Изобретение относится к биотехнологии, а именно к способу пересборки (разборки и сборки) капсида бактериофага, в том числе бактериофага MS2, в том числе рекомбинантного. Способ включает два этапа: разборку капсида природного или рекомбинантного бактериофага, а затем сборку с различными токсическими ионами и их соединениями, например, таллий и другие элементы, в том числе радиоактивные изотопы и без них.
Изобретение позволяет использовать бактериофаги как эффективное средство доставки, в частности, солей таллия или пептидов, в организм реципиента.
Вирусоподобные частицы VLP - это монодисперсные трехмерные структуры, капсиды вирусов, лишенные вирусного генетического материала. Они самостоятельно собираются из множества копий одного или нескольких белковых мономеров в сложные супрамолекулярные структуры и обладают четко определенными внутренними и внешними поверхностями, а также интерфейсами между строительными блоками субъединиц. Обычно они имеют размеры, структуру и антигенность подобно нативным вирусам. Во внутреннем объеме, лишенном генетического материала исходного вируса и физически отделенном от внешней среды, можно размещать нужные молекулы для целевого перемещения в определенные места. Внешняя поверхность капсидов отвечает за клеточное взаимодействие как при инфицировании, так и при иммунологической защите. Интерфейс между субъединицами играет критическую роль в сборке и разборке частицы, влияя как на пути сборки, так и на морфологию получаемых частиц, а также стабильность получившихся структур каркаса.
VLP служат уникальной платформой для синтетической химии благодаря разнообразным реакционным группам, доступным на боковых цепях аминокислот поверхностных белков (например, амины, карбоксилаты, тиолы, амиды, фенолы). Большинство VLP имеют структурные модели с разрешением, близким к атомарному (рентгенокристаллические структуры или крио-ЭМ модели высокого разрешения), и основываясь на этой структурной информации можно предсказать и выбрать на капсиде место, куда следует ввести целевой белок (например, внутри, на границе или снаружи).
Еще одним важным свойством VLP является способность переносить нуклеиновые кислоты, защищая их от деградации нуклеазами.
Также, VLP нагружают лекарственными препаратами, токсичными для клеток (например, противоопухолевые препараты), и благодаря адресной доставке их высвобождение происходит в целевых клетках.
Бактериофаги являются вирусами бактерий, полностью безопасными для эукариот. Они хорошо изучены и удобны для генетических и биохимических манипуляций. На основе бактериофага MS2 создаются новые платформы для доставки различных веществ для терапевтических, иммунологических, генетических и других целей. Предложено много методов приготовления фагоподобных частиц, в том числе MS2 (PLPMS2), подходящих для различных целей. В основном, это создание генетических конструкций, несущих 2 гена CP-белка с последовательностью His-tag для упрощения очистки экспрессированных продуктов [Mikel et al., 2017].
Качество сборки фаговых частиц и их стабильность зависят от температуры, ионной силы и рН раствора, в котором проводят выделение, очистку и сборку фага. Температура плавления фага MS2 составляет 66°С [Plevka et al., 2009].
Частицы MS2 не слипаются в присутствии одновалентных катионов вплоть до концентрации 1,0 М. При этом в растворе с двухвалентными катионами, например, Са++, происходит быстрая агрегация частиц [Fu, Li 2016].
Подобран буфер, в котором наиболее успешно происходит очистка и самосборка рекомбинантных PLPMS2 [Hashemi et al., 2021]. Авторы отказались от ультрацентрифугирования в градиентах цезия или сахарозы, предпочитая осаждение преципитата ПЭГ 6000. Оказалось, что использование 1 М буфера Tris-base pH 8 с добавлением 100 mM NaNO3 при формировании PLPMS2 значительно улучшает качество и стабильность созданных частиц. Полученные таким образом PLP однородны по размерам и форме, и похожи на частицы фага MS2, также в них лучше сохранялась запакованная тестовая РНК.
Из патентов РФ №2735828 и №2599462 известен способ нагрузки бактериофага путем вакуумной сушки, когда проводили сушку смеси препарата фага с растворами таллия.
Метод вакуумной сушки не позволяет произвести модификацию капсида пептидами, а также контролировать упаковку таллия, уменьшить неспецифическую иммуногенность.
Наиболее близким аналогом является способ разборки-сборки бактериофага, известный из патента США №8987173. В известном способе фаг выдерживают 24 часа в буфере ST (50 mM трис, 100 mM NaCl) в присутствии 0,25 М ТМАО и 0,1 М соли таллия, затем раствор центрифугировают при 10,000 g 10 минут. Супернатант смешивали с ПЭГ6000 и NaCl до конечной концентрации 12,5% и 0,5 М, соответственно. Через 2 часа раствор осаждали центрифугированием (17,800 g в течении 45 минут) при 4°С. Осадок растворяли в минимальном количестве ST буфера и вновь осаждали при 10,000g в течении 10 минут. Надосадок фракционировали, и фракции, соответствующие интактным капсулам вируса MS2, собирали и после хранили в 4°С в ST буфере.
Данный способ также не позволяет в достаточной мере контролировать упаковку таллия уменьшить неспецифическую иммуногенность.
Патентуемый способ позволяет контролировать упаковку таллия, уменьшить неспецифическую иммуногенность препарата за счет обеспечения отсутствия РНК бактериофага.
Таким образом, технический результат заявленного изобретения заключается в очистке препарата вирусоподобной частицы бактериофага (ВПЧ) от РНК бактериофага с целью снижения неспецифической иммуногенности. Также технический результат заключается в контроле упаковке таллия, способ позволяет преодолеть барьер минимальной концентрации таллия при получении субстанции препарата, концентрация 0,16 мг/мл, что важно при использовании радиоактивных изотопов в вирусоподобные частицы. При этом заявленный способ позволяет получать рекомбинантные ВПЧ бактериофага.
Для достижения данного эффекта важен этап способа, где к концентрированному MS2 добавляют 2 объема предварительно охлажденной 100% уксусной кислоты. Смесь выдерживают в холодильнике при плюс 4°С в течение 30 минут. Колонку для обессоливания PD-10 подготавливают, уравновесив ее предварительно охлажденной уксусной кислотой на фосфатно-солевом буфере (ФСБ) с рН 7,4. Для этого, первоначально, сливают буфер из колонки. Затем вносят на колонку уксусную кислоту на ФСБ и производят центрифугирование в течение 2 минут при 1000 g. Процедуру повторяют 4 раза. По истечение 30 минут смесь бактериофага и уксусной кислоты осаждают путем центрифугирования при 14000 g в течение 10 минут в охлажденной до плюс 4°С центрифуге. Данный этап обеспечивает разрушение капсида бактериофага до мономерных капсидных белков. Также разрушается и РНК бактериофага.
Данная методика позволяет преодолеть барьер минимальной концентрации таллия при получении субстанции препарата, что важно при использовании радиоактивных изотопов в вирусоподобные частицы. Для достижения данного эффекта важен этап, когда концентрат разобранного MS2 выдерживают в течение 1-24 часов при плюс 4°С в присутствии 25 мМ ТМАО, 10 мМ тРНК дрожжей и раствора таллия, с концентрацией 0,16 мг/мл. Для этого в эппендорфе смешать 2:1 разобранного MS2, 221 мкл 1,8 М ТМАО, 355 мкл тРНК дрожжей, с концентрацией 1 мг/мл, и 27,2 мкл раствора таллия, с концентрацией 1,6 мг/мл.
В результате применения разработанной нами методики представляется возможным получение препарата, обладающего цитотоксическими свойствами за счет присутствия таллия внутри вирусоподобной частицы на основе бактериофага MS2. Способы синтеза PLPMS2 разняться в зависимости от цели их применения.
Для возможности проникновения частиц фага MS2 в клетки эукариот на поверхность капсида химическим методом присоединяются пептиды СРР (cell-penetratedpeptide) из вируса HIV-1, а именно, пептид ТАТ 47-57 с последовательностью YGRKKRRQRRR [Pan et al., 2012].
В работе [Wangetal., 2016] пептид ТАТ присоединяли непосредственно к мономерам и димерам CP белка фага MS2, а далее наблюдали самосборку характерного для MS2 капсида, причем участки ТАТ были ориентированы наружу.
Созданы генетические конструкции, в которых последовательности одноцепочечных вариабельных фрагментов антител (scFvs) слиты с последовательностью белка CP, такие плазмиды экспрессированы, и получившиеся в результате рекомбинантные белки собрались в PLP-частицы, причем scFvs экспонировались на поверхности фаговых частиц [Lino et al., 2017].
Материалы и методы
Бактериофаг MS2 относится к семейству Leviviridae. Это фаг с одноцепочечной смысловой РНК, он заражает только клетки Escherichiacoli, несущие фактор фертильности (Hfr-штаммы или штаммы с плазмидой F'). F-пили являются рецепторами фага [Fu, Li 2016].
Интактная фаговая частица состоит из молекулы РНК, 180 молекул белка оболочки CP и 1 молекулы белка созревания (А-белка), ответственного за прикрепление фага к F-пилям Escherichiacoli. Капсид MS2 икосаэдрической формы с триангуляционным числом равным 3 и имеет размеры от 27 до 34 нм.
Использование бактериофага MS2 в качестве средства доставки различных веществ к клеткам признано очень удобным. Фаг не является инфекционным агентом для эукариот. Фаговые частицы легко создаются с помощью технологии рекомбинантных белков. В капсиды MS2 можно упаковать целевую ДНК или РНК pac-сайтом на 5'-конце, и тем защитить препараты от нуклеазной деградации. Давая возможность эффективно и удобно упаковать все виды веществ и обеспечивая доставку в целевые клетки, частицы фага MS2 имеют большие перспективы в качестве новой платформы для терапевтических, иммунологических и диагностических целей.
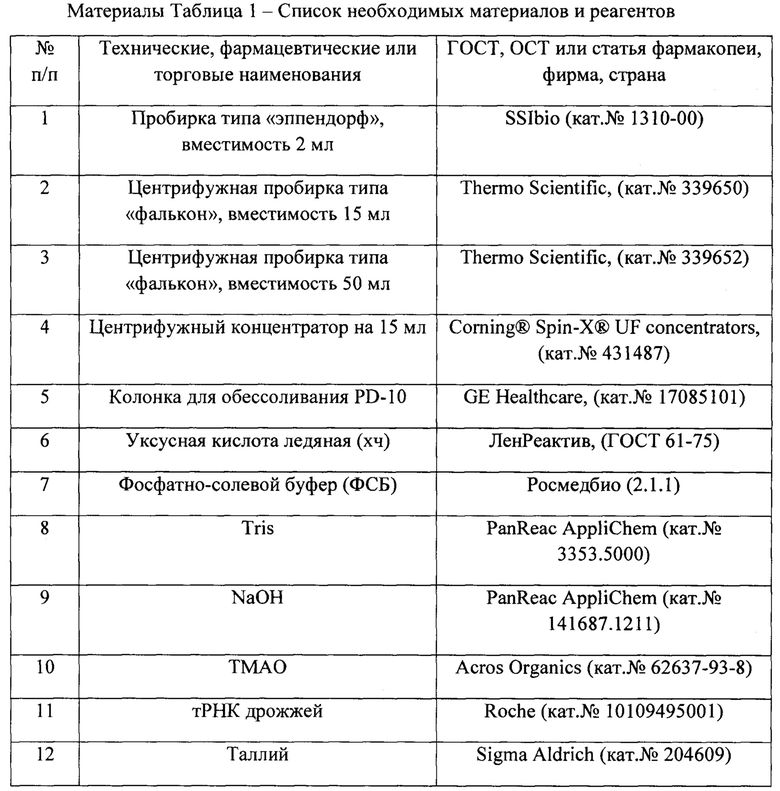

Допускается применение других средств измерения, вспомогательных устройств, посуды, реактивов и материалов с метрологическими и техническими характеристиками не хуже, чем у приведенных.


Приготовление растворов, проведение анализа проводилось при следующих условиях:
- температура окружающего воздуха - плюс (20±5)°С;
- атмосферное давление - (от 84,0 до 106,7) кПа;
- относительная влажность воздуха - не более 80%.
Перед проведением анализа проводят следующие работы: подготовку стеклянной посуды, приготовление растворов необходимых реактивов.
Посуду, используемую для приготовления растворов, замачивают на 3-4 часа в свежеприготовленном 3% растворе клиндезина-специаль (3 мл на 100 мл концентрированной серной кислоты) и отмывают в проточной воде с последующим ополаскиванием дистиллированной водой и высушиванием в сушильном шкафу при температуре плюс 150°С.
Подготовка используемых реагентов:
а) Уксусную кислоту предварительно охладить при плюс 4°С.
б) Подготовить фосфатно-солевой буферный раствор (ФСБ) с рН 7,4. Для этого 5 таблеток ФСБ растворить в 500 мл дистиллированной воды.
в) Для получения 1 мМ уксусной кислоты на 50 мл ФСБ добавить 3 мкл концентрированной уксусной кислоты.
г) Приготовить 50 мл 50 мМ Tris (рН 7). Для этого 1,25 мл 2 М Tris разбавить 48,75 мл дистиллированной воды.
д) Приготовить 50 мл 50 мМ Tris (рН 8,8). Для этого 1,25 мл 2 М Tris разбавить 48,75 мл дистиллированной воды. Довести рН до 8 с помощью 10М NaOH.
е) Для приготовления 5 мл 1,8 М ТМАО на 50 мМ Tris использовать навеску в 1 г.
ж) Подготовить насыщенный раствор таллия: 0,955 г таллия растворить в 10 мл дистиллированной воды. Для растворения поместить фалькон на водяную баню, предварительно прогретую до плюс 60°С. Выдерживать на бане до полного растворения таллия. Полученный насыщенный раствор имеет концентрацию 96 мг/мл.
з) Из насыщенного раствора таллия подготовить стоковый раствор с концентрацией 1,6 мг/мл (т.е. развести насыщенный раствор в 60 раз).
и) Подготовить тРНК дрожжей на 50 мМ Tris с концентрацией 1 мг/мл.
Структура пептидов, использованных для модификации капсида ВПЧ:
Пептид iRGD-P1 первичной структуры GGGcyclo(CRGDKGPDC) - пептид №1
Пептид iRGD-P2 первичной структуры GGGcyclo(CRGDRGPDC) - пептид №2
Пептид iRGD-РЗ первичной структуры GGGcyclo(CRGDKGPEC) - пептид №3
Пептид iRGD-P4 первичной структуры GGGcyclo(CRGDRGPEC) - пептид №4
Далее решение поясняется с помощью примеров. Пример 1
Получение фага MS2
Получение высокого титра фага MS2 проводили по методу, описанному ранее (Княжев В.А., Сивов И.Г., Сергиенко В.И. РНК-трансдукция неинфекционными вирионами фага MS2 // Молекулярная генетика, микробиология и вирусология 2002, №3, стр. 56-63). Контролями служили секвенирование ОТ-ПЦР фрагмента геномной РНК, а также определение титра методом агаровых слоев (Gratia A. Numerical Relations between Lysogenic Bacteria and Particles of Bacteriophage // Ann. Inst. Pasteur 1936, v.57, p.652). Концентрирование вирионов фага, проводили после осветления клеточного лизата Стр.: 8 RU 2 599 462 С1 5 10 15 20 25 30 35 40 45 центрифугированием (15000 об/мин) с последующим осаждением ПЭГ-6000 в присутствии NaCl, как известно специалистам. Концентрирование препарата фага так же проводили обезвоживанием сухим сефадексом G-10.
Пример 2
Процедура разборки/сборки бактериофага
MS2 (с концентрацией 5×1011 БОЕ) сконцентрируют до 1 мл с помощью центрифужного концентратора на 15 мл в течение часа при плюс 4°С, 3600 об/мин.
К 1 мл концентрированного MS2 добавляют 2 объема предварительно охлажденной до температуры 4°С 100% уксусной кислоты (2 мл). Смесь выдерживают в холодильнике при плюс 4°С в течение 30 минут. Готовят колонку для обессоливания PD-10, уравновесив ее предварительно охлажденной 1 мМ уксусной кислотой на фосфатносолевом буфере (ФСБ) с рН 7,4. Для этого, первоначально, сливают буфер из колонки. Затем вносят на колонку 4 мл 1 мМ уксусной кислоты на ФСБ и производят центрифугирование в течение 2 минут при 1000 g. Процедуру повторяют 4 раза.
По истечение 30 минут смесь бактериофага и уксусной кислоты осаждают путем центрифугирования при 14000 g в течение 10 минут в охлажденной до плюс 4°С центрифуге. Супернатант после осаждения наносят на предварительно подготовленную колонку для обессоливания и производят центрифугирование в течение 2 минут при 1000 g. После чего собрают проскок, содержащий разобранный бактериофаг.
5 мл разобранного бактериофага разбавляют в 3 раз 50 мМ Tris с рН 7. Доводят рН до 7,3 с помощью добавления 10 М NaOH. 15 мл разобранного MS2 концентрируют в 7,5 раз с помощью центрифужного концентратора на 15 мл в течение 45 минут при 3600 об/мин в охлажденной до плюс 4°С центрифуге. Таким образом, получают 2 мл разобранного бактериофага.
Первую половину (1 мл) концентрата разобранного MS2 выдерживают в течение 24 часов при плюс 4°С в присутствии 25 мМ ТМАО на 50 мМ Tris с рН 7, 10 мМ тРНК дрожжей на 50 мМ Tris с рН 7. Для этого в эппендорфе на 2 мл смешивают 1 мл разобранного MS2, 221 мкл 1,8 М ТМАО, 355 мкл тРНК дрожжей (с концентрацией 1 мг/мл).
Вторую половину (1 мл) концентрата разобранного MS2 выдерживают в течение 24 часов при плюс 4°С в присутствии 25 мМ ТМАО, 10 мМ тРНК дрожжей и раствора таллия (с концентрацией 0,16 мг/мл). Для этого в эппендорфе на 2 мл смешивают 1 мл разобранного MS2, 221 мкл 1,8 М ТМАО, 355 мкл тРНК дрожжей (с концентрацией 1 мг/мл) и 27,2 мкл раствора таллия (с концентрацией 1,6 мг/мл). По истечение 24 часов пробы с собранным бактериофагом обессоливают с помощью колонок PD-10, предварительно уравновешенных 50 мМ Tris с рН 8,8. Таким образом, в ходе проведенных работ получают 1 мл бактериофага, собранного без таллия и 1 мл - собранного с таллием.
Пример 3
Модификация поверхности вирионов пептидами
А. Синтез пептидов со структурой (NH2)GGGCRGDK/RGPD/EC(COOH). Пептид синтезировали методом твердофазного пептидного синтеза исходя из Fmoc аминокислот на автоматическом пептидном синтезаторе 433А Applied Biosystems на смоле с присоединенным остатком Fmoc-Cys(Acm). Использовали следующие производные аминокислот: Fmoc-Cys(Acm), Fmoc-Arg(Pbf), Fmoc-Asp(OtBu), Fmoc-Gly, Fmoc-Lys(Boc), Fmoc-Pro. Снятие Fmoc-защитной группы с N-концевой альфааминогруппы растущей пептидной цепи проводили 22%-ным раствором 4-метилпиперидина в N-диметилформамиде (Алешина Е.Ю., Пындык Н.В., Мойса А.А., Санжаков М.А., Харыбин О.Н., Николаев Е.Н., Колесанова Е.Ф. Синтез фрагмента Р-амилоида 5RHDSGY10 и его изомеров. Биомед. химия, 2008, т. 54, №2, 154-166). Присоединение аминокислот к растущей пептидной цепи (кроме Fmoc-Cys(Acm)) осуществляли с предварительной активацией Fmoc-аминокислот гексафторфосфатом 2-(1Н-бензотриазол-1-ил)-1,1,3,3-тетраметиламиния в присутствии 1-гидроксибензотриазола и 2,4,6-коллидина согласно процедуре FastMoc, описанной в инструкции к синтезатору. Fmoc-Cys(Acm) присоединяли с активацией in situ, используя в качестве активатора диизопропилкарбодиимид в присутствии 1-гидроксибензотриазола. По окончании синтеза пептид снимали со смолы обработкой смесью трифторуксусной кислоты, три-(изопропил)-силана, 3,6-диокса-1,8-октандитиола и воды (в объемном соотношении 94:1:2,5:2,5) и осаждали метил-трет-бутиловым эфиром. Осадок пептида растворяли в 10%-ном водном ацетонитриле с 0,1% трифторуксусной кислоты, и полученный раствор подвергали очистке методом ВЭЖХ на колонке Zorbax SB-C8, 21,2×250 мм, 7 мкм в градиенте концентрации ацетонитрила в 0,1% водном растворе трифторуксусной кислоты. Фракцию, содержащую целевой пептид, собирали и упаривали под вакуумом и затем проводили удаление защитных Acm-групп с остатков цистеина с одновременным формированием дисульфидного мостика по известной методике (Fernando Albericio et al. Preparation and handling of peptides containing methionine and cysteine // In: Fmoc Solid Phase Peptide Synthesis: A Practical Approach. Eds. W.C. Chang and P.D. White. Oxford University Press, 2000). Пептид подвергали повторной очистке методом ВЭЖХ на той же колонке, фракцию целевого пептида упаривали под вакуумом.
Б. Чистоту препарата синтезированного пептида подтверждали массспектрометрическим анализом с ионизацией электрораспылением и детекцией методом ионной ловушки, а также аналитической ВЭЖХ в соответствии с протоколами, описанными ранее (M.H.V. Van Regenmortel, S. Muller. Synthetic peptides as antigens. Elsevier 1999, pp. 88-90).
В. Конъюгацию пептида с фаговыми частицами проводили с использованием диметиладипимидата в соответствии со стандартной процедурой, известной специалистам (M.H.V. Van Regenmortel, S. Muller. Synthetic peptides as antigens. Elsevier, 1999, pp.88-90).
Пример 4
Создание вирусоподбной таргетной частицы на основе бактериофага MS2
1) Разборка вирусных капсидов MS2
MS2 (с концентрацией 5×1011 БОЕ) сконцентрировали до 1 мл с помощью центрифужного концентратора на 15 мл (Corning 431487) в течение часа при плюс 4°С, 3600 об/мин.
К 1 объему концентрированного MS2 добавили 2 объема предварительно охлажденной 100% уксусной кислоты. Смесь выдерживали
в холодильнике при плюс 4°С в течение 30 минут.
Подготовили колонку для обессоливания PD-10 (GEHealthcare, Германия), уравновесив ее предварительно охлажденной 1 мМ уксусной кислотой (Ленреактив, Россия) на фосфатно-солевом буфере (PBS), (Росмедбио, Россия) с рН 7,4. Для этого, первоначально, слили буфер из колонки. Затем внесли на колонку 4 объема 1 мМ уксусной кислоты на PBS и произвели центрифугирование в течение 2 минут при 1000 g. Процедуру повторили 4 раза.
По истечение 30 минут смесь бактериофага и уксусной кислоты осадили путем центрифугирования при 14000 g в течение 10 минут в охлажденной до плюс 4°С центрифуге.
Супернатант после осаждения нанесли на предварительно подготовленную колонку для обессоливания и произвели центрифугирование в течение 2 минут при 1000 g. После чего собрали проскок, содержащий мономер капсидного белка бактериофага.
2) Получение рекомбинантного капсидного белка, конъюгация с пептидом IRGD или другими таргетными пептидными молекулами
Получение плазмиды pL1118 на основе вектора pR1504 (3), содержащей нуклеотидную последовательность гена капсидного белка MS2 из Escherichia virus и 6-His в N-концевой области.
Полученный штамм продуцент использовали для наработки рекомбинатного белка MS2 в клетках Escherichia coli.
3) Получение рекомбинантного капсидного белка сшитого с пептидом IRGD или другими таргетными пептидными молекулами
Получение плазмиды pL1120 на основе вектора pR1504 (3), содержащей нуклеотидную последовательность гена капсидного белка MS2 из Escherichia virus, участок связывания с клеткой RGD в С-концевой области и 6-His в N-концевой области.
Получение плазмиды pL1145 на основе вектора pR1504 (3), содержащей нуклеотидную последовательность гена капсидного белка MS2 из Escherichia virus, участок связывания с клеткой RGD в петле нуклеотидной последовательности капсидного белка MS2 и 6-His в N-концевой области.
Получение плазмиды pL1146 на основе вектора pR1504 (3), содержащей нуклеотидную последовательность гена капсидного белка MS2 из Escherichia virus, участок связывания с клеткой RGD в петле нуклеотидной последовательности капсидного белка MS2 и 6-His в N-концевой области.
Полученные штаммы продуценты использовали для наработки рекомбинатного белка MS2 сшитого с таргетной молекулой в клетках Escherichia coli.
4) Обработка перед сборкой
Проводили перевод рекомбинантного капсидного белка 1118/1120/1145/1146 в мономерную форму.
К 1 объему очищенного рекомбинантного капсидного белка 1118/1120 добавили 2 объема предварительно охлажденной 100% уксусной кислоты. Смесь выдерживали в холодильнике при плюс 4°С в течение 30 минут.
Подготовили колонку для обессоливания PD-10 (GEHealthcare, Германия), уравновесив ее предварительно охлажденной 1 мМ уксусной кислотой (Ленреактив, Россия) на фосфатно-солевом буфере (PBS), (Росмедбио, Россия) с рН 7,4. Для этого, первоначально, слили буфер из колонки. Затем внесли на колонку 4 мл 1 мМ уксусной кислоты на PBS и произвели центрифугирование в течение 2 минут при 1000 g. Процедуру повторили 4 раза.
По истечение 30 минут смесь рекомбинантного капсидного белка 1118/1120 и уксусной кислоты осадили путем центрифугирования при 14000 g в течение 10 минут в охлажденной до плюс 4°С центрифуге.
Супернатант после осаждения нанесли на предварительно подготовленную колонку для обессоливания и произвели центрифугирование в течение 2 минут при 1000 g. После чего собрали проскок, содержащий рекомбинантный капсидный белок 1118/1120 в виде мономера.
Пример 5
Создание вирусоподобной таргетной частицы на основе бактериофага MS2 согласно примеру 4, где на стадиях 2) и 3) использован пептид ТАТ 47-57 с последовательностью YGRKKRRQRRR.
Пример 6
Упаковка в вирусоподобные частицы, полученные по примеру 4 и 5
1) упаковка таллия
5 мл разобранного бактериофага разбавили в 3 раз 50 мМ Tris с рН 7. Довели рН до 7,3 с помощью добавления 10М NaOH.
15 мл разобранного MS2 сконцентрировали в 7,5 раз с помощью центрифужного концентратора на 15 мл (Corning 431487) в течение 45 минут при 3600 об/мин в охлажденной до плюс 4°С центрифуге. Таким образом, получили 2 мл разобранного бактериофага.
(1 мл) концентрата разобранного MS2 выдерживали в течение 24 часов при плюс 4°С в присутствии 25 мМ ТМАО, 10 мМ тРНК дрожжей и раствора таллия (SigmaAldrich, США)
(с концентрацией 0,16 мг/мл) или РНК (конц. 50 нг/мл и 5 нг/мл)
По истечение 1-24 часов пробы с собранным бактериофагом обессоливали с помощью колонок PD-10, предварительно уравновешенных 50 мМ Tris с рН 8,8. Таким образом, получили 1 мл вирусоподобных частиц с упакованным в них таллием.
2) упаковка радиоактивного изотопа таллия
На этапе разборки вирусоподобных частиц бактериофага MS2 добавляют 2 объема охлажденной ледяной уксусной кислоты к 1 объему бактериофага до конечной концентрации уксусной кислоты 66%. Раствор выдерживают на льду в течение 30 минут. На этапе сборки осаждают центрифугированием при 10 000 × g в течение 10 минут нуклеиновые кислоты. Отбирают супернатант, добавляют фосфатный буфер в присутствии 0.25М ТМАО и соли таллия (конц. 1 мг/мл и 0,1 мг/мл) и РНК (конц. 50 нг/мл и 5 нг/мл) выдерживают 24 часа.
Степень упаковки таллия в бактериофаг определяли методом ИСП-АЭС (Табл. 3).

3) упаковка иных изотопов
Пример 7
Определение чувствительности эндотелиальных и других клеточных культур к модифицированным вирионам, наполненных таллием, и агглютинация клеток модифицированными вирионами.
Клеточные культуры эндотелиальных клеток получали из коллекции НИИ канцерогенеза РОНЦ и размножали согласно способам, известным, например, из патента РФ 2359030 и РФ 2493251. Индукцию образования интегринов на поверхности эндотелиальных клеток и клеток ЗСО проводили эндотелиальным фактором роста (VEGF), согласно описанной процедуре (патент РФ 2377017). Агглютинацию клеток проводили согласно описанной процедуре (US Patent 5401636; US Patent 5541417). Проникновение вирионов фага MS2, наполненных таллием, фиксировали при помощи флуоресцентной микроскопии с красителем PTSA по процедуре, описанной ранее (патент РФ 2305270). Расчет LD50 проводили с помощью программы Probit_Analysis. Ниже приводятся полученные результаты: для HBL100=11,2 (количество частиц на одну клетку вызывающее гибель в 50% случаев); для HepG2=6,1; для MCF10A=4,8; для MCF7=3,2.
Пример 8
Определение чувствительности животных к 1010 поверхностно модифицированным вирионам фага MS2, наполненных солями одновалентного таллия.
Эксперименты выполнены на 130 белых неинбредных крысах обоего пола массой 200-400 г. и 263 белых неинбредных мышах обоего пола массой 17-36 г., содержащихся в условиях вивария с естественным световым режимом на стандартной диете (ГОСТ Р 50258-92), согласно методическим руководствам и нормативным документам - ГОСТ 3 51000.3-96 и 51000.4-96; правила и Международные рекомендации Европейской конвенции по защите позвоночных животных, используемых при экспериментальных исследованиях (1997); правила лабораторной практики (GLP) в РФ, утвержденные приказом Минздрава РФ от 19 июня 2003 г. №267. Погибших животных при трехкратном введении дозы 108 поверхностно модифицированных вирионов фага MS2, наполненных солями одновалентного таллия, не было. Аналогичный эксперимент с новорожденными животными не выявил токсичности в указанной дозе. Предлагаемый способ полисигнальной активации апоптоза клеток ЗСО позволяет обеспечить комплексное, эффективное пролонгированное цитоксическое воздействие на очаговые и метастатические скопления клеток ЗСО при минимизации нежелательных побочных воздействий на здоровые клетки организма и может быть реализован в амбулаторных условиях на базе имеющегося оборудования.
Пример 9
Конъюгация пептида с ВПЧ
Конъюгацию пептида с ВПЧ проводили с использованием диметиладипимидата в соответствии со описанной ниже процедурой. Раствор 5 мкг пептида в 0.1 мл PBS перемешивали с 25 мкг фага 30 мин, затем в течение 1 ч добавляли 0.25 мл 0.25% водного глутарового альдегида. Реакционную массу перемешивали 15 ч, а затем диализовали против PBS с трехкратной сменой буфера. N,N-бис(4-метилфенилсульфонил)-адипамид получали смешивая 10 мМ (СН2)4(СООН)2 с 20 мМ пара-толуенсульфонамидом и избытком оксихлоридом фосфора, инкубировали смесь 1 час на кипящей водяной бане.
Осадок получали фильтрованием и промывали горячим этиловым спиртом. Полученное соединение использовали для конъюгации мономерной формы пептида с фагом.
Пептид (30 мкМ) растворяли в безводном-диоксане с избытком изобутил хлороформата и триэтиламина. Раствор перемешивали при комнатной температуре 30 минут. 50 мкг фага растворили в 30 мл в 0.2 М боратном буфере рН=8. Раствор охлаждали льдом. Для активации пептида в растворе с фагом, 2 мл р-диоксана были добавлены к раствору. Сложение р-диоксана к белковому раствору вызвало небольшую мутность. Активизированный раствор пептида добавляли к фагу по каплям с перемешиванием. Перемешивание было продолжено на льду в течение 30 минут к часу. Чтобы удалить непрореагировавшие молекулы фаг осаждали ледяным 100%-ым этанолом. Преципитат осаждали центрифугированием при 4°С в течение 10 минут при 4500 g. Супернатант, содержащий непрореагировавшие малые молекулы, удаляли. Осадок повторно суспендировали холодным этанолом три раза и осаждали центрифугированием. В заключении, осадок разводили дистиллированной водой до концентрации около 5 мг белка/мл.
Пептид растворяли в безводном DMF и реакцию проводили под азотом. После добавления изобутилхлороформата и tri-n-butylamine смесь размешивалась в 4°С в течение 10 минут, затем в комнатной температуре в течение еще 20 минут. Приблизительно 1 мл активизированного раствора для пептида (содержащий приблизительно 2.5 μmol) был добавлен к каждому разведению ВПЧ (буфер бората мл на 14 мг/2) и размешан в 4-10°С для 1 h. Перемешивание было продолжено в комнатной температуре быстро. Конъюгаты очищали диализом в фосфатносолевом растворе буфере(РВ8). Конъюгаты анализировали по содержанию белка, разливали по небольшим объемам (25 мкл) и хранили при -80°С до использования.
Далее проводили реакцию между амидными группами (высокоэффективная). Реакция была выполнена с перемешиванием под азотом в колбе, охлажденной в водяной бане (приблизительно 19-20°С). Нитрит бутила (417 μL 30 mM в безводном ДМСО) был добавлен каплями в общем объеме 12.5 μmol к фаголизату (3.0 мкг). Для контроля замерзания реакции, которое происходит при 18°С, добавляли 12.5 μmol раствор H2SO4 (625 μL 20 mM в безводном ДМСО). Общий объем реакции не превышал 4.2 мл. После 30 минут раствор с диазонием разделяли. Малое количество из смеси добавили к избытку раствора n-крезолав 0.2М борнокислом буфере рН=8.7. Эту смесь перемешивали при комнатной температуре в течение 3 часов, концентрировали в вакууме, а затем подвергали подготовительному HPLC (гексан/ацетон 9:1). Были выделены несколько изоамирфных продуктов желтого цвета, с характеристиками: EI-MSm/z (относительная интенсивность) М+420 (8), 424 (3), 422 (8), 389 (1), 387 (4), 385 (7), 135 (16), 107 (100). Последний из указанных продуктов (пик 107) соли диазония разделен на две равные части*. Каждая половина (по 2.5 мл) была добавлена каплями к энергично размешиваемому раствору фага - носителя в 0.2М борнокислом буфере рН=8.7 (120 мкг фаголизата в 3 мл буфере), при 19-20°С. После двухчасовой инкубации при комнатной температуре смесь быстро перемешали. Желтый осадок конъюгата очищали диализом с PBS.
После модификации поверхности ВПЧ на основе бактериофага MS2 пептидами и завершения сборки получили модифицированный фаг, состоящий из оболочки, содержащей циклический лиганд iRGD, ковалентно связанный с оболочкой, и сердцевины, содержащей геномную РНК вириона и соль таллия.
Эффективность коньюгации оценивали посредством определения количества пептида P1 (iRGD-P1 первичной структуры GGGcyclo(CRGDKGPDC)), конъюгированного с ВПЧ на основе MS2, методом Эллмана (определение свободных SH-групп).
Чистота препарата подтверждена аналитической ВЭЖХ на обращенной фазе и масс-спектрометрическим анализом с ионизацией электрораспылением (рабочая станция Agilent 1100, колонка YMC-Pack С4 150 × 3.0 mm ID, S - 5 μm, градиент ацетонитрила 2%-98% за 10 мин в 0,3%-ной гептафтормасляной кислоте, скорость элюции 0,7 мл/мин; масс-спектрометрический детектор IonTrap LC7MS 6340 (Agilent)).
Пример 10
Осуществляют аналогично примеру 9, используя пептид iRGD-P2 первичной структуры GGGcyclo(CRGDRGPDC).
Пример 11
Осуществляют аналогично примеру 9, используя пептид iRGD-Р3 первичной структуры GGGcyclo(CRGDKGPEC).
Пример 12
Осуществляют аналогично примеру 9, используя пептид iRGD-P4 первичной структуры GGGcyclo(CRGDRGPEC)
ВПЧ бактериофагов, полученные способами по изобретения могут представлять собой ВПЧ, содержащие следующие последовательности и иметь следующие характеристики:
Вариант 6His-MS2 (pL1144

Вариант 6His-RGD1-MS2 (pL1145)

Вариант 6His-RGD2-MS2 (pL1146)

Вариант 6His-MS2 (pL1118)

Вариант 6His-MS2-IRGD (pL1120)


Молекулярная масса белка 6His-MS2-RGD3 составляет 15,8 кДа.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения вирусоподобной частицы бактериофага | 2024 |
|
RU2821694C1 |
| Ректальные суппозитории на основе модифицированного бактериофага MS2 для таргетной терапии злокачественных солидных опухолей | 2019 |
|
RU2735828C1 |
| СПОСОБ ПОЛИСИГНАЛЬНОЙ АКТИВАЦИИ АПОПТОЗА КЛЕТОК ЗЛОКАЧЕСТВЕННЫХ СОЛИДНЫХ ОПУХОЛЕЙ | 2015 |
|
RU2599462C1 |
| Онколитический способ терапии рака молочной железы. | 2018 |
|
RU2695136C1 |
| СПОСОБ ПОЛУЧЕНИЯ ЧАСТИЦ БАКТЕРИОФАГОВ СЕМЕЙСТВА LEVIVIRUS | 2020 |
|
RU2811106C2 |
| СПОСОБ ОЧИСТКИ БАКТЕРИОФАГОВ | 2011 |
|
RU2590609C2 |
| Способ очистки нитчатого бактериофага М13 | 2015 |
|
RU2610178C1 |
| БЕЛКИ, СЛИТЫЕ С КОШАЧЬИМ АЛЛЕРГЕНОМ, И ИХ ПРИМЕНЕНИЕ | 2006 |
|
RU2414239C2 |
| КОНЪЮГАТЫ ИНТЕРЛЕЙКИНА-1 И ИХ ПРИМЕНЕНИЕ | 2006 |
|
RU2441067C2 |
| КОМПОЗИЦИЯ И СПОСОБ ДЛЯ ИММУНИЗАЦИИ, СПОСОБ ПРОДУЦИРОВАНИЯ НЕПРИРОДНОГО, УПОРЯДОЧЕННОГО И ПОВТОРЯЮЩЕГОСЯ МАССИВА АНТИГЕНОВ | 2002 |
|
RU2294211C2 |
Изобретение относится к биотехнологии. Представлен способ пересборки (разборки и сборки) капсида бактериофага, в том числе бактериофага MS2, в том числе рекомбинантного. Способ включает два этапа: разборку капсида природного или рекомбинантного бактериофага, а затем сборку с различными токсическими ионами и их соединениями, например, таллий и другие элементы, в том числе радиоактивные изотопы и без них. Также представлен способ получения ВПЧ бактериофага, включающий: бактериофаг концентрируют до требуемого объема; к одному объему бактериофага, сконцентрированного на стадии А), добавляют два объема предварительно охлажденной 100% уксусной кислоты; смесь, полученную на этапе Б), выдерживают в холодильнике при плюс 4°С в течение 30 мин; по истечении 30 мин смесь бактериофага и уксусной кислоты осаждают путем центрифугирования при 14000 g в течение 10 мин в охлажденной до плюс 4°С центрифуге; супернатант после осаждения наносят на предварительно подготовленную колонку для обессоливания и производят центрифугирование в течение 2 мин при 1000 g; собирают проскок, содержащий разобранный бактериофаг; разобранный бактериофаг разбавляют в 3 раза 50 мМ Tris с рН 7; смесь, полученную на этапе Ж), доводят до рН 7,3 с помощью добавления 10 М NaOH; смесь, полученную на этапе Ж), концентрируют в 7,5 раз с помощью центрифужного концентратора при 3600 об/мин в охлажденной до плюс 4°С центрифуге; полученный концентрат разобранного бактериофага делят на две части; одну часть концентрата разобранного бактериофага выдерживают в течение 24 ч при плюс 4°С в присутствии 25 мМ ТМАО на 50 мМ Tris с рН 7,10 мМ тРНК дрожжей с концентрацией 1 мг/мл на 50 мМ Tris с рН 7; вторую часть концентрата разобранного бактериофага выдерживают в течение 24 ч при плюс 4°С в присутствии 25 мМ ТМАО, 10 мМ тРНК дрожжей с концентрацией 1 мг/мл и раствора таллия с концентрацией 0,16 мг/мл; по истечении 24 ч пробы с собранным бактериофагом обессоливают, таким образом, получая ВПЧ бактериофага. При этом ВПЧ бактериофага может иметь модифицированный пептидом капсид, в частности пептидом IRGD или пептидом YGRKKRRQRRR, а также может быть получена из рекомбинантных капсидных белков с дальнейшей модификацией - присоединением пептида методом конъюгации. ВПЧ также могут быть получены из рекомбинантных капсидных белков с пришитым пептидом, а таллий представлять собой радиоактивный изотоп. Изобретение позволяет использовать бактериофаги как эффективное средство доставки, в частности, солей таллия или пептидов в организм реципиента. Данный способ также не позволяет в достаточной мере контролировать упаковку таллия, уменьшить неспецифическую иммуногенность. Патентуемый способ позволяет контролировать упаковку таллия, уменьшить неспецифическую иммуногенность препарата за счет обеспечения отсутствия РНК бактериофага. Таким образом, технический результат заявленного изобретения заключается в очистке препарата вирусоподобной частицы бактериофага (ВПЧ) от РНК бактериофага с целью снижения неспецифической иммуногенности. Также технический результат заключается в контроле упаковки таллия, способ позволяет преодолеть барьер минимальной концентрации таллия при получении субстанции препарата, концентрация 0,16 мг/мл, что важно при использовании радиоактивных изотопов в вирусоподобных частицах. При этом заявленный способ позволяет получать рекомбинантные ВПЧ бактериофага. 2 н. и 10 з.п. ф-лы, 3 табл., 12 пр.
1. Способ получения ВПЧ бактериофага, характеризующийся тем, что включает следующие стадии:
A) концентрируют бактериофаг до требуемого объема;
Б) к одному объему бактериофага, сконцентрированного на стадии А), добавляют два объема предварительно охлажденной 100% уксусной кислоты;
B) смесь, полученную на этапе Б), выдерживают в холодильнике при плюс 4°С в течение 30 мин;
Г) по истечении 30 мин смесь бактериофага и уксусной кислоты осаждают путем центрифугирования при 14000 g в течение 10 мин в охлажденной до плюс 4°С центрифуге;
Д) супернатант после осаждения наносят на предварительно подготовленную колонку для обессоливания и производят центрифугирование в течение 2 мин при 1000 g;
Е) собирают проскок, содержащий разобранный бактериофаг;
Ж) разобранный бактериофаг разбавляют в 3 раза 50 мМ Tris с рН 7;
З) смесь, полученную на этапе Ж), доводят до рН 7,3 с помощью добавления 10 М NaOH;
И) смесь, полученную на этапе Ж), концентрируют в 7,5 раз с помощью центрифужного концентратора при 3600 об/мин в охлажденной до плюс 4°С центрифуге;
К) полученный концентрат разобранного бактериофага делят на две части;
Л) одну часть концентрата разобранного бактериофага выдерживают в течение 24 ч при плюс 4°С в присутствии 25 мМ ТМАО на 50 мМ Tris с рН 7, 10 мМ тРНК дрожжей с концентрацией 1 мг/мл на 50 мМ Tris с рН 7; вторую часть концентрата разобранного бактериофага выдерживают в течение 24 ч при плюс 4°С в присутствии 25 мМ ТМАО, 10 мМ тРНК дрожжей с концентрацией 1 мг/мл и раствора радиоактивного металла с концентрацией 0,16 мг/мл;
М) по истечении 24 ч пробы с собранным бактериофагом обессоливают, таким образом, получая ВПЧ бактериофага.
2. Способ по п. 1, характеризующийся тем, что пептид является пептидом iRGD или пептидом YGRKKRRQRRR.
3. Способ по п. 2, характеризующийся тем, что пептид iRGD представляет собой пептид: первичной структуры GGGcyclo(CRGDKGPDC), первичной структуры GGGcyclo(CRGDRGPDC), первичной структуры GGGcyclo(CRGDKGPEC) или первичной структуры GGGcyclo(CRGDRGPEC).
4. Способ по любому из пп. 2, 3, характеризующийся тем, что ВПЧ получают из рекомбинантных капсидных белков с дальнейшей модификацией - присоединением пептида методом конъюгации.
5. Способ по любому из пп. 2-4, характеризующийся тем, что ВПЧ получают из рекомбинантных капсидных белков с пришитым пептидом.
6. Способ по любому из пп. 1, 2, характеризующийся тем, что радиоактивный металл представляет собой таллий, радий или технеций.
7. Способ получения модифицированного ВПЧ бактериофага, характеризующийся тем, что включает следующие стадии:
A) концентрируют бактериофаг до требуемого объема;
Б) к одному объему бактериофага, сконцентрированного на стадии А), добавляют два объема предварительно охлажденной 100% уксусной кислоты;
B) смесь, полученную на этапе Б), выдерживают в холодильнике при плюс 4°С в течение 30 мин;
Г) по истечении 30 мин смесь бактериофага и уксусной кислоты осаждают путем центрифугирования при 14000 g в течение 10 мин в охлажденной до плюс 4°С центрифуге;
Д) супернатант после осаждения наносят на предварительно подготовленную колонку для обессоливания и производят центрифугирование в течение 2 мин при 1000 g;
Е) собирают проскок, содержащий разобранный бактериофаг;
Ж) разобранный бактериофаг разбавляют в 3 раза 50 мМ Tris с рН 7;
З) смесь, полученную на этапе Ж), доводят до рН до 7,3 с помощью добавления 10 М NaOH;
И) смесь, полученную на этапе Ж), концентрируют в 7,5 раз с помощью центрифужного концентратора при 3600 об/мин в охлажденной до плюс 4°С центрифуге;
К) полученный концентрат разобранного бактериофага делят на две части;
Л) одну часть концентрата разобранного бактериофага выдерживают в течение 24 ч при плюс 4°С в присутствии 25 мМ ТМАО на 50 мМ Tris с рН 7, 10 мМ тРНК дрожжей с концентрацией 1 мг/мл на 50 мМ Tris с рН 7; вторую часть концентрата разобранного бактериофага выдерживают в течение 24 ч при плюс 4°С в присутствии 25 мМ ТМАО, 10 мМ тРНК дрожжей с концентрацией 1 мг/мл и раствора радиоактивного металла с концентрацией 0,16 мг/мл;
М) по истечении 24 ч пробы с собранным бактериофагом обессоливают, таким образом, получая ВПЧ бактериофага, где модификация ВПЧ бактериофага представляет собой модификацию пептидом капсида.
8. Способ по п. 7, характеризующийся тем, что пептид является пептидом iRGD или пептидом YGRKKRRQRRR.
9. Способ по п. 8, характеризующийся тем, что пептид iRGD представляет собой пептид: первичной структуры GGGcyclo(CRGDKGPDC), первичной структуры GGGcyclo(CRGDRGPDC), первичной структуры GGGcyclo(CRGDKGPEC) или первичной структуры GGGcyclo(CRGDRGPEC).
10. Способ по любому из пп. 8, 9, характеризующийся тем, что ВПЧ получают из рекомбинантных капсидных белков с дальнейшей модификацией - присоединением пептида методом конъюгации.
11. Способ по любому из пп. 8-10, характеризующийся тем, что ВПЧ получают из рекомбинантных капсидных белков с пришитым пептидом.
12. Способ по любому из пп. 7, 8, характеризующийся тем, что радиоактивный металл представляет собой таллий, радий или технеций.
| US 8987173 B2, 24.03.2015 | |||
| Ректальные суппозитории на основе модифицированного бактериофага MS2 для таргетной терапии злокачественных солидных опухолей | 2019 |
|
RU2735828C1 |
| СПОСОБ ПОЛИСИГНАЛЬНОЙ АКТИВАЦИИ АПОПТОЗА КЛЕТОК ЗЛОКАЧЕСТВЕННЫХ СОЛИДНЫХ ОПУХОЛЕЙ | 2015 |
|
RU2599462C1 |

