Область использования настоящего изобретения представляет собой в основном медицину и ветеринарию. Настоящее изобретение касается новых веществ, оказывающих влияние на рост и пролиферацию клеток, а также использования этих веществ для предотвращения или лечения заболеваний. Кроме того, настоящее изобретение касается метода диагностики заболеваний или скриннинга субъектов, имеющих предрасположенность к заболеванию или склонных к распространению его.
Словарь
Для упрощения приведенного описания необходимо использование ряда новых терминов. Ниже приведены некоторые из них и их значения в контексте настоящего описания:
"Регулятор роста клеток (РР)" - это агент, оказывающий влияние на рост или пролиферацию клеток. Воздействия РР на рост или пролиферацию может состоять или в ускорении роста, или пролиферации клеток, или в подавлении этих процессов.
"Эндогенный регулятор роста клеток (ЭРР)" - это агент, выделяемый или распространяемый клеткой организма и оказывающий влияние на рост или пролиферацию той же клетки или других клеток этого же организма.
"Цитостатический РР/ЭРР - это РР/ЭРР, который наполовину снижает рост или пролиферацию клеток или значительно замедляет эти процессы, причем это происходит без существенного разрушения клеток. Обычно цитостатические ЭРР оказывают тормозящее действие на определенный клеточный цикл одной из циклических фаз клетки.
"Низкомолекулярный эндогенный регулятор роста (НМ-ЭРР)" - это эндогенный регулятор роста, молекулярный вес которого ниже ~ 3.000 дальтон.
"Мышечный фактор (МФ)" - это НМ-ЭРР, выделяемый или распространяемый мышечными клетками.
"Лейкоцитарный фактор (ЛФ)" - НМ-ЭРР, выделяемый или распространяемый лейкоцитами.
"Клетка-источник" - это клетка, которая выделяет или распространяет ЭРР.
"Клетка-мишень" - это клетка, которая испытывает воздействие ЭРР.
"Улучшенная среда (УС)" - это среда, улучшенная ростом в ней клеток-источников. Поэтому эта среда содержит эндогенный фактор роста, выделяемый клетками-источниками.
"Улучшенная среда мышечных клеток" и "улучшенная среда лейкоцитов" - это среда, улучшенная ростом в ней мышечных клеток и лейкоцитов соответственно. Улучшенная среда мышечных клеток и улучшенная среда лейкоцитов содержат соответственно МФ и ЛФ.
Представленные выше термины необходимо истолковывать и понимать со ссылкой на приведенное далее описание.
ПРЕДПОСЫЛКИ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
Среди исследований, включающих раскрытие, характеристику, биологические испытания и клинические исследования эндогенных регуляторов роста клеток, существенную часть составляют исследования ЭРР, связанных с цитокинезом. Все раскрытые до настоящего времени регуляторы этого типа представляют собой напоминающие белок вещества, молекулярный вес которых составляет от нескольких тысяч дальтон до нескольких десятков тысяч дальтон. Хотя у указанных ЭРР - разные клетки-источник и клетки-мишени и разная активность, они выделяют общее действующее вещество, состоящее целиком из напоминающих белок вещества.
Есть сообщения о том, что физические упражнения в значительной степени замедляют рост и развитие опухолевого процесса у лабораторных животных (S.A. Hoffman et al., 1962, Cancer. Res., 22: 597-599; V.E. Baracos, 1989, Сhеm., J. Physiol. Pharmacol. , 67: 864-870). A.Szent-Gyorgyi с соавторами (1963, Science, 140: 1391-1392) сообщают, что экстракты некоторых тканей (включая вилочковую железу, аорту, мышцу и сухожилие) содержат два вещества, одно из которых (названное ими "Ргomine") ускоряет рост асцитной опухоли у мышей, а другое (названное ими "Retine") замедляет такой рост. Последнее описано в этой работе как низкомолекулярное вещество; оно сравнительно нестойко, так как разлагается при комнатной температуре в течение почти одной недели. Более того, на основе способа его выделения выяснили, что это вещество является липофильным. Ингибирование асцитных опухолевых клеток экстрактами мышечной ткани описали и T.Namba с соавторами (1968, British J. of Exp. Pathol. 49: 294-301) и ингибиторную активность обнаружили в мышечном экстракте, который можно было подвергнуть диализу через силиконовую мембрану. Нагревание оказывало влияние на указанную активность этого экстракта, хотя в самом диализате не было обнаружено никакого воздействия этого нагревания.
E. Watta с соавторами (Патент США 4708948) раскрыл высокомолекулярный полипептид, подавляющий рост опухоли; этот полипептид можно получить также из мышечной ткани. M.Djal-detti с соавторами (Патент США 5242692) раскрыл фактор (полученный из мышечных клеток), который подавляет пролиферацию опухолевых клеток. Оказалось, что кажущийся молекулярный вес этого вещества, выделенного из супернатанта мышечной клеточной культуры, составляет (согласно определению электрофорезом в геле) 25.000-30.000 дальтон.
РЕЗЮМЕ ИЗОБРЕТЕНИЯ
Основу настоящего изобретения составляет раскрытие новых веществ, которые выделяются, распространяются или получаются или клетками мышц, или лейкоцитами, и которые биологически активны в подавлении пролиферации опухолевых клеток, при этом в значительной степени они не оказывают влияния на пролиферацию нормальных, неопухолевых клеток. Кроме того, было установлено, что эти вещества эффективно подавляют пролиферацию возбужденных иммунных клеток.
Следовательно, согласно одному из своих аспектов настоящее изобретение обеспечивает в значительной степени очищенный регулятор роста клеток, который представляет собой или:
(a) низкомолекулярный эндогенный фактор роста (НМ-ЭРР), являющийся агентом со следующими характеристиками:
i) его производят, выделяют или распространяют из клеток, в частности клеток мышц или лейкоцитов;
ii молекулярный вес его менее чем ~ 3.000 дальтон;
iii) он не напоминает белок;
iv) он растворим в воде;
v) термически стоек, и
vi) биологически активен в подавлении пролиферации клеток, в частности пролиферации опухолевых клеток или пролиферации возбужденных лимфоцитов; или
(b) представляет собой агент, являющийся производным агента пункта (а) и обладающий биологической активностью в подавлении пролиферации клеток.
Согласно другому своему аспекту настоящее изобретения обеспечивает использование указанного вещества для предотвращения заболеваний или в их терапии. В соответствии с этим аспектом обеспечивается способ предотвращения заболевания или нарушения; этот способ включает введение нуждающемуся субъекту эффективного количества указанного PP. При заданном типе активности РР в подавлении пролиферации его обычно следует вводить субъекту периодически, в течение некоторого времени. Кроме того, в соответствие с этим аспектом обеспечивается состав, содержащий некоторое количество РР. Этот состав может представлять собой фармацевтическую композицию, содержащую терапевтически эффективное количество указанного РР и фармацевтически приемлемый носитель или разбавитель. Эту фармацевтическую композицию можно составить так, чтобы ее можно было использовать для предотвращения некоторого заболевания или нарушения; или составить так, чтобы ее можно было применять в терапии некоторого заболевания или нарушения. Композиция может также представлять собой состав безрецептурного отпуска, например нейтральную композицию, пищевую добавку, гигиенический пищевой препарат и т.д. Наконец, в соответствии с этим аспектом изобретения обеспечивается также применение указанного РР для получения таких составов.
Особенно предпочтительным является использование указанного РР для лечения и предотвращения рака или для подавления (устранения или снижения) активности возбужденных лимфоцитов при лечении или предотвращении многообразных состояний, вызванных гиперактивной иммунной системой (например, лечение, направленное на предотвращение отторжения органов; лечение аутоиммунных заболеваний и т.д.).
В соответствии со следующим аспектом настоящего изобретения обеспечивается метод диагностики рака, или злокачественного состояния субъекта, или метод скриннинга субъектов, склонных к распространению рака или имеющих предрасположенность к нему. Метод включает определение уровня указанного низкомолекулярного эндогенного регулятора роста НМ-ЭРР в жидкости организма указанного субъекта или в супернатанте клеточной культуры, полученной от этого субъекта.
Дополнительный аспект настоящего изобретения представляет собой процесс получения РР (согласно настоящему изобретению) на основе очистки активных фракций соответствующей улучшенной среды.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение основано на раскрытии новых ЭРР, имеющих низкий молекулярный вес. Термин "низкий молекулярный вес" следует понимать как молекулярный вес, который (определенный методом ультрафильтрации) составляет менее ~ 3.000 дальтон, в частности меньше ~2.000 дальтон, а предпочтительно менее ~ 500 дальтон. Специалистам ясно, что эти молекулярные веса - приближенные величины, и их не следует принимать за точные.
Было обнаружено, что низкомолекулярные эндогенные регуляторы роста настоящего изобретения не напоминают белок, т.е. они не являются ни протеинами, ни пептидами, ни каким бы то ни было другим веществом, имеющим протеиновый или пептидный фрагмент, играющий роль в его биологической активности. (Из выводов настоящего изобретения нельзя исключить возможность существования этих НМ-ЭРР в некоторой форме, где они связаны химической или комплексной связью с пептидным или протеиновым фрагментом, не играющим никакой роли в активности НМ-ЭРР как регулятора роста или играющим в ней лишь только ограниченную роль).
В соответствии с настоящим изобретением низкомолекулярные эндогенные регуляторы роста НМ-ЭРР были получены из улучшенной среды клеток мышц и из улучшенной среды лейкоцитов. Однако полагают, что их можно получить, используя и другие источники. Поэтому настоящее изобретение не ограничивается определенным мышечным фактором МФ и определенным лейкоцитарным фактором ЛФ. Напротив, используя сведения, полученные из выводов настоящего изобретения, стандартные навыки и доступные знания, специалист без всяких сложностей найдет другие НМ-ЭРР, попадающие в рамки настоящего изобретения.
Было найдено, что указанные МФ и ЛФ представляют собой опухолево-специфические цитостатические эндогенные регуляторы роста. Им присуща необычная биологическая активность, состоящая в специфическом подавлении ими роста и пролиферации опухолевых клеток; причем они не оказывают ощутимого воздействия на нормальные, неонкогенные клетки. Кроме того, обнаружили, что и мышечный фактор, и лейкоцитарный фактор являются неспецифически опухолевыми факторами (т. е. они эффективны в подавлении роста и пролиферации разнообразных опухолевых клеток) и не являются видоспецифическими (т.е. обладают активностью в подавлении роста и пролиферации опухолевых клеток животных разных видов).
Другими словами, указанные МФ и ЛФ обладают широким спектром активности в подавлении роста и пролиферации раковых клеток. Более того, данные, связанные с настоящим изобретением, означают также, что МФ или ЛФ, полученный от одного вида животного (в частности, млекопитающего), может быть использован для лечения рака у животного другого вида (в частности, млекопитающего).
Необходимо отметить, что хотя, как было обнаружено, МФ и ЛФ представляют собой цитостатические агенты, они, вероятно, могут оказывать некоторое деструктивное влияние на клетки, особенно после продолжительного воздействия. Например, после продолжительного воздействия как МФ или ЛФ, так и их производных опухолевые клетки-мишени могут в конечном счете погибнуть (например, в результате апоптоза).
Было найдено, что кроме активности МФ и ЛФ в подавлении роста и пролиферации опухолевых клеток они активны также в подавлении пролиферации лимфоцитов. Это доказывается ингибированием реакции лимфоцита на митоген или ингибированием смешанной реакции лимфоцитов, что означает, что эти регуляторы роста могут обладать иммуносупрессионной активностью.
Очевидно, что если выделить низкомолекулярный эндогенный регулятор роста из одного вида животных, то можно найти гомологичный НМ-ЭРР в других видах. Например, до сих пор, в соответствии с настоящим изобретением, МФ получали от крысы или человека. Без сомнения, можно также получить гомологичные МФ от других видов животных, в частности от млекопитающих. Аналогично, ЛФ согласно настоящему изобретению получали от человека. Не существует никакого сомнения, что можно также получить гомологичные ЛФ от других видов животных, в частности от млекопитающих. Настоящее изобретение включает и такие гомологи.
Регулятор роста по настоящему изобретению может представлять собой единичную молекулу; или группу молекул, которые действуют вместе аддитивно или сивергически, оказывая влияние на рост и пролиферацию клеток; или же молекулярный комплекс, обладающий такой же активностью.
Если выделить НМ-ЭРР, то можно (например, химической модификацией) получить его производные, которые будут обладать биологической активностью, аналогичной активности исходного НМ-ЭРР. Производные с биологической активностью, аналогичной активности какого-либо НМ-ЭРР или его гомологов, можно идентифицировать, например, используя те же биологические тесты, что и для характеристики указанного НМ-ЭРР. Например, для случая МФ и ЛФ, активных в подавлении роста и пролиферации опухолевых клеток, их производные или гомологи можно найти, испытывая активность подавления роста или пролиферации опухолевых клеток, выросших in vitro, или испытывая способность подавлять смешанную реакцию лейкоцитов. В каждом конкретном случае специалист, без сомнения, должен быть способен выбрать соответствующий биологический анализ.
Это производное может представлять собой молекулу, структура которой аналогична молекулярной структуре указанного НМ-ЭРР, но в ней одна или более химических групп замещены другой группой, продуктом восстановления или окисления указанного низкомолекулярного эндогенного регулятора роста и т.п.
РР настоящего изобретения можно использовать для многообразных терапевтических целей, причем эти регуляторы следует вводить субъекту, нуждающемуся в них, в терапевтически эффективном количестве. Одним из предпочтительных терапевтических показаний использования указанного регулятора роста является лечение или предотвращение рака. В лечебных целях РР может быть введен субъектам, имеющим в анамнезе рак, например, для того, чтобы подавить его рецидив. Такого рода лечение обычно представляет собой контроль за начальной стадией лечения, направленного на удаление или уничтожение опухоли (например, химиотерапии, радиационной терапии и хирургии). Для предотвращения заболевания указанный регулятор роста может быть введен нераковым субъектам или субъектам до предъявления им диагноза какого-либо ракового состояния, в частности субъектам из группы риска, имеющим предрасположенность к заболеванию рака или склонных к распространению его. Субъектами с высокой степенью риска могут быть лица, имеющие генетическую предрасположенность к распространению рака, например субъекты, у которых диагностировано наличие одного из генов, которые, как известно, сочетаются с раком; лица, имеющие рак в семейном анамнезе; лица с высоким риском распространения рака как результата действия разнообразных факторов окружающей среды, таких как облучение, воздействие канцерогенов и т.д. и т.д.
При заданной биологической активности регулятора роста (которая скорее представляет собой подавление пролиферации клеток-мишеней, чем их немедленное разрушение) этот РР обычно следует вводить периодически в течение некоторого времени. Однако, как уже отмечалось выше, возможно, что вслед за продолжительной задержкой роста опухолевых клеток, они в конечном счете погибнут, и поэтому спустя определенное время можно прекратить лечение.
Другое предпочтительное терапевтическое показание указанного регулятора роста представляет собой подавление активности компонентов иммунной системы. Примерами этого является лечение аутоиммунных заболеваний; использование в рамках трансплантационной терапии, например лечение после трансплантации, направленное на предотвращение отторжения органа или ткани и т.д.
Регуляторы роста согласно настоящему изобретению испытывали на разнообразных моделях, включая модели первичного рака у животных, в которых рак индуцируется подкожной, внутримышечной или внутрибрюшинной инокуляцией раковых клеток; и модели метастазов у животных, в которых рак индуцируется посредством внутривенной инокуляцией опухолевых клеток. Было обнаружено, что указанный РР эффективно подавляет распространение рака в обеих моделях. Было обнаружено, что регуляторы роста эффективны для подавления распространения рака как при оральном, так и при парентеральном их введении. Как известно, оральное введение физиологически более удовлетворительно, чем парентеральное. Поэтому вариант орального введения предпочтителен при лечении или предотвращении рака, особенно если схемы лечения или предотвращения включают периодическое введение РР в течение длительного времени.
РР может быть включен в состав фармацевтической композиции, которая будет содержать эффективное количество этого регулятора роста, и физиологически приемлемый носитель, совместимый с указанным РР. Оказалось, что указанный регулятор роста растворим в воде, и, следовательно, физиологически приемлемым носителем для парентерального введения может быть физиологический раствор, а пригодная в пищу водная жидкость - для орального введения. Кроме того, для орального введения указанный РР может быть включен в состав разнообразных дозировочных форм, таких как капсулы, таблетки и т.п. Далее, для того, чтобы вводить РР вместе с носителем или разбавителем, перед применением этот регулятор роста можно предварительно подвергнуть лиофилизации.
Используемый здесь термин "эффективное количество" следует понимать как некоторое количество, эффективное для достижения желаемого эффекта. Например, при терапевтическом лечении рака эффективным количеством является такое количество указанного РР в данной схеме терапевтического лечения, которое достаточно для подавления роста и пролиферации опухолевых клеток, что доказывается, например, уменьшением скорости появления метастазов или сокращением их числа, или уменьшением смертности, вызванной распространением рака. Для предотвращения рака эффективным количеством является такое количество указанного РР при данной схеме превентивного лечения, которое достаточно для задержки появления первичного опухолевого образования.
Согласно настоящему изобретению помимо включения регуляторов роста в фармацевтические композиции, их можно также вводить в композиции других типов, например в пищевые композиции, нейтральные композиции, оздоровительные продукты безрецептурного отпуска и т.д.
Эксперименты, проведенные в связи с настоящим изобретением, показали, что лейкоциты раковых больных выделяют намного меньше низкомолекулярного эндогенного регулятора роста, чем лейкоциты нормальных, здоровых субъектов. Поэтому, определив уровень НМ-ЭРР в жидкости организма (например, в сыворотке, моче и т.д.) или в супернатанте культуры клеток, (например, клеток мышц или лейкоцитов) некоторого субъекта, можно будет как диагностировать рак у него, так и получить данные о злокачественном состоянии. Помимо этого, определение уровня НМ-ЭРР в жидкости организма или в указанном супернатанте может служить основой для скриннинга субъектов, склонных к распространению рака или имеющих расположенность к этому. Уровень НМ-ЭРР можно определить биологическим анализом, который включает определение активности жидкости организма или супернатанта или активности их соответствующих фракций в подавлении пролиферации опухолевых клеток. Кроме этого анализа, присутствие НМ-ЭРР можно также определить аналитическими методами, которые обычно сами по себе известны специалистам. Эти методы включают разнообразные иммунологические пробы на основе использования специфических антител низкомолекулярного эндогенного регулятора роста НМ-ЭРР; анализы на основе использования соответствующих химических реагентов (например, красящих реагентов); спектроскопических анализов или анализов на основе поглощения излучения (например, поглощении света); разнообразных хроматографических методик и т. д.
Согласно другому аспекту настоящего изобретения обеспечивается процесс очистки РР, полученного согласно настоящему изобретению из биологического источника. В соответствии с этим аспектом указанный процесс включает:
(a) рост клеток в таких условиях, при которых эти клетки производят, выделяют или распространяют регуляторы роста клеток в окружающую их среду;
(b) сбор супернатанта от культуры клетки;
(c) отделение фракции супернатанта, содержащей вещества с молекулярным весом выше ~ 3.000 дальтон от фракций супернатанта, содержащих вещества с молекулярным весом ниже ~ 3.000 дальтон и отбор последних.
Фракции, отобранные на стадии (с), можно потом очистить, используя различные методики очистки, в частности хроматографии, например жидкостной хроматографией под высоким давлением (ВЭЖХ).
Далее настоящее изобретение будет проиллюстрировано некоторыми примерами, проводимыми в связи с настоящим изобретением, со ссылкой на чертежи, приведенные в приложении. В этих экспериментах была продемонстрирована активность МФ и ЛФ in vitro и in vivo. Описаны также методы очистки и методики характеристики указанного МФ. Без сомнения, специалист оценит, что эти иллюстрации не следует рассматривать ограничивающими рамки настоящее изобретение, а скорее как примеры всего объема настоящего изобретения, который определен в прилагаемых пунктах патентной формулы. Специалист, без сомнения, сможет на основе приведенного выше и на основе описания воспроизвести настоящее изобретение в полном заявляемом объеме.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На приведенных чертежах:
На фиг. 1-9 показано действие МФ и ЛФ, содержащихся в фильтрате мышечных клеток улучшенной среды УС или в фильтрате лейкоцитов УС, отфильтрованных через мембрану с молекулярным отсечением, рассчитанным на 3.000 дальтон (такой фильтрат будем называть здесь "ультрафильтратом мышечного фактора МФ в 3.000 дальтон" и " ультрафильтратом лейкоцитарного фактора ЛФ в 3.000 дальтон" соответственно). На этих чертежах показаны клетки, выращенные в чашке Петри с 96 микроячейками и инкубированные с ультрафильтратом мышечного фактора МФ в 3.000 дальтон при разных разведениях ("1" - нулевое разведение, "2" - двукратное разведение и т.д.). Во всех этих экспериментах в качестве контрольной использовали неулучшенную среду. На фиг. 1-4 и 6, 8 и 9 и частично на фиг. 7 перфорацию клетки измеряли по включению 3H-тимидина, и ордината указывала величину радиоактивности. На фиг. 5 и частично на фиг. 7 пролиферацию измеряли подсчетом клеток, и ордината указывала число этих клеток.
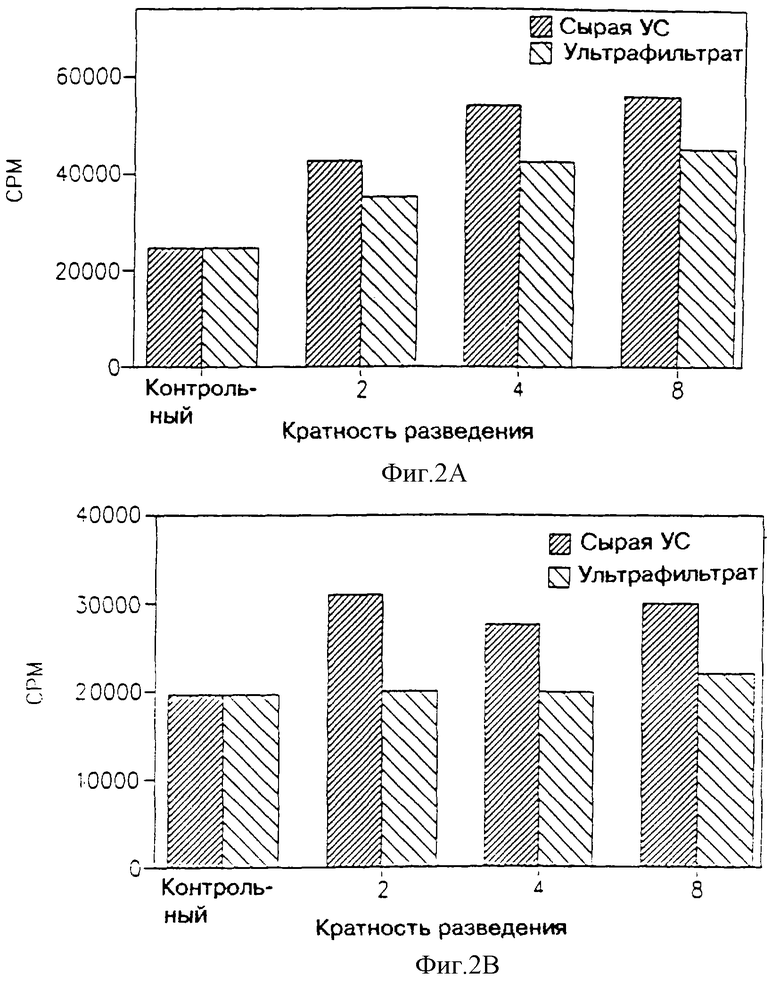
На фиг. 1 показано влияние ультрафильтрата МФ в 2.000 дальтон, полученного от первичной культуры клеток поперечно-полосатых мышц новорожденных крыс, на пролиферацию двух опухолевых клеточных линий: В16, которая представляет собой клеточную линию меланомы грызунов, включая мышей и крыс (фиг. 1А); и НТВ-38, представляющей собой клеточную линию аденокарциномы человека (фиг. 1В). На этих чертежах активность ультрафильтрата МФ в 3.000 дальтон сравнивают с активностью сырой улучшенной среды ("сырая УС").
На фиг. 2 показано влияние ультрафильтрата МФ в 3.000 дальтон, являющегося производным первичной культуры клеток поперечно-полосатых мышц новорожденных крыс, на два типа нормальных, неопухолевых клеток: фибробдаста крыс (фиг. 2А) и клеток костного мозга грызунов (включая мышей и крыс) (фиг. 2В).
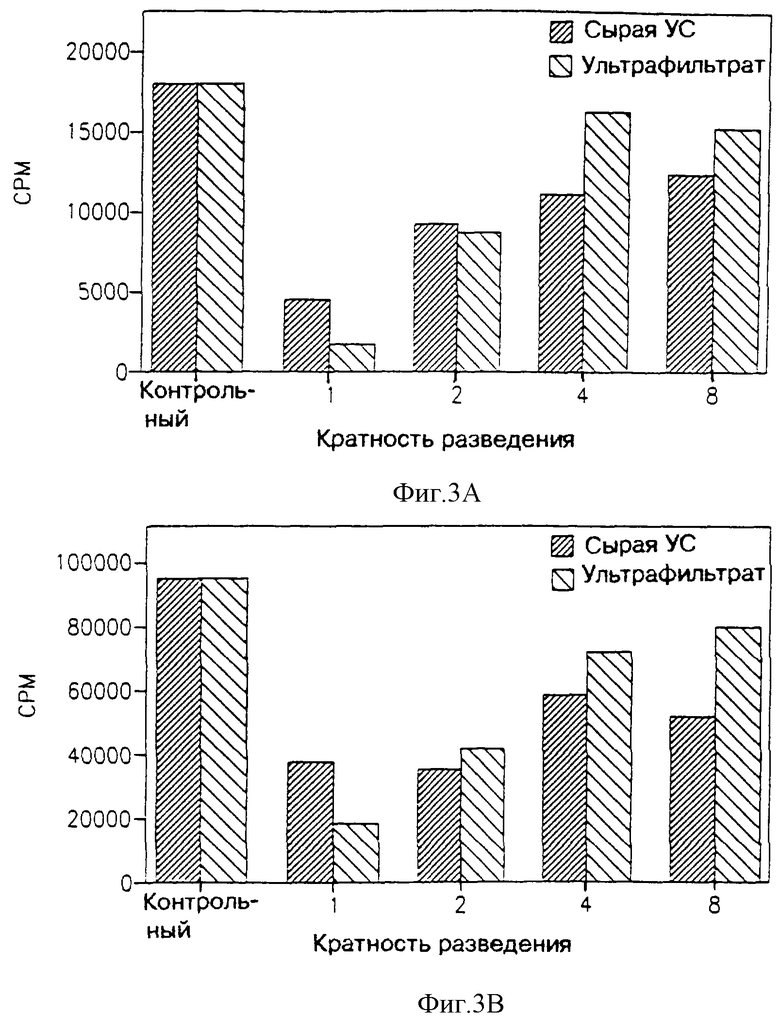
На фиг. 3 показано влияние ультрафильтрата МФ в 3.000 дальтон, являющегося производным линии клеток поперечно-полосатых мышц крыс L-8, на пролиферацию двух типов опухолевых клеточных линий: В16 (фиг. 3А) и линии МСА-105, индуцированной метилхолантреном саркомы легкого грызунов (включая мышей и крыс) (фиг. 3В).
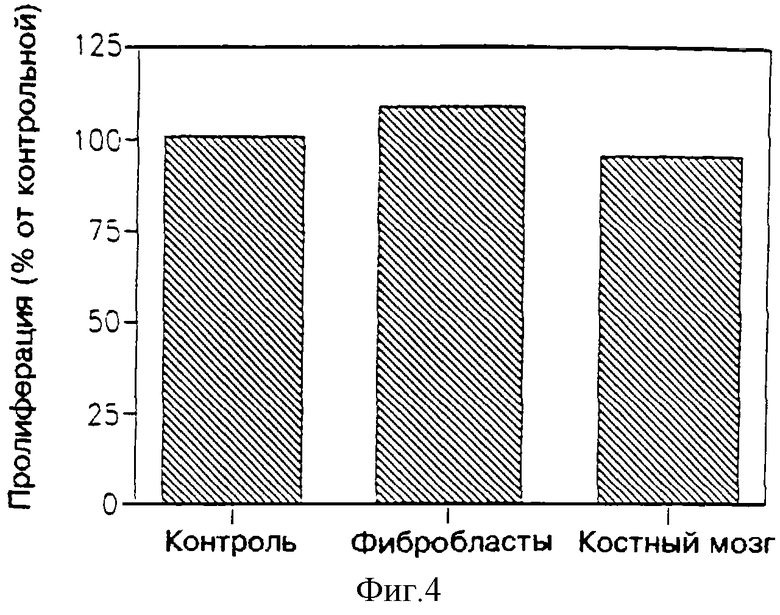
На фиг. 4 показано влияние ультрафильтрата МФ в 3.000 дальтон, являющегося производным клеток L-8, на два типа нормальных клеток: клеток костного мозга грызунов (включая мышей и крыс) и первичной культуры фибробласта крыс.
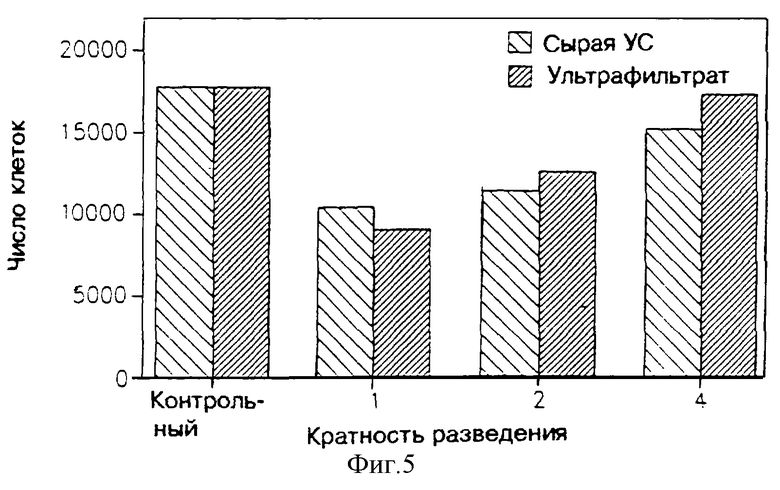
На фиг. 5 показано влияние ультрафильтрата МФ в 3.000 дальтон, являющегося производным линии клеток поперечно-полосатых мышц крыс L-8 на пролиферацию клеток клеточной линии Nba2-11C лимфомы крыс, при испытаниях по росту клеток на основе их подсчета. В этих испытаниях указанные клетки были синхронизированы в фазе G0/G1, потом добавили ультрафильтрат МФ в 3.000 дальтон сырой улучшенной среды УС или контрольную среду, а затем рост клеток стимулировали добавкой гормона роста человека.
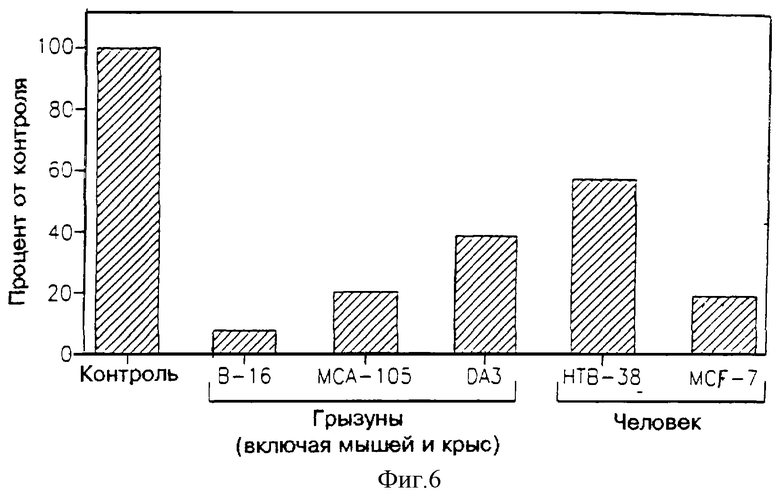
На фиг. 6 показано влияние ультрафильтрата МФ в 3.000 дальтон, являющегося производным клеток L-8, на пролиферацию нескольких опухолевых клеточных линий. Результаты в каждом случае представлены в процентах от контрольной пролиферации в соответствующем эксперименте.
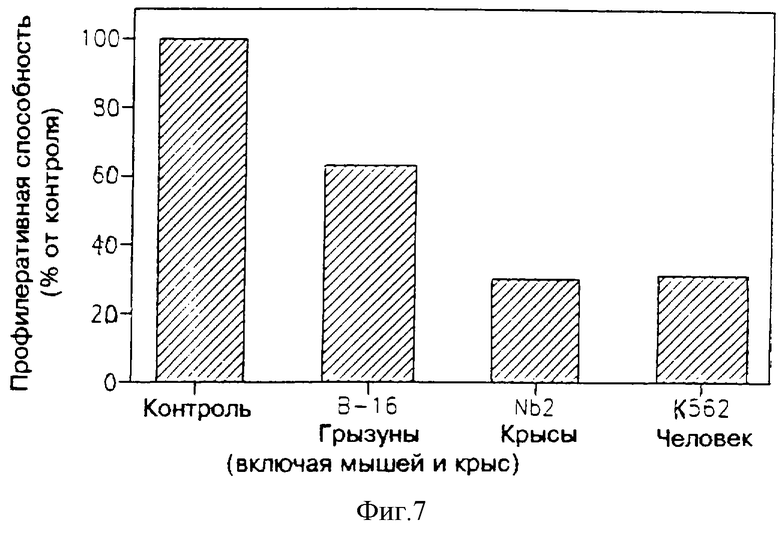
На фиг. 7 показано влияние ультрафильтрата МФ в 3.000 дальтон, являющегося производным миобласта человека. Пролиферацию клеток В-16 и К562 определяли по внедрению 3Н-тимидина; пролиферацию клеток NBT определяли подсчетом клеток.
На фиг. 8 показано влияние ультрафильтрата МФ в 3.000 дальтон, являющегося производным лейкоцитов человека, на пролиферацию опухолевых клеток грызунов (включая мышей и крыс) и человека. Указанная пролиферация дана в процентах от контрольной.
На фиг. 9 показано влияние ультрафильтрата МФ в 3.000 дальтон, являющегося производным клеток L-8, на чувствительность лейкоцитов к фитогемагглюгинину (ФГГ) и на чувствительность в смешанной реакции лимфоцитов.
Фиг. 10 представляет собой фотографию выделенной брюшины двух репрезентативных мышей, у которых внутрибрюшинной инъекцией 2х105 клеток МСА-105 индуцировали рак. После этой инъекции мышь, изображенная справа, дважды в день получала внутрибрюшинную инъекцию 0.5 мл МФ, который содержал фракцию, элюированную из препаративной колонки ВЭЖХ с обращенной фазой С18 (эту фракцию ниже в тексте называют "MF-SP" (см. 7.6.2)); а мышь, изображенная слева, получала инъекцию контрольной среды RPMI.
Фиг. 11 представляет собой фотографию выделенной брюшины двух репрезентативных мышей, у которых внутрибрюшинной инъекцией 5х105 клеток меланомы В-16 индуцировали рак. На фото справа представлена мышь, которая один раз в день получала инъекцию 1 мл ультрафильтрата МФ в 3.000 дальтон (в фосфатно-содевом буферном растворе), являющегося производным клеток L-8; на фото слева представлена мышь, которая один раз в день получала внутрибрюшинную инъекцию контрольной среды, являющейся раствором фосфатно-солевого буфера.
Фиг. 12 представляет собой фотографию выделенных брюшин репрезентативных мышей, которым внутрибрюшинной инъекцией вводили 2.5х105 клеток МСА-105. На фиг. 12А представлены мыши из группы, представители которой ежедневно (начиная со дня инокуляции опухоли) перорально получали по 1 мл ультрафильтрата МФ в 3.000 дальтон, являющегося производным клеток L-8. На фиг. 12В представлены мыши из контрольной группы, которые получали перорально контрольную среду RPMI.
Фиг. 13 представляет собой фотографию, на которой показаны изолированные легкие мышей, которым внутривенно вводили 5х105 клеток меланомы В-16. В верхнем ряду показаны легкие животных из контрольной группы, эти животные ежедневно получали перорально 1 мл контрольного раствора фосфатно-солевого буфера. В нижнем ряду показаны легкие животных экспериментальной группы, включавшей животных, которым давали ежедневно перорально 1 мл ультрафильтрата МФ в 3.000 дальтон, полученного из клеток L-8.
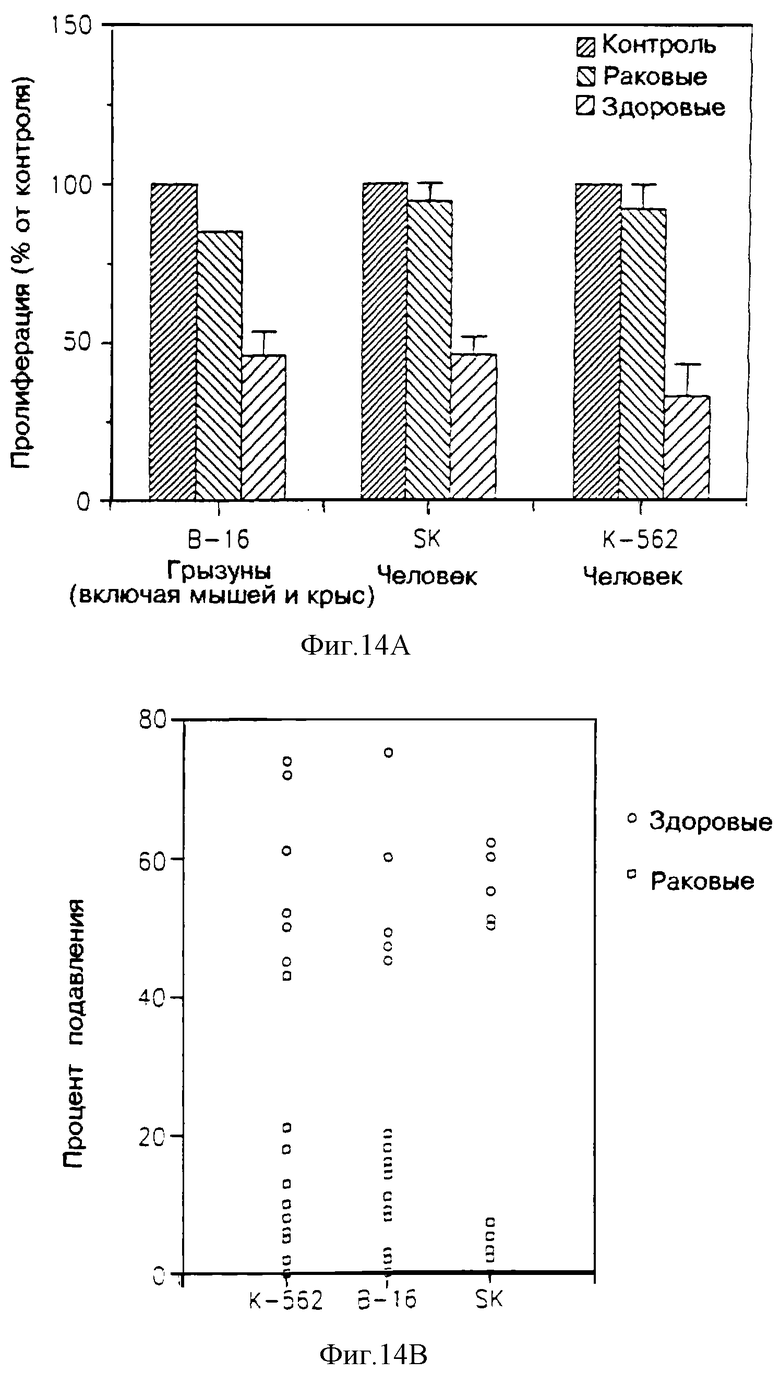
На фиг. 14 показано влияние ультрафильтрата МФ в 3.000 дальтон из супернатанта лейкоцитов крови раковых пациентов и лейкоцитов крови здоровых субъектов, на подавление пролиферации трех опухолевых клеточных линий (В-16, SK и К-562).
На фиг. 14А показана пролиферация этих трех линий клеток относительно контрольной после воздействии ультрафильтратом МФ в 3.000 дальтон; и
На фиг. 14В приведен разброс результатов, указанных как процент подавления, причем подавление эквивалентно пролиферации; 100% пролиферация составляет 0% подавления и т.д. (более чем 100% пролиферации также приписали значение "0"%).
На фиг. 15 приведен 220 нм профиль вымывания раствора, содержащего МФ, после того, как его подвергли повторной хроматографии на колонке ВЭЖХ с обращенной фазой (эту повторную хроматографию проводили также и на той же колонке, что и первую хроматографию).
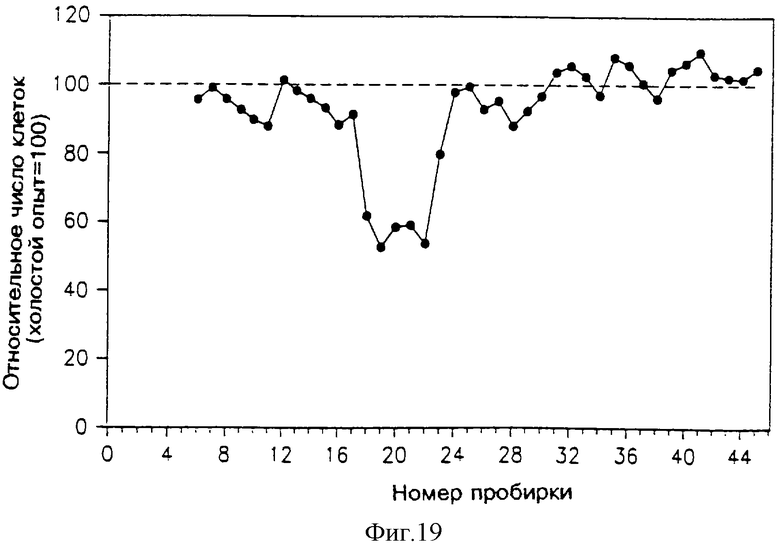
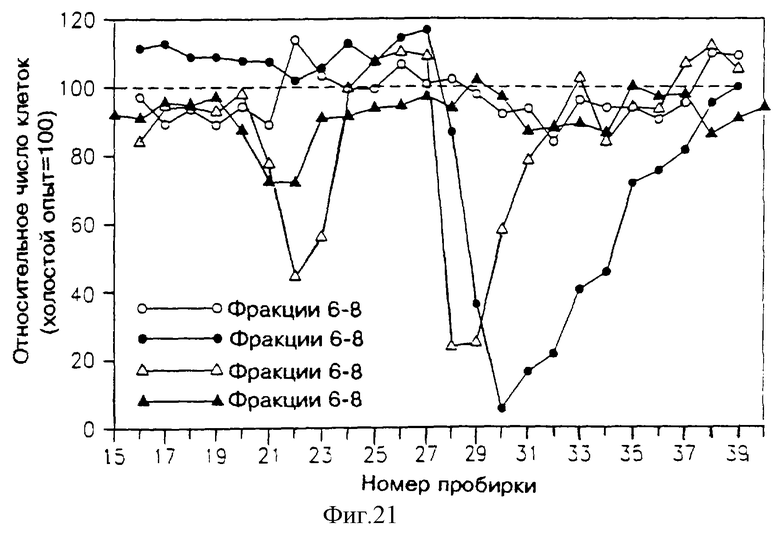
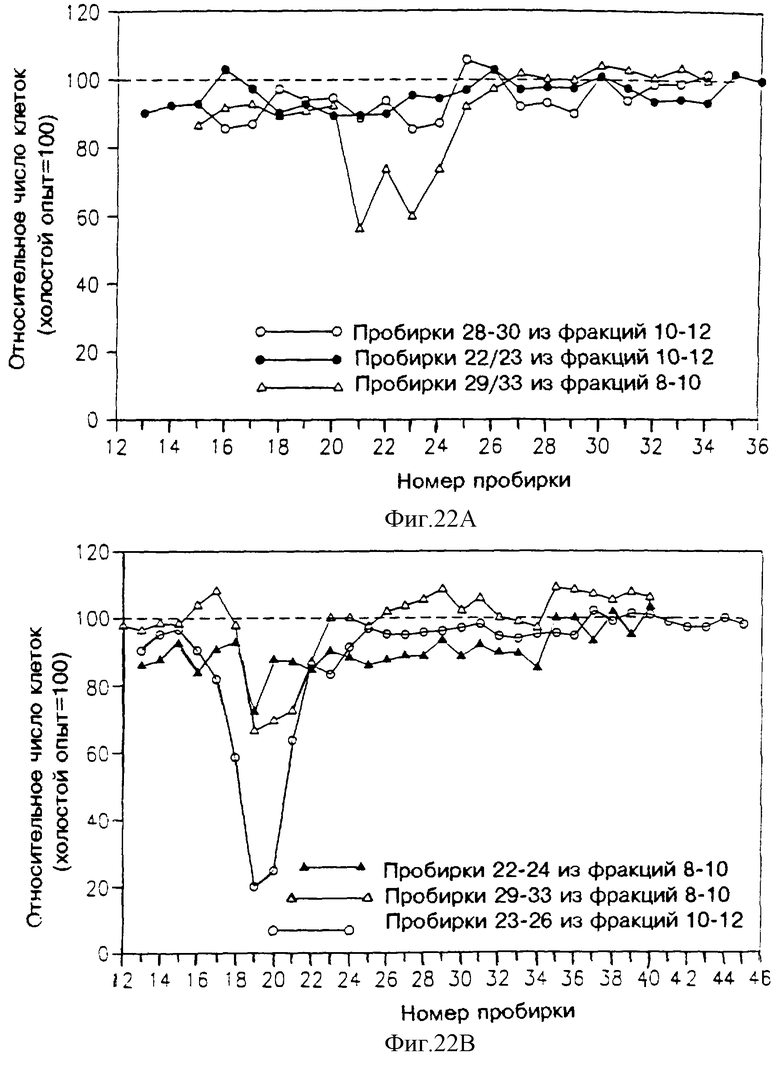
На фиг. 16-21 указана активность различных фракций, элюированных из колонок ВЭЖХ, в подавлении пролиферации клеток Nb2, определяемой путем их подсчета. На каждом из этих чертежей абсцисса указывает номер пробирки (скорость потока и объем каждой пробирки указаны ниже в тексте). Ордината показывает относительное число клеток в сравнении с холостым опытом (клетки, выросшие без добавления какого-либо раствора к питательной среде клеток Nb2, составляющей 100%).
На фиг. 16 и 17 приведена активность различных фракций, элюированных в двух разных сериях опытов из препаративной колонки ВЭЖХ с обращенной фазой (С-18), в подавлении пролиферации клеток Nb2. Растворы, которые подавали в колонки, представляли собой или ультрафильтрат МФ в 3.000 дальтон, полученный из клеток L-8 (в фосфатно-солевом буферном растворе) или контрольный фосфатно-солевой раствор. На фиг. 16 раствор, содержащий МФ, изображен незакрашенными кружочками; контрольный раствор изображен закрашенными кружочками. На фиг. 17 раствор, содержащий МФ, изображен закрашенными треугольниками; контрольный раствор изображен закрашенными кружочками.
На фиг. 18 приведена активность различных фракций, элюированных из аналитической колонки ВЭЖХ с обращенной фазой (С-18), в подавлении пролиферации клеток Nb2. Раствор, который подавали в эту колонку, содержал объединенные фракции 5 и 6, указанные на фиг. 16. Раствор, содержащий МФ, изображен закрашенными кружочками, контрольный раствор - незакрашенными кружочками.
На фиг. 19 приведена активность различных фракций, элюированных из колонки superdex, в подавлении пролиферации клеток Nb2. Раствор, который подавали в эту колонку, представлял собой объединенный раствор из элюата с фиг. 17; этот раствор содержал 250 мл из пробирок 9-10, 520 мл из пробирок 10-11 и 600 мл фракций 11-12.
На фиг. 20 приведена активность различных фракций, элюированных из аналитической колонки ВЭЖХ с обращенной фазой (С-18), в подавлении пролиферации клеток Nb2. Раствор, подаваемый в эту колонку, представлял собой объединенный раствор из пробирок 6-12 с фиг. 17. Раствор, содержащий МФ, изображен закрашенными квадратами, контрольный фосфатно-солевой раствор изображен незакрашенными кружочками.
На фиг. 21 приведена активность различных фракций, элюированных из колонки "эксклюзивного размера" (SE), в подавлении пролиферации клеток Nb2. Растворы, подаваемые в эту колонку, представляли собой различные фракции, элюированные из препаративной колонки ВЭЖХ с обращенной фазой и показанные на фиг. 17: фракция "А", пробирки 6-8 - незакрашенные кружочки; фракция "В", пробирки 8-10 - закрашенные кружочки; фракция "С", пробирки 10-12 - незакрашенные треугольники; фракция "D", пробирки 12-14 - закрашенные треугольники.
На фиг. 22 показана активность различных фракций, элюированных из колонки superdex, в подавлении пролиферации клеток Nb2. Растворы, подаваемые в эту колонку, содержали различные фракции элюата с фиг. 21, а именно:
На фиг. 22А представлены результаты, полученные для следующих подаваемых растворов: пробирки 28-30 фракции С - незакрашенные кружочки; пробирки 22-23 фракции С - закрашенные кружочки; пробирки 29-33 фракции В - незакрашенные треугольники; и
На фиг. 22В представлены результаты, полученные для следующих подаваемых растворов: пробирки 31-32 фракции С - закрашенные кружочки; пробирки 23-26 фракции С - незакрашенные кружочки; пробирки 29-33 фракции В - незакрашенные треугольники.
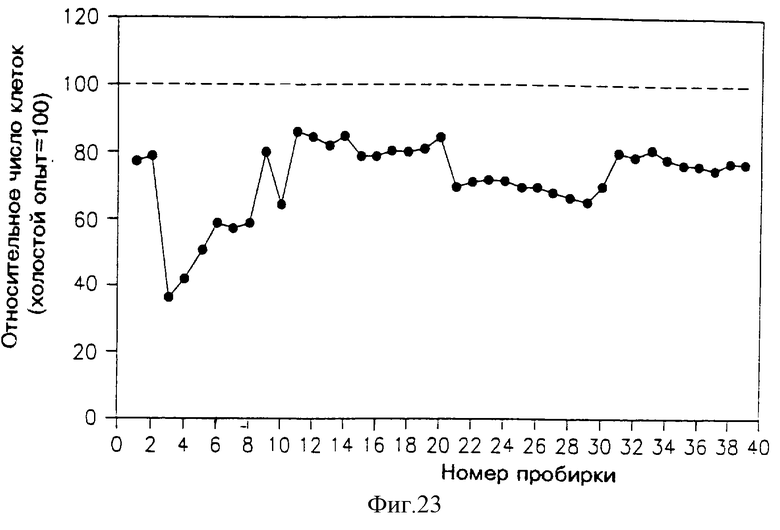
На фиг. 23 приведен профиль активности фракций, элюированных из аналитической колонки ВЭЖХ с обращенной фазой (С-18), в подавлении пролиферации клеток Nb2. Раствор, подаваемый в эту колонку, содержал активные фракции (элюированные из препаративной колонки ВЭЖХ с обращенной фазой при 18-28% содержании ацетонитрила).
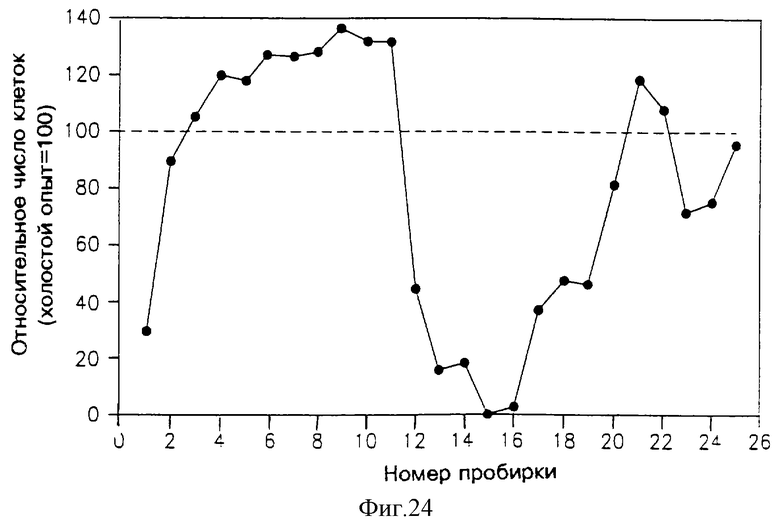
На фиг. 24 приведен профиль активности фракций, элюированных из колонки SE, в подавлении пролиферации клеток Nb2. Растворы, подаваемые в указанную колонку, представляли собой фракции 13-17 из аликвоты с фиг.23.
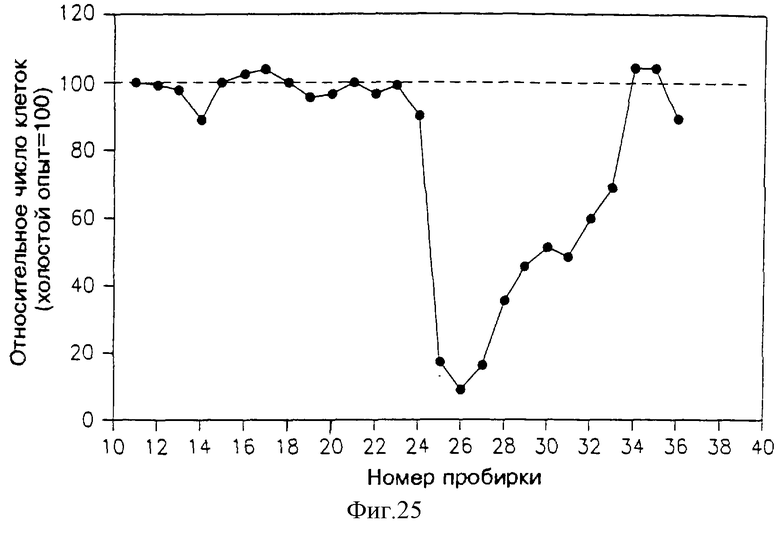
На фиг. 25 приведен профиль активности различных фракций, элюированных из колонки "гидрофильного взаимодействия" (СНО), в подавлении пролиферации клеток Nb2. Растворы, подаваемые в указанную колонку, представляли собой объединенные фракции 13-17 из аликвоты с фиг.24.
Фиг. 26 представляет собой ЯМР-сканирование активных фракций, полученных из колонки SE (фракции, элюированные в интервале 27-32 мин).
ДЕТАЛЬНОЕ ОПИСАНИЕ НАСТОЯЩЕГО ИЗОБРЕТЕНИЯ
В дальнейшем при обсуждении активности активных фракций, полученных из улучшенной среды УС мышечных клеток и улучшенной среды УС клеток лейкоцитов, иногда будут делать ссылку на мышечный фактор "МФ" или на лейкоцитарный фактор "ЛФ". Необходимо понять, что обе эти УС могут содержать более чем один фактор с описанной ниже активностью. Действительно, результаты фракционирования ВЭЖХ (некоторые из которых показаны ниже), полученные в связи с настоящим изобретением, так можно объяснить (хотя возможны и другие объяснения). Так, например, УС мышечной клетки может содержать более одного фактора, подавляющего развитие опухолевого процесса. Приводимую иногда ссылку на "определенный МФ" и ей подобные не следует рассматривать как обозначение того, что улучшенная мышечная среда УС содержит только один НМ-ЭРР, так как эта УС мышечной клетки в действительности может содержать несколько веществ, и все их можно назвать "МФ".
1. Улучшенная среда
1.1. МФ
МФ получали из улучшенной среды (УС) трех типов препаратов мышечных клеток:
1.1.1. Первичные культуры мышц новорожденных крыс
Мышцы задних лап новорожденных крыс в возрасте 24-48 часов были отделены и мелко измельчены. После трипсинизации 0.25% раствором трипсинверсана клетки сначала высевали в чашку Петри с культурой ткани для удаления фибробластов и моноцитов. Количество клеток подсчитали, а потом высеяли в обогащенную модифицированную среду (ОМС). Спустя 5 дней эти культуры содержали сократительные мышечные клетки. Затем указанную среду удалили и добавили среду RPMI или среду фосфатно-солевого буфера. Клетки инкубировали в среде RPMI в течение 24 часа и 8 или 24 часа в среде фосфатно-солевого буфера. После этого супернатант собрали, отцентрифугировали и сохраняли в холодильнике при -20 oС до дальнейшей переработки.
1.1.2. Линия мышечных клеток крысы (L-8)
Линия L-8 (которая в American Type Culture Collection ATCC обозначена CRL 1769) представляет собой линию скелетных мышечных миобластов новорожденных крыс, которая содержит недифференцированные миобласты, пролиферирующих без добавления факторов роста. Клетки L-8 высевали в чашки, и потом они росли в среде RPMI (эту среду здесь и далее будут называть "RPMI"), содержащей 4% глюкозы. Спустя 3 дня после деления клеток супернатант культуры отбросили и его заменили или RPMI или раствором фосфатно-солевого буфера; потом в течение последующих 24 часов проводили инкубирование. Затем супернатанты собрали, и если их немедленная обработка не была необходима, то супенатанты сохраняли в холодильнике при -20oС.
1.1.3. Миобласты человека
Миобласты человека, полученные из биопсии человека, культивировали во вращающихся сосудах до сгущения (см. патент США 5130441). Питательную среду удалили и заменили ее или DMEM или раствором фосфатно-солевого буфера, а клетки инкубировали в этих растворах 24 часа. Супернатант собрали и клетки отделили от него центрифугированием.
1.2. ЛФ
ЛФ получали из улучшенной среды одноядерных клеток. Одноядерные клетки выделяли из венозной крови описанным ниже способом. У донора-человека провели забор 20 мл венозной крови с помощью специального шприца. Гепаринизированную таким образом кровь разбавили фосфатно-солевым буфером в соотношении 1:1, поместили в 15 мл Ficoll-Hypaque (Pharmacia) или Histopaque (Sigma, St. Louis, USA) и центрифугировали 30 минут при 400xg. Внутренний слой, содержащий одноядерные клетки, собрали и промывали трижды фосфатно-солевым буфером.
Одноядерные клетки (2x106 клеток/мл) суспендировали в фосфатно-солевом буфере и инкубировали при 37oС 48 часов во влажной атмосфере, содержащей 5% СО2 и 95% воздуха. Полученную при этом суспензию клеток отцентрифугировали, а супернатант собрали.
2. Ультрафильтрация
Улучшенную среду УС, полученную из приведенных выше источников, подвергли ультрафильтрации, используя фильтры с молекулярным отсечением на 500, 2.000, 3.000 и 10.000 дальтон (Centricon, Amicon, USA).
Было найдено присутствие МФ в ультрафильтрате мембран с молекулярным отсечением на 10.000, 3.000, 2.000 дальтон, так же, как и в ультрафильтрате мембраны с молекулярным отсечением на 500 дальтон (что будет показано далее). ЛФ подвергли ультрафильтрации через мембрану с молекулярным отсечением на 3.000 дальтон и в полученном ультрафильтрате обнаружили ЛФ.
Ультрафильтрат УС мышечных клеток, прошедший через фильтр с молекулярным отсечением на 500, 2.000 дальтон и т.д. здесь будут называть " Ультрафильтрат МФ в 500 дальтон", "Ультрафильтрат МФ в 2.000 дальтон" и т.д.; ультрафильтрат УС лимфоцитов, прошедший через фильтры с молекулярным отсечением на 3.000 дальтон, здесь будут называть "Ультрафильтрат ЛФ в 3.000 дальтон".
3. Подавление in vitro пролиферации опухолевых клеток с помощью МФ и ЛФ
3.1. Методы
3.1.1. Клеточные линии
Влияние МФ и ЛФ на анализ in vitro было изучено на нескольких опухолевых клеточных линиях и нескольких неопухолевых клетках. Испытывались следующие клетки:
(а) опухолевые клеточные линии:
Н-29, которая представляет собой клеточную линию аденокарциномы, полученную из толстой кишки человека (АТСС, обозначена НТВ-38);
МСА-105, которая представляет собой клеточную линию индуцированной метилхлорантеном саркомы легкого грызунов (включая мышей и крыс);
B16-F1, которая представляет собой клеточную линию меланомы грызунов (включая мышей и крыс);
SK-28, которая представляет собой клеточную линию меланомы человека;
К-562, которая представляет собой клеточную линию лейкемии человека;
Клетки DAS, которые являются клеточной линией рака молочной железы;
MCF-7, которые являются клеточной линией рака молочной железы;
Nb2-11C, которая представляет собой клеточную линию гормонально-зависимой лимфомы крысы (т.е. для роста этих клеток необходима добавка гормона роста) (Gertler et al., 1985 Endocrinol.. 116: 1636-1644); и
ND2-SP, которая представляет собой независимую от лактогенного гормона, клеточную линию лимфомы крысы;
(b) неопухолевые клетки:
клетки костного мозга грызунов (включая мышей и крыс);
первичные фибробласты крысы; и IM-9, которая представляет собой клеточную линию лимфоцита человека.
3.1.2. Анализ включения 3Н-тимидина
Испытуемые клетки (клетки, в которых анализировали включение 3Н-тимидина) высевали в чашку Петри с 96 микроячейками, начальная плотность клеток составляла 1x104 клеток/микроячейку. Каждая микроячейка содержала смесь RPMI и испытуемого раствора, который представлял собой или улучшенную среду (УС мышечных клеток или УС лейкоцитов или RPMI или фосфатно-солевой буфер), фракционированную УС, контрольную RPMI или раствор фосфатно-солевого буфера (т.е. неулучшенную среду) или фракционированную контрольную RPMI или раствор фосфатно-солевого буфера. Результаты, полученные для каждого из испытываемых растворов, сравнивали с соответствующим контрольным раствором (например, испытуемый фракционированный раствор в фосфатно-солевом буфере сравнивали с фракционированным фосфатно-солевым буфером и т.д.). После 42 часов инкубирования при 37oС в каждую микроячейку добавили 10 μCi 3Н-тимидина и потом еще 6 часов продолжали инкубирование с этим радиоактивным маркером. После этого количество поглощенного 3Н-тимидина измеряли с помощью жидкого сцинтилляци-онного счетчика.
3.1.3. Определение количества клеток
Клетки Nb2-11C клеточной линии лимфомы крыс синхронизировали и культивировали, как это было описано выше (Gertler et al., 1985. Endocrinol., 116: 1636-1644) за исключением того, что это культивирование проводили в среде RMPI с добавкой 5% сыворотки зародыша теленка. Синхронизацию клеток Nb2-11C в фазе G0/G1 и мониторинг их собственной пролиферации проводили, как это было описано ранее (Gertler et al., там же). Клетки быстро перенесли в среду с добавлением лошадиной сыворотки и инкубировали в течение ночи. После этого клетки разбавили до количества ~ 3х105 клеток/мл и распределили по 0.5 мл в каждую из 24 микроячеек чашки Петри. Затем добавили до 0.5 мл испытуемого раствора и инициировали собственную проилферацию, добавив гормон роста человека до конечной концентрации 2 мг/мл. Клетки инкубировали при 37oС в атмосфере, содержащей 5% CO2, и после их инкубирования в течение 72 часов было определено количество клеток в камере Coulter. Каждый опыт повторяли дважды.
Аналогичным образом проводили испытание клеток Nb2-11C и клеточной линии IM-9.
3.2. Результаты
3.2.1. MF
На фиг. 1 и 2 приведены данные для ультрафильтрата МФ в 3.000 дальтон, полученного от первичной культуры клеток поперечно-полосатых мышц новорожденных крыс; данные для ультрафильтрата МФ в 3.000 дальтон, полученного от клеточной линии L-8, приведены на фиг. 3 и 4. На этих чертежах цифрами по оси абсцисс указано обратное разведение ("1" - неразведенный, "2" - двукратное разведение и т. д. ), а ордината указывает радиоактивность (импульс в минуту); в качестве контрольной среды (левая колонка в каждом случае) использовали среду, обработанную так же, как указанная улучшенная среда УС.
Как видно из фиг. 1 и 3, МФ первичной культуры клеток поперечно-полосатых мышц крыс (фиг. 1) или клеточной линии L-8 поперечно-полосатых мышц крыс (фиг. 3) подавляет пролиферацию опухолевых клеток, в то время как на нормальные клетки (фиг. 3 и 4 соответственно) этот фактор не оказывает такого подавляющего действия. Более того, можно видеть, что МФ фильтруется сквозь мембрану с молекулярным отсечением на 3.000 дальтон, это доказывается тем, что в этом ультрафильтрате сохраняется антипролиферативная активность, присущая сырой улучшенной среде. Как еще можно видеть по фиг. 1 и 3, такое действие МФ уменьшается с увеличением разведения. Это указывает, что указанное воздействие усредняется посредством некоторого специфического фактора указанного ультрафильтрата.
На фиг. 5 показана активность сырой улучшенной среды УС, полученной из линии L-8, и ультрафильтрата МФ в 3.000 дальтон, при различных их разведениях. Как можно видеть, сырая УС проявляет антипролиферативную активность, сохраняемую в указанном ультрафильтрате. Более того, это воздействие уменьшается с увеличением разведения, что вновь указывает на тот факт, что указанное воздействие представляет собой специфическую активность, усредняемую неким фактором.
Влияние ультрафильтрата МФ в 3.000 дальтон (неразведенного), полученного из клеток L-8, приведено также на фиг. 6 (результаты даны в процентах от контроля; контроль в каждом эксперименте принимали за 100%). Как можно видеть, указанный МФ активен в подавлении пролиферации всех испытуемых клеточных опухолевых линий.
На фиг. 7 приведена активность ультрафильтрата МФ в 3.000 дальтон, полученного из миобластов человека, в подавлении пролиферации трех клеточных линий (результаты даны в процентах от контроля). Пролиферацию клеточных линий В-16 и К562 анализировали, определяя внедрения 3Н-тимидина; пролиферацию клеточных линий Nb2 анализировали подсчетом клеток. Можно видеть МФ, полученный от человека, активен в подавлении пролиферации всех испытуемых клеточных опухолевых линий.
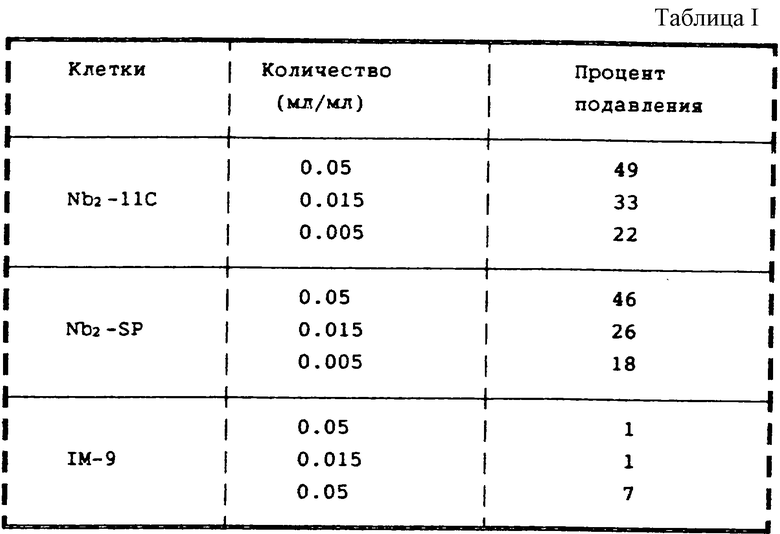
В другом опыте собрали вместе фракции после препаративной ВЭЖХ с обращенной фазой, которые оказались активны в подавлении пролиферации опухолевых клеток (фракции 5 и 6 с фиг.15). Эти фракции упаривали при 45 oС, затем растворили в 5 мл воды, перенесли в пробирки и снова высушили в вакууме, а в заключение растворили в 2 мл воды и простерилизовали. Некоторые количества этих фракций проанализировали, чтобы определить их способность подавлять пролиферацию трех различных клеточных линий Nb2-11C, nb2-SP и IM-9.
Результаты приведены в таблице I (пролиферация, определяемая подсчетом клеток).
Можно видеть, что указанный МФ подавляет пролиферацию как гормонально-зависимой опухолевой клеточной линии Nb2-11C, так и гормонально-независимой опухолевой клеточной линии Nb2-SP. Напротив, указанный МФ практически не оказывает никакого влияния на неопухолевую клеточную линию IM-9 лимфоцитов человека.
На фиг. 8 показано влияние ультрафильтрата ЛФ в 3.000 дальтон на пролиферацию клеточных линий человека и грызунов (крысы, мыши). Снова можно видеть, что указанный ЛФ подавляет пролиферацию всех испытуемых клеточных линий, полученных как от человека, так и от грызунов.
4. Ингибирование in vitro с помощью МФ и ЛФ реакции лимфоцитов на фитогемагглютин и смешанной реакции лейкоцитов
Если лимфоциты двух субъектов культивируют вместе, то антигены альбумина плазмы человека каждого субъекта будут вызывать такую реакцию клетки, которая приведет к пролиферации лимфоцитов (эта пролиферация непосредственно связана с разницей в антигенах альбумина плазмы человека этих двух субъектов). Для того чтобы изучить действие ЛФ или МФ на эту реакцию, в фосфатно-солевом буфере, содержащем 10% сыворотки плода коровы, инкубировали одноядерные клетки от двух доноров (эти клетки были получены так, как это описано выше в пункте 1.2); количество клеток составляло 1х10 6 клеток/мл. К этим клеткам добавили различные разведения ультрафильтрата МФ или ЛФ в 3.000 дальтон. Полученные культуры инкубировали 5 дней при 37 oС во влажной атмосфере, содержащей 5% СО2 и 95% воздуха. В течение последних 6 часов инкубирования в каждую микроячейку добавили 1 μСi 3H-тимидина. Клетки собрали и поглощение 3Н-тимидина определяли с помощью жидкого сцинтилляционного счетчика радиации LKB (LKB, Piscataway, NJ, USA).
Фитогемагглютинин (ФГГ) представляет собой митоген, который связывает сахара лимфоцитов с поверхностью клетки и индуцирует превращение лимфоцитов в форму бластной клетки с последующей пролиферацией клетки. Для изучения влияния МФ или ЛФ на реакции, индуцированные ФГГ, одноядерные клетки (при их концентрации 106 клеток/мл) засевали в чашки Петри с 96 микроячейками. Каждая из этих ячеек содержала 0.2 мл RPMI с добавкой 10% сыворотки зародыша теленка (Israel Industries, Bet-ha-Emek, Israel) и 1 мкг/мл ФГГ (Welcome Laboratories, U. K. ). В некоторых из ячеек эти 0.2 мл RPMI наполовину состояли из нативной RPMI, а наполовину - из ультрафильтрата МФ в 3.000 дальтон из клеток L-8 RPMI. Полученные культуры инкубировали 4 дня в инкубаторе с СО2, а в конце периода инкубации в эти клетки добавили I μСi 3[Н]-тимидина. Инкубирование продолжали еще 24 часа и радиоактивность клеток, собранных с помощью устройства Dyatech, определяли в LKB сцинтилляторе.
На фиг. 9 показано влияние МФ на подавлении реакции лимфоцитов на ФГГ и на смешанную реакцию лейкоцитов (приведены результаты для разведения 1:1, они даны в процентах от контроля). Качественно аналогичные результаты были получены для ЛФ. Более того, воздействие как МФ, так и ЛФ было пропорционально разведению (уменьшение влияния с увеличением разведения) (результаты не приведены).
5. Исследования in vivo
5.1 Индукция опухолевого процесса (МСА 105) внутрибрюшинной инокуляцией; внутрибрюшинная обработка с помощью МФ.
30 мышам C57BL6/J была сделана внутрибрюшинная инъекция 2.5х105 клеток MAC-105. Дважды в день эти мыши получали внутрибрюшинную инъекцию 0.5 мл фракции, полученной в результате ВЭЖХ с обращенной фазой и обозначенной ниже (раздел 6.5.2) как "МФ-SP". На 33 день мышей умертвили и провели оценку опухолевого очага.
На фиг. 10 приведены репрезентативные результаты. На нем показано две мыши с открытой брюшиной; животное, изображенное на фиг. 10А, получало фракцию МФ-SP, а животное на фиг. 10В получало контрольную среду RPMI. Как видно из этих чертежей, у контрольного животного обнаружено значительное распространение опухолевого процесса (стрелка на фиг. 10В), в то время как у животного, получавшего МФ, наблюдался очень незначительный, трудно различимый очаг опухолевого процесса (один показан стрелкой на фиг. 10А).
5.2. Индукция опухолевого процесса (меланома В-16) внутрибрюшинной инокуляцией; обработка внутрибрюшинной инъекцией МФ
40 мышам C57BL6/J делали внутрибрюшинную инъекцию 5x105 клеток меланомы В-16. 20 мышей использовали как контрольные, они ежедневно получали внутрибрюшинную инъекцию раствора фосфатно-солевого буфера. 20 мышей ежедневно обрабатывали 1 мл ультрафильтрата МФ в 3.000 дальтон, полученного из клеток L-8. На 15 день мышей умертвили и оценивали степень распространения опухолевого процесса в брюшине этих животных. Из фиг. 11, на которой изображены открытые брюшины двух репрезентативных мышей из каждой экспериментальной группы, видно, что в то время как у животного контрольной группы имеются многочисленные крупные очаги опухолевого процесса, у животного, прошедшего обработку, имеется всего лишь несколько значительно более мелких очагов.
5.3. Индукция опухолевого процесса (МСА-105) внутрибрюшинной инокуляцией; обработка с использованием МФ как при внутрибрюшинном, так и при пероральным введении
30 мышам C57BL6/J была сделана внутрибрюшинная инъекция 2.5х105 клеток MAC-105. 10 мышам ежедневно проводили пероральное введение 1 мл ультрафильтрата МФ в 3.000 дальтон, полученного из мышечных клеток L-8; 10 мышам ежедневно делали внутрибрюшинную инъекцию того же самого раствора; а 10 мышей представляли собой контрольную группу и они получали перорально RPMI.
На 30 день мышей умертвили, и на их выделенных брюшинах изучали распространение опухолевого процесса. На фиг. 2 показаны две репрезентативные мыши из каждой группы (фиг. 12А - мыши, обработанные МФ; фиг. 12В - контрольные). Как видно из фиг. 12А, на брюшине животных, обработанных МФ, нет никаких признаков развития опухолевого процесса, в то время как на брюшине животных из контрольной группы появляется крупный очаг опухолевого процесса. В этом эксперименте у 90% мышей из контрольной группы распространялся очаг опухолевого процесса, в то время как только у 40% мышей, обработанных (или внутрибрюшинно или перорально) МФ, наблюдалось распространение этого процесса.
5.4. Индукция опухолевого процесса (рак молочной железы DA3) внутримышечной инокуляцией; обработка МФ при внутрибрюшинном и пероральном введении
30 мышам BALB/C делали в лапу внутримышечную инъекцию 1x106 клеток DA3. 10 мышей служили в качестве контрольных и они ежедневно получали инъекцию раствора фосфатно-солевого буфера; 10 мышам ежедневно делали внутрибрюшинную инъекцию 1 мл ультрафильтрата МФ в 3.000 дальтон, полученного из мышечных клеток L-8 (полученных в растворе фосфатно-солевого буфера); а 10 мышей ежедневно получали перорально 1 мл раствора, содержащего МФ. В легких мышей распространялся крупный опухолевый процесс; спустя 3 недели этих мышей умертвили, и в их легких можно было выявить очаги метастазов и сосчитать их. В контрольной группе обнаружили 23.4±6 очага метастазов; в группе животных, обработанных внутрибрюшинно, обнаружили 6.2±1.3 очага метастазов; а в группе животных, обработанной перорально - 2.2±0.6 очага.
5.5 Индукция опухолевого процесса (меланома В-16) внутривенной инокуляцией; обработка МФ при пероральном введении
30 мышам C57BL6/J делали внутривенную инъекцию 5x105 клеток меланомы В-16. 20 мышей получали перорально ежедневно, начиная со дня инокуляции опухолевого процесса, 1 мл ультрафильтрата МФ в 3.000 дальтон, полученного из клеток L-8; 10 мышей служили контролем, они получали перорально только раствор фосфатно-солевого буфера. На 18 день мышей умертвили, а в их легких можно было обнаружить развившиеся черные очаги опухолевого процесса. На фиг. 13 изображены репрезентативные легкие животных из двух этих групп (верхний ряд - контрольный; нижний ряд - обработанный МФ). Можно видеть, что в то время как в легких животных контрольной группы обнаружено много черных очагов метастазов, в экспериментальной группе имеется несколько мелких очагов метастазов.
6. Уровень ЛФ, выделяемого лейкоцитами здоровых особей и раковых пациентов
Одноядерные клетки были выделенными из венозной крови 23 здоровых людей (образцы, полученные из банка крови) и 33 стационарных раковых больных. Процедура выделения была такая, как это описано в пункте 1.2. С культуры одноядерных клеток (2х10 6 клеток/мл) собрали супернатанты, потом каждый из них подвергли ультрафильтрации через фильтр, с молекулярным отсечением на 3.000 дальтон, как это описано в пункте 2.
Полученные ультрафильтраты испытали на их способность подавлять рост раковых клеток от трех раковых клеточных линий: меланомы В-16 грызунов (включая мышей и крыс); меланомы SK человека; и клеточной линии К-562 лейкемии человека.
Результаты приведены на фиг. 14. Из фиг. 14А видно, что в то время как в ультрафильтрате одноядерных клеток здоровых людей наблюдается значительное подавление пролиферации (которая для всех испытуемых клеточных линий была ниже 50%) в ультрафильтрате, полученном от раковых пациентов, наблюдалось лишь слабое, едва заметное подавление пролиферации. Далее, как видно из фиг. 14В. между этими двумя группами абсолютно не наблюдалось перекрывания.
Эти результаты показывают, что уровень ЛФ, выделяемого одноядерными клетками раковых пациентов, значительно ниже, чем уровень, выделяемый лимфоцитами здоровых людей. Эти результаты указывают диагностическую важность исследования уровня НМ-ЭРР настоящего изобретения. Кроме этого, и высокий уровень ЛФ имеет важное терапевтическое значение.
7. Характеристика МФ
7.1. Биологический анализ
УС мышечных клеток фракционировали (это будет детально описано ниже), и полученные фракции анализировали с использованием упомянутых выше клеточных линий. Влияние каждой фракции на рост клеток определяли по поглощению 3H-тимидина или описанными ранее методами подсчета клеток.
7.2. Изучение мнимой протеиновой природы МФ
Для того чтобы определить, представляет ли собой МФ протеиноподобное вещество или нет, УС мышечных клеток (полученных от первичной клеточной культуры мышц крысы) исследовали в серии опытов, включая чувствительность к протеолитическим ферментам, стабильность во время лиофилизации и влияние инкубации при разных температурах. После этих опытов указанную УС привели снова к исходной концентрации соли и протеина. Последнее осуществили разбавлением исходной среды, или если она разбавлена, то при оценке этих результатов принимали во внимание фактор разбавления. Был использован анализ поглощения 3H-тимидина.
7.2.1. Влияние протеолитических ферментов
Испытывали протеолитические ферментные препараты, трипсин и проназа. Для определения влияния трипсина использовали две методики:
(i) раствор, содержащий МФ, инкубировали 4 часа с трипсином (0.5-2 мкг/мл) при 37oС, после чего активность трипсина блокировали добавкой почти двукратного молярного избытка ингибитора соевого трипсина (STI). В качестве контрольной использовали неулучшенную (не содержащую МФ) среду;
(ii) раствор, содержащий МФ, инкубировали 1-4 часа с трипсином (0.5-2 мкг/мл) при 37oС или при комнатной температуре в течение ночи. После инкубирования фермент перенесли на колонку с п-аминобензамидинагарозой. В качестве контрольной использовали среду, не содержащую МФ. Последующие контрольные испытания с одним только трипсином показали, что когда в качестве подвижной фазы использовали бикарбонатиый буфер, то из этой колонки не элюировалось никакой триптической активности.
Для испытания действия проназы мышечные клетки УС привели в контакт с проназой, иммобилизованной на геле сефарозы. В этом случае в качестве контрольной также использовали среду, не содержащую МФ. В таблице II приведены полученные результаты (числами указывают % подавления по сравнению с неулучшенной контрольной средой).
Как показано в таблице II, обработка как трипсином, так и проназой по обеим методикам не оказывает существенного воздействия на активность МФ в подавлении опухолевого процесса.
7.2.2 Лиофилизация
УС мышечных клеток лиофилизировали без ее предварительного диализа, а полученный лиофилизат потом растворили в воде до его первоначального объема. После такой обработки не происходило существенной потери активности МФ в подавлении опухолевого процесса, что указывает на стабильность МФ к лиофилизации.
7.2.3 Тепловая обработка
УС мышечных клеток (полученную из указанной ранее культуры мышечных клеток крыс) обрабатывали в течение разного времени в диапазоне температур от 4 до 100oС. После этого образцы испытывали как с помощью клеток МСА, так и НТВ клеток. Результаты приведены в таблице III (цифрами указано изменение в (%) ингибиторной активности МФ после тепловой обработки).
В других сериях опытов ультрафильтрат улучшенной среды в 3.000 дальтон нагревали вплоть до кипения, что происходило без всякой потери активности указанной низкомолекулярной фракции в подавлении пролиферации опухолевых клеток.
Приведенные выше результаты показывают, что в диапазоне всех анализируемых температур, включая кипение при 100oС, не наблюдалось снижения ингибиторного потенциала.
7.2.4. Резюме
В условиях экспериментов не наблюдалось никакого уменьшения потенциала МФ в подавлении роста опухолевых клеток, это ясно показывает, что этот МФ не является протеином. Напротив, полученные результаты показывают увеличение ингибиторной активности после некоторых видов обработки. Последнее можно объяснить тем фактом, что среда, содержащая МФ, включает протеиноподобный фактор, который оказывает противоположное действие на МФ и который разрушается при такой обработке.
7.3. Размер МФ
Улучшенную среду УС, содержащую МФ, фракционировали ультрафильтрацией на мембранах Amicon с молекулярным отсечением в 10, 2 и 0.5 kD. Полученный остаток в каждом случае дважды фильтровали, используя по меньшей мере один дополнительный объем раствора фосфатно-солевого буфера. Существенная часть ингибиторной активности (выше 90%) была обнаружена в первых двух фильтратах; а для 0.5 kD мембраны приблизительно 80% активности было обнаружено в указанных фильтратах, а некоторая ее часть (20%) сохранялась во втором остатке. Ингибирующая активность оценивалась в каждом случае как на клетках НТВ 38, так и на клетках МСА.
Диализ УС, содержащей МФ, сквозь мембраны с молекулярным отсечением в 12 и 3 kD показал, что этот активный компонент проходил сквозь обе мембраны.
Приведенные выше результаты показывают, что указанный МФ имеет молекулярный вес порядка ~ 500 дальтон или менее.
7.4. Характеристика МФ (полученного из первичной культуры мышечных клеток крыс) с помощью ВЭЖХ с обращенной фазой
Фильтраты мембран на 10 kD подвергли хроматографии на колонке С 18 (4x250 мм) с обращенной фазой. Фильтрат довели до начальной концентрации разбавлением в растворе 0.1% трифторуксусной кислоты, прежде чем его поместили в колонку, которую "проявляли" с помощью 5-35% градиента ацетонитрила (оба эти компонента предварительно испытали, чтобы определить, что они не влияют на ингибиторную активность МФ). Указанную активность анализировали на клетках НТВ-38, вымываемых при 15% ацетонитрила в 0.1% трифторуксусной кислоты. Потом эта активная фракция была подвергнута повторной хроматографии на той же самой колонке и при том же градиенте ацетонитрила и частично очищенного МФ, вымытым в том же положении, что и раньше (время удерживания ~ 21 мин). Третий раз хроматография с обращенной фазой проводилась с этой последней фракцией при тех же условиях, что и первые два опыта.
Был обнаружен единственный ингибиторный пик, и он соответствует положению 220 нм пика поглощения в профиле вымывания, что видно на фиг. 15.
7.5. Высокий уровень очистки МФ (полученного из первичной культуры мышечных клеток крыс)
7.5.1. Протокол эксперимента
УС мышечных клеток крыс подвергли ультрафильтрации на мембранах Amicon 10 kD. Хроматографию фильтрата 10 kD проводили на колонке С-18 (47x300 мм) с обращенной фазой, эта колонка была присоединена к препаративной ВЭЖХ колонке. Полученные фракции анализировали методом клеточной пролиферации, используя НТВ-38 клетки, а полученную активную фракцию повторно хроматографировали на второй колонке С-18 с обращенной фазой (аналитическая колонка: 4x250 мм); эту активную фракцию идентифицировали так же, как это указано выше.
7.5.2. Активность очищенных фракций МФ
Вещество, полученное на последней стадии в разделе 7.5.1. назвали "МФ-Р". а вещество, подученное из первой колонки с обращенной фазой, назвали "МФ-SP" (эту же фракцию испытывали в разделе 5.1). В таблице IV указаны как активности сырой улучшенной среды (УС), фракций МФ-Р и МФ-SP, так и остатка (R) ультрафильтрации (активность выражена в ед/мл, 1 единицу определяют как такое количество вещества, которое в опыте вызывает 50% подавление пролиферации).
Видно, что все анализируемые фракция активны в подавлении пролиферации всех испытуемых клеточных линий. В таблицу IV включена также R фракция, активный фактор которой может быть агентом подавления опухолевой пролиферации, раскрываемым в патенте США 5242692.
7.6. Характеристика методом ВЭЖХ МФ, полученного от клеточной линий L-8.
В разделах 7.6.1-7.6.5 приведены некоторые репрезентативные результаты ВЭЖХ. В разделе 7.6.1 приведен типичный метод очистки данного МФ.
7.6.1. ВЭЖХ с обращенной фазой
На колонке (С-18) ВЭЖХ с обращенной фазой осуществляли хроматографию 160 мл ультрафильтрата МФ в 3.000 дальтон в растворе фосфатно-солевого буфера (полученного после 8 часов инкубирования клеток мышц с этим раствором фосфатно-солевого буфера). Вымываемая жидкость представляла собой градиент воды для ВЭЖХ, полученной в устройстве B-pure (Barnstead, Du-buque, Iowa) и ацетонитрил для ВЭЖХ (G.T. Baker, USA). Градиент вымываемой жидкости составлял от 0 до 60% ацетонитрила за 30 минут. Скорость течения составляла 100 мл/минуту. Собрали две минутные фракции (каждая содержала по 200 мл).
20 мл каждой из фракций, полученных ВЭЖХ, выпарили досуха в концентрирующей центрифуге, а высушенные фракции потом суспендировади в 100 мкл воды. Затем снова высушили досуха и растворили в 2 мл раствора фосфатно-солевого буфера; после чего активность подавления пролиферации исследовали подсчетом клеток. Его проводили, помещая 0.2 мл каждой фракции в микроячейку тестера, содержащую клетки.
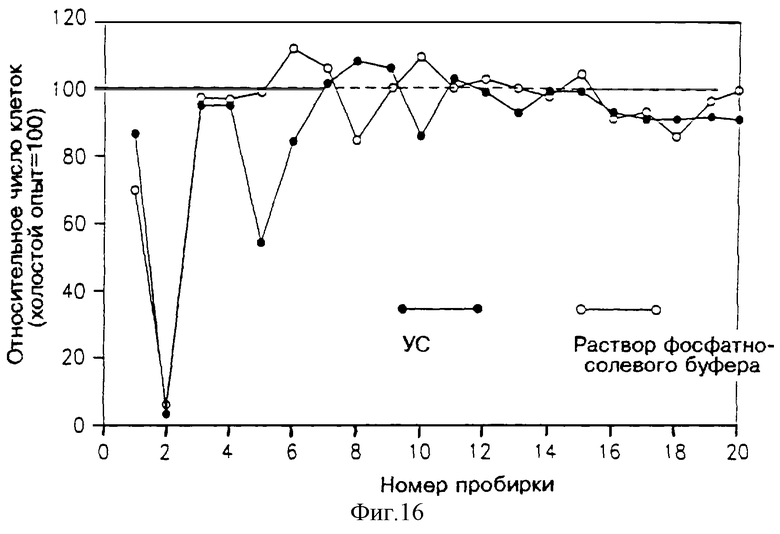
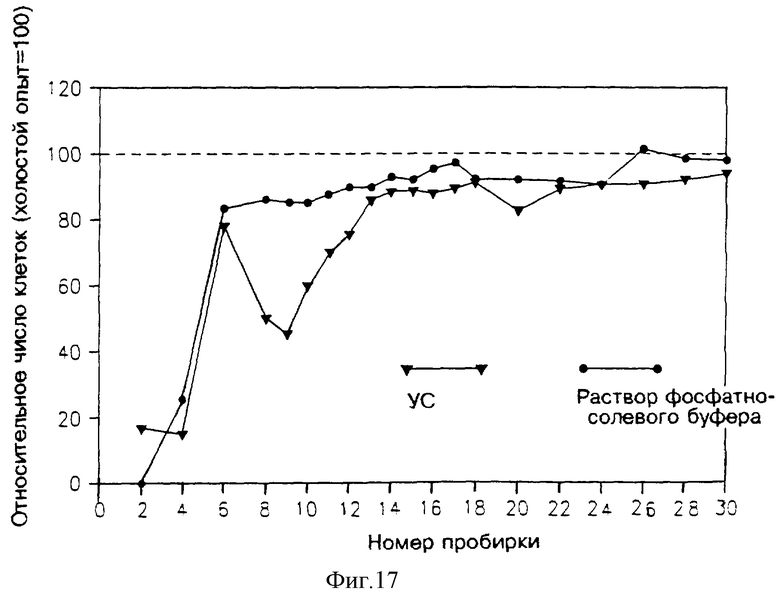
На фиг. 16 и 17 показана активность разных фракций в двух разных опытах. В обоих случаях результаты указывают на присутствие специфического ингибирования при вымывании во фракциях 5 и 6 на фиг. 16, и во фракциях 8-12 на фиг. 17.
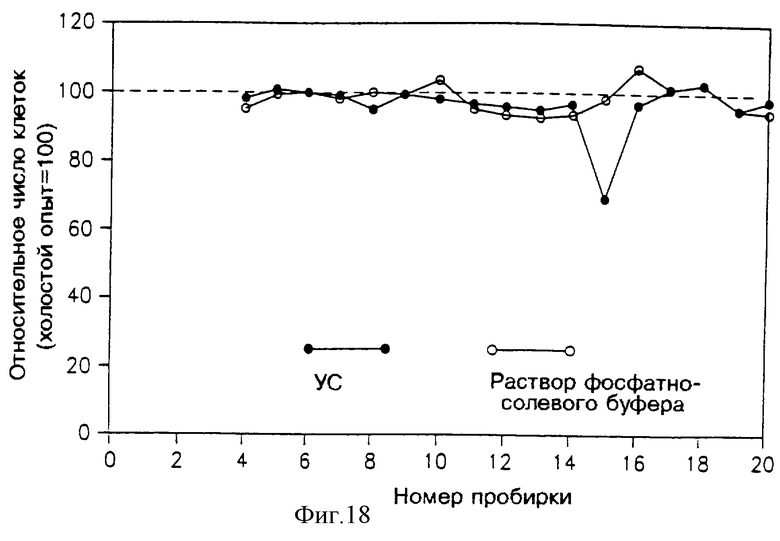
Фракции 5 и 6 с фиг. 16 слили вместе, упаривали при 45oС, растворили в 5 мл воды, перенесли в пробирки и снова высушили в вакууме, растворили в 2 мл воды и простерилизовали. 1 мл из этих концентрированных фракций подвергли хроматографии в аналитической колонке ВЭЖХ с обращенной фазой (С-18) в течение 20 минут при градиенте подвижных текучих сред от 100% воды и 60% ацетонитрила, скорость потока составляла 1 мл/мин. Собрали фракции, вымываемые через 1 минуту (каждая по 1 мл), высушили, растворили в 0.4 мл RPMI, содержащем 5% HS, и простерилизовали. 0.15 мл полученного раствора добавили в каждую из ячеек, содержащую клетки (активность определяли методом подсчета клеток). Для контроля растворы фосфатно-солевого буфера фракционировали ВЭЖХ и аналогичным образом испытали их активность.
На фиг. 18 показана активность различных вымываемых фракций. Полученные результаты указывают на наличие специфического ингибирования во фракции 15.
7.6.2. ВЭЖХ на колонке Superdex
Активные фракции с фиг. 17 разделили на две объединенные фракции; первую фракцию, названную "FR1", получили из комбинации 1200 мл из пробирок 6-8, 550 мл из пробирок 8-9, 300 мл из пробирок 9-10 и 30 мл из пробирок 10-11; вторую фракцию, названную "FR2", получили из 250 мл из пробирок 9-10, 520 мл из пробирок 9-11 и 600 мл из пробирок 11-12. И фракцию FR1, и фракцию FR2 высушили и каждую растворили в 10 мл дистиллированной воды. После того как в результате центрифугирования получили нерастворимый осадок, провели экстракцию этих фракций 10 мл (в случае FR1) и 5 мл (в случае FR2) раствора фосфатно-солевого буфера. Даже после второй экстракции остался значительный осадок.
Полученные результаты показывают, что большая часть активности присутствовала во фракции FR1; активность FR2 составляла только ~ 15-20% от активности FR1. Экстракт нерастворимого препарата раствором фосфатно-солевого буфера давал только ~ 4% активности от активности FR1.
2 мл FR2 высушили и растворили в 0.4 мл воды. 0.2 мл обработали на колонке superdex, уравновешенной раствором фосфатно-солевого буфера. Колонку "проявляли" при 1 мл/мин, используя раствор фосфатно-солевого буфера в качестве подвижной текучей среды, и собрали 1 мл фракций. Сбор фракций начали спустя 0.3 минуты после начала разделения. Элюированные фракции простерилизовали и добавили по 0.2 мл их в каждую ячейку.
На фиг. 19 показана активность различных фракций, элюированных из колонки superdex. Специфическое ингибирование можно наблюдать во фракциях 18-22. Фракции 18-22 собрали вместе и подвергли повторной хроматографии на той же самой колонке при таких же условиях. Результаты этой повторной хроматографии указывают, что в этих фракциях возможно присутствие смеси двух или более активных агентов ("МФ").
Аналогичный профиль вымывания получили также для FR1, хотя эти результаты, по-видимому, показывают, что фракция FR1 примерно в 10 раз более концентрирована по содержанию в ней МФ по сравнению с FR2.
7.6.3. Аналитическая ВЭЖХ с обращенной фазой, проведенная после superdex ВЭЖХ
Фракции FR1 и FR2 объединили и 1 мл аликвоты анализировали на ВЭЖХ колонке (С-18) с обращенной фазой. 1 мл фракций собрали, высушили и растворили в 0.4 мл RPMI, содержащем 5% HS; 0.5 мл этого раствора добавили в каждую ячейку, содержащую клетки. Результаты приведены на фиг. 20. Как видно, существует только один острый пик активности в пробирке 16. Кроме того, наблюдается также слабая ингибиторная активность в пробирке 4. Эти результаты показывают, что в УС мышечных клеток возможно наличие более одного агента, подавляющего опухолевую пролиферацию.
7.6.4. ВЭЖХ "эксклюзивного размера" (SE)
Фракции 6-8 ("А"), 8-10 ("В"), 10-12 ("С") и 12-14 ("D") из препаративной колонки ВЭЖХ с обращенной фазой хроматографировали отдельно в колонке, содержащей частицы силикагеля, покрытые полигидроксиэтиласпартамидом (PolyHYDROXY-ETHYL-A, полученный PolyLC). В качестве подвижной текучей среды использовали изократический раствор (т.е. без какого-либо градиента), который представлял собой водный раствор 50 мкМ муравьиной кислоты.
Собрали фракции в 0.5 мл, их высушили и растворили в 0.8 мл раствора фосфатно-солевого буфера. 0.15 мл каждой из фракций потом добавили в каждую из ячеек, содержащих клетки. На фиг. 21 приведены результаты хроматографии. Эти результаты указывают, что во фракциях 6-8 нет никакой активности, кроме пожалуй слабой активности в пробирке 32. Во фракции В в пробирках 29-37 можно видеть очень ясный и широкий пик активности. Во фракции С имеется 2 ясных пика активности, один в пробирках 21-23, а другой в пробирках 28-31. Наконец, во фракции D в пробирках 21-22 появляется единичный пик активности.
7.6.5. Superdex ВЭЖХ, проводимая после SE ВЭЖХ
Активные фракции, элюированные из колонки SE, собрали и высушили в концентрирующей центрифуге (Hetovac VR-1, производитель Неto, Дания) под высоким давлением, растворили в 0.4 мл, а потом 0.2 мл образцы отделяли на колонках superdex, равновешанных раствором фосфатно-солевого буфера. Собрали фракции в 1 мл и потом 0.2 мл добавили в каждую ячейку, содержащую клетки.
Как видно из фиг. 22А, SE-ВЭЖХ пробирок 29-33 фракции В показывает активность в пробирках 21-24, вымываемых из колонки superdex. Профиль этой активности может указывать на возможность существования более чем одного агента или на то, что этот активный агент существует в нескольких формах.
На фиг. 22В показан специфический ингибиторный пик, вымываемый от 17 до 21 минуты.
7.7.6. ВЭЖХ с "гидрофильным взаимодействием"
Фракции 27-32 из аликвоты SE ВЭЖХ с фиг. 25 собрали вместе, высушили и растворили в 0.2 мл воды. В колонку ВЭЖХ с гидрофильным взаимодействием (СНО), (PolyGlycoplex, произведенную Poly-LC) дважды впрыскивали по 0.085 мл. Затем колонку "проявляли" раствором, содержащим 70% ацетонитрила в воде при скорости фракции 1 мл/мин. 1 мл фракции собрали, высушили и растворили в 0.6 мл раствора фосфатно-солевого буфера, а затем 0.1 мл фракций добавили в каждую из 96 ячеек микротитровальной чашки и проводили анализ содержания солей (раздел 3.3.1)
Профиль активности раствора, вымываемого из этой колонки, приведен на фиг. 23. Можно видеть, что самое сильное специфическое подавление аликвоты из колонки СНО было обнаружено во фракциях 2-8, хотя вторичный пик подавления можно также видеть во фракции 10. Слабую активность также можно было наблюдать в широком пике фракций 21-30.
8. Очистка МФ
8.1. Процедура
Для очистки МФ был разработан метод, включающий следующие стадии:
(a) Получение улучшенной среды:
Улучшенную среду получали из клеток L-8 в растворе фосфатно-солевого буфера, как это описано в пункте 1.1.2.
(b) Ультрафильтрация:
Улучшенную среду подвергли ультрафильрации сквозь мембрану с молекулярным отсечением на 3.000 дальтон, как это описано в пункте 2.
(c) Препаративная ВЭЖХ с обращенной фазой
Полученный ультрафильтрат в 3.000 дальтон хроматографировали на препаративной колонке ВЭЖХ с обращенной фазой (С-18). Обычно эту колонку сначала 20 минут промывали водой для ВЭЖХ, потом в нее загрузили 200 мл ультрафильтрата в 3.000 дальтон и в течение следующих 10 минут промывали водой. Затем эту колонку "проявляли" в течение 30 минут с помощью ацетонитрила: градиент воды от 0% ацетонитрила до 60% ацетонитрила. Скорость подвижной текучей среды составляет ~ 100 мл/мин. Активные фракции вымываются при их выходе из колонки в интервале от 8 до 14 минут.
Полученные активные фракции потом собрали вместе и объединенную фракцию концентрировали в ротационном испарителе (Rotovap, Buchi, Switzerland), а потом в концетрирующей центрифуге.
(d) Аналитическая ВЭЖХ с обращенной фазой
Объединенную, сконцентрированную и высушенную фракцию потом растворили в 5 мл воды, снова высушили в вакууме и растворили в 2 мл воды. 1 мл этой концентрированной фракции хроматографировали 20 минут на аналитической колонке ВЭЖХ (С-18), в качестве подвижной текучей среды использовали градиент ацетонитрила: градиент воды от 0% ацетонитрила до 60% ацетонитрила при скорости потока 1 мл/мин. Колонку загрузили и затем промывали 5 минут водой. После этого промывания колонку "проявляли" градиентом ацетонитрила.
Активная фракция вымывается при выходе фракций из колонки от 12 до 19 минут.
Потом активные фракции концентрировали и высушили в концентрирующей центрифуге.
(e) Хроматография "эксклюзивного размера".
Высушенные фракции смешали с муравьиной кислотой и 200 мл образцы этого раствора загрузили в колонку "эксклюзивного размера", описанную в пункте 7.6.4, проявляющая жидкость и условия перегонки описаны там же. Активные фракции вымывались спустя 27-32 минут.
Следует отметить, что разные колонки, даже те, характеристики которых аналогичны описанным выше, и незначительные изменения в условиях элюирования, могут давать разные профили вымывания, и, следовательно, активные фракции могут вымываться спустя время удерживания, отличное от времени, указанного выше. Однако, испытывая ингибиторную активность каждой фракции, как это было описано выше, специалист может, без какой-либо сложности, изолировать и выделить очищенные фракции, содержащие РР настоящего изобретения.
8.2. Результаты очистки
На фиг. 24 показан профиль ингибиторной активности фракций, элюированных из аналитической колонки ВЭЖХ с обращенной фазой и активность фракций, вымываемых из колонки "эксклюзивного размера" (фиг. 25), причем методика очистки такая, как это было описано выше в разделе 8.1
9. ЯМР-вероятная олигосахаридная природа МФ
Активные фракции, элюированные из колонки "эксклюзивного размера" (такие, как те, что показаны на фиг. 25) высушили, а потом растворили в метаноле (проводилось три следующих друг за другом цикла сушки с последующим растворением в метаноле). Потом метанольный экстракт высушили досуха, а полученный препарат повторно растворили в 0.5 мл дейтерометанола. Остаток после экстракции метанолом растворили в 0.5 мл дейтерированной воды.
На фиг. 26 приведен ЯМР-спектр. Видно, что ЯМР-сканирование водной фракции дает один, вероятно несущественный пик, в то время как ЯМР-сканирование метанольного экстакта дает несколько пиков в диапазоне 3-4х10-6. Эти пики характерны для протонов С-ОН групп, типичных для олигосахаридов.
Полученные результаты допускают вероятность того, что МФ является олигосахаридом.
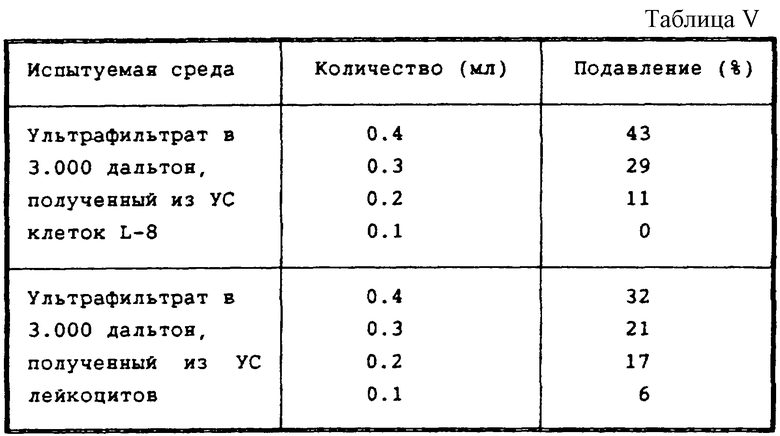
Активность ультрафильтрата в 3.000 дальтон УС лейкоцитов и их фракционирование.
Была испытана активность ультрафильтрата в 3.000 дальтон, полученного из УС лейкоцитов, и ее сравнивали с активностью ультрафильтрата в 3.000 дальтон из УС клеток L-8. Результаты показаны в таблице V.
200 мл ультрафильтрата в 3.000 дальтон, полученного из УС лейкоцитов, разделяли препаративной ВЭЖХ с обращенной фазой; разделение проводили так, как это описано выше (смотри раздел 7.6.6). Собрали фракции в 200 мл. 10 мл аликвоты (из 200 мл) высушили и растворили в 0.6 мл. Потом в каждую из ячеек поместили 0.1 мл.
Для сравнения также фракционировали ультрафильтрат в 3.000 дальтон, полученный из УС клеток L-8.
Результаты приведены в таблице VI.
Как видно, обе аликвоты демонстрируют ингибиторный пик во фракциях 4-6.
| название | год | авторы | номер документа |
|---|---|---|---|
| ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ, СОДЕРЖАЩИЕ АГОНИСТ ИЛИ АНТАГОНИСТ АДЕНОЗИНОВОГО РЕЦЕПТОРА | 2000 |
|
RU2239455C2 |
| ВЫСОКОЭФФЕКТИВНЫЙ СПОСОБ ПОЛУЧЕНИЯ ЛЕКАРСТВЕННОЙ ФОРМЫ АДРЕСНОГО ДЕЙСТВИЯ ДЛЯ ТЕРАПИИ ЗЛОКАЧЕСТВЕННЫХ НОВООБРАЗОВАНИЙ | 2019 |
|
RU2727924C1 |
| ВЕКТОРНЫЙ ПОЛИПЕПТИД - АНАЛОГ ФРАГМЕНТА ТРАНСФОРМИРУЮЩЕГО ФАКТОРА РОСТА АЛЬФА (ТФРАЛЬФА), ЕГО ПРОТИВООПУХОЛЕВЫЙ КОНЪЮГАТ И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ОСНОВЕ КОНЪЮГАТА | 2004 |
|
RU2277930C1 |
| ПРИРОДНЫЕ ПРОТИВООПУХОЛЕВЫЕ ИЛИ ПРОТИВОВИРУСНЫЕ ВЕЩЕСТВА И ИХ ПРИМЕНЕНИЕ | 1998 |
|
RU2205010C2 |
| ПОЛИПЕПТИД, ЯВЛЯЮЩИЙСЯ АНАЛОГОМ РЕЦЕПТОРСВЯЗЫВАЮЩЕГО ФРАГМЕНТА ЭПИДЕРМАЛЬНОГО ФАКТОРА РОСТА С 21-Й ПО 31-Ю АМИНОКИСЛОТУ, ЕГО КОНЪЮГАТ С ДОКСОРУБИЦИНОМ И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ НА ЕГО ОСНОВЕ | 2001 |
|
RU2196604C1 |
| ПОЛИФУНКЦИОНАЛЬНЫЙ БИОСОВМЕСТИМЫЙ ГИДРОГЕЛЬ И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2001 |
|
RU2205034C1 |
| КОМПОЗИЦИИ ПОДВЕРГНУТЫХ РЕСТРИКЦИИ КЛЕТОК, СПОСОБНЫХ К БЫСТРОМУ РОСТУ, КОТОРЫЕ ПРОДУЦИРУЮТ ВЕЩЕСТВА, ПОДАВЛЯЮЩИЕ ПРОЛИФЕРАЦИЮ КЛЕТОК, И ИХ ПРИМЕНЕНИЕ | 1999 |
|
RU2236855C2 |
| Антитела против рецептора фактора роста эндотелия сосудов 1 (VEGFR1) человека | 2023 |
|
RU2838680C1 |
| ЭКСТРАКТЫ АКУЛЬЕГО ХРЯЩА, ОБЛАДАЮЩИЕ ПРОТИВОКОЛЛАГЕНОЛИТИЧЕСКИМ, ПРОТИВОВОСПАЛИТЕЛЬНЫМ, АНТИАНГИОГЕННЫМ И ПРОТИВООПУХОЛЕВЫМ ДЕЙСТВИЕМ, СПОСОБ ПОЛУЧЕНИЯ, СПОСОБЫ ПРИМЕНЕНИЯ И СОДЕРЖАЩИЕ ИХ КОМПОЗИЦИИ | 1996 |
|
RU2181292C2 |
| ТРИТЕРПЕНОВЫЕ КОМПОЗИЦИИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 1999 |
|
RU2244547C2 |













Изобретение относится к биотехнологии, медицине и ветеринарии и может быть использовано для лечения и предотвращения рака, а также состояний, вызванных гиперактивной иммунной системой. Низкомолекулярный эндогенный фактор роста (НМ-ЭРР) имеет молекулярный вес менее 3000 дальтон, растворим в воде, термически стабилен, не протеиноподобен, ингибирует рост клеток. НМ-ЭРР секретирует мышечные клетки и лейкоциты. НМ-ЭРР получают путем культивирования мышечных клеток или лейкоцитов. Супернатант клеточной культуры собирают, фильтруют через мембрану с молекулярным отсеканием 3000 дальтон и очищают хроматографически. Рак или предрасположенность к заболеванию рака диагносцируют по превышению содержания НМ-ЭРР у пациента по сравнению со средним значением содержания НМ-ЭРР у здоровых людей. Лечение или профилактику рака осуществляют путем введения пациенту эффективного количества регулятора роста. Изобретение позволяет обеспечить средство для подавления пролиферации опухолевых клеток и иммунносупрессии. 6 с. и 15 з. п. ф-лы, 26 ил., 6 табл.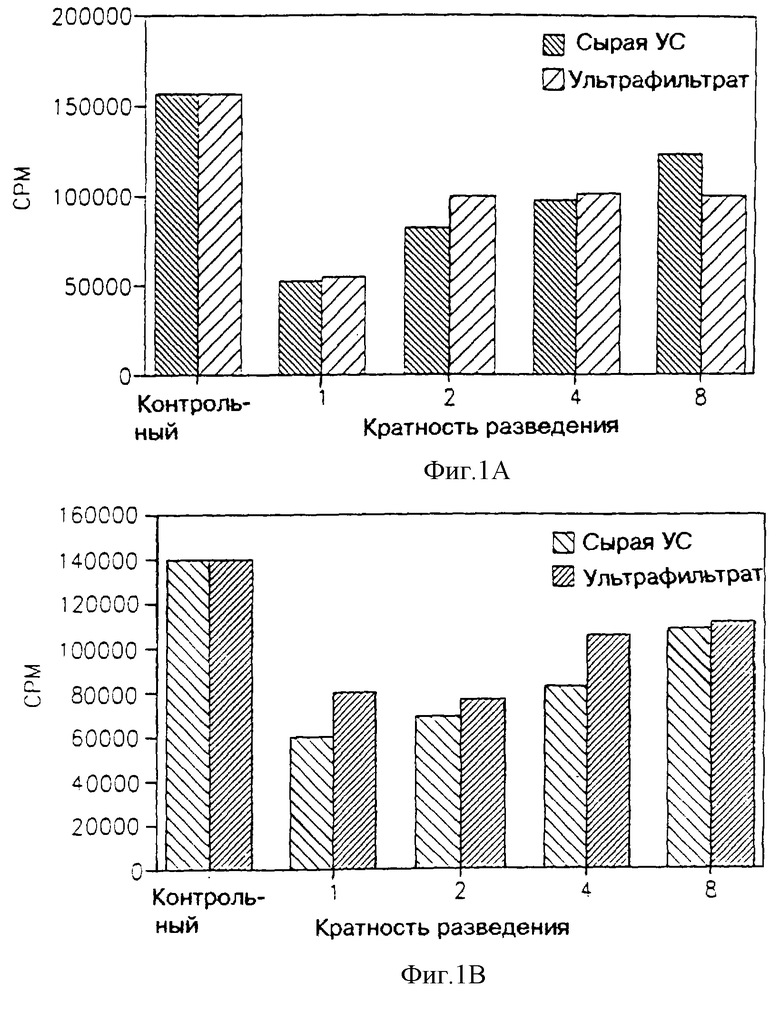
| Приемник градиента давления | 1973 |
|
SU466440A1 |

