Изобретение относится к области молекулярной биологии, может быть использовано для детекции носителей мутации у крыс линии DAT, у которых искусственно, путем редактирования генома с использованием нуклеаз с «цинковыми пальцами» выключен ген, кодирующий транспортер дофамина из семейства натрий- хлорид- зависимых нейротрансмиттеров-DAT.
Транспортер дофамина является одним из основных белков, вовлеченных в метаболизм дофамина в ЦНС, с помощью которого осуществляется обратный захват дофамина из синаптической щели.
Модельное животное (МЖ)- это нечеловеческий вид, используемый в биомедицинских исследованиях, поскольку он может имитировать аспекты биологического процесса/ болезни, обнаруженные у людей. Модельные животные (мыши, крысы …) довольно схожи с людьми по своей физиологии, анатомии, или реакции на патоген, в связи с этим ученые могут экстраполировать результаты исследований, полученных на МЖ, чтобы лучше понять человеческую физиологию и этиологию болезни. Исследователи могут проводить различные эксперименты на модельных животных, которые были бы непрактичны или запрещены с точки зрения этики над людьми.
Крысы являются одними из наиболее распространенных объектов для проведения исследовательских, испытательных экспериментов во всех лабораториях мира. Крысы относятся к роду Rattus, семейству мышевидных (Muridae). Для экспериментальных исследований в лабораториях используют белых крыс, являющихся альбиносами черной и серой пород. Важное преимущество лабораторных крыс заключается в том, что они довольно устойчивы к инфекционным заболеваниям, обладают большой плодовитостью, а их небольшой вес, относительно простое содержание, разведение в лабораторных условиях позволяют проводить с ними массовые опыты.
Крысы необходимы для установления токсичности лекарственных веществ, ядов, находят широкое применение при изучении вопросов питания, проведения биологической стандартизации гормональных препаратов, для постановки научных исследований по биохимии, эндокринологии, витаминологии. Кроме того, лабораторных крыс используют для воспроизведения на них экспериментальных опухолей (саркомы Кричевского и Синельникова) и инфекционных заболеваний (бешенство, амебиаз, грипп свиней ...). Инбредные и гибридные линии наряду с трансгенными и нокаутными линиями широко используются в медикобиологических исследованиях, в том числе связанных с экспериментальными и доклиническими исследованиями лекарств.
Широкое применение нашли крысы в качестве модельных животных при исследовании заболеваний мозга, в том числе нейродегенеративных заболеваний, психических расстройств, алкогольной зависимости.
В этих исследованиях большое внимание уделяется изучению системы нейротрансмиттеров, в том числе дофаминэргических, которые играют важную роль в функционировании ЦНС. Нарушение работы дофаминовых нейронов лежат в основе патогенеза многих нейропсихических расстройств (болезнь Паркинсона, шизофрения, обсессивно-компульсионное расстройство, маниакально-депрессивный психоз), а также формирования зависимости от психоактивных веществ.
Транспортер домафина (DAT) является одним из основных белков, вовлеченных в метаболизм дофамина в ЦНС, с помощью которого осуществляется обратный захват дофамина из синаптической щели.
Линии мышей без функционального DAT (DAT-KO) и со сниженным до 90 % уровнем экспрессии DAT, стали самыми известными, а также одними из наиболее используемых генетически модифицированных животных в нейропсихофармакологии.
Исследования DAT-KO мышей позволили получить информацию о механизмах действия различных психотропных препаратов, взаимодействии важнейших нейромедиаторных систем ЦНС, базовых принципах дофаминовой нейропередачи, патофизиологических механизмах расстройств, связанных с нарушением дофаминовой нейропередачи.
Дальнейшие исследования требовали разработку новых объектов и расширения спектра видов экспериментальных животных из-за ограниченности изучения ряда поведенческих реакций на мышах.
Крысы DAT-KO были созданы на основе аутбредных крыс Wistar Han посредством редактирования генома с помощью нуклеаз с «цинковыми пальцами»: у данной линии генетически модифицированных крыс отсутствует функциональный DAT благодаря делеции 5 нуклеотидов, которая приводит к раннему формированию стоп-кодона [Анализ делеции (5 bp) в гене Slc6a3 в линии крыс DAT / А.М. Тазетдинов А.М., З.Р. Тахирова, П.А. Ахмадиев, З.Р. Хисматуллина, Э.К. Хуснутдинова // Biomics. - 2022.- Т.14(4). - С. 353-358., Pronounced Hyperactivity, Cognitive Dysfunctions, and BDNF Dysregulation in Dopamine Transporter Knock-out Rats. / D. Leo, I Sukhanov., F. Zoratto et al. // The Journal of Neuroscience. - 2018. - V. 38(8). - P. 1959-1972.].
Авторы отмечают, что выключение DAT приводит к тому, что у крыс в возрасте 1 месяц отмечается выраженная гиперактивность, которая сохраняется как минимум до 4 месяцев. Известно, что психостимуляторы (амфетамин, метилфенидат) используют в клинической практике для терапии пациентов, страдающих синдромом дефицита внимания с гиперактивностью (СДВГ).
Результаты проведенных экспериментов показали, что крысы DAT-KO позволяют моделировать как гипер-, так и гиподофаминэргические состояния в доклинических исследованиях. Данные модели обладают как внешней, так и предиктивной валидностью и могут быть использованы для разработки новых фармакологических подходов к лечению СДВГ и болезни Паркинсона.
Известен способ детекции искомой делеции с помощью анализа ПДРФ (полиморфизм длин рестрикционных фрагментов) заключающийся в том, что после продуцирования делеции в 5 нуклеотидов во втором экзоне гена формируется ранний стоп-кодон и исчезает сайт узнавания для эндонуклеазы рестрикции (рестриктазы) BtsI MutI.
Таким образом, после выделения ДНК и постановки полимеразной цепной реакции (ПЦР) синтезируется продукт длиной 175 и/или 170 н.п. в зависимости от генотипа. После обработки ПЦР продукта ферментом рестриктазой возможны три варианта: 1) нормальный тип (без делеции) фрагменты длиной 71 и 104 п.н. (вследствие наличия сайта узнавания); 2) гетерозигота- 71, 104, 170 п.н.; 3) гомозигота по делеции- 170 п.н.
К недостаткам ранее описанного способа детекции делеции, несмотря на относительную простоту дизайна эксперимента, можно отнести:
1) необходимость проверки наличия ПЦР продукта путем эклектрофоретического разделения в полиакриламидном геле перед проведением рестрикции, т.е. проведение дополнительных процедур в виде электрофоретического разделения продуктов ПЦР;
2) не очень высокую специфичность сайта узнавания для рестриктазы BtsI Mut I*;
3) «капризность» ПДРФ анализа, поскольку со временем, при наличии определенных веществ активность фермента снижается, что обуславливает его использование сверх нормы, ведущее к увеличению стоимости проведенного анализа. Игнорирование факта уменьшения активности фермента может привести к неправильной интерпретации полученных данных.
* BtsI Mut I фермент, катализирующий гидролиз двухцепочечной ДНК рядом со специфическим сайтом узнавания (рестриктаза Типа IIS). Сайт узнавания- 5’- …CAGTGNN …3’ 3’-…GTCAC NN…5’.
Известен способ выявления в ДНК опухоли мутаций в гене BRAF, ассоциированных с чувствительностью опухоли к таргетной терапии, с использованием асимметричной ПЦР в реальном времени с двумя флуоресцентными зондами TaqMan, применяемыми в качестве агентов, блокирующих синтез аллелей дикого типа, и пост-амплификационным плавлением дуплексов зонд-однонитевая ДНК. Анализ результатов плавления осуществляют с помощью стандартного модуля плавления. В специально подобранных условиях ПЦР зонды TaqMan подавляют амплификацию аллеля дикого типа сильнее, чем мутантного аллеля из-за неспаренного в последнем случае основания, существенно ослабляющего структуру дуплекса. За счет преимущественной амплификации мутантного аллеля существенно возрастает чувствительность способа [патент № 2631824 МПК C12Q 1/68, дата опубликования 26.09.2017].
Известен способ детекции мутаций H63D и С282Y в гене HFE с помощью ПЦР в реальном времени и флуоресцентной детекцией, основанной на использовании аллель-специфичных олигонуклеотидных зондов с двойной флуоресцентной меткой и 5’ экзонуклеазной активности ДНК-полимеразы. В процессе репликации анализируемой молекулы ДНК (матрицы) ДНК-полимераза с 5’ экзонуклеазной активностью расщепляет связанный с матрицей зонд, вследствие чего происходит удаление друг от друга молекул красителя и тушителя, и тушение прекращается. В результате в каждом цикле ПЦР флуоресценция повышается пропорционально расщеплению зонда, или, что то же самое, пропорционально накоплению специфического продукта [патент № 2304170, МПК C12Q 1/68, дата опубликования 10.08.2007].
Известен способ одновременной идентификации трех основных мутаций (S65C, H63D, C282Y) в гене HFE, связанных с развитием наследственного гемохроматоза посредством проведения проведения аллель-специфической ПЦР в присутствии красителя Sybr Green 1, в которой мутантный и нормальный аллели получают в различных реакционных сосудах, и определения конечного продукта путем анализа кривых температурной диссоциации ампликонов (патент № 2396354, МПК C12Q 1/68, дата опубликования 10.08.2010).
Задачей и техническим результатом изобретения является способ детекции делеции у крыс линии DAT посредством проведения полимеразной цепной реакции в присутствии красителя Eva Green с последующим анализом кривых температурной диссоциации полученных амплификатов и/или проведением вертикального электрофоретического разделения продуктов амплификации в неденатурирующих условиях.
Технический результат достигается постановкой ПЦР с интеркалирующим красителем, с последующим анализом кривой плавлением в приборе с оптической детекцией (Rotor Gene Q (Qiagen, CFX 96 (Biorad)) и/или разделением продуктов амплификации в концентрированном полиакриламидном геле в неденатурирующих условиях.
Суть процедуры детекции делеции сводится к следующему. В амплификаторе/ приборе с оптической детекцией проводят ПЦР для проб, полученных из образцов биологического материала (кровь, фрагмент хвоста), с подобранными праймерами, охватывающими участок, где локализована мутация. Каждая реакция проходит в отдельной пробирке или отдельной ячейке 96-луночной ПЦР панели, где присутствуют все необходимые компоненты для прохождения ПЦР на основе индивидуальной матрицы, внесенной в соответствующие пробирки или лунки.
Исследуемый образец ДНК, полученный с использованием любого известного способа выделения (фенольно-хлороформной экстракции, выделения и очистки на магнитных частицах с применением любого коммерческого набора для выделения ДНК) добавляется по 60 нг матрицы в каждую реакционную смесь.
Одновременно в том же приборе и в тех же условиях проводятся реакции с контрольными ДНК (в отдельных пробирках), в том числе с отрицательным негативным контролем, в которых наличие/отсутствие мутаций заранее подтверждено независимым методом и подтверждено секвенированием по Сэнгеру.
Тест выполняется с использованием олигонуклеотидов, имеющих последовательности (5-3):
F- TCAAGGAGCAGAACGGAGTG; R- ACAGCAAAGCCGATGACTGA.
Размер амплифицируемого фрагмента в зависимости от генотипа (без делеции/с делецией) 133 п.о. и 128 п.о. соответственно.
Реакционная смесь объемом 20 мкл (на 1 образец) состоит из:
10-кратный буфер - 2,0 мкл.
10-кратный раствор Mg2+ - 2,0 мкл.
дАТФ, дЦТФ, дГТФ, дТТФ - каждого по 5 мМ- 0,28 мкл.
специфические праймеры- каждого по 15,0 pMol- 3 мкл каждого (F и R).
Taq ДНК полимераза - 5 ед - 0,12 мкл.
50-кратный флуоресцентный интеркалирующий краситель Eva Green- 0,4 мкл.
Н2O -3,2 мкл.
ДНК - 6 мкл.
Условия амплификации:
1) 95 °C- 5 мин.
2) 94°C- 35 сек.
68°С- 45 сек 32 цикла.
72°С- 45 сек.
3) 72°С- 10 мин.
4) 4°С - хранение.
Реакцию можно проводить на любом амплификаторе с размером ячейки 0,2 мл (200 мкл).
В случае использования обычного амплификатора на этапе ПЦР на следующем этапе необходимо использовать прибор с оптической детекцией с возможностью проведения плавления/ HRM анализа (например, Quant Studio 5 (Thermofisher Scientific), CFX 96 (BioRad), Rotor Gene Q (Qiagen).
Регистрация результатов производится по кривой плавления в амплификаторе с оптическим модулем, позволяющим регистрировать флуоресценцию с длиной волны 520 нм. Устанавливается шаг нагрева модуля от 72°С до 92°С - 0,1°С/2 сек.
Точки диссоциации ампликонов находятся в области 83-88°С.
Поскольку регистрация и анализ кривой плавления проводились на оборудовании Rotor Gene Q (Qiagen), то при использовании другого оборудования (Quant Studio 5 (Thermofisher Scientific), CFX 96 (BioRad), возможны небольшие различия в параметрах оптимального шага нагрева модуля и точки диссоциации ампликонов, что не является критическим- подобрать оптимальные условия для конкретного прибора эмпирически не представляется сложным.
Результат эксперимента учитывается по наличию пика, соответствующего точке диссоциации контрольной ДНК, наличие/отсутствие мутаций в которой заранее подтверждено независимым способом и подтверждено секвенированием по Сэнгеру.
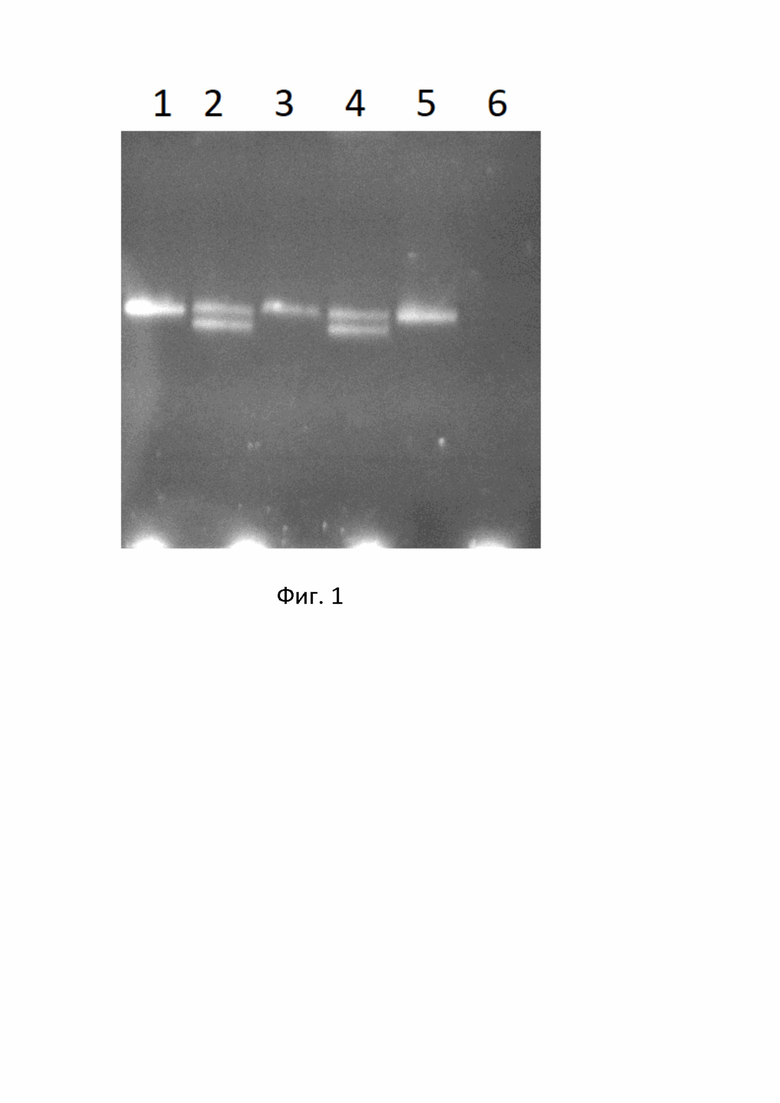
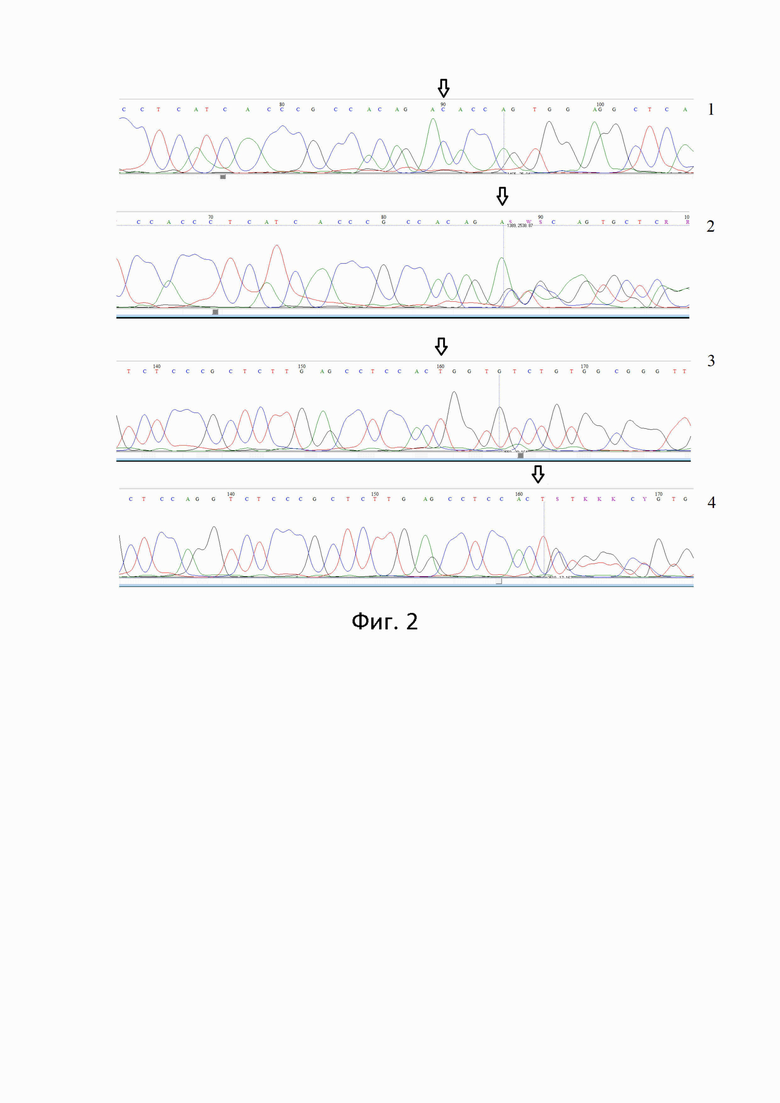
Наличие/отсутствие делеции подтверждено результатами секвенирования по Сэнгеру для гетерозигот и гомозигот (без делеции) с использованием прямого и обратного праймеров (см. электрофореграмма (фиг.1), где образец 1- гомозигота (без делеции) с прямого праймера, образец 2- гетерозигота с прямого праймера, образец 3- гомозигота (без делеции) с обратного праймера, образец 4- гетерозигота с обратного праймера, стрелкой указано местоположение делеции.
Несмотря на возможность вычисления температуры плавления для ампликонов, рекомендуется сопоставление результатов тестов испытуемой ДНК и контрольной, поскольку эмпирические данные редко в точности полностью совпадают с расчетными показателями, кроме того, они могут варьировать в результате изменения условий реакций, которые трудно абсолютно идентично выдержать от теста к тесту.
Кроме того, продукты, полученные после ПЦР, регистрации и анализа кривой плавления, можно дополнительно подвергнуть электрофоретическому разделению в концентрированном полиакриламидном геле (10-12 %) в течение 5-6 часов при напряжении 300-350 В на любом источнике тока и камере (например, на камерах для вертикального электрофореза от ООО «Хеликон», источник тока «Эльф-8» («ДНК-технология»).
Далее проводится стандартная окраска бромистым этидием с проявлением и анализом в гельдокументирующей системе.
Наличие одного/двух полос после электрофоретического разделения продуктов ПЦР свидетельствует соответственно о гомозиготных/гетерозиготных образцах.
При проведении исследований по детекции делеции посредством проведения электрофореза рекомендуется использование контрольных образцов с известным генотипом.
Пример конкретной реализации способа.
1) Формирование выборки из 75 особей линии DAT, c последующим выделением ДНК из крови фенольно/хлороформной экстракцией, с использованием магнитных частиц и набора реагентов «Prepfiler BTA Express» (США) (59 особей) и из фрагментов хвостов с использованием магнитных частиц и набора реагентов «Prepfiler BTA Express» (16 особей).
Определение концентрации проб ДНК с использованием спектрофотометра Nanodrop 2000 (США) и приготовление рабочих растворов ДНК с концентрацией 10 нг/мкл.
Приготовление реакционной смеси из следующих компонентов (см. таблицу 1) с последующим добавлением ДНК матрицы, загрузка в амплификатор Rotor Gene Q (Qiagen), запуск полимеразной цепной реакции (амплификации).
Условия проведения амплификации:

При анализе кривой плавления устанавливается шаг нагрева модуля от 72°С до 92°С - 0,1°С/2 сек.
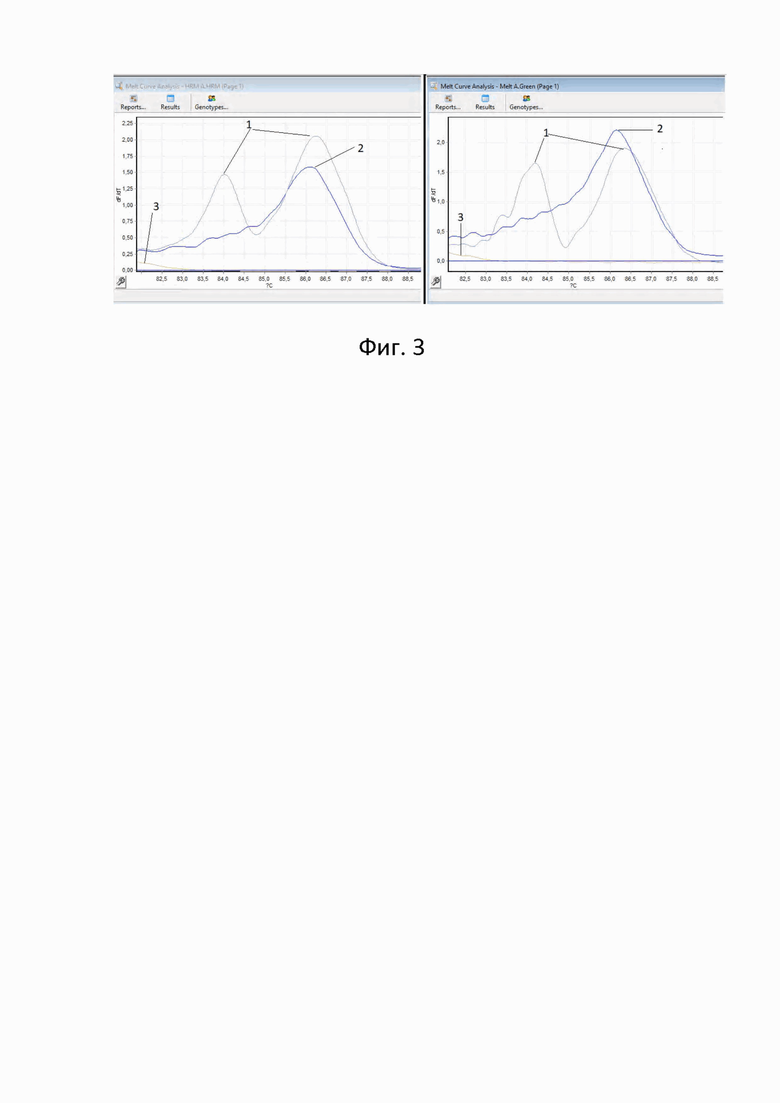
На диаграмме (фиг. 2) представлены результаты проведения анализа кривых температурной диссоциации полученных амплификатов, где образец 1- гетерозигота (с делецией и нормальным аллелем), образец 2- гомозигота (без делеции), образец 3- негативный контроль.
Кроме того, продукты, полученные после ПЦР, регистрации и анализа кривой плавления, можно дополнительно подвергнуть электрофоретическому разделению в 11 % полиакриламидном геле в течение 6 часов при напряжении 320 В в камере для вертикального электрофореза, с использованием источника тока «Эльф-8».
Далее проводится стандартная окраска бромистым этидием с проявлением и анализом в гельдокументирующей системе.
На электрофореграмма (фиг.3) представлены результаты проведения вертикального электрофоретического разделения продуктов амплификации в неденатурирующих условиях, где Дорожки 1, 3, 5- гомозиготы (без делеции), Дорожки 2, 4- гетерозиготы (с делецией и нормальным аллелем), Дорожка 6- негативный контроль.
Таким образом, заявляемое изобретение позволяет эффективно, быстро, надежно выявлять носителей делеции у крыс линии DAT.
| название | год | авторы | номер документа |
|---|---|---|---|
| МОЛЕКУЛЯРНО-ГЕНЕТИЧЕСКИЙ СПОСОБ ОПРЕДЕЛЕНИЯ ЧУВСТВИТЕЛЬНОСТИ ОПУХОЛИ У ПАЦИЕНТОВ С РАКОМ ЛЕГКОГО К ТЕРАПИИ ГЕФИТИНИБОМ | 2010 |
|
RU2454464C2 |
| СПОСОБ ИДЕНТИФИКАЦИИ МУТАЦИЙ S65C, H63D, C282Y И ПОЛИМОРФИЗМА 2 EX + 4T>C В ГЕНЕ HFE | 2008 |
|
RU2396354C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ РИСКА РАЗВИТИЯ АРТЕРИАЛЬНОЙ ГИПЕРТЕНЗИИ | 2015 |
|
RU2600442C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ В ГЕНЕ ГАЛАКТОЗО-1-ФОСФАТУРИДИЛТРАНСФЕРАЗЫ ЧЕЛОВЕКА МУТАЦИИ Q188R, RS75391579 | 2017 |
|
RU2675324C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ЧУВСТВИТЕЛЬНОСТИ ОПУХОЛИ ЛЕГКОГО К ТЕРАПИИ ИНГИБИТОРАМИ ТИРОЗИНКИНАЗ | 2012 |
|
RU2499994C1 |
| СПОСОБ ОДНОВРЕМЕННОЙ ГЕНОДИАГНОСТИКИ ЧЕТЫРЕХ МУТАНТНЫХ АЛЛЕЛЕЙ КАППА-КАЗЕИНА У КРУПНОГО РОГАТОГО СКОТА И ТЕСТ-СИСТЕМА ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2017 |
|
RU2691995C2 |
| НАБОР ПОСЛЕДОВАТЕЛЬНОСТИ ПРАЙМЕРОВ И АЛЛЕЛЬСПЕЦИФИЧЕСКИХ ЗОНДОВ ДЛЯ ОДНОВРЕМЕННОЙ ГЕНОДИАГНОСТИКИ ЧЕТЫРЕХ МУТАНТНЫХ АЛЛЕЛЕЙ КАППА-КАЗЕИНА У КРУПНОГО РОГАТОГО СКОТА | 2017 |
|
RU2646140C1 |
| Способ диагностики мутации 35delG (rs80338939) гена GJB2 | 2020 |
|
RU2739889C1 |
| Способ диагностики мутации 167delT (rs80338942) гена GJB2 | 2020 |
|
RU2739943C1 |
| Способ выявления мутации p.L265P в гене MYD88 | 2020 |
|
RU2756909C1 |
Изобретение относится к области молекулярной биологии, может быть использовано для детекции носителей мутации в линии крыс DAT (нокаут), у которых искусственно выключен ген, кодирующий транспортер дофамина из семейства натрий- хлорид- зависимых нейротрансмиттеров. Предложен простой в исполнении способ идентификации мутации в виде делеции в гене Slc6a3 размером 5 bp у крыс линии DAT посредством проведения ПЦР с использованием праймеров F - TCAAGGAGCAGAACGGAGTG и R - ACAGCAAAGCCGATGACTGA в присутствии красителя Eva Green, с последующим анализом кривых температурной диссоциации полученных амплификатов и/или проведением вертикального электрофоретического разделения продуктов амплификации в неденатурирующих условиях. Изобретение предлагает эффективный, быстрый и надежный способ анализа делеции у крыс линии DAT на предмет её наличия/отсутствия. 3 ил., 1 табл., 1 пр.
Способ детекции делеции в гене Slc6a3 размером 5 bp у крыс линии DAT посредством проведения полимеразной цепной реакции с использованием праймеров F- TCAAGGAGCAGAACGGAGTG и R - ACAGCAAAGCCGATGACTGA в присутствии красителя Eva Green с последующим анализом кривых температурной диссоциации полученных амплификатов и/или проведением вертикального электрофоретического разделения продуктов амплификации в неденатурирующих условиях.
| ADINOLFI A., Behavioral characterization of DAT-KO rats and evidence of asocial-like phenotypes in DAT-HET rats: The potential involvement of norepinephrine system, Behav Brain Res, 2019, vol | |||
| Способ получения гидроцеллюлозы | 1920 |
|
SU359A1 |
| Редукционный или предохранительный клапан с диафрагмой, нагруженной пружиной или грузом | 1925 |
|
SU516A1 |
| REITH M.E.A | |||
| et al., The dopamine transporter gene SLC6A3: multidisease risks, Mol Psychiatry, 2022, vol | |||
| Прибор с двумя призмами | 1917 |
|
SU27A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| ЭЛЕКТРОД ДЛЯ ЭЛЕКТРИЧЕСКОЙ ДУГОВОЙ СВАРКИ И СПОСОБ ЕГО ИЗГОТОВЛЕНИЯ | 1923 |
|
SU1031A1 |
