Область техники изобретения
Изобретение относится к слитым полипептидам. В частности, оно относится к слитым полипептидам, содержащим IgE-связывающий домен и компонент человеческого сывороточного альбумина (HSA), и их солям. Изобретение также относится к полинуклеотидам и функционально эквивалентным по физиологической активности полипептидам, которые являются промежуточными продуктами при получении таких слитых полипептидов, их соответствующим рекомбинантным векторам экспрессии, соответствующим прокариотическим и эукариотическим системам экспрессии и способам синтеза слитых полипептидов.
Предпосылки создания изобретения
Взаимодействие между иммуноглобулином Е (IgE) и его рецепторами играет определенную роль в защите от паразитарных инфекций у людей (М. Сарrоn и A. Capron, Science, 264 [1994] 1876-1877). Однако в промышленно развитых странах, в которых санитарно-гигиенические условия превосходят таковые в менее развитых странах, случаи паразитарных инфекций являются более редкими, чем нарушение системы IgE в результате перепроизводства IgE в ответ на аллергены окружающей среды, которые вызывают аллергии и другие болезненные состояния, опосредованные IgE или IgE-рецептором.
IgE представляет собой первичное антитело, участвующее в инициации непосредственной аллергической реакции, и является основным компонентом в поддержании поздней фазы ответа. IgE синтезируется в В-лимфоцитах и проявляет свои действия после связывания с рецептором, обладающим высокой аффинностью к IgE, т.е. FcεRI, который обнаружен на поверхности клеток-эффекторов аллергии, таких как тучные клетки, базофилы и эозинофилы. IgE также обладает индуцирующими функциями благодаря связыванию с его рецептором на антигенпрезентирующих клетках, таких, как клетки Лангерганса, В-клетки и моноциты (G. C. Mudde и др., Allergy, 50 [1995] 193-199).
Аллергическая реакция или состояние проявляется, когда молекулы IgE связываются с поверхностью клеток-эффекторов аллергии через IgE-рецептор, т. е. FcεRI, и становятся перекрестно сшитыми с аллергеном, вызывая тем самым сигнал инициации дегрануляции цитоплазматических гранул в клетках, сопровождаемый выделением медиаторов аллергии, таких как гистамин, серотонин, простагландины и цитокины, и последующий локальный отек ткани и увеличение количества воспаленных клеток. Иными словами, стимуляция аллергии и связанные с ней состояния обусловлены взаимодействием IgE-рецептора, FcεRI, с находящимися в кровотоке аутоантителами к FcεRI.
FcεRI обычно существует в виде тетрамера, включающего α-,β- и две γ-цепи (т. е. "субъединицы"), хотя на моноцитах и клетках Лангерганса β-cубъединица отсутствует.
Было установлено, что IgE-связывающий сайт FcεRI полностью находится внутри его α-субъединицы (обозначен как FcεRIα) (J. Hakimi и др., J. Biol. Chem. , 265 [1990] 22079-22081; U. Blank и др., J. Biol. Chem. 266 [1991] 2639-2646). Установлено далее, что рекомбинантные "knockout"-мыши, у которых генетическим путем удалена вся α-субъединица, не способны поддерживать аллергическую реакцию при контрольном заражении аллергеном (D. Dombrowicz и др., Cell, 75 [1993] 969-976).
FcεRIα представляет собой высоко гликозилированный полипептид с молекулярной массой приблизительно 60 кДа, включающий гидрофобный трансмембранный домен, а также гидрофильный внеклеточный ("экто-") и цитоплазматический домены, которые расположены на наружной поверхности клетки. Кроме того, IgE-связывающая способность FcεRIα обусловлена его внеклеточной областью (J. Hakimi и др. [1990] , см. выше; Leder и др., US 4962035). Представляется вероятным производство растворимой, секретируемой молекулы путем вырезания трансмембранной области и последовательностей, расположенных за ней по ходу транскрипции (С. Ra и др., Int. Immunol. 5 [1993] 47-54), причем полученная в результате этого укороченная последовательность, состоящая в основном из внеклеточного домена FcεRIα человека, обладает IgE-связывающей активностью in vitro и in vivo (M. Haak-Frendscho и др., Immunol. 151 [1993] 351-358 определили остатки аминокислот в положении 1-204 FcεRIα, слитые с укороченной тяжелой цепью С-области IgGl; С. Ra и др. [1993], см. выше, определили остатки 1-172 FcεRIα, соответствующие остаткам 26-197 последовательности, представленной в SEQ.ID.NO.1). Структурными особенностями этого фрагмента являются два потенциальных дисульфидных мостика и семь потенциальных сайтов гликозилирования (M. Haak-Frendscho и др. [1993], см. выше).
Таким образом, укороченные последовательности FcεRIα, состоящие в основном из внеклеточного домена, потенциально могут вводиться в качестве терапевтических агентов млекопитающему для связывания сывороточного IgE с целью предупреждения его связывания со своим высокоаффинным рецептором на клетках-эффекторах аллергии, а также для подавления биосинтеза de novo IgE в лимфоцитах человека (Y. Yanagihara и др., J. Clin. Invest. 94 [1994] 2162-2165).
Однако эффективное использование IgE-связывающего полипептида, такого как внеклеточный домен FcεRIα, для системного лечения аллергических нарушений у млекопитающих, опосредованных IgE или IgE-рецептором, было затруднено из-за его экстремально короткого времени существования in vivo вследствие быстрого клиренса из циркулирующей плазмы. Учитывая, что заболевания, опосредованные IgE или IgE-рецептором, составляют 10-20% от случаев обращения пациентов к врачу, эффективное лечение должно быть весьма ценным для пациентов, страдающих от таких заболеваний, и должно представлять собой важное достижение в клиническом лечении нарушений, опосредованных IgE или IgE-рецептором, таких как аллергия и связанные с аллергией состояния.
Таким образом, представляется важным получить IgE-связывающий полипептид, обладающий пролонгированной эффективной продолжительностью жизни в сыворотке и вследствие этого обладающий улучшенной клинической ценностью для лечения аллергии, в частности для системного лечения атонического дерматита, атонической астмы и хронической крапивницы, а также улучшенной активностью с точки зрения эффективного и относительно недорого способа лечения.
Краткое изложение сущности изобретения
Согласно изобретению было установлено, что путем слияния IgE-связывающего домена и компонента человеческого сывороточного альбумина (HSA) могут быть получены слитые полипептиды, обладающие удлиненным временем полужизни в сыворотке по сравнению с одним IgE-связывающим доменом без потери IgE-связывающей активности, что приводит к образованию IgE-связывающих полипептидов, предложенных для применения в системном лечении аллергии и других, опосредованных IgE заболеваний. Системно введенный IgE-связывающий полипептид должен связываться с IgE сыворотки, а также с находящимися в кровотоке аутоантителами к IgE-рецептору, FcεRIα, препятствуя их связыванию со связанным с клеткой FcεRIα и тем самым препятствуя и/или ингибируя аллергическую реакцию и связанные с ней проявления.
Кроме того, было установлено, что при использовании слитых полипептидов по изобретению, включающих более одного IgE-связывающего домена на молекулу, могут быть достигнуты значительные улучшения IgE-связывающей активности. Например, было обнаружено, что "димерная" молекула по изобретению обладает значительно увеличенной IgE-связывающей активностью по сравнению с "мономером" по изобретению.
Понятие "димер" в контексте настоящего описания относится к слитому полипептиду по изобретению, несущему два IgE-связывающих домена. Понятие "мономер" относится к слитому полипептиду по изобретению, несущему один IgE-связывающий домен. Мономер или димер может быть сконструирован в виде слияния с одним или с множеством HSA-компонентов. Например, мономерный слитый полипептид по изобретению может включать IgE-связывающий домен, слитый либо на своем амино-, либо на карбоксильном конце с HSA-компонентом. В альтернативном варианте мономер может включать IgE-связывающий домен, слитый на своих обоих концах с HSA-компонентами. Димерный слитый полипептид по изобретению может включать, например, два IgE-связывающих домена, слитых через карбоксильный конец одного и аминоконец другого с подлежащим встраиванию HSA-компонентом. В альтернативном варианте димер может содержать в дополнение к его двум IgE-связывающим доменам множество HSA-компонентов.
Кроме того, было установлено, что димерная молекула по изобретению обладает неожиданно высокой активностью.
Таким образом, изобретение относится к слитым полипептидам и их солям, включающим по крайней мере один IgE-связывающий домен, слитый по крайней мере с одним HSA-компонентом; предпочтительно к мономерным слитым полипептидам, в которых один IgE-связывающий домен слит с одним или с несколькими HSA-компонентами, или к многомерным слитым полипептидам, в которых два или несколько IgE-связывающих доменов слиты по крайней мере с одним HSA-компонентом; более предпочтительно к димерам, несущим два IgE-связывающих домена и по крайней мере один HSA-компонент.
Кроме того, изобретение относится к полинуклеотидам, являющимся промежуточными продуктами полипептидов.
Понятие "слитый" или "слияние" в контексте настоящего описания относится к полипептидам, в которых:
(I) данный функционально активный домен (т.е. IgE-связывающий домен) связан на своем карбоксильном конце посредством ковалентной (предпочтительно пептидной, т.е. амидной) связи либо с аминоконцом другого функционально активного домена (т.е. HSA-компонента), либо с пептидом-линкером, который сам связан посредством ковалентной (предпочтительно пептидной) связи с аминоконцом другого функционально активного домена, и/или
(II) данный функционально активный домен (т.е. IgE-связывающий домен) связан на своем аминоконце посредством ковалентной (предпочтительно пептидной, т. е. амидной) связи либо с карбоксильным концом другого функционально активного домена (т. е. HSA-компонента), либо с пептидом-линкером, который сам связан посредством ковалентной (предпочтительно пептидной) связи с карбоксильным концом другого функционально активного домена.
Аналогично этому понятие "слитый", когда оно используется в отношении полинуклеотидных промежуточных продуктов по изобретению, обозначает, что 3'-конец [или 5'-конец] нуклеотидной последовательности, кодирующей первый функционально активный домен, связан соответственно с 5'-концом [или 3'-концом] нуклеотидной последовательности, кодирующей второй функционально активный домен, либо посредством ковалентной связи, либо опосредованно с помощью нуклеотидного линкера, который сам ковалентно связан предпочтительно на своих концах с полинуклеотидом, кодирующим первый функционально активный домен, и с полинуклеотидом, кодирующим второй функционально активный домен.
Предпочтительно слитый полипептид или его соль представляет собой мономер или димер, причем в случае димера он включает два IgE-связывающих домена, слитых с HSA-компонентом, причем первый IgE-связывающий домен слит на своем карбоксильном конце с аминоконцом HSA-компонента, а второй IgE-связывающий домен слит на своем аминоконце с карбоксильным концом HSA-компонента.
Слитые полипептиды по изобретению также могут содержать дополнительные функционально активные домены в дополнение к IgE-связывающим доменам и HSA-компонентам, например полноразмерные или укороченные человеческие протеины (например, растворимые внеклеточные фрагменты), такие как цитокины, или аминоконцевой фрагмент урокиназы. Эти дополнительные функционально активные домены могут сами по себе служить в качестве линкерных пептидов, например, соединяя IgE-связывающий домен с другим IgE-связывающим доменом или с HSA-компонентом; альтернативно этому они могут быть локализованы еще где-либо в слитой молекуле, например на ее амино- или карбоксильном конце.
Примеры слитых полипептидов по изобретению могут быть представлены следующими формулами:
R1-L-R2, (I)
R2-L-R1, (II)
R1-L-R2-L-R1, (III)
R1-L-R1-L-R2, (IV)
R2-L-R1-L-R1, (V)
где R1 обозначает аминокислотную последовательность IgE-связывающего домена,
R2 обозначает аминокислотную последовательность HSA-компонента,
L в каждом случае назависимо обозначает ковалентную (предпочтительно пептидную) связь или обозначает пептид-линкер, который связан посредством ковалентной (предпочтительно пептидной) связи с концом R1 и/или R2,
причем вышеуказанные фрагменты молекул приведены в прямом направлении, т. е. их левый конец соответствует аминоконцу, а правый конец карбоксильному концу молекулы.
Было обнаружено, что уровни IgE-связывания являются высокими, когда слитый полипептид находится под управлением IgE-связывающего домена; это означает, что IgE-связывающий домен содержит N-концевую область зрелого слитого протеина (как в случае соединений формул I, III и IV, см. выше). Особенно предпочтительным является димер вышеприведенной формулы III, т.е. димер, в котором протеиновая область, которая управляет экспрессией, представляет собой область, входящую в IgE-связывающий домен, слитый на своем карбоксильном конце с аминоконцом HSA-компонента, который в свою очередь слит на своем карбоксильном конце с аминоконцом второго IgE-связывающего домена. Если существует более одного фрагмента R1, или более одного фрагмента R2, или более одного фрагмента L, эти фрагменты могут быть идентичны, но это условие не является обязательным.
Предпочтительно слитые полипептиды по изобретению включают по крайней мере один растворимый секретируемый млекопитающим IgE-связывающий домен, слитый по крайней мере с одним HSA-компонентом, и их фармацевтически приемлемые соли.
Кроме того, изобретение относится к способу предупреждения и/или лечения нарушений, опосредованных IgE или IgE-рецептором, в частности аллергических реакций, таких как атопический дерматит, атопическая астма и хроническая крапивница, предусматривающему введение слитых полипептидов по изобретению или их фармацевтически приемлемых солей пациенту, нуждающемуся в таком лечении, а также изобретение относится к фармацевтическим композициям, включающим слитые полипептиды по изобретению или их фармацевтически приемлемые соли вместе по крайней мере с одним фармацевтически приемлемым носителем или разбавителем.
Кроме того, изобретение относится к физиологически активным функциональным эквивалентам слитых полипептидов по изобретению, которые являются промежуточными продуктами при синтезе полипептидов, полинуклеотидам, которые являются промежуточными продуктами при получении слитых полипептидов или их физиологически активным функциональным эквивалентам, как они определены выше, олигонуклеотидным промежуточным продуктам, необходимым для их конструирования, пептидам, кодируемым такими олигонуклеотидами, рекомбинантным векторам для экспрессии слитых молекул, прокариотическим или эукариотическим (прежде всего относящимся к млекопитающим) системам экспрессии и способам получения с помощью таких систем экспрессии слитых полипептидов или их физиологически активных функциональных эквивалентов.
Описание последовательностей SEQ.ID.NO и относящихся к ним понятий SEQ. ID.NO.1: Аминокислотная последовательность доминантной формы полноразмерного нативного человеческого FcεRIα, включающая сигнальную последовательность
Понятие "пре-IgER" относится к остаткам Met1-Leu204 последовательности SEQ. ID. NO. 1. Понятие "IgER" относится к зрелой форме пре-IgE, которая включает остатки Val26-Leu204 последовательности SEQ.ID.NO.1 (т.е. внеклеточного домена FcεRIα).
SEQ.ID.NO.2: Аминокислотная последовательность нативной формы препро-HSA (обозначенная в настоящем описании как "препро-HSA I"), включающая остатки Met1-Leu609
Доминантная форма зрелого нативного протеина (обозначенная в настоящем описании как "HSA I") представлена остатками Asp25-Leu609 последовательности SEQ.ID.NO.2.
Понятие "препро-HSA II" относится к усеченной на одну аминокислоту (Leu609) на карбоксильном конце нативной последовательности и, следовательно, относится к остаткам Met1-Gly608 последовательности SEQ.ID.NO.2.
Зрелая форма препро-HSA II, обозначенная в настоящем описании как "HSA II", представлена остатками Asp25-Leu608 последовательности SEQ.ID.NO.2.
SEQ. ID. NO.3: Аминокислотная последовательность, кодируемая EcoRI-фрагментом плазмиды R-H-R/SK 50, полученной в примере 5, включает последовательность "пpe-IgER", представленную остатками 1-204; линкер AlaSer(Gly)4Ser (обозначенный далее в настоящем описании как "L1"), представленный остатками 205-211, последовательность HSA II, представленную остатками 212-795, линкер (Gly)3Ser (обозначенный далее в настоящем описании как "L2"), представленный остатками 796-799, и последовательность "IgER", представленную остатками 800-978.
Зрелый димерный слитый полипептид по изобретению, обозначенный в настоящем описании как "IgER-L1-HSA II-L2-IgER" или в альтернативном варианте как "IgER-HSA-IgER-димep", экспрессируемый клетками СНО согласно способу, описанному в примере 7, имеет аминокислотную последовательность Val26-Leu978 последовательности SEQ.ID.NO.3.
SEQ. ID. NO. 4: Нуклеотидная последовательность EcoRI-фрагмента плазмиды R-H-R/SK 50 из примера 5, включающая полинуклеотидную последовательность, кодирующую "пpe-IgER", в положениях 10-621, олигонуклеотид, кодирующий L1, в положениях 622-642, полинуклеотид, кодирующий HSA II, в положениях 643-2394, олигонуклеотид, кодирующий L2, в положениях 2395-2406, и полинуклеотид, кодирующий "IgER", в положениях 2407-2943, с двумя стоп-кодонами в положениях 2944-2949.
Сайты рестрикции на концах кодирующих фрагментов и в областях линкеров находятся в положениях 1-6, 622-627, 637-642, 2387-2393, 2401-2406 и 2950-2955. Последовательность Kozak представляет собой нуклеотидную последовательность в положениях 7-9.
Точковые мутации, отличающиеся от консенсусной нуклеотидной последовательности HSA, находятся в положениях 804, 1239, 1290, 1446, 1815, 2064 и 2079. Поскольку точковые мутации находятся в незначимом положении, они не оказывают влияния на аминокислотную последовательность.
SEQ.ID.NO.5: Нуклеотидная последовательность доминантной формы нативного препро-HSA, соответствующая фиг.12 и аминокислотной последовательности SEQ. ID.NO.2
SEQ. ID.NO.6: Нуклеотидная последовательность доминантной формы полноразмерного нативного человеческого FcεRIα. включающая сигнальную последовательность и соответствующая фиг.13 и аминокислотной последовательности SEQ. ID.NO.1
Описание чертежей
На приложенных чертежах указанные молекулы находятся в прямом положении, т. е. левый конец соответствует аминоконцу (или 5'-концу), а правый конец соответствует карбоксильному концу (или 3'-концу).
Фиг. 1А-В: Период полужизни в сыворотке мышей: Изменение во времени (10-800 мин после инъекции) концентрации (в пикомолях протеина на мл сыворотки) in vivo
(А) свободного протеина HSA I и
(Б) IgER-протеина (обозначенного как "свободная альфа-цепь") и IgER-L1-HSA II-L2-IgER-димерного слитого полипептида (обозначенного как "IgER-HSA-IgER-димep"), полученного из СНО-клеток, как описано в примере 7.
(В) Период полужизни в сыворотке свободного HSA I по фиг.1А в сравнении с периодом полужизни димерного слитого полипептида ("IgER-HSA-IgER") по фиг. 1Б путем нормирования на 1 по отношению к концентрациям в сыворотке через 10 мин после инъекции.
Фиг. 2: Экстравазация, являющаяся результатом пассивной кожной реакции анафилаксии у мышей, которым вводят внутривенно серийные разведения (10, 50 или 500 мкг/кг) полипептида IgER-L1-HSA II-L2-IgER ("IgER-HSA-IgER-димер"), полученного согласно примеру 7 (контрольная группа получала концентрацию 0 мкг/мл); площадь в мм2; интервалы 5, 15 и 30 мин до сенсибилизации с помощью внутрикожной инъекции антителом в виде моноклонального мышиного IgE к динитрофенилу (ДНФ) и последующего контрольного заражения раствором ДНФ-бычий сывороточный альбумин, содержащим 1% красителя Эванса голубого.
Фиг.3: Схематическое изображение трех слитых полипептидов по изобретению (два мономера и один димер), также приведены полипептиды-линкеры:
I. Мономеры
(1) Мономер с лидирующим HSA, содержащий HSA II (обозначенный на чертежах как "HSA"), слитый через L2 ("GGGS") с IgER. Нуклеотиды, кодирующие остатки Leu607Gly608 HSA II, как установлено, содержат уникальный сайт MstII, а нуклеотиды, кодирующие остатки GlySer L2, содержат сайт BamHI (кодируемые аминокислоты подчеркнуты).
(2) Мономер с лидирующим IgE-связывающим доменом, содержащий IgER, слитый на своем карбоксильном конце через L1 (т. е. "ASGGGGS") с HSA II (обозначен на чертеже как "HSA"). Олигонуклеотид, кодирующий L1, содержит сайт NheI и BamHI соответственно (кодируемые аминокислоты AlaSer и GlySer L1 подчеркнуты).
II. Димер
Димер с лидирующим IgE-связывающим доменом, содержащий первый IgER, слитый на своем карбоксильном конце через L1 с аминоконцом HSA II (обозначен на чертеже как "HSA"), карбоксильный конец которого слит со вторым IgER через L2, с сайтами рестрикции в кодирующем полинуклеотиде, как описано выше для мономера.
Фиг. 4: ПЦР-праймеры для укорачивания полноразмерной кДНК человеческого FcεRIα (обозначена далее как кДНК IgE-рецептора) с получением ДНК, кодирующей:
(I) IgER
[сайт BamHI добавлен к 5'-концу кодирующей цепи для FcεRIα с помощью олигонуклеотида 18, а стоп-кодон и сайты EcoRI и SalI добавлены к 3'-концу некодирующей цепи с помощью олигонуклеотида 19],
(II) npe-IgER
[олигонуклеотид 20, добавляющий сайты SstI, EcoRI и Kozak к 5'-концу цепи, кодирующей FcεRIα, олигонуклеотид 31, добавляющий сайты NotI и NheI (и удаляющий второй сайт NheI) в некодирующей цепи FcεRIα],
(III) npe-IgER
[олигонуклеотиды 20 и 19, применяемые в соответствии с описанным выше]:
(А) "конструкция с лидирующим HSA": субклонирование (I) (т.е. IgER), указанного выше, в векторе SK с получением клона рЕК1 клонирующего вектора ТА, использованного для конструирования мономера с лидирующим HSA II,
(Б) "конструкция с лидирующим IgER": субклонирование (II) (т.е. пре-IgER) в векторе SK с получением конструкции IgER/TA 1, применяемой для получения мономера с лидирующим IgER,
(В) "только IgER": субклонирование (III) (т.е. пре-IgER) в векторе SK с получением конструкции IgER/Fl/TA 34, применяемой для экспрессии зрелого IgER, который используется в качестве стандарта.
Двойными звездочками обозначены два стоп-кодона.
Фиг. 5: Пары ПЦР-праймеров для укорачивания полноразмерной кДНК человеческого препро-HSA I с получением кДНК, кодирующей:
(I) препро-HSA II, слитый на карбоксильном конце с L2
[олигонуклеотид 24, добавляющий сайты SpeI, EcoRI и последовательность Kozak к 5'-концу кодирующей цепи, олигонуклеотиды 28 и 29, добавляющие линкер, кодирующий сайты MstII и HindIII на 3'-конце HSA II],
(II) L1, слитый с 5'-концом HSA II
[олигонуклеотид 26, добавляющий сайты NotI, NheI, линкер и BamHI к кодирующей цепи, олигонуклеотид 27, содержащий сайт NcoI в некодирующей цепи],
(III) препро-HSA I
[олигонуклеотид 24, применяемый в соответствии с описанным выше, и олигонуклеотид 25, добавляющий стоп-кодон, сайты EcoRI и HindIII к 3'-концу некодирующей цепи]:
(А) "конструкция с лидирующим HSA": субклонирование (I) в векторе SK с получением рЕК7, использованного для конструирования мономера с лидирующим HSA II,
(Б) "конструкция с лидирующим IgER": субклонирование (II) в векторе SK, с получением конструкции HSA/SK 5, применяемой для получения мономера с лидирующим IgER,
(В) "только HSA": субклонирование (III) в векторе SK с получением конструкции HSA/SK 17, кодирующей зрелый протеин нативного человеческого сывороточного альбумина (т.е. HSA I), который используется в качестве стандарта.
Фиг. 6: Олигонуклеотиды 1-10, 12, 16, 17, 22 и 30 для секвенирования человеческого сывороточного альбумина (HSA). применяемые для секвенирования материалов, клонированных в ТА и полученных после связывания с IgE-связывающими фрагментами.
Фиг. 7: Олигонуклеотиды 11, 13 и 23 для секвенирования IgE-рецептора, применяемые для секвенирования фрагментов, клонированных в ТА и полученных после связывания с HSA-компонентом.
Фиг.8:
(А) Олигонуклеотиды 14 и 15 для введения мутаций в человеческий сывороточный альбумин;
(Б) ПЦР- и линкерные олигонуклеотиды 18-20, 24-29 и 31 для конструирования слитых полипептидов.
Фиг. 9: Конструирование вектора HSA-IgER/SK 49, включающего полинуклеотид, кодирующий препро-HSA II (обозначен на чертеже как "HSA"), слитый на 3'-конце через олигонуклеотид, кодирующий линкер L2 (представленный на фиг.9 "GGG"), с 5'-концом полинуклеотида, кодирующего IgE (обозначен на чертеже как "IgER"), путем лигирования BamHI,SalI-фрагмента вектора рЕК1 с BamHI,SalI-фрагментом рЕК7.
Фиг. 10: Конструирование вектора IgER-HSA/SK 1, включающего полинуклеотид, кодирующий пре-IgER (обозначен на чертеже как "IgER"), слитый на 3'-конце через олигонуклеотид, кодирующий линкер L1 (представленный как "GGG"), с 5'-концом полинуклеотида, кодирующего HSA II ("HSA"), путем лигирования SstI, NheI-фрагмента IgER/TA 1 с SstI-NheI фрагментом HSA/SK 5.
Фиг.11: Конструирование вектора HSA-IgER Pst Sal/SK 37, включающего полинуклеотид, кодирующий HSA II (обозначен на чертеже как "HSA3"), слитый на 3'-конце через олигонуклеотид, кодирующий линкер L2 (не показан), с 5'-концом полинуклеотида, кодирующего IgER (обозначен на чертеже как "IgER"), путем лигирования PstI, SalI-фрагмента HSA-IgER/SK 49 с вектором SK. Также показано конструирование вектора R-H-R/SK 50, включающего полинуклеотид, кодирующий пре-IgER (обозначен на чертеже как "IgER"), слитый на 3'-конце через олигонуклеотид, кодирующий линкер L1 (не показан), с 5'-концом полинуклеотида, кодирующего HSA II ("HSA"), который в свою очередь слит через олигонуклеотид, кодирующий L2 (не показан), с полинуклеотидом, кодирующим IgER ("IgER"). Вектор получают лигированием РstI,КрnI-фрагмента вектора HSA-IgER Pst Sal/SK 37 с PstI, KpnI-фрагментом вектора IgER-HSA/SK l.
Фиг.12: Нуклеотидная последовательность человеческого сывороточного альбумина и аминокислотная последовательность, соответствующие SEQ.ID.NO.2 и SEQ.ID.NO.5.
Фиг. 13: Нуклеотидная последовательность IgER и аминокислотная последовательность, соответствующие SEQ.ID.NO.1 и SEQ.ID.NO.6.
Фиг.14: Нуклеотидная и аминокислотная последовательности EcoRI-фрагмента вектора R-H-R/SK 50, соответствующие последовательностям SEQ.ID.NO.3 и SEQ. ID. NO. 4, кодирующим димерный слитый протеин R1-HSA-R1. Последовательность HSA обозначена курсивом. Линкерные последовательности обозначены маленькими буквами. Точковые мутации, отличающиеся от консенсусной нуклеотидной последовательности HSA, приведены жирным шрифтом маленькими буквами. Поскольку точковые мутации находятся в незначимом положении, они не оказывают влияния на аминокислотную последовательность. Сайты рестрикции на концах фрагментов и в линкерной области подчеркнуты.
Фиг. 15: Данные ДСН-ПААГ-электрофореза очищенного зрелого слитого полипептида из примера 7.
Общее количество, внесенное в гель: 8 мкг (полоса 1), 6 мкг (полоса 2), 4 мкг (полоса 3) и 2 мкг (полоса 4); стандарты молекулярной массы: 97,4, 66,2, 45; 31, 21,5 и 14,4 кДа (полоса 5).
Подробное описание изобретения
Изобретение относится к слитым полипептидам и их солям, включающим по крайней мере один IgE-связывающий домен, слитый по крайней мере с одним компонентом HSA.
Понятие "IgE-связывающий домен" относится к аминокислотной последовательности, обладающей способностью связываться или каким-либо иным образом взаимодействовать с IgE млекопитающего, например с IgE человека, таким образом, чтобы препятствовать связыванию IgE со своим рецептором, FcεRI.
кДНК и установленная аминокислотная последовательность человеческого FcεRIα являются известными (J. Kochan и др., Nucleic Acids Research 16 [1988] 3584; Leder и др., US 4962035). FcεRIα человека кодирует NH2-концевой сигнальный пептид [аминокислотные остатки (включая первый Met) Met1-Ala25], два иммуноглобулинподобных внеклеточных домена (остатки Val26-Leu204), гидрофобную трансмембранную область (Gin205-Ile224) и гидрофильный цитоплазматический хвост (остатки Ser225-Asn257) (относительно SEQ.ID.NO.1). Сигнальный пептид отщепляется во время внутриклеточного процессинга. Полноразмерная аминокислотная последовательность доминантной формы нативного человеческого FcεRIα включена в настоящее описание в виде SEQ.ID.NO.1.
"IgER" в контексте настоящего описания относится к аминокислотной последовательности Val26-Leu204 последовательности SEQ.ID.NO.1 (кодирующей внеклеточный домен), а понятие "пpe-IgER" относится к остаткам Met1-Leu204 последовательности SEQ.ID.NO.1 (кодирующей сигнальную последовательность, расположенную против хода транскрипции от внеклеточного домена).
IgE-связывающий домен представляет собой, например, аминокислотную последовательность, обладающую по крайней мере 80%-ной, более предпочтительно по крайней мере 85%-ной, еще более предпочтительно по крайней мере 95%-ной или наиболее предпочтительно по крайней мере 99%-ной гомологией с IgER, как она определена выше (гомологию рассчитывают как % идентичности между нативной последовательностью и измененной последовательностью в зависимости от общей длины последовательности нативной молекулы после линеаризации последовательностей и при необходимости введения брешей для достижения максимального процента идентичности последовательностей).
В подгруппе слитых полипептидов по изобретению компонент IgE-связывающего домена представляет собой
(Ха)
где (Ха) обозначает:
(а) IgER или
(б) встречающийся в естественных условиях аллель IgE, или
(в) укороченную на карбоксильном конце на 1-12 (например, на 1-10, 1-7 или 1-4) аминокислот последовательность (а) или (б),
(г) вариант (а), (б) или (в).
Под понятием "вариант" понимают:
- последовательность (а), (б) или (в), модифицированную с помощью одной или нескольких делеций (которые отличаются от последовательностей, укороченных на карбоксильном конце) в общей сложности до 10 (например, 1-7 или 1-5) аминокислот,
- инсерции в общей сложности до 10 аминокислот (например, 1-5), которые встроены внутрь аминокислотной последовательности, или в общей сложности до 100 аминокислот, которые присоединены к любому ее концу, или
- консервативные замещения в общей сложности до 15 (например, 1-5) аминокислот.
IgE-связывающий домен предпочтительно представляет собой
(Хb)
где (Хb) обозначает:
(а) IgER,
(б) укороченную на карбоксильном конце на 1-7 аминокислот последовательность IgER,
(в) вариант (а) или (б).
Еще более предпочтительно IgE-связывающий домен представляет собой
(Хс)
где (Хс) обозначает:
(а) IgER,
(б) укороченную на карбоксильном конце на 1-7 аминокислот последовательность IgE (например, последовательность Val26-Ala197 относительно SEQ.ID. NO.1).
Наиболее предпочтительно IgE-связывающий домен представляет собой IgER.
Предпочтительно любую гомологичную, укороченную последовательность или вариант получают методом рекомбинантной ДНК в качестве "секретируемого" полипептида, а именно зрелая последовательность пептида, кодирующего IgE-связывающий домен, может секретироваться из клетки-хозяина, в которой он (или его форма-предшественник, такая как преформа) синтезируется из полинуклеотида.
Вторая основная составляющая рекомбинантных слитых полипептидов по изобретению представляет собой человеческий сывороточный альбумин (HSA), компонент, который является наиболее широко распространенным в плазме протеином, на долю которого приходится 60 мас.% относительно всего содержания протеинов в плазме. Молекула человеческого сывороточного альбумина состоит из одной негликозилированной полипептидной цепи, состоящей из 585 аминокислот с молекулярной массой 66,5 кДа. Характерной особенностью альбумина является сложная структура его дисульфидного мостика (F.F. Clerc и др., J. Chromatogr. 662 [1994] 245-259). Человеческий сывороточный альбумин широко распространен в организме, в частности в компартментах кишечника и крови, где он в основном присутствует как наиболее широко распространенный протеин сыворотки, поддерживая осмотическое давление и объем плазмы. Кроме того, он медленно выводится печенью, а время его полужизни в организме человека in vivo составляет 14-20 дней (Т.A. Waldmann "Albumin Structure, Function and Uses", Pergamon Press [1977] 255-275). Человеческий сывороточный альбумин не обладает ферментативной или иммунологической функцией. Он является естественным носителем, участвующим в эндогенном транспорте и доставке различных природных, а также терапевтических молекул (Н. Lu и др., FEBS Lett. 356 [1994] 56-59; WO 93/15199; ЕР 648499; P. Yeh и др., PNAS USA 89 [1992] 1904-1908; ЕР 413622).
Человеческие клетки синтезируют сывороточный альбумин первоначально в форме препро-полипептида. Сигнальная последовательность из 18 аминокислот (включая первый Met) удаляется, когда протеин проходит через полость эндоплазматического ретикулума, но при этом еще сохраняется 6 аминокислот на N-конце (наиболее часто Arg-Gly-Val-Phe-Arg-Arg), которые затем подвергаются протеолитическому отщеплению во время или сразу после транспорта через секреторный аппарат.
Хорошо известно, что человеческий сывороточный альбумин является полиморфным (D. C. Carter и J.X. Но, Adv. Prot. Chem. 45 [1994] 153-203). Например, альбумин Наскапи имеет Lys372 вместо Glu372, проальбумин Кристчерча имеет измененную про-последовательность, т.е. Glu24 вместо Arg24 (Lattа и др., US 5100784).
Полная аминокислотная последовательность доминантной формы встречающегося в естественных условиях протеина, обозначенная в настоящем описании как "препро-HSA I", известна (A. Dugaiczyk и др., PNAS USA 79 [1982] 71-75) (SEQ. ID. NO. 2). Доминантная форма зрелого протеина ("HSA I") представлена Asp25-Leu609 последовательности SEQ.ID.NO.2.
HSA-носитель слитого полипептида по изобретению может иметь аминокислотную последовательность, обладающую по крайней мере 80%-ной, более предпочтительно по крайней мере 85%-ной, еще более предпочтительно по крайней мере 95%-ной и наиболее предпочтительно по крайней мере 99%-ной гомологией с остатками 25-609 SEQ. ID. NO. 2 (т.е. HSA I) (гомологию рассчитывают как % идентичности между нативной последовательностью и измененной последовательностью в зависимости от общей длины последовательности нативной молекулы после линеаризации последовательностей и при необходимости введения брешей для достижения максимального процента идентичности последовательностей).
В подгруппе слитых полипептидов по изобретению HSA-компонент представляет собой
(Ya)
где (Ya) представляет собой:
(а) HSA I, или
(б) встречающийся в естественных условиях аллель (а), или
(в) укороченную на карбоксильном конце на 1-10 (например, на 1-5 или на 1 или 2) аминокислот последовательность (а) или (б), или
(г) укороченную последовательность (а), заканчивающуюся на аминокислотном остатке n, где n обозначает 369-419 и прежде всего обозначает 373, 387, 388, 389, 390 и 407 (как описано в US 5380712), или
(д) вариант (а), (б), (в) или (г).
Под понятием "вариант" понимают:
- последовательность (а), (б), (в) или (г), модифицированную с помощью одной или нескольких делеций (которые отличаются от последовательностей, укороченных на карбоксильном конце) в общей сложности до 10 (например, 1-7 или 1-5) аминокислот,
- инсерции в общей сложности до 10 (например, 1-5) аминокислот, встроенные внутрь аминокислотной последовательности, или в общей сложности до 100 аминокислот, присоединенных к любому ее концу, или
- консервативные замещения в общей сложности до 15 (например, 1-5) аминокислот.
HSA-компонент более предпочтительно представляет собой
(Yb)
где (Yb) представляет собой:
(а) HSA I, или
(б) встречающийся в естественных условиях аллель (а), или
(в) укороченную на карбоксильном конце на 1-10 (например, на 1-5 или на 1 или 2) аминокислот последовательность (а) или (б), или
(г) вариант (а), (б) или (в).
HSA-компонент еще более предпочтительно представляет собой
(Yc),
где (Yc) представляет собой:
(а) HSA I, или
(б) укороченный на своем карбоксильном конце на 1-10 (например, на 1) аминокислот HSA I (примером такой укороченной последовательности является "HSA II", которая имеет аминокислотную последовательность, представленную Asp25-Gly608 последовательности SEQ.ID.NO.2).
В еще более предпочтительной подгруппе HSA-компонент представляет собюй HSA I или HSA II, предпочтительно HSA II.
Особенно предпочтительным является слитый полипептид из примера 7, в частности без лидерной последовательности, т.е. полипептид Val26-Leu978 SEQ. ID.NO.3.
Предпочтительно любая гомологичная, укороченная последовательность или вариант нативного HSA, применяемые для получения слитого протеина по изобретению, должны быть лишены ферментативной функции и не должны препятствовать связыванию IgE-связывающего домена с сывороточным IgE или ингибировать такое связывание. Также предпочтительно, чтобы время полужизни в сыворотке любого такого гомолога или варианта составляло по крайней мере 14 дней.
Относительно либо IgE-связывающего домена, либо HSA-компонента понятие "консервативные замещения" относится к замещению одной или нескольких аминокислот другими, имеющими аналогичные свойства таким образом, чтобы можно было ожидать, что по крайней мере вторичная структура и предпочтительно третичная структура полипептида практически не изменялись. Например, типичные примеры таких замещений включают замену аланина или валина на глицин, аспарагина на глутамин, серина на треонин и аргинина на лизин. Все эти аминокислоты (кроме глицина) предпочтительно представляют собой встречающиеся в естественных условиях L-аминокислоты.
Предпочтительно каждый IgE-связывающий домен обладает по крайней мере 95%-ной гомологией с IgER, а каждый HSA-компонент обладает по крайней мере 95%-ной гомологией с HSA I.
В других предпочтительных подгруппах:
- каждый IgE-связывающий домен предпочтительно представляет собой (Ха), а каждый HSA-компонент представляет собой (Ya), более предпочтительно (Yb) или еще более предпочтительно (Yc), как определено выше,
- каждый IgE-связывающий домен более предпочтительно представляет собой (Хb), а каждый HSA-компонент представляет собой (Ya), более предпочтительно (Yb) или еще более предпочтительно (Yc), как определено выше,
- каждый IgE-связывающий домен более предпочтительно представляет собой (Хс), а каждый HSA-компонент представляет собой (Ya), более предпочтительно (Yb) или еще более предпочтительно (Yc).
В еще более предпочтительном варианте IgE-связывающий домен представляет собой IgER, а HSA-компонент обозначает (Ya), более предпочтительно (Yb) или еще более предпочтительно (Yc), как определено выше, а в наиболее предпочтительном варианте обозначает HSA I или HSA II, в частности HSA II.
Указанные выше предпочтительные варианты также относятся к полипептидам каждой из формул I, II, III, IV и V, которые представлены в настоящем описании.
Предпочтительными полипептидами по изобретению являются таковые, включающие первую молекулу IgER, которая слита через свой карбоксильный конец с аминоконцом молекулы HSA II, а эта молекула HSA II слита на своем карбоксильном конце с аминоконцом второй молекулы IgER (IgER представляет собой остатки Val26-Leu204 последовательности SEQ.ID.NO.1, а HSA II представляет собой остатки Asp25-Gly608 последовательности SEQ.ID.NO.2).
Особенно предпочтительные полипептиды по изобретению включают димеры формулы III (см. выше), в которых каждый R1 обозначает IgER, a R2 обозначает HSA II. Особенно предпочтительными являются димеры формулы III, в которых каждый символ L обозначает 1-25 аминокислот.
Любой пептидный линкер (обозначенный через "L" в формулах I-V в настоящем описании) предпочтительно независимо определяет складчатость и активность IgE-связывающего домена, причем он лишен способности определять упорядоченную вторичную структуру, которая могла бы взаимодействовать с IgE-связывающим доменом или вызывать иммунологическую реакцию у пациента, и имеет минимальные характеристики гидрофобности или заряда, которые могли бы взаимодействовать с IgE-связывающим доменом.
Пептидный линкер предпочтительно состоит из 1-500 аминокислот, более предпочтительно из 1-250 и еще более предпочтительно из 1-100 (например, из 1-25, 1-10, 1-7 или 1-4) аминокислот. Линкер предпочтительно является линейным, т.е. неразветвленным. Как правило, можно ожидать, что пептидные линкеры, включающие Gly, Ala и Ser, удовлетворяют критериям, предъявляемым к линкеру. Например, линкеры по настоящему изобретению включают GlyGlyGlySer ("L1" в настоящем описании) и AlaSerGlyGlyGlyGlySer ("L2" в настоящем описании). Также могут использоваться линкеры различных других длин и состава последовательностей.
Изобретение также относится к олигонуклеотидам, кодирующим пептидные линкеры по изобретению. Такие олигонуклеотиды должны быть "слиты в рамке считывания" с полинуклеотидами, кодирующими IgE-связывающий домен и HSA-компонент, и предпочтительно включают сайты рестрикции, уникальные для данной молекулы. Под понятием "слиты в рамке считывания" подразумевают, что:
(1) отсутствует сдвиг в рамке считывания IgE-связывающего домена или HSA-компонента, вызванный олигонуклеотидом линкера, и
(2) отсутствует терминация трансляции между рамками считывания между IgE-связывающим доменом и HSA-компонентом.
Изобретение также относится к физиологически активным функциональным эквивалентам слитых полипептидов по изобретению, которые, как правило, являются промежуточными продуктами при синтезе новых полипептидов. Понятие "физиологически активный функциональный эквивалент" предпочтительно относится к более крупной молекуле, включающей слитый полипептид по изобретению, к которому была добавлена такая аминокислотная последовательность, которая является необходимой или желательной для эффективной экспрессии и секреции зрелого рекомбинантного слитого полипептида по изобретению из определенной клетки-хозяина. Такая добавленная последовательность обычно находится на аминоконце зрелого полипептида и обычно содержит лидерную (т.е. сигнальную) последовательность, которая служит для направления их метаболизма по пути секреции и, как правило, отщепляется после или до секреции полипептида из клетки. Сигнальная последовательность может происходить из природной N-концевой области соответствующего полипептида или может быть получена из генов хозяина, кодирующих секретируемые протеины, или она может происходить из любой последовательности, для которой известно, что она усиливает секрецию представляющего интерес полипептида, включая синтетические последовательности и все комбинации между "пре-" и "про-" областью. Стык между сигнальной последовательностью и последовательностью, кодирующей зрелый полипептид, должен соответствовать сайту расщепления в хозяине.
В слитых полипептидах, в которых IgE-связывающий домен "управляет" экспрессией, т. е. расположен против хода транскрипции относительно других кодирующих последовательностей в слитой молекуле, целесообразно использовать лидерную последовательность нативного человеческого FcεRIα (т.е. Met1+Ala2-Ala25 последовательности SEQ.ID.NO.1), и этот вариант был успешно использован для получения зрелого полипептида в системе экспрессии млекопитающего (например, СНО, COS), а также из дрожжей (Pichia pastoris).
Примером физиологически активного функционального эквивалента зрелого димерного слитого полипептида IgER-L1-HSA II-L2-IgER является пре-IgER-L1-HSA II-L2-IgE. Однако дополнительная лидерная последовательность не обязательно представляет собой таковую из FcεRIα человека, а может быть получена из любого приемлемого источника при условии, что она пригодна для воздействия на экспрессию/секрецию зрелого полипептида конкретной клеткой-хозяином. Например, препро-последовательность HSA также может применяться для получения вышеуказанных димерных слитых полипептидов, в частности для экспрессии в дрожжах.
В слитых полипептидах по изобретению, в которых HSA-компонент управляет экспрессией, пригодная лидерная последовательность может представлять собой нативную лидерную последовательность. Например, нативная препро-область HSA может применяться для осуществления секреции зрелого гетерологичного полипептида из клеток млекопитающего (например, СНО, COS) или дрожжей (Pichia pastoris).
Примером физиологически активного функционального эквивалента зрелого слитого полипептида HSA II-L2-IgER является пpeпpo-HSA II-L2-IgER. Однако эффективную экспрессию зрелого слитого протеина в определенных хозяевах могут обеспечивать и другие лидерные последовательности, не обязательно являющиеся нативными для HSA или для других клеток-хозяев (US 5100784, US 5330901).
Изобретение также относится к полинуклеотидам, которые являются промежуточными продуктами при получении рекомбинантных слитых полипептидов по изобретению, включая полинуклеотиды, кодирующие слитые полипептиды или их физиологически активные функциональные эквиваленты, и олигонуклеотиды, кодирующие линкерные пептиды, например, показанные на фиг.8.
Еще одним объектом изобретения является способ получения рекомбинантного слитого полипептида, как он определен выше, или его соли, который включает:
(а) трансформацию клетки-хозяина вектором, включающим ДНК, кодирующую слитый полипептид, как он определен выше, или его физиологически активный функциональный эквивалент,
(б) экспрессию слитого полипептида или его физиологически активного функционального эквивалента в этой клетке, причем физиологически активный функциональный эквивалент полипептида модифицируют (например, путем отщепления сигнальной последовательности) с получением слитого полипептида, как он определен выше, и
(в) выделение образовавшегося в результате полипептида из клетки-хозяина, предпочтительно в виде секретируемого продукта, необязательно в форме его соли.
Еще одним объектом настоящего изобретения являются векторы, предпочтительно плазмиды, предназначенные для экспрессии слитых полипептидов. Эти векторы включают ДНК, кодирующую указанные выше полинуклеотиды или их физиологически активные функциональные эквиваленты. В целом пригодные векторы, которые могут трансформировать микроорганизмы, способные экспрессировать слитые полипетиды, включают векторы экспрессии, которые содержат нуклеотидные последовательности, кодирующие слитые полипептиды, соединенные с регуляторными последовательностями транскрипции и трансляции, такими как промоторы, которые все вместе образуют кассету экспрессии. Промоторы могут происходить из генов конкретного применяемого хозяина, или такие контрольные области могут быть модифицированы, например, in vitro путем сайтнаправленного мутагенеза, путем интродукции дополнительных контрольных элементов или синтетических последовательностей. Таким образом, кассета экспрессии, применяемая в настоящем изобретении, также включает область терминации транскрипции и трансляции, которая является функционально активной в требуемом хозяине и которая расположена на 3'-конце последовательности, кодирующей гибридную макромолекулу. Помимо кассеты экспрессии вектор должен включать один или несколько маркеров, позволяющих осуществлять селекцию трансформированного хозяина. К таким маркерам относятся маркеры, обусловливающие устойчивость к антибиотикам, такие как G418. Эти гены устойчивости могут быть помещены под контроль соответствующих сигналов транскрипции и трансляции, позволяющих осуществлять экспрессию в данном хозяине.
Более конкретно рекомбинантные слитые полипептиды по изобретению можно получать, например, следующим путем.
А. Конструирование векторов экспрессии слитого протеина
Первой стадией при конструировании рекомбинантных слитых полипептидов является субклонирование частей слитых полипептидов в клонирующих векторах. В контексте данного описания понятие "клонирующий вектор" относится к молекуле ДНК, такой как плазмида, космида или бактериофаг, которая может автономно размножаться в прокариотической клетке-хозяине. Клонирующие векторы обычно содержат один или небольшое число сайтов, распознаваемых рестрикционными эндонуклеазами, по которым чужеродные последовательности ДНК могут быть встроены определенным образом без потери необходимой биологической функции вектора, а также маркерный ген, который можно применять для идентификации и селекции клеток, трансформированных клонирующим вектором. Маркерные гены обычно представляют собой гены, придающие устойчивость к тетрациклину или устойчивость к ампициллину. Пригодные клонирующие векторы описаны в Molecular Cloning: A Laboratory Manual, под ред. J. Sambrook и др., 2-е изд. (Cold Spring Harbour Laboratory Press [1989] и могут быть получены, например, из различных источников.
Клон кДНК FcεRIα pGEM-3-110B-1 описан у A. Shimizu и др., Рrос. Nat. Acad. Sci. USA 85 [1988] 1907-1911 и может быть получен из Американской коллекции типовых культур тканей (АТСС штамм 67566). Одноцепочечная кДНК из печени человека может быть приобретена у фирмы Clontech (клон кДНК для ПЦР [PCR-ready Quick Clone cDNA], Cat. D 7113-1). Последовательность HSA может быть получена из GenBank под регистрационными номерами VOO495, JOOO78, LOO132, LOO133. кДНК HSA может быть получена с помощью ПЦР-амплификации с использованием олигонуклеотидов 24 и 25, как описано в примере 2.
Полинуклеотид, кодирующий пригодный IgE-связывающий домен, или ДНК HSA-компонента могут быть получены с помощью полимеразной цепной реакции (ПЦР). В ПЦР используется одноцепочечная кДНК в качестве матрицы и смесь олигонуклеотидных праймеров. ПЦР проводят по хорошо известной методике (см., например, C.R.M. Bangham, "The Polymerase Chain Reaction: Getting Started" в Protocols in Human Molecular Genetics, Human Press [1991], глава I, стр. 1-8). Наборы для ПЦР и материалы, предназначенные для использования в этих наборах, также поставляются в продажу различными поставщиками. Наборы и методы для их применения, кроме того, описаны в US 5487993.
Последовательности ДНК, кодирующие сигнальные пептиды, могут быть добавлены с помощью ПЦР или при необходимости с помощью синтетических олигонуклеотидов, которые кодируют известные сигнальные пептидные последовательности. Последовательности ДНК, кодирующие гетерологичный сигнальный пептид, субклонируют в рамке считывания с последовательностями ДНК, кодирующими N-конец слитого полипептида.
Слияние полинуклеотидов можно проводить путем субклонирования в промежуточных векторах. В альтернативном варианте один ген может быть клонирован непосредственно в векторе, содержащем другой ген. Для соединения последовательностей ДНК могут применяться линкеры и адаптеры.
Субклонирование проводят в соответствии с общепринятыми методами, например с использованием расщепления рестрикционным ферментом с получением пригодных концов, используя обработку щелочной фосфатазой для предупреждения нежелательного соединения молекул ДНК, и лигирования соответствующими лигазами. Такие методы описаны в литературе и известны в данной области.
Б. Экспрессионное клонирование слитых полипептидов
Клонированный слитый протеин затем отщепляют от клонирующего вектора и встраивают в вектор экспрессии. Пригодные векторы экспрессии, как правило, содержат
(1) элементы прокариотической ДНК, кодирующие начало репликации ДНК в бактериях, и маркер селекции по признаку устойчивости к антибиотику для обеспечения роста и отбора вектора экспрессии в бактерии-хозяине,
(2) элементы эукариотической ДНК, контролирующие инициацию транскрипции, такие как промотор, и
(3) элементы ДНК, контролирующие процессинг транскриптов, такие как последовательность терминации транскрипции/полиаденилирования.
Таким образом, еще одним объектом настоящего изобретения являются векторы, предпочтительно плазмидные векторы, которые предназначены для использования при экспрессии слитых полипептидов по изобретению и которые содержат полинуклеотидные последовательности, представленные в данном описании, которые кодируют слитые полипептиды по изобретению. Приемлемые векторы экспрессии, которые могут трансформировать прокариотические или эукариотические клетки, включают векторы экспрессии, которые содержат нуклеотидные последовательности, кодирующие слитые молекулы, соединенные с регуляторными последовательностями транскрипции и трансляции, которые выбирают в зависимости от используемых клеток-хозяев. Например, для экспрессии в Е. coli могут применяться такие векторы, как описанные у M.W. Robertson, J. Biol. Chem. 268 [1993] 12736-12743. Ожидается, что продукт будет связан дисульфидным мостиком, но не будет гликозилирован. Для экспрессии в дрожжах может применяться такой вектор экспрессии, как pHIL-D2, поставляемый фирмой Invitrogen (San Diego, CA, США), набор для экспрессии в Pichia pastoris (регистрационный номер К1710-01). Ожидается, что протеиновый продукт будет связан дисульфидным мостиком и будет гликозилирован. Другие пригодные системы экспрессии дрожжей включают Saccharomyces cerevisiae и Kluveromyces lactis. Для экспрессии в бакуловирусах, заражающих клетки насекомых, пригодны такие векторы, как рАС360, pVL1392 и pVL1393 (фирма Invitrogen, San Diego, CA, США), которые, как ожидается, должны секретировать гликозилированный и связанный дисульфидным мостиком продукт.
Слитый полипетид по изобретению, как правило, представляет собой гликопротеин, в частности при экспрессии в клетках млекопитающих, и изобретение включает слитые полипептиды в любом состоянии гликозилирования или связывания дисульфидными мостиками. В частности, анализ мутаций позволяет предположить, что N-связанное гликозилирование в первом, втором и седьмом положениях N-связанных сайтов гликозилирования α-цепи рецептора IgE (A. Shimizu и др. , PNAS USA 85 [1988] 1907-1911, фиг.2) (что соответствует аминокислотным остаткам 46, 67 и 191 SEQ.ID.NO.1) усиливает биологическую активность молекулы IgER и содержащих ее мономеров и димеров. Наиболее предпочтительной системой экспрессии является система, в которой любые добавленные сахара должны быть максимально схожи с сахарами в нативной молекуле, из которой получен полипептид. Известно, что клетки дрожжей и насекомых модифицируют гликопротеины по-иному, нежели клетки млекопитающих, в то время как Е. coli не добавляет молекулы сахара после секреции. Таким образом, в то время как экспрессия в любой из этих систем экспрессии может привести к получению протеинового продукта, который пригоден для применения в диагностике (например, для обнаружения аллергического состояния), наиболее предпочтительной формой для экспрессии этого продукта для применения в качестве терапевтической молекулы является экспрессия в клетках млекопитающих или возможно экспрессия в молоке трансгенных млекопитающих.
Для экспрессии в млекопитающем-хозяине источниками регуляторных сигналов транскрипции и трансляции, применяемых в кассете экспрессии, могут служить вирусные источники, такие как аденовирусы, вирус папилломы быков или вакуолизирующие обезьи вирусы, в которых регуляторные сигналы связаны с определенным геном, который имеет высокий уровень экспрессии. Приемлемые регуляторные последовательности транскрипции и трансляции также могут быть получены из генов млекопитающих, таких как гены актина, коллагена, миозина и металлотионеина. Регуляторные последовательности транскрипции включают промоторную область, достаточную для обеспечения инициации синтеза РНК в клетке млекопитающего. Примерами типичных клеток млекопитающих являются клетки яичника китайского хомячка (СНО), SV40-трансформированные клетки почки обезьяны (COS), HeLa, BHK, клетки эмбриона мыши NIH Swiss, фибробласты крысы, обезьяны или человека или клетки гепатомы крысы.
Предпочтительным методом экспрессии в клетках млекопитающего является секреция клетками СНО. Ни СНО-, ни человеческие клетки не выделяют α-1,3-связанные остатки галактозы, которые характерны для экспрессии в мышиных клетках, таких как С127- и SP2/0-клетки. Антитела к этой сахарной связи присутствуют в человеческой сыворотке [C.F. Goochee и др., BioTechnol. 9, [1991] 1347-1355] и могут оказывать влияние на время полужизни, доступность и клиренс рекомбинантных продуктов, экспрессируемых этими мышиными клетками, dhfr-клетки СНО являются мутантными по гену дигидрофолатредуктазы (DHFR) и, следовательно, не способны синтезировать de novo пурины, тимидин и глицин. Число копий интегрированных в хромосому плазмид, несущих копии дикого типа гена DHFR, может быть увеличено или амплифицировано при экспозиции клеток, трансформированных этими плазмидами, с целью увеличения уровней метотрексата, аналога фолата, который конкурирует с фолатом при связывании с активным сайтом фермента. Пригодным вектором экспрессии в dhfr-клетках СНО является рМТ2 (R.J. Kaufman и др., EMBO J. 6 [1987] 187-193). Вектор рМТ2 имеет копию гена DHFR дикого типа, которая транскрибируется как одна мРНК с чужеродным геном. Таким образом, при обработке трансформированных dhfr-клеток СНО повышенными концентрациями метотрексата совместно амплифицируются и чужеродный ген и ген DHFR. Ожидается, что продукт, секретируемый из этих клеток, должен быть связан дисульфидным мостиком и быть гликозилированным, что характерно для млекопитающих.
Вектор экспрессии может быть интродуцирован в клетки-хозяева с помощью различных способов, включая трансфекцию фосфатом кальция, опосредованную липосомами трансфекцию, электропорацию и т.п. Предпочтительно трансфектированные клетки отбирают и размножают, при этом вектор экспрессии стабильно интегрируется в геном клетки-хозяина для
образования стабильных трансформантов. Клетки могут культивироваться, например, в среде DMEM (среда Игла, модифицированная по методу Дульбекко). Полипептид, секретируемый в среду, можно выделять стандартными биохимическими методами после непродолжительной экспрессии через 24-72 ч после трансфекции клеток или после создания стабильных линий клеток после селекции, например, по признаку устойчивости к антибиотику.
Общепринятые методы выделения экспрессированного полипептида из культуры включают фракционирование содержащей полипептид части культуры с помощью хорошо известных биохимических методов. Для выделения экспрессированных протеинов, образовавшихся в культуре, могут применяться, например, такие методы, как гель-фильтрация, гель-хроматография, ультрафильтрация, электрофорез, ионообменная или аффинная хроматография, которые известны для фракционирования протеинов. Кроме того, могут применяться общепринятые иммунохимические методы, такие как основанные на иммуноаффинности и иммуноабсорбции. Методы трансформации, культивирования, амплификации, скрининга и получения и очистки продукта являются хорошо известными (см., например, J. Sambrook и др., [1989], см. выше; R.J. Kaufman, Genetic Engineering: Principles and Methods, Plenum Press, том 9, [1987] 156-198).
Слитые полипетиды по изобретению могут применяться в терапии для лечения млекопитающих, в частности человека, например больных, страдающих аллергией. IgE-связанный домен слитых полипетидов конкурирует за IgE с IgE-рецептором, который присутствует в естественных условиях на поверхности тучных клеток, базофилов и клеток Лангерганса, при этом IgE связывается с введенным протеином и теряет способность связываться с этими клетками-эффекторами аллергии, что могло бы опосредовать аллергическую реакцию. IgE-связывающий домен также конкурирует с IgE-рецептором, FcεRI, путем связывания с аутоантителами к FcεRI.
Таким образом, настоящее изобретение относится к фармацевтической композиции, предназначенной для конкурентного связывания IgE (или антител к FcεRI) и/или для ингибирования производства IgE и, следовательно, предназначенной для суппресивного и/или превентивного лечения болезней, опосредованных IgE или IgE-рецептором, в частности для лечения аллергии и состояний, связанных с аллергией, таких как атонический дерматит, атопическая астма и хроническая крапивница.
Под "болезнями, опосредованными IgE или IgE-рецептором" понимают болезни, обусловленные взаимодействием с находящимся на клетках IgE-рецептором, FcεRI, с IgE или с аутоантителами к FcεRI, например типичные аллергические реакции (синдром "гипер-IgE"), такие, как бронхиальная астма, атопическая астма, сенная лихорадка, аллергия на пыльцу, аллергический ринит, атопический дерматит, экзема, анафилаксия и хроническая крапивница, а также неаллергическая болезнь Кимура и другие легочные, дерматологические или аутоиммунные заболевания.
В частности, атопический дерматит, являющийся одним из наиболее выраженных проявлений атопии, представляет собой хроническое воспалительное заболевание кожи, связанное с высокими уровнями IgE в сыворотке и сопровождающееся сенсибилизацией к различным аллергенам окружающей среды (М.-А. Morren и др. , J. Am. Acad. Dermatol. 31 [1994] 467-473). Современное лечение атопического дерматита основано на использовании кремов, содержащих стероиды, а серьезные случаи атопического дерматита успешно лечились циклоспорином А (Н. Granland и др., Br. J. Dermatol. 132 [1995] 106-112), однако побочные эффекты ограничивают такое лечение, делая его возможным лишь для небольшого числа пациентов. Агент, который ингибирует функции IgE, такие как дегрануляция тучных клеток, и опосредованную IgE презентацию антигена В-клетками и другими антигенпрезентирующими клетками, может оказаться наиболее пригодным из всех известных в настоящее время методов лечения атопического дерматита и, кроме того, также может оказаться пригодным для лечения других более легких форм аллергии.
Кроме атопического дерматита и атопической астмы полипептиды по изобретению могут оказаться пригодными для лечения или предупреждения хронической крапивницы (ХК), в возникновении которой определенную играет роль дегрануляция тучных клеток в результате активации FcεRIα. Слитые полипептиды по изобретению могут очищать кровь от аутоантител к FcεRIα. не затрагивая моноклональные антитела к IgE, которые, наоборот, предложены для лечения болезни.
Полипептиды могут применяться в форме фармацевтической композиции, включающей полипептид по изобретению, предпочтительно немодифицированный полипептид, или его фармацевтически приемлемую соль в фармацевтически приемлемом носителе или разбавителе.
Понятие "соль" относится, в частности, к фармацевтически приемлемым солям, полученным из фармацевтически приемлемых, нетоксичных кислот с образованием кислотно-аддитивных солей, например, с аминогруппой полипептидной цепи, или полученным из фармацевтически приемлемых нетоксичных оснований с образованием солей присоединения оснований, например с карбоксильной группой полипептидной цепи. Такие соли могут быть получены в виде внутренних солей и/или солей с амино- или карбоксильным концом полипептида по изобретению.
Приемлемыми кислотно-аддитивными солями являются соли фармацевтически приемлемых нетоксичных органических кислот, полимерных кислот или органических кислот. Примерами приемлемых органических солей являются уксусная, аскорбиновая, бензойная, бензолсульфоновая, лимонная, этансульфоновая, фумаровая, глюконовая, глутаминовая, бромистоводородная, соляная, изотионовая, молочная, малеиновая, яблочная, миндальная, метансульфоновая, слизевая, азотная, щавелевая, памовая, пантотеновая, форфорная, салициловая, янтарная, серная, винная и пара-толуолсульфоновая, а также полимерные кислоты, такие как дубильная кислота или карбоксиметилцеллюлоза. Приемлемые неорганические кислоты включают минеральные кислоты, такие как соляная, бромистоводородная, серная, фосфорная и азотная кислоты.
Примерами приемлемых неорганических оснований для образования солей с карбокисильной группой являются соли щелочных металлов, такие как соли натрия, калия и лития, соли щелочно-земельных металлов, таких как соли кальция, бария и магния, и соли аммония, меди, железистые и железные соли, соли цинка, алюминия, марганцовистые и марганцовые соли. Предпочтительными являются соли аммония, кальция, магния, калия и натрия. Примерами фармацевтически приемлемых органических оснований, пригодных для образования солей с карбоксильной группой, являются органические амины, такие как триметиламин, триэтиламин, три(н-пропил)амин, дицилогексиламин, β-(диметиламино)этанол, трис(гидроксиметил)аминометан, триэтаноламин, β-(диэтиламино)этанол, аргинин, лизин, гистидин, N-этилпиперидин, гидрабамин, холин, бетаин, этилендиамин, глюкозамин, метилглюкамин, теобромин, пурины, пиперазины, пиперидины, кофеин и прокаин.
Кислотно-аддитивные соли полипептидов могут быть получены общепринятым способом путем взаимодействия полипептида с одним или несколькими эквивалентами требуемой неорганической или органической кислоты, такой как соляная кислота. Соли с карбоксильными группами пептида могут быть получены общепринятым способом в результате взаимодействия пептида с одним или несколькими эквивалентами соответствующего основания, такого как основание в виде гидроксида металла, например гидроксида натрия, основания в виде карбоната или бикарбоната металла, такого как карбонат натрия или бикарбронат натрия, или основания в виде амина, такого как триэтиламин или триэтаноламин.
Таким образом, изобретение относится к фармацевтическим композициям, включающим новый слитый полипептид, как он определен выше, или его фармацевтически приемлемую соль вместе с фармацевтически приемлемым носителем или разбавителем. Носитель предпочтительно представляет собой стерильную, не содержащую пирогена, приемлемую для парентерального введения жидкость. В частности, для инъецируемых растворов предпочтительными жидкими носителями или разбавителями (если раствор является изотоничным) являются вода, физиологический раствор, водная декстроза и гликоли. Композиция может быть изготовлена общепринятым методом в виде лиофилизата.
Композиции могут вводиться системно, т. е. парентерально (например, внутримышечно, внутривенно, подкожно или внутрикожно), или внутрибрюшинно. Для парентерального введения слитые полипептиды предпочтительно должны обладать достаточной растворимостью в сыворотке или плазме пациента, например по крайней мере 1 мг полипептида должно растворяться в 1 мл сыворотки или плазмы.
Композиции также могут вводиться известными способами, предназначенными для местного введения. Примеры приемлемых дозируемых форм включают спреи, глазные растворы, назальные растворы и мази. Например, спрей может быть изготовлен растворением пептида в приемлемом растворителе и загрузкой его в пульверизатор, который служит в качестве аэрозоля, который обычно применяется в ингаляционной терапии. Глазной или назальный раствор может быть изготовлен растворением действующего вещества в дистиллированной воде, добавлением любого необходимого вспомогательного вещества, такого как буфер, раствор, создающий изотоничность, наполнитель, консервант, стабилизатор, поверхностно-активное вещество или антисептик, и доведением значения рН смеси до 4-9. Мази могут быть изготовлены, например, путем приготовления композиции из раствора полимера, такого как 2%-ный водный карбоксивиниловый полимер, и основания, такого как 2%-ный гидроксид натрия, смешения с получением геля и смешения с гелем определенного количества очищенного слитого полипептида.
Композиции предпочтительно вводят подкожно или внутривенно, наиболее часто подкожно.
Изобретение также относится к способу лечения аллергических состояний, включающему введение терапевтически эффективного количества слитого полипептида по изобретению или его фармацевтически приемлемой соли пациенту, нуждающемуся в таком лечении. На практике такой способ лечения применяют при аллергическом болезненном состоянии (т.е. когда специально требуется ослабление симптомов), либо в качестве непрерывного или профилактического лечения (т.е. до начала проявления признаков болезни, опосредованной IgE или IgE-рецептором, такой как аллергическая реакция).
Эффективная доза согласно изобретению может варьироваться в широких пределах, например, в зависимости от степени и серьезности состояния, подлежащего лечению, возраста, пола и состояния пациента, продолжительности лечения и активности определенного слитого протеина, т.е. факторов, которые определяются общепринятыми методами.
Поскольку содержание IgE (а также антител к FcεRI) значительно варьируется у отдельных индивидуумов, терапевтически эффективная доза согласно изобретению предпочтительно должна в 5•102-1•104 раз превышать на молекулярном уровне содержание в сыворотке IgE и антител к FcεRI. Пациенту могут вводиться однократные или многократные суточные дозы. Композиция также может применяться для лечения в течение месяцев (или с соответствующими недельными интервалами), а также путем однократных или многократных обработок.
Для человека со средним весом (70 кг) пригодная месячная доза слитого полипептида по изобретению, такого как димер из примера 7, обычно вводимый в виде разделенных доз один или два раза в месяц, может варьироваться от минимальной дозы, составляющей приблизительно 0,5 мг/месяц, до максимальной дозы, составляющей приблизительно 500 мг/месяц, предпочтительно от приблизительно 1 мг до приблизительно 300 мг, более предпочтительно от приблизительно 20 мг до приблизительно 250 мг в месяц. Более высокие диапазоны доз, например от 500 мг в месяц до 2 г в месяц, могут быть показаны пациентам, имеющим высокие концентрации IgE в сыворотке, или на ранних стадиях лечения.
Доза и время введения также могут варьироваться. Начальные обработки могут быть проведены с использованием композиций с более высокими дозами в указанных выше пределах и их можно вводить более часто по сравнению с более поздней стадией лечения болезни. Требуемые дозы могут быть определены путем измерения в сыворотке уровня содержания IgE и антител к FcεRI, как указано выше. Например, на ранней стадии лечения болезни слитый полипептид по изобретению, такой как димер из примера 7, можно вводить исходя из дозы 200-500 мг полипептида в неделю пациенту со средним весом (70 кг). После клиренса сыворотки от IgE и антитела к FcεRI введение активного вещества можно проводить менее часто, например, еженедельно или каждые две недели, дозами в пределах от 50 мкг до 100 мг полипептида на каждое введение.
Композиции по настоящему изобретению могут вводиться либо по отдельности, либо в сочетании с другими соединениями по настоящему изобретению или с другими фармацевтическими агентами, такими как антигистаминные препараты или кортикостероиды.
Изобретение также относится к применению слитых полипептидов по изобретению в диагностическом тесте in vitro с использованием любого стандартного формата, например ELISA, с целью определения уровня IgE или аутоантител к FcεRI (например, у пациентов, страдающих хронической крапивницей) в биологическом образце, полученном у больного человека, например в образцах крови или ткани. HSA-компонент преимущественно облегчает связывание и выявление IgE или аутоантител к FcεRI в анализе на основе ELISA. Количество IgE или аутоантител, присутствующих в образце, может служить мерой аллергической реакции пациента на вещество, действию которого подвергали пациента. Уровни IgE или аутоантител также могут быть измерены с целью определения эффективности противоаллергических терапий и наблюдения за аллергическим статусом пациента в течение определенного промежутка времени.
Изобретение, кроме того, относится к методу проведения генной терапии людей с использованием полинуклеотида, кодирующего слитый полипептид по изобретению, для лечения болезней, опосредованных IgE и IgE-рецептором. Метод генной терапии включает модификацию клеток пациента путем введения ему полинуклеотида, кодирующего слитый полипептид по изобретению, и экспрессии полипептида этими клетками. Например, сначала у пациента могут быть выделены соматические клетки, которые затем генетически модифицируют в культуре путем инсерции полинуклеотида, после чего полученные в результате модифицированные клетки повторно интродуцируют пациенту, что в результате приводит к экспрессии полипептида по изобретению клетками пациента.
В альтернативном варианте клетки могут быть модифицированы in vivo путем непосредственной инсерции векторной ДНК, кодирующей полипептид.
Пригодные для модификации клетки включают эндотелиальные клетки или лейкоциты. Для генной терапии полинуклеотид по изобретению предпочтительно применять под контролем регулируемого (например, индуцибельного) промотора. Экспрессию полипептида можно при этом сделать зависимой от экспозиции пациента экзогенным фактором с использованием известных регулируемых промоторных систем.
Изобретение также относится к применению методов генной терапии для получения соматических рекомбинантных или трансгенных животных (за исключением человека), не принадлежащих к виду Homo sapiens, экспрессирующих слитые полипептиды по изобретению, например, в молоке. Такие модифицированные животные также составляют часть изобретения. Примеры пригодных животных включают мышей, крыс, кроликов, свиней, овец, коз и крупный рогатый скот, причем все эти виды могут быть сделаны трансгенными с помощью стандартных методик (например, см. D. R. Hurwitz и др., Transgenic Research 3 [1994] 365-375). Животные могут использоваться в целях моделирования или для фактического производства протеина. В частности, изобретение относится к трансгенной мыши, козе, корове или свинье, экспрессирующим слитый полипептид по изобретению. Методы получения таких животных хорошо известны. Полинуклеотид, кодирующий слитый протеин по изобретению, может быть интродуцирован в соматические клетки животноных in vitro или in vivo с целью получения соматических рекомбинантных животных, которые модифицированы генетически, но которые не могут передать генетическую модификацию потомству. В альтернативном варианте полинуклеотиды могут быть встроены в клетки эмбрионов для получения трансгенных животных, способных передавать способность экспрессировать протеины по изобретению потомству.
Трансфекцию клеток для генной терапии можно осуществлять общепринятым способом, например путем электропорации, осаждения фосфатом кальция, с помощью способа, основанного на применении липофектина, внутримышечной инъекции, микроинъекции или с помощью "генного ружья". Плазмидные векторы предпочтительно включают маркер, такой как ген неомицина, для селекции в эукариотических клетках стабильных трансфектантов с цитотоксичным аминогликозидом G418.
Заражение проводят путем включения генной последовательности, кодирующей слитый полипептид, в ретровирусный вектор. В данной области техники известны различные методики такого включения. В одной из таких широко применяемых методик для упаковки ретровируса используется дефектный мышиный ретровирус, и амфотропно упакованная линия клеток используется для получения зараженного амфотропного вируса, предназначенного для использования при заражении донорских клеток-мишеней.
Дополнительные характеристики можно определить, например, следующим образом.
Определение in vivo времени полужизни в сыворотке
Общая методика
Самкам мышей линии Charles River SKHl/hr/hr весом приблизительно 25 г внутривенно вводят тестируемые протеины в стерильном ЗФР без Са+2. Mg+2. Мышей разделяют на следующие группы:
- группа (I), которой вводят 130 мкг (1 нмоль) слитого полипептида, например димера IgER-L1-HSA II-L2-IgER, полученного согласно примеру 7, или
- группа (II), которой вводят 60 мкг IgER (2 нмоля), или
- группа (III), которой вводят 65 мкг HSA I (1 нмоль).
Образцы крови объемом 100 мкл берут у каждой мыши через 10 мин, 30 мин, 3 ч, 6 ч и 12 ч после инъекции. Сыворотку получают центрифугированием. Уровни концентраций различных протеинов в сыворотке определяют а) анализом связывания с IgE-рецептором с помощью метода ELISA, б) на основе ингибирования с использованием метода ELISA или в) методом ELISA с помощью HSA-иммуносэндвича, следующим образом.
а) Анализ методом ELISA связывания с IgE-рецептором
Концентрацию IgE в сыворотке определяют, анализируя IgE-связывание следующим методом ELISA с помощью иммуносэндвича: 200 нг человеческого IgE иммобилизуют в лунках планшета типа COSTAR Strip Plate-8 (Cambridge, МА, США) в 100 мкл сенсибилизирующего буфера во влажной камере при 4oС в течение ночи. Каждую лунку дважды промывают 300 мкл ЗФР без Ca+2, Mg, рН 7,2. Планашеты блокируют в течение 1 ч при комнатной температуре с помощью 200 мкл ЗФР без Са+2, Mg+2, содержащего 5% БСА (фирма Sigma). После двукратной промывки 300 мкл ЗФР без Ca+2, Mg+2, содержащим 0,05% Твина 20 (ЗФРТ), добавляют образцы, растворенные в 100 мкл разбавленной в 10 раз (в ЗФР без Ca+2, Mg+2) мышиной сыворотке, и инкубируют в течение 1 ч при комнатной температуре.
Лунки дважды промывают 300 мкл ЗФР и инкубируют с 100 мкл (1 нг) моноклонального антитела к человеческому IgE-рецептору, такому как 5H5/F8, в течение 1 ч при комнатной температуре. Лунки дополнительно дважды промывают 300 мкл ЗФР и инкубируют с 100 мкл козьего антимышиного IgG-HRP (фирма Biorad, разбавлен в соотношении 1:2000 в ЗФР без Ca+2, Mg+2) в течение 1 ч при комнатной температуре. Планшеты трижды промывают 300 мкл ЗФРТ и с помощью 100 мкл субстрата ABTS (фирма Biorad) определяют конъюгаты с перкосидазой хрена (HRP). Реакцию прекращают через 5 мин с помощью 100 мкл 3%-ной щавелевой кислоты. Интенсивность цвета оценивают с помощью фотометра типа EASY READER при длине волны 405 нм.
б) Анализ ингибирования методом ELISA
100 нг FcεRIα иммобилизуют в 96-луночных иммунопланшетах типа Nunc (F96 cert. Maxisorb) в 100 мкл сенсибилизирующего буфера (0,1М NaHCO3, 0,01% NaN3, pH 9,6) во влажной камере при 4oС в течение ночи. Каждую лунку промывают 4 раза 300 мкл ЗФР, содержащим 0,05% Твина 20 (буфер для промывки). В различные наборы лунок добавляют либо отрицательный контроль (50 мкл мышиной сыворотки, разбавленной в соотношении 1:25 в ЗФР, который содержит 0,05% Твина 20, 2% ФТС), серийные разбавления стандартного слитого протеина, например IgER-L1-HSA II-L2-IgE-стандарта (разбавления от 400 нг/мл до 1,6 нг/мл), либо серийные разбавления образцов. Стандарт и образцы разбавляют мышиной сывороткой, разбавленной в соотношении 1:25 ЗФР, содержащим 0,05% Твина 20, 2% ФТС. Сразу после этого добавляют 50 мкл 400 нг/мл конъюгата человеческий IgE-биотин в буфере для разбавления и смешивают. Конечное разбавление мышиной сыворотки в инкубационной смеси составляет 1:50.
После инкубации в течение 2 ч при 37oС планшеты 4 раза промывают 300 мкл буфера для промывки и добавляют 50 мкл конъюгата стептавидин-щелочная фосфатаза (фирма Gibco), разбавленного в соотношении 1:1000 буфером для разбавления, и инкубируют в течение 1 ч при 37oС. Планшеты промывают 4 раза 300 мкл буфера для промывки и после добавления 100 мкл субстрата (1 мг/мл паранитрофенилфосфата в диэтаноламиновом буфере, рН 9,8 [фирма Biorad]) реакцию прекращают после инкубации в течение 30 мин при 37oС с помощью 50 мкл 2М NaOH. Оптические плотности (ОП) измеряют с помощью фотометра, снабженного рабочей станцией, типа BIOMEK-1000 при 406 нм. Количественное определение с помощью стандартных кривых (4-параметрическая аппроксимация логарифмической кривой) осуществляют с помощью программы для расчета Beckman IMMUNOFIT ELISA. Расчет осуществляют по следующей формуле: %связывания = [ОП (значение для образца или стандарта)/ОП (значение для буфера)]•100.
в) Анализ методом ELISA с помощью HSA-иммуносэндвича
500 нг моноклонального антимышиного HSA (HSA-9) иммобилизуют в 96-луночных иммунопланшетах типа Nunc (F96 cert. Maxisorb) в 100 мкл сенсибилизирующего буфера (0,1М NaHCO3, 0,01% NaN3, pH 9,6) во влажной камере при 4oС в течение ночи. Каждую лунку промывают 4 раза 300 мкл буфера для промывки (ЗФР, содержащий 0,05% Твина 20). В качестве отрицательного контроля добавляют 100 мкл разбавленной в соотношении 1:100 мышиной сыворотки (в буфере для разбавления, состоящем из ЗФР, который содержит 0,05% Твина 20, 2% ФТС), или 100 мкл стандарта (в концентрации от 1 мкг/мл до 2 нг/мл человеческого сывороточного альбумина, KABI), или 100 мкл образца, разбавленного в соотношении 1: 100 в мышиной сыворотке. После инкубации в течение 2 ч при 37oС планшеты промывают 4 раза 300 мкл буфера для промывки и добавляют 100 мкл 1 нг/мкл конъюгата кроличий анти-HSA-биотин, разбавленного буфером для разбавления. Конъюгат анти-HSA-биотин очищают с помощью иммуноаффинной хроматографии с CH-Seph4B-HSA и образцы, дающие перекрестные реакции на мышиную сыворотку, удаляют с помощью иммуносорбции с мышиной сывороткой-агарозой. После инкубации в течение 2 ч при 37oС планшеты промывают 4 раза 300 мкл буфера для промывки и добавляют 50 мкл конъюгата стептавидин-щелочная фосфатаза (фирма Gibco), разбавленного в соотношении 1:1000 буфером для разбавления, и инкубируют в течение 1 ч при 37oС. Планшеты промывают 4 раза 300 мкл буфера для промывки и после добавления 100 мкл субстрата (1 мг/мл пара-нитрофенолфосфата в диэтаноламиновом буфере, рН 9,8 [фирма Biorad]) реакцию прекращают после инкубации в течение 15 мин при 37oС с помощью 50 мкл 2М NaOH. Оптические плотности измеряют с помощью фотометра, снабженного рабочей станцией, типа BIOMEK-1000 при 405 нм. Количественное определение с помощью стандартных кривых (4-параметрическая аппроксимация логарифмической кривой) осуществляют с помощью программы для расчета Beckman IMMUNOFIT ELISA.
Результаты
Концентрации димерного слитого полипептида IgER-L1-HSA II-L2-IgER, свободного IgER (обозначена в настоящем описании как "свободная альфа-цепь") или HSA I (обозначен в настоящем описании как "HSA") в зависимости от времени, прошедшем после инъекции, приведены в виде пмолей/мл сыворотки на фиг.1А и 1Б.
На фиг.1Б показано, что свободный рецептор может быть выявлен в мышиной сыворотке только в течение приблизительно 10 мин, в то время как димерный слитый полипептид все еще может быть выявлен через 12 ч после введения.
На фиг.1В показаны кинетики клиренса в отношении HSA и димерного слитого полипептида. После стабилизации распределения между тканью и кровью в первые 10 мин для димера и HSA получены практически идентичные кривые клиренса. Это подтверждает предположение, что время полужизни в сыворотке IgE-связывающего домена может быть в значительной степени пролонгировано путем связывания с HSA.
Ингибирование пассивной кожной анафилаксии (ПКА) у мышей
Общая методика
(а) Введение полипептида
Серийные разбавления 10, 50 или 500 мкг/кг слитого полипептида, например IgER-L1-HSA II-L2-IgER, полученного из клеток СНО согласно примеру 7, внутривенно инъецируют самкам мышей линии Charles River SKHl/hr/hr весом приблизительно 25 г за различные интервалы времени до сенсибилизации.
Формируют 3 группы мышей в зависимости от количества полипептида, инъецируемого до сенсибилизации (т.е. 10 мкг/кг, 50 мкг/кг или 500 мкг/кг). Четвертой контрольной группе мышей вместо полипептида внутривенно вводят 200 мкг/кг ЗФР. Каждую из четырех групп мышей разделяют на 3 подгруппы, которые отличаются по интервалу между внутривенной инъекцией соединения и внутрикожной сенсибилизацией IgE [стадия (б), см. ниже]. Исследование проводили через следующие интервалы: 5 мин, 15 мин и 30 мин.
(б) Внутрикожная сенсибилизация
Мышей подвергают анестезии и сенсибилизируют с помощью 4 внутрикожных инъекций в кожу спины 5 нг каждого моноклонального мышиного антитела в виде IgE к динитрофенилу (ДНФ) в 10 мл ЗФР (фирма BioMakor, Rehvot, Израиль). В контрольной группе физиологический раствор инъецируют внутрикожно в одно место.
(в) Контрольное заражение аллергеном
Через 90 мин после сенсибилизации мышей вновь подвергают анестезии и проводят контрольное заражение с помощью внутривенной инъекции раствора, содержащего 50 мкг конъюгата динитрофенил-бычий сывороточный альбумин (ДНФ-БСА) (фирма Calbiochem-Behring, San Diego, США), содержащим 1% красителя Эванса голубого. ПКА-реакцию оценивают количественно, измеряя диаметр окрашенной тестируемой области после транссудации.
Результаты
Результаты приведены в виде гистограмм на фиг.2 для зрелого слитого полипептида из примера 7 (аминокислоты Val26-Leu978 SEQ.ID.NO.3). В концентрации 500 мкг/кг димерный слитый полипептид полностью блокирует ПКА при исследовании всех интервалов времени обработки. В концентрации 50 мкг/кг обработка за 30 мин до контрольного заражения позволила получить статистическое достоверное уменьшение на 36%. При обработке за 15 мин до контрольного заражения видна тенденция к уменьшению при введении как 10, так и 50 мкг/кг димера. Таким образом, димерный слитый полипептид по изобретению обладает эффективностью в отношении предупреждения ПКА.
Способы и методы, необходимые для осуществления настоящего изобретения, являются известными в данной области. Используемые соединения, реагенты, векторы, линии клеток и т.д., если их получение не приведено специально в данном описании, являются легкодоступными или могут быть получены общепринятым методом из известных и легко доступных продуктов, или эквивалентные им продукты могут быть получены общепринятым способом из известных и легкодоступных продуктов.
Ниже изобретение проиллюстрировано на примерах, не ограничивающих его объем. Все температуры указаны в градусах Цельсия.
Материалы и методы
ПЦР-амплификация
Химический синтез de novo праймеров по изобретению может быть проведен с использованием любого приемлемого метода, такого как, например, фосфотриэфирный или фосфодиэфирный методы.
Методы и системы для амплификации специфической последовательности нуклеиновой кислоты описаны в US 4683195 и US 4683202, а также в Polymerase Chain Reaction, под ред. Н.А. Erlich и др., Cold Spring Harbor Laboratory Press [1989].
После ПЦР ДНК-фрагменты вырезают и очищают с использованием протокола QiaEx (фирма Qiagen Inc. , Chatsworth, CA, США), а затем субклонируют в векторе ТА [набор ТА Cloning® Kit (фирма Invitrogen), (известный из литературы продукт, версия 2.2)]. Праймеры, использованные для проведения ПЦР-амплификации нуклеиновых кислот, представлены на фиг.8. ДНК секвенируют, используя метод с применением секвеназы (фирма USB, Cleveland, ОН, США).
Плазмиды и реагенты
Клон кДНК FcεRIα pGEM-3-110B-l (A. Shimizu и др., Proc. Nat. Acad. Sci. USA 85 [1988] 1907-1911] был получен из Американской коллекции типовых культур тканей (штамм АТСС 67566). Одноцепочечную кДНК из печени человека была получена от фирмы Clontech (готовый для ПЦР быстрый клон (PCR-ready Quick Clone кДНК, Cat. D 7113-1). Последовательность HSA поступает в продажу под регистрационными номерами GenBank VОО495, JООО78, LОО132 и LОО133. Рестрикционные ферменты были получены от фирм Boehringer-Mannheim или Gibco/BRL. ДНК-полимераза Taq была получена от фирмы Perkin-Elmer Cetus (PECI) или от фирмы Boehringer-Mannheim. Вектор SK был получен от фирмы Stratagen.
pHIL-D2 поставляется фирмой Invitrogen (San Diego, CA, США; кат. номер К1710-01 [1994]) (см. "Pichia Expression Kit - Protein Expression - A Manual of Methods for Expression of Recombinant Proteins in Pichia pastoris - Version 3.0" [декабрь 1994]) (далее в настоящем описании "Invitrogen Manual").
Вектор рХМТ3 получали из рМТ2 (Sambrook и др. (ред.) [1989], см. выше) путем клонирования линкера PstI-EcoRI из pUC8 (фирма Pharmacia) в сайтах PstI и EcoRI pMT2 (R.J. Kaufman и др. [1987], см. выше).
Стандартные методы, по которым работали в описанных ниже примерах, описаны у Sambrook и др. (ред.) [1989], см. выше.
Пример А: Клонирующий вектор ТА
Плазмиду pCR2 лигируют по отдельности с каждым из продуктов, полученных в результате ПЦР-амплификации в примерах 1 и 2. Реакции лигирования проводят с использованием ДНК-лигазы Т4 в следующей реакционной смеси:
25мМ Трис-НСl (рН 7,8),
10мМ MgCl2,
1мМ ДТТ,
1мМ АТФ,
50 нг векторной ДНК,
100-200 нг продуктов ПЦР-реакции (неочищенных),
4 ед. ДНК-лигазы Т4 (фирма New England Biolabs).
Каждую реакционную смесь выдерживают при 15oС в течение 18 ч перед трансформацией компетентных клеток.
Пример 1: ПЦР-амплификация и клонирование кДНК FcεRIα
кДНК FcεRIα очищают из pGEM-3-110B-1, используя метод Qiagen.
После этого работают по следующей методике.
(А) Для получения конструкции с лидирующим HSA ПЦР-амплификацию кДНК FcεRIα проводят с олигонуклеотидами 18 и 19 (фиг.4, 8), используя следующую реакционную смесь:
1 мкл (50 нг) кДНК FcεRIα,
50 пмолей каждого из олигонуклеотидов 18 и 19,
5 мкл десятикратного (10х) ПЦР-буфера (PECI),
0,5 мкл 20 мМ маточного раствора дНТФ=200 мкМ конечной концентрации дНТФ,
0,5 мкл (2,5 ед.) ДНК-полимеразы Taq (PECI),
вода до 50 мкл.
В реакционную смесь загружают минеральное масло для предупреждения испарения и проводят термоциклизацию в термоячейке типа Perkin Elmer Cetus DNA, модель 480. Условия циклизации для этой реакции являются следующими: нагревание до 95oС в течение 5 мин, затем 30 циклов при 94oС в течение 1,5 мин, при 53oС в течение 2 мин, при 72oС в течение 3 мин с последующим удлинением в течение 3 мин при 72oС и выдержкой в течение ночи при 4oС.
После электрофореза наличие амплифицированного продукта длиной ~550 пар оснований подтверждают с помощью окрашивания бромидом этидия. Фрагмент субклонируют в ПЦР2-векторе, получая вектор рЕК1, кодирующий IgER (фиг.4, 8Б).
(Б) Для получения конструкции с лидирующим IgE ПЦР-амплификацию кДНК FcεRIα проводят согласно приведенному выше способу (А), за исключением того, что применяют олигонуклеотиды 20 и 31 (фиг.8Б). После электрофореза на 7%-ном агарозном геле наличие амплифицированного продукта длиной ~600 пар оснований подтверждают с помощью окрашивания бромидом этидия. Фрагмент субклонируют в ПЦР2-векторе, получая вектор IgЕR/ТА 1, кодирующий пре-IgER (фиг. 4).
(В) Для получения конструкции, кодирующей пре-IgER, с целью экспрессии зрелого укороченного протеина для использования в анализах в качестве контроля ПЦР-амплификацию кДНК FcεRIα проводят согласно приведенному выше способу (А) с олигонуклеотидами 20 и 19 (фиг.8Б). После электрофореза наличие амплифицированного продукта длиной ~620 пар оснований подтверждают с помощью окрашивания бромидом этидия. ПЦР-фрагмент отщепляют и субклонируют в ПЦР II-векторе, получая вектор IgER/TA 34, кодирующий пре-IgE с расположенным за ним стоп-кодоном (фиг.4).
Пример 2: ПЦР-амплификация и клонирование кДНК человеческого сывороточного альбумина
Для получения последовательности, кодирующей препро-HSA II, ПЦР-амплификацию полноразмерной кДНК человеческого сывороточного альбумина проводят с олигонуклеотидами 24 и 25 (фиг.8Б) по общей методике, описанной в примере 1(А). Полученный в результате клон HSA/TA 1 имел последовательность, наиболее близкую к последовательности из Genbank. В этом клоне выявлено 7 мутаций в незначимом положении и 1 мутация, приводящая к замене лизина на глутаминовую кислоту в основании, находящемся в положении 1333. Семь мутаций в нестрогом положении представляют собой основание, находящееся в положении 309 (замена А на Т), основание, находящееся в положении 744 (замена А на G), основание, находящееся в положении 795 (замена G на А), основание, находящееся в положении 951 (замена G на А), основание, находящееся в положении 1320 (замена С на Т), основание, находящееся в положении 1569 (замена А на С), основание, находящееся в положении 1584 (замена G на А) (относительно HSA/TA 1). Мутацию, приводящую к замене лизина на глутаминовую кислоту, подвергают обратному корректированию на последовательности из Genbank, используя сайтнаправленный мутагенез с олигонуклеотидами, приведенными на фиг. 8А, и набор с мутагеном для мутагенеза in vitro Bio-Rad Mutagene Phagemid (фирма Biorad, кат. номер 170-3581). Этот метод основан на включении остатков урацила в родительскую цепь и их последующий перенос на мутантную цепь (Т.A. Kunkel Proc. Nat. Acad. Sci. USA 82 [1985] 488-492]. Поскольку точковая мутация в незначимом положении не может изменить нативную аминокислотную последовательность кодируемого протеина, вышеуказанные 7 мутаций не корректировали. После электрофореза наличие амплифицированного продукта длиной ~1,8 т.п.н. подтверждают с помощью окрашивания бромидом этидия.
Продукт длиной ~1,8 т.п.н. субклонируют в векторе pCR2 с целью получения мутанта 16 HSA/TA (HSA/TA mut 16), который проверяют с помощью секвенирования ДНК на наличие полной последовательности препро-HSA II. HSA/TA mut 16 субклонируют в векторе Bluescript SK в виде SреI,НindIII-фрагмента, получая HSA/SK 17 (фиг.5).
(A) Для получения конструкции с лидирующим HSA HSA/SK 17 расщепляют с помощью MstII и HindIII для удаления нуклеотидной последовательности, кодирующей 3'-концевую аминокислоту (Leu609), и олигонуклеотид, кодирующий линкер L2, как определено выше, интродуцируют на 3'-конец линеаризованного НSА/SК 17 путем обработки киназой олигонуклеотидов 28 и 29 (фиг.8Б) и отжига и лигирования этих фрагментов в сайты MstII/HmdIII линеаризованного НSА/SК 17, получая рЕК7, кодирующую препро-HSA II, слитый с L2 (фиг.8Б). Минипреп ДНК подтверждают расщеплением с помощью BamHI и секвенированием.
(Б) Для получения конструкции с лидирующим IgE НSА/SК 17, кодирующий препро-HSA II, амплифицируют с помощью ПЦР с олигонуклеотидами 26 и 27 (фиг. 5, 8). Олигонуклеотид 26 удаляет препро-последовательность из HSA и добавляет олигонуклеотид, кодирующий L1, на 5'-конец HSA. Олигонуклеотид 27 заканчивается встречающимся в естественных условиях уникальным сайтом NcoI приблизительно на нуклеотиде в положении 800 кодирующей последовательности HSA. ПЦР-амплификацию проводят согласно способу, описанному в примере 1(А). Фрагмент длиной приблизительно 806 пар оснований выделяют с помощью гель-электрофореза и Qia субклонируют в векторе pCR2, получая клон HSA Nсо/ТА 13, последовательность которого подтверждают с помощью секвернирования ДНК. NcoI-NotI-фрагмент из этого клона субклонируют в ДНК НSА/SК 17, расщепленной с помощью NcoI и NotI, получая HSA/SK 5 (фиг.5), кодирующий L1, связанный с 5'-концом кДНК, кодирующей HSA II.
(B) Для экспрессии только HSA применяют указанную выше конструкцию HSA/SK 17.
Пример 3: Слитая конструкция HSA-IgER/SK 22, кодирующая препро-HSA+линкер+IgЕR
Вектор рЕК7 (содержащий кДНК препро-HSA II + олигонуклеотид для L2) и рЕK1 (содержащий IgER) расщепляют с помощью BamHI и SalI. Полученный фрагмент длиной 1,8 т.п.н. из рЕК7 обрабатывают фосфатазой и затем лигируют с фрагментом длиной 550 т.п.н., полученным из рЕК1. Получают один позитивный минипреп 23, но он оказался загрязнен другой неизвестной плазмидой, которая препятствует восстановлению полосы HSA-IgER. Поэтому из минипрепа 23 очищают SpeI-SalI-фрагмент длиной 2,4 т.п.н., содержащий слияние HSA-IgER, и фрагмент длиной 2,9 т.п.н., содержащий векторную ДНК, и лигируют вместе, получая в результате клон НSА-IgЕR/SК 49 (фиг.9) [было обнаружено, что сайты вектора отсутствовали на любом конце субклонированной области, и поэтому SpeI-SalI-фрагмент длиной 2,4 т. п. н. из HSA-IgER/SK 49 субклонируют в векторе Bluescript SK, расщепленном с помощью SpeI плюс SalI, получая в результате HSA-IgER/SK 22 (не показано на чертежах)].
Последовательность стыка HSA и линкера с ДНК IgE-рецептора и последовательность концов слияния с векторной ДНК представляли собой, как и ожидалось, НSА-IgЕR/SК 22, что подтверждено при анализе последовательности ДНК.
Пример 4: Слитая конструкция IgER-HSA/SK l, кодирующая IgER+препро-HSA II
HSA/SK 5 и IgER/TA 1 расщепляют с помощью SstI и NheI. Фрагмент длиной 600 пар оснований из IgЕR/ТА 1 очищают с помощью гель-электрофореза и лигируют с расщепленным и обработанным фосфатазой НSА/SК 5, получая клон IgЕR-HSА/SК 1 (фиг.10).
Пример 5: Слитая конструкция R-Н-R/SК 50, кодирующая пpe-IgER-L1-HSA II-L2-IgER
Сайт PstI, уникальный для HSA-области IgER-HSA/SK 1, и HSA-IgER/SК 49 применяют для соединения IgER и препро-HSA II через олигонуклеотид для L2. HSA-IgER/SK 49 и Bluescript SK расщепляют с помощью PstI и SalI. Фрагмент длиной 1,2 т.п.н., содержащий 3'-область HSA II, линкер и последовательность IgER, лигируют с ДНК вектора Bluescript, расщепленной с помощью PstI плюс SalI, получая в результате вектор HSA-IgER Pst Sal/SK 37 (фиг.11), который расщепляют с помощью PstI и KpnI, получая фрагмент длиной 1,2 т.п.н.
ДНК IgER-HSA/SK 1 расщепляют с помощью PstI и KpnI и выделяют фрагмент длиной 4,8 т.п.н., содержащий вектор, IgER, линкер и 5'-область HSA, обрабатывают фосфатазой и лигируют с фрагментом длиной 1,2 т.п.н. из вектора HSA-IgER Pst Sаl/SК 37. Получают образовавшуюся при этом димерную конструкцию R-H-R/SK 50 (фиг.11).
Пример 6: Получение мономерных слитых полипептидов HSA II-L2-IgER путем трансфекции и культивирования Pichia pastoris
Плазмиду МВ 2, кодирующую HSA II-L2-IgER получают путем расщепления плазмиды HSA/SK 49 с помощью EcoRI и выделения фрагмента длиной 2,4 т.п.н., кодирующего слитый протеин. Этот фрагмент встраивают в уникальный сайт EcoRI в векторе pHIL-D2, обеспечивающем экспрессию в Pichia pastoris (фирма Invitrogen), после расщепления плазмиды pHIL-D2 с помощью EcoRI и обработки щелочной фосфатазой. Образовавшуюся плазмиду МВ 2 линеаризуют путем расщепления с помощью NotI и трансформируют ею клетки his4 Gs115, как описано в "Invitrogen Manual".
Трансформантов, позитивных по His (His+), подвергают скринингу на способность расти в среде с добавлением метанола. Штаммы, проявившие медленный рост на метаноле, выращивают на минимальной среде с глицерином, как указано в "Invitrogen Manual", до стационарной фазы, переносят центрифугированием в забуференную метанольным комплексом среду и выращивают в течение 4 дней. Супернатант, полученный из клеток, затем анализируют с помощью ELISA на присутствие HSA и на способность связываться с IgE. Способность связываться с IgE продукта, полученного в результате секрециии Р. pastoris, эквивалентна концентрации HSA на молярной основе и полностью эквивалентна биологической активности.
Пример 7: Получение димерного слитого полипептида IgER-L1-HSA II-L2-IgER путем трансфекции и культивирования в клетках СНО
Плазмиду pXMT3-RIα-HSA-RIα (содержащую полинуклеотид, представленный в SEQ.ID.NO.4 и кодирующий пpe-IgER-L1-HSA II-L2-IgER), получают путем расщепления R-H-R/SK 50 (см. пример 5) с помощью EcoRI и выделения EcoRI-фрагмента длиной 3 т. п. н. , кодирующего димерный слитый полипептид. Этот фрагмент встраивают в уникальный сайт EcoRI плазмиды рХМТ3 после расщепления рХМТ3 с помощью EcoRI и обработки щелочной фосфатазой.
Плазмидой трансфектируют клетки СНО DUKX B11. Эти клетки лишены функционального гена dhfr (дигидрофолатредуктаза), необходимого для синтеза нуклеозидов. Поэтому клетки поддерживают в среде альфа+ (среда MEM ALPHA MEDIUM с рибонуклеозидами и дезоксирибонуклеозидами/фирма Gibco), содержащей 10% фетальной телячьей сыворотки (ФТС). Для трансфекции клетки промывают дважды в ЗФР без Са+2, Mg+2 (CMF-ЗФР) и концентрацию клеток доводят до 2•106 клеток/мл CMF-ЗФР. 0,8 мл клеточной суспензии добавляют к 15 мкг плазмидной ДНК. Трансфекцию осуществляют с помощью электропорации, используя устройство BIO RAD GENE Pulcer (напряжение 1000 В; емкость 25 мкФ). После трансфекции клетки культивируют в 15 мл среды альфа+, содержащей 10% ФТС, в течение 3 дней.
Локализация гена dhfr в плазмиде рХМТ3 позволяет производить отбор в отношении рекомбинантных клеток в среде без нуклеозидов. Через 3 дня после трансфекции клетки помещают в альфа-среду (среда MEM ALPHA MEDIUM без рибонуклеозидов и дезоксирибонуклеозидов/фирма Gibco), содержащую 10% диализованной фетальной телячьей сыворотки (ДФТС). После культивирования в течение 2 недель рекомбинантные колонии клеток становятся заметными. Клетки выдерживают в альфа-среде, содержащей 10% ДФТС, в течение 4 дополнительных пассажей до начала амплификации гена.
Амплификацию dhfr и связанных с ним генов проводят в присутствии метатрексата (МТ), получая в результате повышенную экспрессию трансгена. Поэтому селекция рекомбинантных клеток в среде без нуклеозидов сопровождается культивированием в присутствии 20нМ МТ в альфа-среде, содержащей 10% ДФТС. Дополнительную амплификацию получают при ступенчатом увеличении концентрации метотрексата до 100нМ и 500нМ МТ. Протеин получают путем сбора пула Т1/3-500нМ в альфа-среде, содержащей 10% ДФТС (фирма Gibco), при плотности 9•103/см2 в роллер-флаконы. Первый супернатант собирают через 5 дней после посева с последующим переносом в бессывороточную альфа-среду. Второй сбор проводят через 3 дня, получая всего 1 л супернатанта, предназначенного для очистки.
Вторую порцию очищают из 2 л супернатанта, полученного из пула Т1/3-500нМ, адаптированного к росту в бессывороточных условиях. Клетки высевают с плотностью 5•104/мл и супернатант собирают через 6 дней после посева.
Пример 8: Очистка слитого протеина
Супернатанты культур из примера 7 очищают с помощью иммуноаффинной хроматографии на иммобилизованных моноклональных антителах к FcεRI (например, 5H5-F8; мышиный IgG1), полученных и очищенных стандартными методами.
а) Приготовление хроматографического носителя
Моноклональные антитела связывают с CNBr-активированной Сефарозой 4В (фирма Pharmacia, Uppsala, Швеция) с плотностью 10 мг антитела/мл геля согласно инструкциям производителя. Избыток реакционноспособных групп затем блокируют этаноламином и до использования хранят в смоле в ЗФР, дополненном 0,02% NaN3.
б) Аффинная хроматография
Прозрачный супернатант культуры вносят в колонку, содержащую 5 мл антитела и уравновешенную в ЗФР, со скоростью потока 0,5 мл/мин. Абсорбированный продукт элюируют 50мМ лимонной кислотой, 140мМ NaCl, рН 2,70. Значение рН фракций, содержащих протеин, сразу же доводят до 7,0 (NaOH), а затем фильтруют в стерильных условиях.
в) Количественная оценка/характеристика
Концентрацию димерного слитого полипептида определяют путем абсорбции при 280 нм в его нативной конформации в 30мМ (3-[N-морфолино]пропан)сульфоновой кислоте (МПС), рН 7,0, и в его денатурированной форме (6М гуанидин•НСl). Соответствующий коэффициент молярной абсорбции рассчитывают, исходя из количества остатков триптофана, тирозина и цистеина, используя табличные коэффициенты абсорбции этих аминокислот в модельных соединениях и корректируют для различий в оптической плотности между складчатым и нескладчатым протеинами. Слитый протеин содержит 17 остатков триптофана, 40 тирозина и 21 цистеина, что позволяет получить теоретическое значение коэффициента экстинции 150840 М-1см-1. Качество очищенного продукта определяют с помощью стандартного ДСН-ПААГ-электрофореза (электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия) и путем N-концевого автоматического секвенирования с использованием газофазного разложения Эдмана и масс-спектрометрии. На основании данных ДСН-ПААГ-электрофореза (фиг.15) полипептид мигрирует со скоростью, соответствующей, по-видимому, молекулярной массе примерно 140 кДа, что свидетельствует о приблизительно 28%-ном гликозилировании (теоретическая молекулярная масса без гликозилирования: 108863,61 Да).
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА ПРОТИВ РЕЦЕПТОРА FcγRIIB И ИХ ПРИМЕНЕНИЕ | 2005 |
|
RU2404991C2 |
| НОВЫЕ ПЕСТИЦИДНЫЕ ПРОТЕИНЫ И ШТАММЫ | 1995 |
|
RU2196824C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ГЕТЕРОКАРПИНА | 2003 |
|
RU2359033C2 |
| УЛУЧШЕННОЕ АНТИ-IGE АНТИТЕЛО (ВАРИАНТЫ) И СПОСОБ УЛУЧШЕНИЯ ПОЛИПЕПТИДОВ | 1998 |
|
RU2242515C2 |
| НОВЫЕ КЛЕТКИ ПОЗВОНОЧНЫХ И СПОСОБЫ РЕКОМБИНАНТНОЙ ЭКСПРЕССИИ ИНТЕРЕСУЮЩЕГО ПОЛИПЕПТИДА | 2015 |
|
RU2710726C2 |
| ПРОТЕИН, СВЯЗАННЫЙ С ЗАБОЛЕВАНИЕМ | 2002 |
|
RU2384586C2 |
| ВЫСОКОКОНЦЕНТРИРОВАННЫЕ КОМПОЗИЦИИ АНТИТЕЛ И БЕЛКОВ | 2004 |
|
RU2332986C2 |
| ТЕРАПЕВТИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2002 |
|
RU2328506C2 |
| ОПТИМИЗИРОВАННАЯ ЭКСПРЕССИОННАЯ КАССЕТА ДЛЯ ЭКСПРЕССИИ ПОЛИПЕПТИДА С ВЫСОКИМ ВЫХОДОМ | 2013 |
|
RU2682884C2 |
| КОНСТРУКЦИИ АНТИТЕЛ И ХЕМОКИНОВ И ИХ ПРИМЕНЕНИЕ ПРИ ИММУНОЛОГИЧЕСКИХ НАРУШЕНИЯХ | 2001 |
|
RU2252786C2 |











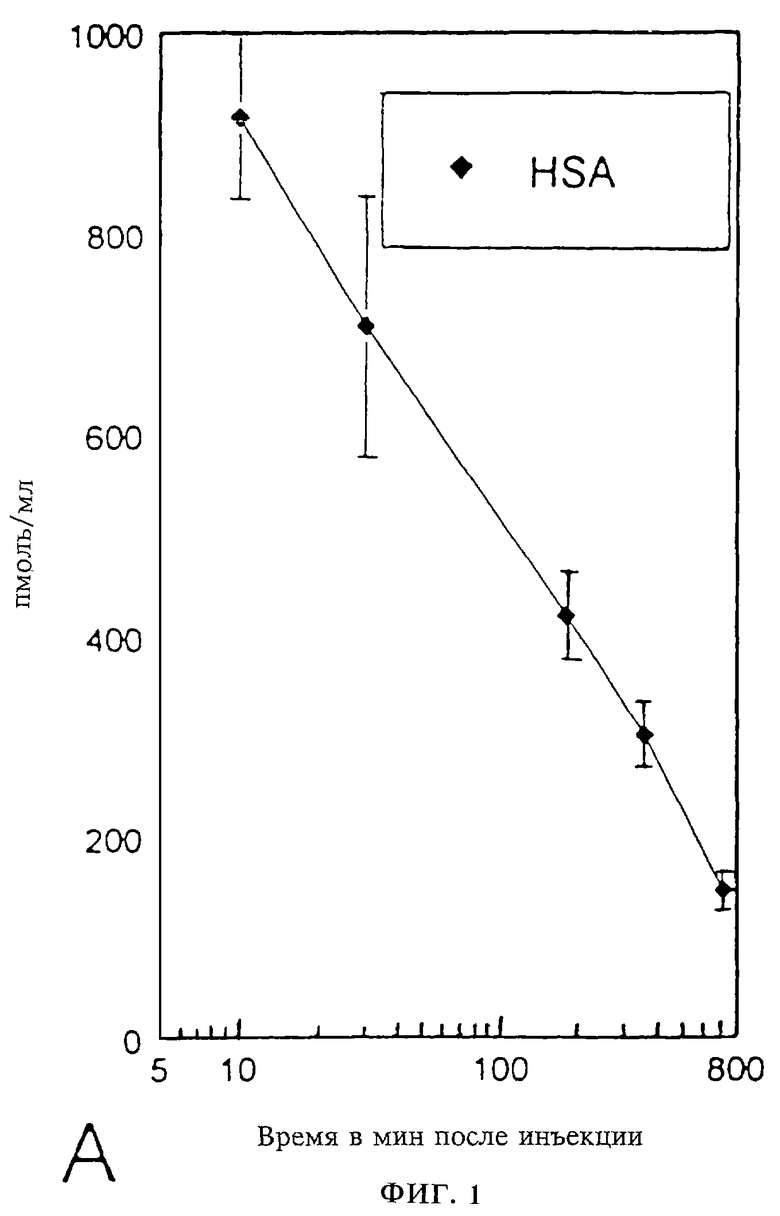
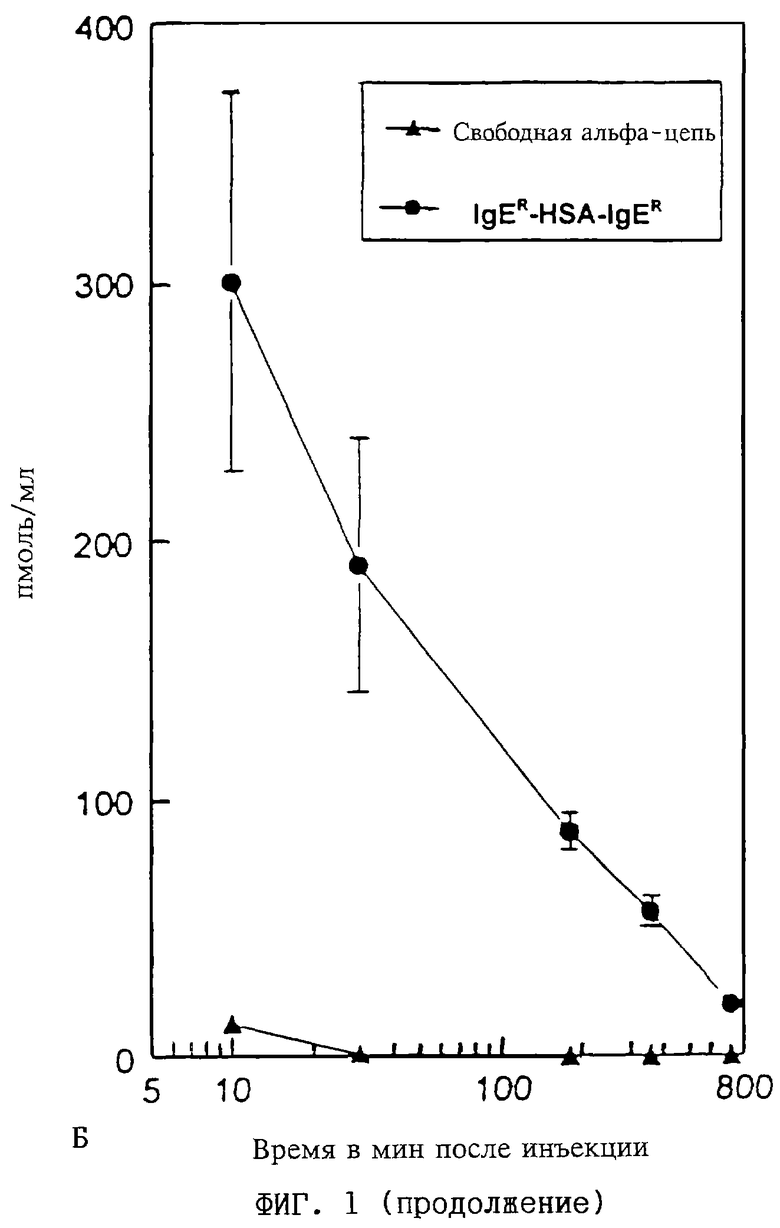
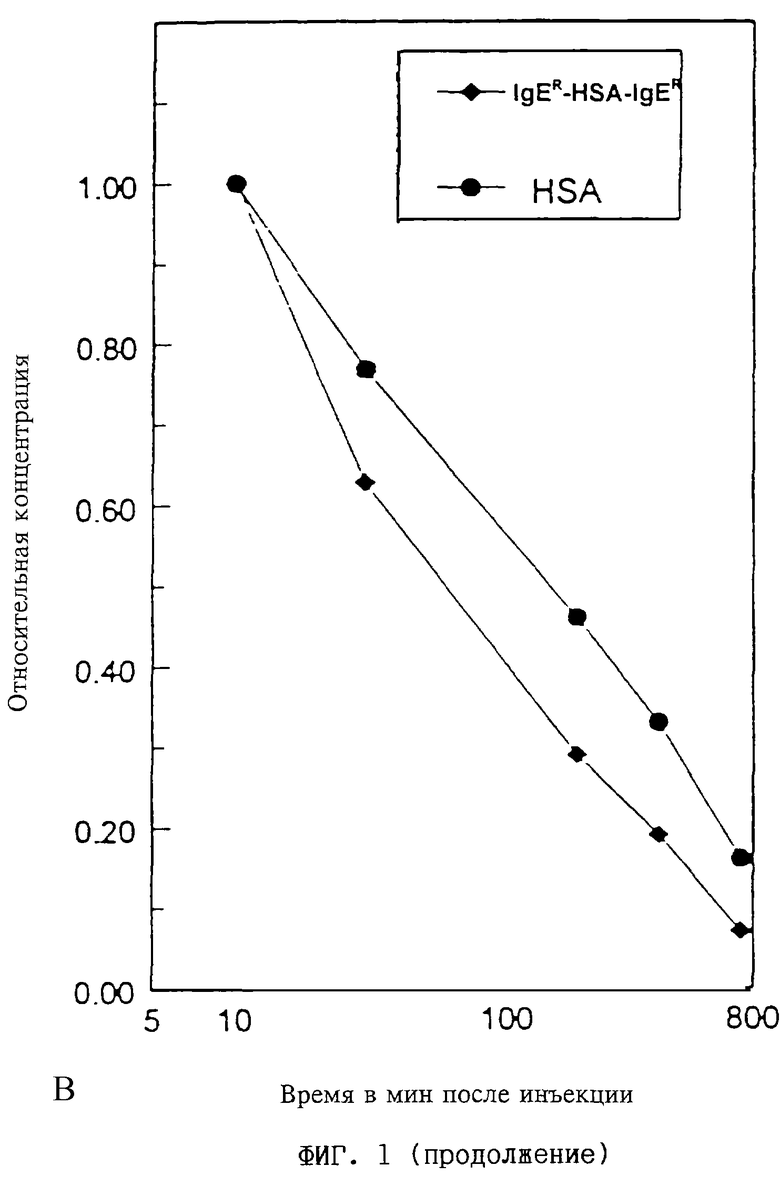
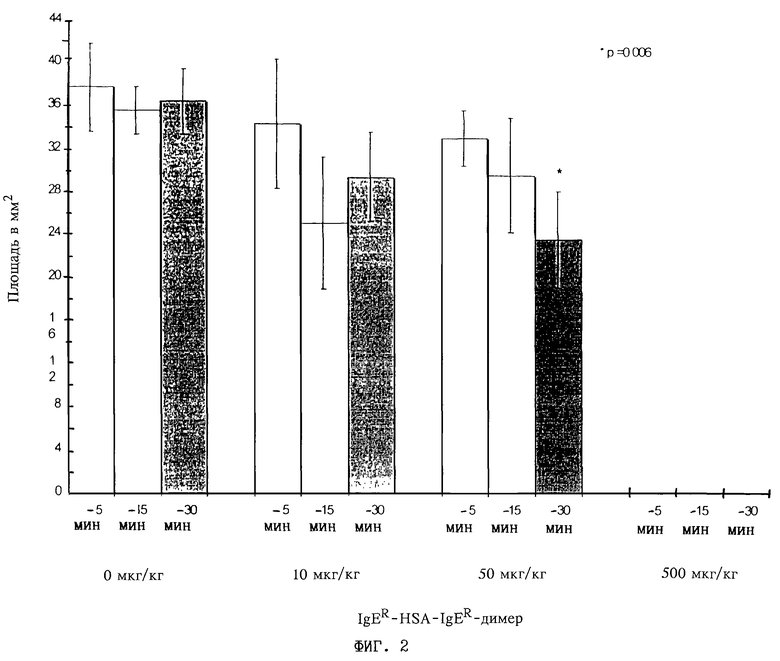

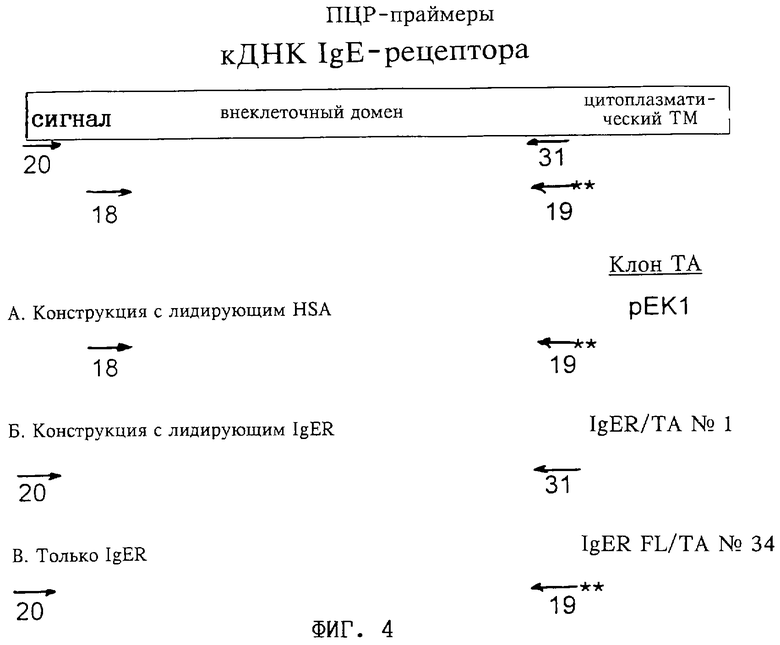
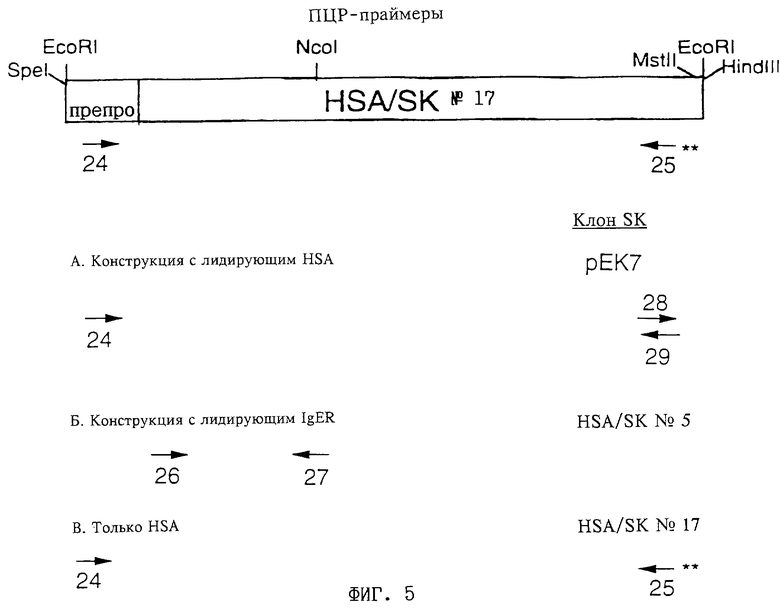





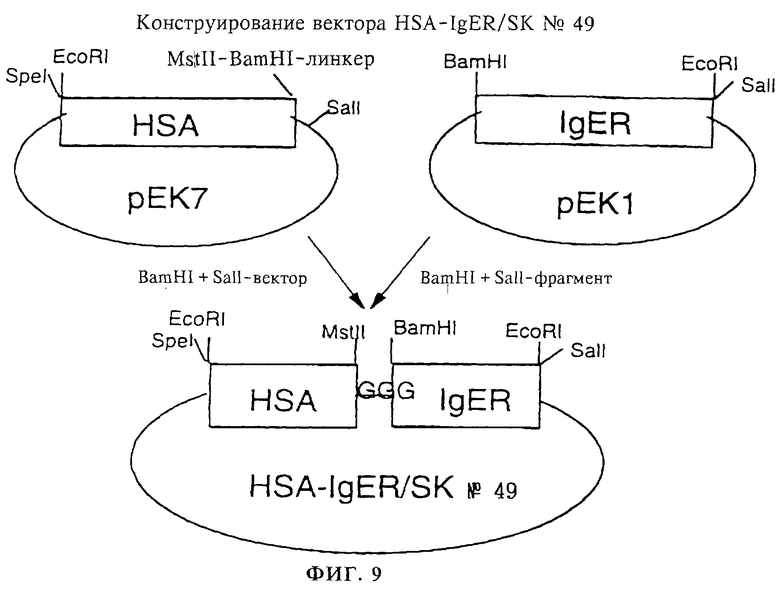
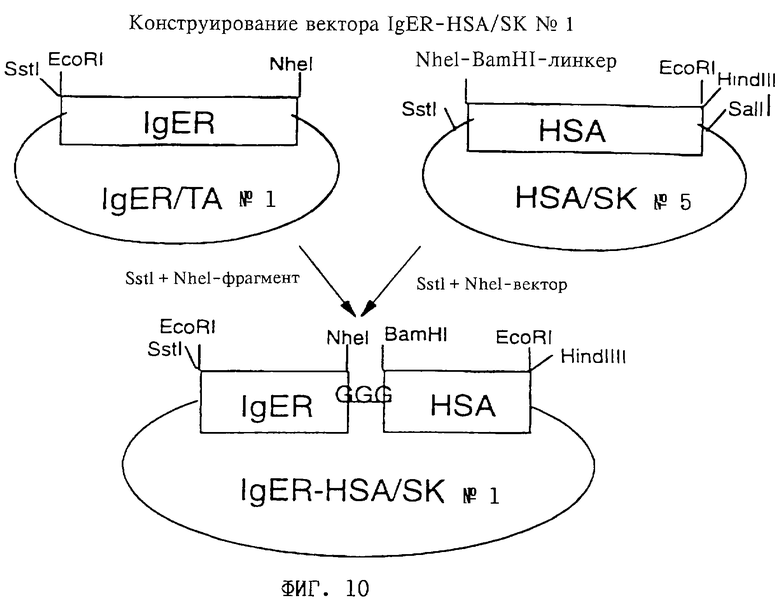
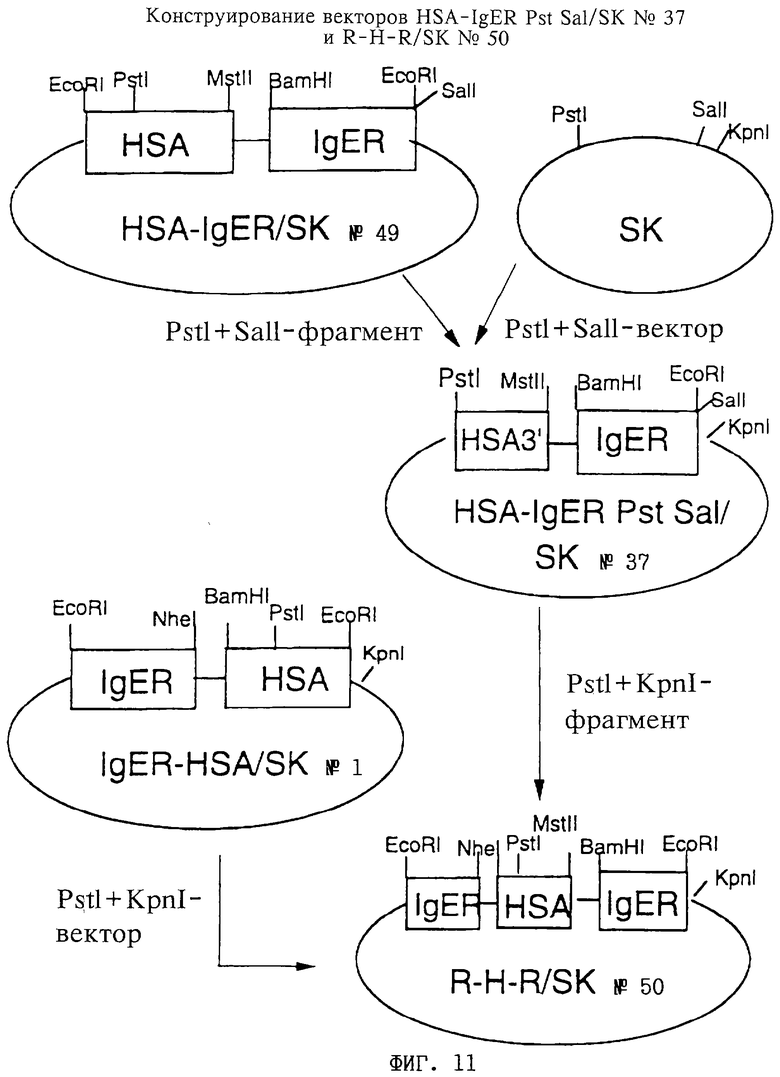







Изобретение относится к генетической инженерии. Слитый полипептид или его соль включает, по крайней мере, один IgЕ-связывающий домен с, по крайней мере, одним компонентом сывороточного альбумина (HSA). IgE-связывающий домен является IgЕR или пре-IgЕR. Аминокислотная последовательность приведена в описании. Экспрессирующий вектор содержит нуклеотидную последовательность, кодирующую слитый полипептид. Фармацевтическая композиция содержит слитый полипептид или его соль в эффективном количестве и фармацевтически приемлемые носители или разбавители. Изобретение позволяет получить фармкомпозицию для лечения опосредованных IgE аллергических заболеваний и связанных с ними нарушений, таких как атопический дерматит, атопическая астма и хроническая крапивница. 3 с. и 3 з.п.ф-лы, 15 ил.
| WO 9013653 А, 15.11.1990 | |||
| SABAN R | |||
| et al | |||
| Allergens, IgE, mediators, inflammatory mechanisms | |||
| Journal of allergy and clinical immunology, VOL.94, NO | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Коросниматель окорочного станка роторного типа | 1974 |
|
SU499112A1 |
| RU 94031472 А1, 20.04.1996 | |||
| US 4962035 А, 09.10.1990. | |||

