Изобретение касается моноклональных антител, специфичных по отношению к гликопротеину P (Pgp). Конкретно, настоящее изобретение касается моноклональных антител к внеклеточному эпитопу человеческого гликопротеина P, их получения и применения в диагностических и клинических целях.
Известны (Kohler G. and Milstein C., Nature, 256, 495, 1975) методы слияния соматических клеток для получения гибридных клеточных линий, продуцирующих моноклональные антитела, которые сделали возможным получать неограниченные количества гомогенных препаратов антител, обладающих определенной специфичностью по отношению к отдельным антигенным детерминантам. Участок белковой антигенной детерминанты, узнаваемой и взаимодействующей с антителом, обозначается здесь как эпитоп и состоит из одной или более специфичных эпитоп-образующих аминокислотных последовательностей.
Гликопротеин P представляет собой белок плазматической мембраны, экспрессирующийся на поверхности клеточных линий или опухолевых клеток человека, проявляющих свойственный им или приобретенный фенотип множественной устойчивости к антибиотикам (МУА). Экспрессия указанного белка придает клеткам способность активно размножаться в присутствии таких концентраций цитостатических антибиотиков, которые токсичны для не-МУА исходных клеток. Изучение трансфекции с использованием клонированных кДНК гликопротеина P продемонстрировало непосредственное участие указанного белка в определении фенотипа МУА (Ueda et al. (1987), PNAS, 84, 3004-3008). Считают, что гликопротеин P служит энергетически-зависимым насосом, выводящим антибиотики.
Аминокислотная последовательность гликопротеина P была выведена на основании нуклеотидных последовательностей генов, способных придавать фенотип МУА реципиентным клеткам (Ueda et al. (1987), PNAS, 84, 3004-3008; Shen et al. (1986), Mol. Cell. Biol. , 6, 4039-4044; Chen et al. (1987), Cell, 47, 381-389). Вторичная структура гликопротеина P была предсказана на основании анализа его первичной структуры и соответствует трансмембранному белку, состоящему из 12-ти гидрофобных трансмембранных спиралей, соединенных 6-ю гидрофильными внеклеточными петлями и 2-мя цитоплазматическими доменами, кодирующими две последовательности связывания АТФ.
Выделено и охарактеризовано несколько различных моноклональных антител к гликопротеину P, специфичность некоторых из них рассматривается Ченциарелли с соавторами (Cenciarelli et al. (1991), Int.J.Cancer, 47, 533-544). Позднее были описаны другие моноклональные антитела, обладающие специфичностью к внеклеточным доменам гликопротеина P (Arceci et al. (1993), Cancer Res., 53, 310-317; Mechtner & Robinson (1992), PNAS, 89, 5824-5828; Georges et al. (1993), JBC, 268, 1792-1798). Тем не менее, моноклональные антитела к гликопротеину P, для которых охарактеризованы эпитоп-образующие аминокислотные последовательности, взаимодействуют либо с цитоплазматическими доменами гликопротеина P, либо с внеклеточными эпитопами, включающими в себя непротяженные участки аминокислотной последовательности белка. Например, Джорджес с соавторами (Georges et al. (1993), JBC, 268, 1792-1798) картировали эпитоп моноклонального антитела MKR-16, описанного Хамада и Цуруо (Hamada and Tsuruo (1986), PNAS, 83, 7785-7789), который включает в себя пептиды, присутствующие на 2-х из 6-ти внеклеточных белковых петель (1 и 4-й), предсказанные на основании первичной аминокислотной последовательности.
Моноклональные антитела к гликопротеину P предложено использовать в диагностических и клинических целях, в том числе для мониторинга уровня экспрессии гликопротеина P и проявляющегося вследствие этого фенотипа множественной устойчивости к антибиотикам (МУА) у клеток человека. Кроме того, указанные антитела могут найти применение в подавлении МУА, обусловленной гликопротеином P (Mechtner & Robinson (1992), PNAS, 89, 5824-5828).
Ни одно из известных моноклональных антител к гликопротеину P не узнает специфические для человека внеклеточные эпитопы гликопротеина P, состоящие из протяженной эпитоп-образующей аминокислотной последовательности. Более того, наблюдается постоянная потребность в антителах, способных узнавать малые количества гликопротеина P, например, при низком уровне его экспрессии МУА-клетками, или в том случае, когда МУА-клетки составляют незначительную часть клеточной популяции. Моноклональные антитела, способные узнавать указанные специфичные для человека эпитопы, а также малые количества гликопротеина P, были бы удобны для иммунологического тестирования.
Таким образом, при создании изобретения ставилась задача получения антител, которые способны узнавать и связываться с высокой специфичностью со структурно протяженными эпитопами гликопротеина P человека, характеризующимися внеклеточной локализацией и состоящими из протяженной эпитоп-образующей аминокислотной последовательности.
С использованием указанных методов слияния соматических клеток были получены гибридомы, секретирующие моноклональные антитела (ММ 4.17), способные к избирательному, высоко специфичному связыванию с ранее неизвестными протяженными внеклеточными эпитопами гликопротеина P человека.
Соответственно, в изобретении предложено моноклональное антитело, взаимодействующее со структурно протяженным и локализованным вне клетки эпитопом гликопротеина P человека, который состоит из протяженной эпитоп-образующей аминокислотной последовательности, имеющее следующие свойства:
- окрашивает более 90% живых клеток CEM-VBL10, как определяется экспериментами по проточной цитометрии;
- вариабельный домен этого антитела имеет определяющие комплементарность районы (ОКР) тяжелой цепи, которые являются такими же, как соответствующие ОКР последовательности 3.
В аминокислотной последовательности, представленной в Последовательностях 2 и 3, ОКР1 соответствует аминокислотным остаткам 32 - 36 включительно, ОКР2 - аминокислотным остаткам 51 - 67 включительно, а ОКРЗ - аминокислотным остаткам 100 - 111 включительно. Остальные участки аминокислотных последовательностей, представленных в Последовательностях 2 и 3, включают в себя каркасные районы (КР1, КР2, КРЗ и КР4 по направлению от N-конца к C-концу), которые могут быть замещены на другие каркасные районы без существенного влияния на способности вариабельного домена связывать антиген.
В настоящем описании вариантом последовательности ОКР считают последовательность, которая по меньшей мере на 70%, в предпочтительном случае - по меньшей мере на 80%, в более предпочтительном случае - по меньшей мере на 90% и в наиболее предпочтительном случае - по меньшей мере на 95% гомологична соответствующей последовательности ОКР, в том числе ОКР1, ОКР2 или ОКРЗ аминокислотных последовательностей, представленных в Последовательностях 2 и 3.
В настоящем описании термин "гликопротеин P человека" соответствует продукту человеческого гена MDR1. Кроме того, понятие "моноклональное антитело" охватывает целые молекулы антител и их способные к связыванию антигена фрагменты, включая фрагменты FAb, F(Ab')2, Fv и одноименные антитела, и не ограничивается продуктами, получаемыми от гибридомных клеточных линий, но включает также молекулы антител и их способные к связыванию антигена фрагменты, полученные с помощью методов рекомбинантной ДНК, в том числе путем экспрессии клонированных последовательностей ДНК, кодирующих антитело.
Узнаваемая антителом эпитоп-образующая аминокислотная последовательность может быть локализована в пределах любой из внеклеточных петель гликопротеина P человека. В предпочтительном случае эпитоп-образующая аминокислотная последовательность локализуется в пределах 4-й петли гликопротеина P человека. В более предпочтительном случае эпитоп-образующая аминокислотная последовательность входит в состав пептида гликопротеина P человека, имеющего следующую последовательность: FTRIDDPETKRQNSNL (Последовательность 1 в прилагаемом перечне последовательностей). В еще более предпочтительном случае эпитоп-образующая последовательность включает в себя по меньшей мере 5 смежных аминокислотных остатков из числа 8-ми аминокислот, представляющих собой остатки 2 - 9 из Последовательности 1 включительно, а в еще более предпочтительном случае - 5 аминокислот, соответствующих аминокислотным остаткам 3 - 7 из Последовательности 1 включительно.
Неожиданно для авторов было обнаружено, что по сравнению с известными моноклональными антителами моноклональные антитела ММ4.17 обладают более высокой степенью сродства к МУА-клеткам и способны выявлять даже минимальные уровни приобретенной множественной устойчивости к антибиотикам у клеток CEM-VBL10 (Cianfriglia et al. (1990), Int.J.Cancer, 45, 95-103; Beck et al. (1979), Cancer Res. , 39, 2070-2077). Кроме того, ММ4.17 способны выявлять приобретенную множественную устойчивость к антибиотикам у клеток CEM-VBL16 (Beck et al. (1979), Cancer Res., 39, 2070-2077). Относительная устойчивость (ОУ) указанной клеточной линии к винбластину равна 6,25. Клеточные линии CEM-VBL10 и CEM-VBL16 представляют собой МУА-производные чувствительной к антибиотикам клеточной линии CEM ((Beck et al. (1979), Cancer Res., 39, 2070-2077), депонирована под номером ATCC CCL 119), полученные посредством селекции в среде, содержащей соответственно 10 нг/мл и 16 нг/мл винбластина.
Таким образом, в наиболее предпочтительной реализации настоящее изобретение включает в себя предусмотренные настоящим изобретением моноклональные антитела, имеющие сродство связывания с гликопротеином P, близкое таковому для ММ4.17, в том числе моноклональные антитела, способные выявлять приобретенную множественную устойчивость к антибиотикам у клеток CEM-VBL10 или CEMVBL16 или сходных клеток, проявляющих МУА.
Сравнение сродства связывания антител ММ4.17 и ранее полученных анти-Pgp моноклональных антител C219 (Kartner et al. (1985), Nature, 316, 820-823), C494 (Kartner et al. (1985), Nature, 316, 820-823), JSB1 (Scheper et al. (1988), Int.J.Cancer, 42, 389-394), MRK-16 (Hamada and Tsuruo (1986), PNAS, 83, 7785-7789) и MC57 (Cenciarelli et al. (1991), Int.J.Cancer, 47, 533-544) с гликопротеином P по отношению к чувствительным к антибиотикам исходным клеткам СЕМ и их МУА-производным клеточным линиям CEM-VBL10 и CEM-VBL100 приведено ниже в Примере 6. Проявляющие МУА клеточные линии CEM-VBL10, CEM-VBL16 и CEM-VBL100 и их получение описаны Беком с соавторами (Beck et al. (1979), Cancer Res., 39, 2070-2077). Сходные клеточные линии, проявляющие МУА, можно получать принципиально так же, как это описано Беком с соавторами.
Предусмотренные настоящим изобретением моноклональные антитела можно получать с помощью стандартных методов. Так, гибридомные клеточные линии можно получать посредством слияния соматических клеток с использованием клеток селезенки, полученных у животных, иммунизированных МУА-клетками человека. Можно использовать любую пригодную клеточную линию человека, проявляющую МУА, включая клеточные линии МУА, полученные из чувствительных к антибиотикам исходных клеточных линий, такие как клеточная линия CEM-VBL100, описанная Беком с соавторами (Beck et al. (1979), Cancer Res., 39, 2070-2077), или сходные клеточные линии МУА. CEM-VBL100 представляет собой МУА-производное чувствительной к антибиотикам клеточной линии СЕМ, депонированной Американской Коллекцией типов культур под номером ATCC CCL 119.
Для иммунизации более удобно использовать пептиды, соответствующие внеклеточным доменам гликопротеина P человека или его фрагментам. Можно использовать любой пригодный пептид внеклеточного домена гликопротеина P человека. Например, в качестве антигенов можно использовать пептиды, соответствующие четвертой петле гликопротеина P или ее фрагментам, такие как пептид, приведенный в Последовательности 1, или его обсуждавшиеся выше фрагменты.
В предпочтительном случае используют способы иммунизации, приводящие к образованию моноклональных антител, проявляющих высокое сродство, такие как способы, описанные Чианфриглия с соавторами (Cianfriglia et al. (1986), Meth. in Enzymol., 121, 193-210), и сходные с ними. Гибридомы, продуцирующие искомые моноклональные антитела, можно выявлять на основании их способности преимущественно связываться с МУА-клетками человека по сравнению с чувствительными к антибиотикам клетками, после чего следует этап селекции гибридом, специфичных по отношению к структурно протяженным эпитопам, характеризующимся внеклеточной локализацией. Для скринирования искомых моноклональных антител используют предпочтительные пептиды, соответствующие структурно протяженным внеклеточным доменам человеческого гликопротеина P или их частям, таким, как рассмотренные выше предпочтительные пептиды.
Моноклональные антитела можно получать из гибридомных клеточных линий с помощью методов культуры клеток и асцитов. Фрагменты антител можно получать с использованием адекватных методов, таких как ферментативное расщепление. Рекомбинантные продукты можно получать с помощью методов, включающих в себя трансформацию или трансфекцию пригодных хозяйских клеток последовательностями ДНК, кодирующими целую молекулу или фрагмент моноклонального продукта, а также культивирование трансформированных хозяйских клеток с получением указанных продуктов.
Указанные антитела могут быть успешно использованы
- для выявления присутствия гликопротеина P в нормальных или опухолевых тканях с применением иммунофлюоресцентных или иммунохимических методов;
- в тестах in vitro, предназначенных для выявления клеток, проявляющих фенотип МУА;
- в методах обогащения популяций специфичных клеток, у которых имеется гликопротеин P (стволовые клетки, ВИЧ-инфицированные клетки, T-лимфоциты).
В дальнейшем изобретение описывается путем иллюстрирования только следующими Примерами со ссылкой на прилагаемый чертеж, который представляет собой график, показывающий результаты анализа экспресии гликопротеина P человека у живых клеток СЕМ и их МУА-вариантов, что определяется с помощью моноклональных антител ММ4.17 (1 = чувствительная к антибиотикам линия СЕМ; 2 = CEM-VBL20; 3 = CEM-VBL40; 4 = CEM-VBL80; 5 = CEM-VBL100. Абсциссы: интенсивность флюоресценции; ординаты: количество клеток), проводимого флюоресцентным методом разделения клеток (FACS).
Пример 1. Иммунизация мышей и гибридизация спленоцитов с клетками миеломной линии Sp2-01/Ag.l4 (ATCC CRL 1581).
Мышей BALB/c иммунизируют МУА-вариантом Т-лимфобластоидной линии человека, уже охарактеризованной и известной под номером CEM-VBL100 (Beck et al. (1979), Cancer Res., 39, 2070-2077). Получение клеточной линии CEM-VBL100 из исходной клеточной линии СЕМ (ATCC CCL 119) описано Беком с соавторами (Beck et al. (1979), Cancer Res., 39, 2070-2077), у которого можно получить указанную клеточную линию и другие МУА-производные линии СЕМ. Клетки линии CEM-VBL100 активно делятся в присутствии 100 нг/мл винбластина или относительно высоких концентраций широкого спектра цитостатических агентов. Для того, чтобы получить моноклональные антитела, обладающие высоким сродством, иммунизацию осуществляют принципиально так же, как это описано Чианфриглия с соавторами (Cianfriglia et al. (1986), Meth. in Enzymol., 121,193-210), с введением адекватных модификаций. Способ иммунизации включает в себя внутрибрюшинное введение 1•107 живых, предварительно необработанных клеток CEM-VBL100, ресуспендированных в фосфатно-буферном растворе (PBS), через каждые 15 дней в течение 10 месяцев. Последнюю иммунизацию проводят, вводя 2•107 клеток (ресуспендированных в PBS) в хвостовую вену и такое же количество клеток - внутрибрюшинно, за три дня до соматического слияния спленоцитов, полученных от обработанных мышей.
После тщательной промывки культуральной средой, не содержащей сыворотки, 25•106 спленоцитов помещают в экспериментальную пробирку с 10•106 миеломными клетками Sp2-01/Ag.14 (ATCC CRL 1581), предварительно обработанными таким же образом, как и лимфоциты.
В дальнейшем смесь клеток селезенки и миеломных клеток обрабатывают так, как это описано Чианфриглия с соавторами (Cianfriglia et al. (1986), Meth. in Enzymol. , 121, 193-210). Кратко, обнаруженные гетерокарионы переносят в планшеты для микротитрования (Costar) для того, чтобы получить после селекции в культуральной среде ГАТ (Г = гипоксантин, А = аминоптерин, Т = тимидин, Gibco) клональные гибридомы. Супернатанты культур из лунок, в которых наблюдается повышенный рост клеток, тестируют на непрямую иммунофлюоресценцию с чувствительной к антибиотикам исходной линией СЕМ (ATCC CCL 119) и одновременно - с вариантом CEM-VBL100 для выявления гибридом, секретирующих иммуноглобулины, избирательно взаимодействующие с иммунизирующими МУА-клетками, но не проявляющих перекрестной реакции с чувствительной к антибиотикам исходной линией. Из отобранных таким образом гибридомных линий выделили линию, известную как MM4.17, которая секретирует иммуноглобулины изотипа IgG2a, k, эффективно взаимодействующие с клетками CEM-VBL100, что показано на основании интенсивного иммунофлюоресцентного сигнала (см. чертеж), выявленного с помощью проточно-цитофлюориметрических исследований. Проточно-цитометрические тесты осуществляли с помощью стандартных методов с флюоресцеин-изотионат-конъюгированным F(Ab')2 анти-мышиным иммуноглобулином G (Cappel, West Chester, PA, USA). После окрашивания клетки фиксировали 1%-ным формальдегидом в PBS, pH 7,2 и анализировали на стендовом проточном цитометре (FACScan, Becton Dickinson), оборудованном 15-нВ аргоновым ионным лазером, испускающим свет с фиксированной длиной волны в 488 нм.
Гибридому MM4.17 засевают в 96-луночные планшеты для микротитрования в концентрации 0,5 клетки на клетку монослоя для обеспечения питательными веществами (питающая подложка), полученными путем промывания брюшной полости мышей линии BALB/c. После культивирования в течение 10-14 дней в каждую лунку добавляют культуральную среду, содержащую HT (гипоксантин-тимидин) (100 мкл), а полученные клоны еще дважды субклонируют так же, как это было описано выше. Субклоны секретируют антитела, имеющие те же свойства, что и антитела, продуцируемые неклонированной линией.
Гибридому ММ4.17 культивируют in vitro с получением из супернатанта больших количеств антител и инъецируют ее сингенным мышам BALB/c, предварительно обработанным неполным стимулятором Фрейнда, способствующим образованию асцитов.
Вместо клеток CEM-VBL100 для иммунизации мышей и отбора гибридомной клеточной линии можно использовать любые сходные МУА-клеточные линии человека.
Пример 2. Специфичность моноклональных антител ММ4.17 по отношению к МУА1-гликопротеину P человека.
Супернатант гибридомной клеточной линии ММ166.4.17 (известной в настоящее время как ММ4.17) связывается только с МУА-вариантами СЕМ-клеток человека (см. Cianfriglia et al. (1990), Int. J. Cancer, 45, 95-103; Beck et al. (1979), Cancer Res., 39, 2070-2077) и совершенно не взаимодействует с исходной чувствительной к антибиотику клеточной линией. После двух циклов клонирования выделяют и охарактеризовывают клонированную гибридомную клеточную линию, секретирующую моноклональные иммуноглобулины антитела IgG2a, k, а из длительно пассируемых безмикоплазмных культур создают банк гибридомных клеток. Размораживают живые образцы из этого банка, и тестируют гибридомные клетки на стабильность, стерильность и специфичность. Создают дополнительные рабочие банки клеток, а из их супернатантов получают сообразные гомогенные партии моноклональных антител в очищенной или грубой форме.
На чертеже представлены профили флюоресценции моноклональных антител ММ4.17 на клетках СЕМ или их МУА-вариантах, проявляющих различную степень относительной устойчивости (ОУ). Эти данные четко свидетельствуют о способности клеток ММ4.17 узнавать очень низкие уровни ОУ (см. Cianfriglia et al. (1990), Int. J.Cancer, 45, 95-103; Wyler et al. (1993), Proceedings of the 84th Annual Meeting of AACR, Abs. No. 1837, page 309) и колебания экспрессии гликопротеина P на клетках МУА. Моноклональные антитела ММ4.17 сильно окрашивают только МУА-варианты различных исследованных клеточных линий человека и не окрашивают сколь бы то ни было существенно как чувствительные, так и устойчивые к антибиотикам клетки, происходящие от грызунов. Специфичность антител ММ4.17 подтверждают также с помощью исследования генетики соматических клеток. Межвидовые гибриды, полученные путем соматической гибридизации чувствительных к антибиотикам клеток грызунов с клетками CEM-VBL100, выщепляют, судя по фенотипу МУА, ген MDR1, а антигенная детерминанта узнается антителами ММ4.17.
Пример 3. Идентификация эпитопа гликопротеина P ,узнаваемого антителами ММ4.17.
Синтезируют пептиды с аминокислотными последовательностями, определенными на основании участков кДНК, кодирующих локализованные вне клетки домены гликопротеина P человека или мышей (Ueda et al. (1987), PNAS, 84, 3004-3008; Shen et al. (1986), Mol.Cell.Biol., 6, 4039-4044; Chenet al. (1987), Cell, 47, 381-389). Указанные пептиды инкубируют с супернатантом культуры гибридомы, секретирующей антитела ММ4.17. Для того чтобы показать, что пептиды, соответствующие остаткам 739-754 четвертой внеклеточной петли гликопротеина P (Последовательность 1), способны взаимодействовать с антителами ММ4.17, используют тест EL1SA. Другие пептиды из предсказанных внеклеточных доменов гликопротеина P не взаимодействуют с антителами. Кроме того, не наблюдается никакого заметного связывания с пептидами мышей, соответствующими эквивалентным участкам предсказанных внеклеточных доменов mdr1 мыши. В частности, несмотря на достаточно высокую степень гомологии между соответствующими пептидными последовательностями человека и мыши, ММ4.17 не дает перекрестной реакции с пептидами, представляющими собой участки четвертой внеклеточной петли mdr1-гликопротеина P мыши.
Для того чтобы определить минимальную последовательность, способную взаимодействовать с антителами ММ4.17, синтезируют перекрывающиеся серии из 70 тетра-декапептидов, представленных в Последовательности 1, и подвергают их тесту ELISA с антителами ММ4.17. Указанные серии состоят из 10 наборов тетра-декапептидов; первый набор включает в себя тетрапептид, соответствующий аминокислотным остаткам 1-4, и декапептид, соответствующий аминокислотным остаткам 1-10 Последовательности 1; второй набор включает в себя тетрапептид, соответствующий аминокислотным остаткам 2-5, и декапептид, соответствующий аминокислотным остаткам 2-11 Последовательности 1, и т.д. для каждого последующего набора тетра-декапептидов. Результаты теста ELISA свидетельствуют о том, что октапептид, состоящий из остатков 2-9, входящих в состав Последовательности 1, представляет собой оптимальный наименьший эпитоп. Утрата первого и даже в еще большей степени последнего треонина приводит к заметной потере способности связываться с антителами ММ4.17. Анализ антител, связывающихся с различными пептидами, четко свидетельствует о том, что:
1. связывание существенно ослабляется в том случае, если оптимальную наименьшую последовательность удлиняют в любом направлении,
2. гексапептид, состоящий из остатков 4-9, входящих в состав Последовательности 1, представляет собой ядро сайта узнавания антителом, поскольку он обнаружен во всех позитивных пептидах,
3. специфичность антител ММ4.17 подчеркивается очень высокими показателями связывания с более короткими пептидами, такими, как пептиды, состоящие из остатков 3-8, 4-9 и 4-8, входящих в состав Последовательности 1.
Пример 4. Специфичность ММ4.17 по отношению к продукту гена MDR1 человека.
МУА-специфичность моноклональных антител ММ4.17 определяли посредством тестирования их связывания с октапептидами, представляющими собой аминокислотные последовательности, предсказанные для других генов, кодирующих P-гликопротеины млекопитающих и выделенные по их гомологии со специфическими эпитоп-образующими аминокислотными последовательностями оптимально минимального белка. Моноклональные антитела ММ4.17 не связываются как с соответствующим пептидом (Последовательность 4) гена MDR3 - высоко изменчивого второго члена семейства человеческих генов, контролирующих МУА (идентичные последовательности обнаружены в гликопротеинах mdr2 мыши и pgp3 хомяка), так и с пептидами P-гликопротеинов mdr1 и mdr3 мыши, а также pgp1 и pgp2 хомяка, которые наиболее сходны с оптимальным минимальным эпитоп-образующим пептидом MDR1.
Пример 5. Определение нуклеотидной последовательности и аминокислотной последовательности вариабильного района тяжелой цепи антител ММ4.17.
Последовательность кДНК, кодирующая вариабильный домен и его гипервариабильную часть тяжелой цепи антител ММ4.17, получали, как это описано (Orlandi et al. (1989), PNAS, 86, 3833-3837). Последовательность кДНК и соответствующая предсказанная аминокислотная последовательность приведены в Последовательности 2, а аминокислотная последовательность в отдельности приведена в Последовательности 3. Определяющими Комплементарность Районами (ОКР) являются остатки 32- 36 включительно (в случае ОКР1), остатки 51-67 (в случае ОКР2) и остатки 100-111 (в случае ОКРЗ) аминокислотной последовательности тяжелой цепи вариабильного домена, представленной в Последовательностях 2 и 3.
Пример 6. Сравнение антител ММ4.17 и первичных моноклональных антител по сродству их связывания с гликопротеином P на МУА- и чувствительных к антибиотикам клетках.
Моноклональные антитела ММ4.17 сравнивали с первичными моноклональными антителами C219, C494, JSB1, MRK-16 и MC57 по связыванию с клетками CEM, CEM-VBL10 и CEM-VBL100 в экспериментах по проточной цитометрии, в которых измеряли окрашивающие способности флюоресцентно-меченных вариантов антител. Полученные результаты представлены в приведенной ниже таблице. Моноклональные антитела использовали в очищенной форме при одной и той же концентрации белка (10 мкг/мл). Полученные результаты четко свидетельствуют о том, что моноклональные антитела ММ4.17 имеют более высокое сродство связывания с клеточными линиями, проявляющими МУА, и являются моноклональными антителами, несомненно связывающимися с клеточной линией CEM-VBL10.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
1. ОБЩАЯ ИНФОРМАЦИЯ:
1. ЗАЯВИТЕЛЬ:
А. НАЗВАНИЕ: Institute Superiore di Sanita
Б. УЛИЦА: Viale Regina Elena 299
В. ГОРОД: 00161 Rome
Д. СТРАНА: Italy
Е. ПОЧТОВЫЙ ИНДЕКС: 00161
Ж. ТЕЛЕФОН: 003964444990
3. ТЕЛЕФАКС: 003964440067
И. ТЕЛЕКС: RM610071
II. НАЗВАНИЕ ИЗОБРЕТЕНИЯ: Моноклональные антитела к гликопротеину P
III. КОЛИЧЕСТВО ПОСЛЕДОВАТЕЛЬНОСТЕЙ: 4
IV. ПРИЕМЛЕМАЯ ДЛЯ КОМПЬЮТЕРНОГО АНАЛИЗА ФОРМА:
А. ТИП НОСИТЕЛЯ: Дискета
Б. КОМПЬЮТЕР: IBM PC-совместимый
В. ОПЕРАТИВНАЯ СИСТЕМА: PC-DOS/MS-DOS
Д. ПРОГРАММНОЕ ОБЕСПЕЧЕНИЕ: Patentin Release #1.0, Version #1.25 (EPO)
V. ДАННЫЕ НАСТОЯЩЕЙ ЗАЯВКИ:
НОМЕР ЗАЯВКИ: WO PCT/EP93/
2. ИНФОРМАЦИЯ О ПОСЛЕДОВАТЕЛЬНОСТИ 1:
1. ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
А. ДЛИНА: 16 аминокислот
Б. ТИП: последовательность аминокислот
Г. ТОПОЛОГИЯ: линейная
II. ТИП МОЛЕКУЛЫ: пептид
III. ГИПОТЕТИЧЕСКАЯ: Нет
V. ТИП ФРАГМЕНТА: внутренний
VI. ИСТОЧНИК ОРИГИНАЛА:
А. ОРГАНИЗМ: Homo sapiens
XI. ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ 1: представлено в конце описания.
2. ИНФОРМАЦИЯ О ПОСЛЕДОВАТЕЛЬНОСТИ 2:
I. ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
А. ДЛИНА: 369 пар оснований
Б. ТИП: нуклеиновая кислота
В. КОЛИЧЕСТВО НИТЕЙ: одна
Г. ТОПОЛОГИЯ: линейная
II. ТИП МОЛЕКУЛЫ: ДНК (геномная)
III. ГИПОТЕТИЧЕСКАЯ: Нет
IV. АНТИСМЫСЛОВАЯ: Нет
V. ТИП ФРАГМЕНТА: внутренний
VI. ИСТОЧНИК ОРИГИНАЛА:
А. ОРГАНИЗМ: Mus musculus
Б. ЛИНИЯ: BALB/C
IX. ОСОБЕННОСТЬ:
А. НАЗВАНИЕ/КЛЮЧ: CDS
Б. ПОЛОЖЕНИЕ: 1...369
XI. ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ 2: представлено в конце описания.
ИНФОРМАЦИЯ О ПОСЛЕДОВАТЕЛЬНОСТИ 3:
I. ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
А. ДЛИНА: 123 аминокислоты
Б. ТИП: последовательность аминокислот
Д. ТОПОЛОГИЯ: линейная
II. ТИП МОЛЕКУЛЫ: белок
XI. ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ 3: представлено в конце описания.
2. ИНФОРМАЦИЯ О ПОСЛЕДОВАТЕЛЬНОСТИ 4:
I. ХАРАКТЕРИСТИКИ ПОСЛЕДОВАТЕЛЬНОСТИ:
А. ДЛИНА: 8 аминокислот
Б. ТИП: последовательность аминокислот
Д. ТОПОЛОГИЯ: линейная
II. ТИП МОЛЕКУЛЫ: пептид
III. ГИПОТЕТИЧЕСКАЯ: Нет
V. ТИП ФРАГМЕНТА: внутренний
VI. ИСТОЧНИК ОРИГИНАЛА:
А. ОРГАНИЗМ: Homo sapiens
XI. ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТИ 4: представлено в конце описания.





Моноклональное антитело специфично и с высокой эффективностью связывает гликопротеин Р человека, а потому может быть использовано для выявления МУА-клеток человека, если они составляют лишь очень малую долю клеточной популяции или вырабатывают человеческий гликопротеин Р лишь в очень малых количествах. Указанное моноклональное антитело может представлять собой целое антитело или антиген-связывающий фрагмент и может быть получено с помощью техники гибридом иди рекомбинантной ДНК. Моноклональное антитело пригодно для выявления или очистки клеток, вырабатывающих человеческий гликопротеин Р, в частности, когда оно присутствует в гетерогенных клеточных популяциях, а также для тестирования множественной устойчивости к антибиотикам у клеток, например у раковых клеток. 4 з.п. ф-лы, 1 табл., 1 ил.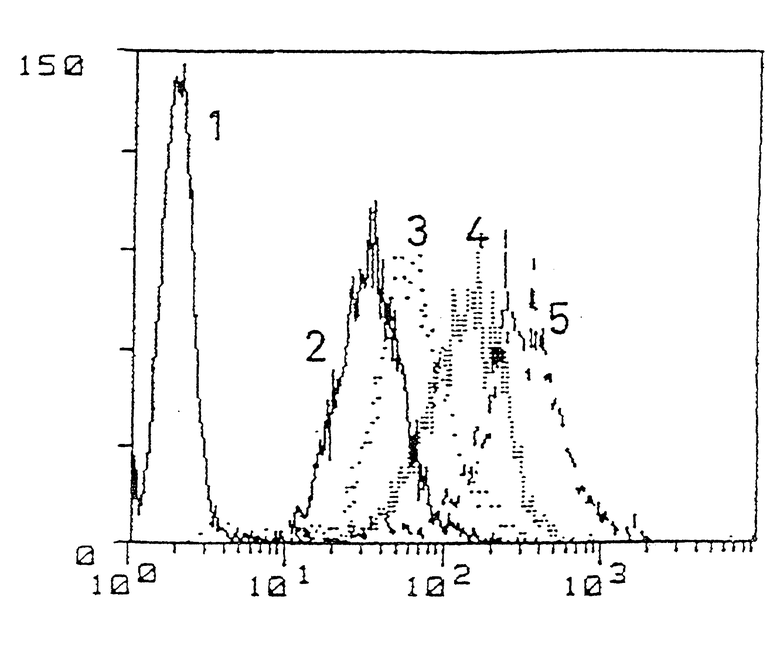
| Способ изготовления фанеры-переклейки | 1921 |
|
SU1993A1 |

