Список последовательностей
[001] Аминокислотные последовательности, перечисленные в сопроводительном списке последовательностей, показаны с использованием стандартного трехбуквенного кода для аминокислот, как определено в 37 CFR 1.822. Список последовательностей представляется в виде текстового файла Unicode, созданного 16 августейший 2016 года, с именем 0351 Sequence Listing.txt и размером 6604 байт, который включен в данный документ в качестве ссылки.
ОБЛАСТЬ ТЕХНИКИ
[002] Описание изобретения относится к способу получения рекомбинантного полипептида, включающему: (а) обеспечение подпитываемого периодического биореактора объемом от 100 до 25000 л, содержащего (i) клетки, способные экспрессировать рекомбинантный полипептид асфотазу альфа (SEQ ID NO: 1) и (ii) культуральную среду, подходящую для проведения такой экспрессии, при этом культуральная среда, содержит от около 25 мкМ до около 300 мкМ цинка; (b) культивирование клеток в условиях, подходящих для экспрессии рекомбинантной асфотазы альфа; причем рН культуральной среды составляет от около 6,7 до около 7,1, и причем цинк добавляют в указанную культуральную среду, так что концентрация цинка в культуральной среде поддерживается в концентрации от около 25 мкМ до около 300 мкМ цинка.
УРОВЕНЬ ТЕХНИКИ
[003] Гипофосфатазия (ГФФ) представляет собой опасное для жизни, генетическое и крайне редкое метаболическое расстройство, которое приводит к неспособности продуцировать функциональную тканенеспецифическую щелочную фосфатазу (TNSALP). Это приводит к накоплению неминерализованного костного матрикса (например, размягчению костей, остеомаляции), характеризующегося гипоминерализацией костей и зубов. Когда растущая кость не минерализуется должным образом, ухудшение роста является результатом, который приводит к искажению суставов и костей. Это, в свою очередь, влияет на двигательную моторику, дыхательную функцию и может даже привести к смерти. Было обнаружено, что различные формы ГФФ включают перинатальную, инфантильную, ювенильную и взрослую ГФФ. Недавно было определено шесть клинических форм, большинство из которых базируется на возрасте при начале симптомов, включая перинатальную, доброкачественную пренатальную, инфантильную, ювенильную, взрослую и одонто-ГФФ. Асфотаза альфа - одобренная, первая в своем классе целевая ферментная заместительная терапия, предназначенная для решения проблемы уровней дефектных эндогенных TNSALP. Для лечения ГФФ с помощью TNSALP см. Whyte et al., 2012 N Engl J Med. 366:904-13.
[004] Асфотаза альфа (STRENSIQ®, Alexion Pharmaceuticals, Inc.) представляет собой растворимый гибридный гликопротеин, состоящий из каталитического домена TNSALP человека, Fc-домена человеческого иммуноглобулина G1 и дека-аспартатного пептида (т.е. D10), используемого в качестве домена, ориентированного на кости. In vitro асфотаза альфа связывается с большим сродством c гидроксиапатитом, чем растворимый TNSALP, лишенный дека-аспартатного пептида, что позволяет фрагменту TNSALP асфотазы альфа эффективно деградировать избыточный местный неорганический пирофосфат (PPi) и восстанавливать нормальную минерализацию. Гидролиз пирофосфатов способствует минерализации кости, и его эффекты сходны между видами, оцененными в неклинических исследованиях. Первоначальные исследования эффективности проводились на мышиной модели ГФФ (мыши Akp2-/-). Мышиная модель Akp2-/-, созданная путем инактивации гена TNSALP (Narisawa et al., 1997 Dev Dyn. 208:432-46), разделяет многие общие черты состояния человека, включая накопление неминерализованной костной матрицы.
КРАТКОЕ СОДЕРЖАНИЕ
[005] В данном документе описаны композиции щелочных фосфатаз (например, асфотазы альфа), которые имеют специфические характеристики (например, конкретные структуры гликанов, конкретные значения общего содержания сиаловой кислоты (TSAC) и т. д.) и производственные процессы, используемые для получения щелочных фосфатаз (например, асфотазы альфа) со специфическими характеристиками. Такие щелочные фосфатазы (например, асфотаза альфа) пригодны для использования в терапии, например, для лечения состояний, связанных с пониженным уровнем и/или функциями белка щелочной фосфатазы (например, недостаточным расщеплением неорганического пирофосфата (PPi)) у субъекта, например, человека.
[006] В одном аспекте настоящее описание обеспечивает способ получения рекомбинантного полипептида, имеющего функцию щелочной фосфатазы. В различных вариантах осуществления изобретения, функция щелочной фосфатазы может включать любые функции щелочной фосфатазы, известные в данной области, такие как ферментативная активность по отношению к природным субстратам, включая фосфоэтаноламин (PEA), неорганический пирофосфат (PPi) и пиридоксаль-5'-фосфат (PLP). Такой рекомбинантный полипептид может содержать асфотазу альфа (SEQ ID NO: 1).
[007] В некоторых вариантах осуществления изобретеия, описанный в данном документе способ дополнительно включает добавление цинка в указанную культуральную среду для получения рекомбинантного полипептида. Цинк может способствовать улучшению активности и/или стабильности рекомбинантного полипептида. В некоторых вариантах осуществления изобретения, цинк может быть добавлен для обеспечения концентрации цинка от около 1 до около 300 мкМ в указанной культуральной среде. В одном варианте осуществления изобретения, цинк может быть добавлен для обеспечения концентрации цинка от около 10 до около 150 мкМ (например, 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140 или 150 мкМ) в культуральной среде. В некоторых вариантах осуществления изобретения, добавляют цинк, чтобы обеспечить концентрацию цинка в культуральной среде от около 25 мкМ до около 150 мкМ, или от около 60 мкМ до около 150 мкМ. В одном конкретном варианте осуществления изобретения, добавляют цинк, чтобы обеспечить концентрацию цинка в культуральной среде около 30, 60 или 90 мкМ цинка. В одном конкретном варианте осуществления изобретения, добавляют цинк, чтобы обеспечить концентрацию цинка в культуральной среде около 28 мкМ. В некоторых вариантах осуществления изобретения, цинк добавляют в указанную культуральную среду в болюсе, непрерывно или полунепрерывно.
[008] В некоторых вариантах осуществления изобретения, описанный в данном документе способ дополнительно включает контроль рН указанной культуральной среды для получения рекомбинантного полипептида. Например, рН может быть установлен от около 6,8 до около 7,0. В одном конкретном варианте осуществления изобретения, рН устанавливают на около 6,9.
[009] В некоторых вариантах осуществления изобретения, описанный в данном документе способ дополнительно включает добавление, по меньшей мере, одной дополнительной болюсной подпитки в исходную культуральную среду, содержащую клетки во время культивирования и/или получения полипептида. Такое добавление свежей культуральной среды может улучшить активность (например, специфическую активность) полученного рекомбинантного полипептида. В одном варианте осуществления изобретения в культуральную среду во время культивирования добавляют по меньшей мере одну, две, три или четыре болюсных подпитки. В одном конкретном варианте осуществления добавляют по меньшей мере четыре болюсных подпитки. В некоторых вариантах осуществления изобретения, болюсное добавление(я) подпитки улучшает специфическую активность рекомбинантного полипептида. Клетки, описанные в данном документе для получения рекомбинантного полипептида, могут представлять собой любые клетки (например, клетки млекопитающих), известные в данной области. В некоторых вариантах осуществления изобретения, клетки выбирают из группы, состоящей из CHO, NSO/1, PER.C6, COS-7, эмбриональной линии почки человека (293 или 293 клетки, субклонированные для роста в суспензионной культуре), BHK, TM4, CVl, VERO -76, HeLa, MDCK, BRL 3A, W138, Hep G2, MMT 060562, TRI, MRC 5, клетки FS4 и клетки Hep G2. В некоторых вариантах осуществления клетки представляют собой клетки СНО.
[0010] В некоторых вариантах осуществления изобретения, клетки выращивают при первой температуре в течение определенного времени для роста клеток, а затем переходят на вторую температуру для экспрессии полипептида. Например, в некоторых вариантах осуществления описан способ, который дополнительно включает культивирование клеток при первой температуре до достижения плотности клеток, по меньшей мере, около 2,5×106 жизнеспособных клеток, а затем переходят на вторую температуру, которая ниже первой температуры для экспрессии рекомбинантного полипептида. Например, в некоторых вариантах осуществления изобретения, первая температура составляет от около 35°С до около 37,5°С. В некоторых вариантах осуществления изобретения, вторая температура составляет от около 29°С до около 35°С. В некоторых вариантах осуществления изобретения, первая температура составляет около 37°С, а вторая температура составляет около 30°С. В некоторых вариантах осуществления изобретения, первая температура составляет около 36,5°С, а вторая температура составляет около 33°С.
[0011] В другом аспекте настоящее описание обеспечивает рекомбинантный полипептид, полученный любым из способов, описанных в данном документе. Такой продуцируемый рекомбинантный полипептид может иметь по меньшей мере одну из специфических характеристик, возникающих в результате способов получения, описанных в данном документе. Такие характеристики могут включать, по меньшей мере, одну, выбранную из группы, включающей: (а) общее содержание сиаловой кислоты (TSAC) между от около 0,9 до 3,5 моль сиаловой кислоты/моль мономера белка; (b) изоэлектрическую фокусировку (IEF) между около 5,2 до около 6,7; (c) основную структуру гликанов, как показано на фиг. 41 или фиг. 42; (d) хроматографический профиль меченного 2-АВ олигосахарида, как показано на фиг. 38 или 39; (e) MALDI-ToF фингерпринтный профиль гликопептида, как показано на фиг. 40 или 44-49; (f) основную полосу на SDS-PAGE в восстановительных условиях, имеющую молекулярную массу около 88-108 кДа и не менее чем около 85% от общего количества полученного рекомбинантного полипептида; (g) основную полосу на SDS-PAGE в невосстанавливающих условиях, имеющую молекулярную массу от около 194 до около 273 кДа и не менее чем около 85% от общего количества, полученного рекомбинантного полипептида; (h) не менее чем около 95,0% для димеров рекомбинантного полипептида и не более чем около 5,0% для агрегатов с помощью жидкостной хроматографии высокого давления (ВЭЖХ) с исключением по размеру; (i) чистоту не менее чем около 95,0% с помощью жидкостной хроматографии высокого давления с обращенной фазой (RP-HPLC); (j) не менее чем около 90,0% для главного пика, не более чем около 6,0% для кислотных пиков и не более чем около 4,0% для основных пиков анион-обменной хроматографии (AEX); (k) процент связывания гидроксиапатита (HA) от около 75 до около 125%; (l) специфическая активность продукта (pNPP) от около 620 до около 1250 единиц/мг; (m) Km от около 13 до около 69 мкМ в анализе гидролиза неорганического пирофосфата (PPi); (n) Kcat от около 65 до около 165 с-1 в анализе гидролиза неорганического пирофосфата (PPi); (o) диапазон pI от около 6,45 до около 6,95 для всех пиков при капиллярном электрофорезе; (p) пики в масс-спектре MALDI-TоF, как показано на фиг. 34A после дегликозилирования; (q) пики в масс-спектре MALDI-TоF, как показано на фиг. 34B после восстановления и дегликозилирования; (r) пики в масс-спектре MALDI-TоF, как показано на фиг. 35; (s) профиль фосфорилирования, как показано на фиг. 36; (t) профиль сиалированных гликанов в отрицательном масс-спектре MALDI-TоF, как показано на фиг. 37A; (u) профиль нейтральных гликанов в положительном масс-спектре MALDI-TоF, как показано на фиг. 37B; (v) молярное отношение магния на моль рекомбинантного полипептида от около 0,03 до около 0,15; (w) молярное отношение кальция на моль рекомбинантного полипептида от около 0,5 до около 1,5; и (x) молярное отношение цинка на моль рекомбинантного полипептида от около 0,5 до около 3,0.
[0012] В одном варианте осуществления изобретения, полученный рекомбинантный белок имеет от около 0,7 до около 1,19 свободного цистеина на половину молекулы.
[0013] В одном варианте осуществления изобретения, полученный рекомбинантный белок имеет фосфорилирование в Ser 93 в процентах от около 13,5% до около 35,7%.
[0014] В одном варианте осуществления изобретения, полученный рекомбинантный белок имеет не менее чем 90,0% главного пика, не более чем 6,0% для кислых пиков и не более чем 4,0% для основных пиков на хроматограмме AEX. В одном конкретном варианте осуществления изобретения, полученный рекомбинантный белок имеет не менее чем 93,7% главного пика, не более чем 4,9% для кислых пиков и не более чем 3,4% для основных пиков на хроматограмме AEX.
[0015] В некоторых вариантах осуществления изобретения, полученный рекомбинантный белок имеет общее содержание сиаловой кислоты (TSAC) между от около 0,9 до около 3,5 моль сиаловой кислоты/моль мономера белка. В одном варианте осуществления изобретения, полученный рекомбинантный белок имеет среднее значение общего содержания сиаловой кислоты (TSAC) от около 1,2 до около 3,0 моль сиаловой кислоты/моль на мономер. В одном варианте осуществления изобретения, полученный рекомбинантный белок имеет среднее значение общего содержания сиаловой кислоты от около 1,9 до около 2,7 моль сиаловой кислоты/моль на мономер. В одном конкретном варианте осуществления изобретения, полученный рекомбинантный белок имеет среднее значение общего содержания сиаловой кислоты от около 1,85 до около 2,28 моль сиаловой кислоты/моль на мономер.
[0016] В одном варианте осуществления изобретения, полученный рекомбинантный белок имеет молярное отношение связывания иона магния менее чем около 0,15. В некоторых вариантах осуществления изобретения, полученный рекомбинантный белок имеет молярное отношение магния от около 0,05 до около 0,10. В одном конкретном варианте осуществления изобретения, полученный рекомбинантный белок имеет молярное отношение связывания иона магния около 0,12.
[0017] В некоторых вариантах осуществления изобретения, полученный рекомбинантный белок содержит, по меньшей мере, около 95,0% димеров рекомбинантного полипептида, около 5,0% или менее полипептидных агрегатов, то есть, как измерено с помощью эксклюзионной ВЭЖХ. В одном варианте осуществления изобретения, полученный рекомбинантный белок содержит, по меньшей мере, около 96,8% димеров рекомбинантного полипептида и около 3,2% или менее полипептидных агрегатов, то есть, как измерено с помощью эксклюзионной ВЭЖХ. В одном конкретном варианте осуществления изобретения, полученный рекомбинантный белок содержит, по меньшей мере, около 97,6% димеров рекомбинантного полипептида и около 2,4% или менее агрегатов, то есть, как измерено с помощью эксклюзионной ВЭЖХ.
[0018] В одном варианте осуществления изобретения, полученный рекомбинантный белок имеет чистоту не менее чем 95,0%, как измерено с помощью RP-HPLC. В одном конкретном варианте осуществления изобретения, полученный рекомбинантный белок имеет чистоту не менее чем 97,6%, как измерено с помощью RP-HPLC.
[0019] В некоторых вариантах осуществления изобретения, полученный рекомбинантный белок имеет процент связывания гидроксиапатита (НА), составляющий от около 75 до около 125%. В одном варианте осуществления изобретения, полученный рекомбинантный белок имеет средний % связывания гидроксиапатита от около 85% до около 97%. В одном конкретном варианте осуществления изобретения, полученный рекомбинантный белок имеет средний % связывания гидроксиапатита от около 90% до около 91%.
[0020] В одном варианте осуществления изобретения, полученный рекомбинантный белок имеет специфическую активность (pNPP) от около 620 до около 1250 единиц/мг. В одном конкретном варианте осуществления изобретения, полученный рекомбинантный белок имеет среднюю специфическую активность (pNPP) от около 904,0 до около 907,7 Ед/мг.
[0021] В некоторых вариантах осуществления изобретения, рекомбинантный полипептид кодируется полинуклеотидом, кодирующим полипептид, содержащий последовательность, указанную в SEQ ID NO: 1, или последовательность, полностью комплементарную SEQ ID NO: 1.
[0022] Рекомбинантный полипептид, описанный в данном документе, может быть получен в промышленном или коммерческом масштабе. Например, в некоторых вариантах осуществления изобретения, подпитываемый периодический реактор имеет объем от 200 до 20 000 л. В некоторых вариантах осуществления подпитываемый периодический реактор имеет объем от 2000 до 20 000 л.
[0023] В другом аспекте настоящее описание представляет собой фармацевтическую композицию, содержащую композицию, включающую описанный в данном документе рекомбинантный полипептид, в комбинации с фармацевтически приемлемым носителем, разбавителем или эксципиентом.
[0024] В другом аспекте настоящее описание обеспечивает способ использования рекомбинантного полипептида или обсуждаемого в данном документе фармацевтического препарата для увеличения расщепления неорганического пирофосфата (PPi) у субъекта.
[0025] В другом аспекте настоящее описание относится к способу лечения субъекта, включающему введение субъекту, страдающему от состояния, связанного с дефицитом щелочной фосфатазы, терапевтически эффективного количества рекомбинантного полипептида или обсуждаемой в данном документе фармацевтической композиции. Такое состояние, связанное с дефицитом щелочной фосфатазы, включает, например, гипофосфатазию (ГФФ) и нейрофиброматоз типа I (NF1). Такая гипофосфатазия (ГФФ) может быть любой из перинатальной, инфантильной, ювенильной или взрослой ГФФ. Такое состояние может быть охарактеризовано неминерализованной костной матрицей и/или гипоминерализацией костей и зубов. Например, такой неминерализованный костный матрикс может приводить к рахиту и/или остеомаляции.
[0026] В некоторых вариантах осуществления такой субъект представляет собой млекопитающее. В некоторых вариантах осуществления изобретения, такой субъект представляет собой человека.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0027] В прилагаемых графических материалах:
[0028] Фигура 1 представляет собой графики, показывающие сравнения роста клеток (вставка A, слева) и жизнеспособность (вставка A, справа) и общую концентрацию глюкозы (вставка B, слева) и лактата (вставка B, справа) между иллюстративным производственным процессом (№ 1) с или без изменения температуры для производства белка. Контроль представляет собой средний результат двух прогонов с изменением температуры.
[0029] На фигуре 2 представлены графики, показывающие сравнения титров, связываемых с белком А (вставка А), объемной активности (вставка В) и специфической активности (вставка С) асфотазы альфа, полученной с использованием иллюстративного производственного процесса (№ 1) с изменением температуры или без него. Контроль представляет собой средний результат двух прогонов с изменением температуры.
[0030] На фиг. 3 представлены графики, показывающие сравнения измерений кислотного пика (%) AEX (анионной обменной хроматографии) (группа A) и SEC (эксклюзионной хроматографии) агрегата (%) (панель B) асфотазы альфа, полученных с использованием иллюстративного производственного процесса (# 1) с изменением температуры или без.
[0031] На фиг. 4 представлены графики, показывающие сравнения LoC в невосстановительных условиях (панель Lab-on-Chip, вставка A), LoC в восстановительных условиях (вставка B) и TSAC (общее содержание сиаловой кислоты, вставка C) асфотазы альфа, полученной с использованием иллюстративного производственного процесса (№ 1) с изменением температуры или без.
[0032] Фиг. 5 представляет собой график, показывающий профили нейрального гликана (с помощью метода время-пролетной ионизации лазерной десорбции с использованием матрицы или MALDI-TOF) асфотазы альфа, полученной иллюстративным производственным процессом (№ 1) с изменением температуры или без. Эталон представляет собой стандартную асфотазу альфа, полученную в предыдущем 20K процессе.
[0033] На фиг. 6 представлены графики, показывающие влияние смещения температуры производства на агрегированные уровни (вставка A) и TSAC (вставка B) произведенной асфотазы альфа. Планки ошибок представляют собой стандартное отклонение при каждом условии.
[0034] Фиг. 7 представляет собой график, показывающий влияние рН культуры на значения TSAC асфотазы альфа, полученной иллюстративномым процессом №2 после 10 или 12 дней культивирования.
[0035] На фиг. 8 представлены графики, показывающие влияние добавления подпитки среды (представленные различными концентрациями гликозы (glc), вставка A) и режима добавления подпитки (вставка B) на TSAC асфотазы альфа в иллюстративном процессе №2.
[0036] На фиг. 9 представлены графики, показывающие влияние добавок цинка на активность асфотазы альфа (вставка А, цинк, добавленный в день 9), и влияние концентрации цинка на рост клеток (вставка В) и жизнеспособность (вставка С) в иллюстративном процессе №2.
[0037] На фиг. 10 представлены графики, показывающие влияние рН и температуры производства на TSAC асфотазы альфа (вставка A), титр белка A (вставка B) и объемную активность (вставка C) в иллюстративном процессе № 3.
[0038] На фиг. 11 представлены графики, показывающие влияние температуры роста на объемную активность (вставка А) и фрагментацию асфотазы альфа (вставка В, измеренная SEC) в иллюстративном процессе № 3.
[0039] На фиг. 12 представлены графики, показывающие влияние температуры и рН производства на агрегацию асфотазы альфа (вставка А, измеренная SEC), % кислотного пика (вставка B, измеренная AEX), фрагментацию асфотазы альфа (вставка C, измеренная SEC) и % основного пика (вставка D, измеренная AEX) в иллюстративном процессе № 3.
[0040] На фиг. 13 представлены графики, показывающие влияние плотности посева и времени температурного сдвига на объемную активность (вставка A) и фрагментацию асфотазы альфа (вставка B, измеренная SEC). Посев выдерживают для плотности посева в иллюстративном процессе № 3.
[0041] На фиг. 14 представлены графики, показывающие влияние подпитки среды (количество и время) на объемную активность асфотазы альфа (вставка A), агрегацию (вставка B, измеренная SEC), % кислотного пика (вставка C, измеренная AEX), % основного пика (вставка D, измеренная AEX) и TSAC (панель E) в иллюстративном процессе № 3.
[0042] На фиг. 15 представлены графики, показывающие влияние рН культуральной среды на рост клеток (вставка А) и жизнеспособность (вставка B) до десяти дней в иллюстративном процессе №4.
[0043] На фиг. 16 представлены графики, показывающие влияние температуры производства среды на рост клеток (вставка А) и жизнеспособность (вставка B) до десяти дней в иллюстративном процессе №4.
[0044] На фиг. 17 представлены графики, показывающие влияние рН культуральной среды на концентрации глюкозы (вставка А) и лактата (вставка В) в культуральной среде до десяти дней в иллюстративном процессе №4.
[0045] На фиг. 18 представлены графики, показывающие влияние температуры производства на концентрации глюкозы (вставка А) и лактата (вставка В) в культуральной среде до десяти дней в иллюстративном процессе №4.
[0046] На фиг. 19 представлены графики, показывающие влияние рН культуральной среды на титр белка А (вставка А), объемную активность (вставка В), специфичесКую производительность белка А (вставка С) и специфическую активность (вставка D) до десяти дней в иллюстративном процессе # 4.
[0047] На фиг. 20 представлены графики, показывающие различное влияние температуры производства на титр белка А (вставка А), объемную активность (вставка В), специфическую производительность белка А (вставка С) и специфическую активность (вставка D) в иллюстративном процессе # 4.
[0048] На фиг. 21 представлены графики, показывающие влияние рН культуральной среды (вставка A) и температуры производства (вставка B) на TSAC асфотазы альфа в иллюстративном процессе №4.
[0049] На фиг. 22 представлены графики, показывающие влияние рН культуральной среды (вставка A) и температуры производства (вставка B) на кислотный пик AEX полученной асфотазы альфа в иллюстративном процессе №4.
[0050] На фиг. 23 представлены графики, показывающие влияние рН культуральной среды (вставка A) и температуры производства (вставка B) на агрегацию асфотазы альфа (измереную SEC) в иллюстративном процессе №4.
[0051] На фиг. 24 представлены графики, показывающие различное влияние рН (вставка А) культуральной среды и температуры производства (вставка B) на измерение главного пика LoC (%, не восстанавливающие условия) произведенной асфотазы альфа в иллюстративном процессе №4.
[0052] На фиг. 25 показаны графики, показывающие влияние рН культуральной среды на нейтральные профили гликана полученной асфотазы альфа (измеренной MALDI-TOF) в иллюстративном процессе №4. Верхняя панель показывает все пики MALDI-ToF для асфотазы альфа, полученные при температуре 33°C в течение 10 дней, но при разном рН культуральной среды. Значение каждого пика рассчитывали и сравнивали в нижней вставке.
[0053] На фиг. 26 показаны графики, показывающие влияние температуры производства на нейтральные профили гликана полученной асфотазы альфа (измеренной MALDI-TоF) в иллюстративном процессе №4. Верхняя вставка показывает все пики MALDI-ToF для асфотазы альфа, полученные при разной температуре производства при рН 6,9 в течение 10 дней. Значение каждого пика рассчитывали и сравнивали в нижней вставке.
[0054] На фиг. 27 представлены графики, показывающие влияние рН культуральной среды (вставка A) и температуры производства (вставка B) на проценты пика капиллярного изоэлектрического фокусирования для iCE280 анализа качества асфотазы альфа в иллюстративном процессе №4.
[0055] На фиг. 28 показаны графики, сравнивающие рост клеток (вставка A) и жизнеспособность (вставка B) для различных процессов (синяя линия: контрольный процесс (выполняется для двух резервуаров биореактора) с одной болюсной подпиткой среды); красная линия: улучшенный процесс (выполняется для шести резервуаров биореактора) с четырехкратной питательной средой; серая линия: в качестве стандарта использовались предыдущие 20K процессов (выполненные для двадцати биореакторных резервуаров). Серая линия представляет собой среднее значение 20K процессов, а пунктирные линии представляют собой среднее±1 x стандартное отклонение.
[0056] На фиг. 29 показаны графики сравнения использования глюкозы (вставка A) и производства лактата (вставка B) для различных процессов (синяя линия: контрольный процесс (выполняется для двух резервуаров биореактора) с одной болюсной подпиткой среды); красная линия: улучшенный процесс (выполняется для шести резервуаров биореактора) с четырехкратной подпиткой среды; серая линия: в качестве стандарта использовались предыдущие 20K процессов (выполненные для двадцати биореакторных резервуаров). Серая линия представляет собой среднее значение 20K процессов, а пунктирные линии представляют собой среднее±1 x стандартное отклонение.
[0057] На фиг. 30 показаны графики сравнения титра белка А (вставка A) и специфической активности (вставка B) асфотазы альфа, полученной различными способами (синяя линия: контрольный процесс (выполняется для двух резервуаров биореактора) с одной болюсной подпиткой среды); красная линия: улучшенный процесс (выполняется для шести резервуаров биореактора) с четырехкратной подпиткой среды; серая линия: в качестве стандарта использовались предыдущие 20K процессов (выполненные для двадцати биореакторных резервуаров). Серая линия представляет собой среднее значение 20K процессов, а пунктирные линии представляют собой среднее±1 x стандартное отклонение.
[0058] На фиг. 31 показаны графики, сравнивающие активный титр (вставка A) и общую объемную активность (вставка B) для различных процессов (синяя линия: контрольный процесс (выполняется для двух резервуаров биореактора) с одной болюсной подпиткой среды); красная линия: улучшенный процесс (выполняется для шести резервуаров биореактора) с четырехкратной подпиткой среды; серая линия: в качестве стандарта использовались предыдущие 20K процессов (выполненные для двадцати биореакторных резервуаров). Серая линия представляет собой среднее значение 20K процессов, а пунктирные линии представляют собой среднее±1 x стандартное отклонение.
[0059] Фиг. 32 представляет собой график, сравнивающий специфическую активность асфотазы альфа из контрольных процессов (серая, одна болюсная подпитка среды) и улучшенных процессов (темно-серый, четырехкратная подпитка среды).
[0060] Фигура 33 представляет собой график, сравнивающий специфическую активность асфотазы альфа из предыдущего процесса Y (нижняя линия без дополнительной добавки цинка) и еще один улучшенный процесс (верхняя линия, показывающая среднее значение условий добавок 30-90 мкМ цинка и время культивирования, продленное до 14 дней).
[0061] На фиг. 34 представлены графики, показывающие данные масс-спектра MALDI-ToF для полученной асфотазы альфа после дегликозилирования (вставка A) и для продуцируемой асфотазы альфа, которая была восстановлена и дегликозилирована (вставка B).
[0062] Фигура 35 представляет собой график, показывающий масс-спектр MALDI-ToF продуцируемой асфотазы альфа. Фигура 35 обеспечивает «TYNTNAQVPDSAGTATAYAYLCGVK» в качестве остатков 83-105 SEQ ID NO: 1, «SAGTA» в качестве остатков 93-97 SEQ ID NO: 1 и «AYLCGV» в качестве остатков 99-104 SEQ ID NO: 1, соответственно, в порядке появления.
[0063] Фигура 36 представляет собой графики, показывающие определение MС/MС сайта фосфорилирования на асфотазе альфа.
[0064] На фиг. 37 представлены графики, показывающие отрицательный масс-спектр MALDI-ToF сиалированных гликанов (вставка A) и положительный масс-спектр MALDI-TOF нейтральных гликанов (вставка B) на продуцируемой асфотазе альфа.
[0065] Фигура 38 представляет собой график, показывающий флуоресцентную хроматограмму олигосахаридов асфотазы альфа.
[0066] Фигура 39 представляет собой график, показывающий флуоресцентную хроматограмму эталонного стандарта асфотазы альфа.
[0067] На фиг. 40 представлены графики, показывающие масс-спектр гликопептидов, полученных из асфотазы альфа.
[0068] На фиг. 41 представлены типичные предлагаемые структуры основных гликанов асфотазы альфа (C7108H11008O2206S56 (димер части белка) или C3554H5506O1103S28 (мономер)). Число NeuAc на гликан (FA2G2, FA2G1 и A2G2) является оценочным числом.
[0069] На фиг. 42 представлены типичные оцененненные структуры гликанов на сайтах гликозилирования асфотазы альфа.
[0070] Фигура 43 представляет собой график, показывающий типичную электроферограмму асфотазы альфа.
[0071] На фиг. 44 представлены графики, показывающие фингерпринты массы гликопептида для асфотазы альфа, полученные из резервуаров 20K и 2K (N123 в T15-16). 2K номера резервуара: # 35, # 36 и # 38; 20K: # 40, # 42 и # 34.
[0072] На фиг. 45 представлены графики, показывающие фингерпринты массы гликопептида для асфотазы альфа, полученные из резервуаров 20K и 2K (N213 в T26-27). 2K номера резервуара: # 35, # 36 и # 38; 20K: # 40, # 42 и # 34.
[0073] На фиг. 46 представлены графики, показывающие фингерпринты массы гликопептида для асфотазы альфа, полученные из резервуаров 20K и 2K (N254 в T33). 2K номера резервуара: # 35, # 36 и # 38; 20K: # 40, # 42 и # 34.
[0074] На фиг. 47 представлены графики, показывающие фингерпринты массы гликопептида для асфотазы альфа, полученные из резервуаров 20K и 2K (N286 в T35). 2K номера резервуара: # 35, # 36 и # 38; 20K: # 40, # 42 и # 34.
[0075] На фиг. 48 представлены графики, показывающие фингерпринты массы гликопептида для асфотазы альфа, полученные из резервуаров 20K и 2K (N413 в T45-46). 2K номера резервуара: # 35, # 36 и # 38; 20K: # 40, # 42 и # 34.
[0076] На фиг. 49 представлены графики, показывающие фингерпринты массы гликопептида для асфотазы альфа, полученные из резервуаров 20K и 2K (N564 в T55). 2K номера резервуара: # 35, # 36 и # 38; 20K: # 40, # 42 и # 34.
[0077] На фиг. 50 представлены графики, сравнивающие молярное отношение ICP ионов металлов для цинка (вставка A), магния (вставка B) и кальция (вставка C) асфотазы альфа, полученного из партий 2K и 20K.
[0078] На фигуре 51А показан график, сравнивающий плотность жизнеспособных клеток (VCD) через время культивирования в разных условиях рН. На фигуре 51В приведен график, сравнивающий жизнеспособность клеток через время культивирования при различных условиях рН.
[0079] На фиг. 52А представлен график, показывающий концентрацию глюкозы в биореакторах через прошедшее время культивирования. На фиг. 52В представлен график, показывающий концентрацию лактата в биореакторах через прошедшее время культивирования.
[0080] Фигура 53 представляет собой график, сравнивающий конкретные профили активности при различных условиях рН.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Определения
[0081] «Около», «Примерно»: Как используется в данном документе, термины «около» и «примерно», применительно к одному или более конкретным условиям культивирования клеток, относятся к диапазону значений, которые похожи на указанную опорную величину для данного состояния или условий культивирования. В некоторых вариантах осуществления изобретения, термин «около» относится к диапазону значений, которые находятся в пределах 25, 20, 19, 18, 17, 16, 15, 14, 13, 12, 11, 10, 9, 8, 7, 6, 5, 4, 3, 2, 1 процентов или менее указанного эталонного значения для этого условия или условий культивирования.
[0082] "Аминокислота": Используемый в данном документе термин «аминокислота» относится к любой из двадцати встречающихся в природе аминокислот, которые обычно используются при образовании полипептидов или аналогов или производных этих аминокислот. Аминокислоты настоящего изобретения могут быть представлены в среде клеточных культур. Аминокислоты, представленные в среде, могут быть представлены в виде солей или в форме гидрата.
[0083] «Периодическая культура»: Используемый в данном документе термин «периодическая культура» относится к способу культивирования клеток, в котором все компоненты, которые в конечном итоге будут использоваться для культивирования клеток, включая среду (см. Определение «среда» ниже), а также сами клетки, обеспечиваются в начале процесса культивирования. Периодическую культуру обычно останавливают в какой-то момент, и клетки и/или компоненты в среде собирают и, необязательно, очищают.
[0084] «Биореактор»: Используемый в данном документе термин «биореактор» относится к любому сосуду, используемому для роста клеточной культуры (например, к культуре клеток млекопитающих). Биореактор может быть любого размера, если он полезен для культивирования клеток. Как правило, биореактор будет составлять по меньшей мере 1 литр и может составлять 10, 100, 250, 500, 1000, 2500, 5000, 8000, 10000, 120000, 20000 литров или более или любой объем между ними. Внутренние условия биореактора, включающие, но не ограничиваясь ими, рН и температуру, обычно контролируются в течение периода культивирования. Биореактор может состоять из любого материала, который подходит для удерживания клеточных культур млекопитающих или других клеточных культур, суспендированных в средах в условиях культивирования настоящего изобретения, включая стекло, пластик или металл. Используемый в данном документе термин «производственный биореактор» относится к конечному биореактору, используемому при получении представляющего интерес полипептида или белка. Объем крупного биореактора для культивирования клеток обычно составляет не менее 500 литров и может составлять 1000, 2500, 5000, 8000, 10000, 120000, 20000 литров или более, или любой объем между ними. Специалист в данной области техники будет знать и сможет выбирать подходящие биореакторы для использования в практике настоящего описания.
[0085] «Плотность клеток»: Используемый в данном документе термин «плотность клеток» относится к числу клеток, присутствующих в данном объеме среды.
[0086] "Жизнеспособность клеток": Термин «жизнеспособность клеток», используемый в данном документе, относится к способности клеток в культуре выживать при заданном наборе условий культивирования или экспериментальных вариациях. Используемый в данном документе термин также относится к той части клеток, которая жива в определенное время по отношению к общему количеству клеток, живых и мертвых, в культуре в то же время.
[0087] «Культура» и «культура клеток»: Эти термины, используемые в данном документе, относятся к популяции клеток, которая суспендирована в среде (см. Определение «среда» ниже) в условиях, подходящих для выживания и/или роста популяции клеток. Как будет ясно специалистам в данной области техники, эти термины, используемые в данном документе, могут относиться к комбинации, содержащей популяцию клеток и среду, в которой популяция суспендирована.
[0088] ʺПодпитываемая периодическая культураʺ: Термин «подпитываемая периодическая культура», как используется в данном документе, относится к способу культивирования клеток, в котором дополнительные компоненты обеспечиваются в культуре в течение некоторого времени после начала процесса культивирования. Предоставленные компоненты обычно содержат питательные добавки для клеток, которые были истощены во время процесса культивирования. Подпитываемую периодическую культуру обычно останавливают в какой-то момент, и клетки и/или компоненты в среде собирают и, необязательно, очищают. Подпитываемая периодическая культура может быть запущена в соответствующем подпитываемом периодическом биореакторе.
[0089] "Фрагмент": Используемый в данном документе термин «фрагмент» относится к полипептиду и определяется как любая дискретная часть данного полипептида, которая является уникальной или характерной для этого полипептида. Используемый в данном документе термин также относится к любой дискретной части данного полипептида, который сохраняет по меньшей мере часть активности полноразмерного полипептида. В некоторых вариантах осуществления изобретения доля сохраняемой активности сохраняется по меньшей мере на 10% от активности полноразмерного полипептида. В различных вариантах осуществления изобретения, доля сохраняемой активности составляет по меньшей мере 20%, 30%, 40%, 50%, 60%, 70%, 80% или 90% от активности полноразмерного полипептида. В других вариантах осуществления изобретения, доля сохраняемой активности составляет по меньшей мере 95%, 96%, 97%, 98% или 99% от активности полноразмерного полипептида. В одном варианте осуществления изобретения, доля сохраняемой активности составляет 100% от активности полноразмерного полипептида. Используемый в данном документе термин также относится к любой части данного полипептида, который включает по меньшей мере установленный элемент последовательности, обнаруженный в полноразмерном полипептиде. В некоторых вариантах осуществления изобретения, элемент последовательности охватывает по меньшей мере 4-5 аминокислот полноразмерного полипептида. В некоторых вариантах осуществления изобретения, элемент последовательности охватывает по меньшей мере около 10, 15, 20, 25, 30, 35, 40, 45, 50 или более аминокислот полноразмерного полипептида.
[0090] «Интегрированная плотность жизнеспособных клеток»: Термин «интегрированная плотность жизнеспособных клеток», как используется в данном документе, относится к средней плотности жизнеспособных клеток в течение культуры, умноженной на количество времени, которое культура культивировалась. Предполагая, что количество продуцируемого полипептида и/или белка пропорционально количеству жизнеспособных клеток, присутствующих в ходе культуры, интегрированная плотность жизнеспособных клеток является полезным инструментом для оценки количества полипептида и/или белка, продуцируемого в течение культивирования.
[0091] «Среда», «среда для культивирования клеток» и «культуральная среда»: Эти термины, используемые в данном документе, относятся к раствору, содержащему питательные вещества, которые питают растущие клетки млекопитающих. Как правило, эти растворы обеспечивают незаменимые и не незаменимые аминокислоты, витамины, источники энергии, липиды и микроэлементы, необходимые клетке для минимального роста и/или выживания. Раствор может также содержать компоненты, которые усиливают рост и/или выживание выше минимальной нормы, включая гормоны и факторы роста. Раствор, например, составлен до концентрации рН и соли, оптимальной для выживания и пролиферации клеток. Среда также может быть «определенной средой» - бессывороточной средой, которая не содержит белков, гидролизатов или компонентов неизвестного состава. Определенные среды не содержат компонентов животного происхождения, и все компоненты имеют известную химическую структуру.
[0092] «Конечный продукт обмена веществ»: Используемый в данном документе термин «конечный продукт обмена веществ» относится к соединению, продуцируемому клеточной культурой в результате нормальных или ненормальных метаболических процессов, которые каким-то образом наносят ущерб культуре клеток, особенно в отношении экспрессии или активности желаемого рекомбинантного полипептида или белка. Например, конечные продукты обмена веществ могут пагубно влиять на рост или жизнеспособность культуры клеток, могут уменьшать количество продуцируемого рекомбинантного полипептида или белка, могут изменять сворачивание, стабильность, гликозилирование или другую посттрансляционную модификацию экспрессированного полипептида или белка, или может быть вредным для клеток и/или экспрессии или активности рекомбинантного полипептида или белка любым другим способом. Иллюстративные конечные продукты обмена веществ включают лактат, который образуется в результате метаболизма глюкозы, и аммония, который образуется в результате метаболизма глютамина. В одном варианте осуществления используются способы замедления производства, уменьшения или даже устранения конечных продуктов обмена веществ в культурах клеток.
[0093] «Осмоляльность» и «осмолярность»: Осмоляльность является мерой осмотического давления растворенных частиц в водном растворе. Частицы растворенного вещества включают как ионы, так и неионизированные молекулы. Осмоляльность выражается в виде концентрации осмотически активных частиц (то есть осмолей), растворенных в 1 кг раствора (1 мОсм/кг Н2О при 38°С эквивалентно осмотическому давлению 19 мм рт. ст.). «Осмолярность», напротив, относится к числу растворенных частиц, растворенных в 1 литре раствора. При использовании в данном документе аббревиатура «мОсм» означает «миллиосмолей/кг раствора».
[0094] «Перфузионная культура»: Термин «перфузионная культура», как используется в данном документе, относится к способу культивирования клеток, в котором дополнительные компоненты предоставляются культуре непрерывно или полунепрерывно после начала процесса культивирования. Предоставленные компоненты обычно содержат питательные добавки для клеток, которые были истощены во время процесса культивирования. Часть клеток и/или компонентов в среде обычно собирают на непрерывной или полунепрерывной основе и необязательно очищают.
[0095] "Полипептид": Используемый в данном документе термин «полипептид» относится к последовательной цепи аминокислот, связанных вместе через пептидные связи. Этот термин используется для обозначения аминокислотной цепи любой длины, но специалист в данной области техники поймет, что этот термин не ограничивается длинными цепями и может относиться к минимальной цепи, содержащей две аминокислоты, связанные вместе через пептидную связь.
[0096] «Белок»: Используемый в данном документе термин «белок» относится к одному или более полипептидам, которые функционируют как дискретная единица. Если один полипептид является дискретной функциональной единицей и не требует постоянной физической связи с другими полипептидами для образования дискретной функциональной единицы, термины «полипептид» и «белок», используемые в данном документе, используются взаимозаменяемо.
[0097] «Рекомбинантно экспрессируемый полипептид» и «рекомбинантный полипептид»: Эти термины, используемые в данном документе, относятся к полипептиду, экспрессируемому из клетки-хозяина, который был генетически сконструирован для экспрессии этого полипептида. Рекомбинантно экспрессируемый полипептид может быть идентичным или подобным полипептиду, который обычно экспрессируется в клетке-хозяине млекопитающего. Рекомбинантно экспрессируемый полипептид также может быть чужеродным по отношению к клетке-хозяину, то есть гетерологичен пептидам, обычно экспрессируемым в клетке-хозяине. Альтернативно, рекомбинантно экспрессируемый полипептид может быть химерическим в тех частях полипептида, которые содержат аминокислотные последовательности, которые идентичны или аналогичны полипептидам, обычно экспрессируемым в клетке-хозяине млекопитающих, тогда как другие части являются чужеродными по отношению к клетке-хозяину.
[0098] «Посев»: Используемый в данном документе термин «посев» относится к процессу загрузки культуры клеток в биореактор или другой сосуд. Клетки могут ранее выращиваться в другом биореакторе или сосуде. Альтернативно, клетки могут быть заморожены и разморожены непосредственно перед их подачей в биореактор или сосуд. Термин относится к любому числу клеток, включая одну клетку.
[0099] "Титр": Используемый в данном документе термин «титр» относится к общему количеству рекомбинантно экспрессируемого полипептида или белка, продуцируемого клеточной культурой, деленному на заданное количество среднего объема. Титр обычно выражают в единицах миллиграммов полипептида или белка на миллилитр среды.
[00100] Акронимы, используемые в данном документе, включают, например, VCD: Плотность жизнеспособных клеток; IVCC: Интеграл концентрации жизнеспособных клеток; TSAC: Общее содержание сиаловой кислоты; HPAE-PAD: Высокоэффективная анион-обменная хроматография с импульсным амперометрическим детектированием; SEC: Гель-проникающая Хроматография; AEX: Анионная обменная хроматография; LoC: Lab-on-Chip; и MALDI-TOF: Матричная лазерная десорбция/ионизация - время полета.
[00101] Настоящее описание представляет способ культивирования клеток (например, клеток млекопитающих, включая, но не ограничиваясь ими, клетки яичника китайского хомячка (CHO)), экспрессирующие рекомбинантный белок. Настоящее описание обеспечивает производственные системы для получения щелочной фосфатазы (например, асфотазы альфа) с помощью культуры клеток. В некоторых вариантах осуществления изобретения, предусматриваются системы, которые сводят к минимуму производство одного или более метаболических продуктов, которые наносят ущерб росту, жизнеспособности клеток и/или производству белка или качеству. В конкретных вариантах осуществления изобретения, клеточная культура представляет собой периодическую культуру, подпитываемую периодическую культуру, культуру или непрерывную культуру. Другие варианты осуществления описания подробно обсуждаются ниже. Специалисты в данной области техники поймут, однако, что различные модификации этих вариантов осуществления изобретения включены в объем описания.
Белки
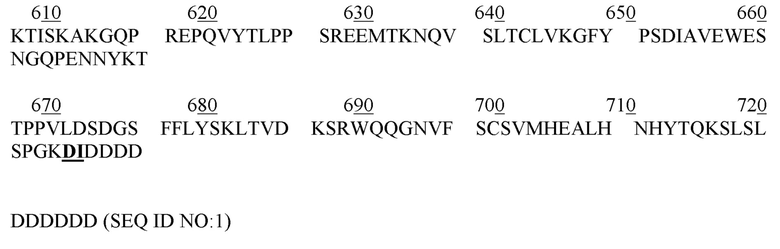
[00102] Настоящее описание относится к экспрессии белка щелочной фосфатазы, асфотазы альфа, в клеточной культуре. В некоторых вариантах осуществления изобретения, такие, как асфотаза альфа, после получения описанными в данном документе способами, могут быть использованы для лечения или профилактики заболеваний или расстройств, связанных с щелочной фосфатазой. Например, такая асфотаза альфа может вводиться субъекту, имеющему уменьшенное количество и/или не работающую эндогенную щелочную фосфатазу, или имеющему сверхэкспрессированные (например, выше нормального уровня) субстраты щелочной фосфатазы. В некоторых вариантах осуществления изобретения, асфотаза альфа в этом описании представляет собой рекомбинантный белок. В некоторых вариантах осуществления изобретения, асфотаза альфа представляет собой гибридный белок. В некоторых вариантах осуществления изобретения, асфотаза альфа в этом описании конкретно нацелена на тип клетки, ткань (например, соединительную, мышечную, нервную или эпителиальную ткань) или орган (например, печень, сердце, почки, мышцы, кости, хрящ, связки, сухожилия и т. д.). Асфотаза альфа представляет собой растворимый Fc- гибридный белок, состоящий из двух полипептидов sTNALP-Fc-D10, каждый из которых содержит 726 аминокислот, как показано в SEQ ID NO: 1. Подчеркнутые аспарагиновые (N) остатки соответствуют потенциальным сайтам гликозилирования (т.е. N 123, 213, 254, 286, 413 и 564). Подчеркнутые жирным аминокислотные остатки (L486-K487 и D715-I716) соответствуют линкерам между доменами sALP и Fc, Fc и D10, соответственно.


[00103] Каждый полипептид или мономер состоит из пяти частей. Первая часть (sALP), содержащая аминокислоты L1-S485, является растворимой частью неспецифического фермента щелочной фосфатазы ткани человека, который содержит каталитическую функцию. Вторая часть содержит аминокислоты L486-K487 в качестве линкера. Третья часть (Fc), содержащая аминокислоты D488-K714, представляет собой часть Fc иммуноглобулина гамма 1 человека (IgG1), содержащего шарнир, домены CH2 и CH3. Четвертая часть содержит D715-I716 в качестве линкера. Пятая часть содержит аминокислоты D717-D726 (D10), которые представляют собой целевой фрагмент кости, который позволяет асфотазе альфа связываться с минеральной фазой кости. Кроме того, каждая полипептидная цепь содержит шесть потенциальных сайтов гликозилирования и одиннадцать цистеиновых (Cys) остатков. Cys102 существует как свободный цистеин. Каждая полипептидная цепь содержит четыре внутрицепочечных дисульфидных связи между Cys122 и Cys184, Cys472 и Cys480, Cys528 и Cys588, Cys634 и Cys692. Две полипептидные цепи связаны двумя межцепочечными дисульфидными связями между Cys493 на обеих цепях и между Cys496 на обеих цепях. В дополнение к этим ковалентным структурным признакам считается, что щелочная фосфатаза млекопитающих имеет четыре сайта связывания металлов на каждой полипептидной цепи, включая два участка для цинка, один сайт для магния и один сайт для кальция.
Асфотаза альфа
[00104] В одном варианте осуществления изобретения, белок щелочной фосфатазы (например, гибридный белок sALP, нацеленный на кости) является асфотазой альфа (т.е. sTNALP-Fc-D10, SEQ ID NO: 1). В частности, асфотаза альфа представляет собой комплексный растворимый гликопротеин с длиной полипептида 726 аминокислот. Асфотаза альфа представляет собой Fc-гибридный белок, состоящий из 3 доменов. От N-конца до C-конца асфотаза альфа содержит: (1) растворимый каталитический домен неспецифической щелочной фосфатазы ткани человека (TNSALP) (UniProtKB/Swiss-Prot Accession No. P05186), (2) домен Fc иммуноглобулина G1 человека (UniProtKB/Swiss-Prot Accession No. P01857) и (3) дека-аспартатный пептид (D10), используемый в качестве домена нацеливания кости (Nishioka et al., 2006 Mol Genet Metab 88: 244-255). Белок связывается в гомодимер из двух первичных белковых последовательностей. Этот гибридный белок содержит 6 подтвержденных комплексных сайтов N-гликозилирования. Пять из этих сайтов N-гликозилирования расположены в домене sALP и один в домене Fc. Другая важная посттрансляционная модификация, присутствующая в асфотазе альфа представляет собой наличие дисульфидных мостиков, стабилизирующих фермент и структуру Fc-домена. Всего 4 внутримолекулярных дисульфидных мостика присутствуют на мономер, и 2 межмолекулярных дисульфидных мостика присутствуют в димере. Один цистеин домена щелочной фосфатазы является свободным.
[00105] Асфотаза альфа может использоваться как фермент-заместительная терапия для лечения гипофосфатазии (ГФФ). У пациентов с ГФФ мутация(и) потери функции в гене, кодирующем TNSALP, вызывает дефицит ферментативной активности TNSALP, что приводит к повышенным циркулирующим уровням субстратов, таким как неорганический пирофосфат (PPi) и пиридоксаль-5'-фосфат (PLP). Введение асфотазы альфа пациентам с ГФФ расщепляет PPi, высвобождая неорганический фосфат для комбинации с кальцием, тем самым способствуя образованию кристаллов гидроксиапатита и минерализации кости и восстанавливая нормальный скелетный фенотип. Более подробную информацию о асфотазе альфа и ее использовании в лечении см. в публикациях PCT WO 2005103263 и WO2008138131, принципы которых включены в настоящее описание посредством ссылки во всей их полноте. В другом варианте осуществления изобретения, асфотаза альфа может быть использована в качестве фермент-заместительной терапии для лечения нейрофиброматоза типа I (NF1). Более подробную информацию о асфотазе альфа и ее использовании (вместе с использованием других щелочных фосфатаз) в лечении NF1 см. в публикации PCT № WO 2013/058833, которая включена в данный документ путем ссылки во всей ее полноте.
Производственный процесс
[00106] Белок щелочной фосфатазы (например, асфотаза альфа) может быть получен с помощью клеток млекопитающего или другими клеток с использованием стандартных способов, известных в данной области. Такие клетки можно выращивать в культуральных чашках, стеклянных колбах или биореакторах. Конкретные способы культивирования клеток и получения рекомбинантных белков известны в данной области, как описано в работе Nelson and Geyer, 1991 Bioprocess Technol. 13:112-143 и Rea et al., Supplement to BioPharm International March 2008, 20-25. Иллюстративные биореакторы включают периодические, подпитываемые периодические и непрерывные реакторы. В некоторых вариантах осуществления изобретения, белок щелочной фосфатазы продуцируется в подпитываемом периодическом биореакторе.
[00107] Потенциальная изменчивость в физико-химической среде процесса клеточной культуры включает, например, изменение рН, температуры, состава среды для культуры клеток, изменение сырого материала, фильтрующий материал среды, разницу в масштабе биореактора, стратегию газирования (воздух, кислород и диоксид углерода) и т. д. Как описано в данном документе, на профили гликозилирования произведенного белка щелочной фосфатазы могут влиять изменения одного или более параметров.
Разработка процессов культивирования клеток
[00108] Для получения рекомбинантного белка в клеточной культуре рекомбинантный ген с необходимыми регуляторными элементами транскрипции сначала переносят в клетку-хозяина. Как правило, переносится второй ген, который придает клеткам-реципиентам избирательное преимущество. В присутствии агента селекции, который обычно применяется через несколько дней после переноса гена, выживают только те клетки, которые экспрессируют ген-селектор. Двумя популярными генами для селекции являются дигидрофолатредуктаза (DHFR), фермент, участвующий в метаболизме нуклеотидов, и глютаминсинтетаза (GS). В обоих случаях выбор происходит в отсутствие соответствующего метаболита (гипоксантин и тимидин, в случае DHFR, глутамин в случае GS), предотвращая рост нетрансформированных клеток. В общем, для эффективной экспрессии рекомбинантного белка не имеет значения, находится ли биофармацевтически-кодирующий ген и гены-селектора в одной и той же плазмиде или нет.
[00109] После селекции выживающие клетки могут переноситься как отдельные клетки во второй сосуд для культивирования, а культуры размножаются для получения клональных популяций. В конечном счете, отдельные клоны оцениваются на экспрессию рекомбинантного белка, причем самые высокие производители сохраняются для дальнейшего культивирования и анализа. Из этих кандидатов выбирают одну клеточную линию с соответствующими характеристиками роста и производительности для производства рекомбинантного белка. Затем запускается процесс культивирования, который определяется потребностями производства.
Клетки
[00110] Любые клетки млекопитающих или тип клеток не-млекопитающих, которые могут быть культивированы для получения полипептида, могут быть использованы в соответствии с настоящим изобретением. Неограничивающие примеры клеток млекопитающих, которые могут быть использованы, включают, например, клетки яичника китайского хомячка +/- DHFR (CHO, Urlaub and Chasin, 1980 Proc. Natl. Acad. Sci. USA, 77: 4216); линия миеломы мыши BALB/c (NSO/1, ECACC № доступа: 85110503); ретинобласты человека (PER.C6 (CruCell, Leiden, Нидерланды)); линия CVl почки обезьяны, трансформированная SV40 (COS-7, ATCC CRL 1651); эмбриональная линия почек человека (293 или 293 клетки, субклонированные для роста в суспензионной культуре, Graham et al., 1977 J. Gen Virol., 36:59); клетки почек детеныша хомяка (BHK, ATCC CCL 10); клетки мыши Сертоли (TM4, Mather, Biol. Reprod., 23:243-251 (1980)); клетки почек обезьяны (CVl ATCC CCL 70); клетки почек африканской зеленой обезьяны (VERO-76, ATCC CRL-I 587); клетки карциномы шейки матки человека (HeLa, ATCC CCL 2); клетки почек собаки (MDCK, ATCC CCL 34); клетки печени крысы буйвола (BRL 3A, ATCC CRL 1442); клетки легких человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); опухоль молочной железы мыши (MMT 060562, ATCC CCL51); клетки TRI (Mather et al., 1982, Annals N.Y. Acad. Sci. 383: 44-68); Клетки MRC 5; FS4-клетки; и линии гепатомы человека (Hep G2). В конкретном варианте осуществления изобретения, культивирование и экспрессия полипептидов и белков происходит из клеточной линии яичника китайского хомячка (CHO).
[00111] Кроме того, любое количество коммерчески доступных и не доступных рекомбинантных клеточных линий, которые экспрессируют полипептиды или белки, может быть использовано в соответствии с настоящим изобретением. Специалист в данной области поймет, что рекомбинантные клеточные линии могут иметь разные требования к питанию и/или могут потребовать различные условия культивирования для оптимального роста и экспрессии полипептида или белка, и будет способен модифицировать условия по мере необходимости.
[00112] Как отмечено выше, во многих случаях клетки будут выбраны или сконструированы для получения высоких уровней белка или полипептида. Часто клетки генетически сконструированы для получения высоких уровней белка, например, путем введения гена, кодирующего представляющий интерес белок или полипептид, и/или путем введения контрольных элементов, которые регулируют экспрессию гена (будь то эндогенного или введенного), кодирующего представляющий интерес полипептида.
Изменение температуры
[00113] Время действия процессов клеточной культуры, особенно не-непрерывных процессов (например, подпитываемые процессы в биореакторах), обычно ограничено оставшейся жизнеспособностью клеток, которая обычно снижается в течение культивирования. Следовательно, желательно увеличить продолжительность жизнеспособности клеток для улучшения продуцирования рекомбинантного белка. Проблемы качества продукта также мотивируют для минимизации снижения плотности жизнеспособных клеток и поддержания высокой жизнеспособности клеток, поскольку клеточная смерть может высвобождать сиалидазы в супернатанте культуры, что может снизить содержание сиаловой кислоты в экспрессированном белке. Проблемы очистки белка предлагают еще одну мотивацию для минимизации снижения плотности жизнеспособных клеток и поддержания высокой жизнеспособности клеток. Обломки клеток и содержание мертвых клеток в культуре могут отрицательно влиять на способность изолировать и/или очистить белковый продукт в конце цикла культивирования. Таким образом, сохраняя жизнеспособность клеток в течение более длительного периода времени в культуре, обеспечивают уменьшение загрязнения культуральной среды клеточными белками и ферментами (например, клеточными протеазами и сиалидазами), которые могут вызывать деградацию и конечное снижение качества желаемого гликопротеина, продуцируемого клетками.
[00114] Для достижения высокой жизнеспособности клеток в клеточных культурах могут применяться многие способы. Один из них включает снижение температуры культуры после первоначального культивирования при нормальной температуре. Например, см. Ressler et al., 1996, Enzyme and Microbial Technology 18: 423-427). Как правило, клетки млекопитающих или других типов клеток, способных экспрессировать интересующий белок, сначала выращивают при нормальной температуре для увеличения количества клеток. Такие «нормальные» температуры для каждого типа клеток обычно составляют около 37°С (например, от около 35°С до около 39°С, включая, например, 35,0°С, 35,5°С, 36,0°С, 36,5°С, 37,0°С, 37,5°С, 38,0°C, 38,5°C и/или 39,0°C). В одном конкретном варианте осуществления изобретения, температуру для получения асфотазы альфа сначала устанавливают около 37°С. Когда достигается достаточно высокая плотность клеток, температура культивирования для всей клеточной культуры затем смещается (например, уменьшается), чтобы способствовать производству белка. В большинстве случаев снижение температуры сдвигает клетки к фазе отсутствия роста G1 клеточного цикла, что может увеличить плотность и жизнеспособность клеток по сравнению с предыдущей средой с более высокой температурой. Кроме того, более низкая температура может также способствовать продуцированию рекомбинантного белка за счет увеличения скорости образования клеточного белка, облегчения посттрансляционной модификации белка (например, гликозилирования), уменьшения фрагментации или агрегации вновь продуцируемых белков, облегчения сворачивания белка и формирования трехмерной структуры (таким образом, сохраняя активность) и/или уменьшая деградацию вновь продуцируемых белков. В некоторых вариантах осуществления изобретения, нижняя температура составляет от около 30°С до около 35°С (например, 30,0°С, 30,5°С, 31,0°С, 31,5°С, 32,0°С, 32,5°С, 33,0°С, 33,5°С, 34,0°С, 34,5°С и/или 35,0°C). В других вариантах осуществления изобретения, температура для получения асфотазы альфа сначала устанавливается от около 35,0°С до около 39,0°С, а затем сдвигается от около 30,0°С до около 35,0°С. В одном варианте осуществления изобретения, температуру для получения асфотазы альфа сначала устанавливают до около 37,0°С, а затем сдвигают до около 30°С. В другом варианте осуществления изобретения, температуру для получения асфотазы альфа сначала устанавливают до около 36,5°С, а затем сдвигают до около 33°С. В еще одном варианте осуществления изобретения, температуру для получения асфотазы альфа сначала устанавливают до около 37,0°С, а затем сдвигают до около 33°С. В еще одном дополнительном варианте осуществления изобретения, температуру для получения асфотазы альфа сначала устанавливают до около 36,5°С, а затем сдвигают до около 30°С. В других вариантах осуществления изобретения, может применяться множество (например, более чем одна) ступеней изменения температуры. Например, температура может быть снижена с 37°C до 33°C, а затем далее до 30°C.
[00115] Время для поддержания культуры при определенной температуре перед смещением к другой температуре может быть определено для достижения достаточной (или желаемой) плотности клеток, сохраняя жизнеспособность клеток и способность продуцировать представляющий интерес белок. В некоторых вариантах осуществление изобретения, культуру клеток выращивают при первой температуре до тех пор, пока плотность жизнеспособных клеток не достигнет около 105 клеток/мл до около 107 клеток/мл (например, 1×105, 1,5×105, 2,0×105, 2,5×105, 3,0×105, 3,5×105, 4,0×105, 4,5×105, 5,0×105, 5,5×105, 6,0×105, 6,5×105, 7,0×105, 7,5×105, 8,0×105, 8,5×105, 9,0×105, 9,5×105, 1,0×106, 1,5×106, 2,0×106, 2,5×106, 3,0×106, 3,5×106, 4,0×106, 4,5×106, 5,0×106, 5,5×106, 6,0×106, 6,5×106, 7,0×106, 7,5×106, 8,0×106, 8,5×106, 9,0×106, 9,5×106, 1×107 клеток/мл и более) до перехода к другой температуре. В одном варианте осуществления изобретения, культуру клеток выращивают при первой температуре до тех пор, пока плотность жизнеспособных клеток не достигнет от около 2,5 до около 3,4×106 клеток/мл перед переходом на другую температуру. В другом варианте осуществления изобретения, культуру клеток выращивают при первой температуре до тех пор, пока плотность жизнеспособных клеток не достигнет от около 2,5 до около 3,2×106 клеток/мл перед переходом на другую температуру. В еще одном варианте осуществления изобретения, культуру клеток выращивают при первой температуре до тех пор, пока плотность жизнеспособных клеток не достигнет от около 2,5 до около 2,8×106 клеток/мл перед переходом на другую температуру.
[00116] В некоторых вариантах осуществления изобретения, культуру клеток выращивают при 37°C до тех пор, пока плотность жизнеспособных клеток не достигнет около 2,5-2,8×106 клеток/мл до смещения до 30°C для производства белка. В других вариантах осуществления изобретения, культуру клеток выращивают при 37°C до тех пор, пока плотность жизнеспособных клеток не достигнет около 2,5-3,4×106 клеток/мл до смещения до 30°C для производства белка.
pH
[00117] Изменение рН среды роста в культуре клеток может влиять на клеточную протеолитическую активность, секрецию и уровни продуцирования белка. Большинство клеточных линий хорошо растут при около рН 7-8. Хотя оптимальный рН для роста клеток относительно немного отличается от разных клеточных штаммов, некоторые нормальные линии клеток фибробластов лучше всего работают при рН 7,0-7,7, а трансформированные клетки обычно лучше всего работают при рН 7,0-7,4 (Eagle, 1973 The effect of environmental pH on the growth of normal and malignant cells. J Cell Physiol 82:1-8). В некоторых вариантах осуществления изобретения, рН культуральной среды для получения асфотазы альфа составляет около pH 6,5-7,7 (например, 6,50, 6,55, 6,60, 6,65, 6,70, 6,75, 6,80, 6,85, 6,90, 6,95, 7,00, 7,05, 7,10, 7,15, 7,20, 7,25, 7,30, 7,35, 7,39, 7,40, 7,45, 7,50, 7,55, 7,60, 7,65 и 7,70). В некоторых вариантах осуществления изобретения, рН культуральной среды для получения асфотазы альфа составляет около pH 7,20-7,60. В других вариантах осуществления изобретения, рН культуральной среды для получения асфотазы альфа составляет около pH 6,9-7,1. В одном конкретном варианте осуществления изобретения, рН культуральной среды для получения асфотазы альфа составляет около pH 6,9. В другом варианте осуществления изобретения, рН культуральной среды для получения асфотазы альфа составляет около pH 7,30. В еще одном варианте осуществления изобретения, рН культуральной среды для получения асфотазы альфа составляет около pH 7,39.
[00118] Все цитируемые в данном документе ссылки включены в качестве ссылки в полном объеме.
[00119] Хотя это описание было описано более подробно с помощью иллюстрации и примера для ясности понимания, специалистам в данной области очевидно, что будут выполняться некоторые незначительные изменения и модификации. Поэтому описание и примеры не должны толковаться как ограничивающие объем описания.
Примеры
Пример 1: Общий процесс производства асфотазы альфа
[00120] Как описано в данном документе, был разработан способ получения щелочных фосфатаз (например, асфотазы альфа (sTNALP-Fc-D10)).
Таблица 1. Различия между иллюстративными производственными процессами (предшествующие)
[00121] Стабильные клеточные линии СНО, экспрессирующие асфотазу альфа, были разработаны с использованием системы экспрессии GS-генов. Вторичные клоны были получены из высокопродуктивных первичных клонов в одном раунде ограниченного клонирования методом серийных разведений и была выбрана конечная клеточная линия.
[00122] Иллюстративный процесс изготовления, процесс X, описан в данном документе. Флакон Master Cell Bankl был разморожен, и весь объем флакона был ресуспендирован. Весь объем переносили в колбу для встряхивания емкостью 250 мл для роста. Образцы брались ежедневно для проверки количества и жизнеспособности (также для всех последующих этапов увеличения). Клетки расширяли с помощью нескольких стадий и инокулировали в 1000 л затравочного биореактора (N-3 низкого уровня), 1000 л затравочного биореактор (N-2 высокого уровня) и 4000 л затравочного биореактора (N-1), а затем 20000 л биореактора. После производства асфотазы альфа процесс очистки урожая использовался для удаления интактных клеток и клеточного мусора с помощью стерильной фильтрации. Затем урожай клеток был ультрафильтрован (Post Harvest UF) для концентрации и буферного разведения. Дальнейшие процессы включают, например, вирусную инактивацию (для химической инактивации вирусных частиц), хроматографию MabSelect SuRe, хроматографию с гидрофобным взаимодействием (HIC), пост HIC UF/DF (UF/DF2), капто-адгезию в смешанной хроматографии, фильтрацию вируса (с помощью исключения размера), состав (UF/DF3) и объемный наполнитель. Были выполнены многочисленные производственные процессы, в том числе, например, 2000 л-масштабные процесы и следующее увеличение до масштаба производства 20000 литров. Иллюстративные различия между процессами 2000 л (2K) и 20000 л (20K) приведены в таблицах 2 и 3. Процесс масштаба 2000 л (2K) имел более выраженную лаг-фазу и более разнообразную позднюю стадию жизнеспособности (данные не показаны).
Таблица 2. Сравнительный анализ параметров инокулята между иллюстративным процессом 2000 л и процессом 20000 л
Пассирование: 0,40×106 жизнеспособных клеток/мл
Пассирование: 0,35×106 жизнеспособных клеток/мл
Таблица 3. Сравнение параметров биореактора производства между иллюстративным 2000 л процессом и 20000 л процессом
30°C, когда плотность клеток составляет 2,5-2,8×106 жизнеспособных клеток/мл
30°C, когда плотность клеток составляет 2,5-3,4×106 жизнеспособных клеток/мл
[0123] Были сопоставлены общие выходы 2K-процессов и 20K-процессов, а также качества соответствующей полученной асфотазы альфа. Аналитические способы, используемые для сравнения характеристик продукта, включают, например, SEC-HPLC, RP-HPLC и другие способы для измерения специфической активности, концентрации белка, pH и общего содержания сиаловой кислоты (TSAC) продуцируемой асфотазы альфа. Кроме того, были проведены тесты на загрязнение и безопасность для измерения, например, остаточной ДНК, остаточного белка A, белков клетки-хозяина, бионагрузки и эндотоксина в асфотазе альфа, полученных из разных процессов. Были также проведены три дополнительных теста, то есть изоэлектрическая фокусировка (IEF), электрофорез в додецилсульфатном полиакриламидном геле (SDS-PAGE) и картирование олигосахаридов для сравнения лекарственных веществ из шкал 2K и 20K партий. Результаты этих трех тестов показали сопоставимые профили для этих партий. В заключение, качество асфотазы альфа между шкалами 2000 л и 20 000 л было сопоставимо по всем партиям.
Пример 2: Влияние температурного сдвига на производительность и качество асфотазы альфа
[0124] Среди различных производственных процессов, принятых и практикуемых для производства sTNALP-Fc-D10, изменение температуры обычно оказывает влияние на производительность и конечное качество асфотазы альфа. Температурный сдвиг от температуры роста (сравнительно высокая температура) до температуры производства (сравнительно низкая температура) был реализован во всех процессах.
[0125] Для сравнения эффектов смещения температуры по сравнению с отсутствием смещения были проведены повторяющиеся прогоны биореактора sTNALP-Fc-D10, в которых был или не было температурного сдвига. Биореакторы Sartorius 2 л и 10 л использовались для различных производственных процессов. В этом иллюстративном процессе сырье, используемое в производственных биореакторах, включало, например, производственную среду, 10% карбонат натрия, CHO подпидка и исходный раствор глютамина, как показано в таблице 4. SFM4CHO относится к бессывороточной среде для CHO.
Таблица 4. Параметр процесса в иллюстративном процессе
Способы
Дизайн экспериментов с клеточной культурой
[0126] Были реализованы два блока экспериментов для оценки влияния температурного сдвига на характеристики производительности и качества асфотазы альфа для иллюстративного процесса. Как показано в таблице 5, два прогона биореакторов (т. е. № 1 и № 2) проводились с температурным сдвигом (от 37°С до 30°С), а два других прогона (т. е. № 3 и № 4) проводились без температурного сдвига от 37°С.
Таблица 5. Условия биореакторов для процесса № 1
[0127] Плотность и жизнеспособность клеток подсчитывали с использованием счетчика клеток (то есть ViCell VR, Beckman Coulter). рН и автономный газ измеряли с использованием pHOx, а основные метаболиты, включая глюкозу и лактат, измеряли с использованием датчика (Nova Profile 100, Nova Biomedical, Уолтем, Массачусетс). Ферментативную активность измеряли стандартным способом с модификацией, согласно которой каждый образец разбавляли только один раз, а не три раза, до измерения ферментативной активности.
Способы сбора урожая и очистки
[0128] Пятьдесят микролитров образцов 10-го дня (240±4 часа) были собраны из всех биореакторов с использованием шприцев. После удаления клеток центрифугированием (3000×g, 5 мин), супернатанты осветляли с использованием 0,22 мкм фильтров горловины флакона и хранили при -80°С до очистки. В этом исследовании был применен один этап очистки высокой пропускной способности белка А. Образцы переводили в буфер (Buffer Exchange) с низким содержанием соли (5 мМ Na3PO4, pH 7,4) до аналитического анализа и анализа характеристики белка.
Аналитические способы и способы характеристики белка
[0129] Критерии качества, проанализированные в этом исследовании, включали агрегацию асфотазы альфа, фрагментацию, распределение заряда, общее содержание сиаловой кислоты (TSAC) и уровни видов нейтральных гликанов. Агрегированный уровень оценивали по проценту совокупных пиков до общего количества белка, определенного в SEC. Уровень фрагмента оценивали по процентному соотношению фрагмента к общему количеству белка, измеренному в LoC. Распределение заряда оценивали по проценту основных пиков, основного пика и кислотного пика к общему белку, соответственно, количественно определяли в AEX. Общее содержание сиаловой кислоты (TSAC) подсчитывали с помощью HPAE-PAD. Обнаружение видов нейтральных гликанов проводили с помощью масс-спектрометрии MALDI-TOF.
Результаты
Производительность клеточной культуры
[0130] В процессе № 1 заданное значение температуры было смещено от 37°C до 30°C в течение 5 часов после того, как плотность жизнеспособных клеток (VCD) достигла 25-32×105 клеток/мл. Без температурного сдвига культура клеток достигла более высокого пикового VCD, но испытала более быстрое снижение жизнеспособности (фиг. 1А). Более высокое общее потребление глюкозы наблюдалось также в условиях без температурного сдвига (фиг. 1В). Добавление болюса глюкозы применяли на 5-й день и 8-й день в условиях без температурного сдвига в соответствии с описанием процесса. Интересно, что лактат все еще потреблялся, но при значительно более низкой скорости в условиях без температурного сдвига, чем условия с температурным сдвигом (фиг. 1В).
[0131] Более высокий пиковый VCD в условиях без температурного сдвига приводил к раннему производству связывающего титра белка А (фиг. 2А). С более быстрым снижением жизнеспособности в условиях без температурного сдвига аналогичные титры связывающего белка А были достигнуты на 10-й день в обоих условиях. Суммарная специфическая продуктивность способная связываться с белком А составляла 6,4 пг/клеток/день для условия температурного сдвига и 6,7 пг/клеток/день для состояния без температурного сдвига. Интересно, что хотя подобный титр связывающего белка А наблюдался в обоих условиях, объемная активность в условиях без температурного сдвига была значительно ниже, чем условие температурного сдвига (фиг. 2В). Кроме того, специфическая активность в условиях без температурного сдвига была значительно ниже, чем условие температурного сдвига на протяжении всей продолжительности культивирования (фиг. 2C). Эти данные показывают, что температурный сдвиг от 37°C до 33°C является оптимальным для поддержания специфической активности для процесса № 1.
Все количественные показатели качества, определенные в процессе № 1, приведены в таблице 6.
Таблица 6. Количественные показатели качества для процесса № 1
[0132] Результаты AEX показали, что меньшее количество кислотных видов было получено при условии без температурного сдвига по сравнению с условием температурного сдвига (фиг. 3А). Однако значительно более высокие агрегататы были количественно оценены с помощью результатов SEC в условиях без температурного сдвига (фиг. 3B). В условиях без температурного сдвига средний LоC процент пика в невосстанавливающих условиях был выше (фиг. 4A, также в колонке «LоC (% Главный, NR)» в Таблице 6), тогда как восстановленный средний LоC процент пика был ниже (Фиг. 4B, также в колонке «LоC (% Главный, R)» в Таблице 6).
[0133] Сиалилирование было увеличено примерно в два раза в условиях без температурного сдвига (фиг. 4C).
[0134] Анализ нейтральных гликанов с помощью MALDI-TOF не обнаружил ни одного вида маннозы более высокого порядка или атипичных видов гликанов, обнаруженных в условиях без температурного сдвига (фиг. 5). Однако условие без температурного сдвига приводило к уменьшению количества A2 (преобладающих афукозилированных видов гликанов в условиях контроля с изменением температуры), более высокого фукозилирования (более высокое отношение FA2 к A2) и увеличения гликанов более высокого порядка, включая FA3G3, FA4G3, FA4G1L1 и FA4G4L1 (фиг. 5).
[0135] Влияние температуры без сдвига на производительность и качество асфотазы альфа в процессе № 1 приведено в таблице 7. Без температурного сдвига полученная асфотаза альфа имела значительно более низкую специфическую активность. Между тем, более высокая температура производства в условиях без температурного сдвига привела к более высокому сиалилированию и к более высокому фукозилированию.
Таблица 7. Влияние не меняющейся температуры на производительность и качество асфотазы альфа
Пример 3: Первоначальная оценка параметров предшествующего процесса для иллюстративного процесса №2
[0136] В этом примере суммируется первоначальная оценка параметров предшествующего технологического процесса для иллюстративного процесса производства асфотазы альфа №2 и их потенциала для воздействия на характеристики критического качества произведенной асфотазы альфа. Некоторые параметры предшествующего процесса включают в себя % агрегации, % фрагментации, сиалилирования, гликозилирования, распределения заряда и специфической активности.
Культура клеток
[0137] Все производственные процессы, упомянутые в этом исследовании, проводились либо во встряхиваемых колбах, либо в биореакторах. После оттаивания клетки выращивали с помощью серии встряхиваемых колб и вращающихся колб до инокуляции производственного биореактора. Производственный биореактор (2 л, 5 л, 10 л или 200 л) был масштабирован с использованием согласованной мощности на объем и объема барботажного газа на объем жидкости в минуту (VVM), если не указано иное. Температуру производственного биореактора контролировали при 36,5°С от 0 до примерно 120 часов и смещали до 33,0°C примерно через 120 часов, если не указано иное. Установленная точка растворенного кислорода поддерживалась на уровне 30%. Примерно 2,7% (об./об.) CPN питания на болюс добавляли через 96 часов (час), 144 часа и 192 часа, когда культуру собирали на 10 день (240 ч±6 ч), а дополнительное болюсное питание давали через 240 часов, когда культуру собирали позже 10-го дня.
Сбор и очистка
[0138] Образцы, собранные из производственных биореакторов, осветляли фильтрацией 0,22 мкм после центрифугирования при 3000 × g в течение 5 минут. Была проведена очистка осветленного отфильтрованного урожая, и степень очистки была продиктована требованиями к исследованию и испытаниям образцов. Для большинства тестируемых образцов была осуществлена либо одна хроматографическая стадия (белок А), либо две последовательные стадии хроматографии (белок А, за которым следует гидрофобная хроматография взаимодействия) (HIC). Кроме того, перед аналитическим тестированием образцы переносили (Buffer Exchange) в буфер с низким содержанием соли (5 мМ Na3PO4, pH 7,4).
Аналитическая характеристика
[0139] Характеристика качества, упомянутые в этом исследовании, были идентифицированы как подмножества более широких показателей качества, таких как чистота, эффективность и структура. Описанные подтипы качества включают в себя % агрегации, % фрагментации, распределения заряда, общего содержания сиаловой кислоты (TSAC), профиля нейтрального гликана и специфической активности. Как уровень агрегата, так и уровень фрагмента определяли с помощью относительных площадей пиков, количественно определяемых с использованием гельпроникающей ВЭЖХ (GP-HPLC). Кроме того, уровень фрагмента определяли с использованием либо SDS-PAGE или капиллярного электрофореза Lab-on-Chip (LoC). Распределение заряда оценивали с использованием анионообменной (AEX) хроматографии. TSAC определяли количественно с помощью высокоэффективной анионообменной хроматографией с импульсным амперометрическим детектированием (HPAE-PAD). Обнаружение нейтральных гликанов проводили с помощью масс-спектрометрии с лазерной десорбцией/ионизацией (MALDI) с матричной поддержкой. Специфическую активность измеряли после ферментативного анализа щелочной фосфатазы на основе паранитрофенилфосфата (pNPP).
Результаты
1. Рост и температура производства
[0140] Обсуждается влияние температуры на фрагментацию продукта. Для асфотазы альфа, продуцируемой выбранным клеточным клоном, выращенным при 36,5 °C, основная полоса асфотазы альфа была рассчитана как 95,9%, причем приблизительно 4% в виде фрагментов, измеренных с помощью SDS-PAGE. Когда температура была сдвинута с 36,5°C до 33,0°C на 5-й день, тот же клон почти не продуцировал фрагменты асфотазы альфа (основная полоса рассчитана как >99%). Другие вторичные клоны показали очень сходные тенденции в восстановлении фрагментов путем изменения температуры от 36,5°C до 33,0°C, хотя не все вторичные клоны смогли снизить уровень фрагмента до менее 1%. Поэтому температура роста и температура производства могут влиять на фрагментацию. Кроме того, время, связанное с проведением температурного сдвига, также может влиять на качество асфотазы альфы.
[0141] Более высокие агрегированные уровни, обнаруженные с помощью GP-HPLC, наблюдались при более низких температурах производства во время вторичного скрининга клона в 2 л биореакторах. На фигуре 6А показан агрегированный уровень асфотазы альфа, продуцируемый 6-ю вторичными клонами при смещении температуры до 33°С по сравнению с материалом, полученным с помощью 10 вторичных клонов, когда температура не сдвигалась (т. е. поддерживалась при 36,5°С). Эти данные свидетельствуют о том, что изменение температуры от 36,5°C до 33,0°C может привести к увеличению агрегированного уровня.
[0142] Кроме того, влияние изменения температуры на TSAC было исследовано во время первичного отбора клонов и начального развития процесса (фиг. 6B). Первичный клон использовался для исследования влияния температуры производства (5-й день и далее) на сиалилирование или TSAC в 2 л биореакторах. В этом исследовании температура производства была сдвинута с 36,5°C до 30,0°C на 5-й день при условии 1, от 36,5°C до 33,0°C на 5-й день в условиях 2, и контроль проводился без температурного сдвига. Образцы собирали, когда жизнеспособность составляла между 60% до 80%, и эти культуры проводились в течение 11-18 дней до сбора урожая в зависимости от жизнеспособности. Как показано на фиг. 6В, внедрение температурного сдвига приводило к снижению TSAC по сравнению с контролем при постоянной температуре при такой же жизнеспособности, а переход к более низкой температуре соответствовал более низкому TSAC. Требуемое минимальное значение TSAC в данном документе составляет 1,8. Поскольку сиалилирование (или TSAC) изменяет профиль заряда продукта и превращает нейтральные гликаны в отрицательно заряженные гликаны, эти три атрибута качества (температура производства, агрегация и TSAC) считаются тесно связанными.
2. pH
[0143] Предварительные результаты показали, что изменение заданного значения pH с 6,90 до 7,00 привело к увеличению TSAC (фиг. 7). Используя выбранный вторичный клон, для исследования влияния рН на сиалилирование асфотазы альфа использовали два 2 л биореактора с заданными значениями рН 6,90 и 7,00 соответственно. Образцы собирали из биореакторов на 10-й день и 12-й день соответственно. Было отмечено, что TSAC снизился с 1,9 на 10 день до 1,4 в день 12 при условии рН 6,90. Хотя та же тенденция снижения значений TSAC наблюдалась в условиях pH 7,00, значения TSAC при рН 7,00 в оба дня (2,6 на 10-й день и 2,1 в день 12) были выше, чем значения при более низком уровне рН. Эти данные свидетельствуют о том, что TSAC снижается с продолжительностью культивирования, а установка более низкого pH приводит к более низкому значению TSAC.
3. Добавление подпитки среды
[0144] Предварительные данные показывают, что отсрочка добавления первого болюса или увеличение концентрации глюкозы в среде может привести к значительному увеличению TSAC по сравнению с контрольным условием (фиг. 8A).
[0145] Кроме того, то, как подпитка среды подается (либо непрерывно, либо путем добавления болюса), также влияет на TSAC. Данные от обоих 2 л и 10 л размеров показывают, что значительно более высокие TSAC наблюдались, когда подпитка была добавлена в болюсном режиме по сравнению с режимом непрерывного добавления (фиг. 8B). Следовательно, CPN подпитка определяется как добавленная в виде болюса. Кроме того, никакого влияния на TSAC не наблюдалось, когда подпитка была доставлена путем добавления через верхнее отверстие или путем подповерхностного добавления (погружения).
4. Добавка сульфата цинка
[0146] Известно, что ионы цинка необходимы для стабильности щелочной фосфатазы (например, асфотаза альфа), поскольку она помогает поддерживать их структуру и активность. Например, два атома цинка связаны с одной молекулой плацентарной щелочной фосфатазы (Helene Le Du et al., 2001 J. Biol. Chem. 276:9158-9165). Исходя из этого соотношения, для титра 1 г/л асфотазы альфа, полученного по иллюстративному производственному процессу, разработанному в малогабаритных моделях, для активности асфотазы альфа требуется приблизительно 20 мкМ цинка.
[0147] Было отмечено, что по мере прогрессирования клеточной культуры белок А-связываемая асфотаза альфа продуцируется непрерывно, но соответствующая объемная активность не синхронизируется с титром белка А, что указывает на то, что продуцируемый белок неактивен (фиг. 9А). Кроме того, хотя объемная активность снижалась с 5-го дня до 9-го дня, активность белка немедленно восстанавливалась после добавления сульфата цинка на 9-й день. Эти данные свидетельствуют о необходимости добавления цинка в производственную среду. В этом исследовании концентрация сульфата цинка была увеличена на 25 мкМ путем добавление болюса на 9-й день. Учитывая, что титр белка А составлял приблизительно 1000 мг/л на 9-й день, это добавление болюса цинка увеличивало отношение иона цинка к асфотазе альфа, по меньшей мере до 2:1.
[0148] В следующем исследовании уровень цитотоксичности сульфата цинка на рост и жизнеспособность клеток исследовали с использованием мини-биореакторной системы в периодическом режиме, в которой сульфат цинка в разных концентрациях добавляли в культуральную среду в день 0. Когда концентрация сульфата цинка была ниже 300 мкМ, наблюдалось небольшое влияние на рост или жизнеспособность (фиг. 9В и 17С). Однако в условиях, когда концентрация цинка была больше или равна 500 мкМ, резко ухудшались как рост, так и жизнеспособность. Поэтому оптимальная концентрация общего количества цинка была ниже 500 мкМ. Таким образом, было подсчитано, что оптимальная концентрация цинка, которая должна быть внесена в день 0, может составлять между около 25 мкМ до около 300 мкМ (с учетом как роста/жизнеспособности клеток, так и проблем с функцией белка). Фактически, концентрация цинка при 150 мкМ может быть даже лучше, чем 300 мкМ, поскольку первая приводит к меньшему ингибированию роста клеток после 5-го дня (фиг. 9А). Также удивительно, что хотя 20 мкМ цинка теоретически достаточно для получения функциональной асфотазы альфа (т.е. 2 ионов цинка на активный фермент), фактическое добавление цинка может потребовать значительно более высоких концентраций цинка (например, 150 мкМ) в щелочных фосфатах (например, асфотаза альфа, TNALP, PALP, GCALP, IAP или их гибридных белков/вариантов белков) производственных процессов.
5. Плотность посева
[0149] Как правило, более высокая плотность посева приводит к более высокой пиковой плотности жизнеспособных клеток, что в первую очередь связано с определенными ограничениями на питательные компоненты в производственной среде. Поскольку подпитка и температурный сдвиг основаны на продолжительности культивирования, плотность посева в сочетании с другими параметрами предшествующего процесса, особенно с временем температурного сдвига, может потенциально влиять на качество асфотазы альфа.
6. Время сбора
[0150] Время сбора в этом иллюстративном процессе составляло 240±6 часов. Было продемонстрировано влияние времени сбора на TSAC (см., например, фигуру 7). Кроме того, время сбора также считается связанным с жизнеспособностью (см., например, фигуру 14), а снижение жизнеспособности к концу культуры указывает на то, что время сбора может потенциально влиять на качество асфотазы альфа.
[0151] Были оценены различные параметры предшествующего процесса, включая контроль плотности посева, температуры, рН, DO, стратегии газирования, перемешивания, CPN подпитки, добавления глюкозы, добавления галактозы, добавления цинка и времени сбора. Были оценены дополнительные параметры предшествующего процесса, включая % агрегации, % фрагментации, сиалилирование, гликозилирование, распределение заряда и специфическую активность, например, как описано ниже.
[0152] Температура оказывает сильное влияние на агрегацию, фрагментацию и сиалилирование. pH также влияет на сиалилирование асфотазы альфа. Стратегия газирования и/или перемешивания, по-видимому, влияют на жизнеспособность. Добавление CPN подпитки, по-видимому, влияет на сиалилирование. Было показано, что добавление цинка является необходимым для поддержания ферментативной активности, и количество добавления цинка (150 мкМ) определяют на основании представленных исследований. Было показано, что время сбора связано с жизнеспособностью, а TSAC снижается при приближении к концу культивирования.
Пример 4: Дальнейшая оценка параметров предшествующего процесса для иллюстративного процесса №3
[0153] Этот пример суммирует дальнейшую характеристику предшествующих процессов, которые были выполнены с целью скрининга параметров технологического процесса биореактора. Для начальных оценок в этом иллюстративном процессе был проверен список параметров предшествующего процесса, включающий рН продуцирования культуры, температуру, плотность посева, стратегию газообразования, перемешивание, CPN подпитку, добавление глюкозы, добавление галактозы и время сбора.
СОКРАЩЕНИЯ
[0154] CPP: параметр критического процесса, параметр процесса (вход процесса), который влияет на критическую характеристику качества и, следовательно, будет контролироваться и контролироваться, чтобы гарантировать, что процесс дает желаемое качество.
CQA: критическая характеристика качества
ВЭЖХ: Высокоэффективная жидкостная хроматография
LoQ: Предел количественной оценки
P/V: Мощность на объем
Материалы и способы
[0155] Были дополнительно исследованы параметры процесса, которые были показаны в начальных экспериментах с потенциальным воздействием. Шесть блоков биореакторов были проведены в следующем процессе характеристики исследования (таблица 8). Три уровня для каждого параметра процесса были протестированы со срединным уровнем как текущее заданное значение (контроль) с учетом возможностей контроля при изготовлении и тестировании диапазона параметров процесса. В блоке с 1 по 5 реализован трехуровневый полный факторный дизайн с двумя репликационными элементами контроля. В блоке 6 были проведены четыре комбинации экстремальных P/V и зарядов VVM плюс два элемента контроля.
Таблица 8. Исследуемые параметры процесса и уровни
(105 клеток/мл)
Культура клеток
[0156] Все производственные процессы, упомянутые в этом исследовании, проводились либо в 2 л, либо в 10 л биореакторах. После оттаивания клетки выращивали с помощью серии встряхиваемых колб и вращающихся колб до инокуляции производственного биореактора. Производственный биореактор (2 л или 10 л) был масштабирован с использованием P/V и барботирования VVM, если не указано иное. В условиях контроля параметры процесса клеточной культуры были следующими. Начальная pH культуры была 6,90 с 0,05 мертвой зоной. Температуру контролировали при 36,5°С от 0 до 120 часов и смещали до 33°С через 120 часов. Установленная точка растворенного кислорода поддерживалась на уровне 30%. Начальная величина P/V составляла 17 Вт/м3 (0-96 часов) и сдвигалась до 81 Вт/м3 перед первым добавлением болюсной подпитки через 96 часов. 2,67% (об./об.) CPN подпитки на болюс добавляли через 96 часов, 168 часов и 192 часа. Глюкоза поддерживалась на уровне≤2 г/л. Если глюкоза была ниже 2 г/л, ежедневное добавление болюса применяли для увеличения концентрации глюкозы в диапазоне от 1,8 до 2,0 г/л. Галактозу добавляли по всей культуре: 2,0 г/л в день 0 в день 4 и 1,0 г/л ежедневно с 5-го дня до 9-го дня. Часть культуры собирали на 10-й день (240±4 часа) и на 11-й день (264±4 часа).
Сбор и очистка
[0157] Образцы, собранные из производственных биореакторов, осветляли фильтром 0,22 мкм после центрифугирования при 3000 × g в течение 5 минут. Была проведена очистка осветленного отфильтрованного урожая. Степень очистки была продиктована требованиями к исследованию и испытаниям образцов. Была реализована либо одна стадия (белок А), либо две последовательные стадии (белок А и HIC). Кроме того, перед аналитическим тестированием образцы переносили в буфер с низким содержанием соли (5 мМ Na3PO4, pH 7,4).
Аналитическая характеристика
[0158] Критерии качества, указанные в этом исследовании, включают в себя % агрегации, % фрагментации, распределение заряда, общее содержание сиаловой кислоты (TSAC) и профиль нейтрального гликана. Как агрегированный уровень, так и уровень фрагмента определяли с помощью относительных площадей пиков, количественно определяемых с использованием эксклюзионной хроматографии (SEC). Кроме того, чистоту асфотазы альфа (основной пик) определяли также с помощью капиллярного электрофореза на основе Lab-on-Chip (LoC). Распределение заряда оценивали с использованием анионообменной хроматографии (AEX). TSAC определяли количественно с помощью высокоэффективной анионообменной хроматографией с импульсным амперометрическим детектированием (HPAE-PAD). Обнаружение видов нейтральных гликанов проводили с помощью время-пролетной ионизации лазерной десорбции с использованием матрицы (MALDI-TOF) масс-спектрометрии. Объемную активность измеряли после ферментативного анализа щелочной фосфатазы на основе паранитрофенилфосфата (pNPP).
Статистический анализ и визуализация данных
[0159] Потенциал тестируемых параметров процесса для воздействия на производительность и каждый атрибут качества оценивался с использованием статистического программного пакета JMP (Кэри, Северная Каролина). Были проанализированы три непрерывных параметра (три уровня для двух испытанных параметров процесса, соответственно, и два уровня для сбора). Сообщалось об индивидуальных значениях p, рассчитанных с помощью двухстороннего t-теста, и 0,05 был выбран в качестве порога значимости. Контурные графики, созданные с помощью JMP, были использованы для визуализации потенциального воздействия тестируемых параметров на каждый атрибут качества.
Результаты
[0160] Поскольку приемлемый диапазон для каждого CQA в этом иллюстративном процессе все еще находился на исследовании, был создан иллюстративный экспериментальный диапазон и использовался для целей сравнения (таблица 9). Исходные данные были получены из всех результатов 10-го дня, полученных в этом исследовании. Для измерения примеси, включая фрагмент и агрегат с помощью SEC, а также базового и кислотного пика с помощью AEX, в качестве верхнего порога использовалось среднее значение плюс двух кратное стандартное отклонение. Поскольку рассчитанный верхний порог для фрагмента, измеренный с помощью SEC и базовый пик, измеренный с помощью AEX, ниже, чем LoQ (1%) анализа SEC и AEX, верхний порог был установлен равным 1% для обоих качеств. Что касается TSAC, то в качестве экспериментального диапазона использовалось среднее значение, минус и плюс двухкратное стандартное отклонение.
Таблица 9. Маломасштабные экспериментальные диапазоны для CQA
[0161] Значимость прямого воздействия каждого параметра процесса на каждый CQA суммируется в таблице 10. p-значение P/V и барботажное воздействие VVM не было рассчитано из-за отсутствия статистической мощности. p-значения, меньшие или равные 0,05, считаются статистически значимыми.
Таблица 10. Значение прямого воздействия каждого параметра процесса на CQA, проанализированное с помощью JMP
сдвига
[0162] В таблице 11 суммируется среднее значение характеристики качества, полученное на уровне минимального и максимального тестируемого параметров процесса. Например, в исследовании добавления глюкозы и галактозы, три биореактора использовали 1,5 г/л в качестве порога добавления глюкозы, и каждый из них имел другой уровень добавления галактозы. Затем значение CQA при 1,5 г/л порогового значения глюкозы рассчитывали, используя среднее значение из этих трех биореакторов. В следующих разделах данные дня 10 используются для обсуждения качества, если не указано иное.
Таблица 11. Среднее значение характеристики качества при протестированных условиях
(0-96 ч);
41 (96-240 ч);
(0-96 ч);
121
(96-240 ч);
Примечание: Когда средний процент основного пика и кислотного пика, измеренный с помощью AEX, был рассчитан для состояния 15 г/л общего добавления галактозы, образец выброса был исключен из-за его низкого пика ответа в анализе. Его биологический повторный образец из другого биореактора остался для целей расчета.
1. pH
[0163] Предыдущие результаты показали, что pH влияет на TSAC продуцируемой асфотазы альфа. Потенциальное влияние рН культуры и температуры производства на производство и качество асфотазы альфа было дополнительно исследовано в 2 л биореакторах. Была реализована полная факториальная конструкция, содержащая три уровня рН культуры (6,75, 6,90 и 7,10) и три уровня температуры производства (30,0°C, 33,0°C и 35,0°C), и образцы из всех условий собирали на 240±4 часа и 264±4 часа, соответственно. Перед аналитическим анализом применяли двухступенчатую очистку (ProA+HIC).
[0164] Установка pH для культуры в тестируемом диапазоне от 6,75 до 7,10 не считалось критическим параметром процесса (CPP) в том, что все протестированные CQA были в пределах небольшого экспериментального диапазона, указанного в таблице 9. Однако было показано, что рН оказывает значительное влияние на несколько протестированных характеристик качества. Повышение заданного значения pH с 6,75 до 7,10 привело к увеличению TSAC с 1,9 до 2,7 при температуре производства после сдвига 33°C (фиг. 10A). рН также оказал значительное влияние на фрагментацию, измеренную с помощью SEC и основным пиком, измеренным с помощью AEX (таблица 10), но воздействие было меньше или равно 0,2% (таблица 11), что было ниже вариации анализа на основе ВЭЖХ равной 0,5%. Кроме того, при контрольном состоянии (рН 6,90 и температуре продукта 33,0°С) была достигнута максимальная производительность как по титру белка А, так и по объемной активности (фиг. 10В и 10С).
[0165] Данные показывают, что воздействие заданного значения рН в диапазоне от 6,75 до 7,10 на CQA находилось в пределах экспериментального диапазона, предполагая, что pH культуры производства вряд ли будет CPP. В различных вариантах осуществления изобретения, щелочная фосфатаза (например, асфотаза альфа) получается при заданном значении рН 6,65, 6,70, 6,75, 6,80, 6,85, 6,90, 6,95, 7,00, 7,05, 7,10, 7,15, 7,2 или выше.
2. Температура
[0166] Предварительные результаты показали, что температура оказывает сильное влияние на несколько параметров, включая агрегацию, фрагментацию и TSAC. Как и в предыдущих процессах использующих температурный сдвиг от 36,5°C до 33,0°C через 120 часов, исследование воздействия температуры расчленялось на три параметра: температура роста (от 0 до 120 часов), температура производства (от 120 часов до сбора) и время сдвига температуры. Эти три параметра, связанные с температурой, были дополнительно исследованы отдельно и суммированы ниже.
2.1 Температура Роста
[0167] Влияние температуры роста было исследовано вместе с заданной точкой DO в 2 л биореакторах. Три уровня температуры роста (35,0°C, 36,5°C и 37,5°C) поддерживались от 0 до 120 часов в разных условиях испытания с последующим температурным сдвигом до 33,0°C через 120 часов. Образцы из всех условий собирали в течение 240±4 часа и 264±4 часа. Перед аналитическим анализом применяли одноэтапную очистку (ProA). Температура роста влияет на темпы роста и жизнеспособность. Более высокая температура роста коррелировала с более быстрыми темпами роста на ранних стадиях, после чего последовало быстрое снижение температуры после температурного сдвига, что привело к более низкому пику VCD. Кроме того, культуры с более высокой температурой роста также имели конечную жизнеспособность на 2-4% ниже в день сбора. Интересно, что температура роста значительно влияет на производительность, а объемная активность достигла пикового значения при температуре роста 36,5°C (фиг. 11A).
[0168] Что касается влияния на качество асфотазы альфа, то, как было показано, температура роста оказывает значительное влияние только на фрагментацию, измеренную с помощью SEC (фиг. 11B). Увеличение температуры роста от 35,0°С до 37,5°С привело к тому, что уровень фрагмента был увеличен от 0,3% до 0,8%. Отмечается, что эти измеренные значения были ниже LoQ (1%) анализа SEC.
[0169] Поскольку воздействие на CQAs с помощью температуры роста находилось в пределах экспериментального диапазона (таблица 9 и таблица 11), температура роста в пределах от 35,0°C до 37,5°C вряд ли будет CPP. В различных вариантах осуществления изобретения, щелочную фосфатазу (например, асфотаза альфа) получают при температуре роста 33,0°C, 33,5°C, 34,0°C, 34,5°C, 35,0°C, 35,5°C, 36,0°C, 36,5°C, 37,0°С, 37,5°С, 38,0°С или 38,5°С. В одном конкретном варианте осуществления изобретения, щелочную фосфатазу (например, асфотаза альфа) получаются при температуре роста 35,0°C, 36,5°C, 37,0°C или 37,5°C.
2.2 Температура производства
[0170] Влияние температуры производства (30,0°C, 33,0°C и 35,0°C) на CQA было исследовано вместе с установленной величиной рН культуры в 2 л биореакторах. Перед аналитическим анализом применяли одноэтапную очистку (ProA).
[0171] Температура производства значительно влияет на CQA, особенно TSAC, агрегацию и фрагментацию, измеренную с помощью SEC, и кислотный пик, измеренный с помощью AEX (таблица 10 и таблица 11). Повышение температуры производства от 30,0°C до 35,0°C привело к среднему увеличению TSAC с 1,6 до 2,7 (фиг. 10A), а снижение температуры производства от 35,0°C до 30,0°C привело к значительному увеличению агрегатов, измеренному с помощью SEC, от 3,8% до 8,2% (фиг. 12А). Кроме того, кислотный пик, измеренный с помощью AEX, увеличился от 3,7% до 8,5% при снижении температуры производства (фиг. 12В). Эти данные свидетельствуют о том, что температура производства выше чем 30,0°C, вероятно, полезна для снижения риска низкого TSAC и предотвращения увеличения агрегации/кислотного пика. Между тем, как представляется, существует очень тонкая положительная корреляция между температурой производства и фрагментацией, измеренная с помощью SEC (фиг. 12C), и основным пиком, измеренным с помощью AEX (фиг. 12D). Эти данные предполагают, что температура производства ниже, чем 35°C может быть предпочтительнее, чтобы снизить риск фрагментации асфотазы альфа.
[0172] С точки зрения воздействия на производительность, температура производства оказывает наибольшее влияние на титр и объемную активность белка А (фиг. 10В и 10С). В диапазоне от 30,0°С до 35,0°С повышение температуры производства коррелирует с увеличением производительности. Однако при рН ниже 7,10 температура производства взаимодействует с pH для повышения производительности, при этом максимальная объемная производительность происходит при температуре производства 33,0°C. Было показано, что пиковая продуктивность как по объемной активности, так и по титру белка А наблюдается при рН 6,90 при температуре производства, которая выше или равна 33,0°С. Хотя, как было показано, температура производства оказывает значительное влияние на несколько CQA (таблица 10), воздействие все еще находилось в пределах экспериментального диапазона (таблица 9 и таблица 11). Поэтому температура производства в диапазоне от 30,0°C до 35,0°C вряд ли будет CPP.
В различных вариантах осуществления изобретения, щелочную фосфатазу (например, асфотазу альфа) получают при температуре производства 29,0°C, 29,5°C, 30,0°C, 30,5°C, 31,0°C, 31,5°C, 32,0°C, 32,5°C, 33,0°C, 33,5°C, 34,0°C, 34,5°C, 35,0°C, 35,5°C, 36°C или более высокой температуре. В одном конкретном варианте осуществления изобретения, щелочную фосфатазу (например, асфотазу альфа) получают при температуре производства 30,0°C, 30,5°C или 35,0°C.
3. Время температурного сдвига
[0173] Влияние времени температурного сдвига на CQA было исследовано вместе с плотностью посева в 2 л биореакторах. Были протестированы три временных интервала температурного сдвига (108 часов, 120 часов и 132 часа) и образцы из всех условий были собраны через 240±4 часа и 264±4 часа. Перед аналитическим анализом применяли одноэтапную очистку (белок A).
[0174] Время температурного сдвига оказывает незначительное влияние на объемную активность (фиг. 13А). При высокой плотности посева отсрочка времени температурного сдвига от 108 до 132 часов, по-видимому, уменьшает объемную активность. Однако при более низкой плотности высева максимальная объемная активность наблюдается при условии более позднего температурного сдвига.
[0175] Было показано, что с точки зрения качества асфотазы альфа, время температурного сдвига оказывает существенное влияние на фрагментацию, как измерено с помощью SEC (фиг. 13B). При условии последнего температурного сдвига (132 часа) средний уровень фрагмента составлял 0,8%, близкий к маломасштабному экспериментальному диапазону для фрагментации (<1,0%). Поэтому более узкое окно времени температурного сдвига, вероятно, полезно для снижения риска фрагментации.
[0176] Поскольку влияние на CQA с помощью времени температурного сдвига находилось в пределах экспериментального диапазона (таблица 9 и таблица 11), время температурного сдвига в диапазоне от 108 до 132 часов вряд ли будет CPP.
[0177] В различных вариантах осуществления изобретения, щелочную фосфатазу (например, асфотазу альфа) получают с помощью процесса, в котором происходит температурный сдвиг от около 100 часов, 105 часов, 108 часов, 110 часов, 115 часов, 120 часов, 125 часов, 130 часов, 132 час, 135 часов, 140 часов, 145 часов или 150 часов после начальной точки роста (например, инокуляции). В одном конкретном варианте осуществления изобретения, щелочную фосфатазу (например, асфотазу альфа) получают с помощью процесса, в котором происходит температурный сдвиг через около 108 часов, 120 часов или 132 часа. В различных вариантах осуществления изобретения, щелочную фосфатазу (например, асфотазу альфа) собирают в момент времени около 200 часов, 210 часов, 220 часов, 230 часов, 240 часов, 250 часов, 260 часов, 264 часа, 270 часов, 280 часов, 288 часов (т. е. 12 дней) или более чем 12 дней. В одном конкретном варианте осуществления изобретения, щелочную фосфатазу (например, асфотазу альфа) собирают в момент времени около 240, 264 или 288 часов.
4. Подпитка среды
[0178] Предыдущие результаты показали, что добавление подпитки влияет на сиалилирование. В качестве контроля для этого иллюстративного процесса три болюса среды добавляли в производственный биореактор на 4-й день, 6-й день и 8-й день, и общее количество было эквивалентно 8% (об./об.) исходного объема культуры. При неизменном количестве болюсов были протестированы три количества подпитки: 67%, 100% (контроль) и 133%. С точки зрения времени добавления болюса были исследованы три стратегии: (1) 3-й день, 5-й день и 7-й день; (2) 4-й день, 6-й день и 8-й день (контроль); (3) 5-й день, 7-й день и 9-й день. Все биореакции были проведены в 2 л биореакторах, и образцы из всех условий собирали в течение 240±4 часа и 264±4 часа. Перед аналитическим анализом применяли одноэтапную очистку (белок A).
[0179] Хотя среднее количество подпитки и время инициации напрямую не влияли на производительность, их взаимодействие, по-видимому, оказало незначительное влияние на объемную активность (фиг. 14A). В условиях контроля была достигнута максимальная объемная активность. Когда была обеспечена подпитка на 33% меньше, отсрочка начала подпитки приводила к более низкой объемной активности.
[0180] Повышенный кислотный пик (10,1%) и агрегация (9,5%), наблюдаемые при более высоком (133%) и более раннем (первый болюс в день 3) состоянии подпитки, близок к маломасштабному экспериментальному диапазону (10,2% для кислотного пика и 9,6% для агрегации, таблица 9), что указывает на то, что по меньшей мере один параметр требует более жесткого контроля в масштабах, например, 15% изменение среднего количества подпитки на болюс и/или относительно короткое временное окно добавки (96±3 часа, 144±3 часа и 192±3 часа). Поскольку воздействие на CQA путем добавления подпитки среды находилось в пределах экспериментального диапазона (таблица 9 и таблица 11), среднее количество подпитки среды в диапазоне от 67% до 133% и время подпитки (96±24 часа, 144±24 часа и 192±24 часа) вряд ли будет CPP.
[0181] В различных вариантах осуществления изобретения, щелочную фосфатазу (например, асфотазу альфа) получают с помощью процесса в котором дополнительные болюсы культуральной среды добавляют в производственный биореактор. Например, можно добавить один, два, три, четыре, пять, шесть или более болюсов культуральной среды. В одном конкретном варианте осуществления изобретения, добавляют три болюса культуральной среды. В различных вариантах осуществления изобретения, такие дополнительные болюсы культуральной среды могут быть добавлены в различных количествах. Например, такие болюсы культуральной среды могут быть добавлены в количестве около 20%, 25%, 30%, 33%, 40%, 45%, 50%, 60%, 67%, 70%, 75%, 80%, 90%, 100%, 110%, 120%, 125%, 130%, 133%, 140%, 150%, 160%, 167%, 170%, 175%, 180%, 190%, 200% или более, исходного объема культуральной среды в производственном биореакторе. В одном конкретном варианте осуществления изобретения, такие болюсы культуральной среды могут быть добавлены в количестве около 33%, 67%, 100% или 133% от исходного объема. В различных вариантах осуществления изобретения, такое добавление дополнительных болюсов может происходить в разное время во время роста клеток или периода производства белка. Например, болюсы могут быть добавлены на 1-й день, 2-й день, 3-й день, 4-й день, 5-й день, 6-й день, 7-й день, 8-й день, 9-й день, день 10, день 11, день 12 или позже в процессе. В одном конкретном варианте осуществления изобретения, такие болюсы культуральной среды могут быть добавлены через день (например, в (1) 3-й день, 5-й день и 7-й день; (2) 4-й день, 6-й день и 8-й день; или (3) 5-й день, 7-й день и 9-й день. На практике частота, количество, момент времени и другие параметры болюсных добавок культуральной среды могут свободно объединяться в соответствии с вышеуказанным ограничением и определяться экспериментальной практикой.
5. Плотность посева
[0182] В этом исследовании влияние плотности посева на CQA было исследовано вместе с временем температурного сдвига в 2 л биореакторах. Были протестированы три плотности посева (4,0×105 клеток/мл, 5,5×105 клеток/мл и 8,0×105 клеток/мл), и образцы из всех условий собирали через 240±4 часа и 264±4 часа. Перед аналитическим анализом применяли одноэтапную очистку (белок A).
[0183] Время температурного сдвига оказывает очень сильное влияние на производительность. Как показано на фиг. 13А, увеличение плотности посева от 4,0×105 клеток/мл до 8,0×105 клеток/мл привело к увеличению объемной активности от 806 Ед/мл до 1012 Ед/мл в среднем.
[0184] Что касается качества асфотазы альфа, было показано, что плотность посева значительно влияет на фрагментацию, как измерено с помощью SEC (фиг. 13B). При самом высоком уровне плотности посева (8,0×105 клеток/мл) средний уровень фрагмента составлял 0,8%, близкий к маломасштабному экспериментальному диапазону для фрагментации (<1,0%). Поэтому более плотный диапазон плотности посева может быть полезен для снижения риска образования фрагментов в масштабах.
[0185] Поскольку воздействие на CQA с помощью плотности посева находилось в пределах экспериментального диапазона (таблица 9 и таблица 11), плотность посева в пределах от 4,0×105 клеток/мл до 8,0×105 клеток/мл вряд ли будет CPP. В различных вариантах осуществления изобретения, щелочную фосфатазу (например, асфотазу альфа) получают способом, в котором клетки высевают с плотностью около 1,0×105 клеток/мл, 1,5×105 клеток/мл, 2,0×105 клеток/мл, 2,5×105 клеток/мл, 3,0×105 клеток/мл, 3,5×105 клеток/мл, 4,0×105 клеток/мл, 4,5×105 клеток/мл, 5,0×105 клеток/мл, 5,5×105 клеток/мл, 6,0×105 клеток/мл, 6,5×105 клеток/мл, 7,0×105 клеток/мл, 7,5×105 клеток/мл, 8,0×105 клеток/мл, 8,5×105 клеток/мл, 9,0×105 клеток/мл, 9,5×105 клеток/мл, 1,0×106 клеток/мл, 1,5×106 клеток/мл, 2,0×106 клеток/мл или более высокой плотности. В одном конкретном варианте осуществления изобретения, в таких процессах клетки высевают в плотности около 4,0×105 клеток/мл, 5,5×105 клеток/мл или 8,0×105 клеток/мл.
6. Время сбора
[0186] Предварительные данные показывают, что отсрочка сбора была связана с жизнеспособностью и сокращением TSAC, поэтому время сбора может оказать потенциальное влияние на другие CQA. Во всех исследованиях характеристик процесса образцы собирали с 10-го дня (240±4 часа) и 11-го дня (264±4 часа) из всех биореакторов. Десять 2 л биореакторов служили в качестве контрольного условия во множественных блоках исследования характеристик процесса. Поскольку все были выполнены в тех же условиях с текущими настройками параметров процесса, они позволили провести исследование времени сбора по производительности и качеству асфотазы альфа. Аналитический результат этих десяти 2 л биореакторов приведен в таблице 12. Отмечается, что образцы из двух биореакторов (№ 18 и № 19) очищали с помощью двухступенчатой очистки (ProA+HIC), а образцы из других восьми биореакторов (№ 10-№ 17) очищали только с помощью ProA. Следует отметить, что HIC способен повысить чистоту образца без воздействия на TSAC.
Таблица 12. Аналитический результат из партий иллюстративных 2 л биореакторов
(День)
[0187] В каждом отдельном исследовании время сбора рассматривалось как третий параметр для статистического анализа. Как показано в каждом отдельном исследовании, отсрочка сбора с 10-го дня до 11 дня привела к увеличению титра белка А и объемной активности на приблизительно 10-15%.
[0188] Значительное влияние времени сбора на TSAC наблюдалось во всех пяти отдельных исследованиях. Задержка сбора за один день привела к снижению TSAC примерно на 0,3-0,4 моля сиаловой кислоты на моль белка в среднем, хотя TSAC в обоих условиях все еще находился в пределах маломасштабного экспериментального диапазона (1,5-2,9), как указано в таблице 9. Из-за значительного влияния времени сбора на TSAC, ранний сбор может быть выполнен для увеличения TSAC.
[0189] Что касается агрегирования, как измерено с помощью SEC, и кислого пика, как измерено с помощью AEX, было показано, что время сбора статистически значимо влияет на три из пяти исследований (таблица 13).
[0190] Однако величина воздействия на CQA была незначительной, менее чем 1% по сравнению с контрольными условиями в каждом отдельном исследовании. Кроме того, все значения упали в маломасштабных экспериментальных диапазонах, которые указаны в таблице 9 (≤9,6% для агрегатов и≤10,2% для кислотного пика).
[0191] В одном из пяти исследований на уровни фрагментов влияли сроки сбора (см. Таблицу 13). Однако задержка сбора на один день привела к увеличению фрагмента только на 0,1-0,2% в двух условиях контроля, а уровень фрагмента на 11-й день все еще был ниже LoQ (1%) анализа SEC. Аналогичным образом, уровень основного пика был повышен с 0,2-0,3% на 10 день до 0,4% на 11-й день с p-значением 0,03 (таблица 13). Однако оба значения были все еще намного ниже LoQ (1%) анализа AEX. Поэтому время сбора, начиная с 10-го дня (240±4 часа) до 11 дня (264±4 часа), вряд ли будет CPP.
Таблица 13. Влияние сроков сбора на CQA от индивидуальных исследований
Вывод
[0192] Тестируемые параметры процесса в этом исследовании включают в себя заданное значение рН культуры, температуру, плотность посева, количество и время подачи подпитки, добавление глюкозы, добавление галактозы и время сбора. В таблице 14 приведены все используемые параметры процесса.
Таблица 14. Сводка параметров процесса
Пример 5. Высокоразрешающее исследование влияния рН и температуры производства на характеристику качества асфотазы альфа для иллюстративного процесса №4
[0193] В дополнение к предыдущим оценкам, это исследование обеспечивает более высокий разрешающий эффект воздействия рН культуры и температуры производства (≥120 часов) на производительность и качество асфотазы альфа в иллюстративном производственном процессе. Протестированные сценарии включают: 1) различные уровни рН (т.е. 6,70, 6,80, 6,90, 7,00 и 7,10) и одинаковая температура производства 33,0°С; и 2) различные уровни температуры производства (т.е. 31,0°C, 32,0°C, 33,0°C, 34,0°C и 35,0°C) и заданное значение pH 6,90. Образцы из всех условий собирали через 240±4 часов, а белок A-очищенные материалы испытывали на несколько характеристик качества (например, процентную фрагментацию, процентную агрегацию, распределение заряда, общее содержание сиаловой кислоты (TSAC) и покрытие нейтральных гликанов. Это исследование подтвердило, что процесс протекающий при заданном значении рН в диапазоне от 6,70 до 7,10 и температуре производства от 31,0°С до 35,0°С, поддерживает все CQA.
Культура клеток
[0194] Все эксперименты, проведенные в этом исследовании, использовали одноразовые биореакторы Mobius 3L (CR0003L200, EMD Millipore). Список параметров процесса, используемых в этом исследовании, представлен в таблице 15. Кроме того, исследовали четыре дополнительных условия рН в культуре с контрольной температурой производства (33,0°С) и четыре дополнительных температуры с pH контрольной культуры (6,90) (таблица 16).
Таблица 15. Параметры процесса клеточной культуры для иллюстративного процесса № 4
Таблица 16. Протестированные условия
[0195] Плотность и жизнеспособность клеток подсчитывали с использованием ViCell VR. рН и автономные газы измеряли с использованием pHOx, а основные метаболиты, включая глюкозу и лактат, измеряли с использованием Nova Profile 100 (Nova Biomedical, Уолтам, Массачусетс). Ферментативную активность измеряли стандартными способами с модификацией, согласно которой каждый образец разбавляли только один раз, а не три раза, до измерения ферментативной активности.
Сбор и очистка
[0196] Пятьдесят микролитров образцов 10-го дня (240±4 часа) были собраны из всех биореакторов с использованием шприцев. После удаления клеток центрифугированием (3000×g, 5 мин), супернатанты осветляли с использованием 0,22 мкм фильтров горловины флакона и хранили при -80°С до очистки. В этом исследовании был применен один этап высокопропускной очистки на основе пластин (белок А). Образцы переводили в буфер с низким содержанием соли (5 мМ Na3PO4, pH 7,4) до аналитического анализа и анализа характеристики белка.
Аналитическая характеристика и характеристика белка
[0197] Критические характеристики качества, проанализированные в этом исследовании, включали процент агрегата, процент фрагмента, распределение заряда, TSAC и распределение нейтральных гликанов. Уровень агрегата оценивали по проценту совокупных пиков до общего количества белка, определенного в SEC. Уровень фрагмента оценивали по процентному соотношению фрагмента к общему количеству белка, определенного в SEC, а также в LoC. Распределение заряда оценивали по проценту основных пиков, основного пика и кислотного пика к общему белку, соответственно, количественно определенному с помощью AEX. Уровень сиалилирования (или TSAC) был определен с помощью HPAE-PAD. Обнаружение видов нейтральных гликанов проводили с помощью масс-спектрометрии MALDI-TOF. Изоэлектрическую фокусировку проводили с использованием системы iCE280.
Результаты
Производительность клеточной культуры
[0198] Как отмечено, как pH культуры, так и температура производства влияют на производительность клеток. Низкое значение рН (рН=6,70) приводило к наименьшей скорости роста и минимальному максимальному VCD, с достижением максимальной плотности 11,1×106 клеток/мл (фиг. 15А) в день сбора (день 10). Более высокие значения рН (рН=7,00 и 7,10) приводили к снижению жизнеспособности на 1-2% по сравнению с другими условиями на 10-й день (фиг. 15В).
[0199] Более высокие температуры производства также были связаны с усилением роста (фиг. 16А) и более быстрым снижением жизнеспособности к концу культивирования (фиг. 16В). Более низкая температура производства привела к более низкому пику VCD, но обеспечила сравнимую жизнеспособность с контрольной точкой 33,0°C.
[0200] Повышение рН культуры также вызывало гораздо более быстрое потребление глюкозы (фиг. 17А) и вызывало как большее образование лактата на ранних стадиях культуры (до температурного сдвига), так и большее общее накопление лактата в течение всего культивирования (фиг. 17B).
[0201] Клетки культуры выращивали в течение пяти дней (120 ч) до смещения температуры роста до температуры производства. Оба профиля глюкозы и лактата были очень сопоставимы между днем 0 и днем 5 (фиг. 18А и фиг. 18В). После температурного сдвига более высокие температуры производства были связаны с более высоким уровнем потребления глюкозы. Небольшое снижение максимальной концентрации лактата наблюдалось для условий температуры производства 33,0°C по сравнению с другими условиями. Кроме того, температура производства 35,0°C показала значительное снижение потребления лактата в течение 9 и 10 дней в культуре по сравнению со всеми другими условиями.
[0202] Ранее наблюдалось, что температура производства имела более драматический эффект, чем рН культуры, как на титр белка А, так и на специфическую активность для диапазона температур производства между 30,0°С до 35,0°С и рН культуры между 6,75 и 7,10. Однако условие низкого рН в этом исследовании (6,70) оказало существенное влияние на титр продуцирования белка А, снижающийся на 35% по сравнению с контрольным условием (рН=6,90) (фиг. 19А) и значительно меньшей объемной активностью по сравнению со всеми другими исследованными условиями рН (фиг. 19B). Культуральный рН, по-видимому, не влиял на специфическую производительность белка А (фиг. 19С). Однако условие низкого рН вызывало наибольшую специфическую активность (фиг. 19D), и наблюдалась общая тенденция зависимости между снижением рН культуры и увеличением специфической активности.
[0203] Условия производства с низкой температурой (31,0°C и 32,0°C) вызвали значительное снижение (26% и 21% соответственно) в титре белка A по сравнению с контрольными условиями (фиг. 20A). Кроме того, повышенная температура производства была связана с более высокой объемной активностью (фиг. 20B) и небольшим увеличением специфической производительности белка А (фиг. 20С). Не наблюдалось никакого воздействия на специфическую активность от изменения температуры производства (фиг. 20D).
[0204] Все характеристики качества этого исследования и предыдущей работы, изучающие влияние рН культуры и температуры производства, приведены в таблице 17.
Таблица 17. Количественные показатели качества для различных рН и различных температур производства
[0205] Как и в предыдущих исследованиях, на TSAC влияли как pH культуры, так и температура производства (фиг. 21). Диапазон маломасштабного эксперимента для TSAC составлял 1,5-2,9, и более высокий TSAC был более желательным в этом диапазоне. Было показано, что увеличение либо pH культуры, либо температуры производства увеличивает TSAC продукта. С одной стороны, повышение значения рН культуры с 6,70 до 7,10 привело к увеличению TSAC примерно от 1,5 до 2,6-2,8, как было в диапазоне маломасштабного эксперимента. С другой стороны, увеличение температуры производства от 30,0°C до 35,0°C привело к увеличению TSAC примерно с 1,4 до 2,6-2,8. Поскольку значение TSAC (1,4) при температуре производства 30,0°C было ниже чем нижний предел (1,5) диапазона маломасштабного эксперимента, в производственном биореакторе, вероятно, следует избегать температуры производства 30,0°C. Эти данные показывают, что TSAC может быть увеличен за счет увеличения рН культуры или температуры производства или того и другого.
[0206] Уровень примеси, который в настоящее время измеряется процентом кислого пика с использованием AEX, представлен на фиг. 22. Повышение температуры производства уменьшило процент кислотного пика в продукте. рН культуры, по-видимому, не коррелирует с примесью, измеряемой с помощью кислотного пика AEX. Хотя температура производства продемонстрировала влияние на кислотный пик AEX, все значения находились в допустимых пределах для очистки (≤10,2%). Основный пик AEX был последовательно ниже 1% LoQ для анализа для всех рассмотренных условий.
[0207] Предыдущее исследование показало сильную корреляцию между кислотным пиком AEX и процентом агрегата, измеренным с помощью SEC. Эта тенденция сохранилась для данного исследования, как показано на фиг. 23. Более высокая температура производства привела к более низкому проценту агрегата. Все проценты агрегата были в пределах диапазона маломасштабного эксперимента (≤9,6%). Культуральный рН не оказывает сильного влияния на процент агрегата. Фрагменты асфотазы альфа, измеренные с помощью SEC, были ниже 1% LoQ для анализа для всех условий.
[0208] Процент основного пика, измеренный с помощью LoC, как было ранее идентифицировано, значительно подвержен влиянию температуры производства. Была обнаружена положительная корреляция между повышением температуры производства и процентом главного пика в невосстанавливающих условиях (фиг. 24B). Не обнаружено явного влияния рН культуры на процентное значение главного пика LоC (в невосстанавливающих условиях) (фиг. 24A). Анализ LоC в восстанавливающих условиях определил 100% главный пик для всех условий.
[0209] Никаких новых нейтральных видов не наблюдалось ни для какого из условий, по сравнению с материалами BDS, описанными в иллюстративном процессе примера 2. Однако наблюдался сдвиг профиля между условием низкого рН и более высоким значением рН, при котором наблюдались гликоформы более высокого порядка в материале, полученном при более высоком рН культуры (фиг. 25). Аналогичная тенденция наблюдалась для температуры производства с гликоформами более высокого порядка, присутствующими в материале, полученном при более высокой температуре производства (34,0°C и 35,0°C) (фиг. 26).
[0210] Определение пика с хроматограмм iCE280 проводилось путем определения самого основного пика как пика 6 и определения следующих четырех пиков как пика 5 по 2 в обратном порядке. Пик 1 затем состоял из всех процентов пика, указанных ниже pI следующего пика после пика 2. Полученные значения pI для идентифицированных пиков приведены в таблице 18. Как рН среды, так и температура производства оказывают влияние на результаты капиллярной изоэлектрической фокусировки (фиг. 27). Увеличение либо рН культуры, либо температуры производства приводило к увеличению процента низких пиков pI, идентифицированных как пик 1. Кроме того, связанный процент высоких пиков pI, пик 5 и 6, уменьшался с увеличением рН культуры или температуры производства. Эти факторы указывают на общий сдвиг в сторону более кислых видов, связанных с повышением рН или температуры производства. Этот сдвиг может быть связан с увеличением сиалилирования, связанным как с повышением рН, так и с температурой производства, что приведет к увеличению процента кислых видов.
Таблица 18. Диапазоны pI для идентифицированных пиков iCE280
Выводы
[0211] Это исследование подтверждает предыдущие результаты, свидетельствующие о том, что рН культуры (6,70-7,10) и температура производства (31,0-35,0°C) вряд ли будут CPP в том, что измеренные CQA находились в пределах малой шкалы, как указано ранее. Как рН культуры, так и температура производства влияют на многие параметры производительности клеточной культуры и характеристики качества. Примечательно, что увеличение либо рН культуры, либо температуры производства приводит к увеличению максимального VCD, но также приводит к быстрому снижению жизнеспособности на поздних стадиях культуры.
[0212] Как рН культуры, так и температура производства могут влиять на производительность с точки зрения титра белка А и объемной активности. Культуральный рН 6,70 приводил к снижению титра белка А и объемной активности. Повышение значения рН в культуре привело к снижению специфической активности. Поэтому, несмотря на то, что рН культуры оказывает положительное влияние на титр белка А, практически нет различий в объемной активности при рН от 6,80 до 7,10. Кроме того, увеличение температуры производства привело к увеличению титра белка А и объемной активности, но не повлияло на конкретную активность.
[0213] Повышение рН культуры или температуры производства также привело к более высоким значениям TSAC. Увеличение последнего параметра также уменьшало образование агрегатов и кислотных пиков. Наконец, pH культуры и температура производства, как представляется, сдвигают профиль изоэлектрического фокусирования продукта. Эти результаты показывают, что рН в диапазоне от 6,70 до 7,10 и температура производства в диапазоне от 31,0°С до 35,0°С являются приемлемыми, и рекомендуется жесткое регулирование обоих параметров процесса для поддержания устойчивости процесса и согласованности в масштабировании (например, для увеличения масштаба процесса до 10 000 л).
Пример 6. Улучшение титра активной асфотазы альфа с помощью дополнительной подачи подпитки
[0214] Предыдущие иллюстративные способы получения асфотазы альфа (sTNALP-Fc-D10), как описано в предыдущих примерах, были низкопроизводительными процессами с титрам активной асфотазы альфы составляющими приблизительно 0,1 г/л при сборе. Альтернативный процесс изготовления иллюстрируется в данном документе для улучшения процесса. Были оценены как предшествующие, так и последующие процессы.
[0215] Потенциальное влияние параметров предшествующего процесса культуры клеток на качество асфотазы альфа и ее продуктивность было оценено в предыдущем исследовании. Были оценены параметры клеточной культуры, включая добавление металлов (Zn2+/Mg2+/Ca2+), время температурного сдвига, количество CHO подпитки и время сбора, чтобы улучшить титр и/или объемную активность без влияния на другие критические характеристики качества (CQA). Данные предыдущего исследования показали, что добавление металлов привело к значительно более низкому TSAC, чем контрольные условия, и поэтому было исключено в дальнейшей оценке. Отсрочка времени температурного сдвига с 54 часов до 78 часов приводила к снижению специфической активности и более высокому титру белка А. В результате общая объемная активность при более поздних условиях температурного сдвига оставалась неизменной. Увеличение количества болюса CHO подпитки от 1X (5 мл/л в день 2) до 4X (5 мл/л в день 2, 4, 6 и 8) привело к значительному увеличению объемной активности без неблагоприятного воздействия на другую протестированную характеристику качества (например, TSAC, распределение заряда и распределение нейтральных гликанов). Отсрочка дня сбора с 10-го дня до 12-го дня была связана с более высокой продуктивностью (например, объемной активностью, титром белка А и специфической активностью) и незначительно меньшим TSAC. Исходя из этих данных, в этом исследовании было выбрано увеличение количества CHO подпитки для дальнейшей оценки в масштабе 10 л.
Материалы и способы
[0216] Предшествующий процесс для производства асфотазы альфа (sTNALP-Fc-D10) проводился с использованием следующей маломасштабной системы посевных ферментеров и процесса клеточной культуры: оттаивание инокулятов клеток с целевой плотностью посева при 5×105 клеток/мл; увеличение инокулята (стадия колбы/цилиндра) с целевой плотностью посева при 3,5×105 клеток/мл; увеличение инокулята (стадия клеточного мешка) с целевой плотностью посева при 3,5×105 клеток/мл; увеличение инокулята (100 л - низкий уровень) с целевой плотностью посева 3,5×105 клеток/мл; увеличение инокулята (100 л - высокий уровень) с целевой плотностью посева 3,5×105 клеток/мл; посев 1000 л биореактора (N3 - низкий уровень) с целевой плотностью посева при 3,5×105 клеток/мл; посев 1000 л биореактора (N2 - высокий уровень) с целевой плотностью посева при 3,5×105 клеток/мл; посев 4000 л биореактора (N1) с целевой плотностью посева 3,5×105 клеток/мл; производство в 20000 л биореакторе с целевой плотностью посева при 5,5×105 клеток/мл. Для этого исследования маломасштабная система посевных ферментеров, используемый в качестве инокулята для биореакторов как для контрольных, так и для улучшенных процессов биореактора, включал: оттаивание инокулята; пассаж 1 (1×100 мл колбу для встряхивания, целевой посев 5×105 клеток/мл); пассаж 2 (2×100 мл колбу для встряхивания, целевой посев 3,5×105 клеток/мл); пассаж 3 (2×1 л вращающее устройство, целевой посев 3,5×105 клеток/мл); пассаж 4 (2×15 л вращающее устройство, целевой посев 3,5×105 клеток/мл); 8×10 л биореактор, целевой посев при 5,5×105 клеток/мл. 10л биореакторами, используемыми в этом исследовании, были биореакторная система Sartorius BIOSTAT® B-DCU II (Sartorius Stedim Biotech) и программное обеспечение для сбора данных MFCS/win 3.0 Module Shell 3.0 (уровень 32) (Sartorius Stedim Biotech). Список сырья, используемого в производстве в биореакторах, включает: среду для роста CM69, 100 мкМ MTX (метотрексат), 200 мМ L-глутамина, среда для роста CM70, CHO подпитка Sigma, 10% карбонат натрия и FoamAway AOF (Gibco).
[0217] Клеточный флакон из клеточного банка разработки клона асфотазы альфа (sTNALP-Fc-D10) оттаивали и пассировали через систему посевных ферментеров через стадию N-1 (что эквивалентно 4000-литровому посевному биореактору в 20000 л процессе, использованном ранее). Культивирование N-1 проводили в двух 15-литровых вращающихся механизмах с рабочим объемом 15 л. Для инокуляции использовали обе культуры N-1. Перемешивание устанавливали со скоростью 200 об/мин с общим газовым потоком 20 л/ч. Газообразование кислорода и углекислого газа регулировали в соответствии с измеренным растворенным кислородом и рН. Восемь 10-литровых биореакторов инокулировали и собирали на 10-й день. Два биореактора получали с использованием контрольной стратегии подпитки (одна подпитка), в то время как шесть других получали подпитки с использованием улучшенной стратегии подпитки (множественные подпитки на 2, 4, 6 и 8 сутки). В таблице 19 суммированы параметры процесса клеточной культуры, используемые для этих биореакторов.
Таблица 19. Параметры процесса клеточной культуры
*: каждая болюсная подпитка содержит 1,2 мМ хлорида цинка, по сравнению с базальной средой, содержащей только около 9 мкМ хлорида цинка. Таким образом, каждая болюсная подпитка увеличивает около 6 мкМ концентрации цинка (0,5 об./об.).
[0218] Производственный процесс в масштабе 20000 л использует стратегию подпитки на основе VCD (один болюс на VCD около 25,0-32,0×105 клеток/мл, обычно около 2 дней культивирования). В этом исследовании время температурного смещения было запрограммировано на 58 часов после инокуляции, чтобы соответствовать критериям VCD (25,0-32,0×105 клеток/мл) на основе усредненного времени удвоения 24,5 часа, рассчитанного по данным производства в 20K биореакторе. Это изменение было реализовано для упрощения процесса и обеспечения более жесткого контроля за сроками подпитки.
Результаты
[0219] Показатели клеточной культуры всех биореакторов приведены на фигурах с фиг. 28 по фиг. 31.
[0220] Динамика роста и жизнеспособность улучшенного в настоящее время процесса были сопоставлены с контрольным и предыдущим 20000 л процессами. VCD в течение культуры был аналогичен контролю и 20000 л-процессам (фиг. 28А, планки ошибок представляют собой±1X стандартное отклонение). Жизнеспособность клеток в контрольных процессах и улучшенном процессе №4 снижалась с одинаковыми скоростями, что было быстрее, чем скорость при предыдущих процессах 20000 л-масштаба (фиг. 28B). Конечная жизнеспособность составляла 73% и 70% соответственно для контрольного и улучшенного процессов, тогда как процесс 20000 л обычно оставался равным около 80%. Это, по-видимому, проблема масштабирования 10 л системы, возможно, из-за скорости перемешивания. Это было протестировано во втором блоке генерации материала, и данные этого прогона показали, что падение жизнеспособности, вероятно, связано с комбинацией скорости перемешивания и того, что параметры счетчика клеток ViCell устанавливаются на стандартный тип клеток СНО, который имеет более широкий приемлемый диапазон диаметра клетки (5-50 мкМ), чем для типа клетки (11-30 мкМ) в предыдущих процессах.
[0221] Показатели использования глюкозы и потребления лактата показаны на фиг. 29. Улучшенный процесс показал сходное использование глюкозы и лактатные профили по сравнению с контролем.
[0222] Титр белка А асфотазы альфа и специфическая активность по всей культуре показан на фиг. 30. Титр белка А как для контрольного, так и для улучшенного процесса был сопоставим. Однако они были ниже, чем титры, обычно наблюдаемые при 20000 л процессе. Это может быть частично объяснено различиями в масштабе, наблюдаемыми с жизнеспособностью, поскольку соотношение мощности к объему, используемое в этом исследовании, оказалось на 30% выше, чем то, которое используется в производстве (подтверждено последующим получением отношения P/V, используемого при производственном масштабе). Специфическая активность (единицы/мг белка) для улучшенного процесса была в среднем на 17% выше (532 Ед/мг против 456 Ед/мг), чем контроль. Эти данные, взятые с данными титра белка А, указывают на то, что при одновременном получении такого же количества белка в каждом процессе, полученный белок был более активным, когда культуру подпитывали больше.
[0223] Активный титр и общая объемная активность (показан на фиг. 31) показывают, что улучшенный процесс генерирует на примерно 22-24% больше активности, чем контрольный процесс в масштабе 10 л. Усовершенствованный процесс генерировал примерно тот же активный титр, что и при масштабе 20000 л, тогда как контрольный процесс был на около 19% ниже.
Вывод
[0224] В этом исследовании подтверждается эффективность клеточной культуры улучшенного производственного процесса (масштаб 10 л), который использует увеличенное количество подпитки, что привело к увеличению активного титра на приблизительно 22-24% при сохранении титра белка А продукта. Сравнение специфической активности асфотазы альфа между контрольными и улучшенными процессами показано на фиг. 32. Очевидно, что все шесть повторностей улучшенных условий процесса (каждый с тремя дополнительными добавками подпитки) показали более высокую специфическую активность, чем контрольные.
[0225] Каждая болюсная подпитка содержала 1,2 мМ хлорида цинка по сравнению с основной средой, содержащей только около 9 мкМ хлорида цинка. Таким образом, каждая болюсная подпитка приводила к увеличению на около 6 мкМ концентрации цинка (0,5 об./об.) во всей культуре. Не ограничиваясь какой-либо конкретной теорией, постулируется, что увеличение специфической активности путем добавления дополнительной подпитки происходит, по меньшей мере частично, из-за добавления цинка.
Пример 7. Дальнейшее улучшение активности асфотазы альфа путем добавления цинка
[0226] На основе производственных процессов, описанных в предыдущих примерах, были проведены новые исследования для дальнейшей оптимизации параметров предшествующего процесса. Иллюстративные процессы Z и Z 'суммируются и сравниваются в таблице 20.
Таблица 20. Различия между иллюстративными производственными процессами (предшествующие)
[0227] Дальнейшая оптимизация была выполнена на основе предыдущего процесса Y. Например, новый процесс Z' включал добавление около 30-90 мкМ сульфата цинка в культуральную среду и увеличение общего времени культивирования от около 240 часов (то есть от 10 дней) до около 288 часов (т. е. 12 дней). Исследование маломасштабной встряхиваемой колбы проводили для испытания трех вариантов процесса Z' (добавление соответственно 30, 60 или 90 мкМ цинка). 30 мкМ добавки вносили путем фронтальной загрузки в культуральную среду. Добавки 60 и 90 мкМ включали добавление цинка в двух равных болюсах на 2-й и 6-й день (при условии 60 мкМ) или трех равных болюсах на 2-й, 6-й и 10-й день (для 90 мкМ условия). Как показано на фиг. 33, добавка цинка (красная линия, показывающая среднее значение трех экспериментов с различными концентрациями цинка) эффективно улучшала специфическую активность асфотазы альфа по сравнению с контрольным процессом Y (синяя линия). Это увеличение достигло своего пика около 288-часового (т. е. 12 дней) временного интервала и впоследствии уменьшилось.
Пример 8. Характеристика производимой асфотазы альфа
[0228] В данном документе проиллюстрирована примерная характеристика изготовленной асфотазы альфа с несколькими ортогональными аналитическими методами. Эти методы использовались для оценки идентичности, чистоты, размера, структуры, гликозилирования и профилей зарядов продуцируемой асфотазы альфа. Некоторые методы также использовались для обеспечения согласованности между партиями. Кроме того, были также охарактеризованы связанные с продуктом вещества и связанные с продуктом примеси.
Структурный анализ асфотазы альфа
Анализ времяпролетной матрично-активированной лазерной десорбции/ионизации (MALDI-TOF) молекулярных масс после удаления олигосахаридов
[0229] MALDI-TOF масс-спектрометрический анализ асфотазы альфа после удаления олигосахаридов использовали для установления идентичности асфотазы альфа. Кроме того, этим способом также будет обнаружено присутствие любых значительных уровней посттрансляционных модификаций с молекулярными массами, существенно отличающимися от теоретической молекулярной массы асфотазы альфа. Удаление олигосахаридов было достигнуто путем переваривания асфотазы альфа с использованием PNGазы F с или без восстановления дисульфидных связей. Образцы обессоливали и смешивали со смесью 90:10 2,5-дигидроксибензойной кислоты и 2-гидрокси-5-метоксибензойной кислоты в ацетонитриле и трифторуксусной кислоте. Образцы высушивали на воздухе, а затем анализировали с использованием масс-спектрометра MALDI-TOF. Масс-спектры были получены в положительном режиме. Инструмент был откалиброван внешне с использованием бычьего сывороточного альбумина (BSA). Масс-спектр асфотазы альфа после дегликозилирования показан на фиг. 34А. Пики с m/z 161140, 80571 и 53748 соответствуют молекуле, которая является одно-, двух- или трехкратно заряженной, соответственно. Измеренная молекулярная масса 161140 Да согласуется с рассчитанной молекулярной массой (161135,2 Да) в пределах лимита точности прибора 1%. Масс-спектр асфотазы альфа после восстановления и удаления олигосахарида показан на фиг. 34B. Пики с m/z 80545, 40267 и 26808 соответствуют молекуле, которая является одно-, двух- или трехкратно заряженной, соответственно. Измеренная молекулярная масса 80545 Да согласуется с рассчитанной молекулярной массой (80577,7 Да) в пределах лимита точности прибора 1%. Результаты подтвердили идентичность асфотазы альфа и продемонстрировали отсутствие значительных уровней посттрансляционных модификаций, отличных от гликозилирования.
Аналитическое ультрацентрифугирование (AUC)
[0230] AUC использовали для определения процента агрегатов, димера и фрагментов, если они присутствуют, и, следовательно, чистоты асфотазы альфа. Кроме того, оно определяет молекулярную массу димера асфотазы альфа, которая является характерной для молекулы из-за уникальной аминокислотной последовательности и посттрансляционных модификаций. Оно считается ортогональным методом для гель-проникающей хроматографии (SEC). Образцы асфотазы альфа разбавляли приблизительно до 1 мг/мл, используя 25 мМ фосфат, 150 мМ NaCl, pH 7,4. Для проведения анализа скорости седиментации использовалась аналитическая ультрацентрифуга. Использовались клетки двойного сектора, оборудованные кварцевыми окнами. Ротор уравновешивали в вакууме при 20°С и примерно через 1 час ротор ускорялся до 36 000 об/мин. Сканы абсорбции при 280 нм были получены с 4,5-минутными интервалами в течение приблизительно 6 часов. Данные сначала анализировали с использованием DCDT для определения наблюдаемой молекулярной массы, а затем с использованием метода (программное обеспечение SEDFIT) для определения процента димеров, высокомолекулярных и низкомолекулярных видов соединений. Молекулярная масса асфотазы альфа составляла 211 кДа, а чистота составляла 96,8% для иллюстративной партии.
MALDI-TOF интактного молекулярного веса
[0231] MALDI-TOF измерение молекулярной массы асфотазы альфа определяет размер молекулы, включая все посттрансляционные модификации, главным образом, гликозилирование. Образцы асфотазы альфа обессоливали и смешивали с матрицей, полученной из смеси 90:10 2,5-дигидроксибензойной кислоты и 2-гидрокси-5-метоксибензойной кислоты. Образцы высушивали на воздухе, а затем анализировали с использованием масс-спектрометра MALDI-TOF. Масс-спектры были получены в положительном режиме. Инструмент был откалиброван снаружи с использованием бычьего сывороточного альбумина (BSA). Масс-спектр асфотазы альфа показан на фиг. 35. Пики с m/z 180157, 90223 и 60540 соответствуют молекуле, которая является одно-, двух- или трехкратно заряженной, соответственно. Другие низкообогащенные пики были сформированы из-за фрагментации, вызванной жесткими настройками инструмента. Измеренная молекулярная масса 180157 Да значительно выше расчетной молекулярной массы 161135,2 Да на основе известной аминокислотной последовательности. Этот результат указывает на обширные посттрансляционные модификации асфотазы альфа, главным образом гликозилирования, поскольку MALDI-TOF и ESI-TOF анализы дегликозилированной асфотазы альфа не обнаруживают значительных уровней модификаций.
Эксклюзионная хроматография с кратными углами рассеяния света (SEC-MALS)
[0232] SEC-MALS используется для разделения белков на основе размеров и затем определения молекулярной массы каждого пика. Асфотазу альфа анализировали с помощью SEC-MALS (данные не показаны). Рассчитанная молекулярная масса пика в окне времени удерживания 7,0-8,2 минуты составляет 194,1 Да, что хорошо согласуется с рассчитанной молекулярной массой асфотазы альфа на основе известной аминокислотной последовательности и обширного гликозилирования, а также в соответствии с молекулярной массой, наблюдаемой с помощью MALDI-TOF.
Анализ металлов
[0233] Для определения содержания цинка и магния использовалась индуктивно-связанная плазменная масс-спектрометрия (ICP-MS) и для определения содержания кальция использовалась индуктивно связанная плазменно-атомная эмиссионная спектрометрия (ICP-AES). Образцы асфотазы альфа расщепляли с использованием микроволнового расщепления с нагреванием с добавлением концентрированной азотной кислоты сначала, а затем с добавлением перекиси водорода. Затем образцы анализировали с использованием прибора ICP-MS для цинка и магния и прибора ICP-AES для кальция. Определенные молярные соотношения (моль ионов/моль мономера асфотазы альфа) составляли 2,27 для цинка, 0,97 для кальция и 0,12 для магния. Соотношения цинка и кальция находились в хорошем соответствии с представленными соотношениями в литературе (Stec et al., 2000 J Mol Biol 299: 1303-1311 и Mornet et al., 2001 J Biol Chem 276: 31171-31178). Mornet et al. (2001) утверждает, что кальций может занимать сайт связывания магния на TNALP и, таким образом, предполагает, что отношение магния в молекуле асфотазы альфы может быть менее чем один (1). Однако протестированное соотношение магния как 0,12 все еще является неожиданно низким.
Определение сайта фосфорилирования
[0234] UPLC-MSe использовался для определения сайта и процента фосфорилирования. Асфотазу альфа при конечной концентрации 0,5 мг/мл денатурировали и восстанавливали в присутствии 6 М гуанидина гидрохлорида в 0,2 М Трис, рН 8,6, используя 40 мМ DTT при 37°С в течение 1 часа. Восстановленный образец был алкилирован добавлением йодууксусной кислоты до конечной концентрации 50 мМ и инкубацией при комнатной температуре в течение 30 минут. Образец затем переносили в буфер 50 мМ бикарбоната аммония с использованием диализной пробирки с отсечкой молекулярной массы 10 кДа. Образец расщепляли с использованием трипсина при конечном соотношении (масса:масса) 1:50 трипсина; белка при 37°С в течение 24 часов. Для анализа переваренных образцов использовалась система UPLC, оборудованная колонкой C18 с обращенной фазой и масс-спектрометром Q-TOF. Триптический пептид, содержащий Ser93, идентифицировали как фосфорилированный, так и нефосфорилированный. Как показано на фиг. 36, спектр MС/MС продемонстрировал, что S93 сдвинут на +80 Да массы фосфата, добавленной фосфорилированием, как это наблюдается с помощью ионов y12 и y13. Относительный процент фосфорилирования определяли с использованием площадей пиков экстрагированных ионных хроматограмм (EIC) как 33,9%.
MALDI анализ свободных гликанов
[0235] MALDI-TOF анализ свободных гликанов использовался для определения видов олигосахаридов и их относительного процента. Олигосахариды высвобождались из эталонного стандарта альфатазы альфа путем расщепления PNGазой F. Выделенные гликаны очищали с использованием колонок с обращенной фазой с верхним наконечником. Выделенные гликаны смешивали 1:1 с матрицей sDHB и затем анализировали MALDI-TOF. Анализы проводились в отрицательном режиме для гликанов с сиаловой кислотой, а затем в положительном режиме для нейтральных гликанов. Масс-спектр, полученный с использованием отрицательного режима, показан на фиг. 37A. Масс-спектр, полученный с использованием положительного режима, показан на фиг. 37B. Виды олигосахаридов определяются путем сопоставления наблюдаемых молекулярных масс с теоретическими молекулярными массами общеизвестных олигосахаридов. Относительный процент определяли делением индивидуальной интенсивности пика на общую интенсивность пиков всех гликанов. Основные олигосахариды приведены в таблице 21 и таблице 22. Основываясь на молекулярных массах, структура сиаловой кислоты определялась как N-ацетилнейраминовая кислота (Neu5Ac или NANA).
Таблица 21. Теоретические и наблюдаемые молекулярные массы сиалированных гликанов (M-H)-
Таблица 22. Теоретические и наблюдаемые молекулярные массы нейтральных гликанов (М+Na)+
Профилирование олигосахаридов меченных 2-аминобензамидом (2-АВ)
[0236] Выделенные из асфотазы альфа олигосахариды метили 2-аминобензамидом (2-АВ), а затем анализировали с помощью ВЭЖХ с нормальной фазой с детектированием флуоресценции для определения типов и относительного процента различных олигосахаридов. Образцы асфотазы альфа восстанавливали с использованием дитиотреитола (DTT) при 37°С в течение 1 часа и затем расщепляли с использованием PNGазы F при 37°C в течение ночи для высвобождения N-связанных олигосахаридов. Белок из расщепленного образца осаждался и отделялся от высвобожденных гликанов холодным этанолом и центрифугированием. Супернатанты, содержащие высвобожденные гликаны, сушили с использованием вакуумной центрифуги, восстанавливали с использованием 1% муравьиной кислоты и инкубировали при комнатной температуре в течение 40 минут, а затем снова сушили. Затем образцы метили с использованием реагентов 2-AB для мечения, следуя инструкциям производителя (Ludger, Оксфордшир, Великобритания). Меченые гликаны очищали с помощью картриджей гликана S (Prozyme, Хейвард, Калифорния), а затем анализировали с использованием ВЭЖХ с детектированием флуоресценции с длиной волны возбуждения 330 нм и длиной волны излучения 420 нм. Флуоресцентная хроматограмма из анализа асфотазы альфа показана на фиг. 38. Точность пиков была установлена путем сбора фракций, масс-спектрометрического анализа молекулярных масс и сопоставления молекулярных масс с молекулярными массами общеизвестных олигосахаридных структур. Олигосахариды, идентифицированные с помощью мечения 2-AB, суммированы в таблице 23. Хроматограмма, полученная в результате анализа эталонного стандарта альфатазы альфа, показана на фиг. 39.
Таблица 23. Гликоформы и их относительный процент с помощью 2-AB мечения
A1
FA1
A2
FA2
M5/A3
A2G1
FA3
FA2G1
FA2G1
FA4
A2G2
FA3G1
FA2G2
FA4G1
FA3G3
A3G3/A4G2
FA3G3
FA3G3
FA4G3
FA4G3
FA4G4
FA4G4L1
FA4G4L2
НЕИЗВЕСТНЫЙ
1,90
11,58
0,26
25,55
5,18
1,25
2,67
4,58
4,59
2,68
2,46
2,25
7,03
3,42
1,71
3,00
2,49
3,01
2,13
0,89
5,35
3,54
1,87
0,61
Профилирование гликопептидов
[0237] Для определения сайт-специфического распределения олигосахарида использовали профиль гликопептида. Образцы асфотазы альфа при конечной концентрации 0,5 мг/мл денатурировали и восстанавливали в присутствии 6 М гуанидина гидрохлорида в 0,2 М Трис, рН 8,6, используя 40 мМ DTT при 37°С в течение 1 часа. Восстановленные образцы алкилировали добавлением йодууксусной кислоты до конечной концентрации 50 мМ и инкубацией при комнатной температуре в течение 30 минут. Образцы затем переносили в буфер 50 мМ бикарбоната аммония с использованием диализной пробирки с отсечкой молекулярной массы 10 кДа. Образцы расщепляли с использованием трипсина при конечном соотношении (масса:масса) 1:50 трипсина; белка при 37 °С в течение 24 часов. Для анализа переваренных образцов использовалась система UPLC, оборудованная колонкой C18 с обращенной фазой и масс-спектрометром Q-TOF. Данные были получены в положительном режиме. Масс-спектры в течение окон времени элюирования усредняли для каждого гликопептида. Шесть гликопептидов были получены при расщеплении трипсином асфотазы альфа, что соответствует шести сайтам гликозилирования на мономер асфотазы альфа. Поскольку пептидный фрагмент является основным фактором, определяющим время удержания, гликопептиды разделяют на основе их уникальной аминокислотной последовательности; поэтому можно получить сайт-специфическое распределение олигосахаридов. Масс-спектры гликопептидов показаны на фиг. 40. Виды олигосахаридов, связанные с каждым сайтом гликозилирования, суммированы в таблице 24.
Таблица 24. Виды олигосахарида, связанные с каждым гликопептидом
FA2
FA2G1
FA3
FA2G2
FA3G1
FA4
FA3G2
FA4G1
FA3G3
3281,42
3443,47
3484,50
3605,53
3646,55
3687,58
3808,61
3849,63
3970,66
3281,40
3443,45
3484,48
3605,51
3646,54
3687,57
3808,60
3849,62
3970,65
FA2
FA2G1
FA3
FA2G2
FA3G1
FA4
FA4G1
FA4G2
FA4G3
2901,18
3063,23
3104,26
3225,28
3266,31
3307,34
3469,39
3631,43
3793,49
2901,16
3063,21
3104,25
3225,27
3266,29
3307,32
3469,38
3631,43
3793,47
A2
A2G1
A3
A2G2
A3G1
A4
2394,99
2557,05
2598,07
2719,10
2760,13
2801,15
2395,01
2557,06
2598,07
2719,11
2760,14
2801,16
A3
FA3
A4
FA3G1
FA4
FA4G1
3699,74
3845,80
3902,82
4007,85
4048,88
4210,93
3699,73
3845,79
3902,80
4007,85
4048,87
4210,93
FA2G1
FA3
FA2G2
FA3G1
FA4
FA3G2
FA4G1
FA4G2
4710,09
4751,11
4872,14
4913,17
4954,19
5075,22
5116,25
5278,29
4710,05
4751,08
4872,10
4913,13
4954,21
5075,18
5116,21
5278,26
A2
FA2
FA2G1
2487,99
2634,05
2796,10
2488,00
2634,06
2796,12
[0238] Основываясь на наблюдаемых молекулярных массах и других результатах анализа (например, данные масс-спектра и данные профилирования олигосахаридов с 2-аминобензамидами (2-AB)), структуры основных гликанов (≥4% относительной численности) асфотазы альфа были определены на фиг. 41 и фиг. 42.
Капиллярное изоэлектрическое фокусирование (cIEF)
[0239] cIEF отделяет белки на основе их изоэлектрических точек и используется для контроля заряда вариантов альфатазы альфа. Образцы асфотазы альфа при конечной концентрации 0,5 мг/мл в буфере, содержащем мочевину, метилцеллюлозу, сахарозу, фармалиты и маркеры pI, анализировали с использованием системы капиллярного электрофореза. Образцы сфокусировали в течение 0,1 минуты при 1500 В, а затем в течение 14 минут при 3000 В. Разделенные варианты белка были обнаружены с использованием УФ-поглощения при 280 нм. Электроферограмма из репрезентативной партии асфотазы альфа показана на фиг. 43. Шесть пиков наблюдались с помощью cIEF. Как правило, пик 1 имеет широкий диапазон pI. Дополнительные пики наблюдались для пиков 4 и 5. Этот результат продемонстрировал, что асфотаза альфа является сильно гетерогенной по отношению к заряду, что ожидалось, из-за наличия множества сайтов гликозилирования с различным количеством и уровнями сиаловых кислот на каждом сайте. pI каждого пика и их относительный процент асфотазы альфа репрезентативной партии приведены в таблице 25.
Таблица 25. pI и относительная процентная доля пиков, наблюдаемая с помощью cIEF
2
3
4
5
6
6,65
6,68
6,73
6,80
6,85
8,66
11,78
16,71
11,24
15,27
Связанное с продуктом вещество
[0240] Ожидается структурная гетерогенность асфотазы альфа, которая экспрессируется в клеточной линии СНО, вследствие посттрансляционной модификации. Как описано выше, гетерогенность асфотазы альфа отражается в вариантах заряда, где шесть пиков наблюдаются при анализе с помощью cIEF. Последовательная картина профилей заряда асфотазы альфа была продемонстрирована в клинических и PV партиях. Как определено с помощью AUC, SDS-PAGE, SEC и AEX с использованием репрезентативного материала, асфотаза альфа имеет чистоту 96%, что указывает на то, что множественные пики, наблюдаемые с помощью cIEF, полностью представляют собой продуцированную асфотазу альфа и что все пики являются продуктами, связанного с этим веществом.
Связанные с продуктом примеси
[0241] Связанные с продуктом примеси представляют собой молекулярные варианты, которые не обладают свойствами, сопоставимыми с желаемой асфотазой альфа, по отношению к активности, эффективности и безопасности (Guidance for Industry, Q6B specifications: test procedures and acceptance criteria for biotechnological/biological products, August 1999, ICH). Связанные с продуктом примеси наблюдались с помощью трех анализов, а характеристики каждой примеси приведены в таблице 26.
Таблица 26. Краткое описание примесей, связанных с производством асфотазы альфа
Характеристика полос SDS-PAGE
[0242] Асфотазу альфа анализировали SDS-PAGE в невосстанавливающих и восстанавливающих условиях. Результаты анализа SDS-PAGE показывают, что полоса 1 соответствует интактной асфотазе альфа, полоса 2 соответствует интактной асфотазе альфа только с одной линией фермента, а полоса 3 соответствует восстановленной форме (мономер) асфотазы альфа (данные не показаны).
[0243] Как и ожидалось, пептиды как из ферментной части, так и из участка IgG1-Fc наблюдались из полосы SDS-PAGE в восстановительных условиях. Этот результат, помимо видимой молекулярной массы полосы SDS-PAGE в восстанавливающих условиях, указывает на то, что основная полоса соответствует асфотазе альфа в ее восстановленной форме. Характеристика полосы суммирована в таблице 27.
Таблица 27. Обзор характеристик полос SDS-PAGE
Предполагаемая усеченная форма, обнаруженная с помощью SDS-PAGE в невосстанавливающих условиях, не была обнаружена с помощью SDS-PAGE в восстанавливающих условиях из-за ее низкой концентрации.
Характеристика пиков SEC
[0244] SEC-HPLC использовали в качестве анализа чистоты с помощью разделение видов с высокой молекулярной массой (агрегатов), димера асфотазы альфа и видов с низкой молекулярной массой. На типичной хроматограмме SEC наиболее распространенный пик соответствует гомодимеру асфотазы альфа. Пик, который элюируется до главного пика, соответствует видам с высокой молекулярной массой. Пик, который элюируется после главного пика, соответствует видам с низкой молекулярной массой. Фракции, соответствующие как видам с высокой молекулярной массой, так и главным пикам, собирали и анализировали с помощью SDS-PAGE с последующим перевариванием в геле и анализом масс-спектрометрии. Согласование наблюдаемых молекулярных масс пептидов с теоретическими молекулярными массами, полученными из известной аминокислотной последовательности, показало, что степень охвата 86,6% для главного пика и 70,7% для видов с высокой молекулярной массой. Поэтому делается вывод о том, что виды с высокой молекулярной массой представляют собой связанные с продуктом примеси.
Характеристика пиков AEX
[0245] AEX использовался для контроля вариантов заряда асфотазы альфа. Пять фракций AEX вместе с исходным материалом анализировали с помощью GP-HPLC и SDS-PAGE в невосстанвливающих и восстанавливающих условиях (данные не показаны). Данные свидетельствуют о том, что основные пики AEX содержат в основном асфотазу альфа и, возможно, небольшой процент видов с высокой молекулярной массой. Кислотный пик AEX содержит высокомолекулярные виды асфотазы альфа. Основные и кислые виды связаны с продуктом, что подтверждается фингерпринтингом пептида с помощью анализа MALDI-TOF.
[0246] Асфотаза альфа хорошо охарактеризована с использованием множества аналитических методов. Ее идентичность была подтверждена N-терминальным секвенированием, аминокислотным анализом и анализом молекулярной массы с помощью MALDI-TOF и ESI-MS. Чистоту продуцируемой асфотазы альфа контролировали с помощью AUC, SDS-PAGE, GP-HPLC и AEX. Размер асфотазы альфа оценивали с помощью анализа MALDI-TOF и SEC-MALS молекулы с олигосахаридами. Кроме того, анализы молекулярной массы асфотазы альфа также выполняли с помощью AUC и SDS-PAGE. Этими ортогональными методами были обнаружены последовательные более высокие молекулярные массы асфотазы альфа, что свидетельствует об обширном гликозилировании молекулы на нескольких сайтах, что было подтверждено анализом гликозилирования. Структуры более высокого порядка асфотазы альфа определялись как с помощью дальнего-УФ, так и ультра-УФ-CD. Гидродинамический размер молекулы также можно определить из анализа AUC и SEC-MALS, что также подтвердило, что асфотаза альфа существует как ковалентный димер. Низкое количество агрегатов было обнаружено с помощью SEC, AUC и SEC-MALS. Структура нативной дисульфидной связи и расположение свободного цистеина при 102 подтверждается с помощью LC-MS-анализа пептидных карт в невосстанавливающих условиях. Присутствие только одного основного свободного цистеина было дополнительно подтверждено оценкой общего количества свободного сульфгидрила. Анализ LC-MS показал, что остаток серина активного сайта частично фосфорилируется. Три металла, включая цинк, кальций и магний, как сообщается в литературе, были обнаружены в препарате асфотазы альфа. Гликозилирование асфотазы альфа тщательно анализировали с помощью анализа свободного гликана с использованием MALDI-TOF, анализа ВЭЖХ с нормальной фазой меченых олигосахаридов 2-AB, LC-MS-анализа гликопептидов и общего измерения сиаловой кислоты. Было обнаружено, что все шесть сайтов гликозилирования на мономер асфотазы альфы ассоциируются с гетерогенной популяцией олигосахаридов. Профиль зарядов, гетерогенность, вызванная главным образом наличием сиаловой кислоты, была проанализирована с помощью cIEF, где наблюдалось шесть пиков. Связанное с продуктом вещество отражается с помощью наблюдаемых шести пиков cIEF. Низкие уровни связанных с продуктом загрязнений были обнаружены с помощью SEC, AEX и SDS-PAGE.
Пример 9. Сопоставимость асфотазы альфа, производимой в масштабах 2000 л (2K) и 20000 л (20K)
[0247] В этом примере лекарственное вещество асфотазы альфа было изготовлено с использованием процессов 2000 л (2K) и 20000 л (20K). Чтобы продемонстрировать сопоставимость между асфотазой альфа, полученной в разных масштабах, три партии асфотазы альфа, полученные с использованием процесса 2K (№ 35, № 36 и № 38) и трех партий асфотазы альфа, полученных с использованием процесса 20K (№ 40, № 42 и № 34) были проанализированы по их физико-химическим свойствам параллельно, где это возможно, с использованием способов, описанных в таблице 28. Конкретные партии 2K и 20K были выбраны на основе клинического применения, а ранее тестированные партии доказали согласованность между партиями для всех партий 2K и 20K.
[0248] Тестируемые физико-химические свойства устанавливали как чистоту асфотазы альфа, варианты заряда, размер, идентичность, структуру, гликозилирование и активность. Результаты тестирований приведены в таблице 29. Сравнимость производства была дополнительно оценена с использованием паралелльного исследования принудительной деградации, где сравнивались пути деградации и кинетика вышеупомянутых партий 2K и 20K. Образцы инкубировали при 40°C в течение 0, 14, 24 и 48 часов и анализировали способами, перечисленными в таблице 30. Результаты тестирований для исследования принудительной температурной деградации приведены в таблице 31.
[0249] Общие результаты показывают, что асфотаза альфа, изготовленная с использованием процессов 2K и 20K, сравнима.
Таблица 28. Физико-химические тесты Асфотазы Альфа
Таблица 29. Физико-химические данные Асфотазы Альфа
* Мономер асфотазы альфа, о котором сообщается с помощью аналитического ультрацентрифугирования, эквивалентен/синоним «димеру» с помощью GP-HPLC и SEC-MALS.
Таблица 29 (Продолж.). Физико-химические данные Асфотазы Альфа
Mw (кДа) Димер
Mw (кДа) виды с высокой молекулярной массой
Гликозилированный
Дегликозилированный
Деглико и восстановленный
Деглико и восстановленный
Таблица 29 (Продолж.). Физико-химические данные Асфотазы Альфа
Таблица 29 (Продолж.). Физико-химические данные Асфотазы Альфа
Таблица 29 (Продолж.). Физико-химические данные Асфотазы Альфа
Таблица 29 (Продолж.). Физико-химические данные Асфотазы Альфа
Таблица 29 (Продолж.). Физико-химические данные Асфотазы Альфа
Таблица 29 (Продолж.). Физико-химические данные Асфотазы Альфа
Таблица 29 (Продолж.). Физико-химические данные Асфотазы Альфа
Таблица 29 (Продолж.). Физико-химические данные Асфотазы Альфа
Таблица 29 (Продолж.). Физико-химические данные Асфотазы Альфа
Таблица 29 (Продолж.). Физико-химические данные Асфотазы Альфа
†: Пептидные моды: Cys102 COMe; Молекулярная формула: C102H159N27O38S1; Моноизотопная масса: 2402,106
‡: Пептидные моды: Cys102 COMe S93 PO4; Молекулярная формула: C102H160N27O41S1P1; Моноизотопная масса: 2482,0723
Таблица 29 (Продолж.). Физико-химические данные Асфотазы Альфа
эталонная Km
эталонная Kcat
Таблица 30. Тесты для принудительной тепмературной деградации асфотазы альфа
невосстанавливающих условиях
Таблица 31. Итог результатов принудительной температурной деградации асфотазы альфа
Чистота
Аналитическое ультрацентрифугирование (AUC)
[0250] Уровни % мономера, определенные аналитическим ультрацентрифугированием, были сопоставимы между партиями 2K и 20K. Все значения находились в диапазоне от 94,5% до 97,6% со средним значением 20K 96,8% и средним значением 2K 95,6%. Разница заключается в изменчивости анализа AUC, как это было ранее продемонстрировано для анализа антитела IgG1 (см. Pekar and Sukumar 2007 Quantitation of aggregates in therapeutic proteins using sedimentation velocity analytical ultracentrifugation: Practical considerations that affect precision and accuracy. Analytical Biochemistry, 367: 225-237), которое также представляет собой гликопротеин с аналогичной молекулярной массой. Данные GP-HPLC (показанные ниже) показывают сопоставимый % мономера, что также указывает на то, что небольшая разница между партиями, обнаруженная с помощью AUC, была обусловлена вариацией внутри анализа (например, разницей между элементами прибора). Небольшой дополнительный пик при 2,5 (с) был обнаружен для асфотазы альфа, полученной из иллюстративного процесса № 35, который был артефактом c(с) анализа. Смещение пика № 35, соответствующее димеру асфотазы альфа (приблизительно 11 (с)), рассматривается в пределах лимита погрешности способа. Данные AUC асфотазы альфа были получены Университетом Коннектикута, Аналитическим Ультрацентрифугирующим Центром, Центром биосервиса Биотехнологий. Этот метод отличает мономер асфотазы альфа от агрегатов, состоящих из димерных или более крупных видов и их относительных % в зависимости от коэффициента непрерывного распределения седиментации.
SDS-PAGE/ LoC (Lab on Chip)
[0251] % главной полосы, определяемый с помощью SDS-PAGE: анализ LABCHIP® (PerkinElmer) GXII Protein для как невосстанавливающих, так и восстановливающих условий был сопоставим между партиями 2K и партиями 20K. Как суммировано в таблице 29, средний % главной полосы составлял 100,0% (невосстанавливающие условия) и 99,8% (восстанавливающие условия) для трех партий 2K и 100,0% (невосстанавливающие условия) и 99,8% (восстанавливающие условия) для партий 20K. Кроме того, для партий 2K и 20K наблюдались сопоставимые образцы полос, что было продемонстрировано электрофореграммами как анализов в невосстанавливающих и восстанавливающих условиях. Этот тест отделяет белок на основе его молекулярной массы и обеспечивает анализ чистоты интактного белка, выраженного в процентах от главной полосы. Анализ проводили согласно LabChip GXII Protein Assay Quick Guide, HT Protein Express LabChip Kit, версия 2 (пересмотренный в марте 2010 года).
SDS-PAGE
[0252] Три партии 2K и три партии 20K были проанализированы параллельно с использованием SDS-PAGE. Анализ методом денситометрии показал, что основная полоса составляла 100% для всех партий как для SDS-PAGE в не восстановливающих, так и в восстанавливающих условиях. В дополнение к главной полосе наблюдалась слабая полоса ниже лимита количественного определения с кажущейся молекулярной массой выше, чем у димера асфотазы альфа, для всех партий. Только одна полоса наблюдалась для всех партий с помощью SDS-PAGE в восстанавливающих условиях. Этот тест отделяет белок на основе его молекулярной массы и обеспечивает анализ чистоты интактного белка, выраженного в процентах от главной полосы. Образец белка отделяли на геле, окрашивали кумасси-синим для визуализации, отмывали окраску и анализировали с помощью денситометрии. Молекулярную массу асфотазы альфа, продуктов разложения и связанных с процессом примесей оценивали по сравнению с известными стандартами молекулярной массы. Количественные измерения отделенных белков выполняются путем сканирования и анализа с помощью денситометрического анализа.
GP-HPLC
[0253] Чистоту асфотазы альфа анализировали параллельно с помощью GP-HPLC. Среднее значение 2K (96,5%) и среднее значение 20K (98,6%) показаны в таблице 29. % асфотазы альфа из партий 20K немного выше, чем у партий 2K. Тем не менее, данные о партиях показали аналогичный % димера, предполагая, что немного более низкий % димера из 2K партий в основном обусловлен различиями в истории образцов, таких как возраст материала. Хроматограмма GP-HPLC также показывала сравнимые продукты из партий 2K и 20K. Одни и те же пики при одинаковых временах удерживания указывают на тот же вид. Этот способ отличает асфотазу альфа от более крупных видов и меньших фрагментов.
Анион-обменная хроматография (AEX)
[0254] % главного пика AEX для всех партий 2K и 20K партий были в узком диапазоне от 93,62% до 96,84%, как показано в таблице 29. Среднее значение 2K составляло 94,17%, а среднее значение 20K составляло 96,38% от площади главного пика. Чуть более высокий % главного пика наблюдался для партий 20K из-за уменьшения пика позднего элюирования. Несколько более высокий уровень главного пика в партиях 20K демонстрирует более высокую чистоту продукта. Хроматограмма AEX показала наложенные пики для партий 2K и 20K с одинаковым временем удерживания, что указывает на то, что продукты из всех партий имеют одинаковые виды. Этот способ отделяет белок в порядке увеличения чистого поверхностного анионного заряда. Разделенные белки детектируются ультрафиолетовой абсорбцией при 280 нм, а затем количественно определяют как процент площади пика.
Профиль заряда
Капиллярное изоэлектрическое фокусирование (cIEF)
[0255] Профили пика и относительный % каждого пика партий 2K и 20K были сопоставимы при анализе асфотазы альфа с помощью cIEF. Все образцы демонстрируют шесть заметных пиков в диапазоне изоэлектрической точки (pI) от ~ 6,57 до ~ 6,85, и эти пики называются пиками 1, 2, 3, 4, 5 и 6 слева направо. Площади пика, обнаруженного в партиях 2K и 20K для каждого заряженного варианта вида, сопоставимы, как показано в таблице 29. Сопоставимость между % пика cIEF от партий 2K и 20K была подтверждена с использованием анализа T-теста всех партий, как показано в таблице 32 (все p-значения были >0,05, что указывает на то, что любые наблюдаемые различия между партиями незначительны). Электрофореграммы cIEF из партий 2K и 20K показали одинаковые профили пиков в тех же диапазонах pI. cIEF обеспечивает полуколичественное разделение зарядово-вариантных видов на основе pI изоформ белка. Отделенные белки детектировали при поглощении 280 нм.
Таблица 32. T-тест % пика сIEF от партий 2K и 20K
Молекулярный размер
Эксклюзионная хроматография - Множественное рассеяние света (SEC-MALS)
[0256] Молекулярные массы асфотазы альфа, определяемые с помощью SEC-MALS партий 2K и 20K, были сопоставимы. Средняя молекулярная масса 2K партий составляет 189,2 кДа, а средняя молекулярная масса 20K партий составляет 189,2 кДа, как показано в таблице 29. Аналогичные профили пика наблюдались для всех партий на УФ-хроматограммах (образец из партии № 36 был запущен позднее, из-за доступности образцов, что привело к несогласованности его профилей пиков с другими образцами). Способ SEC-MALS объединяет колонку SEC для разделения видов белка по размеру с помощью прибора MALS для определения молекулярной массы пиков. Высокие вариации молекулярных масс видов с высокой молекулярной массой (см. Табл. 29), определяемые сигналом светорассеяния и показателем преломления (RI), обусловлены более высокой вариацией метода для определения видов с низким содержанием. На основе УФ-хроматограмм относительное время удерживания видов с высокой молекулярной массой асфотазы альфа одинаково для всех шести партий, что указывает на то, что молекулярные массы видов с высокой молекулярной массой также должны быть одинаковыми.
Анализ интактной молекулярной массы (MALDI-ToF-MS)
[0257] Средняя молекулярная масса 2K партий составляла 180902 кДа, а средняя молекулярная масса 20K партий составляла 180019 кДа, как показано в таблице 29. Молекулярная масса 20K партий и 2K партий находится в пределах 1% массовой точности внешне калиброванного MALDI-ToF и, следовательно, сопоставима. Несколько более высокий тренд молекулярной массы 2K, вероятно, обусловлен наличием немного большего количества больших олигосахаридов, таких как олигосахариды с сиаловой кислотой. Спектры MALDI-ToF интактной молекулярной массы (IMW) для партий асфотазы альфа 2K и 20K также обнаруживали одни и те же профили пика. Партию № 36 анализировали в отдельном случае с использованием другой калибровки прибора, но была включена для полноты сопоставимости. Дополнительные массы, присутствующие в спектрах, обусловлены индуцированной анализом фрагментацией и или несколькими заряженными ионами. Различия в интенсивности, наблюдаемые для идентифицированных видов, являются результатом матричной кристаллизации и лазерных эффектов. Этот способ идентифицирует молекулу на основе молекулярного веса. Испытуемые образцы экстрагировали твердой фазой и смешивали с матричным раствором sDHB и совместно осаждали на мишени MALDI. Этот высушенный образец ионизировали лазером в масс-спектрометре ToF. С каждого образца снимали спектр m/z, а интактный m/z конвертировали в молекулярную массу.
Идентичность
Анализ дегликозилированной/восстановленной и дегликозилированной молекулярной массы (MALDI-ToF-MS)
[0258] Молекулярную массу дегликозилированной асфотазы альфа рассчитывали из первичной аминокислотной последовательности двух идентичных полипептидных аминокислотных последовательностей из 726 аминокислот с использованием молекулярных масс NIST и процентного содержания изотопов с 20 цистеинами, образующими 10 дисульфидных связей. Дегликозилированную молекулярную массу асфотазы альфа рассчитывали как равную 161135,20 Да.
[0259] Молекулярные массы дегликозилированной асфотазы альфа из партий 20K и из партий 2K находились в пределах 1% от рассчитанной дегликозилированной молекулярной массы (точность массы калиброванного извне MALDI-TOF), как показано в таблице 29. Средняя молекулярная масса 20K партий составляла 160881 кДа, а средняя молекулярная масса 2K партий составляла 161050 кДа, как показано в таблице 29. Спектры MALDI-TOF для партий 2K и 20 K выявили аналогичные профили пиков. Дополнительные массы, присутствующие в спектрах, были обусловлены вызванной анализом фрагментацией, неполным дегликозилированием и/или многозарядными ионами. Различия в интенсивности, наблюдаемые для идентифицированных видов, являются результатом матричной кристаллизации и лазерных эффектов.
[0260] Молекулярную массу восстановленной и дегликозилированной асфотазы альфа рассчитывали из первичной аминокислотной последовательности аминокислотной последовательности полипептида из 726 аминокислот с использованием молекулярных масс NIST и процентного содержания изотопов. Рассчитанная молекулярная масса восстановленной и дегликозилированной асфотазы альфа составляет 80577,68 Да.
[0261] Значения молекулярной массы для восстановленных и дегликозилированных партий асфотазы альфа 20K сравниваются с партиями 2K, и все значения были в пределах 1% от рассчитанной восстановленной и дегликозилированной молекулярной массы (точность массы калиброванного извне MALDI-ToF), как показано в таблице 29. Среднее количество партий 2K составляло 80530 Да, а среднее значение 20K партий составляло 80539 Да. Спектры MALDI-TоF для партий 2K и 20K выявили сопоставимые профили пиков. Дополнительные массы, присутствующие в спектрах, обусловлены индуцированной анализом фрагментацией и/или несколькими заряженными ионами. Различия в интенсивности, наблюдаемые для идентифицированных видов, являются результатом матричной кристаллизации и лазерных эффектов. Этот способ идентифицирует молекулу на основе молекулярного веса. Испытуемые образцы экстрагировали твердой фазой и смешивали с матричным раствором sDHB и совместно осаждали на мишени MALDI. Этот высушенный образец ионизировали лазером в масс-спектрометре ToF. С каждого образца снимали спектр m/z, а интактный m/z конвертировали в молекулярную массу.
[0262] Молекулярные массы дегликозилированной асфотазы альфа партий 2K и 20K были сопоставимы, и все значения находились в пределах 100 м.д. (ppm) рассчитанной дегликозилированной молекулярной массы (которая представляет собой точность массы калиброванного извне ESI-ToF-MS), как показано в таблице 29. Средняя молекулярная масса партий 2K составляла 161137,39 Да, а средняя молекулярная масса 20K партий была равна 161137,28 Да, и значения согласовались с рассчитанной дегликозилированной молекулярной массой 161135,20 Да. Восстановленные и дегликозилированные значения молекулярной массы для основного пика партий 2K были сопоставимы со значениями партий 20K, и все значения находились в пределах 100 ppm рассчитанной восстановленной и дегликозилированной молекулярной массы (которая представляет собой точность массы калиброванного извне ESI-ToF-MS), как показано в таблице 29. Средняя молекулярная масса партий 2K составляла 80575,52 Да, а средняя молекулярная масса 20K партий составляла 80574,76 Да. Значения согласуются с рассчитанным восстановленным и дегликозилированным значением молекулярной массы для асфотазы альфа 80577,68 Да. Этот способ идентифицирует молекулу на основе интактной молекулярной массы. Испытуемые образцы инъецировали в колонку С4 ОФ-ВЭЖХ и элюировали градиентом водного:органического растворителя. Элюат затем подвергали электрораспылению в масс-спектрометр ToF, и спектр из верхней половины хроматографического пика подвергали операции деконволюции для обеспечения интактной молекулярной массы.
[0263] Молярные отношения ионов цинка и магния, определяемые методом индуктивно связанной плазменной масс-спектрометрии (ICP-MS) для трех партий 20K и трех партий 2K, были сопоставимы, как показано в таблице 29. Молярные отношения ионов кальция, определяемые методом индуктивно связанной плазменно-атомной эмиссионной спектроскопии (ICP-AES) для партий 20K и 2K, были сопоставимы, как показано в таблице 29. Молекулярные соотношения ионов для цинка и кальция также согласуются с литературой, где два цинка и один кальций были связаны с каждым мономером фермента щелочной фосфатазы (см. E. Mornet et al., 2001 ʺStructural evidence for a functional role of human tissue non-specific alkaline phosphatase in bone mineralization.ʺ JBC, 276: 31171-31178; и B. Stec, KM. Holtz and ER. Kantrowitz. 2000 ʺA revised mechanism for the alkaline phosphatase reaction involving three metal ions.ʺ JMB, 299: 1303-1311). Молярное отношение ионов магния для всех партий было неожиданно ниже ожидаемой величины, указанной в литературе. ICP-AES был выбран для анализа кальция, поскольку ICP-AES не страдает от аргоновых полиатомных помех, наблюдаемых для изотопов кальция в ICP-MS. Данные ICP асфотазы альфа были приобретены на коммерческой основе. Этими способами определяли количество магния, цинка и кальция (мкг/мл), из которого были рассчитаны отношения молярных ионов для асфотазы альфа.
Идентификация и количественная характеристика сайта фосфорилирования с использованием UPLC-MSe
[0264] Сайт и относительный процент фосфорилирования определяли путем картирования триптического пептида UPLC-MS. Известно, что щелочная фосфатаза содержит сериновый остаток в активном сайте, который может образовывать серинфосфатное промежуточное соединение (см. стр. 28-29 Millán, José Luis. 2006 ʺMammalian Alkaline Phosphatases.ʺ Wiley-VCH). Было обнаружено, что тот же остаток 93 серина был фосфорилирован во всех партиях в процентах в диапазоне от 35,7% до 31,2%, как показано в таблице 29. Было обнаружено, что процент фосфорилирования не коррелирует с специфической активностью асфотазы альфа (pNPP), связыванием с HA, Km и Kcat (данные не показаны). Исходя из недостатка корреляции, фосфорилированное промежуточное соединение не является ступенью ограничения скорости активности асфотазы альфа. Подтверждение и идентификацию сайта фосфорилирования на S93 в асфотазе альфа подтверждали с помощью пептидного картирования UPLC-MSe. Для анализа использовалась теоретическая последовательность асфотазы альфа и предсказанные триптические расщепления. Полученные данные LC-MSe собирали и обрабатывали с использованием Waters Biopharmalynx v.1.2. Программное обеспечение Biopharmalynx (Waters) идентифицировало 13-й триптический пептид из первичной последовательности (T13), который содержит S93, серин активного сайта из ферментативной части молекулы, имеющий Cys102, ограниченный с ac, и S93, существующий как фосфорилированный, так и нефосфорилированный. После идентификации в качестве части карты пептида, экстрагированные ионные хроматограммы (EIC) вытягивали для каждого из этих пептидов и сглаживали и интегрировали. Из результирующих площадей пиков EIC отношение площади пика фосфорилированного пептида к общей площади пика дает относительный процент фосфорилирования на S93. Подтверждение +80 Да, связанного с фосфорилированной боковой цепью серина, было получено путем изучения картины фрагментации MSe, сгенерированной для предлагаемого фосфорилированного пептида T13, как показано на фиг. 36. Эта картина фрагментации подтвердила, что S93 сдвинут на +80 Да (масса фосфорилирования), что наблюдается с помощью ионов y12 и y13. Каждую партию асфотазы альфа восстанавливали и алкилировали, затем расщепляли трипсином. Обнаружение проводили с помощью ESI-Q-ToF-MS с использованием масс-спектрометра Waters Acquity UPLC и Synapt Q-ToF, работающего в режиме электрораспыления с положительным ионом.
ГЛИКОЗИЛИРОВАНИЕ
Масс-профилирование N-связанных олигосахаридов (MALDI-ToF-MS)
[0265] Восстановленные олигосахариды из трех партий 2K и трех партий 20K были проанализированы с помощью MALDI-ToF, результаты приведены в таблице 29. Те же самые виды гликанов с аналогичным относительным процентом наблюдались в партиях 20K и в партиях 2K. Этот метод идентифицирует гликаны, связанные с лекарственным веществом, с помощью молекулярных масс. Гликаны были ферментативно расщеплены из лекарственного вещества с использованием переваривания PNGазы-F и последующего подкисления. Гликаны затем экстрагировали твердой фазой и смешивали с супер-3, матричным раствором 4-дигидроксибензойной кислоты и совместно осаждали на мишени MALDI. Этот высушенный образец ионизировали лазером в масс-спектрометре ToF. Собирали спектры m/z (M+Na) +.
Профилирование 2-AB-меченых N-связанных олигосахаридов (ВЭЖХ)
[0266] Типы гликоформ и относительный процент каждой гликоформы анализировали с помощью 2-AB мечения и анализа ВЭЖХ. Результаты показаны в таблице 29. Пик с одинаковым временем удерживания и с аналогичным относительным процентом наблюдался во всех партиях (данные не показаны), предполагающие одинаковые типы олигосахаридов и аналогичные уровни. Партия № 36 была запущена позднее. В качестве примера процент основной гликоформы FA2 был нанесен на график против ферментативной активности. Не наблюдалось корреляции между процентом FA2 и специфической активностью (pNPP), связыванием HA, Km и Kcat (данные не показаны).
[0267] Этот анализ характеризует картину гликозилирования путем определения олигосахаридов, связанных с молекулой лекарственного средства, на основе времени удержания и площади пика флуоресцентно меченных свободных олигосахаридов. Свободные олигосахариды из лекарственного вещества высвобождались ферментативно и метились 2-аминобензамидом (2-АВ) путем восстановительного аминирования. Образцы затем инъектировали в систему ВЭЖХ с колонкой Людгера, а 2AB меченные олигосахариды были разделены и детектированы с детектором флуоресценции; 330 нм возбуждение, 420 нм излучение. Хроматограммы каждой партии сравнивали, и площадь пика каждого разрешенного пика использовалась для определения относительного количества олигосахарида из каждой партии.
Профиль гликопептида с помощью UPLC-QToF-MS
[0268] Олигосахаридный профиль на каждом сайте гликозилирования оценивали с помощью анализа гликопептидов с помощью UPLC-MS. Тканенеспецифичная щелочная фосфатаза (TNAP) имеет пять сайтов N-гликозилирования (см. стр. 54-55 Millán, 2006 Wiley-VCH). Асфотаза альфа содержит дополнительный сайт N-гликозилирования в Fc-участке, обеспечивающий в общей сложности шесть сайтов N-гликозилирования на мономер. На фиг. 44 по фиг. 49 показаны профили фингерпринтов гликопептида для N123 в T15-16, N213 в T26-27, N254 в T33, N286 в T35, N413 в T45-46 и N564 в T55. Фингерпринты гликопептидов UPLC-Q-ToF-MS суммированных N-связанных олигосахаридов дают подробную характеристику N-гликозилирования. Идентификацию видов гликопептидов по масс-анализу и относительное количественное определение каждого вида проводили путем трансформации множественных зарядовых состояний, наблюдаемых в необработанных спектрах фингерпринтов гликопептида, в однократно заряженные и деизотопные спектры для каждого сайта. Результаты этих преобразований приведены в таблице 29. В ней перечислены все массы, наблюдаемые для гликоформ, и их относительный % для гликоформ, составляющих≥5% от общего содержания гликоформ в каждом наблюдаемом N-связанном сайте олигосахарида. В этом анализе асфотазу альфа восстанавливали и алкилировали, затем расщепляли трипсином. Обнаружение проводили с помощью ESI-Q-ToF-MS с использованием масс-спектрометра Waters Acquity UPLC и Synapt Q-ToF, работающего в режиме электрораспыления с положительным ионом. Для анализа использовалась теоретическая последовательность асфотазы альфа и предсказанные триптические расщепления. Молекулярные массы для теоретических гликопептидов рассчитывали с использованием молекулярных масс NIST и процентного содержания изотопов. Гликопептиды наблюдаются как коллекции связанных гликоформ с близким временем элюирования и, следовательно, требуют суммирования спектров для обеспечения полного дополнения гликопептида для конкретного сайта. Для каждого из наиболее распространенных видов пептидов, содержащих каждый из N-связанных олигосахаридных сайтов в асфотазе альфа для каждой тестируемой партии, были получены специфичные для сайта спектры суммирования (фингерпринтов массы гликопептидов).
Общее содержание сиаловой кислоты (TSAC)
[0269] Общее содержание сиаловой кислоты определяли с помощью HPAE-PAD. Как показано в таблице 29, содержание сиаловой кислоты для каждой партии находится в пределах спецификации 0,9-3,5 моль сиаловой кислоты на моль мономера. Учитывая изменение содержания сиаловой кислоты из-за ее относительно меньшего количества, партии 2K и партии 20K были сопоставимы. Не наблюдалось корреляции между значением TSAC и специфической активностью (pNPP), связыванием HA, Km и Kcat.
АКТИВНОСТЬ
Специфическая Активность (pNPP)
[0270] Специфическая активность асфотазы альфа определялась для трех 20K и трех партий 2K. Средняя специфическая активность партий 20K составляет 981,7 Ед/мг и 861,7 Ед/мг для трех партий 2 K. Специфическая активность всех партий была в пределах 620-1250 Ед/мг и была сопоставимой для партий 20K и 2K. Этот способ используется для определения ферментативной активности асфотазы альфа с использованием pNPP в качестве субстрата. Асфотаза альфа представляет собой рекомбинантный белок, который имеет один домен из тканенеспецифичного фермента щелочной фосфатазы человека. Этот домен является каталитически активным и гидролизует сложные эфиры фосфатов. Анализ проводят при насыщении фермента для достижения и поддержания Vmax для продолжительности измерения. рNPP гидролизуют в продукт желтого цвета (максимальное поглощение при 405 нм). Скорость реакции прямо пропорциональна активности фермента. Одна единица (Ед) соответствует 1 мкмоль pNPP, вырабатываемой в минуту в условиях используемого способа. Специфическая активность с помощью pNPP (ферментативная активность на мг белка) была рассчитана на основе ферментативной активности и концентрации белка на A280. Результаты показаны в таблице 29.
Связывание гидроксиапатита
[0271] % связывания гидроксиапатита, определяемый анализом связывания гидроксиапатита (HA) в 2K партиях асфотазы альфа, сравним с партиями 20K со всеми значениями в диапазоне 85-97%. Средний % HA-связывания составляет 91% для обеих партий 20K и 2K (таблица 29). Анализ HA-связывания измеряет ферментативную активность комплекса асфотазы альфа для pNPP при связывании с гидроксиапатитом. Синтетический субстрат, пара-нитрофенил фосфат (pNPP), когда он гидролизуется асфотазой альфа, продуцирует продукт желтого цвета, который может быть количественно измерен при поглощении при 405 нм. Этот анализ проводили при насыщении субстрата для достижения и поддержания Vmax в течение реакции.
PPi ферментный кинетический анализ с использованием физиологического релевантного вещества
[0272] Кинетические параметры Km и Kcat были определены с помощью кинетического анализа PPi. Образцы прогонялись в ряде случаев параллельно с эталонным стандартом. Для сравнения, Km и Kcat отображаются на графике в процентах от эталонного стандартного значения во время тестирования образца (таблица 29). Все значения Km были в пределах 30% от эталонного стандартного значения параллельного запуска и считаются сравнимыми. Все значения Kcat были в пределах 20% от эталонного стандартного параллельного запуска и считаются сравнимыми. Кm % эталона и Kcat % эталона для партий 2K были сопоставимы с партиями 20K. Кинетический анализ PPi измеряет активность очищенной асфотазы альфа (ферментативную активность) с использованием PPi, субстрата щелочной фосфатазы, и определяет кинетические константы Km и Kcat гидролиза PPi в физиологических условиях (37°C, pH 7,4).
ПРИНУДИТЕЛЬНАЯ ТЕМПЕРАТУРНАЯ ДЕГРАДАЦИЯ
[0273] Чтобы дополнительно обеспечить сопоставимость партий 2K и 20K, было проведено параллельное исследование принудительной деградации для сравнения путей и кинетики деградации. Для исследования были использованы три партии 2K (№ 35, № 36 и № 38) и три партии 20K (№ 40, № 42 и № 34), приведенные в таблице 33. Некоторые партии требовали использования материала лекарственного продукта (DP) из-за ограниченной доступности образца BDS.
[0274] Партии № 35, № 36, № 40, № 42 и № 34 разбавляли с использованием буферного раствора асфотазы альфа (25 мМ фосфата натрия и 150 мМ хлорида натрия, рН 7,4) до 40 мг/мл, чтобы соответствовать концентрации белка в партии # 38. Для достижения значительных уровней деградации для установления кинетики деградации, условия принудительной деградации были выбраны на основе предварительного скрининга условий принудительной деградации. Аликвоты (200 мкл) были созданы для каждого анализа теста в каждый момент времени (0, 14, 24 и 48 часов). Аликвоты T0 выдерживали при 5°C до тестирования. Остальные аликвоты образца инкубировали при 40°С в течение 14, 24 и 48 часов. Образцы анализировали с помощью SDS-PAGE, GP-HPLC, AEX, RP-HPLC, картирования пептидов и специфической активности, как указано в таблице 30, для определения сравнимости. Результаты испытаний для исследования принудительной температурной деградации приведены в таблице 31 для трех партий 20K и трех партий 2K.
Таблица 33. Партии, используемые для исследования деградации
SDS-PAGE (В невосстанавливающих условиях и в восстанавливающих условиях)
[0275] T0 и T48 час образцов анализировали с помощью SDS-PAGE (в невосстанавливающих и восстановливающих условиях) на чистоту. % главной полосы SDS-PAGE в невосстанавливающих условиях составлял 100% для всех партий при T=0 и уменьшался до среднего значения 97,3% для партий 20K и 96,8% для партий 2K, как показано в таблице 31. Тенденция деградации сравнима для партий 20K и 2K. Виды с высокой молекулярной массой были обнаружены в T48 часов образца для всех шести партий. Этот вид исчез при восстановлении. Этот результат предполагает, что образование агрегатов, связанных с дисульфидными связями, представляет собой путь деградации для всех партий. Никаких дополнительных полос не наблюдалось при уменьшении SDS-PAGE для партий 20K и 2K при T=0 и T=48 часов, что указывало на отсутствие деградации пептидных связей. Этот тест отделяет белок на основе его молекулярной массы и обеспечивает анализ чистоты интактного белка, выраженного в процентах от главной полосы. Образец белка отделяют на геле, окрашивали кумасси-синим для визуализации, отмывали окраску и анализировали с помощью денситометрии. Молекулярная масса асфотазы альфа, продуктов разложения, и связанных с процессом примесей оценивается по сравнению с известными стандартами молекулярной массы. Количественные измерения отделенных белков выполняются путем сканирования и анализа с помощью денситометрического анализа.
GP-HPLC
[0276] Деградация с образованием агрегатов и фрагментов, вызванных термическим стрессом, контролировалась с помощью GP-HPLC, и результаты показаны в таблице 31. Профили пиков для всех партий при T=0 были сходными, что свидетельствует о наличии одного и того же вида. Термический стресс привел к появлению пика примерно через 6 минут, что соответствовало большим агрегатам, которые со временем увеличивались. Профили пиков всех партий при T=14 часов, T=24 часа и T=48 часов были похожи, что указывает на аналогичный путь деградации. Наклоны снижения основного пика с течением времени для всех партий были схожи с учетом того, что используемые материалы были разных возрастов. Подобный наклон указывает на аналогичную кинетику деградации.
Анион-обменная хроматография (AEX)
[0277] Деградация асфотазы альфа также контролировалась с помощью AEX, который разделяет белки на основе их заряда. Те же самые профили пиков наблюдались для всех партий при T=0, что указывало на тот же вид во всех партиях. Термический стресс привел к увеличению пиков в окне времени удержания приблизительно 22-26 минут. Сопоставимые профили пиков для всех партий наблюдались для всех партий при T=14 часов, T=24 часа и при T=48 часов, что указывает на образование одного и того же вида путем термического разложения. Аналогичные наклоны деградации, контролируемые по уменьшению основного пика, наблюдались для всех партий с учетом небольшой разницы при T=0 (как описано ранее) и разницы в возрасте разных партий.
RP-HPLC
[0278] При анализе с помощью RP-HPLC средний пик в окне времени удержания приблизительно 21,2-21,3 минуты наблюдался для всех партий, указывая на один и тот же вид во всех партиях. Основной пик был единственным основным пиком, наблюдаемым для всех партий при T=0. Термический стресс привел к увеличению пика, элюирующегося сразу же после основного пика. Тот же самый пик от термической деградации, наблюдаемый для всех партий при T=14, T=24 и T=48 часов, указывает на тот же путь деградации. Кинетика деградации, контролируемая по уменьшению основного пика, была сопоставима для всех партий, о чем свидетельствуют сходные наклоны всех партий.
Картирование пептидов
[0279] Время T0, T24 и T48 образцов для партий 20K и 2K было проанализировано картированием пептидов для оценки потенциальной химической деградации. Не было обнаружено существенной разницы между 20K и 2K во все моменты времени (T=0 час и T=48 часов). Этот метод денатурирует, уменьшает и алкилирует белок гидрохлоридом гуанидина, дитиотреитолом и иодацетатом, а затем перевариванием трипсином протеазы. Полученные пептидные фрагменты разделяют градиентным ВЭЖХ с обращенной фазой и детектируют при 210 нм.
Специфическая Активность (pNPP)
[0280] Специфическая активность (pNPP) 20K партий и 2K партий асфотазы альфа определялась ферментативным анализом после инкубации при 40°C в каждый момент времени (T0, T14, T24 и T48). Активность партий 20K была сопоставима с партиями 2K в каждый момент времени. Снижение активности, вызванное термальным стрессом, следует по сходным склонам для всех партий с течением времени. Протеинфосфатаза - фермент, который контролирует удаление фосфатной (PO43-) группы из молекул белка. Анализ фосфатазы pNPP использовали для определения активности фосфатазы и сравнения партий 20K и 2K. Этот способ используется для определения ферментативной активности асфотазы альфа с использованием pNPP в качестве субстрата. Асфотаза альфа представляет собой рекомбинантный белок, который имеет один домен из тканенеспецифичного фермента щелочной фосфатазы человека. Этот домен является каталитически активным и гидролизует сложные эфиры фосфатов. Анализ проводят при насыщении фермента для достижения и поддержания Vmax для продолжительности измерения. рNPP гидролизуют в продукт желтого цвета (максимальное поглощение при 405 нм). Скорость реакции прямо пропорциональна активности фермента. Одна единица (Ед) соответствует 1 мкмоль pNPP, вырабатываемому в минуту в условиях используемого способа. Специфическую активность с помощью pNPP (ферментативная активность на мг белка) рассчитывали на основе ферментативной активности и концентрации белка на A280.
[0281] Установлена сопоставимость асфотазы альфа, изготовленной в масштабе 20K, и асфотазы альфа, изготовленной в масштабе 2K. По возможности, три партии по каждой шкале анализировались параллельно, используя физико-химические методы для оценки чистоты асфотазы альфа, примесей, вариантов заряда, размера, структуры, гликозилирования, дисульфидной связи, свободных тиолов и активности. Кроме того, было проведено параллельное исследование принудительной деградации для оценки путей деградации и кинетики. Результаты показывают, что произведенные продукты в 20K и 2K масштабах были сопоставимы.
[0282] Чистоту партий оценивали аналитическим ультрацентрифугированием, SDS-PAGE/LoC, SDS-PAGE, GP-HPLC и AEX. Результаты показали сопоставимую чистоту для всех партий. Те же самые типы агрегатов на очень низких сравниваемых уровнях были обнаружены во всех партиях с помощью AUC, SDS-PAGE и GP-HPLC.
[0283] Варианты заряда, общие для рекомбинантных белков, оценивались с помощью cIEF. Тот же вид на сопоставимых уровнях наблюдался для всех партий, что свидетельствует о последовательном уровне посттрансляционных модификаций.
[0284] Молекулярный размер партий оценивали с помощью SEC-MALS и интактной молекулярной массы MALDI-ToF. Идентичность была подтверждена дегликозилированной/восстановленной и дегликозилированной молекулярной массой MALDI-ToF и дегликозилированной/восстановленной и дегликозилированной молекулярной массой ESI-ToF. Структуру также оценивали с помощью CD, дисульфидного связывания и свободных тиолов (ЖХ/MС), полного свободного тиолового анализа, анализа содержания металлов (ICP-MS/ICP-AES) и заполнения участка фосфорилирования. Сопоставимые молекулярные массы и сопоставимые гидродинамические размеры были получены для всех партий, что указывает на аналогичные модификации и аналогичную конформацию асфотазы альфа всех партий.
[0285] Гликозилирование асфотазы альфа оценивали с помощью MALDI-ToF свободного гликана, fHPLC 2AB-меченного олигосахарида, ЖХ-МС гликопептидного анализа и общего содержания сиаловой кислоты. Те же олигосахаридные виды на сопоставимых уровнях были получены для всех партий.
[0286] Активность лекарственного вещества оценивали по специфической активности (pNPP), связыванию HA и активности PPi. Результаты показали, что партии 20K асфотазы альфа были сопоставимы с партиями 2K асфотазы альфа.
[0287] Наконец, один и тот же путь деградации и аналогичная кинетика деградации партий 20K и 2K наблюдались путем параллельного исследования принудительной деградации с использованием термического стресса.
[0288] Таким образом, партии 20K асфотазы альфа и партии 2K сопоставимы, что подтверждается расширенной характеристикой с использованием множества ортогональных способов и параллельного исследования принудительной деградации. Таким образом, материал из партий 20K должен иметь сопоставимую безопасность и эффективность с материалом из партий 2K.
Пример 10. Дополнительная сопоставимость асфотазы альфа, изготовленной в масштабах 2000 л (2K) и 20000 л (20K)
[0289] Семь партий асфотазы альфа были получены в масштабе 20000 л, а пять дополнительных партий асфотазы альфа были получены с использованием масштаба 2000 л. Продукты для обоих масштабов были проанализированы и оказались сопоставимыми (данные не показаны).
| название | год | авторы | номер документа |
|---|---|---|---|
| ПРОИЗВОДСТВО ЩЕЛОЧНЫХ ФОСФАТАЗ | 2016 |
|
RU2745528C2 |
| СПОСОБ ПОЭТАПНОГО УДЕРЖИВАНИЯ И ПРОИЗВОДСТВЕННЫЙ МОДУЛЬ ДЛЯ ПОЛУЧЕНИЯ БИОМАКРОМОЛЕКУЛ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2769767C1 |
| БЕЛКОВЫЕ КОМПОЗИЦИИ ПРОТИВ VEGF И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2020 |
|
RU2824045C2 |
| БЕЛКОВЫЕ КОМПОЗИЦИИ ПРОТИВ VEGF И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2020 |
|
RU2795973C1 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ПОЛИПЕПТИДА | 2011 |
|
RU2577972C2 |
| ПРИМЕНЕНИЕ НИЗКОЙ ТЕМПЕРАТУРЫ И/ИЛИ НИЗКОГО pН В КУЛЬТУРЕ КЛЕТОК | 2008 |
|
RU2478702C2 |
| БЕЛКОВЫЕ КОМПОЗИЦИИ ПРОТИВ VEGF И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2020 |
|
RU2778325C1 |
| БЕЛКОВЫЕ КОМПОЗИЦИИ ПРОТИВ VEGF И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2020 |
|
RU2785994C1 |
| БЕЛКОВЫЕ КОМПОЗИЦИИ ПРОТИВ VEGF И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2020 |
|
RU2788949C1 |
| СПОСОБ ОЧИСТКИ КОНЪЮГАТОВ НА ОСНОВЕ IL-15/IL-15R-АЛЬФА | 2015 |
|
RU2701301C2 |
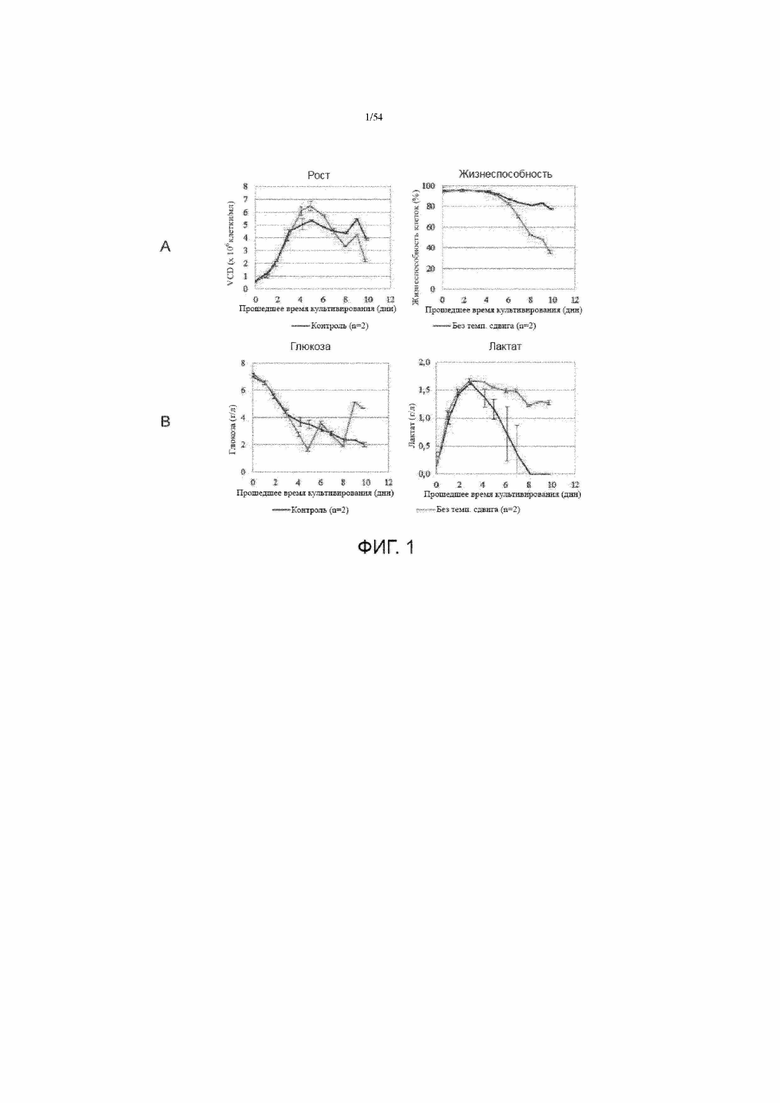

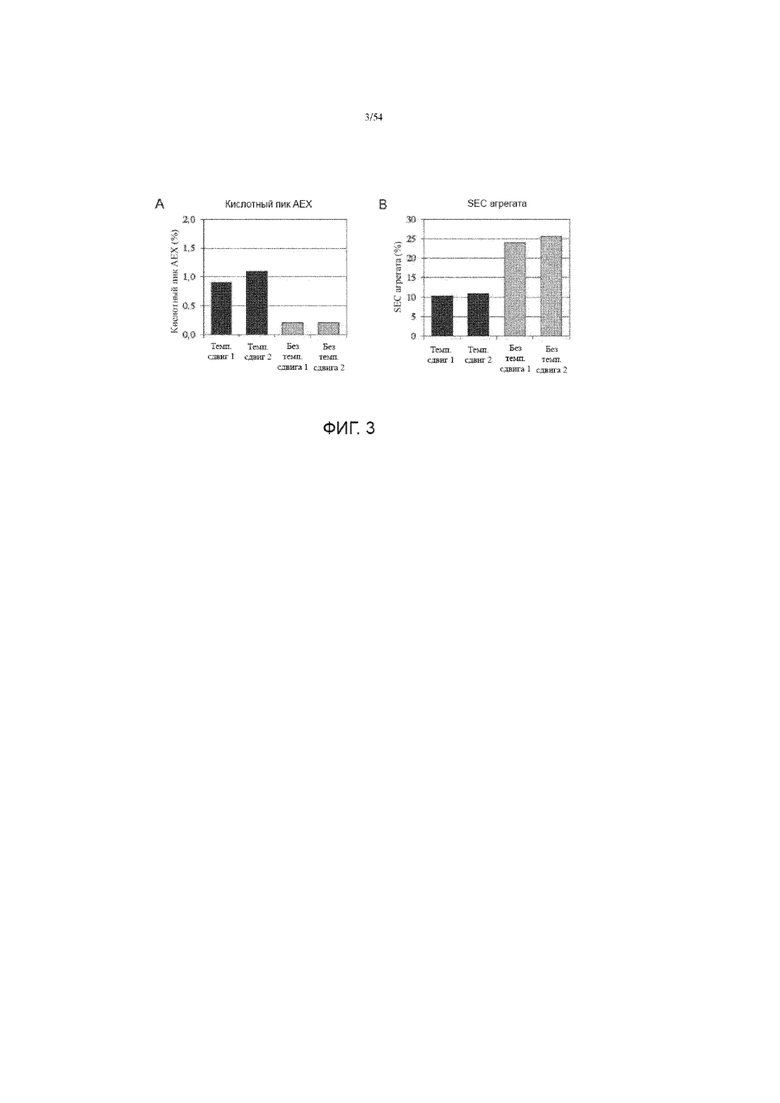
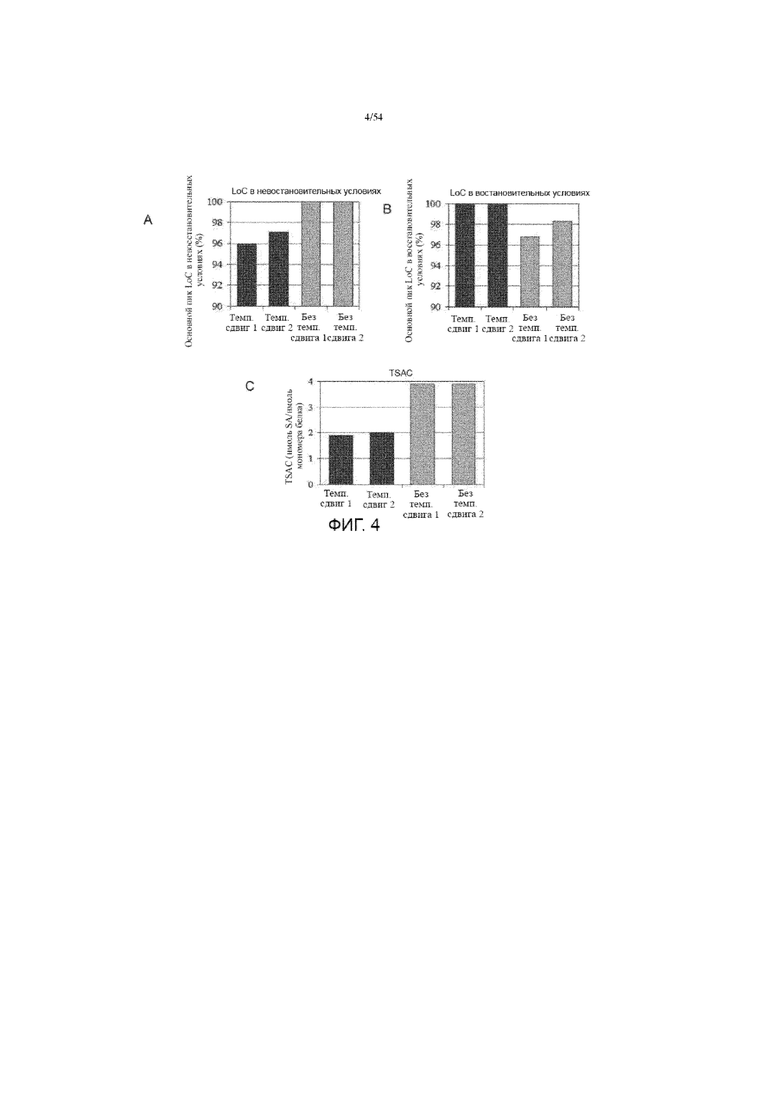
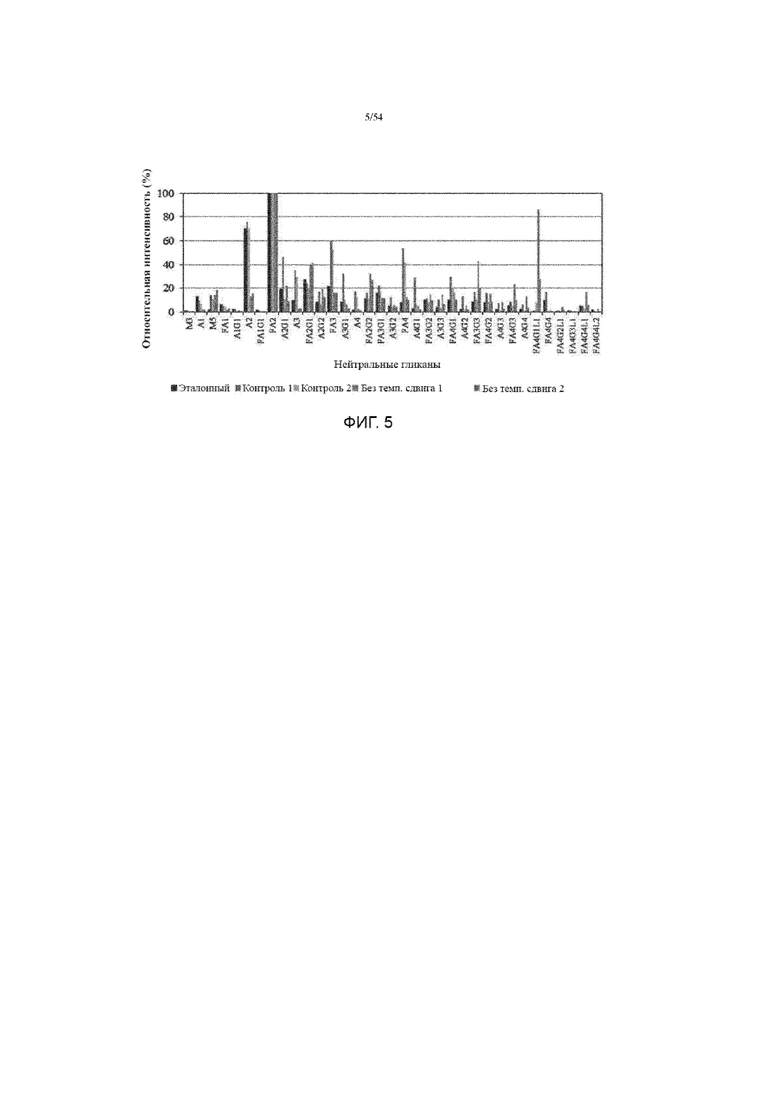
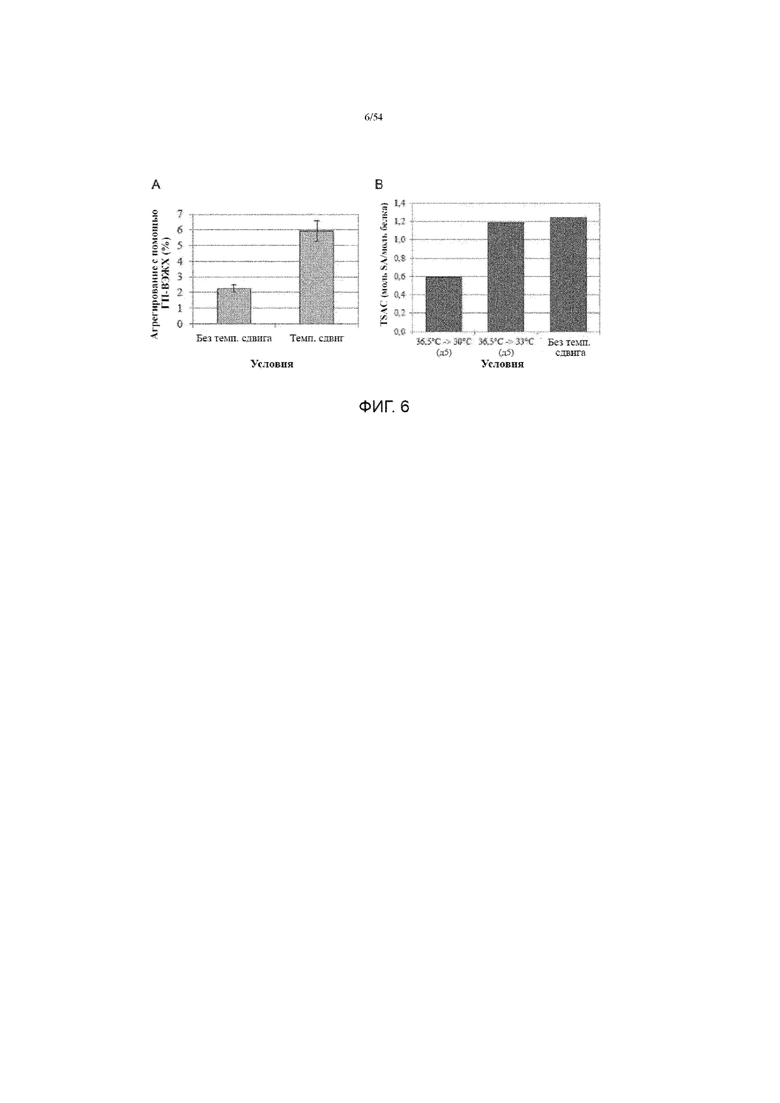
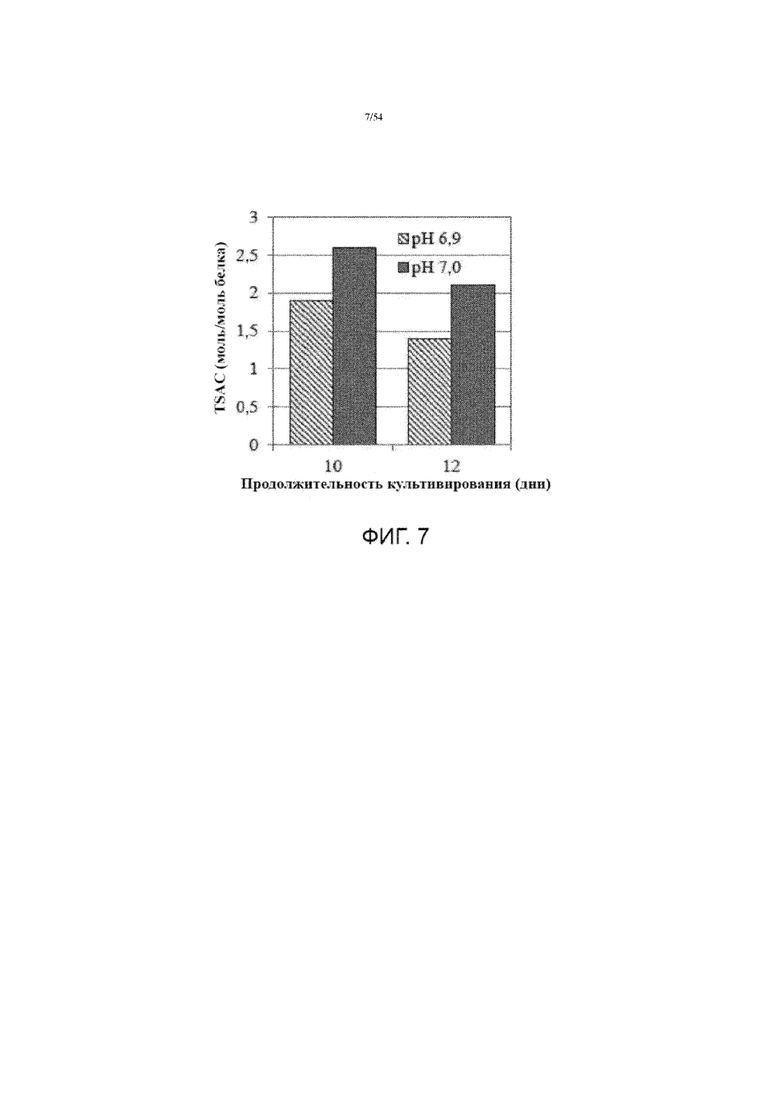
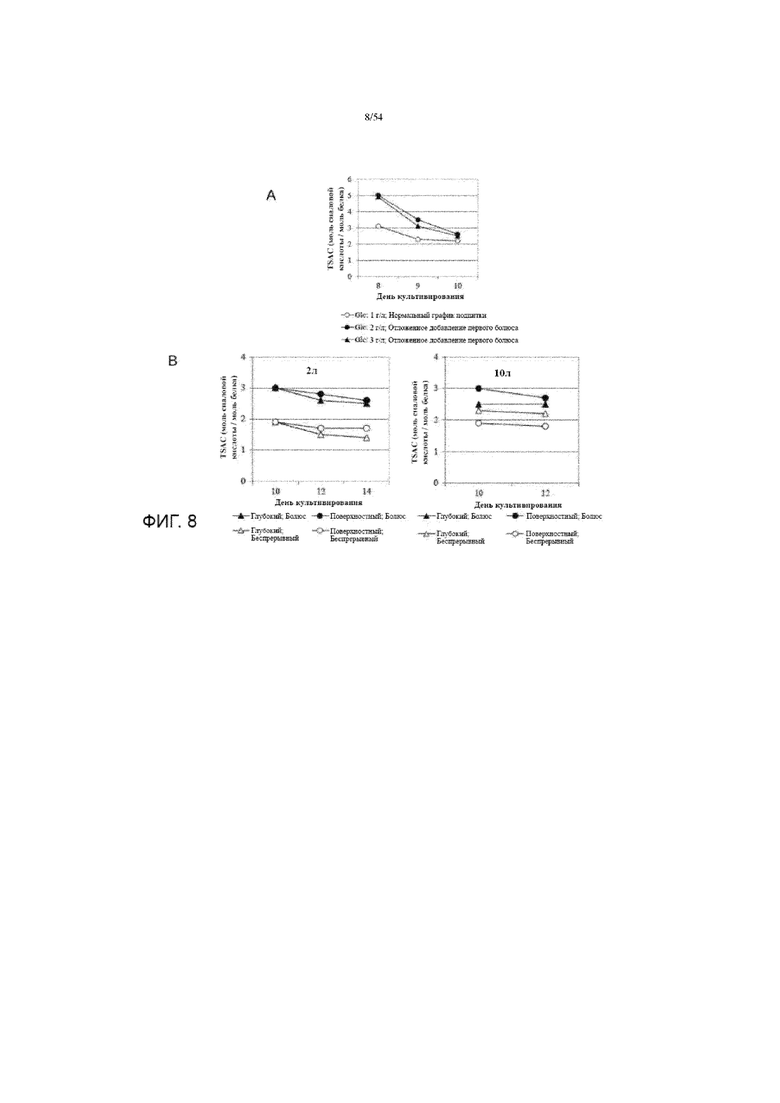

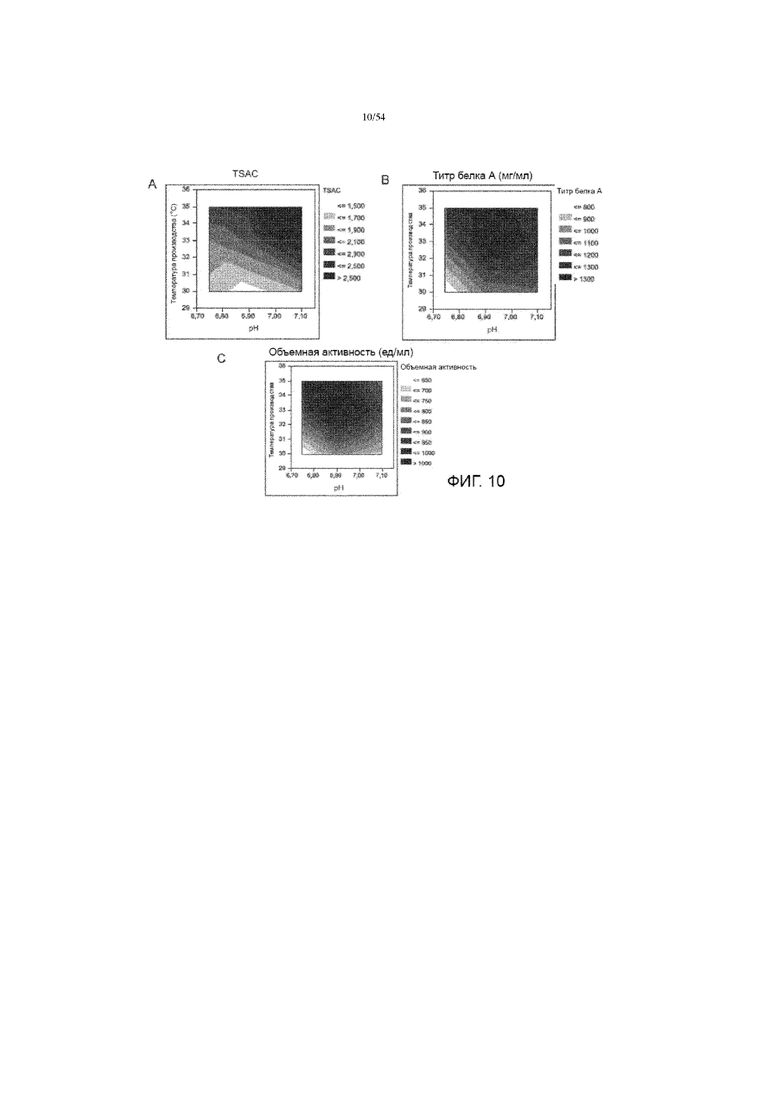
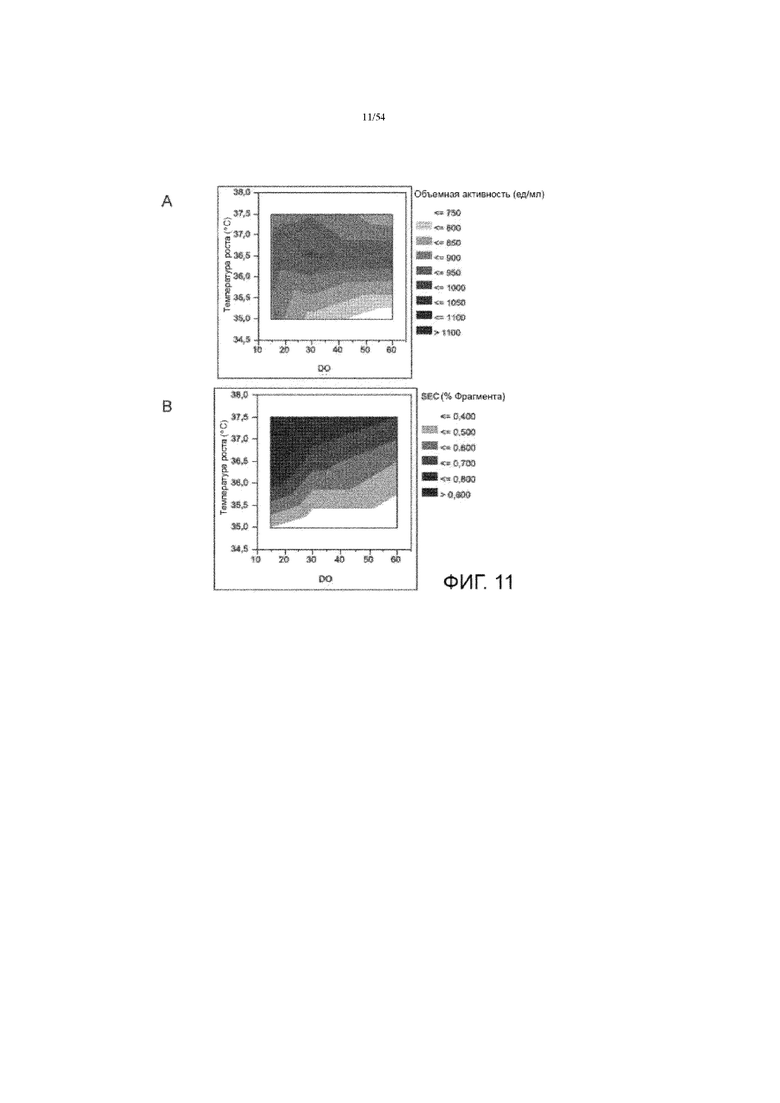

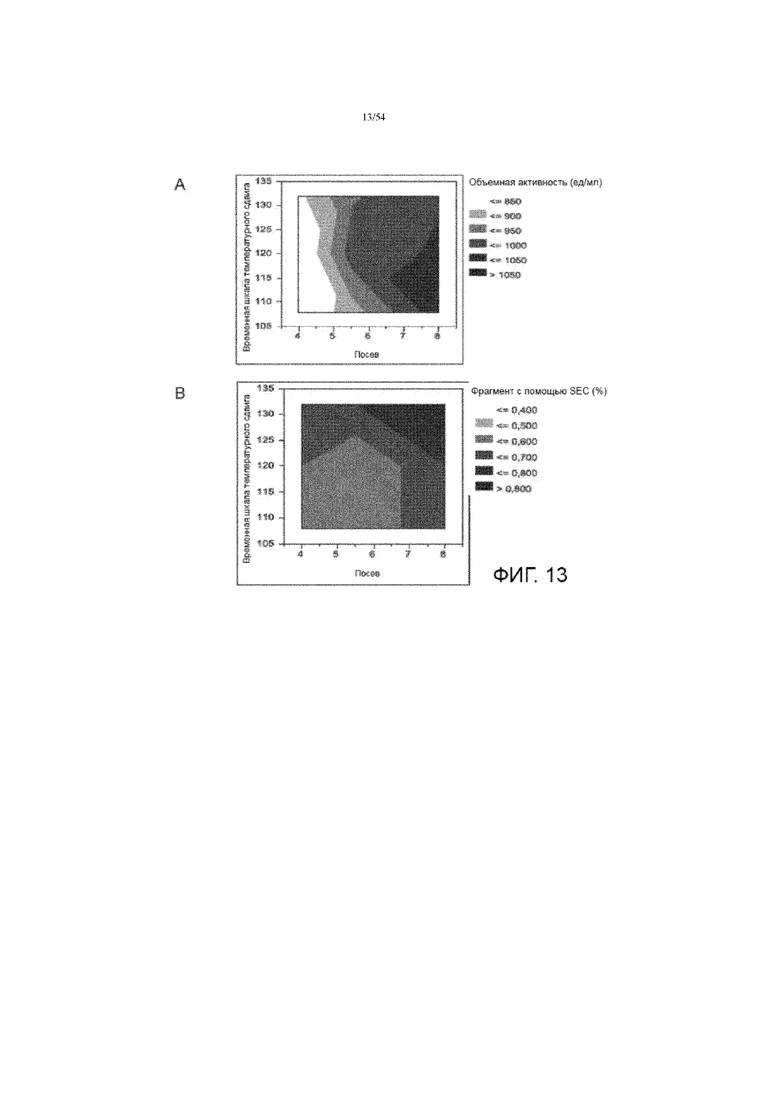


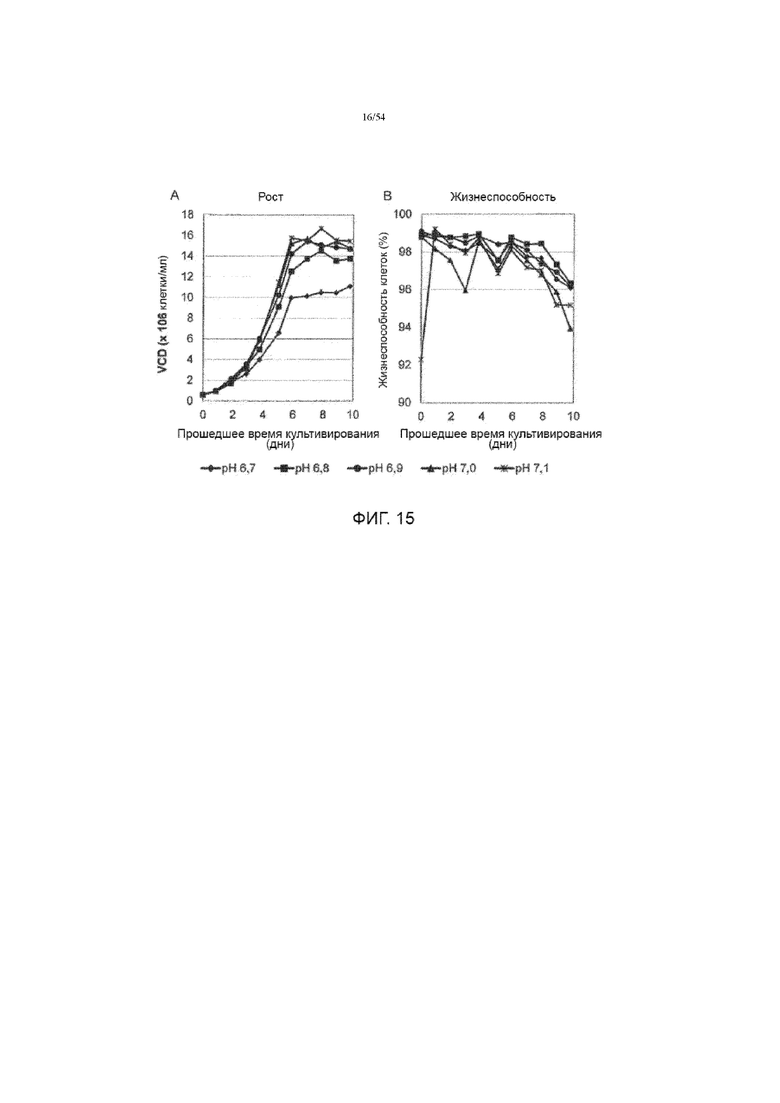
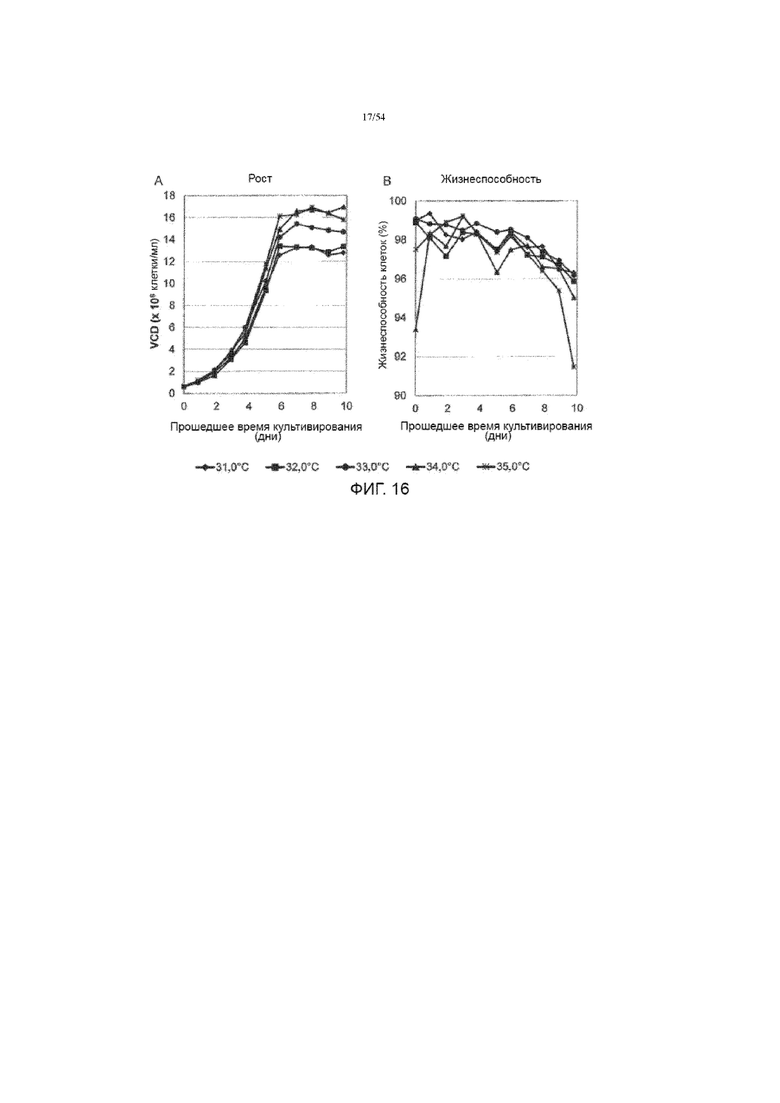
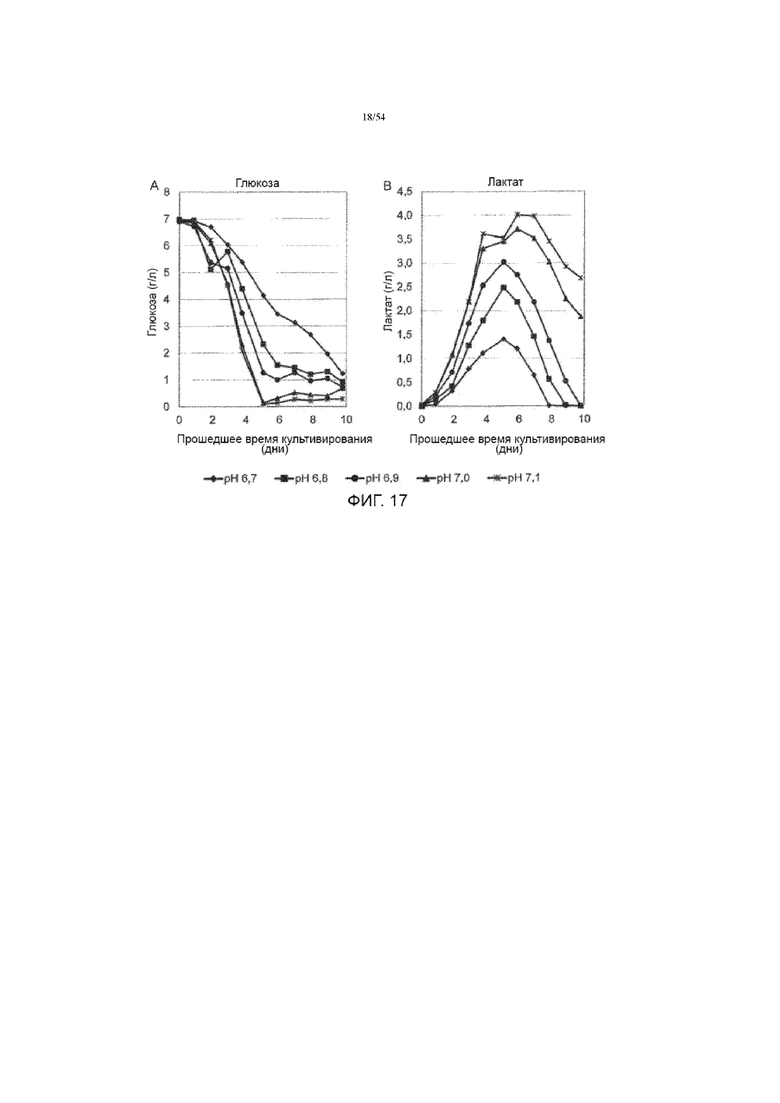
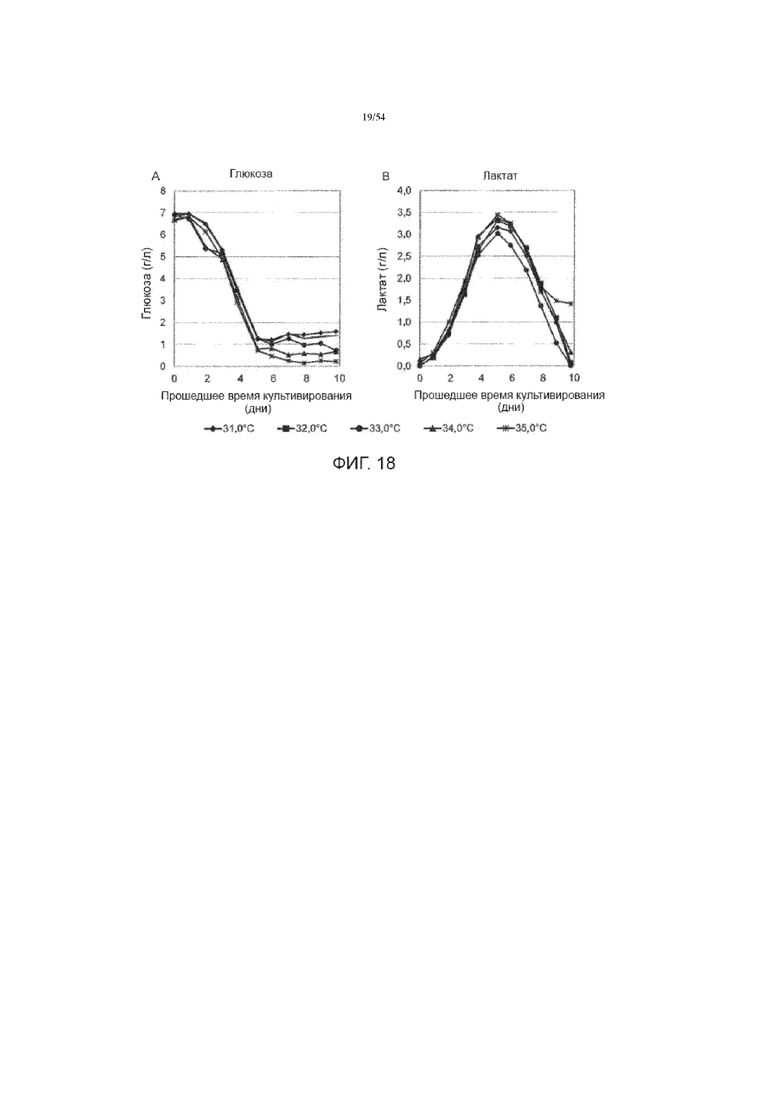
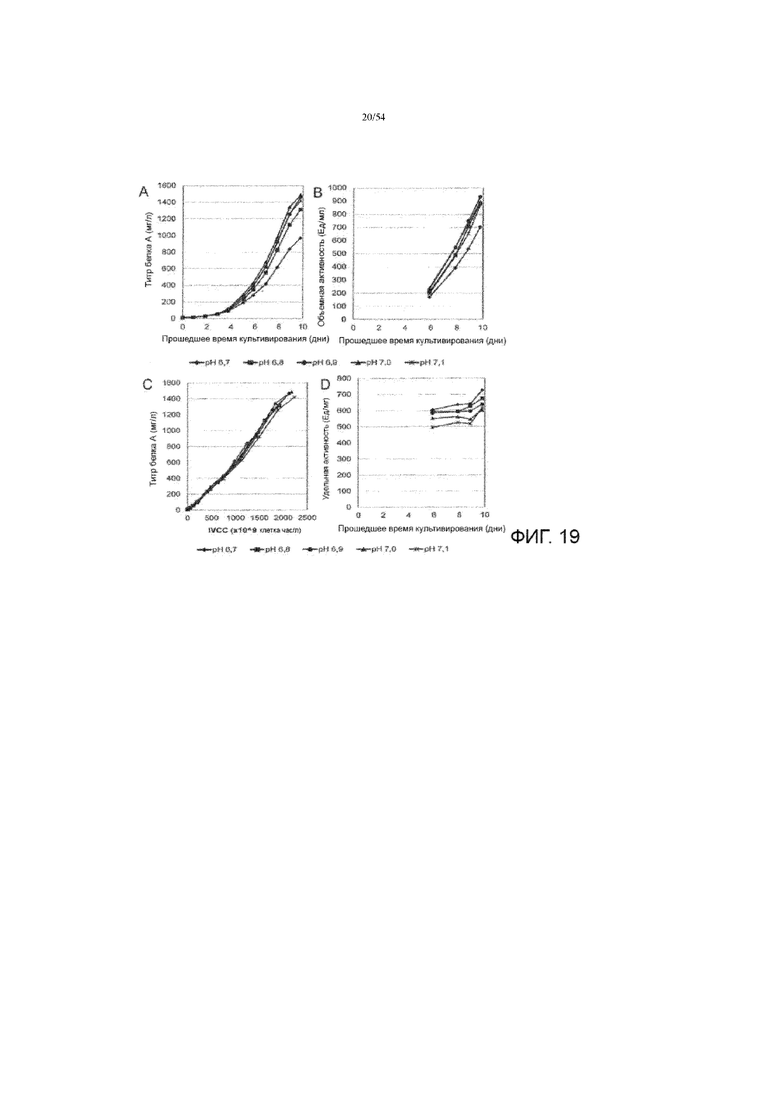

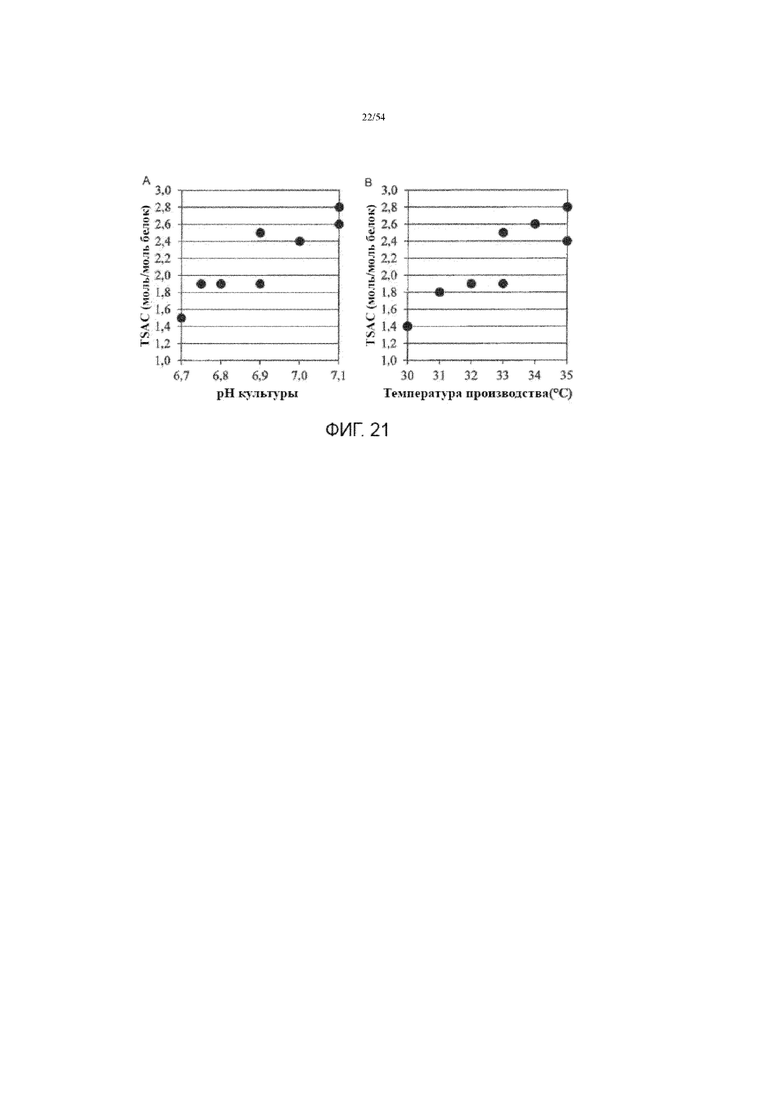
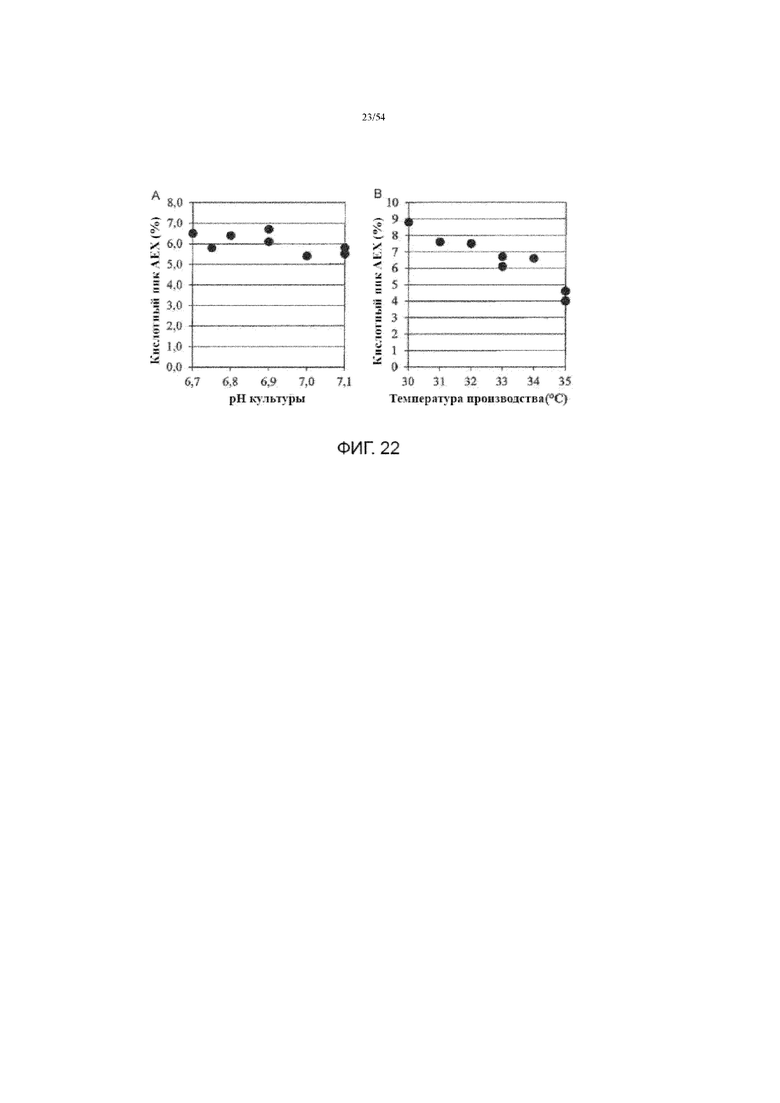
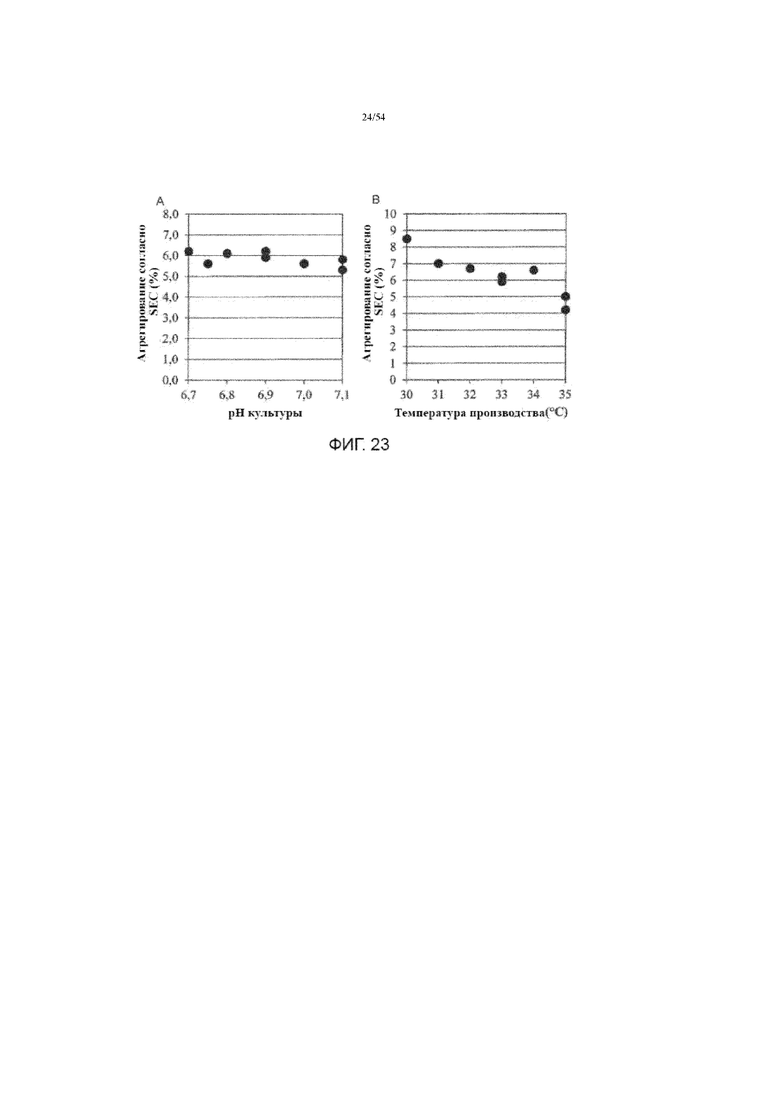
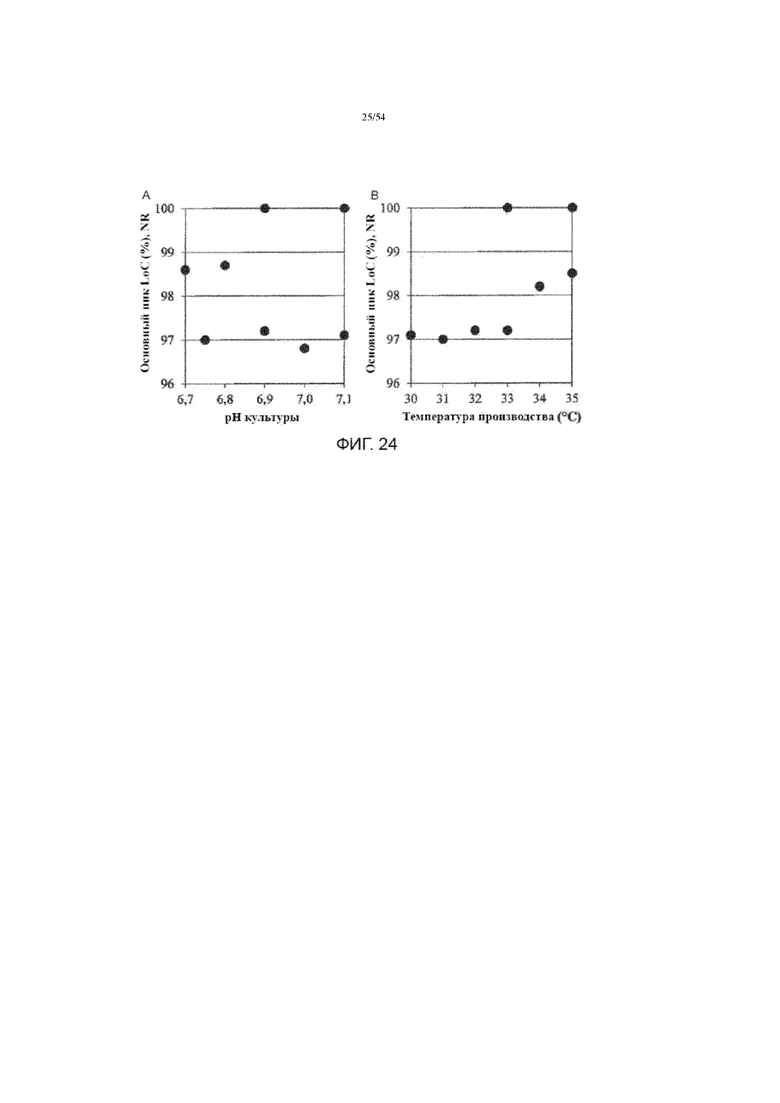
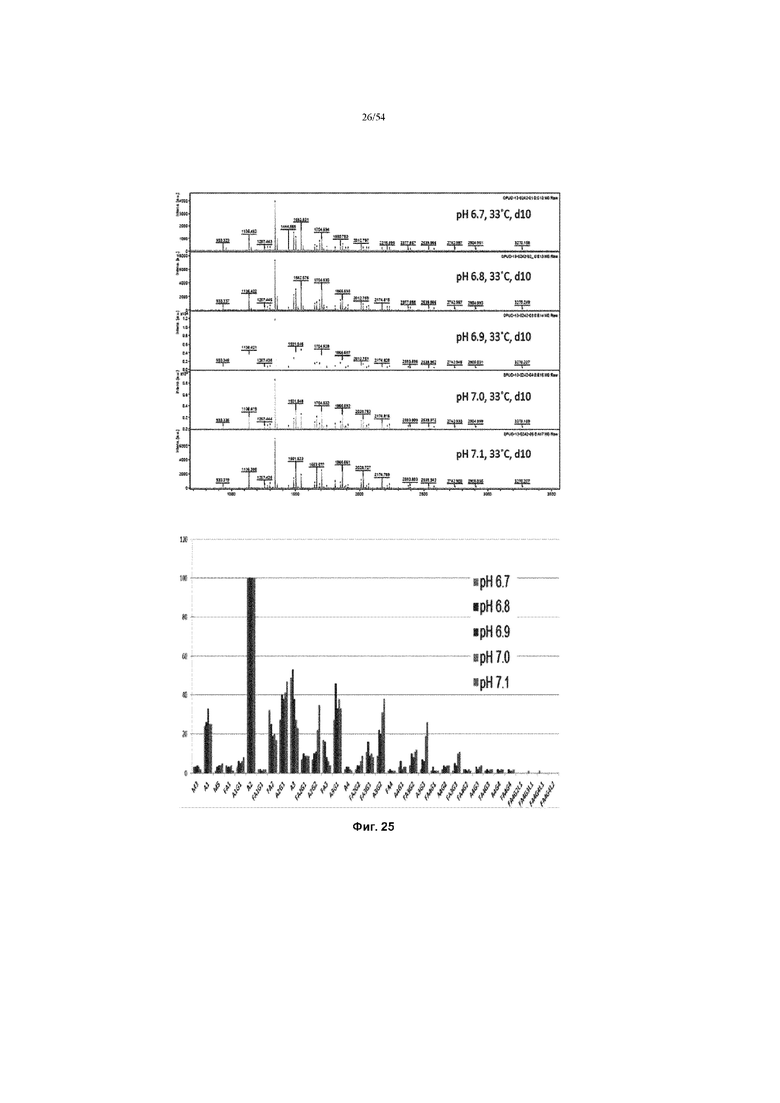
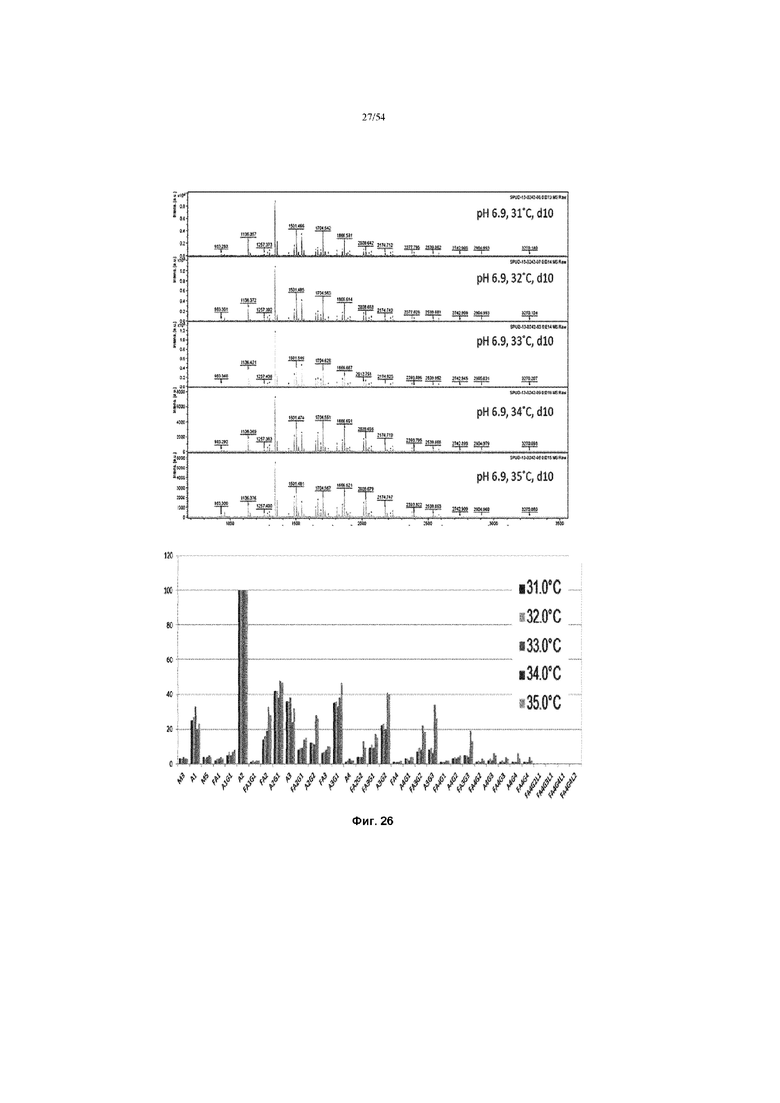
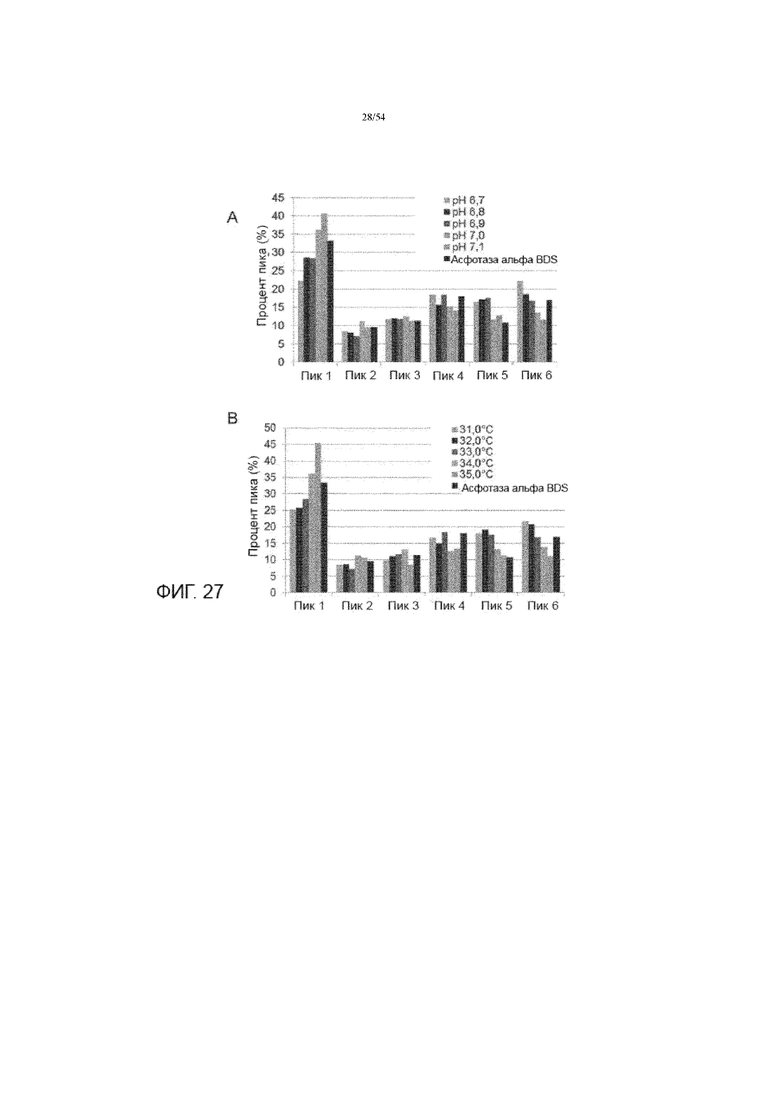

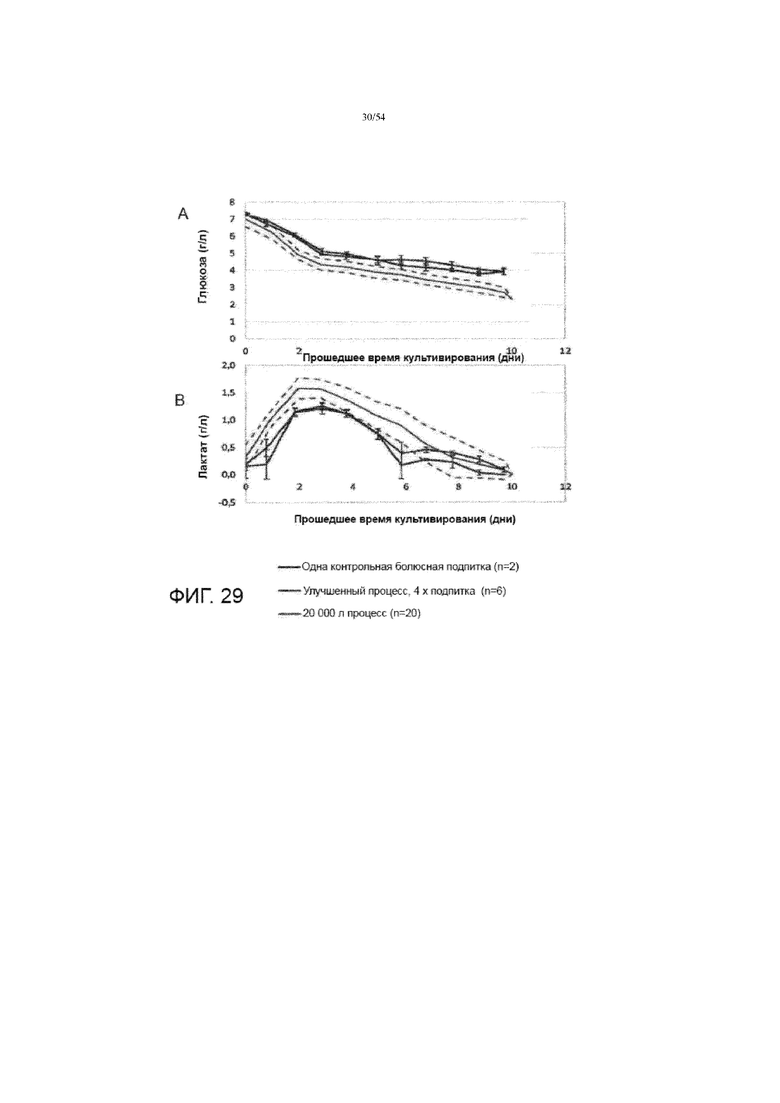

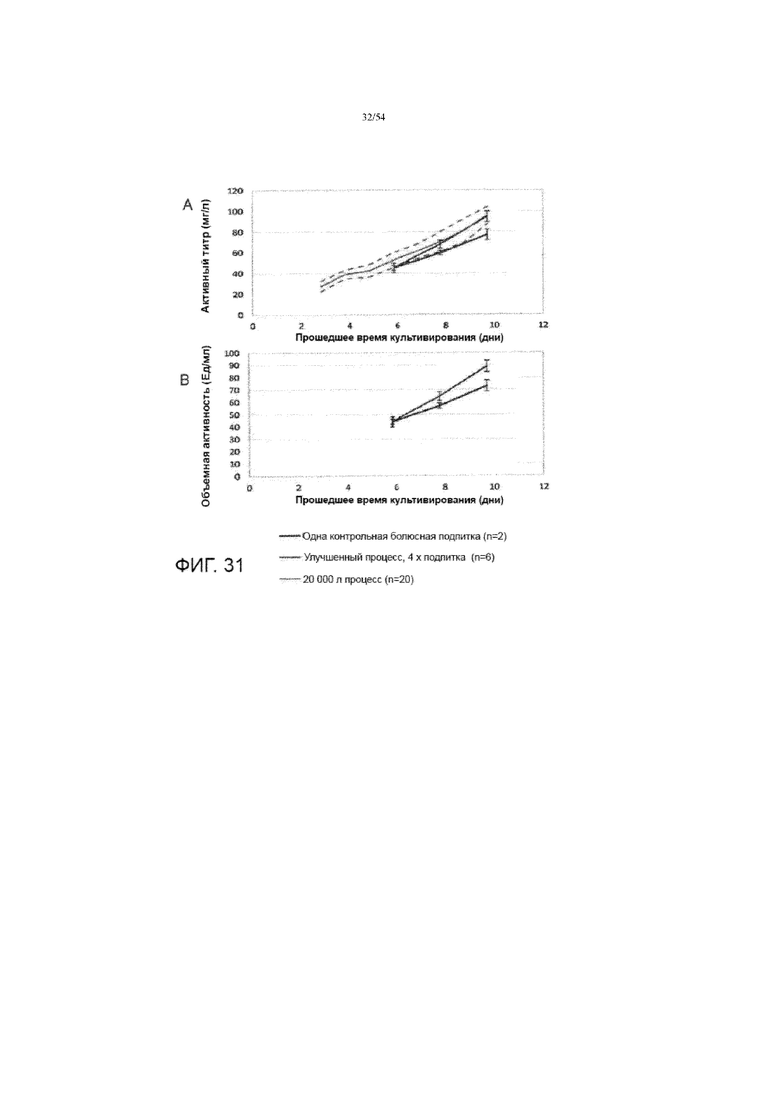

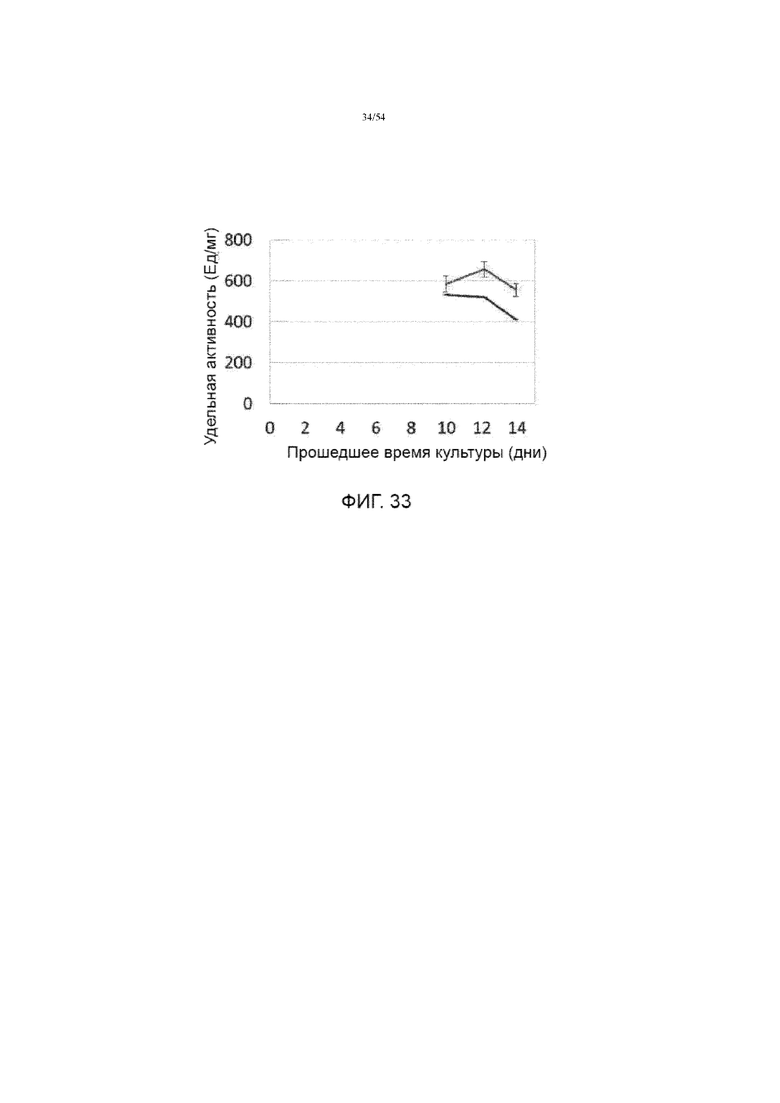
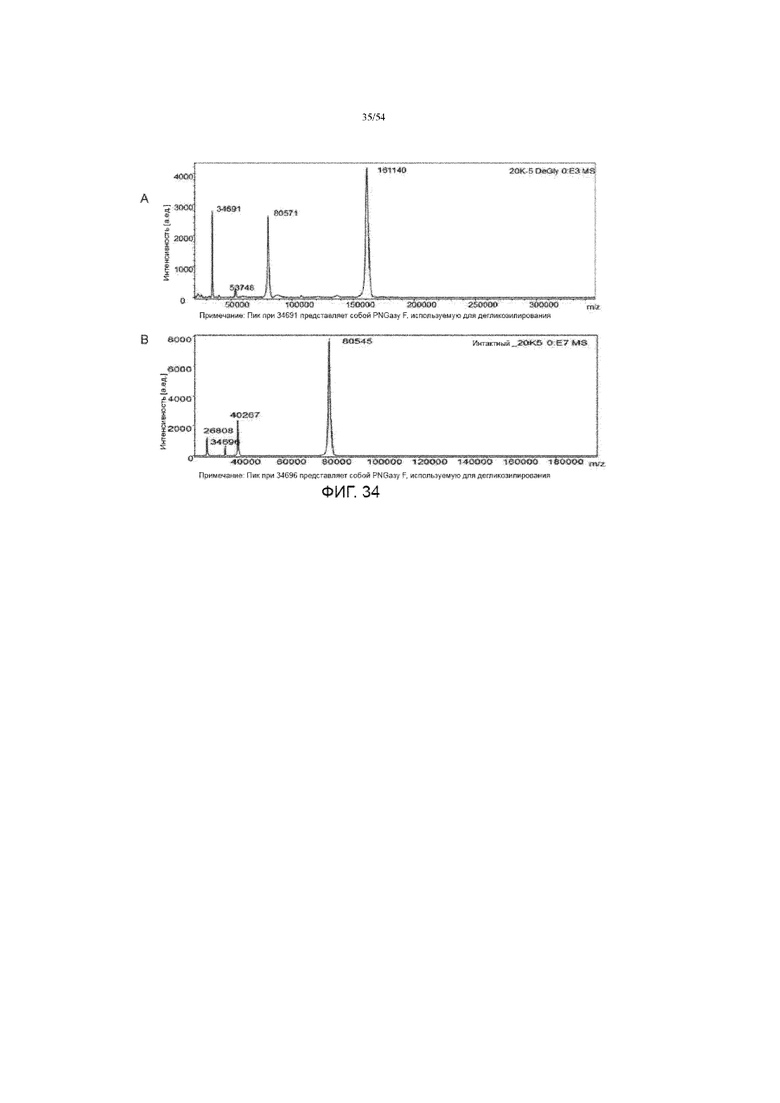
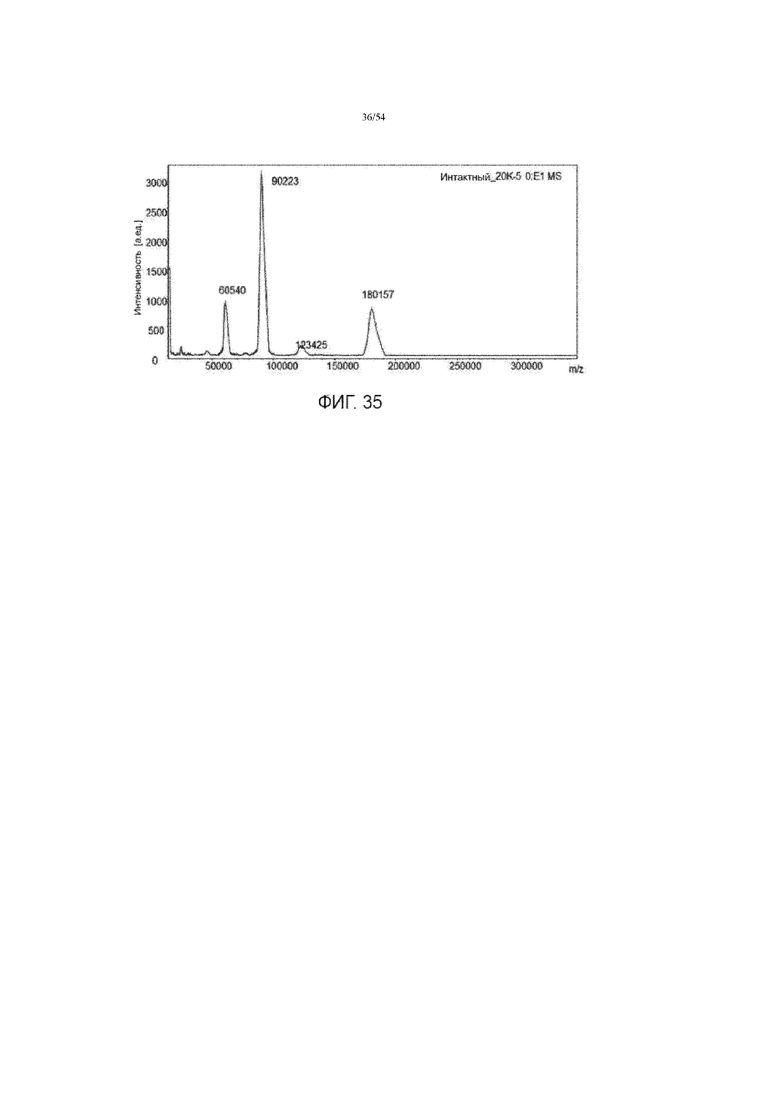
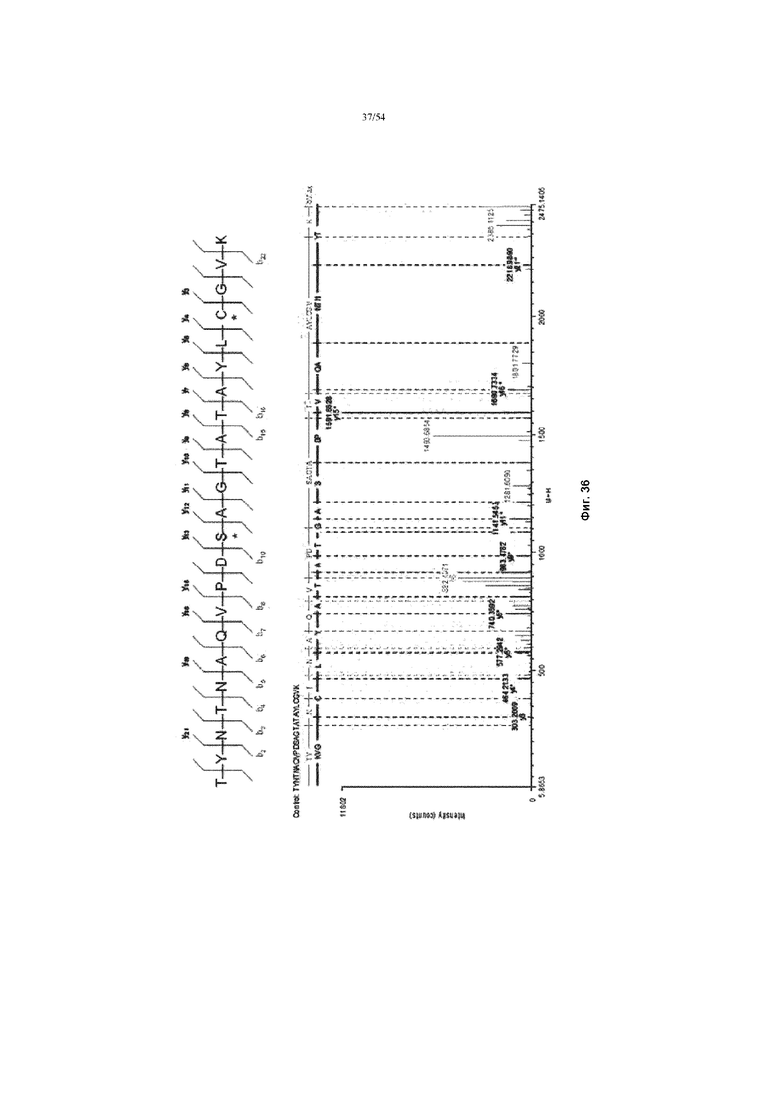
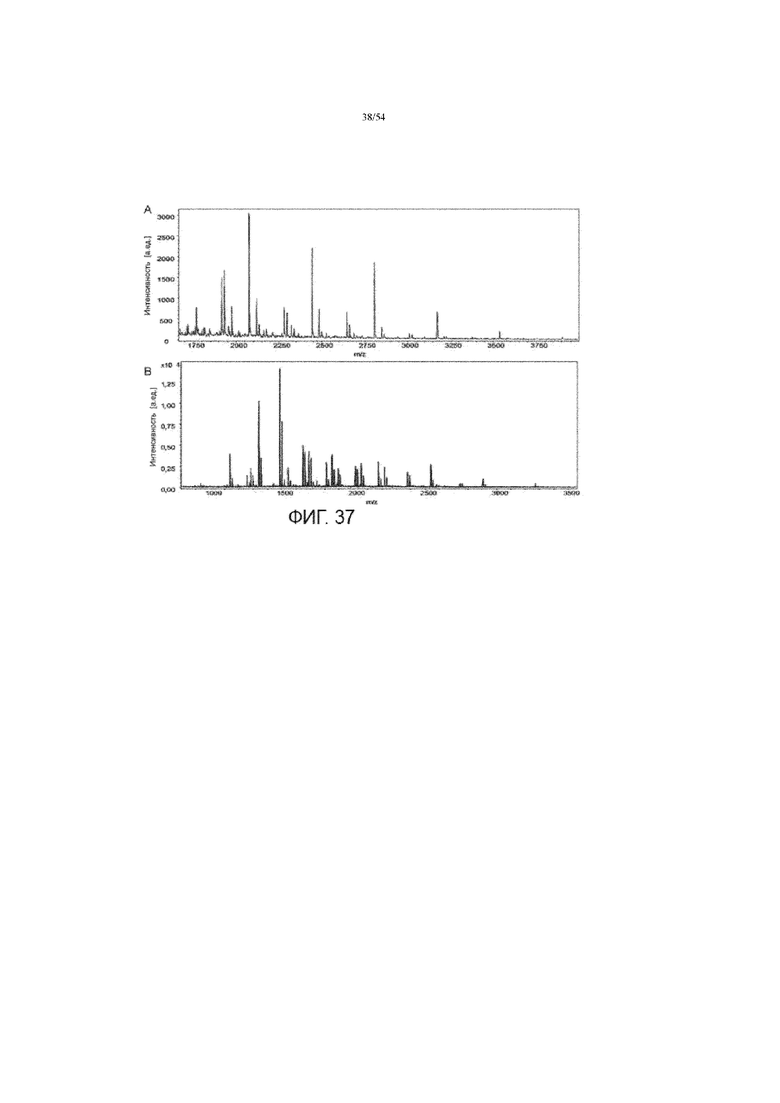

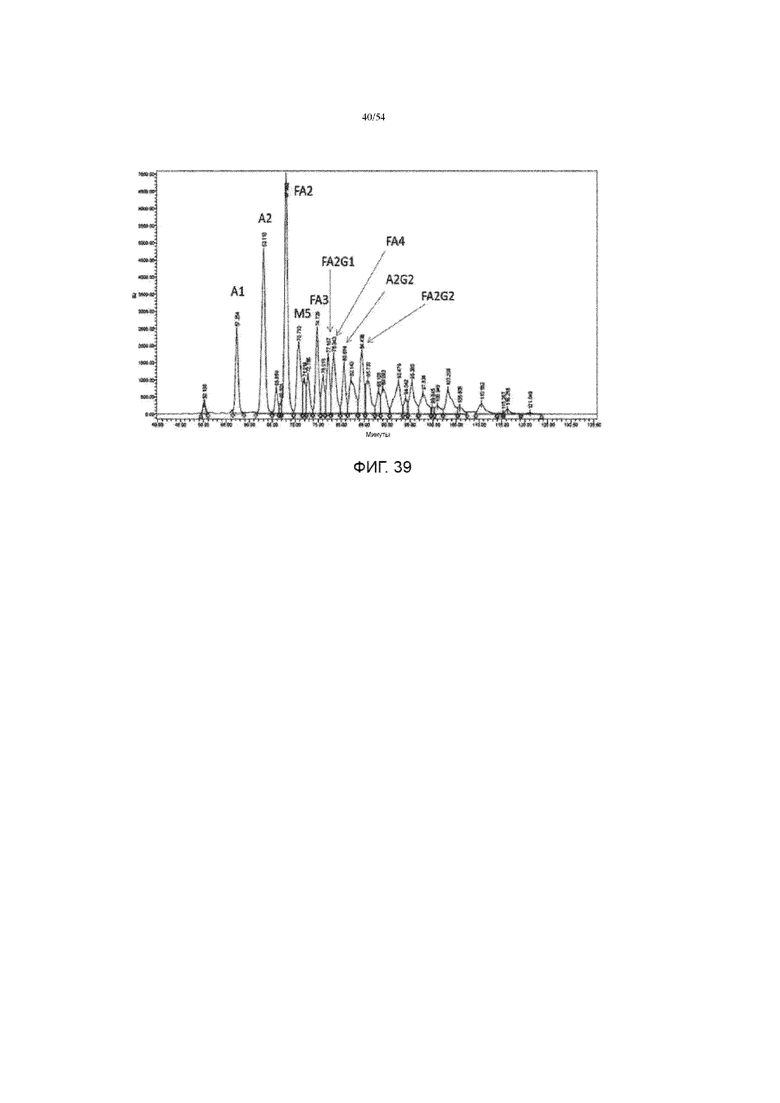
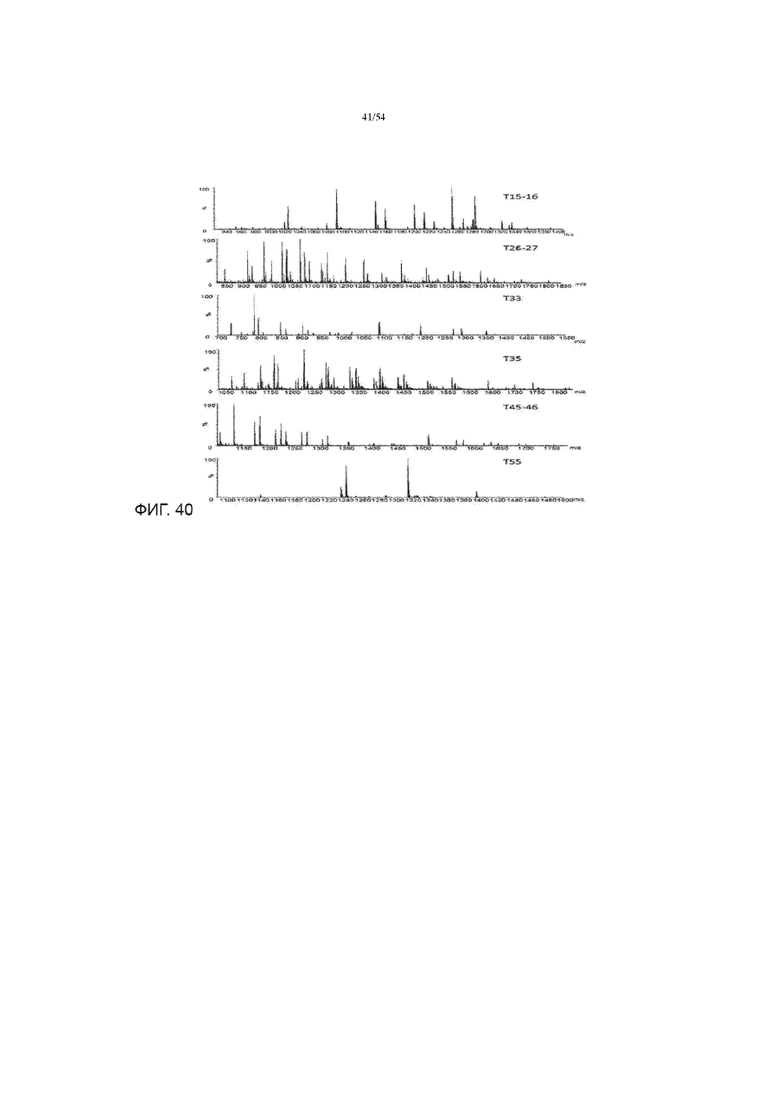


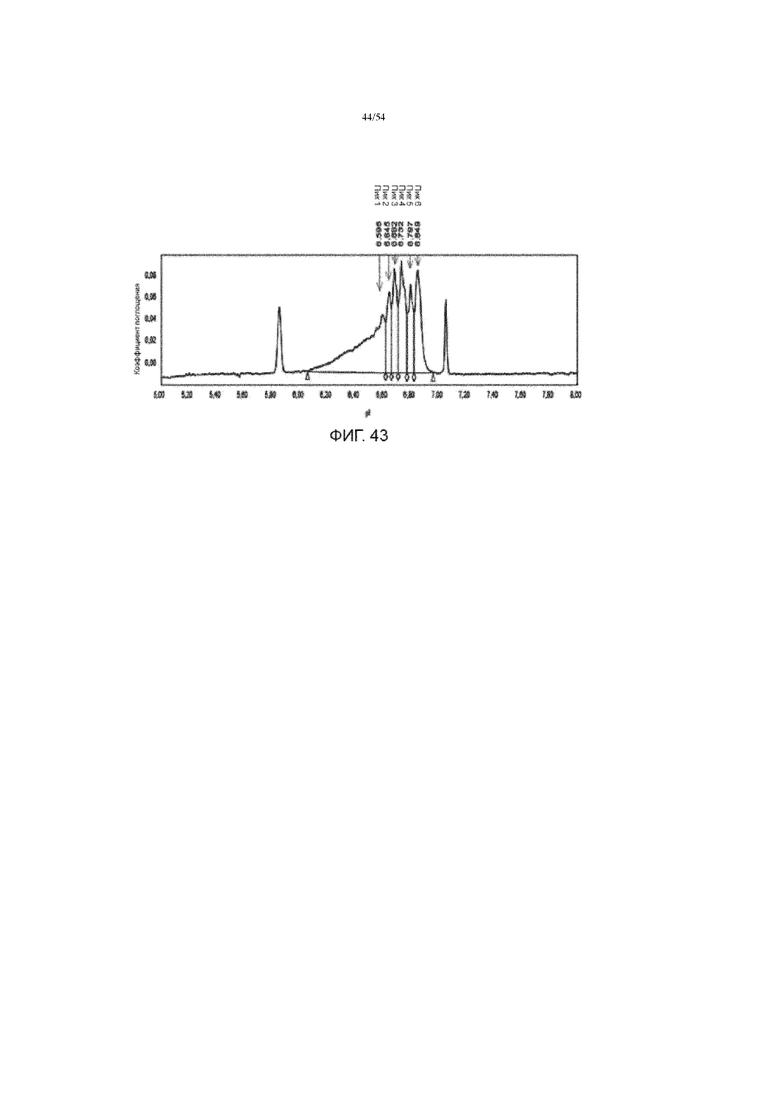
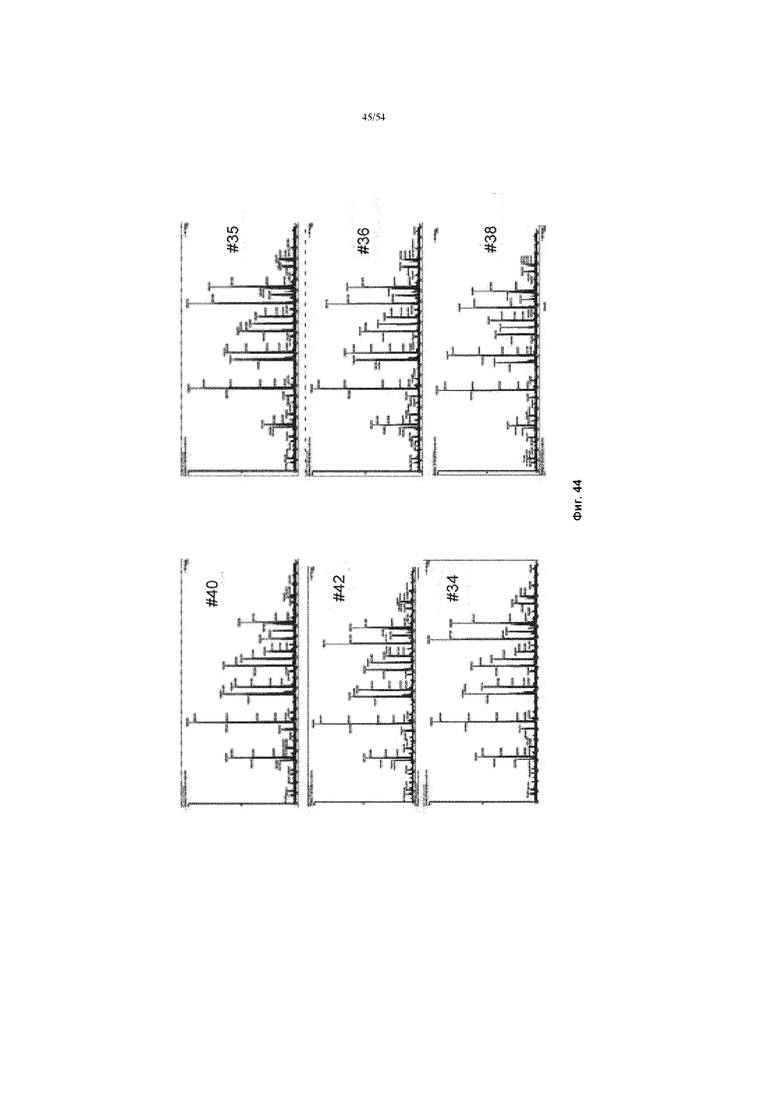

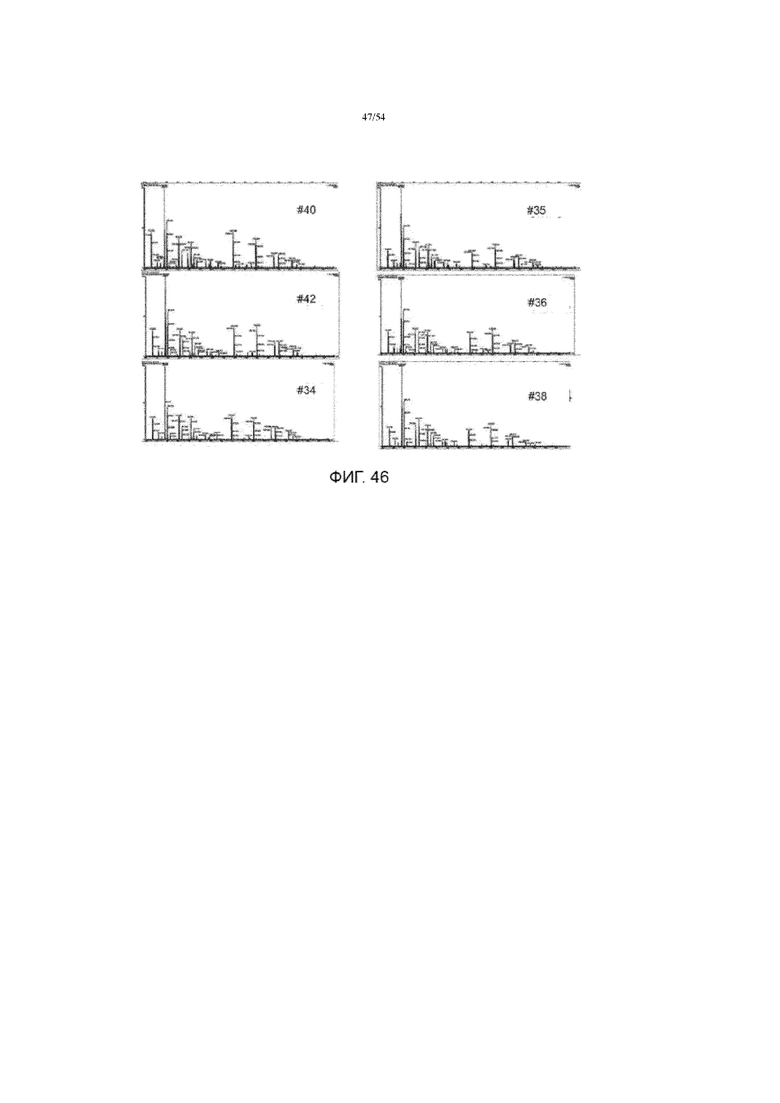




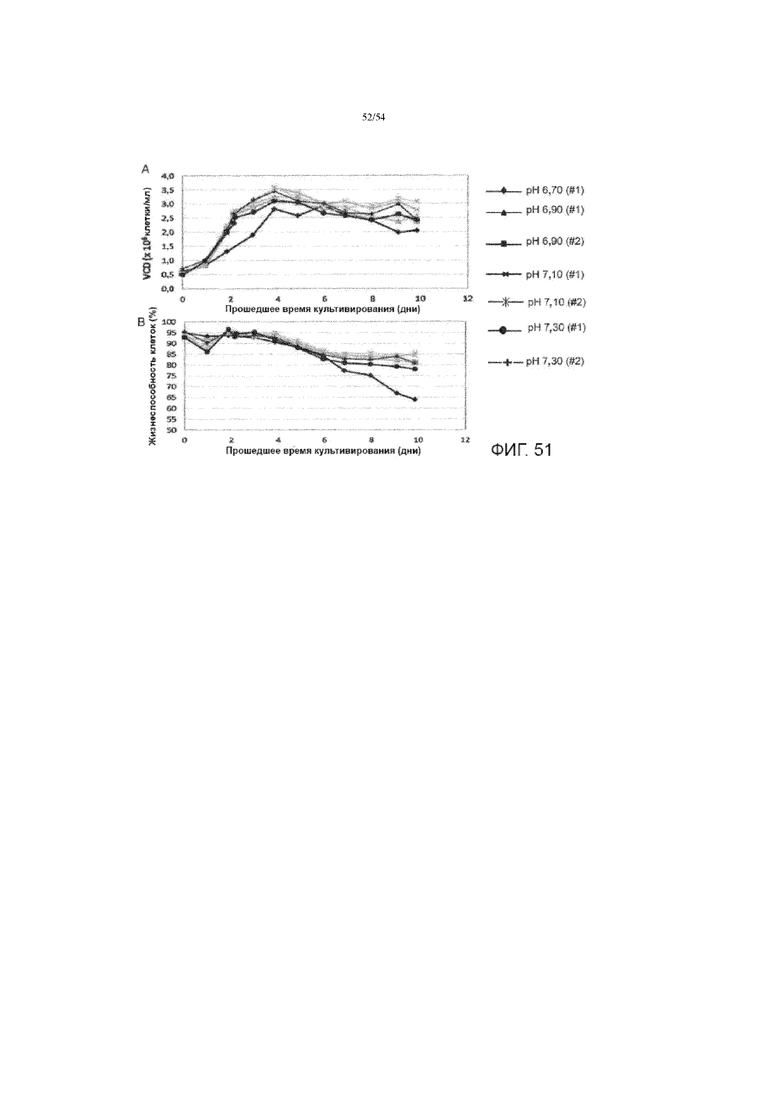
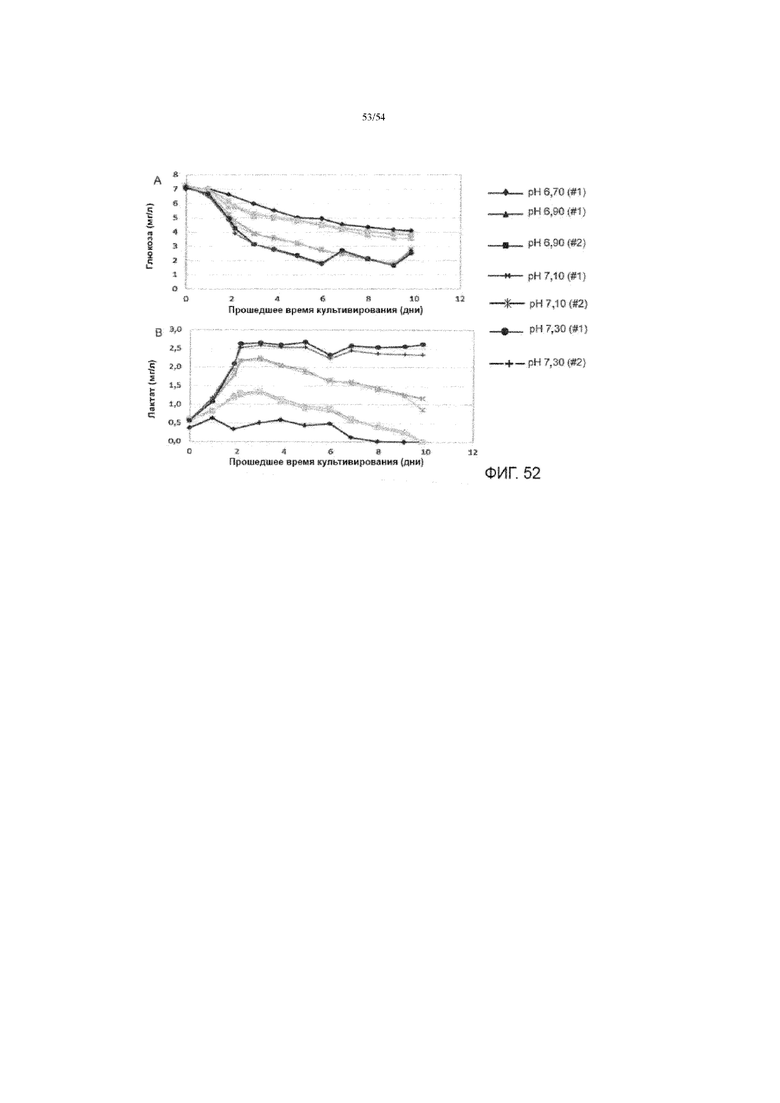
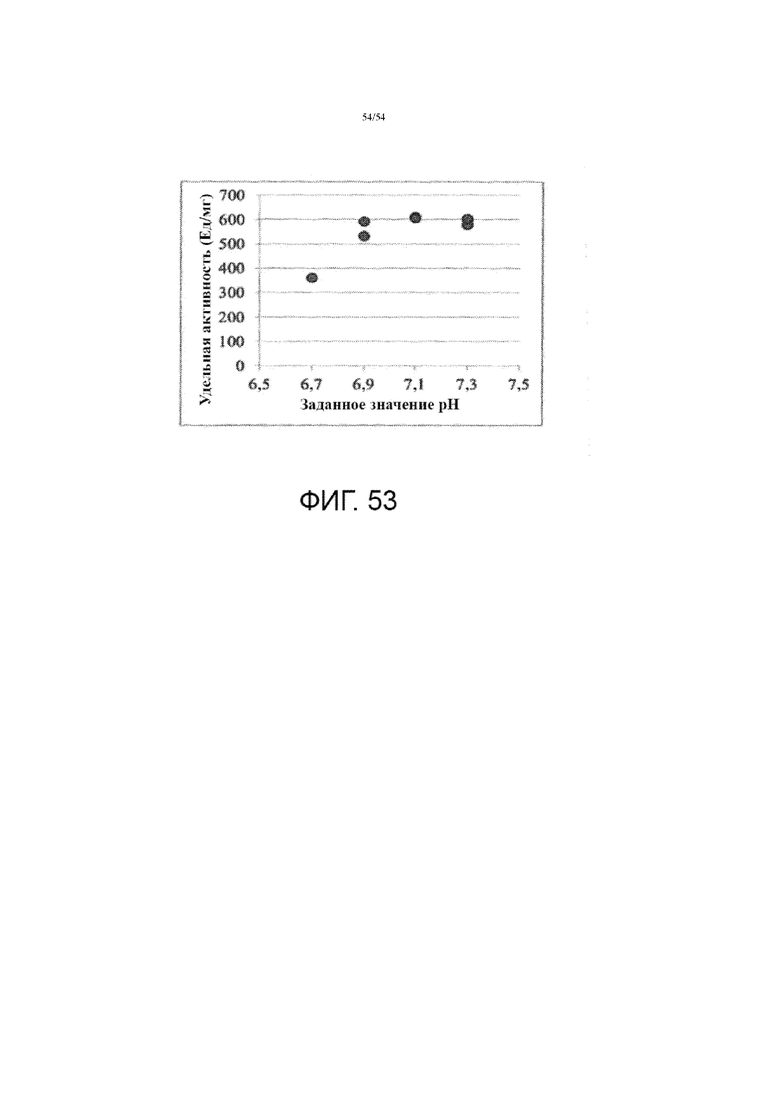
Изобретение относится к биотехнологии. Описан рекомбинантный полипептид, полученный в культуре клеток млекопитающих, содержащий асфотазу альфа (SEQ ID NO: 1) и имеющий среднее значение общего содержания сиаловой кислоты 0,9-3,5 моль сиаловой кислоты/моль мономера белка и еще одну характеристику, выбранную из группы, включающей: изоэлектрическую фокусировку между 5,2 и 6,7, крупную гликановую структуру, как показано на фиг. 41 или фиг. 42, профиль хроматограммы 2-AB-меченого олигосахарида, как показано на фиг. 38 или фиг. 39, MALDI-ToF фингерпринтный профиль гликопептида, показанный на фиг. 40 или фиг. 44-49, и другие. Также приведена фармацевтическая композиция для лечения субъекта, страдающего от состояния, связанного с дефицитом щелочной фосфатазы, содержащая указанный рекомбинантный полипептид. Изобретение позволяет получить полипептид с высокой степенью эффективности. 2 н. и 23 з.п. ф-лы, 53 ил., 33 табл., 10 пр.
1. Рекомбинантный полипептид, полученный в культуре клеток млекопитающих, причем рекомбинантный полипептид содержит асфотазу альфа (SEQ ID NO: 1) и имеет среднее значение общего содержания сиаловой кислоты (TSAC) между приблизительно 0,9 до приблизительно 3,5 моль сиаловой кислоты/моль мономера белка и по меньшей мере одну характеристику, выбранную из группы, включающей:
a) изоэлектрическую фокусировку (IEF) между приблизительно 5,2 и приблизительно 6,7;
b) крупную гликановую структуру, как показано на фиг. 41 или фиг. 42;
c) профиль хроматограммы 2-AB-меченого олигосахарида, как показано на фиг. 38 или фиг. 39;
d) MALDI-ToF фингерпринтный профиль гликопептида, показанный на фиг. 40 или фиг. 44-49;
e) основную полосу на SDS-PAGE в восстанавливающих условиях, имеющую молекулярную массу приблизительно 88-108 кДа и не менее чем приблизительно 85% от общего количества полученного рекомбинантного полипептида;
f) основную полосу на SDS-PAGE в невосстанавливающих условиях, имеющую молекулярную массу приблизительно 194 кДа до приблизительно 273 кДа и не менее чем приблизительно 85% от общего количества полученного рекомбинантного полипептида;
g) не менее чем приблизительно 95,0% для димеров рекомбинантного полипептида и не более чем приблизительно 5,0% для агрегатов, как определено с помощью высокоэффективной жидкостной эксклюзионной хроматографии (ВЭЖХ);
h) чистоту не менее чем приблизительно 95,0%, как определено с помощью высокоэффективной жидкостной хроматографии с обращенной фазой (RP-HPLC);
i) не менее чем приблизительно 90,0% для главного пика, не более чем приблизительно 6,0% для кислотных пиков и не более чем приблизительно 4,0% для основных пиков, как определено с помощью анион-обменной хроматографии (AEX);
j) процент связывания гидроксиапатита (НА) от приблизительно 75 до приблизительно 125%;
k) специфическую активность продукта (pNPP) от приблизительно 620 до приблизительно 1250 единиц/мг;
l) Km от приблизительно 13 до приблизительно 69 мкМ в анализе гидролиза неорганического пирофосфата (PPi);
m) Kcat от приблизительно 65 до приблизительно 165 с-1 в анализе гидролиза неорганического пирофосфата (PPi);
n) диапазон pI от приблизительно 6,45 до приблизительно 6,95 для всех пиков при капиллярном электрофорезе;
o) пики на масс-спектре MALDI-TOF, как показано на фиг. 34A, после дегликозилирования;
p) пики на масс-спектре MALDI-TOF, как показано на фиг. 34B, после восстановления и дегликозилирования;
q) пики на масс-спектре MALDI-TоF, как показано на фиг. 35;
r) профиль фосфорилирования, как показано на фиг. 36;
s) профиль сиалированного гликана на отрицательном масс-спектре MALDI-TоF, как показано на фиг. 37A;
t) профиль нейтральных гликанов на положительном масс-спектре MALDI-TоF, как показано на фиг. 37B;
u) молярное отношение магния на моль рекомбинантного полипептида менее чем приблизительно 0,15;
v) молярное отношение кальция на моль рекомбинантного полипептида от приблизительно 0,5 до приблизительно 1,5; и
w) молярное отношение цинка на моль рекомбинантного полипептида от приблизительно 0,5 до приблизительно 3,0.
2. Рекомбинантный полипептид по п. 1, имеющий от приблизительно 0,7 до приблизительно 1,19 свободного цистеина на половину молекулы.
3. Рекомбинантный полипептид по п. 1 или 2, имеющий фосфорилирование на Ser 93 в процентах от приблизительно 13,5 до приблизительно 35,7%.
4. Рекомбинантный полипептид по любому из пп. 1-3, имеющий не менее чем приблизительно 90,0% главного пика, не более чем приблизительно 6,0% для кислотных пиков и не более чем приблизительно 4,0% для основных пиков на хроматограмме AEX.
5. Рекомбинантный полипептид по п. 4, имеющий главный пик не менее чем приблизительно 93,7%, не более чем приблизительно 4,9% для кислотных пиков и не более чем приблизительно 3,4% для основных пиков на хроматограмме AEX.
6. Рекомбинантный полипептид по п. 1, имеющий среднее TSAC от приблизительно 1,2 до приблизительно 3,0 моль сиаловой кислоты/моль на мономер.
7. Рекомбинантный полипептид по п. 6, имеющий среднее значение TSAC от приблизительно 1,9 до приблизительно 2,7 моль сиаловой кислоты/моль на мономер.
8. Рекомбинантный полипептид по п. 7, имеющий среднее значение TSAC от приблизительно 1,85 до приблизительно 2,28 моль сиаловой кислоты/моль на мономер.
9. Рекомбинантный полипептид по любому из пп. 1-8, имеющий молярное отношение магния от приблизительно 0,03 до приблизительно 0,15.
10. Рекомбинантный полипептид по п. 9, имеющий молярное отношение магния от приблизительно 0,05 до приблизительно 0,10.
11. Рекомбинантный полипептид по п. 9, имеющий молярное отношение магния приблизительно 0,12.
12. Рекомбинантный полипептид по любому из пп. 1-11, содержащий по меньшей мере приблизительно 95,0% димеров рекомбинантного полипептида и приблизительно 5,0% или менее полипептидных агрегатов, как определено с помощью эксклюзионной ВЭЖХ.
13. Рекомбинантный полипептид по п. 12, содержащий по меньшей мере приблизительно 96,8% димеров рекомбинантного полипептида и приблизительно 3,2% или менее полипептидных агрегатов, в частности, как определено методом эксклюзионной ВЭЖХ.
14. Рекомбинантный полипептид по п. 13, содержащий по меньшей мере приблизительно 97,6% димеров рекомбинантного полипептида и приблизительно 2,4% или менее агрегатов, в частности, как определено методом эксклюзионной ВЭЖХ.
15. Рекомбинантный полипептид по любому из пп. 1-14, имеющий по меньшей мере приблизительно 95,0% чистоту, в частности, как определено с помощью RP-HPLC.
16. Рекомбинантный полипептид по п. 15, имеющий по меньшей мере приблизительно 97,6% чистоту, как определено с помощью RP-HPLC.
17. Рекомбинантный полипептид по любому из пп. 1-16, имеющий процент связывания гидроксиапатита (НА) от приблизительно 75 до приблизительно 125%.
18. Рекомбинантный полипептид по п. 17, имеющий средний процент связывания гидроксиапатита от приблизительно 85 до приблизительно 97%.
19. Рекомбинантный полипептид по п. 18, имеющий средний процент связывания гидроксиапатита от приблизительно 90 до приблизительно 91%.
20. Рекомбинантный полипептид по любому из пп. 1-19, имеющий специфическую активность (pNPP) от приблизительно 620 до приблизительно 1250 единиц/мг.
21. Рекомбинантный полипептид по п. 20, имеющий среднюю специфическую активность (pNPP) от приблизительно 904,0 до приблизительно 907,7 Ед/мг.
22. Рекомбинантный полипептид по любому из пп. 1-21, где указанный рекомбинантный полипептид кодируется полинуклеотидом, кодирующим полипептид, содержащий последовательность SEQ ID NO: 1 или последовательность, полностью комплементарную SEQ ID NO: 1.
23. Рекомбинантный полипептид по любому из пп. 1-22, где рекомбинантный полипептид получают в периодически подпитываемом реакторе, имеющем объем от 200 до 20000 л.
24. Рекомбинантный полипептид по п. 23, где подпитываемый периодический реактор имеет объем от 2000 до 20000 л.
25. Фармацевтическая композиция для лечения субъекта, страдающего от состояния, связанного с дефицитом щелочной фосфатазы, содержащая рекомбинантный полипептид, как указывается по любому из пп. 1-24, и по меньшей мере один фармацевтически приемлемый носитель, разбавитель или эксципиент.
| EP 1985697 A1, 29.10.2008 | |||
| WO 2015112017 A1, 30.07.2015 | |||
| WO 2008138131 A1, 20.11.2008 | |||
| US 7943126 B2, 17.05.2011 | |||
| РОДИОНОВА С.С | |||
| Гипофосфатазия у взрослых: клинические случаи и обзор литературы, Остеопороз и остеопатии, 2015, N 2, С.25-27. |

