Ссылка на родственные заявки
По настоящей заявке испрашивается приоритет предварительной заявки на патент США 61/665231, поданной 27 июня 2012 года, и предварительной заявки на патент США 61/784802, поданной 14 марта 2013 года. Содержание заявок включено в настоящее описание в качестве ссылки в полном объеме.
Интересы правительства
Изобретение, раскрытое в настоящей заявке, было осуществлено по меньшей мере частично при поддержке правительства грантами Национальных институтов здравоохранения под номерами AI045761, AI106036, MH079197 и HL106788. Правительство США имеет определенные права на настоящее изобретение.
Область техники, к которой относится изобретение
Настоящее изобретение относится к способам быстрой идентификации экспрессии генов в отдельных клетках.
Уровень техники
Многие патологические состояния провоцируют опосредованные T-клетками иммунные ответы. К таким состояниям относятся злокачественные новообразования, аутоиммунные заболевания, реакции при трансплантации и инфекционные заболевания, включающие, например, туберкулез (TB). При всех этих состояниях активация T-клеток включает взаимодействие T-клеточного рецептора (TCR) со специфичным антигеном (Ag), выступающим в качестве общего фактора, тогда как функциональные последствия такого взаимодействия различны. В той или иной T-клетке сигнальный каскад TCR может стимулировать экспрессию различных наборов генов в зависимости от физической природы Ag, его презентации, репертуара TCR и силы Ag-индуцированной активации. Эти индуцированные различия профилей экспрессии генов в процессе активации, пролиферации и дифференцировки клеток могут, в конечном счете, способствовать проявлению функциональной гетерогенности клонально размноженных эффекторных клеток. Дополнительный элемент клеточного разнообразия возникает в процессе гомеостаза T-клеток. Если Ag установлен, то наиболее размноженные эффекторные клетки погибают вследствие апоптоза, тогда как небольшая фракция клеток выживает в качестве долгоживущих клеток памяти. Ответы, которые возникают при повторном контакте с Ag, регулируются вышеперечисленными факторами, причем они также находятся под влиянием предыдущего процесса дифференцировки. В результате на каждый Ag, с которым встречается организм, иммунный ответ производит функционально различные субпопуляции T-клеток (например, эффекторные клетки - центральные клетки памяти - эффекторные клетки памяти), обладающие общей специфичностью к Ag. В тот или иной момент развития иммунопатологического состояния изменяется относительная репрезентация этих различных субпопуляций; они могут определять или отражать патологический процесс. Субпопуляции T-клеток коррелируют с Ag и патогенной нагрузкой in vivo и, как было показано в исследованиях хронических вирусных инфекций и TB, количественный анализ этих ключевых субпопуляций перспективен для диагностики и оценок защитного иммунитета. Контрольные изменения в субпопуляциях T-клеток также следует отмечать в ходе лечения, поскольку результат лечения связан со специфическим развитием иммунопатологического состояния. Такие характерные показатели прогрессирующих состояний (природных или вызванных лечением) не поддаются измерению в совокупности и могут быть обнаружены только на уровне одиночных T-клеток. Свойства Т-клеток человека полностью проявляются только при их ответе на стимуляцию Ag.
Современные клинические иммунологические анализы крови обычно позволяют распознавать присутствие неинфекционной и инфекционной патологии, но могут не обнаруживать прогрессию патологии. С другой стороны, на уровне одиночных клеток функциональный анализ Ag-специфичных иммунных клеток в периферической крови должен раскрывать множественные, пересекающиеся иммунопатологические состояния, связанные с развитием инфекции и болезни. Такой анализ может повысить прогностическую значимость и способствовать эффективному вмешательству. Убедительным примером является инфекция, вызываемая Mycobacterium tuberculosis. Современные методы позволяют обычно диагностировать TB после того, как больной уже передал инфекцию контактировавшим с ним лицам. По существу диагноз бессимптомной инфекции не требует медицинского вмешательства. Возможность отличать стабильную латентную инфекцию (LTBI) от ранней активной болезни (до того как она становится контагиозной) крайне необходима. Существующие иммунодиагностические анализы не способны удовлетворить эту потребность, поскольку они не позволяют различать Ag-специфичные субпопуляции функциональных T-клеток, которые, как известно, отличаются в зависимости от состояния инфекции. Кроме того, такие анализы не позволяют оценить ответы, которые могут отражать стадиеспецифическую экспрессию микобактериальных Ag.
Характерной особенностью инфекции, вызываемой M.tuberculosis, является огромное количество людей, бессимптомно инфицированных этим патогеном (2 миллиарда). В отсутствие иммунной недостаточности, у 90-95% латентно инфицированных лиц активная форма TB не будет развиваться. Однако, учитывая объем резервуара инфекции, M.tuberculosis все еще вызывает 9,4 миллиона новых случаев активного TB и 1,7 миллиона смертельных случаев ежегодно. Перенос инфекции можно было бы значительно снизить, если бы было возможно выявлять и лечить инфицированных лиц по мере прогрессирования у них болезни с ее переходом в активную форму, до проявления у них симптомов и до того, как они становятся заразными. Обнаружение туберкулезной палочки или бациллярных продуктов чрезвычайно затруднено на ранней стадии болезни из-за крайне низкого числа бацилл. Стандартный микробиологический диагноз активного TB выявляет лиц, которые уже являются носителями туберкулезной палочки в секрете дыхательных органов и поэтому являются заразными.
Методы, основанные на множественном измерении, не подходят для выявления сложного клеточно-опосредованного иммунного ответа и не могут обнаруживать редкие или переходные состояния клеток, связанные со стадией и развитием заболевания. Наиболее распространенными форматами иммунологических тестов для анализа одиночных T-клеток являются метод иммуноферментного спот-анализа (ELISPOT) и количественный проточный цитометрический (FC) анализ. В коммерческом тесте ELISPOT производства Oxford Immunotech (United Kingdom) мононуклеарные клетки периферической крови (PBMC) сначала выделяют из пробы крови, промывают и подсчитывают. Затем в лунки планшета, предварительно покрытые антителами к интерферону-гамма (IFNγ), добавляют установленное количество (например, 250000) PBMC и M.tuberculosis-специфичных Ag и оставляют для инкубирования на ночь (16-20 часов). IFNγ, высвобождаемый из активированных T-клеток, захватывается в лунках. После этого следует инкубирование со вторым антителом (Ab), которое конъюгировано с цветообразующим ферментом, после чего лунки промывают и добавляют цветообразующий субстрат. Окрашивание проявляется в тех лунках, где IFNγ секретировали T-клетки, что приводит к появлению характерного цветного кольца (спота или пятна), окружающего цитокин-секретирующую клетку. Наконец пятна подсчитывают либо невооруженным глазом, либо используют сканирующий спектрофотометр для планшетов. ELISPOT очень чувствителен (предел его чувствительности составляет 10/106 в PBMC). Однако он не способен обеспечить желаемое определение различия в субпопуляции функциональных T-клеток по нескольким причинам. Во-первых, он обычно не позволяет вести одновременную детекцию нескольких мишеней. Во-вторых, формат ограничивает число клеток для анализа (например, 250000 клеток в лунке в коммерческом тесте T-SPOT.TB, указанном выше). Стандартная FC обладает многопараметрической функциональностью. Клетки в цельной крови или PBMC стимулируют путем инкубирования в течение 6 часов или больше с определенными Ag, например, M.tuberculosis-специфическими пептидами или белками, и ингибитором внеклеточной секреции, таким как брефелдин или монензин. Пермеабилизованные T-клетки затем окрашивают, используя меченное флуорофором Ab к цитокину (например, FITC-конъюгированное антитело против IFNγ), и анализируют с помощью FC. Окрашивание белков антителом уменьшает универсальность обычной FC, особенно в случае внутриклеточных белков. Кроме того, анализы ограничены методиками, которые требуют длительной стимуляции T-клеток эндогенными антигенпрезентирующими клетками (APC), что увеличивает длительность цикла и уменьшает клиническую полезность.
Существующие способы анализа цитокинов отдельных клеток не позволяют быстро идентифицировать редко встречающиеся популяции T-клеток, присутствующие в малых образцах периферической крови. Коммерческий тест ELISPOT не различает TB и LTBI. Кроме того, положительный результат у бессимптомного лица (в отсутствие микробиологического подтверждения активного TB) неинформативен в отношении риска повторной активации. Таким образом, несмотря на точное отражение присутствия инфекции M.tuberculosis, положительный результат не считается показанием для терапевтического вмешательства. Это уменьшает пригодность теста, особенно в бедных странах с высоким уровнем распространения туберкулеза, где заражено большинство населения (до 80% в областях Южной Африки). Кроме того, даже в странах с низким уровнем распространения и высокими доходами, таких как США, побочные эффекты лечения LTBI часто приводят к отрицательному результату, когда лечение рекомендуют без указания риска повторной активации. Несомненно, ограничения существующих иммунодиагностических методов имеют огромные последствия для здравоохранения в мировом масштабе.
Таким образом, существует неудовлетворенная потребность в способах, которые позволяют быстро детектировать экспрессию генов в отдельных клетках.
Сущность изобретения
Настоящее изобретение относится к новым быстрым способам детекции различных молекул РНК в отдельных клетках.
Таким образом, в одном из аспектов настоящее изобретение относится к способу детекции копий по меньшей мере одной молекулы РНК, экспрессируемой в отдельных клетках. Способ включает: (a) предоставление образца, содержащего популяцию клеток (например, по меньшей мере 100 или по меньшей мере 10000 клеток), которые экспрессируют специфические рецепторы, способные инициировать сигнальные каскады, вызывающие экспрессию генов, при связывании с ними лигандов или стимулирующих молекул; (b) индукцию экспрессии генов в клетках ex vivo либо сразу, либо после культивирования при инкубировании клеток по меньшей мере с одним соединением, например, пептидом или смесью пептидов, полученных из микроорганизма (такого как M.tuberculosis), которое связывается со специфическими рецепторами или иным образом инициирует сигнализацию и инициирует экспрессию генов; (c) фиксацию и пермеабилизацию клеток; (d) мечение копий по меньшей мере одной молекулы РНК, экспрессируемой клетками, с помощью набора флуоресцентно меченных олигонуклеотидных зондов гибридизации и удаление не связавшихся зондов при промывке; и (e) детекцию клеток с событиями экспрессии, с помощью проточной цитометрии (FC), где событие экспрессии является одним или более измерениями флуоресценции, классифицируемыми путем установки диапазона (гейтирования) интенсивности флуоресценции.
Вышеуказанный способ может использоваться для обнаружения различных молекул РНК в отдельных клетках. В частности, он может использоваться для обнаружения молекул РНК, кодируемых генами цитокинов, таких как IL-2, TNFα или IFNγ.
В способе стадия индуцирования может включать стимуляцию сигнализации T-клеточных рецепторов (TCR) и последующую экспрессию генов антигенпрезентирующими клетками (APC), которые присутствуют в популяции клеток, или искусственными APC (aAPC). Индукцию можно проводить в течение периода от 30 минут до 6 часов. В любом случае, этап индуцирования также может включать костимуляцию по меньшей мере с одним моноклональным антителом (mAb). Моноклональное антитело может быть связано с или добавлено к aAPC или APC.
В одном из вариантов осуществления способа популяция клеток может быть популяцией T-клеток, полученных из PBMC человека. В этом случае по меньшей мере одна молекула РНК может быть молекулой РНК, кодируемой геном цитокина. При этом клетки в популяции клеток могут быть индуцированы APC или aAPC с одной или более нагруженными пептидами молекулами MHC, которые взаимодействуют с TCR.
В способе соединение или стимулирующие молекулы могут быть получены из микроорганизма, например, полученными из M.tuberculosis иммуногенными пептидами. В одном из примеров соединение может содержать пептид или смесь пептидов, например, таких, которые связаны с молекулами MHC класса I или MHC класса II. Эти пептиды или смесь пептидов может быть добавлена к APC или aAPC, указанным выше, в различных концентрациях, например, в пределах от 1 до 20 мкг/мл.
В вышеописанном способе стадия индуцирования может включать стимуляцию сигнализации toll-подобных рецепторов и последующей экспрессии генов в клетке, содержащей такие рецепторы, при воздействии стимула, примеры которого включают цитокины, микробные продукты или синтетические соединения. Примеры микробного продукта включают липид, гликан, гликолипид, сульфолипид, гликопротеин, белок, пептид или нуклеиновую кислоту (например, РНК или ДНК). Стимуляцию можно проводить в течение периода от 30 минут до 72 часов (например, от 30 минут до 6 часов, от 6 часов до 24 часов, от 24 часов до 72 часов). Стимул может присутствовать в концентрации в пределах от 1 до 20 мкг/мл (например, от 1 до 100 нг/мл или от 100 до 1000 нг/мл). Соединение или стимул может являться цитокином, таким как IFNγ, IL-2, IL-15, TNFα или другим цитокином кроме IFNγ, IL-2, IL-15 или TNFα. В способе более одного соединения или стимула можно предоставить в одно и то же время или в разное время.
На стадии (d) способа набор зондов может включать от 20 до 60 зондов, каждый из которых отдельно помечен одной и той же флуоресцентной молекулой.
В другом варианте осуществления детектированные клетки, имеющие события экспрессии, могут быть отделены (например, путем сортировки клетки с активированной флуоресценцией) от клеток, не имеющих события экспрессии. Отделенные клетки могут включать только одну клетку или больше одной клетки (например, 10, 100, 1000, 10000 или 100000 клеток). Экспрессию генов в отделенных клетках можно затем измерять, например, с помощью ОТ-ПЦР или транскриптомного анализа РНК в клетках.
Подробное описание одного или более вариантов осуществления изобретения представлены в описании ниже. Другие признаки, объекты и преимущества изобретения будут очевидны из описания и из формулы изобретения.
Краткое описание фигур
Фиг. 1 - график интенсивности флуоресценции против канала прямого рассеяния A (FSC-A) для показаний FC, описанных в примере 2.
Фиг. 2 - набор из четырех частотных графиков, полученных при FC детекции образцов, описанных в пример 7.
Фиг. 3 - набор из четырех частотных графиков, полученных при FC детекции образцов, описанных в примере 8.
Фиг. 4 - набор из трех частотных графиков, полученных при FC детекции образцов, описанных в примере 9.
Фиг. 5 - набор из двенадцати частотных графиков, полученных при FC детекции образцов, описанных в примере 10.
Фиг. 6A и 6B - (A) набор из двадцати восьми частотных графиков, полученных в результате FC детекции образцов, исследованных с использованием наборов мРНК зондов, как описано в примере 11, и (B) набор из шестнадцати частотных графиков, полученных при FC детекции образцов, исследованных с использованием mAb, как описано в примере 11.
Фиг. 7A и 7B - (A) набор из двух частотных графиков, полученных в результате FC детекции образцов, исследованных с использованием зонда к зеленому флуоресцентному белку до (левая панель) и после (правая панель) дополнительного гейтирования для разделения клеток на цитокин-положительные и цитокин-отрицательные популяции, как описано в примере 12, и (B) набор из двух частотных графиков, полученных в результате FC детекции образцов, исследованных одновременно с использованием четырех наборов мРНК зондов, до (левая панель) и после (правая панель) дополнительного гейтирования для разделения клеток на цитокин-положительные и цитокин-отрицательные популяции, как описано в примере 12.
Фиг. 8A и 8B - (A) таблица, в которой показаны пороговые циклы амплификации двух генов в сортированных (+/-) популяциях клеток, описанных в примере 12, и (B) фотография, на которой показан электрофоретический гель продуктов амплификации, описанных в примере 12.
Подробное описание изобретения
Настоящее изобретение относится к способу измерения и оценки экспрессии одного или более генов, включая РНК транскрипты, которые кодируют белки, например, матричную РНК (мРНК) и пре-мРНК, и которые не кодируют белки.
Как раскрыто в настоящем описании, способ может использоваться для оценки РНК транскриптов в клетках, экспрессия генов в которых изменяется в ответ на различные индукции. Индукция может происходить естественно или ее могут стимулировать любые известные лиганды клеточных рецепторов, в том числе, без ограничения, TCR, и включать, в принципе, любой рецептор или лиганд (например, toll-подобные рецепторы или рецепторы хемокинов и их лиганды) или соединение, которое инициирует сигнальный каскад, и может приводить к синтезу транскриптов и экспрессии некоторых генов, например, по меньшей мере одного гена цитокина, такого как IL-2, TNFα или IFNγ.
В некоторых вариантах осуществления индукция представляет собой стимуляцию соединением или соединениями, которые связывают специфические рецепторы и таким образом инициируют экспрессию генов, например, пептидом или смесью пептидов, связанных с молекулами MHC I или MHC II. Соединение или соединения могут быть получены из микроорганизма, например, стимулирующие молекулы, которые являются полученными из М. tuberculosis иммуногенными пептидами. Индукцию экспрессии генов, например, стимуляцию сигнализации T-клеточных рецепторов и последующей экспрессии генов могут производить APC, которые присутствуют в исследуемой популяции клеток. В альтернативе стимуляцию могут производить искусственные APC (aAPC). Пептид или смесь пептидов, которые связываются с молекулами MHC класса I или класса II, могут быть добавлены к APC или aAPC, предпочтительно в концентрации в пределах от 1 до 20 мкг/мл, с получением нагруженных пептидами молекул MHC, которые взаимодействуют с TCR. Некоторые варианты осуществления способов настоящего изобретения включают костимуляцию по меньшей мере с одним моноклональным антителом в дополнение к APC или aAPC. В случае aAPC, с aAPC могут быть связаны mAb.
Способ настоящего изобретения позволяет выполнять быстрые и чувствительные измерения изменений состава и функции популяции клеток, которые происходят в процессе инфекционного или неинфекционного заболевания и ассоциированной прогрессии патологии. Такие измерения могут описывать свойства неопластических и/или злокачественных клеток гематопоэтического и негематопоэтического происхождения, а также клеток, вовлеченных в аутоиммунное заболевание.
Способ может также использоваться в мониторинге ответа в ходе терапии для лечения болезни. В случае инфекционной болезни иммунный контроль в ответ на антибиотикотерапию может быть определен при сопоставлении прогресса в устранении патогена, например M.tuberculosis, и изменений клеточных ответов на стимуляцию, таких как ответы субпопуляций функциональных T-клеток против антигенных пептидов.
Способы настоящего изобретения в целом могут быть использованы фактически для всех клеток млекопитающих (включая, без ограничения, человека) и низших эукариотов, которые экспрессируют специфические рецепторы, способные инициировать сигнальные каскады при связывании с лигандом/стимулирующей молекулой. Они могут использоваться для популяций бактерий, а также многоклеточных организмов. Некоторые предпочтительные варианты осуществления относятся к применению способов настоящего изобретения для клеток животных, в том числе, без ограничения, для клеток человека. Подходящий источник обеспечивает достаточное количество клеток для обнаружения редких событий и/или для статистического анализа. В некоторых вариантах осуществления это количество составляет всего лишь 1000 или даже 100 клеток. Во многих вариантах осуществления, например, когда клетки являются T-клетками, предпочтителен источник, который обеспечивает по меньшей мере 10000 клеток. Более предпочтительно источник обеспечивает по меньшей мере 100000 клеток или по меньшей мере один миллион клеток. Подходящие источники включают образцы крови и образцы ткани, например, биоптат. Образцы ткани необходимо разделить на индивидуальные клетки, то есть для обработки требуется дезагрегированная ткань.
Как указано выше, способы настоящего изобретения включают быструю индукцию в течение не более 5 дней и предпочтительно меньше, например 4, 3, 2 или 1 день, или 16, 12, 10, 8, 6 или 4 часа. Предпочтительное время индукции составляет от 30 минут до 8 часов, предпочтительно от 30 минут до 6 часов, более предпочтительно от 30 минут до 4 часов и еще более предпочтительно от 30 минут до 2 часов.
Способы настоящего изобретения включают быстрое измерение экспрессии одного или более генов в иммунных и неиммунных клетках, основанное на активации и функциональных последствиях, вызванных потоками ионов, включая, без ограничения, потоки ионов, вызванные ионофорами и/или митогенами, или сигнализацией посредством клеточных поверхностных или внутриклеточных рецепторов, включая, без ограничения, рецепторы распознавания паттерна, включая toll-подобные рецепторы, NOD и NOD-подобные рецепторы, G-белок-сопряженные рецепторы, включая рецепторы хемокинов, рецепторы полипептидных гормонов, рецепторы цитокинов, B-клеточные рецепторы или TCR. Способы по настоящему изобретению, как предполагают, способны улучшить медицинское обслуживание при различных инфекционных и неинфекционных патологиях. Предпочтительные варианты осуществления включают обнаружение экспрессии РНК, включая мРНК и пре-мРНК, одного или более цитокинов в индивидуальных лимфоцитах, будь то выделенные PBMC, T-клетки или пробы цельной крови (а не выделенные PBMC), а также в других типах клеток, как указано ниже. Способы по настоящему изобретению применяются, например, для анализа CD3+CD4+ T-клеток (T-хэлперов 1 или Th1-клеток), которые продуцируют IL-2, IFNγ и TNFα; CD3+CD4+ T-клеток (T-хэлперов 2 или Th2-клеток), которые продуцируют IL-4, IL-5, IL-6, IL-10 и IL-13; CD3+CD8+ T-клеток, которые продуцируют (IL-2, IFNγ, TNFα, MIP-1a и другие хемокины), и субпопуляций других специализированных T-клеток, включая, без ограничения, клетки Th17 и Treg, или субпопуляций других лимфоцитов, включая, без ограничения, NK-клетки, NKT-клетки, субпопуляции макрофагов и дендритных клеток, клеток, формирующих эндотелий и эпителий различных органов тела, или клеток нервной системы, включая, без ограничения, нейроны и глию, а также пре-мРНК и мРНК цитокинов, хемокинов и функциональных молекул (например, смотрите таблицу 2), вырабатываемых этими клетками.
Некоторые способы по настоящему изобретению включают ex vivo стимуляцию клеток или индукцию специфичных молекулярных взаимодействий, приводящих к экспрессии маркеров активации и модулированию клеточных функций. Стимуляция может быть выполнена на клетках либо сразу, либо после культивирования в культуральной среде. Стимуляция может быть естественной, то есть являться простым инкубированием клеток в культуре. Предпочтительно стимуляции может способствовать инкубирование клеток в культуре с одним или более соединениями, которые активируют специфичные рецепторы/лиганды и индуцируют сигнальные каскады. Эти соединения могут быть синтетическими или полученными из микроорганизмов, включающих, без ограничения, бактерии, вирусы и грибы, например, липополисахариды, липоарабиноманнаны, пептидогликаны, миколевые кислоты бактериального происхождения или белки вирусного происхождения, включая, без ограничения, вирус Эпштейна-Барра или цитомегаловирус (например, белки ebvIL-10, cmvIL-10 и UL146, соответственно), или грибковые (например, Cryptococcus- и Aspergillus-ассоциированные) галактоксило- и галактоманнаны и растворимые антигены; полипептидные гормоны, например, тромбоцитарный фактор роста или VEGF; цитокины; и антигены, включая антигенные пептиды. В предпочтительном способе стимуляции используют искусственные антигенпрезентирующие клетки (aAPC; смотрите ниже).
Способы по настоящему изобретению также включают детекцию отдельных клеток, включая, в частности, активированные клетки, имеющие биомаркерный набор характеристик, предпочтительно набор характеристик из множества биомаркеров, а именно одной или более РНК, включая, в частности, молекулы мРНК или пре-мРНК in situ в интактных клетках.
Предпочтительные способы по настоящему изобретению включают обнаружение индуцированной экспрессии генов, например цитокинов, и наиболее предпочтительно множества цитокинов, в отдельных клетках, например, активированных T-клетках, посредством мечения экспрессированных молекул РНК, включая молекулы мРНК или пре-мРНК, в фиксированных, пермеабилизованных клетках, при использовании наборов флуоресцентно меченных гибридизационных зондов и удаления не связавшихся зондов при промывке. Чувствительность к одиночной молекуле получают при использовании множества нуклеотидных гибридизационных зондов, которые обеспечивают множество флуоресцентных меток для каждой РНК-мишени. Они могут быть небольшим количеством зондов, множественно меченных специфическим флуорофором, или меченных зондов, дающих флуоресценцию при резонансном переносе энергии флуоресценции (FRET), когда они гибридизуются с когнатной мРНК или пре-мРНК. В предпочтительных способах настоящего изобретения используют большее количество, например от 10 до 100, более предпочтительно от 20 до 60, еще более предпочтительно от 30 до 50, например, приблизительно 50, более коротких олигонуклеотидных зондов, каждый из которых помечен одним флуоресцентным красителем, которые одновременно связываются с мишеневой последовательностью. Присоединение множества меток к каждой молекуле РНК делает клетку в достаточной степени флуоресцентной, выше фона, чтобы ее можно было обнаружить с помощью методов FC.
Кроме того, способы по настоящему изобретению включают обнаружение и анализ отдельных клеток, экспрессирующих мишеневую молекулу РНК или молекулы РНК, с помощью FC. Некоторые предпочтительные способы настоящего изобретения включают применение количественной FC для обнаружения клеток, которые содержат продукты экспрессии РНК генов цитокинов, индуцированные в требуемых клетках при стимуляции APC или aAPC.
FC включает гидроструйную систему для гидродинамического фокусирования, позволяющую создавать единый файл клеток. Одиночные клетки затем можно проанализировать на эмиссию флуоресценции при одной или нескольких длинах волн. FC является стандартной методикой в лабораториях клинической патологии. Например, иммунное фенотипирование с применением FC является важным инструментом в диагностике и определении стадии различных гематологических неоплазий. Важные этапы FC включают мечение клеток одной или более флуоресцентными молекулами, например флуорофорами, введение меченых клеток в проточный цитофлуориметр, облучение каждой клетки источником света возбуждения, таким как лазер, который испускается при длине волны, которая возбуждает флуоресцентные молекулы, и детекцию испускаемого флуоресцентного излучения при использовании фильтров и зеркал, которые отделяют определенные испускаемые длины волны для каждой используемой флуоресцентной молекулы, с получением данных, которые показывают присутствие и, предпочтительно, количество каждой флуоресцентной молекулы, связанной с каждой клеткой, основанной на возбуждении и испускании.
Как раскрыто в настоящей заявке, FC может использоваться для обнаружения клеток, экспрессирующих РНК, с помощью множества одиночно меченных флуоресцентных гибридизационных зондов. Наиболее предпочтительным вариантом осуществления является обнаружение РНК (мРНК или пре-мРНК), экспрессируемой с генов цитокинов в T-клеточной фракции выделенных PBMC, которые подвергли стимулированию посредством презентации антигенов M.tuberculosis эндогенными APC или, более предпочтительно, aAPC. РНК нескольких цитокинов можно обнаружить при помощи зондов, специфичных к каждому типу РНК, которые помечены различными флуоресцентными молекулами (например, различными флуорофорами) для каждого типа детектируемой РНК.
В способах настоящего изобретения, FC проводят на популяциях клеток (от не более 10000 клеток до более чем одного миллиона клеток). Данные регистрируют для одного или более событий. Событие представляет собой набор измерений для одной клетки. Данные для каждого события распределяют в определяемые пользователем окна, например, сигнал для флуорофора A или сигнал для флуорофора A, который находится между уровнем интенсивности X и уровнем интенсивности Y. Распределение данных FC - процесс, называемый гейтированием, причем события гейтируют или отбирают таким образом, что обычно они превышают определенный порог или ограничены определенными пороговыми пределами. Результаты затем могут быть представлены как абсолютные количества клеток в определенном окне или, предпочтительно, как фракция клеток в одном или более окнах, то есть частота. Например, в анализе состояния болезни результаты могли бы быть такими, что 10% клеток были бы положительными по мРНК A и мРНК B, 7% клеток были бы положительными по мРНК B и мРНК C, и 2% клеток были бы положительными по всем трем мРНК A, мРНК B и мРНК C. Если проанализировано 10000 клеток, минимальный положительный результат состоит в том, что определенное событие происходит в одной из 10000 клеток. С другой стороны, если проанализирован один миллион клеток, минимальный положительный результат состоит в том, что определенное событие происходит в одной из одного миллиона клеток.
Анализ результатов FC может включать дисперсионный анализ (ANOVA), который является набором статистических моделей для анализа дисперсии, обычно для проверки значимого различия между средними значениями для групп или переменных при разделении полной дисперсии на компоненты, обусловленные истинной случайной ошибкой, и компоненты, обусловленные различием между средними.
В настоящем изобретении также раскрыты наборы реагентов для осуществления описанный ранее способов. Наборы по настоящему изобретению включают по меньшей мере одно стимулирующее соединение, например белки или пептиды, которые будут стимулировать экспрессию одного или более цитокинов в качестве характеристики ответа тестируемого заболевания, или aAPC, которые загружены такими пептидами и несут костимулирующие молекулы, как описано ниже, и один или более наборов олигонуклеотидных зондов, каждый из которых специфично гибридизуется с одной последовательностью, присутствующей в мРНК или пре-мРНК, где каждый набор предпочтительно включает по 20-60 олигонуклеотидов длиной по 16-20 оснований, и при этом каждый олигонуклеотид помечен одним флуорофором. Набор, помимо мРНК-специфичных зондов, может дополнительно включать, например, флуорофор-конъюгированные антитела, узнающие клеточные белки, или химические зонды, меченные флуорофорами, связывающиеся с целевыми мишенями, реактивы для фиксации и пермеабилизации анализируемых клеток, реактивы для гибридизации включенных зондов с клетками и реактивы для постгибридизационной обработки, например, промывки, для удаления избыточного, негибридизованного зонда(ов) или других не связавшихся меченых зондов.
В предпочтительном варианте осуществления способ настоящего изобретения включает стимуляцию сигнализации TCR и последующей экспрессии генов AP-клетками, которые присутствуют в популяции клеток, или синтетическими сферами, которые именуются в настоящем описании как aAPC. Синтетические сферы служат в качестве механической подложки, то есть платформы, к которой белки (MHC и/или костимулирующие рецепторы и лиганды) могут быть присоединены посредством химического, электростатического или любого другого типа связывания. Таким образом, например, любая полимерная поверхность с или без химической модификации или дериватизации при необходимости может использоваться в качестве механической подложки или платформы для aAPC.
APC взаимодействуют (или реагируют) с добавляемыми пептидами (или белками) таким образом, что они связываются с (или эндогенно процессируются и презентируются) молекулами MHC и затем могут стимулировать TCR. aAPC взаимодействуют с добавляемыми пептидами, которые связываются с молекулами MHC, и затем эти нагруженные пептидами aAPC могут стимулировать TCR. Таким образом, aAPC несут один или более классов нагруженных пептидами молекул MHC, чтобы взаимодействовать с TCR, с или без одного или более антител или другого типа лигандов, которые взаимодействуют с костимулирующими рецепторами, экспрессируемыми на T-клетках, такими как CD28 и/или CD49d. В некоторых предпочтительных вариантах осуществления стимулирующие пептиды являются полученными из M.tuberculosis иммуногенами. Стадия стимулирования TCR включает инкубирование TCR-экспрессирующих клеток с APC или aAPC, которые включают молекулы MHC класса I или класса II, нагруженные соответствующими пептидами, и в некоторых вариантах осуществления костимулирующими белками или функциональными эквивалентами, такими как антитела, пептиды, различные углевод- и липид-несущие молекулы, которые связывают свои мишени на костимулирующих рецепторах, что усиливает сигнализацию, приводя к активации T-клеток.
Функционально различные субпопуляции T-клеток часто экспрессируются с частотой, близкой к доступным на сегодня пределам чувствительности (обычно 0,01-0,1%). Таким образом, их обнаружение требует мощной стимуляции TCR и применения высокочувствительных методов детекции, чтобы даже редкие субпопуляции T-клеток можно было непосредственно обнаруживать или размножить до титра выше предела чувствительности конкретного метода анализа. Способы настоящего изобретения включают индукцию эффективной стимуляции TCR AP-клетками, которые присутствуют в популяции клеток, или aAP-клетками, синтетической платформой на основе сфер, которая содержит: (i) молекулы MHC класса I или класса II, нагруженные соответствующими пептидами, и в некоторых предпочтительных вариантах осуществления (ii) костимулирующие белки (моноклональное антитело против CD28 и/или против CD49d), которые обеспечивают дополнительные сигналы (Oelke et al., 2003, Nature Medicine 9 (5):619-24]. Практически любой аллель MHC может быть присоединен к платформе на основе сфер, при этом любой костимулирующий сигнал можно представить как присоединенное к сферам специфичное моноклональное антитело (mAb) или лиганд корецептора, природные или определенные химически. В дополнение к своим превосходным свойствам стимуляции, aAPC очень стабильны: готовые aAPC клеточных размеров, со связанными синтетическими пептидами или без них, имеют длительный срок хранения в лиофилизированной форме и с легкостью могут переносить транспортировку.
В способах настоящего изобретения методика стимуляции обычно включает получение требуемых клеток у индивида, инкубирование клеток в культуральной среде и добавление соединений для стимуляции культивируемых клеток в течение различного времени, например, 30 минут, 1 ч, 2 ч, 4 ч, 6 ч, 8 ч, 10 ч, 12 ч, 14 ч, 16 ч или более длительных периодов (например, 1, 2, 3, 4 или 5 дней). Клетки могут быть получены при биопсии твердой ткани или аспирации клеток в жидкости из мишеневого участка, такой как бронхоальвеолярный лаваж, или источником клеток может быть периферическая кровь. Нужные клетки можно культивировать после их отделения от нежелательных клеток, подвергая последние лизису и отделяя интактные мишеневые клетки от остатков лизированных клеток, или отделения мишеневых клеток от нежелательных клеток на основе определенных свойств клеток. Например, PBMC из крови можно культивировать после лизиса эритроцитов (RBC) или после разделения на основе плотности клеток при осаждении в фиколле и удаления фиколла перед инкубированием в культуральной среде. Стимуляцию можно выполнить путем добавления стимулирующего соединения, как описано выше (такого как бактериальный, грибковый или вирусный продукт, или белок или пептид организма-хозяина), в достаточном количестве, чтобы производить ответ, который будет обнаружен, и инкубирования в течение достаточного времени, чтобы производить ответ. Ответ может быть синтезом пре-мРНК или мРНК в культивируемых стимулированных клетках. Ответ можно интерпретировать как набор характеристик ответа мишеневых клеток на данный стимул.
Для получения набора характеристик инфекции M.tuberculosis, например, aAPC на основе HLA-A*0201 могут быть нагружены HLA-A*0201 эпитопами (пептидами), которые, как известно, связываются с человеческим белком MHC класса I HLA-A-0201, полученным из известных антигенов, таких как Rv1886c, Rv3874 и Rv3875, и нагруженная антигеном aAPC на основе A2 может быть смешана с PBMC в отношении 1:1, при стандартных условиях культивирования, для обнаружения CD8+ T-эффекторов. aAPC на основе HLA-DRB1*04 можно нагружать подобным образом и использовать для обнаружения CD4+ T-эффекторов.
Для обнаружения клеток, содержащих копии одной или более определенных РНК, для каждой РНК можно использовать набор из множества флуоресцентно меченных олигонуклеотидных гибридизационных зондов. Олигонуклеотидные зонды могут представлять собой ДНК, РНК или смеси ДНК и РНК. Они могут включать неприродные нуклеотиды, аналоги нуклеотидов и неприродные межнуклеотидные связи. Каждый зонд может быть помечен множеством флуоресцентных молекул, например, множеством копий флуорофора, квантовой точкой или другой флуоресцентной молекулой. Например, в WO/1997/014816 описана детекция мРНК бета- и гамма-актинов с помощью одностадийной гибридизации in situ с использованием пяти зондов на одну мишень, зонды представляют собой одноцепочечную ДНК длиной приблизительно 50 нуклеотидов, меченную флуорофором (флуоресцеин или Cy3) по каждому десятому нуклеотиду, то есть по пять флуорофоров на зонд. В предпочтительных способах настоящего изобретения используется большее количество, например, от 10 до 100, более предпочтительно от 20 до 60, еще более предпочтительно от 30 до 50, например приблизительно 50, коротких олигонуклеотидных зондов, каждый из которых помечен одним флуоресцентным красителем, которые одновременно связываются с мишеневой последовательностью. Присоединение множества меток к каждой молекуле РНК делает клетку достаточно флуоресцентной, выше фона, чтобы ее можно было обнаружить с помощью методов FC. В большинстве вариантов осуществления каждый набор зондов будет иметь одну флуоресцентную молекулу, причем каждую флуоресцентную молекулу можно будет отличить при детекции от других присутствующих флуоресцентных молекул. Если же требуется обнаружить, экспрессирует ли каждая клетка первую РНК или вторую РНК, или нет, оба набора зондов могут быть помечены одной и той же флуоресцентной молекулой.
Фоновая флуоресценция представляет иную проблему для способов настоящего изобретения, в которых используется FC, нежели в микроскопических методах smFISH, в которых используется микроскопическая визуализация отдельных РНК в виде ярких пятен в клетках. В способах настоящего изобретения могут использоваться методы снижения фона. Одна из таких методик заключается в применении FRET между зондами, которые располагаются рядом на РНК мишени. Учитывая, например, что первый зонд и второй зонд, который расположен рядом с 5'-концом первого зонда (в пределах подходящей FRET дистанции, которая известна), примыкающего к 3'-концу второго зонда, можно добавить FRET донор на 5'-конец первого зонда и добавить FRET акцептор на 3'-конец второго зонда. При такой комбинации, клетки возбуждаются при длине волны поглощения донорного флуорофора, например, флуоресцеина, но сигнал детектируют при длине волны испускания акцепторного флуорофора, например, техасского красного. Поскольку флуорофоры по типу техасского красного не возбуждаются непосредственно, негибридизованные зонды или ошибочно гибридизованные зонды не испускают флуоресценцию. Другая методика состоит в применении FRET между красителем для двухцепочечной ДНК и флуорофором набора зондов, как это выполняют в анализе с использованием зондов ResonSense. В этом случае включают краситель для дцДНК, такой как SYBR Green (SYBR Green имеет сходные с флуоресцеином длины волн поглощения и испускания), при этом набор зондов метят флуорофором, который поглощает эмиссию от SYBR Green, например, TMR. При такой комбинации клетки можно возбуждать длиной волны поглощения SYBR Green, а детектировать испускание при длине волны эмиссии флуорофора. Поскольку флуорофор не возбуждается непосредственно, негибридизованные зонды не испускают флуоресценцию.
Хотя мишенями для зондов могут служить все области РНК, при подборе таких зондов следует учитывать несколько факторов. Предпочтительно мишеневая область не должна экспрессироваться в клетке с других областей генома или, более предпочтительно, мишеневая область вообще не должна присутствовать где-либо в геноме. Это можно обеспечить при проверке последовательности желаемой мишени в базах данных, в которых содержатся экспрессируемые гены и полные геномные последовательности изучаемого организма. Эта "фильтрация" последовательностей, потенциально генерирующих фоновый сигнал, может быть выполнена при использовании таких общедоступных компьютерных программ, как RepeatMasker. Зонды могут быть подобраны из непосредственно смежных областей мишеней, или между ближайшими зондами может быть некоторое пространство. Длина зондов может варьировать в зависимости от строгости гибридизации.
Как показано ниже, в примере 1 продемонстрирована индукция и применение зондов при распознавании наборов характеристик экспрессии генов в одиночных клетках.
Более конкретно, анализы проводили с целью исследования дифференцированных клеток THP-1, стимулированных облученной M.tuberculosis. Клетки фиксировали при инкубировании в течение 20 минут с 4% параформальдегидом через 24 часа после возбуждения и затем гибридизовали с зондами к двум мишеням мРНК. Одой из них являлся TNFα, ключевая мишень для анализа активированных T-клеток и макрофагов. Другой мишенью был ACSL1 (ген метаболизма липидов, GenBank, по каталогу № BC050073.1), который, как также ожидали, должен индуцироваться стимулом. Набор зондов, специфичных к TNFα (последовательность Ensembl ID ENST00000376122), включал 48 зондов длиной приблизительно по 20 нуклеотидов каждый, меченных на конце флуорофором тетраметилродамином. Набор зондов, специфичных к ACSL1, включал 48 зондов длиной приблизительно по 20 нуклеотидов каждый, меченных оптически распознаваемым флуорофором, а именно AlexaFluor 596.
В примере 1 экспрессируемые продукты РНК детектировали, используя известную микроскопическую методику, описанную в Raj et al., 2010, Methods in Enzymology, 472:365-386; и Raj et al., 2008, Nature Methods, 5:877-879, а именно smFISH. Эта методика включает обнаружение флуоресцентных пятен в качестве признака гибридизации набора зондов с отдельной молекулой РНК. В примере 1 детектировали пятна, соответствующие обеим молекулам мРНК. Изображения TNFα и ACSL1 накладывали в 3D стеки для каждого канала для одного набора клеток. Расшифровка включала обработку изображений при использовании компьютерной программы для идентификации дифракционно-ограниченных пятен, соответствующих индивидуальным молекулам мРНК, наложение их на изображение DIC клеток и получение поклеточного подсчета количества молекул мРНК, как описано в ссылках. Наблюдали сильную вариацию количества транскриптов для каждого гена в зависимости от клетки, что согласуется с предыдущим наблюдением, что синтез мРНК в клетках млекопитающих крайне стохастический. Результаты, среднее количество пятен для TNFα в стимулированных и нестимулированных клетках от общего количества пятен, которое подсчитывали в 50 последовательно проанализированных клетках, показали, что измерения одиночных клеток намного более информативны, нежели групповые измерения.
Регистрация данных FC для способов настоящего изобретения может быть выполнена обычными методами FC. Свет можно детектировать как в канале прямого рассеяния (FSC), так и в канале бокового рассеяния (SSC). Данные могут быть представлены в виде графиков плотности и контурных диаграмм. Для визуализации мишеневых клеток, с исключением результатов нежелательных частиц, например, дебриса, используются FC гейтирование. Диапазоны и окна, или области, определяют стандартным способом так, что в окне появляются положительные события. Чтобы гейтирование позволяло отличать истинно положительные события от ложноположительных, полезно использовать отрицательный контроль, то есть зонды к чему то, что клетки не могут экспрессировать. В принципе это аналогично белковой FC, где используются изотипический отрицательный контроль и/или неокрашенные клетки.
FC детекция показывает число положительных событий в образце, например, одно положительное событие на 10000 клеток (1/10000), три положительных события на 100000 клеток (3/100000) или два положительных события на один миллион клеток (2/106). Частота события может привести к заключению относительно биологического состояния, например, состояния болезни. В альтернативе, комбинации могут быть такими: например, 10% клеток в образце являются положительными по первому цитокину (цитокин A), а также положительными по второму цитокину (цитокин B). В качестве другого примера, состояние болезни можно было бы охарактеризовать следующей комбинацией: 10% или более клеток являются положительными по цитокинам A и B; 7% или более клеток являются положительными по цитокину B и третьему цитокину (цитокину C); и 2% или больше клеток являются положительными по всем трем цитокинам.
Как раскрыто в настоящем описании, было продемонстрировано, что FC может применяться для считывания данных в способах настоящего изобретения. Эксперимент, описанный ниже в примере 2, был разработан с целью проверки, можно ли с помощью FC обнаруживать сигналы, генерируемые наборами зондов, гибридизованных с РНК мишенями в отдельных клетках. В этом примере описаны этапы обнаружения мРНК GAG ВИЧ в клеточных культурах, экспрессирующих его с лентивирусной конструкции. Более конкретно, клетки 293T трансфицировали тремя плазмидами, которые вместе обеспечивают экспрессию рекомбинантной лентивирусной конструкции, и гибридизовали с зондами, специфичными к мРНК GAG. Результаты FC представлены на Фиг. 1, слева показаны графики интенсивности флуоресценции против канала прямого рассеяния (FSC-A), полученные для клеток, которые не были трансфицированы, и справа показаны клетки, трансфицированные тремя плазмидами из лентивирусной системы упаковки. Оба набора клеток гибридизовали с набором зондов к мРНК GAG. Популяция трансфицированных клеток производила сигналы интенсивности, которые на 1-2 порядка превышали величину, наблюдаемую для сигналов от исключаемых нетрансфицированных клеток (то есть выше установленного порога). В качестве контроля, основанный на изображениях анализ результатов smFISH показал, что приблизительно 25% клеток экспрессировали конструкцию. В анализе FC той же популяции установили, что флуоресцентной была та же фракция клеток. Эти результаты демонстрируют, что экспрессию мРНК с легкостью можно обнаруживать с помощью FC. Кроме того, данные обеспечивают руководство для отделения низкой частоты сильно стимулированных (с яркой флуоресценцией) T-клеток от выбросов в нестимулированной популяции. Анализ с использованием рабочей характеристической кривой (ROC, от англ. receiver operating characteristic, рабочая характеристика приемника) может позволить определить оптимальный порог. ROC-анализ известен специалистам в области разработки тест-систем. Смотрите, например, Zweig et al., 1993, Clinical Chemistry, 39 (8):561-577, и Pepe, 2003, The statistical evaluation of medical tests for classification and prediction, New York, NY: Oxford.
В детекции транскрипционных наборов характеристик активации T-клеток две сложные задачи - лишь малая субпопуляция клеток индуцирована, и индуцированные клетки экспрессируют всего несколько копий мРНК. В экспериментах, описанных в примере 7, авторы изобретения с помощью способов настоящего изобретения сумели продемонстрировать и чувствительное обнаружение низких частотных характеристик, и одновременное обнаружение нескольких мишеней мРНК. Анализ, который включал возбуждение, использование зондов на фиксированных клетках и FC, позволил обнаружить клетки, экспрессирующие пару IL-2/IFNγ с частотой 3,46% (Фиг. 2, правая панель), тогда как клетки, экспрессирующие пары цитокинов IL-2/TNFα и IFNγ/TNFα, были обнаружены с частотой 3,35% и 6,37%, соответственно. Сравнения популяций с одиночными и двойными цитокинами, например, IFNγ (4,76-5,40%) по отношению к IL-2/IFNγ (3,46%), позволили предложить, что не все продуценты IFNγ могли продуцировать IL-2. Напротив, большинство продуцентов IFNγ (5,40%), как оказалось, также экспрессировали TNFα, на что указывал размер IFNγ/TNFα-продуцирующей популяции (6,37%). Этот результат ожидали для популяции T-клеток, неспецифично активированных посредством комбинированной TCR активации и лектин-опосредованной стимуляции (mAb против CD3 и фитогемагглютинин). Таким образом, анализ по настоящему изобретению дал результаты, согласующиеся с известной цитологией T-клеток.
Как показано в примере 8, способ по настоящему изобретению использовали для обнаружения M. tuberuclosis-специфичных клеток, стимулированных ex vivo. PBMC инкубировали в течение 5 ч со смесью ESAT6- и CFP10-производных пептидов (T-SPOT.TB, Oxford Immunotec, UK) при концентрации 5 мкг/мл. Как показано на Фиг. 3, наблюдали приблизительно 2-кратное увеличение клеток, экспрессирующих мРНК цитокинов в Ag-специфичных T-клетках после этой стимуляции. Таким образом, даже в отсутствие костимуляции, обеспечиваемой mAb против CD28, анализ позволил обнаружить низкие частоты циркулирующих M.tuberuclosis-специфичных T-клеток, присутствующих в этом доноре LTBI. Данный результат демонстрирует применимость анализа в соответствии с настоящим изобретением.
В примере 9 описан способ по настоящему изобретению с усиленным сигналом по сравнению с примером 8. Захват корецептора CD28 TCR моноклональным антителом использовали для применения превосходной стимуляции к Ag-специфичным T-клеткам. Как показано в примере 8, PBMC инкубировали в течение 5 ч со смесью ESAT6- и CFP10-производных пептидов (T-SPOT.TB, Oxford Immunotec, UK) в концентрации 5 мкг/мл, но на этот раз с или без mAb против CD28, 53D10. Фиксированные и промытые клетки инкубировали с Cy5-меченными ДНК зондами, специфичными к зеленому флуоресцентному белку (GFP, отрицательный контроль) или IFNγ, и затем анализировали с помощью FC. Частоты продуцентов цитокина показаны на Фиг. 4. В присутствии mAb против CD28 наблюдали более чем 2-кратное увеличение экспрессии мРНК IFNγ в Ag-специфичных T-клетках донора LTBI. Костимуляция CD28 не влияла на неспецифичную гибридизацию с контрольными зондами к GFP и ВИЧ-1 GAG. В примере 9 показана иная стратегия усиления сигнала, а именно посредством передачи костимулирующих сигналов TCR, специфично захваченному когнатным Ag. Кроме того, так как костимуляция, опосредуемая mAb против CD28, не влияла на неспецифичное связывание контрольных зондов (GFP и ВИЧ-1 GAG), костимуляция может использоваться для увеличения силы специфичного сигнала по отношению к фоновому шуму.
Несколько стратегий особенно удобны для оптимизации анализов по настоящему изобретению. Они включают следующее:
i) Кинетика обнаружения сигнала. Кинетику экспрессии аналита мРНК для каждой из мишеней РНК (например, мРНК цитокинов IL-2, IFNγ и TNFα) определяют с клетками данного источника (например, разделенные через фиколл PBMC, полученные из проб крови доноров LTBI+ и LTBI-), которые стимулируют соответствующим индуктором или индукторами (например, смесью антигенов (например, ESAT6- и CFP10-производных пептидов (T-SPOT.TB, Oxford Immunotec, UK), присутствующих в концентрации 5 мкг/мл)), как в примере 8. Экспрессию каждого цитокина затем оценивают отдельно в Ag-стимулированных PBMC в различное время (например, 30 минут, 2, 4, 8, 12 и 16 ч). В частности, в случае цитокинов PBMC, неспецифично стимулированные с использованием mAb против CD3 и фитогемагглютинина (PHA), служат в качестве положительного контроля стимуляции. В анализе для исследования специфичного ответа PBMC, в предпочтительном способе измерения неспецифичного ответа используют и сравнивают PBMC из групп, которые включают лиц, которые, как известно, не поражены и поражены заболеванием, в отношении которого измеряют специфичный ответ.
ii) Эффект дозы. Эффект дозы в отношении стимуляции Ag может быть определен аналогичным способом (например, для каждого из этих трех цитокинов) при изменении концентраций (от 1 до 20 мкг/мл) смеси пептидов ESAT6 и CFP10 в оптимальный момент обнаружения сигнала. Дополнительно, неспецифичная стимуляция (антитело к CD3+PHA) PBMC от любых доноров может быть оптимизирована путем тестирования различных концентраций, отдельно и в комбинации, с обеспечением стандартного положительного контроля для конкретного анализа. В анализе для исследования специфичного ответа PBMC, в предпочтительном способе измерения неспецифичного ответа используют и сравнивают PBMC из групп, которые включают лиц, которые, как известно, не поражены и поражены заболеванием, в отношении которого будут измерять специфичный ответ.
iii) Усиление сигнала по отношению к шуму. Могут быть созданы наборы зондов с большей или более высокой интенсивностью флуорофоров. Эта цель также будет достигнута посредством нескольких стратегий улучшения. Дополнительно, способы мечения зондов могут быть изменены, как описано выше. Строгость гибридизации может быть оптимизирована опытным путем для увеличения отношения сигнал/шум.
Еще одна стратегия усиления сигнала состоит в передаче костимулирующих сигналов к TCR, специфично захваченному когнатным Ag, как описано в отношении примера 9. Условия костимуляции могут быть оптимизированы, как указано выше в отношении кинетики и эффекта дозы, для достижения надежного обнаружения множества молекул РНК (например, трех цитокинов) с помощью анализа, описанного в настоящей заявке.
iv) Определение порога. Начальная оценка порога для обнаружения РНК может служить для стандартизации оптимизированного анализа, хотя клинически принятый порог для конкретного анализа может быть определен на более поздней стадии создания коммерческого теста. В фазе исследования можно оценить порог обнаружения РНК (экспрессия цитокина) в Ag-специфичных T-клетках, полученных у доноров и стимулированных ex vivo. Ag-специфичные ответы можно вычислить, вычитая сигнал от нестимулированных PBMC из значений, полученных с Ag-стимулированными PBMC, за оптимальное время и при оптимальной дозе Ag. Для каждого считывания данных цитокина ROC кривая, как оценивают, показывает отношение между чувствительностью и специфичностью в различных точках пересечения (включая средние значения для доноров плюс два стандартных отклонения), и оптимальные пороговые значения выбирают для идентификации Ag-специфичных ответов.
v) Уменьшение объема крови и продолжительности обработки. В настоящее время анализируют 2×106 клеток/параметр (например, каждый цитокин), что соответствует, в среднем, 0,8 мл крови. С применением лизиса эритроцитов в цельной крови вместо разделения в фиколле при получении PBMC, а также других модификаций обработки крови, например, вакуумного осаждения (96-луночный формат) вместо центрифугирования, можно уменьшить объемы проб и сократить время анализа. Предварительные эксперименты показывают, что время гибридизации можно уменьшить до 2 часов (ч) без существенной потери сигнала.
Разработку анализа можно проводить обычными методами. Например, PBMC трех групп доноров (активный TB, LTBI и неинфицированные контрольные лица) можно стимулировать aAPC на основе HLA-A2 и HLA-DR4, нагруженными пептидами ESAT6 и CFP10, как в примере 4. Каждый анализ позволяет протестировать набор из 4 маркеров реакции на стимуляцию (оцениваемой вместе на основе свойств экспрессии), выбранных из группы, включающей, без ограничения, гены, показанные в таблице 1, при использовании 4-цветной FC для анализа. Это позволяет анализировать все мишени для каждого донора. Маркеры, выбранные в первоначальных тестах, могут быть повторно протестированы в комбинациях, выбранных с целью группировки генов, экспрессируемых на сходных уровнях в каждой группе. Статистический анализ может быть выполнен как описано в примере 4. При необходимости также можно измерять экспрессию мишеней, индуцированных при обычной стимуляции APC, для подтверждения способности aAPC повышать чувствительность обнаружения для множества аналитов внутри одиночных клеток. Кроме того, анализ также может быть протестирован с помощью флуоресцентной микроскопии. Статистический анализ может быть выполнен как описано в примере 5.
Эксперимент, описанный в примере 10, демонстрирует, что та же самая методика, которая именуется как "FISH-Flow", используемая для обнаружения экспрессии цитокинов мРНК в T-клетках, может успешно использоваться в других типах клеток, включая, без ограничения, первичные макрофаги. Кроме того, эксперимент демонстрирует, что ответы на различные стимулы можно распознавать в первичных макрофагах, отслеживая экспрессию одного гена в разное время.
Результаты, показанные на Фиг. 5, показывают, что фоновая экспрессия, оцениваемая по отрицательному контролю GFP, оставалась постоянной, но незначительной (приблизительно 0,1-0,2% гейтированных клеток). Обработка смесью липополисахарида (LPS) и IFNγ или N-пальмитоил-5'-[2,3-бис(пальмитоилокси)-пропил]-(R)-цистеинил-(лизил)3-лизина (Pam3CSK4) с легкостью вызвала экспрессию транскриптов TNFα в этих культивируемых макрофагах до уровня от 6 до 42% всех клеток, являющихся положительными. Четкая экспрессия транскриптов TNFα ожидается в активированных макрофагах и контрастирует с умеренной (меньше 2%) экспрессией мРНК IFNγ, на которую не влиял выбор стимулов и/или длительности возбуждения, и, таким образом, может отражать необычно высокий фон в этих клетках, а не реальный профиль экспрессии. Кинетика экспрессии мРНК TNFα, индуцируемая LPS и IFNγ, вероятно, отличается от кинетики экспрессии, индуцируемой Pam3CSK4, при этом указанная выше стимуляция приводит к стойкому, долговременному повышению уровней мРНК. Напротив, стимуляция той же мРНК при воздействии Pam3CSK4 была значительно менее стойкой.
Пример 11 демонстрирует идентификацию временных профилей экспрессии генов цитокинов, хемокинов и рецепторов хемокинов, которые являются маркерами активации T-клеток, в стимулированных Т-клетках человека, при сравнении с результатами проточной цитометрии, полученными при использовании зондов FISH, специфичных к указанным мРНК (Фиг. 6A) или при использовании mAb, узнающих их белковые продукты (Фиг. 6B). Частоты продуцентов цитокинов согласно FISH-Flow (Фиг. 6A) показали, что экспрессия мРНК IL-2, IFNγ, TNFα, CCL3, CCR7 и cFos в нестимулированных клетках являлась либо незначительной (контроль GFP, IL-2, IFNγ и CCR7), либо очень низкой (TNFα, CCL3 и cFos) и наблюдалась в <1% гейтированных клеток. Неожиданно индуцированная экспрессия этих мРНК, как оказалось, достигает максимума через 2 часа после стимуляции, а затем падает в 1 день и 3 день. Это отличается от параллельных данных, полученных с теми же клетками (в присутствии 1 мкг/мл брефелдина A) при использовании обычного иммуноокрашивания mAb и проточной цитометрии. В данном случае нестимулированные клетки четко показывают базальные уровни (1-3%) экспрессии белка, обнаруживаемые по меньшей мере для некоторых маркеров, таких как IL-2, IFNγ и TNFα, при сравнении с отрицательным контролем - сходным по изотипу (Iso) mAb. Кроме того, все мишени через 2 часа индуцированы и остаются на повышенных уровнях в течение 3-дневного периода исследования. Таким образом, никаких других временных профилей нельзя было обнаружить с использованием mAb, которые связываются с цитокинами. Пример 11 демонстрирует различия в кинетике, что обеспечивает превосходную профильную дискриминацию при использовании FISH-Flow по сравнению с обычным окрашиванием mAb и проточной цитометрией и указывает на то, что вышеуказанный способ обеспечивает существенно более высокую чувствительность для обнаружения активации.
Для полного определения функционального набора характеристик клетки любого типа (то есть способности выполнять специализированные биологические функции) может быть важно знать экспрессию всех других мРНК в активированных клетках при сравнении с неактивированными (неотвечающими) клетками. Исследование, представленное в примере 12, демонстрирует, что FISH-Flow позволяет выполнять мечение клеток, которые экспрессируют одну или множество мишеней в ответ на стимуляцию, и что такие респондеры могут быть выделены из целой клеточной популяции с целью исследования их профилей экспрессии генов.
Стимулированные Т-клетки человека инкубировали либо с Cy5-меченными зондами ДНК, комплементарными к РНК, специфичной к GFP, и анализировали с помощью FC в качестве контроля, либо со смесью нескольких наборов зондов FISH (таблица 3), активных в отношении мРНК IL-2, IFNγ, TNFα и CCL3, при этом все зонды одиночно помечены Cy5. Результаты, полученные с зондами GFP, показаны на Фиг. 7A. Результаты, полученные со смесью зондов, показаны на Фиг. 7B. После гейтирования в случае популяции лимфоцитов (левая панель на каждой фигуре) применяли консервативное дополнительное гейтирование для разделения проанализированных лимфоцитов на две различные популяции (правая панель на каждой фигуре), что позволяет отличить клетки, экспрессирующие IFNγ (IFN+), от клеток, не экспрессирующих IFNγ (IFN-). Гейтирование клеток, гибридизованных с зондами GFP (Фиг. 7A), определило две популяции: 0,1% в популяции IFN+ и 71,2% в популяции IFN-, из всех проанализированных лимфоцитов. Клетки, гибридизованные с зондами IL-2:IFNγ:TNFα:CCL3 (Фиг. 7B), были следующими: 10,2% IFN+ и 33,7% IFN- с таким же гейтированием. Последние, гибридизованные с зондами FISH, клетки (Фиг. 7B) разделяли на положительные и отрицательные популяции с помощью сортировки флуоресцентно-активированных клеток (FACS) для выполнения анализа экспрессии генов в выделенных активированных и неотвечающих клетках. Сортированные популяции клеток анализировали с целью демонстрации различных профилей экспрессии генов, согласующихся с их активностью в отношении определенных зондов FISH. Экспрессию мРНК, кодируемых двумя генами, IFNγ и геном "домашнего хозяйства" GAPDH (контрольным), выбрали, чтобы иллюстрировать вариант осуществления данного аспекта изобретения. Сортированные популяции клеток подвергали ОТ-ПЦР при использовании праймеров, специфичных к одному из этих двух генов. Количественное измерение амплифицированного продукта выполняли при включении красителя SYBR Green в реакционную смесь для амплификации. Результаты, показанные на Фиг. 8A, демонстрируют, что IFNγ-специфичные праймеры обнаруживают мишени в большем количестве в клетках IFN+, которые были идентифицированы с использованием зондов к IL-2, IFNγ, TNFα и CCL3, чем в клетках IFN-, которые были на минимальном уровне помечены указанными зондами. Амплификация сигнала контрольного GAPDH, который не изменяется при активации, как ожидают, будет аналогичным и в активированных клетках, и в клетках-нереспондерах. Немного пониженная амплификация, которую наблюдали в случае GAPDH-специфичных праймеров в активированных клетках, отражает тот факт, что эта сортированная популяция значительно меньше и, таким образом, могла давать меньшее количество мРНК.
Эти результаты (подтвержденные анализом размера, Фиг. 8B) демонстрируют, что экспрессия генов может быть также проанализирована в требуемых субпопуляциях клеток, идентифицированных с применением платформы FISH-flow настоящего изобретения, при использовании методов, известных специалистам в области сортировки и получения субпопуляций клеток, а также методов измерения экспрессии генов, таких как ОТ-ПЦР или транскриптомный анализ. Субпопуляция, которая может быть идентифицирована и изучена, может содержать одну клетку или больше одной клетки, в том числе, без ограничения, 10, 100, 1000, 10000, 100000 или больше клеток.
ПРИМЕРЫ
ПРИМЕР 1. ДЕМОНСТРАЦИЯ ИНДУКЦИИ И НАБОРОВ ЗОНДОВ
Данный пример демонстрирует применение искусственной стимуляции клеток с целью продукции РНК, в данном случае мРНК, и применение наборов одиночно меченных флуоресцентных зондов для гибридизации с экспрессируемыми РНК.
В этом примере методику smFISH использовали для визуализации пятен, соответствующих индивидуальным молекулам мРНК, связанным с зондами. Смотрите Raj et al., 2010, Methods in Enzymology 472:365-386; и Raj et al., 2008, Nature Methods 5:877-879. Условия гибридизации зондов и условия промывки клеток описаны в указанных ссылках. Методика отличалась одним важным моментом. Чтобы избежать потери клеток на стадиях промывки, в промывочные растворы включали 0,5% эмбриональной бычьей сыворотки. Набор нуклеотидных зондов, в данном случае зондов ДНК, был подобран для мРНК TNFα, ключевой мишени для анализа активированных T-клеток и макрофагов, и использован при тестировании дифференцированных клеток THP-1, стимулированных облученной M.tuberculosis. Также был включен второй набор зондов ДНК, меченных флуорофором другого цвета и предназначенных для связывания с мРНК ACSL1 (ген метаболизма липида). Последний ген, как также ожидали, будет индуцироваться стимулом. Набор зондов, специфичных к TNFα человека, включал 48 зондов длиной приблизительно по 20 нуклеотидов каждый, меченных на конце флуорофором тетраметилродамином. Набор зондов, специфичных к гену ACSL1 человека, включал 48 зондов длиной приблизительно по 20 нуклеотидов каждый, меченных оптически распознаваемым флуорофором, а именно alexafluor 594. Последовательности зондов перечислены в списке последовательностей в конце настоящего описания (таблица 3). Хотя зонды, перечисленные в каждом наборе, подходят для детекции мишени мРНК, также могут использоваться альтернативные зонды, отобранные из различных областей мишеней РНК, как описано в Raj et al., 2010, Methods in Enzymology 472:365-386; и Raj et al., 2008, Nature Methods 5:877-879. Как обсуждается выше, детектировали пятна, соответствующие обоим мРНК в клетках. Число молекул РНК TNFα увеличилось с 0,24 на клетку без индукции до 14 на клетку, в среднем, после стимуляции.
ПРИМЕР 2. ИСПОЛЬЗОВАНИЕ FC ДЛЯ ОБНАРУЖЕНИЯ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ ГЕН
В данном эксперименте анализы проводили с целью обнаружения мРНК GAG ВИЧ в культурах клеток, экспрессирующих лентивирусную конструкцию.
Коротко, клетки 293T трансфицировали тремя плазмидами, которые вместе обеспечивают экспрессию рекомбинантной лентивирусной конструкции, и подвергали гибридизации с зондами, специфичными к области мРНК GAG ВИЧ (GenBank, по каталогу № AY835771.1). Последовательности зондов перечислены в конце настоящего описания (таблица 3). Зонды гибридизовали, после чего клетки промывали согласно методикам, описанным в статьях Raj et al., 2010, Methods in Enzymology 472:365-386; и Raj et al., 2008, Nature Methods 5:877-879, которые включены в настоящее описание в качестве ссылки в полном объеме. Опять же, чтобы избежать потери клеток на стадиях промывки, в промывочные растворы включали 0,5% эмбриональной бычьей сыворотки. Затем выполняли FC, чтобы обнаружить клетки, экспрессирующие конструкцию. Гейты устанавливали по флуоресценции клеток, которые не были трансфицированы. В клетках, которые были трансфицированы, приблизительно 25% клеток экспрессировали конструкцию, что было отражено сигналом большей интенсивности по сравнению с уровнем, установленным гейтом. Результаты FC представлены на Фиг. 1, на которой показаны две диаграммы интенсивности флуоресценции ("у.е.") в зависимости от прямого рассеяния ("FSC-A"). На диаграмме слева (Лентивирус-) показаны клетки, которые не были трансфицированы. На диаграмме справа (Лентивирус+) показаны клетки, которые были трансфицированы тремя плазмидами из лентивирусной системы упаковки.
ПРИМЕР 3. ОБНАРУЖЕНИЕ ЭКСПРЕССИИ ГЕНОВ ЦИТОКИНОВ, СТИМУЛИРОВАННОЙ M-TB ПРОИЗВОДНЫМИ ПЕПТИДАМИ
PBMC получали у неинфицированных доноров, доноров с бессимптомной латентной инфекцией (LTBI) и доноров с активной формой TB. T-клетки активировали добавлением смеси пептидов, используемой в анализе секреции IFNγ T-SPOT.TB IFNγ Release Assay (IGRA) (Oxford Immunotec, Marlborough, MA) согласно инструкциям производителя, которая получена из иммунодоминантных микобактериальных Ag Rv3875 (ESAT6) и Rv3874 (CFP10). В разные моменты времена (от 4 до 120 ч) стимулированные клетки подвергали фиксации и пермеабилизации, а затем гибридизовали с один или более наборами зондов, описанных ниже. Клетки анализировали FC для определения частоты клеток, которые экспрессируют мишени, отдельно и в комбинации, при использовании доступных наборов фильтров, которые позволяют различать флуорофоры, используемые для мечения зондов.
Набор зондов для TNFα является набором зондов, описанным в примере 1. Получали два дополнительных набора зондов, один для IL-2 и один для IFNγ, содержащие по 50 зондов (номера последовательностей Ensembl ENST00000226730 и ENST00000229135, соответственно). Зонды во всех трех наборах представляют собой линейные (случайная спираль) олигонуклеотиды длиной 15-25 нуклеотидов, меченные одиночным флуорофором. Три флуорофора, используемые в трех наборах, являются оптически различимыми. Эти три гена цитокинов являются известными маркерами активации и пролиферации T-клеток, которые позволяют дифференцировать подтипы T-клеток. Транскрипты-мишени имеют достаточную длину, чтобы на каждом транскрипте можно было использовать по меньшей мере 50 зондов.
Для обнаружения экспрессии маркера в малой фракции (<0,01%) клеток с помощью FC, регистрировали результаты для 1×106 событий. Чтобы определить статистическое значение различий между группами доноров, порог для разделения клеток-нереспондеров от клеток-респондеров определяли с помощью ROC-анализа (смотрите, например, Zweig et al., 7993, Clinical Chemistry 39(8):561-577, и Pepe, 2003, The statistical evaluation of medical tests for classification and prediction. New York, NY: Oxford), и определяли частоту положительных клеток. Для анализа данных может использоваться смешанная модель дисперсионного анализа (ANOVA). Перед анализом данные преобразовывали (например, логарифмически). В случаях, когда простое преобразование было недостаточным, чтобы обратиться к допущения ANOVA, применяли непараметрический аналог статистического метода.
В активированных клетках была определена обнаружимая частота экспрессии для всех трех маркеров. Частоты клеток, которые экспрессируют различные комбинации маркеров, являются набором характеристик болезни.
ПРИМЕР 4. АНАЛИЗ ДЛЯ М.TUBERCULOSIS-СПЕЦИФИЧНЫХ ЭФФЕКТОРНЫХ T-КЛЕТОК
Данный пример описывает анализы для детекции М.Tuberculosis-специфичных эффекторных T-клеток.
a. Обнаружение CD8+ T-эффекторов. Пациенты с TB проходили HLA-типирование с помощью FC детекции PBMC, иммунно-окрашенных с использованием аллель-специфичных mAb BB7.2 и 0222, или посредством подтверждения ПЦР. aAPC на основе HLA-A*0201 нагружали известными отдельными эпитопами HLA-A*0201 (пептидами), полученными из Rv1886c, Rv3874 и Rv3875.
Эти пептиды получены из иммунодоминантных Ag M.tuberculosis. PBMC выделяли у HLA-A2-типированных пациентов с TB и стимулировали aAPC, нагруженными Ag, на основе A2, в соотношении 1:1, в стандартных условиях культивирования. В альтернативе, выделенные PBMC инкубировали с добавлением пептидов для стимуляции T-клеток эндогенными APC. В различное время, от 4 до 120 ч, клетки фиксировали, пермеабилизовывали и инкубировали с зондами, специфичными к IL-2, IFNγ и TNFα. Отрицательный контроль включал PBMC от здоровых, неинфицированных (IGRA-отрицательных) доноров, стимулированные вышеописанными Ag-нагруженными aAPC, и PBMC от положительных доноров, стимулированные aAPC, нагруженными нерелевантными меланомаспецифическим пептидом Mart 1 (эпитоп 26-35) и пептидом дифференцировки Ag gp100 (эпитоп 44-59). Положительный контроль включал PBMC, неспецифично стимулированные mAb против CD3+PHA.
b. Обнаружение CD4+ T-эффекторов. Сначала конструировали aAPC на основе HLA-DRB1*04, используя методику, описанную выше для сборки aAPC на основе HLA-A*0201. Главным аспектом данного подхода является экспрессия растворимой формы молекулы HLA-DR4, что требует спаривания полипептидов a и b класса II в процессе биосинтеза. Для облегчения спаривания субъединиц растворимых аналогов молекул MHC класса II, в качестве молекулярного каркаса использовали молекулу IgG: она является бивалентной и ее можно с легкостью модифицировать с целью включения целого ряда белковых доменов. Модифицированные цепи a и b клонировали в бифункциональный бакуловирусный вектор экспрессии. Клетки, трансфицированные таким вектором, секретируют приблизительно от 0,5 до 2,0 мг/мл растворимого HLA-DR4-Ig-подобного материала. Для сборки растворимых комплексов MHC класса II HLA-DR4-Ig, при использовании универсального вектора клонирования pZig ДНК, кодирующую внеклеточные домены класса II цепей α и β белков DR4 лигировали с ДНК, кодирующей тяжелую и легкую цепи Ig, используя сайты эндонуклеаз рестрикции; и лигировали химерные конструкции последовательно в модифицированный вектор pAcUW51. Выход, чистоту и целостность секретируемых белков подтверждали биохимическим анализом (ДСН-ПААГ и Вестерн-блоттинг) с HLA-DR4-сецифичным mAb 0222HA (One Lambda). Очищенные растворимые белки HLA-DR4-Ig и mAb против CD28 в соотношении 1:1 конъюгировали с магнитными сферами (M-450 Epoxy Dynabeads, Dynal Invitrogen).
Известными пептидами, полученными из Rv1886c, Rv3874 и Rv3875 и ограниченными аллелем HLA-DRB1*04 (таблица 1), нагружали aAPC на основе HLA-DR4 и использовали для стимуляции PBMC, полученных от неинфицированных доноров, доноров с LTBI и доноров с активным TB. В альтернативе, выделенные PBMC инкубировали с добавлением пептидов для стимуляции T-клеток эндогенными APC. После стимуляции в различные моменты времени, от 4 до 72 ч, клетки гибридизовали с зондами к мРНК IL-2, IFNγ и TNFα и анализировали с помощью FC. Использовали отрицательный и положительный контроль, как описанный выше для CD8+ T-клеток.
HLA-A*0201- и HLA-DRB1*04-ограниченные эпитопы
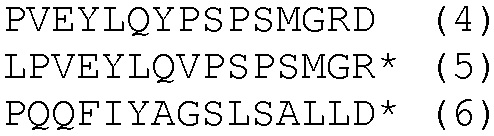


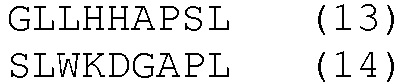


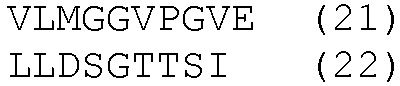
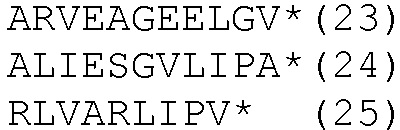
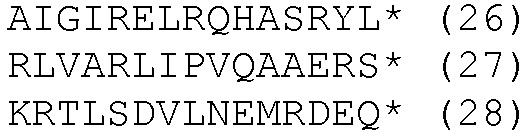
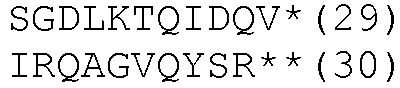
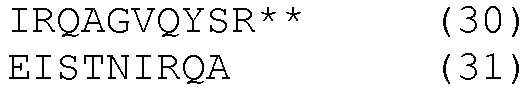
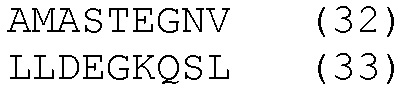
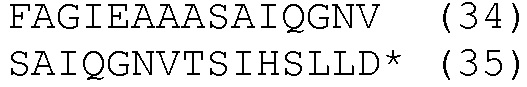
К этим результатам применяли методы статистического анализа, описанные в примере 3. Методы множественной стимуляции в этом примере также требуют смешанной модели ANOVA для сравнений.
Предыдущий тест позволяет обнаруживать как CD4+, так и CD8+ эффекторные T-клетки в крови пациентов с TB на основе стимуляции аллельспецифическими HLA класса I или класса II-ограниченными пептидами. Активированные эффекторные T-клетки, узнающие антигены Rv1886c, Rv3874 и Rv3875 и способные вырабатывать по меньшей мере один цитокин, IFNγ или TNFα, можно обнаруживать с легкостью. Одновременная продукция двух цитокинов (IL-2+IFNγ+ и IL-2+TNFα+) является менее распространенной в полностью дифференцированных T-клетках, которые представляют большинство эффекторов в крови пациентов с активным TB.
ПРИМЕР 5. ЦИТОКИНОВЫЕ ПРОФИЛИ ОДИНОЧНЫХ T-КЛЕТОК, ИНДУЦИРУЕМЫЕ ИНФЕКЦИЕЙ СТАДИЕСПЕЦИФИЧЕСКИХ АG
PBMC от доноров с активным TB, LTBI и неинфицированных доноров стимулировали в различные моменты времени (от 4 до 120 ч) aAPC на основе HLA-A2 и HLA-DR4, нагруженными смесями пептидов, полученных из Rv1886c (активная стадия TB) и Rv1986, Rv2659c и Rv3407 (стадия LTBI). В альтернативе выделенные PBMC инкубировали с добавлением пептидов для стимуляции T-клеток эндогенными APC. Регистрируемым результатом является экспрессия IL-2, IFNγ и TNFα при использовании трех наборов из приблизительно по 50 одиночно меченных флуоресцентных зондов и обнаружение с помощью FC количества экспрессирующих клеток. Для идентификации кандидатных Ag, которые могут позволять различать состояния болезни, использовали однофакторные ANOVA и высокое p-значение (p=0,25) в качестве порогового. Отобранные Ag использовали для создания модели ответов, которая распознает группы доноров, используя обобщения логистической регрессии, такие как политомную или интегральную логистическую регрессию.
Стимуляция с использованием Rv1886c дает характерный, связанный со стадией инфекции, цитокиновый профиль T-клеток: одиночные, двойные и тройные продуценты в случае пациентов с TB и, главным образом, продуценты IL-2 в случае пациентов с LTBI.
ПРИМЕР 6. ИДЕНТИФИКАЦИЯ НАБОРА ХАРАКТЕРИСТИК ФУНКЦИОНАЛЬНЫХ ЭФФЕКТОРНЫХ T-КЛЕТОК И T-КЛЕТОК ПАМЯТИ, СВЯЗАННЫХ СО СТАДИЕЙ ИНФЕКЦИИ, В ОДИНОЧНЫХ T-КЛЕТКАХ
Функциональные маркеры, индуцируемые при активации TCR, могут быть измерены с применением способов настоящего изобретения, чтобы точно описать главные функциональные субпопуляции T-клеток. Для этого анализа были отобраны кандидатные маркеры, индуцируемые в течение 4-6 часов после захвата TCR, и/или гены, которые отражают стадию созревания и функцию известных субпопуляций T-клеток. Полученная группа активации и функциональные маркеры, экспрессируемые в эффекторных T-клетках, эффекторных T-клетках памяти и центральных T-клетках памяти (таблица 2), удовлетворяет таким критериям. Данные в таблице 2 были составлены из Zinkernagel RM., T cell activation, в Mak TW, Saunders ME, editors. New York: Elsevier; 2006. p. 373-401, и из Doherty P., T cell differentiation and effector function, в Mak TW, Saunders ME, editors. New York: Elsevier; 2006. p. 403-432, а также приведенных там ссылок.
Ключевые маркеры при определении набора характеристик T-клеток*
Использование aAPC, нагруженных пептидами (как в примере 3 или 4), для стимуляции через TCR и костимуляции через CD28 или, в альтернативе, mAb против CD28 в сочетании с пептидами (как в примере 3 или 4) для стимуляции эндогенными APC, быстро увеличивает экспрессию ключевых хемокинов и цитокинов (в том числе, без ограничения, ΜΙΡ-1α, IFNγ и TNFα) и других костимулирующих молекул (в том числе, без ограничения, CD40L, CD137), что в свою очередь дополнительно повышает реактивность. Положительная обратная связь усиливает ответы во временных рамках, используемых для детекции, и повышает чувствительность результатов FC для мультипараметрического анализа. Комбинации отобранных маркеров выбирают для обеспечения воспроизводимой идентификации и определения характеристик отдельных эффекторных клеток и клеток памяти, а также их количественного анализа в периферической крови групп доноров. Они служат в качестве функциональных наборов характеристик T-клеток. Четырехцветную FC используют, чтобы определяемый набор(ы) характеристик был пригоден для анализа с применением инструментов, обычно используемых в клинических условиях.
Высокую воспроизводимость по донорам обнаружили для данных, полученных с PBMC, стимулируемых коммерческой смесью пептидов ESAT6 и CFP10 и aAPC на основе HLA-A2 или HLA-DR4. Данные, полученные с этой смесью пептидов, совпадают со стандартным IGRA, основанным на эндогенных APC. Надежная стимуляция, которую обеспечивают aAPC, распространяется на индукцию множественных мишеней, как описано выше в отношении стандартных цитокиновых мишеней. Клетки, экспрессирующие множественные маркеры, также обнаруживали с более высокой частотой, чем обычно наблюдают при стандартной стимуляции APC. Способ в настоящем примере позволяет проводить идентификацию наборов маркеров, которые служат в качестве наборов характеристик отдельных T-клеток, которые присутствуют при различных стадиях инфекции, в данном случае инфекции TB.
ПРИМЕР 7. ОДНОВРЕМЕННОЕ ОБНАРУЖЕНИЕ МРНК ДВУХ ЦИТОКИНОВ В СТИМУЛИРОВАННЫХ PBMC
В данном примере показано, что можно обнаруживать очень малую популяцию флуоресцентных клеток. Коротко, CFSE-меченные PBMC последовательно разводили в MITOTRACKER (Invitrogen)-меченных PBMC. Было установлено, что анализ позволял обнаруживать первые при частоте 0,0013%, что демонстрирует достаточную чувствительность.
Чтобы продемонстрировать обнаружение мРНК IL-2, IFNγ и TNFα, PBMC неспецифично стимулировали через TCR, используя mAb против CD3 и PHA, который, как известно, вызывает экспрессию различных цитокинов в этих клетках. После стимуляции, клетки фиксировали и исследовали с использованием парных комбинаций наборов зондов против IL-2 и IFNγ, IL-2 и TNFα, а также IFNγ и TNFα. Один набор зондов в каждой паре был помечен флуорофором Cy5, а другой был помечен флуорофором TMR. В частности, PBMC стимулировали mAb OKT3 против CD3 и PHA (оба в концентрации 1 мг/мл) в течение 5 дней, а затем фиксировали 4% параформальдегидом и 70% этанолом. Фиксированные и промытые клетки метили путем инкубирования с Cy5- или TMR-меченными зондами РНК, специфичными к IL-2 и IFNγ, снова промывали и анализировали с помощью двухцветной FC (LSRII, Becton Dickinson). Результаты показаны на Фиг. 2. Четыре графика на Фиг. 2 относятся к следующему, слева направо: неокрашенный/фиктивно гибридизованный контроль; одиночный цитокин, IFNγ-Cy5-меченные клетки; одиночный цитокин, IFNγ-TMR-меченные клетки; и двойной цитокин (IL-2, Cy5-меченные и IFNγ, TMR-меченные)-меченные клетки. Гэйтирование демонстрирует популяции клеток, экспрессирующих один или два цитокина, и их частоты.
ПРИМЕР 8. ОБНАРУЖЕНИЕ ЦИТОКИНОВ МРНК В М.TUBERUCLOSIS-СПЕЦИФИЧНЫХ КЛЕТКАХ, СТИМУЛИРОВАННЫХ EX VIVO
В данном примере способ по настоящему изобретению применяли для обнаружения М.tuberuclosis-специфичных клеток, стимулированных ex vivo. Циркулирующие М.tuberuclosis-специфичные клетки обычно присутствуют с низкими частотами у лиц с латентной формой TB инфекции (LTBI). Кровь у донора с LTBI получали и стимулировали в течение 5 часов смесью пептидов, полученных из Ag M.tuberculosis ESAT6 и CFP10 (обычно используемых в тесте T-SPOT.TB). PBMC инкубировали в течение 5 ч либо в одной среде, либо со смесью ESAT6- и CFP10-производных пептидов (T-SPOT.TB, Oxford Immunotec, UK) при концентрации 5 мкг/мл. Фиксированные и промытые клетки инкубировали с Cy5-меченными зондами ДНК, комплементарными к РНК, специфичной к GFP, в качестве отрицательного контроля, или к IL-2 и TNFα, а затем анализировали с помощью FC. Частоты продуцентов цитокинов показаны на Фиг. 3, где три верхних графика относятся к PBMC, которые инкубировали в одной среде, а три нижних графика - к PBMC, которые инкубировали в среде, содержащей смесь Ag.
ПРИМЕР 9. ОБНАРУЖЕНИЕ IFNγ В КОСТИМУЛИРОВАННЫХ PBMC ДОНОРА С LTBI
PBMC инкубировали в течение 5 ч со смесью ESAT6- и CFP10-производных пептидов (T-SPOT.TB, Oxford Immunotec, UK) при концентрации 5 мкг/мл, с или без mAb 53D10 против CD28. Фиксированные и промытые клетки инкубировали с Cy5-меченными ДНК зондами, специфичными к мРНК GFP (GFP, отрицательный контроль) (последовательность GFP соответствует pTREd2EGFP (Invitrogen)) или мРНК IFNγ, а затем анализировали с помощью FC. Частоты продуцентов цитокинов показаны на Фиг. 4. На неспецифичное окрашивание GFP контроля костимуляция CD28 не повлияла.
ПРИМЕР 10. ИНДУКЦИЯ ЦИТОКИНОВ В АКТИВИРОВАННЫХ МАКРОФАГАХ
Данный пример иллюстрирует применение искусственной стимуляции не T-клеток, в данном случае макрофагов человека, полученных из периферической крови при адгезии на пластике и культивируемых в течение 2 недель в стандартных условиях (среда RPMI 1640 с добавкой 10% эмбриональной бычьей сыворотки и антибиотиков).
Коротко, активацию выполняли смесью липополисахарида (LPS) (100 нг/мл) и IFNγ (20 нг/мл) или N-пальмитоил-5'-[2,3-бис(пальмитоилокси)-пропил]-(R)-цистеинил-(лизил)3-лизина (Pam3CSK4) (100 нг/мл) в течение 4 часов или 1 дня. Стимулированные первичные макрофаги снимали с пластика в 0,2-0,3% ЭДТА в PBS, фиксировали и пермеабилизовывали, как описано в примере 7, а затем инкубировали с зондами FISH, специфичными к мРНК IFNγ и TNFα (таблица 3), и анализировали с помощью проточной цитометрии. Как и в примере 8, фиксированные и промытые клетки инкубировали с Cy5-меченными зондами ДНК, комплементарными к РНК, специфичной к GFP (таблица 3), и анализировали с помощью FC в качестве отрицательного контроля. Частоты клеток, детектированных зондами с помощью анализа FISH-flow, описанного выше, например, в примере 2, показаны на Фиг. 5, где верхний ряд соответствует результатам с зондами GFP, средний ряд - с зондами IFNγ, и нижний ряд - с зондами TNFα. Первая колонка в каждом ряду относится к стимуляции с использованием LPS и IFNγ в течение 4 часов, вторая - к стимуляции с использованием LPS и IFNγ в течение 1 дня, третья - к стимуляции с использованием Pam3CSK4 в течение 4 часов, и четвертая - к стимуляции с использованием Pam3CSK4 в течение 1 дня.
ПРИМЕР 11. КИНЕТИКА ЭКСПРЕССИИ ЦИТОКИНОВ И АКТИВАЦИЯ МАРКЕРОВ В АКТИВИРОВАННЫХ T-КЛЕТКАХ
Данный пример демонстрирует идентификацию временных профилей экспрессии генов цитокинов, хемокинов и рецепторов хемокинов, которые являются маркерами активации T-клеток, в ответ на стимуляцию PMA (25 нг/мл) и иономицином (350 нг/мл) Т-клеток человека, полученных из периферической крови в градиенте фиколла. Более конкретно, после различных периодов стимуляции (2 часа, 1 день, 3 дня) клетки фиксировали и пермеабилизовывали, как описано в примере 7, а затем инкубировали с зондами FISH, специфичными к указанным мРНК (таблица 3), или с mAb, узнающими их белковые продукты, с последующим анализом методом проточной цитометрии. Частоты клеток, детектированных с помощью анализа FISH-flow, описанного выше, например, в примере 2, представлены на Фиг. 6A, где верхний ряд соответствует результатам без стимуляции, а следующие ряды - результатам после стимуляции в течение 2 часов, 1 дня и 3 дней, соответственно. Первая колонка в каждом ряду содержит результаты, полученные с зондами GFP; вторая колонка - при использовании зондов IL-2; третья колонка - при использовании зондов IFNγ; четвертая колонка - при использовании зондов TNFα; пятая колонка - при использовании зондов CCL3; шестая колонка - при использовании зондов CCR7, и последняя колонка - при использовании зондов cFos. Верхний ряд соответствует зондам GFP, средний ряд - зондам IFNγ, и нижний ряд - зондам TNFα. Частоты клеток, обнаруженных при использовании mAb с помощью FC, представлены на Фиг. 6B, где верхний ряд соответствует результатам без стимуляции, а следующие ряды - результатам после стимуляции в течение 2 часов, 1 дня и 3 дней, соответственно. Первая колонка в каждом ряду содержит результаты, полученные для отрицательного контроля - сходного по изотипу (Iso) mAb; вторая колонка - при использовании mAb, узнающего IL-2; третья колонка - при использовании mAb, узнающего IFNγ; четвертая колонка - при использовании mAb, узнающего TNFα.
ПРИМЕР 12. РАЗДЕЛЕНИЕ И АНАЛИЗ АКТИВИРОВАННЫХ, МЕЧЕННЫХ ЗОНДАМИ FISH T-КЛЕТОК В ЭКСПРЕССИРУЮЩИХ И НЕ ЭКСПРЕССИРУЮЩИХ ЦИТОКИНЫ ПОПУЛЯЦИЯХ
Данный пример демонстрирует, что вышеописанный FISH-Flow позволяет выполнять мечение клеток, которые экспрессируют одну или множественные мишени в ответ на стимуляцию, и что такие респондеры могут быть выделены из целой клеточной популяции с целью исследования их профилей экспрессии генов.
Часть I: Разделение на популяции
Т-клетки человека, полученные из периферической крови в градиентах фиколла и глобально стимулированные PMA (25 нг/мл) и иономицином (350 нг/мл), инкубировали в течение 2 часов либо с Cy5-меченными зондами ДНК, комплементарными к РНК, специфичной к GFP, и анализировали с помощью FC как контроль, или со смесью нескольких наборов зондов FISH (таблица 3), реактивных с мРНК IL-2, IFNγ, TNFα и CCL3, причем все зонды одиночно помечены Cy5. Частоты клеток, обнаруженных при использовании зондов с помощью анализа FISH-Flow, описанного выше, например, в примере 2, представлены на Фиг. 7A и Фиг. 7B. Результаты, полученные с зондами GFP, показаны на Фиг. 7A. Результаты, полученные со смесью зондов, показаны на Фиг. 7B. После гейтирования популяции лимфоцитов на основе FSC и SSC (левая панель на каждой фигуре), применяли дополнительное консервативное гейтирование, чтобы разделить проанализированные лимфоциты на две различных популяции (правая панель на каждой фигуре) - цитокин-положительную (обозначенную IFN+) и отрицательную (обозначенную IFN-). Гейтирование, установленное для меченных зондами GFP клеток (Фиг. 7A), определило две популяции: 0,1% IFN+ и 71,2% IFN-, из всех проанализированных лимфоцитов. Популяции, меченные зондами IL-2:IFNγ:TNFα:CCL3 (Фиг. 7B), являлись следующими: 10,2% IFN+ и 33,7% IFN- при таком же гейтировании. Последние, меченные зондами FISH клетки (Фиг. 7B), разделяли на положительные и отрицательные популяции с помощью FACS для выполнения анализа экспрессии генов в выделенных клетках, экспрессирующих и не экспрессирующих цитокины, как описано ниже. Данный подход также успешно применяли в отношении стимулированных T-клеток, меченных только IFNγ-специфичным зондом FISH, которые различали и сортировали на субпопуляции IFN+ и IFN-.
Часть II: Анализ экспрессии генов цитокинов в T-клетках, меченных зондами FISH и сортированных на активированные и нереспондерные популяции
Активированные и нереспондерные T-клетки, полученные и сортированные на отдельные популяции, как описано в части I, анализировали, чтобы продемонстрировать различные профили экспрессии генов, согласующиеся с их реактивностью по отношению к специфичным зондам FISH. Экспрессию мРНК, кодируемых двумя генами, IFNγ и геном "домашнего хозяйства" GAPDH (контроль), выбирали, чтобы проиллюстрировать возможность решения данной задачи. РНК, выделенную из сортированных популяций клеток стандартными методами, подвергали ОТ-ПЦР (обратная транскрипция с последующей амплификацией полученной кДНК с помощью полимеразной цепной реакции) при использовании набора QIAGEN OneStep для ОТ-ПЦР, согласно методике, описанной поставщиком, и при использовании праймеров, специфичных к одному из этих двух генов. Последовательности праймеров для реакций амплификации являлись следующими:
прямой праймер IFNγ: 5' GAGCATCCAAAAGAGTGTGGAG (SEQ ID NO: 36),
обратный праймер IFNγ: 5' TTCATGTATTGCTTTGCGTTGG (SEQ ID NO: 37),
прямой праймер GAPDH: 5' CCAATATGATTCCACCCATGGC (SEQ ID NO: 38),
обратный праймер GAPDH: 5' TCCTGGAAGATGGTGATGGGAT (SEQ ID NO: 39).
После инкубирования при комнатной температуре и первичной денатурации в течение 15 мин при 94°C использовали следующие тепловые циклы: 0,5 мин при 94°C, 0,5 мин при 55°C, 1 мин при 72°C. Выполняли сорок циклов. Обнаружение амплифицированных продуктов осуществляли при использовании красителя SYBR Green. Пороговые циклы реакций амплификации показаны на Фиг. 8A. Для подтверждения результатов амплификации и исключения возможности артефактов, размер идентифицированных ампликонов проверяли при использовании IFNγ- и GAPDH-специфичных праймеров с помощью гель-электрофореза. Результаты показаны на Фиг. 8B.

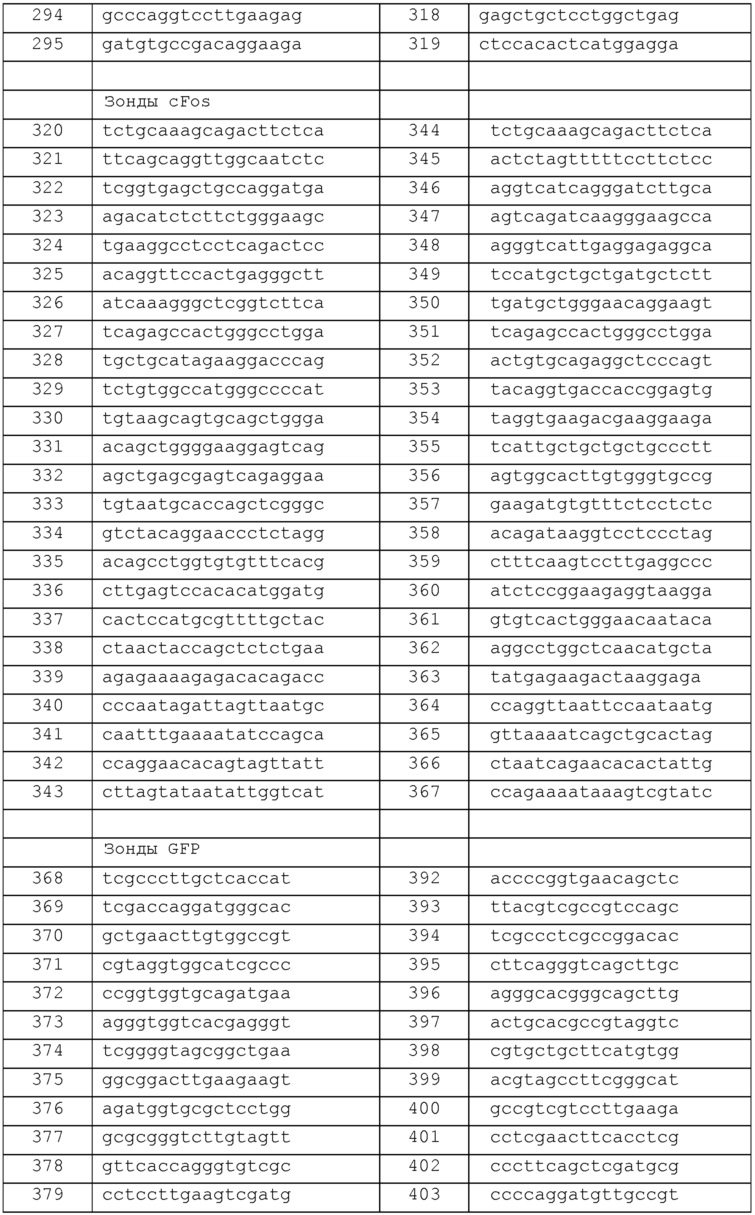

Предшествующие примеры и описание предпочтительных вариантов осуществления следует рассматривать как иллюстрацию, а не как ограничение объема настоящего изобретения, определяемого в соответствии с формулой изобретения. Как будет с легкостью оценено, многочисленные изменения и комбинации признаков, изложенных выше, могут использоваться без отступления от настоящего изобретения, изложенного в формуле изобретения. Такие изменения не следует рассматривать как отступление от объема изобретения, и все такие изменения должны быть включены в объем следующей формулы изобретения. Все ссылки, цитируемые в настоящем описании, включены в настоящую заявку в полном объеме.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ДИАГНОСТИКИ АЛЛЕРГИЧЕСКИХ РЕАКЦИЙ | 2009 |
|
RU2536291C2 |
| АНТИТЕЛА, СПЕЦИФИЧНЫЕ К BTN2, И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2821703C2 |
| СПОСОБЫ ВЫЯВЛЕНИЯ ВОСПАЛИТЕЛЬНОГО ЗАБОЛЕВАНИЯ КИШЕЧНИКА | 2008 |
|
RU2502806C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ ИНФЕКЦИЙ И ОПУХОЛЕЙ | 2007 |
|
RU2540490C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ ИММУННЫХ ЗАБОЛЕВАНИЙ | 2000 |
|
RU2203682C2 |
| СПОСОБ И КОМПОЗИЦИИ ДЛЯ КЛЕТОЧНОЙ ИММУНОТЕРАПИИ | 2012 |
|
RU2688185C2 |
| ЛИГАНД GITR И СВЯЗАННЫЕ С ЛИГАНДОМ GITR МОЛЕКУЛЫ И АНТИТЕЛА И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2004 |
|
RU2369636C2 |
| СВЯЗАННЫЙ С РАКОМ ТОЛСТОГО КИШЕЧНИКА ГЕН ТОМ34 | 2006 |
|
RU2526196C2 |
| СВЯЗАННЫЙ С РАКОМ ТОЛСТОГО КИШЕЧНИКА ГЕН ТОМ34 | 2006 |
|
RU2449020C2 |
| СПОСОБЫ ЛЕЧЕНИЯ РАКА С ПРИМЕНЕНИЕМ АКТИВИРОВАННЫХ T-КЛЕТОК | 2016 |
|
RU2729362C2 |


































































































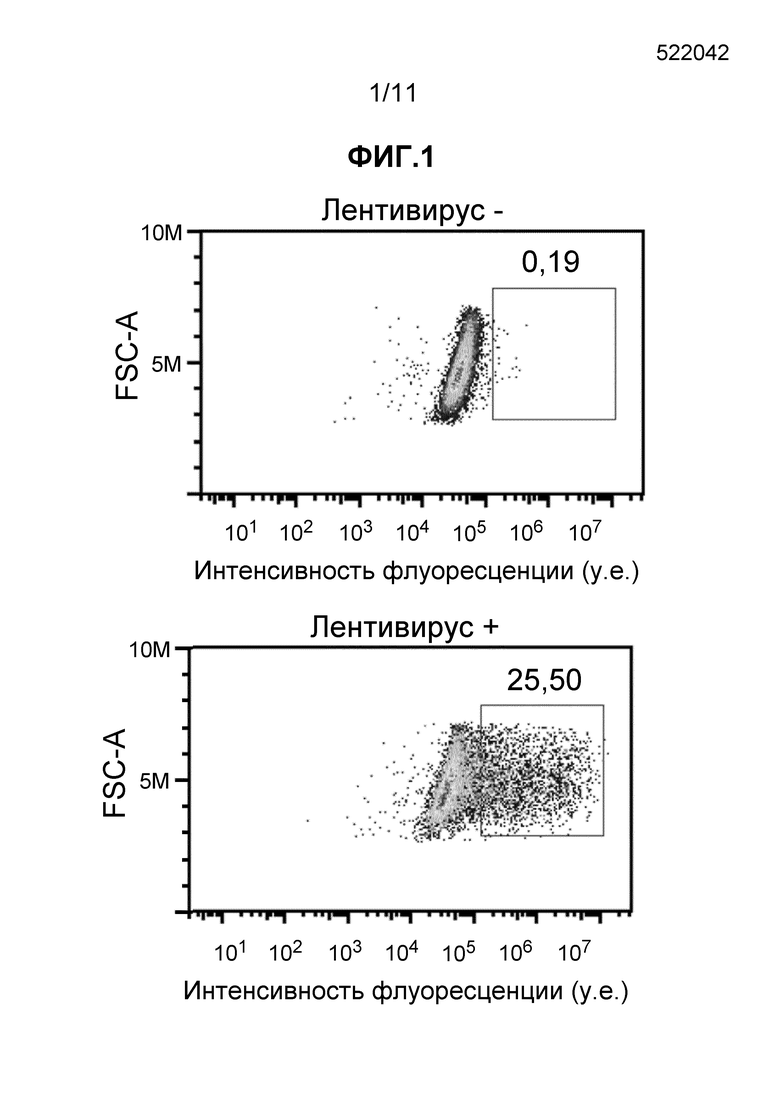
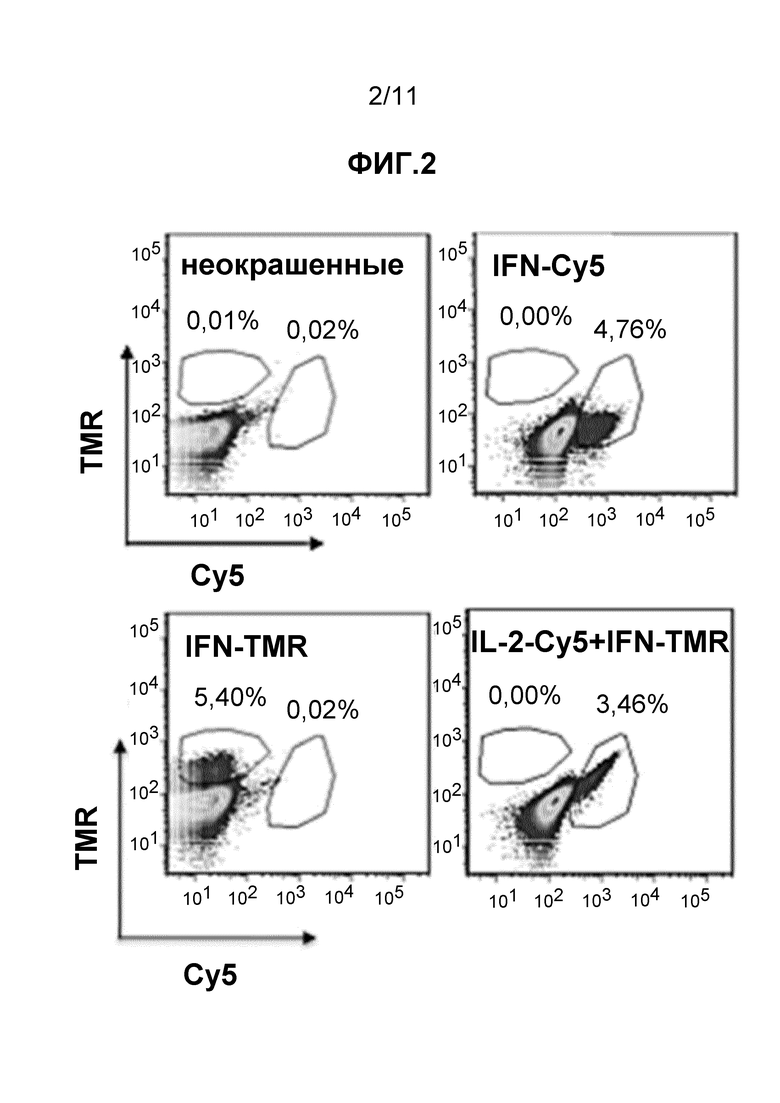
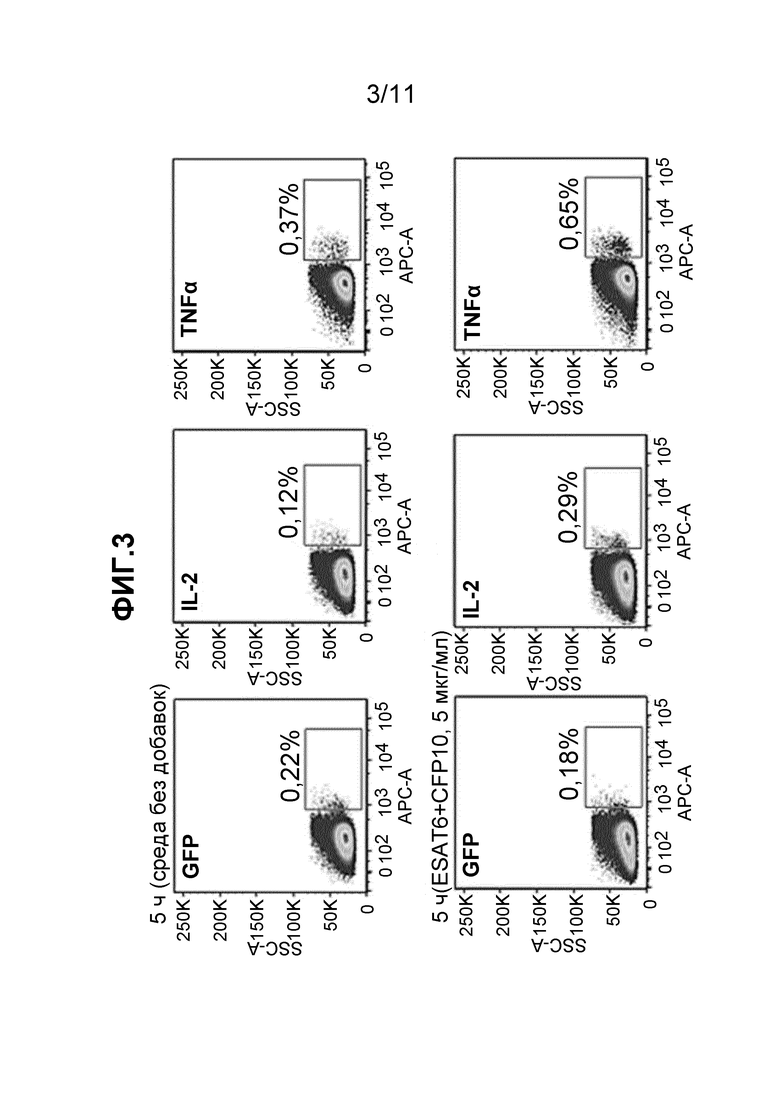
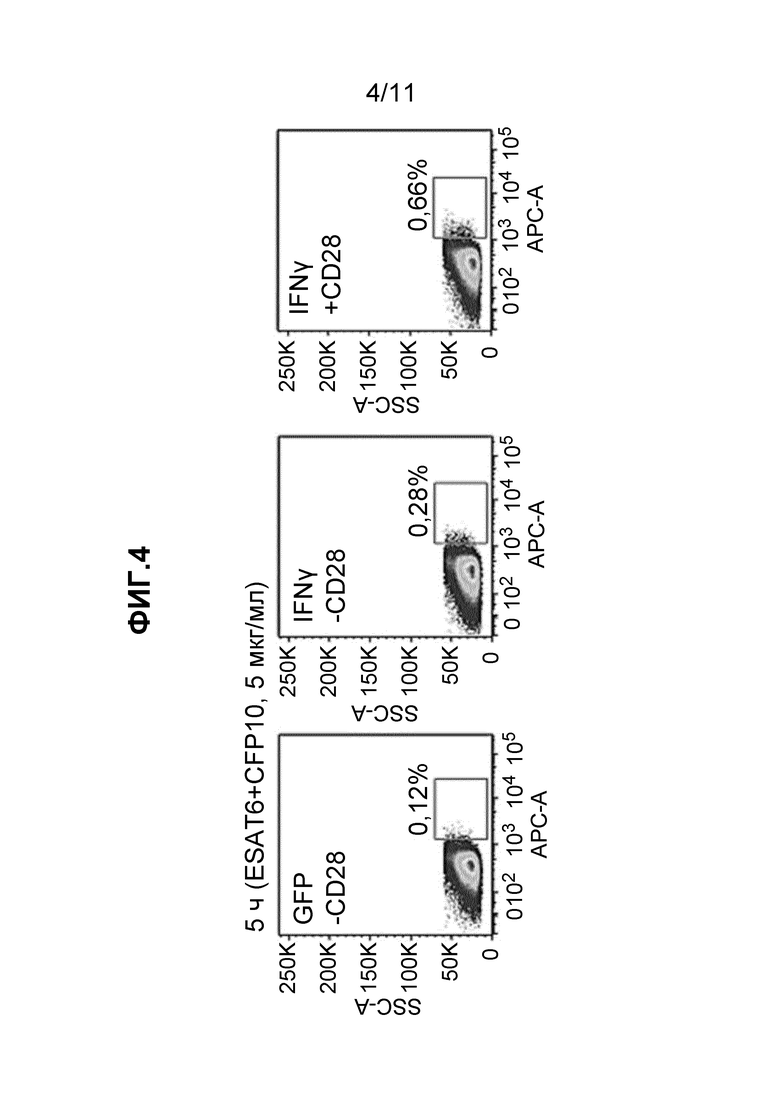
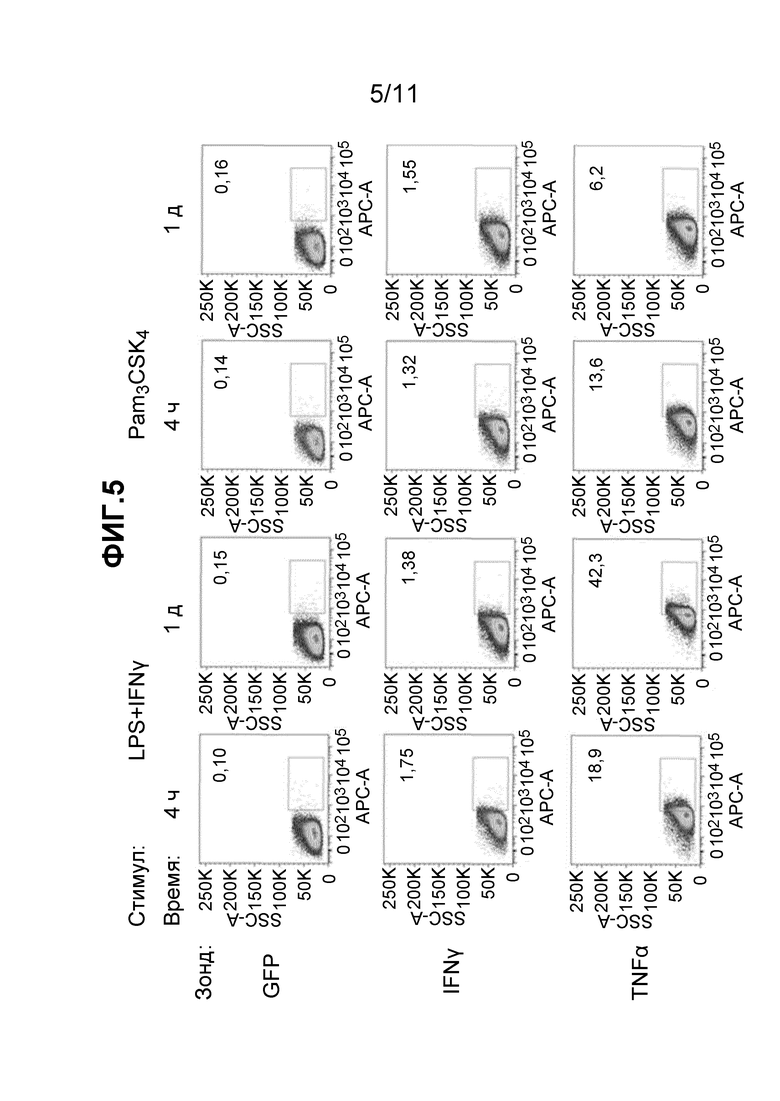
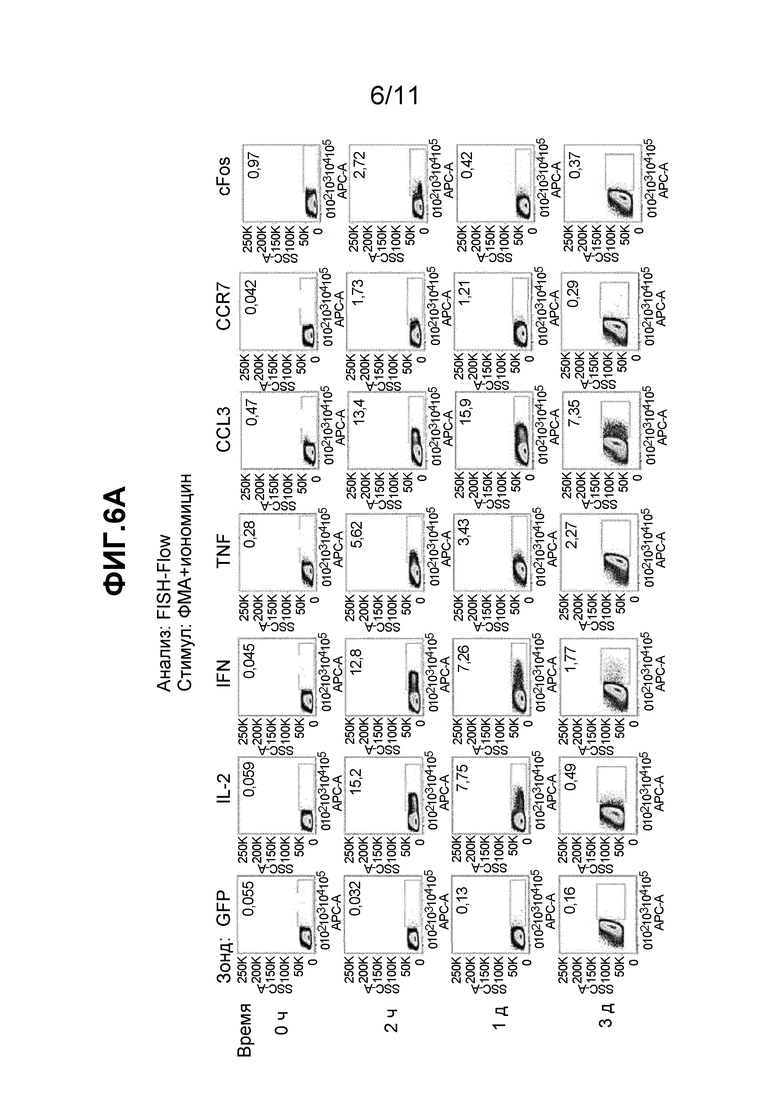

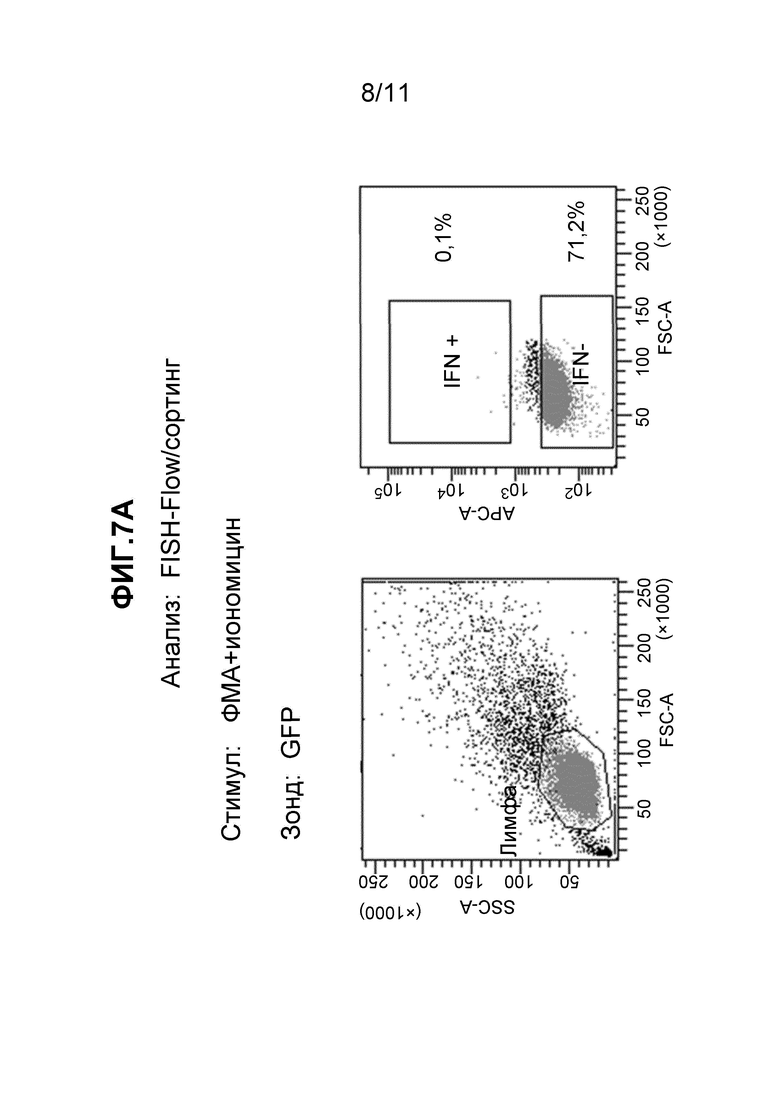
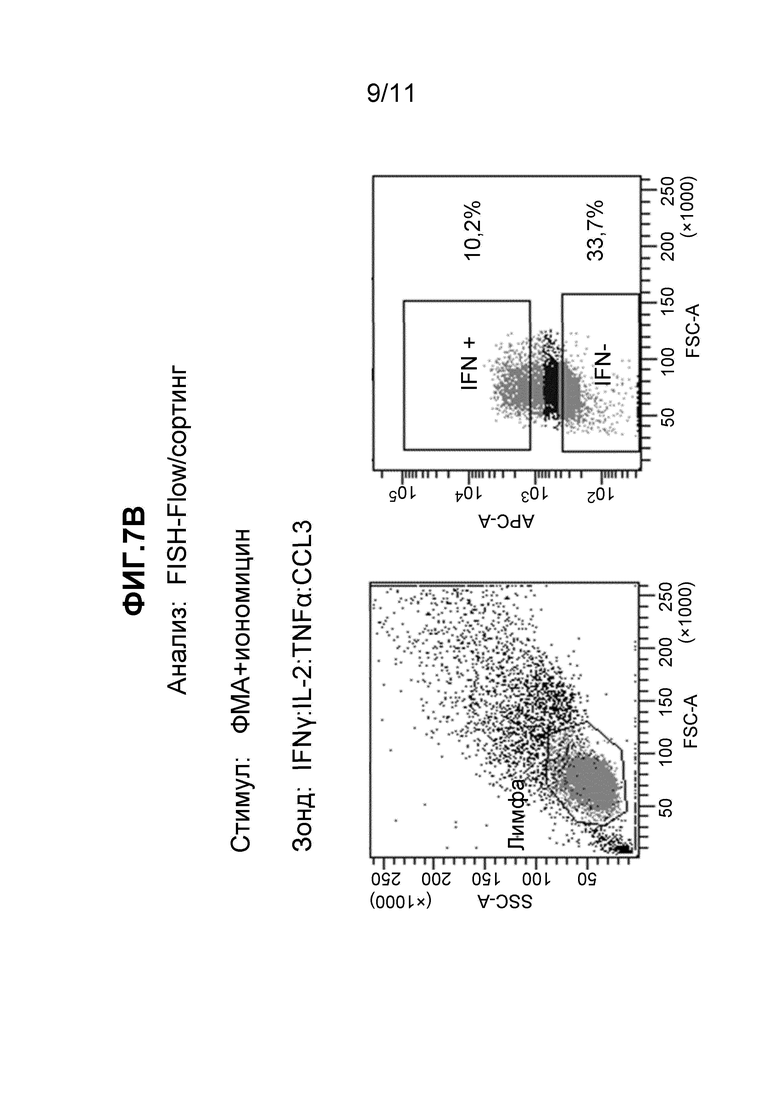

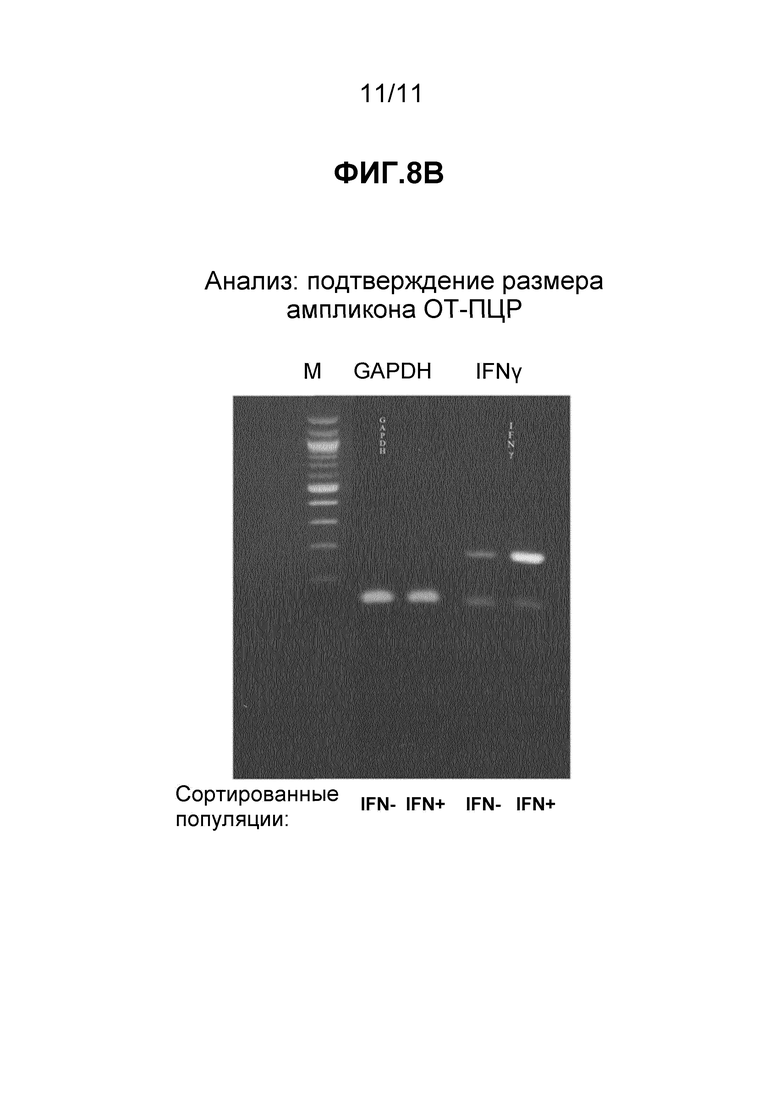
Изобретение относится к биотехнологии. Описан способ анализа активации Т-клеток путем детекции копий по меньшей мере одной молекулы РНК, экспрессируемой в отдельных Т-клетках, включающий предоставление образца, содержащего популяцию клеток, содержащую Т-клетки; индукцию экспрессии генов в Т-клетках ex vivo, либо сразу, либо после культивирования, при инкубировании клеток по меньшей мере с одним соединением, которое связывается специфическими рецепторами и инициирует экспрессию генов; фиксацию и пермеабилизацию клеток; мечение копий по меньшей мере одной молекулы РНК, экспрессируемой Т-клетками, с помощью набора флуоресцентно меченных олигонуклеотидных зондов гибридизации, которые имеют комплементарные последовательности к молекулам РНК, и удаление не связавшихся зондов при промывке; и детекцию клеток с событиями экспрессии с помощью проточной цитометрии (FC), где событие экспрессии является одним или более измерениями флуоресценции, которые группируют с помощью гейтирования интенсивности флуоресценции. Изобретение позволяет быстро обнаружить копии по меньшей мере одной молекулы РНК, экспрессируемой в отдельных клетках. 24 з.п. ф-лы, 8 ил., 3 табл., 12 пр.
1. Способ анализа активации Т-клеток путем детекции копий по меньшей мере одной молекулы РНК, экспрессируемой в отдельных Т-клетках, включающий:
(a) предоставление образца, содержащего популяцию клеток, содержащую Т-клетки;
(b) индукцию экспрессии генов в Т-клетках ex vivo, либо сразу, либо после культивирования, при инкубировании клеток по меньшей мере с одним соединением, которое связывается специфическими рецепторами и инициирует экспрессию генов;
(c) фиксацию и пермеабилизацию клеток;
(d) мечение копий по меньшей мере одной молекулы РНК, экспрессируемой Т-клетками, с помощью набора флуоресцентно меченных олигонуклеотидных зондов гибридизации, которые имеют комплементарные последовательности к молекулам РНК, и удаление не связавшихся зондов при промывке; и
(e) детекцию клеток с событиями экспрессии с помощью проточной цитометрии (FC), где событие экспрессии является одним или более измерениями флуоресценции, которые группируют с помощью гейтирования интенсивности флуоресценции.
2. Способ по п.1, где стадия индукции включает стимуляцию сигнализации T-клеточных рецепторов (TCR) и последующей экспрессии генов с помощью антигенпрезентирующих клеток (APC), которые присутствуют в популяции клеток, или с помощью искусственных APC (aAPC).
3. Способ по п.2, где стимуляция с помощью APC или aAPC индукции происходит в течение периода, выбранного из группы, состоящей из: от 30 минут до 6 часов, от 6 до 24 часов и от 24 до 72 часов.
4. Способ по п.2, где стадия индукции включает костимуляцию с использованием по меньшей мере одного моноклонального антитела (mAb).
5. Способ по п.2, где стадия индукции включает костимуляцию с использованием по меньшей мере одного моноклонального антитела (mAb), которое связывается с aAPC.
6. Способ по п.1, где соединение получено из микроорганизма.
7. Способ по п.1, где соединение включает пептид или смесь пептидов.
8. Способ по п.2, где пептид или смесь пептидов, которые связываются с молекулами MHC класса I или MHC класса II, добавляют к APC или aAPC.
9. Способ по п.8, где пептид или смесь пептидов присутствует в концентрации от 1 до 20 мкг/мл.
10. Способ по п.1, где по меньшей мере одна молекула РНК кодируется геном цитокина.
11. Способ по п.10, где ген кодирует IL-2, TNFα или IFNγ.
12. Способ по п.1, где стимулирующие молекулы являются иммуногенными пептидами, полученными из M. tuberculosis.
13. Способ по п.1, где набор зондов включает от 20 до 60 зондов, каждый из которых однократно помечен одной и той же флуоресцентной молекулой.
14. Способ по п.1, где популяция клеток представляет собой T-клетки, полученные из PBMC человека, по меньшей мере одна молекула РНК кодируется геном цитокина, клетки в популяции клеток индуцируются aAPC с одной или более нагруженными пептидами молекулами MHC, которые взаимодействуют с TCR, и набор зондов включает от 20 до 60 зондов, каждый из которых однократно помечен одной и той же флуоресцентной молекулой.
15. Способ по п.1, где по меньшей мере одну детектированную клетку с событием экспрессии отделяют от клеток, не имеющих событий экспрессии, с помощью сортировки флуоресцентно-активированных клеток.
16. Способ по п.15, где экспрессию генов измеряют в отделенных клетках.
17. Способ по п.16, где измерение экспрессии генов проводят с помощью ОТ-ПЦР или транскриптомного анализа РНК в клетках.
18. Способ по п.1, где стадия индукции включает стимуляцию сигнализации toll-подобных рецепторов и последующей экспрессии генов с помощью микробного продукта или синтетического соединения, где микробный продукт является липидом, гликаном, гликолипидом, сульфолипидом, гликопротеином, белком, пептидом или нуклеиновой кислотой и где стимул присутствует в концентрации в пределах от 1 до 100 нг/мл, от 100 до 1000 нг/мл или от 1 до 20 мкг/мл.
19. Способ по п.18, где стимуляцию проводят в течение периода, выбранного из группы, состоящей из: от 30 минут до 6 часов, от 6 до 24 часов и от 24 до 72 часов.
20. Способ по п.1, где соединение является цитокином.
21. Способ по п.20, где цитокин является цитокином, выбранным из группы, состоящей из IFNγ, IL-2, IL-15 и TNFα.
22. Способ по п.1 или 18, в котором присутствует больше одного соединения.
23. Способ по п.22, где различные соединения подают в разное время.
24. Способ по п.1, где рецепторы выбраны из группы, состоящей из toll-подобных рецепторов, NOD и NOD-подобных рецепторов, сопряженных с G-белком рецепторов, рецепторов полипептидных гормонов, рецепторов цитокинов, B-клеточных рецепторов и T-клеточных рецепторов.
25. Способ по п.1, где популяция включает клетки, выбранные из группы, состоящей из PBMC, лимфоцитов, CD3+CD4+ T-клеток (T-хэлперов 1 или Th1 клеток), CD3+CD4+ T-клеток (T-хэлперов 2 или Th2 клеток), CD3+CD8+ T-клеток, Th17, Treg клеток, NK-клеток, NKT-клеток, макрофагов, дендритных клеток, клеток, формирующих эндотелий и эпителий различных органов тела, и клеток нервной системы.
| Колосоуборка | 1923 |
|
SU2009A1 |
| СОСТАВ ДЛЯ ОБЕЗВОЖИВАНИЯ И ОБЕССОЛИВАНИЯ НЕФТИ | 2000 |
|
RU2186827C2 |
| Жатвенная машина | 1928 |
|
SU13466A1 |
| НОВЫЙ ЦИТОКИН ZALPHA11-ЛИГАНД | 2000 |
|
RU2258710C2 |

