Родственные заявки
[0001] Эта заявка подана 15 января 2020 г. как международная РСТ-заявка на патент, и по ней испрашивается приоритет временной заявки на патент США с № 62/794452, поданной 18 января 2019 г., и временной заявки на патент США с № 62/869248, поданной 1 июля 2019 г., описания которых в их полном объеме включены сюда посредством ссылки.
Область техники
[0002] Здесь раскрывается композиция для приема внутрь, содержащая сфинган, и ее применение в качестве пребиотика.
Предпосылки к созданию изобретения
[0003] Желудочно-кишечный тракт человека является очень сложной микробной экосистемой, которая является, как было установлено, удивительно стабильной (Zoetendal (1998)) Множество различных подходов использовалось для модуляции кишечной флоры таким образом, чтобы это было полезно для здоровья хозяина (смотрите, например, Bielecka (2002) и Steer (2000)). Эти различные подходы включают добавление живых микроорганизмов в пищу (пробиотиков), добавление пищевых ингредиентов или пищевых волокон для избирательной стимуляции полезных бактерий в организме хозяина (пребиотиков), и добавление комбинации как пробиотиков, так и пребиотиков (синбиотиков).
[0004] Пребиотики представляют собой неперевариваемые субстраты, которые избирательно используются микроорганизмами хозяина, приносящими пользу для здоровья (Gibson (2017)). Эффекты пребиотиков в кишечнике можно оценить на основе роста полезных для здоровья бактерий, таких как лактобациллы и бифидобактерии, уменьшения количества кишечных патогенов и повышения или снижения продукции метаболитов бактерий, связанных со здоровьем. Пребиотическая/бифидогенная природа выбранных пребиотиков (таких как инулин, фруктоолигосахариды, галактоолигосахариды, лактулоза и арабиноолигосахарид) была предположена и/или подтверждена в предыдущих исследованиях (смотрите, например, Guimaraes (2018), Karltohn-Senaye (2013), Patel (2013), Saavedra (2002), Tuohy (2001), Tuohy (2002), US8313789B2, US20100092440A1 и WO2004002240A2.)
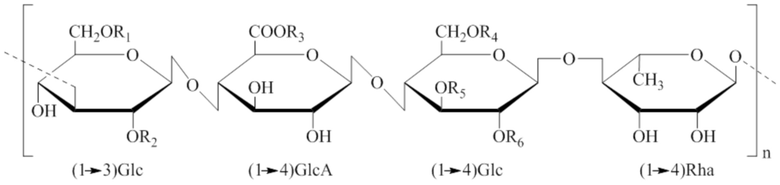
[0005] Как правило, сфинганы представляют собой полисахариды, состоящие из следующего замещенного или незамещенного тетрамерного сахарида, обычно обозначаемого как [(→3)Glc(β1→4)GlcA(β1→4)Glc (β1→4)Rha(α1→)]n. Известные сфинганы включают, например, геллан (S-60), велан (S-130), рамсан (S-194) и диутан (S-657)).
[0006] Геллан (геллановая камедь или S-60) продуцируется штаммами вида Sphingomonas elodea (раньше Pseudomonas elodea), например штаммом ATCC 31461. (Смотрите, например, Morrison (2016), Sworn (2009) и US4326053A.) Обычные формы геллановой камеди включают высокоацильные (также известные под именем «природные»), неосветленные (например, геллан KELCOGEL® LT100), низкоацильные, неосветленные (например, геллан KELCOGEL® LT) и низкоацильные, осветленные (например, геллановые камеди KELCOGEL® и KELCOGEL® F). (Sworn (2009).) Также доступен ряд специальных сортов, например, высокоацильный, не содержащий PHB (полигидроксиоксибутират), осветленный (например, геллан KELGOGEL® HT) и низкоацильный, осветленный (дважды преципитированный) (например, геллан GELRITETM MK). Природная, или высокоацильная, форма геллана включает два ацильных заместителя (ацетат в положении O6 и глицерат в положении O2) в (1→3)Glc-единице, и в среднем один глицерат приходится на тетрамер, и один ацетат - на два тетрамера. (Kuo (1986).) В низкоацильном геллане глицерат и ацетат отсутствуют. Геллановые камеди также могут продуцироваться со средним содержанием глицерата и ацетата. Коммерческим продуктом с пониженным содержанием глицерата и ацетата является геллан KELCOGEL® DGA.
[0007] Геллановая камедь, как правило, действует как гелеобразующий или суспендирующий агент в некоторых продуктах для приема внутрь и присутствует на уровнях, которые колеблются в пределах от 0,02 до 0,5% (в отношении веса к объему) (Смотрите, например, Fallourd (2009), Morrison (2016), Sworn (2009), US6602996B1, US6663911B2, US5342626A, US8513408B2, and US20080008814A1). До ее одобрения в качестве пищевой добавки, в исследованиях оценивалась безопасность геллановой камеди при введении крысам и людям. (Смотрите, например, Anderson (1988) and Edwards (1995); смотрите также Anderson (1990).) Например, Edwards (1995) описывает кормление крыс линии Wistar в течение 28 дней с использованием диеты, которая включала 50 г/кг/день геллановой камеди. (Для справки, 50 г/кг у крыс соответствует эквивалентному для человека количеству, составляющему приблизительно 8 г/кг (смотрите, например, FDA Guidance (2005).) Интересно, что Edwards (1995) пришел к выводу, что геллановая камедь не оказывает устойчивый эффект на короткоцепочечные жирные кислоты (SCFA, такие как ацетат, пропионат и бутират) в слепой кишке или кале. Кроме того, Anderson (1988) приводит исследование, в котором добровольцы-люди принимали внутрь некоторое количество геллановой камеди в соответствии со схемой приема постоянной дозы, составляющей 175 мг/кг/день, в течение 7 дней, а затем 200 мг/кг/день в течение еще 16 дней (Для веса человека в диапазоне от 60 до 75 кг, 175 мг/кг соответствует диапазону от 10,5 до 13 г, тогда как 200 мг/кг соответствует диапазону от 12 г до 15 г.) На основе результатов, представленных там, Anderson (1988) пришел к выводу, что прием геллановой камеди не вызывает вредных диетических или физиологических эффектов. Кроме того, Anderson (1988) пришел к выводу, что геллановая камедь проявляет эффект увеличения каловых масс. В соответствии с эффектом увеличения каловых масс, наблюдаемым Anderson (1988), последующее исследование показало, что геллановая камедь уменьшает диарею у кошек. (US9028861B2.) Со ссылкой на Tetsuguchi (1997), Li (2019) упоминает, без объяснения или доказательства, что геллан в виде олигосахарида по сообщениям оказывает пребиотические эффекты в кишечнике, хотя Tetsuguchi (1997) явно не оценивал пребиотические эффекты геллана в виде олигосахарида в кишечнике. На сегодняшний день нет исследований, которые окончательно продемонстрировали бы, действует ли геллановая камедь или олигосахарид, полученный из геллановой камеди, в качестве пребиотика.
[0008] Велан (велановая камедь или S-130) продуцируется Sphingomonas sp. (например, ATCC 31555). (US4342866A и US5175277A.) Приблизительно две трети (1→4)Glc-единиц велана замещены в положении O3 α-L-рамнопиранозильной группой (т.е. Rha(α1→)), тогда как остальные (1→4)Glc-единицы велана замещены α-L-маннопиранозильной группой (т.е. Man(α1→)). (Stankowski (1992).) Кроме того, (1→3)Glc-единица велана может быть замещена в положении O2 ацетилом. (Stankowski (1992).)
[0009] Рамсан (рамсановая камедь или S-194) продуцируется Sphingomonas sp. (например, ATCC 31961). (US4401760A.) Рамсан замещен в положении O6 (1→3)Glc-единицы на D-Glc(β1→6)-D-Glc(α1→). (Jansson (1986).) Рамсан содержит одну O-ацетильную группу на повторяющуюся единицу, с распределением по вторичным положениям. (Jansson (1986).)
[0010] Диутан (диутановая камедь или S-657) продуцируется Sphingomonas sp. (например, ATCC 53159). (US5175278A и US20130189748A1.) (1→4)Glc-единица диутана замещена в положении O3 на Rha(α1→4)-Rha(α1→), в положении O6 на ацетил и в различной степени в положениях O2 и/или O6 (1→3)Glc-единицы на ацетил. (Diltz (2001).)
Краткое изложение сущности настоящего изобретения
[0011] Здесь раскрывается композиция для приема внутрь, содержащая сфинган, и ее применение в качестве пребиотика.
Сокращения
[0012] Следующий текст включает многочисленные сокращенные термины. Сокращения для выбранных терминов, раскрытых здесь, указаны ниже.
[0013] A: донор A (женщина, 28 лет)
[0014] Ac: ацетат
[0015] B: донор B (женщина, 41 год)
[0016] b-SCFA: жирные кислоты с разветвленными короткими цепями (например, изобутират, изовалерат и изокапроат)
[0017] C: донор C (женщина, 34 года)
[0018] C1: контрольный период 1
[0019] C2: контрольный период 2
[0020] CON(ave): средняя концентрация для контрольных периодов 1 и 2
[0021] CD: болезнь Крона
[0022] DC: дистальный отдел ободочной кишки в качестве ферментера
[0023] DP: степень полимеризации
[0024] Glc: D-глюкопиранозил
[0025] GlcA: D-глюкопиранозилуроновая кислота
[0026] Glyc: L-глицерат
[0027] GPRs: связанный с G-белком рецептор
[0028] HA: высокоацильный
[0029] HA/LA: высокоацильный или низкоацильный
[0030] IBD: воспалительные заболевания кишечника
[0031] IBS: синдром раздраженного кишечника
[0032] IFN: интерферон
[0033] IL: интерлейкин
[0034] LA: низкоацильный
[0035] LCSs: длинноцепочечные сфинганы
[0036] LPS: липополисахарид
[0037] MAMP: ассоциированные с микроорганизмом молекулярные структуры
[0038] Man: L-маннопиранозил
[0039] мМ: миллимолярный (т.е. миллимоль на литр)
[0040] MN: среднечисловая молекулярная масса
[0041] MW: средневзвешенная молекулярная масса
[0042] NaB: бутират натрия
[0043] OTU: операционная таксономическая единица
[0044] PHB: полигидроксиоксибутират
[0045] PC: проксимальный отдел ободочной кишки
[0046] PRR: паттерн-распознающие рецепторы
[0047] Rha: L-рамнопиранозил
[0048] ROS: активные формы кислорода
[0049] SCFA: короткоцепочечные жирные кислоты (например, ацетат, пропионат и бутират)
[0050] SHIME: имитатор микробной экосистемы в кишечнике человека
[0051] SOS: сфинган в виде олигосахарида
[0052] SPS: сфинган в виде полисахарида
[0053] TEER: трансэпителиальное электрическое сопротивление
[0054] Тетрамер: [Glc(β1→4)GlcA(β1→4)Glc(β1→4)Rha], Glc, GlcA,Glc, Rha или Glc2,GlcA, Rha
[0055] Октамер: [Glc(β1→4)GlcA(β1→4)Glc(β1→4)Rha]2 Glc, GlcA,Glc, Rha,Glc, GlcA,Glc, Rha или Glc4,GlcA2,Rha2
[0056] SEC: гель-фильтрация
[0057] TGF: трансформирующий фактор роста
[0058] TLR: toll-подобный рецептор
[0059] TNF: фактор некроза опухолей
[0060] TR1: период обработки 1
[0061] TR2: период обработки 2
[0062] TR3: период обработки 3
[0063] TRT(ave): Средняя концентрация для периодов обработки 1, 2 и 3
[0064] UC: язвенный колит
Краткое описание чертежей
[0065] Фиг. 1a. Полученная в ходе гель-фильтрации хроматограмма для обработанных кислотой (SN9, сплошная линия) и ферментом (SN18, пунктирная линия) сфинганов в виде поли- и олигосахаридов, полученных из высокоацильного геллана, показывающая время элюирования стандартов молекулярной массы пуллуланов (а именно, >50 кДа (6,5 мин, закрашенный квадрат (  )), 12 кДа (8,8 мин, закрашенный кружок (
)), 12 кДа (8,8 мин, закрашенный кружок (  )), 5 кДа (9,3 мин, закрашенный треугольник (
)), 5 кДа (9,3 мин, закрашенный треугольник (  )), 1 кДа (10 мин, пустой квадрат (
)), 1 кДа (10 мин, пустой квадрат (  )), 342 Да (10,65 мин, пустой кружок (
)), 342 Да (10,65 мин, пустой кружок (  )), и 180 Да (11,15 мин, пустой треугольник (
)), и 180 Да (11,15 мин, пустой треугольник (  ))).
))).
[0066] Фиг. 1b. Полученная в ходе гель-фильтрации хроматограмма для обработанных кислотой (SN10, сплошная линия) и ферментом (SN17, пунктирная линия) сфинганов в виде поли- и олигосахаридов, полученных из низкоацильного геллана, показывающая время элюирования стандартов молекулярной массы пуллуланов (а именно, >50 кДа (6,5 мин, закрашенный квадрат ( )), 12 кДа (8,8 мин, закрашенный кружок ( )), 5 кДа (9,3 мин, закрашенный треугольник ( )), 1 кДа (10 мин, пустой квадрат ( )), 342 Да (10,65 мин, пустой кружок ( )), и 180 Да (11,15 мин, пустой треугольник ( ))).
[0067] Фиг. 2a. Средняя продукция ацетата (мМ) за контрольный период (CON(ave.), n=6) и период обработки (TRT(ave), n=9) для ферментера в качестве проксимального отдела ободочной кишки (PC) для трех разных доноров (A, B и C), где * указывает на статистически значимые различия по сравнению с предыдущим периодом, тогда как разные буквы указывают на статистическое различие между разными обработками; p<0,05.
[0068] Фиг. 2b. Средняя продукция ацетата (мМ) за контрольный период (CON(ave.), n=6) и период обработки (TRT(ave), n=9) для ферментера в качестве дистального отдела ободочной кишки (DC) для трех разных доноров (A, B и C), где * указывает на статистически значимые различия по сравнению с предыдущим периодом, тогда как разные буквы указывают на статистическое различие между разными обработками; p<0,05.
[0069] Фиг. 3a. Средняя продукция пропионата (мМ) в ферментере в качестве проксимального отдела ободочной кишки (PC) за контрольный период (CON(ave), n=6) и период обработки (TRT(ave), n=9) для трех разных доноров (A, B и C), где * указывает на статистически значимые различия по сравнению с предыдущим периодом, тогда как разные буквы указывают на статистическое различие между разными обработками; p<0,05.
[0070] Фиг. 3b. Средняя продукция пропионата (мМ) в ферментере в качестве дистального отдела ободочной кишки (DC) за контрольный период (CON(ave), n=6) и период обработки (TRT(ave), n=9) для трех разных доноров (A, B и C), где * указывает на статистически значимые различия по сравнению с предыдущим периодом, тогда как разные буквы указывают на статистическое различие между разными обработками; p<0,05.
[0071] Фиг. 4a. Средняя продукция бутирата (мМ) в ферментере в качестве проксимального отдела ободочной кишки (PC) за контрольный период (CON(ave), n=6) и период обработки (TRT(ave), n=9) для трех разных доноров (A, B и C), где * указывает на статистически значимые различия по сравнению с предыдущим периодом, тогда как разные буквы указывают на статистическое различие между разными обработками; p<0,05.
[0072] Фиг. 4b. Средняя продукция бутирата (мМ) в ферментере в качестве дистального отдела ободочной кишки (DC) за контрольный период (CON(ave), n=6) и период обработки (TRT(ave), n=9) для трех разных доноров (A, B и C), где * указывает на статистически значимые различия по сравнению с предыдущим периодом, тогда как разные буквы указывают на статистическое различие между разными обработками; p<0,05.
[0073] Фиг. 5a. Средняя продукция лактата (мМ) в ферментере в качестве проксимального отдела ободочной кишки (PC) за контрольный период (CON(ave), n=6) и период обработки (n=9) для трех разных доноров (A, B и C), где разные буквы указывают на статистическое различие между разными обработками; p<0,05.
[0074] Фиг. 5b. Средняя продукция лактата (мМ) в ферментере в качестве дистального отдела ободочной кишки (DC) за контрольный период (CON(ave), n=6) и период обработки (n=9) для трех разных доноров (A, B и C), где разные буквы указывают на статистическое различие между разными обработками; p<0,05.
[0075] Фиг.6а. Средняя продукция аммиака (мг/л) в ферментере в качестве проксимального отдела ободочной кишки (PC) за контрольный период (CON(ave), n=6) и период обработки (TRT(ave), n=9) для трех разных доноров (A, B и C), где разные буквы указывают на статистическое различие между разными обработками; p<0,05.
[0076] Фиг. 6b. Средняя продукция аммиака (мг/л) в ферментере в качестве дистального отдела ободочной кишки (DC) за контрольный период (CON(ave), n=6) и период обработки (TRT(ave), n=9) для трех разных доноров (A, B и C), где разные буквы указывают на статистическое различие между разными обработками; p<0,05.
[0077] Фиг.7a. Средняя продукция SCFA с разветвленной цепью (мМ) в ферментере в качестве проксимального отдела ободочной кишки (DC) за контрольный период (CON(ave), n=6) и период обработки (TRT(ave), n=9) для трех разных доноров (A, B и C), где разные буквы указывают на статистическое различие между разными обработками; p<0,05.
[0078] Фиг.7b. Средняя продукция SCFA с разветвленной цепью (мМ) в ферментере в качестве дистального отдела ободочной кишки (DC) за контрольный период (CON(ave), n=6) и период обработки (TRT(ave), n=9) для трех разных доноров (A, B и C), где * указывает на статистически значимые различия по сравнению с предыдущим периодом, а разные буквы указывают на статистическое различие между разными обработками; p<0,05.
[0079] Фиг. 8. Обратная величина индекса разнообразия Симпсона в случае просвета и слизи проксимального (PC) или дистального отдела (DC) ободочной кишки в виде ферментера SHIME в разные моменты времени во время контрольного периода (C1 и C2) и периода обработки (TR1, TR2 и TR3) геллановой камедью для трех разных доноров (n=1). Интенсивность затенения указывает на индекс полного разнообразия, нормализованный для каждого из трех разных доноров (т.е. внутри каждой строки).
[0080] Фиг. 9. Относительное содержание (%) доминирующих типов в просвете или слизи проксимального (PC) или дистального отдела (DC) ободочной кишки в виде ферментера SHIME в разные моменты времени во время контрольного периода (C1 и C2) и периода обработки (TR1, TR2 и TR3) геллановой камедью для трех разных доноров-людей (n=1). N.B. Один образец был явным отклонением и поэтому был удален из этого анализа контрольных образцов, т.е. образец слизистой оболочки в PC донора А на второй контрольной неделе (C2).
[0081] Фиг. 10. Относительное содержание (%) различных семейств, относящихся к определенным типам, в просвете проксимального отдела ободочной кишки (PC) в виде ферментеров SHIME в разные моменты времени во время контрольного периода (C1 и C2) и периода обработки (TR1, TR2 и TR3) геллановой камедью для трех разных доноров-людей (n=1). Интенсивность затенения указывает на абсолютное количество, нормализованное для каждого из разных семейств (т.е. внутри каждой строки). Интенсивность затенения указывает на абсолютное количество, нормализованное для каждого из различных семейств (т.е. в пределах каждого ряда).
[0082] Фиг. 11. Относительное содержание (%) различных семейств, относящихся к определенным типам, в просвете дистального отдела ободочной кишки (DC) в виде ферментеров SHIME в разные моменты времени во время контрольного периода (C1 и C2) и периода обработки (TR1, TR2 и TR3) геллановой камедью для трех разных доноров-людей (n=1). Интенсивность затенения указывает на абсолютное количество, нормализованное для каждого из разных семейств (т.е. в пределах каждого ряда).
[0083] Фиг. 12. Относительное содержание (%) различных семейств, относящихся к определенным типам, в слизи проксимального отдела ободочной кишки (PC) в виде ферментеров SHIME в разные моменты времени во время контрольного периода (C1 и C2) и периода обработки (TR1, TR2 и TR3) геллановой камедью для трех разных доноров-людей (n=1). Интенсивность затенения указывает на абсолютное количество, нормализованное для каждого из разных семейств (т.е. внутри каждой строки). В качестве замечания, один образец был явным отклонением и поэтому был удален из этого анализа контрольных образцов, т.е. образец слизистой оболочки PC донора А на второй контрольной неделе (C2).
[0084] Фиг. 13. Относительное содержание (%) различных семейств, относящихся к определенным типам, в слизи дистального отдела ободочной кишки (PC) в качестве ферментеров SHIME в разные моменты времени во время контрольного периода (C1 и C2) и периода обработки (TR1, TR2 и TR3) геллановой камедью для трех разных доноров-людей (n=1). Интенсивность затенения указывает на абсолютное количество, нормализованное для каждого из разных семейств (т.е. в пределах каждого ряда).
[0085] Фиг. 14. Схематическое представление сокультивирования клеток Caco-2 и THP1. Клетки Caco-2 высевают на полупроницаемую мембрану, которую помещают поверх лунок, засеянных клетками THP1. Это создает апикальный (AP) и базолатеральный (BL) компартмент. Монослой клеток Caco-2 создает барьер для макромолекул и допускает проход путем пассивного переноса небольших молекул между межклеточным пространством и активного переноса микро- и макромолекул через клеточные мембраны. Сокультивирование клеток обоих типов делает возможным косвенное взаимодействие между люминальным содержимым, которое приводят в контакт с клетками Caco-2, и периинтестинальным содержимым, которое контактирует с иммунными клетками (THP1). Кроме того, метаболиты, используемые/преобразуемые эпителиальными клетками, могут модулировать иммунный клеточный ответ, и наоборот.
[0086] Фиг. 15. Каскад передачи сигналов, активируемый при повреждении эпителиального барьера кишечника, что приводит к пролому люминальным содержимом клеточной клетки кишечника. IFN-γ: интерферон гамма; IL: интерлейкины; MCP-1: белок-хемоаттрактант моноцитов 1; ROS: активные формы кислорода; TGF-β: трансформирующим фактором роста бета; TH: Т-клетки-хелперы; TNF-α: фактор некроза опухолей альфа; Treg: регуляторные T-клетки.
[0087] Фиг. 16. Пути передачи сигналов от LPS and TNF-α, ведущие к воспалению. AP-1: белок 1-активатор (фактор транскрипции); IL: интерлейкины, LPS: липополисахариды; NF-κB: ядерный фактор каппа B (фактор транскрипции); TLR4: toll-подобный рецептор 4 (рецептор LPS); TNF-α: фактор некроза опухолей альфа; TNFR: рецептор TNF-α.
[0088] Фиг. 17. Трансэпителиальное электрическое сопротивление (TEER) в контрольных исследованиях CM и NaB. TEER измеряли через 24 часа после обработки сокультур Caco-2/THP1-Blue™, и каждое значение в 24 часа нормализовали к соответствующему ему значению в 0 часов и представляли как процент от исходного значения. Пунктирная линия представляет 100% (исходное значение). Данные нанесены как среднее значение ± стандартная ошибка среднего значения (SEM). (*) представляет статистически значимое различие между CM и NaB. (****) = p<0,0001. CM: полня среда; NaB: бутират натрия.
[0089] Фиг. 18. Базолатерная активность NF-κB в клетках THP1-Blue™ при контрольных исследованиях LPS-, LPS+, LPS+HC и LPS+NaB. Активность NF-κB измеряли через 6 часов обработки LPS сокультур Caco-2/THP1-Blue™ на базолатерной стороне после предварительной обработки в течение 24 часов NaB или средой полного состава на апикальной стороне. Данные нанесены как среднее значение ± SEM. (*) представляет статистически значимое различие по сравнению с LPS+. (*) = p<0,05; (****) = p<0,0001. LPS-: клетки, обработанные средой полного состава (без LPS); LPS+: клетки, обработанные LPS; HC: гидрокортизон; NaB: бутират натрия.
[0090] Фиг. 19. Базолатерная секреция IL-6 (A) и IL-10 (B) при контрольных исследованиях LPS-, LPS+, LPS+HC и LPS+NaB. Цитокины измеряли через 6 часов обработки LPS сокультур Caco-2/THP1-Blue™ на базолатерной стороне после предварительной обработки в течение 24 часов NaB или средой полного состава на апикальной стороне. Данные нанесены как среднее значение ± SEM. (*) представляет статистически значимое различие по сравнению с LPS+. (***) = p<0,001; (****) = p<0,0001. LPS-: клетки, обработанные средой полного состава (без LPS); LPS+: клетки, обработанные LPS; HC: гидрокортизон; NaB: бутират натрия.
[0091] Фиг. 20. Базолатерная секреция IL-1β (A), IL-8 (B), CXCL10 (C), TNF-α (D) и MCP-1 (E) при контрольных исследованиях LPS-, LPS+, LPS+HC и LPS+NaB. Цитокины измеряли через 6 часов обработки сокультур Caco-2/THP1-Blue™ на базолатерной стороне после предварительной обработки в течение 24 часов NaB или полной средой на апикальной стороне. Данные нанесены как среднее значение ± SEM. (*) представляет статистически значимое различие по сравнению с LPS+. (*) = p<0,05; (**) = p<0,01; (***) = p<0,001; (****) = p<0,0001. LPS-: клетки, обработанные средой полного состава (без LPS); LPS+: клетки, обработанные LPS; HC: гидрокортизон; NaB: бутират натрия.
[0092] Фиг. 21. Эффект образцов SHIME на трансэпителиальное электрическое сопротивление (TEER) сокультур Caco-2/THP1-Blue™. Результаты показаны для трех разных доноров отдельно (A) и как среднее значение для трех доноров (B). TEER измеряли через 24 часа после обработки сокультур, и каждое значение для момента времени=24 часам нормализовали к соответствующему ему значению для момента времени=0 часам и представлено как процент от исходного значения. Серая пунктирная линия представляет 100% (исходное значение). Пунктирная линия соответствует экспериментальному контролю CM (полной среде). Данные нанесены как среднее значение ± SEM. Не было обнаружено значимых различий между контролем и обработкой трех разных доноров. PC: образцы из проксимального отдела ободочной кишки; DC: образцы из дистального отдела ободочной кишки.
[0093] Фиг. 22. Эффект образцов SHIME на активность NF-κB в клетках THP-1-Blue™. Результаты показаны для трех разных доноров отдельно (A) и как среднее значение для трех доноров (B). Уровни активности NF-κB измеряли через 6 часов после обработки LPS на базолатеральной стороне сокультур Caco-2/THP-1-Blue™ после предварительной обработки апикальной стороны в течение 24 часов образцами SHIME. Пунктирная линия соответствует экспериментальному контролю LPS+. Данные нанесены как среднее значение ± SEM. Не было обнаружено значимых различий между контролем и обработкой трех разных доноров. PC: образцы из проксимального отдела ободочной кишки; DC: образцы из дистального отдела ободочной кишки.
[0094] Фиг. 23. Эффект образцов SHIME на секрецию IL-6 (A и B) и IL-10 (C и D). Результаты показаны для трех разных доноров отдельно (A и C) и как среднее значение для трех доноров (B и D). Уровни цитокинов измеряли через 6 часов после обработки LPS на базолатеральной стороне сокультур Caco-2/THP-1-Blue™ после предварительной обработки апикальной стороны в течение 24 часов образцами SHIME. Пунктирная линия соответствует экспериментальному контролю LPS+. Данные нанесены как среднее значение ± SEM. (*) представляет статистически значимое различие по сравнению с контролем. (*) = p<0,05. PC: образцы из проксимального отдела ободочной кишки; DC: образцы из дистального отдела ободочной кишки.
[0095] Фиг. 24. Эффект образцов SHIME на секрецию IL-1β (A+B) и TNF-α (C+D). Результаты показаны для трех разных доноров отдельно (A-C) и как среднее значение для трех доноров (B-D). Уровни цитокинов измеряли через 6 часов после обработки LPS на базолатеральной стороне сокультур Caco-2/THP-1-Blue™ после предварительной обработки апикальной стороны в течение 24 часов образцами SHIME. Пунктирная линия соответствует экспериментальному контролю LPS+. Данные нанесены как среднее значение ± SEM. (*) представляет статистически значимое различие по сравнению с контролем. (****) = p<0,0001 PC: образцы из проксимального отдела ободочной кишки; DC: образцы из дистального отдела ободочной кишки.
[0096] Фиг. 25. Эффект образцов SHIME на секрецию IL-8 (A+B), CXCL10 (C+D) и MCP-1 (E+F). Результаты показаны для трех разных доноров отдельно (A-C-E) и как среднее значение для трех доноров (B-D-F). Уровни цитокинов измеряли через 6 часов после обработки LPS на базолатеральной стороне сокультур Caco-2/THP-1-Blue™ после предварительной обработки апикальной стороны в течение 24 часов образцами SHIME. Пунктирная линия соответствует экспериментальному контролю LPS+. Данные нанесены как среднее значение ± SEM. Не было обнаружено значимых различий между контролем и обработкой для трех разных доноров. PC: образцы из проксимального отдела ободочной кишки; DC: образцы из дистального отдела ободочной кишки.
Определения
[0097] Используемый здесь термин «сфинган» относится к высокоацильному сфингану, среднеацильному сфингану, низкоацильному сфингану, высокоацильному сфингану в виде полисахарида, среднеацильному сфингану в виде полисахарида, низкоацильному сфингану в виде полисахарида, высокоацильному сфингану в виде олигосахарида, среднеацильному сфингану в виде олигосахарида, низкоацильному сфингану в виде олигосахарида или их комбинации.
[0098] Используемый здесь термин «высокоацильный» (или «HA»), относится к сфингану, содержащему ацильную группу (например, ацетил и глицерил). Высокоацильный сфинган включает, например, HA геллан, HA велан, HA рамсан, HA диутан и т.д.
[0099] Используемый здесь термин «среднеацильный» (или «IA») относится к сфингану, содержание ацильных групп в котором меньше, чем в высокоацильном сфингане, но больше, чем содержание ацильных групп в низкоацильном сфингане. Среднеацильный сфинган включает, например, IA геллан, IA велан, IA рамсан, IA диутан и т.д.
[0100] Используемый здесь термин «низкоацильный» (или «LA») относится к сфингану, в котором ацильная группа (группы) была по существу удалена. Низкоацильный сфинган включает, например, LA геллан, LA велан, LA рамсан, LA диутан и т.д.
[0101] Встречающийся в природе сфинган может включать, например, геллан (S-60), велан (S-130), рамсан (S-194), диутан (S-657), S-88, S-198 и S-7, состоящие из замещенного или незамещенного тетрамерного сахарида («тетрамера»), обычно обозначаемого как [(→3)Glc(β1→4)GlcA(β1→4)Glc(β1→4)Rha(α1→)]n, где Glc и GlcA представляют собой D-сахара, тогда как Rha представляет собой L-сахар, и, где применимо, Man представляет собой L-сахар. Химические структуры выбранных сфинганов представлены ниже указанием сокращенных терминов для отдельных моносахаридов (например, (1→3)Glc, (1→4)GlcA, (1→4)Glc и (1→4)Rha))
[0102] Используемый здесь термин «M» относится к физиологически приемлемому катиону, в том числе, например, протону (H+), натрию (Na+), калию (K+), кальцию (Ca2+), магнию (Mg2+) или их комбинации.
[0103] Значение «n» относится к целому или дробному числу и относится к числу тетрамерных единиц, которые могут быть замещенными или незамещенными. Понятно, что некоторые встречающиеся в природе сфинганы имеют значение n, которое может коррелировать с молекулярной массой встречающегося в природе сфингана (например, природной геллановой камеди, имеющей MW ≈ 2,5×106 и MN ≈ 2,2×106). (US6242035B1)
[0104] Используемое здесь выражение «степень полимеризации» или DP относится к числу моносахаридных единиц в полисахаридной или олигосахаридной цепи. Например, со ссылкой на химическую структуру, представленную выше, где n равно четырем, DP равна шестнадцати.
[0105] Используемое здесь выражение «сфинган в виде полисахарида» (или «SPS») относится к высоко-/низкоацильному сфингану, имеющему DP, большую 30 и меньшую таковой у природного сфингана. Понятно, что SPS, полученный из высоко-/средне-/низкоацильного сфингана, может включать множество полисахаридов с различными DP.
[0106] Используемое здесь выражение «сфинган в виде олигосахарида» (или «SOS») относится к высоко-/низкоацильному сфингану, имеющему DP, большую двух или равную двум и меньшую или равную тридцати (т.е. 2≥ DP≤30). Понятно, что SOS, полученный из высоко-/средне-/низкоацильного сфингана (или HA/IA/LA сфингана), может включать множество олигосахаридов
Подробное описание настоящего изобретения
[0107] Раскрытые здесь варианты осуществления в целом относятся к композиции для приема внутрь, композиции для приема внутрь и ее применению, способам применения композиции для приема внутрь, способу получения сфингана в виде олигосахарида, и сфингану в виде олигосахарида, полученному указанным способом получения сфингана в виде олигосахарида.
[0108] Первый вариант осуществления направлен на композицию для приема внутрь, содержащую эффективное для проявления пребиотической активности количество сфингана.
[0109] Эффективное для проявления пребиотической активности количество сфингана может составлять от приблизительно 1 г до приблизительно 10 г, и все значениями между ними, так, например, приблизительно 1,1, приблизительно 1,2, приблизительно 1,3, приблизительно 1,4, приблизительно 1,5, приблизительно 1,6, приблизительно 1,7, приблизительно 1,8, приблизительно 1,9, приблизительно 2,0, приблизительно 2,1, приблизительно 2,2, приблизительно 2,3, приблизительно 2,4, приблизительно 2,5, приблизительно 2,6, приблизительно 2,7, приблизительно 2,8, приблизительно 2,9, приблизительно 3,0, приблизительно 3,1, приблизительно 3,2, приблизительно 3,3, приблизительно 3,4, приблизительно 3,5, приблизительно 3,6, приблизительно 3,7, приблизительно 3,8, приблизительно 3,9, приблизительно 4,0, приблизительно 4,1, приблизительно 4,2, приблизительно 4,3, приблизительно 4,4, приблизительно 4,5, приблизительно 4,6, приблизительно 4,7, приблизительно 4,8, приблизительно 4,9, приблизительно 5,0, приблизительно 5,1, приблизительно 5,2, приблизительно 5,3, приблизительно 5,4, приблизительно 5,5, приблизительно 5,6, приблизительно 5,7, приблизительно 5,8, приблизительно 5,9, приблизительно 6,0, приблизительно 6,1, приблизительно 6,2, приблизительно 6,3, приблизительно 6,4, приблизительно 6,5, приблизительно 6,6, приблизительно 6,7, приблизительно 6,8, приблизительно 6,9, приблизительно 7,0, приблизительно 7,1, приблизительно 7,2, приблизительно 7,3, приблизительно 7,4, приблизительно 7,5, приблизительно 7,6, приблизительно 7,7, приблизительно 7,8, приблизительно 7,9, приблизительно 8,0, приблизительно 8,1, приблизительно 8,2, приблизительно 8,3, приблизительно 8,4, приблизительно 8,5, приблизительно 8,6, приблизительно 8,7, приблизительно 8,8, приблизительно 8,9, приблизительно 9,0, приблизительно 9,1, приблизительно 9,2, приблизительно 9,3, приблизительно 9,4, приблизительно 9,5, приблизительно 9,6, приблизительно 9,7, приблизительно 9,8 или приблизительно 9,9.
[0110] В одном аспекте первого варианта осуществления количество сфингана выбрано из: от приблизительно 1 г до приблизительно 10 г, от приблизительно 1 г до приблизительно 9 г, от приблизительно 1 г до приблизительно 8 г, от приблизительно 1 г до приблизительно 7 г, от приблизительно 1 г до приблизительно 6 г, от приблизительно 1 г до приблизительно 5 г, от приблизительно 1 г до приблизительно 4 г, от приблизительно 1 г до приблизительно 3 г, или приблизительно 2 г.
[0111] Композиции первого варианта осуществления могут содержать HA/IA/LA сфинган, например, HA геллан, IA геллан, LA геллан, HA велан, IA велан, LA велан, HA рамсан, IA рамсан, LA рамсан, HA диутан, IA диутан, LA диутан, S-88, S-198, S-7, или их комбинацию.
[0112] Композиции первого варианта осуществления могут содержать HA/IA/LA сфинган в виде полисахарида.
[0113] Как здесь объяснено более подробно, HA/IA/LA сфинган в виде полисахарида может быть получен из HA/LA сфингана с использованием, например, способа, который включает гомогенизацию под высоким давлением, как описано, например, в десятом варианте осуществления. Приводимые в качестве примера HA/IA/LA сфинганы в виде полисахаридов включают, но без ограничения этим: высокоацильный геллан в виде полисахарида, полученного из высокоацильного геллана, (например, геллан KELCOGEL® LT100 и KELGOGEL® HT), среднеацильный геллан в виде полисахарида, полученного из среднеацильного геллана (например, KELCOGEL® DGA), низкоацильный геллан в виде полисахарида, полученного из низкоацильного геллана, (например, геллан KELCOGEL® LT, геллан KELCOGEL®, геллан KELCOGEL® F и геллан GELRITETM MK), высоко-/средне-/низкоацильный велан в виде полисахарида, полученного из высоко-/средне-/низкоацильного велана, высоко-/средне-/низкоацильный диутан в виде полисахарида, полученного из высоко-/средне-/низкоацильного диутана, и высоко-/средне-/низкоацильный рамсан в виде полисахарида, полученного из высоко-/средне-/низкоацильного рамсана.
[0114] Композиции первого варианта осуществления могут содержать HA/IA/LA сфинган в виде олигосахарида, происходящего либо из встречающегося в природе HA/IA/LA сфингана, либо из HA/IA/LA сфингана в виде полисахарида. В одном аспекте, композиции первого варианта осуществления могут содержать HA/IA/LA сфинган в виде олигосахарида, происходящего либо из встречающегося в природе HA/IA/LA сфингана, либо из HA/IA/LA сфингана в виде полисахарида, имеющего молекулярную массу, которая, как определено с помощью гель-фильтрации, составляет от приблизительно 0,3 кДа до 12 кДа. В другом аспекте, композиции первого варианта осуществления могут содержать HA/IA/LA сфинган в виде олигосахарида, происходящего либо из встречающегося в природе HA/IA/LA сфингана, либо из HA/IA/LA сфингана в виде полисахарида, имеющего молекулярную массу, которая, как определено с помощью гель-фильтрации, составляет приблизительно 1 кДа.
[0115] Как здесь объяснено более подробно, HA/IA/LA сфинган в виде олигосахарида может быть получен из встречающегося в природе HA/IA/LA сфингана или HA/IA/LA сфингана в виде полисахарида, например, способом, который включает гидролиз гликозидной связи встречающегося в природе HA/IA/LA сфингана или HA/IA/LA сфингана в виде полисахарида и подвергание гидролизованной композиции ультрафильтрации, гель-фильтрации, преципитации, центрифугированию или их комбинации, как описано, например, в десятом варианте осуществления. Приводимые в качестве примера HA/IA/LA сфинганы в виде олигосахаридов включают, но без ограничения этим:
(i) композицию, содержащую (или состоящую из) Glc, GlcA, Glc, GlcA,Glyc, Glc, GlcA,Rha, Glc, GlcA,Rha, Glyc, Glc, GlcA,Rha,-H2O, Glc, Rha, Glc, Rha+28, Glc2,GlcA, Glc2,GlcA, Rha, Glc2,GlcA, Rha,+28, Glc2,GlcA, Rha,Ac, Glc2,GlcA, Rha,Glyc, Glc2,GlcA, Rha,Glyc,+28, Glc2,GlcA, Rha,Glyc.-H2O, Glc2,GlcA, Rha,-H2O, Glc2,GlcA, Rha2,Glyc, Glc2,GlcA2,Rha, Glc2,GlcA2,Rha2,Ac2,Glyc2,-H2O, Glc2,Rha, Glc3,GlcA, Rha, Glc3,GlcA, Rha2, Glc3,GlcA, Rha2, Glc3,GlcA, Rha2, Glc3,GlcA, Rha2,Glyc, Glc3,GlcA2,Rha, Glc3,GlcA2,Rha, Glyc, Glc3,GlcA2,Rha2,Glyc, Glc3,GlcA3,Rha2, Glc3,GlcA3,Rha2, Glc4,GlcA, Rha2,+43, Glc4,GlcA, Rha2,Ac, Glyc, Glc4,GlcA2,Rha, Glc4,GlcA2,Rha, Ac,Glyc,-H2O, Glc4,GlcA2,Rha, Ac,Glyc2, Glc4,GlcA2,Rha2,Ac, Glyc, Glc4,GlcA2,Rha2,Glyc, Glc4,GlcA3,Rha2, Glc4,GlcA2,Rha3,Ac, Glc4,GlcA3,Rha2/Glc4,GlcA2,Rha2,Glyc2, Glc5,GlcA2,Rha2, Glc5,GlcA2,Rha2, Glc5,GlcA2,Rha2,Ac, Glc5,GlcA4,Rha2, Glc6,GlcA3,Rha3, Glc(Ac/Glyc)x, GlcAx,Glcx, Rhax (где x составляет от 4 до приблизительно 25), Glcx, GlcAx,Glcx, Rhax (где x составляет от 4 до приблизительно 25), или их комбинацию;
(ii) композицию, содержащую (или состоящую из) тетрамер (Glc, GlcA,Glc, Rha), тетрамер (Glc, GlcA,Glc, Rha) с ацетатом и/или глицератом, октамер (Glc, GlcA,Glc, Rha,Glc, GlcA,Glc, Rha), октамер (Glc, GlcA,Glc, Rha,Glc, GlcA,Glc, Rha) с ацетатом и/или глицератом, Glc, GlcA,Glc, Rha, Glc,GlcA, Glc, Rha, или их комбинацию;
(iii) композицию, содержащую (или состоящую из) тетрамер (Glc, GlcA,Glc, Rha), октамер (Glc, GlcA,Glc, Rha,Glc, GlcA,Glc, Rha), пентамер (Glc, GlcA,Glc, Rha,Glc), GlcA, Glc,Rha, Glc, GlcA,Glc, Glc, GlcA, или их комбинацию;
(iv) композицию, содержащую (или состоящую из) Glc(Glc-Glc),GlcA, Glc(Glc-Glc), GlcA, Glc, Glc, Glc, или их комбинацию;
(v) композицию, содержащую (или состоящую из) тетрамер (Glc, GlcA,Glc, Rha), GlcA, Glc,(Rha-Rha), Glc,(Rha-Rha),Rha, GlcA, Glc,Rha, Glc, GlcA,Glc, Rha, Glc, GlcA, Glc;
(vi) композицию, содержащую (или состоящую из) Glc, GlcA, Glc, GlcA,Glyc, Glc, GlcA,Rha, Glc, GlcA,Rha, Glyc, Glc, Rha, Glc, Rha+28, Glc2,GlcA, Glc2,GlcA, Rha, Glc2,GlcA, Rha,+28, Glc2,GlcA, Rha,Ac, Glc2,GlcA, Rha,Glyc, Glc2,GlcA, Rha,Glyc,+28, Glc3,GlcA, Rha, Glc3,GlcA, Rha2, Glc3,GlcA, Rha2, Glc3,GlcA, Rha2, Glc3,GlcA, Rha2,Glyc, Glc3,GlcA2,Rha, Glyc, Glc3,GlcA2,Rha2,Glyc, Glc3,GlcA3,Rha2, Glc4,GlcA, Rha2,Ac, Glyc, Glc4,GlcA2,Rha2,Ac, Glyc, Glc4,GlcA2,Rha2,Glyc, Glc4,GlcA2,Rha3,Ac, Glc4,GlcA3,Rha2/Glc4,GlcA2,Rha2,Glyc2, Glc5,GlcA2,Rha2, Glc5,GlcA2,Rha2,Ac, Glc(Ac/Glyc)x, GlcAx,Glcx, Rhax (где x составляет от 4 до приблизительно 25), или их комбинацию;
(vii) композицию, содержащую (или состоящую из) Glc, GlcA, Glc, GlcA, Rha, Glc, Rha, Glc, Rha+28, Glc2,GlcA, Rha, Glc2,GlcA, Rha,+28, Glc2,GlcA2,Rha, Glc3,GlcA, Rha, Glc3,GlcA, Rha2, Glc3,GlcA2,Rha, Glc3,GlcA3,Rha2, Glc3,GlcA3,Rha2, Glc4,GlcA, Rha2,+43, Glc4,GlcA2,Rha, Glc4,GlcA3,Rha2, Glc5,GlcA2,Rha2, Glc5,GlcA2,Rha2, Glc5,GlcA4,Rha2, Glc6,GlcA3,Rha3, Glcx, GlcAx,Glcx, Rhax (где x составляет от 4 до приблизительно 25), или их комбинацию;
(viii) композицию, содержащую (или состоящую из) Glc, GlcA,Rha,-H2O, Glc, Rha, Glc2,GlcA, Rha,-H2O, Glc2,Rha, или их комбинацию;
(ix) композицию, содержащую (или состоящую из) Glc, GlcA, Glc, GlcA, Glyc, Glc, GlcA, Rhaa, Glc, GlcA,Rha, Glyc, Glc, Rha, Glc, Rha+28, Glc2,GlcA, Glc2,GlcA, Rha, Glc2,GlcA, Rha,+28, Glc2,GlcA, Rha,Ac, Glc2,GlcA, Rha,Glyc, Glc2,GlcA, Rha,Glyc,+28, Glc2,GlcA, Rha,Glyc.-H2O, Glc2,GlcA, Rha2,Glyc, Glc2,GlcA2,Rha2,Ac2,Glyc2,-H2O, Glc3,GlcA, Rha, Glc3,GlcA, Rha2, Glc3,GlcA, Rha2,Glyc, Glc3,GlcA2,Rha, Glyc, Glc3,GlcA2,Rha2,Glyc, Glc3,GlcA3,Rha2, Glc4,GlcA, Rha2,+43, Glc4,GlcA, Rha2,Ac, Glyc, Glc4,GlcA2,Rha, Ac,Glyc,-H2O, Glc4,GlcA2,Rha, Ac,Glyc2, Glc4,GlcA2,Rha2,Ac, Glyc, Glc4,GlcA2,Rha2,Glyc, Glc4,GlcA3,Rha2, Glc4,GlcA2,Rha3,Ac, Glc4,GlcA3,Rha2/Glc4,GlcA2,Rha2,Glyc2, Glc5,GlcA2,Rha2, Glc5,GlcA2,Rha2,Ac, Glc(Ac/Glyc)x, GlcAx,Glcx, Rhax (где x составляет от 4 до приблизительно 25), или их комбинацию;
(x) композицию, содержащую (или состоящую из) любой из образцов с №№ 1-18; или
(xi) композицию, содержащую (или состоящую из) любой из образцов с №№ 9, 10, 17 и 18.
[0116] Как указано выше, некоторые сфинганы могут быть замещены ацильным заместителем, моносахаридом, или дисахаридной боковой цепью (например, (1→4)Glc диутана замещается в положении O3 боковой цепью Rha(α1→4)-Rha(α1→)). Замещенный олигосахарид, имеющий сахаридную боковую цепь, обозначается с помощью скобок, например, GlcA, Glc,(Rha-Rha) и Glc,(Rha-Rha),Rha.
[0117] И ссылка на HA/IA/LA сфинган в виде олигосахарида, как подразумевается, означает любой из приводимых в качестве примера HA/IA/LA сфинганов в виде олигосахаридов или их комбинация.
[0118] Композиции могут быть в форме жидкостей, полутвердых или твердых веществ. Композиции могут быть в форме хлопьев, закусок или другой форме для приема внутрь. Композиции могут быть на фруктовой основе, например соке или смузи, или на основе молочных продуктов, например молоке, мороженом или йогуртах. Композиции могут быть соответственно в форме напитков. Термин «напиток» включает готовую к употреблению жидкую форму, а также концентрат и порошкообразный препарат для растворения. Готовый к употреблению напиток может быть негазированным или газированным.
[0119] Композиции могут быть несладкими или подслащенными сахаром или сильными подсластителями, такими как сукралоза, глицирризинат аммония, ацесульфам-K, аспартам, сахарин, соль (например, натрия, калия, кальция и т.д.) сахарина, цикламат натрия, стевия, другие подсластители, не являющиеся сахаром, и их смеси. Композиции могут также содержать другие обычные добавки, такие как корригенты, красители, стабилизаторы и т.д.
[0120] Композиции могут храниться в виде порошка в запечатанном контейнере или упаковке, которая может включать инструкции по применению.
[0121] Альтернативно, композиции могут быть приготовлены в виде продукта таблетки или капсулы, который может содержать, помимо сфингана, другие приемлемые наполнители, такие как связующее, наполнитель, смазывающее вещество, вызывающий дезинтеграцию агент, способствующее скольжению вещество, добавка для повышения текучести, агент против слеживания, сорбент, консервант, смачивающий агент, подсластитель, ароматизатор, глазировочное средство и т.д. На таблетки может быть нанесено покрытие в соответствии со способами, хорошо известными в данной области техники. Примеры наполнителей включают, но без ограничения этим, карбонат щелочноземельных металлов (например, карбонат магния, карбонат кальция и т.д.); сшитый полимер (например, сшитый поливинилпирролидон (кросповидон) и сшитую карбоксиметилцеллюлозу натрия (кроскармеллозу натрия)); жирную кислоту; коллоидальную двуокись кремния; смазывающее вещество (например, стеариновую кислоту, стеарин, стеарат магния); агент для регулирования pH (например, кислоту (например, соляную кислоту) и основание (например, гидроксид натрия)); растительное волокно (например, кукурузный протеин зеин); полисахарид и его производные (например, крахмал, целлюлозу или модифицированную целлюлозу, такую как микрокристаллическая целлюлоза и простые эфиры целлюлозы, такие как гидроксипропилцеллюлоза и гидроксипропилметилцеллюлоза); белок (например, желатин); сахарид и его производные (например, дисахарид, например, сахарозу, лактозу и т. д.); шеллак; диоксид кремния; натрия крахмал гликолят; сахарный спирт (например, изомальтит, ксилит, сорбит и мальтит); синтетический полимер (например, поливинилпирролидон и полиэтиленгликоль), тальк и воск.
[0122] Композиция может также содержать пробиотик и дополнительный пребиотик.
[0123] Примеры пробиотиков включают, но без ограничения этим, Lactobacillus rhamnosus GG, Bifidobacterium infantis, Lactobacillus acidophilus, Bifidobacterium lactis HN019, Bifidobacterium longum (в том числе штамм 35624), Lactobacillus salivarius, Bifodobacterium bifidum, Lactobacillus plantarum, Lactobacillus paracasei, Bifidobacterium breve, Lactobacillus gasseri KS-13, Bacillus coagulans (GBI-30, 6086), Bacillus subtilis DE111, каждый из которых может использоваться отдельно или в их комбинации.
[0124] Примеры дополнительных пребиотиков включают, но без ограничения этим, инулин, фруктоолигосахарид, галактоолигосахарид, гуаровую камедь, камедь тары, ксантановую камедь, ксантановый полисахарид, ксантановый олигосахарид, конжаковую камедь, камедь карайя, арабиногалактан, лактулозу, псиллиум, пектин, пектиновый полисахарид, пектиновый олигосахарид, трагакант, гуммиарабик, каррагенин и т.п.
[0125] Представленные здесь результаты показывают, что сфинган (A) стимулирует рост полезных бактерий в ободочной кишке человека; (B) снижает уровень пропионата и/или увеличивает уровень бутирата в ободочной кишке человека; (C) улучшает целостность кишечного барьера в ободочной кишке человека; и/или (D) снижает уровни TNF-α и/или IL-8 в ободочной кишке человека. Соответственно, раскрытые здесь варианты осуществления относятся к композиции для приема внутрь для:
(A) стимуляции роста полезных бактерий в ободочной кишке млекопитающего, при этом указанная композиция содержит эффективное для стимуляции роста полезных бактерий количество сфингана и среду для приема внутрь (второй вариант осуществления);
(B) снижения уровня пропионата и/или повышения уровня бутирата в ободочной кишке млекопитающего, при этом указанная композиция содержит эффективное количество сфингана и среду для приема внутрь (третий вариант осуществления);
(C) улучшения целостности кишечного барьера в ободочной кишке млекопитающего, при этом указанная композиция содержит эффективное для улучшения целостности кишечного барьера количество сфингана и среду для приема внутрь (четвертый вариант осуществления); или
(D) снижения уровней TNF-α и/или IL-8 в ободочной кишке млекопитающего, при этом указанная композиция содержит эффективное для снижения уровней TNF-α и/или IL-8 количество сфингана и среду для приема внутрь (пятый вариант осуществления).
[0126] По отношению к любому из второго, третьего, четвертого и пятого вариантов осуществления, рассчитанное количество сфингана (т.е. (i) эффективное для стимуляции роста полезных бактерий количество сфингана (второй вариант осуществления), (ii) эффективное количество сфингана (третий вариант осуществления), (iii) эффективное для улучшения целостности кишечного барьера количество сфингана (четвертый вариант осуществления), и (iv) эффективное для снижения уровней TNF-α и/или IL-8 количество сфингана (пятый вариант осуществления)) может составлять от приблизительно 1 г до приблизительно 10 г сфингана, и все значения между ними, так, например, приблизительно 1,1, приблизительно 1,2, приблизительно 1,3, приблизительно 1,4, приблизительно 1,5, приблизительно 1,6, приблизительно 1,7, приблизительно 1,8, приблизительно 1,9, приблизительно 2,0, приблизительно 2,1, приблизительно 2,2, приблизительно 2,3, приблизительно 2,4, приблизительно 2,5, приблизительно 2,6, приблизительно 2,7, приблизительно 2,8, приблизительно 2,9, приблизительно 3,0, приблизительно 3,1, приблизительно 3,2, приблизительно 3,3, приблизительно 3,4, приблизительно 3,5, приблизительно 3,6, приблизительно 3,7, приблизительно 3,8, приблизительно 3,9, приблизительно 4,0, приблизительно 4,1, приблизительно 4,2, приблизительно 4,3, приблизительно 4,4, приблизительно 4,5, приблизительно 4,6, приблизительно 4,7, приблизительно 4,8, приблизительно 4,9, приблизительно 5,0, приблизительно 5,1, приблизительно 5,2, приблизительно 5,3, приблизительно 5,4, приблизительно 5,5, приблизительно 5,6, приблизительно 5,7, приблизительно 5,8, приблизительно 5,9, приблизительно 6,0, приблизительно 6,1, приблизительно 6,2, приблизительно 6,3, приблизительно 6,4, приблизительно 6,5, приблизительно 6,6, приблизительно 6,7, приблизительно 6,8, приблизительно 6,9, приблизительно 7,0, приблизительно 7,1, приблизительно 7,2, приблизительно 7,3, приблизительно 7,4, приблизительно 7,5, приблизительно 7,6, приблизительно 7,7, приблизительно 7,8, приблизительно 7,9, приблизительно 8,0, приблизительно 8,1, приблизительно 8,2, приблизительно 8,3, приблизительно 8,4, приблизительно 8,5, приблизительно 8,6, приблизительно 8,7, приблизительно 8,8, приблизительно 8,9, приблизительно 9,0, приблизительно 9,1, приблизительно 9,2, приблизительно 9,3, приблизительно 9,4, приблизительно 9,5, приблизительно 9,6, приблизительно 9,7, приблизительно 9,8 и приблизительно 9,9 г.
[0127] В одном аспекте любого из второго, третьего, четвертого и пятого вариантов осуществления, млекопитающим является, например, человек, собака, кошка, крыса, мышь, хомяк, морская свинка, корова, бизон, свинья, овца, лошадь, коза, олень, лама, альпака и т.п.
[0128] В одном аспекте любого из второго, третьего, четвертого и пятого вариантов осуществления, количество сфингана выбрано из: от приблизительно 1 г до приблизительно 10 г, от приблизительно 1 г до приблизительно 9 г, от приблизительно 1 г до приблизительно 8 г, от приблизительно 1 г до приблизительно 7 г, от приблизительно 1 г до приблизительно 6 г, от приблизительно 1 г до приблизительно 5 г, от приблизительно 1 г до приблизительно 4 г, от приблизительно 1 г до приблизительно 3 г, или приблизительно 2 г.
[0129] И, в одном аспекте любого из второго, третьего, четвертого и пятого вариантов осуществления, количество сфингана является достаточным для достижения эффективной концентрации сфингана в ободочной кишке, причем указанная концентрация сфингана в ободочной кишке колеблется в пределах от приблизительно 1 мг/мл до приблизительно 10 мг/мл, и составляет все значения между ними, например, приблизительно 1,5 мг/мл, приблизительно 2 мг/мл, приблизительно 2,5 мг/мл, приблизительно 3 мг/мл, приблизительно 3,5 мг/мл, приблизительно 4 мг/мл, приблизительно 4,5 мг/мл, приблизительно 5 мг/мл, приблизительно 5,5 мг/мл, приблизительно 6 мг/мл, приблизительно 6,5 мг/мл, приблизительно 7 мг/мл, приблизительно 7,5 мг/мл, приблизительно 8 мг/мл, приблизительно 8,5 мг/мл, приблизительно 9 мг/мл или 9,5 мг/мл.
[0130] Композиции по любому из второго, третьего, четвертого и пятого вариантов осуществления могут содержать любой из встречающегося в природе HA/IA/LA сфингана, HA/IA/LA сфингана в виде полисахарида, HA/IA/LA сфингана в виде олигосахарида, или их комбинацию, и необязательно дополнительно содержат пробиотик или дополнительный пребиотик, как описано в первом варианте осуществления.
[0131] Кроме того, раскрытые здесь варианты осуществления относятся либо к способу для, либо применению для производства лекарственного средства или пищевой добавки для:
(A) стимуляции роста полезных бактерий в ободочной кишке млекопитающего, при этом указанный способ включает прием внутрь по эффективной схеме эффективного в отношении стимуляции роста полезных бактерий количества сфингана и среды для приема внутрь (шестой вариант осуществления);
(B) снижения уровня пропионата и/или повышения уровня бутирата в ободочной кишке млекопитающего, при этом указанный способ включает прием внутрь по эффективной схеме композиции, содержащей эффективное количество сфингана и среду для приема внутрь (седьмой вариант осуществления);
(C) улучшения целостности кишечного барьера в ободочной кишке млекопитающего, при этом указанный способ включает прием внутрь по эффективной схеме композиции, содержащей эффективное для улучшения целостности кишечного барьера количество сфингана и среду для приема внутрь (восьмой вариант осуществления);
(D) снижения уровней TNF-α и/или IL-8 в ободочной кишке млекопитающего, при этом указанный способ включает прием внутрь по эффективной схеме композиции, содержащей эффективное для снижения уровней TNF-α и/или IL-8 количество сфингана и среду для приема внутрь (девятый вариант осуществления);
(E) применения композиции по любому из первого, второго, третьего, четвертого и пятого вариантов осуществления отдельно или в комбинации с пробиотиком или дополнительным пребиотиком, описанным здесь, для производства композиции для (i) стимуляции роста полезных бактерий в ободочной кишке млекопитающего (десятый вариант осуществления), (ii) снижения уровня пропионата и/или повышения уровня бутирата в ободочной кишке млекопитающего (одиннадцатый вариант осуществления), (iii) улучшения целостности кишечного барьера в ободочной кишке млекопитающего (двенадцатый вариант осуществления), или (iv) снижения уровней TNF-α и/или IL-8 в ободочной кишке млекопитающего (тринадцатый вариант осуществления); или
(F) В одном аспекте любого из шестого, седьмого, восьмого, девятого, десятого, одиннадцатого, двенадцатого и тринадцатого вариантов осуществления, млекопитающим является, например, человек, собака, кошка, крыса, мышь, хомяк, морская свинка, корова, бизон, свинья, овца, лошадь, коза, олень, лама, альпака и т.п.
[0132] Для этих и других вариантов осуществления, описанных и заявленных здесь, эффективная схема для приема внутрь может включать, например, (i) ежедневный прием, например, один раз, два раза, три раза в день, и т.д.; (ii) еженедельный прием, например каждый день в течение семи дней, через день в течение семи дней и т.д.; (iii) ежемесячный прием, например ежедневный прием в течение желаемого периода времени с последующим периодом покоя, с продолжением ежедневного приема в течение желаемого периода времени.
[0133] По отношению к любому из шестого, седьмого, восьмого, девятого, десятого, одиннадцатого, двенадцатого и тринадцатого вариантов осуществления, рассчитанное количество сфингана (т.е. (i) эффективное для стимуляции роста полезных бактерий количество сфингана (шестой вариант осуществления), (ii) эффективное количество сфингана (седьмой вариант осуществления), (iii) эффективное для улучшения целостности кишечного барьера количество сфингана (восьмой вариант осуществления), и (iv) эффективное для снижения уровней TNF-α и/или IL-8 количество сфингана (девятый вариант осуществления)) может составлять от приблизительно 1 г до приблизительно 10 г сфингана, и все значения между ними, так, например, приблизительно 1,1, приблизительно 1,2, приблизительно 1,3, приблизительно 1,4, приблизительно 1,5, приблизительно 1,6, приблизительно 1,7, приблизительно 1,8, приблизительно 1,9, приблизительно 2,0, приблизительно 2,1, приблизительно 2,2, приблизительно 2,3, приблизительно 2,4, приблизительно 2,5, приблизительно 2,6, приблизительно 2,7, приблизительно 2,8, приблизительно 2,9, приблизительно 3,0, приблизительно 3,1, приблизительно 3,2, приблизительно 3,3, приблизительно 3,4, приблизительно 3,5, приблизительно 3,6, приблизительно 3,7, приблизительно 3,8, приблизительно 3,9, приблизительно 4,0, приблизительно 4,1, приблизительно 4,2, приблизительно 4,3, приблизительно 4,4, приблизительно 4,5, приблизительно 4,6, приблизительно 4,7, приблизительно 4,8, приблизительно 4,9, приблизительно 5,0, приблизительно 5,1, приблизительно 5,2, приблизительно 5,3, приблизительно 5,4, приблизительно 5,5, приблизительно 5,6, приблизительно 5,7, приблизительно 5,8, приблизительно 5,9, приблизительно 6,0, приблизительно 6,1, приблизительно 6,2, приблизительно 6,3, приблизительно 6,4, приблизительно 6,5, приблизительно 6,6, приблизительно 6,7, приблизительно 6,8, приблизительно 6,9, приблизительно 7,0, приблизительно 7,1, приблизительно 7,2, приблизительно 7,3, приблизительно 7,4, приблизительно 7,5, приблизительно 7,6, приблизительно 7,7, приблизительно 7,8, приблизительно 7,9, приблизительно 8,0, приблизительно 8,1, приблизительно 8,2, приблизительно 8,3, приблизительно 8,4, приблизительно 8,5, приблизительно 8,6, приблизительно 8,7, приблизительно 8,8, приблизительно 8,9, приблизительно 9,0, приблизительно 9,1, приблизительно 9,2, приблизительно 9,3, приблизительно 9,4, приблизительно 9,5, приблизительно 9,6, приблизительно 9,7, приблизительно 9,8 и приблизительно 9,9 г.
[0134] В одном аспекте любого из шестого, седьмого, восьмого, девятого, десятого, одиннадцатого, двенадцатого и тринадцатого вариантов осуществления, количество сфингана выбрано из: от приблизительно 1 г до приблизительно 10 г, от приблизительно 1 г до приблизительно 9 г, от приблизительно 1 г до приблизительно 8 г, от приблизительно 1 г до приблизительно 7 г, от приблизительно 1 г до приблизительно 6 г, от приблизительно 1 г до приблизительно 5 г, от приблизительно 1 г до приблизительно 4 г, от приблизительно 1 г до приблизительно 3 г, или приблизительно 2 г.
[0135] И, в одном аспекте любого из шестого, седьмого, восьмого, девятого, десятого, одиннадцатого, двенадцатого и тринадцатого вариантов осуществления, количество сфингана является достаточным для достижения эффективной концентрации сфингана в ободочной кишке, как здесь описано.
[0136] В качестве альтернативы, и по отношению к любому из шестого, седьмого, восьмого и девятого вариантов осуществления, млекопитающим является человек, и рассчитанное количество сфингана (т.е. (i) эффективное для стимуляции роста полезных бактерий количество сфингана (шестой вариант осуществления), (ii) эффективное количество сфингана (седьмой вариант осуществления), (iii) эффективное для улучшения целостности кишечного барьера количество сфингана (восьмой вариант осуществления), и (iv) эффективное для снижения уровней TNF-α и/или IL-8 количество сфингана (девятый вариант осуществления)) может составлять от приблизительно 10 мг/кг до приблизительно 150 мг/кг веса тела человека, принимающего внутрь композицию. Кроме того, предполагается, что количество сфингана включает все значения между указанными пределами, так, например, приблизительно 15 мг/кг, приблизительно 20 мг/кг, приблизительно 25 мг/кг, приблизительно 30 мг/кг, приблизительно 35 мг/кг, приблизительно 40 мг/кг, приблизительно 45 мг/кг, приблизительно 50 мг/кг, приблизительно 55 мг/кг, приблизительно 60 мг/кг, приблизительно 65 мг/кг, приблизительно 70 мг/кг, приблизительно 75 мг/кг, приблизительно 80 мг/кг, приблизительно 85 мг/кг, приблизительно 90 мг/кг, приблизительно 95 мг/кг, приблизительно 100 мг/кг, приблизительно 105 мг/кг, приблизительно 110 мг/кг, приблизительно 115 мг/кг, приблизительно 120 мг/кг, приблизительно 125 мг/кг, приблизительно 130 мг/кг, приблизительно 135 мг/кг, приблизительно 140 мг/кг или приблизительно 145 мг/кг.
[0137] В одном аспекте любого из шестого, седьмого, восьмого и девятого вариантов осуществления, млекопитающим является человек, и количество сфингана выбрано из: от приблизительно 10 мг/кг до приблизительно 150 мг/кг, от приблизительно 10 мг/кг до приблизительно 140 мг/кг, от приблизительно 10 мг/кг до приблизительно 130 мг/кг, от приблизительно 10 мг/кг до приблизительно 120 мг/кг, от приблизительно 10 мг/кг до приблизительно 110 мг/кг, от приблизительно 10 мг/кг до приблизительно 100 мг/кг, от приблизительно 10 мг/кг до приблизительно 90 мг/кг, от приблизительно 10 мг/кг до приблизительно 80 мг/кг, от приблизительно 10 мг/кг до приблизительно 70 мг/кг, от приблизительно 10 мг/кг до приблизительно 60 мг/кг, от 10 мг/кг до приблизительно 50 мг/кг, от приблизительно 10 мг/кг до приблизительно 40 мг/кг или от приблизительно 20 мг/кг до приблизительно 30 мг/кг веса тела человека, принимающего внутрь композицию.
[0138] Композиции любого из шестого, седьмого, восьмого и девятого вариантов осуществления могут содержать любой из встречающегося в природе HA/LA сфингана, HA/LA сфингана в виде полисахарида, HA/LA сфингана в виде олигосахарида, или их комбинацию, и необязательно дополнительно содержит пробиотик или дополнительный пребиотик, как описано в первом варианте осуществления.
[0139] Раскрытые здесь результаты показывают, что сфинган (например, геллановая камедь) повышал уровни Bifidobacteriaceae в модели проксимальной и дистальной частей ободочной кишки человека. На уровне операционной таксономической единицы («OTU»), основные изменения, как установлено, относятся к повышению уровней Bifidobacteriaceae OTU 2 (имеющей отношение к Bifidobacterium adolescentis). Следовательно, в одном аспекте второго, шестого или десятого вариантов осуществления бактериями являются Bifidobacteriaceae. Кроме того, в другом аспекте второго, шестого или десятого вариантов осуществления бактериями являются Bifidobacteriaceae OTU 2. Повышение уровней Bifidobacteriaceae в просвете проксимального отдела ободочной кишки колеблется в пределах от приблизительно 20% до приблизительно 180% во время обработки по сравнению с необработанным контролем, тогда как повышение уровней Bifidobacteriaceae в просвете дистального отдела ободочной кишки колеблется в пределах от приблизительно 330% до приблизительно 590% во время обработки по сравнению с необработанным контролем. В еще одном аспекте второго, шестого или десятого вариантов осуществления, уровни Bifidobacteriaceae повышаются в просвете проксимального отдела ободочной кишки на приблизительно 20% - приблизительно 180% во время обработки по сравнению с необработанным контролем. И, в дальнейшем аспекте второго, шестого или десятого вариантов осуществления уровни Bifidobacteriaceae повышаются в просвете дистального отдела ободочной кишки на приблизительно 330% - приблизительно 590% во время обработки по сравнению с необработанным контролем.
[0140] Кроме того, раскрытые здесь результаты показывают, что сфинган в виде олигосахарида в концентрации, составляющей приблизительно 4 мг/мл, повышал уровни бактерий (например, Blautia, Parabacteroides, Faecalibacterium, Clostridium XVIII) in vitro на основе образцов кала здоровых взрослых. Уровни Blautia in vitro повышались в по крайней мере приблизительно 5 раз по сравнению с необработанным контролем. Уровни Parabacteroides in vitro повышались в приблизительно 2 раза - приблизительно 40 раз по сравнению с необработанным контролем. Уровни Faecalibacterium in vitro повышались в приблизительно 10 раз - приблизительно 190 раз по сравнению с необработанным контролем. Уровни Clostridium XVIII in vitro повышались в приблизительно 12 раз - приблизительно 60 раз по сравнению с необработанным контролем.
[0141] Кроме того, раскрытые здесь результаты показывают, что сфинган в виде олигосахарида в концентрации, составляющей приблизительно 4 мг/мл, повышал уровни бактерий (например, Parabacteroides, Faecalibacterium, Clostridium XVIII) in vitro на основе образцов кала пациентов с воспалительным заболеванием кишечника. Уровни Blautia in vitro повышались в по крайней мере приблизительно 8 раз по сравнению с необработанным контролем. Уровни Faecalibacterium in vitro повышались в по крайней мере приблизительно 8 раз по сравнению с необработанным контролем. Уровни Clostridium XVIII in vitro повышались в приблизительно 20 раз - приблизительно 100 раз по сравнению с необработанным контролем.
[0142] Раскрытые здесь результаты показывают, что принимаемый внутрь сфинган (например, геллановая камедь) повышал уровни пропионата в модели как проксимальной, так и дистальной частей ободочной кишки человека, и что принимаемая внутрь геллановая камедь повышала уровни бутирата в модели как проксимальной, так и дистальной частей ободочной кишки. Следовательно, в одном аспекте третьего, седьмого или одиннадцатого вариантов осуществления, в котором млекопитающим является человек, понижение уровней пропионата в проксимальном отделе ободочной кишки колеблется в пределах от приблизительно 8% до приблизительно 21% во время обработки по сравнению с контролем. В одном аспекте третьего, седьмого или одиннадцатого вариантов осуществления, в котором млекопитающим является человек, понижение уровней пропионата в дистальном отделе ободочной кишки колеблется в пределах от приблизительно 8% до приблизительно 11% во время обработки по сравнению с контролем. В одном аспекте третьего, седьмого или одиннадцатого вариантов осуществления, в котором млекопитающим является человек, повышение уровней бутирата в проксимальном отделе ободочной кишки колеблется в пределах от приблизительно 15% до приблизительно 24%. В одном аспекте третьего, седьмого или одиннадцатого вариантов осуществления, в котором млекопитающим является человек, повышение уровней бутирата в дистальном отделе ободочной кишки колеблется в пределах от приблизительно 4% до приблизительно 13%.
[0143] Четырнадцатый вариант осуществления направлен на способ получения сфингана в виде полисахарида («SPS») и/или сфингана в виде олигосахарида («SOS»).
[0144] Способ получения SPS включает: гидратацию встречающегося в природе HA/IA/LA сфингана в воде и уменьшение молекулярной массы встречающегося в природе HA/IA/LA сфингана с помощью гомогенизации, обработки ультразвуком, облучения, окисления и/или гидролиза.
[0145] Уменьшение молекулярной массы (т.е. уменьшение длины цепи) встречающегося в природе HA/IA/LA сфингана может быть достигнуто, используя гомогенизацию под высоким давлением, с помощью способа, который включает: (i) гидратацию порошка продукта HA/LA сфингана в деионизированной воде с получением раствора гидратированного HA/IA/LA сфингана (приблизительно 1% (в отношении веса к объему)); (ii) пропускание раствора гидратированного HA/IA/LA сфингана через гомогенизатор от 1 до 10 раз при рабочем давлении от приблизительно 8500 фунтов на квадратный дюйм до приблизительно 12000 фунтов на квадратный дюйм (и все значениями между ними) с получением раствора гомогенизированного HA/IA/LA SPS; (iii) добавление достаточного количества подходящего органического растворителя к гомогенизированному раствору с получением преципитата HA/IA/LA SPS; (iv) сбор преципитата HA/IA/LA SPS с помощью центрифугирования; и (v) высушивание и измельчение собранного порошка HA/IA/LA SPS.
[0146] В одном аспекте способа получения HA/IA/LA SPS, HA/IA/LA сфинган может представлять собой, например, высокоацильный геллан, среднеацильный геллан, низкоацильный геллан, высокоацильный диутан, среднеацильный диутан, низкоацильный диутан, высокоацильный рамсан, среднеацильный рамсан и низкоацильный рамсан. В другом аспекте способа получения HA/IA/LA SPS, указанное пропускание совершается 1-10 раз (например, 1, 2, 3, 4 и т.д.) при давлении, составляющем приблизительно 8500 фунтов на квадратный дюйм. В еще одном аспекте способа получения SPS, указанное пропускание совершается 1-10 раз (например, 1, 2, 3, 4 и т.д.) при давлении, составляющем приблизительно 12000 фунтов на квадратный дюйм. В дальнейшем аспекте способа получения SPS, указанное пропускание совершается 10 раз при давлении, составляющем приблизительно 12000 фунтов на квадратный дюйм. И, в еще одном аспекте способа получения SPS, подходящим органическим растворителем является тот, который активирует преципитацию полученного таким образом HA/IA/LA сфингана в виде полисахарида, в том числе, например, изопропанол.
[0147] Способ получения HA/IA/LA SOS включает: приготовление первой композиции, содержащей встречающийся в природе HA/IA/LA сфинган или HA/IA/LA SPS и жидкую среду; гидролиз гликозидной связи HA/IA/LA сфингана или HA/IA/LA SPS с получением второй композиции; подвергание второй композиции ультрафильтрации, гель-фильтрации, преципитации, центрифугированию, или их комбинации с получением третьей композиции, содержащей HA/IA/LA SOS; и, необязательно, выделение или извлечение третьей композиции с помощью подходящего метода, такого как, например, лиофилизация.
[0148] В одном аспекте способа получения HA/IA/LA SOS, указанного гидролиза можно добиться при помощи кислоты, фермента, обработки ультразвуком, гомогенизации под высоким давлением, облучения или их комбинации.
[0149] В одном аспекте способа получения HA/IA/LA SOS, указанного гидролиза можно добиться при помощи водной среды с pH от приблизительно 1 до приблизительно 3. В другом аспекте указанного гидролиза можно добиться при помощи водной среды с pH от приблизительно 1 до приблизительно 3 (или pH, равном приблизительно 2), причем указанная водная среда может содержать подходящую неорганическую или органическую кислоту. Примеры подходящих кислот включают, но без ограничения этим, серную кислоту, соляную кислоту, азотную кислоту, фосфорную кислоту, лимонную кислоту, щавелевую кислоту, муравьиную кислоту, уксусную кислоту, трифторуксусную кислоту, или их комбинацию.
[0150] В одном аспекте способа получения HA/IA/LA SOS, указанного гидролиза можно добиться гидролизом муравьиной кислотой при pH, равном приблизительно 2, и температуре, равной приблизительно 95οC, в течение периода времени, достаточного для гидролиза гликозидной связи в HA/IA/LA сфингане или HA/IA/LA SPS.
[0151] В одном аспекте способа получения HA/IA/LA SOS, указанного гидролиза можно добиться при помощи фермента, причем фермент способен расщеплять одну или более гликозидных связей в сфингане, включающего, но без ограничения этим, гелланaзу, рамногалактуронанэндолиазу (EC 4.2.2.23), рамногалактуронанэкзолиазу (EC 4.2.2.24), гелланлиазу (EC 4.2.2.25), описанную Hashimoto, гелланлиазу, описанную Kennedy (1994), или их комбинацию. Понятно, что выражение «гелланaза» относится к ферменту, который способен расщеплять одну или более гликозидных связей в сфингане.
[0152] В одном аспекте способа получения HA/IA/LA SOS, указанное подвергание включает фильтрование второй композиции через мембрану с отсечением по молекулярной масса, составляющим или приблизительно 5 кДа, или приблизительно 10 кДа, с получением фильтрата, включающего третью композицию.
[0153] Пятнадцатый вариант осуществления направлен на композицию, содержащую сфинган в виде олигосахарида, приготовленного согласно четырнадцатому варианту осуществления.
[0154] Кроме особо оговоренных случаев, все технические и научные термины, используемые здесь, имеют то же значение, в котором они обычно понимаются специалистом со средним уровнем компетентности в данной области техники. Следующие примеры предназначены только для дополнительной иллюстрации вариантов осуществления, заявленных и раскрытых здесь, и, как подразумевается, не ограничивают объем заявленного объекта изобретения.
Примеры
I. Пример 1. Получение HA/LA SPS и SOS
Получение сфингана в виде полисахарида
[0155] Длину цепи встречающегося в природе сфингана можно уменьшить, используя гомогенизацию под высоким давлением, с помощью способа, который включает: (i) гидратацию порошка продукта сфингана (например, геллана, диутана и рамсана) в 1 л деионизированной воды в концентрации, составляющей 1% (в отношении веса к объему), с получением раствора гидратированного сфингана; (ii) механическую обработку раствора гидратированного сфингана в гомогенизаторе APV Model 1000 при приблизительно 12000 фунтов на квадратный дюйм (x 10) с получением гомогенизированных растворов; (iii) добавление достаточного количества изопропилового спирта к гомогенизированному раствору с получением преципитата сфингана в виде полисахарида; (iv) сбор преципитата сфингана в виде полисахарида с помощью центрифугирования; и (v) высушивание и измельчение собранного порошка сфингана в виде полисахарида. Используя этот способ по отношению к выбранным сфинганам (например, высокоацильному геллану, низкоацильному геллану, высокоацильному диутану и высокоацильному рамсану), были получены следующие образцы сфинганов в виде полисахаридов, представленные в таблице 1, которые были подвергнуты гидратации в воде в концентрации, составляющей 0,8% (в отношении веса к объему), для последующих исследований.
Таблица 1. Сводка сфинганов в виде полисахаридов (Образцы с №№ 1-7)
aОбразец с № 6 представляет собой геллан в виде полисахарида, полученного из геллана, который был получен из штамма 438, штамма, происходящего от Sphingomonas elodea дикого типа. Геллан из штамма 438 был выделен путем обработки ферментационного бульона протеазой, EDTA, SDS, лизоцимом и глюкоамилазой; с последующим извлечением геллановой камеди с помощью индуцированной изопропанолом преципитации обработанного и нагретого бульона.
bОбразец с № 7 представляет собой геллан в виде полисахарида, полученного из геллана, который был получен из штамма 438. Геллан из штамма 438 был выделен путем центрифугирования ферментационного бульона для получения осажденных клеток и супернатанта, обработки собранного супернатанта глюкоамилазой и протеазой, и извлечения геллана из нагретого бульона с использованием преципитации изопропанолом.
[0156] Полученный здесь геллан в виде полисахарида отличается от коммерчески выпускаемых продуктов гелланов тем, что длина цепи уменьшена путем гомогенизации под высоком давлением. Действительно, предыдущие исследования показали, что гомогенизация под высоким давлением уменьшает длину цепи (и, таким образом, молекулярную массу)) встречающегося в природе геллана. (Смотрите US6242035B1, который включен посредством ссылки в его полном объеме, причем гомогенизация под высоким давлением природной геллановой камеди (MW ≈ 2,5×106; MN ≈ 2,2×106) дает геллановую камедь с MW, меньшей или равной приблизительно 1,7×106, как определено с помощью гель-фильтрации/рассеивания лазерного излучения с кратными углами.)
Получение сфинганов в виде олигосахаридов (SOS)
[0157] Получение SOS, как правило, включает: (i) приготовление 2% (в отношении веса к объему) раствора встречающегося в природе HA/IA/LA сфингана (или HA/IA/LA SPS); (ii) гидролиз муравьиной кислотой (pH 2) при 95οC, в течение ночи с получением гидролизата; (iii) фильтрование гидролиза, используя мембрану для ультрафильтрации с отсечением по молекулярной массе, составляющим или 5 кДа, или 10 кДа, с получением фильтрата; (iv) лиофилизацию фильтрата с получением лиофилизата; (v) промывку лиофизитата безводным этанолом (x 3) с получением промытого порошка; и (vi) высушивание промытого порошка с получением SOS. (Альтернативно, гидролиз может протекать при использовании: (i) подходящего фермента, такого как гелланaза; (ii) обработки ультразвуком; (iii) гомогенизации под высоким давлением; (iv) облучения; или (v) других известных способов). Используя процедуру кислотного гидролиза (например, с использованием муравьиной кислоты) или ферментативный (например, с использованием гелланазы Japan (EC 4.2.2.25) или гелланазы из штамма 438), были получены следующие образцы HA/LA сфинганов в виде олигосахаридов, представленные в таблице 2a, в которой содержание моносахаридов в процентах относится к содержанию моносахаридов (глюкозы и рамнозы), разделенному на концентрацию образца, состав моносахаридов, содержание олигосахарида и молекулярная масса являются теми, что описаны ниже.
Таблица 2a. Сводка сфинганов в виде олигосахаридов (образцы с №№ 8-18)
Состав моносахаридов: Rha, Glc, GlcA, Glyc в приблизительном соотношении 3:5:2:2 с неизвестной уроновой кислотой, которая присутствует.
Содержание олигосахаридов: гелланоподобные олигосахариды (как ацетилированные, так и глицеринированные), имеющие DP от приблизительно 2 до приблизительно 9.
Молекулярная масса: молекулярная масса образца от приблизительно 0,5 кДа до приблизительно 4 кДа (наблюдается двойной пик, соответствующий размеру меньше и больше приблизительно 1,2 кДа).
Состав моносахаридов: Rha, Glc, GlcA в приблизительном соотношении 3:5:2. Только ничтожные количества глицерата.
Содержание олигосахаридов: гелланоподобные олигосахариды (в основном неэстерифицированные), имеющие DP от приблизительно 2 до приблизительно 12.
Молекулярная масса: молекулярная масса образца от приблизительно 0,5 кДа до приблизительно 4 кДа (наблюдается двойной пик, соответствующий размеру меньше и больше 1,2 кДа).
Состав моносахаридов: соотношение Rha и Glc - приблизительно 1 к приблизительно 2 (ничтожные количества глюкуроновой кислоты и большие количества неизвестного соединения 1 и 2).
Содержание олигосахаридов: приблизительно 50% ненасыщенного GlcA, Glc,Rha, Glc и приблизительно 10% ненасыщенного GlcA, Glc,Rha.
Молекулярная масса: узкое распределение по размеру - приблизительно 1 кДа
Состав моносахаридов: Rha, Glc, GlcA, Glyc в приблизительном соотношении 3:5:2:2 с неизвестной уроновой кислотой, которая присутствует.
Содержание олигосахаридов: гелланоподобные олигосахариды (как ацетилированные, так и глицеринированные), имеющие DP от приблизительно 2 до приблизительно 9, с незначительными количествами ненасыщенных соединений.
Молекулярная масса: наблюдаемая молекулярная масса образца от приблизительно 0,5 кДа до приблизительно 4 кДа (наблюдается двойной пик, соответствующий размеру меньше и больше приблизительно 1,2 кДа, как у SN9).
[0158] Содержание моносахаридов определяли для выбранных образцов SOS путем растворения образца SOS в деионизированной воде и анализа содержания глюкозы и рамнозы, используя систему для ионной хроматографии Thermo Fisher. Общее содержание моносахаридов рассчитывается как общая концентрация глюкозы и рамнозы, разделенная на концентрацию образца.
[0159] Состав моносахаридов (для образцов с № 9-10 и 17-18) определяли следующим образом. Образцы SOS растворяли в 4% -ной серной кислоте до концентрации 3,5 г/л и автоклавировали при 121οC в течение одного часа. Количество моносахаридов определяли с помощью системы Dionex ICS-5000 в соответствии с Zeuner (2016). Глицерат количественно определяли с использованием внешних стандартов. Неизвестное соединение 1 («UNK1»), неизвестное соединение 2 («UNK2») и неизвестную уроновую кислоту 1 («UNK URON1») количественно определяли как единицы глюкуроновой кислоты. В таблице 2b суммируется состав моносахаридов для каждого из образцов с № 9-10 и 17-18
Таблица 2b. Сводка сфинганов в виде олигосахаридов (состав моносахаридов, образцы с №№ 9-10 и 17-18
[0160] Содержание олигосахаридов (для образцов c № 9-10 и 17-19) определяли следующим образом. Идентификацию и определение относительного количества олигосахаридов проводили с помощью жидкостной хромато-масс-спектрометрии с электрораспылительной ионизацией (LC-ESI-MS) на ионной ловушке Amazon SL (Bruker Daltonics, Bremen, Германия) в сочетании с UHPLC UltiMate 3000 (Dionex, Sunnyvale, CA, США). 5 мкл образца в 50% ACN (конечная концентрация=5 г/л) вводили в колонку TSKgel Amide 80 HILIC (150 мм x 2 мм; 2 мкм, TOSOH, Greisheim, Германия). Хроматографию проводили со скоростью 0,2 мл/мин при 45οC в системе с тремя элюентами, состоящими из элюента A (воды), элюента B (ацетонитрила) и C (100 мМ формиата аммония, pH 5). Элюент C все время поддерживался на уровне 5%. Профиль элюирования был следующим (время указано в минутах): 0-5, изократический 75% B; 5-25, линейный градиент до 25% B; 25-30, изократический 5% B; 30-40, изократический 75% B. Электрораспыление работало в отрицательном режиме с режимом UltraScan и диапазоном сканирования от 100 до 2000 m/z (масса/заряд), интеллектуальная настройка параметров - 1000 m/z. Автоматические события тандемной масс-спектрометрии (MS2) были выполнены для двух наиболее распространенных ионов-предшественников. Капиллярное напряжение - 4,5 кВ, смещение концевой пластины - 0,5 кВ, давление в распылителе - 3,0 бар, поток сухого газа - 12,0 л/мин и температура сухого газа - 280οC. Соединения идентифицировали с помощью MS (масс-спектрометрии) и MSn и количественно оценивали по относительной интенсивности при анализе данных 4.2 SR2
Таблица 2c. Сводка сфинганов в виде олигосахаридов (содержание олигосахаридов, образцы с №№ 9-10 and 17-18
[0161] SOS, идентифицированный как Glc2,GlcA, Rha,Glyc, представляет собой тетрамерную единицу геллана с одним глицератом, тогда как SOS, идентифицированный как Glc2,GlcA, Rha,Ac, представляет собой тетрамерную единицу геллана с одним ацетилом. Некоторые SOS включают множество сахарных компонентов (а именно, Glc5,GlcA2,Rha2,Ac) - олигосахарид можно установить исходя из определенного числа сахаридов. Например, Glc5,GlcA2,Rha2,Ac включает две тетрамерные единицы (а именно, Glc-GlcA-Glc-Rha) с дополнительным глюкопиранозилом (Glc) и ацетилом (Ac). Кроме того, Glc6,GlcA3,Rha3 представляет собой олигосахарид, который включает три тетрамерные единицы (а именно, 3Х Glc-GlcA-Glc-Rha). SOS, идентифицированные по утрате воды («-H2O», смотрите, например, Glc2,GlcA, Rha,Glyc.-H2O), представляют собой ненасыщенный продукт лиазы/β-излучения. В некоторых случаях для более длинных гелланоподобных олигомеров (например, Glc4,GlcA3,Rha2/Glc4,GlcA2,Rha2,Glyc2), предлагаются две структуры, поскольку масс-спектральная фрагментация недостаточна, чтобы отличить присутствие одной глюкуроновой кислоты от двух глицератных заместителей. Не все наблюдаемые SOS могли быть структурно идентифицированы. На основе фрагментации некоторые компоненты можно частично идентифицировать благодаря схожести паттернов фрагментации, поэтому они обозначены как «производное с неизвестным отношением m/z» после наиболее сходного с ним идентифицированного соединения. Другие соединения было невозможно идентифицировать вследствие слабой фрагментации или вследствие того, что они были соединениями другого типа, чем ожидаемые SOS, происходящие из геллана. Эти неизвестные SOS обозначены как «неизвестный (наблюдаемое отношение m/z) z1 или z2» в зависимости от того, наблюдался ли один или два заряда. Как свидетельствуют данные гель-фильтрации, ниже, проанализированные образцы могут включать сфинганы в виде полисахаридов (DP >30, но меньше, чем у встречающегося в природе сфингана и сфинганы в виде олигосахаридов (2≥DP≤30).
[0162] Представленную молекулярную массу образцов SOS определяли следующим образом. Высокоэффективную гель-фильтрация выполняли с использованием насоса Ultimate iso-3100 SD с пробоотборником WPS-3000 (Dionex), подключенным к детектору показателя преломления RI-101 (Shodex). 100 мкл образца загружали в колонку TSKgel G3000PW (300×7,5 мм), оснащенную защитной колонкой TSKgel PWH (7,5×7,5 мм) (Tosoh Bioscience). Элюцию выполняли с использованием 100 мМ нитрата натрия со скоростью потока=1,0 мл/мин при 40οC. В качестве эталонов использовали стандарты пуллуланов.
[0163] На фиг. 1а представлена полученная в ходе гель-фильтрации хроматограмма («SEC») для обработанных кислотой (SN9, сплошная линия) и ферментом (SN18, пунктирная линия) сфинганов в виде поли- и олигосахаридов, полученных из высокоацильного геллана, тогда как на фиг. 1b представлена SEC для обработанных кислотой (SN10, сплошная линия) и ферментом (SN17, пунктирная линия) сфинганов в виде поли- и олигосахаридов, полученных из низкоацильного геллана. Как на фиг. 1а, так и на фиг. 1b показано время элюирования стандартов молекулярной массы пуллуланов (а именно, >50 кДа (6,5 мин, закрашенный квадрат ( )), 12 кДа (8,8 мин, закрашенный кружок ( )), 5 кДа (9,3 мин, закрашенный треугольник ( )), 1 кДа (10 мин, пустой квадрат ( )), 342 Да (10,65 мин, пустой кружок ( )), и 180 Да (11,15 мин, пустой треугольник ( ))). Данные SEC для фиг. 1a показывают сопоставимое распределение сфинганов в виде полисахаридов (SPS) и сфинганов в виде олигосахаридов (SOS), полученных из высокоацильного сфингана. Их следует сравнить с данными SEC для фиг. 1b, где распределения SPS и SOS для обработанного кислотой образца (SN10) отличаются от распределений SPS и SOS для обработанного ферментом образца (SN17). Данные SEC также показывают диапазон молекулярной массы от приблизительно 0,5 кДа до приблизительно 4 кДа (и, возможно, до приблизительно 12 кДа) для образцов с №№ 9, 10 и 18. Интересно, что образец (SN17), полученный из низкоацильного сфингана с помощью обработки ферментом, демонстрирует начальную элюцию SOS, имеющих молекулярную массу в диапазоне от приблизительно 0,5 кДа до приблизительно 1 кДа (с узким распределением по размеру пика - приблизительно 1 кДа).
[0164] Содержание олигомеров в образцах SOS определяли с помощью масс-спектрального анализа. Как правило, образец SOS готовили путем растворения SOS в концентрации 0,4% с использованием смеси вода/ацетонитрил (1:1), содержащей 1 мМ NaCl. Образцы фильтровали через 0,22-микронный фильтр перед введением в MSQ plus Single Quad Mass Spec от компании Thermo Fisher. Масс-спектрометр работал в режиме отрицательной ионизации электрораспылением, сканировании от 150 до 1000 m/z. Исходя из интактной массы олигомеров, в образцах SOS были обнаружены различные олигосахариды. В таблице 3 суммированы олигомеры, наблюдаемые для выбранных образцов SOS
Таблица 3. Идентифицированные олигомеры в образцах SOS
II. Пример II. Эффект сфинганов (например, встречающихся в природе сфинганов, SPS и SOS) на активность выбранных проб кишечной микробиоты
[0165] Образцы, содержащие 8 мг/мл (0,8% (в отношении веса к объему) SPS или SOS, разводили в два раза для обеспечения образцов, содержащих 4 мг/мл (0,4% (в отношении веса к объему)) SPS или SOS. Эффект образцов с №№ 1-12 с концентрацией 4 мг/мл на панели из более чем двадцати пяти проб кишечной микробиоты оценивали после ферментации в течение 24 часов, используя платформу для скрининга ферментации in vitro («i-screen»), как описано Fehlbaum (2018). Конкретно, использовали стандартный пул проб каловой микробиоты, полученных от 5-6 здоровых взрослых (состояние здоровья на основе критериев исключения), который предварительно культивировали в течение ночи из замороженного материала. После этого было выполнено разведение в титрационных микропланшетах, в которые образцы были добавлены и впоследствии подвергнуты инкубации в анаэробных условиях в течение 24 ч при 37οC. После инкубации образцы культуры собирали и обрабатывали для дальнейшего анализа. В 96-луночном планшете некоторые лунки использовали для технических контролей, контроля без микробиоты (n=3) и отрицательного контроля только с микробиотой (n=3), оставляя 80 лунок доступными для экспериментов. Ряд образцов сфинганов и образцы для сравнения - экстракты растений (например, пектины, пектиновые олигосахариды и каррагенаны) и биогуммы (например, ксантан и ксантаны-олигосахариды)) анализировали в концентрации, составляющей приблизительно 4 мг/мл, что соответствует дозе, составляющей приблизительно 4 г/день. (Van den Abbeele (2011).)
[0166] Изменения в составе микробиоты определяли с помощью секвенирования нового поколения, которое распознает бактерии на уровне рода и во многих случаях (но не во всех) на уровне вида. Для равномерного распределения образцов в пуле для секвенирования, общую бактериальную загрузку устанавливали с помощью количественной полимеразной цепной реакции («ПЦР»), используя универсальный набор праймер-зонд. Ампликоны области V4 16S рДНК получали с помощью ПЦР, тем самым стандартизируя уровень матричной ДНК и используя уникальные корректирующие ошибки праймеры со штриховым кодированием, и избегая чрезмерной амплификации. Затем ампликоны очищали в геле, определяли количественно и объединяли. Затем анализ последовательности выполняли в устройстве lllumina MiSeq® путем секвенирования спаренных концов (2×250 п.о.). Анализ 3’ последовательностей выполняли с использованием конвейера стандартизированного секвенирования, разработанного организацией прикладных научных исследований Нидерландов. Конвейер предусматривает сборку считываний спаренных концов, качественную фильтрацию, удаление химер и таксономическую классификацию+кластеризацию обработанных считываний.
[0167] Стандартные контроли выполняли в трех повторах. В частности, панель микроорганизмов включала Bacteroides, Coprococcus, Lachnospiraceae unclassified, Megasphaera, Escherichia/Shigella, Clostridium XlVa, Allisonella, Bifidobacterium, Dorea, Collinsella, Mogibacterium, Sutterella, Bilophila, Blautia, Clostridium sensu stricto, Phascolarctobacterium, Faecalibacterium, Clostridium XlVb, Clostridium XI, Acidaminococcus, Gemmiger, Lachnospira, Parabacteroides, Paraprevotella и Butyricicoccus. Эффект определяли относительно необработанного контроля. Для первого анализа с использованием «i-screen» образцов с №№ 1-12 в таблице 4a суммируется наблюдаемый эффект на рост Bifidobacterium и Faecalibacterium, где представленные результаты относятся к необработанному контролю (рост обозначен как 1,.0).
Таблица 4а. Данные по активности выбранных проб кишечной микробиоты, наблюдаемой для образцов с №№ 1-12
[0168] Выделенные жирным шрифтом значения представляют значительные изменения бактериального роста по сравнению с необработанным контролем. В соответствии с результатами, представленными в примере III (ниже), сфинганы в виде полисахаридов (а именно, образцы с №№ 1-3) стимулировали рост Bifidobacterium. Удивительно, что сфинганы в виде олигосахаридов (а именно, образцы с №№ 8-12) стимулировали рост каждой из Faecalibacterium, Blautia, и Parabacterioides в значительной степени по сравнению с необработанным контролем. Хорошо известно, что Bifidobacterium и Faecalibacterium (например, Faecalibacterium prausnitzii) являются бактериями, продуцирующими бутират. Соответственно, полученные с использованием i-screen результаты, показывающие, что сфинганы в виде олигосахаридов стимулируют рост Faecalibacterium, говорят о том, что эти композиции проявляют пребиотическую активность. И, поскольку Faecalibacterium prausnitzii, как известно, ассоциируется с противовоспалительным действием, полученные с использованием i-screen результаты наводят на мысль, что SOS действуют в качестве противовоспалительных средств в результате стимуляции роста Faecalibacterium prausnitzii.
[0169] Дополнительные анализы с использованием i-screen были выполнены на образцах с №№ 9, 10, 17 и 18, используя три различных каловых пула, а именно, два пула, полученных от здоровых взрослых, (H1 и H2) и одного пула, полученного от пациентов с синдромом разраженного кишечника («IBD»), для трех или четырех бактерий (а именно, Blautia, Parabacteroides, Faecalibacterium, Clostridium XVIII). Конкретно, использованные каловые пулы включают: (i) пул H1 был получен от шести здоровых добровольцев (европеоидной расы, 25-60 лет, европейского образа жизни и питания, с самооценкой состояния здоровья, без применения антибиотиков в течение последних 3 месяцев), (ii) пул H2 был получен от 5 здоровых взрослых добровольцев (20-65 лет, без применения антибиотиков в течение последних 3 месяцев, самооценкой состояния здоровья) и (iii) пул IBD был получен от четырех пациентов с IBD, а именно с язвенным колитом. Для дополнительных анализов с использованием i-screen (а именно, первого i-screen (№ 1-2), второго i-screen (№ 3-4) и третьего i-screen (№ 5-16)) образцов с №№ 9-10 и 17-18, в таблице 4b суммируется наблюдаемый эффект на рост трех или четырех бактерий (а именно, Blautia («Blaut.»), Parabacteroides («Para.»), Faecalibacterium ("Faecal."), Clostridium XVIII («ClXVIII»)), где представленные результаты относятся к необработанному контролю (рост обозначен как 1,0).
Таблица 4b. Данные по активности выбранных проб кишечной микробиоты, наблюдаемой для образцов с №№ 9-10 и 17-18.
bОтветные реакции Faecal. Являются теми, которые представлены в таблице 5.
[0170] Что касается элементов 1-4 в таблице 4b, можно видеть, что SOS создавали благоприятные условия для увеличения кратности изменения Blautia, Parabacteroides и Faecalibacterium в каловых пулах здоровых взрослых по сравнению с необработанными контролями. SOS, полученные из низкоацильных гелланов-олигомеров, продемонстрировали наибольшие значения кратности изменения Faecalibacterium (188.91), а также Parabacteroides (21.11) и Blautia (4.78)
[0171] Что касается элементов 5-8 в таблице 4b, были сделаны следующие наблюдения. SOS (обработанные кислотой и ферментом) стимулировали рост Blautia, Parabacteroides и Faecalibacterium в каловых пулах здоровых взрослых по сравнению с необработанными контролями. Эти результаты подтверждают результаты первого и второго i-screen для образцов обработанных кислотой высоко- и низкоацильных гелланов в виде олигомеров. Большие значения кратности изменения в трех родах были получены с обработанными кислотой низкоацильными гелланами в виде олигомеров. Кроме того, высоко- и низкоацильные гелланы в виде олигомеров, полученные с помощью ферментативной обработки, также были эффективны для увеличения роста трех родов в одном и том же каловом пуле.
[0172] Используя каловый пул H2 (элементы 9-12 таблицы 4b), можно видеть, что SOS (обработанные кислотой и ферментом) увеличивали кратность изменения Blautia, Parabacteroides, Faecalibacterium и бактерий из кластера XVIII Clostridium. SOS (обработанные кислотой) создавали благоприятные условия для наибольших значений кратности изменения Faecalibacterium (21.93) и Parabacteroides (7.25). Значения кратности изменения кластера XVIII Clostridium варьировали от 12,66 до 55,91. Для сравнения: большая часть кластеров XVIII и XIVa Clostridium, обнаруженных в кишечнике, продуцирует ацетат (несколько штаммов в кластере XIVa Clostridium также продуцируют бутират вместе с ацетатом), также на основе геномного анализа (метаболической сети) оба кластера не образуют токсины. Narushima (2014).
[0173] Используя каловый пул IBD (элементы 13-16 таблицы 4b), результаты показывают, что SOS (из низкоацильного геллана) создавали благоприятные условия для наибольших значений кратности изменения Parabacteroides. Обработанные кислотой низкоацильные гелланы-олигомеры продемонстрировали наибольшее значение кратности изменения Faecalibacterium. Кратность изменения кластера XVIII Clostridium также увеличивалась с 30,84 до 98,28. Интересно отметить значительный рост Parabacteroides в случае SOS. Parabacteroides расщепляют здоровую пищу с высоким содержанием клетчатки, они защищают от воспалений. Эти бактерии отсутствуют у пациентов, страдающих воспалительными заболеваниями кишечника. Martínez (2010), Noor (2010), Segata (2012), и Zitomersky (2013).
[0174] В таблице 5 суммируется наблюдаемый эффект для второго анализа с использованием i-screen образцов с №№ 9-16 (и образцов для сравнения 1-16, а также добавки Livaux™, инулина и амоксициллина) на панели из восьми бактерий (Lachnospiraceae неклассифицированные («Lachn.U.»), Clostridium XlVa («ClX1Va»), Bifidobacterium («Bifid.»), Coprococcus («Copro.») Blautia («Blaut.»), Phascolarctobacterium, («Phasc.»), Faecalibacterium («Faecal.»), Butyricicoccus («Butyr.»), и Parabacteroides («Para.»)), где представленные результаты относятся к необработанному контролю (рост обозначен как 1,0).
Таблица 5. Эффект выбранных SOS (SN9-SN16) и образцов для сравнения (CS) по отношению к панели из восьми бактерий.
[0175] В таблице 6 суммируется композиционный состав образцов для сравнения 1-16, использованных во втором скрининге.
Таблица 6. Сводка образцов для сравнения («CS») 1-16
[0176] На основе результатов таблицы 5 можно видеть, что все сфингана в виде олигосахаридов продемонстрировали наибольший рост Faecalibacterium по сравнению со всеми образцами для сравнения. В частности, наибольший рост Faecalibacterium продемонстрировали гелланы-олигосахариды (приблизительно в 52 раза), полученные из геллана GELRITETM MK (отсечение по молекулярной массе - 5 кДа (SN10)), рамсаны-олигосахариды (приблизительно в 49 раз), полученные из встречающегося в природе рамсана (SN12), и гелланы-олигосахариды (в 43 разе), полученные из GELRITE™ MK (отсечение по молекулярной массе - 10 кДа (SN14)). Интересно, что продукт Livaux™, который рекламируется как обладающий стимулирующей рост Faecalibacterium активностью (смотрите, например, livaux.com/livaux-gi-problem/), продемонстрировал только 1,14-кратное увеличение активности роста Faecalibacterium по сравнению с необработанным контролем. Относительно низкая активность роста Faecalibacterium для продукта Livaux™ согласуется с опубликованными данными. (US20170326190A1).
[0177] Сравнение результатов из таблицы 4 и таблицы 5 показывает, что в некоторых случаях, вариабельность для выбранных образцов (сравните SN10 (188,31 против 51,54) и SN12 (70,44 против 48,73)). Дополнительный анализ выбранных данных показывает, что коэффициент вариации (а именно, отношение стандартного отклонения к среднему значению) для выбранных сфинганов может варьироваться от приблизительно 7% до приблизительно 32%, а в некоторых случаях до приблизительно 80%.
[0178] На основе результатов таблицы 5 можно видеть, что все сфинганы в виде олигосахаридов продемонстрировали увеличение активности роста Blautia (а именно, 2-5-кратное увеличение по сравнению с необработанным контролем).
[0179] Не приведенные данные показывают, что все сфинганы в виде олигосахаридов продемонстрировали снижение активности роста Escherichia/Shigella (снижение на приблизительно 9-36% по сравнению с необработанным контролем). Они будут отличаться от продукта Livaux™, который продемонстрировал увеличение активности роста Escherichia/Shigella (увеличение на приблизительно 45% по сравнению с необработанным контролем).
III. Пример III. Эффект геллановой камеди на активность и состав кишечного микробиома в просвете и слизистой оболочке желудочно-кишечного тракта человека.
А. Материалы и методы, план эксперимента SHIME и типичное устройство ферментера SHIME
[0180] Аспекты имитатора кишечной микробной экосистемы у человека (или SHIME) известны (смотрите, например, Molly (1993), Possemiers (2004), Possemiers (2017), Van de Wiele (2013), Van den Abbeele (2012), и Van den Abbeele (2013).)
[0181] Типичное устройство ферментера SHIME, представляющее желудочно-кишечный тракт взрослого человека, было описано Molly (1993). Оно состоит из последовательности пяти ферментеров, имитирующих различные части желудочно-кишечного тракта человека (например, желудок (V1), тонкую кишку (V2), восходящую ободочную кишку (V3), поперечную ободочную кишку (V4) и нисходящую ободочную кишку (V5)). Первые два ферментера работают по принципу заполнения и вытягивания для имитации различных стадий поглощения и переваривания пищи, с использованием перистальтических насосов, добавляющих определенное количество подаваемого материала SHIME (140 мл 3 раза в день), а также панкреатической жидкости и желчи (60 мл 3 раза в день), соответственно, в компартмент желудка (V1) и тонкой кишки (V2) и опорожняющих соответствующие ферментеры через определенные промежутки времени. Последние три компартмента имитируют толстую кишку. В этих ферментерах происходит постоянное перемешивание; у них есть постоянный контроль объема и pH. Время удержания и pH различных сосудов выбирают таким образом, чтобы они напоминали условия in vivo в различных частях ободочной кишки. После инокуляции каловой микробиотой эти ферментеры имитируют восходящую (V3), поперечную (V4) и нисходящую (V5) ободочную кишку. Приготовление инокулята, настройки времени удержания, pH, температуры и состав подаваемого материала в ферментере были ранее описаны Possemiers (2004). После стабилизации микробного сообщества в различных частях ободочной кишки, в трех компартментах ободочной кишки устанавливается репрезентативное микробное сообщество, которое различается как по составу, так и по функциональности в разных частях ободочной кишки.
[0182] В кишечном тракте человека обитает большое и сложное сообщество микроорганизмов, которое участвует в поддержании здоровья человека, предотвращая колонизацию патогенами и производя питательные вещества. Микроорганизмы не распределяются по кишечнику случайным образом, и те, которые прилипают к стенке кишечника, играют важную роль в качестве «барьера» против патогенов, управляя иммунными ответами слизистых оболочек и занимая нишу за счет потенциально вредных вселенцев. Однако современные стратегии in vitro не позволяют культивировать часть микроорганизмов, которые прилипают к слизистой оболочке кишечника, и ограничиваются моделированием люминального микробного сообщества. Это означает, что важная часть экосистемы кишечника не принимается во внимание, и потенциально важная информация теряется.
[0183] Для решения этой проблемы система SHIME была модифицирована с учетом колонизации слизистого слоя. (Смотрите, например, Van den Abbeele (2012) и Van den Abbeele (2013).) Модифицированная система SHIME известна как M-SHIME, которая позволяет культивировать как люминальное, так и связанное со слизью микробное сообщество в течение нескольких недель.
[0184] Включение компартмента в виде слизистой оболочки увеличивает ценность и возможности моделирования SHIME и позволяет оценить, способна ли конкретная обработка также модулировать микробное сообщество, связанное со слизистой оболочкой.
I. Адаптированное устройство SHIME для исследования
[0185] Устройство SHIME конфигурации TWINSHIME было адаптировано до конфигурации TripleSHIME, которая включала сосуд (или ферментер) для желудка, тонкой кишки, проксимального отдела ободочной кишки и дистального отдела ободочной кишки для каждого из доноров. Конфигурация TripleSHIME позволяла проводить параллельное сравнение трех различных условий. Была оценена потенциальная ферментация геллановой камеди микробиотой трех разных людей-доноров (донора A: женщины, 28 лет; донора B: женщин, 41 года; донор C: женщины, 34 лет). Части ободочной кишки ограничивались двумя частями по сравнению с тремя частями в TWINSHIME. Время удержания и диапазоны pH были оптимизированы для получения результатов, которые являются репрезентативными для имитации всего желудочно-кишечного тракта. На практике в экспериментах TripleSHIME вместо работы с 2 блоками, каждый из которых состоит из конфигурации AC-TC-DC (восходящая, поперечная и нисходящая ободочная кишка), использовали 3 блока PC-DC. После инокуляции каловой микробиотой взрослого человека эти ферментеры имитируют проксимальный отдел ободочной кишки (PC; pH 5,6-5,9; время удержания=20 часов; объем 500 мл) и дистальный отдел ободочной кишки (DC; pH 6,6-6,9; время удержания=32 часа; объем 800 мл).
[0186] Эксперимент SHIME для этого исследования состоял из трех стадий (стабилизации, контролирования и обработки), которые длились семь недель.
[0187] Период стабилизации: После инокуляции ферментеров в качестве ободочной кишки подходящим образцом кала, двухнедельный период стабилизации позволял микробному сообществу дифференцироваться в различных ферментерах в зависимости от местных условий окружающей среды. В течение этого периода в SHIME подавалась минимальная питательная матрица для поддержания максимального разнообразия кишечной микробиоты, изначально присутствующей в каловом инокуляте.
[0188] Период контролирования: В течение этого двухнедельного контрольного периода стандартный питательный матрикс SHIME дополнительно подавался дозами в модель в течение периода, составляющего 14 дней. Анализ образцов в этот период позволяет определить состав и активность микробного сообщества на исходном уровне в различных ферментерах, которые использовались в качестве контроля для оценки эффектов обработки.
[0189] Период обработки: Во время этого трехнедельного периода ферментер SHIME работал в номинальном режиме, но с использованием диеты, дополненной исследуемым продуктом. Образцы, отобранные из ферментеров в качестве ободочной кишки в этот период, позволяют исследовать специфический эффекте на состав и активность постоянно присутствующего микробного сообщества.
В. Анализ состава и активности микробного сообщества
[0190] Особенностью SHIME является возможность работать с устойчивым сообществом - микробиотой и регулярно отбирать образцы из различных частей кишечника для дальнейшего анализа. Большие объемы частей ободочной кишки позволяют собирать достаточные объемы жидкости каждый день, не нарушая микробное сообщество и не подвергая опасности остальную часть эксперимента.
[0191] Ряд параметров микроорганизмов отслеживается на протяжении всего эксперимента как часть стандартного эксперимента SHIME. Эти измерения необходимы для оценки эффективности модели и позволяют отслеживать основные изменения в составе и активности микробного сообщества, вызванные обработкой пребиотиком.
1. Анализ состава и активности микробного сообщества
[0192] Расход кислоты/основания: Продукция микроорганизмами метаболитов в ферментерах в качестве ободочной кишки изменяет pH. Без постоянного контроля pH (путем добавления кислоты или основания) pH будет превышать установленные интервалы. Расход кислоты/основания постоянно контролируют во время эксперимента SHIME.
[0193] Общее газообразование: Оценка общего газообразования является важным аспектом, связанным с потенциальными проблемами переносимости в случае конечного применения. Однако измерения газообразования в режиме онлайн затруднительны в непрерывных моделях кишечника из-за непрерывного притока и оттока масс. Поэтому общий анализ газообразования, как правило, оценивается в периодических установках.
2. Активность микробного сообщества (3 раза в неделю)
[0194] Короткоцепочечные жирные кислоты (SCFA): Анализировали концентрации уксусной кислоты, пропионовой кислоты и масляной кислоты.
[0195] Лактат: предшественник SCFA и потенциальный противомикробный агент.
[0196] Аммиак и SCFA с разветвленной цепью (изомасляная кислота, изовалериановая кислота и изокапроновая кислота) являются маркерами протеолитической ферментации, с довольно неблагоприятными эффектами на здоровье хозяина.
[0197] Состав микробного сообщества (1 раз в неделю); Отбирались образцы для нацеленного на 16S рРНК секвенирования Illumina.
C. Используемая для исследований геллановая камедь
[0198] Исследуемый продукт включал геллановую камедь пищевого качества, геллановую камедь KELCOGEL® LT100-P («Gellam Gum»). Геллановая камедь KELCOGEL® LT100-P представляет собой природную (высокоацильную) геллановую камедь. Продукт исследовали в дозе 1 г/день in vitro, что соответствует дозе 2 г/день in vivo.
D. Стабильность устройства SHIME
[0199] Во время контрольного периода уровни SCFA были очень стабильными в трех блоках SHIME (в среднем уровни были на 94,4% схожими между последовательными временными точками в контрольном периоде), что ясно указывает на стабильность микробного сообщества по показателю активности и состава. Стабильные условия в ферментерах увеличивают уверенность в том, что любой эффект, наблюдаемый во время обработки, действительно является результатом введенного исследуемого продукта.
E. Общая ферментативная активность
1. Расход кислоты/основания
[0200] Расход кислоты и основания отражает общую активность микроорганизмов на протяжении всего эксперимента SHIME. Для обеспечения сохранения оптимальных окружающих условий, pH в системе SHIME контролируется регуляторами pH в диапазоне от 5,6 до 5,9 в проксимальном отделе ободочной кишки и от 6,6 до 6,9 в дистальном отделе ободочной кишки. После стабилизации микробного сообщества в различных ферментерах (начиная с 2 недель после инокуляции), расход щелочи и кислоты является, как правило, низким. Однако во время обработки бактерии могут продуцировать увеличенные количества SCFA. Как следствие, среда в ферментерах будет подкисляться, что потребует введения основания в соответствующие ферментеры для сохранения их в предварительно установленных диапазонах pH. В результате будет увеличиваться расход кислоты/основания. Измеряя расход кислоты/основания на протяжении эксперимента, можно оценить потенциальный эффект исследуемого продукта на активность микробного сообщества. Однако следует отметить, что расход кислоты/основания является лишь грубым показателем ферментации микроорганизмами, поскольку не все кислоты, продуцируемые посредством ферментации, вызывают схожее снижение pH (кислоты с меньшей pKa, такие как ацетат, эффективно снижают pH), наряду с тем, что превращение кислот друг в друга также может влиять на pH (например, превращение ацетата/лактата в пропионат/бутират увеличивает pH). Фактическое измерение микробных метаболитов (таких как SCFA и лактат) дает более точные показания.
[0201] Данные таблицы 7 показывают, что для общей ферментация исследуемого продукта отмечались схожие тенденции по трем донорам, исследованные как в проксимальном, так и в дистальном отделе ободочной кишки.
Таблица 7. Средний еженедельный расход основания и кислоты (мл/день) во время двух контрольных периодов (C1 и C2) и трех недель обработки (TR1-TR3) для обработки геллановой камедью для трех разных доноров (A, B и C) в ферментерах в качестве проксимального отдела ободочной кишки (PC) и средний расход кислоты/основания за весь контрольный период (n=6) и период обработки (n=9).
[0202] В проксимальном отделе ободочной кишки закисление было очень ограниченным, однако на последней неделе обработки у всех исследуемых донов отмечалась тенденция к увеличению расхода основания. В дистальном отделе ободочной кишки закисление было более выраженным во время контрольного периода по сравнению с проксимальным отделом ободочной кишки. Это объясняется тем фактом, что физический перенос более кислой суспензии из проксимального отдела ободочной кишки (pH=5,6-5,9) в дистальный отдел ободочной кишки автоматически вызывает больший расход основания в этом дистальном отделе ободочной кишки для сохранения pH в правильном интервале (pH=6,6-6,9). Добавление исследуемого продукта приводило к небольшому увеличению расхода основания сразу после начала обработки для всех исследуемых доноров.
Газообразование
[0203] Так как газы являются основной конечной точкой активности ферментации кишечными микроорганизмами, изменения в газообразовании служат индикатором профиля общей ферментации. Поскольку газообразование не отслеживается в непрерывной модели SHIME, учитывая регулярную промывку свободного пространства газообразным азотом (для обеспечения анаэробиоза), газообразование оценивают во время отдельных краткосрочных периодических инкубаций. Во время таких инкубаций одинаковая доза исследуемого продукта подается в микробиоту, происходящую из проксимального отдела ободочной кишки, SHIME во время контрольного периода, таким образом имитируя процессы, происходящие в начале обработки в непрерывной модели SHIME.
[0204] Зависимые от доноров эффекты наблюдали в отношении газообразования (не представленные данные). Тогда как у донора B после обработки исследуемым продуктом наблюдалось незначительное увеличение газообразования, обработка приводила к незначительному уменьшению газообразования у других доноров. В целом, газообразование было наиболее интенсивным во временном интервале от 6 до 24 часов для всех условий. Только во временном интервале от 4 до 6 часов наблюдалось устойчивое (но умеренное) увеличение газообразования у всех доноров после обработки исследуемым продуктом, тогда как другие временные интервалы характеризовались специфическими для доноров различиями.
[0205] В целом, обработка геллановой камедью практически не влияла на газообразование кишечной микробиотой у трех исследуемых доноров.
F. Анализ активности микробного сообщества
Продукция короткоцепочечных кислот (SCFA)
[0206] Следующая информация описывает эффект исследуемого продукта на продукцию SCFA в эксперименте Triple-SHIME. Продукция SCFA является результатом метаболизма углеводов в ободочной кишке и связана с различными последствиями для здоровья. Наиболее распространенными SCFA являются ацетат, пропионат и бутират. Хорошо известно, что SCFA играют решающую роль в здоровье кишечника. Ацетат может использоваться в качестве источника энергии для хозяина и в качестве потенциального субстрата для синтеза липидов в организме. Кроме того, он является важным побочным продуктом синтеза бутирата и может оказывать противомикробное действие против патогенов. Однако укрепляющие здоровье эффекты в основном приписываются пропионату и бутирату, которые действуют как основные источники энергии для эпителия кишечника и продемонстрировали защитные действия против воспалений и рака ободочной кишки. Cumminngs (1987). Известно, что пропионат переносится в печень, где он оказывает снижающий уровень холестерина в плазме эффект и положительно влияет на гликемический контроль. (Смотрите Wright (1990), Demigne (1995) и Wong (2006)).
[0207] Итак, положительные эффекты исследуемых субстратов на продукцию SCFA включают, следовательно, увеличение продукции ацетата, пропионата и/или бутирата. Приведенная ниже информация учитывает прямое сравнение результатов для трех доноров.
[0208] Для оптимального сравнения различных доноров, средние уровни SCFA для всех трех из них представлены для каждой из различных SCFA (за неделю и за период).
2. Продукция ацетата
[0209] Ацетат может продуцироваться широким рядов кишечных микроорганизмов, включая, среди многих других, Bacteroides spp. (тип Bacteroidetes) и Bifidobacteria. Отсюда следовало, что, хотя геллановая камедь значительно повышала уровни ацетата в проксимальном отделе ободочной кишки донора A и C, на уровни ацетата у донора B не оказывалось влияние (фиг. 2a, таблица 8). Наибольшее среднее повышение наблюдалось у донора А (т.е. повышение, составляющее 1,7 мМ или +21%). Напротив, в дистальном отделе ободочной кишки повышенные уровни ацетата после обработки геллановой камедью наблюдались только у донора B (фиг. 2b, таблица 8), со средним повышением, составляющим 2,1 мМ (+ 6%) по сравнению с контрольным периодом.
Таблица 8. Эффект обработки геллановой камедью на продукцию ацетата (в мМ) в ферментерах в качестве проксимального (PC) и дистального (DC) отделов ободочной кишки для трех разных доноров (A, B и C) и на среднюю еженедельную продукцию ацетата во время контрольных периодов (C1 и C2) и недель обработки (TR1-TR3) (смотрите также фиг. 1a-1b).
3. Продукция пропионата
[0210] Пропионат может продуцироваться широким рядов кишечных микроорганизмов, самыми распространенными продуцентами пропионата являются Bacteroides spp. (тип Bacteroidetes), Veillonella (тип Firmicutes) и Akkermansia muciniphila (тип Verrucomicrobia). Для всех трех исследуемых доноров введение геллановой камеди приводило к значительному снижению уровней пропионата в ответ на обработку обеих частей ободочной кишки (фиг. 3a-3b, таблица 9). В проксимальном отделе ободочной кишки сильное немедленное снижение наблюдалось для доноров A и B, тогда как эффект был менее выраженным для донора C (т.е. понижение, составляющее 1,7 мМ (-8%), для донора C по сравнению с -4,3 мМ (-18%) и -4,8 мМ (-20%) для доноров A и B, соответственно). С другой стороны, в дистальном отделе ободочной кишки у всех доноров наблюдалось более постепенное снижение уровней пропионата. Эти результаты являются удивительными с учетом исследований Edwards (1995) и Anderson (1988). Например, Edwards (1995) утверждал, что у крыс линии Wistar геллановая камедь не оказывала устойчивый эффект на содержание SCFA, тогда как Anderson (1988) сообщает, что прием внутрь большого количества геллановой камеди приводил к снижению содержания пропионата в кале у женщин-добровольцев на 23% и повышению содержания пропионата в кале у мужчин-добровольцев на 33%.
Таблица 9. Эффект обработки геллановой камедью на продукцию пропионата (в мМ) в ферментерах в качестве проксимального (PC) и дистального (DC) отделов ободочной кишки для трех разных доноров (A, B и C) и на среднюю еженедельную продукцию пропионата во время контрольных периодов (C1 и C2) и недель обработки (TR1-TR3) (смотрите также фиг. 2a-2b).
4. Продукция бутирата
[0211] Бутират продуцируется членами кластеров IV и XIVa Clostridium (типа Firmicutes). В способе, называемом перекрестная подача, эти микроорганизмы превращают ацетат и/или лактат (вместе с другими субстратами) в связанный со здоровьем бутират. Уровни бутирата постепенно повышались после добавления геллановой камеди в проксимальном отделе и в меньшей степени в дистальном отделе ободочной кишки для всех исследуемых доноров (фиг. 4a-4b, таблица 10). Эффект был наиболее выраженным в проксимальном отделе со значительными повышениями, составляющими 2,3 мМ (+24%), 1,9 мМ (+21%) и 1,4 мМ (+15%) для донора A, донора B и донора C, соответственно. В дистальном отделе ободочной кишки уровни бутирата только у донора значительно повышались после добавления геллановой камеди (т.е. повышались на 1,4 мМ (+13%)).
Таблица 10. Эффект обработки геллановой камедью на продукцию бутирата (в мМ) в ферментерах в качестве проксимального (PC) и дистального (DC) отделов ободочной кишки для трех разных доноров (A, B и C) и на среднюю еженедельную продукцию бутирата во время контрольных периодов (C1 и C2) и недель обработки (TR1-TR3) (смотрите также фиг. 4a-4b).
5. Продукция лактата
[0212] В кишечнике человека обитают как продуцирующие лактат, так и утилизирующие лактат бактерии. Лактат продуцируется молочнокислыми бактериями и снижает pH окружающей среды. Лактат может оказывать сильное противомикробное действие против патогенов, особенно при низких значениях pH. Другой полезный эффект лактата связан с его превращением в бутират и/или пропионат. Таким образом, поскольку различные виды микроорганизмов продуцируют и превращают лактат, увеличение концентрации лактата может быть результатом как увеличения продукции, так и уменьшение превращения. Следовательно, нужно быть осторожным с интерпретацией данных, касающихся результатов по лактату.
[0213] В проксимальном отделе ободочной кишки концентрация лактата увеличивалась на последней неделе обработки для всех исследуемых доноров, достигая значимости только для донора A (таблица 11). Однако для других доноров высокие среднеквадратические отклонения можно было наблюдать на последней неделе обработки, поскольку концентрации лактата постепенно увеличивались на этой неделе, т.е. с 0,19 мМ в начале последней недели обработки до 0,73 мМ в конце неделя для донора B и от 0,13 мМ до 0,46 мМ для донора C. В дистальном отделе ободочной кишки значительно увеличенные концентрации лактата наблюдались на последней неделе обработки для донора C. Для донора A отмечалась тенденция к большим концентрациям лактата после добавления геллановой камеди, тогда как указанная обработка не оказывала влияния на концентрацию лактата в случае донора B.
Таблица 11. Эффект обработки геллановой камедью на продукцию лактата (в мМ) в ферментерах в качестве проксимального (PC) и дистального (DC) отделов ободочной кишки для трех разных доноров (A, B и C) и на среднюю еженедельную продукцию лактата во время контрольных периодов (C1 и C2) и недель обработки (TR1-TR3) (смотрите также фиг. 5a-5b).
6. Продукция аммиака и SCFA с разветвленной цепью
[0214] Продукция как аммиака (NH4+), так и SCFA с разветвленной цепью (b-SCFA=сумма изобутирата, изовалерата и изокапроата) является результатом деградации белка и отражает протеолитическую активность кишечной микробиоты. Поскольку последняя была связана с прямыми и непрямыми пагубными последствиями для здоровья (например, канцерогенезом ободочной кишки), снижение продукции аммиака/b-SCFA считается полезным. На фиг. 6a-6b (таблица 12) представлена средняя продукция аммиака (в мг/мл), связанная с различными обработками в двух частях ободочной кишки, тогда как на фиг. 7a-7b (таблица 13) представлена средняя продукция SCFA с разветвленной цепью (в мМ), связанная с различными обработками в двух частях ободочной кишки.
[0215] На уровни аммиака не повлияла обработка геллановой камедью как в проксимальном, так и в дистальном отделе ободочной кишки для всех исследуемых доноров, за исключением небольшого повышения в проксимальном отделе ободочной кишки на последней неделе обработки для донора С. Эти результаты подтверждались уровнями SCFA с разветвленной цепью, когда к концу обработки наблюдалось лишь небольшое повышение как в проксимальном, так и в дистальном отделах ободочной кишки для всех исследуемых доноров.
Таблица 12. Эффект обработки геллановой камедью на продукцию аммиака (мг/л) в ферментерах в качестве проксимального (PC) и дистального (DC) отделов ободочной кишки для трех разных доноров (A, B и C), а также на среднюю еженедельную продукцию аммиака (мг/л) во время контрольных периодов (C1 и C2) и недель обработки (TR1-TR3) (смотрите также фиг. 6a-6b).
Таблица 13. Эффект обработки геллановой камедью на продукцию SCFA с разветвленной цепью (мМ) в ферментерах в качестве проксимального (PC) и дистального (DC) отделов ободочной кишки для трех разных доноров (A, B и C) и на среднюю еженедельную продукцию SCFA с разветвленной цепью (мМ) во время контрольных периодов (C1 и C2) и недель обработки (TR1-TR3) (смотрите также фиг. 7a-7b)).
G. Анализ состава микробного сообщества
[0216] Нацеленное на 16S рРНК секвенирование Illumina представляет собой молекулярный метод, основанный на амплификации гена 16S рРНК. Поскольку метод секвенирования Illumina основан на ПЦР, последовательности микроорганизмов амплифицируются до достижения уровня насыщения. Поэтому при получении информации о широком спектре (не заданных заранее) OTU (> 100 различных из наиболее доминирующих OTU) результаты представляют в виде пропорциональных значений по отношению к общему числу последовательностей в каждой образце, что позволяет получить полуколичественные результаты. Применяемая здесь методология включает праймеры, которые охватывают две гипервариабельные области (V3-V4) 16S рДНК. Используя подход секвенирования спаренных концов, секвенирование 2×250 п.н. дает ампликоны размером 424 п.н. Такие фрагменты таксономически более пригодны, чем меньшие фрагменты, которые таксономически менее информативны. Помимо обработки данных на уровне типа и семейства, можно идентифицировать конкретные измененные OTU, а также можно рассчитать индекс разнообразия Симпсона в качестве показателя как разнообразия, так и равномерности распределения. Наименьшее возможное значение индекса равно 1, что соответствует сообществу, состоящему только из одной OTU. Максимально возможное значение равно общему числу OTU. Индекс будет больше приближаться к максимальному значению, когда распределение OTU будет более равномерным, тогда как сообщество, в котором доминирует небольшое число OTU, даст значения, близкие к 1. Чем выше индекс, тем больше разнообразие и больше равномерность распределения.
1. Индекс разнообразия
[0217] Обратную величину индекса разнообразия Симпсона рассчитывали в качестве показателя разнообразия, с точки зрения как изобилия, так и равномерности распределения видов. На основе индексов разнообразия следовало, что во время контрольного периода каждый из трех блоков SHIME был колонизирован репродуцируемыми микробными сообществами просвета и слизистой оболочки как в PC, так и в DC. Разнообразие было выше в DC, хотя оно также было значительно выше для люминальной микробиоты по сравнению с микробиотой слизистой оболочки как в PC, так и в DC (таблица 14).
Таблица 14. Средняя обратная величина индекса разнообразия Симпсона в просвете (L) и слизи (M) проксимального (PC) и дистального (DC) отделов ободочной кишки в виде трех блоков SHIME во время контрольного периода (n=6). Кроме того, также отмечались значимые различия (p<0,05) для обратной величины индекса разнообразия Симпсона между L и M или между PC и DC, о чем свидетельствует их р-значения, рассчитанные с использованием критерия Стьюдента.
[0218] Кроме того, что касается эффектов обработки, геллановая камедь увеличивала разнообразие кишечной микробиоты по сравнению с контролем для всех трех исследуемых доноров (фиг. 8). Только разнообразие люминальной микробиоты в PC несколько уменьшилось после обработки геллановой камедью.
2. Уровень типа
[0219] К тому же, состав микробиоты на уровне типа указал на то, что три разных блока SHIME были колонизированы репродуцируемыми микробными сообществами просвета и слизистой оболочки как в проксимальном, так и в дистальном отделах ободочной кишки. В результате были рассчитаны средние значения для каждого из четырех условии окружающей среды, в то время как статистические испытания выполнялись, чтобы понять предпочтение конкретного типа для любого из четырех условий окружающей среды (таблица 15).
Таблица 15. Среднее относительное количество (%) на уровне типа микроорганизмов в просвете (L) и слизи (M) проксимального (PC) и дистального отделов ободочной кишки (DC) в виде трех блоков SHIME во время контрольного периода (n=6). Кроме того, также значимые различия (p<0,05) для определенного типа между L и M или между PC и DC выделены жирным шрифтом и подчеркнуты с использованием их p-значений, рассчитанных, используя критерий Стьюдента.
[0220] Это выявило специфическую для типа колонизацию просвета по сравнению со слоем слизи с: (i) более высокими уровнями Bacteroidetes в просвете (значимыми только в DC); (ii) более высокими уровни Proteobacteria в просвете (значимыми только в PC); и (iii) более высокими уровнями Synergistetes в слизи (присутствующими только в DC). Кроме того, вдоль ободочной кишки наблюдались следующие продольные различия: (i) повышенные уровни Actinobacteria в PC; (ii) повышенные уровни Bacteroidetes в DC (значимые только в просвете); (iii) присутствие Synergistetes в DC; и (iv) более низкие уровни Proteobacteria в PC в слое слизи, тогда как в просвете отмечалась противоположная тенденция.
[0221] Что касается обработки, отсюда следовало, что в основном месте ферментации, т.е. в просвете проксимального отдела ободочной кишки (фиг. 9), геллановая камедь сильно увеличивала уровни Actinobacteria за счет Bacteroidetes и Firmicutes для всех трех исследуемых доноров. Схожие наблюдения были сделаны для люминальных образцов из дистального отдела ободочной кишки (фиг. 9). Кроме того, в дистальном отделе ободочной кишки уровни Synergistetes и Lentisphaerae в просвете повышались после обработки геллановой камедью. В компартменте в виде слизистой оболочки (фиг. 9) вариабельность образцов с течением времени, как правило, была выше. Это могло объясняться более гетерогенным составом биопленки, которая образуется поверх слоя слизи, по сравнению с гомогенной люминальной суспензией. Actinobacteria в слизистой оболочке, как и в просвете, были обогащены как в проксимальном, так и в дистальном отделах ободочной кишки после обработки геллановой камедью (за исключением донора C в PC, который продемонстрировал очень сильную стимуляцию Synergistetes), однако это не сопровождалось уменьшением количества Bacteroidetes и Firmicutes, как в просвете. Фактически, в компартменте в виде слизистой оболочки между индивидуумами наблюдались различия в уровнях Firmicutes после обработки геллановой камедью, т.е. у донора A наблюдалось снижение уровней Firmicutes, тогда как у доноров B и C наблюдалось повышение. Наконец, обработка геллановой камедью, как правило, увеличивала относительное содержание Proteobacteria в образцах слизистой оболочки проксимального отдела ободочной кишки.
3. Уровень семейства и OTU
[0222] На уровне семейства эффекты обработки геллановой камеди будут в основном обсуждаться для основного места ферментации, т.е. просвета проксимального отдела ободочной кишки (фиг. 10). Для других условий в ободочной кишке (просвета дистального отдела ободочной кишки (фиг. 11), слизистой проксимального отдела ободочной кишки (фиг. 12) и слизистой дистального отдела ободочной кишки (фиг. 13)) было сделано много схожих наблюдений, и поэтому будут обсуждаться только специфические и отличные изменения от основного места ферментации.
[0223] Геллановая камедь сильно повышала уровни Bifidobacteriaceae для всех трех исследуемых доноров. Представленная на фиг. 10-11 информация показывает, что уровни Bifidobacteriaceae в просвете проксимального отдела ободочной кишки в качестве ферментеров для двух контрольных периодов в среднем составляли 24,7±5,5%, в то время как уровни Bifidobacteriaceae в просвете проксимального отдела ободочной кишки в качестве ферментеров для трех периодов обработки в среднем составляли 39,0±8,8%. Кроме того, представленная на фиг. 10-11 информация показывает, что уровни Bifidobacteriaceae в просвете дистального отдела ободочной кишки в качестве ферментеров для двух контрольных периодов в среднем составляли 1,85±1,0%, в то время как уровни Bifidobacteriaceae в просвете дистального отдела ободочной кишки в качестве ферментеров для трех периодов обработки в среднем составляли 8,3±2,3%. На уровне OTU было установлено, что основные изменения относятся к повышению уровней Bifidobacteriaceae OTU 2 (имеющей отношение к Bifidobacterium adolescentis). Этот сильный бифидогенный эффект хорошо согласуется со значительным повышением уровней ацетата, наблюдаемым для всех трех доноров после обработки геллановой камедью.
[0224] Обработка геллановой камедью сильно снижала уровни Bacteroidaceae для всех трех исследуемых доноров. Семейство Bacteroidaceae содержит много известных продуцентов пропионата, что объясняет сильное снижение уровней пропионата, которое наблюдалось после добавления геллановой камеди. Кроме того, после обработки геллановой камедью наблюдалось снижение относительного количества Veillonellaceae, что в основном связано со снижением количества Veillonellaceae OTU 1 (имеющей отношение к Megamonas sp.). Поскольку эта OTU является мощным продуцентом пропионата (при потреблении лактата), ее уменьшение, вероятно, способствовало снижению концентрации пропионата, наблюдаемому во время периода обработки.
[0225] Геллановая камедь также немного повышала уровни Lachnospiraceae на протяжении трехнедельного периода обработки для трех исследуемых доноров, что может быть связано с увеличенными концентрациями бутирата, наблюдаемыми в этот же период. Напротив, в просвете дистального отдела оболочной кишки уровни Lachnospiraceae снижались, хотя после обработки геллановой камедь повышались уровни других семейств, продуцирующих бутират, т.е. Acidaminococcaceae, Eubacteriaceae и Ruminococcaceae. Однако на последней неделе обработки уровни Ruminococcaceae снова снижались в дистальном отделе ободочной кишки, в то время как на этой же неделе наблюдалась стимуляция Veillonellaceae. Последнее объясняет увеличенную продукцию пропионата, наблюдаемую в дистальном отделе ободочной кишки на последней неделе обработки, и в основном относится к стимуляции Veillonellaceae OTU 1 (имеющей отношение к Megamonas sp.).
[0226] Другим продуцирующим бутират семейством, которое было исключительно обогащено в условиях слизистой оболочки после обработки геллановой камедью, было семейство Clostridiaceae, с явным увеличением Clostridiaceae OTU 23 (имеющей отношение к Clostridium butyricum) в проксимальном отделе ободочной кишки по сравнению с Clostridiaceae OTU 17 (имеющей отношение к Clostridium tertium) в дистальном отделе ободочной кишки.
[0227] Другим открытием, согласующимся с ранее сделанными после обработки геллановой камедью, было увеличение нескольких семейств внутри типа Proteobacteria, такое как увеличение Enterobacteriaceae и Xanthomonadaceae. Эти семейства в основном известны, так как они содержат несколько условно-патогенных видов, однако в этих семействах также присутствует много комменсалов, которые, как известно, ферментируют белки в различных частях ободочной кишки, но в основном в дистальном отделе ободочной кишки. Действительно, схожие наблюдения были сделаны для дистального отдела ободочной кишки, где несколько семейств типа Proteobacteria слегка увеличились после обработки геллановой камедью. Эти данные могут коррелировать с небольшим повышением уровней SCFA с разветвленной цепью, которое наблюдалось к концу периода обработки.
[0228] Наконец, некоторые специфические для доноров изменения отмечались после обработки геллановой камедью в просвете проксимального отдела ободочной кишки: (i) повышенные уровни Microbacteriaceae для доноров B и C; (ii) повышенные уровни Micrococcaceae для доноров A и C; (iii) повышенные уровни Enterococcaceae, особенно наблюдаемые для донора C (схожие наблюдения были сделаны в дистальном отделе ободочной кишки, что могло объяснять увеличенную концентрацию лактата на последней неделе обработки, которая отмечалась для этого донора); и (iv) повышенные уровни Synergistaceae для донора C. Поскольку Synergistaceae в основном колонизируют дистальные части ободочной кишки, более сильные эффекты наблюдались в образцах из просвета дистального отдела ободочной кишки, где для всех трех исследуемых доноров наблюдалось сильное обогащение Synergistaceae.
H. Краткое изложение результатов примера III
[0229] Расход кислоты/основания, газообразование, продукция SCFA, лактата и аммиака были очень стабильными в трех разных блоках SHIME во время контрольного периода. Это указывало на то, что модель SHIME работала в наиболее оптимальных условиях, приводящих к стабильной микробиоте в ободочной кишке. Эта стабильность является предварительным условием того, что любой эффект, наблюдаемый во время обработки, действительно является результатом введенного исследуемого продукта в концентрации, соответствующей дозе=2 г/день in vivo.
[0230] После начала обработки геллановой камедью расход основания увеличивался в проксимальном отделе ободочной кишки (что указывает на ферментацию микроорганизмами через продукцию SCFA/лактата) на последней неделе обработки для всех исследуемых доноров. К тому же, в дистальном отделе ободочной кишки наблюдалось умеренное немедленное увеличение расхода оснований. Что касается газообразования, наблюдались зависимые от доноров эффекты, с небольшим увеличением газообразования для донора B, тогда как газообразование для других доноров уменьшалось после добавления продукта.
[0231] В то время как расход основания и газообразование являются лишь грубым показателем ферментации микроорганизмами, измерения SCFA дают более подробное представление о процессах сахаролитической ферментации. Они показали, что геллановая камедь в основном ферментировалась в проксимальном отделе ободочной кишки, где сразу же снижала уровни пропионата при постепенном повышении уровней ацетата и бутирата. Микробиота донора А давала наиболее выраженные повышения уровней как ацетата, так и бутирата после обработки геллановой камедью. Также в дистальном отделе ободочной кишки уровни ацетата и бутирата постепенно повышались в ходе обработки, тогда как уровни пропионата постепенно снижались, с последующим повышением на последней неделе обработки. Наибольшее увеличение продукции ацетата наблюдалось для донора B, тогда как донор A давал наибольшее повышение уровней бутирата. Кроме того, концентрации лактата в целом оставались очень стабильными. В проксимальном отделе ободочной кишки уровень лактата значительно повышался только на последней неделе обработки для донора А. В дистальном отделе ободочной кишки отмечали значительно увеличенные концентрации лактата на последней неделе обработки для донора С.
[0232] Что касается маркеров протеолитической ферментации, отсюда следовало, что на уровни аммиака не оказывалось влияние для всех исследуемых доноров, как в проксимальном, так и в дистальном отделах ободочной кишки, за исключением небольшого повышения в проксимальном отделе ободочной кишки на последней неделе обработки для донора C. Эти результаты подтверждались уровнями SCFA с разветвленной цепью, когда только небольшие повышения наблюдались к концу обработки как в проксимальном, так и в дистальном отделах ободочной кишки для всех исследуемых доноров.
[0233] Нацеленный на 16S рРНК анализ секвенирования показал, что модель SHIME поддерживает разнообразную микробиоту просвета и слизистых оболочек как в проксимальном, так и в дистальном отделе ободочной кишки для трех исследуемых доноров. Интересно, что микробиота слизистых оболочек была, в соответствии с данными о взрослых людях, сильно обогащена семействами, содержащими хорошо известные виды, продуцирующие бутират. Помимо этой видоспецифической колонизации слизистого слоя, также были установлены продольные различия в микробной колонизации (проксимального отдела по сравнению с дистальным отделом ободочной кишки).
[0234] Что касается эффектов обработки на состав микробного сообщества, было обнаружено, что геллановая камедь увеличивала разнообразие кишечной микробиоты трех исследуемых доноров по сравнению с контрольным периодом. Кроме того, следовало, что в основном месте ферментации (просвете проксимального отдела ободочной кишки) геллановая камедь сильно повышала уровни Actinobacteria за счет Bacteroidetes и Firmicutes. Повышение уровней Actinobacteria было в основном связано с цветением, вызванным Bifidobacteriaceae, что точно соответствовало повышению уровней ацетата для всех трех исследуемых доноров. Интересно, что бифидогенный эффект после добавления геллановой камеди был связан только с повышениями уровней OTU, имеющей отношение к Bifidobacterium adolescentis. Снижения уровней Bacteroidetes и Firmicutes в основном относились к снижениям уровней Bacteroidaceae и Veillonellaceae для всех трех исследованных доноров. Оба семейства содержат несколько сильных продуцентов пропионата, что коррелирует со снижениями концентраций пропионата, наблюдаемыми в период обработки. Наконец, увеличение продукции бутирата на протяжении 3-недельного периода обработки геллановой камедью было потенциально связано с увеличением количества продуцирующих бутират видов, относящихся к нескольким семействам Firmicutes, таким как Lachnospiraceae в просвете проксимального отдела ободочной кишки, Acidaminococcaceae, Eubacteriaceae и Ruminococcaceae в просвете дистального отдела ободочной кишки, и Clostridiaceae в условиях слизистой оболочки.
IV. Пример IV. Влияние геллановой камеди на функции стенки кишечника
А. Введение
[0235] Микроорганизмы в кишечнике представляют собой биологически активное сообщество, которое находится на границе раздела хозяина с его питательной средой. Как следствие, они сильно влияют на некоторые аспекты физиологии и метаболизма хозяина. Широкий ряд структурных компонентов и метаболитов микроорганизмов напрямую взаимодействует с клетками кишечника хозяина, влияя на усвоение питательных веществ и здоровое состояние эпителия. Как молекулярные паттерны, связанные с микроорганизмами (MAMP), так и метаболиты бактериального происхождения (например, короткоцепочечные жирные кислоты (SCFA)) активируют различные пути передачи сигналов, такие как созревание лимфоцитов, здоровое состояние эпителия, передача нейроэндокринных сигналов, опосредованная рецепторами распознавания паттернов (PRR) и опосредованная рецепторами, связанными с G-белком (GPR), передача сигналов. В свою очередь, эти пути передачи сигналов будут определять тонус воспаления, энергетический баланс, сократительную способность кишечника и регуляцию аппетита (обзор в Ha (2014)). В настоящее время считается, что нарушение регуляции взаимодействий микробиома с хозяином способствует возникновению множества заболеваний (Groschwitz (2009)), включая метаболический синдром и ожирение, воспалительные заболевания кишечника (IBD), такие как болезнь Крона (CD) и неспецифический язвенный колит (UC), синдром раздраженного кишечника (IBS), глютеновую болезнь, диабет, аллергию, астму и аутоиммунные заболевания. Общим для этих нарушений является нарушение регуляции кишечного эпителиального барьера (более проницаемого), инициирующее патологию (Fasano (2011)). Когда функция кишечного барьера нарушена, перемещение молекул больше не находится под контролем, поэтому содержимое просвета может проникать в собственную пластинку и активировать иммунную систему, тем самым приводя к неконтролируемым иммунным ответам (процесс, известный как «дырявый кишечник»). Кишечный эпителиальный барьер образован межклеточными плотными соединениями, сложной белок-белковой сетью, которая механически связывает соседние клетки и закрывает межклеточное пространство. Следовательно, кишечный эпителиальный барьер контролирует равновесие между иммунологической толерантностью и активацией иммунной системы, и поэтому он играет важную роль в патогенезе «дырявого кишечника». Неправильное функционирование или регуляция этих плотных соединений, по-видимому, отвечает за большие межклеточные пространства, позволяющие люминальным элементам проходить через барьер, с последующим локальным и системным воспалением.
В. Модель сокультивирования Caco-2/THP1 in vitro
[0236] Для имитации границы раздела между микробиомом кишечника и хозяином, в последние годы было разработано несколько моделей in vitro, которые включают использование кишечных клеток, подобных эпителиальным клеткам, и иммунных клеток человеческого происхождения. Используемая здесь модель представляет собой модель сокультивирования кишечных клеток, подобных эпителиальным клеткам, (клеток Сасо-2) и моноцитов/макрофагов человека (клеток THP1). (смотрите фиг.14; смотрите также Possemiers (2013) Satsu (2006).) При посеве на подходящие подложки Caco-2 спонтанно дифференцируется в зрелые энтероцитоподобные клетки, характеризующиеся поляризацией, наличием ворсинок, образованием конусов, наличием плотных соединений и переносом инфекции, и экспрессией ферментов щеточной каймы на апикальной поверхности (обзор Sambuy (2005)). Моноциты THP1, выделенные от являющегося человеком пациента с острым лейкозом, дифференцируются в макрофагоподобные клетки после обработки форбол-12-миристат-13-ацетатом (PMA). Активированные PMA клетки THP1 приобретают морфологические особенности, характерные для макрофагов, способны прикрепляться к подложке, развивать ламеллиподии, необходимые для миграции и фагоцитоза, и становятся примированными для ответов toll-подобные рецепторов (TLR). (Dumrese (2009).) Белки плотных соединений удерживают смежные эпителиальные клетки вместе, тем самым образуя практически непроницаемый барьер для макромолекул. «Герметичность» этих соединений можно измерить как трансэпителиальное электрическое сопротивление (TEER), при этом высокое TEER соответствует более плотному барьеру. При утрате барьерной функции трансклеточный перенос жидкости (между клетками) увеличивается, что можно определить как снижение TEER. Когда клетки Caco-2 помещают поверх PMA-активированных клеток THP1, которые секретируют цитокины в супернатант, их монослой разрушается. Возможно, это обусловлено опосредованным цитокинами нарушением плотных соединений и может быть определено как снижение TEER.
[0237] Внутри кишечника химическое, механическое или запускаемое патогенами нарушение барьера может привести к притоку бактерий из просвета в собственную пластинку (фиг. 15). Это будет активировать иммунную систему, которая переключится с физиологического «толерогенного» воспаления на вредное патологическое воспаление. Каскад передачи сигналов воспаления будет инициироваться продукцией сигнальных молекул, таких как провоспалительные цитокины (например, фактор некроза опухолей (TNF)-α и интерлейкин (IL)-1β). TNF-α, вместе с интерфероном (IFN)-γ, продуцируется лейкоцитами и CD4+ TH клетками(-хелперами) 1 типа, важными клеточными защитниками от вторжения микроорганизмов. Эти провоспалительные цитокины будут индуцировать продукцию хемокинов (например, IL-8 и лиганда хемокина (мотива C-X-C) (CXCL)-10) и молекул адгезии), необходимых для рекрутинга нейтрофилов и образования активных форм кислорода (ROS). Образование ROS необходимо для уничтожения вторгающихся бактерий и закрытия брешей в эпителиальной стенке. Однако они также могут вызывать разрушение тканей и воспаление, что приводит к необходимости устранения воспаления путем продукции противовоспалительных цитокинов, таких как IL-6 и IL-10.
[0238] IL-6 обладает как провоспалительными, так и противовоспалительными свойствами. Scheller (2011). IL-6 приводит к рекрутингу моноцитов/макрофагов через активацию белка-хемоаттрактанта моноцитов (MCP)-1, который способствует удалению нейтрофилов. IL-6 также способен ингибировать продукцию провоспалительных цитокинов, таких как IL-1. Кроме того, IL-6 оказывает положительный эффект на регенерацию кишечного эпителия и заживление ран. Dann (2008). С другой стороны, IL-6 вместе с трансформирующим фактором роста (TGF)-β) индуцирует дифференцировку важной субпопуляции CD4+ T-клеток - клеток TH17, которые играют ключевую роль в защите хозяина от экстраклеточных микроорганизмов в слизистой ткани.
[0239] IL-10 представляет собой противовоспалительный цитокин, способный подавлять некоторые типы клеток врожденного и адаптивного иммунного ответа. К тому же, IL-10 индуцирует активацию провоспалительных молекул и усиливает функцию регуляторных Т-клеток (Treg), что будет восстанавливать иммунный гомеостаз. Lyer (2012). Когда эти механизмы выключения нарушены, и иммунный гомеостаз не может быть восстановлен, может возникнуть патология кишечника, которая может привести к хроническому воспалению (как видно, например, при IBD, которое характеризуется чрезмерной активацией TH1-опосредованных ответов, а именно сверхпродукцией TNF-α).
[0240] Что касается воспаления, TNF-α является одним из самых мощных и опасных цитокинов, продуцируемых иммунной системой, так как он оказывает плейотропные эффекты и способен усилить передачу сигналов воспаления (фиг. 16). Без противодействия, TNF-α может привести к хроническому воспалению и даже смерти в случае острого воспаления. По этой причине терапия против TNF-α широко используется при хронических воспалительных состояниях, включая IBD и ревматоидный артрит.
[0241] Модель сокультивирования Caco-2/THP-1 демонстрирует некоторые особенности, также наблюдаемые у пациентов с IBD, и поэтому предлагается в качестве модели, подобной IBD, которую можно использовать для исследования эффекта веществ, которые могут как защитить целостность кишечного эпителиального барьера, так и уменьшить воспаление. Satsu (2006). Как говорилось, в этой модели защита барьерной функции кишечника определяется как увеличение TEER, в то время как противовоспалительный потенциал определяется посредством анализа профиля цитокинов (увеличения противовоспалительных цитокинов и уменьшения провоспалительных цитокинов).
[0242] Относящиеся к ободочной кишке суспензии, собранные из SHIME, приводят в контакт с апикальной стороной сокультур (клеток Caco-2). Эффекты, наблюдаемые в базолатеральной камере (где находятся клетки THP1), затем опосредуются опосредованно сигналами, создаваемыми клетками Caco-2, и/или переносом микро- и макромолекул. Уникальный аспект этого подхода заключается в том, что он позволяет оценить эффект, индуцированный продуктом и полученными в результате ферментации метаболитами, продуцируемыми микробиотой кишечника на стадиях пищеварения (т.е. не только чистым продуктом). Daguet (2016).
C. Цель исследования
[0243] Целью этой части исследования было изучение потенциальных положительных эффектов продукта геллановой камеди и ее метаболитов на функции стенок кишечника у трех разных доноров. Бактерии тесно взаимодействуют со стенкой кишечника, поэтому модуляция активности микроорганизмов, вероятно, повлияет на функции стенки кишечника. Это будет оцениваться путем оценки проницаемости кишечного эпителия и специфических иммунных маркеров in vitro.
D. Материалы и методы
[0244] Образцы, собранные в эксперименте SHIME, описанном выше, использовали для оценки in vitro эффекта ферментированных продуктов на функцию кишечного эпителиального барьера и иммунные маркеры. Они включают образцы из ферментеров в качестве проксимального и дистального отделов ободочной кишки трех разных доноров, отобранные в конце контрольного периода и периода обработки.
E. Клетки Caco-2
[0245] Эксперимент по сокультивированию проводили, как описано ранее. Daguet (2016). Вкратце, клетки Caco-2 (HTB-37; Американская коллекция типовых культур) высевали в 24-луночные полупроницаемые вкладыши. Монослои Caco-2 культивировали в течение 14-21 дней с тремя сменами среды в неделю до получения функционального клеточного монослоя с трансэпителиальным электрическим сопротивлением (TEER). Клетки поддерживали в среде Игла, модифицированной Дульбекко (DMEM), содержащей глюкозу и глютамин и дополненной HEPES и 20% (в объемном отношении) инактивированной (HI) фетальной бычьей сывороткой (FBS).
F. Клетки THP1-Blue™
[0246] Клетки THP1-Blue™ (InvivoGen) поддерживали в среде 1640 Roswell Park Memorial Institute (RPMI), содержащей глюкозу и глютамин, с добавлением HEPES, пирувата натрия и 10% (в объемном отношении) HI-FBS. THP1-Blue™ представляют собой моноциты человека THP1, стабильно трансфицированные репортерной конструкцией, экспрессирующей ген секретируемой щелочной фосфатазы (SEAP), под контролем промотора, индуцируемого фактором транскрипции ядерным фактором каппа B (NF-κB). После активации TLR (например, липополисахаридом (LPS); выделенным из грамотрицательных бактерий) NF-κB активируется и индуцирует экспрессию и секрецию SEAP. Затем активность SEAP можно измерить в супернатантах, используя реагент QUANTI-Blue (InvivoGen). Клетки THP1-Blue™ высевали в 24-луночные планшеты и обрабатывали PMA, который индуцирует дифференцировку клеток в макрофагоподобные клетки, которые способны прикрепляться и примируются для передачи сигналов от TLR.
G. Сокультивирование Caco-2/THPl-blue™
[0247] Перед установкой сокультивирования измеряли TEER монослоев Caco-2 (момент времени=0 час). TEER пустого вкладыша вычитали из всех показаний для учета остаточного электрического сопротивления вкладыша. Затем вкладыши, содержащие Caco-2, помещали поверх PMA-дифференцированных клеток THP1-Blue™ для дальнейших экспериментов, как описано ранее. Possemiers (2013) и Lyer (2012).
[0248] Вкратце, апикальный компартмент (содержащий клетки Caco-2) заполняли стерилизованными фильтрованием (0,22 мкм) относящимися к ободочной кишке суспензиями из SHIME или различными концентрациями живых бактерий. Клетки также обрабатывали апикально бутиратом натрия (NaB) (Sigma-Aldrich) в качестве положительного контроля. Базолатеральный компартмент (содержащий клетки THP1-Blue™) заполняли средой полного состава для Caco-2. Клетки также подвергали воздействию среды полного состава для Caco-2 в обеих камерах в качестве контроля. Клетки обрабатывали в течение 24 часов, после чего измеряли TEER (момент времени=24 часа). После вычитания TEER пустого вкладыша все значения в 24 часа нормализовали к его собственному значению в 0 часов (для учета различий в начальном TEER разных вкладышей) и представляли как процент от исходного значения. Затем базолатеральный супернатант удаляли, и клетки стимулировали базолатерально средой полного состава для Caco-2, содержащей сверхчистый LPS (Escherichia coli K12, InvivoGen). Клетки также стимулировали базолатерально LPS в комбинации с гидрокортизоном (HC) (Sigma-Aldrich) и средой без LPS (LPS-) в качестве контролей. После стимуляции LPS базолатеральные супернатанты собирали для измерения цитокинов (IL-Iβ, IL-6, IL-8, IL-10, TNF-α, CXCL10 и MCP-1 человека с помощью мультиплекса Luminex® (Affymetrix-eBioscience)) и для измерения активности NF-κB, в соответствии с инструкциями производителей. Клетки инкубировали при 37οC во влажной атмосфере воздух/СО2 (95:5, в объемном отношении).
H. Статистические данные
[0249] Экспериментальные контроли представляли сначала на отдельных графиках; они относятся к клеткам, обработанным средой полного состава в качестве контроля (CM или LPS-), клеткам, обработанным липополисахаридом (LPS+) и бутиратом натрия (NaB), и гидрокортизоном (HC) в качестве контролей. Что касается TEER, сравнивали условия CM и NaB, и статистическую значимость рассчитывали, используя непарный двусторонний критерий Стьюдента. Для иммунных маркеров (цитокинов/хемокинов и активности NF-κB) все условия (LPS-, LPS+HC и LPS+NaB) сравнивали с LPS+. Статистическую значимость рассчитывали, используя однофакторный дисперсионный анализ с использованием критерия множественных сравнений Даннета с LPS+. (*), (**), (***) и (****) представляют p <0,05, p<0,01, p<0,001 и p<0,0001, соответственно.
[0250] Результаты, касающиеся образцов SHIME, представлены отдельно. Контрольные образцы (C) и образцы после обработки (T), представленные для обоих ферментеров в качестве ободочной кишки (проксимального (PC) и дистального (DC) отделов ободочной кишки), отбирали в виде трех биологических повторов в эксперименте SHIME. Показаны результаты для трех разных доноров отдельно, а также среднее значение для трех доноров. Для оценки различий в TEER, активации NF-κB и продукции цитокинов между каждым подвергнутым обработке образцом и контролем выполняли обычный односторонний дисперсионный анализ (ANOVA) с использованием критерия множественных сравнений Таки (значимость обозначена звездочкой (*)). (*), (**), (***) и (****) представляют p<0,05, p<0,01, p<0,001 и p<0,0001, соответственно. Вся статистика проводилась с использованием программного обеспечения GraphPad Prism™ версии 7.02 для Windows (GraphPad Software, San Diego, CA, США).
I. Результаты для контролей
1. Трансэпителиальное электрическое сопротивление (TEER)
[0251] После сокультивирования в течение 24 часов среда полного состава (CM) в качестве контроля показала снижение TEER почти на 40% из-за повреждения, вызванного PMA-активированными клетками THP1 на клетках Caco-2 (фиг. 17). Как и ожидалось, бутират натрия (NaB; положительный контроль) был способен защищать клетки Caco-2 от этого повреждения и сохранять TEER монослоя. Peng (2007). Обратите внимание, что LPS добавляется только после измерения TEER в момент времени=24 часа. Однако предварительные эксперименты показали, что используемая доза LPS не оказывает значительного влияния на целостность барьера из клеток Caco-2.
2. Иммунные маркеры
[0252] Полученные для различных иммунных маркеров результаты можно видеть на фиг. 18, фиг. 19 и фиг. 20. Как и ожидалось, LPS был способен увеличивать активацию NF-κB (фиг. 18), а также секрецию всех исследуемых цитокинов (IL-6 и IL-10 (фиг. 19) и IL-Iβ, IL-8, CXCL10, TNF-α и MCP-1 (фиг. 20)). Кроме того, гидрокортизон (HC), будучи кортикостероидом, действует как иммунодепрессант широкого действия, подавляя LPS-индуцированные цитокины и хемокины (фиг. 19 и 20) и ингибируя LPS-индуцированную транскрипционную активность NF-κB (фиг. 18). Напротив, бутират натрия (NaB) показал зависимый от маркера эффект. NaB увеличивал транскрипционную активность NF-κB (фиг. 18), эффект, который, возможно, опосредован ослаблением ингибирующих активностей гистондеацетилаз (HDAC) в отношении негистоновых белков, таких как NF-κB. Glozak (2005) и Vinolo (2011). Кроме того, NaB продемонстрировал явную избирательную посттранскрипционную ингибирующую активность в отношении некоторых иммунных медиаторов. Конкретнее, NaB избирательно увеличивал LPS-индуцированную секрецию IL-6 и IL-10 (участвующую в иммунном гомеостазе) (фиг. 19), хотя он избирательно ингибировал LPS-индуцированную продукцию TNF-α (провоспалительных цитокинов) и IL-8, CXCL10 и MCP-1 (хемокинов, участвующих в рекрутинге иммунных клеток) (фиг. 20).
[0253] В заключение, все контроли в этом эксперименте вели себя так, как ожидалось, и результаты, полученные для образцов SHIME, представлены ниже. Обратите внимание, что в этом эксперименте HC и NaB неожиданно не снижали LPS-индуцированную экспрессию IL-1β.
J. Результаты для образцов SHIME
1. Трансэпителиальное электрическое сопротивление (TEER)
[0254] Образцы SHIME, отобранные на последних неделях контроля и обработки из всех ферментеров в качестве ободочной кишки, разводили (1:5, в объемном отношении) в среде полного состава для Caco-2 после фильтрацию (0,22 мкм) и вносили апикально в сокультуры на 24 часа.
[0255] По сравнению со средой полного состава (CM) в качестве контроля, когда TEER снижалось на приблизительно 40% (фиг. 17), все контрольные образцы и подвергнутые обработке образцы, отобранные из SHIME, могли поддерживать TEER почти на начальном уровне (фиг. 21). Хотя и незначительное, небольшое увеличение TEER наблюдалось после обработки геллановой камедью в образцах из дистального отдела ободочной кишки всех для трех доноров по сравнению с контролем. Учитывая тот факт, что это увеличение постоянно наблюдалось для всех трех доноров, можно сделать вывод, что ферментация геллановой камеди имеет потенциал для улучшения функции кишечного эпителиального барьера в используемой модели in vitro.
2. Иммунные маркеры
[0256] После предварительной обработки апикально в течение 24 часов сокультур Caco-2/THP-1-Blue™ образцами SHIME, базолатеральный супернатант удаляли, и клетки стимулировали LPS. После 6-часовой стимуляции базолатеральный супернатант собирали для измерения цитокинов и хемокинов, секретируемых в среду, и для определения активности NF-κB.
[0257] По сравнению с контролем LPS+ (красная пунктирная линия), все образцы SHIME увеличивали LPS-индуцированную транскрипционную активность NF-κB (фиг. 22). Однако не было статистически значимого различия между контрольными образцами и образцами, подвергнутыми обработке. Следовательно, увеличение активности NF-κB скорее отражает эффект на клетки суспензии SHIME, а не исследуемого соединения.
[0258] Подобно результатам, полученным для активности NF-κB, все образцы SHIME повышали уровни LPS-индуцированных IL-6 и IL-10 по сравнению с контролем LPS+ (фиг. 23). Хотя и незначительно, но небольшое повышение уровней IL-6 и IL-10 по сравнению с контролем постоянно наблюдалось для всех доноров в образцах дистального отдела ободочной кишки. Оно было статистически значимо только для уровней IL-6 у донора A. Это повышение уровней IL-6 и IL-10 также наблюдалось при анализе среднего значения для трех доноров. Интересно, что при применении парного критерия Стьюдента ко всем образцам из проксимального и дистального отделов ободочной кишки отмечалось значительное повышение уровней IL-10 (p<0,05) для трех исследуемых доноров.
[0259] Полученные для IL-1β и TNF-α результаты представлены на фиг. 24. Все образцы SHIME явно увеличивали секрецию IL-1β по сравнению с контролем LPS+ (красная пунктирная линия), однако не было обнаружено различий в уровнях IL-1β между контролем и обработкой, за исключением донора B, когда значительное повышение уровней IL-1β наблюдали после обработки для ферментера в качестве проксимального отдела ободочной кишки. При анализе среднего значения для трех доноров не наблюдалось значительных различий в секреции IL-1β между контролем и обработкой.
[0260] По сравнению с их контролями, уровни LPS-индуцированного TNF-α были снижены в образцах из проксимального и дистального отделов ободочной кишки донора A и донора B, но не донора C. При наблюдении за средним значением для трех доноров наблюдалось небольшое снижение секреции TNF-α в образцах из проксимального отдела ободочной кишки по сравнению с контролями, но статистически значимых различий не наблюдалось.
[0261] Как видно на фиг. 25, секреция IL-8, как правило, снижалась после добавления геллановой камеди по сравнению с контролем для образцов из дистального отдела ободочной кишки двух из трех доноров. Однако эта разница не была значимой.
[0262] Уровни LPS-индуцированного CXCL10, как правило, слегка увеличивались в образцах из дистального отдела ободочной кишки всех доноров после обработки геллановой камедью. В случае образцов из проксимального отдела ободочной кишки только один донор продемонстрировал незначительное уменьшение экспрессии CXCL10 после обработки. Уровни MCP-1, как правило, слегка снижались после обработки для проксимального отдела ободочной кишки. Напротив, явное повышение наблюдалось для ферментеров в качестве дистального отдела ободочной кишки для всех доноров. Однако значимость не была достигнута.
[0263] В заключение, хотя геллановая камедь оказывала незначительный эффект на барьерную функцию кишечного эпителия, она, как правило, увеличивала экспрессию противовоспалительных цитокинов IL-6 и IL-10. В некоторых условиях, как правило, снижалась продукция провоспалительных цитокинов и хемокинов. Однако между обработкой и контролем наблюдались лишь некоторые статистически значимые различия.
[0264] Чтобы иметь обзор изменений, вызванных подвергнутыми обработке образцами по сравнению с контролями, среднее значение для подвергнутых обработке образцов SHIME от трех доноров нормализовали для ферментеров в качестве проксимального и дистального отделов ободочной кишки к среднему значению для контрольных образцов SHIME. и представляли в виде таблицы 16.
Таблица 16. Результаты клеточных экспериментов исходя из среднего значения для подвергнутых обработке образцов SHIME от трех доноров, нормализованного к среднему значению для контрольных образцов SHIME.
[0265] В целом, обосновано заявить, что изменения иммунных маркеров в подвергнутых обработке образцах по сравнению с контролями довольно незначительны. Как видно из таблицы 16, геллановая камедь, по-видимому, усиливает секрецию IL-10 и снижает секрецию TNF-α в обоих ферментерах в качестве ободочной кишки. Секреция IL-8 немного снижена, тогда как IL-6 повышена только для образцов из дистального отдела ободочной кишки. Секреция IL-Iβ, по-видимому, увеличена для образцов из проксимального отдела ободочной кишки, но на эти результаты влияет значительное увеличение IL-1β только у одного донора. Секреция MCP-1 уменьшена в результате обработки геллановой камедью для образцов из проксимального отдела ободочной кишки, но увеличена для образцов из дистального отдела ободочной кишки.
К. Краткое изложение результатов примера IV
[0266] Целью этой части исследования было изучение потенциальных положительных эффектов геллановой камеди и ее метаболитов на функции стенок кишечника с точки зрения модуляции «дырявого» состояния кишечника. Это было сделано путем оценки проницаемости кишечного эпителия и специфических иммунных маркеров in vitro.
[0267] После ферментации в ободочной кишке геллановая камедь, как правило, увеличивала целостность кишечного барьера по показателю TEER. Хотя увеличения были незначительными, последовательные увеличения отмечались для всех трех доноров при расположении образцов дистального отдела ободочной кишки в модели in vitro. Кроме того, продукт, как правило, обладал иммуносупрессорными свойствам, что приводило к тенденции снижения уровней нескольких иммунных медиаторов, включающих провоспалительный цитокин TNF-α и белок-хемоаттрактант IL-8, которые, как известно, играют роль в рекрутинге нейтрофилов. MCP-1, который способствует удалению нейтрофилов, как правило, увеличивался в ферментерах в качестве дистального отдела ободочной кишки после обработки геллановой камедью. С другой стороны, IL-10, подлинный противовоспалительный цитокин, как правило, увеличивался, так же как и IL-6, цитокин, участвующий в заживлении ран. Все эти зарегистрированные изменения в основном наблюдались в ферментерах в качестве дистального отдела ободочной кишки, что свидетельствует о более выраженном эффекте продуктов ферментации геллановой камеди на иммунные клетки хозяина в дистальных частях ободочной кишки.
Список противопоставленных материалов в виде непатентных публикаций
[0268] Anderson et al., Food Addit. Contam. (1990) 7(5): 583-590 ("Anderson (1990)”)
[0269] Anderson et al., The dietary effects of gellan gum in humans, Food Addit. Contam. (1988) 5(3): 237-249 ("Anderson (1988)”)
[0270] Bielecka et al., Food Research International (2002) 35: 125-131 ("Bielecka (2002)”)
[0271] Cummings et al., Amer. J. Clin. Nutrit. (1987) 45: 1243-1255 ("Cummings (1987)”)
[0272] Daguet et al., J. Funct. Foods (2016) 20: 369-379 ("Daguet (2016)”)
[0273] Dann et al., J. Immunol. (2008) 180(10): 6816-6826 ("Dann (2008)”)
[0274] Demigne et al., Brit. J. Nutrit. (1995) 74: 209-219 ("Demigne (1995)”)
[0275] Diltz et al., Location of O-acetyl groups in S-657 using the reductive cleavage method, Carbohydr. Res. (2001) 331(3): 265-270 ("Diltz (2001)”)
[0276] Dumrese et al., FEBS Letters (2009) 583: 1637-1643 ("Dumrese (2009)”)
[0277] Edwards et al., Caecal and faecal shortchain fatty acids and stool output in rats fed on diets containing nonstarch polysaccharides, Brit. J. Nutr. (1995) 73: 773-781 ("Edwards (1995)”)
[0278] Esquivel-Elizondo et al., mSystems. (2017) 2(4): e00051-17 ("Esquivel-Elizondo (2017)”)
[0279] Fallourd et al., Ingredient Selection for Stabilisation and Texture Optimisation of Functional Beverages and the Inclusion of Dietary Fibre, Functional and Specialty Beverage Technology (2009) Pt. 1, Sect. 1, 3-38, at 20 ("Fallourd (2009)”)
[0280] Fasano, A., Physiol. Rev. (2011) 91: 151-175 ("Fasano (2011)”)
[0281] FDA Guidance for Industry: Estimating the Maximum Safe Starting Dose in Initial Clinical Trials for Therapeutics in Adult Healthy Volunteers, July 2005 ("FDA Guidance (2005)”)
[0282] Fehlbaum et al., In Vitro Fermentation of Selected Prebiotics and Their Effects on the Composition and Activity of the Adult Gut Microbiota, Int. J. Mol. Sci. (2018) 19: 3097 ("Fehlbaum (2018)”)
[0283] Gibson et al., Nature Revs. Gastro. Hepatol. (2017) 14: 491-502 ("Gibson (2017)")
[0284] Glozak et al., Gene (2005) 363: 15-23 ("Glozak (2005)").
[0285] Groschwitz et al., J. Allergy Clin. Immunol. (2009) 124(1): 3-20 ("Groschwitz (2009)").
[0286] Guimaraes et al., Food Hydrocolloids (2018) 77: 787-795 ("Guimaraes (2018)").
[0287] Ha et al., W.J. Gastroenterol. (2014) 20(44):16498-16517 ("Ha (2014)").
[0288] Hashimoto et al., Arch. Biochem. Biophys. (1998) 354(1): 31-39 ("Hashimoto").
[0289] Jansson, et al., Structural Studies of a Polysaccharide (S-194) Elaborated by Alcaligenes ATCC 31961, Carbohydr. Res. (1986) 156: 157-163 ("Jansson (1986)").
[0290] Karlton-Senaye et al., Agro Food Ind. Hi Tech. (2013) 24(4): 10-14 ("Karlton-Senaye (2013)").
[0291] Kennedy et al. Microbiology (1994) 140: 3007-3013 ("Kennedy (1994)").
[0292] Kuo et al., Identification and Location of L-Glycerate, an Unusual Acyl Substituent in Gellan Gum, Carbohydr. Res. (1986) 156: 173-187 ("Kuo (1986)").
[0293] Li et al, Bioengineered (2019) 10(1): 240-249 ("Li (2019)").
[0294] Lyer et al., Crit. Rev. Immunol. (2012) 32(1): 23-63 ("Lyer (2012)").
[0295] Martínez et al., PLOS One (2010) 5(11): e15-46, ("Martínez (2010)").
[0296] Molly et al., Appl. Microbiol. Biotech. (1993) 39(2): 254-258 ("Molly (1993)").
[0297] Narushima et al., Gut Microbes (2014) 5(3): 333-339 ("Narushima (2014)").
[0298] Noor et al., BMC Gastroenterol. (2010) 10: 134 ("Noor (2010)").
[0299] Patel et al., Adv. Dairy Res. (2013) 1(2): 1-7 ("Patel (2013)").
[0300] Peng et al., Pediatric Res. (2007) 61: 37-41 ("Peng (2007)").
[0301] Possemiers et al., J. Agric. Food Chem. (2013) 61: 9380-9939 ("Possemiers (2013)").
[0302] Possemiers et al., FEMS Microbiol Ecol. (2004) 49(3): 495-507 ("Possemiers (2004)").
[0303] Saavedra et al., Brit. J. Nutrit. (2002) 87: s241-s246 ("Saavedra (2002)").
[0304] Sambuy et al., Cell Biology and Toxicology (2005) 21: 1-26 ("Sambuy (2005)").
[0305] Satsu et al., Exp. Cell Res. (2006) 312: 3909-3939 ("Satsu (2006)").
[0306] Scheller et al., Biochimica et Biophysica Acta (2011) 1813: 878-888 ("Scheller (2011)").
[0307] Stankowski et al., Location of the O-Acetyl Group in Welan by the Reductive-Cleavage Method, Carbohydr. Res. (1992) 224: 337-341 ("Stankowski (1992)").
[0308] Segata et al., Genome Biol. (2012) 13(6): R42, ("Segata (2012)").
[0309] Steer et al., Nutrit. Res. Revs. (2000) 13: 229-254 ("Steer (2000)").
[0310] Sworn G., Gellan Gum, Chapter 9 (pp. 204-227) in Handbook of Hydrocolloids (2nd. Ed.), (2009) Woodhead Publishing Series in Food Science, Technology and Nutrition ("Sworn (2009)").
[0311] Tetsuguchi et al, J. Nutr. Sci. Vitaminol. (1997) 43(5): 515-527 ("Tetsuguchi (1997").
[0312] Tuohy et al., Brit. J. Nutrit., (2001) 86: 341-348 ("Tuohy (2001)").
[0313] Tuohy et al., Microb. Ecol. Health Dis. (2002) 14: 165-173 ("Tuohy (2002)").
[0314] Van de Wiele et al., The Simulator of the Human Intestinal Microbial Ecosystem (SHIME®), Chapt. 27 (pp. 305-318) in The Impact of Food Bioactives on Health (Verhoeckx et al. Eds.) 2013: Springer, New York ("Van de Wiele (2013)").
[0315] Van den Abbeele et al., ISME J. (2013) 6(4):335-340 ("Van den Abbeele (2013)").
[0316] Van den Abbeele et al. Environ. Microbiol. (2011) 13(10): 2667-2680 ("Van den Abbeele (2011)").
[0317] Van den Abbeele et al., Microb Biotechnol. (2012) 5(1):106-115 ("Van den Abbeele (2012)").
[0318] Vinolo et al., Nutrients (2011) 3: 858-876 ("Vinolo (2011)").
[0319] Wong et al., J. Clin. Gastro. (2006) 40: 235-243 ("Wong (2006)").
[0320] Wright et al., Exp. Biol. Med. (1990) 195: 26-29 ("Wright (1990)").
[0321] Zeuner et al. Enzyme Microb. Technol. (Jan. 2016) 82: 42-50 ("Zeuner (2016)").
[0322] Zitomersky et al., PLoS One (2013) 8(6): e63686, ("Zitomersky (2013)").
[0323] Zoetendal et al. App. Environ. Microbiol. (1998) 64: 3854-3859 ("Zoentendal (1998)").
[0324] Альтернативные варианты осуществления, примеры и модификации, которые все еще будут охватываться настоящим изобретением, могут быть придуманы специалистами в данной области техники, в частности, в свете вышеизложенных идей. Кроме того, должно быть понятно, что используемая для описания настоящего изобретения терминология предназначена в соответствии с сущностью слов для описания, а не ограничения.
[0325] Объект изобретения предварительных заявок на патенты США с №№ 62/794452 и 62/869248 таким образом полностью включен посредством ссылки. Кроме того, приведенная здесь справочная информация включена посредством ссылки во всей своей полноте, если это необходимо. В случае разницы в значении между включенными терминами и раскрытыми здесь терминами, значение раскрытых здесь терминов будет иметь преимущественную силу.
[0326] Специалистам в данной области техники также будет понятно, что различные адаптации и модификации предпочтительных и альтернативных вариантов осуществления, описанных выше, могут быть сформированы без отклонения от объема и сущности настоящего изобретения. Поэтому должно быть понятно, что в пределах объема прилагаемой формулы изобретения настоящее изобретение может быть осуществлено на практике иначе, чем здесь конкретно описано.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ ЛЕЧЕНИЯ С ПРИМЕНЕНИЕМ ОЧИЩЕННОГО (ОБЕСЦВЕЧЕННОГО) СУХОГО СОКА ЛИСТА АЛОЭ ВЕРА | 2017 |
|
RU2738776C2 |
| НЕПЕРЕВАРИВАЕМЫЕ ОЛИГОСАХАРИДЫ ДЛЯ СНИЖЕНИЯ ТОЛСТОКИШЕЧНОЙ ФЕРМЕНТАЦИИ БЕЛКА | 2020 |
|
RU2816647C2 |
| ПИЩЕВАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ 2’-ФУКОЗИЛЛАКТОЗУ И ПИЩЕВОЙ БУТИРАТ | 2020 |
|
RU2820217C2 |
| СПОСОБЫ ОБРАБОТКИ СТВОЛОВ СКВАЖИН РЕЦИКЛИРУЕМЫМИ ФЛЮИДАМИ | 2007 |
|
RU2426861C2 |
| ШТАММ BIFIDOBACTERIUM BIFIDUM, ОБЛАДАЮЩИЙ ГАЛАКТОЗИДАЗНОЙ АКТИВНОСТЬЮ, ГАЛАКТООЛИГОСАХАРИДНАЯ КОМПОЗИЦИЯ ДЛЯ СТИМУЛЯЦИИ РОСТА БИФИДОБАКТЕРИЙ, СИНБИОТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ УЛУЧШЕНИЯ СОСТОЯНИЯ КИШЕЧНИКА, ИХ ПРИМЕНЕНИЕ (ВАРИАНТЫ) ДЛЯ ПОЛУЧЕНИЯ ЛЕКАРСТВЕННЫХ ПРЕПАРАТОВ И СПОСОБ ПОЛУЧЕНИЯ СТИМУЛЯТОРА РОСТА БИФИДОБАКТЕРИЙ | 2004 |
|
RU2313572C2 |
| ПИТАТЕЛЬНАЯ КОМПОЗИЦИЯ, ВКЛЮЧАЮЩАЯ 2'ФУКОЗИЛЛАКТОЗУ И 3'ГАЛАКТОЗИЛЛАКТОЗУ | 2020 |
|
RU2818570C2 |
| СМЕСИ ОЛИГОСАХАРИДОВ ГРУДНОГО МОЛОКА (ОГМ) ДЛЯ ПРИМЕНЕНИЯ У МЛАДЕНЦЕВ ИЛИ ДЕТЕЙ МЛАДШЕГО ВОЗРАСТА В МЕДИЦИНСКИХ ЦЕЛЯХ | 2018 |
|
RU2773404C2 |
| СИНЕРГИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ СОХРАНЕНИЯ ЗДОРОВОГО БАЛАНСА МИКРОФЛОРЫ | 2017 |
|
RU2748651C2 |
| СИНЕРГИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ СОХРАНЕНИЯ ЗДОРОВОГО БАЛАНСА МИКРОФЛОРЫ | 2015 |
|
RU2709464C2 |
| ИСПОЛЬЗОВАНИЕ МИКРОБНЫХ СООБЩЕСТВ ДЛЯ ЛЕЧЕНИЯ ЧЕЛОВЕКА И ЖИВОТНЫХ | 2017 |
|
RU2758387C2 |
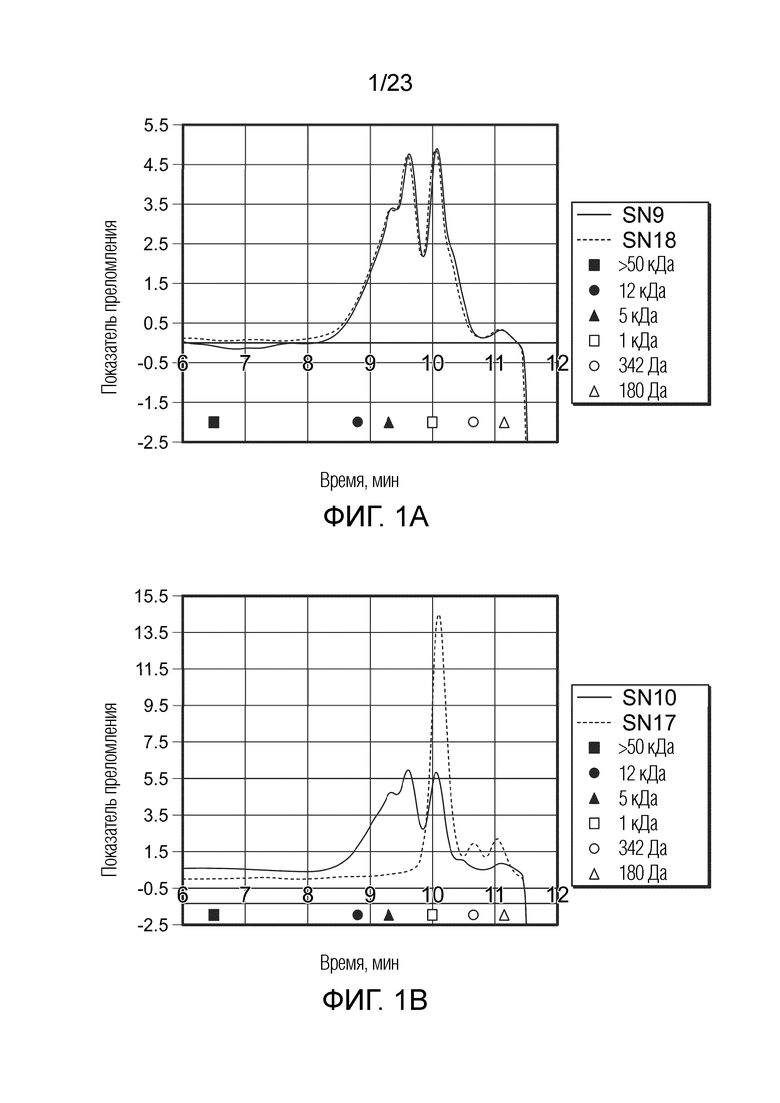
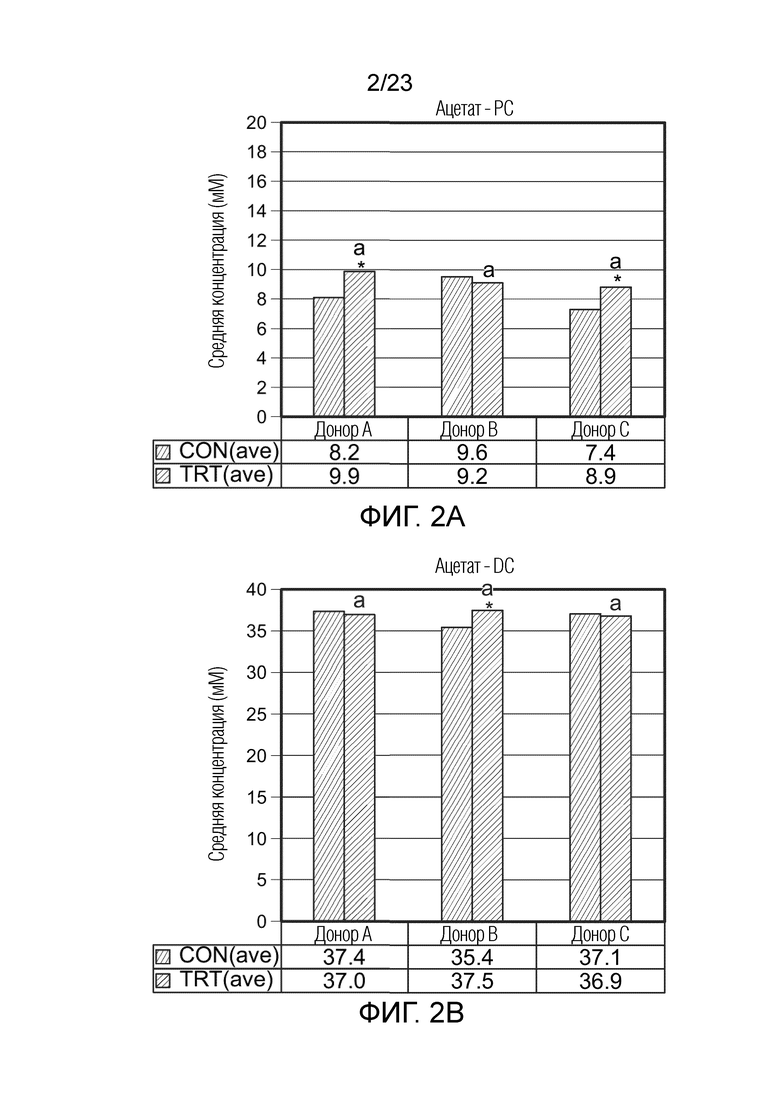
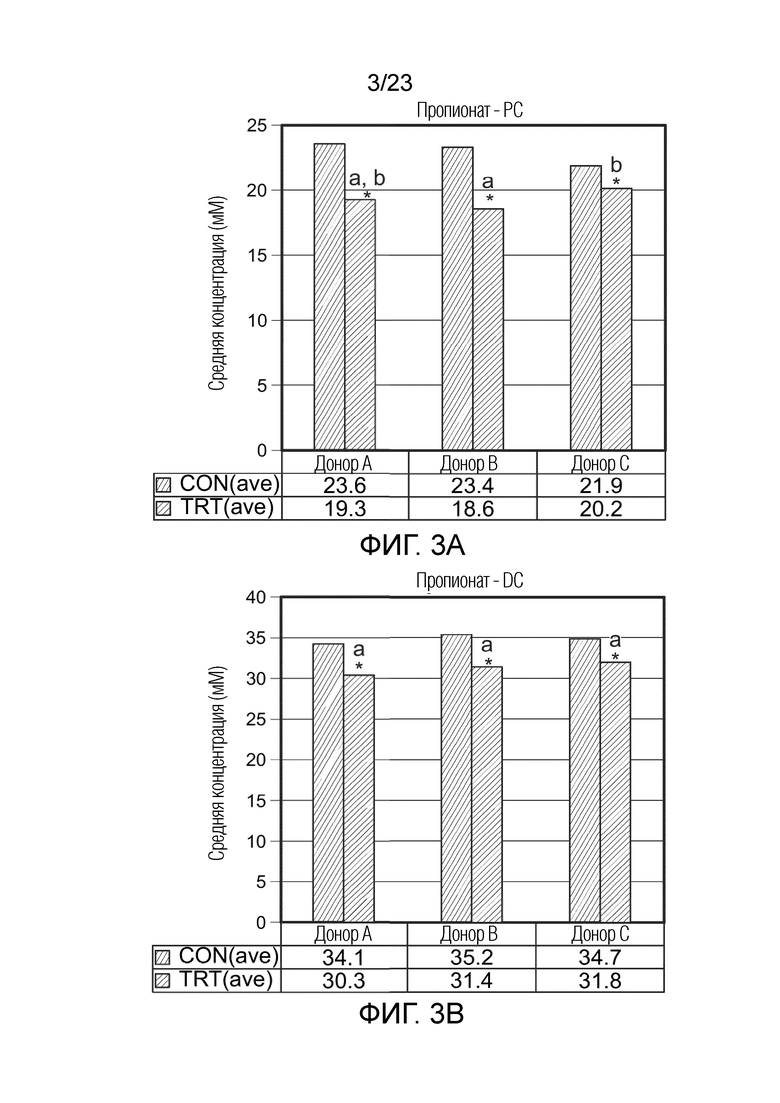

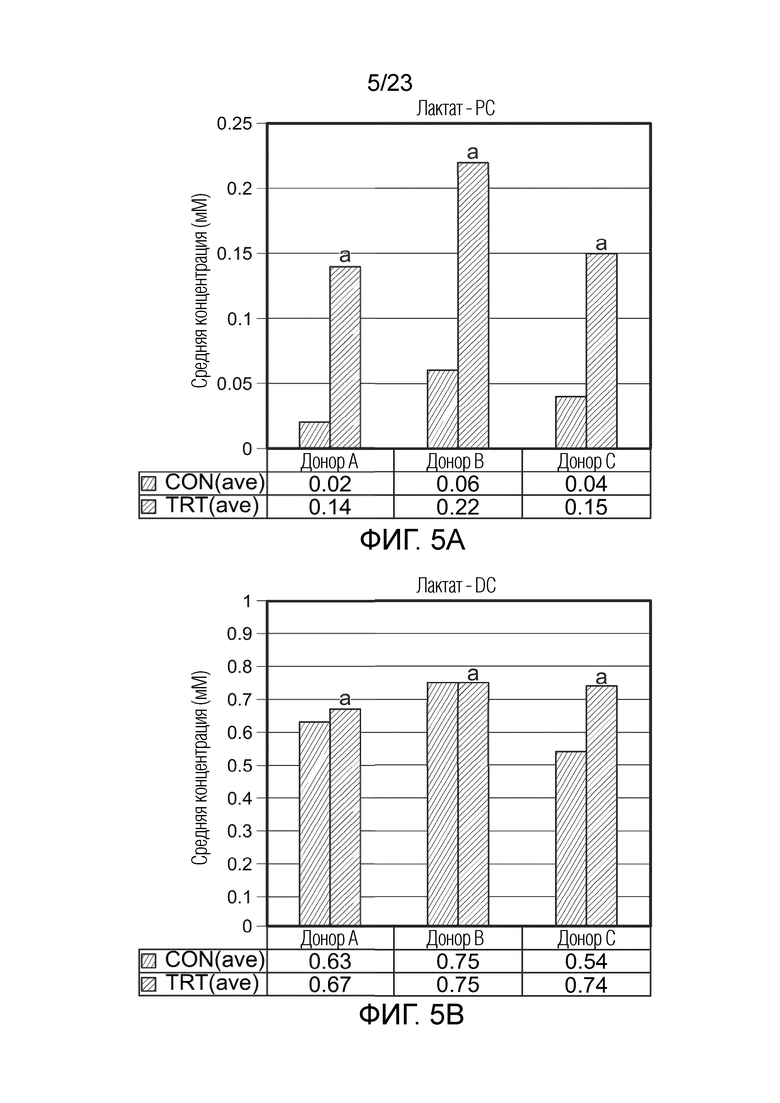

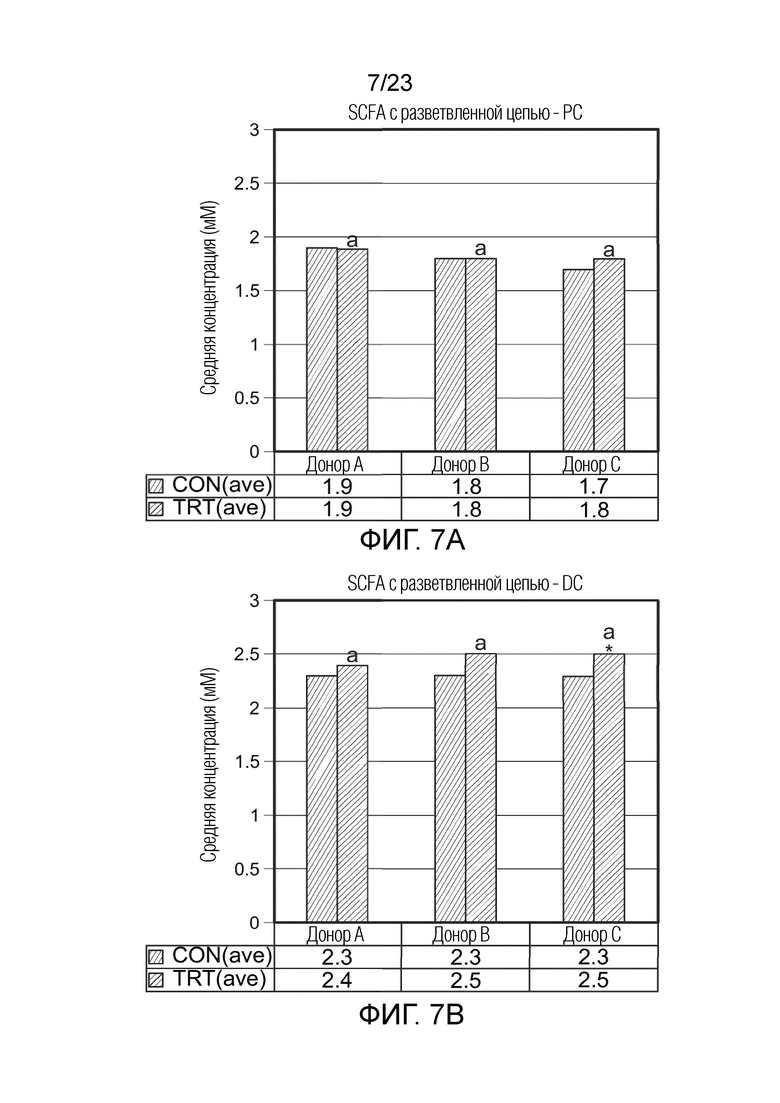
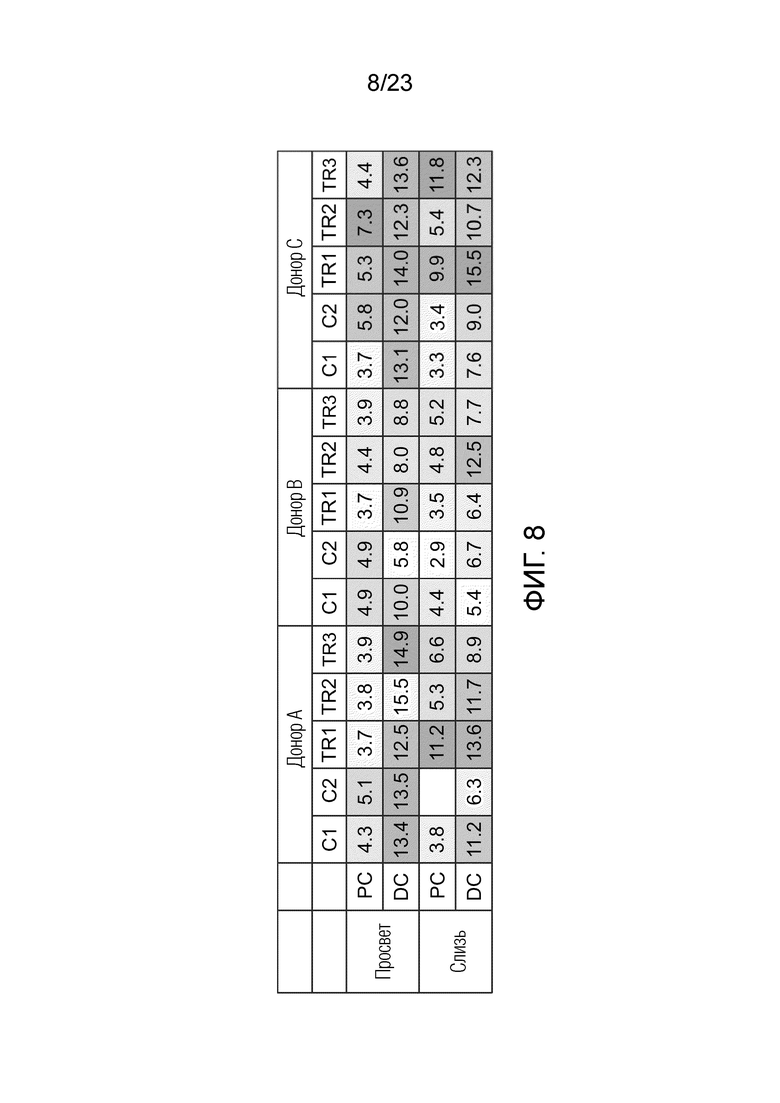





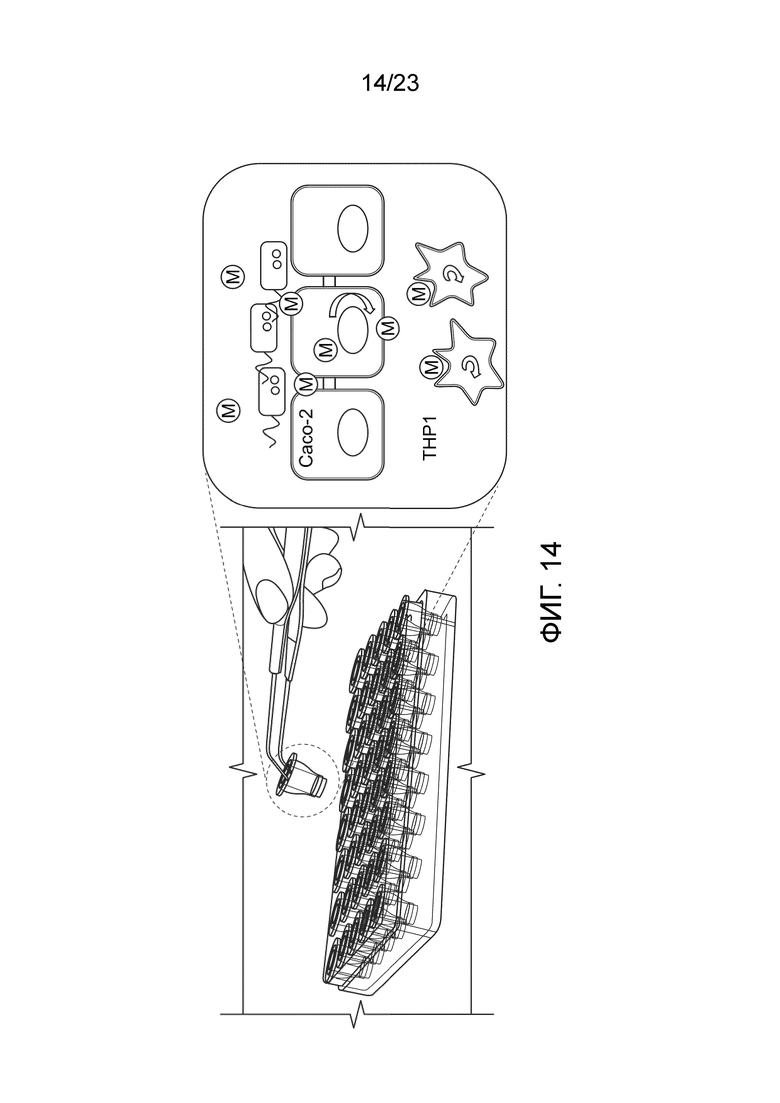
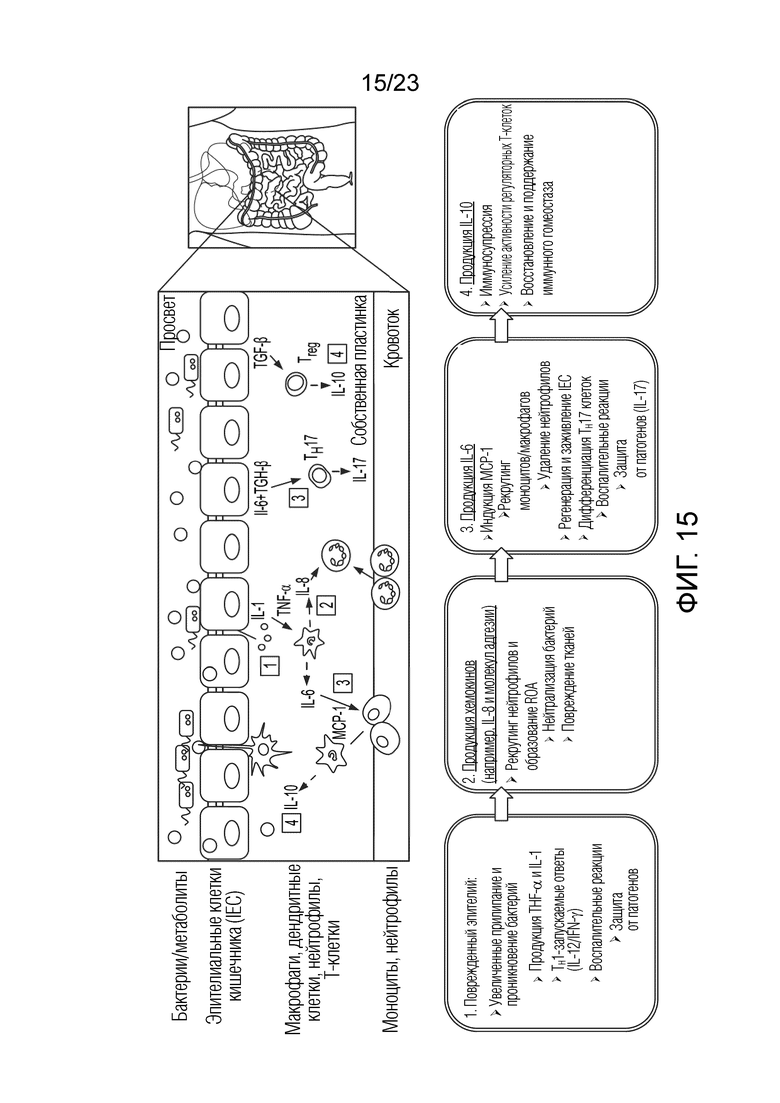
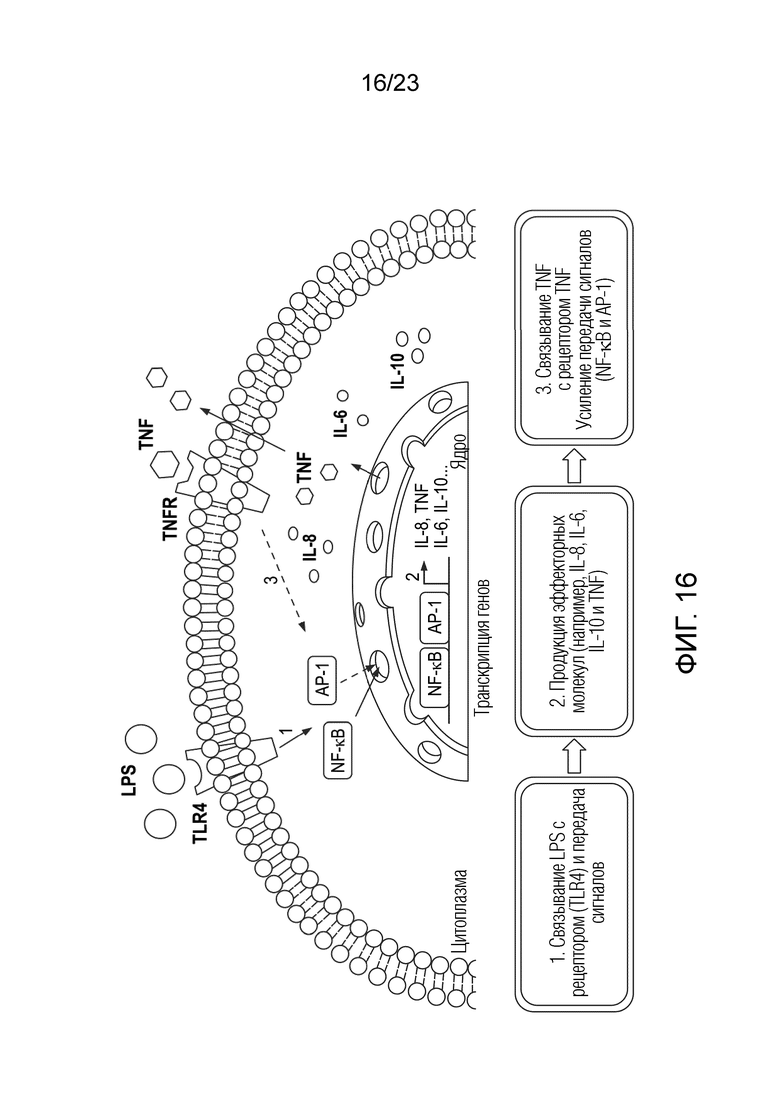

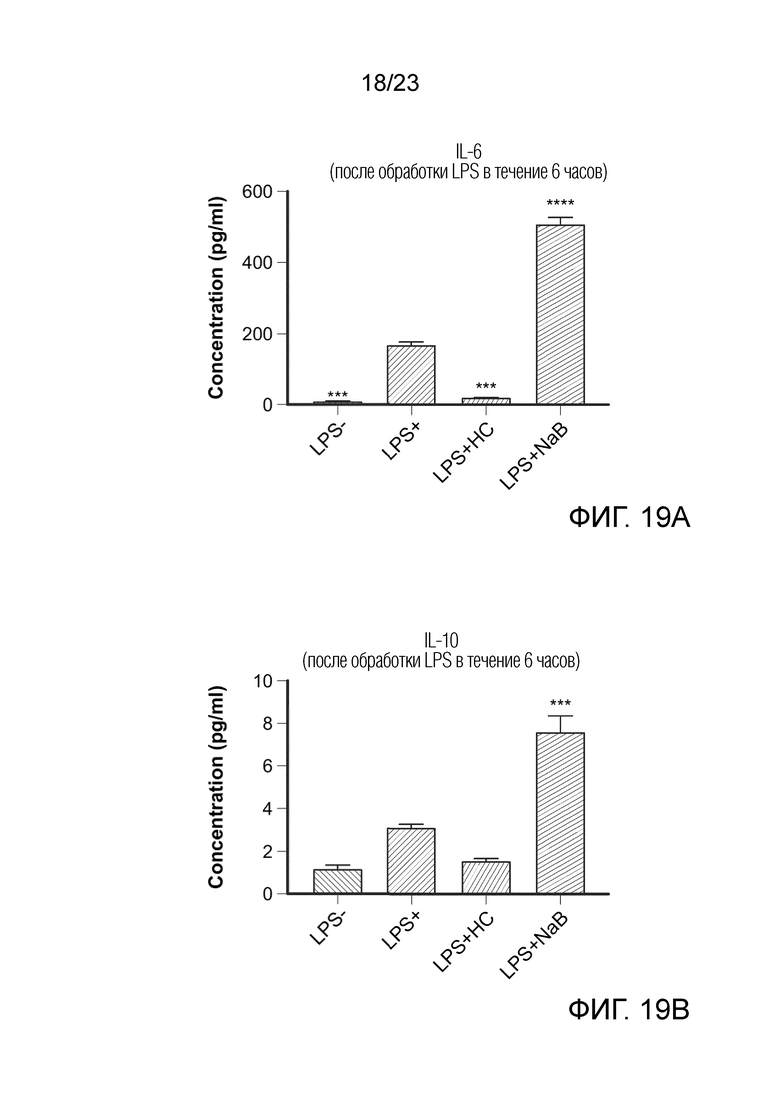
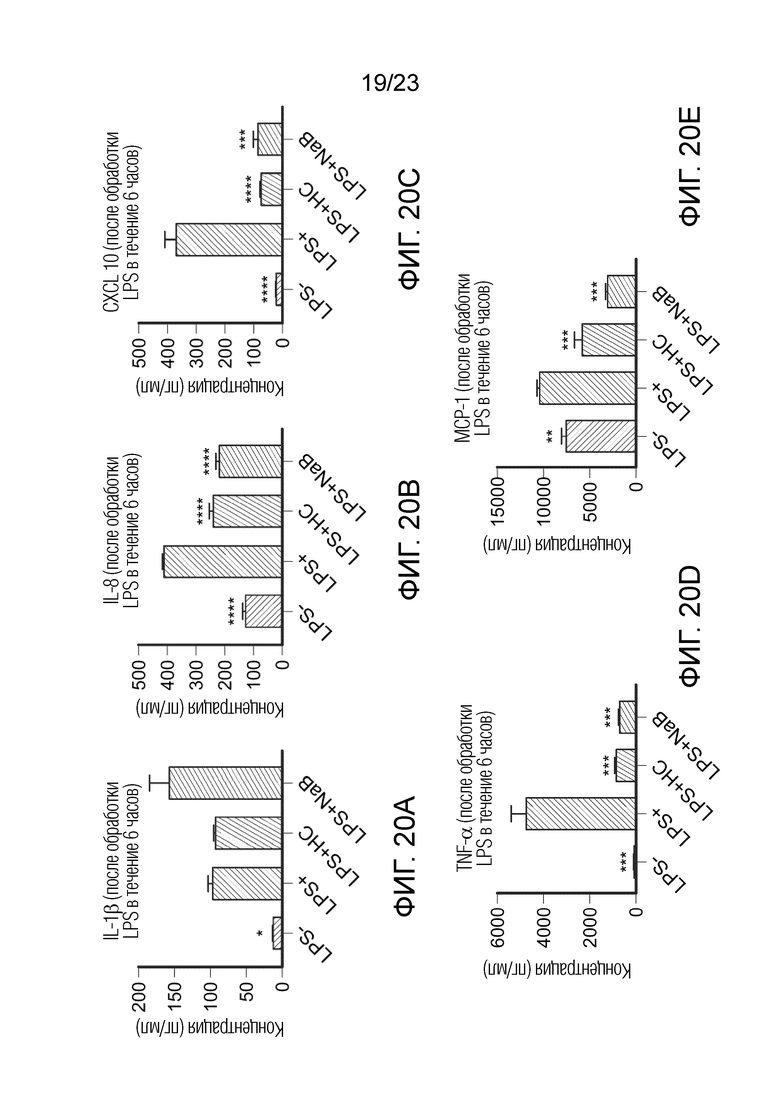
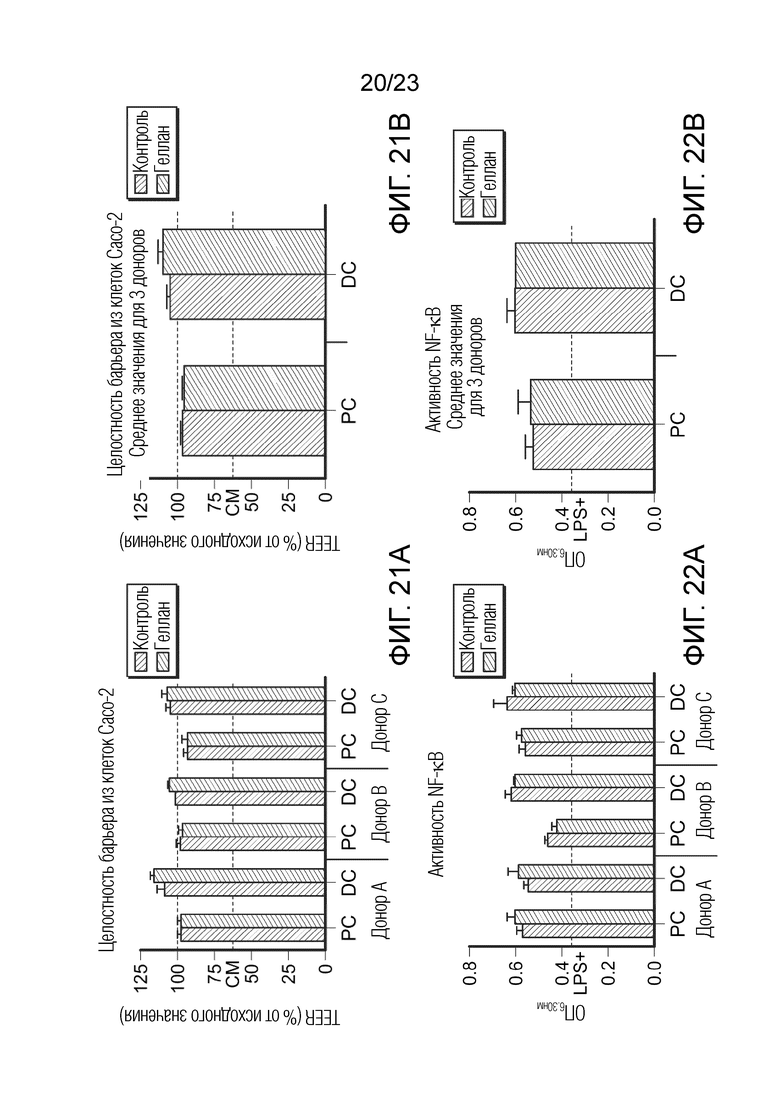
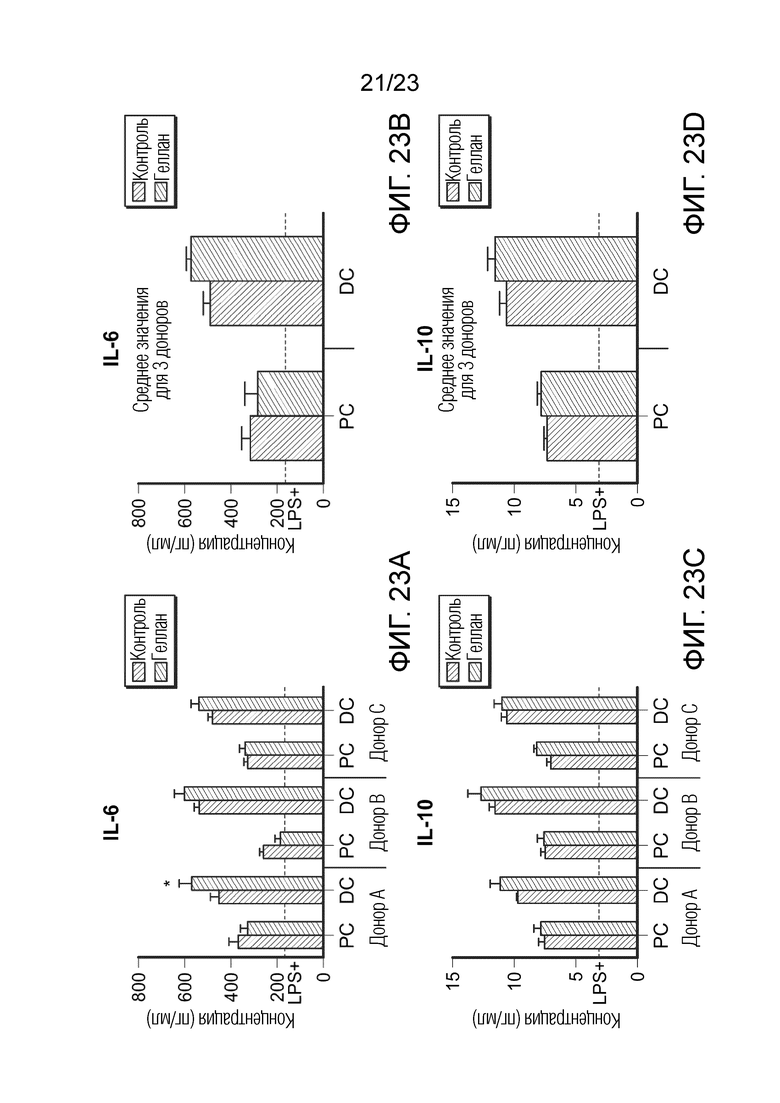
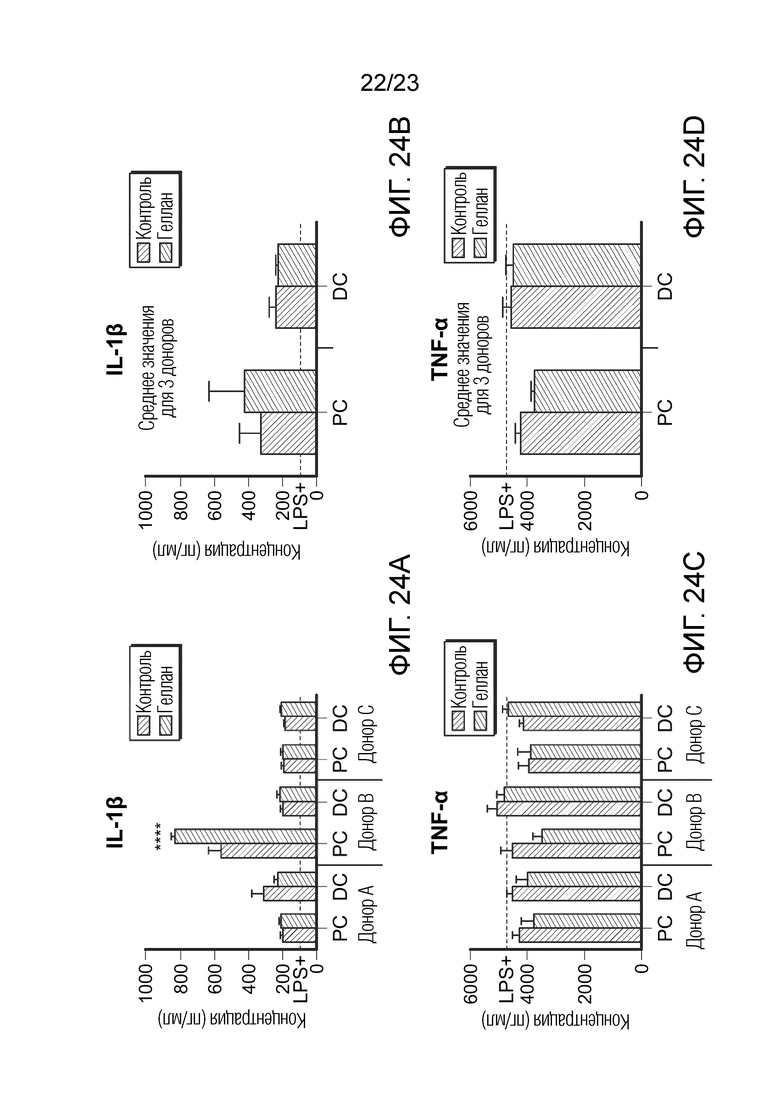
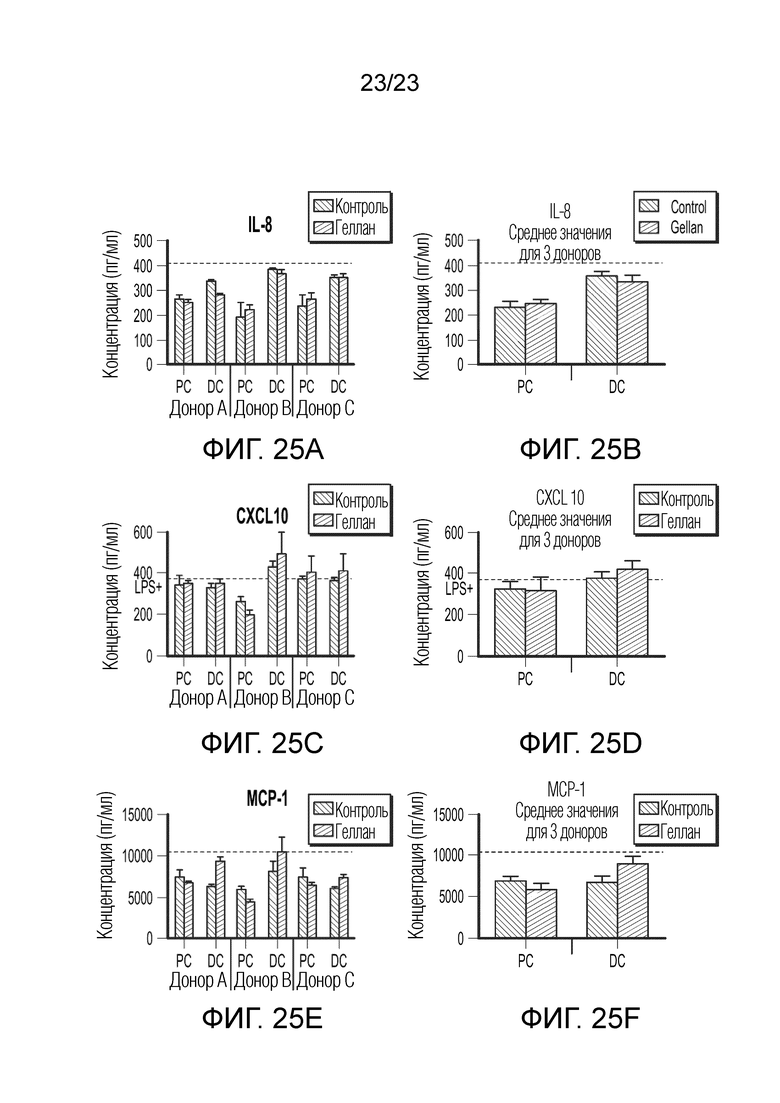
Группа изобретений относится к области медицины и фармацевтики. Предложена пребиотическая композиция, содержащая эффективное для проявления пребиотической активности количество сфингана в виде олигосахарида с DP от 2 до 30. Также предложены способы получения высоко-/средне-/низкоацильного сфингана в виде олигосахарида («SOS») (варианты), способ стимуляции роста полезных бактерий в ободочной кишке млекопитающего, способ снижения уровня пропионата и/или повышения уровня бутирата в ободочной кишке млекопитающего, способ улучшения целостности кишечного барьера в ободочной кишке млекопитающего и способ снижения уровней TNF-α и/или IL-8 в ободочной кишке млекопитающего. Изобретение обеспечивает высокоэффективную пребиотическую композицию. 7 н. и 9 з.п. ф-лы, 25 ил., 16 табл., 4 пр.
1. Пребиотическая композиция, содержащая эффективное для проявления пребиотической активности количество сфингана в виде олигосахарида с DP от 2 до 30.
2. Пребиотическая композиция по п. 1, в которой количество сфингана выбрано из от 1 г до 10 г, от 1 г до 9 г, от 1 г до 8 г, от 1 г до 7 г, от 1 г до 6 г, от 1 г до 5 г, от 1 г до 4 г, от 1 г до 3 г или 2 г.
3. Пребиотическая композиция по п. 1 или 2, в которой сфинган включает высокоацильный сфинган в виде олигосахарида, среднеацильный сфинган в виде олигосахарида, низкоацильный сфинган в виде олигосахарида или их комбинацию.
4. Пребиотическая композиция по п. 1 или 2, в которой сфинган в виде олигосахарида включает высоко-, средне- или низкоацильный сфинган в виде олигосахарида (SOS), полученного способом, который включает
получение первой композиции, содержащей высоко-/средне-/низкоацильный сфинган или высоко-/средне-/низкоацильный сфинган в виде полисахарида и жидкую среду;
гидролиз гликозидной связи высоко-/средне-/низкоацильного сфингана или высоко-/средне-/низкоацильного сфингана в виде полисахарида с получением второй композиции;
осуществление ультрафильтрации, гель-фильтрации, преципитации, центрифугирования второй композиции или их комбинации с получением третьей композиции, содержащей высоко-/средне-/низкоацильный SOS; и,
необязательно, выделение или извлечение третьей композиции.
5. Пребиотическая композиция по любому из пп. 1, 2 и 3, 4, в которой сфинган в виде олигосахарида включает высоко-/средне-/высокоацильный сфинган в виде олигосахарида, выбранного из:
(i) Glc,GlcA, Glc,GlcA,Glyc, Glc,GlcA,Rha, Glc,GlcA,Rha,Glyc, Glc,GlcA,Rha,-H2O, Glc,Rha, Glc,Rha+28, Glc2,GlcA, Glc2,GlcA,Rha, Glc2,GlcA,Rha,+28, Glc2,GlcA,Rha,Ac, Glc2,GlcA,Rha,Glyc, Glc2,GlcA,Rha,Glyc,+28, Glc2,GlcA,Rha,Glyc.-H2O, Glc2,GlcA,Rha,-H2O, Glc2,GlcA,Rha2,Glyc, Glc2,GlcA2,Rha, Glc2,GlcA2,Rha2,Ac2,Glyc2,-H2O, Glc2,Rha, Glc3,GlcA,Rha, Glc3,GlcA,Rha2, Glc3,GlcA,Rha2, Glc3,GlcA,Rha2, Glc3,GlcA,Rha2,Glyc, Glc3,GlcA2,Rha, Glc3,GlcA2,Rha,Glyc, Glc3,GlcA2,Rha2,Glyc, Glc3,GlcA3,Rha2, Glc3,GlcA3,Rha2, Glc4,GlcA,Rha2,+43, Glc4,GlcA,Rha2,Ac, Glyc, Glc4,GlcA2,Rha, Glc4,GlcA2,Rha,Ac,Glyc,-H2O, Glc4,GlcA2,Rha,Ac,Glyc2, Glc4,GlcA2,Rha2,Ac,Glyc, Glc4,GlcA2,Rha2,Glyc, Glc4,GlcA3,Rha2, Glc4,GlcA2,Rha3,Ac, Glc4,GlcA3,Rha2/Glc4,GlcA2,Rha2,Glyc2, Glc5,GlcA2,Rha2, Glc5,GlcA2,Rha2, Glc5,GlcA2,Rha2,Ac, Glc5,GlcA4,Rha2, Glc6,GlcA3,Rha3, Glc(Ac/Glyc)x,GlcAx,Glcx,Rhax (где x составляет от 4 до приблизительно 25), или их комбинацию;
(ii) тетрамера (Glc, GlcA,Glc, Rha), тетрамера (Glc,GlcA,Glc,Rha) с ацетатом и/или глицератом, октамера (Glc,GlcA,Glc,Rha,Glc,GlcA,Glc,Rha), октамера (Glc,GlcA,Glc,Rha,Glc,GlcA,Glc,Rha) с ацетатом и/или глицератом, Glc,GlcA,Glc, Rha,Glc,GlcA, Glc,Rha;
(iii) тетрамера (Glc,GlcA,Glc,Rha), октамера (Glc,GlcA,Glc,Rha,Glc,GlcA,Glc,Rha), пентамера (Glc,GlcA,Glc,Rha,Glc), GlcA,Glc,Rha, Glc,GlcA,Glc, Glc,GlcA;
(iv) Glc(Glc-Glc),GlcA, Glc(Glc-Glc), GlcA,Glc, Glc,Glc;
(v) тетрамера (Glc,GlcA,Glc,Rha), GlcA,Glc,(Rha-Rha), Glc,(Rha-Rha),Rha, GlcA,Glc,Rha, Glc,GlcA,Glc, Rha,Glc, GlcA,Glc;
(vi) Glc,GlcA, Glc,GlcA,Glyc, Glc,GlcA,Rha, Glc,GlcA,Rha,Glyc, Glc,Rha, Glc,Rha+28, Glc2,GlcA, Glc2,GlcA,Rha, Glc2,GlcA,Rha,+28, Glc2,GlcA,Rha,Ac, Glc2,GlcA,Rha,Glyc, Glc2,GlcA,Rha,Glyc,+28, Glc3,GlcA,Rha, Glc3,GlcA,Rha2, Glc3,GlcA,Rha2, Glc3,GlcA,Rha2, Glc3,GlcA,Rha2,Glyc, Glc3,GlcA2,Rha,Glyc, Glc3,GlcA2,Rha2,Glyc, Glc3,GlcA3,Rha2, Glc4,GlcA,Rha2,Ac, Glyc, Glc4,GlcA2,Rha2,Ac,Glyc, Glc4,GlcA2,Rha2,Glyc, Glc4,GlcA2,Rha3,Ac, Glc4,GlcA3,Rha2/Glc4,GlcA2,Rha2,Glyc2, Glc5,GlcA2,Rha2, Glc5,GlcA2,Rha2,Ac, Glc(Ac/Glyc)x,GlcAx,Glcx,Rhax (где x составляет от 4 до приблизительно 25), или их комбинации;
(vii) Glc,GlcA, Glc,GlcA,Rha, Glc,Rha, Glc,Rha+28, Glc2,GlcA,Rha, Glc2,GlcA,Rha,+28, Glc2,GlcA2,Rha, Glc3,GlcA,Rha, Glc3,GlcA,Rha2, Glc3,GlcA2,Rha, Glc3,GlcA3,Rha2, Glc3,GlcA3,Rha2, Glc4,GlcA,Rha2,+43, Glc4,GlcA2,Rha, Glc4,GlcA3,Rha2, Glc5,GlcA2,Rha2, Glc5,GlcA2,Rha2, Glc5,GlcA4,Rha2, Glc6,GlcA3,Rha3, Glc(Ac/Glyc)x,GlcAx,Glcx,Rhax (где x составляет от 4 до 25), или их комбинации;
(viii) Glc,GlcA,Rha,-H2O, Glc,Rha, Glc2,GlcA,Rha,-H2O, Glc2,Rha;
(ix) Glc,GlcA, Glc,GlcA,Glyc, Glc,GlcA,Rha, Glc,GlcA,Rha,Glyc, Glc,Rha, Glc,Rha+28, Glc2,GlcA, Glc2,GlcA,Rha, Glc2,GlcA,Rha,+28, Glc2,GlcA,Rha,Ac, Glc2,GlcA,Rha,Glyc, Glc2,GlcA,Rha,Glyc,+28, Glc2,GlcA,Rha,Glyc.-H2O, Glc2,GlcA,Rha2,Glyc, Glc2,GlcA2,Rha2,Ac2,Glyc2,-H2O, Glc3,GlcA,Rha, Glc3,GlcA,Rha2, Glc3,GlcA,Rha2,Glyc, Glc3,GlcA2,Rha,Glyc, Glc3,GlcA2,Rha2,Glyc, Glc3,GlcA3,Rha2, Glc4,GlcA,Rha2,+43, Glc4,GlcA,Rha2,Ac, Glyc, Glc4,GlcA2,Rha,Ac,Glyc,-H2O, Glc4,GlcA2,Rha,Ac,Glyc2, Glc4,GlcA2,Rha2,Ac,Glyc, Glc4,GlcA2,Rha2,Glyc, Glc4,GlcA3,Rha2, Glc4,GlcA2,Rha3,Ac, Glc4,GlcA3,Rha2/Glc4,GlcA2,Rha2,Glyc2, Glc5,GlcA2,Rha2, Glc5,GlcA2,Rha2,Ac, Glc(Ac/Glyc)x,GlcAx,Glcx,Rhax (где x составляет от 4 до 25), или их комбинации;
или их комбинации.
6. Способ стимуляции роста полезных бактерий в ободочной кишке млекопитающего, причем указанный способ включает прием внутрь по эффективной схеме, которая может включать (i) ежедневный прием внутрь, например один, два, три раза в день и т.д.; или (ii) еженедельный прием внутрь, например каждый день в течение семи дней, через день в течение семи дней и т.д.; или (iii) ежемесячный прием, например ежедневный прием в течение желательного периода времени с последующим периодом отдыха с последующим ежедневным приемом внутрь в течение желаемого периода времени; эффективного для стимуляции роста полезных бактерий количества пребиотической композиции, содержащей пребиотически эффективное количество сфингана в виде олигосахарида по любому из пп. 1-5 и среду для приема внутрь.
7. Способ снижения уровня пропионата и/или повышения уровня бутирата в ободочной кишке млекопитающего, причем указанный способ включает: прием внутрь по эффективной схеме, которая может включать (i) ежедневный прием, например один, два, три раза в день и т.д.; или (ii) еженедельный прием внутрь, например каждый день в течение семи дней, через день в течение семи дней и т.д.; или (iii) ежемесячный прием, например ежедневный прием в течение желательного периода времени с последующим периодом отдыха с последующим ежедневным приемом внутрь в течение желательного периода времени; композиции, содержащей эффективное количество пребиотической композиции, содержащей пребиотически эффективное количество сфингана в виде олигосахарида по пп. 1-5, и среду для приема внутрь.
8. Способ улучшения целостности кишечного барьера в ободочной кишке млекопитающего, причем указанный способ включает: прием внутрь по эффективной схеме, которая может включать (i) ежедневный прием, например один, два, три раза в день и т.д.; или (ii) еженедельный прием внутрь, например каждый день в течение семи дней, через день в течение семи дней и т.д.; или (iii) ежемесячный прием, например ежедневный прием в течение желательного периода времени с последующим периодом отдыха с последующим ежедневным приемом внутрь в течение желаемого периода времени; композиции, содержащей эффективное для целостности кишечного барьера количество пребиотической композиции, содержащей пребиотически эффективное количество сфингана в виде олигосахарида по пп. 1-5, и среду для приема внутрь.
9. Способ снижения уровней TNF-α и/или IL-8 в ободочной кишке млекопитающего, причем указанный способ включает: прием внутрь по эффективной схеме, которая может включать (i) ежедневный прием внутрь, например один, два, три раза в день и т.д.; или (ii) еженедельный прием внутрь, например каждый день в течение семи дней, через день в течение семи дней и т.д.; или (iii) ежемесячный прием, например ежедневный прием в течение желательного периода времени с последующим периодом отдыха с последующим ежедневным приемом внутрь в течение желаемого периода времени; композиции, содержащей эффективное для снижения TNF-α и/или IL-8 количество пребиотической композиции, содержащей пребиотически эффективное количество сфингана в виде олигосахарида по пп. 1-5, и среду для приема внутрь.
10. Способ по пп. 6-9, в котором млекопитающим является человек, и количество сфингана выбрано из от 10 мг/кг до 150 мг/кг, от 10 мг/кг до 140 мг/кг, от 10 мг/кг до 130 мг/кг, от 10 мг/кг до 120 мг/кг, от 10 мг/кг до 110 мг/кг, от 10 мг/кг до 100 мг/кг, от 10 мг/кг до 90 мг/кг, от 10 мг/кг до 80 мг/кг, от 10 мг/кг до 70 мг/кг, от 10 мг/кг до 60 мг/кг, от 10 мг/кг до 50 мг/кг, от 10 мг/кг до 40 мг/кг или от 20 мг/кг до 30 мг/кг массы тела человека, принимающего внутрь композицию.
11. Способ по пп. 6-10, в котором сфинган включает высоко-, средне- или низкоацильный сфинган в виде олигосахарида (SOS), полученного способом, который включает:
получение первой композиции, содержащей высоко-/средне-/низкоацильный сфинган или высоко-/средне-/низкоацильный сфинган в виде полисахарида и жидкую среду;
гидролиз гликозидной связи высоко-/средне-/низкоацильного сфингана или высоко-/средне-/низкоацильного сфингана в виде полисахарида с получением второй композиции;
осуществление ультрафильтрации, гель-фильтрации, преципитации, центрифугирования второй композиции или их комбинации с получением третьей композиции, содержащей высоко-/средне-/низкоацильный SOS; и,
необязательно, выделение или извлечение третьей композиции.
12. Способ по любому из пп. 6-11, в котором сфинган включает высоко-/средне-/высокоацильный сфинган в виде олигосахарида, выбранного из:
(i) Glc,GlcA, Glc,GlcA,Glyc, Glc,GlcA,Rha, Glc,GlcA,Rha,Glyc, Glc,GlcA,Rha,-H2O, Glc,Rha, Glc,Rha+28, Glc2,GlcA, Glc2,GlcA,Rha, Glc2,GlcA,Rha,+28, Glc2,GlcA,Rha,Ac, Glc2,GlcA,Rha,Glyc, Glc2,GlcA,Rha,Glyc,+28, Glc2,GlcA,Rha,Glyc.-H2O, Glc2,GlcA,Rha,-H2O, Glc2,GlcA,Rha2,Glyc, Glc2,GlcA2,Rha, Glc2,GlcA2,Rha2,Ac2,Glyc2,-H2O, Glc2,Rha, Glc3,GlcA,Rha, Glc3,GlcA,Rha2, Glc3,GlcA,Rha2, Glc3,GlcA,Rha2, Glc3,GlcA,Rha2,Glyc, Glc3,GlcA2,Rha, Glc3,GlcA2,Rha,Glyc, Glc3,GlcA2,Rha2,Glyc, Glc3,GlcA3,Rha2, Glc3,GlcA3,Rha2, Glc4,GlcA,Rha2,+43, Glc4,GlcA,Rha2,Ac, Glyc, Glc4,GlcA2,Rha, Glc4,GlcA2,Rha,Ac,Glyc,-H2O, Glc4,GlcA2,Rha,Ac,Glyc2, Glc4,GlcA2,Rha2,Ac,Glyc, Glc4,GlcA2,Rha2,Glyc, Glc4,GlcA3,Rha2, Glc4,GlcA2,Rha3,Ac, Glc4,GlcA3,Rha2/Glc4,GlcA2,Rha2,Glyc2, Glc5,GlcA2,Rha2, Glc5,GlcA2,Rha2, Glc5,GlcA2,Rha2,Ac, Glc5,GlcA4,Rha2, Glc6,GlcA3,Rha3, Glc(Ac/Glyc)x,GlcAx,Glcx,Rhax (где x составляет от 4 до 25), Glcx, GlcAx,Glcx, Rhax (где x составляет от 4 до 25), или их комбинацию;
(ii) тетрамера (Glc,GlcA,Glc,Rha), тетрамера (Glc,GlcA,Glc,Rha) с ацетатом и/или глицератом, октамера (Glc,GlcA,Glc,Rha,Glc,GlcA,Glc,Rha), октамера (Glc,GlcA,Glc,Rha,Glc,GlcA,Glc,Rha) с ацетатом и/или глицератом, Glc,GlcA,Glc, Rha,Glc,GlcA, Glc,Rha;
(iii) тетрамера (Glc,GlcA,Glc,Rha), октамера (Glc,GlcA,Glc,Rha,Glc,GlcA,Glc,Rha), пентамера (Glc,GlcA,Glc,Rha,Glc), GlcA,Glc,Rha, Glc,GlcA,Glc, Glc,GlcA;
(iv) Glc(Glc-Glc),GlcA, Glc(Glc-Glc), GlcA,Glc, Glc,Glc;
(v) тетрамера (Glc,GlcA,Glc,Rha), GlcA,Glc,(Rha-Rha), Glc,(Rha-Rha),Rha, GlcA,Glc,Rha, Glc,GlcA,Glc, Rha,Glc, GlcA,Glc;
(vi) Glc,GlcA, Glc,GlcA,Glyc, Glc,GlcA,Rha, Glc,GlcA,Rha,Glyc, Glc,Rha, Glc,Rha+28, Glc2,GlcA, Glc2,GlcA,Rha, Glc2,GlcA,Rha,+28, Glc2,GlcA,Rha,Ac, Glc2,GlcA,Rha,Glyc, Glc2,GlcA,Rha,Glyc,+28, Glc3,GlcA,Rha, Glc3,GlcA,Rha2, Glc3,GlcA,Rha2, Glc3,GlcA,Rha2, Glc3,GlcA,Rha2,Glyc, Glc3,GlcA2,Rha,Glyc, Glc3,GlcA2,Rha2,Glyc, Glc3,GlcA3,Rha2, Glc4,GlcA,Rha2,Ac, Glyc, Glc4,GlcA2,Rha2,Ac,Glyc, Glc4,GlcA2,Rha2,Glyc, Glc4,GlcA2,Rha3,Ac, Glc4,GlcA3,Rha2/Glc4,GlcA2,Rha2,Glyc2, Glc5,GlcA2,Rha2, Glc5,GlcA2,Rha2,Ac, Glc(Ac/Glyc)x,GlcAx,Glcx,Rhax (где x составляет от 4 до 25), или их комбинации;
(vii) Glc,GlcA, Glc,GlcA,Rha, Glc,Rha, Glc,Rha+28, Glc2,GlcA,Rha, Glc2,GlcA,Rha,+28, Glc2,GlcA2,Rha, Glc3,GlcA,Rha, Glc3,GlcA,Rha2, Glc3,GlcA2,Rha, Glc3,GlcA3,Rha2, Glc3,GlcA3,Rha2, Glc4,GlcA,Rha2,+43, Glc4,GlcA2,Rha, Glc4,GlcA3,Rha2, Glc5,GlcA2,Rha2, Glc5,GlcA2,Rha2, Glc5,GlcA4,Rha2, Glc6,GlcA3,Rha3, Glc(Ac/Glyc)x,GlcAx,Glcx,Rhax (где x составляет от 4 до 25), или их комбинации;
(viii) Glc,GlcA,Rha,-H2O, Glc,Rha, Glc2,GlcA,Rha,-H2O, Glc2,Rha;
(ix) Glc,GlcA, Glc,GlcA,Glyc, Glc,GlcA,Rha, Glc,GlcA,Rha,Glyc, Glc,Rha, Glc,Rha+28, Glc2,GlcA, Glc2,GlcA,Rha, Glc2,GlcA,Rha,+28, Glc2,GlcA,Rha,Ac, Glc2,GlcA,Rha,Glyc, Glc2,GlcA,Rha,Glyc,+28, Glc2,GlcA,Rha,Glyc.-H2O, Glc2,GlcA,Rha2,Glyc, Glc2,GlcA2,Rha2,Ac2,Glyc2,-H2O, Glc3,GlcA,Rha, Glc3,GlcA,Rha2, Glc3,GlcA,Rha2,Glyc, Glc3,GlcA2,Rha,Glyc, Glc3,GlcA2,Rha2,Glyc, Glc3,GlcA3,Rha2, Glc4,GlcA,Rha2,+43, Glc4,GlcA,Rha2,Ac, Glyc, Glc4,GlcA2,Rha,Ac,Glyc,-H2O, Glc4,GlcA2,Rha,Ac,Glyc2, Glc4,GlcA2,Rha2,Ac,Glyc, Glc4,GlcA2,Rha2,Glyc, Glc4,GlcA3,Rha2, Glc4,GlcA2,Rha3,Ac, Glc4,GlcA3,Rha2/Glc4,GlcA2,Rha2,Glyc2, Glc5,GlcA2,Rha2, Glc5,GlcA2,Rha2,Ac, Glc(Ac/Glyc)x,GlcAx,Glcx,Rhax (где x составляет от 4 до 25), или их комбинации;
или их комбинации.
13. Способ по п. 6, в котором сфинган включает встречающийся в природе сфинган, а бактериями являются Bifidobacteriaceae.
14. Способ по п. 6, в котором млекопитающим является человек, и сфинган включает высоко-/низкоацильный сфинган в виде олигосахарида, а бактериями являются Blautia, Parabacteroides, Faecalibacterium, Clostridium XVIII или их комбинация.
15. Способ получения высоко-/средне-/низкоацильныого сфингана в виде олигосахарида («SOS») по любому из пп. 1-5, который включает:
получение первой композиции, содержащей высоко-/средне-/низкоацильный сфинган или высоко-/средне-/низкоацильный сфинган в виде полисахарида и жидкую среду;
гидролиз гликозидной связи высоко-/средне-/низкоацильного сфингана или высоко-/средне-/низкоацильного сфингана в виде полисахарида с получением второй композиции;
осуществление ультрафильтрации, гель-фильтрации, преципитации, центрифугирования второй композиции или их комбинации с получением третьей композиции, содержащей высоко-/средне-/низкоацильный SOS; и
необязательно выделение или извлечение третьей композиции;
причем указанный гидролиз опосредован ферментом, которым является гелланaза, рамногалактуронанэндолиаза (EC 4.2.2.23), рамногалактуронанэкзолиаза (EC 4.2.2.24), гелланлиаза (EC 4.2.2.25) или их комбинация.
16. Способ получения высоко-/средне-/низкоацильного сфингана в виде олигосахарида («SOS») по любому из пп. 1-5, который включает:
получение первой композиции, содержащей высоко-/средне-/низкоацильный сфинган или высоко-/средне-/низкоацильный сфинган в виде полисахарида и жидкую среду;
гидролиз гликозидной связи высоко-/средне-/низкоацильного сфингана или высоко-/средне-/низкоацильного сфингана в виде полисахарида с получением второй композиции;
осуществление ультрафильтрации, гель-фильтрации, преципитации, центрифугирования второй композиции или их комбинации с получением третьей композиции, содержащей высоко-/средне-/низкоацильный SOS; и
необязательно выделение или извлечение третьей композиции, причем указанное осуществление включает фильтрацию второй композиции через мембрану с отсечением по молекулярной масса, составляющим или приблизительно 5 кДа, или приблизительно 10 кДа, с получением фильтрата, включающего третью композицию.
| HUI LI ET AL | |||
| The preparation and characterization of a novel sphingan WL from marine Sphingomonas sp | |||
| WG | |||
| Scientific Reports, 6, Article number: 37899 (2016) | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| WEI JIA ET AL | |||
| Выбрасывающий ячеистый аппарат для рядовых сеялок | 1922 |
|
SU21A1 |
| Способ и приспособление посадки сети на подборные нити при вязке на сетевязальной машине | 1930 |
|
SU31555A1 |
| Carbohydrate Polymers | |||
| Пожарный двухцилиндровый насос | 0 |
|
SU90A1 |

