработанных против IgG (Н + L) мыши, конъюгированных с пероксидазой при помощи периодатного метода (Peroxidase and fluorescein isothiocyanate as antibody markers. A quantitative comparison of two peroxidase conjugates prepared with glutaraldehyde or periodate and a fluorescein conjugate. Broorsma DM, Steefkerk JG, Kors N. J Histochem Cytochem. 1976 Sep;24(9):1017-25.нужна ссылка на источник), в течение 1 ч при 37°С. В качестве субстрата использовали раствор тетраметилбензидина (ТМВ) с перекисью водорода. Оптическую плотность регистрировали при 450 нм.
На 4-й день после второй иммунизации лимфоциты из подколенных узлов гибридизовали с миеломными клетками Sp 2/0-Agl4 по стандартной методике с использованием полиэтиленгликоля с молекулярной массой 4000 (Monoclonal Antibody Production. National Research Council (US) Committee on Methods of Producing Monoclonal Antibodies. Washington (DC): National Academies Press (US); 1999.). Гибридомы отбирали на селективной среде HAT, тестировали в непрямом ИФА и дважды клонировали методом предельных разведений.
Антитела нарабатывали в асцитных жидкостях мышей, для этого мышам, предварительно праймированных пристаном, вводили внутрибрюшинно по 106 клеток каждого клона. Очистку антител из асцитных жидкостей проводили аффинной хроматографией на колонке с белок-G сефарозой по стандартной методике (Jungbauer, А. и др. Comparison of protein A, protein G and copolymerized hydroxyapatite for the purification of human monoclonal antibodies, J. Chromatogr. 1989, 476: 257-268). Чистоту выделенных антител контролировали при помощи SDS-ПААГ электрофореза в 12% полиакриламидном геле в присутствии SDS и DTT в ступенчатой буферной системе Лэмлли, используя Mini PROTEAN 3 Electrophoresis System BIO-RAD, Catalog N 165-3301 (Laemmli, U. K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4, Nature, 1970, 227: 680-685). Чистота всех полученных антител составила не менее 95%.
Всего было получено 2 клона, продуцирующих антитела, распознающие белок PRAME (клоны 5D3 и 6Н8).
Пример 2. Секвенирование антигераспознающих доменов гибридом 5D3 и 6Н8.
Для установления CDR антител 5D3 и 6Н8 необходимо было выявить аминокислотные последовательности полученных антител, что можно было достигнуть при помощи секвенирования генов, кодирующих их.
Для секвенирования проводилась наработка клеток гибридом до количества не менее 1 миллиона. Из этих клеток была выделена РНК по методике, предложенной Chomczynski и Sacchi (Chomczynski P. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction / P. Chomczynski, N. Sacchi // Anal Biochem. - 1987.- 162(1). -P. 156-159). Из на матрице выделенной РНК по общепризнанной методике синтезировалась кДНК. Последовательность кДНК была просеквенирована при помощи сета праймеров, разработанных фирмой Novagen по инструкции, предложенной производителем. Были получены следующие нуклеотидные последовательности лекой и тяжелой цепей антител 5D3 и 6Н8:
Полученные последовательности были загружены в BLAST, в подраздел для выравнивания последовательностей иммуноглобулинов IgBLAST. Инструменты данного сервиса позволили установить первичные белковые структуры вариабельных доменов тяжёлых и легких цепей антител 5D3 и 6Н8. Выравнивание позволило установить аминокислотные замены и определить расположение CDR и FR в антителах 5D3 и 6Н8.
Полученные антитела характеризуются следующими последовательностями:
SEQ ID N0:1 (CDRH1 тяжёлой цепи антитела 5D3)
GYTFTNYG
SEQ ID N0:2 (CDRH2 тяжёлой цепи антитела 5D3)
INTYTGEP
SEQ ID N0:3 (FR3 тяжёлой цепи антитела 5D3)
RFTFS
SEQ ID N0:4 (CDRH3 тяжёлой цепи антитела 5D3)
TRPTGDY
SEQ ID N0:5 (CDRH1 тяжёлой цепи антитела 6Н8)
MSCKTSGYTFPTSW
SEQ ID N0:6 (CDRH2 тяжёлой цепи антитела 6Н8)
INPNSDYT
SEQ ID N0:7 (FR3 тяжёлой цепи антитела 6Н8)
SEDSAVYFC
SEQ ID N0:8 (CDRH3 тяжёлой цепи антитела 6Н8)
ARRGPLALDY
SEQ ID N0:9 (CDRL1 лёгкой цепи антитела 5D3)
QSLLNSRTRKNY
SEQ ID N0:10 (CDRL2 лёгкой цепи антитела 5D3)
WAS
SEQ ID N0:11 (CDRL3 лёгкой цепи антитела 5D3)
KQSYTLYT
SEQ ID N0:12 (CDRH1 лёгкой цепи антитела 6H8)
ENIYSN
SEQ ID N0:13 (CDRH2 лёгкой цепи антитела 6H8)
TAT
SEQ ID N0:14 (CDRH3 лёгкой цепи антитела 6H8)
QHFWGTPLT
SEQ ID N0:15 (тяжёлая цепь 5D3)
QIQLVQSGPELKKPGETVKISCKASGYTFTNYGMNWVKQAPGKGLKWMGWINTYTGEPTYADDFKGRFTFSLETSASTAYLQINNLKNEDTATYFCTRPTGDYWGQGTSVTVSSA
SEQ ID N0:16 (легкая цепь 5D3)
DIVMSQSPSSLAVSAGEKVTMSCKSSQSLLNSRTRKNYLAWYQQKPGQSPKLLIYWASTRESGVPDRFTGSGSGTDFTLTISSVQAEDLAVYYCKQSYTLYTFGGGTKLEIK
SEQ ID N0:17 (тяжёлая цепь 6H8)
QVQLQQSGAELAKPGASVKMSCKTSGYTFPTSWMHWVKQRPGQGLEWIGYINPNSDYTKYNQKFKDKATLTADKSSSTAYMQLSSLTSEDSAVYFCARRGPLALDYWGQGTSVTVSSA
SEQ ID N0:18 (легкая цепь 6H8)
DIQMTQSPASLSVSVGETVTITCRASENIYSNLAWYQQKQGKSPQLLVYTATNLADGVPSRFSGSGSGTQYSL KINSLQSEDFGSYYCQHFWGTPLTFGAGTKLELK
Пример 3. Вестерн-блоттинг белка PRAME при помощи антител 5D3 и 6Н8. Для доказательства способности полученных антител связываться с белком PRAME был проведён эксперимент по выявлению данного белка методом иммуноблоттинга.
Метод иммуноблоттинга (вестерн-блоттинг) позволяет визуализовать иммунные комплексы антиген-антитело после электрофореза препарата антигена в денатурирующих условиях с последующим переносом на мембрану [Towbin Н. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications / H. Towbin, T. Staff helint, J. Gordon // Proc. Natl. Acad. Sci. USA. - 1979.-Vol. 76(9). -P. 4350- 4354]. Главное отличие этого метода от непрямого ИФА заключается в возможности дискриминировать иммунные комплексы с различной молекулярной массой, например, мономерные и олигомерные формы антигена, а также интактный антиген и продукты его деградации.
Электрофорез белков проводили по методу Лэммли. На одну дорожку 10% ПААГ наносили 1 или 2 мкг рекомбинантного белка PRAME (в буфере с ЮОмМ DTT для восстанавливающих условий либо без DTT для невосстанавливающих), либо 20-50 мкг лизата клеток, экспрессирующих белок PRAME. Для калибровки размеров и контроля переноса белка использовали маркер молекулярных масс белка RainbowFull Range (GE Healthcare). После электрофореза перенос белков из геля на Hybond-P PVDF мембрану (GE Healthcare, Великобритания) осуществляли методом полусухого электроблоттинга в течение часа. Мембрану блокировали в 3% БСА в буфере 50мМ трис рН=7,5, 8г/л NaCI, 0,1% Твин 20 в течение 2 часов при комнатной температуре. В качестве первичных антител использовали антитела 5D3hum, 5D3chim, 6H8chim и Fllchim в концентрации 3 мкг/мл в буфере, содержащим 1% БСА, 50мМ трис рН=7,5, 8г/л NaCI, 0,1% Твин 20, инкубировали ночь при +4оС. Затем троекратно отмывали по 15 минут в буфере 50мМ трис рН=7,5, 8г/л NaCI, 0,1% Твин 20 и инкубировали мембрану с вторичным антителом 4G7-HRP (ОАО «ВЦМДЛ», Россия) в разведении 1:50000 в течение часа при комнатной температуре. Снова троекратно отмывали по 15 минут. Для визуализации антител, связавшихся с белком, использовали раствор диаминобензидина 1мг в мл в субстратном буфере (50мМ трис рН=7,5,50мМ имидазол, 0,1% перекись водорода), реакцию останавливали избытком дистиллированной воды. Альтернативно для проявления сигнала на мембране использовали двухкомпонентную систему Clarity Western ECL (Bio-Rad). Затем экспонировали сигнал на люминесцентно-чувствительную пленку (Пленка Hyperfilm ECL) 1-10 минут. Пленку проявляли в проявителе Д-76 (Silberra) 30-60 секунд и фиксировали в фиксаже FIXER А1 (Silberra) 1-5 минут.
Размер молекулярной массы рассчитывали методом линейной регрессии [Фармакопейная статья 1.2.1.0023.15 Электрофорез в полиакриламидном геле. -2015. - Государственная фармакопея Российской Федерации. - XIII изд. Т. 1]. Для этого определяли нормализованные расстояние пробега для каждой полосы белка (маркера или образца антитела) от вершины разрешающего геля. Вычисляли отношение расстояния пробега каждого белка к расстоянию пробега, пройденного фронтом красителя (Rs). Нормализованные расстояния пробега, вычисленные таким образом, называются относительными подвижностями белков и обозначаются, как Rf. Строили график зависимости логарифма относительных молекулярных масс белковых стандартных образцов от полученных значений Rf. Параметры для построения калибровочной кривой представлены в таблице 3.4.1 и на рисунке 3.4.2. Молекулярную массу белка, связанного с антителом, определяли с помощью метода линейной регрессии, если значение, полученное для испытуемых образцов, располагалось на линейной части графика. Для сигналов на мембране, соответствующих связыванию антитела с белком, значения Rf составляли 0,446 и 0,464 для разных концентраций нанесенного белка, что по калибровочной прямой соответствует 59,29+1,21 кДа. При этом рассчитанная теоретически с помощью программы VectorNTIAdvance 10.3.0 молекулярная масса рекомбинантного белка составляет 60 кДа. Результаты представлены на фигуре 1.
Таким образом, антител различают белок, имеющий массу, соответствующую массе белка PRAME, что свидетельствует об их специфичности.
Пример 4. Эпитопный анализ антител 5D3 и 6Н8.
Окончательное установить специфичность антител можно проведя поиск эпитопов, которые они распознают.
Эпитопный анализ сначала проводили методом Вестерн-блоттинга в восстанавливающих условиях с использованием лизатов клеток E.coli, экспрессирующих рекомбинантные белки, содержащие фрагменты белка PRAME. Было выбрано 5 фрагментов белка PRAME, которые были экспрессированы в клетках независимо в составе 5 слитых рекомбинантных белков, каждый из которых содержал аминокислотную последовательность тиоредоксина и соответствующий фрагмент белка PRAME. В качестве контроля использовали клетки, экспрессирующие только тиоредоксин (Тгх) и клетки экспрессирующие N-половину белка PRAME (фрагмент PN, в составе которого находятся аминокислотные остатки с 1 по 220 из первичной структуры природного белка PRAME) без тиоредоксина. N-половина белка была разбита на 4 приблизительно равные части с перекрыванием на 20 аминокислот. Фрагмент PN1 представляет собой полипептидную цепочку, в составе которой находятся аминокислотные остатки природного белка PRAME с 1 по 65. Фрагмент PN2 представляет собой полипептидную цепочку, в составе которой находятся аминокислотные остатки природного белка PRAME с 56 по 125. Фрагмент PN3 представляет собой полипептидную цепочку, в составе которой находятся аминокислотные остатки природного белка PRAME с 115 по 180. Фрагмент PN4 представляет собой полипептидную цепочку, в составе которой находятся аминокислотные остатки природного белка PRAME с 160 по 222. 5-й фрагмент (PN4.3) представлял собой гидрофильный полипептид KRKKNVLRLCCKKLK (аминокислотные остатки 200 по 214 из первичной структуры природного белка PRAME), который с высокой вероятностью мог быть эпитопом для антител 5D3 и 6Н8. Схема фрагментов белка PRAME, слитых с тиоредоксином, представлена на фигуре 2.
Для получения лизатов клетки E.coli ресуспендировали в буфере для нанесения (60 тМ трис-CI, рН=6,8, 10% глицерин, 2% (w/v) додецилсульфат натрия, ЮОмМ DTT, 0.01% бромфеноловый синий) и прогревали 5 минут при 95оС. Полученные лизаты наносили на 12% ПААГ и проводили электрофорез по методу Лэммли. Для контроля переноса и размеров белков использовали маркер молекулярных масс белка RainbowFullRange (GE Healthcare). После электрофореза перенос белков из геля на Hybond-P PVDF мембрану (GE Healthcare) осуществляли методом полусухого электроблоттинга в течение часа. Мембрану блокировали в 3% БСА в буфере 50мМ трис рН=7,5, 8г/л NaCI, 0,1% Твин 20 в течение 2 часов при комнатной температуре. В качестве первичных антител использовали различные антитела 5D3 и 6Н8в концентрации 1,5 мкг/мл в буфере, содержащим 1% БСА, 50мМ трис рН=7,5, 8г/л NaCI, 0,1% Твин 20, инкубировали ночь при +4оС. Затем троекратно отмывали по 15 минут в буфере 50мМ трис рН=7,5, 8г/л NaCI, 0,1% Твин 20 и инкубировали мембрану с вторичным антителом Anti-Mouse-HRP (Dako) в разведении 1:2000 в течение часа при комнатной температуре. Снова троекратно отмывали по 15 минут. Для визуализации антител, связавшихся с белком на мембране, использовали раствор диаминобензидина 1мг в мл в субстратном буфере (50мМ трис рН=7,5, 50мМ имидазол, 0,1% перекись водорода), реакцию останавливали избытком дистиллированной воды. Результаты представлены на фигурах 3 и 4.
Результаты частичного картирования эпитопов антител к белку PRAME методом Вестерн-блот представлены в сводной таблице 1.
Таблица 1-Узнавание фрагментов белка PRAME антителами
наблюдалось
наблюдалось
наблюдалось
наблюдалось
наблюдалось
наблюдалось
наблюдалось
Таким образом, эпитопы PRAME, распознаваемые антителом 5D3, с высокой вероятностью находятся в пределах 160-180 аминокислотных остатков, в то время как эпитопы, распознаваемые антителом 6Н8, с высокой вероятностью находятся в пределах 180-222 аминокислотных остатков.
Следующий этап исследования был проведён методом ИФА. Для данного эксперимента готовили лизаты клеток E.coli, экспрессирующих рекомбинантный тиоредоксин, слитый с N-фрагментами белка PRAME, следующим образом: осадок клеток ресуспендировали в RIPA буфере (10 mM Tris-CI (рН 8.0), 1 тМ EDTA, 1% Triton Х-100, 0.1% sodium deoxycholate, 0.1% SDS, 140 тМ NaCI, 1 тМ PMSF), замораживали при -70°С, затем оттаивали на льду для максимальной экстракции белков, далее центрифугировали в течение 10 минут при 12000 об/мин и температуре 4°С. Супернатанты отбирали, измеряли в них концентрацию белка, аликвоты супернатантов хранили при -20°С.
Клеточные лизаты E.coli PN3, PN4 и PN4.3 сорбировали на планшет в концентрации 1 мкг/мл в ФСБ (0,01М КН2Р04Д1М NaCI), рН 7.2-7.4, по 50 мкл в лунки 96-луночного планшета для ИФА в течение ночи при 4°С. Планшет трижды отмывали ФСБ -Т (0,1% Твин 20) по 270 мкл в лунку. Затем титровали антитела к белку PRAME и контрольное антитело G9 в диапазоне концентраций от 10 нг/мл до 10000 нг/мл с шагом 3 по 50 мкл в лунку в ФСБ -AT (0,01М КН2Р04,0,1М NaCI; 0,2% бычий сывороточный альбумин; 0,05% Твин 20). Планшет инкубировали в течение 1 часа при комнатной температуре. Антивидовой пероксидазный конъюгат антител против IgG мыши разводили в ФСБ -AT в соотношении 1:75000 и вносили по 50 мкл в лунки планшета. Планшет инкубировали в течение 1 часа при комнатной температуре. Планшет трижды отмывали ФСБ -Т по 270 мкл в лунку. Добавляли субстрат ТМБ по 50 мкл/лунку, инкубировали 30 минут при комнатной температуре. Реакцию останавливали добавлением по 50 мкл/лунку 0,5М серной кислоты. Измеряли оптическую плотность в лунках при длине волны 450 нм.
Результаты подтвердили предположение о том, что эпитопы PRAME, распознаваемые антителом 5D3, находятся в пределах 160-180 аминокислотных остатков, в то время как эпитопы, распознаваемые антителом 6Н8, находятся в пределах 180-200 аминокислотных остатков.
Пример 5. Иммуногистохимический анализ экспрессии PRAME при помощи химерных антител 5D3 и 6Н8.
Данный эксперимент был проведён для доказательства пригодности различных вариантов синтетических антител, направленных на эпитопы PRAME 160-180 и 180-200, в работе по выявлению экспрессии белка PRAME в различных гистологических препаратах опухолевых клеток.
Химерные антитела 5D3chim и 6H8chim были получены известными методами путем объединения последовательностей генов, кодирующих вариабельные домены данных антител с последовательностями, кодирующими константные домены IgGl человека. Причём последовательности, кодирующие вариабельные домены лёгких цепей были объединены с последовательностями генов, кодирующих константные домены лёгких цепей, и последовательности, кодирующие вариабельные домены тяжёлых цепей были объединены с последовательностями генов, кодирующих константные домены тяжёлых цепей. Каждая из полученных конструкций была интегрирована в вектор рСЕР4. Пары векторов, кодирующих компоненты антител 5D3chim и 6H8chim были трансфицированы в клетки СНО. Эти клетки продуцировали готовые антитела 5D3chim и 6H8chim среду, из которой они в дальнейшем очищались методом аффинной хроматографии.
Приготовление парафиновых блоков из материала опухоли проводилось следующим образом. Кусочки опухоли фиксировали в забуференном 10% формалине в течение 12 часов при 4°С. После извлечения из формалина, кусочки материала (0,5х 0,5 мм) промыли под проточной водой в течение 12 часов. Далее материал поместили в этиловый спирт, с содержанием не менее 50% на 1 час при комнатной температуре. После этого материал перенесли в 70% этиловый спирт и оставили в нем на ночь при комнатной температуре. На следующем этапе материал обезводили, проведя его по батарее спиртов, содержащих 80%, 96% (I), 96% (II), 100% этилового спирта. Время инкубации материала в каждом из них в течение 1 часа при комнатной температуре. В дальнейшем материал провели через хлороформ (I), хлороформ (II) по 2 часа в каждом при комнатной температуре, после чего поместили в хлороформ-парафиновую смесь (1:1) в термостат с температурой 40°С. После инкубирования материал провели через парафин (I), а затем парафин (II) в течение 1 часа в термостате при 52-54°С. Затем перенесли материал в заливочный парафин при 54-56°С. Пропитанный парафином материал залили в специальные формочки. Затем полученные парафиновые блоки насадили на деревянную подложку (кубики). С помощью микротома были сделаны срезы с блоков толщиной в 3-4 мкм и помещены на предметные стекла с адгезивным покрытием. Далее полученные препараты подсушили на термостолике при температуре не более 37°С.
Иммуногистохимическое окрашивание препаратов было выполнено согласно стандартной методике [Tuffaha M.S.A., Guski Н., Kristiansen G. Immunohistochemistry in Tumor Diagnostics / M.S.A. Tuffaha, H. Guski, G. Kristiansen, Cham: Springer International Publishing, 2018.] [Renshaw S. Immunohistochemistry and Immunocytochemistry: Essential Methods / Wiley-Blackwell, 2017.] [«Правила надлежащей лабораторной практики», утвержденные Приказом Министерства здравоохранения Российской Федерации № 199н от 1 апреля 2016 г.].
Оценку результатов окрашивания провели с применением светового микроскопа Nikon 80i с фотокамерой NikonDs-lFi и программным обеспечением NikonNISBasicResearch (Nikon) при увеличении х100, х200 и х400, получая серию фотографий для дальнейшего анализа. Определение интенсивности окрашивания проводили путем подсчета количества положительно окрашенных CD8+ лимфоцитов не менее чем в 5 репрезентативных полях зрения в областях наиболее интенсивного окрашивания («high- powerfields»). Работы проводились двумя независимыми исследователями. Для определения результатов использовали систему балльной оценки. Для оценки цитоплазматического окрашивания применялась следующая шкала: 0 - отсутствие окрашивания, 1+ - окрашивание слабой интенсивности, 2+ - окрашивание средней интенсивности, 3+-окрашивание высокой интенсивности.
Было проведено исследование ИГХ окрашивания ксенографта линии рака лёгкого А549 при помощи мышиных и химерных антител в разведении 1:50: А - 5D3, Б - 6Н8, В - 5D3chim, F"-6H8chim. Результаты представлены на фиг. 1.
Для подтверждения возможности специфического выявления PRAME на поверхности клеток, полученных из различных видов тканей и различных опухолей, было проведено исследование иммуногистохимического окрашивания при помощи химерных антител на ксенографтах других видов опухолей человека - рака яичников SK-OV-3, аденокарциномы А549, рака толстого кишечника НТ-29, гепатокарциномы Huh-7, рака предстательной железы 22Rvl.
При изучении окрашивания на ксенографте аденокарциномы А549 было получено окрашивание более 80% опухолевых клеток антителами обоих исследованных клонов. При этом наибольшая интенсивность окрашивания наблюдалась при разведении антител 1:50. При большем разведении 1:100 наблюдали снижение интенсивности окраски опухолевых клеток. Интенсивность окраски опухолевых клеток при разведении 1:50 составила 2+ для обоих клонов. На ксенографте аденокарциномы толстого кишечника НТ-29 было получено позитивное окрашивание у более чем 90% опухолевых клеток с интенсивностью 2+ при разведении обоих клонов антител 1:50. Наблюдается окрашивание стромальных клеток ксенографта. При изучении иммуногистохимического окрашивания с химерными антителами на ксенографте гепатокарциномы, не экспрессирующей PRAME по данным ПЦР-анализа, не было выявлено взаимодействия PRAME со всеми клонами антител. На линии рака яичников SK-OV-3 можно было дифференцировать окраску в цитоплазме и на клеточной стенке.
Данный пример подтверждает возможность использования антител 5D3 и 6Н8 в экспериментах по иммуногистохимическому окрашиванию белка PRAME.
Пример 6. Анализ экспрессии поверхностного PRAME при помощи химерных антител 5D3 и 6Н8 методом проточной цитометрии.
Для того чтобы показать возможность связывания антител 5D3 и 6Н8 с поверхностным белком PRAME было выполнено окрашивание клеток линии ТНР1 данными антителами, связывание которых с клетками доказывалось вторичными антимышиными РЕ-мечеными антителами.
Экспрессия гена и белка PRAME в клетках ТНР1 предварительно было доказано методами ПЦР в реальном времени и вестерн-блоттингом. Перед проведением поверхностного окрашивания клетки трижды были отмыты от культуральной среды раствором PBS до получения осадка. Полученный осадок разводился раствором PBS до суспензии, содержащей 500 000 клеток в мл. К 50 мкл суспензии клеток добавлялось по 10 мкл антител 5D3 или 6Н8 или изотипического контроля в концентрации 1 мг/мл. Инкубирование с первичными антителами проводилось в течение 25 минут, после чего проводилась отмывка раствором PBS от несвязавшихся антител. После этого к клеткам добавлялись по 2 мкл вторичных антимышиных РЕ-меченых антител. После инкубирования в течение 25 минут в темноте проводилась отмывка раствором PBS от несвязавшихся вторичных антител и анализ на проточном цитометре ACEA NovoCyte. Полученные результаты показывают, что связывание первичных антител с поверхностным белком PRAME прошло успешно. Интенсивность окрашивания контроля без использования первичных антител составила 2905 единиц, окрашивания с использованием изотипического контроля - 2798 единиц, окрашивания антителом 5D3 - 10511 единиц, окрашивания антителом 6Н8 - 10066 единиц. Результаты представлены на фигуре 2.
Таким образом, антитела 5D3 и 6Н8 могут быть использованы в экспериментах, где ожидается их связывание с белком PRAME, в том случае, если он экспрессируется на поверхности клеток. Связавшиеся антитела могут быть выявлены стандартными иммунохимическими методами. В случае отсутствия PRAME на поверхности клеток неспецифического связывания не происходит. Высокая специфичность антител позволяет их использовать в качестве диагностических антител для выявления экспрессии PRAME на поверхности клеток.
Пример 7. Противоопухолевый эффект использования мышиных антител in vivo.
Поскольку антитела способны распознавать поверхностный белок PRAME, они могут in vivo стимулировать антителозависимую цитотоксичность против PRAME- экспрессирующих опухолей.
20 мышам линии C57BI/6 вводили клетки меланомы мыши B16F10-pCEP4-PRAME, трансфицированные плазмидой с геном PRAME и экспрессирующие соответствующий белок, в дозе 5*104 на мышь. Экспрессия человеческого белка PRAME в трансфицированных клетках B16F10 перед прививанием мышам была подтверждена методом иммуноблота. В качестве контрольной линии использовались клетки меланомы мыши B16F10-pCEP4, трансфицированные пустым вектором. За 3 дня до введения опухолевых клеток, а также на 4, 7, 14, 21 день после введения клеток меланомы, животным внутрибрюшинно вводили 300 мкг мышиных моноклональных антител против PRAME. В контрольной группе использовали мышиные моноклональные антитела 4G1-D2, разработанные против опухолевого антигена GAGE. В каждой группе было по 5 мышей. На 14, 18 и 21 день проводили измерение размеров опухоли в кубических мм. Противоопухолевый эффект оценивали по измерениям размеров опухоли во всех группах. Статистический анализ проводился при помощи критерия Манна-Уитни в программе Statistica 10.
Опухоль B16F10-pCEP4-PRAME реагировала на введение мышам антител 5D3 и 6Н8, распознающих антиген PRAME. Наименьшая скорость роста по сравнению с контролем без применения антител наблюдалась при введении мышам антитела 6Н8 (р=0,0028), и несколько большая - при введении антител 5D3 (р=0,0397). Скорость роста опухоли B16F10- pCEP4-PRAME у мышей, получивших антитело 4G1-D2, была статистически незначимо меньшей по сравнению с контролем (р=0,5762). Результаты представлены на фигуре 3. Таким образом, при введении анти-PRAME антител достигалось снижение скорости роста PRAME-позитивной опухоли.
Таким образом, антитела 5D3 и 6Н8 могут быть основой для создания противоопухолевого препарата, использующегося в таргетной терапии PRAME- экспрессирующих заболеваний.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО 5D3HU, СВЯЗЫВАЮЩЕЕСЯ С ОПУХОЛЕВЫМ АНТИГЕНОМ PRAME, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ АНТИТЕЛА | 2020 |
|
RU2761876C1 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО 6H8HU, СВЯЗЫВАЮЩЕЕСЯ С ОПУХОЛЕВЫМ АНТИГЕНОМ PRAME, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ АНТИТЕЛА | 2020 |
|
RU2768737C1 |
| Антитело, связывающееся с карбоангидразой, и его применение | 2017 |
|
RU2727682C1 |
| АНТИТЕЛА, ПРИГОДНЫЕ В ДИАГНОСТИКЕ РАКА | 2018 |
|
RU2815883C1 |
| ДИАГНОСТИЧЕСКИЕ АНАЛИЗЫ И НАБОРЫ ДЛЯ ДЕТЕКЦИИ ФОЛАТНОГО РЕЦЕПТОРА 1 | 2013 |
|
RU2668824C2 |
| АНТИТЕЛА ПРОТИВ КЛАУДИНА 18.2 ДЛЯ ДИАГНОСТИКИ РАКА | 2013 |
|
RU2661772C2 |
| ЧЕЛОВЕЧЕСКОЕ ОПУХОЛЕСПЕЦИФИЧЕСКОЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО | 2008 |
|
RU2488593C2 |
| Антитела к CD43 и их применение для лечения рака | 2016 |
|
RU2694903C1 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И ПРЕДУПРЕЖДЕНИЯ РАКА | 2009 |
|
RU2533460C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ И/ИЛИ ПРОФИЛАКТИКИ РАКА | 2011 |
|
RU2598258C2 |
Изобретение относится к области биохимии, в частности к антителу или его антигенсвязывающему фрагменту для специфического связывания с белком PRAME. Раскрыто применение указанного антитела для получения лекарственного средства для таргетной терапии PRAME-экспрессирующих опухолевых заболеваний; для выявления экспрессии белка PRAME в образцах тканей и клеток человека; для стимуляции антителозависимой цитотоксичности против PRAME-экспрессирующих опухолей; для снижения скорости роста PRAME-экспрессирующих опухолей. Изобретение позволяет эффективно лечить заболевания, ассоциированные с фактором PRAME. 5 н. и 4 з.п. ф-лы, 3 ил., 1 табл., 5 пр.
1. Антитело или его антигенсвязывающий фрагмент для специфического связывания с белком PRAME человека, характеризующееся следующими свойствами: а) распознающее эпитоп, расположенный в области 160-200 аминокислотных оснований белка PRAME человека; б) не распознающее эпитопы, расположенные за пределами области 160-200 аминокислотных оснований белка PRAME человека; в) по существу, не связывается с белками человека группы PRAME-F, ингибитором рибонуклеаз и белками То11-подобных рецепторов, г) последовательность которого содержит комбинацию CDR, соответствующую одному из следующих вариантов:
1) CDR1 вариабельной области тяжелой цепи с последовательностью GYTFTNYG, CDR2 вариабельной области тяжелой цепи с последовательностью INTYTGEP, CDR3 вариабельной области тяжелой цепи с последовательностью TRPTGDY, CDR1 вариабельной области легкой цепи с последовательностью QSLLNSRTRKNY, CDR2 вариабельной области легкой цепи с последовательностью WAS, CDR3 вариабельной области легкой цепи с последовательностью KQSYTLYT,
2) CDR1 вариабельной области тяжелой цепи с последовательностью MSCKTSGYTFPTSW, CDR2 вариабельной области тяжелой цепи с последовательностью INPNSDYT, CDR3 вариабельной области тяжелой цепи с последовательностью ARRGPLALDY, CDR1 вариабельной области легкой цепи с последовательностью ENIYSN, CDR2 вариабельной области легкой цепи с последовательностью ТАТ, CDR3 вариабельной области легкой цепи с последовательностью QHFWGTPLT.
2. Антитело или его антигенсвязывающий фрагмент по п. 1, распознаваемый эпитоп для которого имеет длину не менее 9 аминокислотных оснований.
3. Антитело или его антигенсвязывающий фрагмент по п. 1, распознающее эпитоп, расположенный в области 160-180 аминокислотных оснований белка PRAME человека.
4. Антитело или его антигенсвязывающий фрагмент по п. 1, распознающее эпитоп, расположенный в области 180-200 аминокислотных оснований белка PRAME человека.
5. Антитело или его антигенсвязывающий фрагмент по п. 1, связывающее пептид, содержащий аминокислотную последовательность, выбранную из KVDGLSTEAEQPFIPVEVLVD и DLFLKEGACDELFSYLIEKVK.
6. Применение антитела или его антигенсвязывающего фрагмента по п. 1 для получения лекарственного средства для таргетной терапии PRAME-экспрессирующих опухолевых заболеваний.
7. Применение антитела или его антигенсвязывающего фрагмента по п. 1 для выявления экспрессии белка PRAME в образцах тканей и клеток человека.
8. Применение антитела или его антигенсвязывающего фрагмента по п. 1 in vivo для стимуляции антителозависимой цитотоксичности против PRAME-экспрессирующих опухолей.
9. Применение антитела или его антигенсвязывающего фрагмента по п. 1 in vivo для снижения скорости роста PRAME-экспрессирующих опухолей.
| WO 2011062634 A2, 26.05.2011 | |||
| WO 2016191246 A2, 01.12.2016 | |||
| АНТИГЕННАЯ КОМПОЗИЦИЯ И ЕЕ ТЕРАПЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ ГИБРИДНОГО БЕЛКА, А ТАКЖЕ СПОСОБ ПОЛУЧЕНИЯ БЕЛКА | 2013 |
|
RU2590701C2 |

