Изобретение относится к растворимым формам, происходящим от мембранного протеина LAG-3 и пригодным в качестве иммунодепрессоров, так же, как к антителам, способным предотвращать специфическую фиксацию протеина LAG-3 на молекулах CMH (главный комплекс гистосовместимости) класса II, в качестве иммуностимуляторов.
В ВОИС - А 91/10682 описывается протеин, обозначаемый LAG-3.
Протеин LAG-3 представляет собой протеин, селективно экспрессируемый активированными NK-клетками и активированными T-лимфоцитами. Сходство последовательности аминокислот, сравнительная экзон-интронная структура и хромосомная локализация показывают, что LAG-3 является родственным CD 4. Первоначальная характеристика гена LAG-3 описывается TRIEBEL и др. (I).
Соответствующая ДНК кодирует трансмембранный протеин из 498 аминокислот типа I, включающий 4 внеклеточных последовательности иммуноглобулинового типа. LAG-3 является членом надсемейства иммуноглобулинов.
Зрелый протеин включает 476 аминокислот (последовательность N 1) с теоретической молекулярной массой 52 кД. Внеклеточная область содержит 8 цистеиновых остатков и 4 потенциальных сайта N-гликозилирования. С помощью Вестерн-блоттинг-анализа показано, что LAG-3 внутри ФГА-бластов или активированных NK-клеток имеет кажущуюся массу Mr = 70000. После обработки с помощью N-гликозидазы - достигают уменьшения размера до 60 кД, демонстрирующего также, что нативный LAG-3 гликолизирован. Подробности описаны в ВОИС-А 91/10682.
BAIXERAS и др. в J. Exp. Med, 176, 327-337 (2), кроме того, описывают, что образование розеток между трансфицированными с помощью LAG-3 клетками (экспрессирующими LAG-3 на своей поверхности) и B-лимфоцитами, экспрессирующими CMH класса II, специфически зависит от взаимодействия LAG-3 с CMH класса II.
Неожиданно оказалось, что этот лиганд CMH класса II обнаруживается в более высоких количествах на активированных лимфоцитах CD8 (ограниченных CMH класса I), чем на активированных лимфоцитах CD4. Ин виво, некоторые одиночные рассеянные клетки LAG-3 (ограниченные CMH класса II) находят в лимфоидной негиперплазической ткани, включающей первичные лимфодные органы, то есть тимус и костный мозг.
Клетки LAG-3 находят в лимфоидных гиперплазических узлах и миндалинах, так же как в одноядерных клетках периферической крови (PBMC) пациентов, получающих инъекции повышенных доз интерлейкина-2.
Эти наблюдения подтверждают, что LAG-3 представляет собой активационный антиген в противоположность CD4, экспрессируемый в субпопуляции покоящихся лимфоцитов и других клеточных типов, особенно в макрофагах.
CMH включает молекулы класса I и класса II, являющиеся мембранными гликопротеинами, которые представляют собой фрагменты протеиновых антигенов к рецепторам T-лимфоцитов (TCR). Молекулы класса I ответственны за презентацию цитотоксическими клетками CD8+ пептидов, происходящих большей частью от синтезированных эндогенным путем протеинов, тогда как молекулы класса II представляют собой вспомогательные лимфоциты CD4+ пептидов, происходящих в первую очередь от чужеродных протеинов, которые вводят эндоцитным, то есть экзогенным, путем. Вспомогательные T-лимфоциты регулируют и амплифицируют иммуноответ, тогда как цитотоксические лимфоциты необходимы для разрушения клеток, каковы бы ни были ткани, экспрессирующие антигены чужеродного ("не своего") типа, например, вирусные антигены. Механизм распознавания вводит в действие внутриклеточные сигналы, приводящие к эффективной активации T-лимфоцитов.
Оказывается, что для инициации иммуноответа на медиацию T-лимфоцитов (CD4+) чужеродные антигены должны быть "захвачены", интернализованы в форме пептидов специализированными клетками: антигенпредставляющими клетками (АПК). Получаемые в результате антигенные пептиды реэкспрессируют на поверхности антигенпредставляющих клеток, где они ассоциируются с молекулами CMH второго класса. Этот комплекс CMH второго класса / пептид специфически распознается T-лимфоцитным рецептором, откуда возникает активация вспомогательных T-лимфоцитов.
Кроме того, создаваемые способами рекомбинации животные модели позволяют подчеркнуть роль, которые играют ин виво молекулы CMH второго класса и их лиганды.
Так, мыши с недостатком молекул CMH второго класса (3) и практически не имеющие периферических T-лимфоцитов CD4+ и содержащие только несколько незрелых лимфоцитов CD4+ в тимусе, полностью неспособны отвечать T-зависимым антигенам.
Мутантные мыши CD4-/- (4) обладают заметно уменьшенной активностью T-лимфоцитов, но показывают нормальные развитие и функцию T-лимфоцитов CD8+, что демонстрирует, что экспрессия CD4 в дочерних клетках и тимоцитах CD4+CD8+ не является обязательной для развития. По сравнению с нормальными мышами, мыши с недостатком CD4 имеют большее количество клеток CD4-CD8-.
Эти клетки с двойным отрицательным зарядом ограничены CMH второго класса и способны распознавать антиген.
Когда их инфицируют с помощью Leishmania, эти мыши показывают популяцию вспомогательных функциональных T-лимфоцитов несмотря на отсутствие CD4. Эти клетки ограничены CMH второго класса и продуцируют γ -интерферон, когда они активированы антигеном. Это указывает на то, что происхождение T-лимфоцитов и их периферическая функция необязательно должны зависеть от функции CD4.
В настоящее время считают, что кодированные участком CMH второго класса протеины вовлекаются в многочисленные аспекты иммунного распознавания, включая туда взаимодействие между различными лимфоидными клетками, как лимфоциты и антигенпредставляющие клетки. Различные наблюдения также показывают, что другие механизмы, не через посредство CD4, принимают участие в активационной функции вспомогательных T-лимфоцитов.
Эти различные наблюдения подчеркивают главную роль, которую играет CMH второго класса и его лиганды в иммунной системе.
Кроме того, известно участие химерных молекул, образованных внецитоплазматическим доменом протеинов, способных фиксироваться на лигандах, и константной областью цепей человеческих иммуноглобулинов (Ig) в получении растворимых форм протеинов и клеточных рецепторов, пригодных особенно в качестве терапевтических агентов.
Так, растворимые формы CD4 проявляют свою эффективность в ингибировании инфекции за счет ВИЧ ин витро зависимым от дозы образом.
Однако, клинические испытания с растворимыми молекулами CD4, особенно CD4 - Ig, не позволили выявить значительное уменьшение вирусных титров. Были "созданы" трансгенные мыши, экспрессирующие вплоть до 20 мкг/мл растворимых CD4 в их сыворотке. Эти мыши не показывают никакого отличия в том, что касается их иммунной функции, по сравнению с контрольными мышами. Вплоть до настоящего времени не установлено никакой прямой связи с CMH второго класса происходящих от CD4 молекул. Это наводит на мысль, что растворимые CD4 не взаимодействуют ин виво с окрашенными молекулами CMH второго класса.
Неожиданно авторы настоящего изобретения обнаружили, что растворимые молекулы, содержащие различные фрагменты внецитоплазматического домена протеина LAG-3, способны связываться с молекулами CMH второго класса и обладать иммуноподавляющим действием.
Внецитоплазматический участок LAG-3, обозначаемый последовательностью N 1, включает домены Д1, Д2, Д3, Д4, охватывающие аминокислоты 1-149, 150-239, 240-330 и 331-412, соответственно.
Предметом изобретения также является полипептидная растворимая (часть) фракция, образованная целиком или частью по крайней мере одного из четырех внеклеточных доменов иммуноглобулинового типа протеина LAG-3 (аминокислоты 1-149, 150- 239, 240-330 и 331-412 последовательности N 1) или пептидной последовательностью, происходящей от этих доменов за счет замены, присоединения и/или делеции одной или нескольких аминокислот, и которая обладает специфичностью, по крайней мере равной или выше таковой LAG-3 в его лиганде.
Настоящее изобретение особенно относится к растворимым полипептидным фракциям, обладающим последовательностью, образованной от нативной последовательности LAG-3 вследствие хорошо известного явления политипии.
Растворимая полипептидная фракция отличается тем, что она включает пептидный участок LAG-3, ответственный за сродство LAG-3 к молекулам CMH второго класса.
Растворимая полипептидная фракция в особенности включает пептидную последовательность, образованную из этих доменов за счет замены, присоединения и/или делеции одной или нескольких аминокислот, и которая обладает специфичностью, равной или выше таковой LAG-3 в его лиганде, например, совокупность двух первых доменов иммуноглобулинового типа LAG-3 или 4 домена иммуноглобулинового типа внецитоплазматического участка LAG-3.
Предпочтительно, растворимая полипептидная фракция образована целиком или частью по крайней мере одного из четырех внеклеточных доменов иммуноглобулинового типа протеина LAG-3 (аминокислоты 1-149, 150-239, 240-330 и 331-412 последовательности N 1), включающего замещение глутаминовой кислотой (Glu) одного или нескольких аргининовых остатков (Arg) в положениях 73, 75 и 76 последовательности N 1.
Предпочтительно, растворимая полипептидная фракция включает петлю, среднее положение атомов которой, образующих основную цепь, представлено позицией аминокислот 46-47 (последовательность N 1), представленной в таблице 1 или таблице 2, или отличается от нее самое большее на 5%.
Растворимая полипептидная фракция, кроме того, предпочтительно включает второй внеклеточный домен (Д2) иммуноглобулинового типа LAG-3 (аминокислоты 150 -239).
Предпочтительно, растворимая полипептидная фракция включает, кроме вышеуказанной пептидной последовательности LAG-3, дополнительную пептидную последовательность на C-конце и/или N-конце, так, чтобы образовать слитый белок. Термин "слитый белок" означает часть любого протеина, позволяющую осуществлять модификацию физико-химических характеристик субфрагмента внецитоплазматического домена протеина LAG-3. Такие слитые белки, например, содержат фрагменты внецитоплазматического домена LAG-3, такие, как указанные выше, связанные с областью связывания -CH2-CH3 тяжелой цепи человеческого иммуноглобулина, предпочтительно иммуноглобулина изотипа IgG4.
Такие слитые белки могут быть димерными или мономерными. Эти слитые белки могут быть получены с помощью хорошо известных специалисту методов рекомбинации, например, с помощью способа, описанного Traunecker и др. (5).
Вообще, способ получения этих слитых белков, включающих иммуноглобулиновую область, соединенную с вышеописанной пептидной последовательностью LAG-3, состоит в том, что в вектор вставляют фрагменты кДНК, кодирующие полипептидные области, соответствующие LAG-3 или производным LAG-3, возможно после амплификации с помощью полимеразной цепной реакции (ПЦР), и кДНК, кодирующую подходящую область иммуноглобулина, соединенную с кДНК, кодирующей соответствующие полипептидные или образованные от LAG-3 области, и после трансфекции фрагменты кДНК экспрессируют в системе экспрессии, особенно в клетках млекопитающих, например, в клетках яичника хомяка.
Слитые белки согласно изобретению также могут быть получены путем расщепления конъюгата LAG-3 - - Ig, сконструированного так, что он содержит соответствующий сайт расщепления.
Предметом изобретения также является терапевтическая композиция с иммуноподавляющей активностью, включающая растворимую полипептидную фракцию согласно изобретению. Эта композиция пригодна для лечения патологий, при которых необходима иммуносупрессия, например, аутоиммунных заболеваний.
Предметом изобретения также является использование антител, направленных против LAG-3, или растворимых полипептидных фракций, образованных от LAG-3, таких, как указанные выше, или фрагментов таких антител, особенно фрагментов Fab, Fab', F(ab')2, для приготовления терапевтической композиции с иммуностимулирующей активностью. Термин "иммуностимулирующий" обозначает молекулярную совокупность, способную стимулировать созревание, дифференцирование, пролиферацию и/или функцию клеток, экспрессирующих LAG-3, то есть активированных T-лимфоцитов или NK-клеток (естественных киллерных клеток). Антитела против LAG-3 (анти-LAG-3-антитела) могут быть использованы в качестве потенциализаторов (усилителей) вакцин или иммуностимуляторов в случае пациентов с подавленной (угнетенной) иммунной системой, таких, как больные, инфицированные ВИЧ или обработанные иммуноподавляющими средствами, или они могут быть использованы для стимуляции иммунной системы за счет удаления из нее клеток, обладающих анормальным состоянием, например, раковых клеток.
Иммуностимулирующая активность анти-LАG-3 -антител является неожиданной там, где анти-CD4-антитела оказывают иммуноподавляющее воздействие.
Такие антитела могут быть поликлональными или моноклональными; однако, предпочтительны моноклональные антитела. Поликлональные антитела могут быть получены хорошо известными методами, например, таким, как описанный BENEDICT А. А. и др. (6). Предпочтительны моноклональные антитела вследствие того, что они специфичны для одного эпитопа и дают результаты с лучшей воспроизводимостью. Методы получения моноклональных антител хорошо известны из уровня техники, в особенности метод, описанный KOHLER и MILSTEIN. Этот метод, так же, как его варианты, описаны YELTON и др. (7).
Предметом изобретения также являются антиидиотипические антитела, направленные против антител согласно изобретению, которые включают "внутренний образ" LAG-3 и, следовательно, способны связываться с CMH второго класса. Такие антитела могут быть использованы особенно в качестве иммуноподавляющих средств и, например, при аутоиммунных патологиях.
Терапевтические композиции согласно настоящему изобретению включают растворимые протеины LAG-3 или антитела, такие, как указанные выше, так же как фармацевтически приемлемый эксципиент. Эти композиции могут быть сформулированы обычными способами. Эксципиент может изменяться в зависимости от выбранного пути введения: орально, парентерально, подъязычно, ректально или через нос.
В случае предназначенных для парентерального введения композиций эксципиент обычно включает стерильную воду, так же, как другие возможные ингредиенты, повышающие растворимость или пригодность для хранения композиции. Парентеральные пути введения могут заключаться во внутривенных, внутримышечных или подкожных инъекциях.
Терапевтическая композиция может быть пролонгированного действия, особенно для обработок в течение длительного периода времени, например, в случае аутоиммунных заболеваний. Вводимая доза зависит от излечиваемого субъекта, особенно от способности его иммунной системы достигать желательной степени защиты. Точные количества вводимого активного ингредиента могут быть легко определены практикующим врачом, который назначает лечение.
Терапевтические композиции согласно изобретению могут включать, кроме растворимого LAG-3 или антител согласно изобретению, другой активный ингредиент, возможно связанный химической связью с LAG-3 или с антителом согласно изобретению. В качестве примера можно назвать растворимые протеины LAG-3 согласно изобретению, связанные с токсином: например, как рицин или дифтерийный анатоксин, способные фиксироваться на молекулах CMH второго класса и убивать клетки, против которых они направлены, например, лейкемические клетки или клетки меланомы; или связанные с радиоактивным изотопом.
Следующие примеры, так же, как прилагаемые стандартизованные чертежи, иллюстрируют подробно изобретение.
Пример 1: Пролиферация линий активированных T-лимфоцитов в присутствии моноклональных антител против LAG-3
Моноклональными анти-LAG-3-антителами, которые используют, являются 17. B4, описанное BAIXERAS и др. (2) и депонированное в Национальной коллекции культур и микроорганизмов под номером 1-1240 10 июля 1992 г. , и 11 E3, описанное HUARD и др. (8).
Эти антитела принадлежат к изотипу IgGI. Эти антитела испытывают в отношении их биологического воздействия на активированные T-лимфоциты, стимулированные специфическими антигенными пептидами или перестроенными антигенами, представляемыми молекулами CMH второго класса, экспрессированными автологичными антигенпредставляющими клетками, экспрессирующими LAG-3.
Моноклональное антитело против CD48, обозначаемое 10 H3, используют в качестве неподходящего антитела IgGI (отрицательный контроль).
Насыщающие концентрации анти-LAG-3- и анти-CD48-антител определяют путем иммунофлуоресценции на ФГА-бластах (фитогемагглютинин) и линиях клеток, трансформированных вирусом Эпштейна-Барра (ЭБВ). В опытах по пролиферации, моноклональные антитела добавляют в количестве, в 5 раз превышающем насыщенную концентрацию.
Используемые линии T-лимфоцитов представляют собой, с одной стороны, клон 154, происходящий от лимфоцитов периферической крови, выращенной против пептида, имитирующего фрагмент гемагглютинина (HA) гриппа, который обладает аминокислотной последовательностью, охватывающей аминокислоты 306 - 329 (пептид p20); с другой стороны, клон 28, T-лимфоцитный клон, происходящий от периферических лимфоцитов одного человеческого донора, выращенный против дифтерийного анатоксина (ДТ). Антигенпредставляющие клетки (АПК), соответствующие клону 154, являются B-лимфоцитами, трансформированными вирусом Эпштейна-Барра того же самого донора (DR 3/DR 11), что и T 154. Антигенпредставляющие клетки, соответствующие клону 28, являются B-лимфоцитами, трансформированными вирусом Эпштейна-Барра одного и того же донора. Этот клон ограничен главным комплексом гистосовме-стимости человека - донора 7.
В случае клона 154, АПК (5•106) инкубируют при 37oC в течение 1,5 часов с изменяемыми дозами пептида p20, затем промывают и облучают (10000 рад. ). Клетки наносят монослоем на микротитрационный планшет с 96 углублениями одновременно с клетками клона 154 (0,5•105 - 10•105 клеток на мл) в соотношении 3: 1. Для клона 28 соотношение отвечающие клетки/стимулирующие клетки составляет 1.
Антигенпредставляющие клетки главного комплекса гистосовместимости человека DR 7/ ЭБВ обрабатывают митомицином или облучают, затем добавляют к T-лимфоцитам в присутствии ДТ (который остается в культуре). Конечная концентрация клеток клона 28 составляет 100000 клеток на мл.
3H-Тимидин (1 микрокюри/углубление) добавляют в интервалы времени, изменяющиеся от дня 2 до дня 10 культивирования.
Каждый эксперимент повторяют трижды.
Результаты выражают в виде среднеарифметического числа импульсов в минуту и после вычитания числа импульсов в минуту, обнаруженных в отрицательном контроле (T-лимфоциты, сокультивированные с незаряженными, в виде иммуногенов, АПК). Опыты по пролиферации осуществляют на планшетах с 96 углублениями. Абсорбцию меченного тритием тимидина в индивидуальных углублениях по 200 мкл измеряют после добавления 1 микрокюри тимидина в последние 18 часов культивирования. Результаты выражают в виде среднего из трех опытов. Типичное отклонение обычно составляет величину менее 12% (немного больше в случае измерений очень незначительного числа импульсов в минуту). С другой стороны, недостаточные жидкости смешанной культуры (клон 154/АПК) собирают, отфильтровывают при использовании мембран с порами размером 0,22 мкм, разделяют на образцы и замораживают при -20oC вплоть до момента титрации с помощью имеющихся в продаже наборов для иммуно-испытаний: набор для титрации IL-2 и INF- α фирмы Иммунотех; набор IFN-γ фирмы Гензим и набор IL-4 фирмы Саумап Chemicals.
Изучение в отношении определения дозы осуществляют для установления профилей (контуров) пролиферации клона 154 в присутствии специфического антигена p20 в изменяемых концентрациях и в присутствии или нет моноклональных анти-LAG-3-антител или моноклональных чужеродных (неподходящих) антител (отрицательный контроль).
Индивидуальные результаты 16 различных опытов показывают, что, какова бы ни была концентрация добавленного антигена, начальная точка вплоть до пика пролиферации не изменяется, однако систематически наблюдают значительное продление пролиферации T-лимфоцитов, инкубируемых с моноклональными анти-LAC2-3-антителами. Фрагменты Fab моноклонального антитела 17B4 получают и используют в опыте по пролиферации клона 154. Профиль пролиферации T-лимфоцитов, активированных антигеном с фрагментами Fab 17B4 (15 мкг/мл), подобен таковому клеток, инкубированных в присутствии целого моноклонального антитела 17B4 (40 мкг/мл) (фиг. 1). Эти результаты показывают, что наблюдаемые биологические эффекты нельзя приписать неспецифической реакции, индуцируемой областью Fc моноклональных анти-LAG-3-антител.
Подобные результаты получают при использовании моноклональных анти-LAG3-антител IIE3.
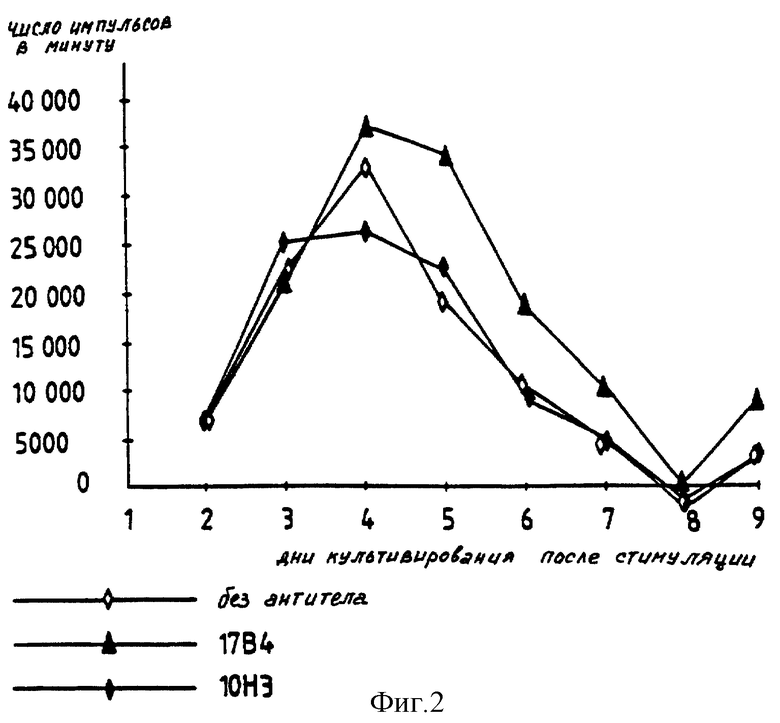
Клон 28 также стимулируют антигеном (столбнячный анатоксин, 10 мкг/мл) в присутствии моноклональных антител 17B4 после сокультивирования с соответствующими антигенпредставляющими клетками (АПК) в присутствии ДТ. Результаты представлены на фиг. 2.
Эффекты моноклональных анти-LAG-3-антител, наблюдаемые в случае клона 28, а именно прологация пролиферации, подобны таковым, наблюдаемым в случае клона 154.
Были проведены опыты, предназначенные для определения разнообразных клеточных событий (les evenements), происходящих после антигенной стимуляции клеток клона 154, инкубированных в присутствии моноклональных анти-LAG-3-антител.
Клетки собирают во время классической антигенной стимуляции клона 154 в присутствии моноклональных анти-LAG-3- или анти-CD48-антител или в присутствие антител и испытывают для экспрессии трансмембранных рецепторов LAG-3 и CBB25 и образцы культуральных надосадочных жидкостей отбирают в различные интервалы времени после стимуляции и испытывают на наличие γ -интерферона, α -фактора некроза опухоли, интерлейкина-4 и интерлейкина-2.
Опыты в случае прямой иммунофлуоресценции с 2 красителями (моноклональные анти-CD3-антитела и моноклональные анти-CD25-антитела) показывают, что рецепторы для интерлейкина-2 слабо, но значительно увеличиваются спустя 5 дней после антигенной стимуляции. Подобные опыты с моноклональными анти-CD3- и IIE3 (анти-LAG-3)-антителами показывают, что LAG-3, начиная со дня активации, сверхэкспрессируется. Кроме того, секреция интерлейкина-2, интерлейкина-4, γ -интерферона и α -фактора некроза опухоли также модулируется за счет инкубации с моноклональными анти-LAG-3-антителами, что свидетельствует о том, что различные события в клетках изменяются вследствие присутствия моноклональных анти-LAG-3-антител и что некоторые события происходят уже спустя 24 часа после стимуляции.
Эти результаты косвенно показывают, что LAG-3 играет роль регулятора в случае клеток CD4+. Тот факт, что моноклональные анти-LAG-3-антитела увеличивают пролиферацию, следовательно действуют как иммуноусилители, свидетельствует о том, что LAG-3 принимает участие в "дезактивации" T-лимфрцитов CD4+ при отрицательной роли LAG-3 в отношении зависимой от антигена стимуляции.
Пример 2: Транзиторная экспрессия слитых белков LAG-3
Растворимые протеины (белки), происходящие от LAG-3, получают по методу рекомбинантной ДНК с помощью соответствующих векторов, включающих ДНК, кодирующую LAG-3, и ДНК, кодирующую фрагмент иммуноглобулина. Система транзиторной экспрессии состоит из трансфицированных клеток Cos. Эта система позволяет продуцировать несколько миллиграммов рекомбинантных слитых белков. Методы рекомбинантной ДНК осуществляют как описано MANIATIS и др. (22). Изменения делают в соответствии с рекомендациями изготовителя.
Конструкция LAG-3 Д1-Д4 Ig и LAG-3 Д1 Д2 Ig
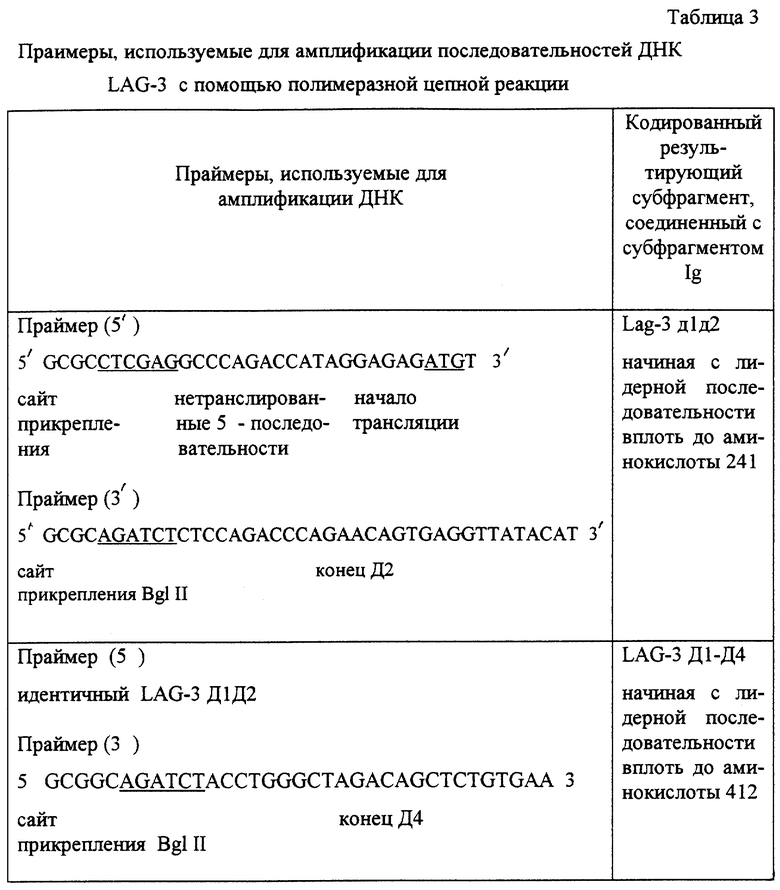
Фрагменты, кодирующие области Д1 Д2 или Д1-Д4, амплифицируют (30 циклов) из фрагментов кДНК (последовательность фолликулярной дендритной клетки, FDC), включающего кДНК LAG-3 (TRIEBEL и др. , /1/), с помощью Tag-полимеразы, лишенной активности 5'-эндонуклеазы и относительно устойчивой к выдерживанию при очень высокой температуре; после амплификации следует денатурация при 98oC (с помощью "термальной кольцевой ДНК, Перкин Элмер Цетус").
Используют специфические праймеры, перечисленные в нижеприводимой таблице.
Полученные в результате амплифицированные фрагменты (739 пар оснований и 1312 пар оснований для LAG-3 Д1Д2 и LAG-3 Д1-Д4, соответственно) вставляют в плазмиду pBS (Стратаген).
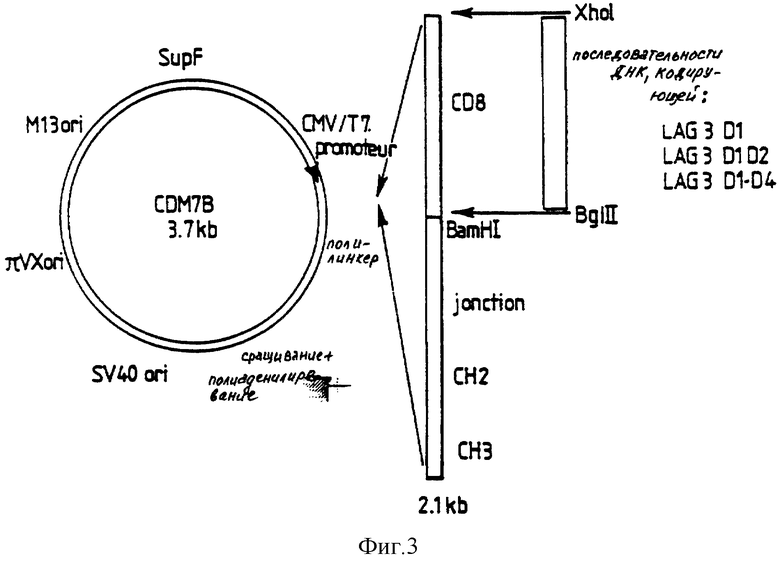
Вставки получают после переваривания с помощью XhoI и BglII и вводят в сайты XhoI/BamHI вектора pCDM7 - CD8 - IgGI (причем pCDM7 происходит от pCDM8, выпускаемого в продажу фирмой Стратаген), как проиллюстрировано на фиг. 3, чтобы обменять последовательности ДНК, кодирующие CD8, на клетки, кодирующие субфрагменты LAG-3. Полученные в результате экспрессирующие векторы содержат последовательности, кодирующие Д1Д2 или Д1-Д4, соединенные с последовательностями ДНК, кодирующими участок соединения -CH2-CH3 цепи человеческого IgHI.
CDM7 представляет собой эукариотный экспрессирующий вектор, происходящий от векторов, описанных SEED и др. (10) для клонирования ДНК и ее экспрессии в E. coli и эукариотных клетках. CDM7 имеет следующие характеристики: (I) промотор человеческого цитомегаловируса для транзиторной экспрессии в клетках млекопитающих; (II) вирусный источник SV40 для аутосомной репликации клеток млекопитающих, экспрессирующих T-антиген; (III) π VX (тип Col El) в качестве плазмидного источника для увеличенного числа копий; (IV) селекция SupF по резистентности к ампициллину и тетрациклину в штаммах E. coli Tetamb и Ampamb; (V) источник репликации MI3 для высвобождения однонитевой РНК (un simple brin); (VI) промотор РНК Т7; (VII) полилинкер для эффективного клонирования гетерологичной ДНК.
Транзиторная экспрессия в клетках Cos
Клетки Cos (5•106) трансфицируют с помощью 30 мкг ДНК соответствующих экспрессирующих векторов (кодирующей либо LAG-3 Д1Д2 Ig, либо LAG-3 Д1-Д4 Ig, либо CD8 Ig) путем электропорации (200 вольт, 1500 мкфарад, 30-40 мсек) с помощью аппарата Cellject (Eurogentech, Льеж, Бельгия). Клетки снова наносят тонким слоем (монослоем) и культивируют при использовании среды, содержащей 5% плодной телячьей сыворотки. Надосадочные жидкости отбирают спустя 6 дней после трансфекции.
За синтезом получающихся слитых белков следят путем анализирования надосадочных жидкостей, так же, как экстрактов трансфицированных клеток по методу Вестерн-блоттинга с моноклональными антителами 17B4. Иммунореактивные материалы обнаруживают в надосадочной жидкости клеток, трансфицированных с помощью ДНК, кодирующей LAG-3 Д1Д2 Ig или LAG-3 Д1-Д4 Ig.
Параллельно, рекомбинантный иммуноадгезин CD8 (CD8 Ig) получают в качестве отрицательного контроля с помощью той же самой системы экспрессии и экспрессирующего вектора pCDM7-CD8 (фиг. 3).
Рекомбинантные протеины LAG-3 Д1Д2 Ig, LAG-3 Д1-Д4 Ig и CD8 Ig очищают по классическому методу на протеин A-сефарозе. Получающийся материал анализируют путем электрофореза в полиакриламидном геле с додецилсульфатом натрия с последующим Coomassie - окрашиванием или анализом по методу Вестерн-блоттинга с помощью человеческого анти-Ig-антитела.
Пример 3: Получение растворимых субфрагментов LAG-3
Для того чтобы получить большие количества рекомбинантных протеинов, была разработана стабильная система экспрессии, состоящая из трансфицированных клеток млекопитающих. Клетками-хозяевами являются клетки яичника хомяка (CHO), зависимые от анкеровки, выделяемые из CHO-клеток с недостатком дигидрофолатредуктазы (dhfr) и, следовательно, требующими для своего роста глицина, пурина и тимидина. Главная роль dhfr в синтезе нуклеиновых предшественников в сочетании с чувствительностью клеток dhfr с недостатком в отношении аналогов тетрагидрофолата, таких, как метотрексат (МТХ), имеет два значительных преимущества. Трансфекция этих клеток экспрессирующими векторами, содержащими ген dhfr, позволяет выделять dhfr - резистентные рекомбинантные клоны, и культура этих клеток на селективных средах, содержащих возрастающие количества МТХ, происходит из амплификации гена dhfr и ДНК, которая ассоциирована с ним.
Конструкция LAG-3 Д1, LAG-3 Д1Д2, LAG-3 Д1-Д4
Фрагменты ДНК, кодирующей области Д1, Д2Д2 или Д1-Д4, амплифицируют с помощью способа полимеразной цепной реакции, идентичного таковому, описанному выше, с помощью праймеров, представленных в таблице 4.
Результирующие амплифицированные фрагменты переваривают с помощью SalI и вставляют в сайт SalI pUC18 (Стратаген).
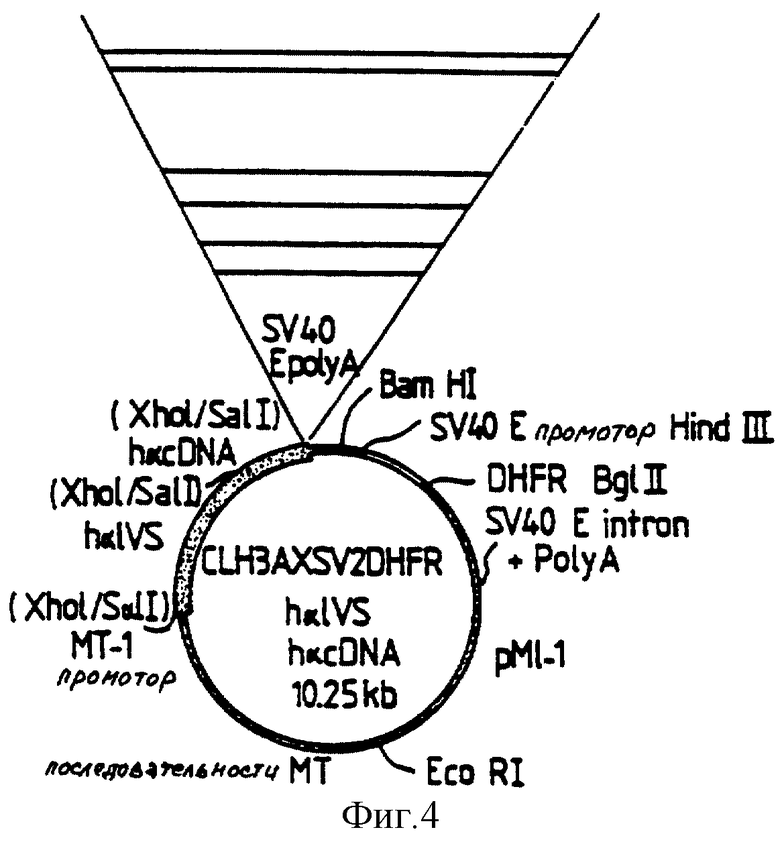
Амплифицированные последовательности идентифицируют и вставки субклонируют в экспрессирующем векторе pCLH3 AXSV2 DHFR h α IVS, как описано COLE и др. (Biotechnology, II, 1014-1024, 1993) (фиг. 4).
Этот вектор представляет собой многофункциональный эукариотный экспрессирующий вектор для экспрессии кДНК и ее амплификации в эукариотных клетках. Он имеет следующие характеристики: (I) мышиный промотор гена металлотионеина-1 и последовательность полиаденилирования SV40 (включающая сайт донорно-акцепторного связывания) для осуществления транскрипции целевого гена; (II) человеческий интрон A, включающий сайт донорно-акцепторного связывания гена субъединицы α -гликопротеина, для достижения высоких степеней транскрипции кДНК; (III) последовательность pML, содержащая источник репликации pBR322 и ген устойчивости к ампицилину, для бактериальной амплификации; (IV) единица транскрипции dhfr SV 40 для осуществления транскрипции последовательностей, используемых для селекции и амплификации трансфектантов.
Стабильная экспрессия в CHO-клетках
Экспрессирующие векторы, кодирующие LAG-3 Д1, LAG-3 Д1Д2 и LAG-3 Д1-Д4, используют для трансфекции клеток CHO DUKX и эти клетки культивируют на селективной среде. Клетки, способные размножаться в этих условиях, собирают и культивируют на среде, содержащей возрастающие количества МТХ. Степень экспрессии определяют с помощью анализа по методу Вестерн-блоттинга при использовании моноклонального антитела 17B4. Клоны, продуцирующие большие количества растворимых рекомбинантных молекул, происходящих от LAG-3, размножают в биореакторах и происходящий от LAG-3 материал очищают путем ионообменной и иммуноаффинной хроматографии.
Анализы по методу Вестерн-блоттинга выявляют в надосадочных жидкостях клеток, трасфицированных с помощью экспрессирующих векторов, кодирующих LAG-3 Д1, LAG-3 Д1Д2, LAG-3 Д1-Д4, полосы с кажущимися молекулярными массами Mr 15-18 кД, 34-36 кД (дублеты) и 55 кД (2 возможные полосы). Соответствующие Mr этих иммунореактивных материалов соответствуют ожидаемым Mr для гликолизированных LAG-3 Д1 Ig (139 аминокислот и предполагаемый сайт гликолизирования), LAG-3 Д1Д2 Ig (239 аминокислот, включающих 3 сайта гликолизирования) и LAG-3 Д1-Д4 Ig (412 аминокислот, включающих 4 сайта гликолизирования).
Пример 4: Специфическое связывание LAG-3 Ig с клетками, экспрессирующими CMH второго класса
Реакционноспособность моноклональных антител и LAG-3 Д1-Д4 Ig изучают путем косвенной (непрямой) иммунофлуоресценции. Целевые клетки (4х10) культивируют в течение 30 минут при 4oC в присутствии LAG-3 Д1-Д4 Ig, CD8 Ig, мышиного моноклонального антитела, (949) человеческого анти-CMH второго класса (DR, DP, DO), конъюгированного с изотиоцианатфторидом (FITC), клона Coulter или мышиного Ig - изотиоцианатфторида: чужеродный иммуноглобулин G, конъюгированный с изотиоцианатфторидом. Клетки промывают и инкубируют при 4oC в течение 30 минут либо с поликлональным козьим F(ab') против челевеческого иммуноглобулина, конъюгированного с флуоресцеином, либо с поликлональным козьим антителом против мышиного иммуноглобулина, конъюгированного с флуоресцеином (клон Coulter).
Для подтверждения связи LAG-3/CMH второго класса, LAG-3 Д1-Д4 инкубируют с положительными или отрицательными клетками CMH второго класса. Четыре линии B-лимфоцитов, экспрессирующих CMH второго класса (L31, Phil ЭВВ, Raji, Sanchez и Personnaz), обрабатывают моноклональным антителом 949 против класса II, или надосадочными жидкостями клеток Cos, трансфицированных ДНК, кодирующей либо LAG-3 Д1-Д4 Ig, либо CD8 Ig. Пять линий клеток, экспрессирующих различные гаплотипы молекул CMH второго класса, распознаются LAG-3 Ig таким же образом, как и моноклональными антителами против класса II (положительный контроль), тогда содержащая CD8 Ig надосадочная жидкость (отрицательный контроль) не связана с этими линиями клеток, как можно было ожидать. Четыре линии отрицательных клеток CMH второго класса (CEM, RJ, HSB2, K562) обрабатывают теми же реагентами, что и указанные выше. Отсутствие реакции с анти-CMH второго класса (отрицательный контроль), а также отсутствие реакции с LAG-3 Д1-Д4 Ig показывает, что связь LAG-3 Д1-Д4 с молекулами CMH второго класса является специфичной.
Дополнительные эксперименты реализуют с помощью (I) мышиных фибробластов, трансфицированных или нетрансфицированных с помощью генов, кодирующих человеческий DR7 или человеческий DP4, (II) мышиных клеток, экспрессирующих или неэкспрессирующих молекулы CMH второго класса, (III) активированных человеческих клеток CD4+ или CD8+, и (IV) линий T-лимфоцитов, экспрессирующих различные гаплотипы молекул CMH второго класса (фиг. 8).
В противоположность CD8 Ig, LAG-3 Д1-Д4 Ig связывается с любыми из клеток, экспрессирующих CMH второго класса, также эффективно, как моноклональное антитело 949 против CMH второго класса. LAG-3 Д1-Д4 Ig связывается с любыми из исследованных гаплотипов DR и DP, с молекулами человеческого CMH второго класса, экспрессируемыми трансфицированными мышиными клетками, с молекулами мышиного CMH второго класса, так же, как с молекулами CMH второго класса, экспрессируемыми T-лимфоцитами CD4+ или CD8+.
Эти результаты впервые служат доказательством того, что растворимые молекулы, происходящие от лиганда CMH второго класса, способны фиксироваться на клетках, экспрессирующих CMH второго класса.
Подобные эксперименты показывают, что LAG-3 Д1Д2 связывается с клетками, экспрессирующими CMH второго класса, также специфически и с такой же эффективностью, что и LAG-3 Д1-Д4.
Активность связи LAG-3 Ig и распределение в клетках лиганда LAG-3 Ig
Способность этого иммуноадгезина связываться с клеточными лигандами определяют с помощью козьей сыворотки, направленной против человеческих иммуноглобулинов, маркированной флуоресцеином.
В этих экспериментах целевые клетки сначала инкубируют с человеческим моноклональным антителом или иммуноадгезином в течение 30 минут при 4oC в RPMI 1640, содержащей 10% плодной телячьей сыворотки (FCS). Клетки затем инкубируют с козьей сывороткой против мышиных иммуноглобулинов, маркированной FITC (Coulter), для моноклональных мышиных антител или с козьей сывороткой против человеческих иммуноглобулинов, маркированной FITC (Tago), для иммуноадгезинов. Флуоресценцию измеряют после двух промывок, анализируя 3000 клеток на цитометре Elite (Coultronics, Hialeah, Финляндия). На фиг. 9 представлены степени фиксации LAG-3 Ig, CD8 Ig, антитела 949 или антитела ОКЗТ (анти-CD3, ATCC), в виде числа подсчитанных клеток в зависимости от логарифма интенсивности измеренной флуоресценции.
LAG-3 Ig фиксируется на мышиных трансфицированных фибробластах для гена молекулы HLA DR4, но не фиксируется на нетрансфицированных клетках. CD8 Ig неспособен связываться с фибробластами HLA DR4 + в тех же условиях. Распределение по клеткам лигандов LAG-3 Ig оценивают на образе клеточных популяций путем иммунофлуоресценции.
LAG-3 Ig обнаруживают во всех положительных испытуемых клетках второго класса, включая туда линии B-клеток, трансформированных вирусом Эпштейна-Барра (производные генетически неродственных доноров, включающих 10 гомозиготных линий типа DR1-DR10), так же как в активированных T-клетках и NK-клетках (естественных киллерных клетках).
На фиг. 9 в качестве примера представлено соединение LAG-3 Ig положительных субклеток DAUDI с антигенами второго класса.
Средняя интенсивность флуоресценции с LAG-3 Ig подобна таковой, наблюдаемой в случае специфического антитела 949 атнигенов второго класса. Фиксация LAG-3 Ig на DR4 (фиг. 9), DR2, DR7 или DPw4 (не представлено), экспрессированных на поверхности мышиных фибробластов, напротив, более слабая, чем таковая, наблюдаемая для антител 949.
Не обнаружено никакого связывания с отрицательными линиями клеток для антигенов второго класса T-происхождения (T-клетки периферической крови, линии CEM, HSB2, REX), B-происхождения (линии Rf 2.2.5), или нелимфоидного происхождения (человеческие линии K562 критро-миклоидного происхождения и линии, происходящие от клеток меланомы - /не представлены/).
Кроме того, LAG-3 Ig фиксируется на ксеногенных молекулах второго класса CMH, таких, как антигены, экспрессированные мышиной лимфомой A 20, и обезьяньих классов II, экспрессированные бластами, стимулированными фитогемагглютинином (данные не представлены).
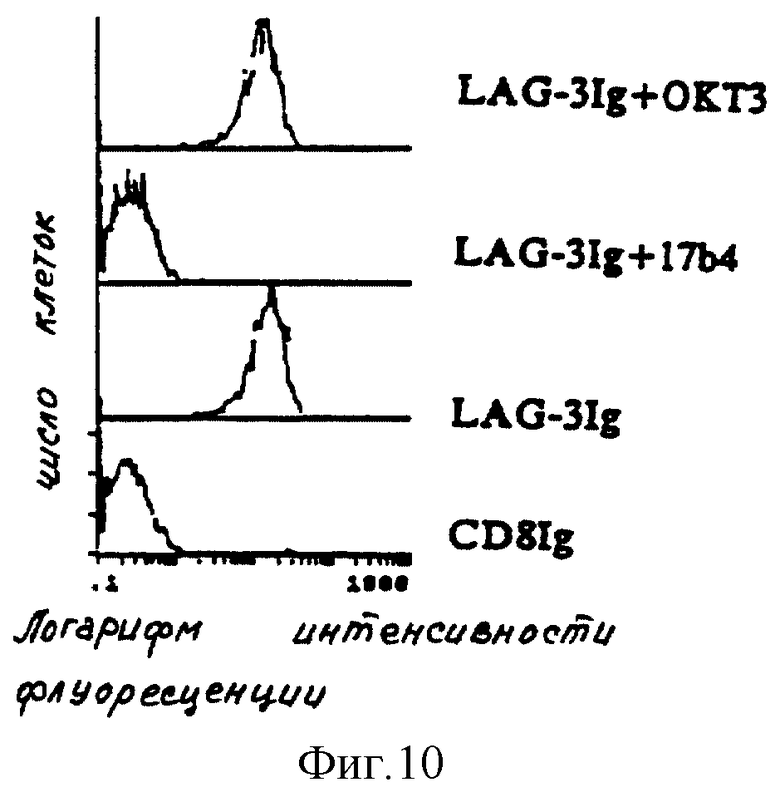
Специфичность фиксации LAG-3 Ig также подтверждают путем использования моноклонального антитела 17B4, способность которого блокировать взаимодействия LAG-3 с CMH второго класса в тестах по клеточной адгезии была показана ранее (фиг. 10).
В этих экспериментах молекулы LAG-3 Ig предварительно инкубируют в течение 30 минут при 4oC либо в одной среде, либо с 17B4 (1 мг/мл), либо с ОКТЗ (1 мг/мл), до введения в соприкосновение с клетками DAUDI.
На фиг. 10 показано, что предварительная инкубация LAG-3 Ig с 17B4 ингибирует фиксацию на клетках класса II+, тогда как никакого ингибирования не наблюдают с контролем ОКТЗ.
Пример 5: Ингибированные взаимодействия LAG-3 с CMH класса II с помощью растворимых фрагментов LAG-3
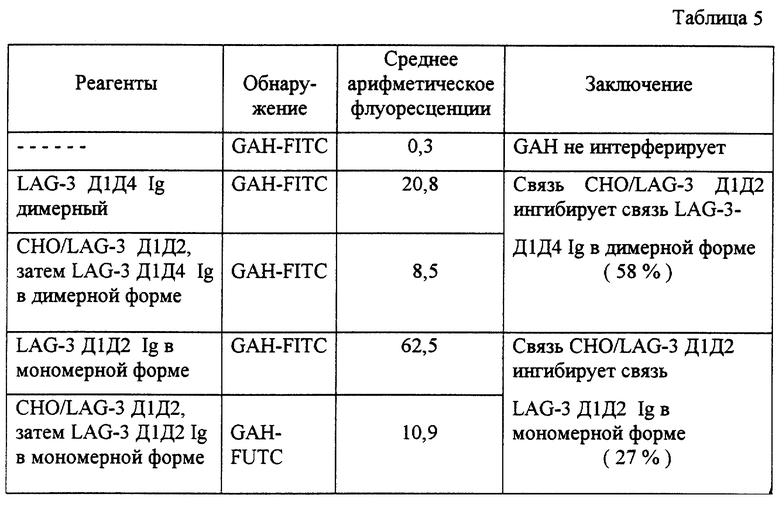
Ингибирование взаимодействия LAG-3 с CMH класса II с помощью растворимых фрагментов LAG-3 может наблюдаться непосредственно при фиксации LAG-3 Ig на CMH второго класса в экспериментах по конкуренции с растворимыми фрагментами.
Для подтверждения того, могут ли растворимые фрагменты LAG-3 Д1Д2, продуцируемые клетками яичника хомяка, заменять связь иммуноадгезинов, происходящих от LAG-3, осуществляют следующие опыты:
Клетки DAUDI инкубируют с растворимыми фрагментами LAG-3 Д1Д2, чтобы могла осуществиться фиксация этих молекул на антигенах второго класса CMH, экспрессированных на поверхности клеток DAUDI.
Во второй стадии, клетки инкубируют в присутствии LAG-3 - Д1Д4 Ig в димерной форме или LAG-3 Д1Д2Ig в мономерной форме.
Фиксацию этих иммуноадгезинов, происходящих от LAG-3, определяют с помощью козьего F(ab')2 против человеческого иммуноглобулина, конъюгированного с флуоресцеином (GAH FITC).
Контрольные группы представлены клетками DAUDI, инкубированными с димерной формой LAG-3 Д1Д2 Ig или мономерной формой LAG-3 Д1Д2 Ig без предварительной инкубации с растворимыми фрагментами LAG-3 Д1Д2.
Результаты представлены в таблице 5, где указывается, что растворимые фрагменты LAG-3 Д1Д2 способны замещать иммуноадгезины, происходящие от LAG-3 в мономерной или димерной форме.
Эти данные подтверждают, что растворимые фрагменты LAG-3 Д1Д2 фиксируются на молекулах второго класса CMH.
Ингибирование взаимодействия LAG-3/CMH второго класса и CD4/CMH второго класса
Образование розеток между клетками Cos, трансфицированными LAG-3 дикого типа, и B-лимфоцитами, трансформированными вирусом Эпштейн-Барра, экспрессирующим молекулы CMH второго класса, показано BAIXERAS и др. (2). Это взаимодействие ингибируется одновременно моноклональными антителами и против человеческой гемоглобиновой клетки (MHC).
Описанный в этой публикации метод видоизменяют, заменяя визуальное наблюдение и подсчет связывающихся с B-лимфоцитами клеток Cos подсчетом остаточной радиоактивности после инкубации меченных с помощью 51Cr B-лимфоцитов с клетками Cos, экспрессирующими LAG-3 (опыт в отношении связывания).
Изучают возможные ингибирующие воздействия растворимых молекул, происходящих от LAG-3, на взаимодействие LAG-3/CMH второго класса, а также на взаимодействие CD4/CMH второго класса.
Клетки Cos трансфицируют соответствующим экспрессирующим вектором (кодирующим LAG-3 дикого типа или CD4). Спустя 2 дня клетки Cos обрабатывают трипсином и снова наносят тонким слоем (монослой) по 0,05 • 106 клеток/углубление на планшеты для культивирования ткани с 12 углублениями. Спустя 24 часа меченные с помощью 51Cr клетки DAUDI (5,5•106) инкубируют на этом монослое клеток Cos (конечный объем 1 мл) в течение 1 часа. Дикие B-клетки затем отсасывают и углубления промывают 5-7 раз, добавляя туда медленно по капле 1 мл среды. Края углублений промывают путем аспирации с помощью пипетки Пастера. Оставшиеся клетки лизируют с помощью 1 мл забуференного фосфатом физиологического раствора (ЗФР), с 1% Тритона, в течение 15 минут при 37oC. Лизаты центрифугируют со скоростью 3000 оборотов в минуту в течение 10 минут и получают 100 мкл результирующей надосадочной жидкости.
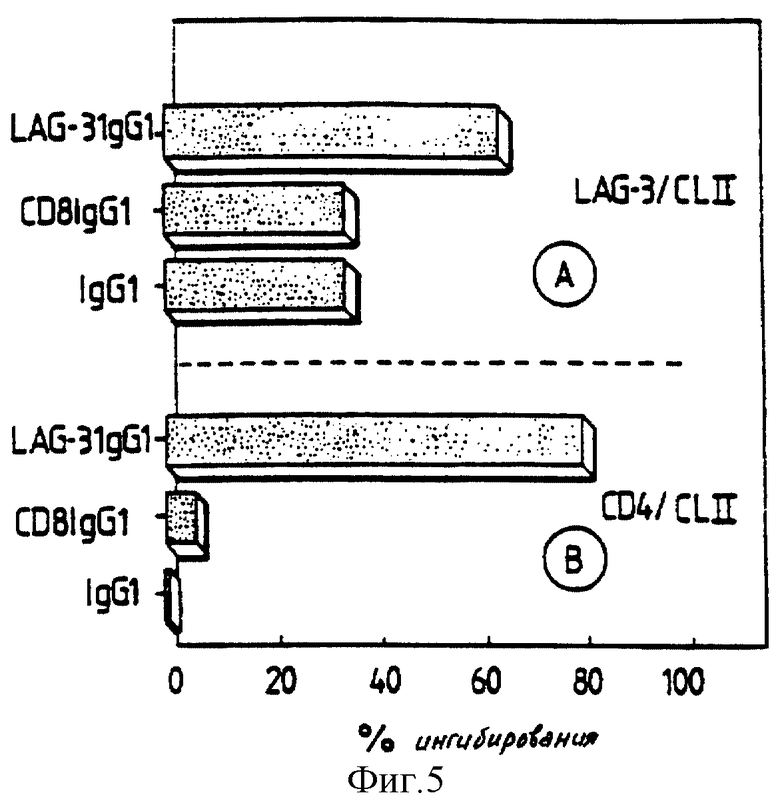
LAG-3 Д1-Д4 Ig используют для ингибирования взаимодействия LAG-3/CMH класса II и CD4/CMH класса II в опыте по связыванию в случае 51Cr. Человеческие CD8 Ig и IgGI испытывают параллельно и используют в качестве отрицательных контролей.
Значительное ингибирование взаимодействия LAG-3/класс II обнаруживают за счет LAG-3 Д1-Д4 Ig (фиг. 5A). Однако, взаимодействие LAG-3/CMH второго класса может быть частично и неспецифически ингибировано с помощью CD8 Ig и человеческих IgGI. С другой стороны, LAG-3 Ig оказывается потенциальным ингибитором взаимодействия CD4/класс II (фиг. 5В) в экспериментальных условиях, где взаимодействие CD4/CMH класса II не модифицировано с помощью CD8 Ig или человеческих IgGI. Это подтверждает тот факт, что взаимодействие LAG-3/класс II более слабое, чем взаимодействие CD4/класс II. Эти результаты служат первым доказательством возможной конкуренции растворимых молекул при взаимодействии CMH второго класса с его лигандами.
Пример 6: Иммунно-супрессивная активность LAG-3 Д1-Д4
Соответствующие опыты проводят с помощью испытаний в отношении пролиферации, описанных выше для биологической активности моноклональных антител против LAG-3.
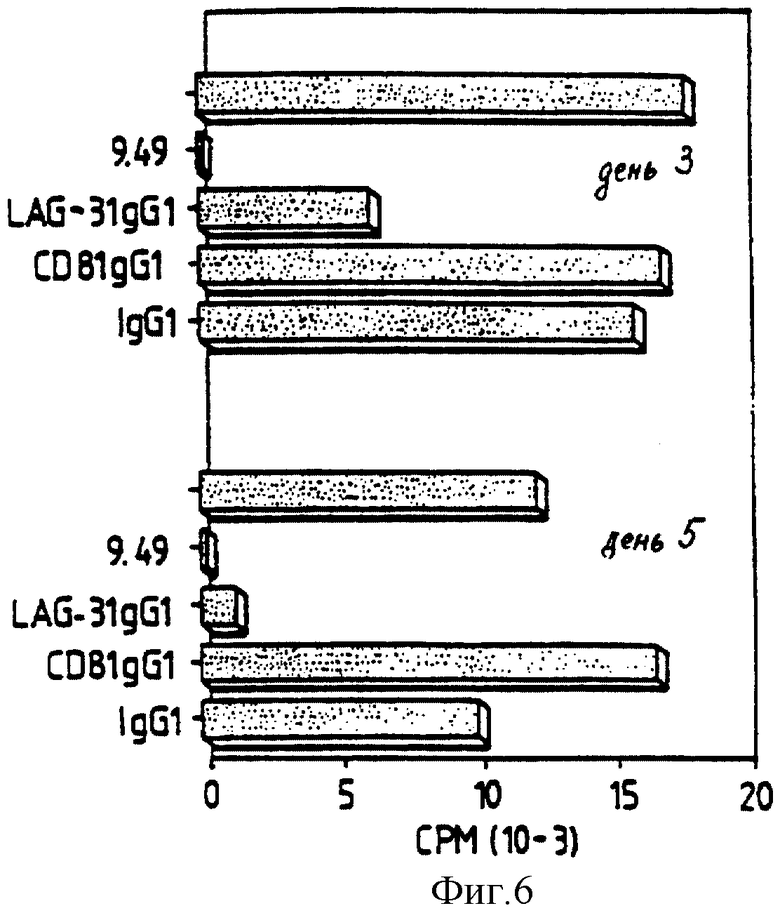
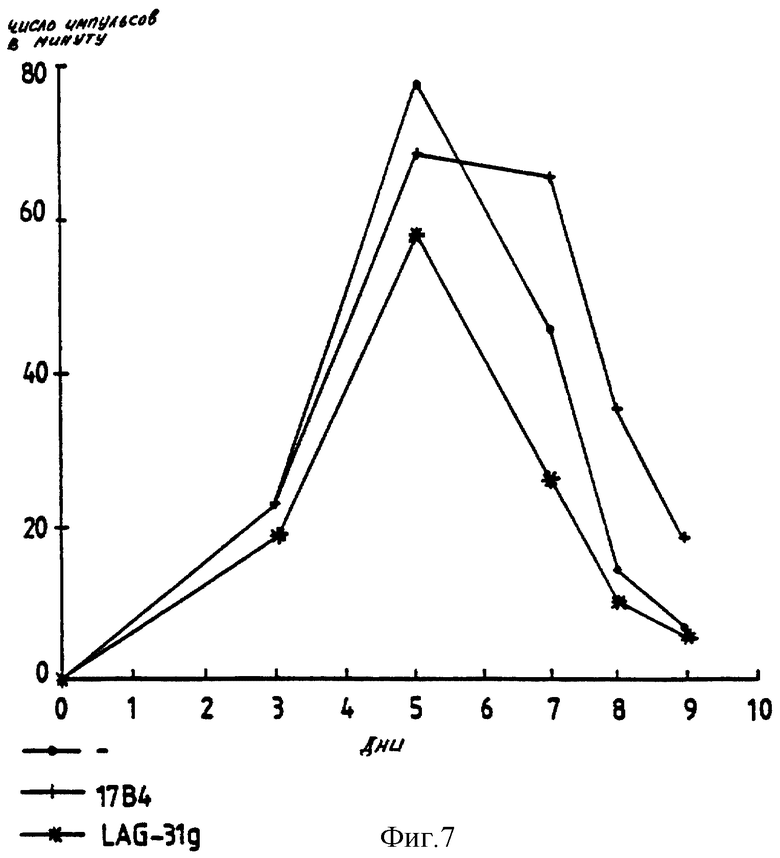
Спустя 3 дня и 5 дней (день 3 и день 5) после антигенной стимуляции, LAG-3 Д1-Д4 Ig показывает сильное ингибирование в отношении пролиферации клона 28, тогда как на человеческие CD8 Ig и IgG не оказывает никакого воздействия (фиг. 6). Подобные эксперименты осуществляют при использовании клона 154 (фиг. 7) и обнаруживают частичное ингибирование в присутствии LAG-3 Ig.
Реализуемый с моноклональными антителами против LAG-3 контроль показывает обратные эффекты, как наблюдалось выше.
Значительное ингибирование пролиферации клеток, инкубированных в присутствии LAG-3 Д1-Д4 Ig, также наблюдается для клона 28.
Эти наблюдения показывают, что LAG-3 Д1-Д4 Ig является потенциальным иммуно-супрессором пролиферации T-лимфоцитов, стимулированных антигеном, и указывают на то, что LAG-3 может действовать как "поглотитель" вторичного иммунного ответа, индуцированного вспомогательными T-лимфоцитами, активированными CD4+.
Роль Lag-3 Ig в отрицательной регуляции иммунных ответов T-клеток
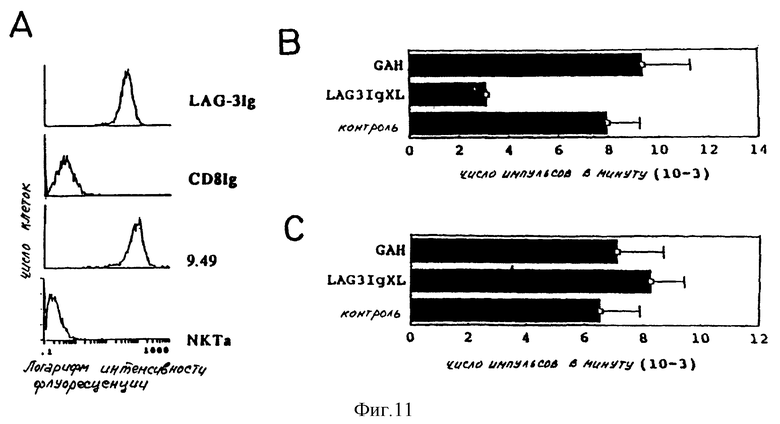
Для того чтобы показать, что растворимая форма LAG-3, имитирующая функции мембранной молекулы, может ингибировать активацию T-клонов CD4 +, стимулированных антигеном, проводят следующие опыты при использовании клона T154: T-клетки предварительно инкубируют с насыщенным количеством LAG-3 Ig (100 нмоль). Клетки затем промывают два раза с помощью холодной RPMI и инкубируют с 10 мкг/мл козьих антител, направленных против человеческих иммуноглобулинов (Tago), при 4oC в течение 30 минут.
После двух новых промывок клетки снова суспендируют в RPMI, содержащий 10% плодной телячьей сыворотки, и инкубируют в течение 2 часов при 37oC перед добавлением "сигнала". Для перекрестного сшивания моноклональных антител используют антимышиные козьи антитела в концентрации 10 мкг/мл (Tago).
На фиг. 11 представлен эксперимент, при котором клон T154 предварительно инкубируют с LAG-3 Ig, связанным ("перекрестно-сшитым") со вторым реагентом (поликлональная специфическая сыворотка для константной области человеческих иммуноглобулинов). Степень фиксации LAG-3 Ig на клетках определяют с помощью иммунофлуоресценции (фиг. 11A). На фиг. 11B показано, что ингибирования более 50% пролиферации клона T154 достигают за счет LAG-3 Ig. В тех же экспериментальных условиях никакого эффекта не наблюдают с контролем CD8 Ig или с LAG-3 Ig без "поперечной сшивки" (на фиг. не представлено).
На фиг. 11C также показано, что никакого эффекта не наблюдают, когда LAG-3 Ig используют для связывания ("поперечное сшивание") молекул второго класса CMH, экспрессированных антигенпредставляющими B-клетками.
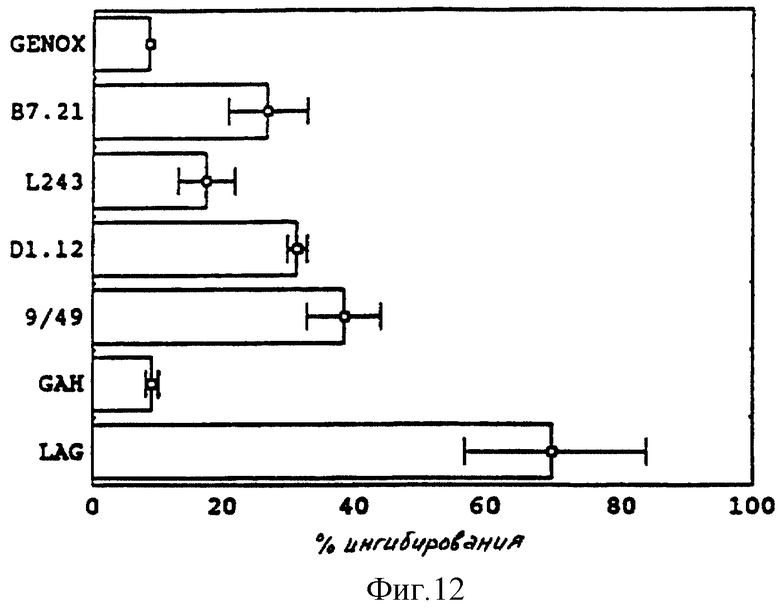
Возможные эффекты связанных ("поперечно-сшитых") моноклональных антител против второго класса на уровне пролиферации T-клеток сравнивают с таковыми LAG-3 Ig. Слабое ингибирование (ниже 50%) наблюдают в случае использования антитела 949 и антитела Д1.12 (анти-DR), связанных с антимышиной козьей поликлональной сывороткой (фиг. 12). Ингибирование пролиферации, следовательно, зависит от эпитопа, причем наиболее значительного эффекта достигают со специфическим эпитопом LAG-3 связи с молекулами класса II.
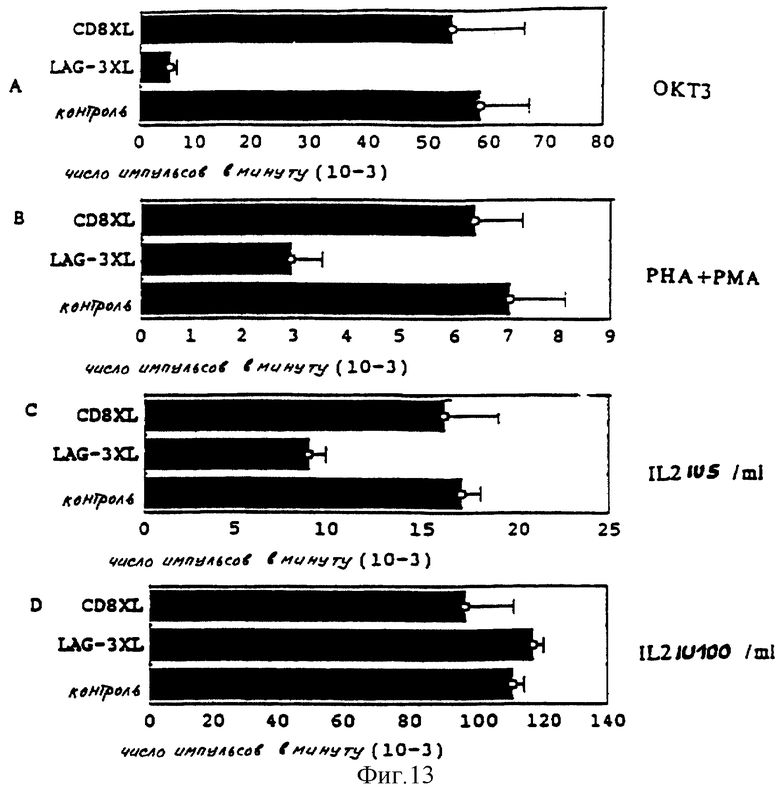
Воздействие LAG-3 Ig на пролиферацию T-клеток также изучают, используя различные сигнальные последовательности, на другом клоне T CD4 +, специфическом клоне TDEL пептида 34-53 основного протеина миелина.
Ингибирование пролиферации наблюдают (n = 2), когда TDEL стимулируют антигеном (не представлено), иммобилизированным ОКТЗ (фиг. 13A), пектинами (PHA /фитогемагглютинин/ + PMA /форболмиристатацетат/) (фиг. 13B) и 5 МЕ/мл интерлейкина-2 (фиг. 13C). Никакого ингибирования не наблюдают со 100 МЕ/мл интерлейкина-2 (фиг. 13D).
В заключение, совокупность этих результатов указывает на то, что LAG-3 и молекулы CMH второго класса, которые представляют собой, каждый из этих факторов, активационные антигены T-клеток, способны к ассимиляции с молекулами-эффекторами, принимающими участие в фазе инактивации ответов T-клеток. Кроме того, эти результаты иллюстрируют значение взаимодействий между T-клетками в контроле иммунного ответа клеток.
Пример 7: Стимуляция клеточной цитотоксичности за счет LAG-3 Ig
Роль LAG-3 Ig в отношении клеточной цитотоксичности изучают при использовании двух типов эффекторных клеток:
- свежевыделенные человеческие лимфоциты периферической крови (PBL);
- клетки линии SIB5 (клон человеческих NK-клеток /естественных киллерных клеток/).
Цитотоксическую активность этих клеток определяют путем подсчета 51Cr, содержащегося в среде за счет предварительно меченых целевых клеток, в присутствии или в отсутствие в среде LAG-3 Ig.
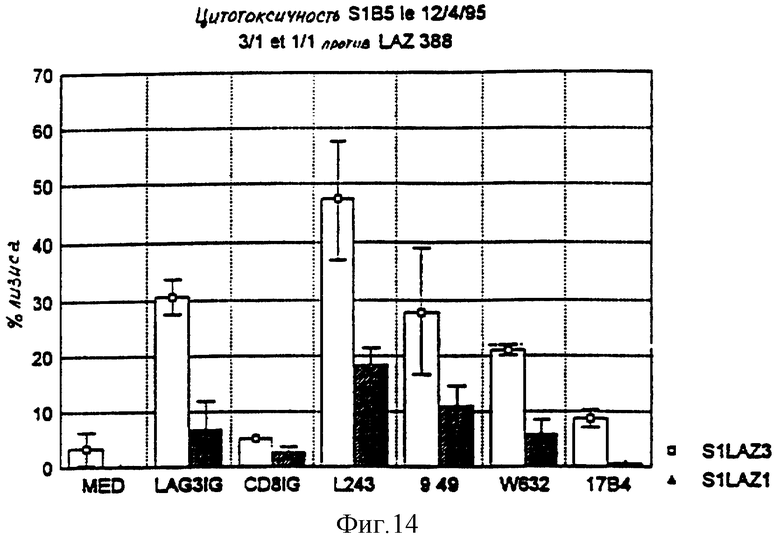
На фиг. 14 показана степень цитотоксичности SIB5 для линии человеческих B-клеток, трансформированных вирусом Эпштейна-Барра и несущих антигены первого и второго классов главного комплекса гистосовместимости (линия LAZ 388), в зависимости от различных реагентов, добавляемых к культурам.
Измерения осуществляются спустя 4 часа после сокультивирования для соотношений эффекторные клетки/целевые клетки (SIB5/LAZ 388) 3: 1 (белые колонки) или 1: 1 (заштрихованные колонки).
Отрицательные контроли представляют собой: одна среда (MED), иммуноадгезин CD8 Ig и моноклональное антитело 17. B4 (анти-LAG-3).
Положительные контроли образованы тремя различными моноклональными антителами:
- антитело L243, направленное против антигенов класса II DR;
- антитело 9.49, направленное против антигенов класса II DR, DP, DO;
- антитело W632, направленное против антигенов класса I главного комплекса гистосовместимости человека.
Антитела против главного комплекса гистосовместимости человека класса I (W632) или класса II (1.243) увеличивают лизис целевых клеток (но не контроля 17B4). Иммуноадгезин LAG-3 Ig повышает лизис; контроль CD8 Ig не оказывает воздействия.
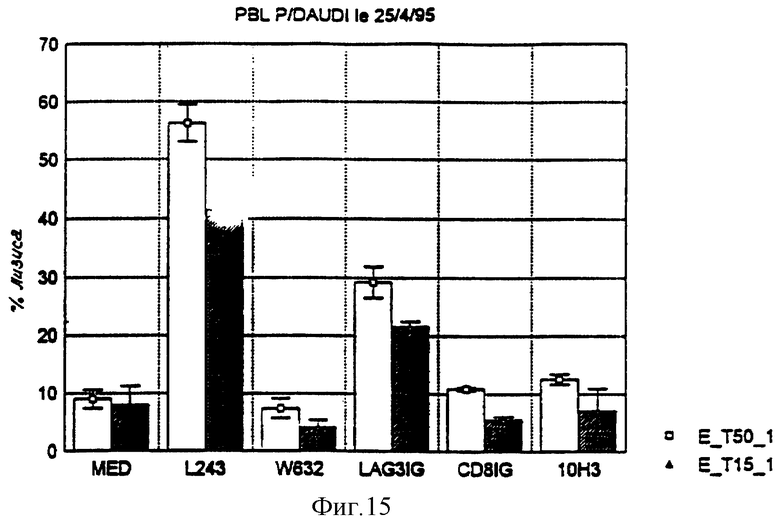
На фиг. 15 представлены результаты эксперимента, аналогичного предыдущему, при котором измеряют цитотоксичность PBL по отношению к клеткам DAUDI (главный комплекс гистосовместимости человека класса I-) для соотношения эффекторы/цели 50: 1 (белые колонки) и 15: 1 (заштрихованные колонки). Добавляемыми в среду реагентами являются те же, что и реагенты, используемые в первом эксперименте, без антитела 9.49 и антитела 17. B4. Антитело 10H3 представляет собой специфический иммуноглобулин изотипа IgGI поверхностного антигена CD45. Его используют в качестве отрицательного контроля.
Никакого изменения не наблюдают при использовании антитела, направленного против антигенов класса I главного комплекса гистосовместимости (W632).
Данные этих двух серий измерений указывают на то, что, по сравнению с отрицательными контролями, LAG-3 Ig активирует цититоксичность NK-клеток. Этот эффект подобен таковому, наблюдаемому с антителами, направленными против молекул второго класса CMH.
Литература
1. TRIEBEL T. et al. , 1990, J. Exp. Med. 171, 1393-1405.
2. BAIXERAS E. et al. , 1992, J. Exp. Med. 176, 327-337.
3. COSGROVE D. et al. , 1991, Cell 66, 1051-1066.
4. RAHEMTULLA A. et al. , 1991, Nature 353, 180-184.
5. TRAUNECKER A. et al. , 1988, Nature 331, 84-86.
6. BENEDICT A. A. et al. , 1967, Methods in Immunology 1, 197-306 (1967).
7. YELTON D. E. et al. , Ann. Rev. of Biochem. 50, 657-680 (1981).
8. HUARD B. et al. , Immunogenetics 39: 213.
9. MANIATIS T. et al. (1982), Molecular cloning: A laboratory manual - Cold Spring Harbor Laboratory, New-York.
10. SEED B. , 1987, Nature 329, 840-842.
11. COLE S. C. et al. Biotechnology II, 1014-1024, 1993.
12. COLE S. C. et al. Biotechnology II, 1014-1024, 1993.






















Изобретение относится к медицине и биотехнологии и включает растворимые полипептидные фракции протеина LAG-3, способ получения, терапевтическую композицию и антитело. Сущность изобретения включает растворимые полипептидные фракции мембранного протеина LAG-3, пригодные для использования в качестве иммуносупрессоров, способ их получения, а также антитела, способные предотвращать специфическую фиксацию протеина LAG-3 на молекулах СМН (главного комплекса гистосовместимости) класса II, для применения в качестве иммуностимуляторов. Преимущество изобретения заключается в том, что заявляемые пептиды обладают способностью связываться с молекулами СМН второго класса и обладать иммуноподавляющим действием, а также неожиданным иммуно-стимулирующим действием анти-СД4-антител. 6 с. и 8 з. п. ф-лы, 15 ил. , 5 табл.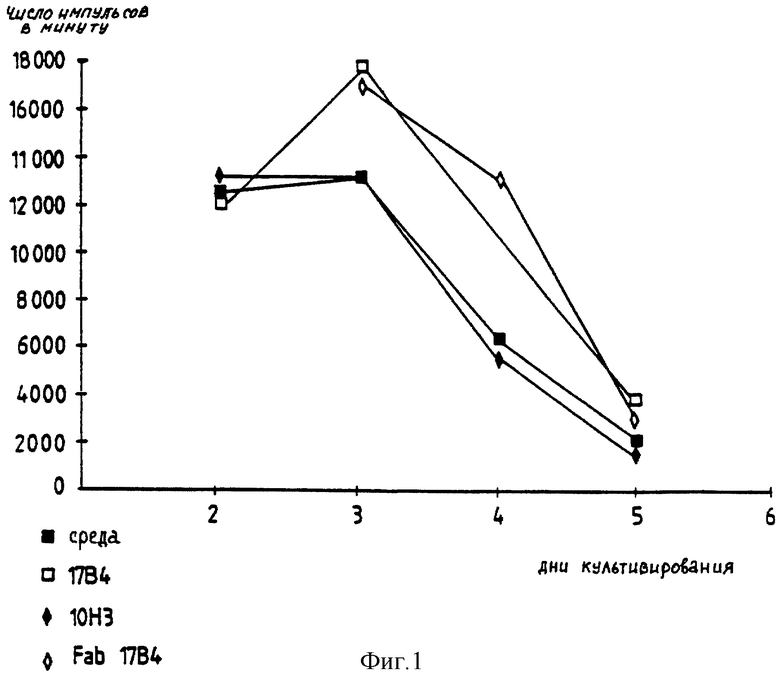
| Автоматический огнетушитель | 0 |
|
SU92A1 |
| Огнетушитель | 0 |
|
SU91A1 |
| LINSLEY et al., T-cell antigen CD-28 mediated adhesion with B cell by interacting with activation antigen B7/BB-1, Proc | |||
| of Natl | |||
| Acad | |||
| Sci USA, 1990, v.87, № 8, p.5031-5035 | |||

