Настоящее изобретение выполнено при частичном финансировании Правительства США, и Правительство США обладает некоторыми правами на это изобретение.
Настоящее изобретение касается общих вопросов, связанных с пассивной слизистой иммунной защитой и поли-Ig-иммунными реактивами и способами. Здесь термин "поли-Ig" распространяется на полимерные классы антител, т.е. на IgA и IgM-антитела. Антитела класса IgM обычно продуцируются на ранней стадии иммунной реакции и не представляет собой важный фактор в защитном слизистом иммунитете. Таким образом, изобретение в целом относится к полимерным Ig-антителам, и в отношении всех аспектов настоящего изобретения обычными и предпочтительными антителами являются антитела класса IgA, которые обычно выделяются в димерной форме и, в меньшей степени, в форме высших Iga-полимеров.
Многие патогенные бактерии и вирусы первоначально получают доступ в организм, проникая через клеточные выстелки /эпителиальные ткани/ желудочно-кишечного, дыхательного или полового трактов. Специализированный класс антител - IgA-антитела - защищают эти поверхности. Антитела IgA-представляют собой димерные или полимерные молекулы, продуцируемые клетками, расположенными в тканях, находящихся под эпителиальными поверхностями. Они транспортируются эпителиальными клетками в слизистые выделения, где они прошивают или покрывают патогенные микроорганизмы, которые еще не попали в организм, препятствуя патогенным микроорганизмам контактировать и прилипать к эпителиальным клеткам. Таким образом, IgA-антитела оказывают воздействие на патогенные микроорганизмы, которые находятся вне организма, и они осуществляют защиту, препятствуя вхождению в организм через эпителиальные поверхности.
Протекающая в естественных условиях IgA-реакция запускается антигеном, попадающим на слизистые поверхности. Антиген поступает в организм через специфические заборные участки /называемые микроскладкой, или M-клетками/, являющиеся эффективными в отношении трансэпителиального переноса антигена в области слизистой выстелки, содержащей специализированные упорядоченные совокупности клеток слизистой иммунной системы. Говоря точнее, как это показано на фиг. 1, антигены A /показанные в виде зачерненных точек/, находящиеся в полости 1, связывают полостную поверхность из M-клеток в месте 2. Антигены попадают внутрь и проникают в клетку в месте 3, освобождаясь в интераэпителиальном кармане 4, содержащем лимфоидные L-клетки /B- и T-клетки/ и клетки по выявлению и обработке антигенов, такие как макрофаговые клетки /M-клетки/.
Антитела класса IgA, присутствующие у хозяина, иммунизированного естественным образом, транспортируются в выделения посредством связывания со специфическим рецептором /называемым поли-Ig-рецептором/ на базальных /внутренних/ поверхностях из эпителиальных и гландулярных клеток в пределах дыхательной и пищеварительной систем, полового тракта и молочных желез. См. литературу Solara, Kraehenbuhl, "Receptor-Mediated Transepitheli al Transport of Polymeric Immunoglobulins, c.c. 269-298, в книге The Mammary Gland Nelville, Daniel, редакторы, изд-во "Пленум паблишинг", г. Кеймбридж /1987/; Mestecky /1987/, I. Clin, Immunol. , 7:265-276. Комплексы из рецептора и IgA-антитела транспортируются через эти клетки и подвергаются экзоцитозу, попадая на полостные /внешние/ клеточные поверхности, на которых рецептор ферментативно отщепляется, высвобождая IgA-антитело, поступающее в выделения вместе с рецепторным фрагментом, называемым секреторным компонентом /SC/. См. литературу: Mostov et Al. /1980/, Proc. Nat'I Acad Sci US. A., 77: 7257-7261; Solari, R., Kraehenbuhl, J.P., Cell. 36:61-71 /1984/; Kuhn, Kraehenbuhi, J. Biol, Chem., 256:12490-12495 /1981/. Сообщают, что секреторный компонент понижает протеолитический распад IgA-антетел. См. литературу: Lindh, I., I. Immunol., 114:284-286 /1975/; Brownet al., I. Clin Invest., 49: 1374 /1974/.
В общем существующие подходы к иммунизации, при которых рассматривается попадание антигенов, сводятся к продуцированию антител IgG-класса, которые циркулируют по всему организму и нейтрализуют патогенные микроорганизмы после попадания их в организм. Введение антигенов в общем случае не сопровождается возникновением существенной IgA-реакции.
Усилия, направленные на активизацию IgA-защиты на слизистых барьерных поверхностях, сводились к проведению оральной иммунизации либо с целью активной защиты иммунизированного млекопитающего, либо с целью пассивной защиты какого-то другого млекопитающего посредством использования слизистого выделения, взятого у уже иммунизированного млекопитающего. См. литературу: Glass et al., New End I. Med, 308:1389-1392 /1983/; Fubara et al., I. Immunol., 111/2/:395-403 /1973/. Были продуцированы моноклональные IgA-антитела и внесены непосредственно на слизистые поверхности дыхательной системы с целью защиты от проникновения патогенных микроорганизмов. См. литературу: Mazanec et al., I. Virol., 61:2624-2625 /1987/.
Раскрыты различные пути производства и стабилизации моноклональных поли-Ig-антител, дающие реактивы для пассивной слизистой иммунной защиты. Говоря точнее, такие реактивы представляют собой существенно изолированные комплексы, образованные между поли-Ig-антителами, являющимися специфическими для выбранного антигена или семейства антигенов, и стабилизаторным белком, включающим в себя по крайней мере одну поли-Ig-связывающую область поли-Ig-рецептора. Поли-Ig-стабилизатор существенно свободен от каких-либо C-концевых трансмембранных или интрацеллюлярных поли-Ig-рецепторных областей. Получающийся комплекс характеризуется наличием протеолитической стабильности в сравнении с незакомплексованными поли-Ig-антителами, и комплекс обладает специфическими связывающими и иммунозащитными прошивающими свойствами, присущими поли-Ig-антителам. В плане первого аспекта изобретения стабилизаторный белок представляет собой рекомбинантный белок. В плане второго аспекта изобретения поли-Ig-антитело представляет собой моноклональное антитело.
Как более подробно будет говориться ниже, одна из предпочтительных форм комплекса характеризуется наличием ковалентных /дисульфидных/ связей между поли-Ig-антителом и стабилизатором. Такие комплексы могут производиться клеткой, способной продуцировать требуемое поли-Ig-антитело, обладающее специфическими свойствами в отношении выбранного антигена или семейства антигенов, причем упомянутая клетка конструируется с учетом продуцирования поли-Ig-стабилизатора. Комплекс продуцируется культурой таких клеток и извлекается из нее.
Желательно, чтобы поли-Ig-антитело представлено собой моноклональное антитело класса IgA. Как это также более подробно рассматривается ниже, предпочтительный стабилизатор может быть охарактеризован посредством ссылки на области, существующие у встречающегося в естественных условиях поли-Ig-рецептора, и предпочтительный стабилизатор включает в себя область I и по крайней мере область IV или V, или же обе области, с областью VI, существенно усеченной /удаленной/. Область I предпочтительно простирается от области V, захватывая последовательность, состоящую примерно из 100-500 аминокислот.
Третий аспект изобретения сводится к рассмотрению рекомбинантного поли-Ig-стабилизатора, включающего в себя поли-Ig-связывающую область поли-Ig-рецептора и преимущественно не содержащего C-концевые трансмембранные и интрацеллюлярные поли-Ig-рецепторные области. К предпочтительным стабилизаторам относятся также стабилизаторы, которые были подвергнуты обсуждению выше при рассмотрении комплекса, отвечающего первым двум аспектам настоящего изобретения.
Четвертый аспект изобретения касается способов тестирования защитной способности поли-Ig-моноклональных антител in vivo посредством введения моноклональных поли-Ig-антител, подвергаемых тестированию, в систему циркуляции млекопитающего и посредством тестирования слизистых выделений млекопитающего на поли-Ig-антитела. Млекопитающее затем подвергают воздействию провокационной пробы с патогенным микроорганизмом /или сперматозоидами/, и определяют защитную способность в отношении патогенного микроорганизма или оплодотворения. Преимущественно моноклональное поли-Ig-антитело вводят в систему кровообращения млекопитающего посредством впрыскивания или вливания. Или же опухоль или опухоливая клетка /т.е. гибридома/ может быть введена подкожно в виде постоянного источника моноклональных поли-Ig-антител.
Пятый аспект изобретения касается способа приготовления описанного выше сульфгидрильного прошитого комплекса посредством использования фиксационно-зависимых эпителиальных клеток, сконструированных для продуцирования поли-Ig-рецептора, посредством культивирования таких эпителиальных клеток в условиях, допускающих образование монослоя клеток на пористой подложке, расположенной таким образом, что оказывается возможным отделение первой среды, находящейся на базальной стороне монослоя, от второй среды, находящейся на апикальной стороне монослоя. Моноклональное IgA-антитело вводят на базальную сторону монослоя, и комплекс образуется в клетке по мере транспортирования антитела по монослою. Комплекс извлекают из среды на апикальной стороне монослоя. Желательно, чтобы первая среда представляла собой питательную среду, пригодную для поддержания роста эпителиальных клеток. Желательно также, чтобы вторая среда была существенно свободной от белков, мешающих извлечению поли-Ig-комплекса. Моноклональные IgA-антитела могут быть введены в первую среду с совместно культивируемыми гибридомными клетками. Или же моноклональное IgA-антитело сначала получают в гибридомной культуре, затем отделяют от культуры и, наконец, вводят в первую среду.
Комплекс, отвечающий первому и второму аспектам изобретения, может быть введен непосредственно в слизистые поверхности млекопитающего с целью защиты от патогенного микроорганизма или от оплодотворения. Желательно, чтобы комплекс вводился орально, назально, вагинально, ректально, через глаз или в среднее ухо. Один предпочтительный способ введения первого млекопитающего, а затем к введению такого слизистого выделения второму млекопитающему. Например, молоко от первого млекопитающего, содержащее комплекс, может быть дано второму млекопитающему. Для генерации выделения поли-Ig-антитело /например, очищенное от гибридомной культуры/ вводят в систему кровообращения первого млекопитающего /например, посредством вливания или впрыскивания/, и молоко или другие слизистые выделения собирают у млекопитающего.
Изобретение обеспечивает возможность проведения особенно эффективной и безопасной пассивной иммунизации, поскольку не затрагивается весь организм. В соответствии с этим иммунологические реактивы животного происхождения /от млекопитающего, не являющегося человеком/ представляются более допустимыми для введения в систему слизистой иммунной защиты, чем для проведения иммунизации всего организма. Неонатальные инфекции, такие как некротический энтероколит и стрептококковые инфекции группы B, являются особенно важными мишенями, что обусловлено отсутствием естественного слизистого иммунитета в этом возрасте.
Шестой аспект изобретения сводится к генерации поли-Ig-продуцирующей гибридомы под воздействием провокационной пробы на млекопитающем с использованием антигена и к извлечению лимфоидных клеток из слизистой ткани млекопитающего, такой как пейровы бляшки, или иной слизистой ткани, богатой лимфоидными клетками. Хотя пейровы бляшки и являются предпочтительными, к другим приемлемым слизистым тканям, богатым лимфоидными клетками, относятся миндалины, аденоиды, аппендикс и ректальная лимфоидная ткань. Лимфоидная клетка сливается с миеломной клеткой. Антиген желательно вводить на слизистую поверхность млекопитающего.
Другие особенности и преимущества изобретения станут очевидными из следующего описания преимущественных вариантов такового.
На фиг. 1 изображено схематическое представление трансцитоза антигена через M-клетки.
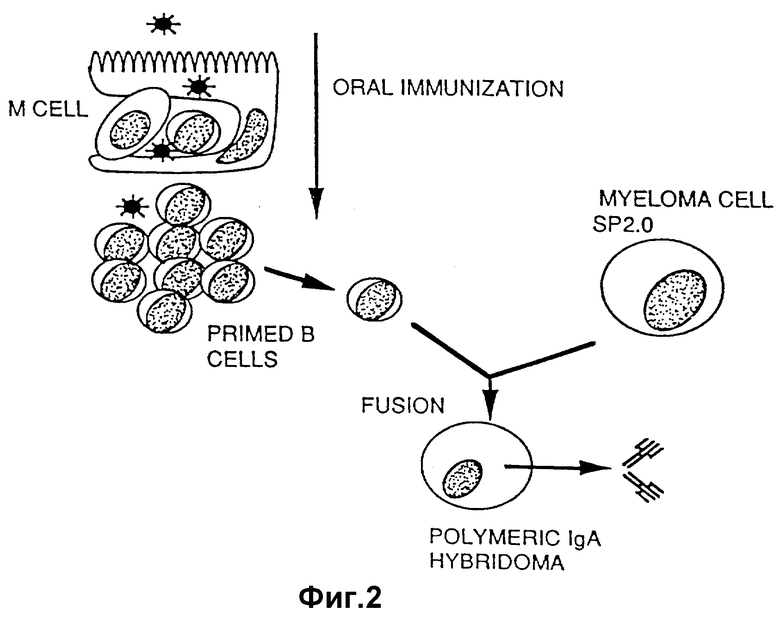
На фиг. 2 изображено схематическое представление этапов образования полимерной IgA-продуцирующей гибридомы.
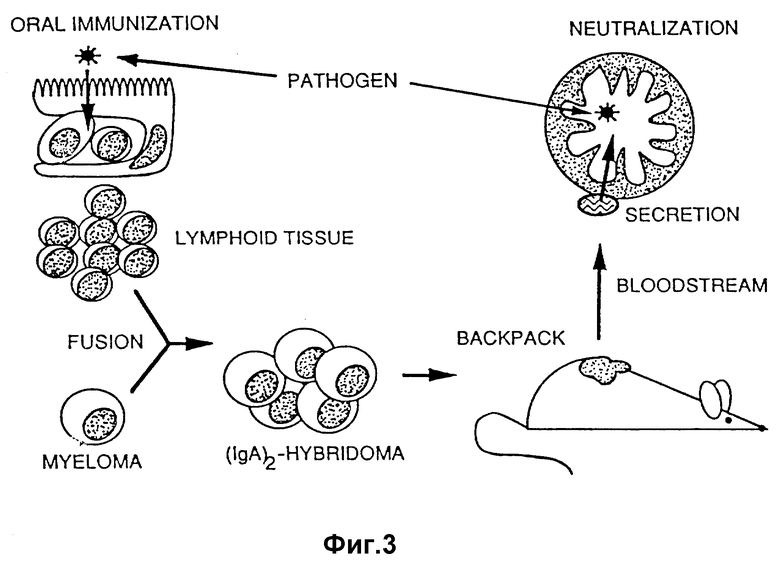
На фиг. 3 изображено схематическое представление способа оценки защитных характеристик полимерного IgA-антитела, продуцируемого гибридомой.
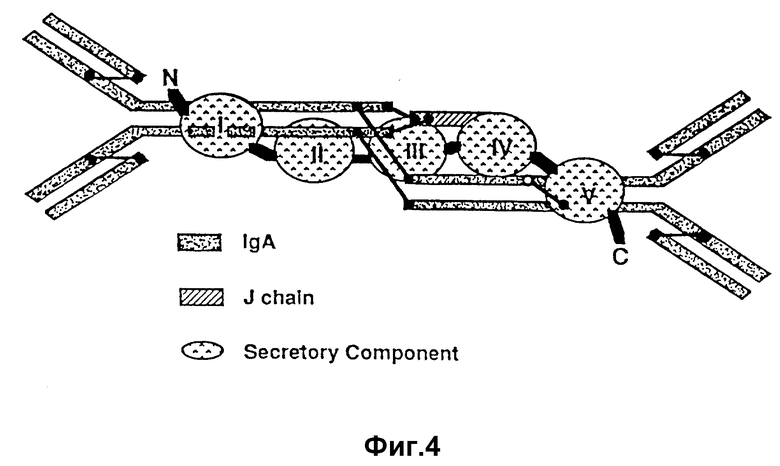
На фиг. 4 изображено схематическое представление структуры комплекса, возникающего между полимерным IgA-антителом и модифицированным поли-Ig-рецептором.
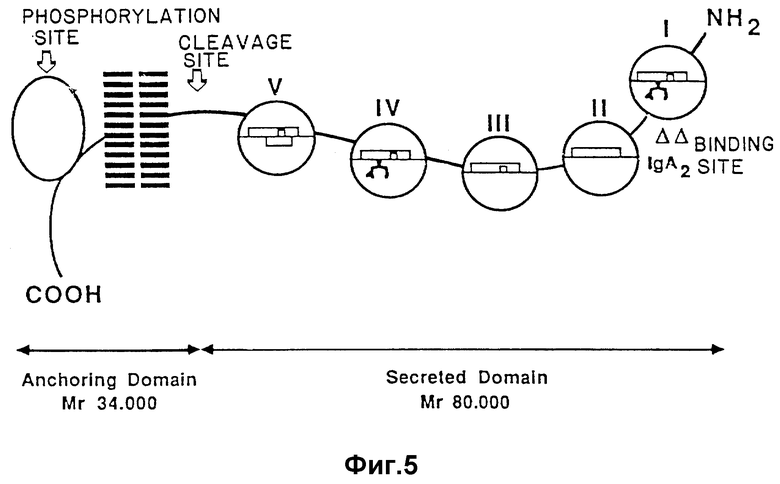
На фиг. 5 изображено схематическое представление областей, существующих у поли-Ig-рецептора.
На фиг. 6A и 6B изображены нуклеотидная последовательность у генетически закодированного поли-Ig-рецептора и выявленная аминокислотная последовательность.
На фиг. 7 изображено выведенная аминокислотная последовательность, установленная по клонированным кДНК двух кроликовых аллелей поли-Ig-рецептора.
На фиг. 8A-8C изображена кДНК, из которой выведена последовательность, показанная на фиг. 7.
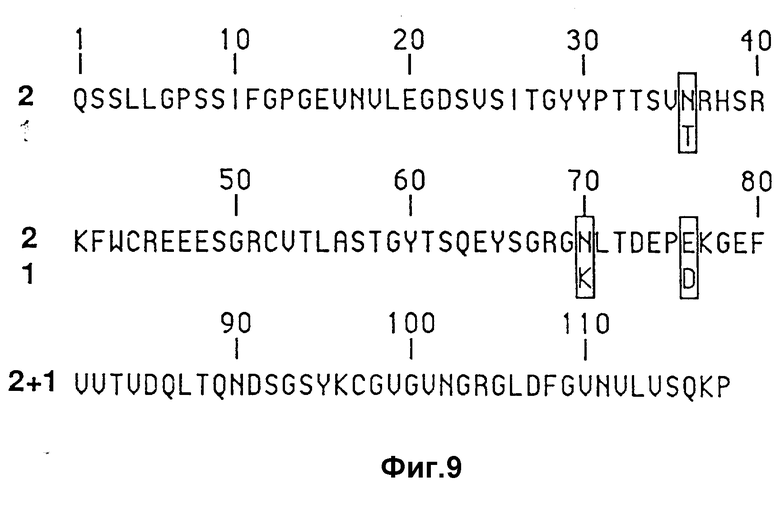
На фиг. 9 изображено сопоставление аминокислотной последовательности области I у поли-Ig-рецепторных аллелей фиг. 7.
На фиг. 10 изображены компоненты различных векторов экспрессии.
На фиг. 11 изображено строение двух специфических векторов экспрессии pLK-нео 6.8 кб и pLM-нео 6.8 кб.
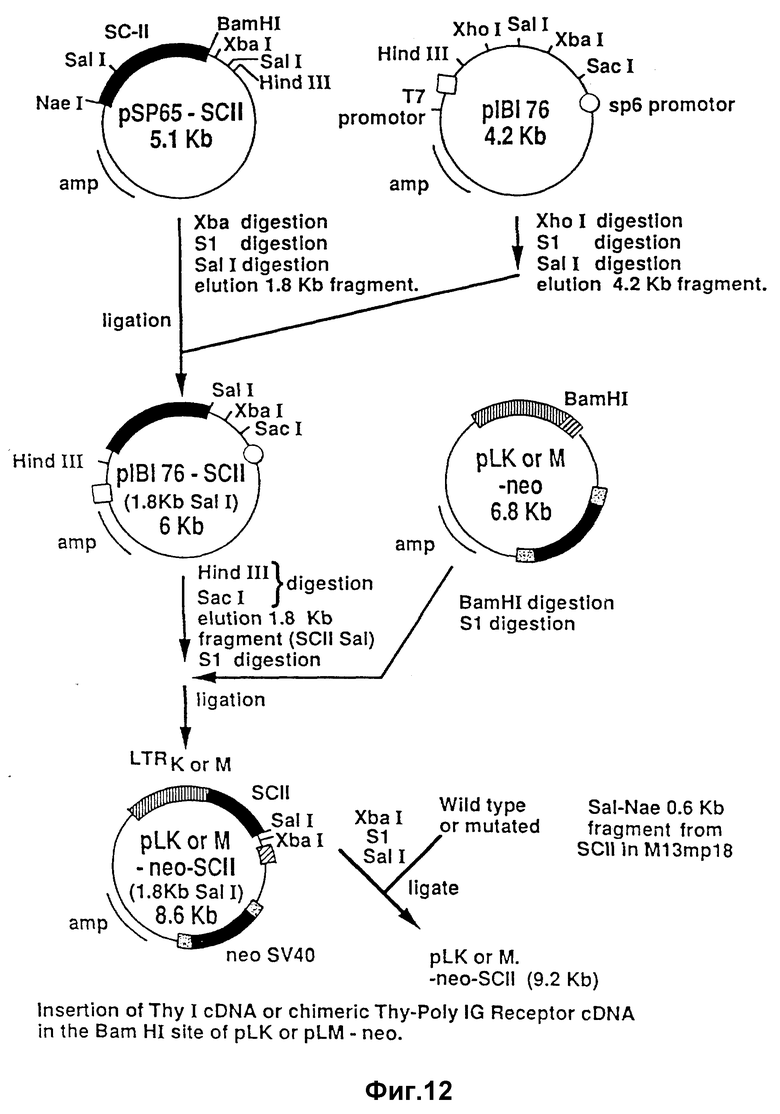
На фиг. 12 изображено введение клонированного поли-Ig-рецепторного гена в векторы, показанные на фиг. 11.
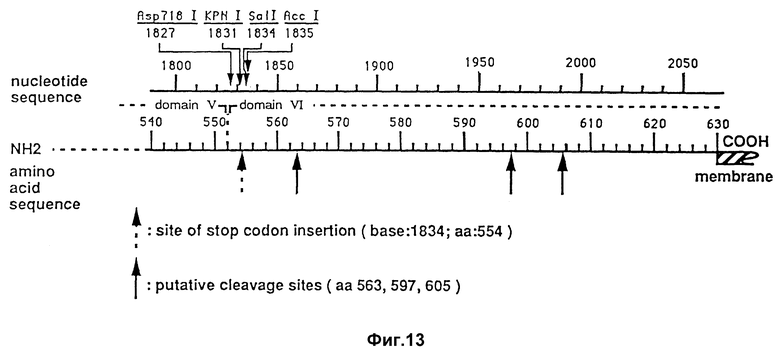
На фиг. 13 изображены различные места ограничения у поли-Ig-рецепторной кДНК.
Примеры.
1. Стабилизаторный белок и поли-Ig-комплекс
Предпочтительный стабилизаторный белок получают из поли-Ig-рецептора, трансмембранной молекулы, посредством которой осуществляется перенос поли-Ig-рецептора. Поли-Ig-рецептор включает следующие области, показанные на фиг. 4 и 5: пять экстрацеллюлярных областей, трансмембранный стягивающий сегмент и C-концевой интрацеллюлярный хвост. При протеолитическом расщеплении рецептора получается свободный пятиобластной белковый "SC", или секреторный компонент. См. литературу: MoS-tov et al., Nature, 308:37-43 /1984/; Kuhnet al. , I. Biol. Chem., 258:6653-6659 /1993/. Между поли-Ig-рецептором и IgA-антителом образуются дисульфидные мостики, которые схематически показаны на фиг. 4.
Поли-Ig-рецептор обычно сохраняется неизменным по млекопитающим видам; и здесь под этот термин подпадают все такие рецепторы независимо от вида млекопитающего, например, мышь, крыса, кролик, морская свинка, свинья, овца, лошадь, корова или человек. Специалистам, работающим в этой области, ясно, что кроликовые поли-Ig-рецепторные последовательности, фигурирующие в этой заявке, могут быть использованы в стандартных гибридизационных клонирующих методиках для идентификации закодированных по кДНК поли-Ig-рецепторов, взятых у других млекопитающих.
Суммируя сказанное выше, можно утверждать, что стабилизаторная белковая часть комплекса может представлять собой рекомбинантный стабилизаторный белок, полученный по крайней мере двумя различными способами. Поли-Ig-рецептор может быть клонирован и выражен в клетке, которая обладает способностью экспортирования его и расщепления его с образованием рекомбинантного стабилизаторного белка. Или же поли-Ig-рецепторный ген может быть сконструирован различными способами /в частности, встраиванием стоп-кодона, как это описано ниже/ с образованием конструкции, которая прямо выражается в виде стабилизаторного белка.
Как отмечали выше, оба пути создания стабилизаторного белка начинаются с использования клонированного поли-Ig-рецепторного гена. Мостов /Moctov/ и др. приводят сведения /Nature, 308:37-43 /1984// о клонировании и последовательности у этого гена. Чертеж 6 взят из указанной статьи, и он более подробно объясняется ниже.
Еще один специфический способ клонирования гена приводится ниже. См. литературу: Schaerer et al., J. Cell. Biol., 110:987-998 /1990/. Цитоплазматическую РНК готовят из молочной железы дающих молоко новозеландских кроликов согласно Сьюарду /Suard и др. /Biochem. J., 201:81-90 /1982//, и поли-A и РНК очищают согласно Шиблеру /Schibler/ и др. /J. Mol. Biol., 142:93-116 /1980/. Поли-A+PHK с усиленной рецепторной трансляционной активностью, достигнутой фракционированием по размерам при линейном сахарозном градиенте в 5-20%, служила в качестве матрицы для синтеза кДНК. Первую кДНК-цепочку синтезировали согласно Вали /Wahli/ и др. /Dev. Biol., 67:371-383 /1978//, а вторую цепочку - согласно ДНК-полимеразному I-РНазному H-методу Гублера /Cubler, Gene, 25:263-269 /1983//. Двухцепочечную и dC-концевую кДНК фракционировали на 1%-ном низкоплавком агарозном геле, и кДНК /кислоты/ с размером в области от 1,5 до 7 кб элюировали и "отжигали" до концевой pBR322, как это описано у Пикока /Peacock/ и др. /Biochem. Biophys. Acta, 655:243-250 /1981//. Клетки DHI подвергали трансформации вместе с "отоженной" ДНК, используя общий метод Ханахэна /Hanahan/ и др. /J. Mol. Biol., 166:577-580 /1983//. Трансформированные бактерии подвергали лизису, и ДНК переносили на нитроцеллозные фильтры /Thomas, Methods Enzymol. , 100:255-266, 1983/ и подвергали испытанию, воздействуя двумя синтетическими олигонуклеотидами. Специфическими зондирующими веществами, использованными в этих испытаниях, были нуклеотиды 186-206 и 2421-2437, взятые из последовательности, которую привели в литературе Мостов /Mostov/ и др. /Nature, 308:37-43 /1984/ и которая была помечена Т4-полинуклеотидной киназой и α P32-АТФ, как это описано в Гофа /Gough, N/ и др. /Nature, 309:763-767 /1984//. Клоны кДНК, гибридирующиеся с обоими зондирующими веществами, далее характеризовали, проводя гибридную селективную трансляцию /Reizman et al. , EMBO J. 2:2161-2168, 1983/. Две образовавшиеся плазмиды представляли собой плазмиду PSC-1, которая соответствовала низкомолекулярной форме поли-Ig-рецептора с отсутствующими областями 2 и 3, как это описано у Дойчера /Deitcher/ и др. /Mol. Cell Biol., 6:2712-2715 /1986//, и плазмиду pSC-11 с закодированной высокомолекулярной формой.
На фиг. 6A и 6B показаны последовательность у поли-Ig-рецепторной mPHK и аминокислотная последовательность, вытекающие из сказанного выше, как это приводят в литературе Мостов /Mostov/ и др. Они описывают фиг. 6A и 6B следующим образом. Остатки нумеруются числами от 5' до 3', начиная с первого нуклеотида после поли-dG-хвоста, расположенного в точках 21, 53. Нуклеотидные номера указываются на правом поле. Полная последовательность насчитывает 3517 оснований и содержит необычную поли-A-добавочную последовательность AUUAAA, начинающуюся с 26 остатков при следовании вверх от поли-A-хвоста и указанную треугольниками, стоящими под каждым остатком. Предсказанная аминокислотная последовательность показана над нуклеотидной последовательностью. Сигнальный белок нумеруется числами от -18 до -1. Первая аминокислота у созревшего поли-Ig-рецептора /GIn/глицин// нумеруется числом 1. Два потенциальных места для аспарагинсвязанного гликозилирования отмечены чертой, проведенной сверху. Предсказанная мембранностягивающая область отмечена пунктирной чертой, проведенной сверху. Цистеиновые пары для пяти областей, а также соответствующий первый цистеин для шестой области указываются зачерченными кружками, расположенными над первым цистеином каждой области, и незачерченными кружками, расположенными над вторым цистеином каждой области.
Обладая клонированным поли-Ig-рецепторным геном, им можно воспользоваться для непосредственной экспрессии требуемого стабилизаторного белка. В частности, рекомбинантные белки, содержащие некоторые поли-Ig-связывающие области у секреторного компонента, будут оказывать стабилизирующее воздействие на полимерный иммуноглобулин в отношении протеолитического распада без проявления существенного вредного эффекта в отношении специфического связывания и защитных функций полимерного IgA-антитела. В частности, области, показанные на фиг. 4 и 5, представляют собой главную поли-Ig-связывающую область I, две промежуточные главные области II и III и C-концевые области IV и V, которые способствуют поли-Ig-стабилизации.
Поли-Ig-рецептор усекается на C-границе с целью удаления трансмембранной и интрацеллюлярной частей белка, которые не являются существенными в отношении стабилизации или защиты. Усеченной поли-Ig-рецептор, пригодный для использования в настоящем изобретении, следовательно, содержит /как минимум/ N-граничную поли-Ig-связывающую область I.
Проведенное ниже обсуждение двух специфических усеченных поли-Ig-рецепторов делается с целью иллюстрации изобретения, а не для ограничения его. Специалистам, работающим в этой области, совершенно ясно, что специфические белковые последовательности, приведенные ниже, могут быть модифицированы с осуществлением обычных замещений и исключений при сохранении функционального назначения и характера изобретения.
На фиг. 7 изображена аминокислотная последовательность, отвечающая клонированным кДНК двух кроликовых аллелей поли-Ig-рецептора. Одна аллель обозначается "Ho. I" и соответствует кДНК, которую в литературе Мостов /Mostov/ с сотр. /Nature, 308:37-43 /1984//, что показано на фиг. 6A и 6B. Другая аллель была клонирована в лаборатории заявителя Краехенбулом /Kraehenbuhl/ по способу, о котором говорили выше, и последовательность у такой аллели обозначается "Ho.2" на фиг. 7. На фиг. 7 знак деления /":"/ используется для обозначения идентичных остатков; знак в виде точки /"."/ используется для обозначения схожих остатков, т.е. A, S, T; D, E; N, Q; R, K; I, L, M, V; F, V.
На фиг. 8A-8C изображены кДНК-последовательности, отвечающие таким клонам. Опять-таки, двоеточие характеризует идентичные основания. Буква "N" относится к неизвестному основанию.
Имеют место следующие семь областей в пределах поли-Ig-рецепторной последовательности /числа относятся к фиг. 6/.
Область - Сегмент
I - 10-118
II - 119-223
III - 224-332
IV - 333-441
V - 442-552
VI - 553-652
VII - 653-755
Область I представляет собой поли-Ig-связывающую область, которая остается у усеченного поли-Ig-рецептора, отвечающего настоящему изобретению.
Специфическая последовательность у области I в случае обоих аллелей /включающих аминокислоты 1-9, которые не играют существенной роли в случае поли-Ig-связывания и, следовательно, не носят критического характера в отношении настоящего изобретения/ показаны на фиг. 9.
Поли-Ig-рецепторная последовательность может быть модифицирована посредством проведения консервативных аминокислотных замещений, исключений и добавлений, которые не изменяют характер поли-Ig-связывания. В самом деле, как видно из фиг. 9, из сопоставления двух последовательностей, отвечающих области I, видно, что имеют место только три аминокислотных изменения в положениях 38, 70 и 76. В каждой из других областей, рассматриваемых в настоящем изобретения, имеют место от двух до шести аминокислотных изменений, причем в области IV изменения приходятся на остатки 337 и 358, а в области V - на остатки 448, 460 и 513.
В сумме, гомология на аминокислотном уровне определенно превышает 80/, и последовательности могут быть модифицированы во всех трех областях посредством проведения аминокислотных замещений или исключением областей.
В частности, применительно к поли-Ig-ассоциации достаточная гомология должна сохраняться в области I, надлежащей интервал /например в 100-500 аминокислот/ должен удерживаться между областями I и V, и область V должна быть пригодной для сохранения ковалентной дисульфидной стабилизации. Таким образом, очень значительное модифицирование может быть допустимым в областях II-IV.
Очень желательно, чтобы усеченный поли-Ig-рецептор включал также область V, которая в силу существования сульфгидрильной связи является ответственной за ковалентную стабилизацию комплекса. См., в общем, литературу /Eiffert et al. , Hoppe Seyler's Z. Phys. Chem., 365:1489-1495 /1984//, где идентифицируются цистеиновые остатки у поли-IgA-антитела /человеческого/, участвующие в образовании дисульфидных мостиков. Важно, кроме того, чтобы интервал, соответствующий областям II-V и определяющий положение области V в отношении образования дисульфидных мостиков, задавался таким, каким он показан на фиг. 5, и чтобы происходило усиление защитного действия в отношении протеолиза.
Для установления надлежащего положения места усечения мембранно-стягивающий сегмент в области VI полного рецептора может быть идентифицирован как промежуточное образование из неизменных преимущественно гидрофобных остатков /например, 630-652 на фиг. 6/ с предшествующим заряженным остатком /например, лизин 629/ и последующей крайне основной последовательностью /например, аргинин-аланин-аргинин-гистидин-аргинин на фиг. 6/. Эту мембранную растягивающую область, а также C-концевой цитоплазматический хвост молекулы исключают усечением.
В случае одного специфического варианта усечение достигается синтезом 16-членного олигонуклеотида ACTAGTOCTAGACTAG со стоп-кодоном TAG во всех трех считываемых кодах. Оликонуклеотид представляет собой полиндром и самогибридизируется. Результирующие сдвоенное образование вводили в описанную выше поли-Ig-рецепторную кДНК /ДНК положение 1834, аминокислота 554/.
Говоря точнее, четыре аминокислоты /T-S-L-D/, закодированные олигомером, добавляли к C-границе области V, в результате чего усечение в точности происходило в месте перехода от области V к области VI. Ограничительный эндонуклеазный участок, выбираемый для вставки олигомера, представляет собой Sall-участок, который является единственным в своем роде и который располагается вблизи предполагаемых участков естественного разрыва /аминокислоты 563, 597 и 605/. Другие удобные участки введения вставки в области V приходятся на Asp /1827/, KpnI /1831/ и ACCl /1835/. В области VI отсутствуют удобные уникальные участки (фиг. 13.)
Описанные выше клонированная поли-Ig-рецепторная кДНК или сконструированная /например, усеченная/ поли-Ig-рецепторная кДНК могут быть в общем случае выражены в какой-либо из широкого разнообразия систем экспрессии, которые хорошо известны специалистам, работающим в этой области. Например, желательные системы экспрессии относятся к специализированным секреторным клеткам /т.е., клеткам, которые естественным образом секретируют большие количества белка/, таким как фибробластовые клетки, включая системы экспрессии Чайниз хамстер оувари /Chinese Hamster Ovary/. Могут быть использованы системы экспрессии эндокринных опухолевых клеток, такие как ATt 20 /ATCC / Американская коллекция типов культур/, циркуляры CRL89 и CRLI795/ или PC-12. Клетки Мейдин-Дарби кейнин кидней /Madin-Darby Canine Kidney /MDCK// являются также желательными клетками хозяина. См. работу Мостова /Moctov/ и др. /1984/, цитированную выше. Выбор носителя эксперссии будет зависеть от клетки хозяина, которая была выбрана.
В случае одного из вариантов усеченный поли-Ig-рецептор может быть экспрессирован в обессмерченных IgA-продуцирующих клетках, таких как образования лимфоцит-миелома. В таком случае предпочтительным носителем экспрессии является носитель, который включает в себя сильный или способный индуцироваться промотор, элемент-усилитель /например, B-лифоцитный специфический усилитель, такой как μ- усилитель или усилитель вирусного начала/, сконструированный стоп-кодон, введенный в поли-Ig-рецепторную кДНК для устранения необходимости протеолитического отделения областей I-V /секреторный компонент/ от клеточного мембранного фиксатора /область VI-VII/. Ниже описаны специфические системы экспрессии.
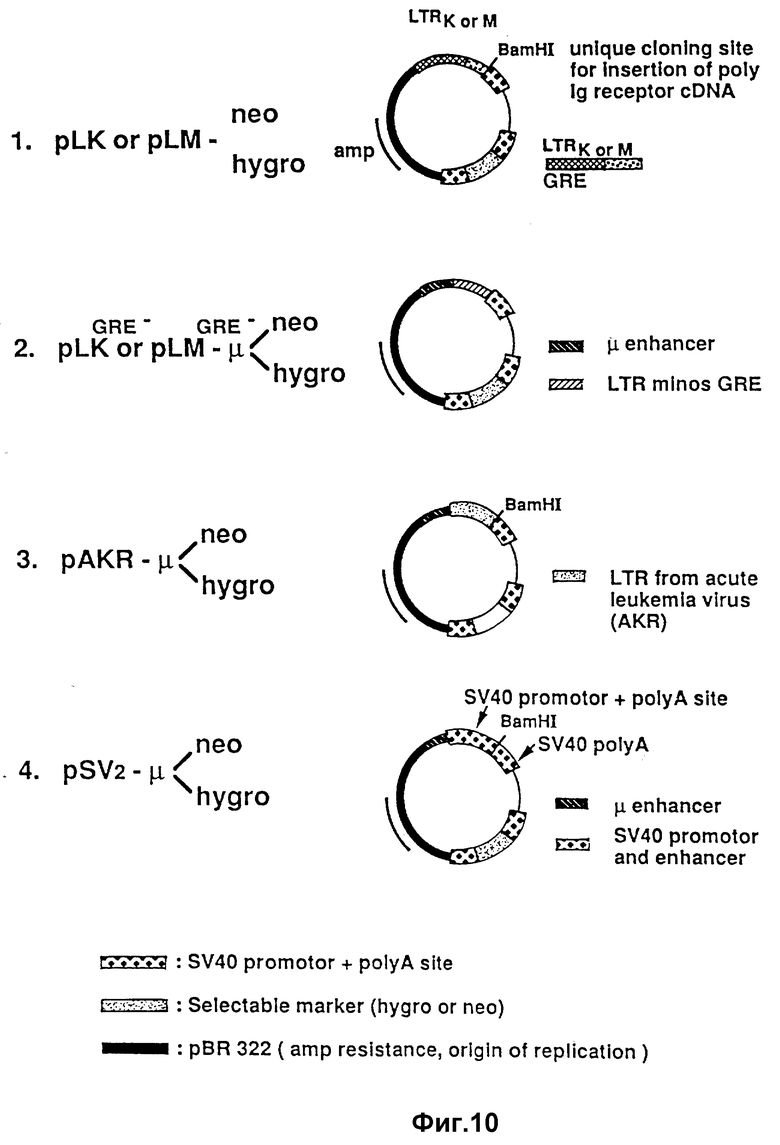
Как это показано в общем виде на фиг. 10, носитель экспрессии может представлять собой вектор, выраженный, к примеру, через pLK-нео или гидро или pLM-нео или гигро с глюкокортикоидными реактивными элементами или через те же векторы, у которых глюкокортикоидные реактивные элементы удалены и заменены на μ- элемент-усилитель, т.е. pAKR-μ-нео и pSV 40-μ-нео. Вектор может содержать следующие элементы.
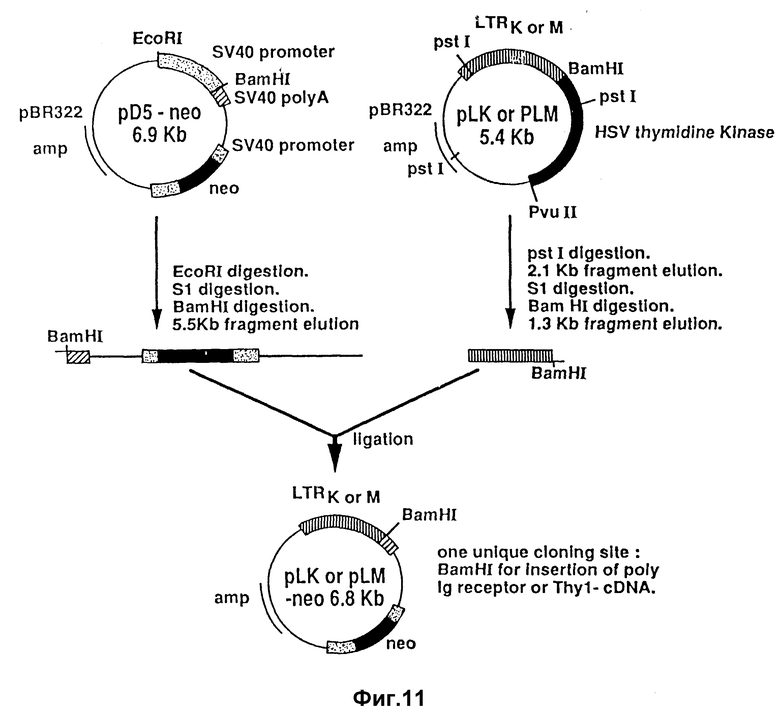
1. Надлежащий промотор для инициирования транскрипции кДНК. LTR-образования, взятые из ретровирусов, являются приемлемыми, включая LTR-образования, выделенные из почечной аденокарциномы, как это описано у Уиллиджера /Williger/ и др. /I. Virol., 60:1-11 /1986/. Почечные LTR-образования являются гомологически подобными LTR-образованиям, выделенным из опухолей молочных желез с некоторыми различиями в U3-области. Оба LTR-образования содержат глюкокортикоидный элемент, с которым связано гормонное стимулирование транскрипции. Например, глюкокортикоид /дексаметазон, фирма "Сигма корпорейшн, г. Сент-Луис, шт. Миссури/ повышает транскрипцию усеченного поли-Ig-рецептора в 10-50 раз. LTR-образование из почки или молочной железы, выделенное из клонирующего вектора /pLK или pLM предоставлял д-р Хейди Диггельманн /Heidi Diggelmann/, Швейцарский институт раковых исследований, в группе которого была сконструирована плазмида; см. работу Уиллиджера /Williger/, и др. , цитированную выше/ последовательным получением pstl, SI и BamH при ферментации, вводили в pSV2-нео-вектор/ промышленно доступный через Биолаборатории/, который был последовательно ферментирован заранее с протеканием EcoRI-, SI- и BamHI-ферментации /для удаления SV40-промотора и замены его на LTR-промотор/ /фиг. 6B/. Вектор pLK-гигро или pLM-гигро конструировали введением гигромицин-резистентного гена /IB/ вместо неомицин-резистентного гена. Плазмида pLKнеоPIRII была внесена в банк Американской коллекции типов культур /АТСС/, г. Роквилл, шт. Мэриленд, под инвентарным номером АТСС 40787. Такая плазмида представляет собой pLEнео-плазмиду, показанную на фиг. 10, с усеченной полимерной иммуноглобулиновой рецепторно-закодированной ДНК /фиг. 8/, введенной в BamHI-участок-плазмиды.
2. Элементы-усилители для усиления транскрипции и, следовательно, продуцирования усеченного поли-Ig-рецептора. Такие элементы включают в себя вирусные элементы /SV40/, глюкокортикоидные реактивные элементы из MMTV/ или специфические усилители из ткани молочной железы, такие как μ- усилитель, ответственный за высокую экспрессию иммуноглобулина в B-клетках. Усилитель μ- типа, извлеченный из мышинного геномного клона, заключающего μ- локус тяжелой цепи /Mercola, M., Wang, X.F., Olsen, Y., and Calame, K., 1983, Science, 221:663-665/, может быть вставлен за различными промоторами.
3. Векторная основа, включающая выбираемые сигнальные гены /например, неомициновый или гигромициновый резистентный ген/, место начала репликации у E.coli с антибиотиковым резистентным геном.
4. Поли-Ig-рецепторная кодирующая кДНК.
5. Синтетический олигомер, содержащий стоп-кодоны в каждой считываемой группе, например, ACTAGTCTAGACTAGT.
Дополнительные подробности по конструированию таких векторов приводятся на фиг. 11. Кислоту кДНК для pSC-I вводили в вектор экспрессии pSV-2 так, как это описано у Садерна и Берга /Southern and Berg, J. Mol. Appl. Gen., 1: 327-341 /1982//. Эта конструкция обозначается как pSV2-SCl.
Результирующий вектор трансфектировали в миеломных клетках, и клетки выделяли для установления наличия эффективной экспрессии у усеченного поли-Ig-рецептора. Для проведения трансфекции использовали стандартные методики электропереноса.
В частности, клетки собирали из ростовой среды /2•106 на образец/ и осаждали при вращении /5•103 об/мин/. Клетки должны выходить из культур в состоянии слипания на 30-70%, и среда должна претерпеть изменения в предшествующие 24 ч. При низкой температуре /ледяная баня/ клетки промывали электропереносящей средой /2 мМ раствор RPMI в глутамине и 0,5%-ная сыворотка стельной коровы с Pen Strp®/. Клетки подсчитывали и вновь суспендировали в электропереносящей среде при содержании 2•106 на 0,75 мл. Примерно 45-50 мкл линеаризованной ДНК /10 мкг линеаризованной кислоты, суспендированной в 45 мкл фосфатно-буферного солевого раствора/ добавляли к аликвоте /0,75 мл/ клеточной суспензии, находящейся в кювете для электропереноса. Кювету ставили на лед на 15 мин, и смесь подвергали пульсирующему воздействию. Клетки затем помещали в подготовленную среду RPMI или среду Игла /Eagle/ и отбирали.
Подвергнутые трансфекции миеломные клетки экспрессируют и выделяют усеченный поли-Ig-рецептор. В противоположность сказанному, миеломные клетки не обладают способностью отщеплять и выделять полный рецептор. Усеченный поли-Ig-рецептор выделяли и очищали от надосадочной жидкости с миеломной культурой, проводя осаждение сульфатом аммония и гелиевое фильтрование.
В одном особенно предпочтительном варианте, описанном более подробно ниже, усеченный поли-Ig-рецептор получали в гибридоме, которая была выбрана для получения желаемого поли-Ig-антитела.
В такой системе ковалентные дисульфидные связи, связывающие усеченный рецептор с поли-Ig-антителом, образуются в живых клетках, и секретируется результирующий стабильный секреторный Ig-комплекс. Эта система может быть сконструирована либо путем трансфекции миеломных клеток, которые будут использованы в качестве партнеров при слиянии с обессмерченными поли-Ig-продуцирующими B-клетками, либо путем трансфекции гибридом, которые уже образованы и экранированы с целью получения требуемого полииммуноглобулина. Результирующая ковалентная дисульфидная связь образуется в любом случае.
Трансфекцию миеломных клеток SP-20 или p3x63/Ag 8U.I /ATCC /Американская коллекция типов культур/, циркуляры CRL 1581, CRL8287, CRL597/, проводили, используя вектор экспрессии pLK, в котором поли-Ig-рецепторная кДНК-транскрипция происходит под управлением почечного MMTV LTP-промотора. См. литературу: Willigr et al., I. Virology, 60:1-11 /1986/. Эффективная транскрипция получается при стимулировании трансфектированных клеток диксаметазоном /10-6М/ в течение, по крайней мере, 12 ч.
Еще одна система, пригодная для получения стабилизаторного белка, сводится к трансфекции носителя экспрессии для экспрессии полного поли-Ig-рецептора в эпителиальной фиксационно-зависимой клеточной линии, отвечающей млекопитающему, которая образует монослой на пористом субстрате. Одной такой примечательной клеточной линией является MDCK-клеточная линия, трансформированная с вектором для экспрессии поли-Ig-рецептора, как это описано выше. В частности, может быть использован pLK-нео-ScII-вектор, содержащий глюкокортикоидное LTRK-образование. Могут быть использованы и другие подходящие эпителиальные клетки хозяина, образующие монослой. Известен гигромициновый резистентный ген, который описали Блоечлинер /Bloechliner/ и Диггельманн /Diggelmann/ /Mol. Cell. Biol., 4:2929-2931 /1984//.
В общем, трансфектированную клеточную линию выращивают с образованием монослоя на пористом субстрате, таком как фильтр /например, пропускающие ячеистые фильтры, 0,45 мкМ, 4,7 см2/, покрытый коллагеном. См. работу Стила /Steele/ и др. /Am. Y. Physiol., 251:C136-C139 /1986//. Клетки будут становиться асимметричными с проявлением базальной и апикальной мембран. Мембрана и монослой располагаются таким образом, чтобы происходило разделение сосуда на области с базальной средой и апикальной средой. Поскольку клетки характеризуются тенденцией получать питание из базальной среды, эта среда должна содержать питательные вещества, необходимые для поддержания роста клеток. Требование к наличию питательных веществ в апикальной среде является менее строгим. Клетки с успехом будут использовать рекомбинантный поли-Ig-рецептор, забирая его из базальной среды и перенося в апикальную мембрану, где рецептор претерпевает ферментативное отщепление с образованием стабилизированного поли-Ig-антитела, ковалентно связанного со стабилизирующим белком. Поли-Ig-A-антитело поступает в базальную среду при культивировании IgA-продуцирующей гибридомы в такой среде. Или же в среду может быть добавлен поли-Ig-рецептор, извлеченный /проведением, например, очистки на основе сродства/ из выделенной гибридомной культуры.
Стабилизированное поли-IgA-антитело может быть извлечено из апикальной среды опять-таки посредством очистки на основе сродства или же посредством использования иных стандартных методов выделения антител. Для упрощения очистки апикальная среда может поддерживаться в состоянии, при котором она является существенно свободной от белковых загрязнений.
3. Конструирование и экранирование
IgA-продуцируюих гибридом.
Еще один аспект настоящего изобретения сводится к возможности легко образовывать поли-Ig-антитела и производить экранировку в отношении поли-Ig-антител /особенно поли-IgА-антител/, обеспечивая защиту от выбранных инфекций, особенно инфекций, обусловленных попаданием вирусных и бактериальных патогенных микроорганизмов в организм через слизистые поверхности. Этот аспект изобретения включает также возможность образовывать поли-Ig-антитела, направленные против поверхностных компонентов сперматозоидов, и экранировать антитела, обеспечивающие защиту от оплодотворения. Кроме того, обеспечивается возможность образования и экранирования поли-Ig-антител, направленных против пищевых и находящихся в воздухе аллергенов.
В плане этого аспекта изобретения у животного-хозяина создается управляемый источник поступления поли-Ig-антител; например, поли-Ig-сектерирующие гибридомы имплантируются животному-хозяину, где они развиваются в опухоль. Поли-Ig-антитела, продуцируемые и высвобождаемые такой опухолью, поступают в систему кровообращения, и циркулирующие димерные моноклональные IgA-антитела поступают из пропускающих капилляров в слизистую ткань, где они находятся и связываются эпителиальными рецепторами и транспортируются в выделении вместе с эндогенным секреторным компонентом. Перитонеальные /асцитные/ опухоли из гибридомных клеток могут /что является известным/ вызывать нежелательные патологические изменения в кишечнике и создавать аномально высокие уровни несекреторных IgA-антител в перитонеальной жидкости кишечника. Согласно сказанному, предпочтительным способом введения IgA-секретирующих гибридомных опухолей является подкожный с введением, например, в верхнюю часть спины.
IgA-продуцирующая гибридома создается посредством обессмерчивания антиген-сенсибилизированного B-лимфоцита, например, извлекаемого из пейеровых бляшек после проведения оральной или слизистой иммунизации. См. литературу: Weltzin et al., I. Cell Biol., 108: 1673-1685 /1989/. Миеломные клетки, используемые для обессмерчивания IgA-продуцирующей клетки, являются широко доступными. Могут быть использованы стандартные способы слияния.
В частности, гибридомы генерируют и тестируют на специфичность IgA-продуцирования в соответствии со следующими специфическими примерами.
Могут быть использованы различные стандартные методы введения провокационной пробы животным с целью продуцирования антиген-сенсибилизированный B-клеток, необходимых для конструирования гибридомы. См. статью Велцина /Weltrin/ и др. , цитированную выше. Один отличительный способ сводится к использованию микрочастиц гидроксиапатита в качестве носителя орально введенного антигена. Подробная методика приготовления сложных образований из антигена и гидроксиапатита, пригодная для проведения слизистой иммунизации, раскрыта в совместной патентной заявке, поданной Амеронгеном /Amerongen/, Ньютра /Neutra/, Велцином /Weltzin/ и Краегенбулем /Kraehenbuhl/ одновременно с настоящей заявкой под названием "Сложные образования из гидроксиапатита и антигена и способы генерации поли-Ig-иммунной реакции". Такая заявка учитывается здесь посредством ссылки.
Микрокристаллы гидроксиапатита, пригодные для транспортирования через эпителий /например, в целом менее примерно 0,1 мкм/, покрывали до насыщения антигеном и смешивали с подходящим носителем пригодным для введения в слизистую оболочку /например, с солевым раствором/. Для оральной иммунизации мыши весом 25 г небольшое количество /eNgN, 30 мкг/ белка, находящегося в 200 мкл фосфатно-солевого буферного раствора, смешивали с надлежащим количеством /например, 1 мг гидроксиапатита на 30 мкг белка /предварительно обработанного ультразвуком и промытого гидроксиапатита. Смесь перемешивали в течение одного часа при 4oC, и гидроксиапатит затем осаждали вращением в течение 2 мин со скоростью 10 000 об/мин в микроцентрифуге. Осадок от центрифугирования промывали и перемешивали или подвергали воздействию ультразвука, проводя новое суспендирование в 200 мкл воды, содержащей 061 М /или менее бикарбоната натрия NaHCO3 /необходимого для нейтрализации желудочной кислоты/. Раствор для введения, предназначенный для оральной иммунизации, может быть приготовлен так, что исключаются условия высокого солевого содержания, при которых имеет место тенденция к высвобождению белка из гидроксиапатита.
Другие приемлемые способы введения провокационной пробы млекопитающему для инициирования слизистой иммунной реакции известны специалистам, работающим в этой области, и они являются приемлемыми для использования в настоящем изобретении.
Один специфический способ продуцирования IgA-секретирующих гибридом сводится к тому, что мононуклеарные клетки из пейеровых бляшек выделяют у животного, подвергнутого воздействию провокационной пробы. В частности, слизистую оболочку и подслизистую основу с 8-10 пейеровых бляшек удаляют и инкубируют в содержащей все компоненты RMPI-среде, содержащей 0,1% коллагеназы, что делают в течение одного часа при 37oC. Ткани истирают между концами двух стерильных микроскопных предметных стекол, концы которых заморожены, и высвобожденные клетки промывают два раза полной RPMI-средой. Из одной мыши извлекают примерно /2-3/•107 жизнеспособных пейеровобляшечных мононуклеарных лейкоцитов.
Миеломные клетки и мышиные мононуклеарные клетки промывали дважды свободной от сыворотки RMPI-средой и сращивали в 37%-ном полиэтиленгликоле 4000 /фирма "Э. Мерк", г. Дармштадт, ФРГ/ при отношении числа лейкоцитов к числу миеломных клеток, равном 5:1. После слияния клетки разбавляли до концентрации 1•106 клеток на 1 мл в полной RPMI-среде и распределяли по пластинкам на 96 ячеек для выращивания тканевых культур, которые за 24 ч до этого были засеяны облученными /800 рад/ мышинными перитонеальными экссудатными клетками с числом 2•104 на ячейку. Гибридомы выбирали в гипоксантиновой, аминоптериновой и тимидиновой среде, используя стандартные методы.
Гибридомы, продуцирующие IgA-антитела, действие которых направлено против требуемых антигенов или клеток, идентифицировали, используя стандартные методы тестирования.
Результирующие гибридомные клетки выращивали, например, в RMPI-среде с культурой 1640, осаждали на центрифуге, вновь суспендировали в фосфатно-буферном солевом растворе и подсчитывали. Плотность клеток доводили до 1•106 клеток на 0,5 мл. Суспендированные клетки /1•106 в 0,1-0,5 мл/ впрыскивали подкожно в верхнюю область спины BALB-C-мышей. По прошествии от одной до трех недель после инъекции появлялись твердые узелки. Когда узелки достигали по оценкам диаметра 3-5 мм брали сывороточные образцы из хвостового кровотока. Для измерения содержания сывороточных IgA-антител использовали устройство ELISA. Секреторные IgA-антитела той же направленности, что и у имплантированной гибридомы, измеряли в кишечных промывках и в молоке, также используя устройство ELISA. Другие слизистые выделения /например, из эпителия и желез оральной/назальной полости, дыхательной и половой систем/ также преимущественно содержали секреторные IgA-антитела.
Этих содержащих опухоль мышей затем использовали в "провокационных" опытах, в которых рассматриваемый вирусный или бактериальный патогенный микроорганизм вводили орально или на иные слизистые поверхности или в которых сперму или сперматозоиды вводили вагинально. Гибридомы, секретирующие полезные IgA-антитела, идентифицировали по мышам, обнаруживающим защитную реакцию в отношении провокационной пробы. Такие тесты могут быть специально использованы для идентификации IgA-антител, наиболее приемлемых для пассивной защиты или контрацепции людей и других видом /см. ниже/. Например, при идентификации приемлемых анти-V. cholerae-секретирующих гибридом новорожденных мышей подвергают имплантированию в день рождения, чем обеспечивается защита до того момента, когда у новорожденных появится резистентность в отношении холеры.
Описанный выше тест может быть использован в опытах по защите новорожденных, в случае которых гибридомы имплантируют беременным BALB/C-мышам во время первой или второй недели 3-недельного периода беременности. Опухолевый рост происходит во время беременности, и циркулирующие димерные IgA-антитела, идущие от опухоли, в достаточном количестве переносятся эпителиальными клетками молочных желез и высвобождаются в виде секреторных IgA-антител, попадая в молозиво и в молоко. Новорожденные получают секреторные IgA-антитела от момента рождения. При желательности введения IgA-антител в течение длительного периода могут быть использованы многократно лактирующие семьи.
Установив путем проведения описанного выше теста in vivo, что моноклональные IgA-антитела могут выступать в роли защитных антител, их тестируют на способность оказывать защитное действие после прямого или пассивного введения в слизистые поверхности. Применительно к этим тестам поли-Ig-антитела со стабилизаторным белком или без него могут быть получены в культуре /как это описано выше/ и введены в слизистые поверхности систем организма, перечисленных ранее.
Особенно важной чертой этого аспекта изобретения является возможность сохранения дееспособности IgA-антител вне организма реципиента на слизистых поверхностях. Так, IgA-антитела не обязательно должны быть иммунологически совместимыми с системой реципиента; им необходимо лишь достичь слизистых поверхностей реципиента и эффективно блокировать поступление патогенных микроорганизмов.
Настоящее изобретение является особенно полезным в плане защиты от Pseudomonas aeruginosa, Hemophilus inluenrae. Vibrio cholerae, Bordetella pertussis, Corynebacterium diphtheriae, Escherichia coli, Salmonella typnhi и typhimurium Clostridium perfringens и других брюшных clostridiae, Shigella dysenteriae, Shigella flexnerii, Neisseria gonorrheae, Trichomonas, Entameba nistolytica, Giardia lamblia, streptococcus, респираторно-синцитиального вируса, ротавируса, реовируса, вируса человеческого иммунодефицита, человеческого Т-клеточного лимфотропного вируса типов I и II, полиовируса, риновируса, вируса гриппа, вирусов герпеса, вируса бородавки человека, патогенных микроорганизмов СПИД 2o, таких как Pneumocystis, и дрожжевых культур, таких как кандиозы. Изобретение является также полезным в отношении защиты от аллергенов, которые контактируют с поверхностями слизистой оболочки дыхательной или пищеварительной системы. Оно является также полезным в плане защиты от беременности посредством связывания сперматозоидов во влагалище и предотвращения их продвижения через шейку и матку.
В каждом случае используют надлежащий известный антиген, например, весь патогенный микроорганизм или специфическую внешнюю часть антигенов, такую как вирусный оболочечный белок или бактериальные поверхностно-клеточные белки, белок фимбрия, липополисахариды, вирусный капсид или белок оболочки, поверхностный компонент протозойной клеточной мембраны, сперматозоидные поверхностные белки или аллергены дыхательной системы. Могут быть также использованы токсоиды, например, CRM-197, инактивированный токсин diphtheria, как это следует из работы Утида /Uchida/ и др. /J. Biol. Chem., 248: 3833-3844 /1973//. Антиген используют по описанной выше методике, генерируя гибридомы, выделяющие требуемые защитные антитела. Приводя всего лишь несколько примеров, можно сослаться на патент W088/08437 /включенный сюда со ссылкой/, в котором раскрывается tcPA-фимбриевый белок, пригодный для образования анти-V. cholerae моноклональных поли-Ig-антител. В патенте США N 4725669 раскрывается HIV/HTLV-III/-оболочечный гликопротеид, пригодный для образования анти-HIV-поли-Ig-моноклональных антител. В следующих патентах и патентных заявках раскрывается получение антител для защиты от стрептококковых инфекций и особенно инфекций группы B streptococaus: патенты США с номерами 4367221, 4367222, 4367223, 4356263, 4207414 /RE 31672/ и WO 87/06267.
Одно приемлемое IgA-моноклональное антитело, направленное против V. cholerae, липополисахарид выделяется гибридомой 2D6, внесенной в банк Американской коллекции типов культур /АТСС/, г. Роквилл, шт. Мэриленд, под инвентарным номером АТСС HB10420.
Другие приемлемые антигены раскрыты в трудах Bacterial Vaccines and Local Immunity Международной конференции /Sclavo International Conf./, проведенной в г. Сиена, Италия, 17-19 ноября 1986 г. Специфические реактивы, пригодные для HIV-антигенов, раскрыты в Программе по исследовательским и эталонным реактивам в области борьбы со СПИД /AIDS Research and Reference Reagent Program/, Национальный институт здравоохранения, июнь 1989 г. Например, препарат gp 120 продает фирма "Микрогенезис, инк". Сперматозоидные поверхностно-клеточные антигены, такие как LDH-C4, являются также известными. См., например, следующую литературу: Shaha et al. and Talwar et al., Vaccine, 7: 97-100 /1989/ и Shaha et al., Int. I. Androl 11:479 /1988/.
Особый интерес при выборе антигенов применительно к практическому использованию изобретения представляет вариант слизистой защиты с прошиванием, при котором устраняется попадание в организм; и в случае этого механизма не требуется, чтобы полимерное антитело убивало или "нейтрализовало" патогенный микроорганизм. В противоположность сказанному, защита /IgC-антителами/ всего организма включает связывание, при котором, чтобы достигалась эффективность, обычно должна происходить нейтрализация патогенного микроорганизма. Таким образом, не каждое IgC-антитело, которое связывается с патогенным микроорганизмом, выполняет защитную функцию, как это иллюстрируется существованием моноклональных антител, которые специфически связывают Vibrio cholerae, но не нейтрализуют вибрион в смысле устранения колонизации, роста и проявления клинических симптомов у его хозяина. Тем самым значительно возрастает роль среды из антигенов и детерминант, пригодных для повышения защитных характеристик у IgA-антител.
3. Пассивная имунная защита.
Гибридомы, идентифицированные описанным выше методом в виде секретирующих защитных поли-Ig-моноклональных антител, культивируют известными способами, и антитело отделяют, например, посредством иммуноабсорбции, и извлекают.
Усеченный Ig-рецепторный комплекс получают либо трансформацией гибридомы совместно с вектором экспрессии, как это было описано выше, либо раздельной экспрессией Ig-рецептора и прибавлением его к моноклональному антителу способом образования монослоя, как это было описано выше.
Результирующее стабилизированное моноклональное антитело используют для создания "пассивной" вакцины, которую вводят непосредственно в слизистую поверхность, на которую может оказать провокационное воздействие рассматриваемый патогенный микроорганизм.
Конкретные носитель и способ введения комплекса находятся в зависимости от характера патогенного микроорганизма. Оральное введение обычно сопровождается приемом буферного солевого раствора или проводится с капсулированием с целью защиты комплекса от воздействия желудочной и панкреатической протеаз, когда это желательно. Назальное введение осуществляется с использованием аэрозоля. Глазной способ введения сочетается с использованием солевого раствора. Половой или ректальный способ введения осуществляется с использованием геля, пены или суппозитория.
Специалисты, работающие в этой области, умеют составлять рецептуру таких носителей известными способами.
4. Активная вакцинация.
Сложные образования из гидроксиапатита и антигенов, типа описанных выше применительно к использованию в качестве провоцирующих доноров гибридомных партнерных клеток слияния, могут быть также использованы для проведения активной вакцинации. В частности, конъюгированный антиген непосредственно наносят на слизистую ткань либо орально /используя способ капсулирования или иные средства доставки/, назальное /посредством аэрозоля/, ректально, либо вагинально /посредством суппозитория или другого носителя/.
5. Хранение препаратов.
Правопреемники патентовладельца, Президент и Член совета Гарвардского колледжа, заявляют, что переданные на хранение препараты с указанными ранее номерами АТСС 40787 и HB10420 были приготовлены согласно Будапештскому договору, заключенному с руководством Американской коллекции типов культур /АТСС/, находящейся в г. Роквилле, шт. Мэриленд. Они, далее, заявляют, что Американская коллекция типов культур /АТСС/ представляет собой организацию, обеспечивающую неизменность хранящегося препарата и легкость доступа к нему общественности, если патент выдан. Все ограничения на доступ общественности к материалу, находящемуся на хранении, полностью снимаются при получении патента. В период прохождения патентной заявки материал будет доступен лицу, указанному Членом комиссии, которое проходит под номерами 37 C.F.R. 1.15 и 35. U.S.C. 122. Отданный на хранение материал будет поддерживаться в течение периода, составляющего по крайней мере пять лет с момента получения самого последнего запроса на предоставление образца находящегося на хранении материала и в любом случае в течение периода, составляющего, по крайней мере тридцать /30/ лет с даты передачи на хранение или в течение периода дееспособности патента, если этот период является более длительным. Правопреемники патентовладельца готовы выполнять свою обязанность по замене хранящегося материала, если организация, осуществляющая хранение материала, окажется не в состоянии предоставлять запрашиваемый образец в силу его состояния.
Другие варианты подпадают под следующие пункты формулы изобретения.









Комплекс моноклональных поли-IgА предназначен для защиты млекопитающих от патогенных микроорганизмов и беременности. Комплекс моноклональных поли-IgА специфичен в отношении патогенных микроорганизмов, аллергенов или сперматозоидов. Комплекс содержит рекомбинатный стабилизаторный белок. Указанный белок включает область I и по крайней мере области IV и V поли-IgА -рецептора и не содержит области VI поли-IgА-рецептора. При этом область I отстоит от области IV и V на расстояние, составляющее около 100-500 аминокислот. Комплекс характеризуется протеолитической стабильностью по сравнению с несвязанными поли-IgА и наличием иммунозащитных и перекрестно-связывающих свойств поли-IgА. Предложен также способ получения комплекса, способ тестирования моноклональных поли-IgА и способ защиты млекопитающих с использованием данного комплекса. Изобретение обеспечивает возможность проведения особенно эффективной и безопасной пассивной иммунизации. 5 с. и 5 з.п. ф-лы. 13 ил.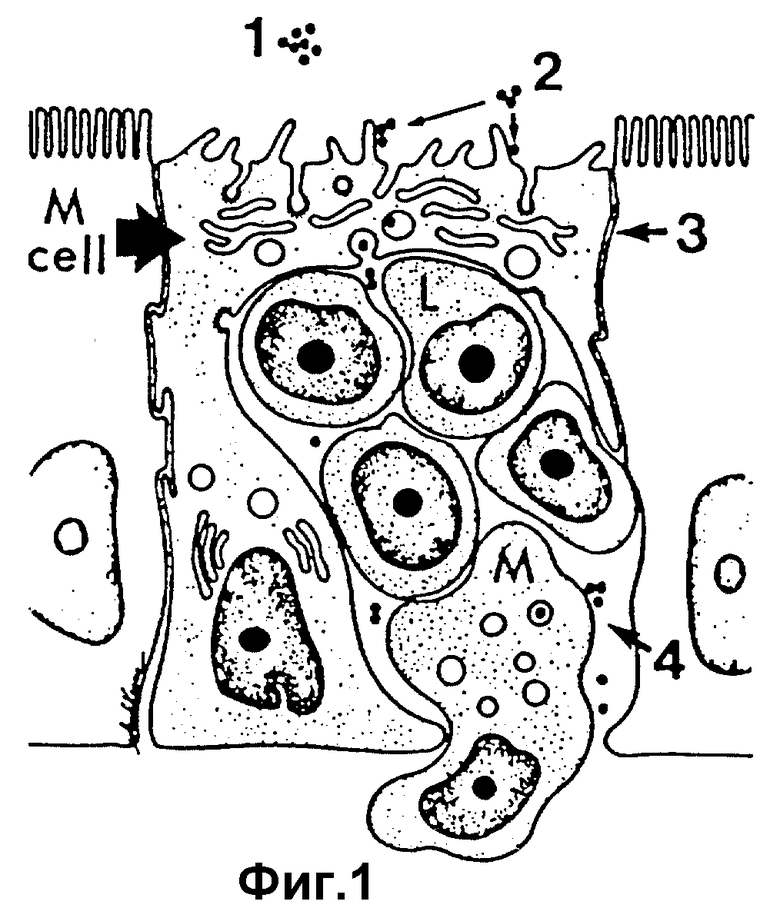
а) введение моноклональных поли-IgA в систему кровообращения млекопитающего, восприимчивого к патогенному микроорганизму, или введение моноклональных поли-IgA млекопитающему посредством подкожной имплантации опухоли или образующей опухоль клетки, продуцирующей моноклональные поли-IgA;
б) введение в слизистую оболочку млекопитающего патогенного микроорганизма или сперматозоида;
в) определение защищенности млекопитающего от патогенного микроорганизма или наличия беременности.
