Изобретение относится к области генетической инженерии, конкретно к биотехнологии, и может быть использовано для трансформации бактерий рода Lactobacillus.
В последние несколько десятков лет активно развивается направление генетической инженерии, связанное с получением гетерологичных белков в клетках модельных организмов, в том числе дрожжей, Е. coli, бактерий родов Bacillus и Lactobacillus [1]. Грамположительные бактерии родов Bacillus и Lactobacillus являются перспективным объектом для генетических манипуляций, поскольку организмы позволяют нарабатывать рекомбинантные белки в больших количествах. Кроме того, в отличие от Е. coli, бактерии родов Bacillus и Lactobacillus способны экспортировать рекомбинантные белки из клеток в питательную среду. Таким образом, значительно упрощается процедуры очистки рекомбинантных белков, поскольку исчезает потребность в разрушении (лизисе) бактериальных клеток с наработанным белком. Еще одно преимущество лактобацилл по сравнению с Е. coli - это отсутствие эндотоксинов в клеточных стенках грамположительных бактерий. Наличие эндотоксинов в белковых препаратах может приводить к развитию пирогенных реакций у людей и животных и затруднять использование белков в фармацевтических целях. Очистка препаратов рекомбинантных белков от эндотоксинов может быть сложным и многоступенчатым процессом, удорожающим стоимость конечного продукта. Как следствие, использование лактобациллы может решить проблему очистки рекомбинантных белков от эндотоксинов.
Бактерии рода Lactobacillus являются безопасными для здоровья человека и могут быть использованы в качестве пробиотических препаратов, нормализующих состав микрофлоры кишечника и повышающих ее активность. Как следствие, лактобациллы могут быть использованы как живой бактериальный вектор для доставки терапевтических белков и белков-антигенов непосредственно к слизистой оболочке кишечника.
Использование лактобацилл в генетической инженерии сталкивается с рядом трудностей. Одной из самых серьезных проблем является сложность получения компетентных клеток лактобацилл, пригодных для трансформации с использованием плазмидной ДНК [2]. Толстая клеточная стенка лактобацилл, состоящая из пептидогликанов, способна выдерживать действие ферментов пищеварительного тракта человека и животных. Однако этот же слой пептидогликанов является препятствием для проникновения чужеродной ДНК в клетку. По этой причине для введения плазмидной ДНК в клетки лактобацилл наиболее часто применяется электропорация, как метод, позволяющий преодолеть толстую клеточную стенку за счет воздействия электрического поля высокой напряженности. Однако проведение электропорации требует использования специальных приборов, создающих кратковременный импульс электрического поля с высокой напряженностью (до нескольких тысяч вольт). Работа с электричеством при таком напряжении сопряжена с риском получения электротравмы и требует специального инструктажа персонала с регулярной проверкой полученных навыков, а также регулярной поверки электропораторов на предмет их безопасности. Таком образом, требования к приборной базе и электробезопасности налагают ограничения на применения электропорации для трансформации клеток лактобацилл плазмидной ДНК.
Альтернативой электропорации являются методы приготовления химических компетентных клеток, основанные на использовании различных химических веществ с целью получения пор в клеточных стенках бактерий. Такие протоколы не требуют использования электропораторов и более пригодны для рутинной практики. Однако для лактобацилл подобные протоколы практически не описаны. Вместе с тем, у лактобацилл отмечено явление физиологической компетентности, то есть способности бактериальных клеток захватывать и использовать чужеродную ДНК из внешней среды. Этот механизм основан на использовании около 15 белков, кодируемых генами сот, которые формируют сложную структуру на клеточной стенке, предназначенную для захвата ДНК из внешней среды. В ходе этого процесса затрачивается большое количество энергии, поэтому активация механизма естественной компетентности бактериальных клеток требует сложной стимуляции и может занимать длительное время. Таким образом, получение химических компетентных клеток лактобацилл продолжает оставаться задачей, требующей серьезных систематических усилий.
Еще одна проблема, возникающая при трансформации лактобацилл - это потребность в инструментах для масштабного скрининга эффективности различных протоколов трансформации. Используемые на сегодняшних день подходы основаны на прямом подсчете колоний-трансформантов, выращенных на чашках Петри после внедрения чужеродной ДНК в клетки лактобацилл. Однако подобный подсчет требует длительного времени и не поддается масштабированию, что ограничивает проведение масштабного сравнения эффективности различных протоколов получения компетентных клеток лактобацилл. Это же обстоятельство затрудняет сравнение эффективности трансформации разных штаммов лактобацилл, а также сравнение влияния разных химических веществ на трансформацию. Таким образом, в связи с растущим интересом к лактобациллам как к перспективному объекту генетической инженерии, а также недостаточной обработанностью протоколов трансформации, существует потребность в развитии высокопроизводительных методов оценки эффективности трансформации лактобацилл.
Наиболее близкой к заявляемому изобретению - прототипом, является методика приготовления компетентных клеток лактобацилл с последующей электропорацией, предложенная Palomino et al [3]. Способ приготовления компетентных клеток заключается в выращивании лактобацилл в питательной среде MRS с 0,9 М или 0,7 М NaCl до оптической плотности при 600 нм равной 2-2,5 с последующей инкубацией бактериальных культур на льду в течение 10 минут, сбором бактериальных клеток центрифугированием при +4°С, троекратной отмывкой клеточного осадка бидистиллированной ледяной водой и ресуспендированием клеток в бидистиллированной ледяной воде. Электропорацию проводят с помощью прибора Gene Pulser (Bio-Rad, США) в 0,2 см кювете, напряжении 2 кВ, сопротивлении 400 Ом, емкости 25 мкФ, длительности импульса 10 мкс. Для электропорации используют 50 мкл компетентных клеток лактобацилл и 50 нг плазмидной ДНК. После электропорации бактериальные клетки смешивают с 0,95 мл питательной среды MRS и инкубируют 3 часа при 37°С. 20 мкл трансформационной смеси распределяют по чашке Петри с агаризованной питательной средой MRS и нужным антибиотиком, после чего инкубируют чашки Петри при 37°С в анаэробных условиях в течение 48-72 часов. После окончания инкубации подсчитывают количество колоний-трансформантов, несущих плазмиду.
Недостатком способа, предложенного Palomino et al, является необходимость длительной инкубации бактериальных культур после трансформации (в течение 48-72 часов), что значительно замедляет проведение анализа. Кроме того, необходимость прямого подсчета колоний-трансформантов на чашках Петри не позволяет масштабировать процесс и проводить высокопроизводительный скрининг эффективности трансформации.
Задачей изобретения является разработка нового способа оценки эффективности методик введения плазмидной ДНК в компетентные бактерии рода Lactobacillus.
Технический результат: ускорение оценки эффективности разных протоколов приготовления компетентных клеток бактерий рода Lactobacillus, а также возможность одновременно скринировать большое число протоколов приготовления компетентных клеток, химических веществ, повышающих эффективность трансформации, и разных штаммов бактерий рода Lactobacillus.
Поставленная задача достигается предлагаемым способом, заключающимся в следующем.
Проводят трансформацию компетентных клеток бактерий рода Lactobacillus плазмидой pLEM415-ldhL-mRFP1 SEQ ID NO: 1, несущей кодирующую последовательность мономерного красного флуоресцентного белка (monomeric red fluorescent protein, mRFP) под контролем конститутивного промотора гена idhL. Компетентные клетки бактерий рода Lactobacillus могут быть приготовлены по разным методикам и/или из разных штаммов бактерий рода Lactobacillus.
Трансформацию компетентных клеток лактобацилл проводят одновременно, для трансформации используют по 20 мкл компетентных клеток, приготовленных по методике Palomino et al и по 20 нг плазмиды pLEM415-ldhL-mRFP1. Каждую вариант условий трансформации анализируют в 4 повторах, к двум из которых добавляют плазмиду pLEM415-ldhL-mRFP1, еще два повтора служат контролями без плазмиды. Перемешанные суспензии компетентных клеток и раствора плазмиды инкубируют в 0,2 см кюветах для электропорации во льду в течение 5 минут. По истечении срока инкубации переносят кюветы в электропоратор Gene Pulser (Bio-Rad, США) и проводят электропорацию при следующих условиях: напряжение 2 кВ, сопротивление 400 Ом, емкость 25 мкФ, длительность импульса 10 мкс. После электропорации бактериальные суспензии немедленно переносят в низкопрофильный ПЦР-планшет. В две пустые лунки ПЦР-планшета добавляют по 30 мкл питательной среды MRS в качестве контроля стерильности. Во все лунки ПЦР-планшета добавляют по 100 мкл питательной среды MRS, после чего ПЦР-планшет инкубируют в термостате при 37°С в течение 1 часа. После инкубации во все лунки ПЦР-планшета добавляют по 12 мкл раствора эритромицина с концентрацией 40 мг/мл, перемешивают содержимое лунок и оставляют инкубироваться при 37°С в течение 16-18 часов. Далее считывают сигнал флуоресценции по каналу FAM. Количество бактериальных клеток с внедренной контрольной плазмидой оценивают по степени роста флуоресценции по сравнению с контрольными пробами без плазмиды.
Определяющим отличием заявляемого способа по сравнению с прототипом является использование флуоресценции для оценки эффективности трансформации, что позволяет ускорить проведение анализа и одновременно скринировать большое число условий методик приготовления компетентных клеток. Количественная оценка по изменению флуоресценции позволит не проводить процедуру подсчета колоний на чашках Петри, замедляющую процесс анализа результатов трансформации.
Плазмида pLEM415-ldhL-mRFP1 несет в себе три ключевых элемента, необходимых для проведения анализа эффективности трансформации. Первый элемент -ген устойчивости к эритромицину, erm (erythromycin resistance methyltransferase, метилтрансфераза устойчивости к эритромицину). Используя S-аденозин-метионин как кофактор, белок-продукт гена erm переносит метальную группу на аденозин в позиции 2058 в 23S рибосомальной ДНК. Эта модификация блокирует сайт связывания эритромицина с рибосомой, в результате чего бактериальные клетки становятся устойчивыми к действию антибиотика. Ген erm в плазмиде pLEM415-ldhL-mRFP1 позволяет использовать эритромицин в качестве селективного маркера для отбора бактериальных клеток, внутрь которых при трансформации попала плазмида. Добавляемый в среду эритромицин блокирует белковый синтез в клетках без плазмиды, в результате чего выживают и размножаются только клетки, несущие плазмиду pLEM415-ldhL-mRFP1.
Второй элемент - это промотор гена idhL. Ген idhL кодирует L-лактат дегидрогеназу, фермент, присутствующий во всех живых клетках и катализирующий превращение лактата в пируват с восстановленного никотинамидадениндинуклеотида (НАДН). Промотор гена idhL является конститутивным и постоянно активирован, что приводит к постоянной наработке белка, находящегося под контролем этого промотора.
Третий элемент - белок mRFP. Это белок-флуорофор массой 25,9 кДа, который при возбуждении флуоресцирует в красно-оранжевом диапазоне. В плазмиде pLEM415-ldhL-mRFP1 кодирующая последовательность белка mRFP находится под контролем конститутивного промотора гена idhL, в результате чего рекомбинантный белок mRFP начинает нарабатываться в клетке сразу же после попадания плазмиды внутрь бактериальной клетки. Таким образом, ген устойчивости к эритромицину егт обеспечивает выживание бактериальных клеток, несущих после трансформации плазмиду pLEM415-ldhL-mRFP1, а нарабатываемый рекомбинантный белок mRFP придает красную окраску бактериальным клеткам с плазмидой pLEM415-ldhL-mRFP1. Эта окраска может быть наблюдаема невооруженным глазом, а также флуоресценция может быть считана с помощью флуориметров или амплификаторв для ПЦР в режиме реального времени. Степень интенсивности окраски напрямую определяется количеством бактериальных клеток с плазмидой pLEM415-ldhL-mRFP1. В результате, использование плазмиды pLEM415-ldhL-mRFP1 позволяет оценивать количество трансформированных клеток бактерий рода Lactobacillus без рассевания бактериальных культур на чашки Петри с питательной средой.
Для лучшего понимания сущности изобретение иллюстрируются следующими примерами осуществления.
Пример 1.
Анализ эффективности трансформации химических компетентных клеток Lactobacillus reuteri и Lactobacillus plantarum.
Компетентные клетки бактерий Lactobacillus reuteri и Lactobacillus plantarum готовили по Palomino et al. Трансформацию компетентных клеток разных лактобацилл проводили одновременно, для трансформации использовали по 20 мкл компетентных клеток и по 20 нг плазмиды pLEM415-ldhL-mRFP1. Каждый штамм бактерий рода Lactobacillus анализировали в 4 повторах, к двум из которых добавляли плазмиду pLEM415-ldhL-mRFP1, еще два повтора служили контролями без плазмиды.
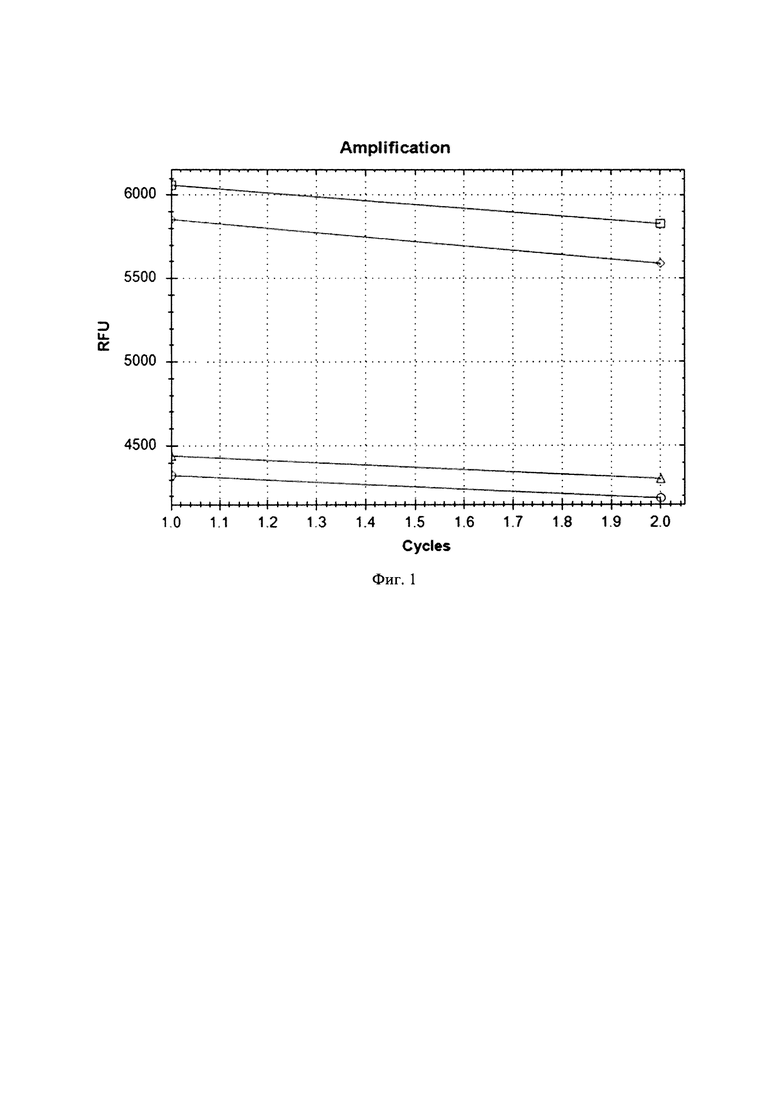
Перемешанные суспензии компетентных клеток и раствора плазмиды инкубировали в 0,2 см кюветах для электропорации во льду в течение 5 минут. По истечении срока инкубации переносили кюветы в электропоратор Gene Pulser (Bio-Rad, США) и проводили электропорацию при следующих условиях: напряжение 2 кВ, сопротивление 400 Ом, емкость 25 мкФ, длительность импульса 10 мкс. После электропорации бактериальные суспензии немедленно переносили в низкопрофильный ПЦР-планшет. В две пустые лунки ПЦР-планшета добавляли по 30 мкл питательной среды MRS в качестве контроля стерильности. Во все лунки ПЦР-планшета добавляли по 100 мкл питательной среды MRS, после чего ПЦР-планшет инкубировали в термостате при 37°С в течение 1 часа. После инкубации во все лунки ПЦР-планшета добавляли по 12 мкл раствора эритромицина с концентрацией 40 мг/мл, перемешивали содержимое лунок и оставляли инкубироваться при 37°С в течение 16-18 часов. Далее считывали сигнал флуоресценции по каналу FAM. Количество бактериальных клеток с внедренной контрольной плазмидой оценивали по степени роста флуоресценции по сравнению с контрольными пробами без плазмиды. Результаты анализа приведены на фиг.1.Из полученных результатов следует, что в пробах как Lactobacillus reuteri, так и Lactobacillus plantarum, содержавших плазмиду pLEM415-ldhL-mRFP1, после электропорации и инкубации с питательной средой увеличилась флуоресценция по каналу FAM. Это говорит о том, что в компетентные клетки обоих видов лактобацилл произошло внедрение плазмиды pLEM415-ldhL-mRFP1 и наработка рекомбинантного белка mRFP1, при возбуждении флуоресцирующего по каналу FAM. При этом размах роста флуоресценции для обоих штаммов был одинаков. Таким образом, трансформация компетентных клеток обоих видов лактобацилл прошла с одинаковой эффективностью, т.к. было наработано одинаковое количество рекомбинантного белка mRFP1.
Пример 2.
Анализ эффективности трансформации химических компетентных клеток L. reuteri.
Компетентные клетки бактерий L. reuteri готовили по методикам Palomino et al и Berthier et al [4]. Трансформацию разных компетентных клеток проводили одновременно, для трансформации использовали по 20 мкл компетентных клеток и по 20 нг плазмиды pLEM415-ldhL-mRFP1. Компетентные клетки L. reuteri, приготовленные по каждой методике, анализировали в 4 повторах, к двум из которых добавляли плазмиду pLEM415-ldhL-mRFP1, еще два повтора служили контролями без плазмиды.
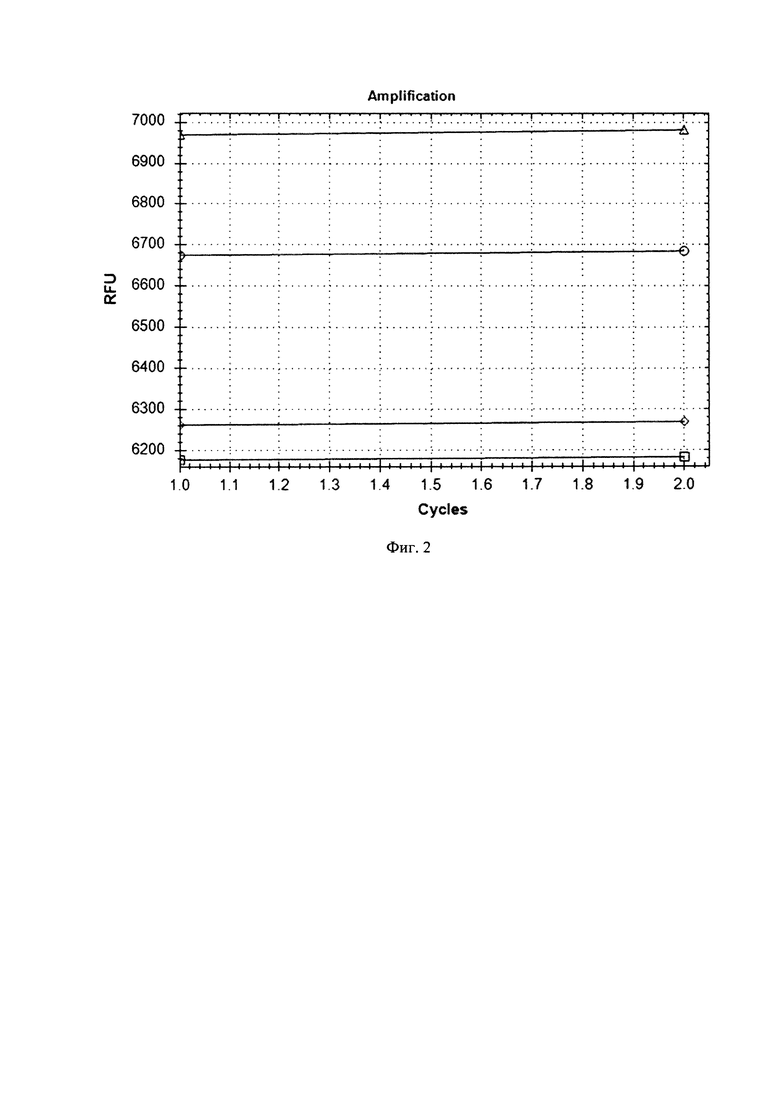
Перемешанные суспензии компетентных клеток и раствора плазмиды инкубировали в 0,2 см кюветах для электропорации во льду в течение 5 минут. По истечении срока инкубации переносили кюветы в электропоратор Gene Pulser (Bio-Rad, США) и проводили электропорацию при следующих условиях: напряжение 2 кВ, сопротивление 400 Ом, емкость 25 мкФ, длительность импульса 10 мкс. После электропорации бактериальные суспензии немедленно переносили в низкопрофильный ПЦР-планшет. В две пустые лунки ПЦР-планшета добавляли по 30 мкл питательной среды MRS в качестве контроля стерильности. Во все лунки ПЦР-планшета добавляли по 100 мкл питательной среды MRS, после чего ПЦР-планшет инкубировали в термостате при 37°С в течение 1 часа. После инкубации во все лунки ПЦР-планшета добавляли по 12 мкл раствора эритромицина с концентрацией 40 мг/мл, перемешивали содержимое лунок и оставляли инкубироваться при 37°С в течение 16-18 часов. Далее считывали сигнал флуоресценции по каналу FAM. Количество бактериальных клеток с внедренной контрольной плазмидой оценивали по степени роста флуоресценции по сравнению с контрольными пробами без плазмиды. Результаты анализа приведены на фиг.2.Из полученных результатов следует, что в пробах компетентных клеток, полученных как по методу Palomino et al, так и Berthier et al, содержавших плазмиду pLEM415-ldhL-mRFP1, после электропорации и инкубации с питательной средой увеличилась флуоресценция по каналу FAM. Это говорит о том, что в компетентные клетки L. reuteri, полученные по обоим методам, произошло внедрение плазмиды pLEM415-ldhL-mRFP1 и наработка рекомбинантного белка mRFP1, при возбуждении флуоресцирующего по каналу FAM. При этом размах роста флуоресценции для компетентных клеток L. reuteri, полученных по методу Palomino et al, был больше, чем для метода Berthier et al. Таким образом, трансформация компетентных клеток по методу Palomino et al прошла с большей эффективностью, т.к. было наработано большее количество рекомбинантного белка mRFP1.
Пример 3.
Анализ эффективности трансформации химических компетентных клеток L. reuteri с добавлением хлорида марганца или хлорида рубидия.
Компетентные клетки бактерий L. reuteri готовили по методике Palomino et al. Трансформацию компетентных клеток проводили одновременно, для трансформации использовали по 20 мкл химических компетентных клеток и по 20 нг плазмиды pLEM415-IdhL-mRFP1. Всего анализировали 5 вариантов условий трансформации: химические компетентные клетки L. reuteri без добавок дополнительных веществ, химические компетентные клетки L. reuteri с добавлением хлорида марганца и химические компетентные клетки L. reuteri с добавлением хлорида рубидия.
Каждый вариант условий трансформации бактерий L. reuteri анализировали в 4 повторах, к двум из которых добавляли плазмиду pLEM415-ldhL-mRFP1, еще два повтора служили контролями без плазмиды. К пробам с хлоридом марганца добавляли по 2 мкл 1 М МnСl2, к пробам с хлоридом рубидия добавляли по 2 мкл 1 М RbCl.
Перемешанные суспензии компетентных клеток и раствора плазмиды инкубировали в 0,2 см кюветах для электропорации во льду в течение 5 минут. По истечении срока инкубации переносили кюветы в электропоратор Gene Pulser (Bio-Rad, США) и проводили электропорацию при следующих условиях: напряжение 2 кВ, сопротивление 400 Ом, емкость 25 мкФ, длительность импульса 10 мкс. После электропорации бактериальные суспензии немедленно переносили в низкопрофильный ПЦР-планшет. В две пустые лунки ПЦР-планшета добавляли по 30 мкл питательной среды MRS в качестве контроля стерильности. Во все лунки ПЦР-планшета добавляли по 100 мкл питательной среды MRS, после чего ПЦР-планшет инкубировали в термостате при 37°С в течение 1 часа. После инкубации во все лунки ПЦР-планшета добавляли по 12 мкл раствора эритромицина с концентрацией 40 мг/мл, перемешивали содержимое лунок и оставляли инкубироваться при 37°С в течение 16-18 часов. Далее считывали сигнал флуоресценции по каналу FAM. Количество бактериальных клеток с внедренной контрольной плазмидой оценивали по степени роста флуоресценции по сравнению с контрольными пробами без плазмиды. Результаты анализа приведены на фиг.3.
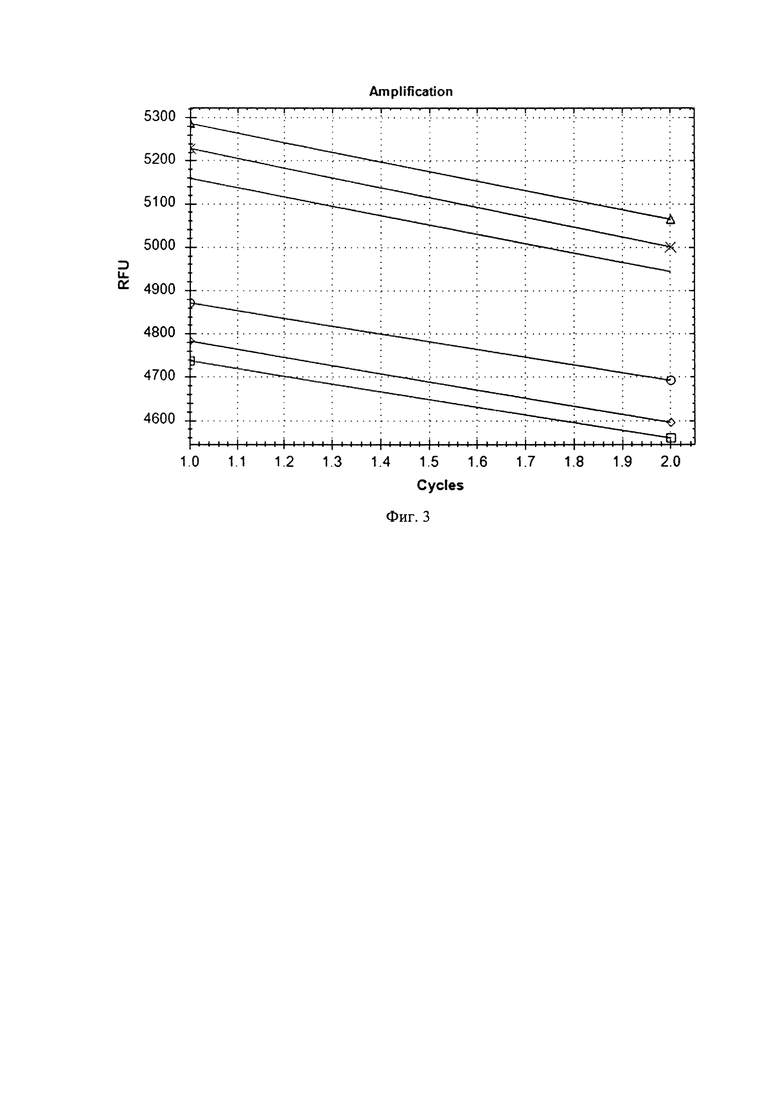
Из полученных результатов следует, что в пробах компетентных клеток с добавлением как MnCl2, так и RbCl, содержавших плазмиду pLEM415-ldhL-mRFP1, после электропорации и инкубации с питательной средой увеличилась флуоресценция по каналу FAM. Это говорит о том, что вне зависимости от добавления MnCl2 и RbCl в компетентные клетки L. reuteri произошло внедрение плазмиды pLEM415-ldhL-mRFP1 и наработка рекомбинантного белка mRFP1, при возбуждении флуоресцирующего по каналу FAM. При этом размах роста флуоресценции был одинаков вне зависимости от наличия или отсутствия MnCl2 и RbCl. Таким образом, трансформация компетентных клеток лактобацилл прошла с одинаковой эффективностью вне зависимости от наличия или отсутствия MnCl2 и RbCl, т.к. было наработано одинаковое количество рекомбинантного белка mRFP1.
Таким образом, предлагаемый способ высокопроизводительной оценки эффективности трансформации компетентных клеток бактерий рода Lactobacillus с помощью плазмиды pLEM415-ldhL-mRFP1 позволяет ускорить анализ эффективности разных протоколов приготовления компетентных клеток бактерий рода Lactobacillus, а также дает возможность одновременно скринировать большое число протоколов приготовления компетентных клеток, химических веществ, повышающих эффективность трансформации, и разных штаммов бактерий рода Lactobacillus.
Источники информации:
1. Garcia-Fruitos Е. Lactic Acid Bacteria: a promising alternative for recombinant protein production. // Microb. Cell Fact. 2012. Vol.11. P. 157.
2. Di Giacomo S. et al. Expanding natural transformation to improve beneficial lactic acid bacteria // FEMS Microbiol. Rev. 2022. Vol.46, №4.
3. Palomino M.M. et al. New method for electroporation of Lactobacillus species grown in high salt // J. Microbiol. Methods. 2010. Vol.83, №2. P. 164-167.
4. Berthier F. et al. Efficient transformation of Lactobacillus sake by electroporation // Microbiology. 1996. Vol.142, №5. P. 1273-1279.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="Способ
высокопроизводительной оценки эффективности трансформации
компетентных клеток бактерий рода Lactobacillus посредством внедрения
плазмидной ДНК с геном флуоресцентного белка.xml" softwareName="WIPO
Sequence" softwareVersion="2.2.0" productionDate="2023-01-23">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>2022133268</ApplicationNumberText>
<FilingDate>2022-12-19</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>1</ApplicantFileReference>
<EarliestPriorityApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>20221133268</ApplicationNumberText>
<FilingDate>2022-12-19</FilingDate>
</EarliestPriorityApplicationIdentification>
<ApplicantName languageCode="ru">Федеральное государственное
автономное образовательное учреждение высшего образования
«Национальный исследовательский Томский государственный
университет»</ApplicantName>
<ApplicantNameLatin> Federalnoe gosudarstvennoe avtonomnoe
obrazovatelnoe uchrezhdenie vysshego obrazovaniia Natsionalnyi
issledovatelskii Tomskii gosudarstvennyi
universitet</ApplicantNameLatin>
<InventionTitle languageCode="ru">Способ высокопроизводительной
оценки эффективности трансформации компетентных клеток бактерий рода
Lactobacillus посредством внедрения плазмидной ДНК с геном
флуоресцентного белка</InventionTitle>
<SequenceTotalQuantity>1</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>7362</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..7362</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q5">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>ggggatgtgctgcaaggcgattaagttgggtaacgccagggttttccca
gtcacgacgttgtaaaacgacggccagtgagcgcgcgtaatacgactcactatagggcgaattgggtacc
gggcccactgagaagttgctctccccagtcgacgtttgttattgctgtttattataccatacccgttttt
ctttctatattagggaataaatagttttctgaaaatttaaacaaaattagcgaactttcatgcaaactat
gctatactggttttcgaaatgaagaattgttttacagaaaggacgtcattatattggcaaacattgaaaa
agatcaccaaaaagttatcctcgtcggcgaaagcttgcatgcctgcaggtcgactctagaggatccccgg
gtaccggtagaaaaaatgactggtggacagcaaatgggtcgggatctgtacgacgatgacgataagccta
ggatggcctcctccgaggacgtcatcaaggagttcatgcgcttcaaggtgcgcatggagggctccgtgaa
cggccacgagttcgagatcgagggcgagggcgagggccgcccctacgagggcacccagaccgccaagctg
aaggtgaccaagggcggccccctgcccttcgcctgggacatcctgtcccctcagttccagtacggctcca
aggcctacgtgaagcaccccgccgacatccccgactacttgaagctgtccttccccgagggcttcaagtg
ggagcgcgtgatgaacttcgaggacggcggcgtggtgaccgtgacccaggactcctccctgcaggacggc
gagttcatctacaaggtgaagctgcgcggcaccaacttcccctccgacggccccgtaatgcagaagaaga
ccatgggctgggaggcctccaccgagcggatgtaccccgaggacggcgccctgaagggcgagatcaagat
gaggctgaagctgaaggacggcggccactacgacgccgaggtcaagaccacctacatggccaagaagccc
gtgcagctgcccggcgcctacaagaccgacatcaagctggacatcacctcccacaacgaggactacacca
tcgtggaacagtacgagcgcgccgagggccgccactccaccggcgcctaagctctacaaataaatcgata
agcttgatatcgaattcctgcagctaattgggtgttcatgacgaccaataatagtatcaggagtccaacc
tgccttgatttgcgcatggatatagttaacttcgatagttggcagttgggtctgcttccgaccacaacga
cgcttatgtcgctgataagtctgaagatattggtcgatggttttaccgtcgttgaggaaacgataaacac
gatagatggtttcttgactacgctgaagtaatttagcagcccgataagctttagtaccttgataccaaaa
atcagctatgagagttaattcacgtgtggtaagatgtttataggtcatttgtgattgcctttcttttgat
taaggatattcaaaagtctatcacaaatggctttttattttttctaacttaattttacaaacgacgaaac
aaaaaaagaggctgattaagttcagtctcttttttaattaattgtgtgattcttcaatttcagccaactt
agatgaagcttgatagatcagtgattctgcgtcactcaatagagataatatatcatcatatgagccatca
tctgcttggctcaaaattcgctttttaaaatcctctaaattatcagccaactgctcgctaatcgtcccat
aaatttttgactgttcaataattgcatcaatgccttttacatctcttaactccataactttcactcccaa
cttacataatcgctaacactacaattccacttatacattaccttttttacaatttcagaatcatcattag
tggcacttacattaatcagatcatcttcattttcatcaaataacaaactatgtctaatatcttttaacaa
tccgccaaaactcaactgcctagatccctgcaatcccttttctagttcgtttaaaataaccaaatcagat
tcttgatcattcgttatataatcagaattttttacttggtacttagcaacttctttagctgaatcaacta
atgccgattgaccatgtttacttttagttcttatcttacgaatatcaataataggatcataatcgagttt
tctagcacgacgccaaagttttatccaatcatcttttgtcaaataattttttgaattaaaataacttgtc
ttcattaacaataataaatgcatatgttgatgaaacgtataattatctcgattgactgtaatttcagaag
atcgtacatagccaatcatatttttagctacttttttatactgaattaagcgtctaactgaactattcat
attttttaaatttatttttaattctccaatttcggaattttcctctgtaagagttaaaaataaataaatc
gcatttggatttttaacatgcgctatatctaaaatttgttgcaactcgtaactattttttaaagatcttc
tccaagaacacaatggacataacctagagtgacagaaccacgtttgatacagtttcattccatccttgcc
tatagcaaattgtaatacttcaccacaatctttaacacgattagctttcttaaattttaaaacctccaaa
tattgcgcataagtaagattagctagtttcttttctcgccatggtcttactttgccactatatttagagg
tatctcttagtctttcacctgtatacacttataaatacctctttactgaaattcatgcagtaaagagagt
tagctatactaattgatataataataattaactactataatagataattgataatattagataactagct
catgcacctaccgatcgccaaatctaaagggactgcatgagttttttatttacgaataaattatacctct
attctattagtaatcaaaatttctacaaacattgatatatcaacgctttaacagcattttaaaaaatgac
gttgtttctataagtatcaagataagtagacacgcgattaaaatcgcgtgtcttagggcgctgtcgcccc
taaaaccccgaccaagcgtcggacgcttgacccccgctagcccttgctagacggcgggcgaaaaataaat
ttttcaccaaataaaagggtcactcactattattcgtcaagggttgccttgcttccctccggtcgcaagc
gtgtgccccttgacgaatggcatctaatagagggctacgccaccagcccacaattagcaaaaaggcaaca
ttcctccgcacggcgggttatgttgcctttttgctttttaaaaattaaacttaaggaatatcaagcttca
tagcgcctaaaatatagggagttgtttcctcccgttaaataatagataactattaaaaatagacaatact
tgctcataagtaatggtacttaaattgtttactttggcgtgtttcattgcttgatgaaactgatttttag
taaacagttgacgatattctcgattgacccattttgaaacaaagtacttatatagcttccaatatttatc
tggaacatctgtggtatggcgggtaagttttattaagacactgtttacttttggtttaggatgaaagcat
tccgctggcagcttaagcaattgctgaatcgagacttgagtgtgcaagagcaaccctagtgttcggtgaa
tatccaaggtacgcttgtagaatccttcttcaacaatcagatagatgtcagacgcacggctttcaaaaac
cacttttttaataatttgtgtgcttaaatggtaaggaatattcccaacaattttatacctctgtttgtta
gggaattgaaactgtagaatatcttggtgaattaaagtgacacgagtattcagttttaatttttctgacg
ataagttgaatagatgactgtctaattcaatagacgttacctgtttacttattttagccagtttcgtcgt
taaatgccctttacctgttccaatttcgtaaacggtatcggtttcttttaaattcaattgttttattatt
tggttgagtactttttcactcgttaaaaagttttgagaatattttatatttttgttcatgtaatctctcc
tgaagtgattacatctgtaaataaatacagaagttaaacgatttgtttgtaattttagttatctgtttaa
aaagtcataagattagtcactggtaggaattaatctaacgtatttatttatctgcgtaatcactgttttt
agtctgtttcaaaacagtagatgttttatctacattacgcatttggaataccaacatgacgaatccctcc
ttcttaattacaaatttttagcatctaatttaacttcaattcctattatacaaaatttttaaacgaattt
tttaaaaatgtattgagcttctgccaattaattttaggaacagcaatgtgactaatccctatcccatctc
gatatatcagtccttgaccaacattgttgttaagccaatctgcagcccgggggatccactagttctagag
cggccgccaccgcggtggagctccagcttttgttccctttagtgagggttaattgcgcgcttggcgtaat
catggtcatagctgtttcctgtgtgaaattgttatccgctcacaattccacacaacatacgagccggaag
cataaagtgtaaagcctggggtgcctaatgagtgagctaactcacattaattgcgttgcgctcactgccc
gctttccagtcgggaaacctgtcgtgccagctgcattaatgaatcggccaacgcgcggggagaggcggtt
tgcgtattgggcgctcttccgcttcctcgctcactgactcgctgcgctcggtcgttcggctgcggcgagc
ggtatcagctcactcaaaggcggtaatacggttatccacagaatcaggggataacgcaggaaagaacatg
tgagcaaaaggccagcaaaaggccaggaaccgtaaaaaggccgcgttgctggcgtttttccataggctcc
gcccccctgacgagcatcacaaaaatcgacgctcaagtcagaggtggcgaaacccgacaggactataaag
ataccaggcgtttccccctggaagctccctcgtgcgctctcctgttccgaccctgccgcttaccggatac
ctgtccgcctttctcccttcgggaagcgtggcgctttctcatagctcacgctgtaggtatctcagttcgg
tgtaggtcgttcgctccaagctgggctgtgtgcacgaaccccccgttcagcccgaccgctgcgccttatc
cggtaactatcgtcttgagtccaacccggtaagacacgacttatcgccactggcagcagccactggtaac
aggattagcagagcgaggtatgtaggcggtgctacagagttcttgaagtggtggcctaactacggctaca
ctagaagaacagtatttggtatctgcgctctgctgaagccagttaccttcggaaaaagagttggtagctc
ttgatccggcaaacaaaccaccgctggtagcggtggtttttttgtttgcaagcagcagattacgcgcaga
aaaaaaggatctcaagaagatcctttgatcttttctacggggtctgacgctcagtggaacgaaaactcac
gttaagggattttggtcatgagattatcaaaaaggatcttcacctagatccttttaaattaaaaatgaag
ttttaaatcaatctaaagtatatatgagtaaacttggtctgacagttaccaatgcttaatcagtgaggca
cctatctcagcgatctgtctatttcgttcatccatagttgcctgactccccgtcgtgtagataactacga
tacgggagggcttaccatctggccccagtgctgcaatgataccgcgagacccacgctcaccggctccaga
tttatcagcaataaaccagccagccggaagggccgagcgcagaagtggtcctgcaactttatccgcctcc
atccagtctattaattgttgccgggaagctagagtaagtagttcgccagttaatagtttgcgcaacgttg
ttgccattgctacaggcatcgtggtgtcacgctcgtcgtttggtatggcttcattcagctccggttccca
acgatcaaggcgagttacatgatcccccatgttgtgcaaaaaagcggttagctccttcggtcctccgatc
gttgtcagaagtaagttggccgcagtgttatcactcatggttatggcagcactgcataattctcttactg
tcatgccatccgtaagatgcttttctgtgactggtgagtactcaaccaagtcattctgagaatagtgtat
gcggcgaccgagttgctcttgcccggcgtcaatacgggataataccgcgccacatagcagaactttaaaa
gtgctcatcattggaaaacgttcttcggggcgaaaactctcaaggatcttaccgctgttgagatccagtt
cgatgtaacccactcgtgcacccaactgatcttcagcatcttttactttcaccagcgtttctgggtgagc
aaaaacaggaaggcaaaatgccgcaaaaaagggaataagggcgacacggaaatgttgaatactcatactc
ttcctttttcaatattattgaagcatttatcagggttattgtctcatgagcggatacatatttgaatgta
tttagaaaaataaacaaataggggttccgcgcacatttccccgaaaagtgccacctaaattgtaagcgtt
aatattttgttaaaattcgcgttaaatttttgttaaatcagctcattttttaaccaataggccgaaatcg
gcaaaatcccttataaatcaaaagaatagaccgagatagggttgagtgttgttccagtttggaacaagag
tccactattaaagaacgtggactccaacgtcaaagggcgaaaaaccgtctatcagggcgatggcccacta
cgtgaaccatcaccctaatcaagttttttggggtcgaggtgccgtaaagcactaaatcggaaccctaaag
ggagcccccgatttagagcttgacggggaaagccggcgaacgtggcgagaaaggaagggaagaaagcgaa
aggagcgggcgctagggcgctggcaagtgtagcggtcacgctgcgcgtaaccaccacacccgccgcgctt
aatgcgccgctacagggcgcgtcccattcgccattcaggctgcgcaactgttgggaagggcgatcggtgc
gggcctcttcgctattacgccagctggcgaaag</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БАКТЕРИАЛЬНЫЕ ШТАММЫ, ВЫДЕЛЕННЫЕ ИЗ СВИНЕЙ | 2012 |
|
RU2677890C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, СОДЕРЖАЩАЯ ГЕНЫ СИНТЕЗА БУТАНОЛА ИЗ Clostridium acetobutylicum (ВАРИАНТЫ), РЕКОМБИНАНТНЫЙ ШТАММ Lactobacillus brevis - ПРОДУЦЕНТ Н-БУТАНОЛА (ВАРИАНТЫ) И СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА Н-БУТАНОЛА | 2008 |
|
RU2375451C1 |
| Способ выявления из естественных сред перспективных пробиотических штаммов | 2021 |
|
RU2772351C1 |
| ШТАММ Lactobacillus rhamnosus, ОБЛАДАЮЩИЙ АНТИБАКТЕРИАЛЬНОЙ И АНТИГРИБКОВОЙ АКТИВНОСТЬЮ В ОТНОШЕНИИ ПАТОГЕННОЙ И УСЛОВНО-ПАТОГЕННОЙ ВАГИНАЛЬНОЙ МИКРОФЛОРЫ | 2013 |
|
RU2570622C2 |
| ШТАММ Lactobacillus rhamnosus, ОБЛАДАЮЩИЙ ПРОТИВОМИКРОБНЫМИ, ИММУНОМОДУЛИРУЮЩИМИ И СПЕЦИФИЧНЫМИ В ОТНОШЕНИИ МАННОЗЫ АДГЕЗИОННЫМИ СВОЙСТВАМИ, А ТАКЖЕ ПРОДУКТ НА ЕГО ОСНОВЕ | 2009 |
|
RU2549699C2 |
| Рекомбинантный вектор, содержащий иммуногенный белок вируса африканской чумы свиней, рекомбинантные бактерии и их применение | 2020 |
|
RU2771177C1 |
| ШТАММ LACTOBACILLUS PARACASEI CNCM I-2116 (NCC 2461), ОБЛАДАЮЩИЙ СПОСОБНОСТЬЮ ПРЕДОТВРАЩАТЬ КОЛОНИЗАЦИЮ КИШЕЧНИКА ПАТОГЕННЫМИ БАКТЕРИЯМИ, ВЫЗЫВАЮЩИМИ ДИАРЕЮ, И ПРЕДОТВРАЩАТЬ ЗАРАЖЕНИЕ ЭПИТЕЛИАЛЬНЫХ КЛЕТОК КИШЕЧНИКА РОТАВИРУСАМИ, ПИЩЕВОЙ ПРОДУКТ И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ПРОФИЛАКТИКИ И/ИЛИ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, СВЯЗАННЫХ С ДИАРЕЕЙ | 2000 |
|
RU2247569C2 |
| НОВЫЕ ПЕПТИДЫ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2010 |
|
RU2583579C2 |
| ПРОБИОТИЧЕСКИЕ ШТАММЫ, ОБЛАДАЮЩИЕ СПОСОБНОСТЬЮ ПОГЛОЩАТЬ ХОЛЕСТЕРИН, СПОСОБЫ И ИХ ПРИМЕНЕНИЕ | 2015 |
|
RU2722038C2 |
| ЛИПОТЕЙХОЕВАЯ КИСЛОТА ИЗ МОЛОЧНО-КИСЛЫХ БАКТЕРИЙ И ЕЕ ПРИМЕНЕНИЕ ДЛЯ МОДУЛЯЦИИ ИММУННЫХ РЕАКЦИЙ, ОПОСРЕДОВАННЫХ ГРАМОТРИЦАТЕЛЬНЫМИ БАКТЕРИЯМИ, ПОТЕНЦИАЛЬНЫМИ ПАТОГЕННЫМИ ГРАМПОЛОЖИТЕЛЬНЫМИ БАКТЕРИЯМИ | 2002 |
|
RU2320356C2 |
Изобретение относится к биотехнологии. Представлен способ трансформации компетентных клеток бактерий рода Lactobacillus плазмидой pLEM415-ldhL-mRFP1, имеющей нуклеотидную последовательность SEQ ID NO: 1, содержащей ген устойчивости к эритромицину erm, конститутивный промотор гена idhL и кодирующую последовательность мономерного красного флуоресцентного белка mRFP. При этом после добавления питательной среды MRS и эритромицина инкубацию бактериальных культур проводят в течение 16-18 часов, а детекцию результатов трансформации осуществляют путем анализа роста флуоресценции по каналу FAM. Способ позволяет ускорить оценку эффективности разных протоколов приготовления компетентных клеток бактерий рода Lactobacillus, а также дает возможность одновременно скринировать большое число протоколов приготовления компетентных клеток, химических веществ, повышающих эффективность трансформации, и разных штаммов бактерий рода Lactobacillus. 3 з.п. ф-лы, 3 ил., 3 пр.
1. Способ оценки эффективности трансформации компетентных клеток бактерий рода Lactobacillus посредством внедрения плазмидной ДНК с геном флуоресцентного белка, включающий трансформацию компетентных клеток бактерий рода Lactobacillus плазмидной ДНК, добавление среды MRS и раствора эритромицина с концентрацией 40 мг/мл, инкубацию бактериальных культур с последующей детекцией результатов трансформации, отличающийся тем, что для трансформации используют плазмиду pLEM415-ldhL-mRFP1, имеющую нуклеотидную последовательность SEQ ID NO: 1, содержащую ген устойчивости к эритромицину erm, конститутивный промотор гена idhL и кодирующую последовательность мономерного красного флуоресцентного белка mRFP, после добавления питательной среды MRS и эритромицина инкубацию бактериальных культур проводят в течение 16-18 часов, а детекцию результатов трансформации осуществляют путем анализа роста флуоресценции по каналу FAM.
2. Способ по п.1, отличающийся тем, что трансформация проводится в низкопрофильном ПЦР-планшете.
3. Способ по п.1, отличающийся тем, что компетентные клетки могут быть приготовлены из разных штаммов бактерий рода Lactobacillus.
4. Способ по п.1, отличающийся тем, что компетентные клетки бактерий рода Lactobacillus готовят путем выращивания лактобацилл в питательной среде MRS с 0,9 М или 0,7 М NaCl.
| PALOMINO, M.M | |||
| et al | |||
| New method for electroporation of Lactobacillus species grown in high salt | |||
| JOURNAL OF MICROBIOLOGICAL METHODS | |||
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
