Область техники, к которой относится изобретение
Настоящее изобретение в общем случае относится к терапии рака. Более конкретно, настоящее изобретение относится к синтетическим полинуклеотидам, кодирующим человеческий ассоциированный с опухолью полипептидный антиген эпидермального фактора роста 2/neu, обозначенный в настоящем описании hHER2.opt, причем полинуклеотиды являются кодон-оптимизированными для экспрессии в клеточной среде человека. Настоящее изобретение также относится к синтетическим полинуклеотидам, кодирующими укороченную форму антигена HER2/neu, в настоящем описании обозначенную как hHER2ECDTM.opt, причем полинуклеотиды являются кодон-оптимизированными для экспрессии в клеточной среде человека. Настоящее изобретение также относится к рекомбинантным векторам и хозяевам, содержащим указанные синтетические полинуклеотиды. Настоящее изобретение также предоставляет аденовирусные векторные и плазмидные конструкции, несущие hHER2.opt, и их использование в вакцинах и фармацевтических композициях для профилактики и лечения рака.
Уровень техники
Эпидермальный фактор роста-2 представляет собой трансмембранный ассоциированный с опухолью антиген, кодируемый HER2/neu протоонкогеном (также называемым c-erbB-2), который является членом семейства рецепторов эпидермального фактора роста рецепторов клеточной поверхности. Ген HER2 первоначально был выделен из нейроглиобластомы крысы (Shih et al., Nature 290:261-264 (1981)), а позже клонирован и описан из человеческих клеток (Coussens et al., Science 230:1132-39(1985); King et al., Science 229:974-76 (1985)).
Кроме того, HER2/neu классифицирован как член семейства HER рецепторных тирозинкиназ, которые состоят из четырех рецепторов, принимающих участие в регуляции клеточного роста и дифференцировке. Рецепторы HER способствуют поддержанию нормального роста клеток путем связывания лигандов фактора роста в виде димеров. В частности, человеческий HER2 формирует гетеродимеры с другими членами семейства EGFR (HER1, HER3 и HER4) (Klapper et al., Adv Cancer Res 77:25-79 (2000)). После димеризации hHER2 и автофосфорилирования тирозина генерируются сайты сцепления для цитоплазматических сигнальных молекул и инициируется рекрутирование вторичных сигнальных молекул. Таким образом, инициируются каскады внутриклеточной сигнализации, которые в результате приводят к активации генов, играющих важную роль в росте клетки.
Низкие уровни экспрессии транскрипта HER2/neu и кодированного белка 185 кДа обычно детектируют во взрослых эпителиальных клетках различных тканей, включая кожу, и молочную железу, и ткани желудочно-кишечного тракта, половых путей и мочеполового тракта (Press et al., Oncogene 5:953-962 (1990)). Более высокие уровни экспрессии HER2/neu также детектируют в соответствующих эмбриональных тканях во время эмбрионального развития (Press et al., см. выше).
Некоторые исследования позволяют рассматривать антиген HER2 в качестве привлекательной мишени для активной специфической иммунотерапии. Во-первых, обычно происходит сверхэкспрессия или амплификация гена HER2/neu в различных злокачественных опухолях, таких как карциномы молочной железы, яичников, мочевого пузыря, толстой кишки и простаты, и аденокарцинома легких (см. Disis и Cheever, Adv. Cancer Research 71:343-371 (1997)). Сверхэкспрессия HER2/neu коррелирует с неблагоприятным прогнозом и с более высокой частотой рецидивов у раковых пациентов (Slamon et al., Science 244:707-712 (1989)). Амплификация человеческого HER2 приводит к увеличенной активности МАР-киназы и клеточной пролиферации и способствует агрессивному поведению раковых клеток (Ben-Levy et al., Embo J 13(14):3302-11 (1994)). Высокий уровень экспрессии HER2, наблюдаемый в клетках опухолей, является полной противоположностью низкому уровню, ассоциированному с нормальными зрелыми тканями.
Кроме того, многие раковые пациенты, страдающие от злокачественных опухолей, ассоциированных со сверхэкспрессией HER2/neu, имели иммунные ответы на белок HER2. Анти-hHER2 цитотоксические Т лимфоциты (CTL) были выделены у пациентов с раком молочной железы и яичников (Ioannides et al., Cell Immunol 151(1):225-34 (1993); Peoples et al., Proc Natl Acad Sci USA 92(14):6547-51 (1995)). Были определены некоторые HLA-A2.1-ассоциированные пептиды hHER2, и пептид-специфичные Т-клетки могут быть генерированы in vitro (Fisk et al., Cancer Res 57(1):8-93 (1997); Yoshino et al., Cancer Res 54(13):3387-90 (1994); Lustgarten et al., Hum Immunol 52(2):109-18 (1997)).
Вышеуказанные данные демонстрируют, что у раковых пациентов активируются анти-ErbB-2 иммунные эффекторные механизмы, и подчеркивают потенциальное преимущество усиления такой иммунной реактивности. Эффективная вакцина, вызывающая иммунный ответ к HER2/neu, должна как усиливать такую невосприимчивость к уровню, который является защитным и/или профилактическим, так и преодолевать самотолерантность.
Исходя из вышеперечисленного, HER2/neu был исследован в качестве мишени при разработке иммунологической терапии злокачественных опухолей. Анти-HER2 моноклональные антитела исследовали как средства терапии рака молочной железы, при этом подходы с использованием каждого из антител демонстрировали различные уровни успеха (обсуждение см. Yarden, Oncology 61 (suppl 2):1-13 (2001)).
Кроме того, сообщалось о вакцинах на основе пептидов и ДНК, нацеленных на HER2/neu. Amici et al. (патент США 6127344) раскрыли способ индуцирования иммунитета к HER2/neu путем введения вектора экспрессии, содержащего кДНК человеческого HER2/neu полной длины, функционально связанную с промотором цитомегаловируса человека. Morris et al. (WO 2004/041065) раскрывают способ вакцинации дендритными клетками, модифицированными аденовирусными векторами, экспрессирующими несигнальный ген HER2/neu. Cheever и Disis раскрывают способы иммунизации людей пептидами HER2 против HER2/neu-ассоциированных видов рака (патент США 5846538). Кроме того, вакцины на основе пептидов HER2/neu были изучены на моделях грызунов (например, см. Disis and Cheever, Adv. Cancer Res. 71:343-71 (1997)).
Развитию и коммерциализации многих вакцин препятствовали трудности, связанные с получением высоких уровней экспрессии экзогенных генов в успешно трансформированных организмах хозяев. Следовательно, несмотря на идентификацию нуклеотидных последовательностей дикого типа, кодирующих белок hHER2, описанный выше, была бы крайне желательной разработка легко воспроизводимого источника человеческого белка HER2, который использует hHER2-кодирующие нуклеотидные последовательности, оптимизированные для экспрессии в предполагаемой клетке хозяина, причем указанный источник предоставляет возможность для разработки противораковой вакцины, которая является эффективной и которой не препятствует самотолерантность.
Сущность изобретения
Настоящее изобретение относится к композициям и способам для выработки или усиления иммунитета к белковым продуктам, экспрессируемым геном человеческого HER2, который ассоциирован с многочисленными видами аденокарциномы, включая рак молочной железы и яичников. В частности, настоящее изобретение предоставляет полинуклеотиды, кодирующие человеческий белок HER2 или укороченную форму человеческого белка HER2, который содержит внеклеточный и трансмембранный домены белка HER2 (далее в настоящем описании hHER2ECDTM), причем указанные полинуклеотиды являются кодон-оптимизированными для экспрессии высокого уровня в человеческой клетке. Настоящее изобретение также предоставляет аденовирусные вектора и вектора на основе плазмид, содержащие синтетические полинуклеотиды, и раскрывает применение указанных векторов в иммуногенных композициях и вакцинах для профилактики и/или лечения HER2-ассоциированного рака. Полинуклеотиды, раскрытые в настоящем описании, являются более эффективными, чем полинуклеотиды HER2 дикого типа, в выработке клеточного и гуморального иммунного ответа на человеческий HER2.
Настоящее изобретение также относится к синтетическим молекулам нуклеиновых кислот (полинуклеотидам), содержащим последовательность нуклеотидов, которые кодируют человеческий антиген эпидермального фактора роста-2 (далее в настоящем описании hHER2), как представлено в SEQ ID NO:2, причем синтетические молекулы нуклеиновых кислот являются кодон-оптимизированными для экспрессии высокого уровня в человеческой клетке (далее в настоящем описании hHER2.opt). Настоящее изобретение также относится к синтетическим молекулам нуклеиновых кислот (полинуклеотидам), содержащим последовательность нуклеотидов, которые кодируют человеческий HER2ECDTM, как представлено в SEQ ID NO:14, причем синтетические молекулы нуклеиновых кислот являются кодон-оптимизированными для экспрессии высокого уровня в человеческой клетке. Молекулы нуклеиновых кислот, раскрытые в настоящем описании, могут быть трансфицированы в выбранную клетку хозяина, причем рекомбинантная клетка хозяина обеспечивает источник для значительных уровней экспрессированного функционального белка hHER2 (SEQ ID NO:2) или белка hHER2ECDTM (SEQ ID NO:14).
Настоящее изобретение также относится к синтетической молекуле нуклеиновой кислоты, которая кодирует мРНК, которая экспрессирует человеческий белок HER2. Предпочтительный аспект этой части настоящего изобретения раскрыт на фиг.1, на которой показана молекула ДНК (SEQ ID NO:1), кодирующая белок hHER2 (SEQ ID NO:2). Предпочтительная молекула нуклеиновой кислоты настоящего изобретения является кодон-оптимизированной для экспрессии высокого уровня в человеческой клетке. Последовательность такого предпочтительного полинуклеотида также содержит мутацию, которая подавляет активность тирозинкиназы (AAA2257GCC, K753A). Нуклеотидные последовательности, которые не содержат такую мутацию, также входят в объем настоящего изобретения.
Настоящее изобретение также относится к синтетической молекуле нуклеиновой кислоты, которая кодирует мРНК, которая экспрессирует человеческий белок HER2ECDTM. Предпочтительный аспект этой части настоящего изобретения раскрыт на фиг.6А, на которой показана молекула ДНК (SEQ ID NO:9), которая кодирует белок hHER2ECDTM (SEQ ID NO:14). Предпочтительная молекула нуклеиновой кислоты настоящего изобретения является кодон-оптимизированной для экспрессии высокого уровня в человеческой клетке.
Настоящее изобретение также относится к рекомбинантным векторам и рекомбинантным клеткам хозяина, как прокариотическим, так и эукариотическим, которые содержат молекулы нуклеиновых кислот, раскрытых в настоящем описании.
Настоящее изобретение также относится к способу экспрессии кодон-оптимизированного человеческого белка HER2 в рекомбинантной клетке хозяина, предусматривающему: (а) введение вектора, содержащего синтетический полинуклеотид, кодирующий человеческий белок HER2, в подходящую клетку хозяина, причем синтетический полинуклеотид является кодон-оптимизированным для оптимальной экспрессии в человеческой клетке; и (b) культивирование клетки хозяина в условиях, которые предоставляют возможность для экспрессии указанного человеческого белка HER2.
Настоящее изобретение также относится к способу экспрессии кодон-оптимизированного человеческого белка HER2ECDTM в рекомбинантной клетке хозяина, предусматривающему: (а) введение вектора, содержащего синтетический полинуклеотид, кодирующий человеческий белок HER2ECDTM, в подходящую клетку хозяина, причем синтетический полинуклеотид является кодон-оптимизированным для оптимальной экспрессии в человеческой клетке; и (b) культивирование клетки хозяина в условиях, которые предоставляют возможность для экспрессии указанного человеческого белка HER2ECDTM.
Другой аспект настоящего изобретения представляет собой способ профилактики или лечения рака, предусматривающий введение млекопитающему вакцинного вектора, содержащего синтетическую молекулу нуклеиновой кислоты, причем синтетическая молекула нуклеиновой кислоты содержит последовательность нуклеотидов, которая кодирует человеческий белок-антиген (hHER2) эпидермального фактора роста-2, как представлено в SEQ ID NO:2, или человеческий белок HER2ECDTM, как представлено в SEQ ID NO:14, причем синтетическая молекула нуклеиновой кислоты является кодон-оптимизированной для экспрессии высокого уровня в человеческой клетке.
Настоящее изобретение также относится к аденовирусному вакцинному вектору, содержащему аденовирусный геном с делецией в участке Е1 и вставку в участке Е1, причем вставка содержит кассету экспрессии, содержащую: (а) кодон-оптимизированный полинуклеотид, кодирующий человеческий белок HER2 или человеческий белок HER2ECDTM; и (b) промотор, функционально связанный с полинуклеотидом.
Настоящее изобретение также относится к вакцинной плазмиде, содержащей плазмидную часть и часть кассеты экспрессии, причем часть кассеты экспрессии содержит: (а) синтетический полинуклеотид, кодирующий человеческий белок HER2 или человеческий белок HER2ECDTM, причем синтетический полинуклеотид является кодон-оптимизированным для оптимальной экспрессии в человеческой клетке; и (b) промотор, функционально связанный с полинуклеотидом.
Другой аспект настоящего изобретения представляет собой способ защиты млекопитающего от рака или лечения млекопитающего, страдающего HER2-ассоциированным раком, предусматривающий: (а) введение млекопитающему первого вектора, содержащего: i) кодон-оптимизированный полинуклеотид, кодирующий человеческий белок HER2 или человеческий белок HER2ECDTM; и ii) промотор, функционально связанный с полинуклеотидом; (b) ожидание в течение заданного периода времени; и (с) введение млекопитающему второго вектора, содержащего: i) кодон-оптимизированный полинуклеотид, кодирующий человеческий белок HER2 или человеческий белок HER2ECDTM; и ii) промотор, функционально связанный с полинуклеотидом.
Как используется в настоящем описании и прилагаемой формуле изобретения, формы единственного числа включают в себя ссылку на формы множественного числа, если только контекстом четко не определено иное.
Как используется в настоящем описании и прилагаемой формуле изобретения, употребляются нижеследующие определения и аббревиатуры:
Термин “промотор” относится к сайту узнавания на нити ДНК, с которой связывается РНК-полимераза. Промотор формирует инициирующий комплекс с РНК-полимеразой для инициации и управления транскрипционной активностью. Комплекс может быть модифицирован посредством активирующих последовательностей, называемых “энхансерами”, или посредством ингибирующих последовательностей, называемых “молчащими”.
Термин “кассета” относится к нуклеотидной или генной последовательности, которая должна быть экспрессирована из вектора, например нуклеотидная или генная последовательность, кодирующая белок HER2 или белок HER2ECDTM. В общем случае, кассета содержит генную последовательность, встроенную в вектор, который в некоторых вариантах осуществления предоставляет регуляторные последовательности для экспрессии нуклеотидной или генной последовательности. В других вариантах осуществления нуклеотидная или генная последовательность предоставляет регуляторную последовательность для своей экспрессии. В других вариантах осуществления вектор предоставляет некоторые регуляторные последовательности, а нуклеотидная или генная последовательность предоставляет другие регуляторные последовательности. Например, вектор может предоставлять промотор для транскрипции нуклеотидной или генной последовательности, а нуклеотидная или генная последовательность предоставляет последовательность терминации транскрипции. Регуляторные последовательности, которые могут быть предоставлены вектором, включают в себя, без ограничений, энхансеры, последовательности терминации транскрипции, акцепторную и донорную последовательности для сплайсинга, интроны, последовательности для связывания с рибосомой и дополнительные поли(А)-последовательности. Кассета в принципе аналогична кассетной ленте; каждая кассета имеет свою собственную последовательность. Таким образом, при смене кассеты вектор будет экспрессировать другую последовательность. Благодаря сайтам рестрикции на 5' и 3' концах кассета может быть легко вставлена, удалена или заменена другой кассетой.
Термин “вектор” относится к некоторым средствам, при помощи которых фрагменты ДНК могут быть введены в организм хозяина или ткань хозяина. Существуют различные типы векторов, включающие плазмиды, вирусы (включая аденовирусы), бактериофаги и космиды.
Термин “первая генерация”, как используется в отношении аденовирусных векторов, описывает указанные аденовирусные вектора, которые являются репликационно-дефектными. Первая генерация аденовирусных векторов обычно имеет делетированный или инактивированный участок гена Е1 и предпочтительно имеет делетированный или инактивированный участок гена Е3.
Обозначение “pV1J-hHER2.opt” относится к плазмидной конструкции, раскрытой в настоящем описании, содержащей ранний (IE) промотор ЦМВ человека с интроном А, кодон-оптимизированный ген человеческого HER2 полной длины, последовательности полиаденилирования и терминации транскрипции, выделенные из гормона роста крупного рогатого скота, и минимальную основу pUC (см. пример 2).
Обозначение “pV1J-hHER2ECDTM.opt” относится к плазмидной конструкции, раскрытой в настоящем описании, содержащей ранний (IE) промотор ЦМВ человека с интроном А, укороченный кодон-оптимизированный ген человеческого HER2, содержащий внеклеточный и трансмембранный домены гена HER2, последовательности полиаденилирования и терминации транскрипции, выделенные из гормона роста крупного рогатого скота, и минимальную основу pUC (см. пример 2).
Обозначение “pV1J-hHER2.wt” относится к конструкции, как описано выше, за исключением того, что данная конструкция содержит ген человеческого HER2 полной длины дикого типа вместо кодон-оптимизированного гена человеческого HER2.
Обозначение “pV1J-hHER2ECDTM.wt” относится к конструкции, как описано выше, за исключением того, что данная конструкция содержит укороченный ген человеческого HER2 дикого типа, причем указанный укороченный ген содержит последовательность нуклеотидов, которая кодирует внеклеточный и трансмембранный домены белка HER2, вместо кодон-оптимизированного гена человеческого HER2 полной длины.
Обозначения “MRKAd5-hHER2.opt”, “MRKAd5-hHER2ECDTM.opt” и “MRKAd5-hHER2.wt” относятся к трем конструкциям, раскрытым в настоящем описании, которые содержат Ad5 аденовирусный геном с делетированными участками Е1 и Е3. В конструкции “MRKAd5-hHER2.opt” участок Е1 замещен кодон-оптимизированным геном человеческого HER2 полной длины в направлении, параллельном Е1, под управлением промотора ЦМВ человека без интрона А, за которым следует сигнал полиаденилирования гормона роста крупного рогатого скота. Конструкция “MRKAd5-hHER2ECDTM.opt”, по существу, является такой, как описано выше, за исключением того, что участок Е1 генома Ad5 замещен кодон-оптимизированной укороченной версией гена человеческого HER2, причем укороченный ген HER2 содержит последовательность нуклеотидов, которая кодирует внеклеточный и трансмембранный домены рецептора HER2. Конструкция “MRKAd5-hHER2.wt”, по существу, является такой, как описано выше, за исключением того, что участок Е1 генома Ad5 замещен последовательностью человеческого HER2 дикого типа полной длины (см. пример 11).
Термин “эффективное количество” обозначает достаточное количество вакцинной композиции, которую вводят для продуцирования адекватных уровней полипептида для получения иммунного ответа. Специалистам в данной области техники известно, что этот уровень может изменяться.
Термин “лечение” относится как к терапевтическому, так и к профилактическому лечению или профилактике. Нуждающиеся в лечении включают тех, кто уже имеет расстройства, а также тех, кто предрасположен к расстройствам, или тех, у кого расстройство должно быть предупреждено.
“Расстройство” представляет собой любое состояние, которое улучшается при лечении способами или вакцинами и иммуногенными композициями, раскрытыми в настоящем описании. Этот термин включает хронические и острые расстройства или заболевания, включающие такие патологические состояния, при которых млекопитающие имеют предрасположенность к рассматриваемому расстройству. Способы и вакцины настоящего изобретения предназначены для лечения расстройств или состояний, связанных с нарушенной HER2/neu-ассоциированной экспрессией или сигнализацией, включая, без ограничений, рак молочной железы, рак ободочной и прямой кишки, рак желудка, рак яичников и рак легких.
“Консервативная аминокислотная замена” относится к замещению одного аминокислотного остатка другим, химически аналогичным, аминокислотным остатком. Примеры таких консервативных замен представляют собой: замену одного гидрофобного остатка (изолейцина, лейцина, валина или метионина) другим; замену одного полярного остатка другим полярным остатком с таким же зарядом (например, аргинина лизином; глутаминовой кислоты аспарагиновой кислотой).
“hHER2.wt” и “hHER2.opt” относятся к человеческому антигену эпидермального фактора роста-2 и кодон-оптимизированному человеческому антигену эпидермального фактора роста-2 соответственно.
“hHER2ECDTM.wt” и “hHER2ECDTM.opt” относятся к укороченному человеческому антигену эпидермального фактора роста-2 и укороченному кодон-оптимизированному человеческому антигену эпидермального фактора роста-2 соответственно. Укороченные формы HER2, “hHER2ECDTM.wt” и “hHER2ECDTM.opt”, содержат внеклеточный и трансмембранный домены человеческого белка HER2.
Термин “млекопитающее” относится к любому виду млекопитающих, включая человека.
Аббревиатура “Ag” относится к антигену.
Аббревиатуры “Ab” и “mAb” относятся к антителу и моноклональному антителу соответственно.
Аббревиатура “ОРС” относится к открытой рамке считывания гена.
Краткое описание чертежей
На фиг.1 показана нуклеотидная последовательность кодон-оптимизированного полинуклеотида (hHER2.opt, SEQ ID NO:1), которая кодирует человеческий белок HER2. См. пример 1. На панели В показана установленная аминокислотная последовательность человеческого белка HER2 (SEQ ID NO:2).
На фиг.2 показана идентификация иммунодоминантных Т-клеточных эпитопов в человеческом белке HER2 при помощи анализа ELISPOT и внутриклеточного окрашивания (ICS). BALB/c мыши, иммунизированные Ad5-hHER2, были проанализированы на индуцирование человеческого HER2-специфичного клеточного иммунитета. Количество Т-клеток, секретирующих IFN-γ против человеческого HER2, определяли способом ELISPOT в спленоцитах из групп мышей (указанных в первом столбце) с использованием пулов или единичных пептидов. Отображенные данные являются репрезентативными для нескольких независимых экспериментов. Значения выражены в виде количества колоний, формирующих пятно, (SFC)/106 общего количества спленоцитов за вычетом фоновых значений, определенных в отсутствие пептидов (обычно менее чем 10 SFC/106 общего количества спленоцитов). Количества, более чем в три раза превышающие фон, измеренный в контрольных экспериментах без антигенных пептидов, рассматривались как положительные значения и указаны жирным шрифтом. Частоту Т- клеток CD4+ или CD8+, секретирующих IFN-γ, измеряли способом ICS. Отображенные данные являются репрезентативными для нескольких независимых экспериментов. Значения выражены в виде 1000×[(IFN-γ CD3+ и CD4+ или CD8+)/(CD3+ и CD4+ или CD8+)]. Значения, превышающие 1%, рассматривались как положительные и указаны жирным шрифтом. Последовательности, охваченные пулом или одиночным пептидом, используемые в исследованиях, указаны слева. Номера относятся к позиции аминокислотного остатка человеческого белка HER2.
На фиг.3 показана экспрессия in vitro hHER2 после трансфекции в (А) человеческие эмбриональные клетки HEK-293 почки и (В) мышиные миобласты С2С7. Данные выражены в виде среднего геометрического каналов флуоресценции за вычетом сигнала, генерируемого пустой плазмидой pV1JnsA. Для клеток С2С7 данные нормализованы по эффективности трансфекции pEGFP ДНК.
На фиг.4 показан иммунный ответ на человеческий HER2 у мышей BALB/c. На панели (А) показано, что кодон-оптимизированный HER2 давал значительно улучшенные значения ELISPOT в сравнении с HER2 дикого типа. Показаны результаты иммунизации четырех групп, каждая из которых содержит двух мышей, плазмидой pV1J-hHER2.wt или pV1J-hHER2.opt (50 мкг/дозу, введенной способом электроинъекции в четырехглавую мышцу). Спустя две недели после последней инъекции определяли частоту IFN-γ-секретирующих Т-клеток в мышиных спленоцитах с помощью анализа IFN-γ ELISPOT, используя пептиды hNeu15.3 (аа 63-71, включая эпитоп CD8+), hNeu301 (аа 1202-1214, включая эпитоп CD8+) и hNeu42 (аа 165-179, включая эпитоп CD4+). Показаны результаты из 2,5×105 и 5×105 спленоцитов с двумя репликами каждого протестированного количества. Средние значения вычислены путем вычитания фонового уровня, определенного в отсутствие пептидов (обычно менее чем 10 SFC/106 общего количества спленоцитов). Результаты выражены в виде количества SFC/106 общего количества спленоцитов. На панели (В) показано, что pV1J-hHER2.opt вырабатывает существенно улучшенный IgG1 и IgG2а гуморальный ответ в сравнении с pV1J-hHER2.wt. Образцы сыворотки собирали на 6 неделе (за день перед первой иммунизацией, предварительное взятие крови) и на 14 неделе (две недели спустя после последней инъекции) из групп, состоящих из 4 мышей, иммунизированных pV1J-hHER2.wt или pV1J-hHER2.opt плазмидной ДНК. Титры анти-hHER2 антител в объединенных сыворотках из каждой группы мышей измеряли при помощи ELISA, используя димерный внеклеточный домен hHER2 (HER2-ECD) в качестве целевого антигена. Для детектирования связанных мышиных антител использовали АР-конъюгированные козьи анти-мышиные IgG1 или IgG2а.
На фиг.5 показано сравнение р185-специфичного Т-клеточного ответа, вырабатываемого у мышей иммунизацией pV1J-HER2 и Ad5-HER2. Мыши BALB/c дикого типа и трансгенные мыши BALB/c, сверхэкспрессирующие крысиный HER2 (обозначенный как NeuT, см. Lucchini et al., Cancer Lett 64(3):203-9 (1992)), были иммунизированы в возрасте 6 и 9 недель либо pV1J-hHER2.wt ДНК (50 мкг/дозу, инъецированную в четырехглавую мышцу) с последующей электростимуляцией, либо Ad5-hHER2.wt. В возрасте 12 недель определяли количество IFN-γ-секретирующих анти- человеческих клеток с помощью анализа ELISPOT из пулов мышей, используя указанные пептиды. Отображенные данные являются репрезентативными для нескольких независимых экспериментов. Значения выражены так же, как на фиг.1.
На панели А фиг.6 показана нуклеотидная последовательность кодон-оптимизированного полинуклеотида (hHER2ECDTM.opt, SEQ ID NO:9), которая кодирует укороченный человеческий белок HER2, причем указанный белок содержит внеклеточный и трансмембранный домены белка HER2. На панели В показан второй полинуклеотид, который кодирует внеклеточный и трансмембранный домены белка HER2, причем второй полинуклеотид содержит нуклеотидную последовательность “дикого типа”, которая не была кодон-оптимизирована (hHER2ECDTM.wt, SEQ ID NO:10).
На фиг.7 показан результат анализа клеточно-опосредованного ответа, индуцированного у макаки-резус, иммунизированной смесью из трех плазмид, экспрессирующих человеческие антигены HER2, CEA и EpCAM, причем указанные плазмиды содержат нуклеотидные последовательности, которые являются кодон-оптимизированными для экспрессии высокого уровня в человеческих клетках. Те же самые животные затем были иммунизированы смесью трех Ad5 векторов, экспрессирующих последовательность дикого типа каждого из трех антигенов. Клеточно-опосредованный иммунный ответ, направленный против высокогомологичного (98,2% сходство последовательностей) белка HER2 макаки-резус, измеряли при помощи IFN-γ ELISPOT ежемесячно в течение одного года. Значения выражены в виде SFC/106PBMC за вычетом фоновых значений, определенных в отсутствие пептидов. Значения, которые существенно отличались (р<0,05) от фона, как измеренные в контрольных экспериментах без антигенных пептидов, и превышали произвольным образом выбранное пороговое значение 55 SFC/106PBMC, указаны жирным шрифтом.
На фиг.8 показано сравнение клеточно-опосредованного иммунного ответа, выработанного у мышей посредством иммунизации pV1J-hHER2.opt и pV1J-hHER2ECDTM.opt. Значения относятся к частоте IFN-γ-секретирующих клеток селезенки, как измерено при помощи ELISPOT. Отображенные данные получены от трех животных и являются репрезентативными для нескольких независимых экспериментов. Значения выражены в виде SFC/106 общего количества клеток селезенки за вычетом фоновых значений, определенных в отсутствие пептидов (обычно менее чем 5 SFC/106 клеток селезенки). Значения, которые существенно отличались (р<0,05) от фона, как измеренные в контрольных экспериментах без антигенных пептидов, и превышали произвольным образом выбранное пороговое значение 25 SFC/106 клеток селезенки, указаны жирным шрифтом.
Подробное описание изобретения
Человеческий эпидермальный фактор роста-2 (hHER2) обычно ассоциирован с несколькими различными типами опухолей, включая карциномы молочной железы, яичников, желудка и толстой кишки. Настоящее изобретение относится к композициям и способам для выработки или усиления иммунитета к белковым продуктам, экспрессируемым геном hHER2, причем нарушенная экспрессия hHER2 ассоциирована с карциномой или ее развитием. Ассоциация нарушенной экспрессии hHER2 с карциномой не требует того, чтобы белок hHER2 экспрессировался в ткани опухоли постоянно во время ее развития, поскольку аномальная экспрессия hHER2 может присутствовать при инициации опухоли и не детектироваться позже во время развития опухоли или наоборот.
С этой целью предоставлены синтетические молекулы ДНК, кодирующие человеческий белок HER2 полной длины или укороченный человеческий белок HER2, называемый в настоящем описании как HER2ECDTM. Указанный укороченный HER2 содержит внеклеточный и трансмембранный домены человеческого белка HER2. Кодоны синтетических молекул ДНК разработаны таким образом, чтобы использовать кодоны, предпочтительные для предполагаемой клетки хозяина, которая в предпочтительных вариантах осуществления является человеческой клеткой. Синтетические молекулы могут быть использованы для разработки вакцин на основе плазмиды или рекомбинантного аденовируса, которые обеспечивают эффективную иммунопрофилактику против HER2-ассоциированного рака путем нейтрализации антитела и клеточно-опосредованного иммунитета. Синтетические молекулы могут быть использованы в виде иммуногенной композиции. Настоящее изобретение предоставляет полинуклеотиды, которые при прямом введении in vivo позвоночным, включая млекопитающих, таких как приматы и люди, индуцируют у животного экспрессию кодируемых белков.
Была опубликована нуклеотидная последовательность человеческого HER2 дикого типа (Coussens et al., Science 230:1132-39 (1985); King et al., Science 229:974-76 (1985)). Настоящее изобретение предоставляет синтетические молекулы ДНК, кодирующие человеческий белок HER2 полной длины или укороченный человеческий белок HER2, HER2ECDTM, содержащий внеклеточный и трансмембранный домены hHER2. Синтетические молекулы настоящего изобретения содержат последовательность нуклеотидов, причем, по меньшей мере, один из нуклеотидов изменен таким образом, чтобы использовать кодоны, предпочтительные для человеческой клетки, таким образом предоставляя возможность для экспрессии высокого уровня hHER2 или hHER2ECDTM в человеческой клетке хозяина. Синтетические молекулы могут быть использованы в качестве источника белка hHER2 или hHER2ECDTM, который может быть использован в противораковой вакцине для обеспечения эффективной иммунопрофилактики против hHER2-ассоциированных карцином путем нейтрализации антительного и клеточно-опосредованного иммунитета. В качестве альтернативы, синтетические молекулы могут быть использованы в качестве основы ДНК вакцины или аденовирусной вакцины.
“Триплет” кодон из четырех возможных нуклеотидных оснований может существовать в более чем 60 различных формах. Поскольку эти кодоны обеспечивают сообщение только для 20 различных аминокислот (а также инициацию и терминацию транскрипции), некоторые аминокислоты могут кодироваться более чем одним кодоном - явление, известное как вырожденность кодона. По неполностью понятым причинам альтернативные кодоны не представлены однообразно в эндогенной ДНК различных типов клеток. Фактически, оказывается, существует изменчивая природная иерархия или “предпочтение” для некоторых кодонов в специфических типах клеток. В качестве одного из примеров, аминокислота лейцин определяется любым из шести ДНК кодонов, включающих CTA, CTC, CTG, CTT, TTA и TTG. Исчерпывающий анализ частот геномных кодонов для микроорганизмов выявил, что эндогенная ДНК E.coli чаще всего содержит CTG лейцин-специфичный кодон, в то время как ДНК дрожжей и миксомицетов чаще всего включает TTA лейцин-специфичный кодон. С точки зрения этой иерархии обычно предполагают, что вероятность получения высоких уровней экспрессии полипептидов, богатых лейцином, при помощи хозяина E.coli до некоторой степени будет зависеть от частоты использования кодона. Например, существует вероятность того, что ген, богатый кодонами ТТА, будет недостаточно экспрессируемым в E.coli, тогда как ген, богатый CTG, возможно будет высокоэкспрессируемым в этом хозяине. Аналогично, предпочтительным кодоном для экспрессии полипептида, богатого лейцином, в дрожжевых клетках хозяина может быть ТТА.
Значение явления предпочтения кодона обнаружено при помощи методов рекомбинантной ДНК, и это явление может служить объяснением многих ранних неудач в достижении высоких уровней экспрессии экзогенных генов в успешно трансформированных организмах хозяев - менее “предпочтительный” кодон может быть неоднократно представлен в интересующем гене, и механизм клетки хозяина, реализующий экспрессию, может не работать также эффективно. Это явление предполагает, что синтетические гены, которые разработаны таким образом, что они включают кодоны, предпочтительные для предполагаемой клетки хозяина, обеспечивают оптимальную форму чужеродного генетического материала для применения методов рекомбинантных ДНК. Таким образом, одним из аспектов настоящего изобретения является ген человеческого HER2, который является кодон-оптимизированным для экспрессии высокого уровня в человеческой клетке. В предпочтительном варианте осуществления настоящего изобретения было обнаружено, что использование альтернативных кодонов, кодирующих такую же белковую последовательность, может устранить ограничения на экспрессию экзогенного белка hHER2 в человеческих клетках. Другим аспектом настоящего изобретения является укороченный ген человеческого HER2, hHER2ECDTM, который является кодон-оптимизированным для экспрессии высокого уровня в человеческой клетке хозяина, причем указанный укороченный ген HER2 содержит нуклеотидные последовательности, которые кодируют внеклеточный и трансмембранный домены человеческого HER2.
Согласно настоящему изобретению последовательность гена человеческого HER2 и последовательность гена человеческого HER2ECDTM были преобразованы в полинуклеотидные последовательности, имеющие идентичные транслируемые последовательности в сравнении с эквивалентами дикого типа, но с альтернативным применением кодона, как описано у Lathe “Synthetic Oligonucleotide Probes Deduced from Amino Acid Sequence Data: Theoretical and Practical Considerations” J. Molec. Biol. 183:1-12 (1985), которая включена в настоящее описание во всей своей полноте в качестве ссылки. Методология в общем случае состоит из идентификации кодонов в последовательности дикого типа, которая обычно не ассоциируется с высокоэкспрессируемыми человеческими генами, и замены их оптимальными кодонами для высокой экспрессии в человеческих клетках. Указанные оптимальные кодоны называются в настоящем описании кодонами “предпочтительными для человека”. Затем новую генную последовательность тестируют на нежелательные последовательности, генерируемые заменами кодонов (например, последовательности “АТТТА”, случайное образование сайтов узнавания сплайсинга интрона, нежелательные сайты рестрикционных ферментов, высокое содержание GC и т.д.). Нежелательные последовательности удаляют путем замены существующих кодонов другими кодонами, кодирующими такую же аминокислоту. Затем синтетические сегменты гена тестируют на улучшенную экспрессию.
Способы, описанные выше, были использованы для создания синтетических генных последовательностей для человеческого HER2 и человеческого HER2ECDTM, что дает в результате ген полной длины и укороченный ген, содержащие кодоны, оптимизированные для экспрессии высокого уровня в человеческих клетках. Хотя вышеописанная процедура в общем предоставляет методологию авторов для разработки кодон-оптимизированных генов для использования в противораковых вакцинах, специалистам в данной области техники ясно, что аналогичная эффективность вакцины и увеличенная экспрессия генов могут быть достигнуты путем небольших изменений в процедуре или путем небольших изменений в последовательности. Специалистам в данной области техники также будет ясно, что дополнительные молекулы ДНК могут быть сконструированы для того, чтобы обеспечить высокие уровни экспрессии hHER2 или hHER2ECDTM в человеческих клетках, причем только часть кодонов молекул ДНК являются кодон-оптимизированными. Молекулы нуклеиновых кислот настоящего изобретения, по существу, не имеют других нуклеиновых кислот.
Следовательно, настоящее изобретение относится к синтетическому полинуклеотиду, содержащему последовательность нуклеотидов, кодирующих человеческий белок HER2, например человеческий белок HER2, приведенный в SEQ ID NO:2, или биологически активный фрагмент или мутированную форму человеческого белка HER2, причем полинуклеотидная последовательность содержит кодоны, оптимизированные для экспрессии в человеке-хозяине. Указанные мутированные формы белка hHER2 включают, без ограничений: консервативные аминокислотные замены, амино-терминальные отсечения, карбокси-терминальные отсечения, делеции или вставки. Любой такой биологически активный фрагмент и/или мутант будет кодировать либо белок, либо фрагмент белка, который, по меньшей мере, по существу, имитирует иммунологические свойства белка hHER2, как представлено в SEQ ID NO:2. Синтетические полинуклеотиды настоящего изобретения кодируют молекулы мРНК, что ведет к экспрессии функционального человеческого белка таким образом, что это является пригодным для разработки терапевтической или профилактической противораковой вакцины.
Предпочтительный полинуклеотид настоящего изобретения представляет собой полинуклеотид, содержащий последовательность нуклеотидов, кодирующую укороченный человеческий белок HER2ECDTM (SEQ ID NO:14), причем полинуклеотидная последовательность содержит кодоны, оптимизированные для экспрессии в человеке-хозяине. Особенно предпочтительный полинуклеотид настоящего изобретения содержит последовательность нуклеотидов, как приведено в SEQ ID NO:9.
Настоящее изобретение также относится к синтетической молекуле нуклеиновой кислоты (полинуклеотид), содержащей последовательность нуклеотидов, которая кодирует мРНК, что ведет к экспрессии человеческого белка HER2, например человеческого белка HER2 полной длины, как приведено в SEQ ID NO:2, или укороченного белка HER2ECDTM, например последовательность HER2ECDTM, как приведено в SEQ ID NO:14. Синтетические молекулы нуклеиновых кислот настоящего изобретения являются кодон-оптимизированными для экспрессии высокого уровня в человеческой клетке хозяина.
Также в объем настоящего изобретения включены кодон-оптимизированные полинуклеотиды, содержащие последовательность нуклеотидов, которая кодирует вариантный полипептид HER2, имеющий, по меньшей мере, 90% идентичность с аминокислотной последовательностью SEQ ID NO:2, которая может включать до Na аминокислотных замен по всей длине SEQ ID NO:2, причем Na представляет собой максимальное количество аминокислотных замен и вычисляется по формуле
Na=Xa-(XaY),
где Ха представляет собой общее количество аминокислот в SEQ ID NO:2, а Y имеет значение 0,90, причем любое произведение Ха и Y, которое не является целым, округляют до ближайшего целого числа перед вычитанием этого произведения из Ха. Аналогично, настоящее изобретение также рассматривает кодон-оптимизированные нуклеотидные последовательности, кодирующие варианты полипептида HER2CDTM, как приведено в SEQ ID NO:14.
Настоящее изобретение также относится к рекомбинантным векторам и рекомбинантым клеткам-хозяевам, как прокариотическим, так и эукариотическим, которые содержат молекулы нуклеиновых кислот, раскрытые в настоящем описании. Синтетические молекулы ДНК, ассоциированные вектора и хозяева настоящего изобретения являются пригодными для разработки противораковой вакцины.
Предпочтительная молекула ДНК настоящего изобретения содержит нуклеотидную последовательность, раскрытую в настоящем изобретении как SEQ ID NO:1 (показанную на фиг.1), которая кодирует человеческий белок HER2, показанный на фиг.2 и приведенный в виде SEQ ID NO:2. Нуклеотидная последовательность, приведенная в SEQ ID NO:1, была кодон-оптимизирована для оптимальной экспрессии в человеческих клетках. Во избежание трудностей, связанных с ПЦР амплификацией, в этом варианте осуществления настоящего изобретения применена конструкция с менее жесткой оптимизацией для последовательности hHER2 между позициями 3601 и 3805, в которой уменьшено содержание GС при сохранении такой же аминокислотной композиции. См. пример 5.
Другая предпочтительная молекула ДНК настоящего изобретения содержит нуклеотидную последовательность, раскрытую в настоящем описании в виде SEQ ID NO:9 (показанную на фиг.6А), которая кодирует человеческий белок HER2ECDTM, приведенный в виде SEQ ID NO:14. Нуклеотидная последовательность, приведенная в SEQ ID NO:9, была кодон-оптимизирована для оптимальной экспрессии в человеческих клетках.
Специалист в данной области техники понимает, что могут быть разработаны другие последовательности HER2, которые будут кодон-оптимизированными для экспрессии высокого уровня в человеческой клетке, при условии, что один или несколько кодонов заменены кодонами, предпочтительными для человека. Предпочтительно, чтобы, по меньшей мере, 80% кодонов, содержащих синтетические нуклеотидные последовательности HER2 настоящего изобретения, являлись кодонами, предпочтительными для человека. Более предпочтительно, чтобы, по меньшей мере, примерно 85% кодонов являлись предпочтительными для человека, и даже более предпочтительно, чтобы, по меньшей мере, примерно 90% кодонов являлись предпочтительными для человека.
Настоящее изобретение также включает биологически активные фрагменты или мутанты SEQ ID NO:1, которые кодируют мРНК, что ведет к экспрессии человеческих белков HER2. Любой такой биологически активный фрагмент и/или мутант будет кодировать либо белок, либо фрагмент белка, который, по меньшей мере, в значительной степени имитирует фармакологические свойства белка hHER2, включая, но не ограничивая, белок hHER2, как приведено в SEQ ID NO:2. Любой такой полинуклеотид включает, но, безусловно, без ограничений: замены нуклеотидов, делеции, вставки, амино-терминальные отсечения и карбокси-терминальные отсечения. Мутанты настоящего изобретения кодируют молекулы мРНК, что ведет к экспрессии функционального белка hHER2 в эукариотической клетке таким образом, что это является пригодным для разработки терапевтической или профилактической противораковой вакцины.
Настоящее изобретение также относится к синтетическим кодон-оптимизированным молекулам ДНК, которые кодируют белок hHER2 или белок hHER2ECDTM, причем нуклеотидная последовательность синтетической ДНК значительно отличается от нуклеотидной последовательности SEQ ID NO:1 или SEQ ID NO:9, но все еще кодирует белок hHER2, как приведено в SEQ ID NO:2, или белок hHER2ECDTM, как приведено в SEQ ID NO:14. Такие синтетические ДНК находятся в пределах объема настоящего изобретения. Следовательно, настоящее изобретение раскрывает вырожденность кодонов, которые в результате могут привести к множеству молекул ДНК, экспрессирующих идентичный белок. В объем настоящего изобретения также включены мутации в ДНК последовательности, которые, по существу, не меняют основные физические свойства экспрессированного белка. Например, замена валина лейцином, аргинина лизином или аспарагина глутамином может не вызывать изменения функциональности полипептида.
Известно, что ДНК последовательности, кодирующие пептид, могут быть изменены таким образом, чтобы кодировать пептид, который имеет свойства, отличные от свойств природного пептида. Способы изменения ДНК последовательностей включают, но не являются ограничивающими, сайт-направленный мутагенез. Примеры измененных свойств включают, без ограничений, изменения аффинности фермента к субстрату или рецептора к лиганду.
Настоящее изобретение также относится к слитым конструкциям hHER2 и hHER2ECDTM, включая, без ограничений, слитые конструкции, которые экспрессируют часть человеческого белка HER2, соединенного с различными маркерами, включая, но никоим образом не ограничивая, GFP (зеленый флуоресцентный белок), MYC эпитоп, GST и Fc. Любая такая слитая конструкция может быть экспрессирована в интересующей клеточной линии и использована для скрининга модуляторов человеческого белка HER2, раскрытого в настоящем описании. Также рассмотрены слитые конструкции, которые сконструированы для усиления иммунного ответа на человеческий HER2, включая, без ограничений: DOM, hsp70 и LTB.
Настоящее изобретение также относится к рекомбинантным векторам, которые содержат молекулы синтетических нуклеиновых кислот, раскрытых в настоящем описании. Такие вектора могут содержаться в ДНК или РНК. Для большинства целей клонирования предпочтительны вектора ДНК. Обычные вектора включают плазмиды, модифицированные вирусы, бакуловирусы, бактериофаги, космиды, синтетические хромосомы дрожжей и другие формы эписомной или интегрированной ДНК, которая может кодировать белок hHER2 или белок hHER2ECDTM. Специалисты в данной области техники легко определят подходящий вектор для переноса или другого применения конкретного гена.
Экспрессионный вектор, содержащий кодон-оптимизированную ДНК, кодирующую белок hHER2, может быть использован для экспрессии высокого уровня hHER2 в рекомбинантной клетке хозяина. Кроме того, экспрессионный вектор, содержащий кодон-оптимизированную ДНК, кодирующую белок hHER2ECDTM, может быть использован для экспрессии высокого уровня hHER2ECDTM в рекомбинантной клетке хозяина. Экспрессионные вектора могут включать, без ограничений, вектора клонирования, модифицированные вектора клонирования, в частности разработанные плазмиды или вирусы. Также, если это необходимо, для экспрессии рекомбинантного hHER2 или hHER2ECDTM в бактериальных клетках может быть использовано множество бактериальных векторов экспрессии. Кроме того, множество векторов экспрессии клеток грибов могут быть использованы для экспрессии рекомбинантного hHER2, или hHER2ECDTM в клетках грибов. Помимо этого, множество векторов экспрессии клеток насекомых может быть использовано для экспрессии рекомбинантного белка в клетках насекомых.
Настоящее изобретение также относится к клеткам хозяина, трансформированным или трансфицированным векторами, содержащими молекулы нуклеиновых кислот настоящего изобретения. Рекомбинантные клетки хозяина могут быть прокариотическими или эукариотическими, включая, без ограничений, бактерии, такие как E.coli, клетки грибов, таких как дрожжи, клетки млекопитающих, включая, но не ограничивая, клеточные линии, полученные из клеток крупного рогатого скота, свиньи, обезьян и грызунов; клетки насекомых, включая, но не ограничивая, Drosophila, и клеточные линии, производные шелкопряда тутового. Такие рекомбинантные клетки хозяина могут быть культивированы в подходящих условиях для продуцирования hHER2 или hHER2ECDTM, или биологически эквивалентной формы. В предпочтительном варианте настоящего изобретения клеткой хозяина является человеческая. Как определено в настоящем описании, термин “клетка хозяина” не включает клетку хозяина в теле трансгенного человека, трансгенного человеческого плода или трансгенного человеческого эмбриона.
Как указано выше, экспрессионный вектор, содержащий ДНК, кодирующую белок hHER2 или белок hHER2ECDTM, может быть использован для экспрессии hHER2 или hHER2ECDTM в рекомбинантной клетке хозяина. Следовательно, другим аспектом настоящего изобретения является способ экспрессии человеческого белка HER2 или человеческого белка HER2ECDTM в рекомбинантной клетке хозяина, предусматривающий: (а) введение вектора, содержащего кодон-оптимизированную нуклеиновую кислоту, которая кодирует человеческий белок HER2 или человеческий белок HER2ECDTM, в подходящую человеческую клетку хозяина; и (b) культивирование клетки-хозяина в условиях, которые позволяют экспрессию указанного человеческого белка HER2 или указанного человеческого белка HER2ECDTM.
Предпочтительный вариант осуществления этого аспекта настоящего изобретения предоставляет способ экспрессии человеческого белка HER2 в рекомбинантной клетке хозяина, предусматривающий: (а) введение вектора, содержащего нуклеиновую кислоту, как приведено в SEQ ID NO:1, в подходящую человеческую клетку хозяина; и (b) культивирование клетки хозяина в условиях, которые позволяют экспрессию указанного человеческого белка HER2.
Другой предпочтительный вариант осуществления этого аспекта настоящего изобретения предоставляет способ экспрессии человеческого белка HER2ECDTM в рекомбинантной клетке хозяина, предусматривающий: (а) введение вектора, содержащего нуклеиновую кислоту, как приведено в SEQ ID NO:9, в подходящую человеческую клетку хозяина; и (b) культивирование клетки хозяина в условиях, которые позволяют экспрессию указанного человеческого белка HER2ECDTM.
После экспрессии hHER2 или hHER2ECDTM в клетке хозяина белок hHER2 или hHER2ECDTM может быть выделен в виде белка в активной форме. Некоторые процедуры очистки белка являются доступными и подходящими для использования. Рекомбинантный белок hHER2 или белок hHER2ECDTM может быть очищен из клеточных лизатов и экстрактов при помощи различных комбинаций или отдельного применения высаливания, ионообменной хроматографии, гель-фильтрации, адсорбционной хроматографии на колонке с гидроксилапатитом и гидрофобной хроматографии. Кроме того, рекомбинантный белок может быть отделен от других клеточных белков при помощи иммуноаффинной колонки, изготовленной с моноклональными или поликлональными антителами, специфичными для белка hHER2 полной длины, или полипептидных фрагментов белка hHER2.
Нуклеиновые кислоты настоящего изобретения могут быть собраны в кассету экспрессии, которая содержит последовательности, разработанные для обеспечения эффективной экспрессии белка в человеческой клетке. В одном из вариантов осуществления настоящего изобретения кассета содержит кодон-оптимизированный ген hHER2 полной длины с относящимися к нему последовательностями управления транскрипцией и трансляцией, функционально связанными с ним, такими как промотор и последовательности терминации. Во втором варианте осуществления настоящего изобретения кассета содержит укороченный ген HER2, HER2ECDTM, который кодирует внеклеточный и трансмембранный домены человеческого белка HER2. В предпочтительных вариантах осуществления промотор представляет собой промотор цитомегаловируса без интронной А последовательности (ЦМВ), хотя специалистам в данной области техники известно, что может быть использовано любое количество других известных промоторов, таких как промотор сильного иммуноглобулина или другой промотор эукариотического гена. Предпочтительный терминатор транскрипции представляет собой терминатор гормона роста крупного рогатого скота, хотя могут быть использованы другие известные терминаторы транскрипции. Комбинация с терминатором ЦМВ-BGH является особенно предпочтительной.
Согласно настоящему изобретению кассету экспрессии hHER2.opt или hHER2ECDTM.opt встраивают в вектор. Вектор предпочтительно представляет собой аденовирусный вектор, хотя могут быть использованы линейная ДНК, связанная с промотором, или другие вектора, такие как аденоассоциированные вирусные или модифицированные вакцинные вирусные, ретровирусные и лентивирусные вектора.
Если выбранный вектор является аденовирусным, то предпочтительно, чтобы вектор представлял собой так называемый аденовирусный вектор первой генерации. Такие аденовирусные вектора характеризуются тем, что они имеют участок нефункционального гена Е1 и предпочтительно делетированный участок аденовирусного гена Е1. В некоторых вариантах осуществления кассету экспрессии вставляют в позицию, в которой обычно расположен аденовирусный ген Е1. Кроме того, такие вектора необязательно имеют нефункциональный или делетированный участок Е3. Предпочтительно, чтобы в используемом аденовирусном геноме были делетированы оба участка, как Е1, так и Е3 (∆Е1∆Е3). Аденовирусы могут быть размножены в известных клеточных линиях, которые экспрессируют вирусный ген Е1, таких как клетки 293, или клетки PERC.6, или клеточные линии, производные клетки 293 или PERC.6, которые временно или постоянно трансформированы для экспрессии дополнительного белка. Например, при использовании конструкций, которые имеют управляемую экспрессию гена, таких как регулируемая тетрациклином промоторная система, клеточная линия может экспрессировать компоненты, включенные в регуляторную систему. Одним из примеров такой клеточной линии является T-Rex-293; в данной области техники известны и другие.
Для удобства манипулирования аденовирусным вектором аденовирусы могут находиться в виде челночной плазмиды. Настоящее изобретение также относится к вектору челночной плазмиды, который содержит плазмидную часть и часть аденовируса, причем часть аденовируса содержит аденовирусный геном, который имеет делетированную Е1 и необязательную делецию Е3, и имеет вставленную кассету экспрессии, содержащую кодон-оптимизированный человеческий HER2 или кодон-оптимизированный hHER2ECDTM. В предпочтительных вариантах осуществления существуют сайты рестрикции, фланкирующие аденовирусную часть плазмиды таким образом, что аденовирусный вектор может быть легко удален. Челночная плазмида может быть реплицирована в прокариотических клетках или эукариотических клетках.
В предпочтительном варианте осуществления настоящего изобретения кассету экспрессии встраивают в аденовирусную плазмиду pMRKAd5-HV0 (см. Eminit et al., WO 02/22080, которая включена в настоящее описание во всей своей полноте в качестве ссылки). Эта плазмида содержит аденовирусный геном Ad5 с делетированными участками Е1 и Е3. Разработка плазмиды pMRKAd5-HV0 улучшила все ранние аденовектора путем расширения 5' цис-действующего участка упаковки далее в ген Е1 для включения элементов, которые, как было обнаружено, являются важными при оптимизации вирусной упаковки, приводя в результате к усилению амплификации вируса. Преимущественно, такой усиленный аденовирусный вектор способен поддерживать генетическую стабильность после многократно пассированного размножения.
Стандартные технологии молекулярной биологии получения и очистки конструкций ДНК дают возможность получения аденовирусов, челночных плазмид и ДНК иммуногенов настоящего изобретения.
Согласно настоящему изобретению определили, что молекулы синтетической кДНК, раскрытые в настоящем описании (например, SEQ ID NO:1 и SEQ ID NO:9), которые являются кодон-оптимизированными для экспрессии высокого уровня в человеческой клетке, экспрессируются с большей эффективностью, чем соответствующая последовательность дикого типа. Кроме того, в настоящем описании показано, что hHER2.opt является более иммуногенным, чем hHER2, и более эффективным при выработке как клеточного, так и гуморального иммунного ответов.
Следовательно, вектора, описанные выше, могут быть использованы в иммуногенных композициях и вакцинах для предупреждения развития аденокарцином, ассоциированных с нарушенной экспрессией HER2, и/или для лечения существующих видов рака. Вектора настоящего изобретения предоставляют возможность для разработки и коммерциализации вакцин путем преодоления трудностей, связанных с получением высоких уровней экспрессии экзогенного HER2 в успешно трансформированных организмах хозяев. При этом одним из аспектов настоящего изобретения является способ профилактики или лечения HER2-ассоциированного рака, предусматривающий введение млекопитающему вакцинного вектора, содержащего синтетическую кодон-оптимизированную молекулу нуклеиновой кислоты, причем синтетическая кодон-оптимизированная молекула нуклеиновой кислоты содержит последовательность нуклеотидов, которая кодирует человеческий белок HER2, как приведено в SEQ ID NO:2, или человеческий белок HER2ECDTM, как приведено в SEQ ID NO:14.
Согласно способу, описанному выше, вакцинный вектор может быть введен любому млекопитающему с целью терапии или профилактики рака. В предпочтительном варианте осуществления настоящего изобретения млекопитающим является человек.
Помимо этого, специалисты в данной области техники могут выбрать любой тип вектора для использования в описанном способе лечения и профилактики. Предпочтительно вектор представляет собой аденовирусный вектор или плазмидный вектор. В предпочтительном варианте осуществления настоящего изобретения вектор представляет собой аденовирусный вектор, содержащий аденовирусный геном с делецией в участке Е1 аденовируса и вставкой в участке Е1 аденовируса, причем вставка содержит кассету экспрессии, содержащую: (а) синтетический кодон-оптимизированный полинуклеотид, кодирующий человеческий белок HER2 или человеческий белок HER2ECDTM; и (b) промотор, функционально связанный с полинуклеотидом.
Настоящее изобретение также относится к аденовирусному вакцинному вектору, содержащему аденовирусный геном с делецией в участке Е1 и вставкой в участке Е1, причем вставка содержит кассету экспрессии, содержащую: (а) синтетический кодон-оптимизированный полинуклеотид, кодирующий человеческий белок HER2 или человеческий белок HER2ECDTM; и (b) промотор, функционально связанный с полинуклеотидом.
В предпочтительном варианте осуществления этого аспекта настоящего изобретения аденовирусный вектор представляет собой Ad5 вектор.
В других предпочтительных вариантах осуществления настоящего изобретения вектор представляет собой Ad6 вектор или Ad24 вектор.
В другом аспекте настоящее изобретение относится к вакцинной плазмиде, содержащей плазмидную часть и часть кассеты экспрессии, причем часть кассеты экспрессии содержит: (а) синтетический кодон-оптимизированный полинуклеотид, кодирующий человеческий белок HER2; и (b) промотор, функционально связанный с полинуклеотидом.
Настоящее изобретение также относится к вакцинной плазмиде, содержащей плазмидную часть и часть кассеты экспрессии, причем часть кассеты экспрессии содержит: (а) синтетический кодон-оптимизированный полинуклеотид, кодирующий человеческий белок HER2ECDTM; и (b) промотор, функционально связанный с полинуклеотидом.
В некоторых вариантах осуществления настоящего изобретения рекомбинантные аденовирусные вакцины, раскрытые в настоящем описании, используются в различных комбинациях, первичная/бустерная, с полинуклеотидной вакциной на основе плазмиды для того, чтобы индуцировать усиленный иммунный ответ. В этом случае два вектора вводят в “первичном и бустерном” режиме. Например, вводят первый тип вектора, затем спустя заданный период времени, например 2 недели, 1 месяц, 2 месяца, шесть месяцев или другой подходящий интервал, вводят второй тип вектора. Предпочтительно вектора несут кассету экспрессии, кодирующую один и тот же полинуклеотид или комбинацию полинуклеотидов. В варианте осуществления, в котором также используется плазмидная ДНК, предпочтительно, чтобы вектор содержал один или несколько промоторов, узнаваемых клетками млекопитающих или насекомых. В предпочтительном варианте осуществления плазмида может содержать сильный промотор, такой как, без ограничений, ЦМВ промотор. Синтетический ген человеческого HER2, ген HER2ECDTM, или другой ген, предназначенный для экспрессии, может быть связан с таким промотором. Примером такой плазмиды может быть плазмида экспрессии V1Jns у млекопитающих, как описано (J. Shiver et al. в DNA Vaccines, M.Liu et al., eds. N.Y. Acad. Sci., N.Y., 772:198-208 (1996), которые включены в настоящее описание во всей своей полноте в качестве ссылки).
Как указано выше, аденовирусная векторная вакцина и плазмидная вакцина могут быть введены позвоночному животному как части одного терапевтического режима для индуцирования иммунного ответа. При этом настоящее изобретение относится к способу защиты млекопитающего от рака, предусматривающему: (а) введение млекопитающему первого вектора, содержащего: i) синтетический кодон-оптимизированный полинуклеотид, кодирующий человеческий белок HER2 или человеческий белок HER2ECDTM; и ii) промотор, функционально связанный с полинуклеотидом; (b) ожидание в течение заданного периода времени; и (с) введение млекопитающему второго вектора, содержащего: i) синтетический кодон-оптимизированный полинуклеотид, кодирующий человеческий белок HER2 или человеческий белок HER2ECDTM; и ii) промотор, функционально связанный с полинуклеотидом.
В одном из вариантов осуществления способа защиты, описанного выше, первый вектор представляет собой плазмиду, а второй вектор представляет собой аденовирусный вектор. В альтернативном варианте осуществления первый вектор представляет собой аденовирусный вектор, а второй вектор представляет собой плазмиду.
Настоящее изобретение также относится к способу лечения млекопитающего, страдающего HER2-ассоциированным раком, предусматривающему: (а) введение млекопитающему первого вектора, содержащего: i) синтетический кодон-оптимизированный полинуклеотид, кодирующий человеческий белок HER2 или человеческий белок HER2ECDTM; и ii) промотор, функционально связанный с полинуклеотидом; (b) ожидание в течение заданного периода времени; и (с) введение млекопитающему второго вектора, содержащего: i) синтетический кодон-оптимизированный полинуклеотид, кодирующий человеческий белок HER2 или человеческий белок HER2ECDTM; и ii) промотор, функционально связанный с полинуклеотидом.
В одном из вариантов осуществления способа лечения, описанного выше, первый вектор представляет собой плазмиду, а второй вектор представляет собой аденовирусный вектор. В альтернативном варианте осуществления первый вектор представляет собой аденовирусный вектор, а второй вектор представляет собой плазмиду.
Количество экспрессируемой ДНК или транскрибируемой РНК, предназначенной для введения в вакцинного реципиента, будет отчасти зависеть от силы используемых промоторов и от иммуногенности экспрессируемого генного продукта. В общем случае, иммунологически или профилактически эффективная доза примерно от 1 нг до 100 мг и предпочтительно примерно от 10 мкг до 300 мкг плазмидного вакцинного вектора вводится непосредственно в мышечную ткань. Эффективная доза для рекомбинантного аденовируса составляет приблизительно 106-1012 частиц и предпочтительно около 107-1011 частиц. Также рассматриваются подкожная инъекция, чрескожная инъекция, чрескожное впрыскивание и другие режимы введения, такие как внутрибрюшинный, внутривенный или введение в виде ингаляции. Также предполагается, что может быть предусмотрена бустерная вакцинация. Также является преимущественным парентеральное введение, такое как внутривенное, внутримышечное, чрескожное или другой способ введения адъювантов, таких как белок интерлейкин 12, одновременно или последовательно при парентеральном введении вакцины настоящего изобретения.
Вакцинные вектора настоящего изобретения могут быть “оголенными”, т.е. несвязанными с какими-либо белками, адъювантами или другими веществами, которые воздействуют на иммунную систему реципиента. В этом случае желательно, чтобы вакцинные вектора находились в физиологически приемлемом растворе, таком как, без ограничений, стерильный физиологический раствор или стерильный забуференный физиологический раствор. В качестве альтернативы, преимущественным может быть введение иммуностимулятора, такого как адъювант, цитокин, белок или другой носитель, с вакцинами или иммуногенными композициями настоящего изобретения. Следовательно, настоящее изобретение включает в себя применение таких иммуностимуляторов в сочетании с композициями и способами настоящего изобретения. Иммуностимулятор, как используется в настоящем описании, по существу, относится к любому веществу, которое усиливает или делает возможным иммунный ответ (антительный и/или клеточно-опосредованный) к экзогенному антигену. Указанные иммуностимуляторы могут быть введены в виде ДНК или белка. Любой из множества иммуностимуляторов может быть использован в сочетании с вакцинами и иммуногенными композициями настоящего изобретения, включая, но без ограничений, GM-CSF, столбнячный токсин IFNα, IL12, B7.1, LFA-3 и ICAM-1. Указанные иммуностимуляторы хорошо известны в данной области техники. Также могут быть использованы вещества, которые содействуют клеточному усвоению ДНК, такие как, без ограничений, ионы кальция. Такие вещества обычно относятся к реагентам, способствующим трансфекции, и фармацевтически приемлемым носителям. Специалисты в данной области техники способны определить конкретный иммуностимулятор или фармацевтически приемлемый носитель, а также подходящее время и режим введения.
Все публикации, указанные в настоящем описании, включены в настоящее описание путем ссылки с целью описания и раскрытия методов и материалов, которые могут быть использованы совместно с настоящим изобретением. Ничто в настоящем описании не должно быть истолковано как признание того, что настоящее изобретение не имеет права противопоставлять такому раскрытию источник информации с более ранним приоритетом на основании более раннего изобретения.
После того как описаны предпочтительные варианты осуществления настоящего изобретения со ссылками на прилагаемые чертежи, должно быть понятно, что настоящее изобретение не ограничено этими определенными вариантами осуществления и что различные изменения и модификации могут быть выполнены специалистами в данной области техники без отступления от объема и сущности настоящего изобретения, как определено в прилагаемой формуле изобретения.
Нижеследующие примеры иллюстрируют, но не ограничивают настоящее изобретение.
ПРИМЕР 1
Кодон-оптимизированная последовательность человеческого HER2.
Полную кодирующую hHER2.opt последовательность синтезировали и собирали при помощи BIONEXUS (Bionexus Inc. Oakland CA.) и клонировали в вектор pCR с “затупленными” концами (Invitrogen, The Netherlands). кДНК hHER2.opt конструировали с использованием олигонуклеотидов и собирали при помощи ПЦР. Для многих экспериментов, раскрытых в настоящем описании, использовали нуклеотидную последовательность hHER2.opt, несущую оптимизированную Козак-последовательность на своем 5'-конце, полную нуклеотидную последовательность, как представлено в SEQ ID NO:11.
Кроме того, АТФ-связывающий остаток лизина 753 замещали аланином (К753А) путем замены кодона ААА кодоном GCA. Эта мутация подавляет тирозинкиназную активность соответствующего белка и элиминирует дальнейшие события сигнализации и, в конечном счете, онкогенную активность человеческого (Messerle et al., Mol Cell Endocrinol 105(1):1-10 (1994)) или крысиного HER2 (Ben Levi et al., см. выше). Кроме того, киназодефицитный мутант К756 может инактивировать активность передачи сигнала коэкспрессируемого онкогена hHER2.wt.
ПРИМЕР 2
Плазмидные конструкции
pV1J-hHER2.wt: последовательность, кодирующую человеческий HER2 дикого типа, амплифицировали при помощи ПЦР из плазмиды pLTR-2/erb-B2 (любезно предоставленную P.Di Fiore, European Institute of Oncology, Milan, Italy; Di Fiore et al., Science 237 (4811):178-82 (1987)) с использованием праймеров hNeu.for1 (5'-CCAGTTTAAACATTTAAATGCCGCCACCATGGAGCTGGCGGCCT-3': (кодирующая SEQ ID NO:3 последовательность подчеркнута)) и hNeu.rev2 (5'-GCCGTCGACTTTACACTGGCACGTCCAGACCCA-3': (SEQ ID NO:4)) и TaKaRa LA Taq полимеразы (TaKaRa Otsu, Shiga, Japan). Продукт амплификации, который включает оптимизированный сайт начала трансляции (Kozak, M., J Mol Biol 196(4):947-50 (1987); Kozak, M., Nucleic Acid Res 15(20):8125-48(1987)), гидролизовали рестрикционными ферментами PmeI и SalI и клонировали в сайты EcoRV и SalI плазмиды экспрессии pV1JnsA для млекопитающих (см. Montgomery et al., DNA Cell Biol 12(9):777-83 (1993)). Таким образом сгенерированная плазмида pV1J-hHER2 содержала последовательность человеческого белка HER2 дикого типа полной длины, транскрипция которой управлялась ранним промотором цитомегаловируса человека с его интронной А последовательностью. Последовательность, кодирующая человеческий HER2 дикого типа полной длины, за которой следовала сигнальная последовательность полиаденилирования гормона роста крупного рогатого скота.
pV1J-hHER2.opt: фрагмент EcoRI-SalI из 3793 п.о. вырезали из плазмиды pCR-hHER2.opt и клонировали в соответствующие сайты плазмиды pV1JnsB (Montgomery et al., см. выше), генерируя плазмиду pV1J-hHER2.opt.
pV1J-hHER2ECDTM.opt: фрагмент из 2168 п.о. амплифицировали при помощи ПЦР с использованием TaKaRa taq с праймерами EcoRV-for (5'-CCAGATATCGAATTCTAGAGCCGCCACCATGGA-3' (SEQ ID NO:12)) и SalI-rev (5'-GCTGTCGACTTTATCAGATCAGGATGCCGAACACCACGCCC-3' (SEQ ID NO:13)) из pV1J-hHER2.opt. Полученный фрагмент гидролизовали рестрикционными ферментами EcoRV и SalI и клонировали в соответствующие сайты плазмиды pV1JnsB (Montgomery et al., см. выше), генерируя плазмиду pV1J-hHER2ECDTM.opt.
ПРИМЕР 3
Кодон-оптимизированная кДНК hHER2
Синтетический ген человеческого HER2 (hHER2.opt, фиг.1) разрабатывали для включения кодонов, предпочтительных для человека (гуманизированных), для каждого аминокислотного (далее в настоящем описании аа) остатка. Во время сборки гена при амплификации посредством ПЦР постоянно делетировалась последовательность из 86 п.о., начиная с позиции 3642, из-за высокого GC содержания последовательности в этом участке. Для решения этой проблемы для последовательности hHER2 между позициями 3601 и 3805 выбирали конструкцию с менее жесткой оптимизацией, что уменьшало GC содержание, при этом сохраняя ту же самую аа композицию.
Кодон-оптимизированную кДНК модифицировали таким образом, чтобы поддерживалась 83,9% идентичность по нуклеотидной последовательности с исходным клоном. Кодон-оптимизированную кДНК клонировали в вектора pV1J (Montgomery et al., см. выше), размещая впереди Козак-оптимизированную последовательность (5'-GCCGCCACC-3', SEQ ID NO:8), и под управлением промотора цитомегаловируса (ЦМВ) человека/интрона А плюс сигнал терминации гормона роста крупного рогатого скота (BGH). Конструкцию назвали pV1J-hHER2.opt (см. пример 2).
ПРИМЕР 4
Экспрессия in vitro плазмидных конструкций
Экспрессию in vitro конструкций pV1J-hHER2.wt и pV1J-hHER2.opt оценивали путем временной трансфекции клеточных линий НЕК-293 почки эмбриона человека и С2С7 мышиных миобластов и детектирования экспрессии человеческого HER2 поточной цитометрией. Сверхспиральную плазмидную ДНК pV1J-hHER2.wt без эндотоксина, кодирующую кассету экспрессии человеческого HER2, используемую для иммунизации, очищали из клеток DH12S E.coli (Invitrogen, Groningen, The Netherlands), используя Qiagen endo-free plasmid Giga Kit (Qiagen, Hilden, Germany).
Плазмиды pV1J-hHER2.wt или pV1J-hHER2.opt трансфицировали при помощи липофектамина (Gibco BRL Invitrogen, Groningen, The Nrtherlands) в клетки HEK-293. Аналогично, клетки С2С7 мышиных миобластов трансфицировали смесью 1:1 или 10:1 плазмид pHygEGFP (BD Biosciences Clontech, PaloAlto, CA) и pV1J-hHER2.wt или pV1J-hHER2.opt.
Трансфекция in vitro клеток HEK-293 и С2С7 показала, что кодон-оптимизированная последовательность значительно улучшает экспрессию hHER2 в сравнении с wt последовательностью (фиг.3А и 3В).
ПРИМЕР 5
Иммунизация мышей
Шестинедельных инбредных самок мышей BALB/c (H-2d, любезно предоставленных G.Forni, University of Turin) содержали в стандартных условиях. Мышей обрабатывали согласно стандартам, учрежденным Европейским союзом. В частности, проводили общую анестезию мышей кетамином (Imalgene 500; Merial Italia, Milano, Italy) по 100 мг/кг веса тела и ксилазином (Xilor, BIO 98; S.Lazzaro, Bologna, Italy) по 5,2 мг/кг, когда это было необходимо для проведения процедур.
Пятьдесят микрограмм плазмидной ДНК электроинъецировали в объеме 50 мкл в четырехглавые мышцы мышей в возрасте 6, 8 и 10 недель, как было описано ранее (Rizzuto et al., Proc. Natl. Acad. Sci. USA 96(11):6417-22 (1999)). 50 мкг оптимизированной или неоптимизированной pCMV_hNeu инъецировали без рассечения кожи в обе четырехглавые мышцы (25 мкг в 50 мкл физиологического раствора/инъекции) и проводили электростимуляцию (ЭС), как было описано ранее (Zucchelli et al., J.Virol. 74(24):11598-607 (2000); Rizzuto et al., см. выше). Кратко, электрический шок состоял из 10 серий из 1000 биполярных импульсов (130 В, 75 мА, 200 мкс/фазу).
Ad инъекцию проводили в четырехглавые мышцы мышей в объемах по 50 мкл. Сыворотки собирали на 7 неделе (1 неделя до первой иммунизации, предварительное взятие крови) и 12 неделе (две недели после последней иммунизации).
ПРИМЕР 6
ELISPOT анализ IFN-γ мышей.
Детектировали мышиные спленоциты, секретирующие IFN-γ, антиген-специфичным способом, используя стандартный чувствительный иммуноферментный анализ (ELISPOT) (Miyahira et al., J. Immunol Methods 181(1):45-54 (1995)). 96-Луночные фильтровальные планшеты MAIP MultiScreen (каталожный номер MAIPS4510; Millipore, Bedford, MA) покрывали аффинно-очищенными крысиными анти-мышиными IFN-γ антителами (IgG1, клон R4-6A2, каталожный номер 18181D, Pharmingen, San Diego, CA), разведенными стерильным PBS. После инкубации в течение ночи планшеты промывали PBST (0,005% Твин в PBS) и инкубировали в среде R10 в течение 2 часов при 37°C для блокирования неспецифичного связывания.
Спленоциты получали удалением селезенки из эйтанизированных мышей в стерильных условиях. Разрушение селезенки осуществляли путем измельчения рассеченной селезенки на металлической сетке. Красные кровяные клетки удаляли осмотическим лизисом путем добавления 1 мл 0,1Х PBS к клеточному осадку и встряхивания не более 15 сек. Затем добавляли один мл 2Х PBS и объем доводили до 4 мл 1Х PBS. Клетки осаждали центрифугированием при 1200 об/мин в течение 10 мин при комнатной температуре и осадок ресуспендировали в 1 мл среды R10 (RPMI 1640, дополненная 10% фетальной телячьей сывороткой, 2мМ L-глутамина, 50 ед. на мл пенициллина, 50 мкг на мл стрептомицина, 10мМ HEPES, 50мкМ 2-меркаптоэтанола). Производили подсчет живых клеток, используя окрашивание Türks.
Спленоциты, выделенные из селезенки двух или более иммунизированных мышей, инкубировали в течение 15 часов в присутствии 6 мкг/мл одного или пула белков с плотностью 2,5-5·105 клеток/лунку. Конканавалин А (ConA) использовали в качестве внутреннего положительного контроля для каждой мыши при 5 мкг/мл. После тщательной промывки PBST добавляли биотинилированные крысиные анти-мышиные IFN-γ антитела (каталожный номер 18112D, PharMingen; San Diego, CA). Планшеты инкубировали при 4°С в течение ночи и затем промывали PBST перед добавлением стрептавидин-щелочной фосфатазы (каталожный номер 13043Е, PharMingen; San Diego, CA). После инкубации в течение 2 часов при комнатной температуре планшеты тщательно промывали PBST и проявляли инкубацией с one-step субстратом нитроголубой тетразолий-5-бром-4-хлор-3-индолилфосфат (каталожный номер 34042, Pierce, Rockford, IL) в течение 5-15 минут для образования пятен. Реакцию останавливали промывкой планшетов водой. ДМСО и конканавалин А (10 мкг/мл) составляли фон и положительный контроль для каждого образца. Производили подсчет пятен при помощи компьютеризованного анализа изображения (AID ELR02, связанный с программным обеспечением 2.6 AID ELISPOT, Strassberg, Germany).
Частоту положительных IFN-γ продуцирующих спленоцитов на общее количество клеток, внесенных в лунку, вычисляли как среднее значение пятен, полученных из повторов с двумя различными концентрациями клеток за вычетом среднего значения, полученного аналогичным способом из пятен, измеренных в контрольных лунках, содержащих несекретирующие спленоциты. Изменения в частоте IFN-γ продуцирующих клеток определяли как превышение 95% доверительного предела, вычисленного из измерений контролей. Отличия со значением р<0,05 считали значимыми.
ПРИМЕР 7
Идентификация иммунодоминантных Т-клеточных эпитопов в человеческом белке HER2.
Триста двенадцать 15-аминокислотных пептидов, перекрывающихся по 11 аминокислотам, разрабатывали для охвата полной последовательности человеческого HER2. Те пептиды, которые также включали семь пептидов, разработанных для решения проблем, связанных с нерастворимостью, были синтезированы SynPep (Dublin, CA). Все пептиды, как было показано способом ВЭЖХ, имели чистоту >90% и были использованы без очистки ВЭЖХ. Пептиды перерастворяли в ДМСО при 35 мг/мл. Те пептиды, которые не растворялись сразу, встряхивали при 37°С, чтобы ускорить растворение. При необходимости добавляли 1-3 дополнительных объема(объемов) ДМСО для полного растворения тех пептидов, которые все еще не переходили в раствор после нескольких часов встряхивания. Перерастворенные пептиды объединяли таким образом, чтобы каждый пептид был одинаково представлен в смеси. Конечную концентрацию каждого пептида в смеси рассчитывали таким образом, чтобы она составляла 1 мг/мл. Каждая смесь была аликвотной и хранилась при -80°С.
Для идентификации иммунодоминантных Т-клеточных эпитопов гена человеческого HER2 в мышах BALB/c (H-2d генетический фон) 6-недельных самок мышей BALB/c иммунизировали при помощи инъекции 109 vp Ad5-hHER2 в четырехглавые мышцы. Вторую инъекцию проводили спустя три недели. Вторую группу мышей инъецировали аналогичным способом физиологическим раствором в качестве отрицательного контроля. Спустя три недели после второй инъекции животных забивали и оценивали частоту IFN-γ-секретирующих Т-клеток в спленоцитах мышей при помощи чувствительного иммуноферментного анализа интерферона-γ (IFN-γ ELISPOT).
Триста одиннадцать пептидов, каждый длиной 15 аминокислот, перекрывающихся по 11 остаткам и охватывающих всю последовательность человеческого белка HER2, объединяли в одиннадцать пулов, обозначенных буквами алфавита от А до К, от N-конца к С-концу. Каждый из этих пулов тестировали на способность к стимуляции IFN-γ-Т-клеток селезенки. Для пептидных пулов А, В и М измеряли при помощи IFN-γ ELISPOT статистически значимое продуцирование IFN-γ мышами, иммунизированными Ad5-hHER2, в сравнении с контролем в отсутствие пептида. Для идентификации индивидуального пептида, ответственного за активность, пептиды из пула А, В и М делили на три подпула, среди которых АIII и AIV, BIII и MI отмечены как положительные. Одиночные пептиды из таких положительных подпулов затем протестировали на их способность запускать секрецию IFN-γ. Перекрывающиеся пептиды hNeu-15 и hNeu-16 показали высокую и сравнимую реактивность. Значительно меньшую реактивность продемонстрировали перекрывающиеся пептиды hNeu-41 и hNeu-42. Также было показано, что другой пептид, hNeu-301, содержал Т-клеточный эпитоп.
Для подтверждения этих данных и идентификации субпопуляций Т-клеток CD4+ или CD8+, ответственных за продуцирование IFN-γ, охарактеризовали IFN-γ-секретирующие Т-клетки при помощи внутриклеточного окрашивания (ICS). Мышиные спленоциты инкубировали с одиночными пептидами в течение 12 часов в присутствии ингибитора секреции брефелдина А, фиксировали, пермеабилизовали и затем окрашивали маркерами для внутриклеточных IFN-γ, Cd3, CD4 и CD8 и анализировали поточной цитометрией. ICS подтвердил реактивность пептида hNeu-15, идентифицируя его как эпитоп, способный активировать клетки CD8+. Пептиды hNeu-15 и hNeu-16 были реактивными в равной степени в ELISPOT анализе, подтверждая, что эпитоп CD8+ должен содержаться в 11 аа остатках, общих для двух пептидов.
Для идентификации целевой девятимерной последовательности авторы протестировали три пептида длиной 9 аа, охватывающих область перекрывания между hNeu-15 и hNeu-16. hNeu-15.3 обеспечил наибольшую реактивность, показывая слабоувеличенную реактивность в сравнении с пептидами hNeu-15 и hNeu-16 длиной 15 аа. Интересно то, что примерно половину реактивности также детектировали с hNeu15.1, что указывает на то, что в такой 11 аа последовательности сосуществуют два перекрывающихся, но отличающихся CD8+ эпитопа.
IFN-γ ICS анализ также подтвердил реактивность hNeu301 и типировал ее как CD8+ эпитоп. Анализ таких CD8+ эпитопов при помощи IFN-γ ELISPOT подтвердил результаты, полученные при помощи ICS. Наконец, низкую реактивность детектировали для пептидов hNeu41 и hNeu42, низкий ответ которых преимущественно составлял CD4+.
ПРИМЕР 8
Внутриклеточное окрашивание цитокина
Внутриклеточное продуцирование IFN-γ измеряли согласно стандартному протоколу BD Pharmingen. Кратко, 2×106 клеток селезенки культивировали в течение 15 часов в среде R10 в присутствии 6 мкг/мл одиночных или пула пептидов и брефелдина А, в качестве ингибитора транспорта белка (набор Cytofix/Cytoperm Plus™ с GolgiPlug™; BD Pharmingen; San Diego, CA). Стафилококковый энтеротоксин В (SEB), 10 мкг/мл (каталожный номер S4881, SIGMA, Saint Louis, MI), и ДМСО тестировали при помощи спленоцитов в качестве положительного контроля и фона соответственно.
Перед окрашиванием поверхностных антигенов Ab анти-мышиные CD16/CD32 использовали для уменьшения неспецифичного иммунофлуоресцентного сигнала (каталожный номер 553142, BD PharMingen, San Diego, CA). Специфичный сигнал получали с APC-анти-мышиными CD3e, РЕ-анти-мышиными CD4 и PerCP-анти-мышиными CD8a (каталожные номера 553066, 553653 и 553036, BD Pharmingen; San Diego, CA). Затем клетки промывали, фиксировали, пермеабилизовали и окрашивали для внутриклеточного IFN-γ, используя FITC-конъюгированные mAb (каталожный номер 554411, BD Pharmingen, San Diego, CA). IFN-γ Т лимфоцитов вычисляли как 1000×[(IFN-γ+, CD3+ и CD4+ или CD8+)/(CD3+ и CD4+ или CD8+)]. Обычно, по меньшей мере, 50000 CD3+ лимфоцитов собирали при помощи одновременного выделения событий CD3+ и малых лимфоцитов. Все образцы получали окрашиванием в пределах 24 часов с использованием поточного цитометра FACSCalibur и программного обеспечения CellQuest (Becton Dickinson, San Jose, CA).
ПРИМЕР 9
Титрование и изотипирование антител
Сыворотки для титрования антител получали взятием крови позади глазничного сплетения. ELISA планшеты (Nunc Maxisorp™, Roskilde, Denmark) оставляли покрытыми козьими анти-человеческими Fc-специфичными IgG (Pierce; каталожный номер 31123) на ночь при 4°С при концентрации 2 мкг/мл в 50мМ NaHCO3 (pH 9,6). Избыток антитела удаляли и неспецифичное связывание блокировали инкубацией в течение 60 мин при 37°С в PBBST5 буфере (BSA 5%, Твин 0,05%). После промывки добавляли супернатант IgB2-клеток в условиях окрашивания и инкубировали при комнатной температуре в течение 2 часов (Chen et al., J Biol Chem 271(13):7620-9 (1996)). IgB2-клетки (любезно предоставленные Dr. Y. Yarden, Weizmann Institute of Science, Rehovot, Israel) представляли собой НЕК-293 клетки, секретирующие димерный слитый белок, образованный внеклеточным доменом HER2 и Fc частью человеческого Ig. Планшеты промывали и последовательно разбавленные сыворотки (от 1:4000 до 1:25600) в PBBST1 буфере (BSA 1%, Твин 0,05%) инкубировали в течение ночи при 4°С. Сыворотки, собранные до иммунизации, использовали в качестве фона. Промывку выполняли в PBBST1. Вторичное антитело (AP-конъюгированные козьи анти-мышиные IgG1 или IgG2a (Pharmingen, 557272 и 553389)) разбавляли 1:40000 в PBBST5 и инкубировали 2-3 часа при комнатной температуре на шейкере. После промывки планшеты проявляли инкубацией с Sigma 106 субстратом для фосфатазы (Sigma; каталожный номер А106) в диэтаноламине. Планшеты считывали при помощи автоматизированного устройства считывания ELISA (Labsistems Multiskan Bichromatic, Helsinki, Finland) и результаты выражали в виде А=А405nm-A620nm. Для каждого образца вычитали фоновый сигнал, детектируемый в сыворотке, собранной до иммунизации.
Титры анти-hHER2 сыворотки вычисляли в виде равного предельного разведения сыворотки, создающей абсорбцию, по меньшей мере, в 3 раза большую, чем абсорбция аутогенной сыворотки, собранной до иммунизации, при таком же разведении.
ПРИМЕР 10
Увеличенная иммуногенность hHER2.opt
Для оценки иммунных ответов, индуцируемых кодон-оптимизированным вектором экспрессии hHER2 и вектором экспрессии hHER2 дикого типа, мышей BALB/c иммунизировали внутримышечно плазмидной ДНК pV1J-hHER2.wt или pV1J-hHER2.opt с последующей ЭС (как описано в примере 5). Мышей инъецировали три раза, в 6, 8 и 10-недельном возрасте. Две недели спустя после последней иммунизации из каждой мыши выделяли спленоциты. Для количественного определения частот IFN-γ-секретирующего hHER2-специфичного CD8 Т-клеточного предшественника, генерируемых иммунизацией плазмидной ДНК, использовали ELISPOT анализ для H-2d укороченных Т-клеточных эпитопов hNeu15.3 и hNeu42. Иммунизация последовательностью HER2 дикого типа вызывала едва детектируемый CD8+ ответ, а реактивность CD4+ пептида отсутствовала. Напротив, оптимизированная последовательность HER2 индуцировала 10-кратное увеличение ответов на пептид CD8+, давая до 286 IFN-γ пятнообразующих клеток (SFC, среднее значение), специфичных для тестируемых эпитопов. Также детектировали более низкую активность CD4+. Пептид-специфичные IFN-γ SFC не были детектированы у мышей, иммунизированных pV1J-nsB (данные не показаны).
Сыворотки из тех же самых мышей тестировали способом ELISA, используя IgB2 белок в качестве субстрата (фиг.4В). Титр hHER2-специфичных антител детектировали у всех мышей, иммунизированных pV1J-hHER2.opt, и значение среднего геометрического Ab титра составляло 46000 или 78000 для IgG1 или IgG2a. Напротив, группа, иммунизированная pV1J-hHER2.wt, показала титр hHER2-специфичных антител со значением среднего геометрического, приблизительно в 100 раз ниже. Таким образом, эти результаты демонстрируют, что кодон-оптимизированная кДНК hHER2 является более эффективной при выработке клеточного и гуморального иммунного ответа, чем последовательность дикого типа.
ПРИМЕР 11
Аденовирусные вектора
MRKAd5-hHER2.wt: Фрагмент ДНК SwaI-SalI из Pv1J-hHER2, содержащий кДНК человеческого HER2, клонировали в соответствующие сайты челночной плазмиды polyMRKΔE1 (Bett et al., Proc Natl Acad Sci USA 91(19):8802-04 (1994)). Полученная плазмида pMRKΔE1_hHER2 содержала промотор ЦМВ человека, управляющий экспрессией кДНК человеческого HER2, за которым следовал сигнал полиаденилирования гормона роста крупного рогатого скота. Проводили рекомбинацию плазмиды pMRKΔE1_hHER2 плазмидой pAd5-HV0 с аденовирусной основой для генерации пре-аденовирусной плазмиды pAd5-hHER2.wt.
MRKАd5-hHER2ECDTM.opt: Плазмиду pCR-hHER2.opt гидролизовали EcoRI. Полученную вставку, длиной 2156 п.о., очищали и клонировали в EcoRI polyMRK-Ad5 челночную плазмиду (см. Emini et al., WO 02/22080, включенную в настоящее описание во всей своей полноте в качестве ссылки).
Плазмиды pAd5-hHER2.wt и pMRKAd5-hHER2.opt линеаризовали гидролизом PacI и трансфицировали в клетки PerC6 для генерации Ad5-hHER2 рекомбинантных аденовирусов. Вирусы выращивали в большом количестве путем множественной амплификации и очищали ультрацентрифугированием в градиенте плотности хлорида цезия (Fallaux et al., Hum Gene Ther 9(13):1909-17 (1998)). Вирусную ДНК экстрагировали гидролизом протеиназой К и целостность генома подтверждали рестрикционным анализом.
Клетки НЕК-293 инфицировали MRKAd5-hHER2.wt или MRKAd5-hHER2ECDTM.opt, используя различные коэффициенты инфицирования (m.o.i.). Экспрессия, обнаруживаемая вестерн-блот анализом, выявила более чем 10-кратное различие между укороченным белком, экспрессированным из кодон-оптимизированной последовательности, в сравнении с белком полной длины, экспрессированным из последовательности wt HER2 (данные не показаны).
ПРИМЕР 12
Сравнение режимов иммунизации
Эффективность аденовируса при индуцировании клеточно-опосредованного иммунного ответа против человеческого HER2 сравнивали с индуцированием иммунного ответа плазмидной ДНК совместно с электростимуляцией, причем и аденовирус, и плазмидная ДНК содержали кассеты экспрессии ЦМВ-HER2. Пятьдесят мкг плазмиды pV1J-HER2 инъецировали в четырехглавые мышцы мышей wt BALB/c или трансгенных мышей BALB/c со сверхэкспрессией крысиного HER2 (указанного в виде NeuT, см. Lucchini et al., Cancer Lett 64(3):203-9 (1992)) с последующей ЭС в возрасте 6 и 9 недель. Две недели спустя после бустерной иммунизации животных забивали, а клетки селезенки собирали и стимулировали пептидами, содержащими иммунодоминантные р185 эпитопы. Клетки, образующие очень небольшие пятна (SFC), детектировали после стимуляции человеческими пептидами, как у мышей BALB/c, так и у мышей neuT (фиг.5). В среднем, ответ был в 50 раз ниже ответа, индуцируемого иммунизацией Ad5-HER2. Вышеприведенные данные показывают, что иммунизация мышей плазмидой pV1J-HER2 индуцировала сопоставимый ответ у мышей BALB/c и neuT для каждого протокола.
ПРИМЕР 13
Иммунизация макак-резусов человеческим HER2 в сочетании с человеческими CEA и EpCAM антигенами
Для оценки эффективности иммунизации макак-резусов (macaca mulatta) человеческим опухолевым антигеном HER2 в сочетании с другими опухолевыми антигенами, группу из 4 макак-резусов (2 самки и 2 самца) иммунизировали смесью плазмидных ДНК векторов, экспрессирующих кодон-оптимизированные последовательности человеческих опухолевых антигенов Ep-CAM, CEA и HER2/neu.
Исследования по иммунизации проводили в Биомедицинском Центре Изучения Приматов (Biomedical Primate Research Centre) (BPRC, Rijswijk, The Netherland). Исследования по иммунизации были спланированы для оценки Т-клеточных ответов, индуцируемых человеческими антигенами против гомологов макак-резусов таких же антигенов.
Макак вакцинировали внутримышечно при помощи инъекций на 0, 2, 4, 6, 8, 10, 12, 14 и 16 неделях с последующей электростимуляцией. Животных инъецировали под анестезией 1 мл раствором (разделенным на две дозы по 0,5 мл/дозу), содержащим 6 мг плазмидной ДНК для животных, весящих 2-5 кг.
Для электростимуляции 2 серии из 100 прямоугольных биполярных импульсов (каждая продолжительностью 1 сек) подавали каждую вторую секунду в течение общего времени обработки, составлявшего 3 сек. Длина импульса составляла 2 мсек/фазу с частотой и амплитудой импульса 100 Гц и 100 мА (режим постоянного тока) соответственно.
Затем те же самые макаки были вакцинированы на 27 и 31 неделях смесью из трех аденовирусов 5 (ΔЕ1-ΔЕ3, “первая генерация”, Р2 уровень), экспрессирующих wt последовательность человеческого HER2, человеческого СЕА или человеческого ЕрСАМ соответственно.
Для измерения иммунного ответа для гомологов макак-резусов человеческого HER2, используя вышеупомянутый протокол, собирали образцы крови каждые четыре недели в течение 1 года. Клеточно-опосредованный иммунный ответ измеряли с помощью IFN-γ ELISPOT анализа. Результаты, показанные на фиг.7, указывают на то, что протокол иммунизации, обсужденный выше, был эффективным при индуцировании специфичного иммунного ответа против эндогенных гомологов макак-резусов человеческого HER2/neu.
ПРИМЕР 14
Сравнение р185-специфичного Т-клеточного ответа, вызываемого у мышей путем иммунизации pV1J-HER2.opt и pV1J-HER2ECDTM.opt
Оценивали эффективность иммунизации мутанта с С-терминальной делецией р185, сохранившего внеклеточный и трансмембранный домены (HER2ECDTM). Плазмидная ДНК, экспрессирующая кодон-оптимизированную последовательность полной длины (pV1J-HER2.opt) или укороченный белок (pV1J-HER2ECDTM.opt), электроинъецировали на 10 и 12 неделях и проводили анализ на 14 неделе. Укороченный белок HER2ECDTM индуцировал анти-185 клеточно-опосредованный ответ, превышающий ответ, индуцированный белком p185 полной длины, как в виде CD4+, так и CD8+ реактивности, как измерено с помощью IFN-gamma ELISPOT анализа (фиг.8). Анализ экспрессии in vitro в мышиных миобластах С2.7 не показал различий в экспрессии между двумя плазмидами (данные не показаны).












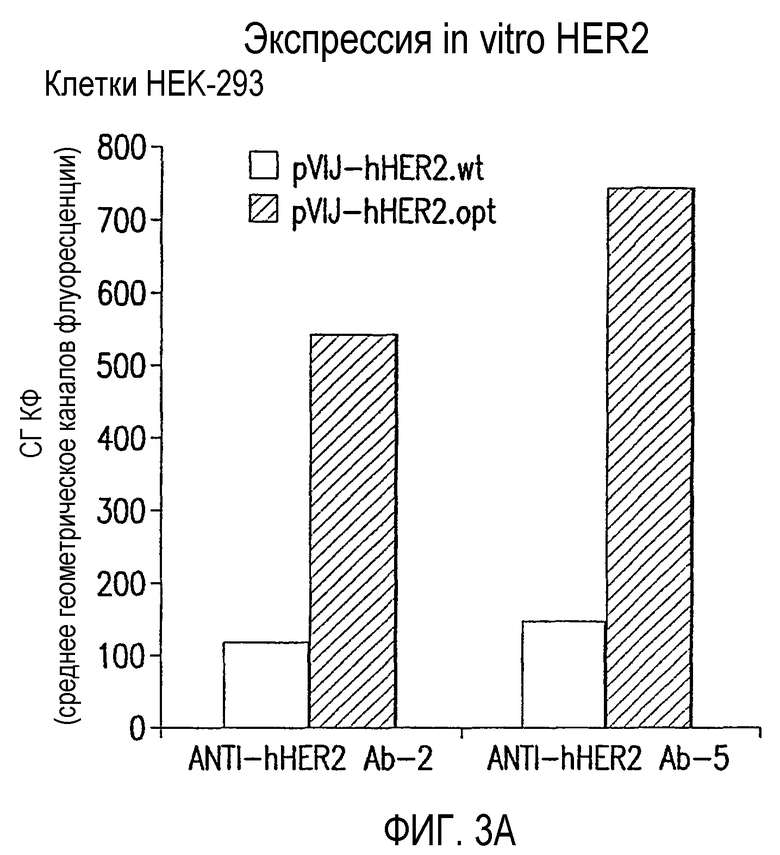
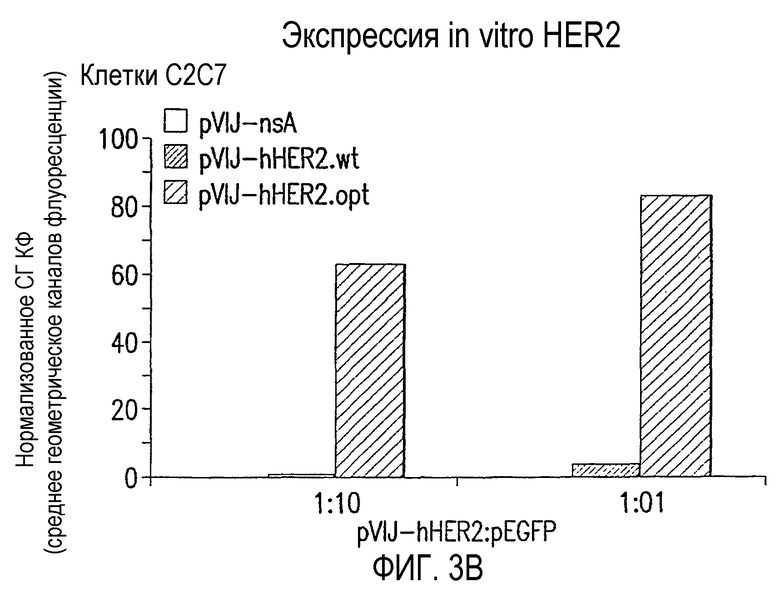
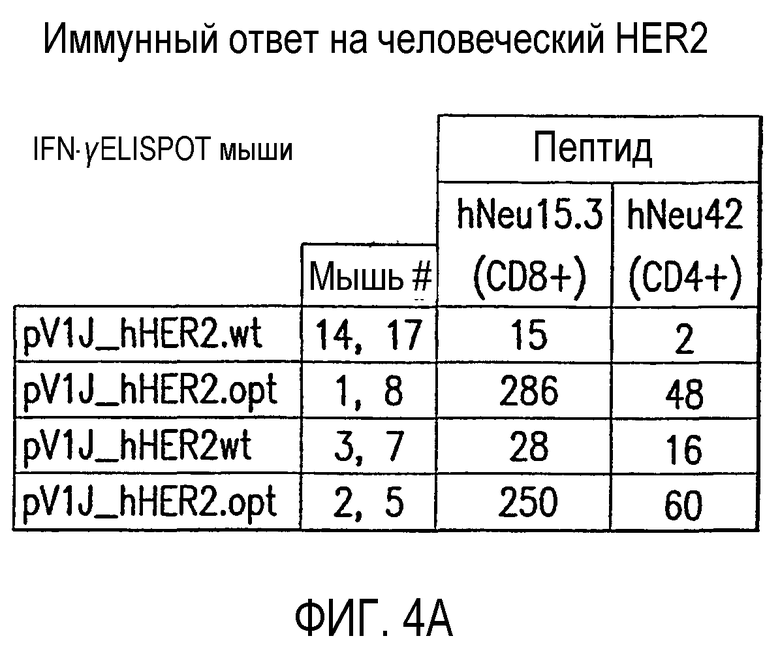

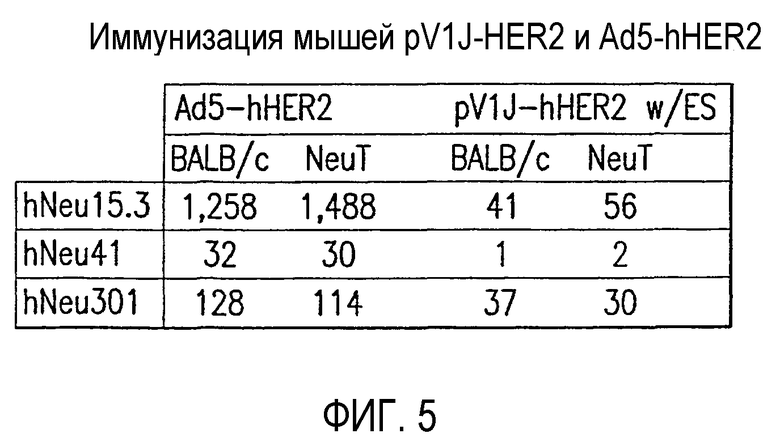


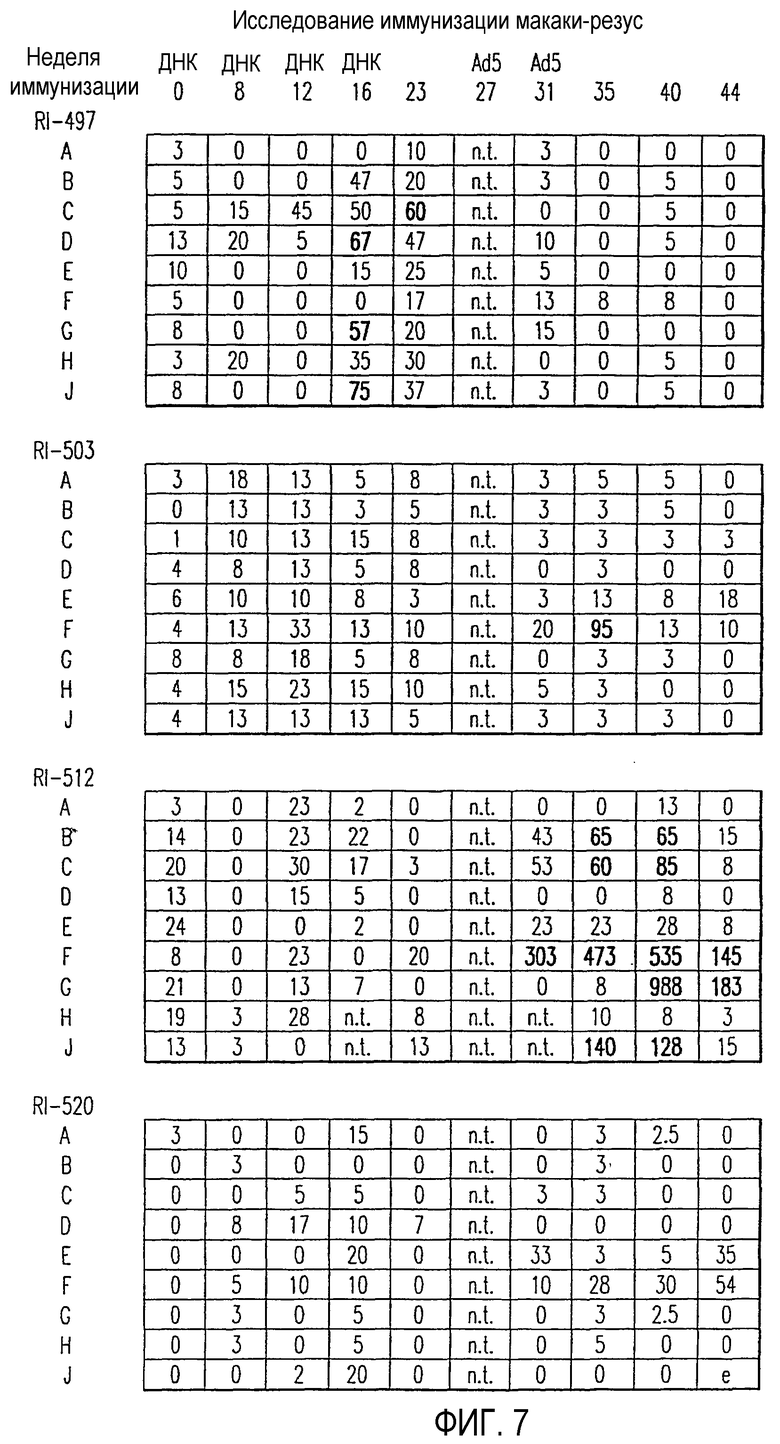
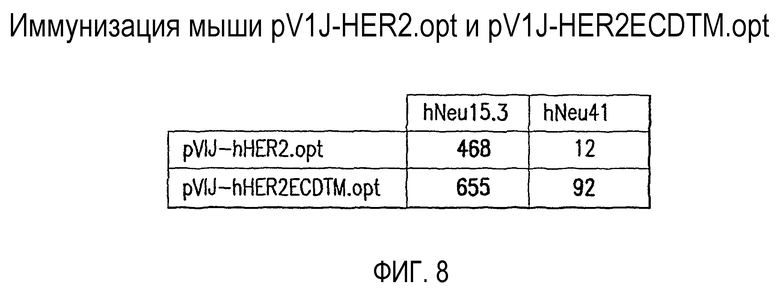
Изобретение относится к области генной инженерии и может быть использовано для оптимизации экспрессии белка антигена эпидермального фактора роста-2 (HER2/neu). Для получения белка HER2/neu используют синтетическую молекулу нуклеиновой кислоты, которая является кодон-оптимизированной для экспрессии в человеческой клетке высокого уровня указанного белка. Изобретение позволяет увеличить продукцию рекомбинантного белка HER2/neu при экспрессии в человеческих клетках. 5 н. и 3 з.п. ф-лы, 10 ил.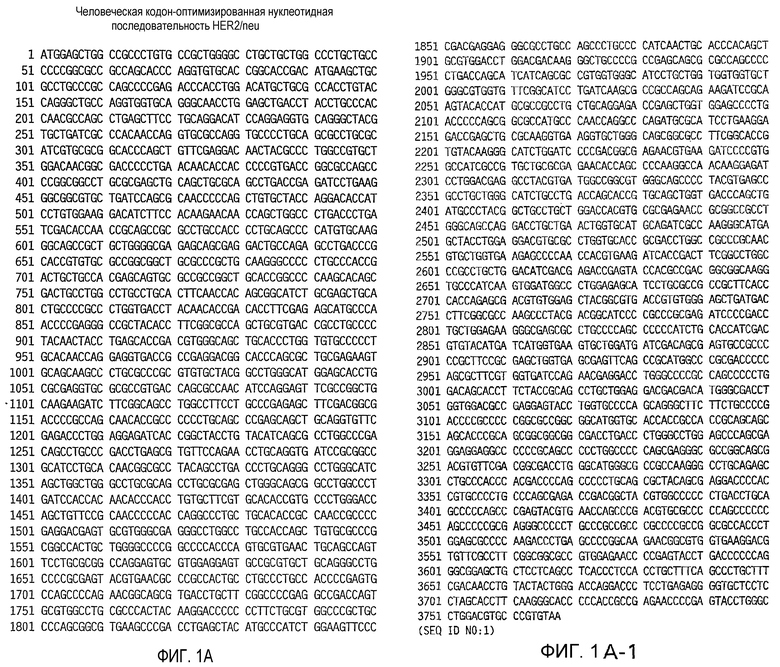
1. Синтетическая молекула нуклеиновой кислоты, которая является кодон-оптимизированной для экспрессии в человеческой клетке высокого уровня человеческого белка антигена эпидермального фактора роста-2 (HER2/neu), где синтетическая молекула нуклеиновой кислоты определяется последовательностью нуклеотидов, представленной в SEQ ID NO:1.
2. Синтетическая молекула нуклеиновой кислоты, которая является кодон-оптимизированной для экспрессии в человеческой клетке высокого уровня укороченной формы человеческого белка антигена эпидермального фактора роста-2 (HER2/neu), которая содержит внеклеточный и трансмембранный домены HER2/neu, где синтетическая молекула нуклеиновой кислоты определяется последовательностью нуклеотидов, представленной в SEQ ID NO:9.
3. Экспрессионный вектор для клеток млекопитающего, содержащий молекулу нуклеиновой кислоты по п.1 или 2, которая функционально связана с промотором.
4. Экспрессионный вектор по п.3, который является аденовирусным вектором.
5. Экспрессионный вектор по п.4, который представляет собой Ad5 вектор.
6. Экспрессионный вектор по п.4, который представляет собой Ad6 вектор или Ad24 вектор.
7. Клетка млекопитающего хозяина, содержащая вектор по п.3, для экспрессии белка HER2/neu или укороченной формы человеческого белка HER2, которая содержит внеклеточный и трансмембранный домены белка HER2.
8. Способ экспрессии человеческого белка HER2/neu или укороченной формы человеческого белка HER2, которая содержит внеклеточный и трансмембранный домены белка HER2 в рекомбинантной клетке млекопитающего хозяина, предусматривающий:
(a) введение экспрессионного вектора, содержащего нуклеиновую кислоту по п.1 или 2, в подходящую клетку млекопитающего хозяина; и
(b) культивирование клетки млекопитающего хозяина в условиях, которые позволяют экспрессию указанного человеческого белка HER2 или указанной укороченной формы человеческого белка HER2, которая содержит внеклеточный и трансмембранный домены белка HER2.
| MESSERLE К | |||
| et al., NIH/3T3 cells transformed with the activated erbB-2 oncogene can be phenotypically reverted by a kinase deficient, dominant negative erbB-2 variant, Mol | |||
| Cell Endocrinol., 1994,v.105, n.1, p.1-10 | |||
| LEDER C | |||
| et al., Enhancement of capsid gene expression: preparing the human papillomavirus type 16 major structural gene L1 for DNA |

