Изобретение относится к области химии, биологии и может быть использовано для скрининга физиологически активных веществ.
Целью изобретения является упрощение, ускорение и повышение точности способа.
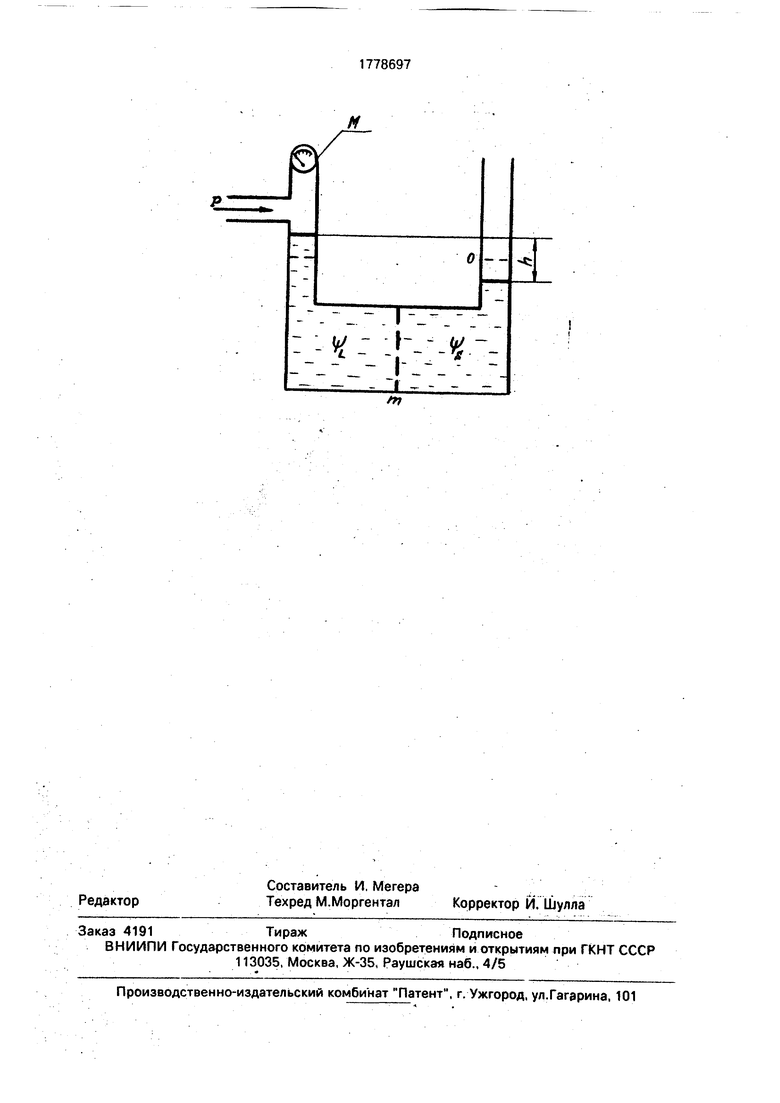
На чертеже представлена модель, объясняющая принцип измерения водного потенциала листа с помощью камеры давления и демонстрирующая механизм увеличения водило потенциала при увеличении проницаемости плазмалеммы. В табл.1 показано влияние физиологически активных соединений на величину водного потенциала листьев пшеницы и осмотический потенциал их ксилемного сока, в табл.2 - изменение параметров водного режима листьев хлопчатника при изменении концентрации раствора физиологически активного соединения, в табл.3 показано перераспределение катионов между симп- ластом и апопластом мезофилла при действии физиологически активного вещества на растение хлопчатника
Предлагаемый способ реализуется следующим образом.
Готовится раствор исследуемого вещества заданной интересуемой концентрации. Этим раствором производится опрыскивание надземной части растения. Затем измеряются параметры водного режима (водный потенциал листа и осмотический потенциал ксилемного экссудата) листа с обработанного исследуемым веществом растения. Эти показатели сравниваются с аналогичными, измеренными на листе контрольного растения, выращенного в тех же условиях, что и опытный вариант.
Измерение водного потенциала листа осуществляют с помощью камеры давления. Осмотический потенциал ксилемного сока, извлеченного из листа камерой давления, определяют микрокриоскопическим осмометром.
Согласно используемой методике для измерения данных показателей водного режима листа лист отделяется от растения лишь к моменту измерения. Лист помещают в камеру давления черешком наружу. В камере давления давление инертного газа (Р) повысл
с
VJ VI оо
о о ч
шается до тех пор, пока оно не уравновесит отрицательное гидростатическое давление, возникшее в ксилеме при срезании листа. В момент появления ксилемного сока на срезе черешка водный потенциал листа fyO равен внешнему давлению газа (L P), регистрируемому манометром Превышение внешнего давления на 1-2 зтм величины водного потенциала листа позволяет извлечь пробу сока из апопласта листа (ксилемы). Оценка осмотического потенциала ($.) этого сока микрокриоскопическим осмометром производится сразу же после определения водного потенциала листа. Определение этих двух параметров (t/. и %) для одного листа производится всего за 4-5 мин.
Уже по изменению водного раствора потенциала 1/ листа по отношению к контролю можно судить о физиологической активности исследуемого вещества: чем больше отличается водный потенциал опытного листа от контрольного, тем выше физиологическая активность исследуемого вещества. Однако такая оценка, обнаруживая эффект, не вскрывает механизм действия исследуемого вещества. Изменение водного потенциала листа в такой ситуации может быть расценено и как следствие изменения эластичности клеточных стенок. Дополнительное измерение осмотического потенциала ксилемного сока, обнаруживающее его изменение по сравнению с контролем, свидетельствует об изменении проницаемости мембран клетки и, в частности, плазмалеммы клеток мезофилла при действии физиологически активного вещества.
Моделирование метода определения водного потенциала листа наглядно демонстрирует причину изменения параметров водного режима листьев. Лист можно схематически представить в виде двух сосудов, разделенных полупроницаемой мембраной (чертеж). Один сосуд соответствует клеткам мезофилла листа с водным потенциалом 1/1. а другой - ксилеме с осмотическим потенциалом $5. До отрыва листа равновесие, существовавшее между клетками мезофилла и ксилемой, создавалось равенством водного потенциала листа и силой натяжения раствора в проводящей системе ксилемы. В момент отрыва листа при перерезании черешка начинает работать закон осмоса и вода ксилемы начинает проникать в клетки мезофилла. Вода из правого сосуда перемещается в левый. В результате уровень жидкости в правом сосуде начинает снижаться, а в левом - повышаться. Это будет продолжаться до тех пор. пока дополнительное давление на жидкость в левом сосуде, образующееся за счет разницы уровней жидкости (h), не скомпенсирует осмотическое
давление растворенного вещества (давление столба h подобно тургорному давлению лимитирует процесс передвижения жидкости из правого сосуда в левый). Если после установления равновесия оказать давление
на поверхность жидкости в левом сосуде сжатым нейтральным газом в такой степени, чтобы вернуть уровни жидкостей в обоих сосудах в исходное состояние (0), то величина этого давления будет соответствовать величине водного потенциала листа $ и может быть измерена манометром (М) (чертеж).
Поскольку у большинства ассимилирующих видов растений Vs 0, давление, регистрируемое манометром при появлении на срезе черешка листа ксилемной жидкости, приравнивается водному потенциалу 1р. Однако, если полупроницаемая мембрана потеряет свойственную ей селективность и
будет пропускать осмотически активные вещества в апопласт листа, то осмотический потенциал ксилемного сока будет сильно отличаться от нуля и при измерении водных потенциалов листьев будет
наблюдаться завышение показателей . Следовательно, завышение показателей водного потенциала листа у обработанных физиологически активными соединениями растений по отношению к контрольному варианту свидетельствует об изменении проницаемости плазмалеммы клеток мезофилла, т.е. об изменении электрохимического градиента на ней, что подтверждается изменением осмотичности ксилемного
сока у листьев опытных растений.
В основе способа лежит определение физиологической активности вещества по реакции на последнее живого растительного объекта, что позволяет повысить точность
способа определения физиологической активности исследуемого вещества.
Проведена серия экспериментов по исследованию физиологической активности ряда классов химических соединений (инсектицидов. поверхностных разобщителей, модификаторов мембран, синтезируемых и испытуемых химическим НИИ по закрытой тематике), которая показала, что механизм действия исследованных физиологически
активных соединений заключается в их воздействии на проницаемость плаэмалеммы, что. в свою очередь, проявляется в изменении водообмена листьев, обработанных ис- следуемым веществом. В табл.1
представлены данные для характерных известных соединении исследуемых классов физиологически активных веществ, описывающие реакцию листьев пшеницы на соот- ветствующее воздействие. Во всех исследуемых случаях наблюдается корреляция в поведении водного потенциала листь- ев и осмотического потенциала их ксилемного экссудата: увеличение 1/V компенсируется уменьшением ifa таким обра- зом, что в сумме абсолютные значения этих параметров равны абсолютной величине водного потенциала листа контрольного растения, т.е. 1/.° $. + t/V. что согласуете я с предлагаемой моделью (чертеж).
Изменяя концентрацию раствора исследуемого вещества, можно менять интенсивность воздействия на живой объект (табл,2) и тем самым установить концентрационные пределы его физиологической ак- тивности по обратимости воздействия на растение вплоть до летальной дозы. Исследуя кинетику описанных выше показателей водного режима по временной шкале длительности воздействия исследуемого веще- ства можно судить о глубине и сложности
перестроек в плазмалемме клеток мезофилла и возможности восстановительных процессов после действия физиологических активных соединений.
Табл.3 приведена в качестве примера перераспределения катионов между симп- ластом и апопластом листа при действии физиологически активного вещества.
Формула изобретения Способ определения физиологической активности вещества, включающий обработку исследуемым веществом растительного материала, исследование мембранного потенциала растительных тканей после обработки и оценку физиологической активности путем сравнения с мембранным потенциалом контроля, отличающийся тем, что, с целью упрощения, ускорения и повышения- точности способа, исследуемым веществом обрабатывают листья, при исследовании мембранного потенциала измеряют водный потенциал листа и осмотический потенциал ксилемного сока этого же листа и чем выше данные показатели, тем выше активность.

| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ДИАГНОСТИКИ ЗАСУХОУСТОЙЧИВОСТИ И ПРОДУКТИВНОСТИ ЗЛАКОВЫХ СЕЛЬСКОХОЗЯЙСТВЕННЫХ КУЛЬТУР | 2006 |
|
RU2339215C2 |
| СПОСОБ ПОЛУЧЕНИЯ МАТЕРИАЛА С КЛЕТОЧНОЙ ПОВЕРХНОСТИ РАСТЕНИЙ | 2020 |
|
RU2839090C1 |
| СРЕДСТВО ДЛЯ ПОВЫШЕНИЯ УРОЖАЙНОСТИ СЕЛЬСКОХОЗЯЙСТВЕННЫХ КУЛЬТУР | 2001 |
|
RU2189960C1 |
| Способ определения скорости флоэмного потока в побегах растений | 1990 |
|
SU1793381A1 |
| Способ определения критического значения дневного водного потенциала листьев древесных растений | 1986 |
|
SU1623581A1 |
| СПОСОБ ОТБОРА ИСХОДНОГО МАТЕРИАЛА ДЛЯ СЕЛЕКЦИИ РАСТЕНИЙ СОИ С КОМПЛЕКСНОЙ УСТОЙЧИВОСТЬЮ К ВОЗБУДИТЕЛЯМ ПЕПЕЛЬНОЙ ГНИЛИ И ФУЗАРИОЗНОГО УВЯДАНИЯ | 2010 |
|
RU2433168C1 |
| НЕТОКСИЧНЫЕ КОМПОЗИЦИИ АГЕНТА ДЛЯ РАСТЕНИЙ, СПОСОБЫ И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2777770C2 |
| СПОСОБ ПОВЫШЕНИЯ УСТОЙЧИВОСТИ РАСТЕНИЙ РАПСА К ИНТЕНСИВНОМУ ХЛОРИДНОМУ ЗАСОЛЕНИЮ | 2013 |
|
RU2522519C1 |
| СПОСОБ ДИАГНОСТИКИ УЗОРЧАТОЙ ТЕКСТУРЫ ДРЕВЕСИНЫ КАРЕЛЬСКОЙ БЕРЕЗЫ | 2015 |
|
RU2596013C1 |
| Способ обратимого образования путей пассивного транспорта анионов хлора в мембранах корневых клеток растения TRIaNea воGотеNSIS каRSт | 1987 |
|
SU1529114A1 |
Использование: биотехнология, физиология и биохимия растений, растениеводство.Сущностьизобретения: физиологическую активность вещества определяют путем воздействия им на растительный материал и измерения биологических показателей при этом в качестве биологических показателей исследуют водный потенциал листа и осмотический потенциал ксилемного сока этого же листа. 1 ил , 3 табл.
Таблица 1
Таблица 2
Таблица 3
Г
: - « г : ---
| Способ определения активности полиеновых антибиотиков | 1981 |
|
SU972400A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Способ определения гормонального действия веществ | 1984 |
|
SU1221598A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| СПОСОБ ОБОРУДОВАНИЯ СКВАЖИН НАПРАВЛЕНИЕМ ПРИ ИХ СТРОИТЕЛЬСТВЕ В МНОГОЛЕТНЕМЕРЗЛЫХ ПОРОДАХ | 1993 |
|
RU2097530C1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
