Изобретение относится к протеину двух ассоциированных субъединиц р40 интерлейкина-12 (IL-12), действующему в качестве антагониста рецепторов интерлейкина-12.
Интерлейкин-12 (IL-12), ранее известный как цитотоксический фактор созревания лимфоцитов (CLMF) или цитотоксический фактор стимуляции естественных клеток-киллеров (NKSF), представляет собой цитокин, который обладает плейотропной активностью, включая стимуляцию пролиферации активированных клеток Т и NK (1, 2), индукцию производства INF-γ при помощи мононуклеарных клеток периферической крови и повышение литической активности клеток NK/LAK (2-4).
IL-12 представляет собой гетеродимерную молекулу с приблизительной молекулярной массой около 75 кДа, состоящую их двух дисульфидно-связанных субъединиц: р35, имеющую приблизительную молекулярную массу около 35 кДа; и p40, имеющую приблизительную молекулярную массу около 40 кДа, (2, 4-6). Субъединица p40 разделяет гомологию аминокислотных последовательностей с рецептором интерлейкина-6 (IL-6R) и поэтому принадлежит к суперсемейству рецепторов цитокина, тогда как р35 имеет отдаленную, но существенную связь с семейством цитокинов IL- 6/G-CSF. Подразумевается, что гетеродимер р35/р40 может представлять комплекс цитокина (р35) и растворимого рецептора цитокина (p40), причем целлюлярный рецептор IL-12 обеспечивает создание функции, аналогичной функции IL-6-сигналтрансдуцирующего протеина, gp130 (7, 8).
Биологическая активность IL-12 опосредована связыванием интактной молекулы IL-12 с рецепторами плазменной мембраны на активированных клетках Т или NK (9, 10); однако вклады отдельных субъединиц в рецепторное связывание и сигнальную трансдукцию остаются неизвестными. Исследования с нейтрализующими антителами к человеческому IL-12 (II) и сайт-специфической модификации (12) предполагают, что субъединица p40 содержит эпитопы, важные для связывания IL-12 с его рецепторами. Кроме того, исследования с человеческими/мышиными химерными молекулами показывают, что р35 ответственен за видоспецифичность гетеродимера для биологической активности.
Краткое описание чертежей.
Фигура 1. Связывание при дозовой зависимости человеческого IL- 12 и COS-экспрессированного rp40 с клетками KIT225/К6, анализируемое с помощью проточной цитометрии. Различные концентрации кондиционированной среды, содержащей очищенный человеческий IL-12 или rp40 [определены с помощью ЕIА (ферментного иммуноанализа) с использованием IL-12 в качестве стандарта], инкубируют с клетками KIT225/К6 и обнаруживают с помощью биотинилированного 8E3 mAb и стрептавидина-РЕ, как описано в разделе "Материалы и Методы". Часть А: кривая "а" обозначает неспецифическое окрашивание клеток, инкубированных только с биотинилированным-8E3 и стрептавидином-РЕ. Кривые "b" и "с" обозначают клетки, инкубированные с 100 и 500 нг/мл человеческого IL-12 соответственно. Часть В: кривая "а" обозначает неспецифическое окрашивание, тогда как кривые "b", "с", "d" и "e" обозначают клетки, инкубированные с 2,5, 12,5, 125 и 500 нг/мл rp40 соответственно.
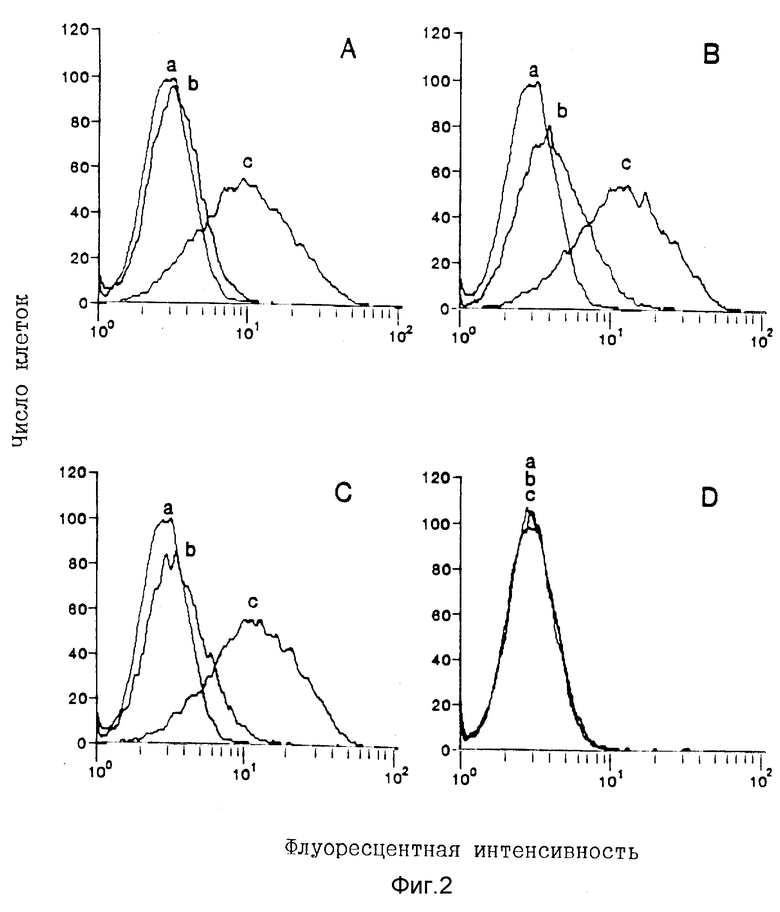
Фигура 2. Специфичность связывания rp40 с клетками KIT225/К6 при обнаружении с помощью анализа FACS (клеточный сортер с возбуждением флуоресценции). Очищенный человеческий IL-12 (А), кондиционированные среды из культур клеток COS, совместно трансфецированных с человеческими кДаНК p35 и p40 (В) или только с человеческой кДаНК p40 (С), разбавляют до 0,5 мкг/мл (определено с помощью ЕIА) и инкубируют с 4A1-нейтрализующим моноклональным античеловеческим IL-12 антителом (b) или с нормальным крысиным IgG (R-IgG) (с) при конечной концентрации 25 мкг/мл при комнатной температуре в течение 1 часа перед добавлением клеток К1Т225/К6. Кондиционированную среду из культуры клеток COS, трансфецированных с плазмидой дикого типа pEF-BOS, используют в качестве контрольной (D). Для измерения неспецифического окрашивания клетки инкубируют только с биотином-8ЕЗ и стрептавидином-PE (а).
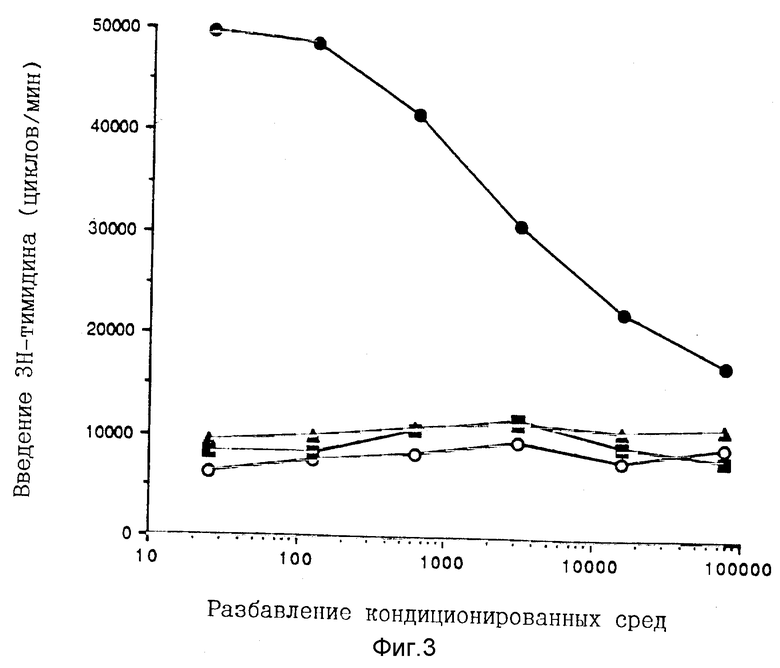
Фигура 3. Пролиферация PHA-активированных человеческих лимфобластов в ответ на кондиционированные среды, содержащие отдельно экспрессированные rp40 и rp35, либо совместно экспрессированные rp35/rp40. Человеческие PHA(фитогемагглютинин)- бласты культивируют с последовательными разбавлениями кондиционированных сред из культур клеток COS, трансфецированных с человеческими p35 и p40 (-•-), /только кДаНК p40  только кДаНК p35
только кДаНК p35  или плазмидой дикого типа pEF-BOS (-O-). Эффект в результате введения [3H] тимидина измеряют через 48 часов, как описано в разделе "Материалы и методы".
или плазмидой дикого типа pEF-BOS (-O-). Эффект в результате введения [3H] тимидина измеряют через 48 часов, как описано в разделе "Материалы и методы".
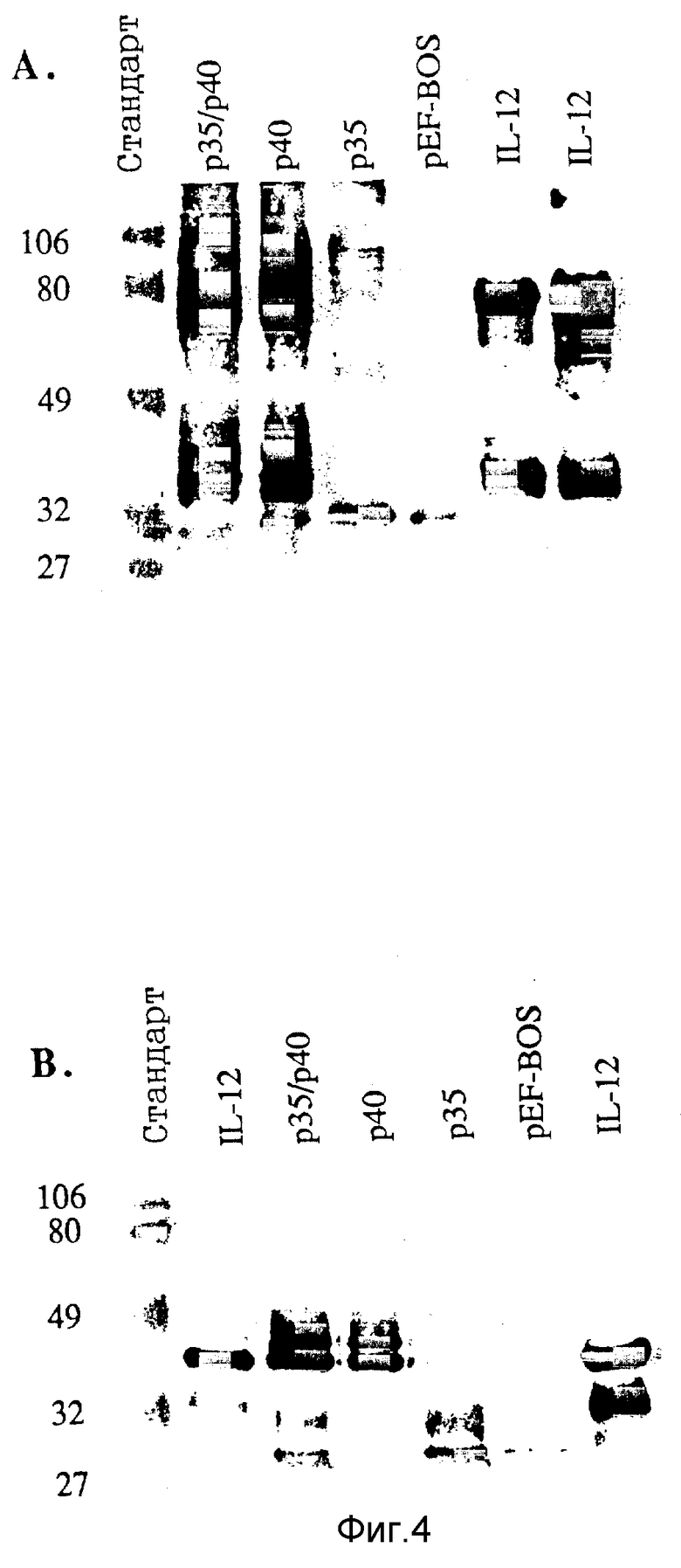
Фигура 4. Вестерн-блоттинг COS-экспрессированных человеческих гетеродимерных протеинов rp35, rp40 и rp35/rp40. Кондиционированные среды (0,5 мл) иммунопреципицируют с 5 мкг протеина lgG, выделенного из козлиной античеловеческой антисыворотки IL-12, выделяют с помощью ЭФ в ПААГ с Na-ДДС (электрофорез в полиакриламидных гелях с додецилсульфатом натрия) в невосстановительных (А) или восстановительных (В) условиях и анализируют иммуноблоттингом с использованием кроличьей античеловеческой антисыворотки IL-12 и пероксидаза-конъюгированного ослиного антикроличьего lgG.
Пробы, нагpуженные к каждой дорожке, указаны на фигуре. Человеческий IL-12 из клеток CHO загружают с двумя различными дозами (50 нг и 200 нг соответственно) для сравнения. Положения стандартов молекулярной массы (х 10-3) показаны слева.
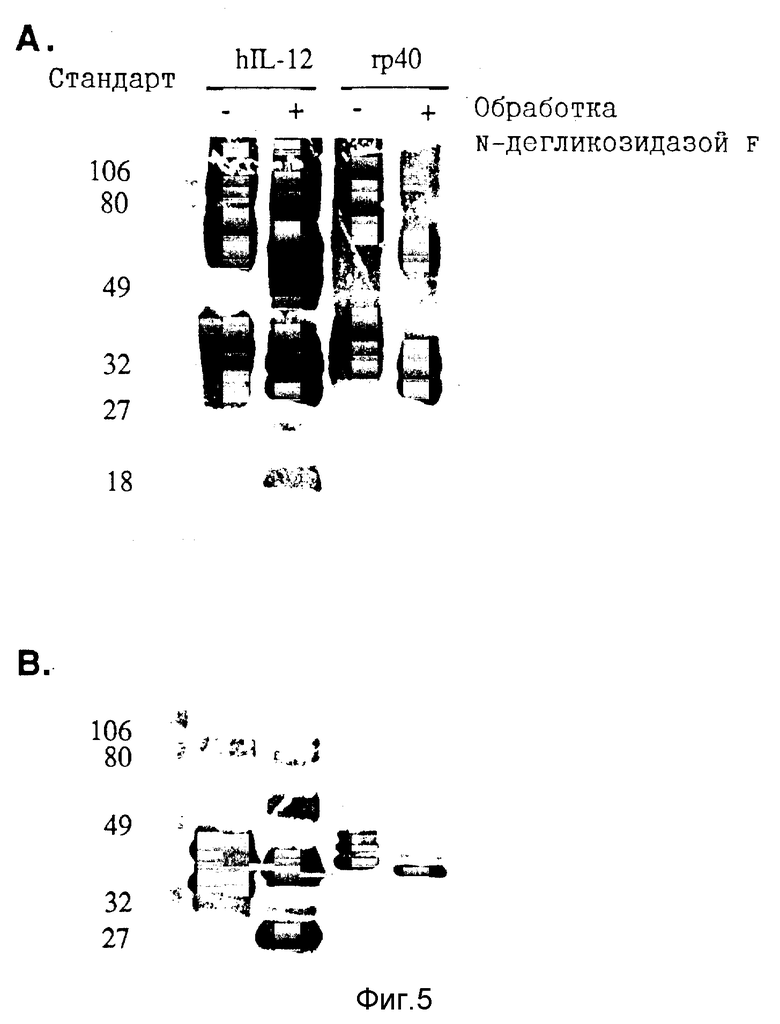
Фигура 5. Дегликозилирование COS-экспрессированных протеинов человеческого rp40. Очищенный человеческий IL-12 (0,5 мкг) и COS-экспрессированные протеины человеческого rp40, иммунопреципицированные с козлиными античеловеческими антисыворотками IL-12, дегликозилируют с помощью N-дегликозидазы F, как описано в разделе "Материалы и методы". Дубликатные пробы дегликозилированных протеинов выделяют с помощью ЭФ в ПААГ с Na-ДДС (электрофорез в полиакриламидных гелях с додецилсульфатом натрия) в невосстановительных (А) или восстановительных (В) условиях и анализируют иммуноблоттингом, как описано в разделе "Материалы и методы". Положения стандартов молекулярной массы (х 10-3) показаны слева.
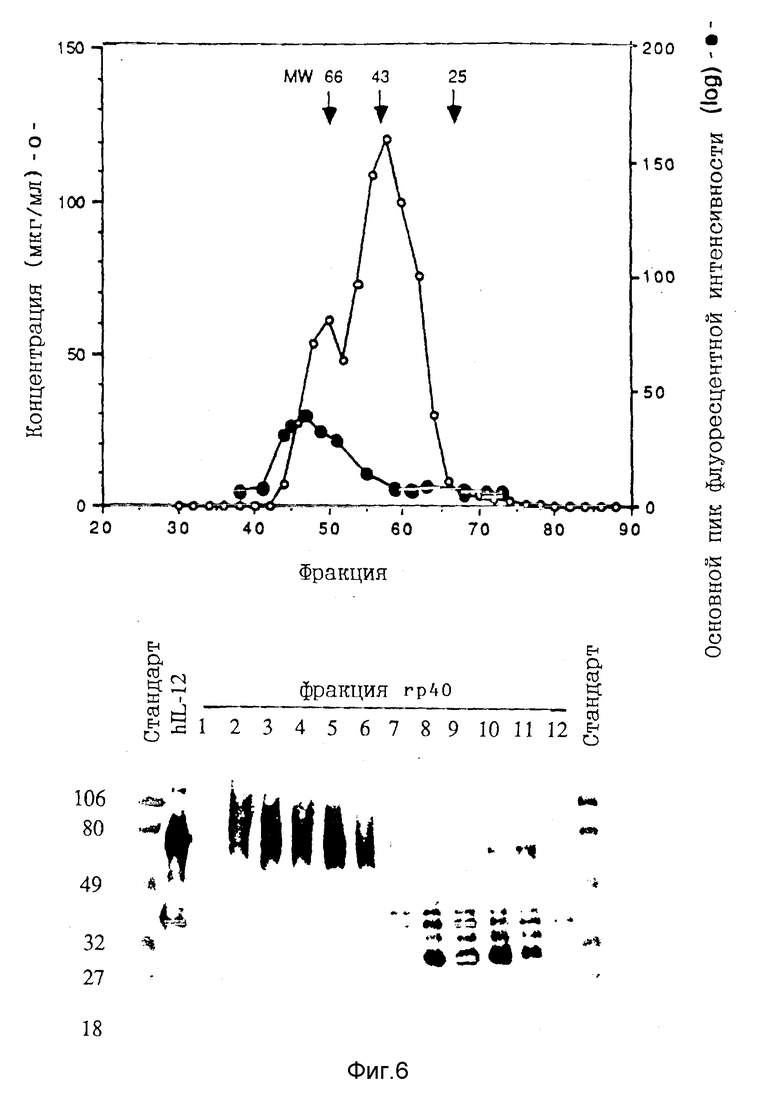
Фигура 6. ЖХВД-фракционирование видов rp40. Рекомбинантные протеины p40 частично очищают иммуноаффинной хроматографией и наносят на гель-фильтрационную колонку HiLoad Superdex 75. Фракции оценивают в EIA p40 и анализе на связывание клеток KIT225/К6 FACS (клеточный сортер с возбуждением флуоресценции). Данные EIA (-O-) откладывают в виде мкг/мл (используя человеческий IL-12 в качестве стандарта), а данные по связыванию (-•-) откладывают в виде основного пика флуоресцентной интенсивности (верхняя панель на фигуре). EIA-положительные фракции оценивают невосстановительным ЭФ в ПААГ с Na-ДДС и Вестерн-блоттингом (нижняя панель). Дорожки 1-12 обозначают протеины (около 50 нг) из фракций 40, 44, 46, 48, 50, 52, 54, 58, 60, 62, 64 и 70 соответственно.
Фигура 7. Ингибирование связывания [125I]человеческого IL-12 с человеческими PHA-бластами под действием COS-экспрессированных протеинов rp40. Различные концентрации очищенного человеческого гетеродимера IL-12
(-•-), COS-экспрессированного гомодимера rp40 (-0-) или мономера rp40 определено анализом EIA с использованием IL-12 в качестве стандарта) инкубируют с 1 • 106 PHA-бластов в присутствии 100 пМ [125I]-человеческого IL-12 в течение 1,5 часов при комнатной температуре. Данные обозначают специфическое связывание [125I]-IL-12 и выражены в виде процента от количества [125I]-IL-12, связанного с клетками в присутствии показанных концентраций немеченого IL-12 или протеинов rp40 в сравнении с общим специфическим связыванием в отсутствие немеченого IL-12.
определено анализом EIA с использованием IL-12 в качестве стандарта) инкубируют с 1 • 106 PHA-бластов в присутствии 100 пМ [125I]-человеческого IL-12 в течение 1,5 часов при комнатной температуре. Данные обозначают специфическое связывание [125I]-IL-12 и выражены в виде процента от количества [125I]-IL-12, связанного с клетками в присутствии показанных концентраций немеченого IL-12 или протеинов rp40 в сравнении с общим специфическим связыванием в отсутствие немеченого IL-12.
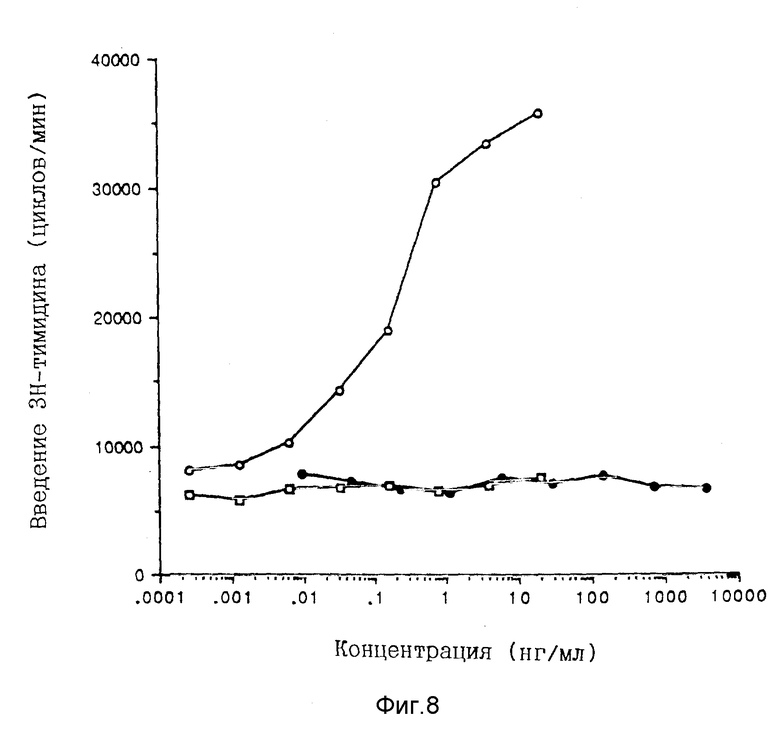
Фигура 8. COS-экспрессированный человеческий гомодимер p40 индуцирует незначительную пролиферацию человеческих PHA-бластов. Последовательные разбавления очищенного нативного человеческого IL-12 (-O-), частично очищенного COS-экспрессированного человеческого гомодимера rp40 (-•-) или буфера PBS (-
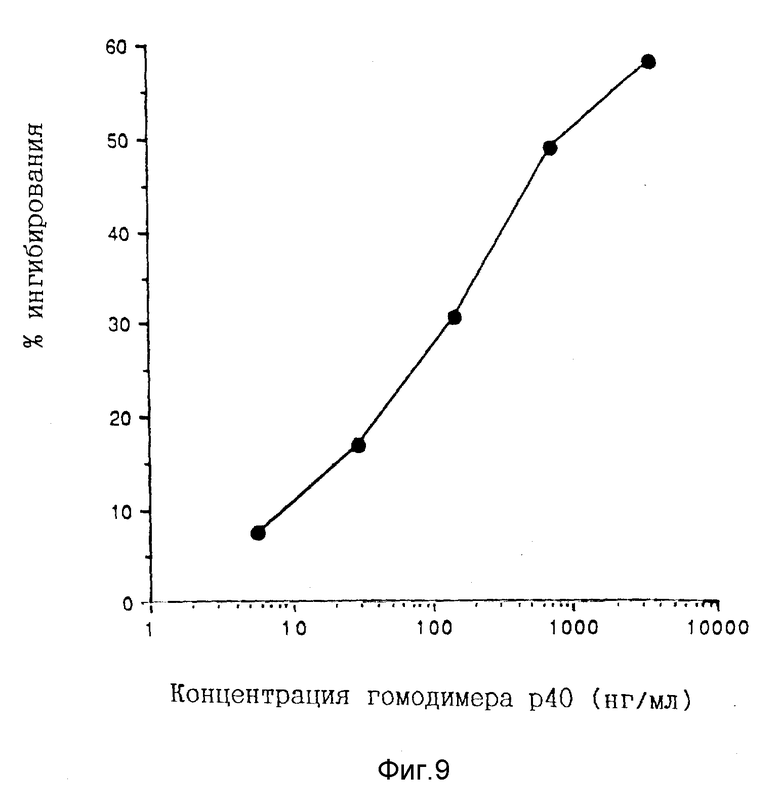
Фигура 9. Ингибирование биологической активности IL-12 под действием COS-экспрессированного гомодимера p40. Различные концентрации COS-экспрессированного человеческого гомодимера rp40 смешивают с 0,1 нг/мл нативного человеческого IL-12 перед инкубированием с 2 • 104 PHA-бластов. Нейтрализацию биологической активности IL-12 COS-экспрессированным гомодимером p40 измеряют в результате проведения 48-часового анализа на пролиферацию в соответствии с описанием в разделе "Материалы и методы". Данные выражены в виде % ингибирования введения [3H] тимидина в присутствии указанной концентрации гомодимера p40 в сравнении с введением [3H]тимидина в присутствии эквивалентной величины разбавления буфера PBS. Концентрацию p40 определяют с помощью сэндвич-EIA с использованием нативного человеческого IL-12, как описано в разделе "Материалы и методы".
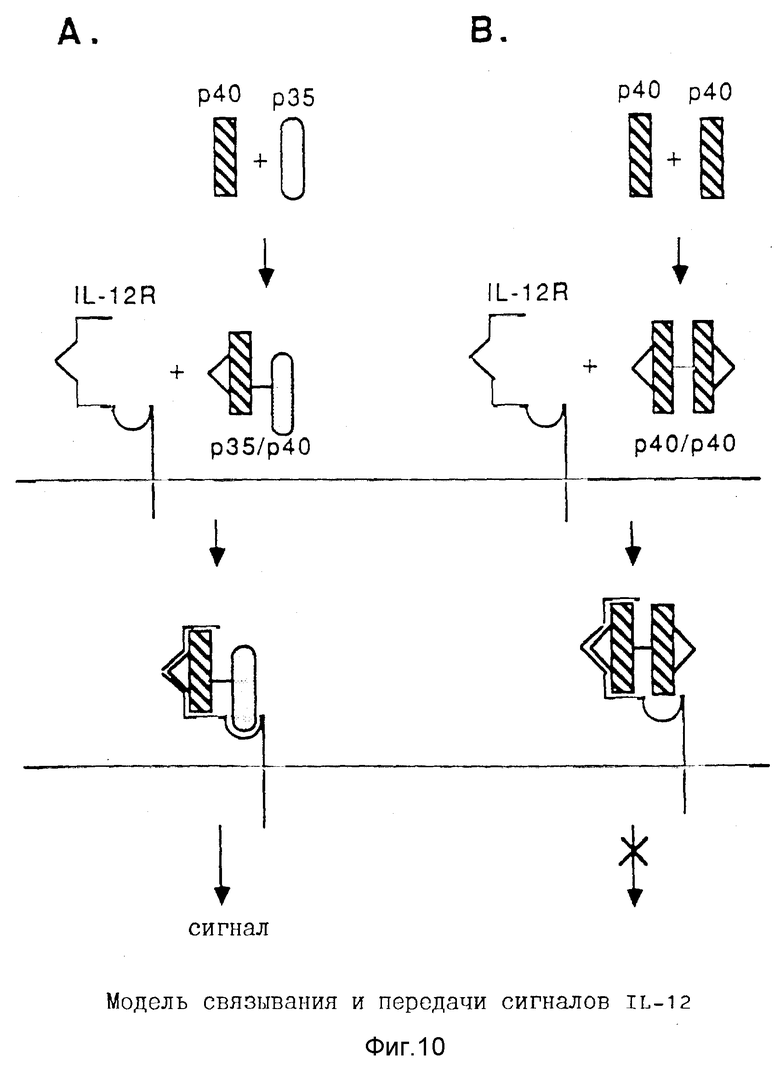
Фигура 10. Модели связывания гетеродимера p35/р40 и гомодимера р40/р40 IL-12 с рецептором IL-12 и сигнальная трансдукция. Субъединица p40 IL-12 должна ассоциироваться с субъединицей p35 или с другой молекулой p40 для правильной конформации эпитопов, необходимых для связывания с рецептором IL-12. Однако только гетеродимер (А), но не гомодимер (В), действует как полный агонист для индуцирования передачи сигналов.
Настоящее изобретение направлено на получение гомодимерных протеинов субъединиц p40 интерлейкина-12, способных связываться с рецептором интерлейкина-12, но не способных опосредовать целлюлярную пролиферацию.
Термин "гомодимер" включает ассоциацию двух субъединиц p40 одна с другой. Ассоциация субъединиц p40 имеет ковалентный или нековалентный характер и может быть достигнута in vivo, например, путем рекомбинантной экспрессии субъединицы p40 в пригодных клетках-хозяинах путем посттрансляционных модификаций, или in vitro, например, химическими средствами, такими как перекрестносшивающие агенты.
Термин "субъединица p40" включает натуральную и рекомбинантную субъединицу p40 интерлейкина-12, а также и ее производные. Термин включает фрагменты субъединицы p40, но также и слитные протеины, то есть производные субъединицы p40, содержащие аминокислотную последовательность натуральной p40 или ее частичные последовательности вместе с аминокислотными последовательностями, имеющими происхождение от других протеинов. Протеин в соответствии с настоящим изобретением может по выбору включать инициатор метионин.
Термин "субъединица p40" также включает аналоги p40 неприродного происхождения, имеющие аминокислотные последовательности, аналогичные аминокислотной последовательности p40 или ее фрагментов. Такими аналогами субъединицы p40 являются протеины, в которых одна или более аминокислот натуральной p40 или ее фрагментов замещены или делецированы без потери упомянутой активности гомодимера p40. Такие аналоги могут быть получены известными способами химии пептидов или известными способами генной инженерии, например сайт-направленным мутагенезом.
Кроме того, термины "гомодимерные протеины p40" и "субъединицы p40" также включают "функциональные производные". Данный термин относится к производным гомодимерного протеина p40 и к субъединице p40, которые могут быть приготовлены из функциональных групп, встречающихся в виде боковых цепей на остатках или N- или С- концевых групп, при помощи известных методов, и они включены в настоящее изобретение, пока остаются фармацевтически приемлемыми, то есть пока они не разрушают активность протеина и не придают токсические свойства содержащим их композициям. Эти производные могут включать, например, полиэтиленгликолевые боковые цепи, которые могут маскировать антигенные сайты и продлевать время пребывания гомодимерного протеина p40 в общих водах организма. Другие производные включают алифатические сложные эфиры карбоксильных групп, амиды карбоксильных групп при взаимодействии с аммиаком или с первичными или вторичными аминами, N-ацилпроизводные свободных аминогрупп аминокислотных остатков, образованных с ацильными составляющими (например, алканоиловые или карбоксильные ароильные группы) или О-ацилпроизводные свободных гидроксильных групп (например, остатков серила или треонила), образованных с ацильными составляющими.
Предпочтительным вариантом изобретения является гомодимерный протеин p40, состоящий из двух субъединиц p40 интерлейкина-12, предпочтительно ассоциированных по крайней мере одной дисульфидной связью. Молекулярная масса этого соединения составляет около 80 кДа. Предпочтительная субъединица p40 приведена под наименованием SEQ ID NO:1.
Гомодимерные протеины p40 в соответствии с настоящим изобретением способны связываться с рецептором интерлейкина-12, однако они не в состоянии опосредовать целлюлярную пролиферацию: они действуют в качестве антагонистов рецепторов интерлейкина-12. Эту биологическую активность можно измерить стандартными способами, известными в данной области техники (см. заявку на Европейский патент 0443827), например, как описано ниже.
В соответствии с настоящим изобретением гомодимерный протеин p40 получают в чистой форме. На основании последовательности субъединицы p40 интерлейкина-12 (SEQ ID NO:1), которую получают известными методами (см. заявку на Европейский патент N 0443827), могут быть получены биологически активные аналоги и фрагменты субъединиц p40 и гомодимерных протеинов p40 соответственно. Эти биологически активные протеины могут быть получены биологически с применением стандартных методов генной инженерии, либо могут быть химически синтезированы в аминокислотном синтезаторе или при помощи ручного синтеза с использованием хорошо известных методов жидкостного или твердофазного синтеза пептидов. Сходным путем могут быть получены аналоги, фрагменты и протеины, содержащие аминокислотную последовательность p40 вместе с другими аминокислотами. Все эти протеины затем можно исследовать на соответствующую биологическую активность.
Таким образом, настоящее изобретение относится к гомодимерным протеинам p40, их использованию и способам их получения.
Практика настоящего изобретения будет использовать, если не указано что-либо иное, традиционные методики молекулярной биологии, микробиологии, рекомбинантных ДНК и иммунологии, которые известны среднему специалисту в данной области. Такие методики объяснены в полном объеме в литературе. См., например, работы Sambrook, Fritsch & Maniatis, MOLECULAR CLONING; A LABORATORY MANUAL (1989); DNA CLONING, VOLUMES I AND II (D.N.Glover ed., 1985); OLIGONUCLEOTIDE SYNTHESIS (M.J.Gait ed., 1984); NUCLEIC ACID HYBRIDIZATION (B. D.Hames & S.J.Higgins eds., 1984); TRANSCRIPTION AND TRANSLATION (B.D.Harnes & S.J.Higgins eds., 1984); ANIMAL CELL CULTURE (R.I.Freshney ed., 1986); IMMOBILIZED CELLS AND ENZYMES (IRL Press, 1986); B.Perbal, A PRACTICAL GUIDE TO MOLECULAR CLONING (1984); the series, METHODS IN ENZYMOLOGY (Academic Press, Inc.); GENE TRANSFER VECTORS FOR MAMMALIAN CELLS (J.H.Miller and M.P. Calos eds. , 1987, Cold Spring Harbot Laboratory), Methods in Enzymology, Vol. 154 and Vol.l55 (Wu and Grossman, and Wu, eds., respectively); IMMUNOCHEMICAL METHODS IN CELL AND MOLECULAR BIOLOGY (Mayer and Walker, eds., 1987, Academic Press, London), Scopes, PROTEIN PURIFICATION: PRINCIPLES AND PRACTICE, second Edition (1987, Springer-Verlag, N.Y.), and HANDBOOK OF EXPERIMENTAL IMMUNOLOGY, Volumes I-IV (D.M.Weir and C.C.BIackwell eds., 1986).
ДНК-последовательности и ДНК-молекулы, кодирующие субъединицу p40 настоящего изобретения, могут быть экспрессированы с использованием широкого круга комбинаций хозяев/векторов. Например, пригодные векторы могут включать сегменты хромосомных, нехромосомных и синтетических ДНК-последовательностей. Примерами таких векторов являются вирусные векторы, такие как различные известные производные SV40, бактериальные векторы, такие как плазмиды из E. coli, включая pCRI, pBR322, рМВ9 и RP4, фаговые ДНК, такие как многочисленные производные фага λ, М13 и других волокнистых фагов однонитевой ДНК, а также векторы, пригодные в дрожжах, такие как плазмида 2 μ, векторы, пригодные в эукариотных клетках, более предпочтительно векторы, пригодные в животных клетках, такие, которые содержат SV40, последовательности ДНК аденовирусного и/или ретровирусного происхождения. Пригодные векторы также могут быть получены из комбинаций плазмидных и фаговых ДНК, например плазмид, которые модифицированы с тем, чтобы содержать фаговую ДНК или другие ее производные.
Экспрессирующие векторы, которые могут быть использованы для получения рекомбинантных протеинов гомодимерных p40, отличаются содержанием по крайней мере одной последовательности контроля экспрессии, которая оперативно связана с последовательностью ДНК p40, инсерцированной в вектор с тем, чтобы контролировать и управлять экспрессией клонированной последовательности ДНК p40. Примерами пригодных последовательностей контроля экспрессии являются система lac, система trp, система tac, система trc, основная операторная и промоторная области фага λ, контрольная область протеина покрытия fd, гликолитические промоторы дрожжей, например, промотор для 3-фосфоглицерат-киназы, промоторы дрожжевой кислой фосфатазы, например, Pho 5, промоторы дрожжевых α- созревающих факторов, а также промоторы, полученные из вируса полиомы, аденовируса, ретровируса и обезьяннего вируса, например ранние и поздние промоторы SV40, и другие последовательности, известные для контроля экспрессии генов прокариотных или эукариотных клеток и их вирусов, а также комбинации указанных промоторно-операторных последовательностей.
ДНК, кодирующая субъединицу p40, известна (2, 4-6, 13). ДНК может быть получена традиционными методиками клонирования или полимеразно-цепьевой реакцией (ПЦР) с использованием праймеров, комплементарных началу и концу последовательностей кодирования кДаНК субъединицы p40 (6, 13).
Настоящее изобретение также предлагает клетки-хозяева и векторы экспрессии для получения гомодимера p40. Способ предусматривает культивирование пригодной клетки или клеточной линии, которая трансформирована ДНК-последовательностью, кодирующей при экспрессии мономер p40 под управлением известных регуляторных последовательностей. Пригодными клетками или линиями клеток могут быть эукариотные клетки, такие как клетки COS, клетки SF9 для системы экспрессии Baculovirus или прокариотные клетки, такие как E.coli. Отбор других клеток-хозяев, векторов экспрессии и способов трансформации, культивирования, амплификации, скрининга и получения продуктов известны в литературе (17). Пригодными векторами экспрессии являются pEF-BOS (16a) для экспрессии COS и pACDZ-l для системы экспрессии Baculovirus (16c).
Настоящее изобретение также предлагает способы восстановления протеинов гомодимера p40, например, при помощи иммуноаффинной, гель-фильтрационной хроматографии, а также гель-электрофореза. Кроме того, гомодимер p40, полученный ферментацией прокариотных и эукариотных хозяев, трансформированных с помощью ДНК-последовательностей настоящего изобретения, затем можно подвергнуть очистке до существенной гомогенности известными методами, например центрифугированием при различных скоростях, преципитацией с помощью сульфата аммония, диализом (при нормальном и пониженном давлениях), препаративным изоэлектрическим фокусированием, препаративным гель-электрофорезом или различными хроматографическими методами, такими как гель-фильтрация, жидкостная хроматография высокого разрешения (ЖХВР), ионообменная хроматография, обращенно-фазовая хроматография и аффинная хроматография (например, на колонке SepharoseTM Blue CL-6B или на моноклональных антителах, связанных с носителем и направленных к гомодимеру IL-12).
Медикаменты, содержащие гомодимер IL-12, тоже составляют цель настоящего изобретения, также как и способ изготовления таких медикаментов, предусматривающий доставку гомодимера IL-12 и, если желательно, одного или более из числа других терапевтически ценных веществ в форме врачебного введения.
Протеин гомодимера p40 или соответствующие фармацевтические композиции могут вводиться перорально, например, в форме таблеток, таблеток с покрытием, драже, твердых или мягких желатиновых капсул, растворов, эмульсий или суспензий. Введение может быть осуществлено ректально, например, с использованием суппозиториев, локально или перкутанно, например, с использованием мазей, кремов, гелей или растворов, либо парентерально путем инъекции или периодической перфузии в течение времени. Соединение может быть введено внутривенно, внутрибрюшинно, внутримышечно или подкожно.
Для приготовления таблеток, таблеток с покрытием, драже или твердых желатиновых капсул соединения настоящего изобретения могут быть смешаны с фармацевтически инертными, неорганическими или органическими наполнителями. Примеры пригодных наполнителей для таблеток, драже или твердых желатиновых капсул включают лактозу, маисовый крахмал или его производные, тальк или стеариновую кислоту или ее соли.
Пригодные наполнители для использования с мягкими желатиновыми капсулами включают, например, растительные масла, парафины, жиры, полутвердые или жидкие полиоли и т.д. В соответствии с природой активных компонентов может возникнуть ситуация, когда для мягких желатиновых капсул не требуется никакого наполнителя.
Для получения растворов и сиропов используемые наполнители включают, например, воду, полиоли, сахарозу, инвертазу, а также глюкозу.
Фармацевтически приемлемые носители и препараты для парентерального введения включают стерильные или водные, или неводные растворы, суспензии и эмульсии. Примерами неводных растворителей являются пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло, а также инъецируемые органические сложные эфиры, такие как этил-олеат. Водные носители включают воду, спиртовые/водные растворы, эмульсии или суспензии, включая солевые или забуференные среды. Парентеральные носители включают раствор хлорида натрия, декстрозу Рихтера, декстрозу и хлорид натрия, лактированные растворы Рихтера или фиксированные масла. Внутривенные носители включают жидкостные и питательные наполнители, электролитные наполнители, например, на основе декстрозы Рихтера и тому подобное. Также могут присутствовать консерванты и другие добавки, такие как, например, противомикробные средства, антиокислители, хелатообразователи, инертные газы и тому подобное. Смотри, в основном, Remington's Pharmaceutical Science. 18th Ed., Mack Eds., 1990.
Для суппозиториев, а также местного или перкутанного применения используемые наполнители включают, например, натуральные или тяжелые масла, парафины, жиры и полутвердые или жидкие полиоли.
Фармацевтические композиции могут также включать консервирующие агенты, солюбилизирующие вещества, стабилизаторы, смачивающие средства, эмульгаторы, подслащивающие вещества, красители, отдушки, соли для изменения осмотического давления, буферы, покровные агенты или антиокислители. Они также могут содержать другие терапевтически ценные агенты.
Гомодимер p40, назначаемый человеку с целью вызвать биологический ответ, должен вводиться предпочтительно внутримышечно или внутривенно 2 или 3 раза в неделю. Ожидаемая дозировка составляет от 0,1 до 2 мг/кг массы тела, хотя доза вводимого гомодимера p40 может быть определена специалистом без проведения затруднительного экспериментирования.
Предлагаемое изобретение также относится к способу получения медицинского средства или фармацевтической композиции, содержащей протеин гомодимера p40 в соответствии с изобретением.
Гомодимер p40 IL-12 пригоден в качестве антагониста IL-12 для блокировки биологической активности IL-12 при патологических иммунных реакциях. Результаты исследований in vitro и in vivo предполагают, что IL-12 играет важную роль в развитии Т-клеток- хелперов Th1-типа, которые промотируют клеточно-опосредованные иммунные ответы (22, 24), в индуцировании производства гамма-интерферона под действием зрелых клеток Т и/или NK (25) и в содействии специфическим ответам цитолитических Т-лимфоцитов (26). Избыточная активность клеток Th1 (27, 28) и/или избыточное производство гамма-интерферона (27-31) могут вовлекаться в патогенез некоторых иммунных расстройств и септического шока, указывая на то, что гомодимер p40 IL-12 представляет собой полезное вещество в лечении таких нарушений, как ревматоидный и другие воспалительные артриты, сахарный диабет I типа, рассеянный склероз, системная красная волчанка, септический шок и так далее. Кроме того, гомодимер p40 IL-12 пригоден в предотвращении или задержке отторжения аллотрансплантата и реакции "трансплантат против хозяина" (РТПХ). При использовании гомодимера p40 IL-12 с целью предотвращения или обращения патологических иммунных расстройств, он может быть объединен с другими антагонистами цитокина, такими как антитела к рецептору IL-2, растворимый рецептор TNF или антагонист рецептора IL-1.
Материалы и методы
Линии клеток.
Как было ранее обнаружено, KIT225/К6, IL-2-зависимый субклон, полученный из линии Т-клеток человека KIT225 (14), экспрессирует рецепторы IL-12 (15). Клетки KIT225/К6 культивируют в среде RPMI 1640 (BioWhittaker, Walkersville, MD), пополненной 2 мМ L-глутамина (Sigma, St. Louis, МО), 100 единиц/мл пенициллина, 100 мкг/мл стрептомицина (Gibco, Grand Island, NY), 15% FCS (JRH Biosciences, Lenexa, KS) и 100 единиц/мл человеческого rlL-2 (Hoffmann-La Roche Nutley, NJ). Клетки COS (ATCC CRL 1650 или 1651) культивируют в среде DMEM (Gibco) вместе с 4500 мг/литр глюкозы, 2 мМ L-глутамина, 50 единиц/мл пенициллина, 50 мкг/мл стрептомицина и 10% FCS (JRH Biosciences).
Экспрессия субъединиц IL-12.
Конструкции экспрессии IL-12 для COS-экспрессии строят в векторе pEF-BOS, который содержит промотор хромосомного гена 1 α (EF-1α) человеческого фактора элонгации полипептидной цепи (16а). кДаНК-фрагменты, содержащие полную кодирующую область кДаНК человеческих или мышиных p40 или p35, генерированных путем полимеразно-цепьевой реакции (ПЦР) с использованием праймеров, комплементарных началу и концу последовательностей, кодирующих кДаНК субъединицы, как описано (6, 13), субклонируют отдельно в вектор pEF-BOS в клонирующем сайте Xba 1 путем лигирования по тупым концам (17). Продукты лигирования трансформируют в штамм E.coli DH-5 альфа (BRL-Gibco), и полученные колонии подвергают скринингу с помощью ПЦР на правильную ориентацию вставок, используя прямой праймер в пределах промотора pEF-BOS и обратный праймер в пределах последовательностей, кодирующих субъединицы. Положительные клоны отбирают и амплифицируют в пригодном штамме E.coli, например, МС1161. Плазмидные ДНК получают с использованием плазмидного набора QIAGEN (Qiagen, Chatsworth, СА) и трансфецируют в клетки COS с использованием метода DEAE-декстра-на/хлорохина (17). ДНК при концентрации 2 мкг/мл смешивают с 10% Nutridoma-SP (Boehringer Mannheim, Indianapolis, IN), 0,5 мг/мл DEAE-декстрана и 0,05 мг/мл хлорохина в среде DMEM (модифицированная по способу Дульбекко питательная среда Игла) и подают в клетки COS, которые высевали в течение 16 часов. Через 2,5-3 часа инкубирования клетки обрабатывают с помощью 10% DMSO (диметилсульфоксид) в безсывороточной среде DMEM в течение 3 часов с последующей промывкой в среде DMEM, после чего культивируют в среде DMEM/10% FCS. Супернатанты собирают с культур трансфецированных клеток COS через 72 часа. Совместную экспрессию субъединиц p40 и p35 осуществляют путем смешивания двух плазмидных ДНК в отношении масс 1:1 в реагентах трансфекции. Супернатанты, полученные из культур COS, трансфецированных с помощью ДНК плазмиды дикого типа pEF-BOS, используют в качестве контрольных веществ.
Конструкцию человеческой p40 IL-12 для экспрессии в системе Baculovirus строят в векторе pACDZ-l (16b, 16c) в сайте BamHI с использованием описанного выше подхода. Рекомбинантный бакуловирус, экспрессирующий цепь p40, получают с помощью совместной трансфекции клеток SF9 (АТСС CRL 1711) с ДНК бакуловируса дикого типа и p40- экспрессирующей плазмидой pACDZ-l. Клонирование с ограниченным разбавлением в титрационных микропланшетах используют для выделения одного рекомбинантного бакуловируса, экспрессирующего субъединицу p40 человеческого IL-12.
Анализы на связывание и пролиферацию рецепторов IL-12.
Связывание COS-экспрессированных молекул IL-12 с клетками, переносящими рецептор IL-12, измеряют анализом FACS (клеточный сортер с возбуждением флуоресценции), в основном в соответствии с описанием Desai et al. (10). Короче говоря, 1 • 106 клеток KIT225/К6, суспендированных в 25 мкл буфера FACS [PBS (фосфат- забуференный солевой раствор)/2% FCS/0,05% азида натрия] инкубируют с препаратами IL-12 (25 мкл) при комнатной температуре в течение 40 минут с последующим инкубированием с биотинилированным mAb 8ЕЗ, неингибирующим античеловеческим IL-12, p40-специфическим моноклональным антителом (5 мкг/мл, 50 мкл) (11)в течение 30 минут, а затем со стрептавидином-РЕ (1,5 мкг/мл, 50 мкл; Fisher-Biotech, Pittsburgh, PA) в течение 20 минут. Окрашенные клетки анализируют на проточном цитомере FACScan (Becton Dickinson). Специфичность связывания определяют предварительным инкубированием препаратов IL-12 (0,5 мкг/мл) с 4А1 (25 мкг/мл), крысиным ингибирующим моноклональным антителом к человеческому IL-12, перед прибавлением клеток. Контрольные пробы инкубируют с нормальным крысиным lgG (25 мкг/мл). Свойства рецепторного связывания COS-экспрессированных молекул IL-12 также оценивают в конкурентном анализе связывания с рецептором [125I]IL-12, осуществляемом в основном в соответствии с вышеприведенным описанием (11). 0,1 мл аликвоты последовательных разбавлений культуральных супернатантов очищенного IL-12 смешивают с 0,05 мл аликвотами связующего буфера (RPMI-1640, 5% FCS, 25 мМ HEPES, pH 7,4), содержащего [125I] IL-12 (2 • 106 циклов в минуту). Смесь добавляют к 0,1 мл активированных бластов (1 х 107 клеток/мл), а затем инкубируют во встряхиваемой водяной бане при температуре 25oC в течение 1,5 часа. Неспецифическое связывание определяют включением в анализ 20 мкг/мл немеченого IL-12. Инкубирования осуществляют дублированно. Клеточно-связанную радиоактивность отделяют от свободного [125I]IL-12 центрифугированием 0,1 мл аликвот содержимого анализа (дублированно) через 0,1 мл силиконового масла в течение 90 секунд с интенсивностью 10000 х г. Наконечник, содержащий осадок после центрифугирования клеток, срезают и клеточно-связанную радиоактивность определяют в счетчике гамма-квантов.
Биологическую активность COS-экспрессированных молекул IL-12 оценивают в анализах на пролиферацию с использованием четырехдневных РНА-активированных человеческих лимфобластов, описанных ранее (4, 13).
Анти-IL-12 антитела и ферментативный сэндвич-иммуноанализ (EIA).
Козлиные и кроличьи антисыворотки к человеческому IL-12 получают от животных, иммунизированных очищенным человеческим rIL-12, который экспрессирован в клетках СНО (35). Фракцию lgG выделяют из 100 мл антисывороток при аффинной хроматографии на колонке Protein-G Sepharose (Pharmacia LKB, Piscataway, NJ) в соответствии с рекомендациями изготовителя. Античеловеческие IL-12-антитела очищают от фракций lgG на иммуноаффинной колонке AvidGel F (BioProbe International) с IL-12-конъюгированным гидразидом (размеры колонки 1,5 х 2,0 см, 0,55 мг протеина на мл смолы). Биотинилирование антител с использованием Biotin X-NHS (Calbiochem, San Diego, CA) осуществляют в соответствии с описанием в работе (18). Моноклональные антитела 4А1 и 8Е3 являются крысиными антителами, которые специфичны к субъединице p40 человеческого IL- 12 (заявка на Европейский патент N 0433827, отсылка 11).
Сэндвич-анализ EIA в отношении IL-12 с использованием mAb 4А1 в качестве захваченного антитела и пероксидаза-конъюгированного 8ЕЗ в качестве обнаружительного антитела осуществляют так, как описано ранее (11). Этот анализ позволяет обнаружить гетеродимер IL-12 и субъединицу p40, а не субъединицу p35. Поэтому для обнаружения p40 и p35 разработан второй сэндвич-EIA в отношении IL-12 с использованием поликлональных антител. В этом анализе 96-луночные планшеты EIA (Nunc MaxiSorp, Thousand Oaks, CA) накрывают аффинно-очищенным козлиным античеловеческим IL-12 антителом (2 мкг/мл, 50 мкл/лунку) при температуре 4oC в течение ночи и блокируют с помощью 1% BSA (бычий сывороточный альбумин) в PBS, pH 7,4, в течение 1 часа при комнатной температуре. Серийные разбавления IL- 12 и культуральные супернатанты наносят на планшеты и инкубируют при комнатной температуре в течение 2,5 часа. Планшеты затем инкубируют с биотинилированным, аффинно-очищенным кроличьим античеловеческим IL-12 антителом (500 нг/мл, 50 мкл/лунку), а затем с пероксидаза-конъюгированным стрептавидином (1 мкг/мл, 50 мкл/лунку, Sigma, St.Louis, МО). Цвет проявляется при использовании 100 мкл 1 мМ ABTS [2,2'-азинобис(3-этилбензтиазолинсульфокислота)] /0,1% (в отношении объемов) H2O2, и спектральную поглощательную способность при 405 нм определяют с помощью кинетического планшет-ридера Vmax (Molecular Devices, Palo Alto, CA). Все значения основаны на стандартной кривой IL-12 без каких-либо коррекций, вычисляемых для определения различий в молекулярной массе мономеров или димеров.
Иммунопреципитация
Иммунопреципитацию COS-экспрессированных субъединиц и гетеродимеров IL-12 осуществляют, как описано (17). Вкратце, 0,5 мл супернатантов от трансфецированных культур COS инкубируют с 5 мкг протеина lgG, выделенного из козлиной антисыворотки к IL-12, при температуре 4oC на вращающемся смесителе в течение ночи. Иммунные комплексы адсорбируют на колонку Protein G-Sepharose (50% суспензия, 10 мкл, Pharmacia LKB) при температуре 4oC в течение 2 часов, и гранулы промывают дважды с помощью 1 мМ буфера NET-Gel [50 мМ Трис-HCl, pH 7,5, 150 мМ NaCl, 0,1% (в отношении объемов) Nonidet Р-40, 1 мМ ЭДТК, 0,25% (в отношении массы к объему) желатины и 0,02% (в отношении массы к объему) азида натрия] и один раз с помощью 1 мл 10 мМ Tris-HCl (pH 7,5) с содержанием 0,1% (в отношении объемов) Nonidet Р-40. Связанные протеины диссоциируются из гранул при нагревании в течение 3 минут при температуре 95oC и в восстановительном (10% 2-МЕ) или в невосстановительном буфере додецилсульфатной пробы.
ЭФ в ПААГ с Na-ДДС и Вестерн-блоттинг.
Электрофорез в полиакриламидных гелях с додецилсульфатом натрия (ЭФ в ПААГ с Na-ДДС) осуществляют в соответствии с методом Laemmli (19). Вестерн-блоттинг осуществляют путем электрофоретического переноса протеинов к нитроцеллюлозной мембране (0,2 мкм) (MSI, Westboro, МА). Перенесенные мембраны блокируют путем инкубации в буфере PBST [PBS с 0,05% (в отношении объемов) Tween-20], содержащем 5% (в отношении массы к объему) обезжиренного сухого молока, после чего зондируют с помощью анти-IL-12 кроличьей антисыворотки (разбавление 1:500). После трех промывок с помощью буфера PBST мембраны инкубируют при комнатной температуре с пероксидаза-конъюгированными ослиными антикроличьими антителами к IgG (разбавление 1:1000) (Jackson Immuno Research, West Grove, PA). Цвет проявляется при использовании 4-хлоро-1-нафтола (BioRad, Richmond, СА) в 20 мМ буфере Tris-HCI, pH 7,5, содержащем 0,1% (в отношении объемов) H2O2.
Очистка COS-экспрессированной p40>
Один литр кондиционированных сред, содержащих приблизительно 3 мкг/мл человеческой рекомбинантной p40 (rp40), вводят в mAb 4А1- конъюгированную иммуноаффинную колонку NuGel (NHS) (2,5 х 10 см, содержит 1,6 мг антитела на мл геля) (35) со скоростью растекания фронта растворителя 2 мл/мин, и колонку промывают обильно с помощью PBS, содержащего 0,5М NaCI и 0,2% Tween 20, до тех пор, пока мониторинг спектральной поглощательной способности при 280 нм не станет менее 0,01. Затем связанные протеины элюируют с помощью 100 мМ глицина/150 мМ NaCI (pH 2,8) со скоростью растекания фронта растворителя 2 мл/мин, и фракции 20 мл собирают и сразу же нейтрализуют с помощью 1/10 объемн. 1 М Трис-HCl (pH 8,0). EIA-положительные фракции группируют, диализируют против PBS в течение ночи при температуре 4oC, концентрируют ультрафильтрацией с использованием мембран YM 10 (Amicon, Beverly, МА) до 5 мл и вводят в колонку HiLoad Superdex 75 (Pharmacia LKB) (1,6 x 60 см), уравновешенную с помощью фосфат-забуференного раствора по способу Дульбекко. Колонку элюируют со скоростью растекания фронта растворителя 1 мл/мин с помощью того же буфера и 1 мл фракции собирают. Протеины из каждой фракции исследуют с помощью EIA, ЭФ в ПААГ с Na-ДДС и Вестерн-блоттинга. Дегликозилирование.
500 нг очищенного человеческого IL-12 или иммунопреципицированного протеина rp40 денатурируют путем нагревания при температуре 95oC в течение 5 минут в 0,25М растворе Na2HPO4 (pH 7,2), 0,5% Na-ДДС с использованием или без использования 1% 2-МЕ. Пробы охлаждают до комнатной температуры, доводят до 1% Nonidet Р-40, 20 мМ ЭДТК и затем обрабатывают с использованием 0,1 единицы N-гликозидазы-Р (Boehringer Mannheim, Indianapolis, IN) при температуре 37oC в течение 24 часов. Дегликозилированные протеины исследуют с помощью ЭФ в ПААГ с Na-ДДС и Вестерн-блоттинга.
Анализ аминоконцевой последовательности COS-экспрессированной p40.
Иммуноаффинно-очищенные протеины rp40 разделяют на 10% невосстановительном геле Na-ДДС и переносят электрофоретически на мембрану PVDF ImmobilonTM (Millipore, Bedford, MA). Полосы с молекулярной массой около 80 и около 40 кДа, идентифицированные окрашиванием голубым Кумасси, подвергают автоматизированному расщеплению по Эдману на газофазном секвенаторе Applied Biosystems, Модель 470А, с применением анализа в диалоговом режиме производных фенилтиогидантоиновой (РТН) аминокислоты, как описано ранее (20).
Экспрессия и характеристика субъединиц человеческого IL-12/
Субъединицы p35 и p40 человеческого IL-12, либо гетеродимер p35/p40 человеческого IL-12 экспрессируют путем трансфекции самостоятельной кДаНК субъединицы или совместной трансфекции обеих кДаНК при отношении 1:1 (в отношении масс) в клетках COS. Секрецию рекомбинантных протеинов оценивают двумя различными анализами EIA. EIA на основе p40-специфического моноклонального антитела способен обнаруживать субъединицу p40 и гетеродимер p40/p35. IL-12- специфический поликлональный EIA также способен обнаруживать субъединицу p35. С использованием IL-12 в качестве стандарта определяют, что концентрация протеинов rp40 и rp35/rp40 в кондиционированных средах составляет 0,5-3,0 мкг/мл, тогда как при экспрессии только одной rp35 концентрация составляет приблизительно 0,2 мкг/мл. Остается непонятным, является ли экспрессия p35 низкой или мала чувствительность поликлонального EIA при обнаружении p35.
Рекомбинантные протеины COS-экспрессированных молекул IL-12 первоначально исследуют на их способность ингибировать связывание [125I] человеческого IL-12 с PHA-активированными человеческими лимфобластами. Супернатанты rp40 при разбавлении 1:2 проявляют 30- 40% ингибирования связывания [125I] IL-12 в результате трех самостоятельных экспериментов, тогда как супернатанты rp35 являются неактивными. Связывание rp40 с рецептором IL-12 далее характеризуют проточной цитометрией с использованием клеток KIT225/К6, которые экспрессируют рецепторы IL-12 (IL-12R) (15). Дозово-зависимое связывание человеческого IL-12 и rp40 с клетками KIT225/К6 наблюдают в диапазоне от 2,5 до 500 нг/мл (Фиг. 1). Специфичность связывания проявляется в достижении свыше 80% ингибирования связывания в процессе предварительной инкубации IL-12 или rp40 с ингибирующим крысиным античеловеческим p40-моноклональным антителом, 4A1 (Фиг.2). Нормальный крысиный lgG не оказывает никакого воздействия на связывание IL-12 или rp40.
Кондиционированные среды, содержащие протеины COS-экспрессированной субъединицы IL-12, оценивают в анализе на пролиферацию человеческих PHA-бластов (Фиг. 3). Среда, содержащая rp35/rp40, поддерживает пролиферацию Т-клеток дозово-зависимым образом с очевидной ЕС50 (эффективной концентрацией), равной 8 нг/мл. Супернатанты rp40 не индуцируют пролиферацию при концентрациях, эквивалентных концентрации супернатантов rp35/rp40.
Характеристика видов 40 кДа и 80 кДа rp40.
Субъединицы рекомбинантного человеческого IL-12 иммунопреципицируют с козлиной антисывороткой к человеческому IL-12 и характеризуют с помощью ЭФ в ПААГ с Na-ДДС и Вестерн-блоттинга. Анализ rp40, экспрессированной клетками COS, трансфецированными только с кДаНК p40, выявляет два набора множественных полос при невосстановительных условиях с гетерогенными молекулярными массами 70-85 кДа и 35-45 кДа (Фиг.4A). При восстановительных условиях идентифицируют только три близко разнесенные полосы массой около 38-49 кДа, что говорит о том, что протеины с молекулярной массой 80 кДа являются дисульфидно-связанными гомодимерами rp40 (Фиг.4B). Обработка иммунопреципитатов rp40 с помощью N-дегликозидазы-F сдвигает оба молекулярных вида вниз к более мелким продуктам при невосстановительных условиях (Фиг.5A) и превращает восстановленные тройные полосы в один 35 кДа-продукт, аналогичный субъединице p40 дегликозилированного человеческого IL- 12 (12), что показывает, что множественные полосы rp40 экспрессированной в клетках COS обусловлены гетерогенностью гликозилирования.
В противоположность этому иммунопреципитация протеина rp35 выявляет только одну полосу с молекулярной массой 35 кДа при восстановительных условиях (Фиг. 4B). При невосстановительных условиях обнаружен ряд слегка окрашенных полос с молекулярной массой 60-70 кДа, что говорит о том, что rp35 также может частично образовывать димеры. Однако поликлональное козлиное анти-1L-12 антитело плохо распознает протеины rp35 (Фиг.4A). Совместная экспрессия p35 и p40 дает рисунок полос, представляющий собой смесь тех, которые видны, когда каждую субъединицу экспрессируют самостоятельно (Фиг.4).
Для подтверждения идентичности двух видов rp40 протеины rp40 частично очищают иммуноаффинной хроматографией на 4А1. Только 60% EIA-положительного вещества восстанавливают путем элюирования с помощью 100 мМ глицина, содержащего 150 мМ NaCI с pH 2,8. Протеины, иммунно-очищенные на колонке 4A1, затем разделяют с помощью ЭФ в ПААГ с Na-ДДС, электрофоретически переносят на мембрану PVDF и подвергают аминокислотному микросеквенированию. Одна широкая полоса с молекулярной массой около 80 кДа и две полосы с молекулярной массой 35-40 кДа дают NH2-терминальные последовательности, идентичные последовательностям нативной р40 человеческого IL-12, очищенной от клеток NC-37 (4, 12) (Таблица 1). Никакого следа последовательности p35 не идентифицировано при использовании вида rp40. Этот результат подтверждает, что протеин с молекулярной массой 80 кДа является гомодимером p40.
Иммуноаффинно-очищенные протеины р40 далее фракционируют с помощью гель-фильтрационной хроматографии на колонке Superdex-75. Два пика EIA-положительных протеинов идентифицируют с молекулярной массой соответственно 80 кДа и 40 кДа (Фиг.6А). ЭФ в ПААГ с Na-ДДС и Вестерн-блоттинг фракций подтверждают отделение димера от мономера rp40 (Фиг. 6В). Отношение мономера к димеру варьируется от эксперимента к эксперименту, однако в среднем приблизительно 30% COS-экспрессированной rp40 представляет собой гомодимер p40.
Фракции от колонки Superdex-75 исследуют на связывание с клетками KIT225 с помощью анализа FACS. Связующая активность коррелирует только с 80 кДа р40 EIA-положительным протеином (Фиг.6). Пиковые фракции 80 и 40 кДа группируют отдельно, концентрируют и исследуют в конкурентном анализе на связывание с радиолигандными рецепторами (Фиг.7). Пул протеинов 80 кДа ингибирует связывание [125I]человеческого IL-12 с PHA-бластами с IC50 (ингибирующей концентрацией, приводящей к потере активности на 50% активности), равной 80 нг/мл, что аналогично IC50 гетеродимера человеческого IL-12 (20 нг/мл). Однако наклон конкурентной кривой под действием гомодимера с молекулярной массой 80 кДа отличается от наклона гетеродимера IL-12, что говорит о различии в связующем взаимодействии с рецептором. Пул протеинов с массой 40 кДа ингибирует связывание [125I] человеческого IL-12 с IC50 почти в сотни раз выше, что по всей вероятности обусловлено малой величиной сочетания с гомодимером p40 (Фиг.6B).
Способности мономера и димера rp40 поддерживать пролиферацию PHA-бластов также исследованы (Фиг.8). Никакого пролиферационного ответа не обнаружено с любыми видами rp40 даже при концентрации в 10000 раз выше, нежели концентрации человеческого IL-12, необходимой для снижения на 50% максимального ответа. Димер rp40 исследуют на его способность нейтрализовать IL-12-зависимую пролиферацию PHA-бластов. Протеин с массой 80 кДа при различных концентрациях смешивают с 0,1 нг/мл человеческого IL-12 и прибавляют к PHA-бластам. Дозово-зависимое ингибирование IL-12- индуцированной пролиферации PHA-бластов достигается при IC50, равной 1 мкг/мл (Фиг.9).
Для пояснения функциональной роли отдельных субъединиц и локализации эпитопов, опосредующих биологические и связующие активности, отдельные субъединицы, экспрессирующие либо отдельно, либо в сочетании одна с другой в клетках COS, экспрессируют и исследуют на экспрессированные протеины в анализе на связывание и биологическом анализе, а также путем Вестерн-блоттинга. Протеин rp35 является неактивным при концентрациях до 100 нг/мл в анализе на связывание и биоанализе; однако протеин rp40 репродуцированно проявляет связующую активность без биологической активности. Анализ кондиционированных сред из культур клеток COS, трансфецированных только с кДаНК р40, выявляет, что такие среды содержат и мономерную p40, и 80 кДа молекулу, реакционноспособную с анти-р40 антителами. Частичная очистка rp40 путем иммуноаффинной хроматографии выявляет, что 80 кДа-протеин, а не 40 кДа-протеин, связывается с IL-12R.
Исследуют возможность того, что 80 кДа-протеин является не гомодимером субъединицы p40, а гетеродимером, состоящим из одной субъединицы p40 IL-12 и второго 35-40 кДа-экзогенного протеина COS-происхождения. В частности, отчеты о том, что многие клеточные линии экспрессируют мРНК p35 IL-12 (21), повышают возможность того, что 80 кДа-протеин может быть p40 человеческого IL-12, ассоциированной с p35 IL-12 COS-происхождения. Вестерн-блоттинг с использованием p35-специфического антитела и эксперименты по дегликозилированию (Фиг.5) подтверждают, что 80 кДа-протеин может быть редуцирован в мономер p40. Отсутствие биологической активности, несмотря на хорошую связующую активность, предполагает, что второй протеин не является субъединицей р35 IL-12 COS-происхождения. Кроме того, экспрессия p40 в бакуловирусной системе привела к получению биологически неактивной формы 80 кДа субъединицы p40, способной связываться с рецептором. Весьма сомнительно, чтобы клетки насекомых продуцировали IL-12-образный протеин p35. Наиболее важно, что подтверждение идентичности 80 кДа-протеина в качестве гомодимера p40 получено аминокислотным микросеквенированием протеина, демонстрирующим одну N-терминальную последовательность, соответствующую субъединице p40 IL-12.
В анализе на конкурентное связывание обнаружено, что гомодимер p40 связывается с IL-12R почти также сильно, как и с гетеродимерным IL-12, что говорит о том, что ключевые связующие эпитопы IL-12 расположены в субъединице p40. Хотя значения IC50 для гетеродимера и гомодимера являются одинаковыми, 20 и 80 нг/мл соответственно, наклоны конкурентных кривых разные. Это предполагает, что есть различие во взаимодействии двух лигандов с рецептором. Наиболее вероятно, что p40-связующие эпитопы являются конформационными и индуцируются путем ассоциации с p35 или второй субъединицей p40.
Ранее излагалось, что субъединица p40 IL-12 продуцируется в избыточном количестве гетеродимерного IL-12 как под действием активированных В лимфобластоидных линий, так и под действием человеческой РВМС, стимулированной с целью получения IL-12 (12, 23). Возможно, что гомодимер p40 образуется в клетках, которые экспрессируют гетеродимеры p40/p35.
На основе наблюдений относительно роли субъединиц IL-12 в осуществлении связывания и передачи сигналов, на Фиг. 10 проиллюстрирована модель связывания IL-12 с его рецептором. Субъединица p40 содержит эпитопы рецепторного связывания, которые, тем не менее, активны только тогда, когда p40 ассоциируется со вторым протеином, то есть p35 или другой молекулой p40. Обе димерные молекулы связываются специфически с IL-12R, тем не менее только димер, содержащий p35, действует в качестве агониста, опосредуя сигналы целлюлярной трансдукции (Фиг. 10А). Напротив, димер p40/p40 ведет себя как антагонист, суппрессируя IL-12-опосредованные ответы (Фиг.10В).
Перечень последовательностей
(1) Общая информация:
(I) Заявитель:
(А) Имя: F.Hoffmann-La Roche AG
(В) Улица: Grenzacherstrasse 124
(С) Город: Basle
(D) КАНТОН: BS
(E) Государство: Switzerland
(F) Почтовый индекс: (ZIP): СН-4002
(G) Телефон: 061 -688 51 08
(Н) Телефакс: 061 - 688 13 95
(I) Телекс: 962292/965542 hlr ch
(II) Название изобретения: Гомодимер p40 Интерлейкина-12
(III) Количество последовательностей: 2
(IV) Машиночитаемая форма:
(А) Тип носителя: Гибкий диск
(В) Компьютер: Apple Macintosh
(С) Операционная система: System 7.1 (Mac)
(D) Программное обеспечение: Word 5.0
(VI) Данные по предшествующей заявке:
(А) Номер заявки: US 08/087,832
(В) Дата подачи: 2 июля 1993 года
(2) Информация о последовательности SEQ ID NO:1
(I) Характеристики последовательности:
(А) Длина: 306 аминокислот
(В) Тип: аминокислота
(D) Топология: линейная
(II) Тип молекулы: протеин
(XI) Описание последовательности SEQ ID NO:1 приведено в конце описания.
Информация о последовательности SEQ ID NO:2:
(I) Характеристики последовательности:
(А) Длина: 10 аминокислот
(В) Тип: аминокислота
(D) Топология: линейная
(II) Тип молекулы: протеин
(XI) Описание последовательности SEQ ID NO:2 приведено в конце описания.
Литература приведена в конце описания.










Изобретение относится к биотехнологии, в частности к протеину, содержащему две субъединицы р40 интерлейкина-12, которые ассоциированы одна с другой предпочтительно при помощи по крайней мере одной дисульфидной связи, имеющие мол. м. около 80 кДа. Гомодимер р40 с мол.м. 80 кДа действует в качестве антагониста рецептора интерлейкина-12. Субъединица р40 имеет аминокислотную последовательность SEQ.IDNO:1, представленную в описании. Гомодимер р40 способен связываться с рецептором интерлейкина-12, не опосредуя целлюлярную пролиферацию. 3 з.п. ф-лы, 10 ил., 1 табл.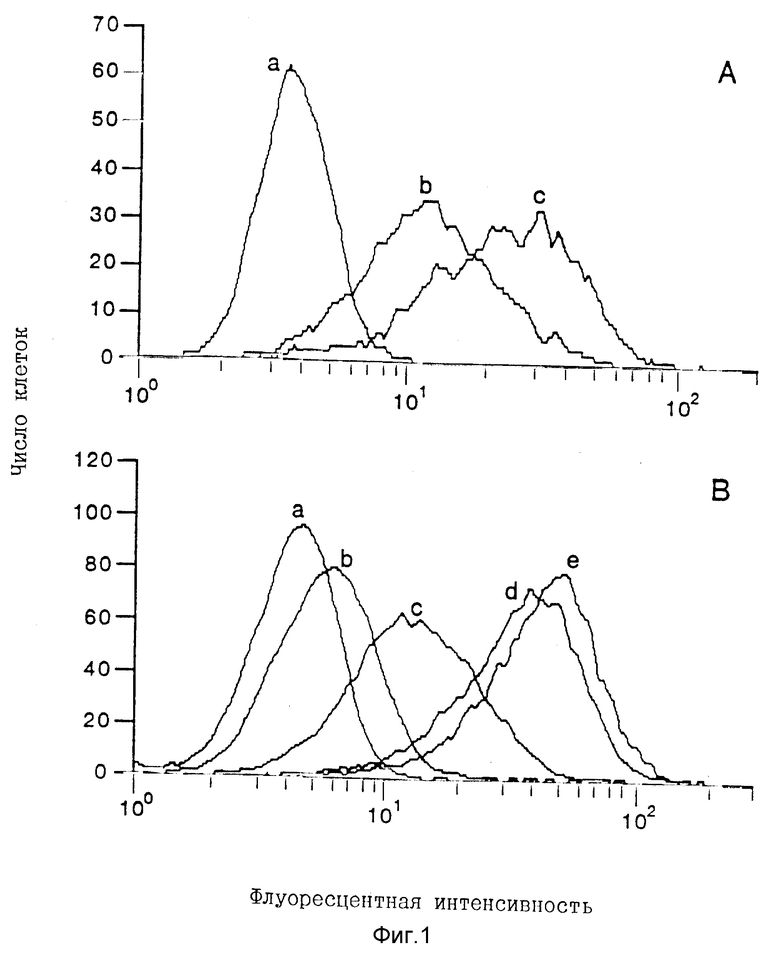
| J | |||
| Jmmunology | |||
| Циркуль-угломер | 1920 |
|
SU1991A1 |
| J | |||
| Jmmunology | |||
| Пуговица для прикрепления ее к материи без пришивки | 1921 |
|
SU1992A1 |
| J | |||
| Jmmunology | |||
| Способ изготовления фанеры-переклейки | 1921 |
|
SU1993A1 |
