Настоящее изобретение относится к способу получения бактериофагов, экспонирующих на своей поверхности чужеродные белки или пептиды, который не требует вмешательства в фаговый геном и не требует применения штаммов фагов, дефицитных по конкретным генам (данный способ не требует генетически модифицированных штаммов). Получаемые препараты бактериофагов имеют широкий спектр применения, в частности в очистке бактериофагов для изготовления продуктов, требующих высокой степени чистоты, например медицинских продуктов, а также в изготовлении бактериофагов, экспрессирующих конкретные белки или пептиды, для иных целей, т.е. в изготовлении вакцины.
Во время литического цикла, в ходе терапии, бактерии разрушаются бактериофагами, которые в них реплицируются. Потомство бактериофага, в значительно возросшем количестве, затем высвобождается в окружающую среду и лизирует очередные популяции бактерий.
В случае лизатов бактериофагов для биотехнологических целей имеет значение не только численность потомства бактериофага, но и высвобождаемые бактериальные составляющие, такие как нуклеиновые кислоты, белки и компоненты клеточной стенки. Стенка грамотрицательных бактерий состоит в значительной степени (примерно на 70%) из липополисахаридов (так называемые пирогены, эндотоксины), пептидов и белков.
Эффективное удаление пирогенов и активных белков из лизатов бактерий является ключевым требованием для производства препаратов бактериофагов, предназначенных для лечения бактериальной инфекции. Эндотоксины являются сильными стимуляторами иммунной системы и индуцируют продукцию интерлейкинов, TNF, NO и т.п.
Способы выведения и выделения эндотоксинов основаны на экстракции с помощью органических растворителей, таких как водный раствор фенола (Westphal О., Lueritz О., Bister F. Uber die Extraktion von Bakterien mit Phenol/wasser. Z. Naturforsch. 7: 148-155, 1952), алифатического амина и смеси кислот (Заявка на патент США 2007/0020292 А1), методах экстракции и хроматографии (Заявка на патент США 2007/0031447 A1). Удаление эндотоксинов из биологических препаратов осуществляли за счет взаимодействия ионов металлов с белками (патент US 6942802 В2, 13 сентября 2005; WO 02083710A1; WO 04003215 A1), путем осаждения эндотоксинов с помощью спирта и бивалентных противоионов (US 5039610). Использование бивалентных ионов в комбинации со спиртами, смолами и детергентами является объектом многих патентов, например для ЕРО 407037 В1. Удаление эндотоксинов осуществляли с помощью белков лимфы краба (US 5760177). Описано множество способов колоночной хроматографии. Они основаны на аффинности липополисахаридов к используемым гаптенам, например к полимиксину (Petsch D, Beeskow ТС, Anspach FB, Deckwer WD, (1997) Membrane adsorbers for selective removal of bacterial endotoxine. J. Chromatogr В Biomed Sci Appl. 693(1):79-91), кальций-силикатной смоле (Hang JP, Wang Q, Smith TR, Hurst WE, Sulpizio T, (2005) Endotoxine removal using a synthetic adsorbent of crystalline calcium silicate hydrate. Biotechnol Prago 21(4):1220-5), синтетическим полимерам (Hirayama Ch, Sakata M, (2002) Chromatographic removal of endotoxine from protein solutions by polymer particles. Journal of Chramatography B, 781:419-432) или полианионным подложкам (Boratynski J, Syper D, Weber-Dabrowska B, Łusiak-Szelachowska M, Pozniak G, Gorski A. Preparation of endotoxine-free bacteriofages Celi Mol Biol Lett. 2004; 9(2):253-9).
До сих пор сохраняется необходимость в способе очистки лизатов бактериофагов, в частности от эндотоксинов, который мог бы применяться для промышленного производства препаратов бактериофагов, предназначенных для применения в лечении бактериальных инфекций у человека. Исходные титры неочищенных лизатов могут содержать LPS с активностью 104-105. Несмотря на существование множества способов очистки лизатов бактериофагов, описанных выше, они в существенной мере не отвечают требованиям промышленного производства препаратов для вышеуказанных целей.
Способ фагового отображения для экспрессии заданного белка или пептида бактериофаговом капсиде основан на (i) введении генов, кодирующих такие белки или пептиды в бактериофаговый геном (разработка ГМО) или (ii) использовании штамма бактериофагов, дефицитного в отношении выбранного гена, кодирующего белок, который не является жизненно важным для бактериофага, и последующем восполнении этого недостатка с помощью рекомбинантных белков, экспрессируемых из конструкций вектора, как правило, в бактериальной системе (создание штамма с делецией выбранного гена). В обоих случаях отображение белка или пептида требует предварительной генетической модификации штамма хозяина.
Неожиданно, настоящее изобретение предлагает способ, удовлетворяющий вышеуказанным требованиям, разработанный на основе индустриальных способов получения очищенных фаговых препаратов, и в то же самое время позволяющий осуществить отображение чужеродных пептидов или белков на фаговом капсиде без внесения изменений в геном фагов (в том числе, фагов дикого типа).
Объектом настоящего изобретения является способ получения бактериофагов, отличающийся тем, что:
a) культуру бактериального хозяина выращивают в подходящей среде, засевают бактериофагом и получают бактериофаговый лизат для дальнейшей очистки или иного применения,
b) бактериофаговый лизат очищают с помощью аффинной хроматографии,
c) и из конечного элюата получают очищенный препарат бактериофага,
при этом на стадии a) используемый штамм бактериального хозяина состоит из бактериальных клеток, содержащих последовательность, кодирующую белки слияния, которые являются компонентами бактериофагового капсида, присутствующего в конечном лизате, с (i) полипептидом, обладающим сродством к хроматографической смоле, используемой на стадии b), если целью является получение очищенного препарата бактериофага, или (ii) другим полипептидом с требуемой активностью, если целью является презентация других активных мотивов.
Предпочтительно, чтобы бактериофаговый штамм, модифицированный и/или очищенный по данному способу, мог представлять собой любой заданный штамм дикого типа, не подвергавшийся генной модификации. Особенно предпочтительно, чтобы культура была засеяна штаммом бактериофага, который не подвергался генетической модификации с целью осуществления метода фагового отображения, т.е. бактериофага дикого типа, или модифицированным штаммом для иных целей, при этом указанный бактериофаг отображает чужеродные полипептиды на своей поверхности.
Полипептид, который позволяет провести очистку бактериофагов, обладающий сродством к хроматографической смоле, предпочтительно выбирают из группы, включающей His-тэг и GST. Пример полипептида, который является производным фагового структурного белка, кодируется последовательностью, кодирующей белок gp23*, зрелую форму главного капсидного белка gp23* (после протеолитического расщепления в ходе созревания головки). Эта последовательность показана на Фиг.1.
Предпочтительно в ходе стадии a) используемый штамм бактериального хозяина представляет собой бактериальный штамм, чувствительный к литической активности амплифицируемого бактериофага.
Предпочтительно в ходе стадии a) получаемый фаговый лизат стерилизуют фильтрацией с помощью стерилизующего фильтра с диаметром пор 0.22 мкм.
Аффинная хроматография является общепризнанным, очень эффективным способом очистки для белков, получаемых из различных источников. Неожиданным оказалось то, что применение этого метода в способе по настоящему изобретению выделения цельных бактериофаговых капсидов (сложных и крупных белковых комплексов) позволило сохранить антибактериальную активность выделенных бактериофагов, несмотря на то, что такие капсиды были существенно модифицированы, с целью обеспечения возможности использовать их в аффинной хроматографии. В примере осуществления мы применяли метод фагового отображения для ведения связывающих мотивов в фаговый капсид, и затем мы основывались на связывании бактериофагов, модифицированных таким образом, с аффинной смолой, выбранной надлежащим образом, для выбранного связывающего мотива.
Настоящее описание сопровождается следующими фигурами.
На Фиг.1 показана последовательность фрагмента 23 гена, кодирующего зрелую форму основного белка капсида gp23* (пост-протеолитическое расщепление).
На Фиг.2 показана кассета экспрессии: белок gp23*, слитый с GST, где ожидаемая масса продукта 23*GST составляет 78 кДа; последовательности из вектора pDEST15 (Invitrogen) выделены жирным, характеристические центры рекомбинации для вектора pDEST15 подчеркнуты, обычным шрифтом обозначены остальные элементы оператора, со следующими дополнительными выделениями: одинарное подчеркивание: n промотор; двойное подчеркивание: RBS; пунктир: GST; зигзаг: n терминатор; курсив обозначает конструкцию, содержащую ген 23*, включающий AcTev сайты расщепления (одинарное подчеркивание), 3xSer (двойное подчеркивание), ген 23* (пунктир).
На Фиг.3 показана кассета экспрессии - белок 23*, слитый с His-тэгом, где ожидаемая масса продукта 23*His-тега составляет 50 кДа; последовательности из вектора pDEST15 (Invitrogen) выделаны жирным, характеристические центры рекомбинации для вектора pDEST15 подчеркнуты, обычным шрифтом обозначены оставшиеся элементы оператора, со следующими дополнительными выделениями: одинарное подчеркивание: n промотор; двойное подчеркивание: RBS; пунктир: His-тэг; зигзаг: n терминатор; курсив обозначает конструкцию, содержащую ген 23*, включающий AcTev сайты расщепления (одинарное подчеркивание), 3xSer (двойное подчеркивание), ген 23* (пунктир).
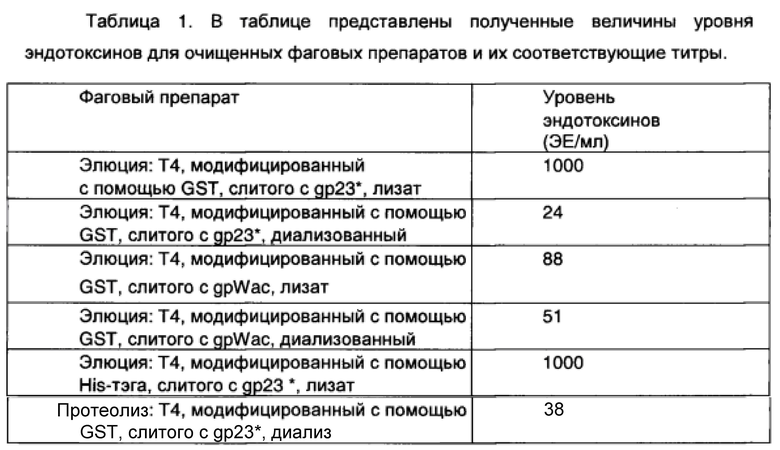
На Фиг.4 показана экспрессия белка gp23* и gpWac, слитого с GST и His-тэгом в клетках E.coli Rosetta (панели A, B, C, D); панель A - гель-электрофоретическое изображение - профиль белка после лизиса клеток, экспрессирующих белок 23*, слитый с His-тэгом: 1 - маркер веса (Fermentas SM0661), 2, 3, 4 - не имеют отношение к белку, 5 - 23*His-тэг через 3 ч после индукции с помощью IPTG (стрелка указывает на продукт); панель B - гель-электрофоретическое изображение - профиль белка после лизиса клеток, экспрессирующих Wac белок, слитый с His-тэгом: 1 - маркер веса (Fermentas SM0661), 2, 3, 4, 5 - не имеют отношение к белку, 6 - WacHis-тэг через 3 ч после IPTG-индукции (стрелка указывает на продукт); панель C - гель-электрофоретическое изображение - профиль белка после лизиса клеток, экспрессирующих Wac белок, слитый с GST: 1 - маркер веса (Fermentas SM0661), 2, 3, 4 - не имеют отношение к белку, 5 - WacGST через 3 ч после IPTG-индукции (стрелка указывает на продукт); панель D - гель-электрофоретическое изображение - профиль белка после лизиса клеток, экспрессирующих белок gp23*, слитый с GST: 1 - маркер веса (Fermentas SM0661), 2, 3 - не имеют отношение к белку, 4 - 23*GST через 3 ч после IPTG-индукции (стрелка указывает на продукт).
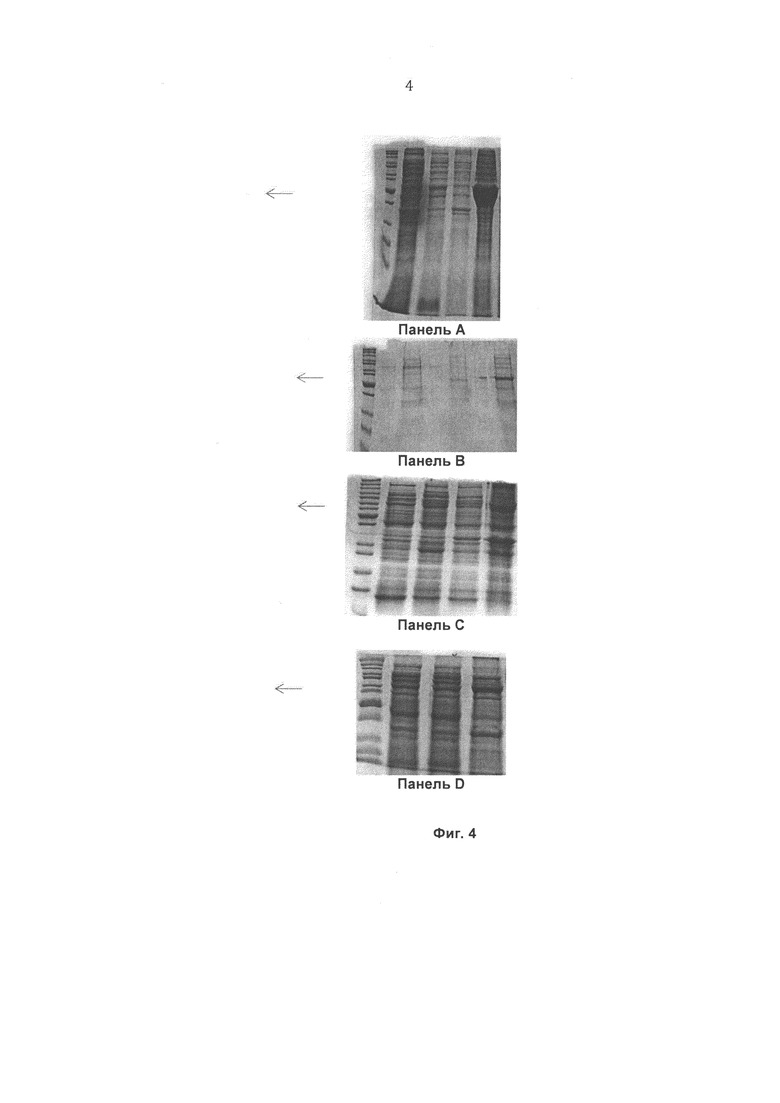
На Фиг.5 показаны результаты, полученные для препарата Т4 бактериофага с помощью конкурентного фагового отображения с использованием штамма Е.coli, экспрессирующего рекомбинантный белок gpWac, и очистки на глутатионовой смоле (dializa wstępna, фракция 2).
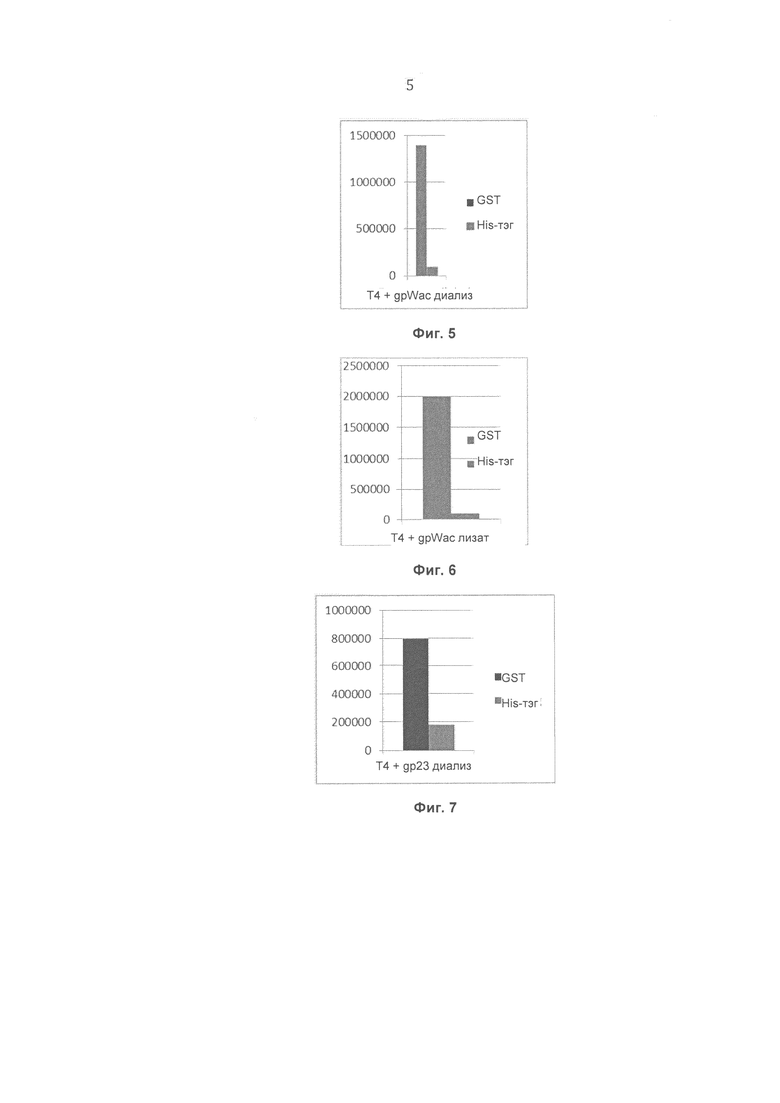
На Фиг.6 показаны результаты, полученные для препарата Т4 бактериофага с помощью конкурентного фагового отображения с использованием штамма Е.coli, экспрессирующего рекомбинантный белок gpWac, и очистки на глутатионовой смоле (без диализа, фракция 1).
На Фиг.7 показаны результаты, полученные для препарата Т4 бактериофага с помощью конкурентного фагового отображения с использованием штамма Е. coli, экспрессирующего рекомбинантный белок gp23*, и очистки на глутатионовой смоле (начальный диализ, фракция 1).
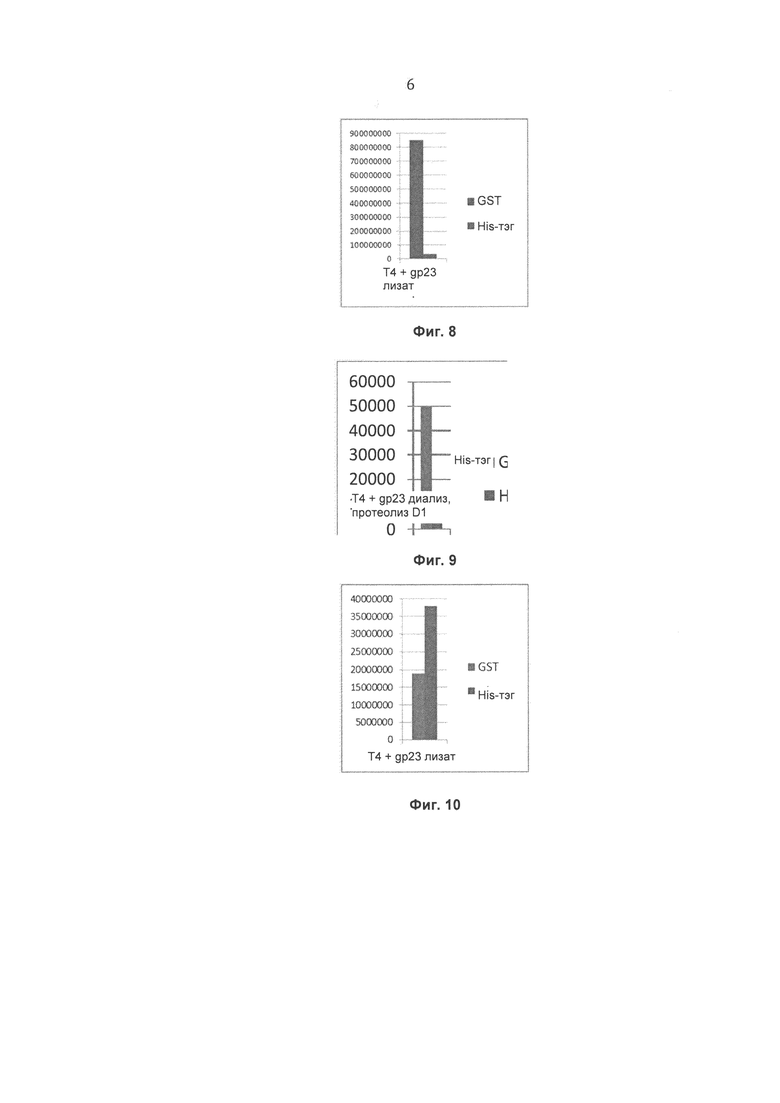
На Фиг. 8 показаны результаты, полученные для препарата Т4 бактериофага с помощью конкурентного фагового отображения, с использованием штамма Е. coli, экспрессирующего рекомбинантный белок gp23*, и очистки на глутатионовой смоле (без диализа, фракция 1).
На Фиг. 9 показаны результаты, полученные для препарата Т4 бактериофага с помощью протеолитического расщепления (1 сутки), с помощью фагового отображения с использованием штамма Е. coli, экспрессирующего рекомбинантный белок gp23*, и очистки на глутатионовой смоле (диализ).
На Фиг. 10 показаны результаты, полученные для препарата Т4 бактериофага с помощью протеолитического расщепления (1 сутки), с помощью конкурентного фагового отображения с использованием штамма Е. coli, экспрессирующего рекомбинантный белок gp23*, и очистки на никель-агарозной смоле NiNTA (без диализа, фракция 1).
Пример 1.
Данный способ основан на приготовлении и применении родительского бактериофагового штамма без генетических альтераций. Используемый хозяин Т4 бактериофага представлял собой экспрессионный штамм Escherichia coli, трансформированный плазмидами экспрессии, содержащими корректный ген 23* или корректный ген wac, слитый с последовательностью, кодирующей выбранный пептидный мотив для экспрессии на капсиде. Такая бактериофаговая культура способна конкурентно включать белки в рекомбинантную экспрессионную плазмиду, которая также экспрессируется в бактерии: gp23*, слитый со связывающим мотивом, или gpWac, слитый со связывающим мотивом. Это приводит к стабильным капсидным структурам, содержащим рекомбинантный белок, и, таким образом, содержащим и отображающим на своей поверхности мотив, обладающий сильным сродством к связывающим смолам.
Можно применять два альтернативных способа выделения бактериофагов из смолы: (i) конкурентная элюция, подразумевающая вытеснение с помощью соединений, способных взаимодействовать со связывающим мотивом на капсиде и/или связывающей смолой (глутатион, имидазол), или (ii) протеолитическое высвобождение с помощью протеазы, распознающей редкие мотивы. Вторая стратегия требует того, чтобы при создании конструкции экспрессионной пазмиды для экспрессии рекомбинантного белка в клетке была введена последовательность, которая распознается соответствующей протеазой. В случае протеолитического связывания у бактериофагового капсида отсутствует связывающий мотив.
Подробный пример осуществления способа по настоящему изобретению приведен ниже.
Клетки бактериального хозяина получали с помощью экспрессионного штамма Escherichia coli, которые были трансформированы экспрессионными плазмидами, содержащими корректный ген 23* или wac, слитый с последовательностью, кодирующей выбранный связывающий мотив. В данном примере осуществления мы использовали плазмиду pDEST15 (Invitrogen), содержавшую кассету экспрессии, которая позволяла получать белок, включающий gp23* или gpWac, слитый с GST (Фиг.2), или кассету экспрессии, кодирующую белок, включающий gp23* или gpWac, слитый с His-тэгом (Фиг.3).
Эффективность трансформации клеток Е.coli Rosetta определяли по экспрессии белков gp23* или gpWac, слитых с тэгами GST и His-тэгом (Фиг.4).
Рекомбинантные клетки бактериального хозяина культивировали при температуре 37°С до ОП600 0.7 в среде LB (LB-Broth, высокосолевой), включающие: энзиматический гидролизат казеина 10 г/л, дрожжевой экстракт 5 г/л, хлорид натрия 10 г/л, pH 7.5. Затем клетки переносили в свежую LB (при соотношении 1:100 к LB); как вариант: культуру выдерживали до OD600=0.1 и вносили IPTG до концентрации 0.0025 М и 1:100 фагового лизата НАР1 (~3×109 БОЕ/мл). Таким образом, индукция экспрессии и засевание происходили одновременно. Инфицированные клетки культивировали при 37°C, 160 об/мин в течение 8-12 часов.
В описанном здесь примере осуществления рекомбинантные Escherichia coli засевали с помощью Т4 бактериофагов дикого типа, одновременно индуцируя экспрессию Нос белков, и в тот же самое время осуществляли фаговый лизис и индукцию экспрессии рекомбинантных белков gp23* или gpWac.
Фаговый лизат фильтровали, возможно, диализовали против фосфатного буфера, идентичного тому, который используется для элюирования с колонок на мембрану с порами 300 кДа, после чего их инкубировали с подходящей агарозной смолой: глутатион-сефарозой или агарозой с ионами металлов, которые образуют комплексы с имидазольными остатками гистидина (т.е. агар NiNTA). Лизаты инкубировали с подходящей смолой в течение ночи при температуре 4°C при осторожном покачивании. После удаления не связавшейся фракции применяли стандартный способ очистки с помощью аффинной хроматографии. Смолу промывали буфером: 50 мМ Na2HPO4, 300 мМ NaCl, pH 7.5 (смола GST) или 50 мМ Na2HPO4, 300 мМ NaCl, 50-100 мМ имидазол, pH 7.5 (никелевая смола). Затем бактериофаги элюировали или протеолитически высвобождали из смолы.
В случае GST возможны два способа элюирования:
- конкурентное элюирование с помощью 40 мМ восстановленного глутатиона (фракции 1 или 2);
- элюирующий буфер: 40 мМ восстановленный глутатион, 50 мМ Tris, pH 8.0. Перед сбором каждой фракции смолу инкубировали с буфером в течение 20 минут;
- протеолитическое высвобождение с помощью AcTEV протеазы;
- применение протеолитического высвобождения требует предшествующего теоретического анализа последовательности или эмпирического теста на чувствительность бактериофага к протеазной активности (восстановление с контролируемым титром); в данном случае мы использовали теоретический анализ, основанный на анализе последовательности внешних Т4 капсидных белков;
- протеазный буфер: 50 мМ Tris, 0.5 мМ EDTA, 1 мМ ОН, pH 8.0. Фермент добавляли в количестве 5 мкл на 1 мл złoża (активность - 10 Ед/мкл). Протеолиз проводили в течение 1 дня при 4°C.
В случае смолы NiNTA фаг вытесняли с помощью имидазола в 100-500 мМ градиенте (1 или 2 фракции).
- элюирующий буфер: 100-500 мМ имидазол (в зависимости от фракции), 50 мМ Na2HPO4, 300 мМ NaCl, pH 7.5. Промывку и элюирование проводили при комнатной температуре.
Методы анализа результатов
Специфичность модифицированных бактериофагов в отношении аффинной смолы анализировали на основании сравнения профиля элюции модифицированных Т4 фагов с тэгом, специфичным для данной смолы, и тэгом, не соответствующим этой смоле. Профиль элюции определяли путем оценки фагового титра в индивидуальных фракциях. В качестве контроля определяли титр фага в последней фракции элюирующего буфера. Эксперимент рассматривали как успешный, основываясь на допущении, что титр в указанной фракции составлял не более 1% связанных фагов, которые были определены в ранее элюированных фракциях. Эффективность очистки оценивали, определяя уровень эндотоксинов, элюированных (или высвобожденных) во фракциях.
Результаты
Полученные результаты представлены в виде Фигур и Таблиц, включенных в настоящее описание.
Приведенные кривые представляют сравнение выбранных элюированных фракций. В случае экспериментов по конкуренции, которые проводили с целью подтверждения специфичности фага по отношению к смоле, начальные титры контрольных и потенциально специфичных образцов были идентичными. Равные объемы лизатов инкубировали с таким же объемом смолы, промывали и элюировали в тех же условиях.
На Фиг. 5 представлено сравнение препаратов Т4 бактериофага с помощью конкурентного фагового отображения с использованием штамма Е. coli, экспрессирующего рекомбинантный белок gpWac, и очистки на глутатионовой смоле (начальный диализ, фракция 2). Аффинность образца, содержащего специфический тэг (GST), по отношению к смоле превышает аффинность образца, включающего неспецифичный тэг (His-тэг), в 15 раз, что свидетельствует о специфическом связывании фага, модифицированного тэгом GST.
На Фиг. 6 представлено сравнение препаратов Т4 бактериофага с помощью конкурентного фагового отображения с использованием штамма Е. coli, экспрессирующего рекомбинантный белок gpWac, и очистки на глутатионовой смоле (без диализа, фракция 1). Аффинность образца, содержащего специфический тэг (GST), по отношению к смоле превышает аффинность образца, включающего неспецифичный тэг (His-тэг), в 20 раз, что свидетельствует о специфическом связывании фага, модифицированного тэгом GST.
На Фиг. 7 представлено сравнение препаратов Т4 бактериофага с помощью конкурентного фагового отображения с использованием штамма Е. coli, экспрессирующего рекомбинантный белок gp23*, и очистки на глутатионовой смоле (начальный диализ, фракция 1). Аффинность образца, содержащего специфический тэг (GST), по отношению к смоле превышает аффинность образца, включающего неспецифичный тэг (His-тэг), в 4.5 раз, что свидетельствует о специфическом связывании фага, модифицированного тэгом GST.
На Фиг. 8 представлено сравнение препаратов Т4 бактериофага с помощью конкурентного фагового отображения с использованием штамма Е. coli, экспрессирующего рекомбинантный белок gp23*, и очистки на глутатионовой смоле (без диализа, фракция 1). Аффинность образца, содержащего специфический тэг (GST), по отношению к смоле превышает аффинность образца, включающего неспецифичный тэг (His-тэг), более чем в 24 раза, что свидетельствует о специфическом связывании фага, модифицированного тэгом GST.
На Фиг. 9 представлено сравнение препаратов Т4 бактериофага с помощью конкурентного фагового отображения с использованием штамма Е. coli, экспрессирующего рекомбинантный белок gp23*, и очистки на глутатионовой смоле (диализ). Аффинность образца, содержащего специфический тэг (GST), по отношению к смоле превышает аффинность образца, включающего неспецифичный тэг (His-тэг) в 10 раз, что свидетельствует о специфическом связывании фага, модифицированного тэгом GST.
На Фиг. 10 представлено сравнение препаратов Т4 бактериофага с помощью конкурентного фагового отображения с использованием штамма Е. coli, экспрессирующего рекомбинантный белок gp23*, и очистки на глутатионовой смоле (без диализа, фракция 1). Аффинность образца, содержащего специфический тэг (His-тэг), по отношению к смоле превышает аффинность образца, включающего неспецифичный тэг (GST), в 2 раза, что свидетельствует о специфическом связывании фага, модифицированного с помощью His-тэга.
Выводы
Предложенный способ позволяет осуществить одностадийную и в то же время эффективную очистку фаговых препаратов, с сохранением антибактериальной активности бактериофагов как в случае стратегии, основанной на вытеснении бактериофагов из смолы, так и для протеолитического высвобождения. Данный способ не требует отдельных стадий для удаления белковых и небелковых (здесь: LPS) загрязнений. Модификация фагового капсида подходящими связывающими мотивами также может облегчать очистку других штаммов бактериофагов с помощью аффинной хроматографии. Предложенный способ способствует презентации выбранных пептидов или белков на фагах, которые не были генетически модифицированы в целях фагового отображения, т.е. штаммов дикого типа, встречающихся в природе, или других (т.е. лабораторных штаммов для различных целей).








Предложенное изобретение касается способа очистки бактериофагов. Охарактеризованный способ включает:
а) культивирование штамма бактериального хозяина в подходящей среде, засев культуры бактериофагом и получение бактериофагового лизата,
b) очищение лизата с помощью аффинной хроматографии,
c) использование полученного фильтрата для приготовления очищенного бактериофага,
при этом на стадии a) используемый штамм бактериального хозяина состоит из бактериальных клеток, содержащих последовательность, кодирующую белок слияния, включающий чужеродный полипептид, обладающий аффинностью к хроматографической смоле, применяемой на стадии b), и полипептид, относящийся к структурным фаговым белкам бактериофага, присутствующий в конечном лизате, и указанный полипептид, обладающий аффинностью по отношению к хроматографической смоле, выбирают из группы, включающей His-тэг и GST. Представленное изобретение позволяет осуществить одностадийную и в то же время эффективную очистку фаговых препаратов для терапевтических целей и обеспечивает сохранение антибактериальной активности бактериофага как в случае вытеснения бактериофага из смолы, так и в случае его протеолитического высвобождения. 4 з.п. ф-лы, 10 ил., 1 табл., 1 пр.
1. Способ очистки бактериофагов, характеризующийся тем, что:
a) штамм бактериального хозяина культивируют в подходящей среде, культуру засевают бактериофагом и получают бактериофаговый лизат,
b) лизат очищают с помощью аффинной хроматографии,
c) полученный фильтрат используют для приготовления очищенного бактериофага,
при этом на стадии a) используемый штамм бактериального хозяина состоит из бактериальных клеток, содержащих последовательность, кодирующую белок слияния, включающий чужеродный полипептид, обладающий аффинностью к хроматографической смоле, применяемой на стадии b), и полипептид, относящийся к структурным фаговым белкам бактериофага, присутствующий в конечном лизате, и указанный полипептид, обладающий аффинностью по отношению к хроматографической смоле, выбирают из группы, включающей His-тэг и GST.
2. Способ по п. 1, характеризующийся тем, что в ходе стадии а) культуру штамма бактериального хозяина ведут на питательной среде с pH около 7.2, содержащей мясной экстракт, энзиматический гидролизат казеина, дрожжевой гидролизат, пептон и NaCl.
3. Способ по п. 1, характеризующийся тем, что на стадии а) используемый штамм бактериального хозяина представляет собой бактериальный штамм, чувствительный к литической активности амплифицируемого бактериофага.
4. Способ по п. 1, характеризующийся тем, что в ходе стадии а) культуру засевают бактериофаговым штаммом, который не был генетически модифицирован с целью применения метода фагового отображения.
5. Способ по п. 1, характеризующийся тем, что в ходе стадии а) конечный фаговый лизат стерилизуют фильтрацией через стерилизующий фильтр с диаметром пор 0.22 мкм.
| JENNIFER JIANG et al., Display of a PorA Peptide from Neisseria meningitidis on the Bacteriophage T4 Capsid Surface, INFECTION AND IMMUNITY, Nov | |||
| Электрическое сопротивление для нагревательных приборов и нагревательный элемент для этих приборов | 1922 |
|
SU1997A1 |
| Разборное приспособление для накатки на рельсы сошедших с них колес подвижного состава | 1920 |
|
SU65A1 |
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| Аппарат для электрической сварки в притык | 1926 |
|
SU4770A1 |
| ANDREW K | |||
| UDIT et al., Immobilization of bacteriophage Qb on metal-derivatized surfaces via polyvalent display of hexahistidine tags, Journal of Inorganic | |||

