Настоящее изобретение относится к новым природно-процессируемым RA антигенным пептидам, которые являются предположительными маркерами эрозивного и неэрозивного RA. Указанные антигенные пептиды представлены HLA-DR молекулами класса II основного комплекса гистосовместимости человека. Кроме того, настоящее изобретение относится к антигенным полипептидам, связанным с молекулами класса II основного комплекса гистосовместимости, а также к антителам, реагирующим с указанными антигенными пептидами, нуклеиновым кислотам, кодирующим указанные антигенные пептиды, конструкциям нуклеиновых кислот и клеткам-хозяевам для экспрессии указанных антигенных пептидов. Антигенные пептиды, заявленные в соответствии с настоящим изобретением, а также полипептиды, из которых их получают, могут быть использованы в качестве маркеров для диагностики RA и в качестве терапевтических агентов в составе анти-RA вакцин.
Ревматоидный артрит (RA, от англ. Rheumatoid Arthritis), обычно обозначаемый термином хронический полиартрит, представляет собой системное аутоиммунное заболевание, которое является одной из самых тяжелых форм воспалительного поражения суставов (Feldmann, M. et al., Cell 85 (1996) 307-310; Dedhia, H.V. & DiBartolomeo, A., Critical care clinics 18 (2002) 841-854). В типичных случаях RA вызывает суставную боль, приводит к деформации и выраженной ригидности суставов. Данное заболевание может манифестировать и не с суставного синдрома, особенно у пациентов, позитивных по аутоантителу, обозначаемому как "ревматоидный фактор" (RF, от англ. Rheumatoid Factor) (Mageed, R.A., in: van Venrooij, W.J. & Maini, R.N. eds., Manual из biological markers of disease, Kluwer Academic Publishers (1996) 1-18). RA чаще встречается у представителей белой европеоидной расы, при этом на предрасположенность к RA влияют генетические факторы и факторы окружающей среды. Указанные факторы оказывают существенное влияние на возникновение и прогрессирование этого аутоиммунного заболевания. Приблизительно 4% общей популяции имеют повышенную генетическую предрасположенность к RA, из которых у 20% (приблизительно 1% общей популяции) в результате воздействия неизвестных ненаследственных факторов развивается RA. Кроме того, было показано, что RA значительно чаще встречается у женщин; так, риск заболеть у женщин в три раза выше, чем у мужчин, что свидетельствует о том, что половые гормоны также могут быть вовлечены в патогенез.
На начальных стадиях RA прогрессирует медленно. Наиболее часто встречающимися ранними симптомами являются потливость ладоней, утренняя скованность в суставах пальцев и симметричное поражение суставов воспалительного характера. Кроме того, могут появляться ревматоидные узелки, что является признаком поражения внесуставных тканей. Упрощенная схема патогенеза RA может быть представлена следующим образом: иммунная система продуцирует антитела к здоровым тканям. Указанные антитела атакуют суставной хрящ, что приводит к воспалению сустава и, на более поздних стадиях, к его разрушению. Это разрушение стимулирует иммунную систему, которая начинает продуцировать еще больше аутоантител. Кроме того, происходит образование цитокинов, таких как фактор некроза опухолей альфа (TNF-α) и интерлейкин-1 (IL-1), которые дополнительно усиливают воспалительную реакцию (Houssiau, F.A., Clin Rheumatol 14 Suppi 2 (1995) 10-13). В ответ на инфильтрацию иммунными клетками, такими как макрофаги и Т-клетки, начинается отек синовиальной оболочки. Указанные клетки активно участвуют в механизме клеточной гибели и поддерживают воспаление в суставе (Fox, D.A., Arthritis Rheum 40 (1997) 598-609; Choy, E.H. & Panayi, G.S., N Engi J Med 344 (2001) 907-916). Все эти процессы формируют порочный круг, включающий образование аутоантител, воспаление и разрушение сустава.
В типичных случаях RA протекает хронически, у 85-90% всех пациентов наблюдается медленное прогрессирование заболевания. Злокачественные формы заболевания, приводящие к полной потере функции суставов вплоть до инвалидности, встречаются в 10-15% случаев. При таких формах RA у пациентов отмечается постоянное воспаление суставов и наличие ревматоидных узелков. Такие пациенты страдают от мучительной хронической боли, а постоянное воспаление приводит к выраженной ригидности суставов пальцев рук и необратимым деформациям и вывихам суставов.
Диагностика
Появляется все больше доказательств того, что терапевтическое воздействие на ранних стадиях заболевания может предотвратить разрушение суставов (Egmose, С.et al., J Rheumatol 22 (1995) 2208-2213; Van der Heide, A. et al., Ann Intern Med 124 (1996) 699-707). С тех пор как лечение модифицирующими заболевание антиревматическими лекарственными средствами (DMARDs, от англ. Disease-Modifying Antirheumatic Drugs) стало считаться оправданным и удалось достичь удовлетворительных соотношений риск/преимущество и цена/эффективность, стало обязательным сразу после начала заболевания проводить дифференциальную диагностику RA и других форм артрита (Kirwan, J.R. & Quilty, В., Clin Exp Rheumatol 15 (1997) 15-25). Диагноз устанавливается на основании четких критериев, выявленных из анамнеза заболевания, клинического обследования и лабораторных данных. Американская Ассоциация Ревматизма опубликовала список критериев, которые позволяют собрать объективные доказательства в пользу RA (Amett, F.C., et al., Arthritis Rheum 31 (1987) 315-324). Однако до настоящего времени не существует ни одного теста, специфичного для RA. Для постановки диагноза RA используются некоторые биологические и биохимические маркеры, например С-реактивный белок (CRP, от англ. C-reactive Protein), скорость оседания эритроцитов (СОЭ), антинуклеарные антитела (ANA, от англ. Antinuclear Antibody) и RF. Однако указанные маркеры не являются специфичными, поскольку они могут появляться и при других воспалительных или аутоиммунных заболеваниях. В частности, RF представляет собой аутоантитело, которое присутствует в сыворотке приблизительно у 50% больных RA. С тех пор как было доказано, что RF может выявляться в сыворотке и при других воспалительных заболеваниях, таких как синдром Шегрена, эндокардит или хронический гепатит, RF перестал использоваться как диагностический маркер RA. Перечисленные выше биологические и биохимические маркеры сами по себе имеют диагностическое значение и, кроме того, могут использоваться для определения степени активности заболевания и прогноза, а также для лечения и классификации пациентов с RA (Nakamura, R.M., J Clin Lab Anal 14 (2000) 305-313).
Недавно был разработан набор критериев диагностики RA, который включает клинические и биохимические показатели, позволяющие на ранних стадиях заболевания провести дифференциальную диагностику между самоограничивающимся RA, персистирующим эрозивным RA и персистирующим неэрозивным RA (Visser, H. et al., Arthritis Rheum 46 (2002) 357-365). Самоограничивающаяся форма RA характеризуется естественной ремиссией: при осмотре пациентов не отмечается признаков артрита в течение определенного периода времени. Эрозивный артрит устанавливают на основании наличия эрозивных изменений на рентгенограммах кисти и/или стопы. В частности, применение антител, которые распознают циклические цитруллинированные пептиды, может оказаться эффективным, а также позволяет предположить важную роль цитруллинированных антигенов в патогенезе эрозивного RA (Schellekens, G.A. et al., J Clin Invest 101 (1998) 273-281; Vincent, С.et al., J Rheumatol 25 (1998) 838-846). Ранняя диагностика эрозивного RA позволяет как можно раньше начать применение DMARDs, что будет способствовать своевременному контролю за патологическим процессом и улучшению исходов заболевания (Symmons, D.P.M. et al., J Rheumatol 25 (1998) 1072-1077; Anderson, J.J. et al., Arthritis Rheum 43 (2000) 22-29). Аналогично, ранняя диагностика самоограничивающейся формы RA и неэрозивного RA позволяет предотвратить ненужную медикаментозную терапию потенциально токсичными лекарственными препаратами (Fries, J.F. et al., Arthritis Rheum 36 (1993)297-306).
Терапия
Целью противовоспалительной терапии является облегчение болевого синдрома для того, чтобы улучшить повседневную активность пациентов. В настоящее время полностью вылечить RA невозможно, но с помощью современных методов лечения можно замедлить прогрессирование заболевания или даже остановить его развитие. Вследствие индивидуальных различий каждому конкретному пациенту требуется своя индивидуально подобранная терапия, и, как упоминалось выше, желательно поставить диагноз как можно раньше. Терапия RA является комплексной и заключается в пожизненном применении лекарственных средств, а также физиотерапии и лучевой терапии. Основными терапевтическими агентами для лечения RA являются DMARDs (например, метотрексат, сульфалазин, гидроксихлорохин, лефлуномид, азатиоприн), кортизон, нестероидные противовоспалительные средства (НПВС) или моноклональные антитела к провоспалительным цитокинам TNF-α, IL-1β или их рецепторам (http://rheuma-online.de). Указанные лекарственные агенты имеют общий механизм действия, который заключается в уменьшении воспалительной реакции путем подавления иммунной системы. Основными недостатками являются их низкая специфичность по отношению к RA, побочные эффекты и неспособность воздействовать на основную причину развития RA.
Аутоиммунизация
Аутоиммунизация запускается тогда, когда инициируется специфический адаптивный иммунный ответ, направленный против собственных антигенов (аутоантигенов) и проявляющийся на начальном этапе образованием автономных Т или В клеток. Естественным результатом адаптивного иммунного ответа, направленного против чужеродного антигена, является элиминация указанного антигена из организма. В том случае, когда адаптивный иммунный ответ развивается на собственный антиген, указанный антиген в большинстве случаев не полностью элиминируется из организма и поддерживает постоянный иммунный ответ. Как следствие, эффекторные механизмы иммунной защиты приводят к хроническим воспалительным процессам в тканях. Механизмы повреждения тканей при аутоиммунных заболеваниях по существу не отличаются от механизмов, которые работают при иммунной защите организма и при развитии реакций гиперчувствительности. И хотя в настоящее время не очень понятно, что является триггерным фактором для аутоиммунизации, недавно были обобщены факторы, которые, по всей видимости, вносят свой вклад в развитие аутоиммунных заболеваний и "отбор" мишеней для аутоантигенов (Marrack, P. et al., Nat Med 7 (2001) 899-905).
Аутоиммунные заболевания контролируются свойствами определенных генов каждого индивидуального фактора или фактора внешней среды. Гены хозяина определяют предрасположенность к аутоиммунизации по крайней мере на трех уровнях. Во-первых, некоторые гены влияют на общую реактивность иммунной системы и, следовательно, могут предопределять развитие у индивидуума определенных аутоиммунных заболеваний или даже развитие нескольких заболеваний разных типов. Во-вторых, такая измененная иммунореактивность приводит к образованию определенных аутоантигенов, которые регулируются генами, влияющими на распознавание антигенных пептидов Т-клетками. В-третьих, другие гены определяют способность тканей-мишеней регулировать иммунную атаку, в частности, оказывая влияние на активность эффекторных клеток иммунной системы, которые инициируют иммунную агрессию. Последние два набора генов определяют, какие антигены будут являться мишенями для аутоантител и, следовательно, какие органы будут подвергаться атаке и повреждаться.
Кроме того, сигналы окружающей среды влияют на развитие аутоиммунизации на тех же трех уровнях, оказывая воздействие на общую реактивность иммунной системы, антиген-специфичность и состояние потенциальных тканей-мишеней. И, наконец, существует перекрестное взаимодействие факторов окружающей среды и генетических факторов.
Основной комплекс гистосовместимости (МНС, от англ. Main Histocompatibility Complex)
Популяционные исследования, генотипирование и современные достижения молекулярной биологии показали, что носительство определенных генов, кодирующих белки основного комплекса гистосовместимости (МНС), является высоким риском развития RA (Stastny, P., Tissue Antigens 4 (1974) 571-579; Wordsworth, P. et al., PNAS 86 (1989) 10049-10053; Wordsworth, P.&Bell, J., Springer Semin Immunopathol 14 (1992) 59-78). В частности, носительство определенных аллелей HLA класса II, таких как HLA-DRB1*0101, *0401, *0404 и *0405, некоторыми этническими группами повышает восприимчивость к RA (Reveille, J., Curr Opin Rheumatol 10 (1998) 187-200). Так, более чем у 90% RF-позитивных пациентов выявляется носительство одного из указанных предрасполагающих аллелей. HLA молекулы класса II представляют собой МНС-поверхностные белки, которые связывают антигенные пептиды внутри клетки и экспонируют их на поверхности антиген-презентирующих клеток для взаимодействия с рецепторами CD4+Т-лимфоцитов, что инициирует клеточный иммунный ответ (Banchereau, J.& Stemman, R.M., Nature 392 (1998) 245-252). RA-ассоциация определенных HLA молекул класса II и наличие большого количества активированных CD4+Т-клеток, по-видимому, является схемой, запускающей развитие заболевания, при которой ассоциированные с заболеванием HLA-DR молекулы экспонируют связанные с заболеванием (например, синовиальные) аутоантигены и вызывают стимуляцию и размножение синовиальных Т-клеток, которые в дальнейшем поддерживают воспалительный процесс (Striebich, С.С.et al., J Immunol 161 (1998) 4428-4436).
HLA-DR (коротко: DR) белки класса II МНС представляют собой гетеродимеры, состоящие из мономорфных α-цепей и чрезвычайно полиморфных β-цепей, которые связывают белковые антигены в пептидсвязывающем желобке. Указанный желобок обычно имеет четыре основных кармана для связывания боковых цепей в соответствующих положениях 1, 4, 6 и 9 пептида (Stem, L.J. et al., Nature 368 (1994) 215-221). Аллельные варианты HLA молекул класса II обладают различной способностью связывать антигенные пептиды. Это объясняет тот факт, что индивидуумы, отличающиеся по HLA аллелям, имеют различный набор антигенных пептидов, что обусловливает различия в интенсивности иммунного ответа (Messaoudi, I. et al., Science 298 (2002) 1797-1800).
Пептиды, которые связываются МНС молекулами класса II, обычно длиннее и более гетерогенные по размеру (11-25 аминокислот), чем пептиды, которые связываются МНС молекулами класса I (8-10 аминокислот). Это различие обусловлено тем, что пептидсвязывающий желобок белков класса II является открытым и, в то время как пептиды удерживаются в центре желобка, их концы могут распространяться за его пределы различным образом (Jones, E.Y., Curr Opin Immunol 9 (1997) 75-79). В итоге молекулы класса II обычно связывают набор перекрывающихся пептидов, которые имеют общую центральную последовательность, называемую "Т-клеточный эпитоп", но различаются по длине.
Более чем десять лет назад было обнаружено, что DRβ цепи, кодируемые RA-ассоциированными DRB1 аллелями, несмотря на их полиморфизм, все имеют общий отрезок идентичных или почти идентичных аминокислот в положениях 67-74, известный как "перекрестный эпитоп" (Gregersen, Р.К. et al., Arthritis Rheum 30 (1987) 1205-1213). До того времени, как было установлено, что иммунитет к аутоантигенам является центральным звеном в патогенезе RA, существовало предположение о том, что перекрестный эпитоп может участвовать в патологическом связывании соответствующих DR молекул с помощью по крайней мере двух механизмов: во-первых, путем отбора релевантных аутоантигенных пептидов для экспонирования, и, во-вторых путем отбора подходящих аутореактивных специфичных Т-клеток во время онтогенеза. Трехмерная пространственная структура DR молекул также подтверждает, что перекрестный эпитоп располагается в центре α-спирали, фланкируя пептидосвязывающий желобок с одной стороны (Stem, L.J. et al., Nature 368 (1994) 215-221). Следовательно, перекрестный эпитоп стратегически расположен таким образом, что он может взаимодействовать как со связанным пептидом, так и с Т-клеточным рецептором.
Однако одним из неразрешенных вопросов ревматологии является вопрос о том, какие артритогенные антигены и эпитопы в организме человека являются триггерными и запускают развитие RA. Хотя в настоящее время были идентифицированы аутоантитела различной специфичности в сыворотке и синовиальной жидкости пациентов, до сих пор неясно, действительно ли антитела, которые высвобождаются при разрушении хряща, запускают патологический процесс или же их высвобождение просто является следствием воспаления (Corrigall, V.M. & Panayi G.S., Crit Rev Immunol 22 (2002) 281-293). Кроме того, очень трудно установить патогенетические механизмы, при которых антиген присутствует в организме, включая суставы, но патологический процесс при этом направлен главным образом или исключительно на суставы.
Аутоантигены
При исследовании сыворотки или, реже, Т-клеток, выделенных от больных с хроническим RA, удалось определить большое число возможных RA аутоантигенов. Одним из точно установленных суставоспецифичных антигенов, который, предположительно, относится к DR молекулам, является коллаген типа II (CII), основной белок суставного хряща. Антитела к CII в высоких концентрациях обнаруживают в сыворотке и суставной жидкости пациентов с RA, хотя до сих пор не ясно, являются ли указанные анти-CII-антитела патогенными при RA (Banerjee, S. et al., Clin Exp Rheumatol 6 (373-380). Snowden et al. показали, что периферические Т-клетки крови, выделенные из организма пациентов с RA, начинают пролиферировать в присутствии CII и наиболее интенсивно у пациентов с анти-CII-антителами. Однако указанный ответ наблюдался только у 50% пациентов (Snowden, N. et al., Rheumatology 40 (1997) 1210-1218). На моделях мышей иммунизация CII индуцировала развитие артрита у мышей, экспрессирующих аллели DRB 1*0401 и *0101 МНС класса II (Rosloniec, E.F., et al., J Exp Med 185 (1997) 1113-1122; Rosloniec, E.F. et al., J Immunol 160 (1998) 2573-2578). Иммунодоминатный эпитоп в обоих *0401 и *0101 аллелях трансгенных мышей был направлен по отношению к пептидам с остатками 261-273 CII человека (Fugger, L. et al., Ew J Immunol 26 (1996) 928-933). Аналогичный эпитоп CII демонстрировал способность стимулировать Т-клеточный ответ у пациентов с RA, в особенности на ранних стадиях болезни. Т-клетки синовиальной жидкости обладали наибольшей реактивностью (Kirn, H.Y. et al., Arthritis Rheum 42 (1999) 2085-2093).
Хотя были обнаружены другие хрящевые белки, которые потенциально могут представлять собой RA антигены, ОР4-связывающие эпитопы были обнаружены только у хрящевого гликопротеина 39 человека (HCgp39). Указанный белок секретируется синовиальными клетками и суставными хондроцитами и его количество в плазме и суставной жидкости увеличивается во время воспаления (Vos, К. et al., Ann Rheum Dis 59 (2000) 544-548). Как и в случае с CII, введение HCgp39 индуцирует развитие артрита у мышей. Также у пациентов с RA определяли иммунный ответ Т-клеток периферической крови на введение HCgp39 (Verheijden, G.F. et al., Arthritis Rheum 40 (1997) 1115-1125). Доминантный эпитоп, который распознают Т-клетки у пациентов-носителей DR4, находится между остатками 263-275 и идентичен иммунодоминантному эпитопу, обнаруживаемому у DRB 1*0401-трансгенных мышей после иммунизации нативным HCgp39 (Соре, А.Р. et al., Arthritis Rheum 42 (1999) 1497-1507). Хотя иммунный ответ на этот белок и не является специфичным для этого заболевания, он коррелирует с активностью заболевания у пациентов с RA (Baeten, D. et al., Arthritis Rheum 43 (2000) 1233-1243). Антитела к HCgp39, однако, определяются в сыворотке пациентов с воспалительными заболеваниями, такими как воспалительные заболевания кишечника и системная красная волчанка (СКВ), хотя и в меньших концентрациях, чем при RA.
При попытке выявить антиген-специфичные Т-клетки у больных RA, для определения синовиальных CD4+Т-клеток, реагирующих с CII или HCgp39 у DR4+пациентов, использовали растворимые пептидные DR4 тетрамерные комплексы (Kotzin, B.L. et al., PNAS 97 (2000) 291-296). CII-DR4 комплекс специфично связывается с CII-реактивными Т-клеточными гибридомами, но не с определяемой фракцией синовиальных CD4+клеток. Практически аналогичные результаты были получены с HCgp39-DR4 комплексом, что позволило сделать предположение о том, что основные олигоклональные популяции CD4+Т-клеток, присутствующие в суставах при RA, не являются специфичными для доминантного CII и HCgp39 доминант, описанных выше.
В целом, несмотря на наличие некоторых существенных признаков ассоциации CII и HCgp39 с патогенезом RA, доказательств того, что они являются основными антигенами при RA, недостаточно. Прямых доказательств того, что CII или HCgp39 белки презентируются рестриктированными по МНС класса II антиген-представляющими клетками с последующей стимуляцией и активацией синовиальных CD4+Т-клеток, явно недостаточно. Кроме того, большой проблемой при использовании модельных животных является то, что их восприимчивость к RA до конца неизвестна, поскольку CII-индуцированный артрит у иммунизированных крыс или мышей по многим параметрам отличается от RA.
Природно-процессируемые пептиды, ассоциированные с МНС класса II
Альтернативный подход для идентификации RA-специфичных аутоантител и Т-клеток заключается в применении анализа последовательностей природно-процессируемых пептидных антигенов, которые связываются с молекулами МНС класса II. При помощи моноклональных антител молекулы МНС класса II, определяющие предрасположенность к RA, могут быть выделены из соответствующих клеток. RA-ассоциированные пептидные антигены могут быть элюированы кислотами из очищенных HLA молекул класса II. Смесь небольших белков может быть разделена с помощью HPLC, а белковые последовательности могут быть определены с помощью секвенирования по Эдману или масс-спектрометрии. Поскольку способы очистки белков и методики секвенирования имеют свои ограничения, до настоящего времени удалось только выделить белковые последовательности из молекул МНС, изолированных из культивированных В-клеточных линий или большого объема ткани, и осуществить анализ лишь некоторых белков, присутствующих в большом количестве (Kropsher et al., J. Exp.Med. 175 (1992) 1799-1803; Chicz, R.M. et al., J Exp Med 178 (1993) 27-47). Создание высокоразрешающих микрокапиллярных колонок для HPLC и более чувствительных масс-спектрометров позволило осуществлять более эффективный анализ МНС-связанных белков (Dongre, A.R. et al., Eur J Immunol 31 (2001) 1485-1494; Engelhard, V.H. et al., Mol Immunol 39 (2002) 127-137).
В настоящем изобретении для исследования пептидного антигенного набора HLA-DR4 молекул, выделенных из аутологичных дендритных клеток (DCs, от англ. Dendritic Cells), которые предварительно обрабатывали сывороткой или синовиальной жидкостью пациентов с RA, использовали методику выделения модифицированных пептидов и их секвенирования. Основным достоинством этой современной методики является использование DCs человека, которые участвуют в RA-ассоциированном процессинге антигенов и их презентации, а не моделей трансгенных животных или искусственных В-клеточных линий.
DCs в большом количестве присутствуют в синовиальной жидкости и тканях при RA и происходят из циркулирующих незрелых предшественников (Thomas, R. et al., J Immunol 152 (1994) 2613-2623). Они являются наиболее активными антиген-представляющими клетками, которые экспрессируют большое количество МНС молекул и разнообразные вспомогательные молекулы (Mellman, I. et al., Trends Cell Biol 8 (1998) 231-237). Последние исследования в этой области показали, что ex vivo дифференцирующиеся DCs человека и макрофаги, которые фенотипически аналогичны антиген-представляющим клеткам синовиальной жидкости из пораженных RA суставов, обладают способностью генерировать и презентировать иммунодоминантные эпитопы из СИ и HCgp39 (Tsark, B.C. et al., J Immunol 169 (2002) 6625-6633). DC обладает способностью праймировать CD4+хелперные Т-клетки и эффективно активировать цитотоксические CD8+Т-клетки (Ridge, Т. et al., Nature 393 (1998) 474-478). Следовательно, пептиды, связывающиеся с МНС молекулами класса II и презентируемые DCs, играют первостепенную роль в патогенезе заболеваний, при которых в иммунный ответ вовлекаются Т-клетки.
Кроме того, проблема, связанная с отсутствием достаточного количества информации, касающейся RA антигенных пептидов, ассоциированных с МНС класса II, может быть решена путем получения новых природно-процессируемых RA антигенных пептидов, ассоциированных с МНС класса II, и полипептидов, из которых можно получать указанные антигенные пептиды, и их использования в качестве маркеров RA.
Настоящее изобретение относится к новым природно-процессируемым антигенным пептидам, которые являются предположительными маркерами эрозивного и неэрозивного RA. Указанные антигенные пептиды представлены HLA-DR молекулами МНС класса II, полученными из дендритных клеток, которые обрабатывают сывороткой или синовиальной жидкостью пациентов с установленным диагнозом эрозивного или неэрозивного RA. Антигенные пептиды МНС класса II, заявленные в соответствии с настоящим изобретением, включают: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка, выбранную из группы, включающей SEQ ID NOs. 49-57 и SEQ ID NOs. 103-122, или (б) по крайней мере аминокислотную последовательность пептидсвязывающего участка, выбранную из группы, включающей SEQ ID NOs. 49-57 и SEQ ID NOs. 103-122, с дополнительными N-и С-концевыми фланкирующими последовательностями соответствующей последовательности, выбранной из группы, включающей SEQ ID NOs. 1-39 или SEQ ID NOs. 58-102; причем заявленную последовательность получают из индуцируемой γ-интерфероном лизосомальной тиолредуктазы, интегрина бета-2, фосфатидилинозитол-4,5-бифосфат-3-киназы; активатора плазминогена урокиназного типа, участка V-III тяжелой цепи иммуноглобулина (Ун2б), белка DJ-1, аполипопротеина В-100, 26S протеасомной не АТФ-азной регуляторной субъединицы 8, рецептора интерлейкина 1, фибромодулина, GM-CSF/IL-3/IL-5 рецептора, сортирующего нексина 3, тяжелой цепи Н4 ингибитора интер-α-трипсина, комплемента С4, комплемента C3 (α-цепь), комплемента C3 (β-цепь), протеина 3, подобного обогащенному глутаминовой кислотой связывающему SH3-домен протеину, индуцируемого интерлейкином-4 протеина 1, гемопексина, Hsc70-интерактивного протеина, константной цепи (Ii), отвечающего на протеин 2 рецептора ретиноевой кислоты, фибронектина, катепсина В, трипептидил-пептидазы II, легумаина, рецептора фактора активации тромбоцитов, поли-альфа-2,8-сиалилтрансферазы и ras-родственного пептида Rab-11B. Настоящее изобретение также относится к указанным антигенным пептидам и белкам, из которых их получают, и их использованию в качестве маркеров эрозивного и неэрозивного RA. Кроме того, настоящее изобретение относится к указанным антигенным пептидам, связанным с молекулами МНС класса II, антителам, реагирующим с указанными антигенными пептидами, нуклеиновым кислотам, кодирующим указанные антигенные пептиды, и конструкциям нуклеиновых кислот, клеткам-хозяевам и способам экспрессии указанных антигенных пептидов. Также настоящее изобретение относится к способам выделения и идентификации RA антигенных пептидов.
Фиг.1: Схематическое изображение анализа тканевых образцов, опосредованного дендритными клетками (DC-опосредованный анализ). Дендритные клетки (DCs), наиболее специализированные антиген-представляющие клетки (APCs, от англ. Antigen-Presenting Cells), обрабатывают средой, содержащей антиген (например, синовиальной жидкостью), при оптимальных условиях для захвата и процессинга антигена. В качестве контроля используют DCs, культивируемые при тех же условиях, но в отсутствие антигенов синовиальной жидкости. После созревания DCs нагруженные антигеном молекулы МНС класса II очищают и соответствующие антигенные пептиды, ассоциированные с МНС класса II, выделяют и идентифицируют.
Фиг.2А: Хроматограмма основного пика (ION-TRAP MS) антигенных пептидов, ассоциированных с МНС класса II, которые были выделены из дендритных клеток, предварительно обработанных сывороткой больного RA. Пептиды непосредственно элюируют с RP-C18-HPLC колонки в масс-спектрометр с ионной ловушкой для немедленной MS/MS идентификации. Числа обозначают время удержания (наибольшие величины) и молекулярную массу (наименьшие величины) наиболее явных белковых пиков в смеси в соответствующее время.
Фиг.2 Б: ION-TRAP MS спектр антигенных пептидов за время удержания, составляющее 65.4 мин. Отмеченный пик дополнительно фрагментируют и согласовывают с двухзарядной ионной формой пептида из ингибитора интеральфа-трипсина ITIH4 (см. Таблицу 3).
Фиг.2В: ION TRAP MS/MS спектр двухзарядного пептидного иона при m/z, составляющем 977.1. Фрагментируемые массы вместе с массой родительского иона анализируют относительно невыборочной базы данных человека с использованием алгоритма SEQUEST. Найденная последовательность MPKNVVFVIDKSGSMSGR (однобуквенный код) соответствует доминантному эпитопу ITIH4 (271-288) ингибитора интер-альфа-трипсина. Положения заданных серий N-концевых и С-концевых Y-ионов маркированы.
Фиг.3: Суммарные данные по дифференциальной связывающей активности тестируемых вероятных RA антигенов относительно связывания с аллелем HLA-DRB1*0401. Предполагаемый HLA-DRB1* 0401-связывающий участок помещен в серую рамку. Как меру аффинности, определяют концентрацию пептида, необходимую для снижения связывания заданного количества биотинилированного НА (307-319) пептида при 50% (1С5о) при конкурентом связывании. Реципрокная величина (1/1С5о) коррелирует с аффинностью пептида. В качестве репортерной молекулы в исследовании используют НА (307-319) пептид, гемагглютинин вируса гриппа (Rothbard, J.B. et al., Cell 52 (1988) 515-523).
Антигенные пептиды, заявленные в соответствии с настоящим изобретением, представляют собой пептиды, ассоциированные и представляемые молекулами МНС, в связи с чем они обладают способностью активировать или повышать чувствительность Т-клеток. Антигенные пептиды, представляемые молекулами МНС класса II, таким образом, являются антигенными пептидами, ассоциированными с молекулами МНС класса II, или антигенными пептидами МНС класса II; при этом антигенные пептиды, представляемые молекулами МНС класса I, являются антигенными пептидами, ассоциированными с молекулами МНС класса I, или антигенными пептидами МНС класса I.
Пептиды, получаемые из белков, закодированных в геноме организма или АРС, обозначают как "аутопептиды". Предполагается, что основная функция аутопептидов, представляемых DCs в периферических органах лимфатической системы, заключается в обеспечении толерантности Т-клеток к аутопептидам. Толерантность подразумевает несостоятельность иммунного ответа на антиген; в том случае, когда такой антиген образуется в тканях самого организма, толерантность обозначают как аутотолерантность.
Антигены, получаемые из собственного организма, называют "аутоантигенами". Адаптивный иммунный ответ, направленный против аутоантигенов, называют аутоиммунным ответом. Аналогично, адаптивный иммунитет, специфичный по отношению к аутоантигенам, называют аутоиммунитетом. Термин аутореактивность объединяет иммунные ответы, направленные против аутоантигенов. RA, предположительно, обусловлен аутоиммунным ответом, в который вовлекаются аутореактивные Т-клетки и/или аутореактивные антитела. К иммунногенным пептидам относятся (без ограничений указанными) антигенные пептиды, способные вызывать или стимулировать клеточный или гуморальный иммунный ответ. Указанные пептиды также могут реагировать с антителами.
Пептиды, получаемые из белков, закодированных в геноме бактерий, вирусов или других чужеродных возбудителей, и отличающиеся от аутопептидов, называют "чужеродными антигенами" или "чужеродными" пептидами. Они обладают способностью вызывать Т-клеточный ответ, направленный против чужеродных белков, из которых они были получены.
RA антигенные пептиды представляют собой аутопептиды, которые выполняют роль аутоантигенов и, как следствие заболевания, ошибочно запускают аутореактивность к собственным тканям организма.
Настоящее изобретение относится к антигенным пептидам МНС класса II, включающим: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка, выбранную из группы, включающей SEQ ID NOs. 49-57 или SEQ ID NOs. 103-122, или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка, выбранную из группы, включающей SEQ ID NOs. 49-57 или SEQ ID NOs. 103-122, с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности, выбранной из группы, включающей SEQ ID NOs. 1-39 или SEQ ID NOs. 58-102. Предпочтительно антигенные пептиды МНС класса II имеют в длину менее 26 аминокислот, более предпочтительно их длина составляет от 11 до 25 аминокислот. Еще более предпочтительно, чтобы длина антигенных пептидов, заявленных в соответствии с настоящим изобретением, составляла от 11 до 19 аминокислот. Наиболее предпочтительно, чтобы в состав антигенного пептида, заявленного в соответствии с настоящим изобретением, входил пептидсвязывающий участок, включающий четыре "якорные" аминокислоты.
Настоящее изобретение относится к антигенным пептидам МНС класса II, включающим: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 49 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 49 с дополнительными С- и N-концевыми фланкирующими последовательностями соответствующей последовательности, выбранной из группы, включающей SEQ ID NOs. 1-3.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, включающим: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 103 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 103 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующих последовательностей SEQ ID NOs. 58 и 59.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, включающим: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 104 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 104 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности SEQ ID NOs. 60.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, включающим: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 105 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 105 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности SEQ ID NOs. 61.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, включающим: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 106 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 106 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности SEQ ID NOs. 62.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, включающим: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 107 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 107 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности SEQ ID NOs. 63.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, включающим: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 50 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 50 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности SEQ ID NOs. 5.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, включающим: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 108 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 108 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующих последовательностей SEQ ID NOs. 64-67.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, включающим: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 109 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 109 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности SEQ ID NOs. 68.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, включающим: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 110 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 110 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующих последовательностей SEQ ID NOs. 69 и 70.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, включающим: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 111 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 111 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности SEQ ID NOs. 72.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, включающим: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 112 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 112 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности SEQ ID NOs. 73.
Антигенные пептиды, ассоциированные с МНС класса II, заявленные в соответствии с настоящим изобретением, получают из индуцируемой γ-интерфероном лизосомальной тиолредуктазы (SEQ ID NOs. 1-3), интегрина бета-2 (SEQ ID NOs. 58-59), фосфатидилинозитол-4,5-бифосфат-3-киназы (SEQ ID NO:60); активатора плазминогена урокиназного типа (SEQ ID NO:61), участка V-III тяжелой цепи иммуноглобулина VH26) (SEQ ID NO:62), белка DJ-1 (SEQ ID NO:63), аполипопротеина В-100 (SEQ ID NOs. 4 и 5), 26S протеасомной не АТФ-азной регуляторной субъединицы 8 (SEQ ID NOs. 64-67), рецептора интерлейкина 1 (SEQ ID NO:68), фибромодулина (SEQ ID NOs.: 69 и 70), GM-CSF/IL-3/IL-5 рецептора (SEQ ID NOs. 71 и 72), сортирующего нексина 3 (SEQ ID NO: 73), тяжелой цепи Н4 ингибитора интер-α-трипсина (SEQ ID NOs.: 6-12), комплемента С4 (SEQ ID NOs.: 13-18), комплемента C3 (α-цепь) (SEQ ID NOs.: 19-23, 74 и 75), комплемента С3 (β-цепь) (SEQ ID NOs. 76 и 77), протеина 3, подобного обогащенному глутаминовой кислотой связывающему SH3-домен протеину (SEQ ID NOs.: 24-27), индуцируемого интерлейкином-4 протеина 1 (SEQ ID NOs.: 28-30), гемопексина (SEQ ID NOs. 31-35 и 78), Hsc70-интерактивного протеина (SEQ ID NOs. 36-39), константной цепи (Ii) (SEQ ID NOs. 79-83), отвечающего на протеин 2 рецептора ретиноевой кислоты (SEQ ID NOs. 84-86), фибронектина (SEQ ID NOs. 87-91), катепсина В (SEQ ID NO: 92), трипептидил-пептидазы II (SEQ ID NOs. 93 и 94), легумаина (SEQ ID NO.: 95), рецептора фактора активации тромбоцитов (SEQ ID NO: 96), поли-альфа-2,8-сиалотрансферазы (SEQ ID NO:97) и ras-родственного пептида Rab-11B (SEQ ID NOs. 98 и 102).
Единичный пептидсвязывающий желобок молекул МНС класса II имеет длину приблизительно 25 А, но в отличие от молекул МНС класса I обе стороны желобка являются открытыми (Stem LJ et al., Nature 1994; 368, 215-221). Таким образом, природно-процессируемые антигенные пептиды, выделенные из молекул МНС класса II человека, имеют минимальную длину, составляющую приблизительно 11 остатков, и могут достигать максимальной длины в 25 остатков (Chicz RM et al., J Exp Med 1993; 178, 27-47).
Стабильность МНС-пептидного взаимодействия обеспечивается более чем дюжиной водородных связей, вовлекающих остов пептида, и комплеменарностью между специфичными карманами связывающего желобка и соответствующим образом расположенными боковыми цепями аминокислот пептида. Аминокислотные остатки пептида, соответствующие определенным карманам, обозначают как "якорные" аминокислоты. С учетом большинства HLA-DR аллелей указанные якорные аминокислоты локализованы в соответствующих положениях Р1, Р4, Р6 и Р9. Комбинация аминокислот в указанных 4 якорных положениях обеспечивает высокостабильное связывание с соответствующим HLA-DR аллельным продуктом и различается от аллеля к алеллю. Пептидсвязывающий участок, как определено в настоящем описании, представляет собой последовательность из девяти аминокислот, в состав которой входят четыре якорные аминокислоты. Пептидсвязывающий участок антигенного пептида МНС класса II, заявленного в соответствии с настоящим изобретением, представлен SEQ ID NO. 49 для пептидов, полученных из индуцируемой интерфероном-γ лизосомальной тиолредуктазы (SEQ ID NOs. l-3); SEQ ID NO. 103 для пептидов, полученных из интегрина бета-2 (SEQ ID NOs. 58 и 59); SEQ ID NO. 104 для пептидов, полученных из фосфатидилинозитол-4,5-бифосфат-3-киназы (SEQ ID NO:60); SEQ ID NO. 105 для пептидов, полученных из активатора плазминогена урокиназного типа (SEQ ID NO: 61); SEQ ID NO. 106 для пептидов, полученных из V-III участка тяжелой цепи иммуноглобулина (VH26) (SEQ ID NO: 62); SEQ ID NO. 107 для пептидов, полученных из DJ-1 белка (SEQ ID NO:63); SEQ ID NO. 50 для пептидов, полученных из аполипопротеина В-100 (SEQ ID NOs. 4 и 5); SEQ ID NO. 108 для пептидов, полученных из 26S протеасомной не АТФ-азной регуляторной субъединицы 8 (SEQ ID NOs. 64 и 67); SEQ ID NO. 109 для пептидов, полученных из рецептора интерлейкина-1 (SEQ ID NO: 68); SEQ ID NO. 110 для пептидов, полученных из фибром одулина (SEQ ID NOs.: 69 и 70); SEQ ID NO. 111 для пептидов, полученных из GM-CSF/IL-3/IL-5 рецептора (SEQ ID NOs. 71 и 72); SEQ ID NO. 112 для пептидов, полученных из сортирующего нексина 3 (SEQ ID NO: 73); SEQ ID NO. 51 для пептидов, полученных из Н4 тяжелой цепи ингибитора интер-α-трипсина (SEQ ID NOs. 6-12); SEQ ID NO. 52 для пептидов, полученных из комплемента С4 (SEQ ID NOs. 13-18); SEQ ID NO. 53 для пептидов, полученных из комплемента С3 (α-цепь) (SEQ ID NOs. 19-23, 74 и 75); SEQ ID NO.113 для пептидов, полученных из комплемента C3 (β-цепь) (SEQ ID NOs. 76 и 77); SEQ ID NO. 54 для пептидов, полученных из протеина 3, подобного обогащенному глутаминовой кислотой связывающему SH3-домен протеину (SEQ ID NOs.: 24-27); SEQ ID NO. 55 для пептидов, полученных из индуцируемого интерлейкином-4 протеина 1 (SEQ ID NOs. 28-30); SEQ ID NO. 56 для пептидов, полученных из гемопексина (SEQ ID NOs. 31-35 и 78); SEQ ID NO. 57 для пептидов, полученных из Hsc70-интерактивного протеина (SEQ ID NOs. 36-39); SEQ ID NO. 114 для пептидов, полученных из константной цепи (Ii) (SEQ ID NOs. 79-83); SEQ ID NO. 115 для пептидов, полученных из отвечающего на протеин 2 рецептора ретиноевой кислоты (SEQ ID NOs. 84-86); SEQ ID NO. 116 для пептидов, полученных из фибронекина (SEQ ID NOs. 87-91); SEQ ID NO. 117 для пептидов, полученных из катепсина В (SEQ ID NO: 92); SEQ ID NO. 118 для пептидов, полученных трипептидил-пептидазы II (SEQ ID NOs. 93 и 94); SEQ ID NO. 119 для пептидов, полученных из легумаина (SEQ ID NO: 95); SEQ ID NO. 120 для пептидов, полученных из рецептора фактора активации тромбоцитов (SEQ ID NO: 96); SEQ ID NO. 121 для пептидов, полученных из поли-альфа-2,8-сиалилтрансферазы (SEQ ID NO:97), и SEQ ID NO. 122 для пептидов, полученных из ras-родственного пептида Rab-11B (SEQ ID NOs. 98 и 102).
Пептидсвязывающий участок также может иметь по крайней мере одну, по крайней мере две, по крайней мере три, по крайней мере четыре или же по крайней мере пять модификаций аминокислотной последовательности и при этом все равно сохранять связывающую способность немодифицированного пептидсвязывающего участка. Предпочтительно модифицированный пептидсвязывающий участок содержит по крайней мере три из четырех якорных аминокислот немодифицированного пептидсвязывающего участка. Аминокислотная модификация может представлять собой консервативную аминокислотную замену, как описано ниже.
Дополнительная энергия связывания обеспечивается водородными связями, в образовании которых участвуют остатки, расположенные кпереди от Р1 якорной аминокислоты и кзади от Р9 якорной аминокислоты. Соответственно, в большинстве природно-процессируемых пептидов нонамерный центральный участок (Р1-Р9) на С- и N-концах фланкирован 3-4 остатками. Следовательно, большинство пептидов имеют в своем составе 15-17 остатков. Более длинные пептиды выступают за пределы желобка, таким образом обеспечивая доступ экзопептидазам, которые отщепляют оба конца.
Итак, антигенные пептиды МНС класса II, заявленные в соответствии с настоящим изобретением, включают а) по крайней мере аминокислотную последовательность пептидсвязывающего участка, выбранную из группы, включающей SEQ ID NOs. 49-57 и SEQ ID NOs. 103-122, или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка, выбранную из группы, включающей SEQ ID NOs. 49-57 и SEQ ID NOs. 103-122, с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности, выбранной из группы, включающей SEQ ID NOs. 1-39 и SEQ ID NOs. 58-102, предпочтительно с N- и С-концевыми фланкирующими аминокислотными последовательностями, обеспечивающими дополнительную энергию связывания.
Предпочтительно антигенные пептиды МНС класса II, заявленные в соответствии с настоящим изобретением, имеют связывающую способность, соответствующую таковой молекул МНС класса II и находящуюся уровне между одной десятой и десятикратной IC50 соответствующего пептида, выбранного из группы, включающей: SEQ ID NOs. 1-39 и SEQ ID NOs. 58-102. Связывающая способность пептида измеряется путем определения концентрации, необходимой для уменьшения связывания меченого репортерного пептида на 50%. Указанная величина называется IC50. Антигенные пептиды МНС класса II, заявленные в соответствии с настоящим изобретением, поддерживают свою связывающую способность по отношению к родственным молекулам МНС класса II до тех пор, пока значения IC50 находятся на уровне между одной десятой и десятикратной IC50 соответствующего заданного пептида.
Поскольку пептидный тримминг каждый раз происходит индивидуальным образом как перед, так и после связывания с пептидсвязывающим желобком, наличие нескольких усеченных вариантов с общим нонамерным коровым участком является общей чертой пептидов, связанных с МНС класса II. Было показано, что С- или N-концевые усеченные варианты одного эпитопа могут индуцировать дивергентные Т-клеточные ответы (Arnold et al., (2002) J. Immunol. 169, 739-749).
Можно указать несколько параметров, которые могут повлиять на относительную избыточность усеченных вариантов определенного эпитопа, например избыток или и целостность родственного антигена, антиген-ассоциированные белки, избыток протеаз, типы доступных протеаз или доступность конкурирующих антигенов и/или пептидов. Поскольку доступность антигена является основным параметром, который может коррелировать с происхождением образца, соотношение определенных усеченных вариантов эпитопа может иметь диагностическое значение.
Пептид, заявленный в соответствии с настоящим изобретением, представляет собой пептид, который либо имеет неприродно-процессируемый эквивалент (например, такой как мутированный пептидный антиген), либо был выделен, то есть был отделен или очищен от компонентов, которые окружают его в естественных условиях, например в тканях, таких как ткань поджелудочной железы, печени, селезенки, яичников, яичек, мышечная ткань, суставная ткань, нервная ткань, гастроинтестинальная ткань, или в естественных жидкостях организма, таких как кровь, сыворотка, синовиальная жидкость или моча. Обычно пептид считается "выделенным", когда препарат, в состав которого входит пептид, заявленный в соответствии с настоящим изобретением, содержит по крайней мере 70% сухого вещества указанного пептида и менее чем 30% белков и природно-существующих органических молекул, с которыми он связан в естественных условиях. Предпочтительно препарат пептида, заявленного в настоящем изобретении, содержит по крайней мере 80%, более предпочтительно по крайней мере 90%, еще более предпочтительно по крайней мере 99% сухого вещества указанного пептида. Поскольку пептид, получаемый с помощью химического синтеза, по своей природе очищен от компонентов, которые окружают его в естественных условиях, такой синтетический пептид считается "выделенным".
Настоящее изобретение также относится к аналогам антигенных пептидов, заявленных в соответствии с настоящим изобретением. Термин "аналог" объединяет пептиды, которые обладают функциональной активностью указанных антигенных пептидов, включая связывающую способность IC50 и способность быть распознанными антителами и клетками иммунной системы. Аналоги имеют практически такое же значение IC50, как и соответствующие пептиды. Термин "аналог" также включает консервативные замены или химические производные пептидов.
Термин "аналог" относится к любому полипептиду, имеющему аминокислотную последовательность, практически идентичную последовательности, представленной здесь, в которой один или более аминокислотный остаток был консервативно заменен остатком, в функциональном отношении идентичным предыдущему; при этом указанный полипептид обладает такой же функциональной активностью, что и пептиды, описанные здесь. Примерами консервативных замен являются замена одной неполярной (гидрофобной) аминокислоты, такой как фенилаланин, тирозин, изолейцин, валин, лейцин или метионин, на другую; замена одной полярной (гидрофильной) аминокислоты на другую, например аргинина на лизин, глутамина на аспарагин, треонина на серин и наоборот; замена одной основной аминокислоты, такой как лизин, аргинин или гистидин, на другую, или замена одной кислой аминокислоты, такой как аспарагиновая кислота или глутаминовая кислота, на другую.
Термин "консервативная замена" также относится к использованию химического производного аминокислоты вместо обычной аминокислоты. Термин "химическое производное" относится к полипептиду, имеющему одну или более аминокислоту с модифицированной с помощью химической реакции функциональной боковой группой. Примерами таких модифицированных молекул являются, например, молекулы, у которых свободные амино-группы были модифицированы с получением аминогидрохлоридов, р-толуол-сульфонильных групп, карбобензокси-групп, t-бутоксикарбонильных групп, хлоруксусных групп, ацетил-групп или формил-групп. Свободные карбоксильные группы могут быть модифицированы для получения солей, метиловых или этиловых эфиров или других эфиров или гидразидов. Свободные гидроксильные группы могут быть модифицированы для получения O-ацил или O-алкил производных. Азот имидазола аминокислоты гистидина может быть модифицирован с получением N-им-бензилгистидина. Также к химическим производным относятся такие белки или пептиды, в состав которых входит одно или более производное любой из двадцати существующих в природе аминокислот. Например: 4-гидроксипролин может замещать пролин; 5-гидроксилизин может замещать лизин; 3-метилгистидин может замещать гистидин; гомосерин может замещать серин, а орнитин или цитруллин может замещать лизин.
Антигенные пептиды МНС класса II, заявленные в соответствии с настоящим изобретением, и белки, из которых их получают, могут использоваться в качестве маркеров RA и в качестве терапевтических агентов в составе анти-RA вакцин. Термин "маркер", используемый здесь, относится к биомолекуле, предпочтительно белку или полипептиду, который выявляется у группы пациентов с установленным заболеванием, например RA, и достигает избытка, который значительно увеличивается или уменьшается по сравнению с контрольной группой.
Маркер, заявленный в соответствии с настоящим изобретением, может использоваться в качестве прогностического маркера, позволяющего прогнозировать восприимчивость к заболеванию, например прогнозировать восприимчивость к RA; в качестве диагностического маркера, позволяющего установить наличие заболевания, например для диагностики RA; в качестве дифференциального диагностического маркера, на основании которого можно проводить дифференциальную диагностику различных форм заболевания, например различных форм RA; в качестве прогностического маркера для прогнозирования исхода заболевания, например для прогнозирования RA, и в качестве ответного маркера для определения эффективности лечения, например в качестве ответного маркера при лечении RA.
Согласно другому варианту осуществления настоящего изобретения антигенные пептиды МНС класса II включают: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка, выбранную из группы, включающей SEQ ID NOs. 49-57 и SEQ ID NOs. 103-122, или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка, выбранную из группы, включающей SEQ ID NOs. 49-57 и SEQ ID NOs. 103-122, с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности, выбранной из группы, включающей SEQ ID NOs. 1-39 и SEQ ID NOs. 58-102, и используются в качестве маркеров эрозивного и/или неэрозивного RA.
Согласно другому варианту осуществления настоящего изобретения антигенные пептиды МНС класса II включают: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 49 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NOs. 49. с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности, выбранной из группы, включающей SEQ ID NOs. 1-3, и используются в качестве маркеров эрозивного и/или неэрозивного RA.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, которые включают: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 103 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 103 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующих последовательностей SEQ ID NOs. 58 и 59 и используются в качестве маркеров эрозивного и/или неэрозивного RA.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, которые включают: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 104 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 104 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности SEQ ID NO. 60 и используются в качестве маркеров эрозивного и/или неэрозивного RA.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, которые включают: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 105 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 105 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности SEQ ID NO. 61 и используются в качестве маркеров эрозивного и/или неэрозивного RA.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, которые включают: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 106 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 106 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности SEQ ID NO. 62 и используются в качестве маркеров эрозивного и/или неэрозивного RA.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, которые включают: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 107 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 107 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности SEQ ID NO. 63 и используются в качестве маркеров эрозивного и/или неэрозивного RA.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, которые включают: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 50 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 50 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности SEQ ID NO. 5 и используются в качестве маркеров эрозивного и/или неэрозивного RA.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, которые включают: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 108 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 108 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующих последовательностей SEQ ID NOs. 64-67 и используются в качестве маркеров эрозивного и/или неэрозивного RA.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, которые включают: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 109 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 109 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности SEQ ID NO. 68 и используются в качестве маркеров эрозивного и/или неэрозивного RA.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, которые включают: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 110 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 110 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующих последовательностей SEQ ID NOs. 69 и 70 и используются в качестве маркеров эрозивного и/или неэрозивного RA.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, которые включают: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 111 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ED NO. 111 c дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности SEQ ID NOs. 72 и 59 и используются в качестве маркеров эрозивного и/или неэрозивного RA.
Кроме того, настоящее изобретение относится к антигенным пептидам МНС класса II, которые включают: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 112 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 112 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности SEQ ID NO. 73 и используются в качестве маркеров эрозивного и/или неэрозивного RA.
Настоящее изобретение, как описано выше, также относится к антигенным пептидам МНС класса II, связанным с молекулами МНС класса II.
Мультимеры (например, димеры, тримеры, тетрамеры, пентамеры, гексамеры или олигомеры) молекул МНС класса II, содержащие ковалентно или нековалентно связанные пептиды, заявленные в соответствии с настоящим изобретением, при их конъюгации с определяемой меткой (например, флуоресцентной молекулой, радиоактивной меткой или ферментом, катализирующим реакцию, результатом которой является образование продукта, который поглощает или испускает световое излучение с определенной длиной волны) могут использоваться для определения у субъекта (например, у человека) количества Т-клеток, несущих на своей поверхности рецепторы, специфичные и, следовательно, связывающие указанные комплексы. Относительно высокое содержание таких Т-клеток может являться диагностическим признаком заболевания или свидетельствовать о том, что Т-клетки участвуют в создании иммунитета к этому заболеванию. Кроме того, постоянный мониторинг относительного количества мультимер-связывающих Т-клеток может быть полезным для контроля за течением заболевания или за эффективностью терапии. Такие методы анализа были разработаны при использовании тетрамеров молекул МНС класса I, включающих пептиды, полученные из вируса HIV-1 или вируса гриппа-15 (Altaian et al., (1996), Science 274:94-96; Ogg et al. (1998) Science 279:2103-21061), поэтому ожидается, что соответствующие полимеры МНС класса II также будут полезными. Указанные комплексы могут быть получены с помощью химического перекрестного связывания очищенных молекул МНС класса II, объединенных в присутствии желаемого пептида, или с помощью модификации уже известных рекомбинантных методик, используемых для получения молекул МНС класса II, содержащих один определенный пептид (Kazono et al. (1994) Nature 369:151-154; Gauthier et al. (1998) Proc. Natl. Acad. Sci. U.S.A. 95:11828-118331). Мономеры молекул МНС класса II указанных полимеров могут представлять собой нативные молекулы, сформированные из полноразмерных альфа- и бета-цепей. Альтернативно, они могут представлять собой молекулы, в состав которых входят внеклеточные домены альфа- и бета-цепей или домены альфа- и бета-цепей, которые формируют "стенки" и "дно" пептидосвязывающего желобка.
Настоящее изобретение также относится к антителу, его фрагментам или производным, которые направлены против описанных выше антигенных пептидов МНС класса II или реагируют с ними. Общая методика получения антител хорошо известна и описана, например, в работе Kohler и Milstein, 1975, Nature 256, 494 или в работе J.G.R. Hurrel, Monoclonal Hybridoma Antibodies: Techniques and Applications, CRC Press Inc., Boco Raron, FL (1982). Антитела могут быть поликлональными или предпочтительно моноклональными или же представлять собой лишь фрагменты антител, такие как F(ab')2, Fab, Fv или scFv. Антитела, заявленные в соответствии с настоящим изобретением, также могут быть гуманизированными (Merluzzi S. et al., (2000) Adv. Clin. Path,, 4(2):77-85) или представлять собой антитела человека (Aujame L. et al., Hum. Antibodies, (1997), 8(4):155-168).
Настоящее изобретение также относится к молекуле нуклеиновой кислоты, кодирующей антигенные пептиды МНС класса II, заявленные в соответствии с настоящим изобретением и включающие: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка, выбранную из группы, включающей SEQ ID NOs. 49-57 и SEQ ID NOs. 103-122, или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка, выбранную из группы, включающей SEQ ID NOs. 49-57 и SEQ ID NOs. 103-122, с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности, выбранной из группы, включающей SEQ ID NOs. 1-39 и SEQ ID NOs. 58-102. Предпочтительно молекула нуклеиновой кислоты представляет собой молекулу ДНК.
Кроме того, настоящее изобретение относится к молекуле нуклеиновой кислоты, кодирующей антигенные пептиды МНС класса II, заявленные в соответствии с настоящим изобретением и связанные с молекулами МНС класса II.
Настоящее изобретение также относится к рекомбинантной конструкции нуклеиновой кислоты, включающей молекулы нуклеиновой кислоты, как описано выше, функционально связанные с вектором экспрессии. Вектор экспрессии, подходящий для использования в соответствии с настоящим изобретением, содержит по крайней мере один контролирующий экспрессию элемент, функционально связанный с нуклеотидной последовательностью, кодирующей антигенный пептид или антигенный пептид, связанный с молекулой МНС класса II. Рекомбинантная конструкция экспрессии может представлять собой ДНК-конструкцию.
Элементы, контролирующие экспрессию, встраивают в вектор для контроля и регуляции экспрессии нуклеотидной последовательности, кодирующей антигенный пептид, заявленный в соответствии с настоящим изобретением. Примерами элементов, контролирующих экспрессию, являются, без ограничений указанными, lac система, оператор и промотор фага лямбда, дрожжевые промоторы и промоторы, полученные из вируса полиомы, аденовируса, ретровируса или SV40. Дополнительные предпочтительные или необходимые оперативные элементы включают, без ограничений указанными, лидерную последовательность, терминирующие кодоны, сигналы полиаденилирования и любые другие последовательности, необходимые или предпочтительные для адекватной транскрипции и последующей трансляции нуклеотидной последовательности в системе хозяина. Специалисту в данной области очевидно, что правильная комбинация необходимых или предпочтительных элементов, контролирующих экспрессию, будет зависеть от выбранной системы-хозяина. Также очевидно, что вектор экспрессии должен включать дополнительные элементы, необходимые для переноса и последующей репликации этого вектора экспрессии, содержащего нуклеотидную последовательность, в систему-хозяин. Примерами таких элементов являются, без ограничений указанными, ориджины репликации и селектируемые маркеры. Специалисту в данной области также понятно, что указанные векторы можно легко сконструировать при помощи стандартных методик ("DNA Isolation and Sequencing", Bruce A. Roe, Judy S. Crabtree and Akbar S. Khan, Published by John Wiley&Sons, 1996); кроме того, они являются коммерчески доступными.
Настоящее изобретение также относится к организму-хозяину или клетке-хозяину, в которую встраивают рекомбинантную конструкцию нуклеиновой кислоты, содержащую молекулы нуклеиновой кислоты, как описано выше, функционально связанную с вектором экспрессии. К клеткам-хозяевам, трансформированным конструкциями нуклеиновой кислоты, заявленными в соответствии с настоящим изобретением, относятся эукариотические клетки, такие как клетки животных, растений, насекомых и дрожжевые клетки, а также прокариотические клетки, такие как E.coli. К способам, с помощью которых конструкция нуклеиновой кислоты, содержащая нуклеотидную последовательность, может быть встроена в клетку, относятся, без ограничений указанными, микроинъекция, электропорация, трансдукция или трансфекция с помощью ДЭАЭ-декстрана, липофекция, применение фосфата кальция и другие способы, хорошо известные специалистам в данной области (Sambrook et al. (1989) in "Molecular Cloning. A Laboratory Manual", Cold Spring Harbor Press, Plainview, New York).
Согласно предпочтительному варианту осуществления настоящего изобретения применяются эукариотические векторы экспрессии, которые функционируют в эукариотических клетках. Примерами таких векторов являются, без ограничений указанными, ретровирусные векторы, вектор экспрессии на основе вируса коровьей оспы, аденовирусные векторы, векторы на основе вируса простого герпеса, векторы на основе вируса птичьей оспы, плизмиды или бакуловирусные векторы переноса. Предпочтительными эукариотическими клеточными линиями являются, без ограничений указанными, COS клетки, СНО клетки, HeLa клетки, NIH/3T3 клетки, 293 клетки (АТСС# CRL15731), Т2 клетки, дендритные клетки, моноциты или В-клетки, трансформированные вирусом Эпштейна-Барр-15.
Антигенный пептид, заявленный в соответствии с настоящим изобретением, может быть получен, например, путем выделения из природного источника (например, элюирования из молекул МНС класса II); путем экспрессии рекомбинантной нуклеиновой кислоты, кодирующей пептид; или с помощью химического синтеза. Пептид, который продуцируется системой клеток, отличной от источника, из которого он происходит в природе, является "выделенным", поскольку он будет отделен от компонентов, которые окружают его в естественных условиях. Рекомбинантный пептид, экспрессируемый организмом-хозяином, может быть получен в виде неочищенного лизата или может быть очищен с помощью стандартных способов, применяющихся для очистки белков и хорошо известных специалистам в данной области, к которым относятся дифференцированная преципитация, хроматография с исключением по размеру, ионообменная хроматография, изоэлектрическое фокусирование, электрофорез в геле, аффинная и иммуноаффинная хроматография и другие. Степень очистки или выделения может быть измерена с помощью любого подходящего способа, например масс-спектрометрии или HPLC анализа. Пептиды могут быть получены синтетическим путем с помощью методик, описанных Memfield, (1986) Science 232: 341-347, and Barany and Merrifield, The Peptides, Gross and Meienher, eds (N.Y., Academic Press). Химический синтез может быть осуществлен в растворе или на твердой фазе или с помощью автоматизированного синтезирующего устройства (Stewart and Young, Solid Peptide Synthesis, 2nd ed., Rockford 111., Pierce Chemical Co. (1984)).
Таким образом, настоящее изобретение также относится к способу получения антигенных пептидов МНС класса II, включающих: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка, выбранную из группы, включающей SEQ ID NOs. 49-57 и SEQ ID NOs. 103-122, или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка, выбранную из группы, включающей SEQ ID NOs. 49-57 и SEQ ID NOs. 103-122, с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности, выбранной из группы, включающей SEQ ID NOs. 1-39 и SEQ ID NOs. 58-102. Указанный способ включает этап культивирования клеток-хозяев, содержащих конструкцию рекомбинантной молекулы нуклеиновой кислоты, как описано выше, в условиях, обеспечивающих экспрессию указанного пептида, и этап выделения пептида из клеток или клеточной среды.
Настоящее изобретение также относится к способу выделения и идентификации RA антигенных пептидов, ассоциированных с МНС класса II, в фемтомолярных количествах, включающему: а) получение незрелых дендритных клеток в количестве, обеспечивающем от 0,1 до 5 мкг молекул МНС класса II; б) обеспечение контакта клеток (а) с сывороткой или синовиальной жидкостью и индицирование созревания дендритных клеток путем добавления TNF альфа; в) выделение комплексов антигенный пептид-молекула МНС класса II из клеток с помощью способов, включающих солюбилизацию клеток и изолирование комплексов молекул МНС класса II с антигенными пептидами путем иммунопреципитации или иммуноаффинной хроматографии; г) отмывание изолированных комплексов молекул МНС класса II с антигенными пептидами водой в пробирке для ультрафильтрации; д) элюирование ассоциированных антигенных пептидов с молекул МНС класса II при температуре 37°С разведенной трифторуксусной кислотой и е) разделение, определение и идентификацию выделенных пептидов с помощью жидкостной хроматографии и масс-спектрометрии. Кроме того, на этапе (е) жидкостная хроматография включает первый этап линейной элюции с материала в обратной фазе объемом, достаточным для элюирования большинства примесей, до элюирования самого пептида. Кроме того, указанный способ может также включать этап (ж) анализа идентифицированных пептидов с помощью известных баз данных и специальной программы, разработанной для осуществления сравнительного анализа полученных данных с многочисленными известными базами данных.
Количество ткани или жидкости организма, необходимое для получения, например, 100 нг молекул МНС класса II, зависит от количества клеток, экспрессирующих МНС класса II и от соотношения экспрессии молекул МНС класса II: например, 100 нг МНС класса II эквивалентны около 2·105 зрелых DCs или 5·106 моноцитов периферической крови или около 5·107 мононуклеарных клеток периферической крови (РВМС), которые могут быть выделены примерно из 50 мл крови.
Для очистки комплексов молекул МНС класса II и антигенных пептидов от клеток или тканей мембраны клеток или тканей необходимо солюбилизировать. Лизис клеток может быть осуществлен с помощью методов, хорошо известных специалистам в данной области, например с помощью циклов замораживания-оттаивания и с помощью детергентов, а также при комбинации указанных способов. Предпочтительным способом лизиса является солюбилизация с помощью детергентов, предпочтительно ТХ-100, NP40, n-октилгликозида, Zwittergent, Lubrol, CHAPS, более предпочтительно ТХ-100 или Zwittergent 3-12. Продукты распада клеток и клеточных ядер необходимо удалить из клеточного лизата, содержащего растворенные комплексы рецептор-пептид, с помощью центрифугирования. Таким образом, комплексы молекул МНС класса II и антигенных пептидов выделяют из клеток с помощью солюбилизации детергентом.
Кроме того, комплексы молекул МНС класса II и антигенных пептидов очищают от клеточного лизата с помощью иммунопреципитации или иммуноаффинной хроматографии. Для иммунопреципитации и иммуноаффинной хроматографии используют антитела, специфичные для молекул МНС класса II и подходящие для применения согласно указанным способам. Предпочтительно используют моноклональные специфические антитела, ковалентно или нековалентно (например, посредством Протеина А) связанные с бусинами, например сефарозными или агарозными бусинами. Панель антител к HLA, используемая в соответствии с указанными способами, включает: анти-HLA-DR антитела: L243, TU36, DA6.147, предпочтительно L243; анти-HLA-DQ антитела: SPVL3, TU22, TU169, предпочтительно TU22 и TU169; анти-HLA-DP антитела В7/21 и анти-HLA-А, В, С антитела W6/32 и В9.12.
Моноклональные антитела, специфичные по отношению к различным молекулам МНС класса II, могут быть получены промышленно (например, Pharmingen, Dianova) или очищены от супернатанта соответствующих клеток гибридомы с помощью Протеин А или Протеин G аффинной хроматографии. Очищенные моноклональные антитела могут быть объединены с помощью различных способов, хорошо известных специалистам в данной области, предпочтительно с помощью ковалентного связывания амино-групп антител с CNBr-активированной сефарозой.
Иммуновыделение молекул МНС может осуществляться путем инкубации бусин с нанесенными на них антителами с клеточным лизатом при вращении в течение нескольких часов или с помощью хроматографии путем пропускания клеточного лизата через микроколонку. Отмывание бусин может осуществляться в пробирках Эппендорф или на микроколонке. Эффективность иммунопреципитации можно оценить с помощью SDS-PAGE и Вестерн-блоттинга при использовании антител, распознающих денатурированные молекулы МНС (анти-HLA-DRальфа; 1В5; анти-HLA класс I; HC10 или НСА2).
Изолированные комплексы молекул МНС класса II и антигенных пептидов перед элюцией отмывают водой или слабосолевым буфером для того, чтобы удалить остатки детергента. В качестве слабосолевого буфера можно использовать Tris, фосфатный или ацетатный буфер с концентрацией от 0,5 до 10 мМ, предпочтительно в концентрации 0,5 мМ. Согласно более предпочтительному варианту осуществления настоящего изобретения комплексы молекул МНС класса II и антигенных пептидов отмывают ультрачистой водой (программируемая шкала), которая традиционно используется для HPLC анализа, предпочтительно ультрачистой водой (программируемая шкала) от MERCK. Этап отмывания можно осуществить с помощью ультрафильтрации. Ультрафильтрацию можно осуществить в пробирке для ультрафильтрации с отсечением по мол. массе 30 кД, 20 кД, 10 кД или 5 кД, предпочтительно 30 кД и объемом пробирки от 0,5 до 1,0 мл (пробирки "Ultrafree"; Millipore). Отмывание в пробирках для ультрафильтрации может осуществляться от 4 до 12 раз, предпочтительно от 6 до 10 раз, в объеме, в 10-20 раз превышающем объем бусин, несущих комплексы рецептор-пептид, предпочтительно в объеме, в 15 раз превышающем объем бусин. Элюированные пептиды могут быть отделены от оставшихся молекул МНС класса II с помощью тех же пробирок для ультрафильтрации. Элюированные пептиды затем могут быть лиофилизированы.
При элюции пептидов с молекул МНС класса II получают комплексную смесь природно-процессируемых пептидов из источника потенциального антигена и из полипептидов внутриклеточного и внеклеточного происхождения. Только после элюции можно разделить пептиды и осуществить анализ их последовательностей.
Антигенные пептиды, заявленные в соответствии с настоящим изобретением, могут быть элюированы с помощью различных способов, хорошо известных специалистам в данной области, предпочтительно, при использовании разведенной кислоты, например разведенного ацетонитрила (Jardetsky TS et al., Nature 1991 353, 326-329), разведенной уксусной кислоты с последующим нагреванием (Radensky AY et al., Nature 1991, 353, 622-626; Chicz RM et al., Nature 1992, 358, 764-768) или разведенной трифторуксусной кислоты при 37°С (Кторзпизег Н et al., J Exp Med 1992, 175, 1799-1803). Более предпочтительно пептиды элюируют при 37°С разведенной трифторуксусной кислотой.
Выделенные антигенные пептиды затем разделяют, детектируют и идентифицируют. Детекция подтверждает, что аминокислотную последовательность отдельных пептидов в смеси выделенных антигенных пептидов можно установить с помощью способов, подходящих для определения и секвенирования пептидов в фемтомолярных количествах. Идентификация позволяет определить, из каких белков или полипептидов были получены антигенные пептиды и какие последовательности этих белков и полипептидов входят в их состав.
На первом этапе комплексная смесь элюированных пептидов может быть разделена любым из известных возможных хроматографических способов, например с помощью хроматографии с обратной фазой, анионообменной хроматографии, катионообменной хроматографии или комбинации указанных способов. Предпочтительно разделение осуществляют с помощью хроматографии с С18-обратной фазой или с помощью обратно-фазовой/катионообменной двухмерной HPLC, обозначаемой как MudPit (Washbum MP et al., Nat BiotechnoL, (2001) 19, 242-247).
Разделение осуществляют с помощью HPLC с использованием микрокапиллярных пробирок из кварцевого стекла, которые соединяют с генератором электрораспыления масс-спектрометра или с устройством для микрофракционирования, которое размещает фракции на планшете для MALDI анализа.
Жидкостная хроматография включает фракционирование пептида с помощью сильного ионообменного материала и гидрофобного материала с обратной фазой. Для элюции пептидов с ионообменного материала и материала с обратной фазой последовательно применяют различные схемы, включающие элюцию с помощью соли и с помощью органических растворителей, например ацетонитрила. Элюцию с материала с обратной фазой осуществляют в несколько этапов линейными градиентами различной длины и наклона. Загрязнение образца, подлежащего фракционированию, может представлять собой любое загрязнение, которое конкурирует с определением белковых пиков в масс-спектрометре. Таким образом, для того чтобы предотвратить совместную элюцию, перед элюцией пептидов необходимо элюировать примеси достаточным объемом растворителя. В зависимости от того, какая колонка используется для жидкостной хроматографии, объем растворителя, достаточный для элюции примесей перед элюцией пептидов, может в 100-200 раз превышать объем колонки.
Подходящими являются различные методики масс-спектрометрии (MS), предпочтительно MALDI-post source decay (PSD) MS или тандемная масс-спектрометрия с ионизацией распылением (ESI-MS, от англ., Electrospray lonization Tandem Mass Spectrometry), наиболее предпочтительно ESI-MS с ионной ловушкой.
Последовательности отдельных пептидов можно определить с помощью методик, хорошо известных специалистам в данной области. Предпочтительно анализ последовательностей осуществляют с помощью фрагментации пептидов и последующим анализом фрагментов на компьютере с использованием алгоритмов, например MASCOT или SEQUEST. Оба указанных компьютерных алгоритма для перекрестного корреляционного анализа теоретически и экспериментально (с помощью тандемной масс-спектрометрии) полученных данных используют базы данных белковых и нуклеотидных последовательностей.
Выделенные и идентифицированные антигенные пептиды, заявленные в соответствии с настоящим изобретением, могут быть подтверждены с помощью МНС-связывающего участка, МНС-связывающей способности и/или с помощью Т-клеточного распознавания.
МНС-связывающий участок
Пептиды, ассоциированные с определенными молекулами МНС (аллельные варианты), имеют общие структурные характеристики, обозначаемые как связывающие участки, необходимые для формирования стабильных комплексов с молекулами МНС. Пептидные лиганды, элюированные с молекул МНС класса I, являются относительно короткими и имеют в своем составе от 8 до 11 аминокислот. Кроме того, 2 или 3 боковые цепи пептида имеют отношение к связыванию. Положение соответствующих боковых цепей аминокислот в аллельных вариантах HLA различно, обычно два так называемых "якорных" аминокислотных остатка располагаются в положениях 2 и 9. Что касается определенного якорного положения, только одна и две аминокислоты в норме могут функционировать как якорные аминокислоты, например лейцин или валин V в положении 2 в случае HLA-A2.
Что касается молекул МНС класса II, длина пептидов варьирует от 11 до 25 аминокислот, поскольку более длинные пептиды связываются лучше, так как оба конца пептидсвязывающего желобка являются открытыми. Большинство HLA молекул класса II имеют вплоть до четырех якорных аминокислотных остатка в соответствующих положениях Р1, Р4, Р6 и Р9, расположенных в нонамерном коровом участке. Указанный коровый участок, однако, располагается на различном расстоянии от N-концевого участка пептида. В большинстве случаев коровому участку предшествуют 2-4 N-концевых остатка. Следовательно, в большинстве HLA молекул класса II Р1 якорные остатки расположены в положениях 3, 4 или 5. Пептиды, элюированные с молекул HLA-DR класса II, имеют общий большой гидрофобный Р1 якорный аминокислотный остаток, представленный тирозином, фенилаланином, триптофаном, метионином, лейцином, изолейцином или валином.
Строго определенный тип якорных аминокислотных остатков и их положение формируют пептидсвязывающий участок, который в настоящее время известен для большинства наиболее часто встречающихся аллельных продуктов HLA класса II. Компьютерная программа, позволяющая подтвердить последовательности пептидов, называется "Tepitope" и предоставляется компанией Vaccinome.
МНС-связывающая способность
Способность пептидов, идентифицированных с помощью способа, заявленного в соответствии с настоящим изобретением, связывать соответствующие молекулы МНС класса II, может быть протестирована с помощью методик, хорошо известных специалистам в данной области, например путем выделения молекул МНС класса II и синтетических пептидов с аминокислотными последовательностями, идентичными тем, которые были идентифицированы с помощью способа, заявленного в соответствии с настоящим изобретением (Kropstnner H et al., J. Exp.Med. 1992; 175, 1799-1803; Vogt AB et al., J Immunol. 1994; 153, 1665-1673; Sloan VS et al., Nature 1995; 375, 802-806). Альтернативно, для верификации идентифицированного эпитопа можно применить клеточное исследование связывания с использованием клеточных линий, экспрессирующих МНС класса II, и биотинилированных пептидов (Amdt SO et al., EMBO J., 2000; 19, 1241-1251).
В обоих случаях соответствующую связывающую способность пептида измеряют путем определения концентрации, необходимой для уменьшения связывания меченого репортерного пептида на 50%. Указанную величину обозначают как IC50. При связывании пептидов со значительной аффинностью по отношению к соответствующим молекулам МНС класса II величины IC50 не превышают величины IC50 для определенных контрольных пептидов более чем в 10 раз.
Аналогичные исследования связывания могут быть осуществлены для того, чтобы протестировать способность пептидов связываться с альтернативными МНС молекулами класса II, то есть молекулами МНС класса II, отличными от тех, с которых элюируют указанные пептиды с помощью способа, заявленного в соответствии с настоящим изобретением. Диагностические методы с использованием указанных пептидов или терапевтические методы, заявленные в соответствии с настоящим изобретением, с использованием как самих пептидов, так и пептидов, полученных на их основе, могут применяться по отношению к субъектам, экспрессирующим такие альтернативные молекулы МНС класса II.
Распознавание Т-клетками
Процедура верификации эпитопов может включать изучение способности пептидов, идентифицированных с помощью способа, заявленного в соответствии с настоящим изобретением, активировать популяции CD4+Т-клеток. Синтезируют пептиды, имеющие аминокислотные последовательности, идентичные тем, что были идентифицированы согласно настоящему изобретению, или соответствующие коровой последовательности, полученной из "гнездовой" группы пептидов, идентифицированных согласно настоящему изобретению. Затем изучают способность синтезированных пептидов активировать CD4+Т-клетки, полученные от (а) субъектов, экспрессирующих представляющие интерес МНС молекулы класса II и имеющих по крайней мере один симптом заболевания; и (б) контрольных субъектов, экспрессирующих представляющие интерес МНС молекулы класса II, у которых нет симптомов заболевания. Дополнительная контрольная группа может включать людей, имеющих симптомы заболевания, но не экспрессирующих представляющие интерес МНС молекулы класса II.
При некоторых заболеваниях (например, таких, в патогенез которых вовлечен аутоиммунный компонент) измеренная реактивность CD4+Т-клеток у тестируемых субъектов [кроме контрольной группы, описанной в пункте (б)] доказывает предположение о том, что релевантный пептид является эпитопом, активирующим CD4+Т-клетки, которые, в свою очередь, инициируют, способствуют развитию и усугубляют течение соответствующих заболеваний. При других заболеваниях (например, онкологических или инфекционных заболеваниях без аутоиммунного компонента) аналогичный характер реактивности (такой же, как и в предыдущем примере) свидетельствует о том, что релевантный пептид является эпитопом, активирующим CD4+Т-клетки, которые могут обеспечивать иммунитет к указанным заболеваниям или, по крайней мере, способствовать уменьшению клинических проявлений.
Ответ CD4+Т-клеток может быть измерен с помощью различных in vitro способов, хорошо известных специалистам в данной области. Например, возможно культивировать цельные мононуклеарные клетки периферической крови (РВМС) как в присутствии, так и в отсутствие представляющего интерес синтетического пептида, а затем измерить их пролиферацию, например, путем включения в ДНК [3Н]-тимидина. То, что пролиферирующие Т-клетки являются именно CD4+Т-клетками, можно подтвердить как путем отделения CD4+Т-клеток от РВМС перед исследованием, так и путем добавления ингибирующих антител, которые связываются с CD4+молекулой Т-клеток и таким образом ингибируют пролиферацию последних. В обоих ситуациях пролиферативный ответ ингибируется только в том случае, если CD4+Т-клетки и являются пролиферирующими клетками. Альтернативно, CD4+Т-клетки могут быть очищены от РВМС, а их пролиферативный ответ на присутствие пептидов изучен в присутствии АРС, экспрессирующих соответствующие МНС молекулы класса II. Указанными АРС могут быть В-лимфоциты, моноциты, макрофаги, или дендритные клетки, или цельные РВМС. В качестве АРС также могут быть использованы иммортализованные клеточные линии, полученные из В-лимфоцитов, моноцитов, макрофагов или дендритных клеток. АРС могут экспрессировать эндогенные МНС молекулы класса II, представляющие интерес, или же экспрессировать трансфицированные полинуклеотиды, кодирующие указанные молекулы. Во всех случаях перед исследованием АРС можно лишить способности к пролиферации путем обработки, например, ионизирующим излучением или митомицином-С.
Альтернативой исследованию клеточной пролиферации может быть исследование, основанное на измерении продукции цитокинов CD4+Т-клетками с помощью способов, хорошо известных специалистам в данной области. К цитокинам относятся, без ограничений указанными, интерлейкин-2 (IL-2), гамма-интерферон (EFN-γ), интерлейкин-4 (IL-4), TNF-альфа, интерлейкин-6 (IL-6), интерлейкин-10 (IL-10), интерлейкин-12 (IL-12) или TGF-бета. К способам измерения их продукции относятся, без ограничений указанными, ELISA и биоисследования, в которых изучают реактивность (например, пролиферацию) клеток, реагирующих на продукцию соответствующего цитокина, в присутствии тестируемого образца.
Альтернативно, можно непосредственно визуализировать продукцию цитокинов CD4+лимфоцитами с помощью внутриклеточного иммунофлуоресцентного окрашивания и проточной цитометрии.
Кроме того, антигенные пептиды МНС класса II, заявленные в соответствии с настоящим изобретением, могут использоваться для диагностики RA. Таким образом, еще одним вариантом осуществления настоящего изобретения является использование антигенных пептидов, заявленных в соответствии с настоящим изобретением, в качестве маркеров RA.
Предпочтительно в качестве маркеров RA используют антигенные пептиды МНС класса II, включающие (а) по крайней мере аминокислотную последовательность пептидсвязывающего участка, выбранную из группы, включающей SEQ ID NOs. 49-57 и SEQ ID NOs. 103-122, или (б) по крайней мере аминокислотную последовательность пептидсвязывающего участка, выбранную из группы, включающей SEQ ID NOs. 49-57 и SEQ ID NOs. 103-122 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности, выбранной из группы, включающей SEQ ID NOs. 1-39 или SEQ ID NOs. 58-102.
Согласно другому варианту осуществления настоящего изобретения антигенные пептиды, заявленные в соответствии с настоящим изобретением, могут использоваться в качестве маркеров ответа для оценки эффективности терапевтического режима. По существу, можно определить исходный уровень антигенных пептидов, затем ввести определенный лекарственный агент и последовательно отслеживать уровень антигенных пептидов, при этом изменение указанного уровня позволит оценить эффективность проводимой терапии.
Кроме того, антигенные пептиды, выявляемые только на определенных стадиях или фазах заболевания, предпочтительно RA, могут использоваться в качестве маркеров определенной стадии заболевания. По существу, проводят регулярное измерение уровня антигенных пептидов, ассоциированных с определенной стадией заболевания, что позволяет получить информацию, касающуюся стадии заболевания и прогрессирования заболевания.
Настоящее изобретение также относится к использованию полипептидов, из которых получают RA антигенные пептиды, в качестве маркеров для диагностики и контроля за течением заболевания, предпочтительно RA, и, в особенности, эрозивной формы RA в противоположность неэрозивной форме RA. Основанием для такого использования соответствующих белков является тот факт, что DCs присутствуют в большинстве тканей, где они осуществляют захват экзогенных антигенов с помощью специфических рецепторов и посредством специализированных внутриклеточных механизмов (например, макропиноцитоза) с последующим представлением процессированного антигена в виде пептида на молекулах МНС класса II. Предшествующие исследования показали, что частота обнаруживаемого пептидного эпитопа в случае МНС молекул класса II, например RA антигенных пептидов, в большинстве случаев отражает избыток белка, из которого получают указанный определенный пептид.
Таким образом, не только RA антигенные пептиды, но и соответствующие белки могут служить маркерами RA.
Таким образом, согласно другому варианту осуществления настоящего изобретения в качестве маркеров RA используют полипептиды, выбранные из группы, включающей индуцируемую интерфероном-γ лизосомальную тиолредуктазу (SEQ ID NO. 40), интегрин-бета-2 (SEQ ID NO.123), фосфатидилинозитол-4,5-бифосфат-3-киназу (SEQ ID NO:124), активатор плазминогена урокиназного типа (SEQ ID NO: 125), V-III участки тяжелой цепи иммуноглобулина (VH26) (SEQ ID NO: 126), DJ-1 белок (SEQ ID NO:127), аполипопротеин В-100 (SEQ ID NO: 41), 26S протеасомную не АТФ-азную регуляторную субъединицу 8 (SEQ ID NO. 128), рецептор интерлейкина-1 (SEQ ID NO: 129), фибромодулин (SEQ ID NOs.: 130), GM-CSF/IL-3/IL-5 рецептор (SEQ ID NOs. 131), сортирующий нексин 3 (SEQ ID NO: 132), H4 тяжелую цепь ингибитора интер-α-трипсина (SEQ ID NO. 42), комплемент С4 (SEQ ID NO: 43), комплемент С3 (SEQ ID NO: 44), протеин 3, подобный обогащенному глутаминовой кислотой связывающему SH3-домен протеину (SEQ ID NO: 45), индуцируемый интерлейкином-4 протеин 1 (SEQ ID NO: 46), гемопексин (SEQ ID NO. 47), Hsc70-интерактивный протеин (SEQ ID NO: 48), константную цепь (Ii) (SEQ ID NO: 133), отвечающий на протеин 2 рецептор ретиноевой кислоты (SEQ ID NO: 134), фибронектин (SEQ ID NO: 135), катепсин В (SEQ ID NO: 136), трипептидил-пептидаза II (SEQ ID NOs. 137), легумаин (SEQ ID NO: 138), рецептор фактора активации тромбоцитов (SEQ ID NO: 139), поли-альфа-2,8-сиалилтрансферазу (SEQ ID NO:140), ras-родственный пептид Rab-11 В (SEQ ID NO: 141). Предпочтительно полипептид используют в качестве маркера эрозивного RA. Также является предпочтительным использовать полипептид в качестве маркера неэрозивного RA. Особенно предпочтительным является использование индуцируемого интерлейкином-4 белка в качестве маркера RA. До настоящего времени полипептид, представленный на Фиг.1, был неизвестен как маркер RA, но он считается одним из наиболее важных кандидатов на эту роль.
Диагноз RA может быть поставлен на основании изучения экспрессии и/или состава полипептидного или пептидного маркера RA, что можно осуществить с помощью различных способов, включая твердофазный иммуноферментный анализ (ELISA), Вестерн-блоттинг, иммунопреципитацию и иммунофлуоресценцию. В тестируемом образце, полученном от пациента, определяют наличие измененной экспрессии и/или структуры полипептида или пептида, заявленного в соответствии с настоящим изобретением. Измененная экспрессия полипептида или пептида может представлять собой, например, количественно измененную экспрессию (то есть изменение количества продуцируемого полипептида); изменение структуры полипептида представляет собой качественное изменение экспрессии полипептида (например, экспрессия мутантного полипептида или пептида, полученного в результате альтернативного сплайсинга).
Оба указанных изменения (количественное и качественное) также могут иметь место. Термин "изменение" экспрессии или структуры полипептида, используемый здесь, относится к изменению экспрессии или структуры полипептида или пептида в тестируемом образце по сравнению с экспрессией или структурой полипептида или пептида в контрольном образце. Контрольный образец представляет собой образец, который соответствует тестируемому образцу (например, он получен из аналогичного типа клеток), но выделен из организма индивидуума, не страдающего RA. Изменения экспрессии или структуры пептида или полипептида в тестируемом образце при сравнении с контрольным образцом свидетельствуют о наличии RA или о предрасположенности к RA. Для анализа указанных изменений экспрессии или структуры пептида или полипептида, заявленного в соответствии с настоящим изобретением, могут быть использованы различные способы, включая спектроскопию, колориметрию, электрофорез, изоэлектрическое фокусирование и иммунологический анализ (например, см., David et al., Pat. No. 4,376, 110), такой как иммуноблоттинг (см. также Current Protocols in Molecular Biology, в частности, главу 10). Например, согласно одному из вариантов осуществления настоящего изобретения, можно использовать антитело, способное связываться с полипептидом (например, описанным выше), предпочтительно с полипептидом, меченным определяемой меткой. Антитела могут быть поликлональными или более предпочтительно моноклональными. Возможно использование интактного антитела или его фрагмента [например, Fab или F(ab')2). Термин "меченый", используемый в отношении образца или антитела, означает прямое введение метки в образец или антитело путем присоединения (то есть физического связывания) детектируемого соединения к образцу или антителу, а также непрямое введение метки в образец или антитело с помощью реакции с другим реагентом, который можно непосредственно определить. Примерами непрямого введения метки являются определение первого антитела с использованием второго антитела, меченного флуоресцентной меткой, или конечное мечение ДНК образца биотином таким образом, что он может быть определен с помощью стрептавидина, меченного флуоресцентной меткой.
Для измерения уровня или количества пептида или полипептида в тестируемом образце и сравнения полученных результатов с контрольным образцом можно применить Вестерн-блоттинг с использованием антитела, которое специфически связывается с пептидом или полипептидом, заявленным в соответствии с настоящим изобретением, как описано выше. Предпочтительно уровень пептида или полипептида в тестируемом образце измеряют с помощью гомогенного или гетерогенного иммунологического анализа. Если уровень или количество полипептида в тестируемом образце выше или ниже уровня или количества полипептида в контрольном образце и указанное различие является существенным, это свидетельствует об измененной экспрессии полипептида и наличии RA или предрасположенности к нему.
Таким образом, настоящее изобретение также относится к диагностической композиции, включающей антитело, реагирующее с антигенным пептидом МНС класса II, заявленным в соответствии с настоящим изобретением.
Согласно другому варианту осуществления настоящего изобретения антигенные пептиды, заявленные в соответствии с настоящим изобретением, или белки, из которых их получают, могут использоваться для профилактики и лечения заболевания, предпочтительно RA.
Одним из аспектов настоящего изобретения является терапевтический аспект, когда один или более идентифицированный антигенный пептид используется для вакцинации пациентов против RA. В течение курса вакцинации антигенный пептид будет индуцировать антиген-специфичную Т-клеточную толерантность у пациента, которая непосредственно будет способствовать регрессу заболевания или замедлению развития заболевания.
Перспективным подходом для индуцирования специфической иммунной толерантности, который может найти применение в будущих клинических исследованиях, является использование ДНК-вакцин, способствующих развитию толерантности. Было показано, что ДНК-вакцины, способствующие развитию толерантности и кодирующие отдельные аутоантигены, снижают Т-клеточный пролиферативный ответ (Ruiz, P. et al., J Immunol 162 (1999) 3336-3341), в то время как ДНК-вакцины, способствующие развитию толерантности и доставляющие аутоантиген совместно с IL-4, также индуцируют протективный Тн2 ответ (Garren, H. et al., Immunity 15 (2001) 15-22). Примерами неполинуклеотидных специфических толеризующих способов терапии, которые находятся в стадии разработки, являются способы, основанные на применении белковых антигенов, природно-процессируемых пептидов, измененных пептидных лигандов, других биомолекул, таких как ДНК, или белки, или пептиды, имеющие посттрансляционные модификации, а также антигенов, вводимых перорально и вызывающих так называемую "пероральную толерантность" (см. обзор Robinson, W.H. et al., Clin Immunol 103 (2002) 7-12). Одним из возможных побочных эффектов такой терапии, способствующей развитию толерантности, являются аутоиммунные реакции.
Для этой цели можно вводить пациенту соответствующие RA антигенные пептиды в количестве, достаточном для того, чтобы связать МНС молекулы и спровоцировать периферическую толерантность Т-клеток.
Альтернативно, антигенные пептиды, заявленные в соответствии с настоящим изобретением, могут использоваться для получения вакцин, основанных на DCs. В этом случае аутологичные DCs, полученные из моноцитов пациента, могут быть обработаны соответствующими пептидами или рекомбинантными белками, включающими соответствующие пептидные последовательности.
Таким образом, настоящее изобретение относится к фармацевтическим композициям, содержащим антигенные пептиды МНС класса II, включающие (а) по крайней мере аминокислотную последовательность пептидсвязывающего участка, выбранную из группы, включающей SEQ ID NOs. 49-57 и SEQ ID NOs. 103-122, или (б) по крайней мере аминокислотную последовательность пептидсвязывающего участка, выбранную из группы, включающей SEQ ID NOs.
49-57 и SEQ ID NOs. 103-122 с дополнительными N- и С-концевыми фланкирующими последовательностями соответствующей последовательности, выбранной из группы, включающей SEQ ID NOs. 1-39 или SEQ ID NOs. 58-102; антитело, реагирующее с указанным антигенным пептидом, или полипептид, выбранный из группы, включающей SEQ ID NOs. 40-48 и SEQ ID NOs. 123-141, и, необязательно, фармацевтически приемлемый наполнитель, растворитель или носитель. Антигенный пептид должен присутствовать в количестве, достаточном для того, чтобы вызвать толерантность специфических лимфоцитов. Указанное количество зависит от используемого пептида, способа введения, тяжести заболевания, которое предстоит вылечить, а также от общего состояния пациента и обычно находится в пределах от 1 до 50 мг/мл, например, для пептидов, связанных с дендритными клетками.
Приемлемым наполнителем, растворителем или носителем может быть фосфатно-буферный солевой раствор для исследований in vitro и физиологической солевой раствор для применения in vivo.
Термин "вакцинация", используемый здесь, относится как к активной иммунизации, то есть к введению пептидов in vivo с целью стимуляции иммунной толерантности in vivo непосредственно у пациента, так и к пассивной иммунизации, то есть использованию пептидов для стимуляции толерантности CD4+Т-лимфоцитов in vitro или для стимуляции аутологичных или аллогенных дендритных клеток, которые затем повторно вводят пациенту.
Настоящее изобретение также относится к антигенным пептидам, антителам, нуклеиновым кислотам, клеткам-хозяевам, способам, композициям и их применению, как описано выше, в особенности с учетом представленных примеров.
Более полное представление о сущности настоящего изобретения можно получить из приведенных ниже специфических примеров, которые представлены здесь только для иллюстрации изобретения и не ограничивают его сущности, если дополнительно не оговаривается иное, с учетом представленных ниже фигур.
Примеры
Примеры, представленные ниже, проиллюстрированы фигурами, описанными выше, и основаны на методике, обобщенной на Фиг.1 и подробно раскрытой ниже. Если дополнительно не оговаривается иное, для всех примеров использовались промышленные реагенты согласно инструкциям производителя.
Методика изобретения
Дендритные клетки и их культивирование
Исследование осуществляют с использованием дендритных клеток человека, которые были дифференцированы из моноцитов, как описано ниже. Моноциты получают из периферической крови человека. Кровь забирают у здоровых доноров, имеющих следующие гаплотипы: (1) HLA-DRB1*0401, *03011, (2) HLA-DRB1*0401, *0304, (3) HLA-DRB1*0401, *0301, (4) HLA-DRB1*0401, *0701, HLA-DRB1*0401, *0407.
Все клетки культивируют в среде RPMI 1640 (RPMI), обогащенной 1 мМ пирувата, 2 мМ глутамина и 10% инактивированной нагреванием эмбриональной телячьей сыворотки (Gibco BRL, Rockville, MD).
Выделение мононуклеарных клеток периферической крови (PBMCs)
Периферическую кровь получают из банка крови Мангейма (Германия) в виде стандартной композиции, представленной лейкоцитарной пленкой, от здоровых доноров. Для предотвращения свертывания используют гепарин (200 МЕ/мл крови, Liquemine, Roche). Мононуклеарные клетки периферической крови (PBMCs) выделяют с помощью центрифугирования в LSM® (1,077-1,080 г/мл; ICN, Aurora, ОН) при 800g (комнатная температура) в течение 30 минут. PBMCs собирают в интерфазе и дважды отмывают PRMI, содержащей 20 мМ Hepes (500g в течение 15 мин, 300g в течение 5 минут). Для удаления эритроцитов PBMCs обрабатывают ALT буфером (140 мМ хлорида аммония, 20 мМ Tris, pH 7.2) в течение 3 минут при 37°С. PBMCs дважды отмывают RPMI, содержащей 20 мМ Hepes (200g в течение 5 минут).
Получение дендритных клеток из моноцитов периферической крови
Моноциты выделяют из PBMCs с помощью позитивной сортировки с использованием анти-CDH магнитных бусин (Miltenyi Biotech, Auburn, CA) в соответствии с инструкциями производителя. Моноциты культивируют в RPMI, обогащенной 1% заменимыми аминокислотами (Gibco, BRL, Rockville, MD), 50 нг/мл рекомбинантного гранулоцитарно-макрофагального колониеобразующего фактора человека (GM-CSF, от англ. Granulocyte Macrophage-Colony Stimulating Factor, S.A.
1,1·107 Ед/мг) (Leucomax; Novartis, Basel Switzerland) и 3 нг/мл рекомбинантного IL-4 человека (S.A. 2,9·104 Ед/мкг) (R&D Systems, Minneapolis, MN). Для получения незрелых дендритных клеток моноциты высевают в количестве 0,3·106 /мл в 6-луночные планшеты и культивируют в течение 5 дней.
Качество незрелых дендритных клеток, полученных из моноцитов, регулярно оценивают с помощью проточной цитометрии; оно считается приемлемым в том случае, когда клетки имеют следующий фенотип: CD1a (высокий), CD3 (нег.), CD14 (низкий), CD19 (нег.), CD56 (нег.), CD80 (низкий), CD83 (нег.), CD86 (низкий) и HLA-DR (высокий). В отличие от этого зрелые дендритные клетки (см. ниже) имеют следующий фенотип: CD1a (низкий), CD80 (высокий), CD83 (высокий), CD86 (высокий) и HLA-DR (высокий). Моноклональные антитела к CD1a, CD3, CD14, CD19, CD56, CD80, CD83, а также соответствующие изотипические контроли приобретают у фирмы Pharmingen (San Diego, CA).
Обработка дендритных клеток сывороткой или синовиальной жидкостью
Сыворотку или синовиальную жидкость в течение 30 минут облучают 137Cs (70 TBq). Для подпитки дендритных клеток сывороткой или антигеном, выделенным из синовии, 6·106 незрелых дендритных клеток обрабатывают или 1 мл сыворотки или 0,6 синовиальной жидкости. Одновременно индуцируют созревание дендритных клеток добавлением 10 нг/мл рекомбинантного фактора некроза опухолей альфа человека (TNFα; S.A. 1,1·105 МЕ/мкг). В качестве контроля 6·106 незрелых дендритных клеток инкубируют только с TNFα.
Через 24 часа культивирования дендритные клетки собирают с помощью центрифугирования при 300g в течение 10 минут. Клетки отмывают PBS и переносят в пробирки Эппендорф. После центрифугирования при 400g в течение 3 мин супернатант полностью удаляют, а клетки замораживают при -70°С.
Получение бусин с анти-HLA класса II
Анти-HLA-DR моноклональное антитело (mAb) L243 (АТСС, Manassas, VA) получают путем культивирования соответствующей клеточной линии гибридомы мыши. MAb L243 очищают с помощью Протеин А сефарозы (Pharmacia, Uppsala, Sweden) и наносят на CNBr-активированные сефарозные бусины (Pharmacia) до конечной концентрации 2,5 мг/мл согласно инструкциям производителя. L243 бусины хранят в PBS, содержащем 0,1% Zwittergent 3-12 (Calbiochem, La Jolla, CA).
Очистка HLA-DR пептидных комплексов в наномасштабе
Осадки замороженных дендритных клеток ресуспендируют в десятикратном объеме ледяного лизирующего буфера [1%-ный Triron-X-100, 20 мМ Tris, pH 7,8, 5 мМ MgCl2, содержащего ингибиторы протеазы, такие как химостатин, пепстатин, PMSF и лейпептин (Roche, Mannheim, Germany)] и лизируют в горизонтальном шейкере при 1000 rpm, 40C в течение 1 ч. Клеточный лизат очищают от клеточного дебриса и ядер с помощью центрифугирования при l0000g, 4°C в течение 10 мин. Лизат инкубируют совместно с L243 бусинами (5-10 мкл L243 бусин на 100 мкл клеточного лизата) в горизонтальном шейкере при 1000 rpm, 4°C в течение 2 ч. Иммунопреципитированные HLA-DR комплексы, связанные с L243 бусинами, осаждают с помощью центрифугирования при l000g, 4°C в течение 1 мин и отмывают четыре раза 500 мкл 0,1% Zwittergent 3-12 (Calbiochem) в PBS.
Эффективность очистки HLA-DR-пептидных комплексов контролируют, анализируя соответствующие клеточные лизаты до и после иммунопреципитации и аликвоты бусин с помощью Вестерн-блоттинга с использованием анти-HLA-DRα-специфичного mAb 1B5 (Adams, Т.Е. et al., Immunology 50 (1983) 613-624).
Элюция HLA-DR-ассоциированных пептидов
HLA-DR-пептидные комплексы, связанные с L243 бусинами, ресуспендируют в 100 мкл Н2О (HPLC-градиент; Merck, Darmstadt, Germany), переносят в пробирки для ультрафильтрации, Ultrafree MC, с отсечением по размеру 30 кД (Millipore, Bedford, МА) и отмывают 10 раз 100 мкл Н2О (HPLC-градиент) путем центрифугирования в течение 1-2 минут при l0000g при комнатной температуре. Для элюции связанных пептидов добавляют 60 мкл 0,1%-ной трифторуксусной кислоты (Fluka, Buchs, Switzerland) в Н2О (HPLC-градиент) и инкубируют в течение 30 минут при 37°С. Элюированные пептиды собирают в новые пробирки Эппендорф путем центрифугирования в системе Ultrafree при 10000g в течение 3 мин при комнатной температуре и немедленно лиофилизируют в вакуумной центрифуге Speed-Vac™.
Фракционирование с помощью двумерной жидкостной хроматографии (LC. от англ. Liquid Chromatography) в нанопотоке
Для секвенирования комплексных смесей пептидов с высокой пропускной способностью используют многомерную технологию для идентификации пептидов (MudPIT, от англ. Multidimensional Protein Identification Technology) (Washbum, M.P. et al., Nat Biotechnol 19 (2001), 242-247), которая основана на фракционировании с помощью жидкостной хроматографии с последующим масс-спектрометрическим анализом последовательностей.
Для этой цели лиофилизированные пептиды, элюированные с HLA молекул, ресуспендируют в буфере, содержащем 5% (об./об.) ацетонитрила (ACN), 0,5% (об./об.) уксусной кислоты, 0,012% (об./об.) гептафтормасляной кислоты (HFBA) и 1% (об./об.) муравьиной кислоты. Смесь пептидов фракционируют на микрокапиллярной колонке из слитого кварцевого стекла (внутренний диаметр 100 мкм · 375 мкм), полученной с помощью лазерного выталкивателя Model P-2000 (Sutler Instrument Co., Novato, CA). Микроколонку наполняют 3 мкм/С18 материала с обратной фазой (С18-АСЕ 3 мкм [ProntoSIL 120-3-С18 ACE-EPS, Leonberg, Germany]), затем 3 см 5 мкм катионообменного материала (Partisphere SCX; Whatman, Clifon, USA).
Осуществляют полностью автоматическое градиентное разделение пептидов на LC Packings UltiMate HPLC (LC Packings, San Francisco, USA), включающее 8 этапов, при этом используют следующие буферы: 5% ACN/ 0,012% HFBA/ 0,5% уксусной кислоты (буфер А); 80% ACN/ 0,012% HFBA/ 0,5% уксусной кислоты (буфер В); 250 мМ ацетата аммония/ 5% ACN/ 0,012% HFBA/ 0,5% уксусной кислоты (буфер С) и 1,5 мМ ацетата аммония/ 5% ACN/ 0,012% HFBA/ 0,5% уксусной кислоты (буфер D). Первый этап, продолжительность которого составляет 116 минут, включает: 75 минутный градиент буфера В от 0% до 40%, 10-минутный градиент буфера В от 40% до 80%, 6 минут выдерживания в 80%-ном буфере В и 10 минут уравновешивания 100%-ным буфером А. Следующие 5 этапов (каждый продолжительностью 164 минуты) характеризуются следующей схемой: 5 минут 100%-ный буфер А, 5-минутный градиент буфера С от 0% до х%, 5 минут 100%-ный буфер А, 30-минутный градиент буфера В от 0% до 10%, 55-минутный градиент буфера В от 10% до 35%, 20-минутный градиент буфера В от 35% до 50%, 10-минутный градиент буфера В от 50% до 80%, 6 минут выдерживания в 80%-ном буфере В и 10 минут уравновешивания 100%-ным буфером А. Процент буфера С (х) на этапах 2-6 составляет: 20, 40, 60, 80 и 90%. 30 минутный градиент буфера В от 0 до 10%, который является первым этапом линейной элюции с материала с обратной фазой, необходим для того, чтобы адекватно разделить элюцию пептидов и элюцию основного загрязнителя (m/z=945), который приводит к потере большинства гидрофильных пептидных пиков. Этап 7 включает следующие фазы: 5 минут 100%-ный буфер А, 20 минут 100%-ный буфер C, 5-минутный градиент буфера В от 0% до 10%, 35-минутный градиент буфера В от 10% до 35%, 50-минутный градиент буфера В от 35% до 50%, 10-минутный градиент буфера В от 50% до 80%, 5 минут выдерживания в 80%-ном буфере В и 10 минут уравновешивания 100%-ным буфером А. Этап 8 идентичен этапу 7, за исключением того, что вместо буфера С используют буфер D.
Масс-спектрометрия MS/MS с ионной ловушкой
HPLC колонку напрямую присоединяют к масс-спектрометру с ионной ловушкой Finnigan LCQ Deca XP Plus (Thermo Finnigan, San Jose, USA), оснащенному нано-LC источником ионизирующего электрораспыления. Масс-спектрометрию в режиме MS/MS осуществляют согласно инструкции производителя. Пептиды идентифицируют с помощью программы SEQUEST (патенты США 6 017 693 и 5 538 897).
Масс-спектрометрия MALDI-ТИЗ
Пептиды, нанесенные на планшет AnchorChip, кристаллизуют совместно с матриксом [5 мг/мл; α-циано-гидрокси-коричная кислота (Merck, Darmstadt, Germany), 50%-ный ацетонитрил, 0,1%-ная трифторуксусная кислота). Для количественного анализа полного набора пептидов образцы анализируют на масс-спектрометре Ultraflex™ MALDI-ТИЗ (Broker, Bremen, Germany) согласно инструкции производителя.
Идентификация последовательностей с помощью SEQUEST и дифференцированный анализ данных
Данные MS/MS фрагментации анализируют с помощью программы SEQUEST (Thermo Finnigan, San Jose, USA). Из внутренней базы данных по белкам, созданной на основе публичной базы данных Swiss-Prot TrEMBL, SEQUEST выбирает для каждого спектра все пептидные последовательности, которые соответствуют молекулярной массе родительского иона и измеряет степень соответствия между экспериментальным спектром и теоретическим, in silico, генерированным спектром. В перечень включаются только кандидатные последовательности, дающие наивысший счет.
Пептидные последовательности, полученные с помощью SEQUEST анализа, а также сопутствующую информацию о точной массе, параметрах счета и происхождении пептидов сохраняют в соответствующей подходящей базе данных и подвергают дополнительной обработке. Для гарантированного сохранения только значимых последовательностей с удовлетворительным SEQUEST счетом вводят определенные ограничения. Два наиболее важных ограничения включают: (i) сохранение только тех последовательностей, которые имеют коэффициент взаимной корреляции (СС, от англ. Correlation Coefficient), превышающий определенную величину, и (ii) из оставшихся последовательностей сохранение тех, которые имеют предопределенный дельта коэффициент взаимной корреляции (СС). Для обоих указанных критериев минимальные отбираемые величины основаны на эмпирических знаниях об интерпретации результатов SEQUEST.
Набор данных определяют в виде суммы данных из конкретного диапазона спектра. Техническое решение базы данных и компьютерной программы позволяет осуществлять запрос как по одному набору данных, так и производить сравнение многочисленных баз данных. Указанное решение базы данных и компьютерной программы позволяют осуществлять анализ образцов, который не обеспечивает программа SEQUEST. Например, возможные запросы по одному набору данных позволяют получить информацию о распределении счета среди сохраненного спектра последовательностей, о наличии дополнительных вариантов последовательностей по длине или о наличии общих субпоследовательностей, а также о том, из какого белка получена пептидная последовательность. Поскольку наличие усеченных вариантов одного эпитопа является общим свойством пептидов, связанных с МНС молекулами класса II, наличие вариантов по длине в наборе данных позволяет получить дополнительное строгое доказательство того, что в этом спектре присутствует эпитоп.
Наиболее важной особенностью анализа многочисленных баз данных является то, что он дает возможность выявить общее подмножество последовательностей, которое удовлетворяет заданным критериям. Указанные критерии должны основываться на сходстве последовательностей; например, среди всех последовательностей, которые находятся в базе данных, отбираются только те последовательности, которые имеют по крайней мере одну общую субпоследовательность. Указанный сравнительный анализ различных баз данных обеспечивает дифференцированный подход (RA образцы против контрольных образцов) и, таким образом, позволяет оптимизировать поиск потенциальных пептидных маркеров RA.
Показатели счета попарного сходства последовательностей вычисляют с помощью обычной компьютерной программы, которая реализует стандартный алгоритм по сравнению строк. Впоследствии указанные показатели счета используют для объединения родственных последовательностей (последовательностей, имеющих общую субпоследовательность) во вполне разделенные кластеры, что осуществляют с помощью дополнительно разработанной компьютерной программы, которая основана на хорошо известном алгоритме (иерархический кластерный анализ, UPGMA).
Полученные кластеры (например, кластеры усеченных вариантов пептидов) затем используют для идентификации родственных последовательностей в различных базах данных.
Кроме того, программа для анализа данных дает возможность быстро и воспроизводимо осуществлять следующее:
- из выходных данных, полученных с помощью SEQUEST, отбирать те последовательности, которые удовлетворяют достоверным эмпирическим критериям;
- сохранять данные в базе данных, разработанной вручную специально для исследования;
- получать информацию о всех последовательностях, сохраненных в каждой базе данных. Указанная информация является ценной, поскольку позволяет определить важность отдельных последовательностей в заданной базе данных и, следовательно, в многочисленных базах данных;
- обеспечивать дифференцированный подход, а именно изучать актуальные последовательности одного образца по сравнению с другим(и), благодаря сравнительному анализу многочисленных баз данных.
Очистка HLA-DR молекул
HLA-DR молекулы очищают от 1010 EBV-трансформированных В-клеточных линий или Т2-трансфектантов с помощью аффинной хроматографии с использованием анти-DR моноклонального антитела L243, как описано выше (Kropshofer Н. et al., PNAS 92 (1995) 8313-8317).
Анализ связывания пептидов in vitro
НА(307-319), PKYVKQNTLKLAT, представляет собой иммунодоминантный эпитоп гемагглютинина вируса гриппа, который хорошо связывается с HLA-DR молекулами и используется в качестве репортерного пептида в анализе связывания пептидов in vitro (Rothbard, J.B. et al., Cell (1988) 52:515-523).
Очищенные растворенные в детергенте HLA-DR4 молекулы (200 нМ) инкубируют совместно с биотинилированным НА (307-319) пептидом (200 нМ) и градуированным количеством конкурирующего пептида (100 нМ - 10 мкМ) в течение 24 часов при 37°С в связывающем буфере (50 нМ фосфата натрия, 50 нМ цитрата натрия, рН 4,8, 0.1% Zwittergent 3-12) в общем объеме 50 мкл. Конкурирующие пептиды происходят из возможных RA антигенов, идентифицированных в данном исследовании, а также используют синтетические пептиды фирмы Medprobe (Lund, Sweden).
Затем 3·10 мкл 10-кратно разводят в PBS, содержащем 0,05%-ный Твин-20 и 1%-ный BSA, и инкубируют в течение 2 ч в микротитрационном планшете (Nalge Nunc), на который предварительно наносят анти-DR моноклональное антитело L243 (в течение ночи). Сразу после этого получают образцы путем инкубации с 0,1 мкг/мл EU-меченым стрептавидином (Wallay Oy, Turku, Finland) в течение 45 минут согласно инструкции производителя. После интенсивного отмывания 0,05%-ным Твином-20 в PBS измеряют флуоресценцию европия с помощью флуориметра с временным разрешением (VICTOR 1420, Wallac/Perkin Elmer Life Sciences) для подсчета связывания биотинилированного НА(307-319) пептида с HLA-DR молекулами (Aradt, S.O. et al., EMBO J. 19 (2000) 1241-1251).
Пример 1
В данном примере для идентификации новых маркеров HLA-DR-ассоциированных пептидов, выделенных из сыворотки и синовиальной жидкости пациентов с неэрозивным RA, используют методику, показанную на Фиг.1.
6·106 незрелых дендритных клеток обрабатывают 1 мл сыворотки (5 образцов) или 0,6 мл синовиальной жидкости (2 образца) пациентов с неэрозивным RA и культивируют в течение 24 часов в присутствии 10 нг/мл TNFα. В качестве контроля культивируют 6·106 дендритных клеток в присутствии TNFα (10 нг/мл) и 1 мл PBS, но без добавления сыворотки. В дополнительном эксперименте 6·106 дендритных клеток обрабатывают 1 мл сыворотки от двух здоровых людей и культивируют в течение 24 часов в присутствии TNFα (10 нг/мл).
Дендритные клетки лизируют детергентом ТХ-100 и HLA-DR молекулы выделяют с помощью mAb L243. HLA-DR-ассоциированные пептиды элюируют 0,1%-ным TFA и анализируют с помощью высоко пропускной 2D-LC-MS/MS методики. Идентификацию пептидов осуществляют с помощью алгоритма SEQUEST. Пептидные последовательности, полученные с помощью анализа SEQUEST, и дополнительную информацию о точной массе, параметрах счета и происхождении пептидов сохраняют в соответствующей подходящей базе данных и подвергают дополнительной обработке.
Пептидные последовательности, полученные из необработанных DCs (контроль 1) и из DCs, обработанных сывороткой здоровых людей (контроль 2), сравнивают с пептидными последовательностями, полученными из DCs, обработанных сывороткой пациентов с неэрозивным RA. Среди RA-специфичных последовательностей для дальнейшего анализа отбирают только те пептиды, которые повторно выявляются по крайней мере в трех из пяти образцов неэрозивного RA.
В каждом образце сыворотки идентифицируют приблизительно 600±150 индивидуальных пептидных последовательностей (коэффициент взаимной корреляции СС>3.0 и ΔСС>0.15). В образцах синовиальной жидкости количество индивидуальных пептидных последовательностей значительно меньше (400±30). Приблизительно 80-85% пептидов, обнаруживаемых в RA образцах, также можно выделить и из контрольных образцов, что свидетельствует о высокой воспроизводимости метода. В большинстве случаев можно выделить несколько вариантов одного эпитопа разной длины, что является характерной особенностью антигенов, связанных с МНС класса II, и подтверждает достоверность результатов (Jones, E.Y., Curr Opin Immunol 9 (1997) 75-79). Другим подтверждением качества полученных данных является тот факт, что некоторые идентифицированные пептиды или белки уже были описаны как ассоциированные с молекулами МНС класса II: эпитопы, полученные из повсеместно распространенных белков, таких как Hsp70, энолазы, аннексии II, катепсин С или коллаген II, а также из молекул МНС (HLA-A, -В, -С, -Е, -G и 2-микроглобулин) и CLIP (Chicz, R.M. et al., J Exp Med 178 (1993) 27-47; Sinigaglia, F. & Hammer, J., Curr Opin Immunol 6 (1994) 52-56; Amold-Schild, D. et al., J Immunol 162 (1999) 3757-3760; Vogt, A.B. & Kropsher, H., Trends Biochem Sci 4 (1999) 150-154) также часто определяются.
RA-специфичные пептидные последовательности дополнительно подтверждают по связыванию с аллелем DRB1*0401, определяющим предрасположенность к RA, с помощью программы TEPITOPE (Hammer, J. et al., Adv Immunol 66 (1997) 67-100). Указанная программа позволяет провести количественное и качественное определение Т-клеточных эпитопов.
Данные, полученные в ходе исследования, касаются только эпитопа, который встречается исключительно в образцах от больных с неэрозивным RA (за одним исключением) (Таблица 1).
Индуцируемая гамма-интерфероном лизосомальная тиолредуктаза
Очень интересный эпитоп, идентифицированный в трех из семи образцов сыворотки и синовии больных с неэрозивным RA, получают из индуцируемой гамма-интерфероном лизосомальной тиолредуктазы (GILT): 16-мерной GILT (192-207) с аминокислотной последовательностью SEQ ID NO: 3 (Таблица 1). Дополнительные варианты указанного эпитопа (по длине) в трех других образцах подтверждают релевантность указанного эпитопа (Таблица 1): 14-мерная GILT (192-205; SEQ ID NO: 1) и 17-мерная GILT (192-208; SEQ ID NO:2).
При анализе наиболее короткого варианта, GILT (192-205), было установлено, что эпитоп содержит подходящий связывающий участок аллеля DRB1*0401, определяющего предрасположенность к RA, причем 196М выполняет функцию Р1 якоря, 199М - Р4 якоря, а 201 А - Р6 якоря. Согласно TEPITOPE подсчету эпитоп имеет показатель связывания (пороговая величина), составляющий 1%, который аналогичен показателю связывания эпитопа гемагглютинина вируса гриппа (307-319), представляющего собой сильный DRB1*040 -связывающий агент (Таблица 1) (Rothbard, J.B. et al., Cell 52 (1988) 515-523).
GILT конститутивно экспрессируется антиген-представляющими клетками, такими как дендритные клетки, макрофаги и В-клетки, а также обеспечивает разворачивание подвергшихся эндоцитозу антигенов в МНС класса II-содержащих компартментах (МПС) путем энзиматического расщепления дисульфидных связей (Phan, U.T. et al., J Biol Chem 275 (2000) 25907-25914). Было показано, что в В-клетках происходит прямое связывание GILT с HLA-DR молекулами (Arunachalam, В. et al., J Immunol 160 (1998) 5797-5806). Также было установлено, что относительно длинный второй эпитоп GILT связывается с HLA-DR3 молекулами: 22-мерная GILT (38-59), имеющая аминокислотную последовательность SPLQALDFFGNGPPVNYKTGNL (Chicz, R.M. et al., J Exp Med 178 (1993) 27-47).
Помимо GILT (192-207) в некоторых RA образцах был идентифицирован другой эпитоп того же белка, он же был выявлен и в контрольных образцах: GILT (210-227) с аминокислотной последовательностью QPPHEYVPWVTVNGKPLE. Указанный эпитоп был идентифицирован совместно с тремя вариантами (по длине): 16-мерная GILT (210-225), 17-мерная GILT (210-226) и 19-мерная GILT (210-228).
Как свидетельствует название фермента, экспрессия GILT может индуцироваться провоспалительным цитокином интерфероном-гамма (IFN-γ) в клетках различных типов, включая макрофаги, эндотелиальные клетки и фибробласты (Luster, A.D. et al., J Biol Chem 263 (1988) 12036-12043). Поскольку ifn-γ обнаруживается в воспаленных суставах пациентов с RA, может наблюдаться избыточная экспрессия GILT в синовии и сыворотке и, как следствие этого, захват ее DCs как экзогенного антигена. GILT (192-207) может быть получена на основе экзогенной GILT. Другой эпитоп GILT, который также выявляется в контрольных образцах, может быть получен из эндогенной GILT, экспрессируемой DCs. Альтернативно, оба варианта, а именно GILT (192-207) и GILT (210-227), могут быть получены на основе эндогенной GILT в том случае, если процессинг GILT и представление происходящего из GILT эпитопа DCs были значительно изменены в результате контакта с RA-ассоциированным материалом.
Идентифицированный эпитоп индуцируемой интерфероном-гамма лизосомальной тиолредуктазы GILT (192-205), который был детально описан выше, дополнительно анализируют в исследовании связывания in vitro с использованием синтетической GILT (192-205) и очищенных HLA-DR молекул (Фиг.3). Согласно подсчету с помощью TEPITOPE пептид связывает HLA-DR4 с высокой аффинностью в сравнении с вирусным НА (307-319) пептидом.
Интегрин бета-2
Подробный анализ выявил наличие эпитопа, который был идентифицирован в двух из семи образцов сыворотки и синовии пациента с неэрозивным RA и который получают из бета-субъединицы интегрина (ITB2): 17-мерный ITB2 (315-331; SEQ ID NO: 58) с аминокислотной последовательностью NIQPIFAVTSRMVKTYE (Таблица 1). Был обнаружен один вариант по длине: 19-мерный ITB2 (313-331; SEQ ID NO: 59), что подтверждает достоверность идентифицированного эпитопа (Таблица 1).
Пептидная последовательность включает умеренный HLA-DRB1*0401 связывающий участок с 3161 и 3191, выполняющими функцию якорных аминокислот Р1 и Р4 соответственно (показатель связывания, рассчитанный с помощью TEPITOPE, составляет 2%).
Интегрины представляют собой семейство поверхностных клеточных рецепторов, которые играют важную роль в эмбриогенезе, заживлении ран, иммунном ответе и клеточной адгезии. ITB2 представляет собой гетеродимерный рецептор для внутриклеточных и сосудистых молекул адгезии (ICAMs и VCAMs) и экспрессируется исключительно на поверхности лейкоцитов. ICAMs и VCAMs относятся к суперсемейству иммуноглобулинов, которые играют главную роль в гуморальном и клеточном иммунном ответе. Многие цитокины, такие как интерферон-γ, IL-1 и TNFα, регуляция которых повышена при воспалительных заболеваниях, таких как RA, индуцируют экспрессию ICAMs на поверхности эндотелиальных клеток. Было показано, что экспрессия ICAM-1 и VCAM-1 в синовиальной ткани пациентов с RA выше, чем у пациентов с остеоартритом (Furuzawa-Carballeda, J. et al., Scand J Immunol 50 (1999) 215-222).
Связывание 1ТВ2 и других интегринов с ICAMs позволяет лейкоцитам инфильтрировать ткани-мишени, например место воспаления. Для того чтобы осуществлять свою функцию неадгезивным образом или циркулировать в тканях, лейкоциты конститутивно экспрессируют 1ТВ2 и другие интегрины с низкой способностью к связыванию лиганда. Взаимодействие ITB2/ICAM-1 является возможной мишенью для воздействия при воспалении, аутоиммунных заболеваниях и раке (Yusuf-Makagiansar, H. et al., Med Res Rev 22 (2002) 146-167), что является строгим подтверждением достоверности того факта, что идентифицированный ITB2 эпитоп является потенциальным маркером RA.
Фосфатидилинозитол-4.5-бифосфат-3-киназа
Другой эпитоп с умеренным HLA-DRB1*0401 -связывающим участком был идентифицирован в двух образцах сыворотки пациентов с неэрозивным RA: 17-мерный PI3K (792-808; SEQ ID NO: 60) с аминокислотной последовательностью NKVFGEDSVGVIFKNGD, выделенный из фосфатидилинозитол-3-киназы (PI3K) (Таблица 1). В указанном пептиде 794V может выполнять функцию гидрофобной якорной аминокислоты PI, 797E - отрицательно заряженной якорной аминокислоты Р4, a 799S - обычной якорной аминокислоты DR4-P6 (показатель связывания, рассчитанный с помощью TEPITOPE, составляет 2%).
PI3K широко экспрессируется клетками многих типов и фосфорилирует липиды, главным образом фосфатидилинозитол-4,5-бифосфат. Этот фермент выступает в роли ключевого трансдуктора сигнала для рецепторов факторов выживания, включая факторы роста, цитокины и интегрины (обзор Toker, A. & Cantley, L., Nature 387 (1997) 673-676). Кроме того, PI3K играет важную роль в пути передачи сигнала Toll-рецепторов (TLRs), которые распознают широкий спектр микробных продуктов, объединенных термином патоген-ассоциированные молекулярные продукты (PAMPs) (Fukao, Т. & Koyasu, S., Trends Immunol 24 (2003) 358-363). Стимуляция TLRs с помощью PAMPs, таких как липополисахарид (LPS) (эндотоксин), запускает синтез различных цитокинов, включая IL-2, который является ключевым цитокином в TLR-опосредованных Thi ответах. Было показано, что PI3K является эндогенным супрессором TLR-опосредованного синтеза IL-2 и ограничивает избыточное перемещение Thi. Таким образом, было сделано предположение о том, что PI3K является негативным регулятором естественных иммунных ответов, что предотвращает развитие пролонгированного активного иммунитета, который может оказаться вредным для организма-хозяина. Информация о том, что аутоиммунные ответы при RA основаны на Th1 цитокинах, позволяет связать PI3K с патогенезом RA. Хотя специфический патоген, непосредственно связанный с развитием RA, до настоящего времени неизвестен, есть основания предполагать, что RA развивается в два этапа, при этом первичный ответ, индуцированный чужеродными антигенами, последовательно переходит в самоподдерживающийся аутоиммунный ответ (Klinman, D., Arthritis Rheum 48 (2003)590-593).
Активатор плазминогена урокиназного типа
Другой эпитоп, определяемый только в образцах от пациентов с неэрозивным RA, был выделен из активатора плазминогена урокиназного типа (uPA): 16-мерный uPA (328-343; SEQ ID NO: 61) с аминокислотной последовательностью YPEQLKMTVVKLISHR (Таблица 1). Исходя из определения HLA-DRB1*0401 -связывания, последовательность содержит умеренный связывающий участок: 332L, 335Т и 337V представляют собой предположительные якорные аминокислоты Р1, Р4 и Р6 соответственно (показатель связывания, полученный с помощью TEPITOPE, составляет 2%).
Активаторы плазминогена (PAs) представляют собой высокоспецифичные сериновые протеазы, отвечающие за превращение плазмина в плазминоген. Два типа PAs, урокиназного типа (uPA) и тканевого типа (tPA), были идентифицированы для млекопитающих. Активность PAs контролируется природными ингибиторами (PAIs). Плазмин вовлечен в воспалительные реакции с участием индуцирующих цитокинов, таких как TGFβ, а также в процесс деградации хряща и фибрина. Признаки постоянной коагуляции в суставе, пораженном RA, совместно с усиленным синтезом uPA и его активностью в суставных тканях позволили сделать предположение о том, что система РА/плазмин ассоциирована с клинической тяжестью артрита (обзор Busso, N. & Hamilton, J.A. Arthtritis Rheum 46 (2002) 2268-2279). На основании исследований in vitro было установлено, что в воспаленных суставах пациентов с RA присутствуют несколько типов клеток, в частности моноциты и фибробласты, которые могут экспрессировать PAs и PAIs, что, следовательно, способствует повышению указанных показателей в in vitro образцах. Активность PAs (то есть uPA) стимулируется цитокинами, такими как IL-1 и TNFα, регуляция которых в сыворотке и синовиальной жидкости пациентов с RA повышена. Приведенные выше данные являются доказательством того, что uPA (328-343) может быть предположительным пептидным маркером RA.
V-II (VH26) участок тяжелой цепи иммуноглобулина
Одним из идентифицированных эпитопов, обнаруживающих очень сильный связывающий участок HLA-DRB1*0401, является эпитоп, выделенный из генного сегмента VH26 тяжелой цепи иммуноглобулина: 16-мерный VH26 (95-110; SEQ ID NO: 62) с аминокислотной последовательностью KNTLYLQMNSLRAEDT (Таблица 1). Очень высокий показатель связывания (1%), полученный с помощью TEPITOPE, подтверждается тем, что функцию мощного якоря Р1 выполняет 99Y, умеренного якоря Р4 - 102М, а функцию типичного HLA-DR4 Р6 якоря выполняет 104S.
Иммуноглобулины (Ig) ответственны за связывание антигена и стимулируют дополнительные иммунные реакции, например, путем связывания с изотип-специфичными Fc рецепторами. Интересно, что генный сегмент VH26 человека, по-видимому, кодирует высокоавидный синовиальный ревматоидный фактор и, таким образом, оказывает влияние на прогрессирование RA (Wong, A. et al., Autoimmunity 20 (1995) 191-199). Тот факт, что идентифицированный эпитоп VH26 (95-110) был обнаружен в сыворотке двух серопозитивных пациентов с RA (Таблица 1), коррелирует с приведенным выше наблюдением.
DJ-1 белок
В двух образцах неэрозивного RA с помощью длительных исследований был выявлен эпитоп, который выделяют из белка, называемого DJ-1: 16-мерный DJ-1 (135-150; SEQ ID NO: 63) с аминокислотной последовательностью NGGHYTYSENRVEKDG (Таблица 1). Согласно TEPITOPE подсчету пептид содержит относительно слабый HLA-DRB1*0401 - связывающий участок с 139Y в роли Р1 якоря, 124 S в роли Р4 якоря и 144N в роли возможного Р6 якоря (показатель связывания: 8%).
DJ-1 относится к семейству ThiJ/PfpI белков, члены которого в процессе эволюции были перенесены от Archaea к Eukarya. ThiJ/PfpI белки имеют общий ThiJ домен, который по своей структуре может быть отнесен к домену глутаминаминотрансферазы I типа (Lee, S.J. et al., J Biol Chem 25 (2003) Epub ahead из print).
DJ-1, который экспрессируется преимущественно тканями яичек и в меньшей степени тканями других органов, впервые был идентифицирован как новый потенциальный онкогенный продукт, который трансформирует NIH3T3 клетки мышей совместно с ras. Тем временем были обнаружены дополнительные физиологические функции DJ-1, включая наличие строгой корреляции с болезнью Паркинсона и фертилизацией спермы. Предполагается, что DJ-1 выполняет множество функций - авторами настоящего изобретения были впервые представлены данные о связи DJ-1 с RA.
Пример 2
В данном примере используется та же методика, которая детально описана в Примере 1. Для идентификации потенциальных маркеров, специфичных в отношении эрозивного RA, используют сыворотку (шесть образцов) и синовиальную жидкость (два образца) от пациентов с диагностированным эрозивным RA.
Пептидные последовательности, обнаруживаемые в образцах от пациентов с эрозивным RA, сравнивают с последовательностями, идентифицированными в необработанных DCs (контроль 1) и DCs, обработанных сывороткой здоровых людей (контроль 2). Среди RA-специфичных последовательностей для дальнейшего анализа отбирают только те, которые повторно выявляются по крайней мере в трех из шести образцах эрозивного RA.
В данном исследовании был обнаружен один эпитоп, который выявляется, за одним исключением, только в образцах эрозивного RA.
Аполипопротеин В-100
Эпитоп, который главным образом выявляется в сыворотке пациентов с эрозивным RA (в четырех образцах RA из восьми) выделяют из аполипопротеина В-100: 16-мерный АроВ (227-2892) с аминокислотной последовательностью SEQ ID NO: 4 (Таблица 2). Кроме того, выявляют вариант указанного эпитопа по длине (Таблица 2): 17-мерный АроВ (2877-2893; SEQ ID NO:5). Можно предположить наличие следующего DRB1*0401-связывающего участка: 288L в качестве Р1 якоря, 288D в качестве Р4 якоря и 2886N в качестве Р6 якоря (показатель связывания 3%).
В ранних исследованиях EBV-B-клеток была обнаружена связь эпитопа АроВ (2885-2900), который частично совпадает с эпитопом, описанным здесь, с HLA-DR4 (Chicz, R.M. et al., J Exp Med 178 (1993) 27-47).
Аполипопротеин В-100 входит в состав липопротеинов очень низкой плотности (ЛОНП) и липопротеинов низкой плотности (ЛПНП) и выполняет роль сигнала распознавания для клеточного связывания и захвата ЛПНП частиц АроВ/Е рецептором (Yang, C.Y. et al., Nature 323 (1986) 738-742). Интересно, что повышенное соотношение холестерина ЛПНП к холестерину ЛПВП наблюдается у пациентов с недавно диагностированным RA (Park, Y.B. et al., J Rheumatol 26 (1999) 1701-1704). Нарушенный липидный профиль при активном RA можно улучшить с помощью применения DMARDs без использования антигиперлипидемических лекарственных средств (Park, Y.B. et al., Am J Med 113 (2002) 188-193). Поскольку стали регистрироваться повышенные показатели смертности от сердечно-сосудистых заболеваний среди пациентов с хроническими воспалительными заболеваниями, такими как RA (Symmons, D.P. et al., J Rheumatol 25 (1998) 1072-1077), было сделано предположение о том, что локальный воспалительный процесс при RA может приводить к нарушению липидного спектра крови и, следовательно, увеличивать риск атеросклероза. Вопрос о том, насколько компоненты метаболизма липидов являются причиной патогенеза RA или же на них только влияют происходящие при RA иммунные реакции, остается открытым. Однако тот факт, что у больных RA отмечается измененный липидный профиль, позволяет подтвердить предположение о том, что эпитоп АроВ может представлять собой потенциальный сывороточный маркер RA.
В контрольных образцах от двух здоровых людей был идентифицирован вариант АроВ (2877-2892) по длине, но не АроВ (2877-2893). Поскольку Аполипопротеин В входит в состав 1% всех белков плазмы, наличие АроВ эпитопов в контрольных образцах не является удивительным. Полученные результаты позволяют предположить, что только вариант АроВ по длине (2877-2893; SEQ ID NO: 5) является специфичным для эрозивного RA.
26S протеасомная не АТФ-азная регуляторная субъединица 8
Эпитоп, который также довольно часто определяется в большинстве образцов от пациентов с эрозивным RA, как в сыворотке, так и в синовиальной жидкости, выделяют из 26S протеасомной не АТФ-азной регуляторной субъединицы 8 (PSMD8): 15-мерная PSMD8 (218-232; SEQ ID NO: 64) с аминокислотной последовательностью GPNNYYSFASQQQKP (Таблица 2). Также были идентифицированы три дополнительных варианта по длине: 16-мерная PSMD8 (218-233; SEQ ID NO: 65), 17-мерная PSMD8 (218-234; SEQ ID NO: 66) и 18-мерная PSMD8 (218-234; SEQ ID NO: 67) (Таблица 2). Наличие вариантов по длине подтверждает достоверность идентифицированного эпитопа как происходящего из молекул МНС класса II антигенного пептида. Пептид содержит участок умеренного связывания DRB1*0401 (TEPITOPE показатель связывания: 3%). Связывание с HLA-DR4 можно подтвердить с помощью исследования связывания in vitro с использованием синтетического PSMD8 (218-233) пептида и очищенных HLA-DR4 молекул (Фиг.3). Согласно величине IC50 (по отношению к репортерному пептиду НА (308-319), PSMD8 (218-233; SEQ ID NO:65) связывается с HLA-DR4 с умеренной аффинностью, что подтверждает данные, полученные с помощью TEPITOPE. 15-мерная PSMD8 (218-232; SEQ ID NO: 64) была идентифицирована в одном из необработанных контрольных образцов.
Около 1% цитоплазматического белкового пула представлено протеасомой, которая участвует в АТФ-зависимой деградации повсеместно распространенных белков. Кроме того, протеасома отвечает за процессинг некоторых факторов транскрипции (например, ядерного фактора-кВ), контроль клеточного цикла и генерацию антигенов, рестриктированных по МНС класса I. Регуляторные субъединицы протеасомы необходимы для обеспечения селективности деградации пептидов. Известно, что не АТФ-азная регуляторная субъединица 8, в частности, необходима для активации белка 28, контролирующего деление клеток (Cdc28), а также является незаменимым регулятором клеточного цикла у дрожжей.
Важно, что значительное повышение уровней циркулирующих протеасом (cProteasomes) выявляются в сыворотках, полученных от пациентов с различными системными аутоиммунными заболеваниями, включая RA (Egerer, К. et al., J Rheumatol 29 (2002) 2045-2052). По всей видимости, существует тесная связь между уровнем циркулирующих протеасом и активностью заболевания и концентрацией С-реактивного белка у пациентов с тяжелым RA. Обсуждается вопрос о том, что циркулирующие протеасомы являются триггерными факторами последующих иммунных ответов, что указывает на управляемый антигенами механизм. Концентрация высвобождаемого протеасомного антигена, по-видимому, отражает степень повреждения клеток при аутоиммунных заболеваниях. На основании своих открытий Egerer et al. сделали заключение о том, что циркулирующие протеасомы могут представлять собой новые маркеры тяжести заболевания при аутоиммунных процессах. Поскольку эпитоп PSMD8 (218-232; SEQ ID NO: 64) был идентифицирован в образцах сыворотки пациентов с эрозивным RA, исследования, проведенные авторами настоящего изобретения, подтверждают это заключение.
Рецептор интерлейкина-1
Другим потенциальным маркером эрозивного RA является рецептор интерлекина-1 (IL-1R), который был идентифицирован в двух образцах сыворотки пациентов с эрозивным RA:IL-1R (79-94; SEQ ID NO: 68) с аминокислотной последовательностью EKLWFVPAKVEDSGHY (Таблица 2). IL-1R (79-94; SEQ ID NO: 68) с 83F в роли PI якоря, 86А в роли Р4 якоря и 88V в роли Р6 якоря имеет в своем составе сильный связывающий участок аллеля DRB1*0401, определяющего предрасположенность к RA (TEPITOPE показатель связывания: 1%). В исследовании связывания in vitro было показано, что синтетический пептид связывается с молекулами HLA-DR4 с высокой аффинностью (Фиг.3), аналогичной таковой НА (309-319) пептида.
Интерлейкин-1 (IL-1) представляет собой провоспалительный цитокин, который участвует в разнообразных инфекционных иммунных ответах, а также в иммунном ответе при RA и других заболеваниях (Dinarello, С., Blood 87 (1996) 2095-2147). Он связывается со своим специфическим рецептором, который передает сигнал, запускающий пролиферацию клеток или стимулирующий синтез белка. Повышенная продукция IL-1 отмечается у пациентов с RA и играет ключевую роль в клинической манифестации этого заболевания (Dayer, J.M. Rheumatology 42 (2003) ii3-ii19). IL-1 рассматривается как основной медиатор при RA, который действует через активацию макрофагов и Т- и В-лимфоцитов.
Кроме того, IL-1 участвует в воспалительных реакциях, индуцируя экспрессию молекул клеточной адгезии, других цитокинов и хемокинов. IL-1 также является одним из главных факторов, участвующих в разрушении кости и хряща при RA, поскольку он стимулирует образование металлопротеиназ матрикса. Следовательно, комплекс IL-1/IL-IR является первой мишенью для противоспалительной терапии. Использование рекомбинантного антагониста рецептора IL-1 человека (IL-1RA) утверждено для лечения пациентов с RA.
Фибромодулин
В трех образцах от пациентов с эрозивным RA был обнаружен эпитоп, выделенный из секретируемого матриксом белка фибромодулина (FM): 13-мерный FM (178-190; SEQ ID NO: 70) с аминокислотной последовательностью LRELHLDHNQISR (Таблица 2). Кроме того, был идентифицирован 14-мерный FM (177-190; SEQ ID NO: 69), который также один раз был обнаружен в необработанном контрольном образце. Эпитоп имеет сильный DRB1*0401-связывающий участок с 181L в роли Р1 якоря, 184D в роли Р4 якоря и 186N в роли Р6 якоря (TEPITOPE показатель связывания: 1%).
Фибромодулин относится к семейству небольших обогащенных лейцином протеогликанов (SLRPs), которые связываются с TGFβ и коллагенами и другими молекулами внеклеточного матрикса. Исследования in vitro показали, что SLRPs регулируют фибриллогенез коллагена, процесс, необходимый для развития, репарации тканей и метастазирования. Для того чтобы лучше понять, какие функции выполняют SLRPs in vivo, получают мышей, дефицитных по SLRPs; было показано, что у таких мышей развивается широкий спектр заболеваний (например, остеопороз и остеоартрит), большинство из которых непосредственно обусловлены нарушенным фибриллогенезом коллагена (Ameye, L. & Young, M.F., Glycobiology 12 (2002) 107R-116R). Поскольку процессы формирования и деградации коллагена значительно активируются в воспаленных суставах при RA, может наблюдаться и повышение уровня фибромодулина. Это предположение также подтверждается обнаружением эпитопа FM в RA образцах синовиальной жидкости (в трех из четырех образцах синовиальной жидкости).
Рецептор GM-CSF/IL-3/IL-5
В пяти из восьми образцах от пациентов с эрозивным RA можно идентифицировать эпитоп из β-цепи множественного рецептора цитокинов (CYRB): 15-мерный CYRB (359-373; SEQ ID NO: 71) с аминокислотной последовательностью ETMKMRYEHIDHTFE и его вариант по длине, 17-мерный CYRB (359-375; SEQ ID NO: 72) (Таблица 2). В исследовании связывания in vitro было показано, что синтетический пептид CYRB (359-375) связывается с молекулами HLA-DR4 с умеренной аффинностью (Фиг.3). Это соответствует данным, полученным с помощью TEPITOPE, которые свидетельствуют о наличии в составе эпитопа умеренно связывающего DRB1*0401 участка (TEPITOPE показатель связывания: 3%). Эпитоп явно в избытке присутствовал в образцах RA и при этом был обнаружен только в одном образце сыворотки здорового человека.
Рецептор GM-CSF/IL-3/IL-5 представляет собой мембранный белок I типа, который дифференцированно экспрессируется клетками системы гемопоэза (обзор Geijsen, N. et al., Cytokine Growth Factor Rev 12 (2001) 19-25). Его лиганд, GM-CSF, секретируется CD4+клетками и является важным стимулом для образования дендритных клеток из клеток-предшественников в костном мозге. Важная роль дендритных клеток в инициации и развитии иммунного ответа позволяет сделать предположение о том, что они также играют важную роль и в развитии аутоиммунных воспалительных заболеваний, таких как RA, транспортируя аутоантиген к дренирующему лимфатическому узлу, где DCs встречаются с "необученными" Т-клетками и праймируют их. Было показано, что активность GM-CSF связана с провоспалительными эффектами при RA, что подтверждает предположение о том, что идентифицированный эпитоп рецептора CYRB (359-373; SEQ ID NO: 71) является потенциальным маркером для диагностики RA.
Сортирующий нексин 3
Другой эпитоп, который, возможно, является показательным для RA, выделяют из белка, обозначаемого как сортирующий нексин 3 (SNX3): 16-мерный SNX3 (142-157; SEQ ID NO: 73) с аминокислотной последовательностью HMFLQDEIIDKSYTPS (Таблица 2). Аминокислоты 144F, 147D и 1491 указанного эпитопа могут выполнять функции Р1, Р4 и Р6 якорных аминокислот соответственно в пептидсвязывающем желобке аллеля DRB1*0401, определяющего предрасположенность к RA (TEPITOPE показатель связывания: 2%).
Сортирующие нексины - это разнообразная по своему составу группа клеточных белков переноса, которые имеют общий фосфолипид-связывающий участок (обзор Worby, С.А. & Dixon, J. В., Nat Rev Mol Cell Biol 3 (2002) 919-31). Способность указанных белков связывать специфические фосфолипиды, так же как и их склонность к формированию белковых комплексов указывает на их участие в регуляции переноса через мембрану и сортировке белков. Сортирующий нексин 3, в частности, присутствует в цитозоле и эндосомах и, по всей видимости, принимает участие в регуляции переноса через мембрану от ранних эндосом к рециркулирующим эндосомам. Играет ли сортирующий нексин 3 какую-либо роль в патогенезе RA, например, влияя на маршрут представления антигена, в настоящее время неизвестно. Идентификация эпитопа SNX3 (142-157; SEQ ID NO:73) в настоящем исследовании предполагает наличие связи между сортирующими нексинами и аутоиммунизацией.
Пример 3
Все пептидные последовательности, идентифицированные в примерах 1 и 2 в образцах от пациентов с неэрозивным и эрозивным RA, в настоящем примере используют для поиска общих маркеров, значимых для обоих типов RA. RA-специфичные последовательности снова сравнивают с пептидными последовательностями контрольных образцов (необработанные DCs и DCs, обработанные сывороткой двух здоровых людей) и для дальнейшего анализа отбирают только те пептиды, которые повторно выявляются по крайней мере в трех из пятнадцати образцах RA (эрозивного и неэрозивного RA).
Ингибитор интер-альфа-трипсина
В десяти из одиннадцати образцах сыворотки (эрозивный и неэрозивный RA) выявляется эпитоп, выделяемый из тяжелой цепи Н4 ингибитора интер-альфа-трипсина: ITIH4 (271-287) с аминокислотной последовательностью SEQ ID NO: 8 (Таблица 3). Помимо указанного самого длинного варианта ITIH4 эпитопа, можно идентифицировать шесть вариантов того же ITIH4 эпитопа по длине: 19-мерный ITIH4 (271-289; SEQ ID NO: б), 18-мерный ITIH4 (271-288; SEQ ID NO: 7), 16-мерный ITIH4 (274-289; SEQ ID NO: 12), 15-мерный ITIH4 (273-287; SEQ ID NO: 10), 15-мерный ITIH4 (274-288; SEQ ID NO: 11) и 14-мерный ITIH4 (274-287; SEQ ID NO: 9).
При анализе самого короткого варианта ITIH4 (274-287) было установлено, что эпитоп имеет в своем составе сильный связывающий участок аллеля DRB1*0401, определяющего предрасположенность к RA: 277F выполняет функцию Р1 якоря, 280D - Р4 якоря и 282S - Р6 якоря (показатель связывания: 1%).
ITIH4 относится к семейству интер-альфа (IαI) ингибиторов, которые представляют собой группу ингибиторов сериновых протеаз, которые связываются с гиалуроновой кислотой (НА) и, по-видимому, участвуют в реакциях острой фазы (Salier, J.P. et al., Biochemical Journal 315 (1996) 1-9).
НА представляет собой полисахарид, обнаруживаемый во всех тканях организма, в частности в рыхлой соединительной ткани, например в суставной жидкости (Evered, D. & Whelan, J. eds., The Biology из Hyaluronan, John Wiley & Sons (1989)). НА выполняет важную структурную функцию в хрящевой ткани и других тканях, где она стабилизирует внеклеточный матрикс, формируя агрегаты с протеогликанами. Также было установлено, что НА выполняет важные биологические функции, которые заключаются в регуляции активности клеток посредством связывания с белками на поверхности клеток, такими как CD44 и ICAM-1 (Knudson, С. В. & Knudson, W., FASEB J 7 (1993) 1233-1241; Hall, C.L. et al., J Cell Biol 126 (1994) 575-588). RA сопровождается значительным повышением общего уровня НА в суставной жидкости, а также в сыворотки крови; на этом основано предположение о том, что циркулирующая НА поступает из пораженных ревматоидным артритом суставов (Engstrom-Laurent, A. et al., Scand J Clin Lab Invest 45 (1985) 497-504).
Комплексы НА и некоторыми членами семейства IαI в значительном количестве определяются в синовиальной жидкости пациентов с RA (Jessen, Т.Е. et al., Biological Chemistry Hoppe-Seyler 375 (1994) 521-526). Роль комплекса IαI-НА в воспалительных реакциях может заключаться в модифицировании взаимодействия CD44-HA, которое опосредует активацию и инвазию лейкоцитов (Isacke, С.М. & Yarwood, H., hit Biochem Cell Biol 34 (2002) 718-721). Кроме того, синовиальная жидкость пациентов с RA содержит большое количество TSG-6, противовоспалительного гликопротеина, принадлежащего к семейству гиаладгеринов, которые относятся к НА-связывающим белкам (Wisniewski, H.G. et al., J Immunol 151 (1993) 6593-6601). Было показано, что комплекс TSF-6 и членов семейства IαI ингибирует активность плазмина, центральной молекулы, участвующей в активации ферментов, ассоциированных с воспалением (Wisniewski, H.G. et а1., J Immunol 156 (1996) 1609-1615). Регуляция активности плазмина некоторыми плазменными белками острой фазы, а именно TSG-6 и белками из семейства IαI, может играть важную роль в патогенезе RA и приводить к повышению уровней НА, TSG-6 и белков из семейства IαI в синовиальной жидкости воспаленных суставов.
Указанные наблюдения, а также идентификация множественных вариантов одного эпитопа по длине и наличие сильного HLA-DR4-связывающего участка убедительно свидетельствует в пользу того, что присутствующий в образцах сыворотки эпитоп ITIH4 является маркером RA.
Комлемент С4
В восьми из одиннадцати образцах RA сыворотки (эрозивный и неэрозивный RA) был идентифицирован другой доминантный эпитоп, выделяемый из комплемента С4: 15-мерный С4 (1697-1711) с аминокислотной последовательностью SEQ ID NO: 13 (Таблица 3). Можно обнаружить также пять вариантов указанного эпитопа по длине (Таблица 3): 12-мерный С4 (1697-1708; SEQ ID NO: 18), 13-мерный С4 (1698-1710; SEQ ID NO:17), 14-мерный С4 (1697-1710; SEQ ID NO: 15). 16-мерный С4 (1697-1712; SEQ ID NO: 14) и 18-мерный С4 (1697-1714; SEQ ID NO: 16). Кроме того, указанный эпитоп имеет очень сильный DRB1*0401-связывающий участок: 1700Y выполняет функцию Р1 якоря, 170D - Р2 якоря и 170N - Р6 якоря (показатель связывания: 1%).
С4, составляющий приблизительно 0,5% белковой массы плазмы, играет основную роль в активации центрального каскада реакций системы комплемента. Белок синтезируется в виде одноцепочечного предшественника и перед секрецией ферментативно расщепляется с формированием тримера из неидентичных α-, β- и γ-цепей. Идентифицированный эпитоп С4 (1697-1711) располагается на самом отдаленном С-концевом участке γ-цепи. α-цепь С4 дополнительно протеолитически расщепляется активированным С1 с формированием С4а анафилоксина, который является медиатором локальных воспалительных реакций (Moon, K.E. et а1., J Biol Chem 256 (1981) 8685-8692).
В целом, комплементный каскад участвует в индуцировании и прогрессировании воспалительных реакций и представляет собой основную систему защиты от различных патогенных агентов, включая бактерии, вирусы и другие антигены (Morgan, B.P., Methods Mol Biol 150 (2000) 1-13). Неадекватная активация, однако, может привести к повреждению тканей и манифестации заболевания (Speth, С.et al., Wien Klin Wochenshr 111 (1999) 378-391).
Неоднократно сообщалось о том, что активация системы комплемента имеет значение в патогенезе RA; основанием для таких утверждений является обнаружение повышенных уровней метаболитов комлемента, включая С4 и С4а, в плазме, синовиальной жидкости и синовиальной ткани пациентов с RA (Neumann, E. et al., Arthtritis Rheum 46 (2002) 934-945). Кроме того, коллаген-индуцированный артрит (CIA) у мышей характеризуется наличием продуктов активации комплемента (Linton, S.M. & Morgan, B.P., Mol. Immunol 36 (1999) 905-914). Развитие CIA можно предотвратить с помощью анти-С5-моноклональных антител (Wang, Y. et al., PNAS 92 (1995) 8955-8959) или с помощью растворимого CR1, ингибитора системы комлемента, вводимых с помощью генной терапии (Dreja, H. et al., Arthritis Rheum 43 (2000) 1698-1709). Активацию факторов комлемента в суставах индуцируют, возможно, различные присутствующие иммунные комплексы; было сделано предположение о том, что стимуляция внутренней иммунной системы инфекционными агентами и цитокинами может влиять на инициацию RA (Friese, M.A. et al., Clin Exp Immunol 121 (2000) 406-414).
Два из шести эпитопов С4, 15-мерный и 18-мерный, также были идентифицированы в двух контрольных образцах (Таблица 3), что свидетельствует о том, что только некоторые варианты указанного эпитопа С4 по длине являются RA-специфичными, а именно антигенные пептиды с SEQ ID NOs: 14, 15, 17 и 18.
Комплемент С3
Другой эпитоп, который был обнаружен в образцах эрозивного и неэрозивного RA, выделяют из комлемента С3 (альфа цепь): 14-мерный С3 (1431-1444) с аминокислотной последовательностью SEQ ID NO: 21 (Таблица 3). Также в сыворотке были идентифицированы шесть вариантов того же эпитопа по длине (Таблица 3): 13-мерный С3 (1431-1443; SEQ ID NO: 23), 14-мерный C3 (1429-1442; SEQ ID NO: 74), 15-мерный C3 (1431-1445; SEQ ID NO: 22), 15-мерный C3 (1429-1443; SEQ ID NO: 20), 17-мерный C3 (1427-1443; SEQ ID NO: 75) и 19-мерный C3 (1426-1444; SEQ ID NO: 19). При анализе самого короткого варианта, C3 (1431-1443), был установлен следующий DRB1*0401-связывающий участок: 1434Y в роли Р1 якоря, 1437D в роли Р4 якоря и 1439А в роли Р6 якоря.
В образцах эрозивного и неэрозивного RA также был обнаружен дополнительный эпитоп, выделяемый из комплемента C3 (бетα-цепь): 19-мерный C3 (157-175) с аминокислотной последовательностью SEQ ID NO: 76 (Таблица 3). Один дополнительный вариант по длине этого же эпитопа был идентифицирован в сыворотке (Таблица 3): 20-мерный C3 (157-176; SEQ ID NO: 77).
Комлемент C3, составляющий приблизительно 1-2% от общей белковой массы плазмы, играет основную роль в активации системы комлемента и относится к семейству белков острой фазы. Его процессинг с помощью C3 конвертазы с образованием C3a анафилоксина и C3b является центральным звеном как классического, так и альтернативного пути активации комплемента (Barrington, R. et al., Immunol Rev 180 (2001) 5-15). После активации C3b может ковалентно связываться (посредством реактивного тиоэфира) с углеводами на поверхности клеток или иммунными агрегатами (Isaac, L. & Isenman, D.E., J Biol Chem 267 (1992) 10062-10069). Идентифицированный эпитоп C3 (1431-1444) располагается на С-концевом участке C3b.
Как уже обсуждалось в отношении эпитопа комплемента С4 (1697-1711), появляется все больше доказательств того, что компоненты системы комлемента играют важную роль в патогенезе RA. Результаты настоящего исследования, в котором в сыворотке пациентов с RA были идентифицированы два основных эпитопа, выделенных из комплемента C3 и С4, подчеркивают тесную связь между активацией системы комлемента и патогенезом RA. Это совпадение является сильным аргументом в пользу того, что присутствующие C3/С4 эпитопы являются потенциальными сывороточными маркерами RA.
Протеин 3, подобный обогащенному глутаминовой кислотой связывающему SH3-домен протеину
Другим эптопом, который достаточно часто выявляется в сыворотке пациентов с RA (в шести из одиннадцати образцах эрозивного и неэрозивного RA), является эпитоп, выделяемый из протеина 3, подобного обогащенному глутаминовой кислотой связывающему SH3-домен протеину (SH3BGRL3): SH3BGRL3 (15-26) с аминокислотной последовательностью SEQ ID NO: 25 (Таблица 3). Также было идентифицировано три варианта указанного эпитопа по длине (Таблица 3): 14-мерный SH3BGRL3 (13-26; SEQ ID NO: 26), 14-мерный SH3BGRL3 (15-28; SEQ ID NO: 27) и 16-мерный SH3BGRL3 (13-28; SEQ ID NO: 24). DRB1*0401 -связывающий участок представлен: 171 в роли Р1 якоря, 20Q в роли Р4 якоря и 22S в роли Р6 якоря (показатель связывания 4%).
SH3BGRL3 представляет собой протеин с мол. массой 10кД, который принадлежит семейству SH3BGR. Предполагаемая функция белка неизвестна, однако отмечена его роль в качестве модулятора биологической активности глутаредоксина (Mazzocco, M. et al., Biochem Biophys Res Commun 285 (2001) 540-545). Таким образом, SH3BGRL3 не описан применительно к RA.
Интересно отметить, что выявлен второй эпитоп того же самого протеина, который высокоизбыточен во всех образцах RA и контрольных образцах: 16-мерный SH3BGRL3 (29-44) с аминокислотной последовательностью DGKRIQYQLVDISQDN. Кроме того, были обнаружены многочисленные варьирующие по длине варианты этого же эпитопа во многих образцах. При изучении наиболее короткого варианта, SH3BGRL3 (31-42), было установлено, что эпитоп содержит практически сходные с DRB1*0401 якорные остатки по сравнению с SH3BGRL3 (15-26): 331 служит в качестве Р1 якоря, 36Q - в качестве Р4 якоря и 38V - в качестве Р6 якоря (счет связывания - 2). Это сходство подтверждается сравнительным счетом связывания.
Наличие второго эпитопа SH3BGRL3 подтверждает достоверность эпитопа SH3BGRL3 (15-26), поскольку оба пептида происходят из одного протеина, однако только один из них, а именно эпитоп SH3BGRL3 (15-26), по-видимому, генерируется RA-специфическим образом. Сходное наблюдение уже описано для GILT в Примере 1.
Среди четырех различающихся по длине вариантов SH3BGRL3 наиболее длинноразмерный вариант, SH3BGRL3 (13-28), также идентифицирован в контрольном образце здорового индивидуума (Таблица 3). Однако этот вариант определенной длины обнаруживался только один раз, что свидетельствует о значительном обогащении эпитопом SH3BGRL3 при RA.
Индуцируемый интерлейкином-4 (ИЛ-4) протеин 1
Во всех исследованных синовиальных жидкостях (эрозивный и неэрозивный RA) и в восьми из 11 сывороток (эрозивный и неэрозивный RA) идентифицирован только высоко доминантный эпитоп, происходящий из гомологичного человеческому индуцируемого ИЛ-4 протеина 1(Fig1): Fig1 (293-309) с аминокислотной последовательностью SEQ ID NO: 28 (Таблица 3): 16-мерный Fig1 (293-308; SEQ ID NO: 30) и 19-мерный Fig1 (293-311; SEQ ID NO: 29). Более того, аминокислотная последовательность обнаруживает типичный DRB1*0401 связывающий участок: 299V служит в качестве Р1 якоря, 302Е - в качестве Р4 якоря и 304S - в качестве Р6 якоря (счет связывания 1%).
Два различающихся по длине варианта одного и того же эпитопа, Fig1 (293-308) и Fig1 (293-309) были идентифицированы в одном необработанном образце и в одном контрольном образце от здорового человека (Таблица 3). Однако наличие эпитопа Fig1 практически во всех образцах RA, но не во всех контрольных образцах строго свидетельствует об обогащении при RA.
Ген fig1 человека впервые идентифицирован в стимулируемых ИЛ-4 В-клеточных культурах (Chu, C.C. & Paul, W.E., PNAS 94 (1997)) 2507-2512). Этот ген локализован на хромосоме 19q13.3-19q13.4, на участке, для которого предварительно показана роль в развитии чувствительности к аутоиммунным заболеваниям, в том числе системной красной волчанке, артриту, множественному склерозу и инсулин-зависимому сахарному диабету (Becker K.G. et al., PNAS 95(1998) 9979-9984). Поскольку экспрессия указанного гена значительно ограничена иммунными тканями и его регуляция зависит от ИЛ-4, ключевого модулятора иммунного ответа, fig1, таким образом, является наиболее вероятным кандидатом на роль гена, определяющего чувствительность к аутоиммунным заболеваниям (Chavan, S.S. et al., Biochim Biophys Acta 1576 (2002) 70-80). Лимитированная по HLA-DR4 презентация эпитопа Fig1 свидетельствует, в первую очередь, о том, что протеин Fig1 продуцируется и, возможно, вовлечен в развитие RA. Полипептид Fig1 не был известен как маркер RA до настоящего времени и считается, что он является одним из важнейших кандидатов на указанную роль.
Гемопексин
Другой кандидат на роль маркера RA, часто обнаруживаемый в образцах сыворотки (10 из 11 образцов) и образцах синовиальной жидкости (два из 4 образцов) (эрозивный и неэрозивный RA), является производным гемопексина (НРХ): НРХ (351-367) с аминокислотной последовательностью SEQ ID NO: 32 (Таблица 3). Обнаружены некоторые различающиеся по длине варианты, которые подтверждают значимость указанного эпитопа (Таблица 3): 13-мерный НРХ (351-363; SEQ ID NO: 33), 14-мерный НРХ (350-363; SEQ ID NO: 34), 15-мерный НРХ (351-365; SEQ ID NO: 35), 18-мерный НРХ (351-368; SEQ ID NO: 31) и 18-мерный НРХ (350-367; SEQ ID NO: 78). Более того, эпитоп содержит очень сильно связывающий DRB1*0401 участок: 3551 служит в качестве Р1 якоря, 358D - в качестве Р4 якоря и 360V - в качестве Р6 якоря (счет связывания 1%).
Два различающихся по длине варианта того же самого эпитопа, а именно НРХ (351-367; SEQ ID NO: 32) и НРХ (351-365; SEQ ID NO: 35), также могут быть идентифицированы в контрольных образцах здорового человека (Таблица 3). Это свидетельствует о том, что только варианты определенной длины специфичны для RA, а именно антигенные пептиды SEQ ID NOs. 31, 33, 34 и 78.
НРХ представляет собой гликопротеин плазмы крови с мол. массой 60 кД с высокой связывающей аффинностью к гему (Muller-Eberhard, U., Methods Enzymol 163 (1988) 536-565). Он главным образом экспрессируется в печени и принадлежит к белкам острой фазы, синтез которых индуцируется при воспалительных процессах. Ревматоидный артрит представляет собой хроническое воспалительное аутоиммунное воспалительное заболевание, для которого характерны повышенные уровни белков острой фазы, в том числе С-реактивного белка и сывороточного амилоида A (Nakamura, R., J Clin Lab Anal 14 (2000) 305-313). НРХ отвечает на цитокины ИЛ-1 и ИЛ-6, которые регулируются повышенным образом у пациентов с RA (Feldmann, М. & Maini, R.N., Rheumatology 38, Suppi 2 (1999) 3-7).
НРХ является основным носителем для транспортировки гема в плазме крови, и его принципиальная роль заключается в предотвращении опосредованного гемом окислительного стресса и потери связанного с гемом железа (Tolosano, Е. & Altruda, F., DNA Cell Biol 21 (2002) 297-306). Он может защитить клетки от окислительного стресса индуцирование экспрессии внутриклеточных антиоксидантов, таких как оксигеназа гема, металлотионеины и ферритин. Металлотионеины и цитозольные протеины экспрессируются в основном в синовиальных фибробластах (Backman, J. Т. et al., Virchows Arch 433 (1998) 153-160). Существует значительное экспериментальное свидетельство в пользу наличия окислительного стресса в синовиальных тканях у больных с RA (см. обзор Schett, G. et al., Arthritis Res 3 (2000) 80-86). Более того, сообщалось о том, что НРХ индуцирует пролиферацию Т-лимфоцитов человека (Smith, A. et al., Exp Cell Res 232 (1997) 246-254). Указанные данные подтверждают вероятность того, что НРХ принадлежит к белкам, которые регулируются повышенным образом в сыворотке крови и синовиальной жидкости больных с RA, свидетельствуя в пользу того, что НРХ (351-367) является кандидатом на роль RA-специфического маркера.
Hsc70-интерактивный протеин
Эпитоп, который главным образом идентифицируется в образцах сыворотки (4 из 11 образцов) (эрозивный и неэрозивный RA) и который также связан с ответом на стресс, происходит из Hsc70-интерактивного протеина Hip: Hip (83-98) с аминокислотной последовательностью SEQ ID NO: 38 (Таблица 3).
Идентифицированы два различающихся по длине варианта указанного эпитопа (Таблица 3): 18-мерный Hip (83-100; SEQ ID NO: 36) и 15-мерный Hip (84-98; SEQ ID NO: 39). Идентифицирован также дополнительный вариант в одном образце синовиальной жидкости (Таблица 3): 15-мерный Hip (85-99; SEQ ID NO: 37). При изучении наиболее короткого варианта Hip (84-98) было установлено, что эпитоп содержит связывающий DRB1*0401 участок, дающий счет связывания 8%: 891 служит в качестве Р1 якоря, 92D - в качестве Р4 якоря и 94D - в качестве Р6 якоря.
В цитозоле эукариотических клеток протеины Hip и Нор ассоциированы с Hsc70 с целью участия в регуляции хаперонной активности Hsc70 (Frydman, J. & Hohfeld, J., Trends Biochem Sci 22 (1997) 87-92). Протеин Hip с мол. массой 42 кД связывается с АТФазным доменом Hsc70. Постулировано, что Hip может увеличивать время полужизни комплекса хаперон-субстрат, создавая молекулярную основу для эффективного взаимодействия Hsc70 с регулируемой понижающим образом хапероновой системой. Показано, что Hsc70 и Hsc90 взаимодействуют в процессе сборки протеина in vitro (Jakob, U. & Buchner, J., Trends Biochem Sci 19 (1994) 205-211; Freeman, B.C. & Morimoto, R.I., EMBO J 15 (1996) 2969-2979) и играют определенную роль в температурной денатурации (Schneider, С.et al., PNAS 93 (1996) 14536-14541). Тот факт, что Hsc70 и Hsc90 ассоциируются с адаптацией к стрессу, неоднозначным образом связывает Hip с ответами на стресс, в том числе индукцию белков теплового шока в синовиальной ткани больных RA (см. обзор Schett, G. et al., Arthritis Res 3 (2001) 80-86).
Связывающие свойства ITIH4. С4. C3. SH3BGRL3. Fig1. HPX и Hip
Для дальнейшего изучения связывающих свойств описанных выше антигенных пептидов ITIH4, С4, C3, SH3BGRL3, Fig1, HPX и Hip с молекулами HLA-DR осуществляют исследования связывания in vitro (Фиг.3): в соответствии с их значениями IC50 против репортерного пептида НА (293-309) синтетические пептиды ITIH4 (274-287), SH3BGRL3 (13-26) и Fig1(293-309) связываются с HLA-DR с высокой аффинностью (Фиг.3). Умеренное связывание с HLA-DR определяют для синтетических пептидов С4 (1696-1709) и HPX (351-365). Синтетический пептид C3 (1431-1444) связывается слабо с HLA-DR, что согласуется со счетом TEPITOPE (9%). Не определяют связывание для пептида Hip (84-98) (данные не приведены).
Инвариантная цепь (Ii)
Среди всех кандидатов на маркерные пептиды, которые, по-видимому, могут быть индикаторами эрозивного и неэрозивного RA, существует эпитоп, который происходит из ассоциированной с HLA-DR инвариантной цепи (Ii): 15-мерная Ii (110-124; SEQ ID NO: 83) с аминокислотной последовательностью ATPLLMQALPMGALP (Таблица 3). Более того, идентифицированы четыре различающихся по длине варианта указанного эпитопа: 16-мерная Ii (109-124; SEQ ID NO: 81), 17-мерная Ii (109-125; SEQ ID NO: 80), 18-мерная Ii (109-126; SEQ ID NO: 79) и 21-мерная Ii (109-129; SEQ ID NO: 82) (Таблица 3). Наличие различающихся по длине вариантов соответствует идентифицированной пептидной последовательности, обнаруживающей участок умеренного связывания DRB1*0401 (счет связывания TEPITOPE составляет 5%), причем 114L служит в качестве Р1 якоря, 117А - в качестве Р4 якоря и 119Р - в качестве Р6 якоря. Эпитоп был идентифицирован в двух образцах эрозивного и в трех образцах неэрозивного RA. Два различающихся по длине варианта Ii было обнаружено в одном необработанном образце и в двух образцах от здоровых индивидуумов.
Пептиды, нагруженные молекулами МНС класса II, регулируются двумя вспомогательными молекулами пути класса II, а именно инвариантной цепью Ii и HLA-DM (см. обзор Bakke, О. & Nordeng T.W., Immunol Rev 172 (1999) 171-187 и Kropshofer, Н. et al., Immunol Today 18 (1997) 77-82). Тримеры Ii связываются с созревающими молекулами МНС класса II в шероховатом эндоплазматическом ретикулуме (ЭР) и блокируют пептид-связывающий желобок, стабилизируя молекулу МНС класса II и предотвращая связывание лигандов, доступных в ЭР. Комплексы "молекулы класса II/Ii" транспортируются через аппарат Гольджи к эндосомам, где Ii расщепляется с образованием гнездового набора ассоциированных с молекулами класса II пептидов Ii (CLIP). Высвобождение CLIP позволяет эндосомальным пептидам связываться с молекулами МНС класса II, которые презентируются рецепторам Т-клеток на клеточной поверхности. Идентифицированный эпитоп Ii (110-124) перекрывается с С-концевым участком CLIP. Интересно отметить, что описанное в литературе редуцированное взаимодействие CLIP с RA-ассоциированными аллелями HLA-DR (Patil, N.S. et al., J Immunol 167 (2001) 7157-7168), указывает на то, что такое взаимодействие может вносить определенный вклад в патофизиологию аутоиммунитета при RA.
Отвечающий на рецептор ретиноевой кислоты протеин 2
В четырех из одиннадцати образцов сыворотки (эрозивный и неэрозивный RA) был идентифицирован эпитоп, происходящий из отвечающего на рецептор ретиноевой кислоты протеина 2 (RARRES2):22-мерный RARRES2 (40-61; SEQ ID NO: 86) с аминокислотной последовательностью HPPVQWAFQETSVESAVDTPFP (Таблица 3). В С-концевом участке эпитопа могут быть идентифицированы два различающихся по длине варианта: 23-мерный RARRES2 (40-62; SEQ ID NO: 84) и 24-мерный RARRES2 (40-63; SEQ ID NO: 85) (Таблица 3). 45W служит в качестве PI якоря, 48Q - в качестве Р4 якоря и 50Т - в качестве Р6 якоря; в эпитопе обнаруживается участок умеренного связывания DRB1*0401 (счет связывания TEPITOPE составляет 3%).
RARRES2 представляет собой небольшой протеин с мол. массой 18, 6 кД, который главным образом экспрессируется в эндотелии и эпидермисе. Экспрессия, по-видимому, является гормонзависимой и выявлены ответы на ретиноевую кислоту в коже и некоторые остеотрофные гормоны в происходящих из костного мозга стромальных клетках (Nagpal, S. et al., J Invest Dermatol 109 (1997) 91-95 и Adams, A.E. et al., J Cell Biochem 74 (1999) 587-595). Функция RARRES2 практически неизвестна, и его ассоциация с RA, например, за счет ослабления остеокластогенеза является предполагаемой.
Фибронектин
Другой HLA-DR4-ассоциированный пептид, который избирательно выявляется в трех из четырех образцах синовиальной жидкости (эрозивный и неэрозивный RA), происходит из фибронектина (Fn), основного гликопротеина плазмы крови и внеклеточного матрикса: 15-мерный Fn (1881-1895; SEQ ID NO: 90) с аминокислотной последовательностью IYLYTLNDNARSSPV (Таблица 3). Эпитоп обнаруживает очень хорошее связывание с DRB1*0401, причем 1885Y служит в качестве гидрофобного Р1 якоря, 188N - в роли Р4 якоря и 1890N - в роли Р6 якоря (счет связывания TEPITOPE составляет 1%). Дополнительно идентифицированы четыре различающихся по длине варианта: 16-мерный Fn (1881-1896; SEQ ID NO:91), 16-мерный Fn (1880-1895; SEQ ID NO:88), 17-мерный Fn (1881-1897; SEQ ID NO:89) и 17-мерный Fn (1880-1896; SEQ ID NO:87), что строго подтверждает значимость идентифицированного Fn-эпитопа (Таблица 3).
Фибронектин играет важную роль в адгезии клеток, подвижности клеток и опсонизации. Он связывает коллаген и фибрин и опосредует адгезию фибробластов к фибриллам коллагена. Фибронектин строго экспрессируется в выстилающем синовиальную полость слое у больных как RA, так и остеоартритом и коррелирует с гиперплазией больных суставов. Идентификация эпитопа Fn (1881-1895) только в образцах синовиального происхождения согласуется с этими данными и позволяет предположить возможную роль высокоизбыточного фибронектина в развитии аутоиммунитета.
Катепсин В
В трех из пятнадцати образцов RA (эрозивный и неэрозивный RA) выявляется эпитоп, происходящий из катепсина В (CatB): 15-мерный CatB (227-241; SEQ ID NO: 92) с аминокислотной последовательностью YNSYSVSNSEKDIMA (Таблица 3). Эпитоп обнаруживает умеренное связывание с DRB1*0401, причем 230Y служит в качестве гидрофобного Р1 якоря, остатки серина в положениях 233 и 235 - в роли возможных Р4 и Р6 якорей (счет связывания TEPITOPE составляет 6%).
Катепсин В представляет собой тиоловую пептидазу и, по-видимому, принимает участие во внутриклеточной деградации и круговороте протеинов, например коллагена. Он локализован в лизосомах клеток различных типов, включая лейкоциты. Показано, что катепсин В вносит определенный вклад в разрушение хряща при остеоартрите и патологическом протеолизе при RA и раке (Cunnane, G. et al., Arthritis Rheum 44 (2001) 1744-1753). Энзиматическая активность катепсина В и других лизосомальных пептидаз коррелирует с прогрессией RA, что подтверждает значимость эпитопа CatB (227-241) как возможного маркерного пептида RA (Sohar, N. et al., Biol Chem 383 (2002) 865-869).
Трипептидил-пептидаза II
Другой происходящий из пептидазы эпитоп был обнаружен в трех из пятнадцати образцов RA (эрозивный и неэрозивный RA), который происходит из трипептидил-пептидазы II (ТРР2): 15-мерный ТРР2 (970-984; SEQ ID NO: 93) с аминокислотной последовательностью AGSLTLSKTELGKKA (Таблица 3). Дополнительно выявляется отличный по длине вариант ТРР2 (970-985; SEQ ID NO: 94). Пептидазный эпитоп содержит типичный связывающий DRB1*0401 участок, причем 973L, 976S и 978Т служат в качестве Р1, Р4 и Р6 якорей соответственно (счет связывания TEPITOPE составляет 3%).
Аналогично катепсину В Трипептидил-пептидаза II вовлечена в расщепление лизосомальных протеинов. Идентификация эпитопа ТРР2 (970-984) в контексте RA свидетельствует, в первую очередь, о том, что ТРР2 может быть задействован в ослаблении деградации белков при воспалительных процессах и RA.
Легумаин
Легумаин (LGMN) завершает набор из трех пептидаз, которые обнаруживаются в данном исследовании. Идентифицированный эпитоп LGMN (99-112; SEQ ID NO: 95) с аминокислотной последовательностью VPKDYTGEDVTPQN (Таблица 3) обнаруживает типичный участок связывания с аллелем HLA DRB1*0401, причем 103Y служит в качестве Р1 якоря, 106Е - в качестве Р4 якоря, a 108V - в качестве P6 якоря (счет связывания TEPITOPE составляет 1%).
Легумаин выявляется в эндосомальных и лизосомальных фракциях антиген-презентирующих клеток, например дендритных клеток (Schwarz, G. et al., Biol Chem 383 (2002) 1813-1816). Он служит исключительно для гидролиза аспарагиниловых связей. Показано, что он играет важную роль в процессировании бактериальных антигенов для молекул МНС класса II (Manoury, В. et al., Nature 396 (1998) 695-699). Остается неизвестным, вовлечен ли легумаин в развитие аутоиммунных заболеваний, таких как RA, но тот факт, что три лизосомальные пептидазы вместе с GILT и 268-протеасомой идентифицированы в процессе данного исследования, позволяет предположить, что механизмы расщепления протеинов для процессирования антигена могут быть существенным образом изменены при RA.
Рецептор фактора активации тромбоцитов
В трех из одиннадцати образцов сыворотки от больных RA (эрозивный и неэрозивный RA) был идентифицирован эпитоп, происходящий из рецептора фактора активации тромбоцитов (PAFR): 13-мерный PAFR (264-276; SEQ ID NO: 96) с аминокислотной последовательностью DSKFHQAINDAHQ (Таблица 3). В эпитопе согласно счету связывания TEPITOPE (3%) обнаруживается участок умеренного связывания DRB1*0401, причем 267F служит в качестве Р1 якоря, 270А - в качестве Р4 якоря и 272N - в качестве Р6 якоря (Таблица 3).
Фактор активации тромбоцитов (PAF) представляет собой провоспалительный липидный медиатор, который связывается с соединенным с G-белком седьмым трансмембранным рецептором на поверхности целого ряда клеток различных типов. За счет связывания с рецептором PAF трансдуцирует плейотропные функции, в том числе подвижность клеток, сокращение гладких мышц, а также синтез и высвобождение цитокинов (см. обзор Honda, Z. et al., J Biochem 131 (2002) 773-779). Фармакологические исследования и получение PAFR (-/-) мышей позволяет предположить, что функции PAF различны и вовлечены в аллергические реакции, воспаление, активность нейронов, репродукцию и развитие атеросклероза. Интересно отметить, что PAF обнаруживается в повышенных концентрациях в синовиальной жидкости больных с RA и показана его роль в индукции неоангиогенеза, который часто наблюдается при ревматоидном синовите (Lupia, E. et al., Eur J Immunol 26 (1996) 1690-1694). Это дополнительно свидетельствует в пользу того, что эпитоп PAFR (264-276) является ассоциированным с RA маркерным пептидом.
Поли-альфа-2.8-сиалилтрансфераза
Другой эпитоп, который был обнаружен в трех из одиннадцати образцов сыворотки (эрозивный и неэрозивный RA), происходит из полисиалилтрансферазы (PST): 15-мерный PST (333-347; SEQ ID NO: 97) с аминокислотной последовательностью MPLEFKTLNVLHNRG (Таблица 3). Он содержит умеренно связывающий DRB1*0401 участок (счет связывания TEPITOPE составляет 2%), причем 337F, 340L и 342 V служат в качестве PI, P4 и Р6 якорей соответственно.
Полисиаловая кислота представляет собой углевод, состоящий из линейного гомополимера альфа-2,8-связанных остатков сиаловой кислоты. Гликан, главным образом, связан с молекулой адгезии нейронов (N-CAM) и вовлечен во многие процессы морфогенеза нейронов за счет модулирования адгезионных свойств N-CAM. Мембранный протеин полисиалилтрансфераза катализирует поликонденсацию остатков сиаловой кислоты и высокоэкспрессируется в головном мозге, легких и почках плода, в сердце, селезенке и тимусе взрослого человека и менее эффективно экспрессируется в лейкоцитах периферической крови (Nakayama, J. et al., Proc Nati Acad Sci (1995) 7031-7035). Повышенные уровни PST выявляются в сыворотке больных с метастазирующими опухолями, а повышенная активность данного фермента ассоциирована с ревматоидным артритом (Berge, P.O. et al., Klin Wochenschr 60 (1982) 445-449). Идентификация эпитопа PST (333-347) в образцах от больных RA свидетельствует, в первую очередь, о том, что этот фермент может быть задействован в развитии RA.
Ras-родственный протеин Rab-11B
Последний эпитоп, который был обнаружен в образцах сыворотки от больных RA, происходит из Ras-родственного протеина Rab-11B:13-мерный Rab-11B (51-63; SEQ ID NO: 102) с аминокислотной последовательностью RSIQVDGKTIKAQ (Таблица 3). Пептидная последовательность была подтверждена по чувствительности связывания аллеля RA DRB1*0401 при использовании программного продукта TEPITOPE (Hammer, J. et al., Adv Immunol 66 (1997) 67-100). Этот эпитоп и четыре дополнительных различающихся по длине варианта, а именно 14-мерный Rab-11 В (50-63; SEQ ID NO:100), 15-мерный Rab-11B (49-63; SEQ ID NO:98), 17-мерный Rab-11B (49-65; SEQ ID NO:101) и 18-мерный Rab-11 В (49-66; SEQ ID NO:99), обнаруживаются в трех образцах от больных RA (эрозивный и неэрозивный RA) (Таблица 3). 15-мерный Rab-11B (49-63) был идентифицирован в одном из двенадцати контрольных образцов.
Протеины Rab представляют собой небольшие GTPазы, которые играют важную роль в мембранном переносе как эндоклеточных, так и экзоклеточных путей. Rab-11B играет существенное значение в транспорте интернализованного трансферрина из рециклизующегося компартмента к плазматической мембране. Более того, Rab-11B выявлен в остеокластах крысы и может играть дополнительную роль в резорбции кости. Эпитоп Rab-11B (51-63) указывает на то, что Rab-11 В вовлечен в развитие RA.
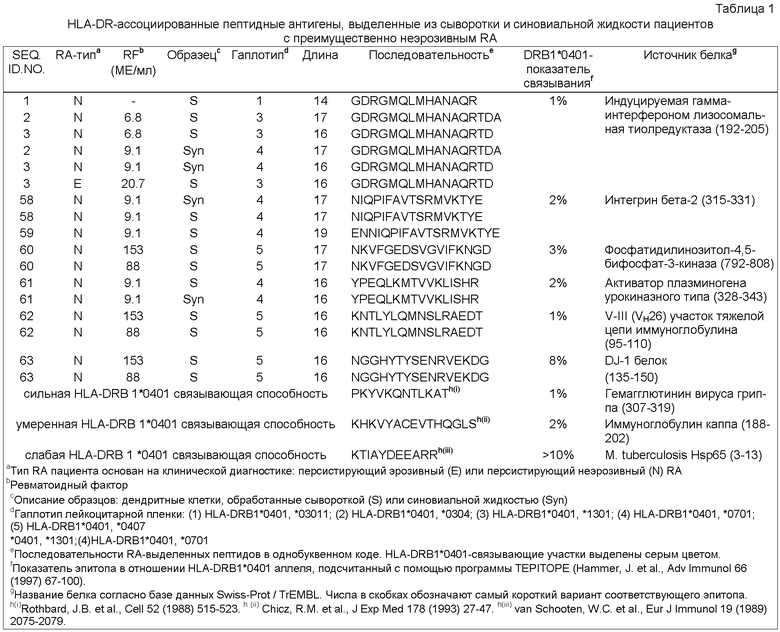
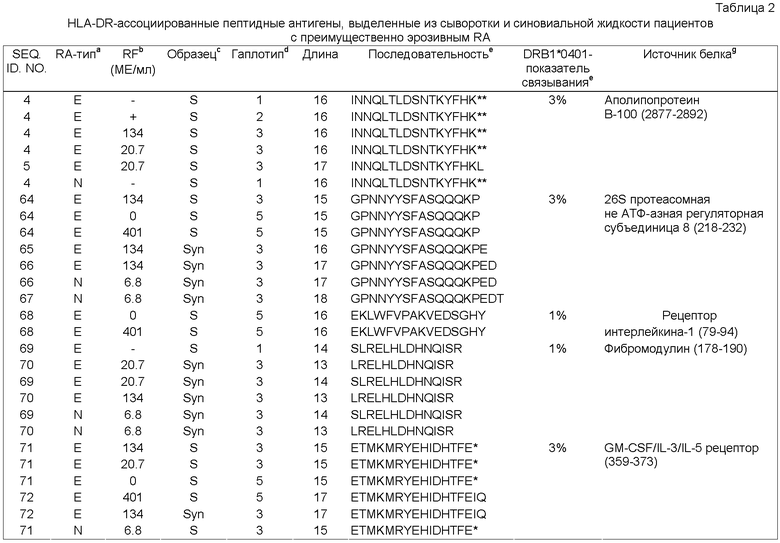
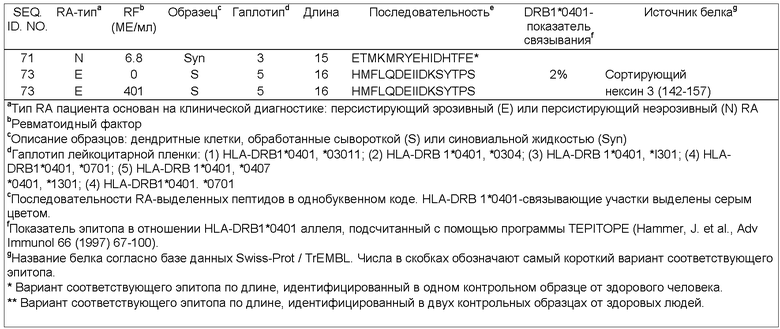
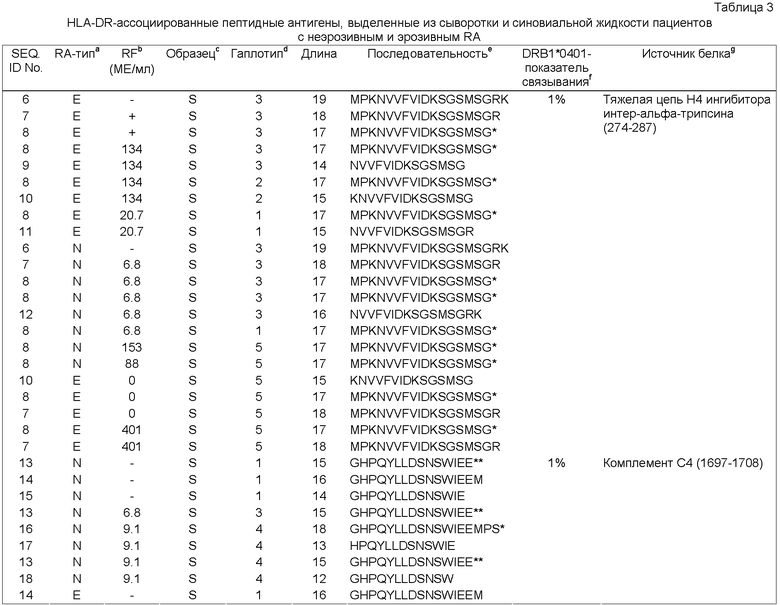
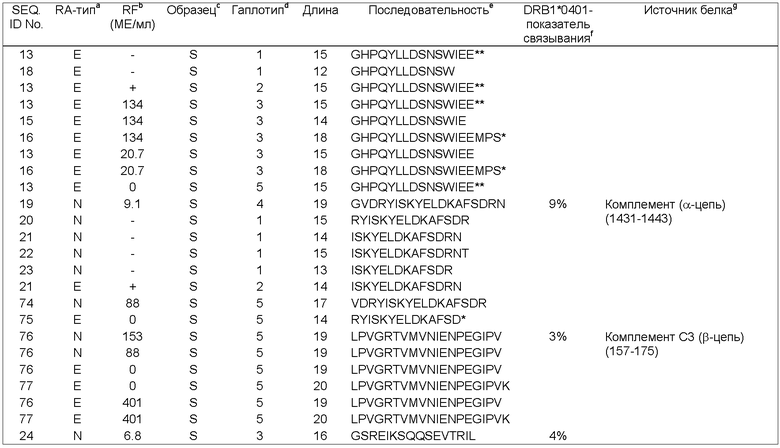
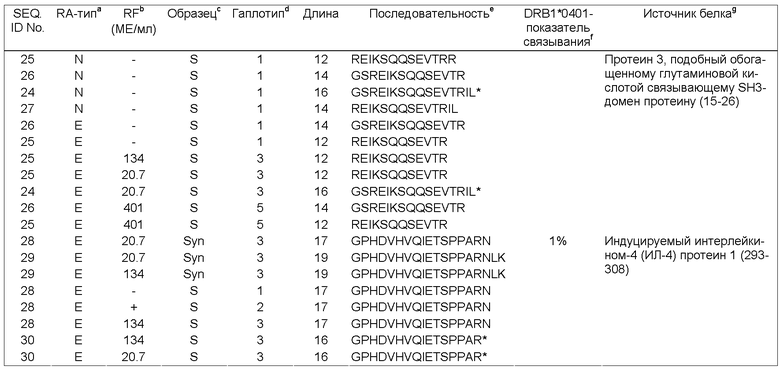
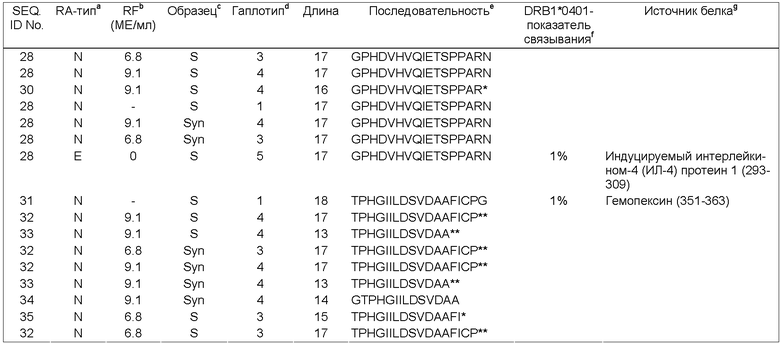
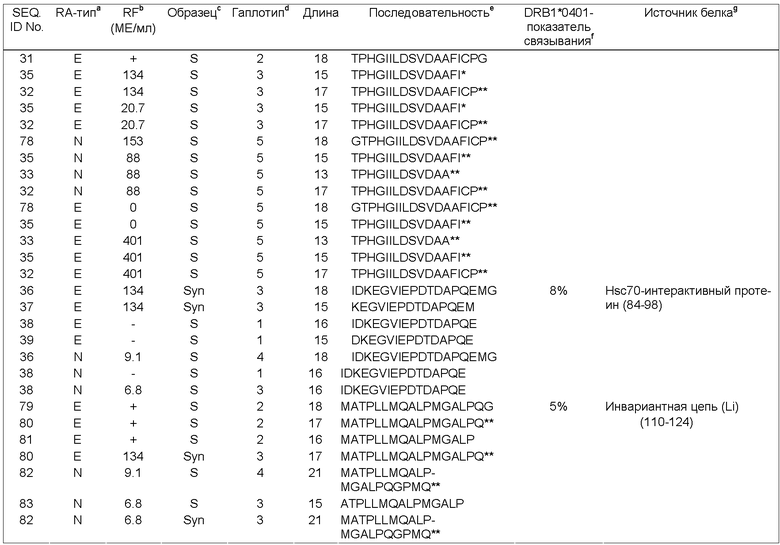

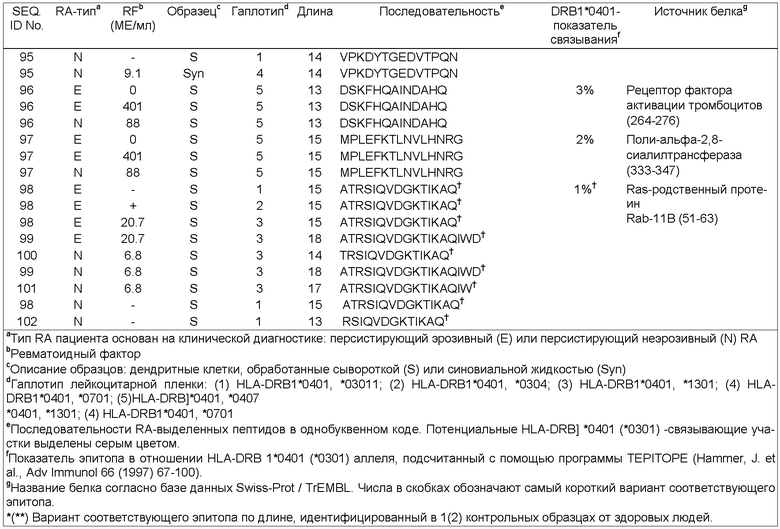
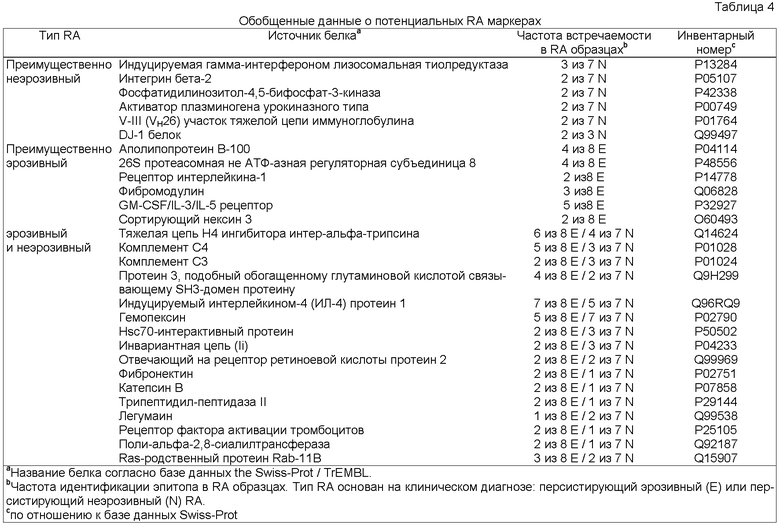
Изобретение относится к биотехнологии. Настоящее изобретение относится к новым антигенным пептидам класса II главного комплекса гистосовместимости, которые связаны с молекулой МНС класса II. Данное изобретение позволяет расширить арсенал технических средств, используемых в ранней диагностике ревматоидного артрита. 1 з.п. ф-лы, 5 ил., 4 табл.
1. Антигенные пептиды МНС класса II, являющиеся маркерами ревматоидного артрита и предрасположенности к нему и включающие: а) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 49 или б) по крайней мере аминокислотную последовательность пептидсвязывающего участка SEQ ID NO. 49 с дополнительными N- и С-концевыми фланкирующими последовательностями, соответствующей последовательности, которая выбрана из группы, включающей SEQ ID NOs. 1-3.
2. Антигенные пептиды МНС класса II по п.1, связанные с молекулой МНС класса II.
| Tsark Е.С | |||
| et al | |||
| Differential МНС class II-mediated presentation of rheumatoid arthritis autoantigens by human dendritic cells and macrophages | |||
| // J Immunol | |||
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| Универсальный двойной гаечный ключ | 1920 |
|
SU169A1 |
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| ВЕСЫ | 1927 |
|
SU6625A1 |
| Santambrogio L | |||
| et al | |||
| Extracellular antigen processing and presentation by immature dendritic cells | |||
| // Proc Natl Acad Sci USA | |||
| Металлический водоудерживающий щит висячей системы | 1922 |
|
SU1999A1 |
| Приспособление в пере для письма с целью увеличения на нем запаса чернил и уменьшения скорости их высыхания | 1917 |
|
SU96A1 |
| Прибор для получения стереоскопических впечатлений от двух изображений различного масштаба | 1917 |
|
SU26A1 |
| КАРТОТЕКА | 1928 |
|
SU15056A1 |
| RU 2003105806 от 20.04.2004. | |||

