Изобретение обеспечивает способ детектирования мишеневого полинуклеотида в пробе, например, при мониторинге реакции амплификации, предпочтительно количественным образом, а также зонды и наборы для использования в этом способе. Способ также пригоден для детектирования свойств таких последовательностей, как полиморфизмы или аллельная вариация, и, таким образом, может быть использован в диагностических методах.
Известные флуоресцентные мониторинговые методы с использованием полимеразной цепной реакции (ПЦР) включают как специфичные к цепи, так и общие для ДНК интеркалярные методы, которые можно использовать с несколькими устройствами второго поколения для термического циклирования в ПЦР.
Общие способы используют интеркалярные ДНК-красители, которые проявляют повышенную флуоресцентность, когда связываются с двухцепочной ДНК. Повышение флуоресцентности, происходящее за счет увеличения объемной концентрации ДНК во время амплификации, можно использовать для оценки протекания реакции и для определения числа копий молекулы-мишени. Кроме того, при мониторинге флуоресценции с контролируемой сменой температуры можно получить кривые плавления ДНК, например, в конце термического циклирования в ПЦР.
Общие для ДНК способы контролируют подъем в объемной концентрации ДНК без каких-либо временных затруднений. Однократное определение флуоресценции можно провести в одной и той же точке в каждой реакции. Анализ кривой плавления в конечной точке можно использовать для того, чтобы отличить артефакты от ампликона и чтобы отличить ампликоны. Пики продуктов можно видеть при концентрациях, которые невозможно визуально оценить при электрофорезе в агарозном геле.
Для того чтобы получить данные по плавлению с высоким разрешением, опыты по плавлению должны проводиться на существующем оборудовании медленно, до пяти минут. Однако при постоянном мониторинге флуоресцентной амплификации можно получить изображение 3D гистерезиса плавления и гибридизации. Это изображение 3D является зависимым от ампликона и может обеспечить достаточной информацией для распознания продукта.
Было установлено, что анализ кривой плавления ДНК вообще является мощным инструментом в оптимизации термальной цикличности ПЦР. Путем определения температур плавления ампликонов возможно снизить температуры денатурации в более поздних циклах ПЦР до этой температуры. Оптимизация амплификации, начиная с продуктов реакции первого поколения, в большей степени, чем геномной ДНК, снижает образование артефактов, имеющее место в более поздних циклах. Температуры плавления олигонуклеотидных праймеров и их комплиментарных последовательностей можно использовать для определения температур их отжига, снижая необходимость в эмпирической оптимизации.
Однако общие интеркалярные способы являются только полуспецифичными к цепи и, следовательно, не являются очень полезными, когда необходимо специфичное детектирование цепи.
В специфичных к цепи способах используются дополнительные реакционные компоненты нуклеиновых кислот для прослеживания протекания реакций амплификации. В этих способах можно применять передачу энергии флуоресценции (FET) в качестве основы для детектирования. Один или более зондов нуклеиновых кислот метят флуоресцентными молекулами, одна из которых способна действовать в качестве донора энергии, и другая из них является молекулой-акцептором энергии. Они иногда определяются как соответственно репортерная молекула и гасящая молекула. Молекула-донор возбуждается при определенной длине волны света, для которой обычно имеется волна эмиссии флуоресценции. Молекула-акцептор также возбуждается при этой длине волны так, что она может принимать энергию эмиссии от молекулы-донора путем различных механизмов передачи энергии, зависимыми от расстояния. Характерным примером возможной передачи энергии флуоресценции является резонансная передача энергии флуоресценции или "FRET". Обычно молекула-акцептор принимает энергию эмиссии от молекулы-донора, когда они находятся в тесной близости (например, одинаковая или соседняя молекула). Основой FET или FRET детектирования является мониторинг изменений при длинах волн эмиссии донора и акцептора.
Обычно применяют два типа FET или FRET зондов, в которых используется гидролиз зондов нуклеиновой кислоты для отделения донора от акцептора и в которых используется гибридизация для изменения пространственного взаимоположения молекул донора и акцептора.
Гидролизные зонды имеются в продаже под названием TaqManTM зондов. Они состоят из ДНК олигонуклеотидов, которые метят молекулами донора и акцептора. Эти зонды предназначаются для связывания со специфической областью в одной из цепей продукта ПЦР. После отжига праймеров ПЦР к этой цепи фермент Taq достраивает ДНК с помощью 5'-3'-полимеразы. Фермент Taq также проявляет активность 5'-3'-экзонуклеазы. Зонды TaqManTM защищаются в 3'-конце фосфорилированием для предотвращения от запуска наращивания под действием Taq. Если зонд TaqManTM гибридизируется с продуктом-цепью, тогда достраивающаяся молекула Taq также может гидролизовать зонд, высвобождая донор от акцептора, как основу для детектирования. Сигнал в этом случае является кумулятивным, поскольку концентрация молекул свободного донора или акцептора увеличивается с каждым циклом реакции амплификации.
Тот факт, что получение сигнала зависит от проявления реакции гидролиза зонда означает, что имеется временная сложность, связанная с этим методом. Кроме того, присутствие зонда может нарушать гладкое протекание ПЦР.
Кроме того, было установлено, что гидролиз может стать неспецифичным, особенно когда необходимы большие количества циклов амплификации, например более 50 циклов. В этих случаях неспецифический гидролиз зонда приведет к чрезмерно повышенному сигналу.
Это означает, что такие методы не очень совместимы со способами быстрой ПЦР, которые становятся все более известными с разработкой быстрых термоциклеров с использованием горячего воздуха, таких как RapidCyclerTM и LightCyclerTM от Idaho Technologies Inc. Описываются другие устройства для быстрой ПЦР, например, в совместно поданных заявках на патент Великобритании 9625442.0 и 9716052.7. О достоинствах быстрого циклирования по сравнению с обычным термическим циклированием сообщается в других источниках. Эти методы особенно полезны, например, в системах для детектирования биологического "оружия", где важна скорость результата, если необходимо избежать гибели или тяжелого повреждения.
Кроме того, гидролизные зонды не обеспечивают существенной информацией в отношении гистерезиса плавления, поскольку получение сигнала, в общем, зависит от гидролиза зонда в большей степени, чем от температуры плавления ампликона.
Гибридизационные зонды являются полезными в ряде случаев. Молекулярными сигналами являются олигонуклеотиды, которые имеют комплиментарные 3'-5'-последовательности, такие, что они образуют шпилькообразные петли. Концевые флуоресцентные метки находятся в тесной близости для проявления FRET, когда образуется шпилькообразная структура. После гибридизации молекулярных сигналов с комплиментарной последовательностью флуоресцентные метки отделяются так, что не возникает FRET, и это создает основу для детектирования.
Можно также использовать пары меченых олигонуклеотидов. Они гибридизируются в тесной близости по отношению к продукту-цепи ПЦР, соединяя вместе молекулы донора и акцептора так, что может возникать FRET. Усиленная FRET является основой для детектирования. Варианты этого типа включают использование меченого праймера амплификации с одним смежным зондом.
Использование двух зондов или зонда типа молекулярного сигнала, которое включает две меченые молекулы, увеличивает стоимость способа. Кроме того, для этого способа необходимо присутствие приемлемо длинной известной последовательности так, что два зонда, которые являются достаточно длинными для специфического связывания в тесной близости друг к другу, являются известными. Это может быть проблемой при некоторых диагностических применениях, где длина консервативных последовательностей в организме, которые можно использовать для конструирования эффективного зонда, такого как вирус HIV, может быть относительно короткой.
Кроме того, применение пары зондов включает более сложную экспериментальную схему. Например, сигнал, обеспечиваемый плавлением зонда, является функцией плавления обоих зондов. Постановка исследования при незначительном несоответствии или, когда одному из зондов необходимо связываться в области сплайса (например, для детектирования РНК по сравнению с ДНК в пробе, где последовательность на любой стороне интрона может использоваться в качестве сайта для зонда), может дать неправильные результаты, если плавится первым другой зонд.
Заявители разработали способ детектирования присутствия определенных последовательностей нуклеиновой кислоты, который можно адаптировать для определения количества мишеневой последовательности в пробе.
Таким образом, изобретение обеспечивает способ детектирования присутствия мишеневой последовательности нуклеиновой кислоты в пробе, указанный способ включает: а) постановку с указанной пробой реакции амплификации с использованием ряда нуклеотидов, по меньшей мере один из которых мечен флуоресцентной меткой; b) контактирование продукта амплификации с зондом в условиях, при которых зонд будет гибридизироваться с указанной мишеневой последовательностью, указанный зонд включает реакционноспособную молекулу, которая способна поглощать флуоресценцию от указанного, меченного флуоресцентной меткой нуклеотида или передавать ему энергию флуоресценсии и с) мониторинг флуоресценции указанной пробы.
Используя способ этого типа, флуоресцентная метка включается только в продукт амплификации. Когда зонд гибридизируется с любой специфичной мишеневой последовательностью, полученной в результате реакции амплификации, реакционноспособная молекула поглощает энергию эмиссии от меченых нуклеотидов или передает энергию меченым нуклеотидам с помощью FET или FRET, изменяя, таким образом, сигнал от флуоресцентных нуклеотидов. Соответственно реакционноспособная молекула способна поглощать флуоресценцию от меченых нуклеотидов и, таким образом, флуоресценция от них уменьшается. Это уменьшение может детектироваться, и это указывает на связывание зонда.
Наиболее предпочтительно, если реакционноспособная молекула является молекулой-акцептором, которая излучает флуоресценцию при определенной длине волны. В этом случае усиление флуоресценции от молекулы-акцептора, которая происходит при другой длине волны, чем для меченого нуклеотида, будет также указывать на связывание зонда.
Присутствие меченого таким образом продукта амплификации может детектироваться мониторингом флуоресценции от молекулы-акцептора на зонде, который специфически связывается только с мишеневой последовательностью. В этом случае сигнал от продукта амплификации может быть отличен от фонового сигнала флуоресцентной метки и также от неспецифического продукта амплификации.
Тот факт, что сигнал частично связан с продуктом амплификации и частично связан с зондом означает, что система высокоспецифична в отношении детектирования специфичных мишеневых последовательностей в реакционных смесях, которые содержат большие количества фоновой ДНК. Это является результатом того, что сигнал от неспецифического продукта амплификации может эффективно отделяться от измеряемого сигнала.
Способ этого типа можно выполнять, используя недорогие реактивы. Непарные меченые зонды являются более экономичными по сравнению с теми, которые включают как молекулы акцептора, так и донора.
Как здесь использовано, выражение "ряд нуклеотидов" относится к группе нуклеотидов, которые являются достаточными для построения нуклеиновых кислот, таких как ДНК и РНК. Таким образом, они включают аденозин, цитозин, гуанин и тимин или урацил. Один или более из них является меченным флуоресцентной меткой. Меченый урацил доступен от Boehringer Mannheim. Пригодные флуоресцентные метки включают флуоресцин.
Использование меченого урацила может быть особенно предпочтительным в том плане, что его применение создает стратегию для предотвращения загрязнения или для переноса из одной реакции амплификации в последующие, проводимые в реакционных сосудах. Ферменты, которые расщепляют содержащие урацил нуклеиновые кислоты, такие как урацил-N-гликозилат, можно использовать на стадии инкубации перед циклированием для обеспечения того, что любые остаточные ампликоны расщепляются до начала термического циклирования в последующем применении.
Соответственно более, чем один нуклеотид, и наиболее предпочтительно все нуклеотиды являются мечеными так, чтобы усреднять уровень сигнала от продукта амплификации и, таким образом, сигнала FET или FRET.
Амплификацию соответственно проводят, используя известные реакции амплификации, такие как полимеразная цепная реакция (ПЦР) или лигазная цепная реакция (ЛЦР), метод замены цепей (SDA) или NASBA, но предпочтительно это ПЦР.
Предпочтительно прослеживается флуоресценция как нуклеотида, так и молекулы-акцептора и рассчитывается взаимосвязь между эмиссиями.
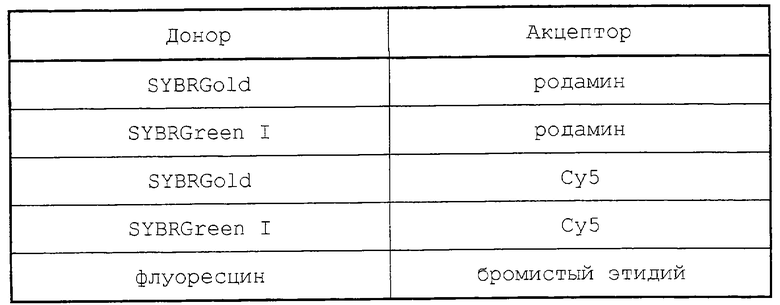
Пригодными реакционноспособными молекулами (такими как молекулы-акцепторы) являются родаминовые краски или другие краски, такие как Сy5. Они могут быть присоединены к зонду обычным способом. Положение реакционноспособной молекулы вдоль зонда является несущественным, хотя обычно они будут располагаться в концевой области зонда.
Для того чтобы имела место FET, как, например, FRET, между реакционноспособной молекулой и эмиссией флуоресценции нуклеотидов, эмиссия флуоресценции элемента (реакционноспособной молекулы или меченого нуклеотида), который действует в качестве донора, должна быть при более короткой длине волны, чем для элемента-акцептора.
Пригодные по этой причине комбинации расположены в таблице.
Предпочтительно, чтобы молекулы, используемые в качестве донора и/или акцептора, давали острые пики, и было бы небольшое перекрытие длин волн эмиссии или отсутствие его. В этих обстоятельствах может отсутствовать необходимость отделять "специфичный для цепи пик" от сигнала, образуемого продуктом амплификации. Простое определение только одного специфичного для цепи сигнала (т. е. обеспеченного реакционноспособной молекулой) будет предоставлять информацию в отношении величины FET или FRET, вызванной мишеневой реакцией. Это требование может выполнить комбинация бромистого этидия/флуоресцина. В этом случае специфичная для цепи реакция будет поддаваться количественному определению по уменьшению флуоресценции при 520 нм, соответственно выраженному как 1/флуоресценция.
Однако, где имеется спектральное перекрытие в сигналах флуоресценции от молекул донора и акцептора, это может учитываться при интерпретации результатов, например, путем эмпирического определения взаимосвязи между спектрами и использовании этой взаимосвязи для нормализации сигналов от двух сигналов.
Способ по изобретению является чрезвычайно разносторонним по его применимости. Этот способ можно использовать для получения как количественных, так и качественных данных, касающихся мишеневой последовательности нуклеиновой кислоты в пробе, как более детально обсуждается ниже. В частности, изобретение не только обеспечивает количественную амплификацию, но также оно может быть использовано дополнительно или альтернативно для получения характеризующих данных, таких как температуры дестабилизации дуплекса или точки плавления.
В способе по изобретению пробу можно подвергнуть воздействию условий, при которых зонд гибридизируется с пробами во время или после того, как реакция амплификации завершена. Способ позволяет осуществить детектирование гомогенным образом, при котором амплификацию и мониторинг можно проводить в одном контейнере со всеми реактивами, внесенными изначально. Нет необходимости в последующих стадиях добавления реактивов. Отсутствует какая-либо необходимость осуществлять способ в присутствии твердых подложек (хотя это возможно, как дополнительно обсуждается ниже).
Например, когда зонд присутствует в течение всей реакции амплификации, сигнал флуоресценции может позволить прослеживать протекание реакции амплификации. Это могут обеспечить средства для определения количества мишеневой последовательности, присутствующей в пробе.
Во время каждого цикла реакции амплификации цепи-ампликоны, содержащие мишеневую последовательность, связываются с зондом и, таким образом, образуют сигнал акцептора. Поскольку количество ампликона в пробе увеличивается, то будет расти и сигнал акцептора. При построении графика зависимости скорости увеличения по отношению к циклам может быть определена начальная точка увеличения.
Зонд может включать молекулу нуклеиновой кислоты, такую как ДНК или РНК, которая будет гибридизироваться с мишеневой последовательностью нуклеиновой кислоты, когда последняя является одноцепочной. В этом случае стадия b) будет включать использование условий, которые будут обеспечивать мишеневую одноцепочную нуклеиновую кислоту. Альтернативно зонд может включать такую молекулу, как пептид нуклеиновой кислоты, который специфически связывается с мишеневой последовательностью, находящейся в двухцепочной форме.
В частности, используемая реакция амплификации, такая как ПЦР или ЛЦР, будет включать стадию, когда проба подвергается условиям, при которых любая мишеневая последовательность нуклеиновой кислоты, присутствующая в пробе, становится одноцепочной.
Тогда для зонда будет возможным гибридизоваться во время протекания реакции амплификации, обеспеченной соответствующими условиями гибридизации.
В предпочтительном воплощении зонд можно сконструировать таким образом, что эти условия имеют место во время каждого цикла реакции амплификации. Таким образом, в некоторой точке во время каждого цикла реакции амплификации зонд будет гибридизироваться с мишеневой последовательностью и давать сигнал, как результат FET или FRET. По мере протекания амплификации зонд будет отделяться или плавиться из мишеневой последовательности и, таким образом, генерируемый реакционноспособной молекулой сигнал будет либо снижаться, либо повышаться в зависимости от того, является она молекулой донором или акцептором. Например, там, где она является акцептором, в каждом цикле амплификации образуется пик флуоресценции от реакционноспособной молекулы. Интенсивность пика будет увеличиваться по мере протекания амплификации, поскольку все больше мишеневой последовательности становится доступной для связывания с зондом.
Проводя мониторинг флуоресценции реакционноспособной молекулы из пробы во время каждого цикла, можно прослеживать протекание реакции амплификации различными путями. Например, можно анализировать данные, обеспеченные пиками плавления, например, рассчитав площадь под пиками плавления и построив график зависимости этих данных от числа циклов.
Мониторинг флуоресценции соответственно проводят с использованием известного флуорометра. Сигналы от него, например, в виде напряжений от фотоумножителя посылаются в процессор данных и преобразуются в спектр, соответствующий каждой пробирке с пробой. В одно и то же время можно оценивать множество пробирок, например 96 пробирок. Таким путем можно собрать данные при частых интервалах, например каждые 10 мс, в течение всей реакции.
Спектры, полученные таким образом, можно расшифровать, например, используя соответствие предварительно выбранных флуоресцентных частей, таких как краски, в отношении образования пиков, характерных для каждой сигнальной части (например, нуклеотида с меткой и/или реакционноспособной молекулы). Можно определить площади под пиками, которые представляют значение интенсивности для каждого сигнала и, если необходимо, выразить в виде частей друг от друга. Разница интенсивности сигнала и/или отношения позволят фиксировать изменения FET или FRET в течение всей реакции или при различных реакционных условиях, таких как температуры. Изменения, как отмечалось выше, относятся к связыванию зонда с мишеневой последовательностью. Общая площадь под дифференциальными пиками позволит вычислить значения интенсивности FET ИЛИ FRET эффектов.
Эти данные обеспечивают одно средство для оценки количества мишеневой нуклеиновой кислоты, присутствующей в пробе.
Зонд может быть свободным в растворе или быть иммобилизованным на твердой подложке, например на поверхности гранул, таких как магнитные шарики, пригодные для разделения продуктов, или на поверхности детекторного устройства, такого как волновод поверхностного плазменного резонансного детектора. Выбор будет зависеть от природы конкретного выбранного метода и от конкретного используемого средства детектирования.
Зонд может быть сконструирован таким образом, что он гидролизуется под действием ДНК-полимеразы, используемой в реакции амплификации, тем самым высвобождая молекулу-акцептор. Это обеспечивает кумулятивный сигнал, поскольку количество свободной реакционноспособной молекулы, присутствующей в системе, увеличивается с каждым циклом. Однако не является необходимым в этом способе, чтобы расход зонда осуществлялся таким образом, поскольку сигнал не зависит от гидролиза зонда.
Для того чтобы достичь полностью обратимого сигнала, который непосредственно относится к количеству продукта амплификации, присутствующему на каждой стадии реакции, и/или где скорость реакции имеет самое большое значение, например, при быстрой ПЦР, предпочтительно, чтобы зонд был сконструирован таким образом, что он высвобождался интактным из мишеневой последовательности и, таким образом, он может вновь принять участие в реакции. Это может быть, например, во время фазы наращивания в реакции амплификации. Однако, поскольку сигнал не зависит от гидролиза зонда, зонд может быть сконструирован таким образом, что он гибридизируется и плавится из мишеневой последовательности на любой стадии во время цикла амплификации, включая фазу отжига и плавления в реакции. Такие зонды будут обеспечивать сведение до минимума интерференции с реакцией амплификации.
Когда используются зонды, которые связываются во время фазы наращивания, их высвобождения в интактном виде можно достичь при использовании недостающего фермента 5'-3'-экзонуклеазы, такого как Stoffle-фрагмент Taq или Pwo. Это может быть полезным, когда является необходимой быстрая ПЦР, поскольку избегают стадий гидролиза.
Когда он используется таким образом, важно обеспечить, чтобы зонд не подвергался достройке во время стадии наращивания реакции. Поэтому 3'-конец зонда блокируется соответственно фосфорилированием.
Данные, полученные таким образом, можно интерпретировать различными путями. В самой простой форме увеличение флуоресценции молекулы акцептора во время или в конце реакции амплификации указывает на увеличение в количестве присутствующей мишеневой последовательности и предполагает тот факт, что реакция амплификация протекает и, следовательно, мишеневая последовательность действительно присутствует в пробе. Однако, как отмечалось выше, количественное определение также возможно при мониторинге реакции амплификации. Наконец, является возможным получить данные по характеристике и, в частности, анализ точки плавления либо в конечной точке определения, либо в течение всей реакции для того, чтобы получить информацию о последовательности, что будет обсуждаться дополнительно ниже.
Таким образом, предпочтительное воплощение изобретения включает способ детектирующий амплификации нуклеиновой кислоты, включающий: постановку амплификации нуклеиновой кислоты на мишеневом полинуклеотиде в присутствии: а) полимеразы нуклеиновой кислоты; b) по меньшей мере одного праймера, способного гибридизироваться с указанным мишеневым полинуклеотидом; с) ряда нуклеотидов, по меньшей мере один из которых мечен флуоресцентной меткой и d) олигонуклеотидного зонда, который способен связываться с указанной мишеневой полинуклеотидной последовательностью и который содержит реакционноспособную молекулу, которая способна поглощать флуоресценцию от указанного меченого нуклеотида или передавать ему флуоресценцию и мониторинг изменений в флуоресценции во время реакции амплификации. Соответственно реакционноспособная молекула является молекулой-акцептором, которая может поглощать энергию от меченого нуклеотида.
Амплификацию соответственно проводят, используя пару праймеров, которые сконструированы таким образом, что амплифицируется только мишеневая нуклеотидная последовательность внутри цепочки ДНК, как это хорошо понятно в этой области. Полимераза нуклеиновой кислоты является соответственно термостабильной полимеразой, такой как Taq-полимераза.
Приемлемые условия, при которых может проводиться реакция амплификации, являются хорошо известными в этой области. Оптимальные условия могут быть различными в каждом случае в зависимости от вовлеченного конкретного ампликона, природы используемых праймеров и применяемых ферментов. Оптимальные условия могут быть определены в каждом случае специалистом. Типичные температуры для денатурации составляют порядка 95oС, типичные температуры отжига - порядка 55oС и температуры наращивания - порядка 72oС.
В частном случае изобретения зонд можно использовать для количественного определения транскриптов РНК, например в опытах по экспресии, что можно использовать при разработке лекарственных препаратов. В частности, это воплощение пригодно для опытов по экспрессии в тканях эукариотических организмов. ДНК-коды белков в клетках-эукариотах могут включать интроны, не кодирующие области последовательности ДНК, и экзоны, которые кодируют последовательность белков. Некодирующие последовательности интронов удаляются из РНК-последовательностей, которые получаются из ДНК-последовательностей, во время клеточного сплайсинга. Праймеры ПЦР обычно являются мишеневыми для кодирующих областей, и когда используется ПЦР с обратной транскриптазой в общих экстрактах нуклеиновых кислот, продукты будут результатом как ДНК-зависимой амплификации, так и РНК-зависимой амплификации. Таким образом, при использовании в опытах по экспрессии одна ПЦР будет включать амплификацию, полученную от геномной ДНК и экспрессированной РНК.
Зонд, который сконструирован для связывания через интроны, в смежных концевых областях кодирующих экзонов, будет иметь место ограниченное взаимодействие за счет области интрона. У сплайсированной РНК эти области удалены и, следовательно, смежные концевые области кодирующих экзонов образуют непрерывную последовательность, что позволяет эффективно связывать зонд.
Напротив, зонд может детектировать только продукт амплификации геномной ДНК, если он так сконструирован, что связывается с областью интрона. Сигнал, образуемый таким зондом, будет относиться только к концентрации ДНК и не к концентрации РНК в пробе.
Таким образом, в дополнительном воплощении зонд является специфичным либо для сплайс-области РНК или интрона ДНК так, что детектируется и/или количественно определяется только одна из амплифицированной РНК или амплифицированной ДНК.
Альтернативно или дополнительно способ по изобретению можно использовать в методах гибридизации для определения свойств последовательности. Таким образом, в дополнительном аспекте изобретение обеспечивает способ определения свойства последовательности, где указанный способ включает: а) амплификацию указанной последовательности с использованием ряда нуклеотидов, по меньшей мере один из которых мечен флуоресцентной меткой; b) контактирование продукта амплификации с зондом в условиях, при которых зонд будет гибридизироваться с указанной мишеневой последовательностью, указанный зонд включает реакционноспособную молекулу, которая способна поглощать флуоресценцию от указанного меченого флуоресцентной меткой нуклеотида или передавать энергию флуоресценсии и с) мониторинг флуоресценции указанной пробы и определение конкретного условия реакции, свойства указанной последовательности, при которых изменяется флуоресценция в результате гибридизации зонда с пробой или дестабилизации дуплекса, образованного между зондом и мишеневой последовательностью нуклеиновой кислоты.
Приемлемые условия реакции включают температуру, электрохимию или ответную реакцию на присутствие определенных ферментов или химических веществ. Прослеживая изменения в флуоресценции, по мере изменения этих свойств, можно получить информацию, характеризующую определенную природу последовательности. Например, в случае температуры можно определить температуру, при которой зонд отделяется от последовательностей в пробе в результате нагревания. Это может быть очень полезным, например, для детектирования и, если желательно, также для количественной оценки полиморфизмов и/или аллельной вариации при генетическом диагнозе. "Полиморфизм" включает транзиции, трансверсии, инсерции, делеции инверсий, которые могут иметь место в последовательности, особенно в природе.
Гистерезис плавления будет различным, если мишеневая последовательность различается только по одной паре оснований. Таким образом, например, где проба содержит только один аллельный вариант, температура плавления зонда будет иметь особую ценность, которая будет отличной от установленной для пробы, которая содержит только другой аллельный вариант. Проба, содержащая оба аллельных варианта, показывает две точки плавления, соответствующие каждому из аллельных вариантов. Аналогичные соображения можно применить в отношении электрохимических свойств или присутствия некоторых ферментов или химических веществ. Зонд может быть иммобилизован на твердой поверхности через которую может быть приложен электрохимический потенциал. Мишеневая последовательность будет связываться с или отталкиваться от зонда при определенных электрохимических значениях в зависимости от определенной природы последовательности.
Кроме того, кинетика гибридизации зонда позволит определить в абсолютных значениях концентрацию мишеневой последовательности. Изменения флуоресценции пробы могут позволить рассчитать скорость гибридизации зонда с пробой. Увеличение скорости гибридизации будет соответствовать количеству мишеневой последовательности, присутствующей в пробе. Поскольку концентрация мишеневой последовательности увеличивается по мере протекания реакции амплификации, гибридизация зонда будет происходить быстрее. Таким образом, этот параметр также можно использовать как основу для количественного определения. Этот способ обработки данных полезен тем, что он не зависит от интенсивности сигнала для обеспечения информацией.
Дополнительные аспекты по изобретению включают наборы для использования в способе по изобретению. Эти наборы будут включать зонд, специфичный для мишеневой нуклеиновой последовательности, который содержит реакционноспособную молекулу, в частности молекулу акцептора. Если желательно, зонд можно иммобилизовать на подложке, такой как гранулы, например магнитные шарики, или подложке, используемой в детекторе, такой как волновод исчезающей волны детекторного устройства.
Кроме того, наборы могут включать один или более меченый флуоресцентной меткой нуклеотид, который/которые совместимы с указанной реакционноспособной молекулой. Другие потенциальные компоненты набора включают реактивы, используемые в реакциях амплификации, такие как ДНК-полимераза.
Чертеж схематично показывает взаимодействия, которые имеют место в способе по изобретению.
В иллюстрированной реакции амплификации молекула ДНК (1), приготовленная для амплификации путем ее контактирования с парой амплификационных праймеров (2), (3) и рядом нуклеотидов (4), некоторые из которых мечены флуоресцирующей меткой (5). Молекула ДНК (1) является одноцепочной (фиг. 1В), посредством чего праймеры (2, 3) связываются как прямые и обратные праймеры в реакции амплификации, как это хорошо известно.
Во время протекания последующей реакции амплификации выстраивается продукт-ампликон (6) (фиг. 1С). Нуклеотиды, меченные и немеченные, включаются в продукт по мере его образования. Обычно продукт-ампликон (6) содержит только мишеневую последовательность, как определено с помощью праймеров.
Когда этот продукт плавится во время последующей фазы амплификации, зонд (7), включающий молекулу-акцептор (8), связывается с мишеневой последовательностью (фиг. ID). Взаимодействие FRET между флуоресцентными нуклеотидами и молекулой-акцептором (8) дает сигнал при длине волны, специфичной для акцептора.
Сигнал акцептора (8) можно затем записать, используя обычные устройства для детектирования флуоресценции.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕАКТОРЫ | 1997 |
|
RU2177830C2 |
| МЕЧЕННОЕ ЛЮЦИФЕРАЗОЙ АНТИТЕЛО И СПОСОБ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ОСУЩЕСТВЛЕНИЯ АНАЛИЗА НА СПЕЦИФИЧЕСКОЕ СВЯЗЫВАНИЕ И НАБОР ДЛЯ ПРИМЕНЕНИЯ В АНАЛИЗЕ НА СПЕЦИФИЧЕСКОЕ СВЯЗЫВАНИЕ | 1995 |
|
RU2199125C2 |
| АНАЛИЗ РЕСПИРАТОРНОЙ ИНФЕКЦИИ | 2012 |
|
RU2642304C2 |
| ПРОТИВОЧУМНАЯ ВАКЦИНА | 1996 |
|
RU2197988C2 |
| СПОСОБЫ И ПРИБОРЫ ДЛЯ ОБНАРУЖЕНИЯ И ИДЕНТИФИКАЦИИ ЗАКОДИРОВАННЫХ ГРАНУЛ И БИОЛОГИЧЕСКИХ МОЛЕКУЛ | 2007 |
|
RU2487169C2 |
| ЭКСПРЕСС-АНАЛИЗЫ АКТИВАЦИИ Т-КЛЕТОК, ОСНОВАННЫЕ НА ИЗМЕРЕНИИ РНК С ИСПОЛЬЗОВАНИЕМ ПРОТОЧНОЙ ЦИТОМЕТРИИ | 2013 |
|
RU2671707C2 |
| РАСТЕНИЕ МАИСА DBN9936 И СПОСОБ ПРИМЕНЕНИЯ В ДЕТЕКТИРОВАНИИ ЕГО ПОСЛЕДОВАТЕЛЬНОСТИ НУКЛЕИНОВОЙ КИСЛОТЫ | 2016 |
|
RU2707527C2 |
| СПОСОБЫ, КОМПОЗИЦИИ И НАБОРЫ ДЛЯ ОПРЕДЕЛЕНИЯ ВИРУСА ГЕПАТИТА А | 2012 |
|
RU2595822C2 |
| СПОСОБЫ ПРОГНОЗИРОВАНИЯ РИСКА ИНТЕРСТИЦИАЛЬНОЙ ПНЕВМОНИИ | 2014 |
|
RU2670148C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ЗИГОТНОСТИ ГЕНА FAD3 В КАНОЛЕ | 2012 |
|
RU2630998C2 |
Изобретение относится к способам детектирования присутствия мишеневой последовательности НК, детектирования амплификации НК, определения свойств последовательности НК, детектирования полиморфизма и/или аллельной вариации, а также раскрывает набор для осуществления вышеуказанных способов. Способы предусматривают постановку реакции амплификации с использованием нуклеотидов, которые мечены флуоресцентной меткой, контактирование продукта амплификации с зондом, который включает реакционноспособную молекулу, которая способна поглощать флуоресценцию от меченого нуклеотида, мониторинг флуоресценции. Набор для способов включает в себя зонд, содержащий реакционноспособную молекулу, меченные флуоресцентной меткой нуклеотиды и ДНК-полимеразу. Данные способы позволяют быстро детектировать в пробе присутствие и свойства НК. 5 с. и 19 з.п.ф-лы, 1 ил., 1 табл.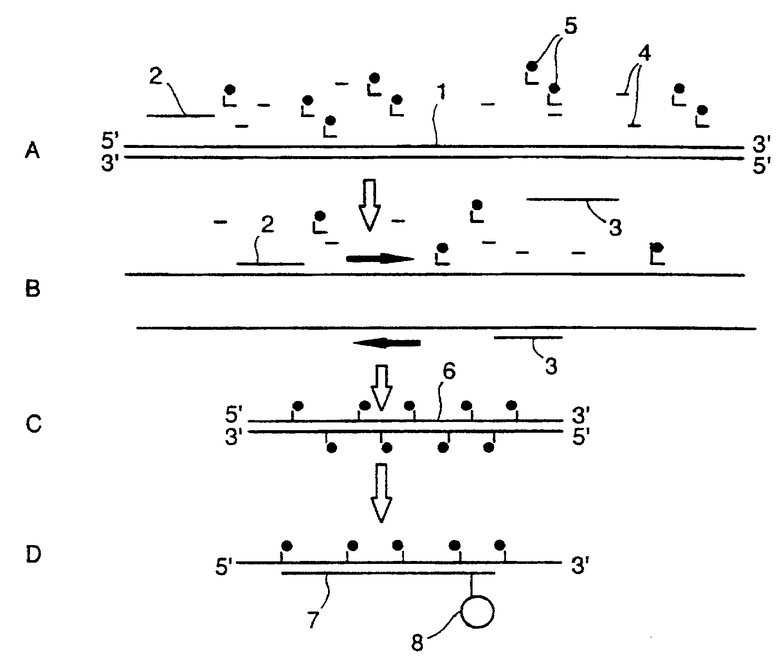
| Воздушно-гидравлическая установка | 1978 |
|
SU794261A1 |
| US 5804375, 08.09.1998 | |||
| US 5861256, 19.01.1999 | |||
| US 5716784, 10.02.1998. | |||

