Изобретение относится к области генетической инженерии и медицины и может быть использовано для создания лекарственных препаратов для лечения инсулинзависимого сахарного диабета.
Инсулин - пептидный гормон поджелудочной железы - синтезируется специализированными клетками, называемыми островками Лангерганса, в виде предшественника - препроинсулина, состоящего из 109 аминокислотных остатков. В процессе биосинтеза в результате отщепления сигнального пептида из 23 аминокислот высвобождается проинсулин. В дальнейшем проинсулин подвергается дальнейшему процессингу, приводящему к образованию собственно инсулина в результате специфического "выщепления" 35 аминокислот C-пептида. Зрелый биологически активный инсулин состоит из двух цепей, соединенных двумя дисульфидными связями. Цепь A содержит 21, а цепь B - 30 аминокислотных остатков.
Усилия многочисленных коллективов в последние 30 - 35 лет направлены на разработку промышленных методов получения инсулина человека, поскольку использование самого близкого по структуре к человеческому свиного инсулина зачастую приводит к нежелательным побочным эффектам. Хотя химический синтез инсулина человека осуществлен еще в 60-х годах, он не может служить основой для промышленного получения гормона вследствие его чрезвычайно низкой экономичности. В настоящее время известно два основных способа получения этого пептидного гормона человека.
Один из методов основан на использовании "полусинтетического" метода превращения свиного инсулина в человеческий. Эти белки отличаются единственной аминокислотой на C-конце B-цепи. В этом положении в свином инсулине находится Ala, а в инсулине человека - Thr. Наиболее технологичным вариантом метода является использование иммобилизованной протеиназы I из Achromobacter Lyticus для удаления C-концевого аланина инсулина свиньи с последующей конденсацией дез-Ala30-инсулина с третбутиловым эфиром треонина. Однако наиболее перспективным способом получения инсулина человека является способ, основанный на технологии рекомбинантных ДНК.
Ген, кодирующий биосинтез проинсулина человека, был получен около 20 лет назад как синтезом кДНК на матрице мРНК, выделенной из поджелудочной железы человека [1,2] , так и химико-ферментативным синтезом [3,4]. Однако попытки достичь высокого уровня биосинтеза проинсулина клетками кишечной палочки не увенчались успехами, что связано с быстрой деградацией кодируемого искусственным геном продукта. К настоящему времени разработана методология экспрессии гена проинсулина человека в клетках E. coli. Она заключается в биосинтезе клетками бактерий проинсулина в составе гибридных белков в виде нерастворимых "тел включения". В качестве лидерных последовательностей, защищающих рекомбинантный белок от протеолитической деградации, использовали гомоолигопептиды [5] , бычий протимозин [6], глутатион-S-трансферазу [7], C-пептид проинсулина [8] и др. Отщепление проинсулина от лидерного пептида достигается обработкой бромцианом по остатку метионина [9, 10]. Такая обработка становится возможной благодаря отсутствию метионина в последовательности проинсулина человека. Последующая ренатурация проинсулина и энзиматическая трансформация в инсулин при помощи трипсина и карбоксипептидазы B представляет собой хорошо отработанный в технологическом отношении процесс. Основным недостатком описанного подхода является использование высокотоксичного соединения - бромциана в крупномасштабном производстве.
Недавно было обнаружено, что гибридный белок, состоящий из двух усредненных иммуноглобулинсвязывающих (IgG) доменов белка A из S. aureus и проинсулина человека, соединенных через остатки Arg, LysArg или Lys, после выделения из "тел включения" и ренатурации может быть с высоким выходом и специфически расщеплен смесью трипсина и карбоксипептидазы B [11], что позволяет исключить из технологии высокотоксичный продукт. Образующиеся при этом инсулин и C-пептид могут быть изолированы при помощи обращеннофазовой хроматографии. Очистка гибридного белка производится с помощью аффинной хроматографии на IgG-сефарозе. Следует отметить, что, несмотря на очевидные преимущества, используемый в работе [11] подход обладает существенным недостатком, поскольку не поддается промышленному масштабированию из-за использования IgG-сефарозы.
Наиболее близким по технической сущности к предлагаемому изобретению является рекомбинантная плазмидная ДНК pTrpZZ-R-proinsulin [11], кодирующая гибридный белок массой около 27 кДа, в котором тандем двух IgG-связывающих доменов соединен через остаток аргинина с аминокислотной последовательностью проинсулина человека.
Задачей предлагаемого изобретения является конструирование плазмиды, детерминирующей синтез гибридного белка, в котором единственный IgG-связывающий домен белка A соединен через пептидный линкер His6AspGlyArg с аминокислотной последовательностью проинсулина человека. Преимущества предлагаемого конструкта заключаются, во-первых, в упрощении аффинной очистки гибридного белка за счет использования более технологичного аффинного сорбента, содержащего остатки способной к образованию металлохелатных комплексов иминодиуксусной кислоты, и, во-вторых, в увеличении доли проинсулина в гибридном белке за счет удаления одного IgG-связывающего домена.
Поставленная задача решается конструированием рекомбинантной плазмидной ДНК pPINS07 и штамма Escherichia coli JM109/pPINS07, обеспечивающего синтез гибридного полипептида с уровнем экспрессии не ниже 25-30%.
Исходной плазмидой для конструирования нового гена, кодирующего гибридный белок, содержащий проинсулин человека, послужила плазмида pPINS03. Эта плазмида была сконструирована на основе векторной плазмиды рКК223-3, у которой был удален сайт рестриктазы BamHI, расположенный перед tac-промотором, при помощи достройки "липких" концов после частичного гидролиза и последующего лигирования полученных "тупых" концов. Полученная таким образом плазмида рКК223-3* была использована в качестве вектора для одновременного клонирования по сайтам EcoRI и HindIII фрагментов, кодирующих домен B белка A из S. aureus и проинсулин человека. Искусственный ген, кодирующий проинсулин человека, фланкированный сайтами рестриктаз BamHI и HindIII, был получен химико-ферментативным синтезом. Фрагмент ДНК, кодирующий IgG-связывающий домен белка A, был синтезирован при помощи полимеразной цепной реакции на матрице ДНК из S. aureus (штамм Cowan) с использованием специфических праймеров. Перед клонированием для генерации липких концов амплификат был обработан рестриктазами EcoRI и BamHI. Полученная в результате плазмида pPINS03 содержит между сайтами EcoRI и HindIII искусственный ген, кодирующий гибридный белок, в котором аминокислотная последовательность IgG-связывающего домена B белка A из S. aureus через аминокислотный линкер Gly-Ser-Met соединена с аминокислотной последовательностью проинсулина человека. Для увеличения уровня биосинтеза гибридного белка выбор кодонов проинсулина был сделан с учетом частоты их встречаемости в высокоэкспрессируемых генах Е. coli. Плазмида pPINS03 обеспечивает в клетках Е. coli JM109 после индукции изопропилтио-β,D-галактопиранозидом (IPTG) высокий уровень биосинтеза гибридного белка. Однако существенным недостатком такой системы экспрессии является необходимость использования высокотоксичного бромциана для расщепления гибридного белка. Чтобы устранить этот недостаток с помощью олигонуклеотиднаправленного мутагенеза была получена новая рекомбинантная плазмида pPINS05, в которой аминокислотные последовательности IgG-связывающего домена белка A и проинсулина человека соединены через пептидный линкер GlySerArg. Сайт-направленный мутагенез проводили с помощью полимеразной цепной реакции. Для этого на матрице плазмидной ДНК проводили 20 циклов амплификации (94oC, 30 с; 48oC, 45 с; 72oC, 45 с) с мутагенизирующим праймером
AACAAAGGTACCCGTTTTGTTAACCAAC-
ACC и якорным праймером TCCGCCAA-
AACAGAAGCT. Продукт амплификации гидролизовали рестриктазами BamHI и HindIII и клонировали в вектор, полученный гидролизом плазмиды pPINS03 теми же эндонуклеазами. В результате получили рекомбинантную плазмидную ДНК pPINS05. Полученная таким образом плазмида pPINS05 детерминирует биосинтез гибридного белка, от которого проинсулин человека может быть освобожден ферментативным гидролизом смесью трипсина и карбоксипептидазы B.
Для упрощения выделения и очистки гибридного белка в его состав были введены гексагистидиновые последовательности, позволяющие очистить его в одну стадию с помощью аффинной хроматографии на металлохелатных колонках. С этой целью ДНК плазмиды pPINS05 была подвергнута исчерпывающему гидролизу рестриктазой BamHI и полученные фрагменты лигировали с синтетическим олигонуклеотидным дуплексом: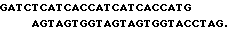
Лигазную смесь использовали для трансформации компетентных клеток Е. coli JM109. Полученная в результате рекомбинантная плазмида pPINS07 была проанализирована гидролизом эндонуклеазами HaeIII и HpaII, а также совместным гидролизом рестриктазами EcoRI и HindIII. Структура гена, кодирующего гибридный белок, была подтверждена секвенированием по модифицированному методу Максама-Гилберта [12] (фиг. 1).
Рекомбинантная плазмидная ДНК pPINS07, кодирующая гибридный полипептид с аминокислотной последовательностью проинсулина человека, характеризуется следующими признаками:
имеет молекулярную массу 3,3 МДа (5,051 т.п.о.);
кодирует гибридный белок, в котором последовательность домена B стафилококкового белка A с C-концевым гексагистидиновым линкером соединена через трипептид GlySerArg с аминокислотной последовательностью проинсулина человека;
состоит из: EcoRI/HindIII-фрагмента плазмиды рКК223-3*, содержащей tac-промотор транскрипции, ген β-лактамазы (bla), участок инициации репликации (ori), терминатор транскрипции рибосомого оперона E. coli [16], EcoRI/BamHI-фрагмента, включающего искусственный ген, кодирующий IgG-связывающий домен белка A из S. aureus и гексагистидиновый домен, и BamHI/HindIII-фрагмента, кодирующего последовательновть аргинил-проинсулина человека;
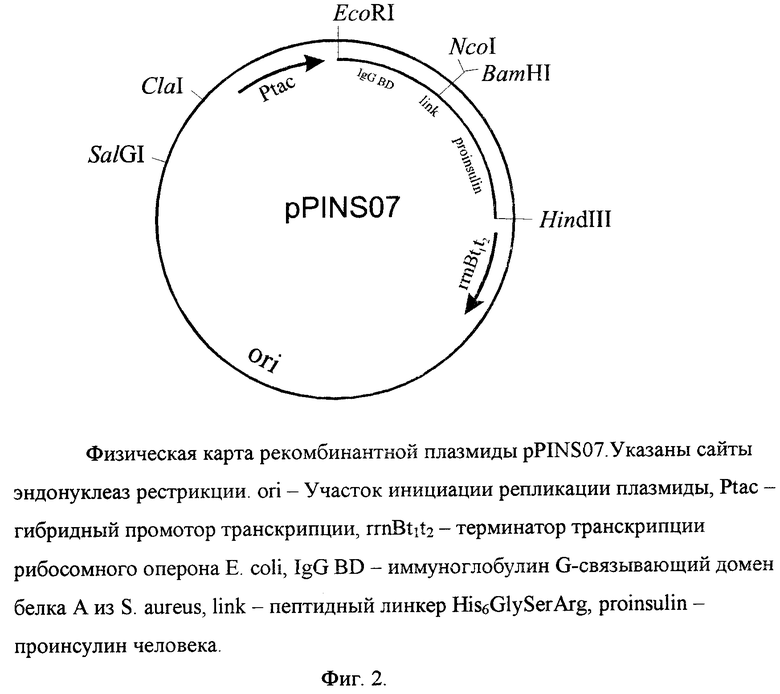
содержит: гибридный tac-промотор транскрипции, искусственный ген, кодирующий гибридный полипептид, в котором последовательность домена B стафилококкового белка A с C-концевым гексагистидиновым линкером соединена через трипептид GlySerArg с аминокислотной последовательностью проинсулина человека; в качестве генетического маркера ген β-лактамазы (bla), определяющий устойчивость трансформированных плазмидой pPINS07 клеток бактерий к ампициллину; уникальные сайты узнавания рестрикционными эндонуклеазами, имеющими следующие координаты: EcoRI - 1, NcoI - 217, BamHI - 221, PstI - 342 и 417, HindIII - 492, SalI - 4513, ClaI - 4792.
Преимуществом предложенной плазмидной конструкции является то, что содержащийся в ней гибридный ген кодирует белок с последовательностью проинсулина человека, которому предшествует последовательность одного IgG-связывающего домена B белка A с гексагистидиновым линкером, что значительно упрощает выделение рекомбинантного белка за счет использования высокотехнологичной аффинной хроматографии на металлохелатных носителях.
Для получения штамма-продуцента гибридного полипептида с проинсулином человека трансформируют компетентные клетки Escherichia coli JM109 рекомбинантной плазмидной ДНК pPINS07.
Полученный штамм Escherichia coli/pPINS07 характеризуется следующими признаками.
Морфологические признаки: клетки мелкие палочковидной формы, грамотрицательные, неспороносные, 1 х 3,5 мкм, подвижные.
Культуральные признаки: при росте на агаризованной среде LB колонии круглые, гладкие, полупрозрачные, блестящие, серые. Край ровный, диаметр колоний 1 - 3 мм, консистенция пастообразная. Рост в жидких средах (LB, минимальная среда с глюкозой) характеризуется ровным помутнением, осадок легко седиментирует.
Физико-биохимические признаки: клетки растут при 4 - 42oC, оптимум pH 6,8-7,6. В качестве источника азота используют как минеральные соли аммония, так и органические соединения: аминокислоты, пептон, триптон, дрожжевой экстракт. В качестве источника углерода при росте на минимальной среде используют глицерин, углеводы, аминокислоты.
Устойчивость к антибиотикам: клетки штамма-продуцента проявляют устойчивость к ампициллину (до 300 мг/мл), обусловленную наличием в плазмиде гена β-лактамазы (bla).
Полученный штамм депонирован в коллекции микроорганизмов ИБХ РАН под номером 013.
На фиг. 1 представлена нуклеотидная последовательность и кодируемая ею аминокислотная последовательность EcoRI/HindIII-фрагмента плазмиды pPINS07; на фиг. 2 - физическая карта плазмиды pPINS07.
Изобретение иллюстрируется следующими примерами.
Пример 1. Конструирование промежуточной рекомбинантной плазмидной ДНК pPINS05.
10 мкг плазмидной ДНК рКК223-3 обрабатывают в 200 мкл буфера, содержащего 10 мМ Трис-HCl, pH 7,5, 50 мМ NaCl, 10 мМ MgCl2 и 10 мМ меркаптоэтанол, гидролизовали 7 ед. рестриктазы BamHI в условиях частичного гидролиза 30 мин при 20oC. Из полученного гидролизата выделяют линейную форму ДНК при помощи электрофореза в 1%-ном геле легкоплавкой агарозы. Полученную ДНК обрабатывают 30 мин при 15oC 10 ед. фрагмента Кленова ДНК-полимеразы I Е. coli в 200 мкл буфера, содержащего 20 мМ Трис-HCl, pH 7,5, 50 мМ NaCl, 10 мМ MgCl2, 10 мМ меркаптоэтанол и 50 мкМ дезоксинуклеозид-5'-трифосфаты. Полученную таким образом ДНК лигируют в 100 мкл буфера для лигирования по тупым концам [13]. 5 мкл лигазной смеси используют для трансформации компетентных клеток Е. coli JM109 [14]. Трансформанты высевают на LB-агар, содержащий 100 мкг/мл ампициллина. Из выросших клонов выделяют плазмидную ДНК и анализируют рестриктным анализом при помощи эндонуклеаз HaeIII и HpaII. Отбирают ДНК рКК223-3*, не содержащую сайта рестриктазы BamHI в положении 256.
Для клонирования IgG-связывающего домена B белка A проводят полимеразную цепную реакцию на матрице 10 нг ДНК из S. aureus (штамм Cowan) в присутствии олигонуклеотидных праймеров CCACCGA-
ATTCTATGGCTGATAACAAAGAACAACAA и
CCACCGGATCCTTTGTTATCAGCTTTTG-
GTGCTTC в 100 мкл буфера для амплификации [15]. Проводят 25 циклов амплификации (95oC, 45 с; 52oC, 45 с; 72oC, 45 с), продукт амплификации гидролизуют рестриктазами EcoRI и BamHI и фрагмент величиной около 200 п.о., кодирующий IgG-связывающий домен B, очищают электрофорезом в 6% ПААГ.
5 мкг плазмидной ДНК рКК223-3* обрабатывают совместно рестриктазами EcoRI и HindIII и из полученного гидролизата выделяют с помощью электрофореза в 1%-ном агарозном геле векторную часть плазмиды (4,5 т.п.о.).
1 мкг полученного вектора лигируют с 0,02 мкг EcoRI/BamHI-фрагмента 200 п. о. , содержащего ген IgG-связывающего домена B, и 0,02 мкг синтетического BamHI/HindIII-фрагмента ДНК величиной 273 п.о., кодирующего проинсулин человека, в 50 мкл буфера для лигирования [13] 10 час при 8oC в присутствии 20 ед. Т4 ДНК-лигазы. 3 мкл лигазной смеси используют для трансформации компетентных клеток Е. coli JM109 [14]. Трансформанты высевают на LB-агар, содержащий 100 мкг/мл ампициллина. Из выросших клонов выделяют плазмидную ДНК pPINS03 и анализируют рестриктным анализом при помощи эндонуклеаз HaeIII и HpaII, а также совместным гидролизом рестриктазами EcoRI+BamHI и BamHI+HindIII. Окончательно строение плазмиды pPINS03 подтверждали определением нуклеотидной последовательности между сайтами рестриктаз EcoRI и BamHI.
Для замены метионинового кодона в гибридном гене на аргининовый проводят полимеразную цепную реакцию на матрице плазмиды pPINS03. Для этого 0,01 мкг ДНК плазмиды амплифицируют в 50 мкл буфера [15], содержащего 25 мМ Трис-HCl, pH 8,8, 10 мМ (NH4)2SО4, 1,2 мМ MgCl2, 2 мМ меркаптоэтанол, 0,1 мМ каждого дезоксинуклеозид-5-трифосфата и 50 пмоль мутагенизирующего (AACAAAGGTACCCGTTTT-
GTTAACCAACACC) и якорного (TCCG-
CCAAAACAGAAGCT) праймеров, в присутствии 2,5 ед. Taq-ДНК-полимеразы. Проводят 20 циклов амплификации (94oC, 30 с; 48oC, 45 с; 72oC, 45 с). Продукт амплификации гидролизуют рестриктазами BamHI и HindIII и лигируют с 50 мкг вектора, полученного расщеплением плазмиды pPINS03 теми же рестриктазами в буфере для лигирования [13]. 10 мкл лигазной смеси используют для трансформации компетентных клеток Е. coli JM109 [14]. Трансформанты высевают на LB-агар, содержащий 100 мкг/мл ампициллина. Из выросших клонов выделяют плазмидную ДНК и анализируют определением нуклеотидной последовательности фрагмента ДНК между сайтами рестриктаз BamHI и HindIII. Полученная таким образом плазмида pPINS05 содержит tac-промотор, вслед за ним EcoRI-сайт, далее между сайтами EcoRI и BamHI структурный ген домена B стафилококкового белка A и структурный ген проинсулина человека между сайтами BamHI и HindIII, соединенный с доменом белка A кодоном аргинина. Клетки Е. coli JM109, трансформированные плазмидой pPINS05, способны синтезировать гибридный полипептид с проинсулином человека при индукции tac-промотора IPTG.
Пример 2. Конструирование рекомбинантной плазмидной ДНК pPINS07.
5 мкг плазмидной ДНК pPINS05 гидролизуют рестриктазой BamHI в соответствии с методикой F, описанной в работе [13]. Полученный векторный фрагмент после очистки электрофорезом в 1%-ном геле легкоплавкой агарозы лигируют с 50-кратным молярным избытком олигонуклеотидного дуплекса: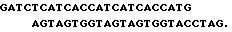
5 мкл лигазной смеси используют для трансформации компетентных клеток Е. coli JM109 [14] . Трансформанты высевают на LB-агар, содержащий 100 мкг/мл ампициллина. Из выросших клонов выделяют плазмидную ДНК и анализируют при помощи гидролиза рестриктазами EcoRI+BamHI и BamHI+HindIII. Из 16 проанализированных клонов 9 показали нужный набор рестриктных фрагментов. Окончательно строение рекомбинантной плазмиды pPINS07 подтверждают определением нуклеотидной последовательности фрагмента ДНК между сайтами рестриктаз BamHI и HindIII.
Пример 3. Определение продуктивности штамма-продуцента гибридного полипептида с проинсулином человека.
В 20 мл жидкой среды LB, содержащей 100 мкг/мл ампициллина, инокулируют индивидуальную колонию клеток Е. coli JM109, содержащих плазмиду pPINS07 и выращивают при 37oC на качалке при 180 об/мин в течение 4 ч до мутности 0,8. Затем добавляют IPTG до концентрации 0,5 мМ и продолжают инкубацию в тех же условиях в течение 6 ч. Отбирают пробу 2 мл и центрифугируют 5 мин при скорости 6000 об/мин, после чего клетки суспендируют в 200 мкл буфера, содержащего 125 мМ Трис-HCl, pH 6,8, 20% глицерин, 3% додецилсульфат натрия, 3% меркаптоэтанол и 0,01% бромфеноловый синий, нагревают 10 мин на кипящей водяной бане. Отбирают образцы 2,5 мкл, 5 мкл, 7,5 мкл, 10 мкл и 15 мкл и анализируют электрофорезом в 13%-ном полиакриламидном геле, содержащем 0,1% додецилсульфат натрия [17]. Гель окрашивают Кумасси R-250 и сканируют на лазерном денситометре Ultrascan XL. По данным сканирования гибридный полипептид составляет 25 - 30% суммарного клеточного белка.
Источники информации:
1. Bell G.I., Swain W.F., Pictet R., Cordell B., Goodman H. M., Rutter W.J. // Nature, 1979, v. 282, p. 525 - 527.
2. Sures I., Goeddel D.V., Gray A., Ulrich A. // Science, 1980, v. 208, p. 57-59.
3. Williams D.C., Van Frank R.M., Murth M.L., Burnett J.P. // Science, 1982, v. 215, p. 687-689.
4. Ovchinnikov Y. A., Efimov V.A., Ivanova I.N., Reverdatto S.V., Skiba N.P., Chakhmakhcheva O.G. // Gene, 1984, v. 31, p. 65-68.
5. Sung W. L. , Yao F.L., Zanab D.M., Narang S.A. // Proc. Natl. Acad. Sci. USA, 1986, v. 83, p. 561-565.
6. Tang J., Xue Y., Fan X., Fu Y. // Clin J. Biotechnol., 1993, v. 9, p. 71 - 78.
7. Berg H., Walter M., Mauch L., Seissler J., Northemann W.J. // Immunol. Methods, 1993, v. 164, p. 221-231.
8. Wei G., Hu M.H., Tang L.G. // Biochem. Mol. Biol. Int., 1995, v. 35, p. 37 - 46.
9. McGregor W.C. // Ann. N.Y. Acad. Sci., 1983, v. 413, p. 231- 237.
10. Cowley D.J., Mackin R.B. // FEBS Lett., 1997, v. 402, p. 124-130.
11. Johansson P., Nilsson L., Samuelsson E., Moks Т., Stahl S., Uhlen M. // Eur. J. Biochem., 1996, v. 236, p. 656-661.
12. Чувпило С.А, Кравченко В.В. // Биоорган. химия, 1983, т. 9, с. 1634 - 1637.
13. Sambrook J., Fritsch E.F., Maniatis T. // Molecular Cloning. A Laboratory Manual. 2nded. Cold Spring Harbor, NY, 1989.
14. Hanahan J. // J. Mol. Biol., 1983, v. 227, p. 557 - 580.
15. Taylor G. R. In: Polymerase Chain Reaction. A practical Approach, v. 1, McPherson M.J., Quirke P., Taylor G.R., eds. Oxford Univ. Press. Oxford. 1994.
16. Brosius J. , Dull T.J., Sleeter D.D., Noller H.F. // J.Mol. Biol., 1981, v. 148, p. 107 -127.
17. Laemmli U.K. // Nature, 1970, v. 227, p. 680-687.

Изобретение относится к биотехнологии, в частности к генетической инженерии. Может быть использовано для получения рекомбинантного проинсулина человека. Предложена рекомбинантная плазмидная ДНК, содержащая искусственный ген, кодирующий гибридный полипептид, состоящий из одного IgG-связывающего домена белка А из Staphylococcus aureus, пептидного линкера His6GlySerArg и проинсулина человека, гибридный tac-промотор и терминатор рибосомного оперона Е. coli. Предложен штамм Escherichia coli - продуцент гибридного полипептида с последовательностью проинсулина человека. Биосинтез гибридного полипептида индуцируется изопропилтиогалактопиранозидом, и уровень его биосинтеза составляет не ниже 25% суммарного клеточного белка. 2 с.п. ф-лы, 2 ил.
| Johansson P | |||
| et al | |||
| Eur.J.Biochem | |||
| Предохранительное устройство для паровых котлов, работающих на нефти | 1922 |
|
SU1996A1 |
| Обжигательная печь | 1936 |
|
SU55945A1 |
| US 4518690 A, 21.05.85. | |||

