Изобретение относится к новым бактериальным штаммам и их использованию в качестве средств или источника средств, противодействующих росту микроорганизмов, вызывающих гниль. Конкретно, настоящее изобретение представляет новые изоляты бактерий видов Pseudomonas fluorescens, Serratia liquefaciens, Serratia plimuthica, Bacillus subtilis, Bacillus pumilia, Bacillus polymyxa, которые особенно эффективны для подавления роста микроорганизмов, вызывающих заболевания плодов после сборки урожая, причиной которых являются грибы Botrytis cinerea и Alternaria brassicicola. Кроме того, представлены антибиотики, полученные из видов Bacillus.
Потери после сбора урожая во время хранения сельхозпродуктов растительного происхождения вызываются, между прочим, потерей воды, увяданием (старением) листьев, проростом и гниением, последнее, в частности, вызывается грибами и бактериальными возбудителями. Такие потери после сбора урожая могут быть значительно снижены путем хранения при низких температурах (например 1 - 20oC) и высокой относительной влажности (например, 95%) (Robinson et al., 1975). При этих условиях хранения B. cinerea является преобладающим видом грибов, обнаруживаемым на сохраняемой капусте, и главной причиной потерь (Geeson, 1978, Brown et al., 1975); причем он является условно-патогенным возбудителем для широкого ряда листовых овощей, поражающим ослабленные, пораненные или состарившиеся листовые ткани, и также известно, что он поражает различные фрукты (смотрите патент США 5041384). Здоровые листовые ткани, однако, как описано, высокорезистентны к поражению Botrytis (New-hook, 1951). Исходная устойчивость подвергаемых воздействию наружных листьев также, вероятно, является причиной того, что обнаружение гнили капусты, вызванной Botrytis, обычно начинается через 2 - 3 месяца хранения на холоде и часто ограничивается наружными подсохшими, состарившимися листьями капусты (Wale 1980).
Для предупреждения грибковой порчи обычной практикой во многих странах является опрыскивание капусты на поле системными фунгицидами и погружение собранных кочанов капусты в растворы фунгицидов перед хранением (Brown et al. , 1975). Так как все больше и больше раскрываются онкогенные свойства многих из наиболее часто используемых фунгицидов и из-за того, что устойчивость большинства фунгицидов повышается при хранении при низких температурах, использование фунгицидов приобретает возрастающее значение. К тому же было сообщено об устойчивости к фунгицидам (Spotts & Cervantes 1986), и подавление главного вызывающего порчу микроорганизма B. cinerea фунгицидами, такими как беномил, как было показано, приводит в результате к повышенному обсеменению (размножению) A. brassicicola, который вызывает более глубокую гниль кочанов капусты, чем B.cinerea (Wale & Epton).
Борьба с грибковыми возбудителями после сбора урожая с помощью бактерий и дрожжей-антагонистов, таких как Bacillus subtilis, Pseudomonas cepacia, Pseudomonas syringae, Enterobacter aerogenes, Enterobacter cloacae and Debaryomyces hansenii была описана для ряда сохраняемых овощей и фруктов, включая яблоки, абрикосы, вишни, цитрусовые, виноград, нектарины, персики, груши, перец, хурму, сливы, картофель и помидоры (смотрите Wilson & Wisniewski, 1989, по недавнему обзору).
Патент США 4,764,271 (кл. A 01 N 63/00, 1988) описывает отдельный штамм Bacillus subtilis, который, как было обнаружено, может использоваться для ингибирования роста коричневой плесени на фруктах. Свободный от клеток фильтрат, полученный из культуральной ростовой среды, проявил антибиотическую активность.
Аналогичный штамм Bacillus subtilis известен также из документа SU 1,706,504 (кл. A 01 N 63/00, 1992).
Однако в большинстве исследований изучалось антагонистическое действие только при температурах около 20oC, а не при температурах между 1 и 10oC, которые используются при коммерческом хранении большинства фруктов и овощей. К тому же в очень немногих публикаций указывается, что были испытаны персистенция антагонистов на растительных поверхностях или сравниваемых микробных популяций на сохраняемых органах растений с антагонистической микрофлорой.
Настоящее изобретение представляет новые бактериальные изоляты видов, которые обладают свойствами подавления роста видов грибов на продуктах после сбора урожая; этими видами являются виды Pseudomonas, Serratia и Bacillus. Наибольшее преимущество в том, что изоляты Pseudomonas и Serratia данного изобретения обладают свойством возможности подавления роста грибков при температурах сохранения на холоде, например, ниже 10oC, обычно от 0 до 4oC. Кроме того, изоляты Bacillus обладают свойством выделения антибиотически активной фракции, также представленной в изобретении, которую саму по себе можно выделить в чистом или полуочищенном виде и использовать для подавления роста таких видов грибов.
Нужно осознать, что изоляты данного изобретения были выбраны из исходной популяции из многих тысяч подобных штаммов, которые неэффективны. Однако из представляемого доступа к помещенным на хранение штаммам, упомянутым выше, и данных и протоколов, относящихся к этому, представленных ниже, опытным специалистам будет понятно, что возможно будет выделить дополнительные родственные штаммы со сходными антагонистическими свойствами, и поэтому объем данного изобретения охватывает также родственные штаммы, насколько в таком случае может быть выяснено путем сравнивания с представленными штаммами и данными и протоколами, представленными здесь.
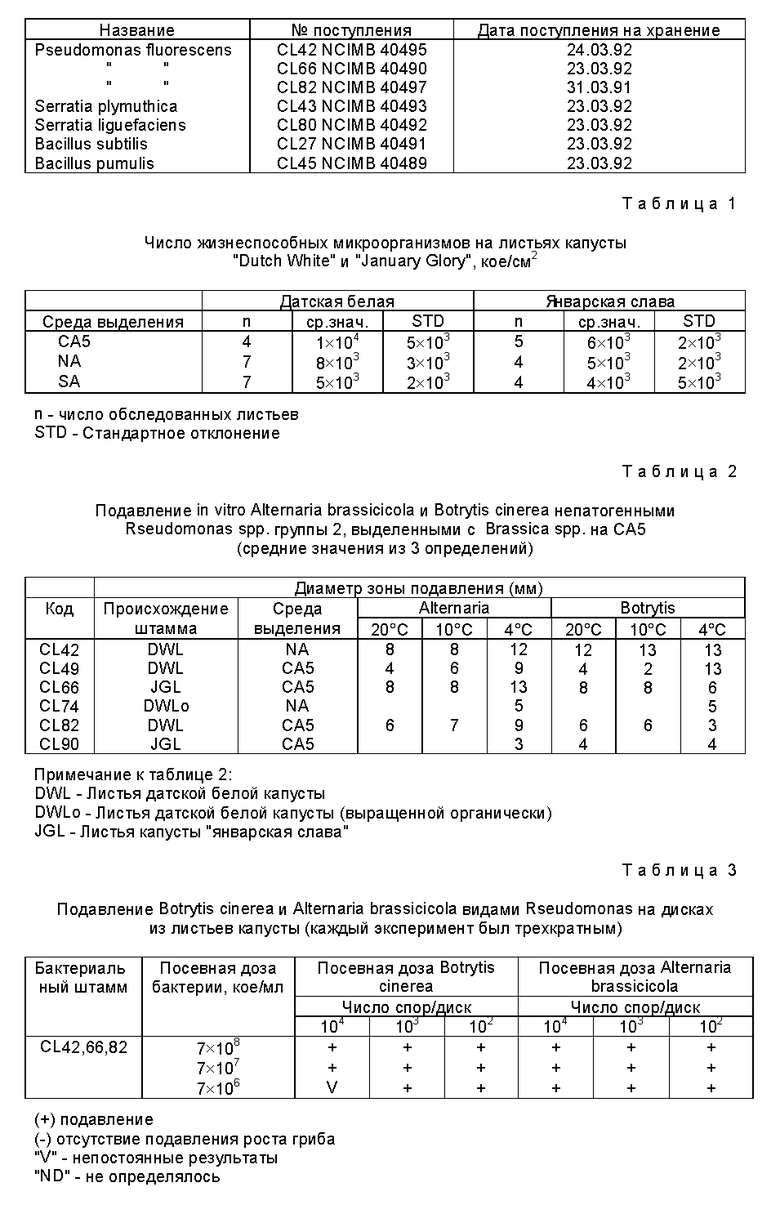
Образцы новых бактериальных штаммов данного изобретения были помещены на хранение по Будапештскому договору о международном признании помещения на хранение микроорганизмов в целях патентных процедур в Национальную коллекцию промышленных и морских бактерий (UK National Collection of Industrial and Marine Bacteria) 23 St Machar Drive, Aberden, AB2, IRY, UK, под номерами поступлений, приведенными в конце описания.
Настоящее изобретение представляет выделенные бактерии, имеющие идентифицирующие характеристики изолятов, выбираемых из группы, состоящей из NCIMB 40489, NCIMB 40490, NCIMB 40491, NCIMB 40492, NCIMB 40493, NCIMB 40494, NCIMB 40495 и NCIMB 40497. Такие изоляты обладают общими характеристиками подавления роста грибов видов A. brassicicola и/или B.cinerea на тканях капусты: на агаре: в водных смесях.
Особенно предпочтительный аспект данного изобретения представляет бактериальный изолят, имеющий идентифицирующие характеристики штамма, выбираемого из группы, состоящей из NCIMB 40490, NCIMB 40492, NCIMB 40493, NCIMB 40495 и NCIMB 40497, причем указанный изолят способен подавлять рост A. brassicicola и/или B. cinerea на листьях капусты при температуре от 20oC или менее, более предпочтительно при 10oC или менее, наиболее предпочтительно при 4oC или менее. Более предпочтительно, чтобы изолят относился к видам, выбираемым из группы, состоящей из Pseudomonas fluorescens, Serratia plymuthica, Serratia liquefaciens.
Другой предпочтительный аспект данного изобретения представляет бактериальный изолят, имеющий идентифицирующие характеристики штамма, выбираемого из группы, состоящей из NCIMB 40489 и NCIMB 40491, причем указанный штамм способен подавлять рост A. brassicicola и/или B.cinerea на листьях капусты при температурах около 20oC. Более предпочтительно, что изолят способен к продукции антибиотической фракции при культивировании в бульоне из гомогенизированных листьев капусты с концентрацией 50 г или более на литр воды, причем указанный антибиотик способен подавлять рост указанных A. brassicicola и/или B.cinerea на листьях капусты при температурах около 20oC.
Дополнительный аспект этого изобретения представляет водные суспензии бактерий из одного или более бактериальных штаммов этого изобретения, причем предпочтительные суспензии содержат достаточное количество кое/мл для подавления роста грибов, имеющих отношение к вышеперечисленным фруктам и/или овощам, которые погружались в нее, и, кроме того, биологически чистые культуры таких бактерий при высоком числе кое/мл для приготовления суспензий для такого погружения.
Данное изобретение тем самым дополнительно представляет способ предупреждения грибковых заболеваний после сбора урожая овощей и/или фруктов, включающий погружение указанных продуктов в суспензию бактерий, которые описаны выше. Предпочтительно суспензия будет содержать от 104 и до 109, более предпочтительно между 105 и 108 кое/мл. При использовании группы не-Bacillus предпочтительным выбором является хранение на холоде.
Еще один дополнительный аспект этого изобретения представляет антибиотики, получаемые путем культивирования выделенных бактерий типа NCIMB 40489 и/или NCIMB 40491 в среде, содержащей гомогенизированные капустные листья и воду в соотношении 5 г до 100 г/литр. Кроме того, представлены композиции, отличающиеся тем, что они включают такие антибиотики в качестве активных ингредиентов.
Теперь будут приведены примеры штаммов, их эффективность против роста грибов, способы отбора подобных таким изолятов и способы их использования при применении этого способа в производственном масштабе только для иллюстрации путем обращения к следующим чертежам и примерам.
Фигура 1. Диаметр зоны подавления (мм) вокруг колоний бактерий на капустном агаре, содержащем различные концентрации тканей капусты, засеянном A.brassicicola и B.cinerea. ▪ = CL42;  = CL66;
= CL66;  = CL82.
= CL82.
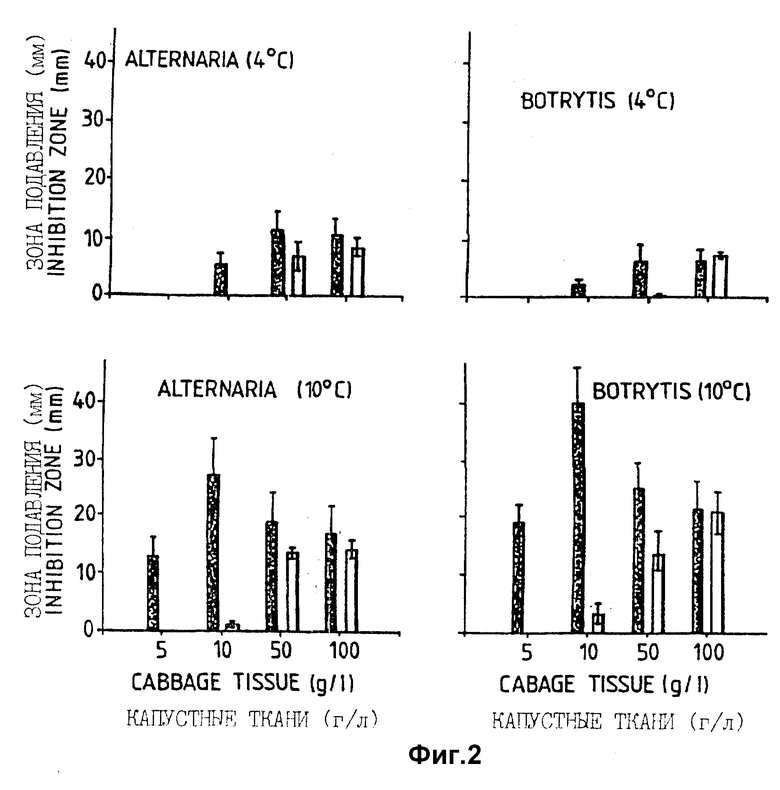
Фигура 2. Средний диаметр зоны подавления (мм) (вертикальные линии = стандартное отклонение) вокруг колоний бактерий на капустном агаре, содержащем различные концентрации тканей капусты, засеянном A.brassicicola и B.cinerea (среднее из 3 определений на изолят). ▪ = Serratia plymuthica (CL43);  = Serratia liquefaciens, CL57; изоляты 58, 59, 61, 62 и 80 продемонстрировали сходные результаты.
= Serratia liquefaciens, CL57; изоляты 58, 59, 61, 62 и 80 продемонстрировали сходные результаты.
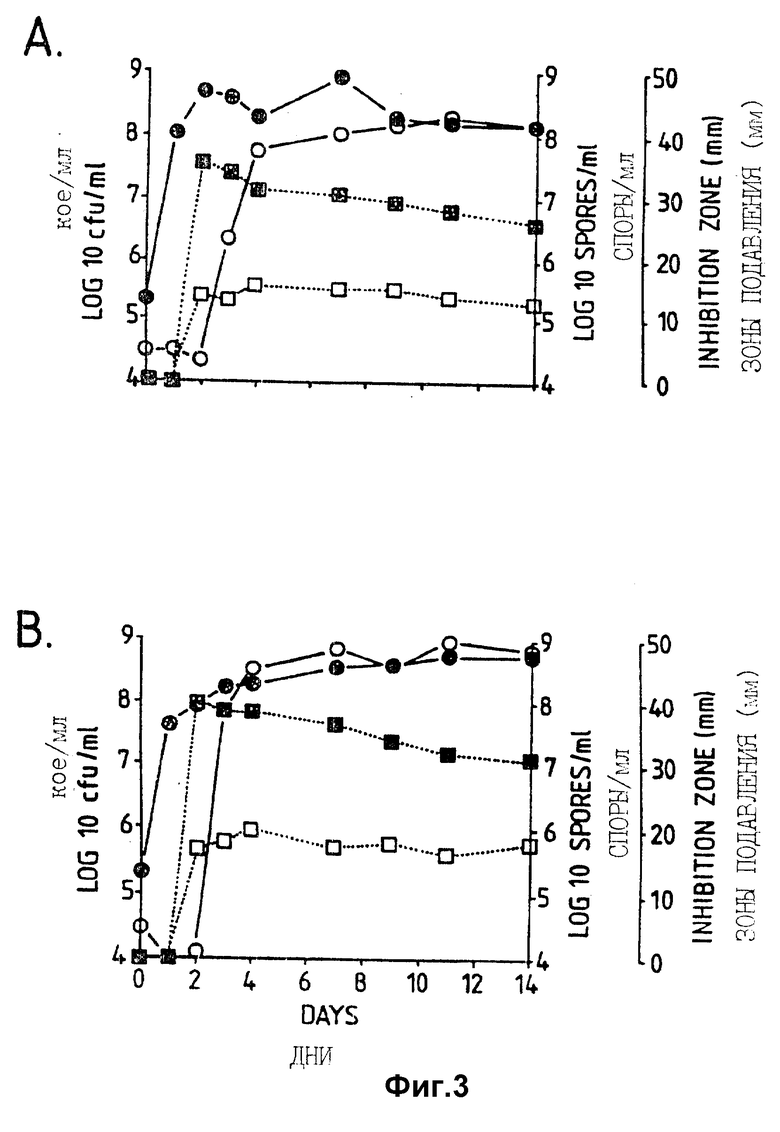
Фигура 3. Рост штамма CL27 Bacillus subtilis, образование спор и проявление антагонистической активности против A.brassicicola и B.cinerea в капустном бульоне (A: CB5, B: CB10) в серийной культуре. противогрибковая активность определялась на чашках Sens-acute, содержащих засеянную грибком CA5. 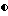 = кое;
= кое;  = споры; ▪ = активность против B.cinerea;
= споры; ▪ = активность против B.cinerea;  = активность против A.brassicicola.
= активность против A.brassicicola.
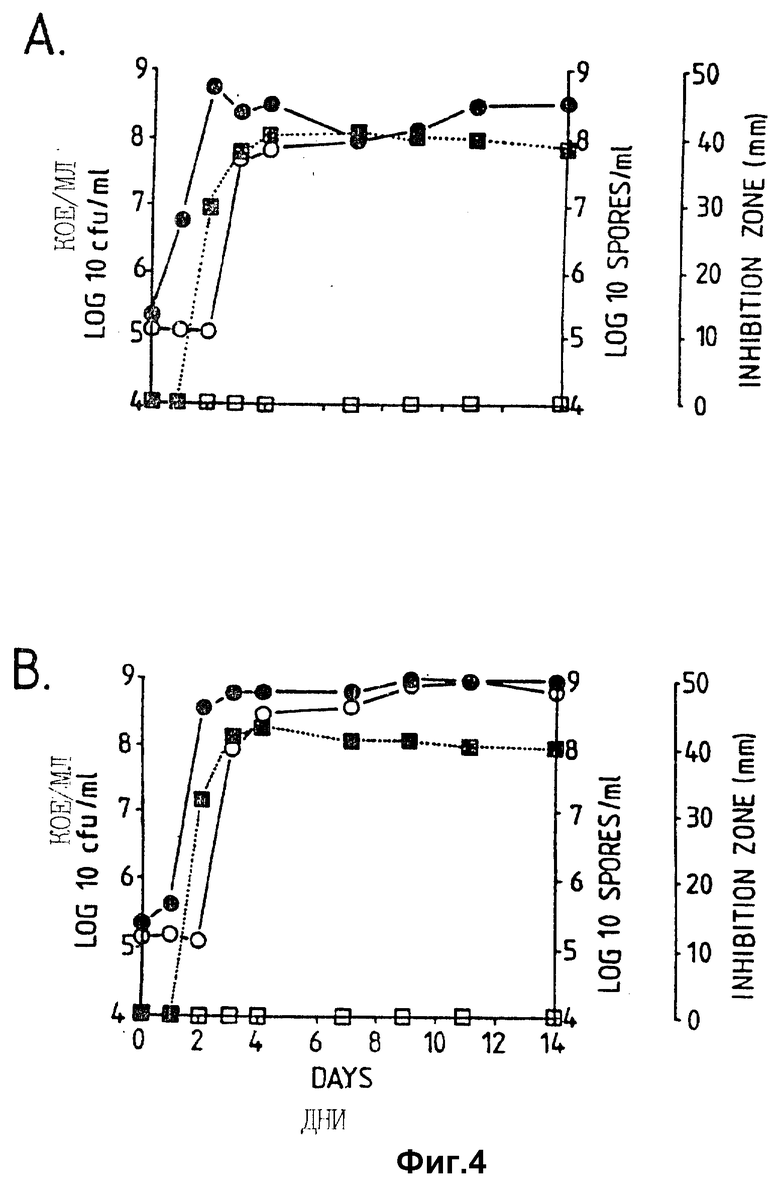
Фигура 4. Рост штамма CL45 Bacillus pumilus, образование спор и проявление антагонистической активности против A.brassicicola и B.cinerea в серийной культуре в капустном бульоне (A: CB5, B:CB10). Противогрибковая активность определялась на чашках Sens-acute, содержащих засеянную грибком CA5. 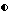 = кое,
= кое,  = споры; ▪ = активность против B.cinerea;
= споры; ▪ = активность против B.cinerea;  = активность против A.brassicicola.
= активность против A.brassicicola.
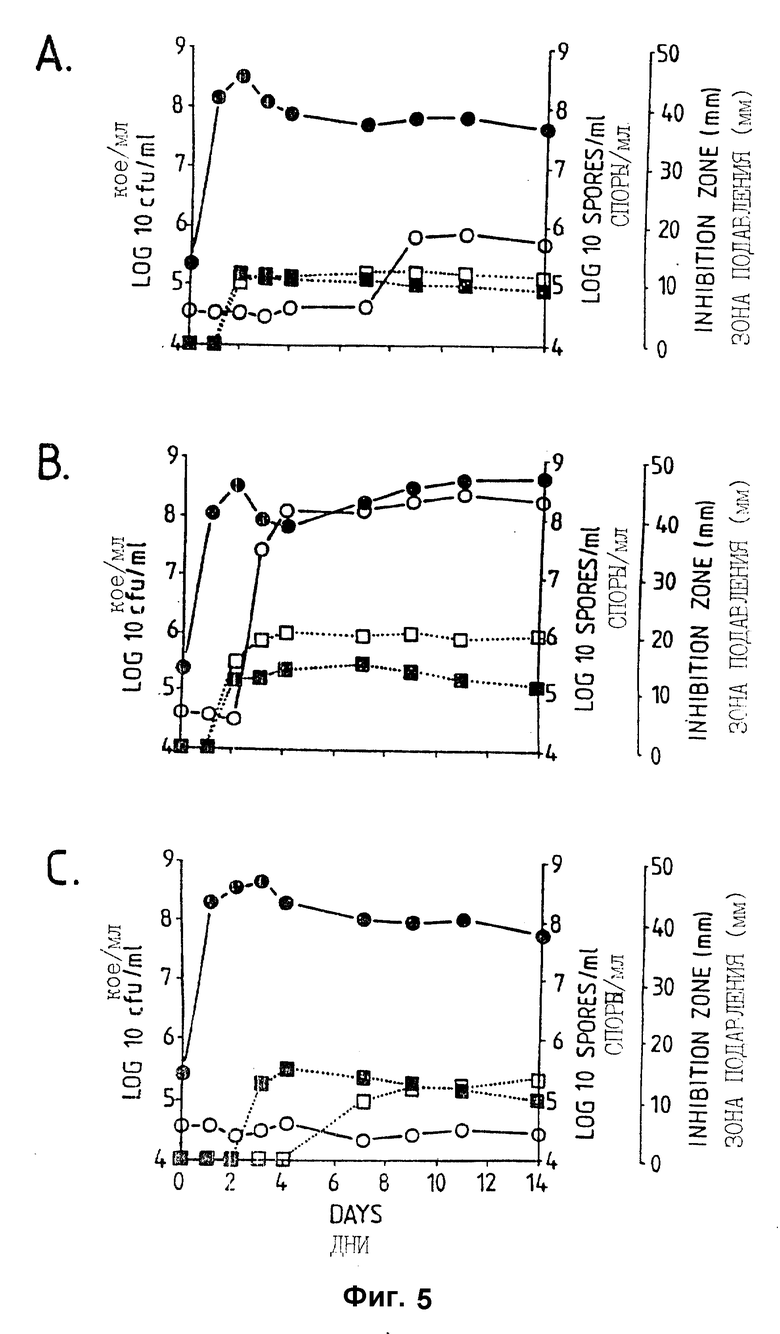
Фигура 5. Рост штамма Bacillus subtilis CL27, образование спор и проявление антагонистической активности против A.brassicicola и B.cinerea в серийной культуре в питательном бульоне (A: NB, B:NB + Mg, C:NB + 1% глюкозы). Противогрибковая активность определялась на чашках Sens-acute, содержащих засеянную грибком CA5.  = кое;
= кое;  = споры; ▪ = активность против B.cinerea;
= споры; ▪ = активность против B.cinerea;  = активность против A.brassicicola.
= активность против A.brassicicola.
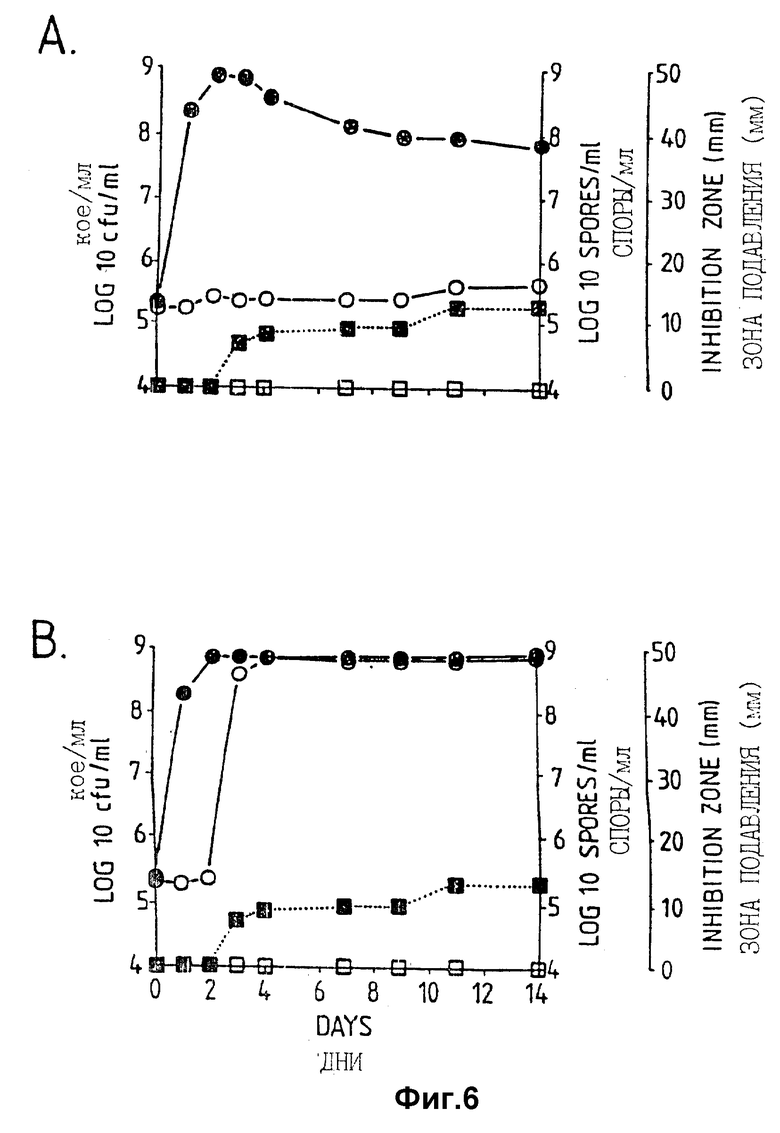
Фигура 6. Рост штамма Bacillus pumilis CL45, образование спор и проявление активности против A. brassicicola и B.cinerea в серийной культуре в питательном бульоне (A:NB, B:NB + Mg). Противогрибковая активность определялась на чашках Sens-acute, содержащих засеянную грибком CA5. 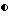 = кое;
= кое;  = споры; ▪ = активность против B.cinerea;
= споры; ▪ = активность против B.cinerea;  = Активность против A.brassicicola.
= Активность против A.brassicicola.
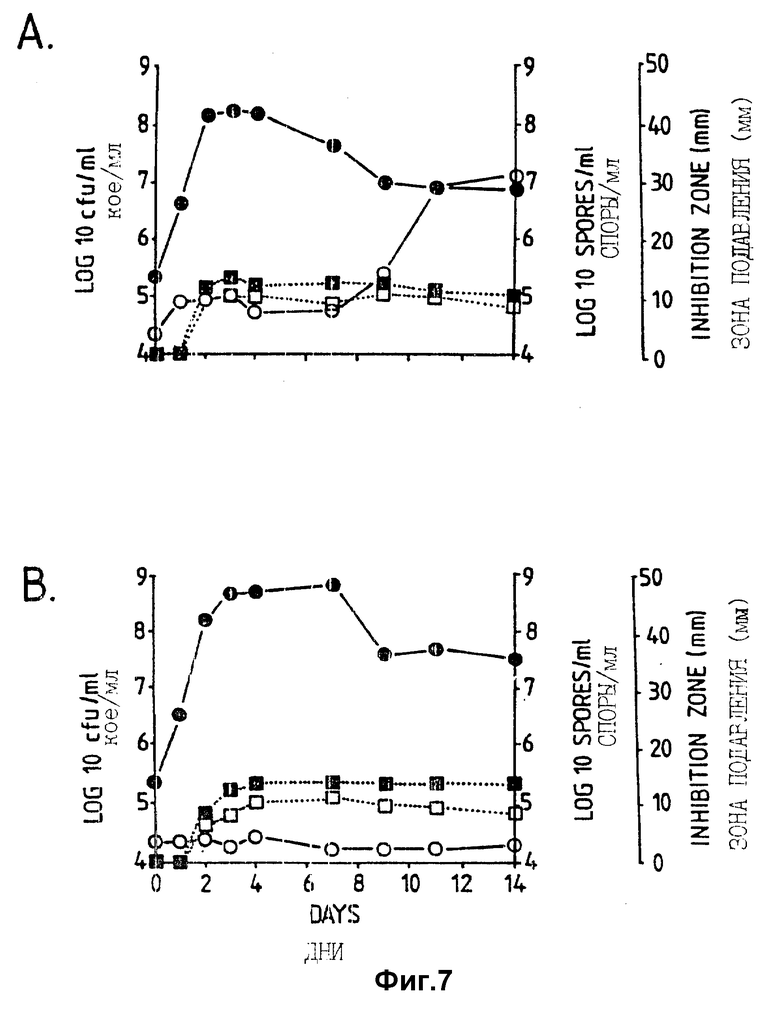
Фигура 7. Рост штамма CL27 Bacillus subtilis, образование спор и проявление активности против A.brassicicola и B.cinerea в серийной культуре в синтетической солевой среде (A:DM, B:DM + 1% глюкозы). Противогрибковую активность определяли на чашках Sens-acute, содержащих засеянную грибком CA5. 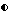 = кое;
= кое;  = споры; ▪ = активность против B.cinerea;
= споры; ▪ = активность против B.cinerea;  = Активность против A.brassicicola.
= Активность против A.brassicicola.
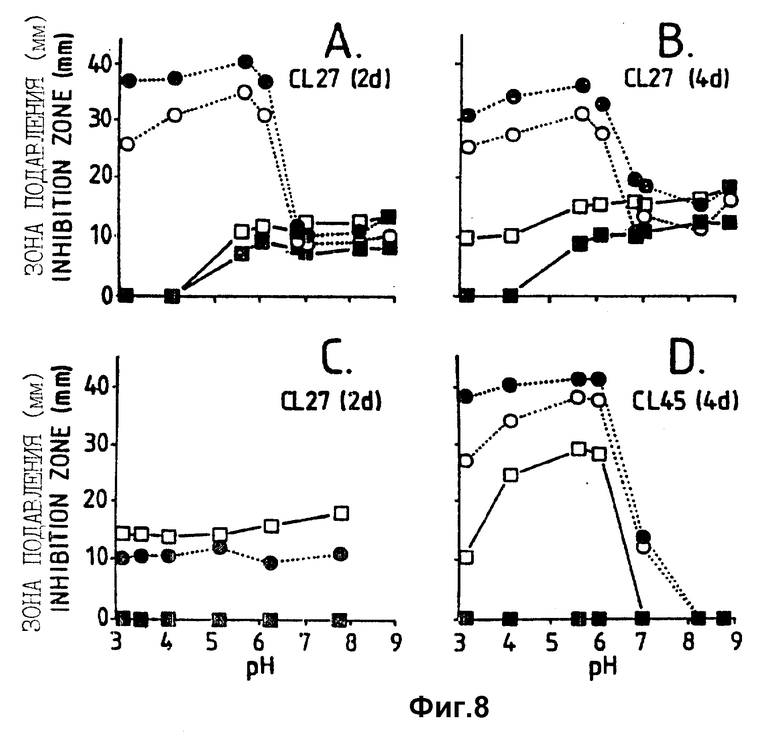
Фигура 8. Воздействие pH среды на активность антибиотиков, из штаммов Bacillus, CL27 и CL45, культивированных серийно и экстрагированных через 2 дня (A и C) или 4 дня (B и D) после посева. В A, B и D подавление возбудителя определяли на чашках Sens-acute для микроопределения, содержащих среду CA5, засеянную B. cinerea, в C - на PDA. Среда для серийной культуры:  = CB10;
= CB10;  = CB5; ▪ = NB;
= CB5; ▪ = NB;  = NB + M.
= NB + M.
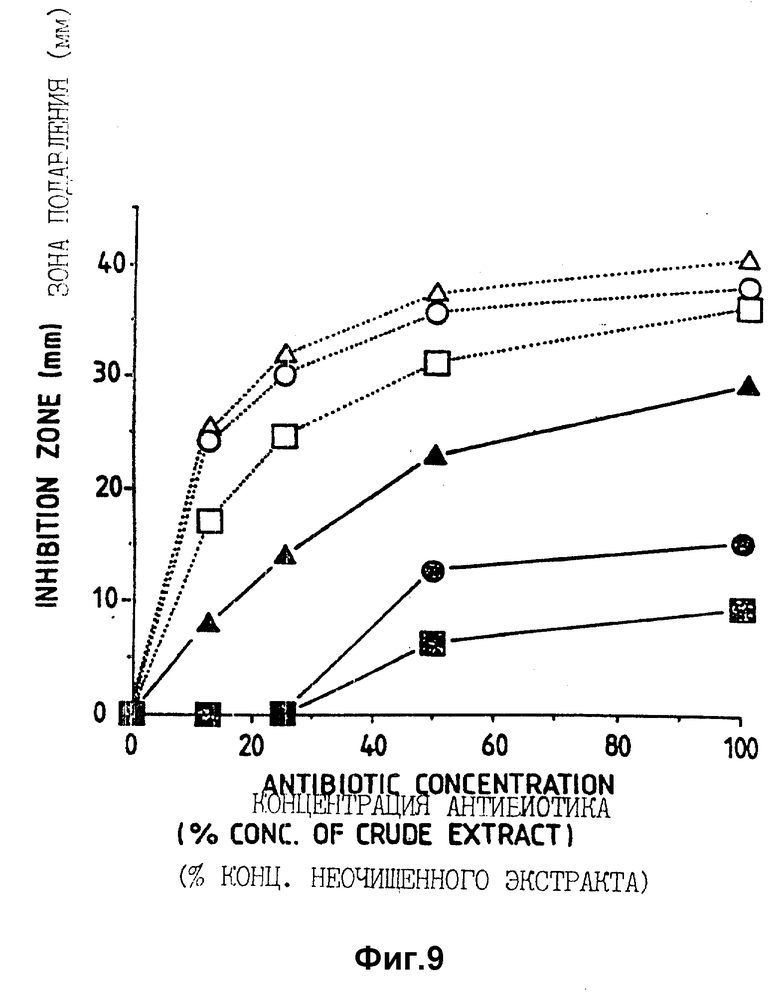
Фигура 9. Воздействие типа питательной среды и концентрации в исследуемой среде (бульон с тканями капусты или картофельной декстрозой, уплотненный 10 г/л агара) на размер образуемых зон подавления. Среду засевали спорами B.cinerea. Использованный неочищенный экстракт собирали через 3 дня от серийных культур штамма CL27 B.subtilis в CB5. Различные концентрации антибиотиков получали путем разбавления неочищенного экстракта (канц. 100%) стерильной CB5. Использованные исследуемые среды:  = 50 мг/мл;
= 50 мг/мл;  = 100 мг/мл;
= 100 мг/мл;  = 200 мг/мл гомогенизированной ткани капусты;
= 200 мг/мл гомогенизированной ткани капусты;  = 1/4 концентрации PDB;
= 1/4 концентрации PDB;  = 1/2 концентрации PDB; ▪ = полная концентрация PDB (картофельный декстрозный бульон).
= 1/2 концентрации PDB; ▪ = полная концентрация PDB (картофельный декстрозный бульон).
Пример 1. Выделение антагонистических штаммов из тканей капустного листа.
Квадрат 10х10 см асептичсеки вырезался из самого наружного листа качана капусты и помещался в пластиковый мешочек с 25 мл стерильного раствора Рингера. Затем ткань помещали в stomacher на 10 минут для удаления поверхностных микроорганизмов. Соответствующие разведения засевали на чашки с питательным агаром, капустным агаром (гомогенизированная ткань капустного листа или 10 г (CA1), 20 г (CA2), 50 г (CA5) или 100 г (CA10) и 10 г агара на литр воды), с капустным агаром, засеянным спорами Botrytis (как описано ниже), агаром Сабуро (Oxoid) и картофельным декстрозным агаром (Oxoid). Антагонисты обнаруживали с помощью зон подавления, образованных или когда листовые смывы непосредственно высевали на чашках с агаром, засеянным Botrytis, или после репликации колоний на чашки с агаром, засеянным Botrytis.
Идентификация основной микрофлоры на капустных листьях.
Число колоний нитчатых грибов подсчитывали непосредственно на чашках для выделения. Чтобы установить число дрожжей, флюоресцирующих псевдомонад и Enterobacteriaceae, с чашек для выделения по случайной выборке снимали 140 колоний и идентифицировали. Для идентификации дрожжей и излучения подвижности бактерий использовали фазово-контрастную микроскопию. Так как все обнаруженные бактерии были грамотрицательными, изоляты, демонстрирующие подвижность и флюоресценцию на среде Кинга, B, классифицировали как флюоресцирующие псевдомонады, а те, которые росли в анаэробных условиях (High and Leifson, 1953), как Enterobacteriaceae. Другие бактериальные изоляты не были идентифицированы.
Число спор Bacillus устанавливали по числу колоний, образовавшихся после 20-кратного концентрирования промывных вод листа путем центрифугирования при 3600 об/мин и прогревания (80oC в течение 20 мин).
Идентификация бактерий-антагонистов.
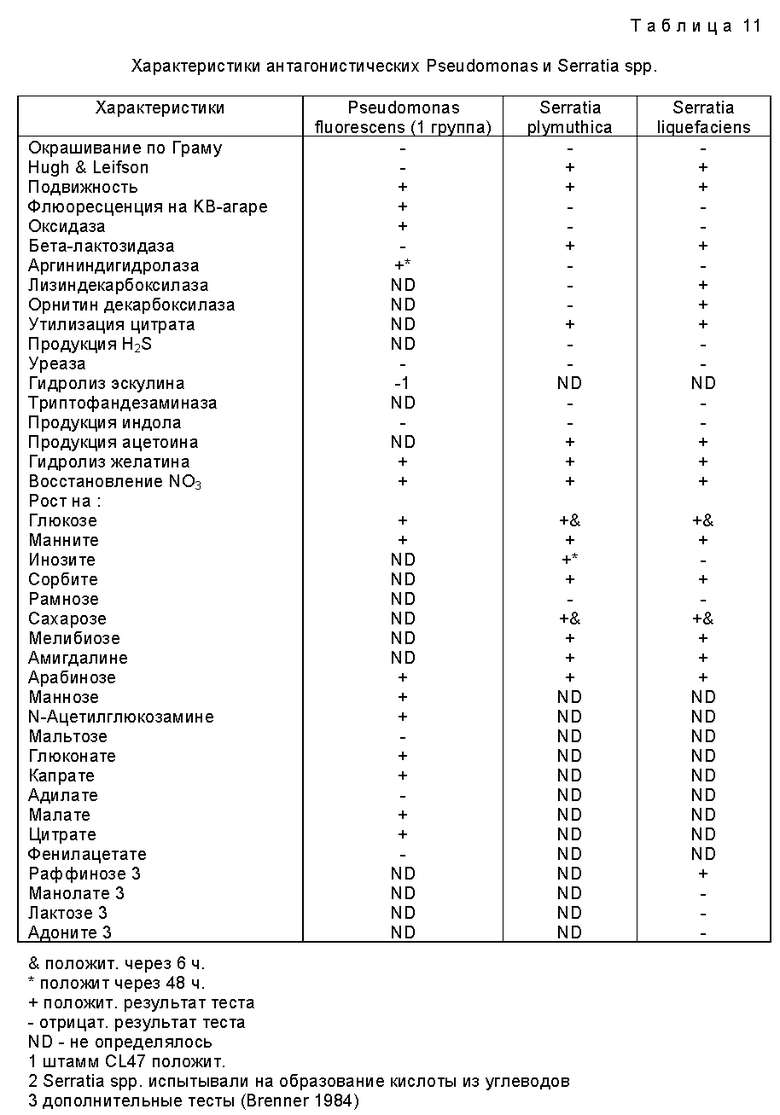
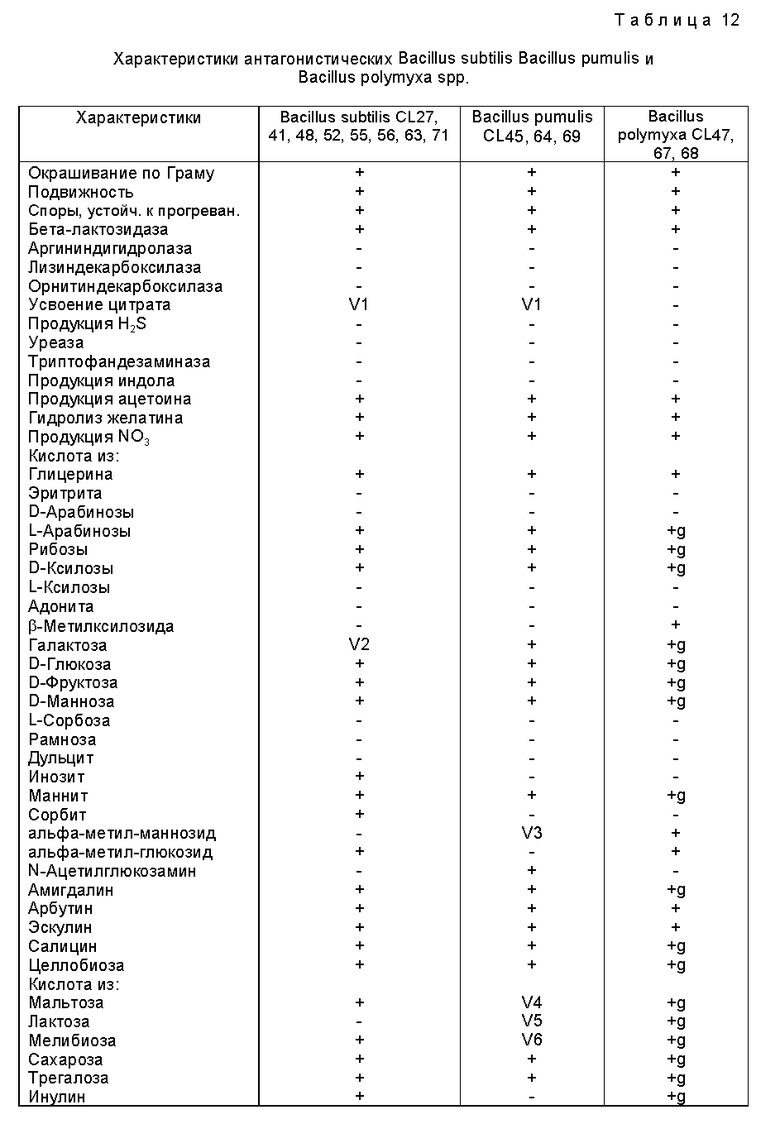
Для первоначальной идентификации использовали окрашивание по Граму (модификация Jensen), форму, подвижность (при фазово-контрастной микроскопии), тест на оксидазу (Kovacs, 1956), прогревание (20 мин при 80oC) и тест окисления/ферментации (Hugh and Leifson, 1953). Оксидаза-отрицательные, образующие кислоту грамотрицательные палочки (Enterobacteriaceae) далее испытывали с помощью API 20E, оксидаза-положительные, не образующие кислоту грамотрицательные палочки - с помощью API 20NE, а устойчивые к прогреванию грамполжительные палочки (Bacillus spp.) - с помощью AP1 20E и 50CHB тест-полосок (API-BioMerieux, UK). Результаты анализировали с помощью компьютерной программы идентификации AP1, а идентификация Serratia liguefaciens (Grimes and Hennerty) подтверждалась путем испытания на образование кислоты из раффинозы, маннолята, лактозы и адонита при использовании методов, описанных Brenner (1984).
Приготовление посевного материала грибков.
Все исходные культуры грибков выращивали на капустном агаре 5 (CA5; 50 г гомогенизированной ткани капусты, 10 г агара, 1 литр дистиллированной воды) при 4oC в темноте. Для образования спор использовали агар на солодовом экстракте (MA). Botrytis cinerea выращивали в течение 5 дней при 20oC в темноте с последующим облучением УФ, чтобы вызвать серообразование, и Alternaria brassicicola выращивали в течение 14 дней при 20oC перед использованием в экспериментах. Споры собирали путем заливки 10 мл стерильного раствора Рингера на чашку с выращенным грибком и суспендирования спор с помощью бактериологичекой петли. Суспензии спор фильтровали через двойной слой муслина и доводили до концентрации 4•106 спор/мл, используя гемацитометр.
Приготовление посевного материала бактерий.
Посевной материал готовили путем суспендирования бактериального подроста из 24-часовой культуры на NA в растворе Рингера 1/4 концентрации. Мутность суспензии затем доводили до абсорбции, равной 1,0 при 625 нм при использовании спектрофотометра ULTRO SPEC 4051 (LKB BIOCHROM Ltd., Cambridge, UK), дающей бактериальную концентрацию, равную 7 • 108 кое/мл для Pseudomonas fluorescens и 5 • 108 кое/мл для изолятов Serratia (число кое получали путем высева на чашки с NA серийных разведений суспензии). Более низкие уровни посевной дозы готовили путем соответствующего разведения в растворе Рингера 1/4 концентрации.
Исследование "in vitro" на противогрибковую активность
20 мл жидкой CA5 охлаждали до 40oC (споры Botrytis cinerea очень чувствительны к нагреванию) и засевали 0,2 мл суспензии грибков, содержащей 4 • 105 спор/мл. После того, как агар застывал, чашки высушивали в шкафу с ламинарным потоком в течение 90 мин, затем на них засевали сразу же петлю бактериальной суспензии, содержащей 2 • 108 кое/мл. При испытании антибиотической активности в неочищенных экстрактах (бесклеточные фильтраты культур) из серийных культур Bacillus spp. засеянную грибком среду наливали в платы для микроопределения Sens-acute (острой чувствительности) (Международная патентная заявка N PCT/GB 90/01067; Proteus Molecular Design Ltd., Marple, Cheshire, UK). После того, как агар застывал, в агаре вырезали 4 мм лунки и заполняли 30 мкл неочищенного экстракта. Диаметры зон подавления вокруг колоний или ячеек измеряли через 2 дня после посева бактерий или заполнения лунок бесклеточными экстрактами Botrytis и через 4 дня после внесения засеянной Alternaria среды.
Исследования "in vivo" (листовые диски) на противогрибковую активность.
Листовые диски (15 мм диаметром) вырезали из внутренних листьев капусты (5 наружных листьев отбрасывали), используя пробочное сверло. Диски переносили в 25 мл универсальные флаконы и нагревали в течение 20 минут при 50oC. Эта обработка, как было обнаружено, необходима, чтобы разрушить природную устойчивость несостарившихся тканей внутренних листьев капусты, чтобы сделать возможной инфекцию, вызванную спорами B. cinerea и A.brassicicola. Диски оставляли для остывания и затем взбалтывали в бактериальной суспензии, содержащей 7•108, 7•107 или 7•106 кое/мл для P.fluorescens изолятов и 5•108, 5•107 или 5•106 кое/мл для штаммов Serratia. Это давало в результате примерно 107, 106 или 105 кое/диск при определении числа кое на листовых дисках сразу после заражения. Это определение выполнялось путем отмывания (10 мин при 6-й скорости с использованием качалки с цапфами Gallencamp) дисков в 2 мл стерильного раствора Рингера 1/4 концентрации для удаления бактерий с растительных поверхностей и посева соответствующих разведений на чашки с питательным агаром.
После заражения бактериями диски переносили на плату для исследования культур тканей (8,5•13 см) с 24 ячейками диаметром 15 мм. Под засеянный прогретый диск помещали непрогретый диск, чтобы имитировать ситуацию в сохраняемом кочане капусты, где чувствительные состарившиеся наружные листья покрывают устойчивые метаболически активные внутренние листья. Исследуемые чашки затем помещали открытыми в камеру с ламинарным потоком до тех пор, пока поверхности дисков не высыхали. Затем в цент помещали 2,5 мкл грибковой суспензии, содержащей 103, 104 или 105 спор. Исследуемые чашки помещали над водой в семенной поднос (15х20 см), и поднос запечатывали пластиковой пленкой. Влажность в подносах измеряли, используя пробу на влажность Vaisala 100 (Vaisala OY, PL 26, Хельсинки, Финляндия), и было обнаружено, что она составляла от 97 до 99% на протяжении всего эксперимента. Подносы инкубировали при 40oC в темноте в течение 10 недель.
Персистенция вновь засеянных антагонистов на листовых поверхностях.
Чтобы определить персистенцию вновь засеянных антагонистических бактерий на листовых поверхностях, непрогретые листовые диски (диаметром 1,5 см) погружали в бактериальную суспензию и инкубировали в чашках для исследования, которые описаны выше. Перед удалением микроорганизмов с поверхности из диска вырезали квадрат 1х1 см, чтобы избежать попадания в образец бактерий с обрезных краев диска. Квадрат помещали в 2 мл раствор Рингера и взбалтывали в течение 10 минут, используя качалку Gallencamp с цапфами при скорости 6, чтобы удалить качалку м поверхности. Число антагонистических бактерий в смыве листа определяли по их антагонизму in vitro против A.brassicicola путем посева смывов с листьев на чашки с CA5, засеянным A.brassicicola и подсчета колоний с зонами подавления роста.
Основная и антагонистическая микрофлора, обнаруженная на Brassica spp.
При определении числа жизнеспособных микроорганизмов на поверхности капустного листа было обнаружено, что листья несут примерно 103 - 104 кое/см2 (таблица 1). Из них примерно от 1 до 15% дрожжей, 65 - 80% составили флюоресцирующие псевдомонады, 5-20% Enterobacteriaceae. Менее 1% поверхностных микроорганизмов были нитчатыми грибами и Bacillus spp., которые могли быть выделены только с помощью метода селективного обогащения, описанного выше. Число жизнеспособных спор Bacillus находилось между 1 и 20 кое/20 см2. Число поверхностных микроорганизмов было ниже на внутренних листьях капусты, и таблица 1 показывает числа, обнаруженные на 6-ом листе.
61% антагонистических бактериальных штаммов, выделенных с Brassica spp., были оксидаза-положительными флюоресцирующими псевдомонадами, 30% были Bacillus и 9% были видами Serratia. Разные группы описаны в отдельных разделах ниже.
Пример 2. Pseudomonas fluorescens: основные свойства.
Флюоресцентные псевдомонады, по-видимому, являются основной частью природной микрофлоры листьев капусты, так как они повторно выделялись в большом количестве. Все выделенные флюоресцентные псевдомонады принадлежали к видам Pseudomonas fluorescens. Все штаммы выделены с выращенной общепринятым образом и органически капусты сортов "датская белая" или "январская слава", брокколи или розеток брюссельской капусты при использовании ряда различных сред для выделения (таблица 2). Присутствовали две основных группы антагонистических псевдомонад, причем это псевдомонады, продуцирующие флюоресцирующие пигменты на питательном агаре (группа 1: CL 6, 7, 9, 10, 11, 12, 15, 16, 18, 21, 29, 39, 40) и псевдомонады, которые не продуцируют эти пигменты (группа 2: CL 42, 66, 74, 82, 13, 14, 17, 22, 23, 30, 31-37, 39).
Подавление in vitro.
Первая группа псевдомонад давала небольшие зоны подавления (< 4 мм) на агаре CA5, засеянном B.cinerea и A.brassicicola, тогда как группа 2 включала: а) штаммы CL 31 - CL 37 и CL 49, которые вызывали гниение капустных листьев (не испытуемые далее на антагонизм in vitro) и б) все штаммы CL 42, 49, 66, 74, 82, 90), которые давали большие зоны подавления (таблица 2). Зоны подавления, образованные различными штаммами псевдомонад из группы 1 при разных температурах и концентрациях питательных веществ в среде были в основном небольшими, (1-4 мм, отдельные результаты не показаны). Однако при сравнении зон подавления штаммов из группы 2 были обнаружены штаммоспецифические примеры (таблица 2). Размер зон подавления на капустном агаре уменьшался при повышении концентрации питательных веществ в среде (от 5 до 100 г/л капустной ткани) для помещенных на хранение штаммов CL 42 и CL 82. Однако штамм CL 66 продемонстрировал увеличение зоны подавления при снижении концентрации питательных веществ на капустном агаре, засеянном Alternaria, но уменьшение зон подавления на капустном агаре, засеянном Botrytis. Важно, что зоны подавления при 4oC были больше при всех концентрациях питательных веществ с обоими грибами вокруг колоний CL 82, а вокруг колоний CL 42 на агаре, засеянном Alternaria, чем при 10oC.
Подавление in vitro.
Pseudomonas - антагонисты также подавляли инфицирование и развитие заболевания из ткани капусты при использовании для определения антагонизма in vivo испытания с дисками из капустных листьев. Штаммы P.fluorescens CL 42, CL 66 и CL 82 продемонстрировали самое лучшее сдерживание роста обоих грибов на дисках из капустных листьев - предупреждение развития симптомов заболевания на дисках, если они погружались в суспензию 7•106 кое/мл; CL 73 давал только среднее сдерживание.
Конец эксперимента (6 недель после того, как на контрольных дисках появлялся рост гриба).
Пример 3. Serratia spp.
Основные характеристики. 9% антагонистических бактериальных штаммов, выделенных Brassica spp. были Serratia spp. (Enterobacteriaceae), причем восемь штаммов были Serratia liquefaciens и один штамм был Serratia plymuthica. Все штаммы были выделены с естественно и общепринятым способом выращенной капусты "датская белая" или "январская слава" ("Dutch White, January Glory") с использованием ряда различных сред для выделения (таблица 4). Зоны подавления на засеянной Botrytis CA5 были больше при 20oC и 10oC, но меньше при 4oC, чем на засеянной Alternaria CA5. Нужно отметить, что Botrytis росли быстрее при всех температурах (развитие видимого роста на среде), чем Alternaria. Зоны подавления штаммов S. liquefaciens CL 57, 58, 59, 61, 62, 73 и 80, как было обнаружено, имели сходный размер, но меньший, чем у зон S. plymuthica, штамм CL 43.
Смотрите условные обозначения к таблице 2.
Размер зон подавления всех штаммов S.Liquefaciens, как было обнаружено, сходен при всех концентрациях питательных веществ в среде (от 5 до 100 г/л ткани капусты) за исключением того, что зоны подавления, образовавшиеся вокруг колоний CL 43 при 10oC с 10 г/л ткани капусты в среде, имеют больший размер (фигура 2). Зоны подавления штамма S. plymuthica CL 43, однако, были в основном больше и CL 43 был единственным штаммом, проявляющим значительный антагонизм при низких концентрациях питательных веществ (5 и 10 г/л, фигура 2).
Смотрите условные обозначения к таблице 3.
Эксперимент заканчивается через 6 недель после того, как на контрольных дисках появился рост гриба.
Подавление in vivo.
Serratia - антагонисты также подавляли инфицирование и развитие заболевания на ткани капусты, когда для обнаружения антагонизма in vivo использовали испытание с дисками из капустных листьев. Когда капустные диски погружались в бактериальную суспензию с 2•109 (результаты не представлены), 2•108, 2•107 или 2•106 кое, повреждение ткани и развитие видимого роста Botrytis cinerea подавлялось с различной степенью в зависимости от использованного вида бактерий и примененной посевной дозы грибка (таблица 5).
Персистенция Pseudomonas и Serratia на капустных листьях
При изучении персистенции вновь засеянных антагонистов P.fluorescens и Serratia spp. были способны персистировать в большом количестве на поверхности капустных листьев при низких температурах (таблица 6).
Пример 4. Основные характеристики Bacillus spp.
30% всех бактерий, проявляющих антагонизм против Botrytis cinerea, как было обнаружено, принадлежат в Bacillus spp.
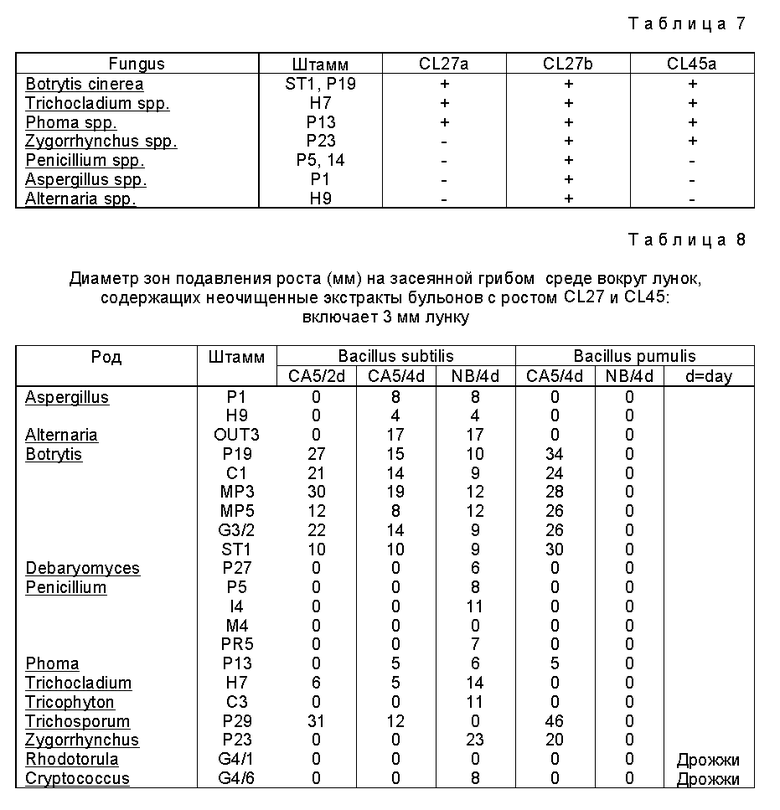
Идентифицировали 40 штаммов Bacillus и обнаружено, что они являлись B. subtilis (8 изолятов), B.pumilus (3 изолята) или B.polymyxa (3 изолята).
Продукция антибиотика in vitro (полуколичественная оценка)
Изоляты Bacillus, выделенные с Brassica spp., не росли при 4oC и 10oC (за исключением штаммов Bacillus CL 47, 52, 64 и 67, которые давали очень слабый рост через 4 недели, на 10 и 6 неделю при 4oC. При 20oC большинство штаммов продуцировали большие зоны подавления на среде CA5 (таблица 7). Однако Bacillus не росли или не давали больших зон подавления при более низких концентрациях питательных веществ в среде (таблица 7).
Пример 5. Продукция антибиотика в серийной культуре Bacillus spp.
В качестве сред роста использовали капустный бульон (50 г (CB 5) или 100 г (CB10) гомогенизированных капустных листьев, 1 литр дистиллированной воды), питательной бульон (NB; Oxoid), питательный бульон + 0,005 г/л MgSO4(NB+Mg), питательный бульон + 1% глюкозы (NB + 1% глюкозы), синтетическую среду (DM: 2 г NH4Cl, 6 г Na2HPO4, 3 г KH2PO4, 3 г NaCl, 0,05 г MgSO4•7H2O, 5 мг L-метионина, 10 мг глицина, 2,5 мг L-аспарагиновой кислоты, 10 г агара, 1 литр дистиллированной воды) и DM + 1% глюкозы. 100 мл среды переносили в 250 мл конические колбы Эрленмейера и автоклавировали и засевали 3-дневной культурой штаммов Bacillus на питательном агаре. Аминокислоты стерилизовали фильтрованием и добавляли в среду после автоклавирования. Через разные отрезки времени после посева из сред отбирали образцы по 4 мл асептически. Число кое определяли путем посева разведений (раствором Рингера 1/4 концентрации) образцов среды на питательный агар в чашках Петри. Число спор Bacillus определяли путем смешивания 0,1 мл абсолютного спирта с 0,1 мл культуральной среды в ячейках микроячеечных плат (0,3 мл ячейки, Nunc Ltd., UK) в течение 20 минут (чтобы убить вегетативные клетки) перед посевом разведений на чашки с питательным агаром.
Неочищенные экстракты антибиотиков из различных сред получали путем центрифугирования жидкости в течение 30 мин при 3000 с использованием центрифуги Mistral 3000 (Fisons, UK). Супернатант фильтровали через 0,2 мкм нитроцеллюлозный фильтр (Sartorius, Gottingen, Германия) для получения бесклеточных фильтров. Для получения низких концентраций антибиотиков эти неочищенные экстракты затем разводили капустным бульоном. 30 мкл различных разведений неочищенного экстракта помещали в лунки диаметром 4 мм в чашках с CA5, засеянной B.cinerea или A.brassicicola. При определении действия pH на активность антибиотиков в неочищенных экстрактах pH исследуемой среды доводили до значений от 3 до 9 с помощью HCl и NaOH перед помещением неочищенных экстрактов в лунки.
Продукция антибиотиков in vitro (количественное определение).
Зоны подавления образовывались также вокруг бесконечных фильтратов культуры Bacillus spp., выращенных в течение 5 дней в капустном бульоне (CB5) или питательном бульоне (NB) в качестве среды роста (таблица 8). Бесклеточные фильтраты серийных культур штаммов CL27 и CL 41 Bacillus с капустного агара обладали очень высокой активностью против Botrytis cinerea и более низкой активностью против Alternaria brassicicola (таблица 8). Фильтраты питательного бульона обладали активностью только против A.brassicicola, если для выполнения B. cinerea исследований использовали чашки Петри, но демонстрировали сходную активность против B.cinerea и A.brassicicola, если исследование выполняли на более чувствительных чашках для микроопределения Sens-acute: смотрите Hampson et al.
Hampson et al., 1992 г. дает детальное описание различий между чашками Sens-acute для микроопредления и чашками Петри. Фильтраты питательного бульона и капустного бульона (CB5) от штамма CL 45 были активны только против B.cinerea (таблица 8).
Пример 6. Воздействие среды культивирования.
Изучалась продукция антибиотиков в серийной культуре после различных периодов инкубации и на различных жидких средах (NB, NB + M, NB + 1% глюкозы, CA5, CA10, DM и DM + 1% глюкозы). Фильтраты культур CL 27 и CL 45 снова проявили очень сильную активность (зоны подавления около 40 мм) против B.cinerea при выращивании в капустном бульоне (фиг. 3 и 4), но проявили значительно более низкую активность (зоны подавления 15 мм) при выращивании в питательном бульоне (фиг. 5 и 6) или синтетической среде (фиг. 7). В капустном бульоне (CA5 и CA10) как CL 27, так и CL 45 проявили - Botrytis активность через 2 дня, примерно за 24 часа до того, как могли быть обнаружены первые вновь образованные споры Bacillus. На NB и DM антибиотическая активность также развивалась через 2 дня, но увеличение числа спор можно было обнаружить только через 9 дней. Добавление 1% глюкозы подавляло спорообразование у Bacillus, но не проявление антигрибковой активности в NB и DM (фиг. 5 и 7).
Как и перед эти штамм CL 27, но не CL 45 проявлял активность против Alternaria brassicicola. Активность была самой высокой у бесколеточных фильтратов культуры из NB+Mg и серийных культур в CB10 (зоны подавления 20 мм) и сходной во всех других средах (зоны подавления между 15 и 10 мм) (фиг. 3 - 7). Подобно активности против Botrytis, активность против Alternaria предшествовала спорообразованию на 24 часа в CA5 и CA10 и на 7 дней в NB и DM, тогда как добавление 1% глюкозы не останавливало развития противогрибковой активности (фиг. 3 - 6).
Пример 7. Действие pH и питательных веществ на антибиологическую активность.
На активность противогрибковых антибиотиков, образовавшихся в жидкой серийной культуре Bacillus subtilis (определяемую по диметру зоны подавления на среде CA5, засеянной грибом) влияли pH (фиг. 8) и концентрация и тип питательных веществ в исследуемой среде (фиг. 9).
Воздействие pH: антибиотики, продуцируемые в капустном бульоне (CB5 и CB10) через 2 и 4 дня и CL 45 в NB + Mg через 2 дня, проявляли наивысшую активность (формировали наибольшие зоны подавления) против Botrytis при pH между 5,6 и 6,0. Активность слегка сжималась, если pH понижался до 3,1 и снижалась до очень низких значений, если pH повышался до значений выше 6,0 (фиг. 8). Антибиотики, продуцируемые CL 27 на NB + Mg имели сходную активность при pH между 5,6 и 8,9, но сниженную активность, когда pH был ниже 5,6. В NB активность можно было обнаружить, если pH был выше 6,0, и она слегка повышалась при повышении pH. Когда активность антибиотиков испытывали на засеянном Botrytis картофельном агаре, экстракты из питательного бульона и CB5 не проявляли активности, экстракты из CA10 показали низкую активность при значениях pH ниже 6, но сходную активность при более высоком pH, и экстракты из питательного бульона с марганцем проявили повышенную активность по сравнению с тем, когда использовался засеянный Botrytis капустный агар (фиг. 8C).
Уровни питательных веществ: при повышенных концентрациях питательных веществ, в виде капустной ткани или картофельного экстракта и декстрозы, в испытуемой среде активность антибиотиков также снижалась (фиг. 9). На картофельно-декстрозном агаре образовывались или только очень небольшие зоны или они отсутствовали (фиг. 9).
Стабильность антибиотиков при низких температурах.
Образовавшиеся антибиотики, как было обнаружено, очень устойчивы при 4oC, так как невозможно было определить снижение активности в неочищенных бесклеточных фильтрах культур штаммов CL 27, CL 41 и CL 45, сохраняемых стерильными в течение 4 месяцев. Фильтраты анализировали через месячные интервалы во время хранения при 4oC.
Пример 8. Выделение и определение свойств антибиотиков, продуцируемых Bacillus subtilis, штаммом CL 27 и Bacillus pumilis, штаммом CL 45.
Бактерии выращивали в течение 7 дней в серийной культуре или в капустном бульоне (10 г гомогенизированной ткани капустных листьев) или питательном бульоне (+0,005 г Mn на литр). Бесклеточные экстракты культур готовили, как описано выше. 0,04 мл бесклеточного фильтрата культуры помещали на силикагелевые 60 ТСХ пластинки (Art. 5271, Merck) и разделяли, используя систему растворителей, описанную Swinburne et al. (Trans. Br. Mycol. Soc, 65 (2), 211-217), то есть бутан-1-ол:уксусная кислота:H2O, 3:1:1. Пластинки высушивали в камере с ламинарным потоком и затем в сушильном шкафу в течение 5 часов при 70oC, чтобы удалить растворитель и остаток уксусной кислоты.
Суспензии грибов готовили или в питательном бульоне или в капустном бульоне (5 г/л гомогенизированных капустных листьев). Высушенные пластинки ТСХ опрыскивали суспензией спор или B.cinerea или A.brassicicola и инкубировали в течение 3 дней или 4 дней (в зависимости от эксперимента) при 20oC в темноте, чтобы дать возможность роста грибов на пластинках. Зоны подавления роста визуализировали по методам Lund and Lyon (J. Chromatography 100, 192-196 (1975)) и использовали для расчета значений Rf для продуцируемых антибиотиков.
Антибиотики - Значения
Антибиотик против Botrytis, продуцируемый CL 27a на капустном бульоне: Предполагаемый не-пептид (отрицательный TDM): активность при pH < 5,6 - 0,39
Антибиотик против Botrytis/Alternaria, продуцируемый CL 27b на капустном бульоне: Предлагаемый пептид (положительный TDM): активность при PH > 5,6 - 0,56
Антибиотик против Alternaria, продуцируемый CL 27c в питательном бульоне: Предположительно пептидной природы (положительный TDM): активность при pH > 5,6 - 0,61
Антибиотик против Botrytis, продуцируемый CL 45a в капустном бульоне. Предлагаемый не-пептид (отрицательный TDM): активность при pH < 5,6 - 0,39
TDM = 4,4'-тетраметилдиаминодифенилметановый реагент: положительный = синий/зеленый. Антибиотики CL 27 образуются в капустном бульоне (CB5), питательном бульоне и питательном бульоне с марганцем: антибиотики CL 45 - только на капустном бульоне (CB5).
Спектр активности антибиотиков (испытание на пластинках, как описано выше).
Когда пластинки для ТСХ, на которые наносились бесклеточные фильтры культуры со среды CB5, засеянной или CL 27 или CL 45, покрывали засеянной Botrytis cinerea средой, развивались большие зоны подавления роста (30-40 мм) на Rf, равном от 0,38 до 0,40. При излучении фильтратов из среды CB5, засеянной штаммом CL 27, можно было обнаружить дополнительную небольшую полоску (3 мм) без роста гриба в положении со значением Rf, равным 0,56 (не присутствующую, если испытывалась CB5, засеянная CL 45). Когда при исследовании фильтратов из NB и NB+Mn, засеянных CL 45 производили покрытие таким же образом, можно было обнаружить только небольшую полоску подавления со значением Rf, равным 0,567 (не было зон подавления, когда излучались среды NB и NB+Mn, засеянные CL 45).
Когда покрытие выполнялось средой, засеянной Alternaria brassicicola, были обнаружены 2 зоны подавления (Rf 0,56 и 0,61) из посевов на среды роста NB, NB+Mn и CB5 CL 27. Таким образом видно, что CL 27 продуцирует, по крайней мере, три антибиотика, один, активный против B.cinerea (Rf 0,39), продуцируемый только на капустном экстракте и активный при pH, меньше 6; второй слабо активный против B.cinerea и активный против A. brassicicola (Rf 0,56), продуцируемый на всех испытуемых средах и активный при pH выше 5,6; и третий, активный только против A.brassicicola (Rf 0,61) тоже продуцируемый на всех испытуемых средах. Сходные значения для антибиотиков из CL 45 и CL 27 наводят на мысль, что они представляют собой одно и то же, но характер активности наводит на мысль о противоположном.
Пример 9. Характеристики антагонистических бактериальных штаммов этого изобретения.
Все бактериальные штаммы, выделенные с Brassica spp., демонстрирующие in vitro антагонизм против Botrytis cinerea, как было обнаружено, являются видами Pseudomonas fluorescens (группа I), Bacillus или Serratia. Флюоресцирующие псевдомонады и Enterobacteriaceae, как было обнаружено, являются основными обитателями капустных листьев, тогда как Bacillus spp. были обнаружены лишь в очень небольших количествах на листьях Brassica. Флюоресцирующие псевдомонады и Bacillus spp. были описаны ранее, как потенциальные средства биологической борьбы с заболеваниями фруктов и овощей после уборки урожая, тогда как подобная активность у Serratia plymuthica Serratia liquefaciens ранее не была показана (смотрите Wilson & Wisniewski, 1989 по недавнему обзору по биологической борьбе с грибковыми заболеваниями фруктов и овощей после уборки урожая).
Pseudomonas fluorescens.
Из всех испытанных бактериальных штаммов штаммы флюоресцирующих псевдомонад CL42, CL 66 и CL 82 продемонстрировали самую высокую активность in vivo на дисках из капустных листьев при 4oC, подавляя инфицирование Botrytis при всех испытанных размерах посевных доз гриба. Псевдомонады, такие, как P. сераcia, P. putida и P. syringae были описаны, как средства биологической борьбы с болезнями после сбора урожая для ряда сохраняемых фруктов и овощей, и описывается, что их способ действия состоит или в продукции антибиотика или в создании конкуренции за питательные вещества (Colyer and Mount 1984, James and Gutterson 1986, Gurusiddaiah et al. 1986, Janisiewicz 1987, Janisiewicz and Roitman 1988). Важно отметить, что P.syringae и P.cepacia могут быть патогенными для растений и что некоторые псевдомонады, включая P.Sceptical, были описаны, как условно-патогенные возбудители болезней у человека (Agrios 1988, Bergan 1981).
Некоторые антагонистические псевдомонады, выделенные данными изобретениями, как было также обнаружено, являются патогенными для некоторых овощей, например, капусты. Поэтому антагонистические псевдомонады должны идентифицироваться до уровня вида, чтобы исключить использование возможных человеческих возбудителей болезней, и должны испытываться на воздействие на растения перед использованием в качестве средств борьбы с болезнями после уборки урожая. Так как выделенные антагонистические штаммы были способны персистировать на капустных листьях, они оцениваются как идеальные потенциальные кандидаты для биологической борьбы с болезнями во время хранения Brassica spp.
Serratia spp.
Serratia spp. , выделенные с листьев капусты хорошо росли и проявляли сильный антагонизм in vitro при низких температурах. Однако только штамм CL 42 S. plymuthica продемонстрировал высокий антагонизм in vivo против B.cinerea на дисках из капустных листьев. Все штаммы S.liquefaciens продемонстрировали от среднего до плохого противодействия болезням in vivo. Serratia spp. ранее не были идентифицированы в качестве антагонистов грибкам. Serratia spp. не были описаны как возбудители болезней растений, но S.marcescens была описана как условно-патогенный возбудитель, вызывающий инфекции у человека (Brenner, 1984). Поэтому такие микроорганизмы должны использоваться только, если уже было показано, что они непатогенны для человека. Enterobacteriaceae образуют основную часть естественной микрофлоры капустных листьев, и так как выделенные антагонистические штаммы способны персистировать на листьях капусты, они обладают возможностями служить в качестве средств биологической борьбы с болезнями, конкретно, например, во время хранения Brassica spp.
Bacillus spp.
Bacillus spp. считались непригодными в качестве потенциальных средств биологической борьбы с болезнями для хранения на холоде, так как они не росли или очень плохо росли при низких температурах. Об этом ранее не сообщалось, так как большинство работавших, описывающих использование видов Bacillus для биологической борьбы с грибковыми возбудителями болезней фруктов и овощей испытывали антагонизм in vitro и in vivo при температурах около 20oC (Pusey and Wilson, 1984, Singh and Deverall, 1984, Utkhede and Sholberg, 1986, Gueldner et al., 1988). Их плохой рост при низких температурах мог, однако, служить объяснением различных результатов, описанных в полевых и предварительных испытаниях с целью коммерческого применения Bacillus subtilis в качестве основы биологических средств борьбы с болезнями (Baker et al., 1985, Pusey et al., 1986 and 1988). Известно, что Bacillus spp. продуцируют пептидные антибиотики с антигрибковой активностью во время спорообразования (смотрите Katz and Demain, 1977 для просмотра) и пептидные антибиотики были идентифицированы как активные вещества при использовании Bacillus subtilis в качестве средства биологической борьбы с грибковыми заболеваниями после сбора урожая (McKeen et al., 1986, Gueldner et al., 1988).
Продукция таких антибиотиков путем ферментации и их использование в качестве "биопестицидов" является предпочтительным использованием таких Bacillus, если невозможно выделить устойчивые к холоду антагонистические Bacillus spp. Однако антибиотики и микроорганизмы могли бы также использоваться для продления срока хранения, когда фрукты и овощи изымаются из хладохранилища. Слабая активность неочищенных экстрактов антибиотиков при определении на картофельно-декстрозном агаре (PDA) указывает на то, что обнаруженные антибиотики являются новыми, так как PDA, использовался большинством других авторов для отбора Bacillus spp., которые продуцируют противогрибковые антибиотики (Katz and Demain 1977, Pusey and Wilson, 1984, Singh and Devereal, 1984, McKeen et al., 1986, Utkhede and Sholberg, 1986, Wilson and Wisniewski, 1989).
Подробные характеристики всех штаммов этого изобретения представлены в таблицах 11 и 12.
Пример 10. Активность in vivo всех штаммов против B. cinerea на поверхности молодых листьев растений.
CL 82, CL 80 и CL 27 испытывали на способность сдерживать рост B.cinerea на поверхности листьев in vivo при использовании испытания на молодых растениях в камерах с созданием высокой влажности. Только CL 27 продемонстрировал значительное сдерживание, сходное с результатом, достигаемым с помощью фунгицидов. Применение бесклеточных фильтратов культуры CL 27 в бульоне давало сходное сдерживание с тем, которое обеспечивалось клеточной суспензией, показывая таким образом эффект антибиоза. Этот результат показывает, что CL 27 и его производные могут использоваться в качестве средства биологической борьбы с болезнями, применяемого перед уборкой урожая. Для определения эффективности такого применения после уборки урожая выполняли исследования по персистенции.
Микрорастения астильбы высаживали в компостные горшочки 3х3 см и их опрыскивали через недельные интервалы спорами грибков и антагонистами при влажности от 86 до 95%. В течение первых 6 недель все обработки давали сходный успех, но через 10 недель только фунгицид и обработка с использованием CL 27 или его бульона давали значительное сдерживание. CL 27 давал лучшее сдерживание, чем фунгицид при этих испытаниях.
Пример 11: Действие штаммов Serratia и Pseudomonas на сохраняемые продукты.
Выращивали датскую белую капусту (вар. Морган) и обрабатывали инсектицидом и гербицидами обычным образом, но не обрабатывали фунгицидами. После сбора кочаны погружали или в суспензию фунгицида (1 г/л Роврала и 2 г/л Ридомила) или в бактериальные суспензии 107 - 108 кое/мл. Примерно 14 кг капусты (10 - 12 кочанов) переносили в пластиковые подносы и запечатывали в пластик, чтобы исключить перекрестное обсеменение; на каждую обработку использовали 5 повторностей подносов. Испытания проводились в холодном хранилище при 4-6o и при 1-3oC и влажности, измеряемой через 7-дневные интервалы и, как обнаружено, находившейся на уровне 92-96% во всех случаях. В одном из испытаний после фунгицидов или бактерий наносили 106 спор/мл B.cinerea; на каждый поднос напыляли 5 мл суспензии спор.
Состояние капусты оценивалось следующим образом: 0 = нет видимого роста; 1 = 1 - 10%, 2 = 11 - 25%, 3 = 26 - 50%, 4 = 51 - 75%, 5 = 76 - 90%, 6 = 91 - 99%, 7 = 100% поверхности покрыто ростом гриба и измерялось через 6-недельные интервалы. Когда наблюдалось 80% покрытия ростом грибка у необработанных контролей (когда происходит обрезка при коммерческом хранении), производилась оценка деструкции при потере веса при удалении инфицированных листьев. Кочаны снова хранили в течение 10-14 недель и затем обрезали после 42 недель хранения. Испытание при 1-3oC не дает возможности определения деструкции, поэтому проводилось экстраполирование инфицированной площади листьев на потери при обрезке.
Использование CL 80 и CL 82 обеспечивало защиту, сходную с защитной фунгицидами во всех случаях, причем CL 82 давал лучшее подавление при обработке с добавлением B.cinerea, чем с A.brassicicola, в противоположность результатами in vitro. CL 82 персистировал на капусте в более высоком количестве, чем другие антагонисты, и таким образом является предпочтительным образцом изобретения. Из других штаммов CL 43 был наиболее эффективным в холодном хранилище.
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛИПОСОМЫ, СОДЕРЖАЩИЕ МАТЕРИАЛЫ, СОСТОЯЩИЕ ИЗ ЧАСТИЦ | 1994 |
|
RU2145212C1 |
| ВЫРАВНИВАЮЩИЙ МЕХАНИЗМ | 1991 |
|
RU2091516C1 |
| ПЛЮЩИЛЬНЫЙ МЕХАНИЗМ, УСТРОЙСТВО ДЛЯ ОТДЕЛЕНИЯ ВОЛОКНА ИЗ ВОЛОКНИСТОГО РАСТИТЕЛЬНОГО МАТЕРИАЛА | 1994 |
|
RU2098521C1 |
| УСТРОЙСТВО ДЛЯ РАЗДЕЛЕНИЯ ВОЛОКНА И КОСТРЫ И МЯЛКА-ВОЛОКНООТДЕЛИТЕЛЬ | 1994 |
|
RU2099446C1 |
| ШТАММ БАКТЕРИЙ PSEUDOMONAS SPECIES ДЛЯ ПОЛУЧЕНИЯ ПРЕПАРАТА ПРОТИВ ВОЗБУДИТЕЛЕЙ ЗАБОЛЕВАНИЙ РАСТЕНИЙ | 1997 |
|
RU2130262C1 |




Изобретение относится к биотехнологии, в частности к производству средств защиты овощей и фруктов от возбудителей грибковых заболеваний после сбора урожая. Для этой цели получены штаммы бактерий Pseudomonas fluorescens NСIМВ 40490, NСIМВ 40495, NCIМВ 40497, Serratia liquefaciens NСIМВ 40492, Serratia plumuthica NCIМВ 40493. Кроме того, получены штаммы бактерий Bacillus pumilis NCIMB 40489 и Bacillus subtilis NCIМВ 40491, используемые не только для вышеуказанной цели, но и для производства антибиотика, ингибирующего возбудителя вышеуказанных заболеваний. На основе вышеуказанных штаммов получена водная суспензия микроорганизма, содержащая штаммы в концентрации 104-109 кое/мл, предпочтительно 105-108 кое/мл. Способ предотвращения грибкового заболевания овощей и/или фруктов после сбора урожая предусматривает погружение овощей и/или фруктов в водную суспензию, содержащую один из вышеуказанных штаммов бактерий. На основе штаммов бактерий Bacillus 40489 и 40491 получен антибиотик, имеющий Rf около 0,39, или 0,56, или 0,61, проявляющий активность против Botrytis cinerea и Alternaria при рН 3,0 - 9,0. Антибиотик получают путем культивирования штамма бактерии на среде, содержащей капустные листья и воду в пропорции 5 - 100 г листьев на 1 л воды. Выделяют антибиотик из фильтрата культуральной жидкости с использованием силикагелевой колонки или пластины и элюента бутан-1-ол:уксусная кислота:вода в соотношении 3: 1: 1. Антибиотик и предложенные штаммы позволяют сохранить урожай фруктов и овощей в процессе хранения, предотвращая их гниение. 10 с. и 6 з. п.ф-лы, 13 табл., 9 ил.
| US 4764371, 16.08.88 | |||
| US 4663162, 05.05.87 | |||
| Бюллетень ВНИИСХМ, 1970, N 14, вып | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Способ окисления боковых цепей ароматических углеводородов и их производных в кислоты и альдегиды | 1921 |
|
SU58A1 |
