Перекрестные ссылки на связанные заявки
По настоящей заявке испрашивается приоритет предварительной заявки США №60/851183, поданной 12 октября 2006 года, содержание которой включено в настоящий документ в качестве ссылки в полном объеме.
Область изобретения
Настоящее изобретение в основном относится к лечению злокачественных заболеваний. Более конкретно, настоящее изобретение относится к полинуклеотидам, кодирующим слитые белки, которые содержат по меньшей мере часть ассоциированного с опухолью полипептида теломеразной обратной транскриптазы (TERT). Настоящее изобретение также относится к рекомбинантным векторам и хозяевам, содержащим указанные полинуклеотиды, к очищенным слитым белкам и способам индукции или усиления иммунной реакции в ответ на белковый продукт гена TERT с использованием композиций и молекул, описываемых в настоящем документе.
Предпосылки изобретения
Вакцинация стала стандартной процедурой для профилактики множества инфекционных заболеваний. Применение вакцин для других заболеваний, таких как злокачественные опухоли, является в настоящее время привлекательной возможностью ввиду прогресса в молекулярной инженерии и лучшего понимания иммунологии опухолей.
Злокачественные опухоли представляют собой одну из ведущих причин смертности в мире. Несмотря на множество связанных со злокачественными опухолями исследований, традиционные способы лечения, сочетающие хирургию, радиационное излучение и химиотерапию, часто не обеспечивают эффективное лечение злокачественных опухолей на поздних сроках. Также недоступными остаются надежные способы профилактики.
Злокачественная опухоль, как правило, включает нарушение функции генов, участвующих в регуляции клеточного цикла или клеточной пролиферации, таких как гены факторов роста и их рецепторов, онкогены и гены опухолевых супрессоров. Продукты многих из этих генов экспрессированы на поверхности широкого спектра опухолевых клеток и, таким образом, являются опухолеспецифическими антигенами (TAA). Во многих экспериментальных моделях показано, что введение кодирующих TAA генов непосредственно индивиду вызывает защитный иммунный ответ против TAA, делая эти молекулы мишенью вакцинотерапии. Однако поскольку многие из этих генных продуктов также экспрессируются в нормальных клетках, хотя и на более низких уровнях, было показано, что многие иммунологические способы лечения, направленные на TAA, являются неэффективными.
Гены, кодирующие некоторые опухолеспецифические антигены (TAA), выделены, охарактеризованы и встроены в генетические векторы, такие как плазмидные ДНК и вирусные векторы. Один из опухолеспецифических антигенов, который вовлечен в патогенез злокачественной опухоли, представляет собой теломеразу (TERT).
Теломераза представляет собой ДНК-полимеразу, функция которой в норме заключается в сохранении длины теломер на концах хромосом. В течение роста нормальной клетки к 5'-концу ДНК присоединяется РНК-праймер и начинается репликация. После удаления РНК-праймера на участке образующейся дочерней цепи ДНК остается пропуск. Таким образом, репликация линейной цепи ДНК обычными полимеразами приводит к уменьшению длины теломеры в каждом следующем раунде репликации. Такое уменьшение длины теломеры отвечает за нормальное старение или созревание клеток у нормальных соматических клеток человека.
Теломераза представляет собой рибонуклеопротеин, содержащий компонент РНК и каталитический белковый компонент (теломеразную обратную транскриптазу). Каталитический компонент теломеразы человека описан Meyerson et al. (Cell 1197: 785-95 (1990) и Nakamura et al. (Science 277: 955-59 (1997)). Фермент TERT использует свой компонент РНК в качестве матрицы для синтеза ДНК теломеры, таким образом, позволяя теломерам поддерживать их длину на протяжении последовательных стадий клеточного роста. Такое поддержание длины теломер в течение многих циклов пролиферации позволяет клеткам избежать нормального старения и стать бессмертными, позволяя опухоли расти и метастазировать в течение длительных периодов времени. Так как теломераза дает клеткам репликационное бессмертие, теломеразная активность выявлена в линиях злокачественных клеток и широком диапазоне типов опухолей (Kim et al. Science 266: 2011-15 (1994)). Наоборот, в нормальных тканях человека и в культурах нормальных соматических клеток теломераза не активна или только временно экспрессируется на низких уровнях. Сочетание сверхэкспрессии теломеразы в большинстве типов злокачественных опухолей, а также низкая или отсутствующая экспрессия в нормальных клетках, делает TERT мишенью для терапии и/или профилактики заболеваний, ассоциированных с нарушенной клеточной пролиферацией, таких как злокачественная опухоль.
В настоящее время возможно получение специфичной для теломеразы вакцины, так как клонированы и охарактеризованы каталитический и РНК компоненты теломеразы (см., например, патент США 6166178). Однако получение и промышленное внедрение многих вакцин затруднено из-за сложностей, связанных с получением высоких уровней экспрессии желаемого иммуногена в успешно трансформированных организмах-хозяевах. Получение эффективных вакцин на основе ДНК также затруднено из-за отсутствия возможности получить иммунный ответ достаточной силы у индивидуумов, получаемых в клинических условиях лечение для регрессии опухоли. Таким образом, несмотря на идентификацию нуклеотидных последовательностей дикого типа, кодирующих описанные выше белки теломераз, крайне желательно получить вакцину, которая могла бы при введении млекопитающему вызывать усиленный специфичный для TERT иммунный ответ по сравнению с полноразмерной кДНК TERT дикого типа. Также желательно разработать способы лечения или профилактики ассоциированных с TERT злокачественных опухолей, в которых используются молекулы нуклеиновой кислоты или белки, которые безопасно и эффективно усиливают специфичный для TERT иммунный ответ.
Сущность изобретения
Как указано выше, экспрессия гена опухолеспецифического антигена теломеразной обратной транскриптазы (TERT), как правило, связана с развитием или наличием аденокарцином, включая колоректальные карциномы. В связи с этим настоящее изобретение относится к композициям и способам индукции или усиления иммунного ответа к белковым продуктам, экспрессируемым геном TERT человека (hTERT). В частности, настоящее изобретение относится к полинуклеотидам, кодирующим слитые белки, которые содержат белок hTERT или его вариант, слитый со значительной частью субъединицы B термолабильного энтеротоксина (LTB) E. coli. Настоящее изобретение также относится к рекомбинантным векторам, включающим, без ограничения перечисленным, аденовирусные и плазмидные векторы, содержащие указанные полинуклеотиды, и к клеткам-хозяевам, содержащим указанные рекомбинантные векторы. Также в настоящем описании представлены очищенные слитые белки, кодируемые полинуклеотидами по изобретению. Слитые белки hTERT-LTB и полинуклеотиды, кодирующие указанные слитые белки, могут использоваться в качестве вакцин для профилактики и/или лечения ассоциированной с теломеразой злокачественной опухоли. Указанные вакцины можно использовать как монотерапию или как часть схемы лечения, которая включает введение второй вакцины, такой как полинуклеотидная, клеточная, белковая или пептидная вакцина, или включает проведение лучевой терапии или химиотерапии.
В предпочтительных вариантах осуществления настоящего изобретения нуклеотидная последовательность, кодирующая hTERT, и/или нуклеотидная последовательность, кодирующая LTB, содержит кодоны, которые оптимизированы для высоких уровней экспрессии в клетке-хозяине человека. Другими словами, в определенных вариантах осуществления характер использования кодонов полинуклеотидной последовательности совпадает с характером использования кодонов высокоэкспрессированных генов млекопитающих и/или человека.
В другом аспекте изобретение относится к экспрессирующей конструкции, содержащей нуклеотиды, кодирующие hTERT-LTB. В предпочтительных вариантах осуществления этой части изобретения конструкция содержит перед кодирующей последовательностью гена TERT лидерную последовательность TPA для гарантии секреции слитого белка TERT-LTB.
В дополнительных предпочтительных вариантах осуществления настоящего изобретения теломеразную каталитическую активность антигена теломеразы инактивируют, чтобы кодируемый слитый белок TERT был более безопасным при вакцинации, чем TERT дикого типа. Ферментативную активность слитого белка TERT можно инактивировать введением в нуклеотидную последовательность, кодирующую TERT, мутаций/делеций. В конкретных вариантах осуществления изобретения, представленных в качестве примеров, нуклеотиды мутированы с изменением конкретных аминокислот D712A и V713I в последовательности белка TERT человека, и D702A и V703I в последовательности белка TERT мыши.
Настоящее изобретение, кроме того, относится к аденовирусным и плазмидным векторам, содержащим нуклеотидную последовательность, кодирующую слитый белок hTERT-LTB. В настоящем изобретении также описано использование аденовирусных и плазмидных векторов в иммуногенных композициях и вакцинах для профилактики и/или лечения ассоциированной с hTERT злокачественной опухоли.
Также представлены аденовирусные векторы, содержащие нуклеотидную последовательность, кодирующую белок TERT или вариант белка TERT человека. Варианты, подходящие для настоящего аспекта по изобретению, содержат мутации, функцией которых является элиминация теломеразной каталитической активности. В предпочтительных вариантах осуществления этой части изобретения аденовирусный вектор представляет собой вектор Ad6. В других предпочтительных вариантах осуществления вектор Ad представляет собой вектор Ad5.
Настоящее изобретение, кроме того, относится к способам профилактики развития злокачественной опухоли у пациента или лечения пациента с ассоциированной с теломеразой опухолью путем индукции иммунного ответа на белок TERT за счет введения вакцины или фармацевтической композиции, содержащей слитые последовательности TERT или слитые белки TERT, предоставленные изобретением. В предпочтительных вариантах осуществления способов по настоящему документу иммунный ответ усилен по сравнению с ответом, индуцируемым TERT дикого типа.
Как используется в настоящем описании и в приложенной формуле изобретения, если в контексте явно не указано иначе, формы единственного числа включают указание на множественное число.
Как используется в настоящем описании и в приложенной формуле изобретения, используют следующие определения и сокращения:
Термин "промотор" относится к сайту распознавания на цепи ДНК, с которой связывается РНК-полимераза. Промотор образует инициирующий комплекс с РНК-полимеразой для инициации и запуска транскрипционной активности. Комплекс может быть модифицирован активирующими последовательностями, называемыми "энхансерами", или ингибирующими последовательностями, называемыми "сайленсерами".
Термин "кассета" относится к нуклеотидной или генной последовательности, которая экспрессируется с вектора, например, нуклеотидная или генная последовательность, кодирующая слитую последовательность hTERT(AI)-LTB. Как правило, кассета содержит генную последовательность, которая может быть встроена в вектор, который в некоторых вариантах осуществления содержит регуляторные последовательности для экспрессии нуклеотидной или генной последовательности. В других вариантах осуществления регуляторные последовательности для своей экспрессии содержит сама нуклеотидная или генная последовательность. В дополнительных вариантах осуществления вектор содержит некоторые регуляторные последовательности, а нуклеотидная или генная последовательность содержит другие регуляторные последовательности. Например, вектор может содержать промотор для транскрипции нуклеотидной или генной последовательности, а нуклеотидная или генная последовательность содержит последовательность терминации транскрипции. Регуляторные последовательности, которые могут содержаться в векторе, включают, но не ограничиваются ими, энхансеры, последовательности терминации транскрипции, последовательности акцепторных участков и донорных участков сплайсинга, интроны, последовательности связывания рибосом и последовательности добавления поли(A).
Термин "вектор" относится к некоторым средствам, с помощью которых фрагменты ДНК могут быть введены в организм хозяина или в ткань хозяина. Существуют различные типы векторов, включая плазмиды, вирусы (в том числе аденовирусы), бактериофаги и космиды.
Как используется по отношению к аденовирусным векторам, термин "первое поколение" описывает аденовирусные векторы, которые дефектны по репликации. У аденовирусных векторов первого поколения, как правило, удалена или инактивирована генная область E1 и, предпочтительно, удалена или инактивирована генная область E3.
Обозначение "pV1JnsA/TPA-mTERT(AI)-LTBopt" относится к описываемой в настоящем документе плазмидной конструкции, содержащей предранний (IE) промотор CMV с интроном A, полноразмерный ген TERT мыши с оптимизированными кодонами, слитый с геном LTB с оптимизированными кодонами, и полученные из бычьего гормона роста последовательности полиаденилирования и терминации транскрипции (см. пример 2). Дополнительно, со стороны 5' от нуклеотидной последовательности, кодирующей mTERT-LTB, находится лидерная последовательность, кодирующая сигнальную последовательность активатора тканевого плазминогена (TPA) человека. Обозначение "AI" указывает на то, что в последовательность для функционального устранения теломеразной каталитической активности TERT введены мутации. Обозначение "pV1JnsA/mTERT(AI)opt" относится к конструкции, в основном такой же, как описано выше, за исключением того, что конструкция содержит оптимизированную для мыши нуклеотидную последовательность TERT, которая не слита с LTB или с TPA.
Обозначение "pV1JnsA/TPA-hTERT(AI)-LTBopt" относится к описываемой в настоящем документе плазмидной конструкции, содержащей предранний (IE) промотор CMV с интроном A, полноразмерный ген TERT человека с оптимизированными кодонами, слитый с геном LTB с оптимизированными кодонами, и полученные из бычьего гормона роста последовательности полиаденилирования и терминации транскрипции (см. пример 2). Дополнительно, со стороны 5' от нуклеотидной последовательности, кодирующей hTERT-LTB, находится лидерная последовательность, кодирующая сигнальную последовательность активатора тканевого плазминогена (TPA) человека. Последовательность hTERT в данной конструкции содержит мутации для функционального устранения теломеразной каталитической активности.
Обозначения "Ad6/TPAmTERT(AI)-LTBopt" и "Ad6/hTERT(AI)" относятся к двум описываемым в настоящем документе конструкциям, которые содержат геном аденовируса Ad6 с делецией областей E1 и E3. В конструкции "Ad6/TPAmTERT(AI)-LTBopt" область E1 заменена геном TERT-LTB мыши с оптимизированными кодонами в параллельной E1 ориентации под контролем промотора CMV человека без интрона A, с последующим сигналом полиаденилирования бычьего гормона роста. Кодирующая TERT последовательность содержит мутации для устранения теломеразной каталитической активности. Конструкция со стороны 5' от кодирующей TERT(AI)-LTB нуклеотидной последовательности дополнительно содержит последовательности, кодирующие сигнальную последовательность активатора тканевого плазминогена (TPA) человека. Конструкция "Ad6/hTERT(AI)" в основном является такой же, как описано выше, за исключением того, что область E1 генома Ad6 заменена кДНК последовательностью TERT, где указанная последовательность содержит мутации для устранения ферментативной активности.
Сокращение "LTB", как правило, относится к субъединице B термолабильного энтеротоксина E. coli или значительной ее части, включая субъединицы, которые укорочены на C-конце или N-конце, но сохраняют адъювантную активность, а также субъединицы, содержащие внутренние аминокислотные вставки, делеции или замены, но сохраняющие адъювантную активность (Fingerut et al. Vaccine 23:4685-4696(2005)).
Как используют в настоящем документе, "слитый белок" относится к белку, имеющему по меньшей мере два гетерологичных полипептида, ковалентно связанных, где один полипептид получен из одной белковой последовательности или домена, а другой полипептид получен из второй белковой последовательности или домена. Слитые белки по настоящему изобретению содержат полипептид TERT или его фрагмент или вариант и второй полипептид, содержащий значительную часть LTB. Полипептид TERT, его фрагмент или вариант могут представлять собой TERT человека или гомолог TERT других видов, например мыши. Полипептиды, составляющие слитый белок, предпочтительно связаны N-концом с C-концом. Слияние полипептида TERT и полипептида LTB можно проводить в любом порядке. В некоторых вариантах осуществления настоящего изобретения C-конец полипептида TERT слит с N-концом LTB, как проиллюстрировано на фигурах 1 и 2. Однако также рассматриваются слитые белки, в которых субъединица LTB слита с N-концом полипептида TERT.
Термин "слитый белок TERT" или "слитый белок TERT-LTB" используют в настоящем документе взаимозаменяемо как общий термин, который относится к слиянию, как описано выше, которое включает полипептид TERT или его фрагмент или вариант, слитый с полипептидом LTB. Термин "слитая последовательность TERT-LTB" относится к последовательности нуклеиновой кислоты, в которой по меньшей мере часть гена TERT слита со значительной частью субъединицы LTB термолабильного энтеротоксина E. coli. Термин "слитый белок TERT-LTB" относится к полипептиду, кодируемому слитой последовательностью TERT-LTB, как описано. Следует понимать, что термины "слитая последовательность TERT-LTB" и "слитый белок TERT-LTB" также относятся к их фрагментам, их гомологам и их функциональным эквивалентам (совместно обозначаемым как "варианты"), таким как варианты, в которых одна или несколько аминокислот вставлены, удалены или заменены другой аминокислотой(ами). Слитые последовательности TERT-LTB по настоящему изобретению после введения млекопитающему, такому как человек, могут стимулировать иммунный ответ хелперных T-клеток или цитотоксических T-клеток по меньшей мере так же, как последовательность hTERT "дикого типа". В предпочтительных вариантах осуществления изобретения слитая последовательность TERT-LTB может усиливать иммунный ответ по сравнению с hTERT дикого типа.
Термин "лечение" относится к терапевтическому лечению и профилактическим или превентивным мерам. Нуждающиеся в лечении включают тех, у кого уже имеется нарушение, а также тех, кто предрасположен к нарушению, или тех, у кого необходимо предупредить нарушение. Как используют в настоящем описании, "лечение" также включает снижение вероятности возникновения нарушения, а также снижение тяжести нарушения у тех, кто уже болен.
"Нарушение" представляет собой любое состояние, которое можно облегчить путем лечения молекулами по настоящему изобретению, включая молекулы нуклеиновой кислоты, описываемые в настоящем документе, и слитые белки, кодируемые указанными молекулами нуклеиновой кислоты. В термин "нарушение" включены хронические и острые нарушения или заболевания, включая патологические состояния, которые вызывают у млекопитающего предрасположение к рассматриваемому нарушению. Молекулы по настоящему изобретению предназначены для использования в качестве лекарственных средств для нарушений или патологических состояний, характеризуемых нарушенной клеточной пролиферацией, включая, но не ограничиваясь перечисленным, рак молочной железы, колоректальный рак и рак легких.
Термин "эффективное количество" обозначает, что введено достаточное количество композиции вакцины для продуцирования достаточных уровней полипептида так, чтобы в результате получить иммунный ответ. Специалисту в данной области понятно, что такой уровень может изменяться.
"Консервативная аминокислотная замена" относится к замене одного аминокислотного остатка другим, химически сходным, аминокислотным остатком. Примеры таких консервативных замен представляют собой: замену одного гидрофобного остатка (изолейцин, лейцин, валин или метионин) другим; замену одного полярного остатка другим полярным остатком того же заряда (например, аргинин лизином; глутаминовая кислота аспарагиновой кислотой).
"hTERT" относится к антигену теломеразной обратной транскриптазы человека или нуклеотидам, кодирующим антиген теломеразной обратной транскриптазы человека. "hTERTopt" относится к нуклеотидной последовательности с оптимизированными кодонами, кодирующей антиген теломеразной обратной транскриптазы человека.
"По существу сходная" означает, что данная нуклеиновая кислота или аминокислотная последовательность идентична по меньшей мере на 75%, предпочтительно по меньшей мере на 85%, более предпочтительно по меньшей мере на 90% и еще более предпочтительно по меньшей мере на 95% эталонной последовательности. В настоящем изобретении, как следует из контекста текста, эталонная последовательность может представлять собой подходящие части нуклеотидной или аминокислотной последовательности TERT человека дикого типа, как приведено в SEQ ID NO:11 и 12, соответственно, или нуклеотидной или аминокислотной последовательности LTB, как приведено в SEQ ID NO:13 (оптимизированная последовательность) и 14. Эталонная последовательность также может представлять собой, например, последовательность TERT дикого типа макака-резус или мыши. Таким образом, последовательность белка TERT, которая "по существу сходна" c белком дикого типа TERT человека или его фрагментом, по меньшей мере на 75% идентична эталонному фрагменту TERT дикого типа человека по длине фрагмента, предпочтительно на 85%, более предпочтительно на 90% и еще более предпочтительно на 95%. Является ли данная аминокислотная последовательность или нуклеотидная последовательность mTERT, hTERT или LTB "по существу сходной" с эталонной последовательностью, можно определить, например, путем сравнения информации последовательностей с использованием программного обеспечения для анализа последовательностей, такого как компьютерная программа GAP, версии 6.0, доступная от University of Wisconsin Genetics Computer Group (UWGCG). В программе GAP используют способ выравнивания Needleman и Wunsch (J. Mol. Biol. 48:443, 1970) с изменениями Smith и Waterman (Adv. Appl. Math. 2:482, 1981).
"Значительная часть" гена, его варианта, фрагмента или субъединицы означает часть, которая составляет по меньшей мере 50%, предпочтительно 75%, более предпочтительно 90% и еще более предпочтительно 95% эталонной последовательности.
"Ген" относится к молекуле нуклеиновой кислоты, нуклеотидная последовательность которой кодирует молекулу полипептида. Гены могут представлять собой непрерывные нуклеотидные последовательности или они могут включать такие вставочные сегменты, как интроны, промоторные области, сайты сплайсинга и повторяющиеся последовательности. Ген может представлять собой РНК или ДНК. Предпочтительный ген представляет собой ген, кодирующий полипептид по изобретению.
Термин "нуклеиновая кислота" или "молекула нуклеиновой кислоты" относится к рибонуклеиновой кислоте (РНК) или дезоксирибонуклеиновой кислоте (ДНК), зондам, олигонуклеотидам, их фрагментам или частям и праймерам.
"TERT дикого типа" или "белок дикого типа" или "белок д.т." относится к белку, содержащему природную последовательность аминокислот, или его варианту. Аминокислотная последовательность дикого типа TERT человека приведена в SEQ ID NO:12. Аминокислотная последовательность дикого типа TERT человека описана ранее (патент США №6166178; патент США 6261836; патент США №6927285; патентная заявка США 2003-0096344; Meyerson et al, Cell 90: 785-95 (1997); Nakamura et al, Science 277: 955-59 (1997)).
"Ген TERT дикого типа" относится к гену, содержащему нуклеотидную последовательность, кодирующую природный белок TERT, включая белки человеческого происхождения или белки, полученные из другого организма, включая, но не ограничиваясь перечисленным, другие млекопитающие, такие как крыса, мышь и макак-резус. Нуклеотидная последовательность гена TERT человека доступна в данной области (выше).
Обозначение "TERT(AI)" относится к последовательности теломеразной обратной транскриптазы, которая содержит мутации, устраняющие или уменьшающие теломеразную каталитическую активность.
Термин "млекопитающее" относится к любому млекопитающему, включая человека.
Аббревиатура "Ag" означает антиген.
Аббревиатура "ORF" означает открытую рамку считывания гена.
Краткое описание чертежей
На фиг.1 приведены нуклеотидная последовательность (SEQ ID NO:1) и аминокислотная последовательность (SEQ ID NO:2) иллюстративной слитой последовательности TPA-hTERT(AI)-LTBopt.
На фиг.2 приведена нуклеотидная последовательность (SEQ ID NO:3) и аминокислотная последовательность (SEQ ID NO:4) иллюстративной слитой последовательности TPA-mTERT(AI)-LTBopt.
На фиг.3 приведены результаты анализа ELIspot IFNγ у мышей BALB/c, вакцинированных повторными инъекциями плазмиды pV1J/TPA-mTERT(AI)-LTBopt. В каждой группе 5 мышей использовали для мониторинга иммунного ответа, направленного против mTERT или LTB, с использованием пулов пептидов. Нанесенные на диаграмму данные получены у 6 разных мышей (незакрашенная окружность). Указаны значения геометрических средних (закрашенные окружности). У мышей, обработанных препаратом отрицательного контроля, T-клеточного ответа на mTERT не выявлено (не показано). См. пример 7.
На фиг.4 приведены результаты внутриклеточного окрашивания на IFNγ в спленоцитах вакцинированных мышей BALB/c. CD4+ и CD8+ T-клеточный ответ на мышиный TERT измеряли с использованием пулов перекрывающихся пептидов, которые покрывают весь белок. Также мониторингу подвергали иммунный ответ на LTB. Нанесенные на диаграмму данные получены у 6 разных мышей (закрашенные треугольники). Указаны значения геометрических средних (прямая линия). У мышей, обработанных препаратом отрицательного контроля, T-клеточного ответа на TERT не выявлено (не показано). См. пример 7.
На фиг.5 приведены результаты высвобождения 51Cr при CTL цитолизе эффекторными T-клетками клеток-мишеней 4T1, импульсно меченных специфичным для CD8+ T-клеток пептидом mTERTaa167 (AYQVCGSPL; SEQ ID NO:20). Эффекторные клетки получены из спленоцитов двух иммунизированных мышей BALB/c, и их повторно стимулировали in vitro специфичным пептидом. Результаты представлены в виде процента специфического цитолиза при различных соотношениях эффектор/мишень. См. пример 8.
На фиг.6 показана индукция иммунного ответа на TERT мыши. Мышей C57BL/6 иммунизировали 5 еженедельными инъекциями плазмиды pV1J/TPA-mTERT(AI)-LTBopt. Иммунный ответ оценивали по спленоцитам мышей анализом ELIspot IFNγ с использованием пулов пептидов TERT мыши. Ответ антигенспецифических CD4+ и CD8+ T-клеток приведен в виде значений геометрических средних 6 иммунизированных мышей, также приведено стандартное отклонение. См. пример 7.
На фиг.7 показана индукция специфического иммунного ответа на mTERT у мышей BALB/c. Мышей иммунизировали 5 еженедельными инъекциями конструкций pV1J-mTERT(AI)opt или pV1J-TPAmTERT(AI)-LTBopt. Иммунный ответ определяли путем внутриклеточного окрашивания на IFNγ в PBMC с использованием CD8+ эпитопа, соответствующего пептидной последовательности mTERTaa167 (AYQVCGSPL; SEQ ID NO:20). Нанесенные на диаграмму данные получены у 8 разных мышей (закрашенные треугольники). Приведены значения геометрических средних (прямые линии).
На фиг.8 показано, что при инъекции аденовирусных векторов, экспрессирующих mTERT(AI)-LTBopt, усиливается специфический иммунный ответ на TERT. Группы из 10 мышей BALB/c подвергали 5 еженедельным инъекциям pV1J/TPA-mTERT(AI)-LTBopt (50 мкг/инъекцию). Через две недели после последней инъекции плазмидной ДНК мышей повторно стимулировали одной дополнительной инъекцией плазмидной ДНК (ДНК-ЭП) или 1010 в.ч. Ad6/TPA-mTERT(AI)-LTBopt (Ad). Иммунный ответ на TERT подвергали мониторингу посредством ICS на IFNγ в PBMC. Нанесенные на диаграмму данные получены у 10 разных мышей (закрашенные ромбы). Также приведены значения геометрических средних каждой группы (прямые линии). См. пример 10.
На фиг.9 приведен протокол, используемый для анализа индуцированного DMH канцерогенеза и вакцинации у мышей BALB/c. DMH вводили в/в еженедельно. Вакцинацию pV1J/TPA-mTERT(AI)-LTBopt проводили в указанные моменты времени. Анализ кишечных очагов проводили на 12 неделе. См. пример 11.
На фиг.10 показано противоопухолевое действие pV1J/TPA-mTERT(AI)-LTBopt при индуцированном DMH канцерогенезе. Мышей BALB/c обрабатывали DMH и проводили ДНК-ЭП, как указано на фигуре 9. Анализ ACF и образования аденомы в кишечнике проводили на 12 неделе от начала обработки. Количество ACF и очагов аденомы у вакцинированных мышей сравнивали с количеством ACF и очагов аденомы, выявленным у невакцинированных контрольных животных. См. пример 11.
На фиг.11 приведен протокол, используемый для анализа индуцированного DMH канцерогенеза и вакцинации у мышей BALB/c. DMH вводили в/в еженедельно. Вакцинацию посредством ДНК и Ad, содержащих pV1J/TPA-mTERT-LTB, проводили в указанные моменты времени. Анализ кишечных очагов проводили на 12 неделе. См. пример 11.
На фиг.12 показано противоопухолевое действие примирования ДНК/повторного стимулирующего введения Ad, содержащих pV1J/TPA-mTERT(AI)-LTBopt, на количество опухолевых очагов. Мышей BALB/c обрабатывали DMH и проводили ДНК-ЭП, как указано на фигуре 9. Анализ ACF и образования аденомы в кишечнике проводили на 30 неделе от начала обработки. Количество ACF и очагов аденомы у вакцинированных мышей сравнивали с количеством ACF и очагов аденомы, выявленным у невакцинированных контрольных животных. См. пример 11.
На фиг.13 показано противоопухолевое действие примирования ДНК/повторного стимулирующего введения Ad, содержащих pV1J/TPA-mTERT-LTB(AI)opt, на объем и стадию дифференцировки опухолевых очагов. Мышей BALB/c обрабатывали DMH и проводили ДНК-ЭП, как указано. Объем (панель A) и стадию гистологической дифференцировки (панель B) аденом кишечника, присутствующих у вакцинированных и контрольных мышей, анализировали через 30 недель после начала обработки. Опухоли классифицировали следующим образом: G1: хорошо дифференцированная аденокарцинома; G2: умеренно дифференцированная аденокарцинома; G3: плохо дифференцированная аденокарцинома. См. пример 11.
На фиг.14 показана индукция иммунного ответа на TERT человека. Трансгенных мышей HHD иммунизировали двумя инъекциями с двухнедельным интервалом (ДНК-ДНК) плазмиды pV1J/TPA-hTERT(AI)-LTBopt или одной инъекцией плазмиды pV1J/TPA-hTERT(AI)-LTBopt, с последующей одной инъекцией Ad6hTERT(AI) 1010 в.ч. (ДНК/Ad) через две недели. Иммунный ответ оценивали в PBMC мыши анализом ICS на IFNγ. PBMC мышей инкубировали c CD8 иммунодоминантным для аллеля HLA-A2 пептидом hTERT865 (SEQ ID No:22). Нанесенные на диаграмму данные получены у 10 разных мышей (закрашенные треугольники). Приведены значения геометрических средних (прямая линия). См. примеры 12 и 13.
На фиг.15 приведены результаты CTL анализа меченных Cr51 клеток-мишеней HeLa-HHD. CD8+ T-клетки, полученные у одной иммунизированной мыши, тестировали в анализе CTL против клеток HeLa-HHD, экзогенно нагруженных пептидом hTERT865 или без пептида, но в присутствии ДМСО, используемого для разведения пептида. Клетки HeLa-HHD также инфицировали вектором Ad, кодирующим hTERT, или Ad, кодирующим GFP, в качестве контроля. См. пример 14.
На фиг.16 показана индукция клеточного иммунного ответа на hTERT у макак-резус, иммунизированных посредством ДНК-ЭП. Анализ ELIspot проводили на PBMC от иммунизированных hTERT макак-резус (pV1J/TPA-hTERT-LTB; ДНК-ЭП/ДНК-ЭП). Анализ проводили в двух повторах и приведено среднее значение из этих повторов. Показан анализ, проведенный после второй ДНК-ЭП (панель A). Также показан анализ, проведенный после пятой ДНК-ЭП (панель B). См. пример 15.
На фиг.17 показано, что у макак-резус, которых вакцинировали ДНК-ЭП и повторно стимулировали аденовирусом, экспрессирующим hTERT, по сравнению с CMI, вызываемым у макак, вакцинированных только ДНК-ЭП, усилен клеточный иммунный ответ на hTERT. ELIspot проводили на PBMC от иммунизированных hTERT макак-резус в конце программы иммунизации, которая включала пять инъекций ДНК-ЭП (pV1J/TPA-hTERT-LTB) с последующими двумя инъекциями Ad (Ad6-hTERT). Анализ проводили в двух повторах и показаны средние значения из этих повторов. См. пример 15.
Подробное описание изобретения
Экспрессию каталитического компонента теломеразы, известной как теломеразная обратная транскриптаза (TERT), как правило, можно обнаружить в широком диапазоне типов опухолей. Сочетание сверхэкспрессии теломеразы в большинстве типов опухолей, а также низкой или отсутствующей экспрессии в нормальных клетках, делает TERT мишенью для терапии и/или профилактики заболеваний, ассоциированных с нарушенной клеточной пролиферацией, такой как злокачественная опухоль. В связи с этим настоящее изобретение относится к композициям и способам индукции или усиления иммунного ответа к белковым продуктам, экспрессируемым в виде опухолеспецифического антигена TERT, где нарушенная экспрессия TERT связана с карциномой или ее развитием. Ассоциация нарушенной экспрессии TERT с карциномой не требует экспрессии белка TERT в опухолевой ткани в каждый момент времени ее развития, так как аномальная экспрессия TERT может иметь место во время инициации опухоли и не выявляться позже при прогрессировании опухоли и наоборот.
Таким образом, настоящее изобретение относится к полинуклеотидам, векторам, клеткам-хозяевам и кодируемым белкам, содержащим последовательность TERT или ее вариант, для применения в вакцинах и фармацевтических композициях для лечения и/или профилактики злокачественной опухоли. Полинуклеотиды по настоящему изобретению содержат нуклеотидную последовательность, кодирующую белок TERT или его вариант, слитую с нуклеотидной последовательностью, кодирующей субъединицу B термолабильного энтеротоксина (LTB) E. coli, которая способна эффективно оказывать адъювантное действие на иммунный ответ на ассоциированный антиген TERT.
В некоторых вариантах осуществления изобретения нуклеотидные последовательности, описываемые в настоящем документе, перед кодирующей последовательностью гена TERT дополнительно содержат лидерную последовательность TPA для гарантии секреции слитого белка TERT-LTB. В предпочтительных вариантах осуществления этого аспекта изобретения слитая последовательность TPA-hTERT(AI)-LTB кодирует аминокислотную последовательность, приведенную в SEQ ID NO:2. Предпочтительная нуклеотидная последовательность приведена в SEQ ID NO:1. В дополнительных предпочтительных вариантах осуществления слитая последовательность TPA-mTERT(AI)-LTB кодирует аминокислотную последовательность, приведенную в SEQ ID NO:4. Предпочтительная нуклеотидная последовательность приведена в SEQ ID NO:3.
Нуклеотидные последовательности TERT по настоящему изобретению могут быть человеческого происхождения или могут представлять собой гомолог TERT других видов, например мыши. Нуклеотидная последовательность TERT дикого типа человека приведена в SEQ ID NO:12 и была опубликована ранее (см., например, патент США №6166178; патент США 6261836; патент США №6927285; патентная заявка США 2003-0096344; Meyerson et al, Cell 90:785-95 (1997); Nakamura et al, Science 277:955-59 (1997)). Часть TERT слитой последовательности TERT может быть полноразмерной или представлять собой любой вариант, достаточный для индукции специфичного для TERT T-клеточного иммунного ответа у млекопитающего. Варианты TERT по настоящему изобретению включают, но не ограничиваются перечисленным, последовательности, которые укорочены на C- или N-конце, последовательности с консервативными заменами и последовательности с внутренними делециями или вставками. Предпочтительные варианты TERT по настоящему изобретению содержат мутации, которые функционально устраняют теломеразную каталитическую активность. Кодируемые слитые белки TERT по настоящему изобретению при введении реципиенту вакцины или нуждающемуся в этом пациенту способны индуцировать клеточный иммунный ответ.
Как указано выше, в предпочтительных вариантах осуществления настоящего изобретения теломеразная каталитическая активность антигена теломеразы инактивирована (в настоящем документе обозначено "TERT(AI)"), так что кодируемый слитый белок TERT для целей вакцинации является более безопасным, чем TERT дикого типа. Ферментативную активность слитого белка TERT можно инактивировать путем введения мутаций/делеций в кодирующую нуклеотидную последовательность TERT. Таким образом, настоящее изобретение относится к молекулам нуклеиновой кислоты, кодирующим слитый белок TERT-LTB, где нуклеотидная последовательность кодирует вариант белка hTERT, где указанный вариант содержит одну или несколько мутаций относительно аминокислотной последовательности hTERT дикого типа, приведенной в SEQ ID No:12, и где указанные мутации приводят к устранению теломеразной каталитической активности кодируемого белка hTERT. В конкретных вариантах осуществления изобретения, представленных в качестве примеров, нуклеотиды мутированы с изменением конкретных аминокислот D712A и V713I в последовательности белка TERT человека (как приведено в SEQ ID NO:10) и D702A и V703I в последовательности белка TERT мыши. Нуклеотидная последовательность, кодирующая мутантную последовательность TERT человека, приведенную в SEQ ID NO:10, приведена в SEQ ID NO:9.
В предпочтительных вариантах осуществления настоящего изобретения часть TERT слитой последовательности TERT-LTB представляет собой TERT человека или ее вариант, такой как hTERT(AI). В других предпочтительных вариантах осуществления часть TERT представляет собой TERT мыши или ее вариант, такой как mTERT(AI). Варианты TERT и варианты кодирующей нуклеотидной последовательности TERT по настоящему изобретению содержат мутации, устраняющие теломеразную ферментативную активность.
Таким образом, настоящее изобретение относится к синтетическим полинуклеотидам, содержащим нуклеотидную последовательность, кодирующим слитый белок TERT-LTB, где указанный слитый белок содержит белок TERT, или биологически неактивный фрагмент, или мутантную форму белка, слитые с белком LTB или его вариантом, который может эффективно усиливать иммунный ответ на белок TERT. Указанные мутантные формы белка TERT включают, но не ограничиваются перечисленным: консервативные аминокислотные замены, укорочения на N-конце, укорочения на C-конце, делеции или добавления. Любой такой вариант, фрагмент и/или мутант TERT кодируют белок или фрагмент белка, который по меньшей мере по существу имитирует иммунологические свойства белка TERT, приведенного в SEQ ID NO:12. Предпочтительные варианты TERT являются каталитически неактивными, например вариант TERT, приведенный в SEQ ID NO:10.
Синтетические полинуклеотиды по настоящему изобретению кодируют молекулы мРНК, которые экспрессируют слитый белок TERT(AI)-LTB, способный стимулировать или усиливать иммунный ответ на ассоциированный белок TERT так, чтобы быть подходящим для получения терапевтической или профилактической противораковой вакцины. Показано, что часть LTB слитых последовательностей TERT-LTB по настоящему изобретению резко усиливает иммуногенность совместно вводимых антигенов (см., например, Simmons et al. Scand. J. Immunol. 53:218-26 (2001)).
Также для использования по настоящему изобретению предусмотрены нуклеотидные последовательности, кодирующие варианты или мутанты LTB, включая, но не ограничиваясь перечисленным: нуклеотидные замены, делеции, добавления, укорочения на аминоконце и укорочения на карбоксиконце. В некоторых случаях для уменьшения или устранения токсичности кодируемого белка может быть выгодным введение в кодирующую LTB нуклеотидную последовательность конкретных точечных мутаций. В предпочтительных вариантах осуществления настоящего изобретения последовательность LTB, слитая с последовательностью TERT, укорочена на ее сигнальную последовательность.
Часть LTB или ее вариант слитых последовательностей TERT-LTB по настоящему изобретению можно сливать с аминоконцом или карбоксиконцом последовательности TERT. Кроме того, последовательность LTB и последовательность TERT можно сливать N-концом с N-концом, C-концом с C-концом, C-концом с N-концом или N-концом с N-концом. В предпочтительных вариантах осуществления настоящего изобретения C-конец полипептида TERT слит с N-концом LTB.
Настоящее изобретение относится к синтетической молекуле нуклеиновой кислоты (полинуклеотиду), содержащей нуклеотидную последовательность, кодирующую мРНК, экспрессирующую новый слитый белок TERT-LTB; например, нуклеотидные последовательности, кодирующие слитые белки, приведенные в SEQ ID NO:6 и 8. Особенно предпочтительная слитая последовательность TERT по настоящему изобретению представляет собой слитую последовательность hTERT(AI)-LTB, приведенную в SEQ ID NO:5. Молекулы нуклеиновой кислоты по настоящему изобретению по существу не содержат других нуклеиновых кислот.
Специалисту в данной области понятно, что для целей вакцинирования подходящими являются другие нуклеотидные последовательности, такие как нуклеотидные последовательности, которые кодируют белковую последовательность hTERT(AI)-LTB, приведенную в SEQ ID NO:6. Настоящее изобретение также относится к нуклеотидным последовательностям, которые кодируют неактивный слитый белок hTERT-LTB, который по существу сходен с SEQ ID NO:6, но не в точности одинаков, особенно вследствие аминокислотных различий в месте соединения последовательности TERT(AI) и последовательности LTB, которые были получены соответственно стратегии клонирования.
Настоящее изобретение также относится к рекомбинантным векторам и рекомбинантным клеткам-хозяевам, как прокариотическим, так и эукариотическим, которые содержат молекулы нуклеиновой кислоты, раскрытые в данном описании. Синтетические молекулы ДНК, ассоциированные векторы и хозяева по настоящему изобретению подходят для получения противораковой вакцины.
Молекулы нуклеиновой кислоты по настоящему изобретению, приведенные в качестве примеров, содержат нуклеотидную последовательность, выбранную из группы, состоящей из: SEQ ID NO:5 и 7, как показано на фигурах 1-2, которая кодирует слитые белки hTERT-LTB и mTERT-LTB по настоящему изобретению, приведенные в качестве примеров.
Настоящее изобретение также относится к биологически активным фрагментам или мутантам SEQ ID NO:5 и 7, которые кодируют мРНК, экспрессирующую слитые белки TERT-LTB, приведенные в качестве примеров. Любой такой биологически активный фрагмент и/или мутант кодирует белок или фрагмент белка, который по меньшей мере по существу имитирует иммунологические свойства белка hTERT, включая, в качестве неограничивающего примера, белок hTERT, как приведено в SEQ ID NO:12. Такие полинуклеотиды включают, но не ограничиваются перечисленным: нуклеотидные замены, делеции, добавления, укорочения на аминоконце и укорочения на карбоксиконце. Предпочтительные мутантные нуклеотидные последовательности по настоящему изобретению кодируют молекулы мРНК, которые экспрессируют в эукариотической клетке ферментативно неактивный слитый белок TERT-LTB, который способен вызывать профилактический или терапевтический иммунный ответ у нуждающегося в этом пациента так, чтобы быть подходящим для разработки противораковой вакцины.
Также в объем настоящего изобретения входят мутации в последовательности ДНК, которые по существу не изменяют конечные физические свойства экспрессируемого белка. Например, замена валина лейцином, аргинина лизином или аспарагина глутамином может не вызывать изменения желаемой функциональности полипептида, такой как способность вызывать иммунный ответ.
Как указано выше, настоящее изобретение, кроме того, относится к рекомбинантным векторам, которые содержат молекулы нуклеиновой кислоты, раскрытые в данном описании. Эти векторы могут состоять из ДНК или РНК. Для большинства задач клонирования предпочтительными являются ДНК-содержащие векторы. Типичные векторы включают плазмиды, модифицированные вирусы, бакуловирусы, бактериофаги, космиды, искусственные дрожжевые хромосомы и другие формы эписомальной или интегрированной ДНК, которая может кодировать слитый белок TERT-LTB. Определение подходящего вектора для переноса конкретного гена или другого применения находится в пределах компетенции специалистов в данной области.
Также настоящим изобретением предусмотрены очищенные слитые белки TERT-LTB, кодируемые нуклеиновыми кислотами, раскрытыми в данном описании, в частности слитые белки TERT(AI)-LTB. В вариантах осуществления данного аспекта изобретения, представленных в качестве примеров, слитый белок TERT-LTB содержит аминокислотную последовательность, выбранную из группы, состоящей из: SEQ ID NO:6 и 8.
В настоящее изобретение включены последовательности ДНК, которые гибридизуются с комплементарными SEQ ID NO:5 и 7 в строгих условиях. В качестве неограничивающего примера способ с использованием условий высокой строгости представляет собой следующее. Предгибридизацию фильтров, содержащих ДНК, проводят в течение периода приблизительно от 2 часов до периода в течение ночи приблизительно при 65°C в буфере, состоящем из 6× SSC, 5× раствора Денхардта и 100 мкг/мл денатурированной ДНК из спермы лосося. Фильтры гибридизуют в течение приблизительно 12-48 часов при 65°C в смеси для предгибридизации, содержащей 100 мкг/мл денатурированной ДНК из спермы лосося и 5-20×106 имп./мин меченного 32P зонда. Отмывку фильтров проводят при 37°C в течение приблизительно 1 часа в растворе, содержащем 2× SSC, 0,1% SDS. После этого перед авторадиографией проводят отмывку в 0,1× SSC, 0,1% SDS при 50°C в течение 45 минут. Другие способы с использованием условий высокой строгости могут включать стадию гибридизации, проводимую в 5× SSC, 5× растворе Денхардта, 50% формамиде приблизительно при 42°C в течение приблизительно 12-48 часов, или стадию отмывки, проводимую в 0,2× SSPE, 0,2% SDS приблизительно при 65°C в течение приблизительно 30-60 минут. Указанные в приведенных выше способах реагенты для проведения гибридизации в строгих условиях хорошо известны в данной области. Подробности состава этих реагентов можно найти в Sambrook et al., Molecular Cloning: A Laboratory Manual 2nd Edition; Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, (1989) или Sambrook and Russell, Molecular Cloning: A Laboratory Manual, 3rd Edition. Cold Spring Harbor Laboratory Press, Plainview, NY (2001). В дополнение к указанному, в данной области также хорошо известны другие условия с высокой строгостью, которые могут быть использованы.
Экспрессирующий вектор, содержащий кодирующую слитый белок TERT-LTB молекулу нуклеиновой кислоты, можно использовать для экспрессии слитого белка TERT-LTB в рекомбинантной клетке-хозяине на высоком уровне. Экспрессирующие векторы могут включать, но не ограничиваясь перечисленным, клонирующие векторы, модифицированные клонирующие векторы, особым образом сконструированные плазмиды или вирусы. Также, при желании, можно использовать множество бактериальных экспрессирующих векторов для экспрессии рекомбинантных слитых последовательностей TERT-LTB в бактериальных клетках. Кроме того, можно использовать множество экспрессирующих векторов для клеток грибов для экспрессии рекомбинантных слитых последовательностей TERT-LTB в клетках грибов. Кроме того, можно использовать множество экспрессирующих векторов для клеток насекомых для экспрессии рекомбинантного белка в клетках насекомых.
Настоящее изобретение также относится к клеткам-хозяевам, трансформированным или трансфицированным векторами, содержащими молекулы нуклеиновой кислоты по настоящему изобретению. Рекомбинантные клетки-хозяева могут быть прокариотическими или эукариотическими, включая, но не ограничиваясь перечисленным, бактерии, такие как E. coli, клетки грибов, таких как дрожжи, клетки млекопитающих, включая, но не ограничиваясь перечисленным, клеточные линии, происходящие от коров, свиней, обезьян и грызунов; и клетки насекомых, включая, но не ограничиваясь перечисленным, клеточные линии, полученные от Drosophila и тутового шелкопряда. Такие рекомбинантные клетки-хозяева можно культивировать в подходящих условиях с получением слитого белка TERT-LTB или биологически эквивалентной формы. В предпочтительном варианте осуществления настоящего изобретения клетка-хозяин является человеческой. Как определено в настоящем описании, термин "клетка-хозяин" не предназначена для включения клетки-хозяина в организме трансгенного человека, плода человека или эмбриона человека.
Как указано выше, экспрессирующий вектор, содержащий ДНК, кодирующую слитый белок TERT-LTB, можно использовать для экспрессии слитого белка TERT в рекомбинантной клетке-хозяине. Таким образом, другой аспект изобретения относится к способу экспрессии слитого белка TERT-LTB в рекомбинантной клетке-хозяине, включающему: (a) введение вектора, содержащего нуклеиновую кислоту, содержащую нуклеотидную последовательность, кодирующую слитый белок TERT-LTB, в подходящую клетку-хозяина человека, где слитый белок TERT-LTB содержит белок TERT или его неактивный вариант, слитый со значительной частью белка LTB, и где слитый белок способен индуцировать иммунный ответ у млекопитающего; и (b) культивирование клетки-хозяина в условиях, позволяющих экспрессию указанного слитого белка TERT-LTB.
В предпочтительных вариантах осуществления способа экспрессии слитого белка TERT-LTB, описанного выше, в последовательности LTB удалена ее сигнальная последовательность.
После экспрессии слитой последовательности TERT-LTB в клетке-хозяине слитый белок TERT-LTB может быть восстановлен с получением очищенного слитого белка TERT-LTB. Доступными и подходящими для использования являются несколько способов очистки белка. Рекомбинантный белок может быть очищен из клеточных лизатов и экстрактов различным сочетанием или индивидуальным применением высаливания, ионообменной хроматографии, гель-хроматографии, адсорбционной хроматографии на гидроксилапатите и хроматографии гидрофобного взаимодействия. Кроме того, рекомбинантный слитый белок TERT-LTB может быть выделен из других клеточных белков с использованием иммуноаффинной колонки, полученной с моноклональными или поликлональными антителами, специфичными для белка TERT или полипептидных фрагментов белка TERT.
Молекулы нуклеиновой кислоты, содержащие слитые последовательности TERT-LTB, и кодируемые слитые белки по настоящему изобретению сконструированы для усиления специфичного для TERT иммунного ответа по сравнению с полноразмерной кДНК дикого типа, кодирующей TERT, для применения в разработке вакцины. Для дополнительного усиления иммуногенных свойств слитых последовательностей TERT-LTB по настоящему изобретению в некоторых вариантах осуществления, описываемых в настоящем документе, полинуклеотиды, кодирующие слитые белки TERT-LTB, содержат оптимизированные кодоны для последующей высокоэффективной экспрессии в клетке-хозяине, как описано ниже. В этих вариантах осуществления по меньшей мере часть кодонов слитых последовательностей TERT-LTB сконструирована так, чтобы использование кодонов было предпочтительным у проектируемой клетки-хозяина, которая в предпочтительных вариантах осуществления является клеткой человека. Оптимизированные слитые последовательности TERT-LTB можно использовать для конструирования основанных на ДНК рекомбинантных аденовирусов или плазмид вакцин, которые обеспечивают эффективную иммунопрофилактику ассоциированных с TERT злокачественных опухолей посредством клеточного иммунитета. В качестве иммуногенной композиции можно использовать синтетические молекулы. Настоящее изобретение относится к слитым полинуклеотидам TERT-LTB с оптимизированными кодонами, которые при непосредственном введении in vivo позвоночным, включая млекопитающих, таких как приматы и люди, индуцируют экспрессию кодируемых белков у животного.
Как указано выше, в некоторых вариантах осуществления настоящего изобретения, синтетические молекулы содержат нуклеотидную последовательность, где некоторые из нуклеотидов изменены так, чтобы использовать кодоны, предпочтительные для клетки человека, таким образом, обеспечивая высокоэффективную экспрессию слитого белка в клетке-хозяине человека. Синтетические молекулы можно использовать в качестве источника слитого белка TERT-LTB, который может быть использован в противораковой вакцине с обеспечением эффективной иммунопрофилактики против ассоциированных с TERT карцином посредством клеточного иммунитета. Молекулы нуклеиновой кислоты, раскрытые в настоящем описании, могут также служить в качестве основы для противораковой вакцины на основе ДНК.
"Триплетный" кодон из четырех возможных нуклеотидных оснований может существовать более чем в 60 вариантных формах. Поскольку эти кодоны обеспечивают информацию только для 20 различных аминокислот (а также для инициации и терминации транскрипции), некоторые аминокислоты могут быть закодированы более чем одним кодоном, феномен, известный как вырожденность. По причинам, не вполне понятным, альтернативные кодоны не одинаково представлены в эндогенной ДНК различных типов клеток. Действительно, для определенных кодонов в определенных типах клеток, по-видимому, существует вариабельная естественная иерархия или "предпочтение". В качестве одного из примеров, аминокислота лейцин кодируется любым из шести кодонов ДНК, включающих CTA, CTC, CTG, CTT, TTA и TTG. Полный анализ частоты кодонов в геноме для микроорганизмов выявил, что эндогенная ДНК E. coli обычно содержит кодирующий лейцин кодон CTG, тогда как ДНК дрожжей и миксомицет обычно включает кодирующий лейцин кодон TTA. Ввиду этой иерархии, как правило, полагают, что вероятность получения высоких уровней экспрессии богатого лейцином полипептида в хозяине E. coli в некоторой степени зависит от частоты использования кодона. Например, вероятно, что ген, богатый кодонами TTA, будет слабо экспрессироваться в E. coli, тогда как богатый CTG ген, вероятно, будет высоко экспрессирован в этом хозяине. Аналогично, предпочтительным кодоном для экспрессии богатого лейцином полипептида в дрожжевых клетках-хозяевах может быть TTA.
Значение феномена предпочтения кодонов для способов рекомбинантной ДНК очевидны, и этот феномен может служить объяснением множества предыдущих неудач в достижении высоких уровней экспрессии экзогенных генов в успешно трансформированных организмах-хозяевах - во вставленном гене часто может встречаться менее "предпочтительный" кодон, и механизм экспрессии клетки-хозяина может действовать неэффективно. Этот феномен позволяет предположить, что синтетические гены, которые сконструированы так, чтобы включать предпочтительные кодоны для проектируемой клетки-хозяина, обеспечивают оптимальную форму чужеродного генетического материала для практического осуществления способов рекомбинантной ДНК. Таким образом, один из аспектов данного изобретения относится к слитому гену TERT-LTB, у которого кодоны оптимизированы для экспрессии в клетке человека. В предпочтительном варианте осуществления данного изобретения выявлено, что использование альтернативных кодонов, кодирующих ту же самую белковую последовательность, может устранять сложности в экспрессии экзогенного слитого белка TERT-LTB в клетках человека.
В соответствии с определенными вариантами осуществления настоящего изобретения молекулы нуклеиновой кислоты, которые кодируют слитые белки TERT-LTB, конвертируют в полинуклеотидную последовательность с идентичной транслированной последовательностью, но с альтернативным использованием кодонов, как описано в публикации Lathe, "Synthetic Oligonucleotide Probes Deduced from Amino Acid Sequence Data: Theoretical and Practical Considerations" J. Molec. Biol. 183:1-12 (1985), таким образом, включенной в качестве ссылки. Как правило, способ включает идентификацию в последовательности дикого типа кодонов, которые часто не ассоциированы с высокоэкспрессированными генами человека, и замену их оптимальными кодонами для высокой экспрессии в клетках человека. Затем новую генную последовательность проверяют на нежелательные последовательности, полученные вследствие этой замены кодонов (например, последовательности "ATTTA", непреднамеренное получение сайтов распознавания сплайсинга интронов, нежелательные сайты распознавания рестрикционных ферментов и т.д.). Нежелательные последовательности устраняют путем замены существующих кодонов другими кодонами, кодирующими ту же аминокислоту. Затем синтетические генные сегменты тестируют на улучшенную экспрессию.
Следует понимать, что этот способ необязательно приводит к полинуклеотидной последовательности, в которой все кодоны являются оптимальными кодонами в соответствии с использованием кодонов в высокоэкспрессированных генах человека и/или млекопитающих. Однако предпочтительно, чтобы в вариантах осуществления изобретения, где предусмотрены варианты полинуклеотидов TERT и/или LTB с оптимизированными кодонами, значительная часть полученных в результате кодонов совпадала с использованием кодонов в высокоэкспрессированных генах человека и/или млекопитающих.
Описанные выше способы использовали для получения синтетических генных последовательностей, которые кодируют слитые белки TERT-LTB, получая в результате ген, содержащий кодоны, оптимизированные для высокоэффективной экспрессии в клетках человека. Хотя приведенный выше способ предоставляет обзор репрезентативных способов конструирования генов с оптимизированными кодонами для применения в противораковых вакцинах, специалисту в данной области понятно, что при незначительных изменениях способа или при незначительных изменениях последовательности можно достичь сходной эффективности вакцины или увеличенной экспрессии генов.
Специалисту в данной области также понятно, что могут быть сконструированы дополнительные молекулы нуклеиновой кислоты, которые обеспечат высокие уровни экспрессии слитой последовательности TERT-LTB в клетках человека, где только часть кодонов молекулы ДНК являются оптимизированными. Например, в некоторых вариантах осуществления настоящего изобретения кодоны, содержащиеся в части TERT слитой последовательности TERT-LTB, оптимизированы для высокоэффективной экспрессии в клетках человека, а кодоны, содержащиеся в части LTB слитой последовательности TERT-LTB, по существу сходны с LTB дикого типа. В других вариантах осуществления настоящего изобретения кодоны, содержащиеся в части LTB слитой последовательности TERT-LTB, оптимизированы для высокоэффективной экспрессии в клетках человека, а кодоны, содержащиеся в части TERT слитой последовательности TERT-LTB, по существу сходны с геном TERT дикого типа. В других вариантах осуществления настоящего изобретения как часть TERT, так и часть LTB слитой последовательности TERT-LTB состоят из оптимизированных для высокоэффективной экспрессии в клетках человека кодонов, например, последовательность hTERT-LTBopt, приведенная в SEQ ID NO:5. В настоящем изобретении предусмотрены слитые последовательности TERT-LTB, в которых оптимизирована только часть кодонов в части TERT и/или LTB слитой последовательности TERT-LTB.
Нуклеиновые кислоты по настоящему изобретению можно собрать в экспрессирующую кассету, которая содержит последовательности, сконструированные для обеспечения эффективной экспрессии белка в клетке человека. Кассета предпочтительно содержит ген, кодирующий слитый белок TERT-LTB, с соответствующими последовательностями транскрипционного и трансляционного контроля, функционально связанными с ним, такими как последовательности промотора и терминатора. В предпочтительном варианте осуществления промотор представляет собой последовательность промотора цитомегаловируса с интроном A (CMV), хотя специалистам в данной области понятно, что можно использовать любой ряд других известных промоторов, таких как сильный промотор гена иммуноглобулина или другого эукариотического гена. Предпочтительный терминатор транскрипции представляет собой терминатор бычьего гормона роста, хотя также можно использовать другие терминаторы транскрипции. Особенно предпочтительно сочетание CMV-терминатор BGH.
По настоящему изобретению слитую экспрессирующую последовательность TERT-LTB кассету вставляют в вектор. Вектор, предпочтительно, представляет собой аденовирусный или плазмидный вектор, хотя также можно использовать линейную ДНК, связанную с промотором, или другие векторы, такие как аденоассоциированный вирус или вектор из модифицированного вируса коровьей оспы, ретровирусный или лентивирусный вектор.
В предпочтительном варианте осуществления изобретения вектор представляет собой аденовирусный вектор (в настоящем описании взаимозаменяемо используемый вместе с “аденовектором”). Основой аденовекторов могут быть различные серотипы аденовирусов, такие как серотипы, выявляемые у людей или животных. Примеры аденовирусов животных включают аденовирусы коров, свиней, шимпанзе, мышей, собак и птиц (CELO). Предпочтительные аденовекторы представляют собой аденовекторы на основе серотипов человека, более предпочтительно серотипов группы B, C или D. Примеры серотипов аденовирусов человека группы B, C, D или E включают типы 2 ("Ad2"), 4 ("Ad4"), 5 ("Ad5"), 6 ("Ad6"), 24 ("Ad24"), 26 ("Ad26"), 34 ("Ad34") и 35 ("Ad35"). В особенно предпочтительных вариантах осуществления настоящего изобретения экспрессирующий вектор представляет собой вектор аденовируса типа 6 (Ad6).
Если выбранный вектор представляет собой аденовирус, предпочтительно, чтобы вектор представлял собой так называемый аденовирусный вектор первого поколения. Эти аденовирусные векторы характеризуются наличием нефункциональной генной области E1 и, предпочтительно, удаленной аденовирусной генной областью E1. Кроме того, в первом поколении векторов может находиться нефункциональная генная область E3 или она может быть удалена (Danthinne et al. Gene Therapy 7:1707-1714 (2000); F.L. Graham, Immunology Today 21(9): 426-428 (2000)). Нет необходимости полностью удалять области E1 и E3 в аденовекторах. Предпочтительнее удалять достаточную часть области E1, чтобы сделать вектор некомпетентным по репликации в отсутствие белков E1, замещаемых в трансположении; и делеция E1 или сочетание делеций E1 и E3 являются по существу достаточно большими, чтобы встроить экспрессирующую гены кассету.
В некоторых вариантах осуществления экспрессирующая кассета встроена в положение, где обычно расположена аденовирусная генная область E1. Кроме того, такие векторы необязательно содержат нефункциональную область E3 или она удалена. Предпочтительно, в геноме используемого аденовируса удалены обе области E1 и E3 (ΔE1ΔE3). Количество аденовирусов можно увеличить в известных клеточных линиях, которые экспрессируют вирусные гены E1, таких как клетки 293 или клетки PERC.6, или в клеточных линиях, полученных из клеток 293 или PERC.6, которые транзиторно или стабильно трансформированы и экспрессируют дополнительный белок. В качестве примера, при использовании конструкций с контролируемой экспрессией генов, таких как регулируемая тетрациклином промоторная система, клеточная линия может экспрессировать компоненты, вовлеченные в систему регуляции. Один из примеров такой клеточной линии представляет собой T-Rex-293; другие известны в данной области.
Для удобства использования аденовирусного вектора аденовирус может находиться в форме челночной плазмиды. Настоящее изобретение также относится к челночному плазмидному вектору, который содержит плазмидную часть и аденовирусную часть, где аденовирусная часть содержит аденовирусный геном с делецией E1 и необязательной делецией E3, и содержит встроенную экспрессирующую кассету, содержащую кодирующую слитый белок TERT-LTB нуклеотидную последовательность. В предпочтительных вариантах осуществления имеется сайт рестрикции, фланкирующий аденовирусную часть плазмиды так, что аденовирусный вектор может быть легко удален. Челночная плазмида может быть реплицирована в прокариотических клетках или эукариотических клетках.
В предпочтительном варианте осуществления изобретения экспрессирующая кассета встроена в аденовирусную плазмиду Ad6 (ΔE1ΔE3) (см. Emini et al, US20040247615, таким образом, включенный в качестве ссылки). Этот вектор содержит геном аденовируса Ad6 с удаленными областями E1 и E3. В других предпочтительных вариантах осуществления изобретения экспрессирующая кассета встроена в аденовирусную плазмиду pMRKAd5-HV0 (см. Emini et al, US20030044421, таким образом, включенный в качестве ссылки). Эта плазмида содержит геном аденовируса Ad5 с удаленными областями E1 и E3. Конструкция плазмиды pMRKAd5-HV0 улучшена по сравнению с предыдущими аденовекторами путем расширения 5' действующей в цис-положении пакующей области дополнительно в генную область E1 со встраиванием элементов, для которых выявлено, что они важны для оптимизации упаковки вируса, с получением в результате усиленной амплификации вируса. Преимущественно, эти улучшенные аденовирусные векторы способны поддерживать генетическую стабильность после размножения при многократном пассировании.
Аденовирусы, челночные плазмиды и иммуногены ДНК по настоящему изобретению могут быть получены стандартными способами получения и очистки конструкций ДНК, известными в молекулярной биологии.
В соответствии с настоящим изобретением было определено, что генетическая вакцинация плазмидной ДНК, кодирующей TPA-mTERT(AI)-LTBopt, может нарушать иммунологическую толерантность мышей BALB/c и B6 (см. пример 7). Также в настоящем описании показано, что иммунизация плазмидой TPA-mTERT(AI)-LTBopt может вызывать цитотоксический иммунный ответ у мышей BALB/c (см. пример 8). Кроме того, было продемонстрировано, что эта конструкция способна индуцировать более сильный CD8+ иммунный ответ, чем ответ, вызванный только mTERT(AI) (см. пример 9), и способна контролировать рост опухоли (пример 11). Таким образом, описываемые в настоящем документе данные демонстрируют, что слияние кодирующей последовательности TERT с кДНК LTB приводит к усилению специфичного для TERT иммунного ответа.
Также в настоящем изобретении показано, что hTERT(AI)-LTBopt может индуцировать рестриктированный по HLA-A2 CD8+ иммунный ответ (см. пример 12) и что этот иммунный ответ можно усилить путем повторной иммунизации с использованием Ad6-hTERT(AI).
Таким образом, описанные выше векторы можно использовать в иммуногенных композициях и вакцинах для профилактики развития опухолей, ассоциированных с нарушенной экспрессией TERT, и/или для лечения существующих злокачественных опухолей. Векторы по настоящему изобретению позволяют разрабатывать и коммерчески применять вакцины, устраняя сложности получения высоких уровней экспрессии экзогенного TERT в успешно трансформированных организмах-хозяевах и обеспечивая получение слитого белка TERT-LTB, который при введении млекопитающему, такому как человек, может вызывать усиленный иммунный ответ.
В связи с этим один из аспектов настоящего изобретения относится к способу профилактики или лечения ассоциированной с TERT злокачественной опухоли, включающему введение млекопитающему вакцинного вектора, содержащего полинуклеотид, содержащий нуклеотидную последовательность, кодирующую слитый белок TERT-LTB, содержащий неактивный белок TERT или его вариант, слитый со значительной частью LTB, и где слитый белок способен вызывать у млекопитающего иммунный ответ.
В соответствии с описанным выше способом вакцинный вектор можно вводить для лечения или профилактики злокачественной опухоли у любого млекопитающего, в том числе, в качестве неограничивающих примеров, рака легких, рака молочной железы и колоректального рака. В предпочтительном варианте осуществления изобретения млекопитающим является человек.
Кроме того, специалист в данной области может выбрать любой тип вектора для использования в описанном способе лечения и профилактики. Предпочтительно, вектор представляет собой аденовирусный вектор или плазмидный вектор. В предпочтительном варианте осуществления изобретения вектор представляет собой аденовирусный вектор, содержащий аденовирусный геном с делецией в области E1 аденовируса и вставкой в области E1 аденовируса, где вставка содержит экспрессирующую кассету, содержащую: (a) нуклеотидную последовательность, кодирующую слитый белок TERT-LTB, где слитый белок TERT-LTB содержит неактивный белок TERT или его вариант, слитый со значительной частью LTB, и где слитый белок способен вызывать иммунный ответ у млекопитающего; и (b) промотор, функционально связанный с полинуклеотидом.
Настоящее изобретение также относится к аденовирусному вакцинному вектору, содержащему аденовирусный геном с делецией в области E1 и вставкой в области E1, где вставка содержит экспрессирующую кассету, содержащую: (a) нуклеотидную последовательность, кодирующую слитый белок TERT, где слитый белок TERT содержит неактивный белок TERT или его вариант, слитый со значительной частью LTB, и где слитый белок способен вызывать иммунный ответ у млекопитающего; и (b) промотор, функционально связанный с полинуклеотидом.
В предпочтительном варианте осуществления этого аспекта изобретения аденовирусный вектор представляет собой вектор Ad6.
В другом предпочтительном варианте осуществления изобретения аденовирусный вектор представляет собой вектор Ad5.
В другом предпочтительном варианте осуществления аденовирусный вектор представляет собой вектор Ad24.
Также для использования по настоящему изобретению предусмотрен аденовирусный вакцинный вектор, содержащий геном аденовируса, который в природе инфицирует виды, отличные от человека, включая, в качестве неограничивающего примера, аденовирусные векторы шимпанзе. Предпочтительный вариант осуществления этого аспекта изобретения представляет собой вакцинный вектор шимпанзе Ad3.
В другом аспекте изобретение относится к вакцинной плазмиде, содержащей плазмидную часть и часть экспрессирующей кассеты, где часть экспрессирующей кассеты содержит: (a) нуклеотидную последовательность, кодирующую слитый белок TERT, где слитый белок TERT содержит неактивный белок TERT или его вариант, слитый со значительной частью LTB, и где слитый белок способен вызывать иммунный ответ у млекопитающего; и (b) промотор, функционально связанный с полинуклеотидом. Примером подходящей плазмиды могла бы быть экспрессирующая плазмида млекопитающих V1Jns, как описано (публикация J. Shiver et. al. in DNA Vaccines, M. Liu et al. eds., N.Y. Acad. Sci., N.Y., 772:198-208 (1996), которая включена в описание в качестве ссылки).
В некоторых вариантах осуществления настоящего изобретения основанные на аденовирусах и плазмидах рекомбинантные полинуклеотидные вакцины, раскрытые в настоящем описании, используют в различных сочетаниях примирования/повторного стимулирующего введения для индукции усиленного иммунного ответа. В данном случае два вектора вводят по схеме "примирование и повторная стимуляция". Например, один или несколько раз вводят первый тип вектора, затем через предопределенный период времени, например 2 недели, 1 месяц, 2 месяца, шесть месяцев или другой подходящий период, один или несколько раз вводят второй тип вектора. Предпочтительно, векторы несут экспрессирующие кассеты, кодирующие те же полинуклеотиды или сочетание полинуклеотидов.
В варианте осуществления, где также используют плазмидную ДНК, предпочтительно, чтобы вектор содержал один или несколько промоторов, распознаваемых клетками млекопитающих или насекомых. В предпочтительном варианте осуществления плазмида содержит сильный промотор, такой как, в качестве неограничивающего примера, промотор CMV. Специалисту в данной области ясно, что с целью контроля экспрессии нуклеотидных последовательностей TERT-LTB по настоящему изобретению можно выбрать любой из ряда других известных промоторов. Дополнительные примеры промоторов включают природные промоторы, такие как промотор EF1 альфа, промотор вируса саркомы Рауса и ранние/поздние промоторы SV40 и промотор п-актина; и искусственные промоторы, такие как синтетический мышечный промотор и химерный мышечный/CMV промотор (Li et al, Nat. Biotechnol. 17:241-245 (1999); Hagstrom et al, Blood 95:2536-2542 (2000)). Синтетический слитый ген TERT-LTB или другой экспрессируемый ген может быть связан с таким промотором.
Как указано выше, аденовирусную векторную вакцину и плазмидную вакцину можно вводить позвоночному в качестве части единой схемы лечения для индукции иммунного ответа. В связи с этим настоящее изобретение относится к способу защиты млекопитающего от ассоциированной с TERT злокачественной опухоли, включающему: (a) введение млекопитающему первого вектора, содержащего: i) нуклеотидную последовательность, кодирующую слитый белок TERT, где слитый белок TERT содержит неактивный белок TERT или его вариант, слитый со значительной частью LTB, и где слитый белок способен вызывать иммунный ответ у млекопитающего; и ii) промотор, функционально связанный с полинуклеотидом; (b) ожидание прохождения предопределенного периода времени; и (c) введение млекопитающему второго вектора, содержащего: i) нуклеотидную последовательность, кодирующую слитый белок TERT, где слитый белок TERT содержит неактивный белок TERT или его вариант, слитый со значительной частью LTB, и где слитый белок способен вызывать иммунный ответ у млекопитающего; и ii) промотор, функционально связанный с полинуклеотидом.
В одном из вариантов осуществления описанного выше способа защиты первый вектор представляет собой плазмиду, а второй вектор представляет собой аденовирусный вектор. В альтернативном варианте осуществления первый вектор представляет собой аденовирусный вектор, а второй вектор представляет собой плазмиду. В некоторых вариантах осуществления настоящего изобретения первый вектор вводят пациенту более одного раза перед введением второго вектора.
В описанном выше способе первый тип вектора можно вводить более одного раза, разделяя каждое введение вектора предопределенным периодом времени. За такими сериями введений первого типа вектора после прохождения предопределенного периода времени может следовать одно или несколько введений второго типа вектора. Аналогично лечению первым типом вектора второй тип вектора также можно вводить один или несколько раз после предопределенных периодов времени.
Настоящее изобретение также относится к способу лечения пациента, страдающего от ассоциированной с TERT злокачественной опухоли, включающему: (a) введение млекопитающему первого вектора, содержащего: i) нуклеотидную последовательность, кодирующую слитый белок TERT, где слитый белок TERT содержит белок TERT(AI) или его вариант, слитый со значительной частью LTB, и где слитый белок способен вызывать иммунный ответ у млекопитающего; и ii) промотор, функционально связанный с полинуклеотидом; (b) ожидание прохождения предопределенного периода времени; и (c) введение пациенту второго вектора, содержащего: i) нуклеотидную последовательность, кодирующую слитый белок TERT, где слитый белок TERT содержит белок TERT(AI) или его вариант, слитый со значительной частью LTB, и где слитый белок способен вызывать иммунный ответ у млекопитающего; и ii) промотор, функционально связанный с полинуклеотидом.
В одном из вариантов осуществления описанного выше способа лечения первый вектор представляет собой плазмиду, а второй вектор представляет собой аденовирусный вектор. В альтернативном варианте осуществления первый вектор представляет собой аденовирусный вектор, а второй вектор представляет собой плазмиду. В других предпочтительных вариантах осуществления описанного выше способа первый вектор вводят пациенту более одного раза перед введением пациенту второго вектора.
В предпочтительных вариантах осуществления описанных выше способов векторы содержат нуклеотидную последовательность, кодирующую слитый белок TERT(AI)-LTB, где слитый белок TERT содержит неактивный белок TERT, слитый со значительной частью LTB.
Количество экспрессируемой ДНК или транскрибируемой РНК для введения реципиенту вакцины частично зависит от силы используемых промоторов и от иммуногенности экспрессируемого генного продукта. Как правило, иммунологически или профилактически эффективную дозу плазмидного вакцинного вектора приблизительно от 1 нг до 100 мг и, предпочтительно, приблизительно от 10 мкг до 300 мкг вводят непосредственно в мышечную ткань. Эффективная доза рекомбинантного аденовируса составляет приблизительно 106-1012 частиц и, предпочтительно, приблизительно 107-1011 частиц. Также предусмотрены подкожная инъекция, внутрикожное введение, проникновение через кожу и другие способы введения, такие как интраперитонеальная, внутривенная, внутримышечная или ингаляционная доставка.
В предпочтительных вариантах осуществления настоящего изобретения вакцинные векторы вводят реципиенту посредством внутримышечной инъекции.
Вакцинные векторы по изобретению могут быть чистыми, т.е. неассоциированными с какими-либо белками или другими средствами, которые воздействуют на иммунную систему реципиента. В этом случае желательно, чтобы вакцинные векторы находились в физиологически приемлемом растворе, таком как, без ограничения перечисленным, стерильный физиологический раствор или стерильный буферный физиологический раствор. Альтернативно, эффективным может быть введение средства, которое содействует клеточному захвату ДНК, такого как, без ограничения приведенным, ион кальция. Эти средства, как правило, называют способствующими трансфекции реагентами и фармацевтически приемлемыми носителями. Специалисты в данной области способны определить конкретный реагент или фармацевтически приемлемый носитель, а также подходящее время и способ введения.
Все указанные в настоящем описании публикации включены в качестве ссылки для описания и раскрытия методик и материалов, которые можно использовать в связи с настоящим изобретением. Ничто в настоящем документе не следует истолковывать как допущение того, что изобретению может быть противопоставлено описание более раннего изобретения.
Имея описание предпочтительных вариантов осуществления изобретения со ссылкой на сопровождающие рисунки, следует учитывать, что изобретение не ограничено этими конкретными вариантами осуществления и что специалист в данной области может осуществлять различные изменения и модификации без отклонения от объема или сущности изобретения, определенных приложенной формулой изобретения.
Следующие примеры иллюстрируют, но не ограничивают изобретение.
ПРИМЕР 1
Конструирование слитых белков TERT
Для определения того, может ли слияние антигена теломеразной обратной транскриптазы (TERT) с субъединицей LTB термолабильного энтеротоксина E. coli (Fingerut et al. Vaccine 23(38): 4685-96 (2005); Rigano et al. Plant Cell Rep. 22(7): 502-8 (2004)) усиливать иммуногенность только TERT, были сконструированы векторы, кодирующие полноразмерную теломеразную обратную транскриптазу с модификациями. Во-первых, кодоны последовательности ДНК были оптимизированы так, чтобы включать кодоны, предпочтительные для клеток-хозяев человека. Кроме того, для гарантии того, что кодируемый антиген безопасен для применения в вакцине, в нуклеотидную последовательность TERT были введены мутации для инактивации теломеразной каталитической активности кодируемого белка. В частности, в последовательность TERT человека были введены мутации D712A и V713I, а в последовательность TERT мыши были введены мутации D702A и V703I (Arai et al. Two independent regions of human telomerase reverse transcriptase are important for its oligomerization and telomerase activity. J. Biol. Chem. 277 (10): 8538-44 (2002)).
Слитые последовательности TERT конструировали путем связывания C-конца описанных выше модифицированных нуклеотидных последовательностей TERT с нуклеотидной последовательностью, кодирующей модифицированную LTB (нукл. от 64 до 375), в которой удалена последовательность, кодирующая сигнальный пептид. Кроме того, со стороны 5' от нуклеотидной последовательности, кодирующей TERT, для гарантии секреции белка была введена лидерная последовательность, кодирующая сигнальную последовательность активатора тканевого плазминогена (TPA) человека (Haddad et al. Comparative study of DNA-based immunization vectors: effect of secretion signals on the antibody responses in mice. FEMS Immunol. Med. Microbiol. 18(3): 193-202 (1997)). Кодоны всей последовательности были оптимизированы c использованием компьютерных алгоритмов так, чтобы включать кодоны, предпочтительные для клеток-хозяев человека. Синтетический ген с оптимизированными кодонами собирали с помощью синтетических олигонуклеотидов. В конечной конструкции между последовательностью TERT и последовательностью LTB вводили две аминокислоты (S и R) соответственно стратегии клонирования.
Нуклеотидные последовательности, кодирующие слитые молекулы TERT, клонировали в вектор pV1JnsA под контроль промотора цитомегаловируса человека (CMV)/интрон A с сигналом полиаденилирования бычьего гормона роста (BGH). Плазмида pV1J/TPA-hTERT-LTBopt несет кДНК инактивированного TERT человека с оптимизированными кодонами, слитую с кодирующими последовательностями сигнальной последовательности TPA на N-конце и с LTB на C-конце. Аналогично, плазмида pV1J/TPA-mTERT-LTBopt несет кДНК инактивированного TERT мыши с оптимизированными кодонами, слитую с TPA и LTB.
ПРИМЕР 2
Плазмидные и аденовирусные конструкции
pV1JnsA/TPA-mTERT(AI)-LTBopt: Плазмиду 041046pucKana, содержащую последовательность TPA-mTERT(AI)-LTB, получали из GENEART (Geneart GmbH, Regensburg, Germany). Плазмиду расщепляли BglII и SalI и полученный фрагмент клонировали в сайт BglII/SalI в плазмиде pV1JnsA (Montgomery et al, DNA Cell Biol, 12(9):777-83(1993)).
pV1JnsA/mTERT(AI)opt: pV1JnsA/TPA-mTERT(AI)-LTB расщепляли XbaI для удаления кодирующей последовательности LTB (находящейся между сайтами XbaI). Полученная конструкция pV1JnsA/TPA-mTERT(AI) после последней а.к. mTERT и перед стоп-кодоном несла 3 а.к. (S; R; N). Удаление кодирующей последовательности TPA проводили в pV1JnsA/TPA-mTERT(AI) путем расщепления с использованием BamHI и EcoRV. Кодирующую последовательность TPA заменяли продуктом ПЦР, полученным амплификацией кодирующей последовательности mTERT со смысловым праймером (5'-GATCTGATGATATCGCCACCATGACCAGAGCCCCCAGATG-3; SEQ ID NO:15) и антисмысловым праймером (5'-AGGGGGGATCCGCACACCTGGTAGGCGCAGCTGGGG-3'; SEQ ID NO:16), и клонировали обратно в расщепленный BamHI и EcoRV вектор.
pV1JnsA/TPA-hTERT(AI)-LTBopt: Соответствующий TPA-hTERT(AI)-LTB синтетический фрагмент, полученный в GENEART, клонировали в pV1JnsA с использованием сайтов рестрикции BglII/SalI.
Ad6-TPAmTERT(AI)-LTBopt: Плазмиду 041046pucKana, содержащую последовательность TPA-mTERT(AI)-LTBopt, получали из GENEART. Плазмиду расщепляли BglII и SalI и фрагмент клонировали по BglII/SalI в плазмидный челночный вектор pNEBAd6-CMVpA. Плазмиду pNEBAd6-CMVpA-TPA-mTERT(AI)-LTB, расщепленную EcoRI/HindIII, рекомбинировали с линеаризованной ClaI плазмидой pMRK Ad6 ΔEΙ ΔE3. Плазмиду расщепляли PacI с высвобождением ITR Ad и трансфицировали в клетки Perc-6. Амплификацию вектора Ad6 проводили серийными пассажами и очищали очисткой на стандартном градиенте CsCl.
Ad6-hTERT(AI): Плазмиду pCRsript, содержащую последовательность hTERT дикого типа (последовательность hTERT дикого типа получали путем обратной транскрипции мРНК опухолевых клеток человека), расщепляли BglII/XbaI, достраивали фрагментом Кленова и клонировали в pV1JnsA, расщепленную EcoRV и BglII. pV1JnsA-hTERT подвергали мутагенезу с использованием олигонуклеотида (5'-CTGTACTTTGTCAAGGTGGCTATCACGGGCGCGTACG-3'; SEQ ID NO:17) и набора для множественного сайт-специфического мутагенеза Stratagene quikchange (Stratagene, LA Jolla, CA; Cat: 200513) с получением pV1JnsA-hTERT(AI). Плазмиду расщепляли BglII и SalI и фрагмент клонировали по BglII/SalI в плазмидный челночный вектор pNEBAd6-CMVpA. Плазмиду pNEBAd6-CMVpA-hTERT(AI) расщепляли PacI/PmeI и рекомбинировали с линеаризованной ClaI плазмидой pMRK Ad6 ΔEΙ ΔE3. Плазмиду разрезали PacI с высвобождением ITR Ad и трансфицировали в клетки Perc-6. Амплификацию вектора Ad6 проводили серийными пассажами и очищали очисткой на стандартном градиенте CsCl.
ПРИМЕР 3
Анализ ELISPOT IFNγ
Спленоциты мышей, секретирующие IFNγ антиген-зависимым образом, детектировали с использованием стандартного иммуноферментного спот-анализа (ELISPOT) (Miyahira et al. J Immunol Methods 181(1): 45-54 (1995)). 96-Луночные планшеты MAIP (Millipore Corp., Billerica, MA) покрывали 100 мкл/лунку очищенными антителами крыс к IFNγ мышей (IgG1, клон R4-6A2, Pharmingen), разведенными до 2,5 мкг/мл в стерильном PBS. После отмывки с помощью PBS проводили блокирование планшетов 200 мкл/лунку среды RIO в течение 2 часов при 37°C.
Спленоциты получали, удаляя селезенку у подвергнутых эвтаназии мышей в стерильных условиях, с последующим разрушением селезенки растиранием на металлической сетке. Эритроциты удаляли путем осмотического лизиса, добавляя к клеточному осадку 1 мл 0,1× PBS и интенсивно перемешивая в течение приблизительно 15 сек. Затем добавляли один мл 2× PBS и объем доводили до 4 мл с помощью 1× PBS. Клетки осаждали центрифугированием при 1200 об/мин в течение 10 мин при комнатной температуре и осадок ресуспендировали в 1 мл среды R10. Живые клетки подсчитывали с использованием окрашивания Turks.
Спленоциты наносили на планшеты в количестве 5×105 и 2,5×105 клеток/лунку в двух повторах и инкубировали в течение 20 часов при 37°C с суспензией каждого пептида в концентрации 1 мкг/мл. Для каждой мыши в качестве внутреннего положительного контроля использовали конканавалин A (ConA) в концентрации 5 мкг/мл. После отмывки с помощью PBS 0,05% Tween 20 планшеты инкубировали в течение ночи при 4°C с 50 мкл/лунку конъюгированных с биотином антител крыс к IFNγ мышей (RatIgG1, клон XMG 1.2, PharMingen), разбавленных 1:2500 в буфере для анализа. После интенсивной отмывки планшеты проявляли, добавляя 50 мкл/лунку NBT/B-CIP (Pierce) до отчетливого визуального проявления пятен. Реакцию останавливали тщательной отмывкой планшетов дистиллированной водой. Планшеты высушивали на воздухе, а затем подсчитывали пятна с использованием автоматического устройства для считывания планшетов ELISPOT.
ПРИМЕР 4
Внутриклеточное окрашивание цитокинов
От одного до двух миллионов спленоцитов или PBMC мышей, полученных путем забора крови из ретроорбитальной области в ЭДТА, ресуспендировали в 1 мл RPMI с 10% FCS, инкубировали с пулом пептидов (конечная концентрация каждого пептида составляла 5-6 мкг/мл) и брефелдином A (1 мкг/мл; BD Pharmingen кат. №555028/2300kk) при 37°C и 5% CO2 в течение 12-16 часов. Затем клетки отмывали буфером FACS (PBS с 1% FBS, 0,01% NaN3) и инкубировали с очищенным блоком Fc к CD16/CD32 мышей (BD Pharmingen кат. №553142) в течение 15 мин при 4°C. Затем клетки отмывали и окрашивали антителами к поверхности: конъюгированные с PE антитела к CD4 мышей (BD Pharmingen, кат. №553049), конъюгированные с PercP антитела к CD8 мышей (BD Pharmingen, кат. №553036) и конъюгированные с APC антитела к CD3e мышей (BD Pharmingen, кат. №553066) в течение 30 минут при комнатной температуре в темноте. После отмывки клетки фиксировали и нарушали проницаемость мембран раствором Cytofix-Cytoperm Solution (BD Pharmingen, кат. №555028/2300kk) в течение 20 мин при 4°C в темноте. После отмывки раствором PermWash Solution (BD Pharmingen, кат. №555028/2300kk) клетки инкубировали с антителами IFNγ-FITC (BD Pharmingen). Затем клетки отмывали, фиксировали 1% формальдегидом в PBS и анализировали на проточном цитометре FACS-Calibur, используя программное обеспечение CellQuest (Becton Dickinson, San Jose, CA).
ПРИМЕР 5
Анализ цитотоксических T-лимфоцитов (CTL)
Спленоциты получали, удаляя селезенку у подвергнутых эвтаназии мышей в стерильных условиях, с последующим разрушением селезенки растиранием на металлической сетке. Эритроциты удаляли путем осмотического лизиса, добавляя к клеточному осадку 1 мл 0,1×PBS и интенсивно перемешивая в течение приблизительно 15 сек. Клетки селезенки в количестве 2×106 клеток/мл в RIO помещали в 24-луночный планшет для клеточных культур (2 мл клеток/лунку) в присутствии иммуногенного пептида в конечной концентрации 10 мкг/мл. Планшеты инкубировали при 37°C, 95% влажности, 5% CO2 в течение 6 суток. На 3 сутки культивирования добавляли рекомбинантный IL-2 человека в конечной концентрации 10 мкг/мл.
Клетки-мишени, растущие в экспоненциальной фазе, собирали и доводили до 1×106 клеток/мл/пробирку в присутствии иммуногенного пептида в конечной концентрации 10 мкг/мл. Клетки метили 50-100 мкКи 51Cr/пробирку при 37°C в течение 2 часов. Клетки-мишени отмывали три раза по 10 мл среды, центрифугируя при 250 g и комнатной температуре в течение 5 мин, и доводили до 1×105 клеток/мл.
Анализ CTL проводили, добавляя эффектор/клетки-мишени с соотношением э/м 100:1, 50:1, 25:1 и 12,5:1. Через 4 часа инкубации планшеты центрифугировали при 1200 об/мин в течение 5 минут и собирали супернатант (30 мкл/лунку). Планшеты сушили в течение ночи и производили подсчет на планшетном счетчике бета-излучения.
ПРИМЕР 6
Иммунизация мышей
Самок мышей C57BL/6 (H-2b) приобретали у Charles River (Lecco, Italy). Мыши HLA-A2.1 (HHD) были любезно предоставлены F. Lemmonier (Institute Pasteur, Paris, France). Мышей BALB/c (H-2d) приобретали у Charles River (Lecco, Italy). Мышей иммунизировали в возрасте 8 недель. Пятьдесят микрограммов плазмидной ДНК в объеме 50 мкл посредством электропорации вводили в четырехглавую мышцу бедра мышей, как описано ранее (Rizzuto et al. Proc. Natl. Acad. Sci. U.S.A. 96(11):6417-22 (1999)). Электропорацию (EP) проводили, как описано ранее (Zucchelli et al. J. Virol. 74(24): 11598-607 (2000); Rizzuto et al, выше). В кратком изложении, электрошок состоял из 10 серий с 1000 прямоугольных биполярных импульсов (90 В/см, 75 мА, 200 мксек/фазу). Инъекции Ad проводили в четырехглавые мышцы мышей в объеме 50 мкл. Клеточный иммунный ответ анализировали в указанное время.
ПРИМЕР 7
Генетическая вакцинация кодирующей TPA-mTERT(AI)-LTB плазмидной ДНК нарушает иммунологическую толерантность
Для определения того, может ли вакцинация мышей TPA-mTERT(AI)-LTB нарушать иммунологическую толерантность, группы по 20 мышей BALB/c иммунизировали 5 еженедельными инъекциями плазмиды pV1J/TPA-mTERT(AI)-LTB с последующей электропорацией, как описано в примере 6. Через одиннадцать суток после последней инъекции у отдельных мышей получали спленоциты, которые использовали для анализа клеточного иммунного ответа анализом ELISPOT IFNγ и путем внутриклеточного окрашивания цитокинов (ICS).
Антиген-специфическую секрецию IFNγ из стимулированных спленоцитов измеряли с использованием трех пулов 15-мерных пептидов mTERT, перекрывающихся по 11 а.к. и охватывающих весь белок mTERT. Пул mTERT-1 состоял из 94 отдельных пептидов, перекрывающих область mTERT от а.к. 1 до а.к. 388. Пул mTERT-2 состоял из 106 отдельных пептидов, перекрывающих область mTERT от а.к. 377 до а.к. 811. Пул mTERT-3 состоял из 78 отдельных пептидов, перекрывающих область mTERT от а.к. 801 до а.к. 1122. В качестве отрицательного контроля также измеряли продукцию цитокинов после стимуляции спленоцитов с использованием ДМСО в той же концентрации, которую использовали для растворения пептидов TERT. Иммунный ответ, индуцированный против адъюванта LTB, также детектировали с использованием пула из 24 отдельных 15-мерных пептидов, перекрывающихся 11 а.к. и покрывающих всю последовательность LTB. Результаты ELIspot указывают на то, что вакцинация мышей TPA-mTERT(AI)-LTBopt вызывала клеточный иммунный ответ (CMI) против N-концевых и центральных областей белка TERT, содержащихся в пулах пептидов TERT-1 и TERT-2 (фиг.3). Также посредством ELIspot выявляли CMI против LTB (фиг.3).
Для лучшей характеристики T-клеточного ответа, вызываемого плазмидой pV1J/TPA-mTERT(AI)-LTBopt, и для идентификации субпопуляции CD8+ и CD4+ T-клеток, ответственной за продукцию IFNγ, в спленоцитах вакцинированных мышей проводили внутриклеточное окрашивание на IFNγ и анализировали с помощью FACS. Полученные с использованием ICS данные демонстрируют, что генетическая иммунизация мышей BALB/c, вакцинированных TPA-mTERT(AI)-LTBopt, вызывала значительный CD8+ T-клеточный ответ против N-концевой области белка TERT (содержащейся в пептидном пуле mTERT-1, см. фиг.4). CD4+ T-клеточный ответ картировали в центральной области TERT (пептидный пул mTERT-2). В отличие от этого против C-концевой области белка (пептидный пул mTERT-3) у этих мышей CMI не выявляли. Также выявляли CD4+ T-клеточный ответ против LTB (фиг.4).
Для дальнейшего картирования CD8+ T-клеточного ответа использовали средний пул 15-мерных пептидов mTERT. Каждый средний пул содержал десять 15-мерных пептидов, каждый из которых содержался в отдельном среднем пуле. Таким образом, положительная реакция 2 средних пулов могла бы однозначно идентифицировать один индивидуальный 15-мерный пептид как иммуногенный. Поскольку длина CD8+ эпитопов всегда составляет 9 а.к., любой CD8+ пептид содержится в двух соседних 11-мерных пептидах. Результаты показали, что CD8+ T-клеточный иммунный ответ был локализован в области TERT, содержащейся в следующих 15-мерных пептидах mTERT: mTERT-41 (TERTaa161; LVPPSCAYQVCGSPL (SEQ ID NO:18) и mTERT-42 (TERTaa165; SCAYQVCGSPLYQIC; SEQ ID NO:19). Перекрывая два 15-мерных пептида (mTERT41 и mTERT-42), синтезировали и тестировали в ICS три возможных реактивных нонамера. Наконец, у мышей BALB/c в следующей последовательности (mTERTaa167; AYQVCGSPL; SEQ ID NO:20) был идентифицирован иммунодоминантный CD8+ эпитоп. Эти результаты демонстрируют, что вакцинация мышей BALB/c с использованием pV1J/TPA-mTERT-LTB может нарушать иммунологическую толерантность на антиген mTERT.
Для подтверждения данных, полученных на мышах BALB/c, мышей C57BL/6 (B6) также иммунизировали 5 еженедельными инъекциями плазмиды pV1J/TPA-mTERT(AI)-LTBopt. В спленоцитах вакцинированных мышей проводили внутриклеточное окрашивание IFNγ и анализировали путем FACS. Подобно результатам, полученным у мышей BALB/c, толерантность к mTERT нарушалась у всех мышей B6. Кроме того, путем ICS после стимуляции пептидами TERT были выявлены и CD4+, и CD8+ T-клетки, продуцирующие IFNγ (см. фиг.6).
CD8+ иммунный ответ, вызванный вакцинацией ДНК у мышей C57BL/6, в основном активировался по отношению к центральной области белка TERT (пептидный пул mTERT-2). Более слабый CD8+ иммунный ответ также детектировали по отношению к пептидному пулу mTERT-1. CD8+ иммунный ответ у мышей B6 был картирован с использованием средних пулов, как описано выше. У B6 идентифицировали два CD8+ эпитопа. Первую иммуногенную последовательность картировали в а.к. последовательности mTERT198 (VGRNFTNL; SEQ ID NO:21), а второй CD8+ эпитоп картировали в последовательности mTERT486 (SLGKYGKL; SEQ ID NO:22). Кроме того, CD4+ иммунный ответ был локализован в N-концевой области TERT (пептидный пул mTERT-1). Иммунного ответа против C-концевой области mTERT не выявлено.
Аналогично результатам, полученным у мышей BALB/c, описанных выше, иммунизация и анализ последующего иммунного ответа у мышей B6 подтвердили, что иммунизация TPA-mTERT(AI)-LTBopt может нарушать иммунную толерантность к антигену mTERT. Таким образом, взятые вместе результаты, полученные у мышей BALB/c и B6, указывают на то, что слитая последовательность TPA-mTERT(AI)-LTBopt представляет собой самостоятельный сильный иммуноген вне зависимости от используемой линии мышей.
ПРИМЕР 8
Цитолитическая активность специфичных к TERT CD8 + T-клеток
Для детекции цитолитической активности специфичных к TERT CD8+ T-клеток, вызванной иммунизацией мышей BALB/c посредством pV1J/TPA-mTERT(AI)-LTBopt, использовали анализ цитотоксических T-лимфоцитов (CTL). Мышей иммунизировали 5 еженедельными инъекциями плазмиды pV1J/TPA-mTERT(AI)-LTBopt с последующей электропорацией. Спленоциты мышей, полученные у 2 отдельных мышей, собирали через 10 суток после последней иммунизации. Эти спленоциты стимулировали в течение одной недели иммуногенным пептидом mTERTaa167 (AYQVCGSPL; SEQ ID NO:20) для получения активированных in vitro T-клеток. В качестве мишени использовали сингенную линию опухолевых клеток BALB/c 4T1 (Aslakson et al. Cancer Res. 52(6):1399-405 (1992)). 4T1 клетки нагружали иммунореактивным пептидом и метили Cr51 для использования в качестве мишени для анализа CTL. Активированные T-клетки совместно инкубировали с клетками-мишенями 4T1 в различных соотношениях эффекторы/мишень. Активированные T-клетки, полученные у мыши №1 и мыши №2, в отношении клеток-мишеней при загрузке иммуногенным пептидом mTERTaa167 (AYQVCGSPL; SEQ ID NO:20) продемонстрировали цитолитическую активность. Две мыши при соотношении эффекторы/мишень 50/1 продемонстрировали различную степень цитотоксической активности в диапазоне от 80% (мышь №1) до 25% (мышь №2).
Результаты указывают на то, что у мышей BALB/c после ДНК-иммунизации посредством TPA-mTERT(AI)-LTBopt в качестве собственного антигена индуцирован цитотоксический иммунный ответ (фиг.5).
ПРИМЕР 9
Сравнительная иммуногенность конструкции TPA-mTERT(AI)-LTBopt
Для сравнения иммуногенности секретируемого белка mTERT(AI), слитого с LTB, кодируемого pV1J/TPA-mTERT(AI)-LTBopt, с несекретируемым вариантом mTERT(AI) без слияния с LTB, была сконструирована производная плазмида pV1J, несущая mTERT(AI), как описано в примере 2.
Группы из 6 мышей BALB/c иммунизировали 5 еженедельными инъекциями плазмид pV1J/TPA-mTERT(AI)-LTBopt или pV1J/mTERT(AI)opt. Иммунный ответ подвергали мониторингу путем внутриклеточного окрашивания на IFNγ с использованием mTERTaa167 последовательности CD8+ иммунодоминантного эпитопа, ранее идентифицированного для мышей BALB/c (AYQVCGSPL; SEQ ID NO:20).
Результаты указывают на то, что TPA-mTERT(AI)-LTBopt представляет собой лучшую конструкцию индукции CD8+ иммунного ответа против теломеразы мышей, чем mTERT(AI) (фиг.7). Наблюдаемое в случае ДНК-иммунизации различие в CD8+ T-клеточном ответе, индуцированном в иммунизированной посредством TPA-mTERT-LTB(AI)opt группе, статистически отличалось от ответа, полученного с использованием mTERT(AI)opt (p=0,04 t-критерий Стьюдента).
ПРИМЕР 10
Сравнение схем иммунизации
Клеточный иммунный ответ против mTERT, вызванный иммунизацией мышей BALB/c ДНК TPA-mTERT(AI)-LTBopt совместно только с электрической стимуляцией, сравнивали с иммунным ответом, индуцированным разнородными схемами иммунизации при примировании/повторном стимулирующем введении, используя ДНК TPA-mTERT(AI)-LTBopt с электрической стимуляцией в качестве примирования и Ad6 TPA-mTERT(AI)-LTBopt в качестве конечного повторного стимулятора иммунного ответа.
Группу из 10 мышей иммунизировали 5 еженедельными инъекциями плазмиды pV1J/TPA-mTERT(AI)-LTBopt. Через одну неделю проводили повторную стимулирующую инъекцию, состоящую из одной дополнительной инъекции 50 мкг ДНК с электропорацией (EP) или 1010 в.ч. Ad6/TPA-mTERT(AI)-LTBopt. Для оценки полученного в результате клеточного иммунного ответа проводили внутриклеточное окрашивание на IFNγ PBMC мышей. В качестве антигена использовали пептид, содержащий специфичный для CD8+ T-клеток эпитоп mTERT167 (AYQVCGSPL; SEQ ID NO:20), который был ранее картирован в N-концевой области TERT. Результаты указывают на то, что амплитуда иммунного ответа, вызванного сочетанием ДНК-ЭП/Ad, была в 4,6 раз больше, чем амплитуда, которую наблюдали у мышей, у которых использовали только схему ДНК-ЭП (см. фиг.8).
ПРИМЕР 11
Вакцинация TPA-mTERT-LTB контролирует рост опухоли
Для определения того, может ли специфичный для mTERT иммунный ответ оказывать противоопухолевое действие, мышей BALB/c до вакцинации обрабатывали химическим канцерогеном диметилгидразином путем интраперитонеальной (и/п) инъекции. Обработка мышей этим канцерогеном приводила к нарастающему прогрессированию опухоли в кишечнике, характеризуемому нарушением образования крипт, аденомой и, наконец, карциномой. Химическая обработка этих мышей не ослабляла их иммунитет, так как иммунизация TPA-mTERT(AI)-LTBopt вызывала иммунный ответ, сравнимый с ответом, ранее наблюдаемым у необработанных мышей (данные не показаны).
Для оценки воздействия генетической иммунизации на ранние стадии канцерогенеза, индуцированного DMH, мышей BALB/c обрабатывали DMH и иммунизировали ДНК с последующей электропорацией (ДНК-ЭП) pV1J/TPA-mTERT(AI)-LTBopt в соответствии со схемой, описанной на фиг.9.
Двадцать мышей подвергали эвтаназии через 12 недель после начала обработки DMH для подсчета количества образованных ACF (фокусов нарушенных крипт) и очагов аденомы. У вакцинированных мышей по сравнению с невакцинированными мышами наблюдали значительное снижение присутствующего количества ACF и количества очагов аденомы (фиг.10).
Также проводили мониторинг эффективности вакцинации mTERT на поздней стадия развития опухоли. Для усиления и поддержания иммунного ответа на mTERT мышей подвергали повторным ДНК-ЭП TPA-mTERT(AI)-LTBopt и Ad6TPA-mTERT(AI)-LTBopt (фиг.11). Иммунный ответ, вызванный у этих мышей, был сравним с иммунным ответом, который выявляли у мышей BALB/c, вакцинированных по той же схеме ДНК/Ad и не обработанных DMH.
Двадцать мышей подвергали эвтаназии через 30 недель после начала обработки DMH для подсчета количества образованных ACF (фокусов нарушенных крипт) и очагов аденомы. Подобно результатам, наблюдаемым на ранней стадии развития опухоли, у вакцинированных мышей по сравнению с невакцинированными мышами наблюдали значительное снижение количества ACF и очагов аденомы (фиг.12).
Наконец, опухоли, обнаруженные у вакцинированных мышей, имели меньший объем по сравнению с опухолями, обнаруженными у невакцинированных мышей (фиг.13A). Кроме того, гистологический анализ показал, что опухоли, присутствующие у вакцинированных мышей, были на менее поздней стадии по сравнению с опухолями у невакцинированных животных (фиг.13B).
ПРИМЕР 12
Характеристика pV1JTPA-hTERT(AI)-LTBopt
Иммуногенный потенциал pV1J/TPA-hTERT(AI)-LTBopt оценивали в качестве чужеродного антигена, поскольку мыши, трансгенные по hTERT, недоступны. Использовали мышей C57BL/6, трансгенных по HLA-A2 человека (трансгенные мыши HHD), так что иммуногенные пептиды TERT могли быть представлены в контексте HLA класса I человека. Трансгенных мышей HHD вакцинировали путем ДНК-ЭП, проводя каждые две недели 2 инъекции по 50 мкг плазмиды pV1J/TPA-hTERT(AI)-LTBopt. Индуцированный иммунный ответ оценивали посредством ICS PBMC мышей. В качестве антигена использовали иммуногенный пептид hTERT, соответствующий эпитопу hTERT865 RLVDDFLLV (SEQ ID NO:23). Эта последовательность эпитопа описана ранее как сильно связывающаяся с HLA-A2 и как индуктор активности CTL в экспериментах примирования T-клеток in vitro (Dupont et al. Cancer Research 65(12): 5417-27 (2005)).
Результаты, полученные у 10 мышей, показаны как среднее, 12% CD8+ клеток при стимуляции иммуногенным пептидом hTERT865 продуцировали IFNγ (фиг.14). Результаты указывают на то, что кандидат для вакцины pV1JTPA-hTERT(AI)-LTBopt является иммуногенным и индуцирует рестриктированный по HLA-A2 CD8 иммунный ответ.
ПРИМЕР 13
Характеристика иммунного ответа у вакцинированных мышей после повторной стимуляции Ad6-hTERT(AI)
Конструкцию Ad6hTERT(AI) собирали, как описано в примере 2. Эта конструкция содержала кодоны дикого типа, за исключением двух мутаций, которые были введены для устранения теломеразной каталитической активности. Также была получена конструкция Ad6 со слитой последовательностью hTERT-LTB, но ее не смогли сохранить, поэтому дальнейшее тестирование проводили только с Ad6hTERT(AI).
Конструкцию Ad6hTERT(AI) тестировали на ее стимулирующее воздействие на иммунный ответ, индуцированный ДНК-ЭП у трансгенных мышей HHD. Группы из 10 мышей вакцинировали путем ДНК-ЭП с 1 инъекцией 50 мкг плазмиды pV1J/TPA-hTERT(AI)-LTBopt и повторно стимулировали второй инъекцией ДНК или посредством 1010 в.ч. Ad6hTERT через две недели после первой инъекции ДНК. Индуцированный иммунный ответ измеряли с использованием ранее идентифицированного иммунодоминантного для аллеля HLA-A2 пептида hTERT865. Реактивные CD8+ T-клетки, продуцирующие IFNγ, детектировали с высокой частотой. Как показано на фиг.14, амплитуда иммунного ответа, вызванного сочетанием ДНК-ЭП/Ad, была в 3 раза больше, чем амплитуда, наблюдаемая у мышей, которых обрабатывали только по схеме ДНК-ЭП.
Результаты продемонстрировали, что специфичный для hTERT иммунный ответ, примированный инъекцией ДНК pV1J/TPA-hTERT(AI)-LTBopt, усиливался в результате стимулирующей повторной инъекции Ad6-hTERT(AI).
ПРИМЕР 14
Характеристика активности CTL, индуцированной иммунизацией hTERT
CD8+ T-клеточный ответ на hTERT, вызванный у вакцинированных мышей, дополнительно характеризовали тестированием цитотоксической активности (CTL) активированных против клеток-мишеней T-клеток. Спленоциты иммунизированных мышей стимулировали в культуре в течение 6 суток в присутствии пептида hTERT865 (SEQ ID NO:23; см. пример 12) и IL-2. Затем тестировали активированные CD8+ T-клетки на их литическую активность против клеток-мишеней в анализе высвобождения Cr51. В качестве клеток-мишеней использовали клетки HeLa, стабильно трансфицированные для экспрессии химерной молекулы MHC класса I, присутствующей у мышей HHD (HeLa-HHD). Клетки-мишени HeLa-HHD для эндогенной сверхэкспрессии антигена TERT экзогенно загружали иммуногенным пептидом hTERT865 или на 24 часа раньше инфицировали вектором Ad6, кодирующим hTERT. В качестве контроля клетки-мишени инфицировали вектором Ad6, кодирующим GFP. Результаты указывают на то, что CD8+ T-клетки, индуцированные путем описанной вакцинации, были способны уничтожать клетки-мишени, сверхэкспрессирующие антиген hTERT (фиг.15).
ПРИМЕР 15
Характеристика иммуногенности вакцины TERT человека у макак-резус
Иммуногенность системы генетической вакцинации на основе примирования ДНК, повторной стимуляции Ad посредством hTERT оценивали у макак-резус. Так как гомология последовательностей TERT человека и TERT макаки-резус составляет 96%, ожидалось, что главную роль в определении эффективности вакцинации будет играть толерантность.
Протокол вакцинации, состоящий из пяти инъекций ДНК-ЭП (pV1J/TPA-hTERT-LTB, 5 мг/инъекцию), проводили каждые две недели. Через четыре недели после последней обработки ДНК-ЭП обезьян повторно стимулировали вектором Ad, экспрессирующим hTERT (Ad6-hTERT, 1011 в.ч.), 2 раза с двухнедельным интервалом. Каждую неделю собирали данные о массе тела и клинических признаках. Отрицательных эффектов на массу тела животных или возникновения клинических признаков не наблюдали.
Индуцированный иммунный ответ подвергали мониторингу анализом ELISPOT IFNγ PBMC с использованием пептидных пулов hTERT, аналогично анализу, описанному в примере 7, и пептидного пула, покрывающего последовательность LTB. Иммунный ответ считали положительным, если полученный сигнал по меньшей мере в четыре раза превосходил фоновую реактивность (ДМСО). Анализ ELIspot индуцированного CMI указывает на то, что после первых 2 обработок ДНК-ЭП детектируемый иммунный ответ возникал у четырех из четырех вакцинированных обезьян (фиг.16A). После пятой ДНК-ЭП наблюдали небольшое усиление ответа (фиг.16B). Иммунный ответ также измеряли в конце полного протокола иммунизации, после двух повторных стимуляций Ad. Результаты указывают на то, что повторные стимуляции Ad индуцировали стойкое увеличение амплитуды CMI (фиг.17). Таким образом, этим экспериментом подтвердили эффективность способа гетерологического примирования/повторной стимуляции.




























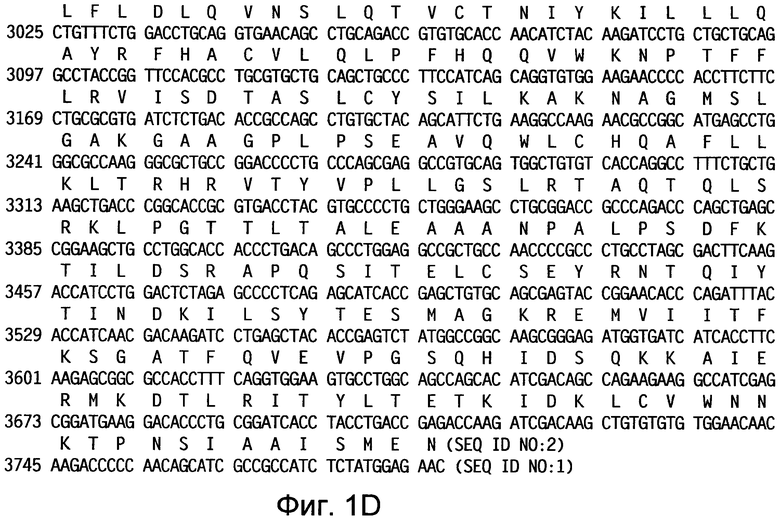




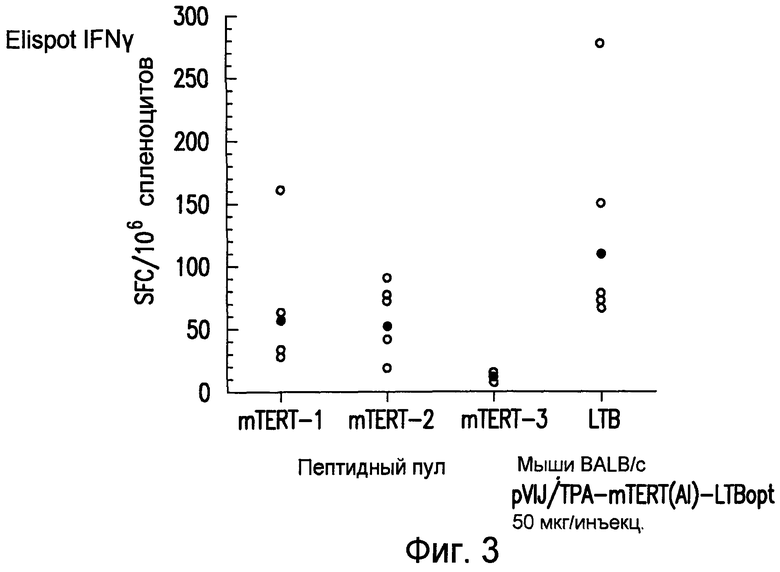
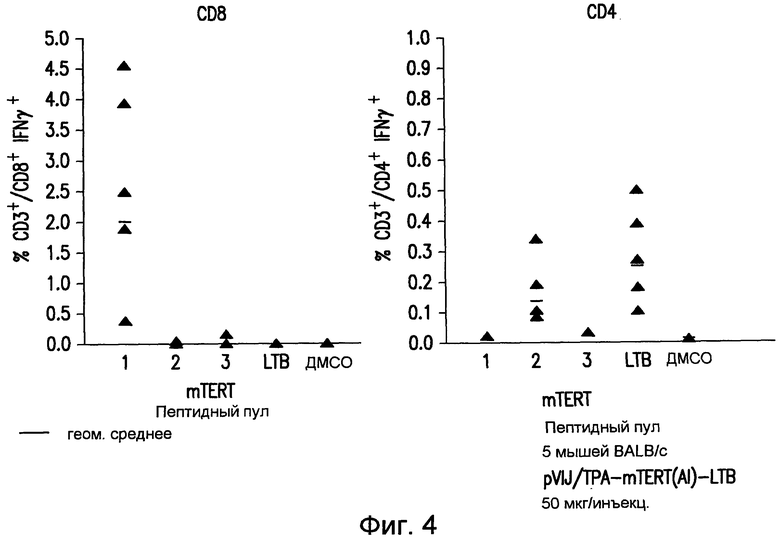
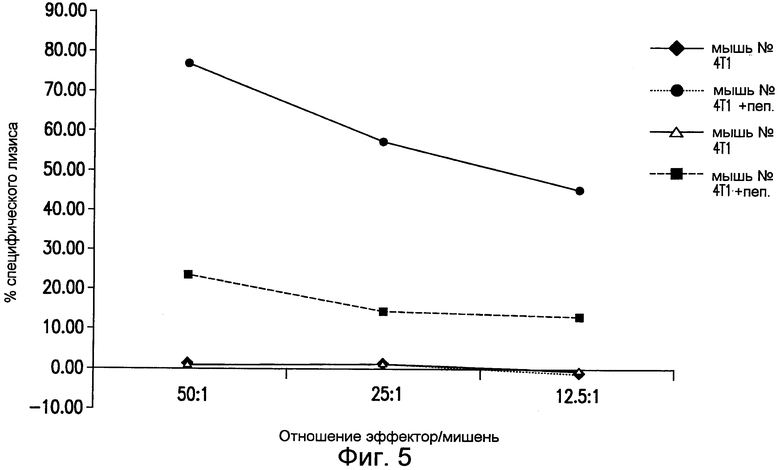

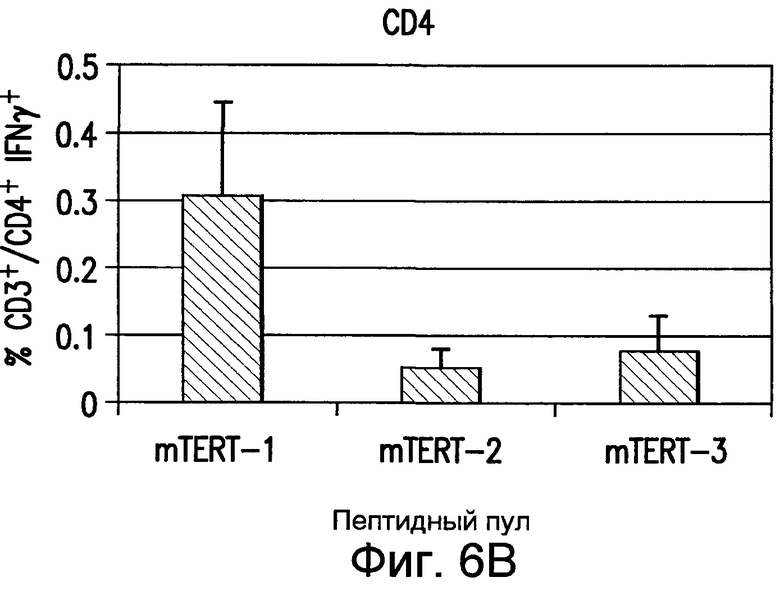
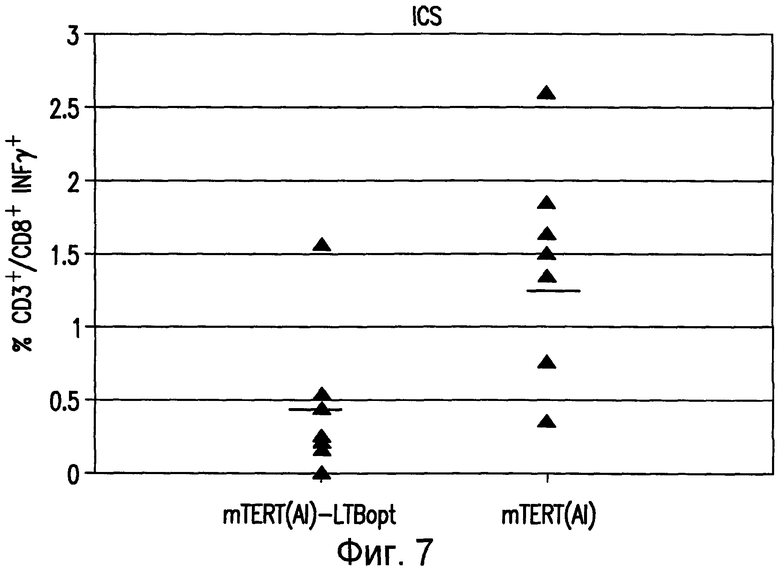
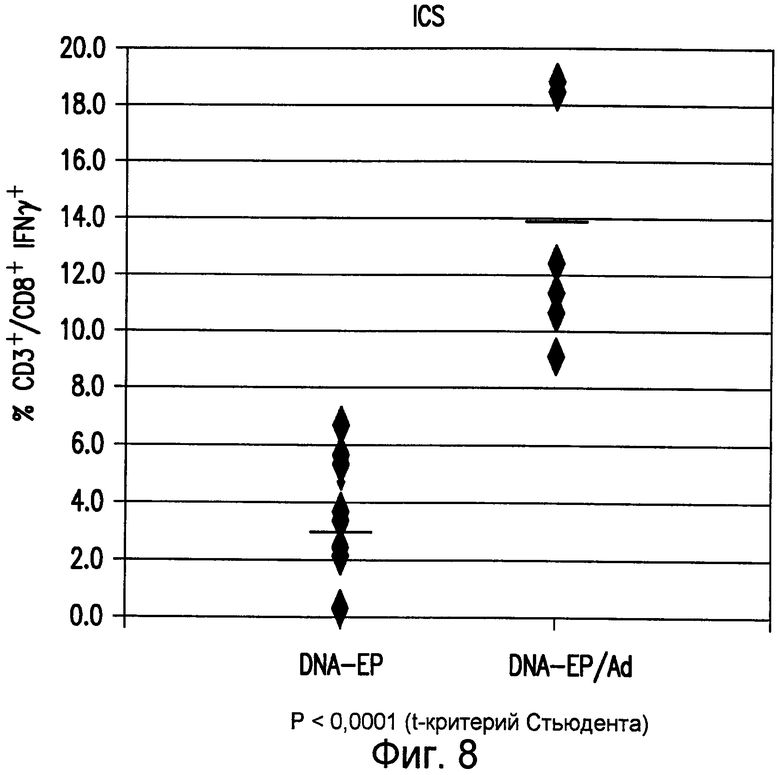
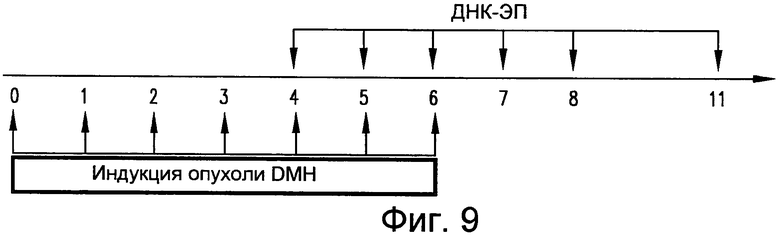
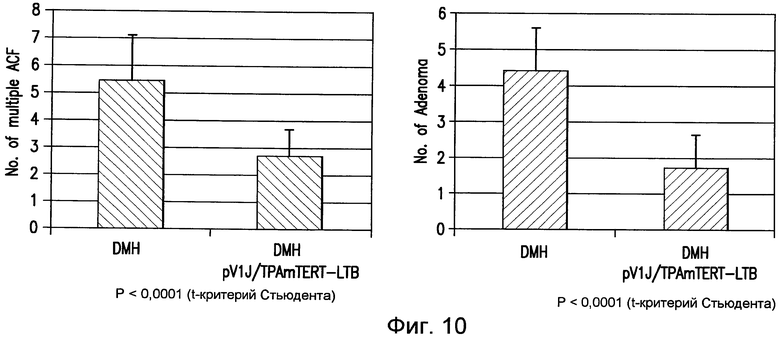
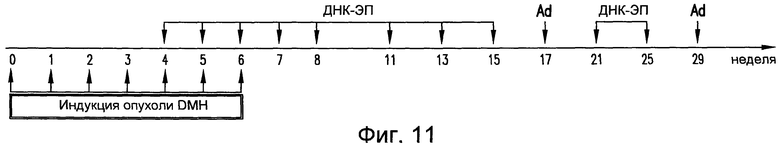
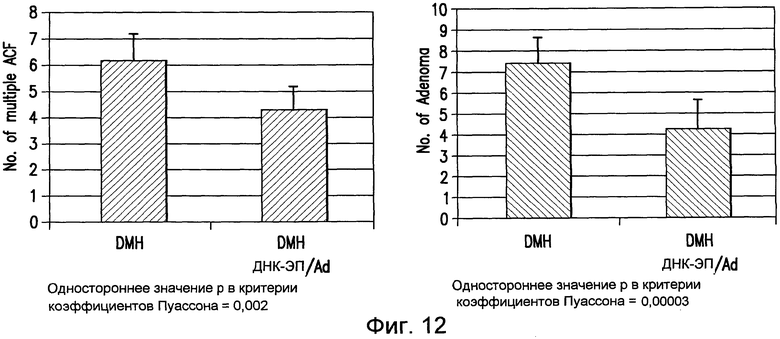
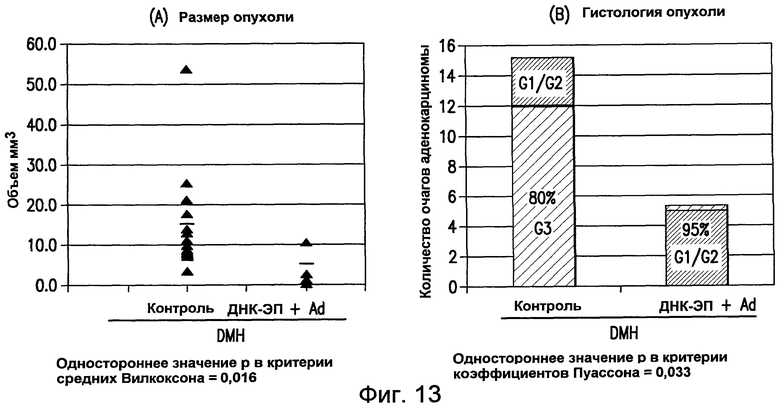
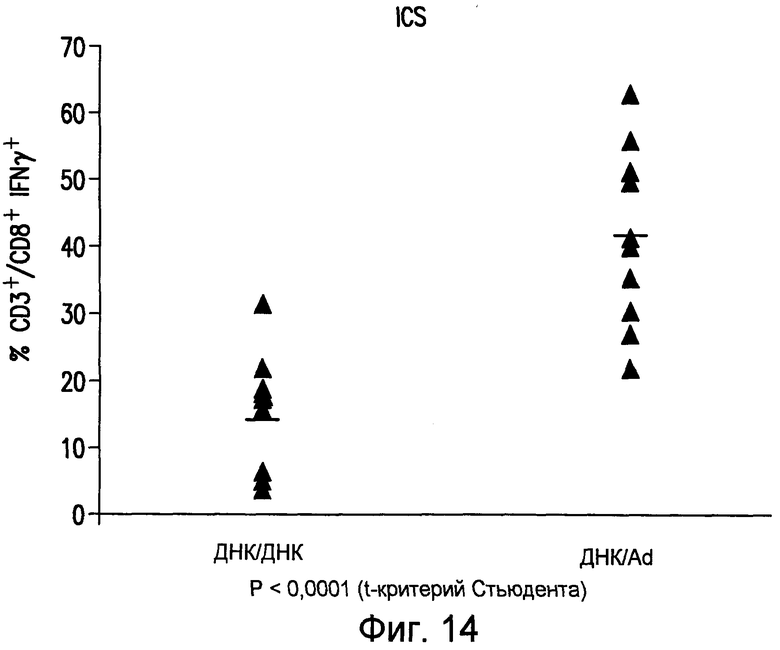
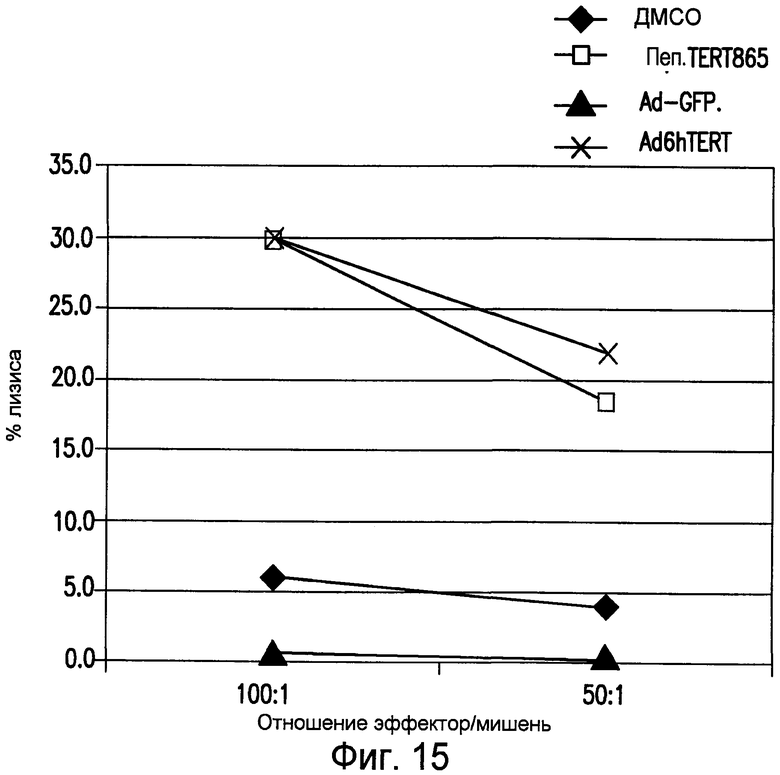
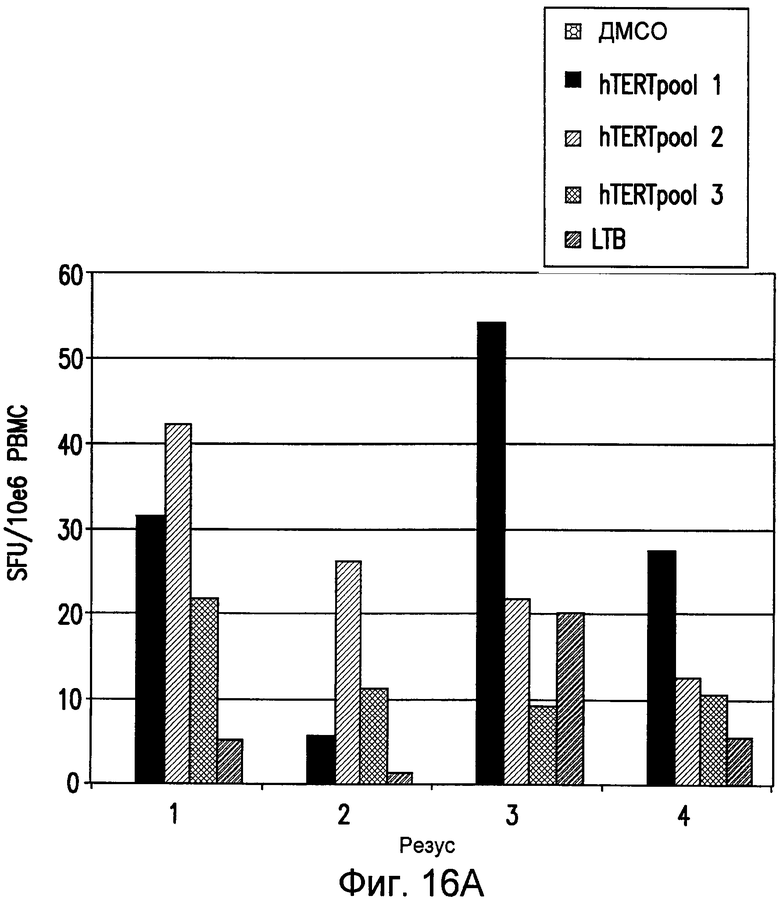
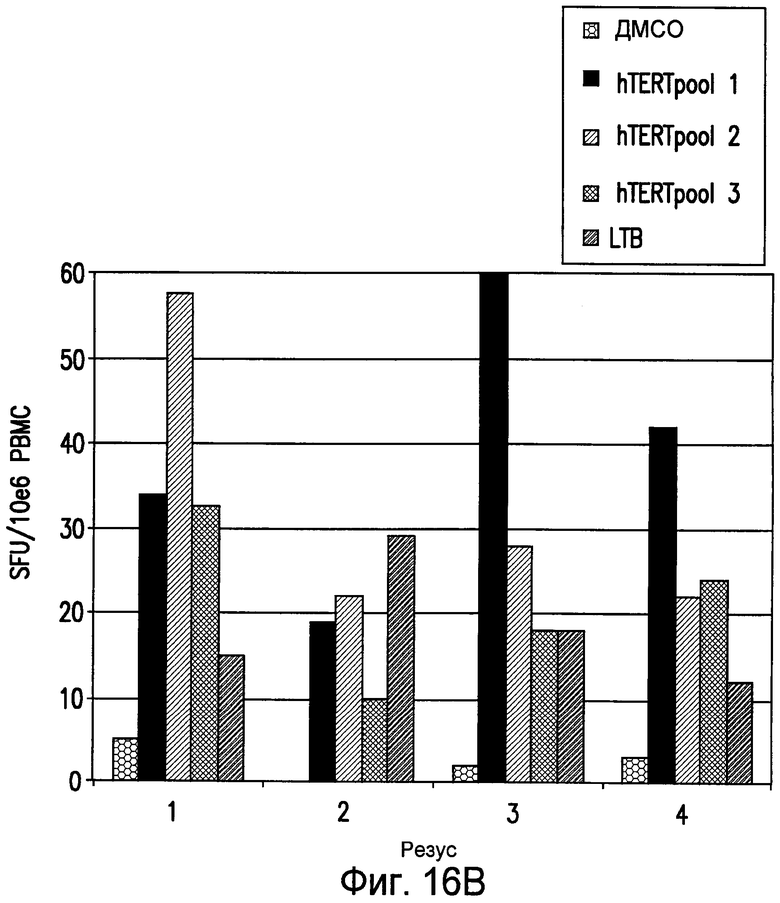
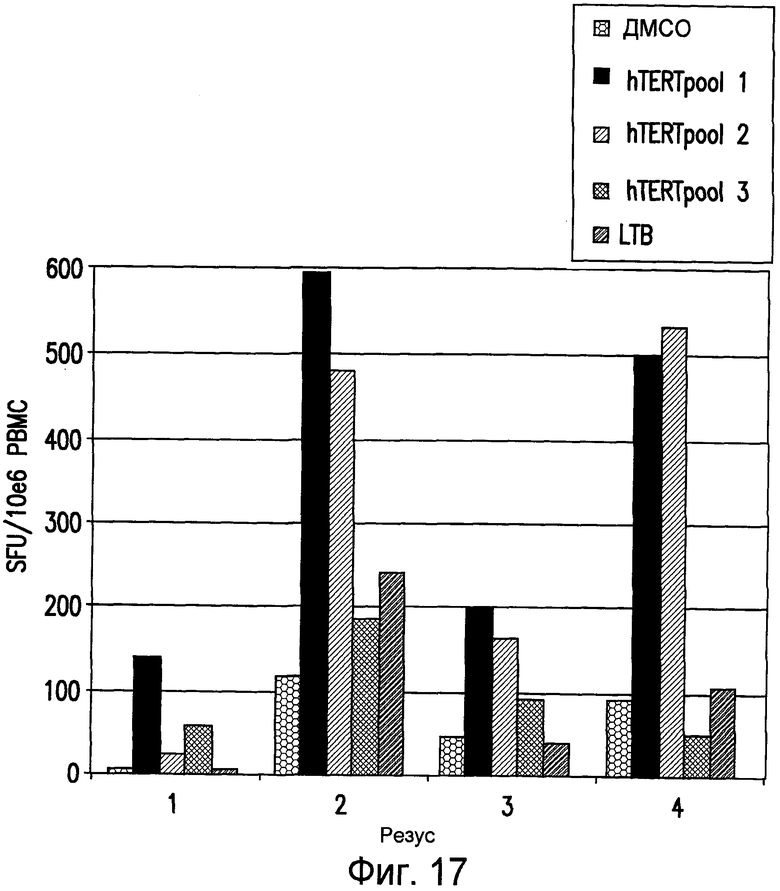
Настоящее изобретение относится к области генетической инженерии и может быть использовано в медицине для индукции или усиления иммунного ответа организма. Предложен полинуклеотид, кодирующий слитый белок теломеразной обратной транскриптазы (TERT), в котором неактивный вариант полипептида TERT объединен со значительной частью субъединицы В термолабильного энтеротоксина (LTB). Показано, что слитый белок по изобретению вызывает у млекопитающего иммунный ответ, который в предпочтительных вариантах осуществления является более сильным, чем иммунный ответ, вызываемый изолированной TERT дикого типа. Настоящее изобретение раскрывает получение слитого белка с вариантом hTERT и необходимые для этого генетические конструкции, в частности аденовирусные векторы и плазмиды, а также клетки-хозяева, обеспечивающие экспрессию полинуклеотида, кодирующего слитый белок по изобретению. 5 н. и 2 з.п. ф-лы, 25 ил., 14 пр.
1. Молекула нуклеиновой кислоты для индукции иммунного ответа против теломеразной обратной транскриптазы человека (hTERT), содержащая нуклеотидную последовательность, кодирующую слитый белок hTERT-LTB, где молекула нуклеиновой кислоты содержит последовательность нуклеотидов SEQ ID NO: 1.
2. Очищенный слитый белок hTERT-LTB для индукции иммунного ответа против hTERT, кодируемый молекулой нуклеиновой кислоты по п.1.
3. Вектор экспрессии, содержащий молекулу нуклеиновой кислоты по п.1.
4. Вектор по п.3, где вектор представляет собой плазмидный вектор или аденовирусный вектор.
5. Вектор по п.16, где вектор представляет собой плазмиду pV1J.
6. Клетка-хозяин, содержащая вектор экспрессии по п.3, для экспрессии нуклеиновой кислоты, кодирующей hTERT-LTB.
7. Способ получения слитого белка hTERT-LTB в рекомбинантной клетке-хозяине, включающий:
(a) введение вектора экспрессии, содержащего молекулу нуклеиновой кислоты по п.1, в подходящую клетку-хозяина и
(b) культивирование клетки-хозяина в условиях, обеспечивающих экспрессию указанной нуклеиновой кислоты, кодирующей hTERT-LTB, с получением слитого белка hTERT-LTB.
| WO 2004002408, 08.01.2004 | |||
| WO 2005077977, 25.08.2005 | |||
| RU 2000116260, 24.11.1998. |

