ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее раскрытие относится к варианту Corynebacterium glutamicum с улучшенной способностью к продуцированию L-лизина и к способу продуцирования L-лизина с его применением.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
L-лизин представляет собой незаменимую аминокислоту, которая не синтезируется в человеческом или животном организме и должна поставляться извне, и он обычно продуцируется посредством ферментации с использованием микроорганизмов, таких как бактерии или дрожжи. При продукции L-лизина можно использовать полученные в природе штаммы дикого типа или варианты, модифицированные для наличия у них повышенной способности к продуцированию L-лизина. Недавно для того, чтобы улучшить эффективность продукции L-лизина разработали разные рекомбинантные штаммы или варианты с превосходной способностью к продуцированию L-лизина и способы продуцирования L-лизина с их использованием посредством применения технологии генной инженерии к таким микроорганизмам, как Escherichia coli и Corynebacterium, и т.д., которые широко используются при продукции L-аминокислот и других полезных веществ.
Согласно корейским патентам №10-0838038 и 10-2139806 нуклеотидные последовательности генов, кодирующих белки, включающие ферменты, связанные с продукцией L-лизина, или их аминокислотные последовательности, модифицируют для увеличения экспрессии генов или для удаления не являющихся необходимыми генов и, посредством этого, улучшают способность к продуцированию L-лизина. Кроме того, в публикации корейского патента №10-2020-0026881 раскрыт способ замены существующего промотора гена промотором с сильной активностью для того, чтобы увеличить экспрессию гена, кодирующего фермент, участвующий в продукции L-лизина.
Как было описано, разрабатывают целый ряд способов для увеличения способности к продуцированию L-лизина. Тем не менее, поскольку существуют десятки типов белков, таких как ферменты, транскрипционные факторы, транспортные белки и т.д., которые прямо или опосредованно участвуют в продукции L-лизина, необходимо проводить много исследований по тому, увеличивается ли или нет способность к продуцированию L-лизина согласно изменениям активностей данных белков.
Документы предшествующего уровня техники
Патентные документы
Корейский патент №10-0838038
Корейский патент №10-2139806
Публикация корейского патента №10-2020-0026881
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Техническая проблема
Целью настоящего раскрытия является предложение варианта Corynebacterium glutamicum с улучшенной способностью к продуцированию L-лизина.
Кроме того, другой целью настоящего раскрытия является предложение способа продуцирования L-лизина с использованием данного варианта.
Техническое решение
Авторы настоящего изобретения исследовали разработку нового варианта с улучшенной способностью к продуцированию L-лизина с использованием штамма Corynebacterium glutamicum и, в результате, они обнаружили то, что продукция L-лизина увеличивается посредством замены нуклеотидной последовательности в конкретном положении в промоторе гена aspB, кодирующего аспартатаминотрансферазу, который участвует в поставке аспартата - предшественника лизина - в биосинтетическом пути L-лизина, посредством этого осуществляя настоящее раскрытие.
Согласно одному аспекту настоящего раскрытия предложен вариант Corynebacterium glutamicum с улучшенной способностью к продуцированию L-лизина посредством усиления активности аспартатаминотрансферазы.
Термин «аспартатаминотрансфераза» (аспартаттрансаминаза) в том виде, как он здесь используется, относится к ферменту, который катализирует реакцию превращения оксалоацетата до аспартата в пути биосинтеза L-лизина.
Согласно конкретному воплощению настоящего раскрытия аспартатаминотрансфераза может происходить из штамма рода Corynebacterium. В частности, штамм рода Corynebacterium может представлять собой Corynebacterium glutamicum, Corynebacterium crudilactis, Corynebacterium deserti, Corynebacterium callunae, Corynebacterium suranareeae, Corynebacterium lubricantis, Corynebacterium doosanense, Corynebacterium efficiens, Corynebacterium uterequi, Corynebacterium stationis, Corynebacterium pacaense, Corynebacterium singulare, Corynebacterium humireducens, Corynebacterium marinum, Corynebacterium halotolerans, Corynebacterium spheniscorum, Corynebacterium freiburgense, Corynebacterium striatum, Corynebacterium canis, Corynebacterium ammoniagenes, Corynebacterium renale, Corynebacterium pollutisoli, Corynebacterium imitans, Corynebacterium caspium, Corynebacterium testudinoris, Corynebacaterium pseudopelargi или Corynebacterium flavescens, но не ограничивается ими.
Термин «усиление активности» в том виде, как он здесь используется, означает то, что уровни экспрессии генов, кодирующих белки, такие как целевые ферменты, транскрипционные факторы, транспортные белки и т.д., увеличиваются посредством введения вновь или усиления данных генов по сравнению с генами штамма дикого типа или штамма перед модификацией. Такое усиление активности также включает случай, где активность самого белка увеличивается посредством замены, вставки или делеции нуклеотида кодирующего гена или их комбинации по сравнению с активностью белка, которым исходно обладал микроорганизм, и случай, где общая ферментативная активность в клетках выше, чем активность штамма дикого типа или штамма перед модификацией из-за повышенной экспрессии или повышенной трансляции кодирующих их генов и их комбинации.
Согласно конкретному воплощению настоящего раскрытия усиление активности аспартатаминотрансферазы может быть индуцировано сайт-специфичной мутацией в промоторе гена, кодирующего аспартатаминотрансферазу.
Согласно конкретному воплощению настоящего раскрытия промотор гена, кодирующего аспартатаминотрансферазу, может быть представлен нуклеотидной последовательностью SEQ ID NO: 1.
Термин «промотор» в том виде, как он здесь используется, относится к специфической области ДНК, которая регулирует транскрипцию гена посредством включения сайта связывания РНК-полимеразы, которая инициирует транскрипцию мРНК интересующего гена и обычно находится выше сайта начала транскрипции. Промотор у прокариот определяется как область около сайта начала транскрипции, где связывается РНК-полимераза, и он обычно состоит из двух коротких нуклеотидных последовательностей в областях пар оснований -10 и -35 выше сайта начала транскрипции. В настоящем раскрытии мутация промотора заключается в том, что промотор улучшается для того, чтобы иметь более высокую активность по сравнению с промотором дикого типа и может увеличивать экспрессию генов, расположенных ниже, посредством индуцирования мутаций в промоторной области, расположенной выше сайта начала транскрипции.
Согласно конкретному воплощению настоящего раскрытия усиление активности аспартатаминотрансферазы может быть обусловлено заменой одного или более чем одного основания в областях от -100 до -10 выше от сайта начала транскрипции в последовательности промотора гена, кодирующего аспартатаминотрансферазу.
Более конкретно, мутация промотора по настоящему раскрытию может представлять собой последовательную или непоследовательную замену одного или более чем одного основания в областях от -100 до -10, предпочтительно последовательную или непоследовательную замену одного, двух, трех, четырех или пяти оснований в областях от -95 до -15, областях от -95 до -55, областях от -55 до -40 или областях от -30 до -15.
Согласно одному типичному воплощению настоящего раскрытия С, который представляет собой нуклеотидную последовательность в области -22 промоторной последовательности гена aspB, кодирующего аспартатаминотрансферазу штамма Corynebacterium glutamicum, заменяется на G с получением варианта Corynebacterium glutamicum, имеющего новую промоторную последовательность гена aspB. Такой вариант Corynebacterium glutamicum может включать измененный промотор гена aspB, который представлен нуклеотидной последовательностью SEQ ID NO: 2.
Кроме того, согласно одному типичному воплощению настоящего раскрытия Т, который представляет собой нуклеотидную последовательность в области -45 промоторной последовательности гена aspB, кодирующего аспартатаминотрансферазу штамма Corynebacterium glutamicum, заменяется на А с получением варианта Corynebacterium glutamicum, имеющего новую промоторную последовательность гена aspB. Такой вариант Corynebacterium glutamicum может включать измененный промотор гена aspB, который представлен нуклеотидной последовательностью SEQ ID NO: 3.
Согласно одному типичному воплощению настоящего раскрытия Т, который представляет собой нуклеотидную последовательность в области -88 промоторной последовательности гена aspB, кодирующего аспартатаминотрансферазу штамма Corynebacterium glutamicum, заменяется на А с получением варианта Corynebacterium glutamicum, имеющего новую промоторную последовательность гена aspB. Такой вариант Corynebacterium glutamicum может включать измененной промотор гена aspB, который представлен нуклеотидной последовательностью SEQ ID NO: 4.
Термин «улучшенная способность к продукции» в том виде, как он здесь используется, означает повышенную продуктивность в отношении L-лизина по сравнению с продуктивностью родительского штамма. Родительский штамм относится к штамму дикого типа или варианту штамма, который подвергся мутации, и он включает субъекта, который непосредственно мутирован или преобразован рекомбинантным вектором и т.д. В настоящем раскрытии родительский штамм может представлять собой штамм Corynebacterium glutamicum дикого типа или штамм, мутировавший из дикого типа.
Согласно конкретному воплощению настоящего раскрытия родительский штамм представляет собой вариант, в котором индуцируются мутации в последовательностях генов (например, генов lysC, zwf и hom), участвующих в продукции лизина, и он может представлять собой штамм Corynebacterium glutamicum (далее именуемый «штамм DS1 Corynebacterium glutamicum»), депонированный в Корейский центр культивирования микроорганизмов 2 апреля 2021 г. с номером доступа KCCM12969P.
Вариант Corynebacterium glutamicum по настоящему раскрытию, имеющий улучшенную способность к продуцированию L-лизина, может включать мутировавшую промоторную последовательность гена, кодирующего аспартатаминотрансферазу.
Согласно конкретному воплощению настоящего раскрытия данный вариант может включать любую из нуклеотидных последовательностей, представленных SEQ ID NO: 2-4, в качестве промоторной последовательности гена аспартатаминотрансферазы.
Согласно одному типичному воплощению настоящего раскрытия данный вариант может включать мутацию промотора гена aspB, кодирующего аспартатаминотрансферазу, демонстрируя, посредством этого, повышенную способность к продуцированию L-лизина по сравнению с родительским штаммом, и, в частности, может демонстрировать 3%-ное или большее, в частности, от 3%-ного до 40%-ного и, более конкретно, от 5%-ного до 30%-ного увеличения продукции L-лизина по сравнению с родительским штаммом, продуцируя посредством этого от 65 г до 90 г L-лизина, предпочтительно от 70 г до 80 г L-лизина на 1 л культуры данного штамма.
Получение варианта Corynebacterium glutamicum по конкретному воплощению настоящего раскрытия может быть достигнуто посредством рекомбинантного вектора, включающего вариант, в котором заменена часть последовательности промотора гена, кодирующего аспартатаминотрансферазу у родительского штамма.
Термин «часть» в том виде, как он здесь используется, означает не всю нуклеотидную последовательность или последовательность полинуклеотида, и может содержать от 1 до 300, предпочтительно от 1 до 100 и более предпочтительно от 1 до 50 нуклеотидов, но не ограничиваясь ими.
Термин «вариант» в том виде, как он здесь используется, относится к варианту промотора, в котором заменено одно или более чем одно основание в областях от -100 до -10 в промоторной последовательности гена аспартатаминотрансферазы, участвующего в биосинтезе L-лизина.
Согласно конкретному воплощению настоящего раскрытия варианты, в каждом из которых нуклеотидная последовательность в областях -22, -45 или -88 в последовательности промотора гена аспартатаминотрансферазы заменена на G, А или А, могут иметь нуклеотидную последовательность SEQ ID NO: 2, SEQ ID NO: 3 или SEQ ID NO: 4 соответственно.
Термин «вектор» в том виде, как он здесь используется, представляет собой экспрессионный вектор, способный экспрессировать целевой белок в подходящей клетке-хозяине, и он относится к генетической конструкции, включающей существенные регуляторные элементы, которые связаны функциональным образом таким образом, что экспрессируется вставка гена. Здесь термин «связанный функциональным образом» означает то, что ген, требующий экспрессии, и его регуляторная последовательность функционально связаны друг с другом с индукцией экспрессии гена, и «регуляторные элементы» включают промотор для осуществления транскрипции, любую последовательность оператора для осуществления контроля транскрипции, последовательность, кодирующую подходящий сайт связывания рибосомы с мРНК, и последовательность, контролирующую терминацию транскрипции и трансляции. Такой вектор может включать плазмидный вектор, космидный вектор, бактериофаговый вектор, вирусный вектор и т.д., но не ограничивается ими.
«Рекомбинантный вектор» в том виде, как здесь используется данный термин, трансформируют в подходящую клетку-хозяина, и затем он реплицируется независимо от генома данной клетки-хозяина или может быть интегрирован в сам геном. В данном отношении «подходящая клетка-хозяин», где данный вектор является реплицируемым, может включать репликатор, который представляет собой конкретную нуклеотидную последовательность, в которой инициируется репликация.
Для трансформации выбирают подходящую технологию введения вектора, в зависимости от клетки-хозяина для экспрессии целевого гена в данной клетке-хозяине. Например, введение вектора может осуществляться электропорацией, тепловым шоком, осаждением с фосфатом кальция (CaPO4), осаждением с хлоридом кальция (CaCl2), микроинъекцией, способом с полиэтиленгликолем (PEG), способом с DEAE-декстраном, способом с катионными липосомами, способом с ацетатом лития-DMSO (диметилсульфоксид) или их комбинациями. При условии, что трансформированный ген может экспрессироваться в клетке-хозяине, он может быть включен без ограничения, независимо от того, вставлен ли или не вставлен данный ген в хромосому клетки-хозяина или локализуется вне хромосомы.
Клетки-хозяева включают клетки, трансфицированные, трансформированные или инфицированные рекомбинантным вектором или полинуклеотидом по настоящему раскрытию in vivo или in vitro. Клетки-хозяева, включающие рекомбинантный вектор по настоящему раскрытию, представляют собой рекомбинантные клетки-хозяева, рекомбинантные клетки или рекомбинантные микроорганизмы.
Кроме того, рекомбинантный вектор по настоящему раскрытию может включать селективный маркер, который предназначен для отбора трансформанта (клетки-хозяина), трансформированного данным вектором. В среде, обработанной селективным маркером, только клетки, экспрессирующие данный селективный маркер, могут выживать, и, таким образом, могут быть отобраны трансформированные клетки. Селективный маркер может быть представлен канамицином, стрептомицином, хлорамфениколом и т.д., но не ограничивается ими.
Гены, вставленные в рекомбинантный вектор для трансформации по настоящему раскрытию, могут быть введены в клетки-хозяева, такие как микроорганизмы рода Corynebacterium, из-за кроссинговера посредством гомологичной рекомбинации.
Согласно конкретному воплощению настоящего раскрытия клетка-хозяин может представлять собой штамм рода Corynebacterium, например, штамм Corynebacterium glutamicum.
Кроме того, согласно другому аспекту настоящего раскрытия предложен способ продуцирования L-лизина, включающий стадии а) культивирования варианта Corynebacterium glutamicum в среде; и б) выделения L-лизина из данного варианта или среды, в которой культивируется данный вариант.
Культивирование может осуществляться согласно подходящей среде и культуральным условиям, известным в данной области, и любой специалист в данной области может легко корректировать и применять данную среду и культуральные условия. В частности, данная среда может представлять собой жидкую среду, но не ограничивается ей. Способ культивирования может включать, например, периодическое культивирование, непрерывное культивирование, культивирование с подпиткой или их комбинации, но не ограничивается ими.
Согласно конкретному воплощению настоящего раскрытия данная среда должна удовлетворять требованиям конкретного штамма правильным образом, и она может быть подходящим образом модифицирована специалистом в данной области. Относительно культуральной среды для штамма рода Corynebacterium может быть сделана ссылка на известный документ (Manual of Methods for General Bacteriology. American Society for Bacteriology. Washington D.C., USA, 1981), но не ограничиваясь им.
Согласно конкретному воплощению настоящего раскрытия данная среда может включать разные источники углерода, источники азота и компоненты-микроэлементы. Источники углерода, которые можно использовать, включают сахариды и углеводы, такие как такие как глюкоза, сахароза, лактоза, фруктоза, мальтоза, крахмал, целлюлоза и т.д., жиры и масла, такие как соевое масло, подсолнечное масло, касторовое масло, кокосовое масло и т.д., жирные кислоты, такие как пальмитиновая кислота, стеариновая кислота и линолевая кислота, спирты, такие как глицерин и этанол, и органические кислоты, такие как уксусная кислота. Данные вещества можно использовать индивидуально или в смеси, но не ограничиваясь ими. Источники азота, которые можно использовать, включают пептон, дрожжевой экстракт, мясной экстракт, солодовый экстракт, жидкий кукурузный экстракт, соевый жмых, мочевину или неорганические соединения, например, сульфат аммония, хлорид аммония, фосфат аммония, карбонат аммония и нитрат аммония. Данные источники азота также можно использовать индивидуально или в смеси, но не ограничиваясь ими. Источники фосфора, которые можно использовать, могут включать калия дигидрофосфат или дикалия гидрофосфат, или соответствующие натрийсодержащие соли, но не ограничиваются ими. Кроме того, культуральная среда может включать соли металлов, такие как сульфат магния или сульфат железа, которые требуются для роста, но не ограничиваются ими. Кроме того, культуральная среда может включать незаменимые ростовые вещества, такие как аминокислоты и витамины. Кроме того, в данной культуральной среде можно использовать подходящие предшественники. Среду или индивидуальные компоненты можно добавлять в культуральную среду во время культивирования подходящим способом периодически или непрерывно, но не ограничиваясь ими.
Согласно конкретному воплощению настоящего раскрытия рН культуральной среды можно корректировать добавлением в культуральную среду микроорганизма подходящим способом во время культивирования таких соединений, как гидроксид аммония, гидроксид калия, аммиак, фосфорная кислота и серная кислота. Кроме того, во время культивирования пенообразование можно подавлять с использованием пеногасителя, такого как сложный полигликоле вый эфир жирной кислоты. Кроме того, для поддержания культуральной среды в аэробном состоянии в данную культуральную среду можно инъецировать кислород или кислородсодержащий газ (например, воздух). Температура данной культуральной среды обычно может составлять от 20°С до 45°С, например, от 25°С до 40°С. Культивирование можно продолжать, пока не продуцируется желательное количество полезного вещества. Например, время культивирования может составлять от 10 часов до 160 часов.
Согласно конкретному воплощению настоящего раскрытия на стадии выделения L-лизина из культивируемого варианта и среды, в которой культивируется данный вариант, продуцированный L-лизин может быть отобран или выделен из культуральной среды с использованием подходящего способа, известного в данной области, в зависимости от способа культивирования. Например, можно использовать центрифугирование, фильтрование, экстракцию, распыление, сушку, упаривание, осаждение, кристаллизацию, электрофорез, фракционное растворение (например, осаждение сульфатом аммония), хроматографию (например, ионообменная, аффинная, гидрофобная и гель-фильтрация) и т.д., но способ не ограничивается ими.
Согласно конкретному воплощению настоящего раскрытия на стадии выделения лизина культуральную среду центрифугируют на низкой скорости для удаления биомассы, и полученный супернатант можно разделять посредством ионообменной хроматографии.
Согласно конкретному воплощению настоящего раскрытия стадия выделения L-лизина может включать способ очистки L-лизина. Полезные эффекты
Вариант Corynebacterium glutamicum по настоящему раскрытию может улучшать продукционный выход L-лизина посредством увеличения или усиления экспрессии гена, кодирующего аспартатаминотрансферазу, по сравнению с родительским штаммом.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
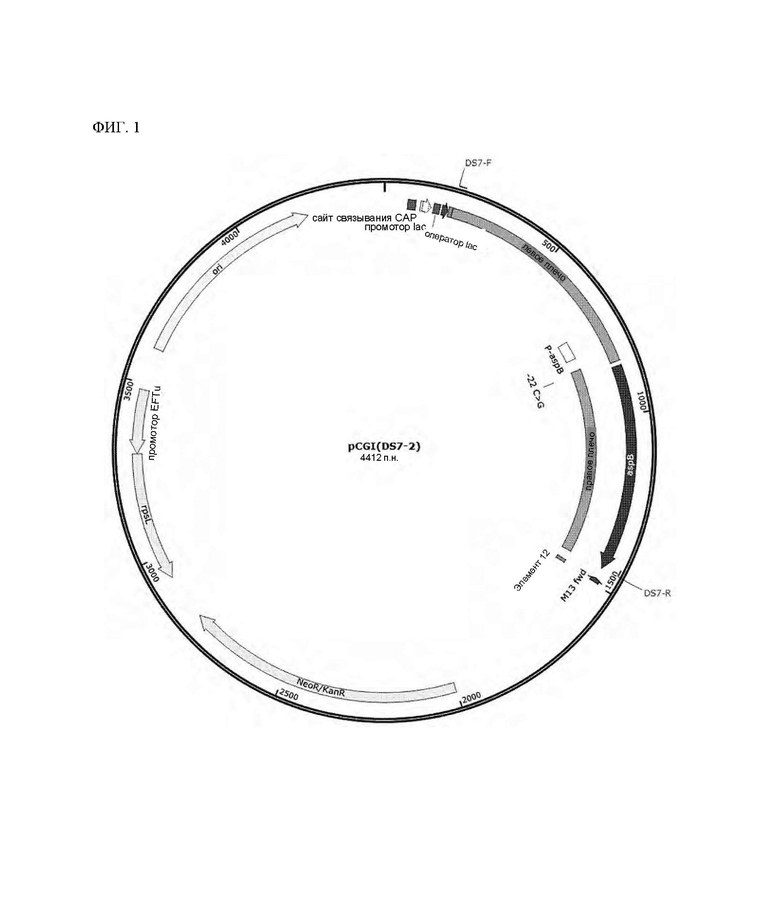
На ФИГ. 1 показана конструкция рекомбинантного вектора pCGI(DS7-2) для замены последовательности в положении 22 промоторной области с С на G согласно одному типичному воплощению настоящего раскрытия;
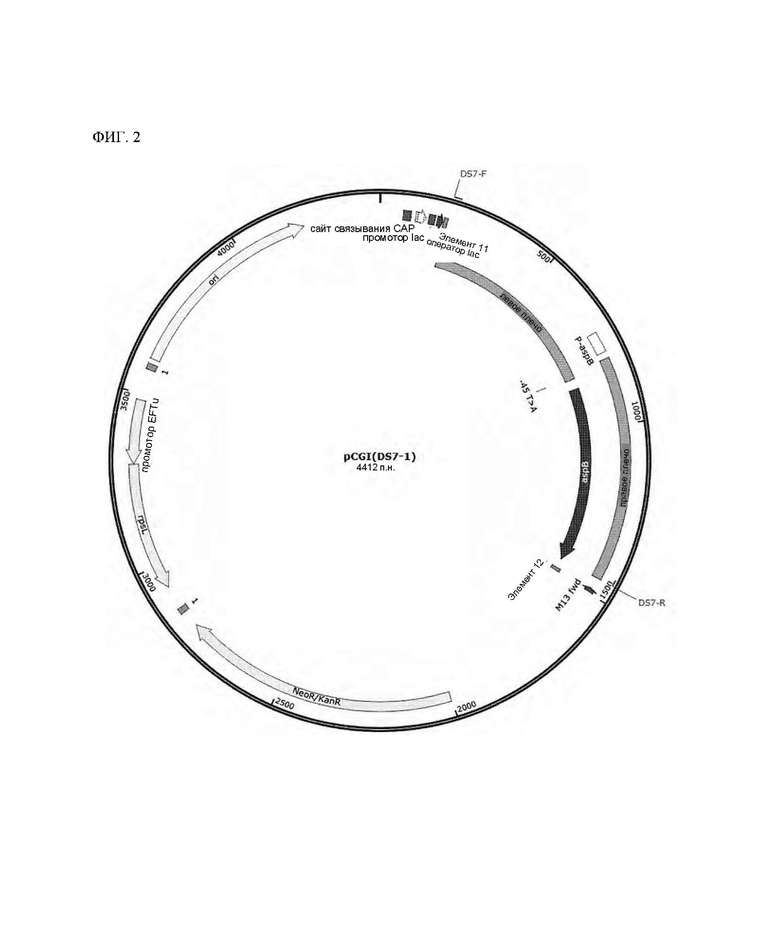
На ФИГ. 2 показана конструкция рекомбинантного вектора pCGI(DS7-l) для замены последовательности в положении 45 промоторной области с Т на А согласно одному типичному воплощению настоящего раскрытия; и
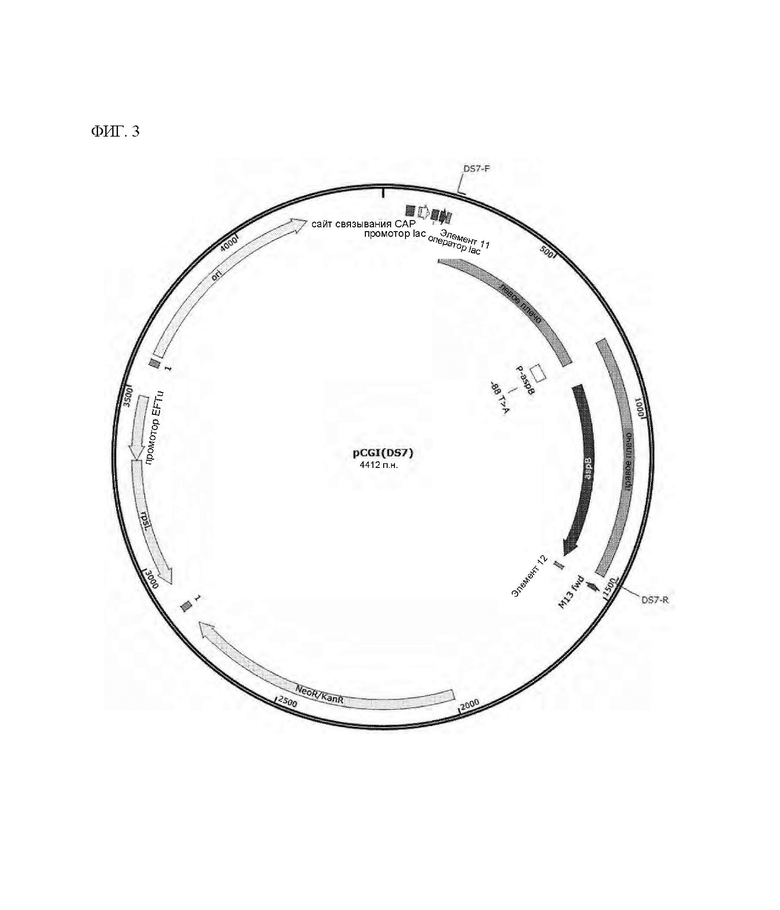
На ФИГ. 3 показана конструкция рекомбинантного вектора pCGI(DS7) для замены последовательности в положении 88 промоторной области с Т на А согласно одному типичному воплощению настоящего раскрытия.
Далее настоящее раскрытие будет описано более подробно. Однако данное описание просто приводится для того, чтобы помочь пониманию настоящего раскрытия, и объем настоящего раскрытия не ограничивается данным типичным описанием.
Пример 1. Получение варианта Corynebacterium glutamicum
Для получения варианта Corynebacterium glutamicum с повышенной аспартатаминотрансферазной активностью для индукции случайной мутации использовали штамм DS1 Corynebacterium glutamicum.
1-1. Мутагенез
Штамм DS1 Corynebacterium glutamicum инокулировали в колбу, содержащую 50 мл жидкой среды СМ (5 г глюкозы, 2,5 г NaCl, 5,0 г дрожжевого экстракта, 1,0 г мочевины, 10,0 г полипептона и 5,0 г говяжьего экстракта, рН 6,8), и добавляли N-метил-N'-нитро-N-нитрозогуанидин (NTG), который представляет собой мутаген, в конечной концентрации 300 мкг/мл, с последующим культивированием при 30°С со встряхиванием при 200 об/мин в течение 20 часов. Затем, для индуцирования дополнительной мутации осуществляли воздействие УФ (ультрафиолетовое излучение) в течение 20 минут. После завершения культивирования культуру центрифугировали при 12000 об/мин в течение 10 минут для удаления супернатанта, и получаемое в результате один раз промывали физиологическим раствором и три раза или более промывали фосфатным буфером. Это суспендировали в 5 мл фосфатного буфера, распределяли на твердой среде СМ (в жидкую среду СМ дополнительно добавляли 15 г/л агара и 8% лизина) и культивировали при 30°С в течение 30 часов для выделения 100 колоний.
1-2. Отбор вариантов с улучшенной способностью к продуцированию L-лизина
Каждые 5% 100 выделенных колоний инокулировали в колбу, содержащую 10 мл жидкой среды для продукции, показанной ниже в Таблице 1, и культивировали со встряхиванием при 200 об/мин при 30°С в течение 30 часов. Поглощение каждой культуры измеряли при ОП 610 нм, и сравнивали продукцию L-лизина для отбора 10 колоний, продуцирующих 75,0 г/л или более L-лизина, по сравнению со штаммом DS1 Corynebacterium glutamicum, в котором мутация не была индуцирована, и его нуклеотидные последовательности анализировали для идентификации сайтов мутации в промоторе гена aspB. В результате проверки нуклеотидных последовательностей вариантов DS1 Corynebacterium glutamicum, в которых были индуцированы данные мутации, идентифицировали три типа мутаций: С>G в положении -22, Т>А в положении -45 и Т>А в положении -88.

Затем проводили эксперимент для проверки увеличения продуктивности в отношении L-лизина из-за трех типов мутаций промотора в гене aspB.
Пример 2. Получение варианта Corynebacterium glutamicum
Для получения варианта Corynebacterium glutamicum с усиленной аспартатаминотрансферазной активностью использовали штамм DS1 Corynebacterium glutamicum и DH5a E. coli (HIT Competent cells™, кат. № RH618).
Штамм DS1 Corynebacterium glutamicum культивировали при температуре 30°C в среде СМ или твердой среде (добавление 15 г/л агара, по необходимости) (рН 6,8), имеющей следующий состав: 5 г глюкозы, 2,5 г NaCl, 5,0 г дрожжевого экстракта, 1,0 г мочевины, 10,0 г полипептона и 5,0 г говяжьего экстракта в 1 л дистиллированной воды.
DH5a Е. coli культивировали при температуре 37°С в среде LB, имеющей следующий состав: 10,0 г триптона, 10,0 г NaCl и 5,0 г дрожжевого экстракта в 1 л дистиллированной воды.
Использовали антибиотики - канамицин и стрептомицин продукты Sigma а анализ секвенирования ДНК проводили в Macrogen Co., Ltd.
2-1. Получение рекомбинантного вектора
Для увеличения продуктивности в отношении лизина посредством усиления поставки в штамм аспартата, предшественника лизина, намеревались усилить активность аспартатаминотрансферазы. Способ, используемый в данном Примере, индуцировал специфичную мутацию в промоторе гена aspB для того, чтобы увеличить экспрессию гена aspB, кодирующего аспартатаминотрансферазу. Нуклеотидную последовательность в положении -22 промотора гена aspB заменяли на G, и фрагмент из 1256 п.н. вокруг гена aspB в геноме каждого варианта, полученного в Примере 1, амплифицировали посредством ПЦР (полимеразная цепная реакция), и клонировали в рекомбинантный вектор pCGI (см. [Kim et al., Journal of Microbiological Methods 84 (2011) 128-130]). Данную плазмиду называли pCGI(DS7-2) (см. ФИГ. 1). Для конструирования данной плазмиды использовали праймеры, показанные в Таблице 2 ниже, для амплификации каждого фрагмента гена.

ПЦР проводили с использованием приведенных выше праймеров при следующих условиях. От 25 до 30 циклов проводили с использованием термоциклера (ТР600, TAKARA BIO Inc., Япония) в присутствии 1 единицы смеси ДНК-полимеразы pfu-X (Solgent) с использованием 1 пМ олигонуклеотида и 10 нг хромосомной ДНК варианта DS1 Corynebacterium glutamicum (мутация происходила в положении -22 промотора), отобранного в Примере 1, в качестве матрицы в реакционном растворе, в который добавляли 100 мкл каждого дезоксинуклеотидтрифосфата (дАТФ, дЦТФ, дГТФ, дТТФ). ПЦР проводили при следующих условиях: (1) стадия денатурации: при 94°С в течение 30 секунд, (2) стадия отжига: при 58°С в течение 30 секунд, и (3) стадия элонгации: при 72°С в течение 1 минуты 2 минут (время полимеризации 2 минуты на 1 т.п.н.).
Каждый фрагмент гена, полученный таким образом, клонировали в вектор pCGI с использованием самосборочного клонирования. Данный вектор трансформировали в Е. coli DH5a, производили посев на чашки с LB-агаром, содержащим 50 мкг/мл канамицина, и культивировали при 37°С в течение 24 часов. Образующиеся наконец колонии выделяли, и подтверждали, правильно ли присутствовала данная вставка в векторе, который выделяли и использовали при рекомбинации штамма Corynebacterium glutamicum.
2-2. Получение варианта
Штамм DS7-2, который представляет собой вариант штамма, получали с использованием вектора pCGI(DS7-2). Данный вектор получали в конечной концентрации 1 мкг/мл или больше, и первичную рекомбинацию индуцировали в штамме DS1 Corynebacterium glutamicum с использованием электропорации (см. документ [Tauch et al., FEMS Microbiology letters 123 (1994) 343-347]). В это время подвергнутый электропорации штамм затем распределяли на твердой среде СМ, содержащей 20 мкг/мкл канамицина, с выделением колоний, и затем подтверждали посредством ПЦР и анализа секвенирования оснований, правильно ли данный вектор был вставлен в индуцированное положение в геноме. Для индукции вторичной рекомбинации выделенный штамм инокулировали в жидкую среду СМ, культивировали в течение ночи или дольше и затем распределяли на среде с агаром, содержащей 40 мг/л стрептомицина для выделения колоний. После проверки устойчивости к канамицину в конечных выделенных колониях посредством анализа секвенирования оснований подтвердили, были ли введены мутации в промотор гена aspB в штаммах без устойчивости к антибиотику (см. документ [Schafer et al., Gene 145 (1994) 69-73]). Наконец, получали вариант Corynebacterium glutamicum (DS7-2), в который вводили мутировавший промотор гена aspB.
Пример 3. Получение варианта Corynebacterium glutamicum
Вариант Corynebacterium glutamicum получали таким же способом, как и в Примере 2, за исключением того, что производили замену нуклеотидной последовательности в положении -45 промотора гена aspB от Т до А, и вариант Corynebacterium glutamicum DS1 (мутация происходила в положении -45 промотора), отобранный в Примере 1, использовали в качестве ДНК-матрицы.
Здесь для конструирования плазмиды использовали праймеры, показанные в Таблице 2 ниже, для амплификации каждого фрагмента гена, и штамм DS7-1, который представляет собой вариант штамма, получали с использованием полученного вектора плазмиды pCGI(DS7-1) (см. ФИГ. 2). Наконец, получали вариант Corynebacterium glutamicum (DS7-1), в который вводили мутировавший ген aspB.
Пример 4. Получение варианта Corynebacterium glutamicum
Вариант Corynebacterium glutamicum получали таким же способом, как и в Примере 2, за исключением того, что производили замену нуклеотидной последовательности в положении -88 промотора гена aspB с Т на А, и вариант Corynebacterium glutamicum DS1 (мутация происходила в положении -88 промотора), отобранный в Примере 1, использовали в качестве ДНК-матрицы.
Здесь для конструирования плазмиды использовали праймеры, показанные в Таблице 2 ниже, для амплификации каждого фрагмента гена, и штамм DS7, который представляет собой вариант штамма, получали с использованием полученного вектора плазмиды pCGI(DS7) (см. ФИГ. 3). Наконец, получали вариант Corynebacterium glutamicum (DS7), в который вводили мутировавший ген aspB.
Экспериментальный пример 1. Сравнение продуктивности в отношении L-лизина между вариантами
Продуктивность в отношении L-лизина сравнивали между родительским штаммом DS1 Corynebacterium glutamicum, штаммами DS7-2, DS7-1 и DS7, которые представляют собой варианты, продуцирующие лизин, полученные в Примерах 2-4.
Каждый штамм инокулировали в 100 мл колбу, содержащую 10 мл лизиновой среды, имеющей композицию, как показано в Таблице 1, и культивировали со встряхиванием при 180 об/мин при 30°С в течение 28 часов. После завершения культивирования анализ L-лизина проводили посредством измерения продукции L-лизина с использованием ВЭЖХ (высокоэффективная жидкостная хроматография) (Shimazu, Япония). Результаты показаны в Таблице 3.
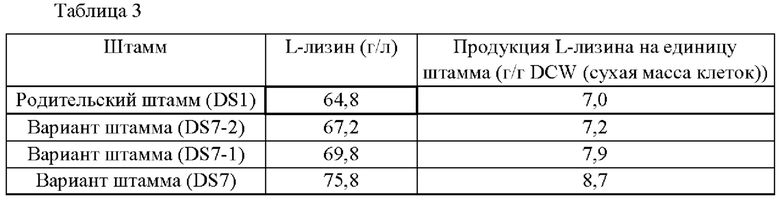
Как показано в Таблице 3, подтвердили то, что варианты Corynebacterium glutamicum - штаммы DS7-2, DS7-1 и DS7 - демонстрировали примерно 3,7%-ное, 7,7%-ное и 16,9%-ное увеличение продуктивности в отношении L-лизина, соответственно, по сравнению с родительским штаммом DS1 Corynebacterium glutamicum из-за замены специфических положений (области -22, -45 или -88) в последовательности промотора гена aspB оптимальной нуклеотидной последовательностью для усиления биосинтетического пути лизина. В частности, подтвердили то, что штамм DS7 демонстрировал наивысшую продуктивность по сравнению с другими вариантами. Данные результаты показывают то, что усиленная экспрессия гена aspB может стимулировать поставку предшественника лизина, стимулируя, посредством этого, способность данного штамма к продуцированию L-лизина.
Выше настоящее раскрытие было описано со ссылкой на его предпочтительные типичные воплощения. Специалистам в области, к которой относится настоящее раскрытие, будет понятно то, что настоящее раскрытие можно осуществлять в модифицированных формах без отступления от важных характеристик настоящего раскрытия. Соответственно, раскрытые здесь типичные воплощения следует рассматривать в иллюстративном аспекте, а не в ограничивающем аспекте. Объем настоящего раскрытия показан не в приведенном выше объяснении, а в приложенной формуле изобретения, и все различия в пределах эквивалентного ей объема следует интерпретировать как включенные в настоящее раскрытие.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> CJ CheilJedang Corporation
<120> ВАРИАНТ CORYNEBACTERIUM GLUTAMICUM, ИМЕЮЩИЙ УЛУЧШЕННУЮ СПОСОБНОСТЬ
К ПРОДУКЦИИ L-ЛИЗИНА, И СПОСОБ ПРОДУЦИРОВАНИЯ L-ЛИЗИНА С ЕГО ПРИМЕНЕНИЕМ
<130> OPA23236
<150> KR 10-2021-0055536
<151> 2021-04-29
<150> KR 10-2021-0066151
<151> 2021-05-24
<160> 6
<170> KoPatentIn 3.0
<210> 1
<211> 134
<212> DNA
<213> Artificial Sequence
<220>
<223> последовательность промотора гена aspB
<400> 1
agggtcagcg agctctgacg ggtgcgcggg atcccctaaa acgtctagag tagtggcttg 60
aggtcactgc tctttttttg tgcccttttt tggtccgtct attttgccac cacatgcgga 120
ggtacgcagt tatg 134
<210> 2
<211> 134
<212> DNA
<213> Artificial Sequence
<220>
<223> мутантная последовательность промотора гена aspB
<400> 2
agggtcagcg agctctgacg ggtgcgcggg atcccctaaa acgtctagag tagtggcttg 60
aggtcactgc tctttttttg tgcccttttt tggtccgtct attttgccag cacatgcgga 120
ggtacgcagt tatg 134
<210> 3
<211> 134
<212> DNA
<213> Artificial Sequence
<220>
<223> мутантная последовательность промотора гена aspB
<400> 3
agggtcagcg agctctgacg ggtgcgcggg atcccctaaa acgtctagag tagtggcttg 60
aggtcactgc tctttttttg tgccctattt tggtccgtct attttgccac cacatgcgga 120
ggtacgcagt tatg 134
<210> 4
<211> 134
<212> DNA
<213> Artificial Sequence
<220>
<223> мутантная последовательность промотора гена aspB
<400> 4
agggtcagcg agctctgacg ggtgcgcggg atcccctaaa acgactagag tagtggcttg 60
aggtcactgc tctttttttg tgcccttttt tggtccgtct attttgccac cacatgcgga 120
ggtacgcagt tatg 134
<210> 5
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> DS7-F
<400> 5
cgagaattac cggcgatg 18
<210> 6
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> DS7-R
<400> 6
tcaccgtgaa accagtcg 18
<---
Изобретение относится к биотехнологии. Предложен промотор, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 4. Также предложены микроорганизм Corynebacterium glutamicum, продуцирующий L-лизин и содержащий нуклеотидную последовательность, представленную SEQ ID NO: 4, и способ продуцирования L-лизина с использованием указанного микроорганизма. Изобретение обеспечивает улучшенный выход продукции L-лизина по сравнению с родительским штаммом. 3 н.п. ф-лы, 3 ил., 3 табл., 4 пр.
1. Промотор, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 4.
2. Микроорганизм Corynebacterium glutamicum, продуцирующий L-лизин, содержащий нуклеотидную последовательность, представленную SEQ ID NO: 4.
3. Способ продуцирования L-лизина, включающий следующие стадии:
а) культивирование микроорганизма по п. 2 в среде; и
б) выделение L-лизина из данного микроорганизма или среды, в которой культивируется данный микроорганизм.
| WO 2011108808 A2, 09.09.2011 | |||
| WO 2018226964 A2, 13.12.2018 | |||
| Микроорганизм Corynebacterium sp. с повышенной способностью продуцировать L-лизин и способ получения L-лизина с его помощью | 2014 |
|
RU2667425C2 |
| JIANZHONG XU ET AL | |||
| Станок для шлифования топоров, ножей и т.п. изделий | 1926 |
|
SU13032A1 |
| Journal of Molecular Catalysis B: Enzymatic, Volume | |||

