Настоящее изобретение относится к заявке, не являющейся временно зарегистрированной в соответствии с 37 CFR § 1.53 (b) (1), по которой испрашивается приоритет в соответствии с 35 U.C.S. § 119 (e) серийной заявки на патент США № 60/606851, поданной 2 сентября 2005 года, полное содержание которой приведено в настоящем описании в качестве ссылки в полном объеме.
Область техники, к которой относится изобретение
Настоящее изобретение относится к антителам, которые предпочтительно связываются с FcγRIIB человека в сравнении с FcγRIIA человека, а также к применению указанных антител.
Предпосылки создания изобретения
Антитело связывается с антигеном и нейтрализует его, предотвращая его связывание с эндогенной мишенью (например, рецептором или лигандом) или индуцируя эффекторные ответы, которые приводят к удалению антигена. Для эффективного удаления и/или разрушения чужеродных антигенов в организме антитело должно обладать как высокой аффинностью к антигену, так и эффекторными функциями. Антитела, обладающие специфичностью (такие как, например, биспецифические антитела), могут использоватся для опосредования комплементарных или синергических ответов на множественные антигены.
Эффекторные функции антитела опосредованы Fc-участком антитела. Эффекторные функции подразделяются на две категории: (1) эффекторные функции, которые проявляются после связывания антитела с антигеном (указанные функции включают участие комплементного каскада или клеток, несущих Fc-рецептор (FcR)); и (2) эффекторные функции, которые работают независимо от связывания с антигеном (такого рода функции определяют персистентный характер антитела в кровотоке и его способность проходить через клеточные барьеры путем трансцитоза) (см., например, Ward and Ghetie, 1995, Therapeutic Immunology 2:77-94). Взаимодействия антител и комплексов антитело-антиген с клетками иммунной системы вызывают такие ответы, как, например, антителоопосредованная клеточная цитотоксичность (ADCC) и комплементзависимая цитотоксичность (CDC) (см. обзор Daërоn, 1997, Annu. Rev. Immunol. 15:203-234; Ward et al., выше; Ravetch et al., 1991, Annu. Rev. Immunol. 9: 457-492; и Ravetch et al., 2000, Science 290: 84-89).
Поскольку Fc-рецепторы опосредуют эффекторную функцию антитела посредством связывания с Fc-участком соответствующего рецептору антитела, FcR определяются по их специфичности к иммуноглобулиновым изотипам: Fc-рецепторы, специфичные для IgG антител, обозначаются как FcγR; Fc-рецепторы, специфичные для IgE антител, обозначаются как FcεR; Fc-рецепторы, специфичные для IgA антител, обозначаются как FcαR и т.п.
Идентифицированы три подкласса FcγR: FcγRI (CD64), FcγRII (CD32) и FcγRIII (CD16). Каждый подкласс FcγR кодируется двумя или тремя генами, которые подвергаются альтернативному РНК-сплайсингу, что ведет к образованию множественных транскриптов и определяет наличие большого разнообразия изоформ FcγR. Три гена, кодирующих молекулы FcγRI подкласса (FcγRIa, FcγRIb и FcγRIc), сгруппированы в кластер на участке 1q21.1 длинного плеча хромосомы 1; гены, кодирующие изоформы FcγRII (FcγRIIa, FcγRIIb и FcγRIIc), локализованы на участке 1q23-24; и два гена, кодирующие изоформы FcγRIII человека (FcγRIIIa и FcγRIIIb), сгруппированы в кластер на участке 1q22. FcγRIIc формируется в результате неравного генетического перекреста между FcγRIIA и FcγRIIB и состоит из внеклеточного участка FcγRIIB и цитоплазматического участка FcγRIIA.
FcγRIIA кодирует трансмембранный рецептор FcγRIIA1. Альтернативный сплайсинг РНК приводит к образованию FcγRIIA2, который не содержит трансмембранный участок. Аллельные варианты гена FcγRIIA определяют возможность появления высокоотзывчивых молекул (HR) или низкоотзывчивых молекул (LR), которые отличаются по своей способности связываться с IgG. Молекулы HR и LR FcγRIIA отличаются двумя аминокислотами, соответствующими положениям 27 и 131. FcγRIIB кодирует сплайсинговые варианты FcγRIIB1, FcγRIIB2 и FcγRIIB3. FcγRIIB1 и FcγRIIB2 отличаются вставкой из 19 аминокислот в цитоплазматическом домене FcγRIIB1; FcγRIIB3 идентичен FcγRIIB2, но не содержит информацию для предположительного сайта сигнала расщепления пептидазой.
Рецепторы также различаются по их аффинности относительно IgG. FcγRI обладают высокой аффинностью к IgG; значение Ka = 108-109 M-1 (Ravetch et al., 2001, Annu. Rev. Immunol. 19: 275-290) и могут связываться с мономерным IgG. Тогда как FcγRII и FcγRIII демонстрируют относительно слабую аффинность в отношении мономерного IgG, Ka≤107 M-1 (Ravetch et al., выше) и эффективно взаимодействуют только с многомерными иммунными комплексами. Различные подтипы FcγR экспрессируются на разных типах клеток (см. обзор Ravetch J.V. et al., Annu. Rev. Immunol. 9: 457-492). Например, только FcγRIIIA экспрессируется на NK-клетках. Связывание антител с указанным рецептором приводит к появлению ADCC активности, характерной для NK-клеток. FcγRIIIB человека обнаружен только на нейтрофилах, тогда как FcγRIIIA обнаружен на макрофагах, моноцитах, природных клетках-киллерах (NK) и в субпопуляции T-клеток. С другой стороны, рецепторы FcγRII с низкой аффинностью для мономерного IgG представляют собой наиболее широко распространенные FcR, которые обычно экспрессируются совместно на одних и тех же клетках. FcγRII (кодируемый CD32) экспрессируется в значительной степени на В-клетках, моноцитах, ганулоцитах, тучных клетках и тромбоцитах, тогда как некоторые T-клетки экспрессируют рецептор на меньших уровнях (Mantzioris, B. X. Ravetch et al., 1993, J. Immunol. 150: 5175-5184; и Zola, H. et al., 2000, J. Biol. Regul. Homeost. Agents, 14: 311-316). Например, рецептор FcγRII человека встречается преимущественно на B-клетках, миелоидных клетках и тучных клетках (Ravetch J. V. et al., 2000, Science 290: 84-89).
Изоформы FcγRIIA и FcγRIIB содержат очень близкие внеклеточные домены (идентичность аминокислотной последовательности составляет примерно 92%), но различаются своими цитоплазматическими участками, что приводит к их функциональным различиям, определяющим их как «активирующие рецепторы» (FcγRIIA) и «ингибирующие рецепторы» (FcγRIIB). Рецепторы FcγRI и FcγRIII также функционируют в качестве активирующих рецепторов. Указанные активирующие рецепторы несут в цитоплазматическом домене тирозинсодержащий мотив активации иммунорецептора из 19 аминокислот (ITAM). ITAM последовательности запускают активацию семейств тирозинкиназ src и syk, которые, в свою очередь, активируют множество клеточных медиаторов, таких как киназы P13K, PLCγ и Tec. В результате таких стадий активации повышается внутриклеточное высвобождение кальция из эндоплазматического ретикулюма и открывается сопряженный с данной функцией кальциевый канал, образуя тем самым длительный кальциевый ответ. Указанные потоки кальция важны для экзоцитоза гранулярного содержимого, стимуляции фагоцитоза, ответов по типу ADCC и для активации специфических ядерных факторов транскрипции.
Клеточные ответы, опосредованные активацией FcγR, регулируются ингибирующим рецептором FcγRIIB при поддержании периферической толерантности за счет регуляции порогов активации ответа и в конечном счете терминации IgG-опосредованной стимуляции эффектора (Ravetch J.V. et al., Annu. Rev. Immunol. 19: 275-290 (2001)). Такая регуляция инициируется сшиванием активирующих рецепторов с ингибирующими рецепторами FcγRIIB через антиген-IgG-антительный иммунный комплекс (см., например, Ravetch J.V. et al., 2000, выше). Сшивка с ITAM-содержащим активирующим рецептором ведет к фосфорилированию тирозина в пределах тирозинсодержащего мотива ингибирования иммунорецептора из 13 аминокислот (ITIM) в цитоплазматическом домене FcγRIIB. Такая «активация»
FcγRIIB инициирует рекрутмент специфической SH-2-содержащей инозитполифосфат-5-фосфатазы (SHIP). SHIP катализирует гидролиз мембранного инозитлипида PIP3, препятствуя тем самым активации киназ PLCγ и Tec и устраняя длительный кальциевый поток, опосредованный потоком кальция через капацитанс-сопряженный канал. Тогда как FcγRIIB выполняет отрицательную регуляцию ITAM-содержащих активирующих рецепторов (Daërоn, M. et al., Immunity. 3: 635-646), было показано, что он оказывает отрицательную регуляцию тирозинкиназного рецептора (RTK) c-kit в процессе контроля RTK-опосредованной клеточной пролиферации (Malbec, О. et al., 1999, J. Immunol. 162: 4424-4429).
Антитела, которые связываются с рецепторами FcγRII, описаны в литературе (Looney et al., (1986) J. Immunol. 136: 1641-1647; Zipf et al., (1983) J. Immunol. 131: 3064-3072; Pulford et al., (1986) Immunology 57: 71-76: Greenman et al., (1991), Mol. Immunol. 28: 1243-1254; Ierino et al., (1993) J. Immunol. 150: 1794-1803; Weinrich et al., (1996) Hybridoma, 15: 109-116; Sonderman et al., (1999) Biochemistry, 38: 8469-8477; Lyden, T. W. et al., (2001) J. Immunol. 3882-3889; а также международная публикация № WO 2004/016750, опубликованная 26 февраля 2004 года). Высокоаффинный рецептор IgER1, FcεRI, вовлечен в сигнальный путь индуцированного антигеном высвобождения гистамина при связывании IgE, например, в ходе аллергической реакции (von Bubnoff, D. et al., (2003) Clinical & Experimental Dermatology. 28(2): 184-187). Было показано, что рецепторы FcγRIIB взаимодействуют с FcεRI и ингибируют его активность через домен FcγRIIB ITIM (Daeron, M. et al., (1995) J. Clin. Invest. 95: 577-585; Malbec, O. et al., (1998) J. Immunol 169: 1647-1658); и Tam, S. W. et al., (2004) Allergy 59: 772-780). Антитела, которые специфически связываются с FcγRIIB человека, необходимы не только для исследовательских целей, но также для влияния на активность FcγRIIB и FcεRI при лечении заболеваний.
Краткое описание сущности изобретения
Настоящее изобретение относится к антигенсвязывающему полипептиду или антителу, которые селективно связываются с FcγRIIB человека. Антигенсвязывающий полипептид или антитело по настоящему изобретению связываются с FcγRIIB человека со значительно более высокой аффинностью, чем с другими FcγR человека, и в некоторых вариантах осуществления они по существу не способны к перекрестной реакции с FcγRIIA человека.
В некоторых вариантах осуществления антигенсвязывающий полипептид или антитело по настоящему изобретению, которые селективно связываются с FcγRIIB человека, содержат по меньшей мере один или несколько CDR (гипервариабельные участки) SEQ ID NO: 1, 2, 3, 4, 5 и 6, а в других вариантах осуществления включает CDR тяжелой цепи SEQ ID No: 1, 2 и 3 и/или CDR легкой цепи SEQ ID No: 4, 5 и 6. В других вариантах осуществления антитело по настоящему изобретению включает один или несколько CDR, которые представляют собой вариант одного или нескольких CDR из SEQ ID No: 1, 2, 3, 4, 5 и 6, где по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 97%, по меньшей мере на 98% или по меньшей мере на 99% идентичен аминокислотной последовательности одного или нескольких CDR SEQ ID NO: 1, 2, 3, 4, 5 и 6. В других вариантах осуществления вариант антигенсвязывающего полипептида или антитела связывается с FcγRIIB с аффинностью, находящейся в пределах от ниже примерно в 10 раз до примерно по меньшей мере в 2 раза, по меньшей мере в 3 раза, по меньшей мере в 5 раз, по меньшей мере в 10 раз, по меньшей мере в 50 раз выше аффинности антитела 5А6 в отношении FcγRIIB, при этом оно по существу не способно к перекрестной реакции с FcγRIIA человека. В других вариантах осуществления антигенсвязывающий полипептид или антитело по настоящему изобретению включает вариабельный домен тяжелой цепи SEQ ID NO: 7 и/или вариабельный домен легкой цепи SEQ ID NO: 8.
В некоторых вариантах осуществления антигенсвязывающий полипептид или антитело по настоящему изобретению представляет собой моноклональное антитело, химерное антитело или гуманизированное антитело, или фрагмент моноклонального, химерного или гуманизированного антитела. В некоторых вариантах осуществления антигенсвязывающий полипептид или антитело по настоящему изобретению, включающий моноклональное, химерное или гуманизированное или полиспецифичные антитела или их фрагменты, получают из антитела, продуцируемого гибридомной клеточной линией с номером доступа в АТСС PTA-4614.
Антигенсвязывающие полипептиды или антитела по настоящему изобретению вводят с терапевтическими антителами или химиотерапевтическими агентами в способах лечения заболевания или расстройства, которое подвергается лечению терапевтическим антителом или химиотерапевтическим агентом.
Настоящее изобретение относится к выделенным биспецифическим антителам, содержащим антитело или антигенсвязывающий полипептид, который селективно связывается с FcγRIIB, включая описанные выше, и второе антитело или антигенсвязывающий полипептид, которые специфически связываются с активирующим рецептором, таким как FcεRI. В некоторых вариантах осуществления биспецифические антитела содержат вариант шарнирной области тяжелой цепи, неспособный к образованию дисульфидной связи между тяжелыми цепями.
Биспецифические антитела по настоящему изобретению можно использовать в способах ингибирования иммунных ответов и подавления высвобождения гистамина, например, связанного с аллергией, астмой и воспалением. В некоторых вариантах осуществления настоящего изобретения биспецифические антитела по настоящему изобретению можно использовать для активации рецептора FcγRIIB в клетках млекопитающих путем совместной агрегации рецептора FcγRIIB с активирующим рецептором в клетке. В некоторых вариантах осуществления клетки млекопитающего представляют собой клетки человека; в других вариантах осуществления клетки человека представляют Т-клетки, В-клетки, тучные клетки, базофилы, антигенпрезентирующие клетки, макрофаги и/или моноциты. В тех вариантах осуществления изобретения, в которых происходит ингибирование белка, опосредованного ITIM, такое ингибирование обычно затрагивает Т-клетки, В-клетки, тучные клетки, базофилы и антигенпрезентирующие клетки. В тех вариантах осуществления изобретения, когда ингибирование опосредовано FcγRIIB, такое ингибирование обычно затрагивает тучные клетки, базофилы, антигенпрезентирующие клетки, моноциты, макрофаги и В-клетки. В некоторых вариантах осуществления настоящего изобретения описываемые в нем биспецифические антитела используются для инактивации, ингибирования активности или для отрицательной регуляции экспрессии рецептора FcεRI. В тех вариантах осуществления, в которых FcεRI ингибируется или подвергается отрицательной регуляции, ингибирование или отрицательная регуляция в типичном случае происходит в тучных клетках, базофилах и антигенпрезентирующих клетках млекопитающего.
В одном из аспектов настоящее изобретение относится к композиции, содержащей выделенное анти-huFcγRIIB/анти-huFcεRI биспецифическое антитело в фармацевтическом носителе. В другом варианте осуществления настоящее изобретение относится к композиции, содержащей выделенное анти-huFcγRIIB/анти-huFcεRI биспецифическое антитело и выделенное анти-IgE антитело. Используемое соотношение биспецифического антитела анти-huFcγRIIB/анти-huFcεRI к анти-IgE антителу в объединенной композиции может быть определено известным способом для каждого пациента. Обычно это соотношение изменяется от примерно 0,01:1 до примерно 100:1. Антитела в композиции могут быть моноклональными, человеческими, гуманизированными или химерными.
В другом аспекте настоящее изобретение относится к терапевтическому способу лечения иммунного расстройства млекопитающего путем введения анти-huFcγRIIB/анти-huFcεRI биспецифического антитела. В одном из вариантов осуществления млекопитающее представляет собой пациента человека, такого как пациент, нуждающийся в лечении аллергического расстройства, астмы и/или воспаления. В другом варианте осуществления терапевтический способ также включает введение млекопитающему, страдающему иммунным расстройством, аллергией или астмой, нуждающемуся в ингибировании высвобождения гистамина, анти-huFcγRIIB/анти-huFcεRI биспецифического антитела по настоящему изобретению. В еще одном из вариантов осуществления анти-huFcγRIIB/анти-huFcεRI биспецифическое антитело по настоящему изобретению вводят в сочетании с анти-IgE антителом, где указанное введение может быть разделено некоторым интервалом времени или может быть одновременным. В одном из вариантов осуществления анти-IgE антитело представляет собой моноклональное антитело. В еще одном из вариантов осуществления анти-IgE антитело представляет собой Xolair®. В еще одном из вариантов осуществления биспецифическое антитело вводят в сочетании с анти-IgE антителом в качестве составной части терапевтического способа лечения имеющегося иммунного расстройства (например, в составе терапевтического режима), где биспецифическое антитело вводят отдельно (не в одно и то же время) от анти-IgE антитела. В другом варианте осуществления биспецифическое антитело по настоящему изобретению и анти-IgE антитело вводят в одно и то же время. Используемое отношение биспецифического антитела анти-huFcγRIIB/анти-huFcεRI к анти-IgE антителу при комбинированном введении (где введение осуществляют в раздельные периоды времени или в одно и то же время) может быть легко определено для каждого пациента. Для целей настоящего изобретения указанное соотношение может составлять от примерно 0,01:1 до примерно 100:1, и в каждом конкретном случае для конкретного пациента используемое соотношение определяют в рамках данного диапазона. Используемое соотношение может представлять, например, 0,05:1, 0,1:1, 0,5:1, 1:1, 1:0,5, 1:0,1 и 1:0,05, и не исключается такое значение используемого соотношения, которое может быть определено стандартными клиническими методами.
Настоящее изобретение, кроме того, относится к выделенной нуклеиновой кислоте, кодирующей антитело, вектор или клетку-хозяин, содержащую указанную нуклеиновую кислоту, к способу получения антитела, в котором предусмотрено культивирование клетки-хозяина и, необязательно, в котором также предусмотрено выделение антитела из культуры клеток-хозяев (например, из клетки-хозяина или культуральной среды, содержащей клетки-хозяева).
Краткое описание чертежей
На фиг.1 схематически изображен нативный IgG. Дисульфидные связи показаны жирной линией между доменами CH1 и CL и двумя доменами CH2. V обозначает вариабельный домен; C обозначает константный домен; L обозначает легкую цепь и H обозначает тяжелую цепь.
На фиг.2А проиллюстрировано выравнивание предпочтительных аминокислотных последовательностей FcγRIIA человека (SEQ ID NO: 9); FcγRIIB2 человека (SEQ ID NO: 10). На фиг.2В показана аминокислотная последовательность FcγRIIB1 (SEQ ID NO: 11).
На фиг.3 проиллюстрировано выравнивание нативной последовательности Fc-участка антитела человека. Приведенные последовательности представляют собой нативную последовательность IgG1 человека (SEQ ID NO: 31), не относящуюся к А-аллотипу; нативную последовательность IgG2 человека (SEQ ID NO: 32); нативную последовательность IgG3 человека (SEQ ID NO: 33); и нативную последовательность IgG4 человека (SEQ ID NO: 34).
На фиг.4 показана диаграмма, иллюстрирующая относительную степень связывания антител с GST-huFcγRIIB в сравнении со слитыми белками GST-huFcγRIIA и GST-huFcγRIII.
На фиг.5 проиллюстрирована специфичность связывания путем выявления иммунофлуоросцентного связывания антител с клетками CHO, экспрессирующими GPI-huFcγRIIB, в сравнении с клетками CHO, экспрессирующими GPI-huFcγRIIA.
На фиг.6-9 представлены кривые аффинности связывания, относящиеся к связыванию разных анти-FcγRII (CD32) МАт с GST-huFcγRIIB, GST-huFcγRIIA(Н131) или GST-huFcγRIIA(R131).
На фиг.10 показаны аминокислотные последовательности легкой и тяжелой цепей моноклонального антитела 5А6.2.1.
На фиг.11-15 показано, что 5А6 не блокирует связывание гексамера E27-IgE с huFcγRIIA, а 5А6 блокирует связывание гексамера E27-IgE с huFcγRIIB.
На фиг.16 проиллюстрирован непрямой иммунофлуоресцентный анализ связывания 5А6 MAb на нативной линии эритролейкоза К562, экспрессирующей FcγRIIA (ATCC № CCL-243).
На фиг.17 продемонстрированы эффекты перекрестного связывания FcγRIIB с активированными рецепторами, выявляемые на основе количественного определения уровня блокирования высвобождения гистамина.
На фиг.18 показаны результаты анти-Fab вестерн-блоттинга для оценки экспрессии компонентов антител p5A6.11.Knob (knob анти-FcγRIIB) и p22E7.11.Hole (hole анти-FcεRI).
На фиг.19 показаны результаты анти-Fc вестерн-блоттинга для оценки экспрессии компонентов антител p5A6.11.Knob (knob анти-FcγRIIB) и p22E7.11.Hole (hole анти-FcεRI).
На фиг.20 показаны результаты анти-Fab вестерн-блоттинга для оценки экспрессии компонентов антител с последовательностями дикого типа или с вариантными последовательностями шарнирной области.
На фиг.21 показаны результаты анти-Fc вестерн-блоттинга для оценки экспрессии компонентов антител с последовательностью дикого типа или с вариантными последовательностями шарнирной области.
На фиг.22 показан результат анализа методом изоэлектрофокусирования 5A6Knob, 22E7Hole, смеси 5A6Knob и 22E7Hole при комнатной температуре и смеси, нагретой до 50°C в течение 5 минут.
На фиг.23 показаны результаты анализа проходящей фракции через аффинную колонку с FcγRIIB для биспецифического антитела 5A6Knob/22E7Hole и антител 22E7Hole и 5A6Knob.
На фиг.24 показаны результаты анализа методом изоэлектрофокусирования смеси 5A6Knob, 22E7Hole и смеси 5A6Knob и 22E7Hole, нагретой до температуры 50°С в течение 10 минут.
На фиг.25 показана последовательность нуклеиновой кислоты (SEQ ID NO: 35), кодирующая промотор щелочной фосфатазы (phoA), STII сигнальную последовательность и полностью (вариабельный и константный домены) легкую цепь антитела 5A6.
На фиг.26 показана последовательность нуклеиновой кислоты (SEQ ID NO: 36), кодирующая промотор щелочной фосфатазы (phoA), сигнальную последовательность STII и полностью (вариабельный и константный домены) легкую цепь антитела 22E7.
На фиг.27 показана последовательность нуклеиновой кислоты (SEQ ID NO: 37), кодирующая последние 3 аминокислоты в сигнальной последовательности STII и примерно 119 аминокислот из вариабельного домена тяжелой цепи антитела 5A6 мыши.
На фиг.28 показана последовательность нуклеиновой кислоты (SEQ ID NO: 38), кодирующая последние 3 аминокислоты в сигнальной последовательности STII и примерно 123 аминокислоты из вариабельного домена тяжелой цепи антитела 22E7 мыши.
На фиг.29 и 30 показаны результаты анализа ELISA, иллюстрирующие двойную связывающую специфичность бесшарнирного биспецифического антитела 5А6/22Е7.
На фиг.31-33 показаны результаты определения по методу ELISA, демонстрирующие способность биспецифического антитела 5А6/22Е7 образовывать перекрестные связи huFcγRIIB с FcεRI.
На фиг.34 приведен график данных анализа ELISA уровня высвобождения гистамина, демонстрирующий блокирование антиген-индуцированного высвобождения гистамина в клетках RBL-huFcεRI+FcγRIIB1 при проведении предварительной инкубации биспецифического антитела 5А6/22Е7 с huFcεRI ECD и huFcγRIIB ECD.
На фиг.35 приведены графики данных FACS уровней связывания биспецифического антитела 5А6/22Е7 в присутствии huFcεRI ECD и huFcγRIIB ECD с клетками RBL-huFcεRI+FcγRIIB1.
На фиг.36 приведен график, иллюстрирующий по данным тестирования методом ELISA уровень высвобождения гистамина в клетках RBL-huFcεRI+FcγRIIB2 при проведении предварительной инкубации биспецифического антитела 5А6/22Е7 с huFcεRI ECD и huFcγRIIB ECD.
На фиг.37 показаны графики, иллюстрирующие по данным тестирования FACS уровни связывания биспецифического антитела 5А6/22Е7 в присутствии huFcεRI ECD и huFcγRIIB ECD с клетками RBL-huFcεRI+FcγRIIB2.
На фиг.38 приведены графики, построенные на основе результатов анализа FACS, которые иллюстрируют блокирование связывания биспецифического антитела 5А6/22Е7 с клетками RBL huFcεRI в присутствии huFcεRI ECD, huFcγRIIB ECD или обоих ECD.
На фиг.39 приведены графики, построенные на основе результатов анализа FACS, которые иллюстрируют блокирование связывания биспецифического антитела 5А6/22Е7 с клетками RBL huFcγRIIB в присутствии huFcεRI ECD, huFcγRIIB ECD или обоих ECD.
На фиг.40 приведены графики, построенные на основе результатов анализа FACS, которые иллюстрируют блокирование связывания биспецифического антитела 5А6/22Е7 с клетками RBL huFcεRI+FcγRIIB1 в присутствии huFcεRI ECD, huFcγRIIB ECD или обоих ECD.
На фиг.41 приведены графики, построенные на основе результатов анализа FACS, которые иллюстрируют блокирование связывания биспецифического антитела 5А6/22Е7 с клетками RBL huFcεRI+FcγRIIB2 в присутствии huFcεRI ECD, huFcγRIIB ECD или обоих ECD.
На фиг.42 приведен график, построенный на основе результатов тестирования высвобождения гистамина анализом ELISA, который демонстрирует ингибирование антиген-индуцированного высвобождения гистамина в клетках RBL huFcεRI+FcγRIIB1 в присутствии биспецифического антитела 5А6/22Е7 в субнасыщающих концентрациях.
На фиг.43 показаны результаты анализа методом проточной цитометрии связывания биспецифического антитела 5А6/22Е7 с клетками RBL huFcεRI+FcγRIIB1.
На фиг.44 показан график результатов анализа ELISA по высвобождению гистамина, который демонстрирует ингибирование индуцированного антигеном высвобождения гистамина в клетках RBL huFcεRI+FcγRIIB2 в присутствии биспецифического антитела 5А6/22Е7 в субнасыщающих концентрациях.
На фиг.45 показаны результаты анализа методом проточной цитометрии связывания биспецифического антитела 5А6/22Е7 с клетками RBL huFcεRI+FcγRIIB2.
На фиг.46А и 46В показаны результаты анализа проточной цитометрии данных титрования при определении связывания биспецифического антитела 5А6/22Е7 с клетками RBL huFcεRI, клетками RBL FcγRIIB, клетками RBL huFcεRI+huFcγRIIB1 и клетками RBL huFcε+FcγRIIB2.
На фиг.47 показан график, иллюстрирующий уровни биспецифического антитела, выявляемые методом ELISA в культуральных клеточных средах в случае клеток RBL FcεRI, клеток RBL FcεRI+FcγRIIB1 и клеток RBL FcεRI+FcγRIIB2 в течение семи дней после обработки IgE в присутствии или в отсутствие биспецифического антитела, которые указывают на то, что истощения по антителам не происходит.
На фиг.48 показан график, иллюстрирующий уровни IgE, выявляемые методом ELISA в клеточных культуральных средах для клеток RBL FcεRI, клеток RBL FcεRI+FcγRIIB1 и клеток RBL FcεRI+FcγRIIB2 в течение семи дней после обработки IgE, в присутствии или в отсутствие биспецифического антитела, которые указывают на то, что истощения по антителам не происходит.
На фиг.49 и 50 представлены результаты анализа проточной цитометрии IgE-индуцированной позитивной регуляции поверхностной экспрессии FcεRI в клетках RBL FcεRI.
На фиг.51 и 52 представлены результаты анализа проточной цитометрии IgE-индуцированной позитивной регуляции поверхностной экспрессии FcεRI в клетках RBL FcεRI+FcγRIIB1.
На фиг.53 и 54 представлены результаты анализа проточной цитометрии IgE-индуцированной позитивной регуляции поверхностной экспрессии FcεRI в клетках RBL FcεRI+FcγRIIB2.
На фиг.55 представлены результаты анализа проточной цитометрии, демонстрирующие эффект биспецифического антитела в направлении отрицательной регуляции поверхностной экспрессии FcεRI в клетках RBL FcεRI после удаления IgE.
На фиг.56 представлены результаты анализа проточной цитометрии, демонстрирующие эффект биспецифического антитела в направлении отрицательной регуляции поверхностной экспрессии FcεRI в клетках RBL FcεRI+FcγRIIB1 после удаления IgE.
На фиг.57 представлены результаты анализа проточной цитометрии, демонстрирующие эффект биспецифического антитела в направлении отрицательной регуляции поверхностной экспрессии FcεRI в клетках RBL FcεRI+FcγRIIB2 после удаления IgE.
На фиг.58-61 показаны результаты анализа методом RT-ПЦР экспрессии мРНК huFcεRIα, FcγRIIB1, FcγRIIB2, huRPL19 (контроль) и FcεRIα крысы в тучных клетках RBL huFcεRI (обозначенных как huFcERIa), клетках RBL huFcεRI+FcγRIIB1 (обозначенных как huFcGRIIb1) и клетках RBL huFcεRI+FcγRIIB2 (обозначенных как huFcGRIIb2) и в базофилах человека от трех разных доноров.
На фиг.62 представлены результаты анализа, в рамках которого анти-IgE-индуцированное высвобождение гистамина в первичных базофилах человека ингибируется анти-FcγRIIB-анти-FcεRI биспецифическим антителом 5А6/22Е7.
На фиг.63 представлены в графическом виде результаты анализа проточной цитометрии, демонстрирующие эффект биспецифического антитела на отрицательную регуляцию IgE-индуцированной поверхностной экспрессии FcεRI в клетках RBL FcεRI+FcγRIIB2 в том случае, когда анти-FcγRIIB-анти-FcεRI биспецифическое антитело 5А6/22Е7 добавляют на нулевой день, третий день и четвертый день.
На фиг.64 представлены результаты тестирований, в которых IgE/антигениндуцированное высвобождение цитокина в клетках RBL FcεRI+FcγRIIB2 ингибируется анти-FcγRIIB-анти-FcεRI биспецифическим антителом 5А6/22Е7. На каждой диаграмме: антиген/IgE, один (NP(11)-OVA+IgE) - темно-серые прямоугольники; антиген/IgE + биспецифическое антитело (NP(11)-OVA+IgE+BsAb) - светло-серые прямоугольники.
На фиг.65 представлены результаты тестирований, в которых IgE/антигениндуцированная каскадная стимуляция арахидоновой кислотой в клетках RBL FcεRI+FcγRIIB1 ингибируется анти-FcγRIIB-анти-FcεRI биспецифическим антителом 5А6/22Е7.
Подробное описание изобретения
I. Определения
Аллергия относится к определенной группе заболеваний, при которых иммунные ответы на антигены окружающей среды вызывают воспаление ткани и дисфункцию органов. Аллергеном может быть любой антиген, который вызывает аллергию. Например, им может выступать сама антигенная молекула или ее источник, так как, например, пыльца, перхоть животных, яд насекомых или пищевой продукт. IgE играет центральную роль в аллергических расстройствах. Высокоаффинные рецепторы IgE (FcεRI) локализованы на тучных клетках и в базофилах, которые служат в качестве антигенных мишеней, стимулирующих дальнейшее высвобождение медиаторов воспаления, продуцирующих многие проявления аллергического заболевания.
IgE-опосредованное воспаление происходит в том случае, когда антиген связывается с антителами IgE, которые занимают рецептор FcεRI на тучных клетках. В течение нескольких минут такое приводит к дегрануляции тучных клеток, которые высвобождают некоторые уже сформированные медиаторы. Затем дегранулированная клетка начинает синтезировать и высвобождать дополнительные медиаторы de novo. Результатом является двухфазный ответ: первоначальный быстрый эффект на кровеносные сосуды, гладкие мышцы и железистую секрецию (быстрая гиперчувствительность), за которой следует позже, через несколько часов, клеточная инфильтрация в вовлеченный в реакцию сайт. IgE-опосредованное воспаление представляет собой механизм, лежащий в основе атопической аллергии (такой как сенная лихорадка, астма и атопический дерматит), системных анафилактических реакций и аллергической крапивницы. В норме оно может выполнять функцию первой линии иммунологической защиты, поскольку вызывает быстрое спазмирование сосудов, облегчает поступление циркулирующих растворимых факторов и клеток в место контакта с антигеном. Многие из наиболее разрушительных свойств аллергического заболевания связаны с действием хемоаттриктирующих лейкоцитов.
Термины «антитело» и иммуноглобулин используются взаимозаменяемо в широком смысле и включают моноклональные антитела (то есть полноразмерные антитела или интактные моноклональные антитела), поликлональные антитела, полиспецифичные антитела (например, биспецифичные антитела, при условии, что они обрадают желательной биологической активностью) и могут также включать некоторые фрагменты антител (как будет далее описано более подробно), такие как антигенсвязывающиеся полипептиды, где указанные полипептиды могут быть фрагментами антител. В одном из вариантов антитела и иммуноглобулины по настоящему изобретению содержат сниженное (меньшее) количество дисульфибных связей. В одном из вариантов антитела и иммуноглобулины по настоящему изобретению включают шарнирную область, в которой по меньшей мере один цистеиновый остаток делают неспособным формировать дисульфидную связь, где дисульфидная связь, предпочтительно, является межмолекулярной, предпочтительно, между двумя тяжелыми цепями. Цистеин шарнирной области можно сделать неспособным формировать дисульфидные связи с помощью любого из большого числа подходящих методов, известных в данной области, ряд из них приведен в настоящем описании и которые включают, но ими не ограничиваются, делецию цистеинового остатка или замещение цистеинового остатка другой аминокислотой.
Антитела (иммуноглобулины) можно распределить на разные классы, в зависимости от аминокислотной последовательности константных доменов тяжелой цепи. Было описано пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM. Они могут быть дополнительно подразделены на подклассы (изотипы), например, IgG-1, IgG-2, IgA-1, IgA-2 и т.п. Константные домены тяжелой цепи, соответствующие каждому классу иммуноглобулинов, обозначают как α, δ, ε, γ и µ для IgA, D, E, G и M соответственно. Субъединичные структуры и трехмерные конфигурации представителей разных классов иммуноглобулинов хорошо известны и описаны, например, в работе Abbas et al., 2000, Cellular and Mol. Immunology, 4th ed. Антитело может быть частью более крупной слитой молекулы, сформированной посредством ковалентной или нековалентной ассоциации антитела с одним или несколькими другими белками или пептидами.
Термины «полноразмерное антитело», «интактное антитело» и «все антитело» используются в настоящем описании взаимозаменяемо и относятся к антителу в его по существу интактной форме, а не к фрагментам антител. В частности, указанные термины относятся к антителу с тяжелой цепью, содержащей Fc-участки. Вариант антитела по настоящему изобретению может представлять собой полноразмерное антитело. Полноразмерное антитело может быть человеческим, гуманизированным, химерным и/или созревшим по аффинности.
«Созревшее по аффинности» антитело представляет собой антитело, содержащее одно или несколько изменений в одном или нескольких CDR, что приводит к повышению аффинности антитела в отношении антигена, в сравнении с исходным антителом, которое не содержит такое(ие) изменение(ия). Предпочтительно, созревшие по аффинности антитела обладают наномолярной, даже пикомолярной аффинностью для мишеневого антигена. Созревшие по аффинности антитела могут быть получены известными процедурами. См., например, работу Marks et al., 1992, Biotechnology 10: 779-783, где описывается созревание по аффинности путем перегруппировки вариабельного домена тяжелой цепи (VH) и вариабельного домена легкой цепи (VL). Случайный мутагенез CDR и/или остатков каркасной области также описан в литературе: например, Barbas, et al., 1994, Proc. Natl. Acad. Sci. USA 91: 3809-3813; Shier et al., 1995, Gene 169: 147-155: Yelton et al., 1995, J. Immunol. 155: 1994-2004; Jackson et al., 1995, J. Immunol. 154(7): 3310-9; и Hawkins et al., 1992, J. Mol. Biol. 226: 889-896.
«Антитело-агонист» представляет собой антитело, которое связывается и активирует антиген, такой как рецептор. В основном способность к активации рецептора у агониста-антитела, по меньшей мере качественно, аналогична (и по существу может быть количественно аналогична) нативному лиганду-агонисту рецептора.
Термин «фрагменты антитела» включает лишь часть интактного антитела, где указанная часть сохраняет по меньшей мере одну или может сохранять большинство или все функции, связанные с частью, которая присутствует в интактном антителе. Фрагмент антитела по настоящему изобретению может включать достаточную часть константного участка для димеризации (или полимеризации) тяжелых цепей, которые обладают сниженной способностью к образованию дисульфидной связи, например, в том случае, когда по меньшей мере один из цистеинов шарнирной области, в норме участвующих в образовании дисульфидной связи между тяжелыми цепями, изменен, как приведено в настоящем описании. В одном из вариантов осуществления фрагмент антитела включает антигенсвязывающий сайт или вариабельные домены интактного антитела и, таким образом, сохраняет способность связываться с антигеном. В другом варианте осуществления фрагмент антитела, например фрагмент, который включает Fc-участок, сохраняет по меньшей мере одну из биологических функций, которые в норме связаны с участком Fc, в случае его присутствия в интактном антителе, такую как FcRn-связывание, модуляция периода полувыведения антитела, ADCC функция и/или связывание с комплементом (например, в том случае, когда антитело имеет профиль гликозилирования, необходимый для ADCC функции или связывания комплемента). Примеры фрагментов антитела включают линейные антитела, одноцепочечные молекулы антител и полиспецифические антитела, образованные из антительных фрагментов.
Термины «антителоопосредованная клеточная цитотоксичность» и «ADCC» относятся к клеточной реакции, при которой неспецифические цитотоксические клетки, которые экспрессируют FcR (такие как природные клетки-киллеры (NK), нейтрофилы и макрофаги), распознают связанное на клетке-мишени антитело и впоследствии вызывают лизис клетки-мишени. NK-клетки, как первичные клетки для ADCC, экспрессируют только FcγRIII, тогда как моноциты экспрессируют FcγRI, FcγRII и FcγRIII. Экспрессия FcR на гематопоэтических клетках описана в таблице 3 на странице 464 Ravetch et al., 1991, Annu. Rev. Immunol. 9: 457-92. Для оценки ADCC активности интересующей молекулы может быть проведен тест на ADCC in vitro, такой как тест, описанный в патентах США 5500362 или 5821337. Эффекторные клетки, которые могут использоваться в таких анализах, включают мононуклеарные клетки периферической крови (МКПК) и природные клетки-киллеры (NK). Альтернативно или дополнительно, ADCC активность интересующей молекулы может быть оценена in vivo, например, на модели животных, такой как описана Clynes et al., 1998, PNAS (USA) 95: 652-656.
«Химерный комплекс антитело-иммуноадгезин» включает молекулу, которая объединяет по меньшей мере один связывающий домен антитела (определенный в настоящем описании) по меньшей мере с одним иммуноадгезином (определенным в данном описании). Примеры химерных комплексов антитело-иммуноадгезин включают биспецифические химерные комплексы CD4-IgG, описанные, например, Berg et al., 1991, PNAS (USA) 88: 4723, и Chamow et al., 1994, J. Immunol 153: 4268.
«Аутоиммунное заболевание» в контексте настоящего описания обозначает незлокачественные заболевания или расстройства, возникающие и направленные против собственных тканей индивидуума. Приведенные в настоящем описании аутоиммунные заболевания конкретно исключают злокачественные или раковые заболевания или состояния, в частности, исключают В-клеточную лимфому, острый лимфобластный лейкоз (ОЛЛ), хронический лимфоцитарный лейкоз (ХЛЛ), лейкемический ретикулез и хронический миелобластный лейкоз. Примеры аутоиммунных заболеваний или расстройств включают, но ими не ограничиваются, воспалительные реакции, такие как воспалительные заболевания кожи, включающие псориаз и дерматит (например, атопический дерматит); системную склеродерму и склероз; реакции, связанные с воспалительным заболеванием кишечника (такие как болезнь Крона или язвенный колит); респираторный дистресс-синдром (включая респираторный дистресс-синдром взрослых; ARDS); дерматит; менингит; энцефалит; увеит; колит; гломерулонефрит; аллергические состояния, такие как экзема и астма, и другие состояния, включающие инфильтрацию Т-клеток и хронические воспалительные реакции; атеросклероз; недостаточность лейкоцитарной адгезии; ревматоидный артрит; системную красную волчанку (SLE); сахарный диабет (например, сахарный диабет типа I или инсулинзависимый диабет); рассеянный склероз; синдром Рейно; аутоиммунный тироидит; аллергический энцефаломиелит; синдром Шегрена; ювенильный диабет; и иммунные реакции, связанные с острой и задержанной гиперчувствительностью, опосредованные цитокинами и Т-лимфоцитами, обычно выявляемые при туберкулезе, саркоидозе, полимиозите, грануломатозе и васкулите; злокачественную анемию (болезнь Аддисона); болезни, вовлекающие лейкоцитарный диапедез; воспалительное расстройство центральной нервной системы (ЦНС); синдром поражения множественных органов; гемолитическую анемию (включая, но ими не ограничиваясь, криоглобунемию или позитивную анемию Кумбса); тяжелую псевдопаралитическую миастению; заболевания, опосредованные образованием комплекса антиген-антитело; заболевание, опосредованное участием антигломерулярной базальной мембраны; антифосфолипидный синдром; аллергический неврит; болезнь Грейвса; миастенический синдром Ламберта-Итона; буллезный пемфигоид; пузырчатку; аутоиммунные полиэндокринопатии; болезнь Рейтера; синдром негнущегося человека; болезнь Бехчета; гигантоклеточный артериит; иммунный комплексный нефрит; IgA нефропатию; IgM полиневропатию; иммунную тромбоцитопеническую пурпуру (ITP) или аутоиммунную тромбоцитопению и т.п.
Термины «биологически активный» или «функциональный» применительно к иммуноглобулину обозначают иммуноглобулин, способный проявлять один или несколько видов природной активности в структурном, регуляторном, биохимическом или биофизическом планах. Например, биологически активное антитело может обладать способностью специфически связываться с антигеном, и такое связывание может проявлять или изменять клеточное или молекулярное событие, такое как сигнальная трансдукция или ферментативная активность. Биологически активное антитело может также блокировать активацию лиганда рецептора или действовать в качестве агониста антитела. Способность антитела проявлять одно или несколько своих природных активностей зависит от нескольких факторов, включающих соответствующую складчатую структуру и сборку полипептидных цепей.
Термин «аффинность связывания» в основном относится к силе суммарных общих нековалентных взаимодействий между одним сайтом связывания молекулы (например, антитела) и его партнером по связыванию (например, антигеном или рецептором FcRn). Аффинность молекулы Х в отношении ее партнера Y может в основном выражаться константой диссоциации (Kd). Аффинность может быть измерена обычными методами, известными в данной области, включая методики, приведенные в настоящем описании. Низкоаффинные антитела связываются с антигеном (или рецептором FcRn) слабо и имеют тенденцию к легкой диссоциации, тогда как высокоаффинные антитела связываются с антигеном (или рецептором FcRn) более прочно и дольше остаются в связанном состоянии.
Термин «блокирующее» антитело или «антитело-антагонист» представляет собой антитело, которое ингибирует или снижает биологическую активность того антигена, с которым оно связывается. Такое блокирование может осуществляться, например, посредством создания препятствий: связыванию лиганда с рецептором, образованию комплекса с рецептором, проявлению тирозинкиназной активности рецептора тирозинкиназы в рецепторном комплексе и/или фосфорилированию тирозинкиназного(ых) остатка(ов) в рецепторе или около рецептора. Например, антитело-антагонист FcγRIIB связывается с FcγRIIB и ингибирует способность IgG связываться с FcγRIIB, что приводит к ингибированию иммуноэффекторного ответа. Предпочтительные блокирующие антитела или антитела-антагонисты по существу или полностью ингибируют биологическую активность антигена.
Термины «рак» и «раковый» относятся или описывают физиологическое состояние у млекопитающего, которое обычно характеризуется нерегулируемым клеточным ростом. Примеры рака включают, но ими не ограничиваются, карциному, лимфому, бластому, саркому и лейкоз. Конкретные примеры таких видов рака включают плоскоклеточный рак, мелкоклеточный рак легкого, немелкоклеточный рак легкого, аденокарциному легкого, плоскоклеточную карциному легкого, рак брюшины, гепатоцеллюлярный рак, рак желудочно-кишечного тракта, рак поджелудочной железы, глиобластому, рак шейки матки, рак яичника, рак печени, рак мочевого пузыря, гепатому, рак молочной железы, рак толстой кишки, колоректальный рак, эндометриальную карциному или карциному матки, карциному слюнной железы, рак почки, рак печени, рак предстательной железы, рак вульвы, рак щитовидной железы, печеночную карциному и разные типы рака головы и шеи.
Термин «химерный» применительно к антителам относится к таким антителам, в которых часть тяжелой и/или легкой цепи идентична или гомологична соответствующим последовательностям в антителах, полученных из конкретного вида или принадлежащих к конкретному классу или подклассу антител, тогда как остаток цепи(ей) идентичен(ны) или гомологичен(ны) соответствующим последовательностям в антителах, полученных от другого вида или принадлежащих к другому классу или подклассу антител, а также относится к фрагментам таких антител, при условии, что они сохраняют желательную биологическую активность (см., например, патент США № 4816567, и Morrison et al., 1984, Proc. Natl. Acad. Sci. USA 81: 6851-6855).
В контексте настоящего описания выражения «клетка», «клеточная линия» и «клеточная культура» используются взаимозаменяемо и все эти обозначения включают в себя соответствующее потомство. Таким образом, слова «трансформанты» и «трансформированные клетки» включают первичные клетки и культуры, полученные из них, независимо от количества переносов. Следует также понимать, что все потомство не может быть полностью идентично по содержанию ДНК в связи с наличием слабых или невредных мутаций. Данным термином также охватывается мутантное потомство, которое имеет ту же самую функцию или такую же биологическую активность, как следует из скрининга исходной трансформированной клетки. В том случае, когда будут подразумеваться конкретные определения, это будет ясно из контекста.
Выражение «контрольные последовательности» относится к последовательностям ДНК, необходимым для экспрессии функционально связанной кодирующей последовательности в конкретном организме хозяина. Контрольные последовательности, которые подходят для прокариотов, включают, например, промотор, необязательно последовательность оператора и сайт связывания рибосомы. Известно, что эукариотические клетки используют промоторы, сигналы полиаденилирования и энхансеры.
Термин «расстройство» относится к любому состоянию, при котором может быть достигнут эффект при лечении терапевтическим антителом. Такое расстройство включает хроническое и острое расстройство или заболевание, включая патологическоие состояния, которые создают предрасположенность млекопитающего к рассматриваемому расстройству. В одном из вариантов расстройство представляет собой рак или аутоиммунное заболевание.
Термин «внеклеточный домен» в контексте настоящего описания обозначает такой участок трансмембранного полипептида, как, например, FcR, который является внешним для клетки.
Термины «рецептор Fc» или «FcR» используются для описания рецептора, который связывается с Fc-участком антитела. Предпочтительный FcR представляет собой нативную последовательность FcR человека. Кроме того, предпочтительный FcR представляет собой такой участок, который связывается с антителом IgG (гамма-рецептор) и включает рецепторы подклассов FcγRI, FcγRII и FcγRIII, в том числе аллельные варианты и альтернативно сплайсинговые формы указанных рецепторов. Другие FcR, включая те из них, которые могут быть идентифицированы в будущем, охватываются термином «FcR» по настоящему описанию. Термин также включает неонатальный рецептор, FcRn, который отвечает за перенос материнских IgG в плод (см. Guyer et al., 1976, J. Immunol. 117: 587, и Kim et al., 1994, J. Immunol. 24: 249).
Термин «Fc-участок» в контексте настоящего описания обозначает С-концевой участок тяжелой цепи иммуноглобулина. «Fc-участок» представляет собой нативную последовательность Fc-участка или вариант Fc-участка. Несмотря на то, что границы Fc-участка тяжелой цепи иммуноглобулина могут варьировать, Fc-участок тяжелой цепи IgG человека обычно определяется как протяженный от аминокислотного остатка в положении Cys226 или от Pro230 до его карбоксильного конца. Fc-участок иммуноглобулина в основном включает два константных домена, CH2 и CH3, как показано на фиг.1. «Функциональный Fc-участок» обладает «эффекторной функцией» нативной последовательности Fc-участка. Примеры эффекторных функций включают связывание C1q; комплементзависимую цитотоксичность; связывание Fc-рецептора; антителозависимую клеточную цитотоксичность (ADCC); фагоцитоз; отрицательную регуляцию рецепторов клеточной поверхности (например, B-клеточного рецептора; BCR); и т.п. Для осуществления таких эффекторных функций в основном необходимо, чтобы Fc-участок был объединен со связывающим доменом (например, вариабельным доменом антитела), и они могут быть оценены с использованием различных тестов, таких как, например, тесты, приведенные в настоящем описании. Выражение «нативная последовательность Fc-участка» включает аминокислотную последовательность, идентичную природной аминокислотной последовательности Fc-участка. Нативная последовательность в Fc-участках человека показана на фиг.3 и включает нативную последовательность Fc-участка IgG1 человека (не относящуюся к А и аллотипам А), нативную последовательность Fc-участка IgG2 человека; нативную последовательность Fc-участка IgG3 человека; и нативную последовательность Fc-участка IgG4 человека, а также их природные варианты. Выражение «вариант Fc-участка» включает аминокислотную последовательность, которая отличается от нативной последовательности Fc-участка по меньшей мере одной «модификацией аминокислоты», как приведено в настоящем описании. Fc-участок варианта может содержать по меньшей мере одно замещение аминокислоты в сравнении с нативной последовательностью Fc-участка или с Fc-участком исходного антитела, и может содержать, например, от примерно одного до примерно десяти аминокислотных замещений или от примерно одного до примерно пяти аминокислотных замещений в нативной последовательности Fc-участка или в Fc-участке исходного антитела. Fc-участок варианта может обладать по меньшей мере приблизительно 80% идентичностью с нативной последовательностью Fc-участка и/или с Fc-участком исходного антитела и может характеризоваться по меньшей мере приблизительно 90% идентичностью с ними, или может характеризоваться по меньшей мере приблизительно 95% идентичностью с ними.
Термин «FcγRIIA», если особо не указано иное, относится к FcγRIIA человека (huFcγRIIA), полипептиду, кодируемому геном FcγRIIA человека, и включает, но ими не ограничивается, FcγRIIA1 и FcγRIIA2, а также их аллельные варианты. FcγRIIA человека представляет собой «активирующий» FcR и включает мотив активации тирозинсодержащего иммунорецептора (ITAM) в своем цитоплазматическом домене. Наиболее предпочтительный FcγRIIA человека представляет собой FcγRIIA1 человека, включающий аминокислотную последовательность SEQ ID NO: 9 или ее аллельные варианты, включая их высокоотзывчивые (HR) и низкоотзывчивые (LR) аллельные варианты.
Термин «FcγRIIB», если особо не указано иное, относится к полипептиду, кодируемому геном FcγRIIB человека, и включает, без ограничения, FcγRIIB1, FcγRIIB2, FcγRIIB3 и их аллельные варианты. Предпочтительный FcγRIIB представляет собой «ингибирующий» рецептор FcR, который включает мотив ингибирования тирозинсодержащего иммунорецептора (ITIM) (I/V/LxYxxL/V) (Sathish et al., 2001, J. Immunol. 166: 1763) в своем цитоплазматическом домене. Предпочтительный FcγRIIB человека представляет собой FcγRIIB2 человека (huFcγRIIB2) или FcγRIIB1 (huFcγRIIB1), включающий аминокислотную последовательность SEQ ID NO: 10 или SEQ ID NO: 11, соответственно, и их аллельные варианты. Последовательности FcγRIIB1 и B2 отличаются друг от друга вставкой из 19 аминокислот в цитоплазматическом домене FcγRIIB1, LPGYPECREMGETLPEKPA (SEQ ID NO: 29).
Выражение «Fc-зависимое состояние» в контексте настоящего описания включает воспаление типа II, IgE-опосредованную аллергию, астму, анафилаксию, аутоиммунное заболевание, IgG-опосредованную цитотоксичность или сыпь.
Термин «шарнирная область» и его вариации, используемые в настоящем описании, включают в себя значения, известные в данной области и которые описаны в ряде работ (Janeway et al., 1999, Immuno Biology: The Immune System in Health and Disease, Elsevier Science Ltd., NY. 4th ed.; Bloom et al., 1997, Protein Science, 6: 407-415; Humphreys et al., 1997, J. Immunol. Methods, 209: 193-202).
Термин «гомология» в контексте настоящего описания определяется как процент остатков в варианте аминокислотной последовательности, которые идентичны после выравнивания последовательностей и введения гэпов, по необходимости, для достижения максимального процента гомологии. Методы и компьютерные программы, применяемые для выравнивания, известны в данной области. Одна из таких компьютерных программ включает «Align 2», принадлежащую Genentech, Inc., зарегистрированную пользователем документации в США (United States Copyright Office, Washington, DC 20559) 10 декабря 1991 года.
Термин «клетка-хозяин» (или «рекомбинантная клетка-хозяин») в контексте настоящего описания относится к клетке, которая была генетически изменена или которая способна к генетическому изменению за счет добавления экзогенного полинуклеотида, такого как рекомбинантная плазмида или вектор. Следует понимать, что термины относятся не только к конкретной клетке, но и к потомству такой клетки. Поскольку в последующих поколениях могут происходить некоторые модификации, за счет мутации или воздействия окружающей среды, такое потомство может фактически быть неидентичным исходной клетке, но также охватывается областью термина «клетка-хозяин» в контексте настоящего описания.
Выражение «эффекторные клетки человека» включает лейкоциты, которые экспрессируют один или несколько FcR и осуществляют эффекторные функции. Предпочтительно, клетки экспрессируют по меньшей мере один FcγRIII и осуществляют эффекторную функцию ADCC. Примеры лейкоцитов человека, которые вовлекаются в ADCC, включают мононуклеарные клетки периферической крови (МНПК), природные клетки-киллеры (NK), моноциты, цитотоксические Т-клетки и нейтрофилы, при этом МНПК и NK-клетки являются предпочтительными. Эффекторные клетки могут быть выделены из нативного источника, например, из крови или МНПК (мононуклеарные клетки периферической крови) по приведенным в настоящем описании методикам.
«Гуманизированные» формы антител, отличных от человека (например, мышиных), представляют собой химерные антитела, которые содержат минимальную последовательность, полученную из иммуноглобулина, взятого из источника, отличного от человека. По большей части, гуманизированные антитела представляют собой иммуноглобулины человека (реципиентные антитела), в которых остатки из гипервариабельного участка реципиента замещены остатками гипервариабельного участка, взятого из вида, отличного от человека (донорское антитело), такого как мышь, крыса, кролик или примат, отличный от человека, обладающий желательной специфичностью, аффинностью и функциональной способностью. В некоторых случаях остатки Fv каркасной области (FR) иммуноглобулина человека замещают соответствующими остатками из вида, отличного от человека. Кроме того, гуманизированные антитела могут включать остатки, которые не найдены в реципиентном антителе или в донорском антителе. Такие модификации вводят для дополнительного улучшения характеристик антитела. В основном, гуманизированное антитело включает по существу все из по меньшей мере одного, и в типичном случае двух вариабельных доменов, в которых все или по существу все гипервариабельные петли соответствуют таковым в иммуноглобулине из источника, отличного от человека, и все или по существу все области FR представляют собой области, взятые из последовательности иммуноглобулина человека. Гуманизированное антитело необязательно также включает по меньшей мере часть константной области иммуноглобулина (Fc), в типичном случае иммуноглобулина человека. Дополнительные детали описаны в ряде имеющихся в литературе работ (Jones et al., Nature. 321: 522-525 (1986); Riechmann et al., 1995, Nature 332: 323-329 (1988); and Presta, Curr. Op. Struct. Biol. 2: 593-596 (1992)).
«Антитело человека» представляет собой антитело, которое имеет аминокислотную последовательность, соответствующую таковой в антителе, образуемом в организме человека, и/или которое было получено с использованием методик для получения антител человека, приведенных в настоящем описании. В данное определение не входит понятие гуманизированного антитела, которое включает остатки антигенсвязывающей области из источника, отличного от человека.
В контексте настоящего описания термин «гипергликемические расстройства» относится ко всем формам диабета и расстройствам, возникающим в результате инсулинрезистентности, таких как диабет типа I и типа II, а также тяжелая форма инсулинрезистентности, гиперинсулинемия и гиперлипидемия, например при ожирении и при инсулинрезистентном диабете, таком как синдром Менденхалла, синдром Вернера, лепрейконизм, липоатрофный диабет и другие виды липодистрофий. Конкретное гипергликемическое расстройство, рассматриваемое в настоящем описании, представляет собой диабет, в особенности диабет типа I и типа II. Термин «диабет» сам по себе относится к прогрессирующему заболеванию, связанному с нарушением углеводного обмена, который включает неадекватную продукцию или использование инсулина и характеризуется гипергликемией и глюкозурией.
Термин «гипервариабельный участок» в контексте настоящего описания относится к аминокислотным остаткам антитела, которые ответственны за связывание с антигеном. Гипервариабельный участок включает аминокислотные остатки из «участка, определяющего комплементарность» или «CDR», определяемые при сопоставлении последовательностей, например остатки 24-34 (L1), 50-56 (L2) и 89-97 (L3) в вариабельном домене легкой цепи и 31-35 (Н1), 50-65 (Н2) и 95-102 (Н3) в вариабельном домене тяжелой цепи (см., Kabat et al., 1991, Sequences of Proteins of Immunological Interest, 5th ed. Public Health Service, National Institutes of Health, Bethesda, MD) и/или те участки из гипервариабельный петли (HVL), которые определяются по структурным характеристкам, например остатки 26-32 (L1), 50-52 (L2) и 91-96 (L3) в вариабельном домене легкой цепи и 26-32 (Н1), 53-55 (Н2) и 96-101 (Н3) в вариабельном домене тяжелой цепи (см., Chothia and Leskl, 1987, J. Mol. Biol. 196: 901-917). Остатки «каркасной области» или «FR» представляют собой остатки вариабельного домена, которые отличаются от остатков гипервариабельного участка, определенного в настоящем описании.
Иммунные и воспалительные заболевания включают ревматоидный артрит, остеоартрит, ювенильный хронический артрит, спондилоартропатии, системный склероз (склеродерму), идиопатические воспалительные миопатии (дерматомиозит), системный васкулит, саркоидоз, аутоиммунную гемолитическую анемию (иммунную панцитопению, пароксизмальную ночную гемоглобинурию), аутоиммунную тромбоцитопению (идиопатическую тромбоцитопеническую пурпуру, иммунную тромбоцитопению), тиреоидит (болезнь Грейва, тиреоидит Хашимото, ювенильный лимфоцитарный тиреоидит, атрофический тиреоидит), аутоиммунные воспалительные заболевания (например, аллергический энцефаломиелит), рассеянный склероз, инсулинзависимый сахарный диабет, аутоиммунный увеоретинит, тиреотоксикоз, аутоиммунное заболевание щитовидной железы, злокачественную анемию, реакцию отторжения аутотрансплантата, сахарный диабет и болезнь почек иммунной природы (гломерулонефрит, канальцево-интерстициальный нефрит), демиелинизирующие заболевания центральной и периферической нервных систем, такие как рассеянный склероз, идиопатическая демиелинизирующая полиневропатия или синдром Гийллиана-Барра и хроническая воспалительная демиелинизирующая полиневропатия; гепатобилиарное заболевание, такое как инфекционный гепатит (гепатит A, B, C, D, E и заболевания, вызванные другими негепатотропными вирусами), аутоиммунный хронический активный гепатит, первичный билиарный цирроз, грануломатозный гепатит и склерозирующий холангит, глютенчувствительную энтеропатию и болезнь Уипла; аутоиммунные или иммуноопосредованные болезни кожи, включающие буллезные заболевания кожи, мультиформную эритему и контактный дерматит, псориаз; аллергические заболевания, такие как астма, аллергический ринит, атопический дерматит, весенний конъюнктивит, экзема, пищевая гиперчувствительность и крапивница; иммунологические болезни легких, такие как эозинофильная пневмония, идиопатический легочный фиброз и пневмонит по типу гиперчувствительности; заболевания, связанные с трансплантацией, включающие реакцию отторжения трансплантата и болезнь «трансплантат против хозяина».
В контексте настоящего описания термин «иммуноадгезин» обозначает молекулы антительного типа, которые объединяют «связывающий домен» гитерологичного белка «адгезина» (например, рецептор, лиганд или фермент) с эффекторными функциями константного домена иммуноглобулина. Структурно иммуноадгезины включают слияние аминокислотной последовательности адгезина с желательной связывающей специфичностью, которая отличается от сайта распознавания и связывания антигена (антигенсвязывающий сайт) антитела (то есть «гетерологичной»), и последовательности константного домена иммуноглобулина. Последовательность константного домена иммуноглобулина в иммуноадгезине предпочтительно получают из γ1, γ2 или γ4 тяжелых цепей, поскольку иммуноадгезины, включающие данные участки, могут быть очищены хроматографией на белке А. (см., например, Lindmark et al., 1983, J. Immunol. Meth. 62: 1-13).
«Выделенное антитело» представляет собой антитело, которое было идентифицировано и отделено и/или восстановлено из компонентов природного окружения. Контаминирующие компоненты его природного окружения представляют собой материалы, которые могут препятствовать диагностическому или терапевтическому использованию антитела и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества. В некоторых вариантах антитело может быть очищено (1) до более 95 вес.% антитела, определяемого по методу Лоури, и более предпочтительно, до более 99 вес.% антитела, (2) до степени, достаточной для получения по меньшей мере 15 остатков N-концевой или внутренней аминокислотной последовательности при использовании центрифужного секвенатора или (3) до гомогенности, по данным анализа в SDS-PAGE, в восстановительных или невосстановительных условиях, с использованием красителя Кумаси синего или, предпочтительно, серебряного красителя. Выделенное антитело включает антитело in situ в рекомбинантных клетках, поскольку по меньшей мере один компонент природного окружения антитела не будет присутствовать. Однако обычно для выделения антитела проводят по меньшей мере одну стадию очистки.
«Выделенная» молекула нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты, которая была идентифицирована и отделена по меньшей мере от одной контаминирующей молекулы нуклеиновой кислоты, с которой она обычно связана в природном источнике данной нуклеиновой кислоты для антитела. Выделенная молекула нуклеиновой кислоты отличается по форме или по другим характеристикам от молекулы, которая встречается в природе. В этой связи, выделенная молекула нуклеиновой кислоты отличается от молекулы нуклеиновой кислоты, имеющейся в природных условиях. Однако выделенная молекула нуклеиновой кислоты включает молекулу нуклеиновой кислоты, содержащуюся в клетках, которая обычно экспрессирует антитело, где по меньшей мере одна молекула нуклеиновой кислоты находится в хромосоме в месте, отличном от места, характерного для клеток в природных условиях.
Термин «млекопитающее» включает любое животное, классифицированное как млекопитающее, включая человека, коров, лошадей, собак и кошек. В одном варианте осуществления настоящего изобретения указанное млекопитающее представляет собой человека.
Термин «моноклональное антитело» в контексте настоящего описания относится к антителу, получаемому из популяции по существу гомогенных антител, например индивидуальных антител, включая популяции, которые идентичны, за исключением возможных встречающихся в природе мутаций, и которые могут присутствовать в минорных количествах. Моноклональные антитела представляют собой высокоспецифические антитела, направленные против одного антигенного сайта. Кроме того, в отличие от обычных (поликлональных) препаратов антител, которые в типичном случае включают разные антитела, направленные против разных детерминант (эпитотов), каждое моноклональное антитело направлено против одной детерминанты на антигене. Определение «моноклональный» указывает на характер антитела, получаемого из по существу гомогенной популяции антител, и его не следует трактовать как указывающий на необходимость продукции антитела каким-то конкретным способом. Например, моноклональные антитела, используемые в соответствии с настоящим изобретением, могут быть получены по методу гибридом, впервые описанному Kohler et al., 1975, Nature. 256: 495, или могут быть получены с использованием методик рекомбинантной ДНК (см., например, патент США № 4816567). «Моноклональные антитела» могут быть также выделены из библиотек фаговых антител с использованием методик, описанных, например, Clackson et al., 1991, Nature 352: 624-628; и Marks et al., 1991, J. Mol. Biol. 222: 581-597.
Моноклональные антитела в контексте настоящего описания включают конкретно «химерные» антитела (иммуноглобулины), в которых часть тяжелой и/или легкой цепей идентична или гомологична соответствующим последовательностям в антителах, полученных из конкретных видов или принадлежащих к конкретному классу или подклассу антитела, тогда как остаток цепи(ей) идентичен или гомологичен соответствующим последовательностям в антителах, полученных из другого вида или принадлежащих к другому классу или подклассу антител, а также фрагменты таких антител, при условии, что они демонстрируют желательную биологическую активность (патент США № 4816567; Morrison et al., 1984, Proc. Natl. Acad. Sci. USA 81: 6851-6855).
В контексте настоящего описания нуклеиновая кислота рассматривается как «оперативно связанная», если она поставлена в функциональную взаимосвязь с другой последовательностью нуклеиновой кислоты. Например, ДНК для предпоследовательности или секреторного лидера оперативно связана с ДНК для антитела, если она экспрессируется в виде предбелка, который участвует в секреции антитела; промотор или энхансер оперативно связан с кодирующей последовательностью, если он воздействует на транскрипцию последовательности; или сайт связывания рибосомы оперативно связан с кодирующей последовательностью, если он расположен так, что облегчает трансляцию. В основном, термин «оперативно связанные» означает, что последовательности ДНК связывают таким образом, что они становятся непрерывными и в случае секреторного лидера становятся непрерывными и находятся в одной фазе считывания. Однако, в случае энхансера непрерывность может быть не соблюдена. Связывание сопровождается лигированием в подходящих сайтах рестрикции. Если таких сайтов не существует, используют синтетические олигонуклеотидные адапторы и линкеры в соответствии с традиционной практикой.
Для целей настоящего описания термин «фармацевтическая композиция» обозначает такую фармацевтическую композицию, которая адаптирована и является приемлемой для введения млекопитающим, в особенности человеку. Таким образом, композиция может использоваться для лечения заболевания или расстройства у млекопитающего. Кроме того, белок в композиции может быть подвержен одной или нескольким стадиям выделения или очистки, так что контаминант(ы), который мог препятствовать его терапевтическому использованию, от него отделяется. В основном, фармацевтическая композиция включает терапевтический белок и терапевтически приемлемый носитель или разбавитель. Композиция обычно является стерильной и может быть лиофилизирована. Ниже следует подробное описание фармацевтических препаратов.
Термины «полинуклеотид» или «нуклеиновая кислота» используются в настоящем описании взаимозаменяемо и относятся к полимерам из нуклеотидов любой длины и включают ДНК и РНК. Нуклеотиды могут представлять собой дезоксирибонукдеотиды, рибонуклеотиды, модифицированные нуклеотиды или основания и/или их аналоги, или любой субстрат, который может быть включен в полимер с помощью ДНК- или РНК-полимеразы или в ходе реакции синтеза. Полинуклеотид может включать модифицированные нуклеотиды, такие как метилированные нуклеотиды и их аналоги. В случае их наличия, модификация нуклеотидной структуры может быть введена до или после сборки полимера. Последовательность нуклеотидов может быть нарушена компонентами ненуклеотидной природы. Полинуклеотид после синтеза может быть далее модифицирован, например, посредством конъюгации с меткой. Другие типы модификации включают, например, «кэпы», замещение одного или нескольких природных нуклеотидов аналогом, межнуклеотидные модификации, такие как, например, модификации, вводимые с помощью незаряженных связок (например, с использованием метилфосфонатов, сложных фосфотриэфиров, фосфоамидатов, карбаматов и т.п.) или с использованием заряженных связок (например, фосфоротиоатов, фосфородитиоатов и т.п.), связок, содержащих дополнительные фрагменты, такие как, например, белки (например, нуклеазы, токсины, антитела, сигнальные пептиды, ply-L-лизин и т.п.), с использованием интеркаляторов (например, акридина, псоралена и т.п.), с использованием хелаторных связок (например, с использованием металлов, радиоактивных металлов, бора, окисляющих металлов и т.п.), с использованием алкилаторов, с модифицированными связками (например, с использованием альфа-аномерных нуклеиновых кислот и т.п.), а также немодифицированных форм полинуклеотида(ов). Кроме того, может быть замещена любая гидроксильная группа, обычно присутствующая в сахаре, например, фосфонатными группами, фосфатными группами, а также может быть защищена стандартными защитными группами или может быть активирована с получением дополнительных связей к дополнительным нуклеотидам или может быть конъюгирована с твердыми или полутвердыми подложками. 5'- и 3'-концевые группы OH могут быть фосфорилированы или замещены аминами или органическими кэпинг-фрагментами, содержащими от 1 до 20 атомов углерода. Другие гидроксилы могут быть также дериватизированы с помощью стандартных защитных групп. Полинуклеотиды могут также содержать аналогичные формы рибозных или дезоксирибозных сахаров, которые в основном известны в данной области, включая, например, 2'-О-метил-, 2'-О-аллил-, 2'-фтор-, или 2'-азидорибозу, карбоциклические сахарные аналоги, α-аномерные сахара, эпимерные сахара, такие как арабиноза, ксилоза или ликсоза, пиранозные сахара, фуранозные сахара, седогептулозу, ациклические аналоги и абазиновые нуклеозидные аналоги, такие как метилрибозид. Одна или несколько сложных фосфодиэфирных связей могут быть замещены альтернативными связующими группами. Указанные альтернативные связующие группы включают, но ими не ограничиваются, варианты, в которых фосфат замещается P(O)S («тиоатом»), P(S)S («дитиоатом»), (O)NR2 («амидатом»), P(O)R, P(O)OR', CO или CH2 («формацеталем»), где каждый R или R' обозначает независимо H или замещенный или незамещенный (С1-20)-алкил, необязательно содержащий эфирную (-О-) связь, арил, алкенил, циклоалкил, циклоалкенил или аралдил. Необязательно, чтобы все связи в полинуклеотиде были идентичны. Предшествующее описание применимо ко всем полинуклеотидам, рассматриваемым в настоящей заявке, включая РНК и ДНК.
Термин «олигонуклеотид» в контексте настоящего описания в основном относится к коротким, в основном одноцепочечным, в основном синтетическим полинуклеотидам, которые в основном, но необязательно, содержат менее примерно 200 нуклеотидов в длину. Термины «олигонуклеотид» и «полинуклеотид» не яввляются взаимоисключающими. Приведенное выше описание для полинуклеотидов в равной мере и полностью применимо к олигонуклеотидам.
Фразы «сигнальная последовательность секреции» или «сигнальная последовательность» относятся к последовательности нуклеиновой кислоты, кодирующей короткий сигнальный пептид, который может использоваться для направления нового синтезированного интересующего белка через клеточную мембрану, обычно внутреннюю мембрану, или и через внутреннюю, и через внешнюю мембраны прокариотов. В данном случае интересующий белок, такой как легкая или тяжелая полипептидная цепь иммуноглобулина, секретируется в периплазму прокариотических клеток-хозяев или в культуральную среду. Сигнальный пептид, кодируемый сигнальной последовательностью секреции, может быть эндогенным для клеток-хозяев или такие пептиды могут быть экзогенными, включая сигнальные пептиды, которые являются нативными для экспрессируемого полипептида. Сигнальные последовательности секреции в типичном случае присутствуют на аминоконце экспрессируемого полипептида и в типичном случае удаляются энзиматически между фазами биосинтеза полипептида и его секреции из цитоплазмы. Таким образом, сигнальный пептид обычно не присутствует в зрелом белковом продукте.
Термин «домен связывания рецептора» используется для обозначения любого нативного лиганда для рецептора, включая молекулы клеточной адгезии или любой участок или производное такого нативного лиганда, сохраняющие по меньшей мере качественную способность к связыванию рецептора для соответствующего нативного лиганда. Указанное определение, в числе других, конкретно включает связывающиеся последовательности лигандов для указанных выше рецепторов.
В контексте настоящего описания термин «терапевтическое антитело» представляет собой антитело, которое эффективно при лечении заболевания или расстройства у млекопитающего, имеющего данное заболевание или расстройство, или предрасположенное к ним. Репрезентативные терапевтические антитела включают 5А6 анти-FcγRIIB антитело по настоящему изобретению и биспецифическое анти-FcγRIIB/анти-FcεRI антитело по настоящему изобретению, а также антитела, включающие rhuMAb 4D5 (HERCEPTIN®) (Carter et al., 1992, Proc. Natl. Acad. Sci. USA 89: 4285-4289; патент США № 5725856); анти-CD20 антитела, такие как химерное анти-CD20 «C2B8», описанное в патенте США № 5736137 (RITUXAN®), химерный или гуманизированный вариант 2Н7 антитела, описанный в патенте США № 5721108, B1 или Тозитумомаб (Tositumomab) (BEXXAR®); анти-IL-8 (St John et al., 1993, Chest. 103: 932 и международная публикация № WO95/23865); анти-VEGF антитела, включающие гуманизированные и/или созревшие по аффинности анти-VEGF антитела, такие как гуманизированное анти-VEGF антитело huA4.6.1 AVASTINTM (Kim, et al., 1992, Growth Factors, 7: 53-64, международная публикация № WO9630046 и WO 98/45331, опубликованная 15 октября 1998 года); анти-PSCA антитела (WO01/40309); анти-CD40 антитела, включающие S2C6 и их гуманизированные варианты (WO00/75348); анти-CD11a (патент США № 5622700, WO98/23761; Steppe, et al., 1991, Transplant Intl. 4: 3-7, и Hourmant et al., 1994, Transplantation 58: 377-380); анти-IgE (Presta et al., 1993, J. Immunol., 151: 2623-2632; и международная публикация № WO 95/19181); анти-CD18 (патент США № 5622700, выданный 22 апреля 1997 года, или WO97/26912, опубликованная 31 июля 1997 года); анти-IgE (патент США № 5714338, выданный 3 февраля 1998 года, или патент США № 5091313, выданный 25 февраля 1992 года, WO93/04173, опубликованная 4 марта 1993 года, или заявка на международный патент № PCT/US98/13410, зарегистрированный 30 июня 1998 года, патент США №5714338); антитело против Apo-2 рецептора (WO98/51793, опубликованная 19 ноября 1998 года); анти-TNF-α антитела, включающие сА2 (REMICADE®), CDP571 и MAK-195 (см. патент США №5672347, выданный 30 сентября 1997 года, Lorenz et al., 1996, J. Immunol. 156(4): 1646-1653, и Dhainaut et al., 1995, Crit. Care Med. 23(9): 1461-1469); антитканевый фактор (TF) (Европейский патент № 0420937 В1, выданный 9 ноября 1994 года); α4-β7 интегрин против молекулы человека (WO98/06248, опубликованная 19 февраля 1998 года); анти-EGFR (химеризованное или гуманизированное 225 антитело, описанное в WO 96/40210, опубликованной 19 декабря 1996 года); анти-CD3 антитела, такие как OKT3 (патент США № 4515893, выданный 7 мая 1985 года); анти-CD25 или анти-tac антитела, такие как CHI-621 (SIMULECT®) и (ZENAPAX®) (см. патент США № 5693762, выданный 2 декабря 1997 года); анти-CD4 антитела, такие как cM-7412 антитело (Choy et al., 1996, Arthritis Rheum. 39(1): 52-56); анти-CD52 антитела, такие как CAMPATH-1H (Riechmann et al., 1988, Nature 332: 323-337); антитела против Fc-рецептора, такие как М22 антитело, направленное против Fc (RI, описанное, например, в работе Graziano et al., 1995, J. Immunol. 155(10): 4996-5002); антитела против карциноэмбрионического антигена (CEA), такие как hMN-14 (Sharkey et al., 1995, Cancer Res. 55 (23 Suppl): 5935s-5945s); антитела против эпителиальных клеток молочной железы, включающие huBrE-3, hu-Mc 3 и CHL6 (Ceriani et al., 1995, Cancer Res. 55(23): 5852s-5856s; и Richman et al., 1995, Cancer Res. 55(23 Suppl): 5916s-5920s); антитела, которые связываются с клетками карциномы толстой кишки, такие как С242 (Litton et al., 1996, Eur.J. Immunol. 26(1): 1-9); анти-CD38 антитела, например, АТ 13/5 (Ellis et al., 1995, J. Immunol. 155(2): 926-937); анти-CD33 антитела, такие как Hu M195 (Jurcic et al., 1995, Cancer Res. 55 (23 Suppl): 5908s-5910s) и CMA-676 или CDP771; анти-CD22 антитела, такие как LL2 или LymphoCide (Juweid et al., 1995, Cancer Res. 55(23 Suppl): 5899s-5907s); анти-EpCAM антитела, такие как 17-1А (PANOREX®); анти-GpIIb/IIIa антитела, такие как абциксимаб (abciximab) или с7Е3 Fab (REOPRO®); анти-RSV антитела, такие как MEDI-493 (SYNAGIS®); анти-CMV антитела, такие как PROTOVIR®; анти-HIV антитела, такие как PRO-542; антитела против гепатита, такие как анти-Hep B антитело OSTAVIR®; анти-CA 125 антитело OvaRex; антитело против идиотипического GD3 эпитопа BEC2; анти-αvβ3 антитела VITAXIN®; антитело против карциномы почечных клеток человека, такое как ch-G250; ING-1; антитело 17-1А против компонента человека (3622W94); антитело против колоректальной опухоли человека (А33); антитело R24 против меланомы человека, направленное против GD3 ганглиозида; антитело против плоскоклеточной карциномы человека (SF-25); и антитела против лейкоцитарного антигена человека (HLA), такие как Smart ID10 и анти-HLA DR антитело Oncolym (Lym-1).
Термин «терапевтически эффективное количество» относится к количеству композиции по настоящему изобретению, которое эффективно для «ослабления» или «лечения» заболевания или расстройства у субъекта или млекопитающего. В одном варианте, если подлежащее лечению заболевание представляет собой заболевание, опосредованное участием В-клеток, это количество означает такое количество, которое приводит к снижению числа В-клеток (истощение по В-клеткам) у млекопитающего.
Термин «лечение» в контексте настоящего описания относится к эффективному способу «лечения» или «воздействия» на заболевание или расстройство у субъекта или млекопитающего. В основном лечение заболевания или расстройства относится к уменьшению одного или нескольких симптомов или медицинских проблем, связанных с заболеванием или расстройством. В некоторых вариантах антитела и композиции по настоящему изобретению могут использоваться для предупреждения начала или возобновления заболевания или расстройства у субъекта или млекопитающего. Например, у субъекта с аутоиммунным заболеванием антитело по настоящему изобретению может использоваться для предупреждения или лечения внезапных обострений болезни. Последовательное лечение или введение относится к лечению по меньшей мере на ежедневной основе без прерывания лечения на один или несколько дней. Перемежающееся лечение или введение, или лечение или введение в перемежающемся режиме, относится к лечению, которое не является непрерывным, но, скорее, циклическим по своей природе. Режим лечения в контексте настоящего изобретения может быть последовательным или перемежающимся.
Термин «вариантная» или «измененная» тяжелая цепь в контексте настоящего описания в основном относится к тяжелой цепи со сниженной способностью к образованию дисульфидных связей, например, когда по меньшей мере один цистеиновый остаток становится неспособным к образованию дисульфидной связи. Предпочтительно, такой по меньшей мере один цистеиновый остаток находится в шарнирной области тяжелой цепи.
Термин «вектор» в контексте настоящего изобретения относится к молекуле нуклеиновой кислоты, способной транспортировать другую молекулу нуклеиновой кислоты, к которой она присоединена. Одним типом вектора является «плазмида», кольцевая двуцепочечная петля ДНК, внутри которой могут быть лигированы дополнительные сегменты ДНК. Другой тип вектора представляет собой фаговый вектор. Еще один тип вектора представляет вирусный вектор, в случае которого дополнительные сегменты ДНК могут быть лигированы при введении их в состав вирусного генома. Некоторые векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, имеющие бактериальный ориджин репликации, и эписомальные векторы млекопитающих). Другие векторы (например, неэпипосомальные векторы млекопитающих) могут быть интегрированы в геном клетки-хозяина при их введении в клетку-хозяин и, таким образом, они будут реплицироваться вместе с геномом-хозяином. Кроме того, некоторые векторы способны направлять экспрессию генов, с которыми они оперативно связаны. Такие векторы в тексте настоящего описания обозначаются как «рекомбинантные векторы экспрессии» (или просто «рекомбинантные векторы»). В основном, векторы экспрессии, используемые в методиках рекомбинантных ДНК, зачастую имеют форму плазмид. В контексте настоящего описания термины «плазмида» и «вектор» используются взаимозаменяемо, поскольку плазмида является наиболее часто используемой формой вектора.
Антитело, которое «селективно связывается с FcγRIIB человека», связывается с FcγRIIB человека со значительно большей аффинностью, чем оно связывается с другими FcγR человека. В некоторых вариантах антитело, которое селективно связывается с FcγRIIB человека, связывается и с FcγRIIB1, и с FcγRIIB2, и демонстрирует небольшое или вообще отсутствие связывания с FcγRIIA, FcγRI и FcγRIII, и их аллельными вариантами. Относительная способность к связыванию и/или аффинность по связыванию может быть продемонстрирована в рамках множества методик, известных в данной области, включающих, но ими не ограничиваясь, иммуноферментный твердофазный анализ (ELISA) и флуоресцентный сортировщик иммунных клеток (FACS). В основном это означает, что антитело по настоящему изобретению связывается с FcγRIIB с реакционной способностью, которая по меньшей мере на приблизительно один логарифм выше по концентрации, чем оно связывается с FcγRIIA, по данным анализа метода ELISA.
Предпочтительно, антитело, которое связывается с FcγRIIB человека, селективно относительно FcγRIIA человека, по существу, не способно к перекрестной реакции с FcγRIIA человека.
В контексте настоящего описания антитело, которое «по существу не способно к перекрестной реакции с FcγRIIA человека», представляет собой антитело, для которого уровень связывания с FcγRIIA человека будет составлять менее 10% от уровня его связывания с FcγRIIB, альтернативно, меньше 8%, альтернативно, меньше 6%, альтернативно, меньше 4%, альтернативно, меньше 2%, альтернативно, меньше 1%, по уровню связывания с FcγRIIA человека относительно уровня связывания с FcγRIIB, при определении по методу сортировки активированных клеток на основе флуоресценции (FACS) или при тестировании по методу радиоиммунопреципитации (RIA).
В контексте настоящего описания антитело, которое «является антагонистом связывания Fc-участка с FcγRIIB человека», блокирует или препятствует связыванию Fc-участка (например, Fc-участка антитела, такого как IgG или иммуноадгезин, или другая Fc-содержащая конструкция) с FcγRIIB человека. Такая антагонистическая активность может быть определена, например, по методу ELISA.
II. Способы осуществления изобретения
А. Получение анти-FcγRIIB антитела
(i) FcγRIIB антиген
Растворимый FcγRIIB человека или его фрагменты, необязательно конъюгированные с другими молекулами, могут использоваться в качестве иммуногенов для получения антител. Примеры иммуногенов включают белки слияния, содержащие внеклеточный домен FcγRIIB1 или FcγRIIB2 с белком-носителем или меткой аффинности, такой как GST или His6. Альтернативно или дополнительно, клетки, экспрессирующие FcγRIIB человека, могут использоваться в качестве иммуногена. Такие клетки могут быть получены из природного источника или могут представлять собой клетки, трансформированные рекомбинантными методиками для экспрессии FcγRIIB человека. Специалистам в данной области известны другие формы FcγRIIB человека, применяемые для получения антител.
(ii) Поликлональные антитела
Поликлональные антитела предпочтительно получают у животных путем множественных подкожных (п/к) или внутрибрюшинных (в/б) инъекций соответствующего антигена и адъюванта. Для этой цели может использоваться конъюгация соответствующего антигена с белком, который является иммуногеном для подлежащего иммунизации вида, например с гемоцианином из моллюска Negathura crenulata, сывороточным альбумином, бычьим тиреоглобулином или соевым ингибитором трипсина, с использованием бифункционального или дериватизирующего агента, например сложного малеимидобензоилсульфосукцинимидного эфира (конъюгация идет через цистеиновые остатки), N-гидроксисукцинимида (конъюгация идет через лизиновые остатки), глютаральдегида, янтарного ангидрида, SOCl2 или R1N=C=NR, где R и R1 обозначают разные алкильные группы.
Животных иммунизируют антигеном, иммуногенными конъюгатами или производными при объединении, например, 100 мкг или 5 мкг белка или конъюгата (для кролика или мыши соответственно) с 3 объемам полного адъюванта Фрейнда с последующей инъекцией раствора внутрикожно во множественные сайты. Примерно через один месяц животных подвергают бустингу с использованием 1/5-1/10 от исходного количества пептида или конъюгата в полном адъюванте Фрейнда путем подкожной инъекции во множественные сайты. Через 7-14 дней у животных отбирают кровь и сыворотку исследуют на титр антител. Животных подвергают бустингу до достижения по титру стадии плато. Предпочтительно, животных подвергают бустингу конгъюгатом одного и того же антигена, но конъюгированного с другим белком и/или при использовании другого сшивающего реагента. Конъюгаты могут быть также получены в рекомбинантной клеточной культуре в виде белков слияния. Кроме того, соответствующим образом могут использоваться агрегирующие агенты, такие как глинозем, для усиления иммунного ответа.
(iii) Моноклональные антитела
Моноклональные антитела могут быть получены с использованием метода гибридом, описанного Kohler et al., 1975, Nature, 256:495, или могут быть получены по методикам рекомбинантной ДНК (см., например, патент США №4816567).
В соответствии с гибридомной методикой мышь или другое соответствующее животное-хозяин, такое как хомяк или обезьяна макака, иммунизируют, как было описано выше, для проявления действия лимфоцитов, которые продуцируют или способны продуцировать антитела, специфически связывающиеся с белком, используемым для иммунизации. Альтернативно, лимфоциты могут быть иммунизированы in vitro. Далее лимфоциты сливают с миеломными клетками, с использованием подходящего агента для слияния, такого как полиэтиленгликоль, с образованием гибридомной клетки (Goding, 1986, Monoclonal Antibodies: Principles and Practice, pp. 59-103 (Academic Press)).
Полученные таким образом гибридомные клетки высевают и растят в подходящей культуральной среде, которая предпочтительно содержит один или несколько субстратов, ингибирующих рост или выживание неслитых исходных миеломных клеток. Например, если исходные миеломные клетки не содержали фермент гипоксантингуанинфосфорибозилтрансферазу (HGPRT или HPRT), культуральная среда для гибридом будет в типичном случае включать гипоксантин, аминоптерин и тимидин (HAT среда), где указанные вещества будут препятствовать росту HGPRT-дефицитных клеток.
Предпочтительные миеломные клетки включают такие клетки, которые эффективно сливаются, поддерживают на высоком уровне стабильную продукцию антитела выбранными антитело-продуцирующими клетками и чувствительны к среде, такой как среда HAT. В числе них предпочтительные миеломные клеточные линии включают миеломные линии мышей, такие как линии, получаемые из MOPC-21 и MPC-11 опухолей мышей, доступных от Центра в Сан Диего, поставляющего клеточные линии (Salk Institute Cell Distribution Center, San Diego, California USA), и клетки SP-2 или X63-Ag8-653, доступные от Американской Коллекции типовых культур (American Type Culture Collection, Rockville, Maryland USA). Миеломные клеточные линии человека и гетеромиеломные мыши-человека также были описаны в контексте их применения для целей получения моноклональных антител человека (Kozbor, 1984, J. Immunol., 133:3001; Brodeur et al., 1987, Monoclonal Antibody Techniques and Applications, pp. 51-63 (Marcel Dekker, Inc., New York)).
Культуральную среду, в которой растят гибридомные клетки, анализируют на предмет выявления продукции моноклональных антител, направленных против данного антигена. Предпочтительно, специфичность по связыванию моноклональных антител, продуцируемых гибридомными клетками, определяют иммунопреципитацией или в тесте на связывание in vitro, таком как радиоиммуноанализ (РИА) или иммуноферментный твердофазный анализ (ELISA).
После идентификации гибридомных клеток как продуцирующих антитела с желательной специфичностью, аффинностью и/или активностью, соответствующие клоны могут быть далее подвергнуты субклонированию по процедурам предельных разведений и далее их растят с использованием стандартных методик (Goding, supra). Подходящие для данной цели культуральные среды включают, например, среду
DMEM или RPMI-1640. Дополнительно гибридомные клетки могут выращиваться in vivo в виде асцитных опухолей у животных.
Моноклональные антитела, секретированные субклонами, соответствующим образом отделяют от культуральной среды, асцитной жидкости или сыворотки с использованием для этого традиционных методов очистки иммуноглобулинов, такими как, например, хроматография с белком А-сефарозой, хроматография на гидроксилапатите, гель-электрофорез, диализ или аффинная хроматография.
ДНК, кодирующую моноклональные антитела, выделяют и секвенируют в рамках известных процедур (например, с использованием олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими тяжелую и легкую цепи моноклональных антител). Гибридомные клетки служат в качестве предпочтительного источника такой ДНК. ДНК, после ее выделения, может быть введена в векторы экспрессии, которые далее используют для трансфекции клеток-хозяев, таких как клетки E. coli., COS клетки обезьяны, клетки яичника китайского хомяка (CHO) или миеломные клетки, которые в ином случае не продуцируют иммуноглобулиновый белок, с достижением синтеза моноклональных антител в рекомбинантных клетках-хозяевах. Ниже более подробно описана рекомбинантная процедура получения таких антител.
В еще одном варианте осуществления настоящего изобретения антитела или фрагменты антител могут быть выделены из библиотек фаговых антител, создаваемых с использованием методик, описанных в литературе (McCafferty et al., 1990, Nature, 348: 552-554). Clackson et al., 1991, Nature, 352: 624-628; и Marks et al., 1991, J. Mol. Biol., 222: 581-597) описывают процедуру выделения антител мыши и человека, соответственно, с использованием фаговых библиотек. Последующая очистка приводит к образованию антител человека с высокой аффинностью (в нМ диапазоне) путем перегруппировки цепей (Marks et al., 1992, Bio/Technology,10: 779-783), а также посредством комбинаторной инфекции и рекомбинации in vivo, используемой в качестве основной стратегии конструирования очень крупных фаговых библиотек (Waterhouse et al., 1993, Nuc. Acids. Res. 21: 2265-2266). Таким образом, указанные методики представляют собой жизнеспособную альтернативу традиционной стратегии получения моноклональных антител, представляющей собой гибридомный метод выделения моноклональных антител.
ДНК может быть также модифицирована, например, путем проведения замещения кодирующей последовательности для константных доменов тяжелой и легкой цепей вместо гомологичных последовательностей мыши (патент США № 4816567; Morrison et al., 1984, Proc. Natl. Acad. Sci. USA, 81: 6851) или путем ковалентного присоединения к кодирующей последовательности иммуноглобулина полностью или части кодирующей последовательности для неиммуноглобулинового материала (например, для белковых доменов).
В типичном случае такой материал, не относящийся к иммуноглобулину, замещают константными доменами антитела или подвергают замещению вариабельными доменами одного антигенобъединенного сайта антитела с образованием химерного бивалентного антитела, включающего один антиген-объединенный сайт со специфичностью в отношении одного антигена и другой антиген-объединенный сайт со специфичностью в отношении другого антигена.
(iv) Гуманизированные и человеческие антитела
Гуманизированное антитело содержит один или несколько аминокислотных остатков, которые происходят из источника, отличного от человеческого. Аминокислотные остатки из источника, отличного от человеческого, часто называют «импортными» остатками и в типичном случае отбирают из «импортного» вариабельного домена. Гуманизация может быть проведена в основном по методике Winter and co-workers Jones et al., 1986, Nature, 321: 522-525; Riechmann et al., 1988, Nature, 332: 323-327; Verhoeyen et al., 1988, Science, 239: 1534-1536 путем замещения одной или нескольких CDR или CDR последовательностей грызунов на соответствующие последовательности антитела человека. Соответственно, такие «гуманизированные» антитела будут представлять собой химерные антитела (патент США № 4816567), в которых по существу менее одного интактного вариабельного домена антитела человека замещены соответствующей последовательностью, взятой из вида, отличного от человека. Практически, гуманизированные антитела в типичном случае представляют собой антитела человека, в которых некоторые остатки CDR и, возможно, остатки FR замещены остатками, взятыми из аналогичных сайтов молекул, отличных от человеческих, например из антител грызунов.
Выбор вариабельных доменов молекулы человека как легкой, так и тяжелой цепей, для использования с целью получения гуманизированных антител очень важен для снижения антигенности. В соответствии с так называемым методом наилучшей подгонки, или «best-fit», последовательность вариабельного домена в антителе грызунов подвергают скринингу с использованием полной библиотеки известных последовательностей вариабельных доменов человека. Далее последовательность человека, которая более всего соответствует последовательности грызунов, выбирают в качестве каркасной области молекулы человека (FR) для создаваемого гуманизированного антитела (Sims et al., 1987, J. Immunol, 151: 2296; Chothia et al., 1987, J. Mol. Biol.,196; 901). В другом способе используют определенную каркасную область, полученную из консенсусной последовательности всех антител человека определенной подгруппы легкой и тяжелой цепей. Одна и та же каркасная область может использоваться для нескольких разных гуманизированных антител (Carter et al., 1992, Proc. Natl. Acad. Sci. USA 89: 4285; Presta et al., 1993, J. Immunol, 151: 2623).
Важно также, чтобы антитела, подвергаемые гуманизации, сохраняли высокую аффинность в отношении антигена и другие полезные биологические свойства. Для достижения данной цели, в соответствии с предпочтительным методом, гуманизированные антитела получают по способу, включающему анализ исходных последовательностей и различных концептуальных гуманизированных продуктов с использованием трехмерных моделей исходной и гуманизированной последовательностей. Трехмерные иммуноглобулиновые модели в основном доступны и известны специалистам в данной области. Доступны также компьютерные программы, которые иллюстрируют и показывают в виде рисунков вероятные трехмерные конформационные структуры выбранных предполагаемых иммуноглобулиновых последовательностей. Осмотр таких рисунков позволяет провести анализ вероятной роли остатков в функционировании предполагаемой иммуноглобулиновой последовательности, например анализ остатков, которые будут влиять на способность предполагаемой молекулы иммуноглобулина связываться с антигеном. В рамках данного способа остатки FR могут быть отобраны из реципиентной и импортной последовательностей и объединены таким образом, чтобы была достигнута желательная характеристика, такая как повышенная аффинность для антигена-мишени. В целом, остатки CDR оказывают непосредственное и очень существенное воздействие на связывание с антигеном.
Альтернативно, в настоящее время имеется возможность создавать трансгенных животных (например, мышей), которые при иммунизации способны продуцировать полный репертуар антител человека, в отсутствие эндогенной продукции иммуноглобулинов. Например, было показано, что гомозиготная делеция в гене присоединяющего участка (JH) тяжелой цепи антитела в химерной и зародышевой линиях мутантных мышей приводит к полному ингибированию эндогенной продукции антитела. Перенос генной совокупности для иммуноглобулина зародышевой линии человека в такую зародышевую линии мутантных мышей будет приводить к продукции антител человека при антигенной провокации (см., например, Jakobovits et al., 1993, Proc. Natl. Acad. Sci. USA, 90: 2551; Jakobovits et al., 1993, Nature, 362: 255-258; Bruggermann et al., 1993, Year in Immuno, 7: 33; и Duchosal et al., 1992, Nature, 355: 258). Антитела человека также могут быть получены из фаговых библиотек (Hoogenboom et al., 1991, J. Mol. Biol. 227: 381; Marks et al., 1991, J. Mol. Biol., 1991, 222: 581-597; Vaughan et al., 1996, Nature Biotech, 14: 309).
(v) Полиспецифические антитела
Полиспецифические антитела характеризуются специфичностью по связыванию, относящейся по меньшей мере к двум разным антигенам. В норме такие молекулы связываются только с двумя антигенами (то есть это биспецифические антитела, БсАт), но в контексте настоящего описания и антитела с дополнительными специфичностями, такие как триспецифические антитела, также охватываются данным выражением. Примеры БсАт включают такие антитела, которые имеют один антигенсвязывающий сайт, направленный против FcγRIIB, и другой антигенсвязывающий сайт, направленный против B-клеточного рецептора (BCR), CD79α и/или CD79β, антигена, экспрессируемого на опухолевой клетке, IgE рецептора (FcεR), IgE, сопряженного с IgER, такого как рецептор на тучных клетках и/или базофилах, IgG рецепторов RI (FcγRI) и RIII (FcγRIII), таких как рецепторы на NK, моноцитах и макрофагах, с-набора рецепторов тирозинкиназы. В некоторых вариантах BsAb включают первую связывающую специфичность для FcγRIIB и вторую связывающую специфичность для активирующего рецептора, содержащего цитоплазматический мотив ITAM. Структура мотива ITAM включает два тирозина, разделенных спейсером из 9-11 аминокислот. Общая консенсусная последовательность представляет собой XxxL/I(x)6-8YxxL (Isakov, N., 1997, J. Leukoc. Biol., 61; 6-16). Примеры активирующих рецепторов включают FcεRI, FcγRIII, FcγRI, FcγRIIA и FcγRIIC. Другие активирующие рецепторы включают, например, CD3, CD2, CD10, CD161, DAP-12, KAR, KARAP, FcεRII, Trem-1, Trem-2, CD28, p44, p46, рецептор В-клеток, LMP2A, STAM, STAM-2, GPVI и CD40 (см., например, Azzoni, et al., 1998, J. Immunol, 161: 3493; Kita, et al., 1999, J. Immunol, 162: 6901; Merchant, et al., 2000, J. Biol. Chem, 74: 9115; Pandey, et al., 2000, J. Biol. Chem, 275: 38633; Zheng, et al., 2001, J. Biol. Chem, 276: 12999; Propst, et al., 2000, J. Immunol, 165: 2214).
В одном варианте BsAb включают первую связывающую специфичность для FcγRIIB и вторую связывающую специфичность для FcεRI. Биспецифические антитела могут быть получены в виде антител полной длины или фрагментов антител (например, биспецифических антител F(ab')2). Биспецифические антитела могут быть получены дополнительно в виде антител по типу «выступы во впадинах», knobs-in-holes, или безшарнирных антител. Биспецифические антитела приведены Segal et al., 2001, J. Immunol, 248: 1-6.
Способы получения биспецифических антител известны в данной области. Традиционная продукция биспецифических антител полной длины основана на совместной экспрессии двух пар тяжелой цепи-легкой цепи иммуноглобулина, где указанные две цепи обладают разными специфичностями (Millstein et al., 1988, Naure, 305: 537-539). В результате статистической пересортировки тяжелой и легкой цепей иммуноглобулина указанные гибридомы (квадромы) становятся способными продуцировать потенциальную смесь из 10 разных антительных молекул, среди которых только одна содержит корректную биспецифическую структуру. Очистка корректной молекулы, производимая обычно с включением стадии аффинной хроматографии, представляет собой достаточно трудоемкий процесс и дает небольшой выход. Аналогичные процедуры описаны в WO 93/08829 и в Traunecker et al., 1991 EMBO J., 10: 3655-3659.
В соответствии с другим подходом вариабельные домены антитела с желательными специфичностями по связыванию (объединенные сайты антитело-антиген) сливают с последовательностями константных доменов иммуноглобулина. Указанное слияние может производиться с использованием константного домена тяжелой цепи иммуноглобулина, включающего по меньшей мере часть шарнира, участки CH2 и CH3. Предпочтительно иметь первый константный участок тяжелой цепи (CH1), содержащий сайт, необходимый для связывания легкой цепи, который присутствует по меньшей мере в одной из объединяемых молекул. ДНК, кодирующие объединяемые тяжелые цепи иммуноглобулина и, при желании, легкие цепи иммуноглобулина, встраивают в отдельные векторы экспрессии и подвергают совместной трансфекции в соответствующий организм-хозяин. Такая процедура обеспечивает высокую гибкость в корректировке общих пропорций трех антительных фрагментов в тех вариантах, когда оптимальный выход дает использование в конструкции неравных частей трех антительных цепей. Однако возможно встраивать кодирующую последовательность для двух или трех антительных цепей в одном векторе экспрессии, когда экспрессия по меньшей мере двух антительных цепей, взятых в равных пропорциях, приводит к высокому выходу или когда пропорции не имеют особого значения.
В другом варианте осуществления данного подхода используемые биспецифические антитела состоят из гибридной тяжелой цепи иммуноглобулина с первой связывающей специфичностью в одном плече и гибридной пары тяжелая цепь-легкая цепь иммуноглобулина (обеспечивающей вторую связывающую специфичностью) в другом плече. Было обнаружено, что такая асимметричная структура облегчает отделение желательного биспецифического соединения от нежелательных сочетаний иммуноглобулиновых цепей, поскольку наличие легкой цепи иммуноглобулина только в одной половине биспецифической молекулы позволяет осуществить такое отделение по упрощенному способу. Данный подход описан в WO 94/04690. Дополнительные детали указанной методики, применяемой для получения биспецифических антител, описаны Suresh et al., 1986, Methods in Enzymology, 121: 210.
В соответствии с другим подходом, описанным в WO 96/27011, интерфейс между парой антительных молекул может быть сконструирован таким образом, чтобы максимизировать процент гетеродимеров, которые выделяют из рекомбинантной клеточной культуры. Предпочтительный интерфейс включает по меньшей мере часть CH3 константного домена антитела. В рамках данного метода, одну или несколько небольших боковых цепей аминокислот из интерфейса первой антительной молекулы заменяют более крупными боковыми цепями (например, тирозина или триптофана). Компенсаторные «полости» идентичного или близкого размера относительно крупной боковой цепи(ей) создают на интерфейсе второй антительной молекулы путем замещения крупных боковых цепей аминокислот более мелкими (например, аланина или треонина). Это обеспечивает механизм повышения выхода гетеродимера относительно других, нежелательных конечных продуктов, таких как гомодимеры.
Биспецифические антитела включают сшитые или «гетероконъюгатные» антитела. Например, одно из антител в гетероконъюгате может быть связано с авидином, второе - с биотином. Такие антитела создаются с той целью, чтобы клетки иммунной системы могли воздействовать на нежелательные клетки (патент США № 4676980), а также с целью лечения ВИЧ-инфекции (WO 91/00360, WO92/200373 и EP 03089). Гетероконъюгатные антитела могут быть созданы с использованием традиционных методик сшивки. Соответствующие сшивающие агенты известны в данной области и описаны, например, в патенте США № 4676980, где приведено также множество других методик для сшивания.
Описываются также антитела, содержащие более двух валентностей. Например, могут быть получены триспецифические антитела по методу Tutt et al., 1991, J. Immunol, 147: 60.
(vi) Антитела с вариантными шарнирными областями
Антитела по настоящему изобретению могут также включать вариантные тяжелые цепи, например, как описано в серийной заявке № 10/697995, зарегистрированной 30 октября 2003 года. Антитела, включающие вариантные тяжелые цепи, содержат изменение по меньшей мере одного цистеинового остатка, образующего дисульфидную связь, так что такой цистениновый остаток становится уже не способным образовывать дисульфидную связь. В одном аспекте указанный(ые) цистеин(ы) локализован(ы) в шарнирной области тяжелой цепи (и такая шарнирная область в контексте настоящего описания может обозначаться как «вариантная шарнирная область», и такое антитело может также называться «бесшарнирным»).
В некоторых аспектах такие иммуноглобулины не содержат полный репертуар цистеиновых остатков тяжелой цепи, которые в норме способны образовывать дисульфидные связи либо в межмолекулярном варианте (такие, как связи между двумя тяжелыми цепями), либо во внутримолекулярном (такие, как связи между двумя цистеиновыми остатками в одной полипептидной цепи). В основном и предпочтительно, дисульфидная связь, образованная цистеиновым(ими) остатком(ами), который(ые) был(и) изменен(ы) (то есть придали неспособность к образованию дисульфидных связей) представляет собой такую связь, которая не присутствует в антителе и не приводит к существенному снижению нормальных физико-химических и/или биологических характеристик иммуноглобулина. Предпочтительно, но необязательно, цистеиновый остаток, который становится неспособным образовать дисульфидные связи, представляет собой цистеин в шарнирной области тяжелой цепи.
Антитело с вариантными тяжелыми цепями или вариантной шарнирной областью в основном получают путем экспрессии в клетке-хозяине антитела, в котором удалены по меньшей мере одна, по меньшей мере две, по меньшей мере три, по меньшей мере четыре или от двух до одиннадцати дисульфидных связей между тяжелыми цепями, с последующим выделением указанного антитела из клетки-хозяина. Экспрессия указанного антитела может быть осуществлена на основе полинуклеотида, кодирующего антитело, где указанное антитело включает вариант тяжелой цепи со сниженной способностью к образованию дисульфидных связей, с последующим выделением указанного антитела из клетки-хозяина, включающей данный полинуклеотид. Предпочтительно, указанная тяжелая цепь включает вариант шарнирной области тяжелой цепи иммуноглобулина, где по меньшей мере один цистеин из вариантной шарнирной области становится неспособным к образованию дисульфидной связи.
Ожидается, что любой цистеин в тяжелой цепи иммуноглобулина можно сделать неспособным к образованию дисульфидной связи аналогично тому, как было описано для цистеинов в шарнирной области, при условии, что такое изменение не будет существенно снижать биологическую функцию иммуноглобулина. Например, IgM и IgE не включают шарнирную область, но каждый из них содержит дополнительный домен в тяжелой цепи; по меньшей мере один (в некоторых вариантах все) цистеин в тяжелой цепи можно сделать неспособным к образованию дисульфидной связи, в соответствии с методиками настоящего изобретения, при условии, что существенно не будет снижаться биологическая функция тяжелой цепи и/или антитела, которое включает такую тяжелую цепь.
Цистеиновые остатки из шарнирной области тяжелой цепи хорошо изучены (см., например, Kabat, 1991, «Sequences of proteins of immunological interest», выше). Как известно в данной области, количество цистеиновых остатков в шарнирной области варьирует в зависимости от класса и подкласса иммуноглобулина (см., например, Janeway, 1999, Immunobiology, 4th Ed., (Garland Publishing, NY)). Например, в IgGI человека два цистеиновых остатка в шарнирной области разделены двумя пролинами и они могут нормально спариваться со своими партнерами на соседней тяжелой цепи через межмолекулярные дисульфидные связи. Другие примеры включают IgG2 человека, которые содержат 4 цистеина в шарнирной области, IgG3, которые содержат 11 цистеинов в шарнирной области, и IgG4, который содержит 2 цистеина в шарнирной области.
Соответственно, методики по настоящему изобретению включают экспрессию в клетку-хозяин тяжелой цепи иммуноглобулина, включающей вариант по шарнирной области, где по меньшей мере один цистеин в варианте шарнирной области становится неспособным к образованию дисульфидной связи, что позволяет тяжелой цепи образовать комплекс с легкой цепью с образованием биологически активного антитела, с последующим выделением такого антитела из клетки-хозяина.
Альтернативные варианты осуществления настоящего изобретения включают такие варианты, в которых по меньшей мере 2, 3 или 4 цистеина становятся неспособными к образованию дисульфидных связей, где от примерно двух до примерно одиннадцати цистеинов становится неспособными к образованию дисульфидных связей и где все цистеины в вариантной шарнирной области становятся неспособными к образованию дисульфидных связей.
Легкие цепи и тяжелые цепи, составляющие антитела по настоящему изобретению, получаемые по описываемым в изобретении методикам, могут кодироваться одним полинуклеотидом или разными полинуклеотидами.
Цистеины, обычно вовлекаемые в образование дисульфидных связей, можно сделать неспособными к образованию дисульфидных связей с помощью множества методик, известных в данной области, или таких методик, которые очевидны как подходящие для этой цели или могут стать известными специалистам в данной области, на основе описанных в данной заявке критериев. Например, цистеин в шарнирной области может быть заменен другой аминокислотой, такой как серин, который неспособен к образованию дисульфидной связи. Замещение аминокислот может быть осуществлено по стандартным методикам молекулярной биологии, включающим, например, направленный мутагенез последовательности нуклеиновой кислоты, кодирующей шарнирную область, подлежащую модификации. Подходящие методики включают методики, описанные Sambrook et al., 1989, Molecular Cloning: A Laboratory Manual, 2nd Ed. Другие методики получения иммуноглобулина с вариантом по шарнирной области включают синтез олигонуклеотида, который кодирует шарнирную область, где кодон для цистеина заменяют кодоном для замещающей аминокислоты. Такой олигонуклеотид может быть далее лигирован внутри векторного скелета, включающего другие соответствующие антительные последовательности, такие как вариабельные участки и Fc-последовательности, как это будет приемлемо.
В другом варианте осуществления настоящего изобретения цистеин в шарнирной области может быть делетирован. Аминокислотная делеция может быть введена по стандартным методикам молекулярной биологии, таким как направленный мутагенез последовательности нуклеиновой кислоты, кодирующей шарнирную область, подлежащую модификации. Подходящие методики включают методики, описанные Sambrook et al., выше. Другие методики получения иммуноглобулина с вариантом по шарнирной области включают синтез олигонуклеотида, содержащего последовательность, которая кодирует шарнирную область, где кодон для цистеина, подлежащего модификации, делетирован. Такой олигонуклеотид может быть далее лигирован внутри векторного скелета, включающего другие подходящие антительные последовательности, такие как вариабельные участки и Fc-последовательности, как это будет приемлемо.
(vii) Биспецифические антитела, образуемые в рамках стратегии «продвижения в полость»
В некоторых вариантах биспецифические антитела по настоящему изобретению формируют с использованием стратегии так называемого «продвижения в полость», или иначе стратегии создания «выступов во впадинах» (knobs into holes), которая служит для создания интерфейса между первым и вторым полипептидом, используемыми для гетероолигомеризации. Предпочтительный интерфейс включает по меньшей мере часть CH3 домена из константного домена антитела. Мутация по типу «выступы в впадине» (knobs into holes) в CH3 домене Fc-последовательности, как было показано, существенно снижает образование гомодимеров (см., например, Merchant et al., 1998, Nature Biotechnology, 16: 677-681). «Выступы» конструируют путем замены малых боковых цепей аминокислот из интерфейса первого полипептида более крупными боковыми цепями (например, тирозина или триптофана). Компенсаторные «полости» идентичного или аналогичного размера для таких выступов могут быть необязательно созданы на интерфейсе второго полипептида путем замены крупных боковых цепей аминокислот более мелкими (например, аланина или треонина). В том случае, когда в интерфейсе первого или второго полипептида имеются соответствующим образом расположенные и имеющие определенные размеры выступ или полость, необходимо только создать подходящие полость или выступ соответственно на соседнем интерфейсе. Выступ и полость могут быть получены с помощью синтеза, такого как синтез измененной нуклеиновой кислоты, кодирующей полипептиды, или путем пептидного синтеза. Дополнительное описание выступов и впадин приведено в патентах США №№ 5731168, 5807706, 5821333.
В некоторых вариантах технология создания «выступов и впадин» используется для усиления гетеродимеризации с целью создания биспецифических анти-FcγRIIB антител и антител против «активирующего рецептора» (например, IgER) полной длины. В одном варианте получают конструкции для анти-FcγIIB компонента (например, p5A6.11.Knob) путем внедрения мутации «knob» (T366W) в Fc-участок, и для анти-IgER компонента (например, p22E7.11.Hole) - путем введения «hole» мутации (T366S, L368A, Y407V). В другом варианте получают конструкцию для анти-FcγIIB компонента (например, p5A6.11.Hole) путем внедрения «hole» мутации в его Fc-участок, и для анти-IgER компонента (например, p22E7.11.Knob) путем внедрения «knob» мутации в его Fc-участок, например, с помощью процедур, приведенных в настоящем описании, или процедур, описанных в литературе (Merchant et al., (1998) выше) или в патентах США №№ 5731168, 5807706, 5821333.
Общий метод получения гетеромультимера с использованием стратегии «продвижения в полость» включает экспрессию в одной или разных клетках-хозяевах полинуклеотида, кодирующего первый полипептид, который был изменен относительно исходного полинуклеотида с целью кодирования выступа, и второго полинуклеотида, кодирующего второй полипептид, который был изменен относительно исходного полинуклеотида с целью кодирования полости. Полипептиды экспрессируют в одной клетке-хозяине с последующим восстановлением гетеромультимера из культуры клеток-хозяев, или в отдельных клетках-хозяевах с последующим восстановлением и очисткой, что приводит к образованию гетеромультимера. В некоторых вариантах образуемый гетеромультимер представляет собой мультимерное антитело, например биспецифическое антитело.
В некоторых вариантах получение антител по настоящему изобретению объединяет стратегию выступов и полостей с конструкцией, включающей вариант по шарнирной области, с получением бесшарнирных биспецифических антител.
B. Векторы, клетки-хозяева и рекомбинантные методики
Настоящее изобретение также относится к выделенным полинуклеотидам, кодирующим антитела, приведенные в настоящем описании, к векторам и клеткам-хозяевам, включающим полинуклеотиды, а также к рекомбинантным методикам, используемым для получения антител.
Для рекомбинантной продукции антител полинуклеотид, кодирующий антитело, выделяют и встраивают в реплицирующийся вектор для дальнейшего клонирования (амплификации ДНК) или для экспрессии. ДНК, кодирующую антитело, выделяют и секвенируют в соответствии с известными процедурами, например, при использовании олигонуклеотидных зондов, способных к специфическому связыванию с генами, кодирующими антитело. Доступно множество векторов. Векторный компонент в основном включает, но ими не ограничивается, один или несколько следующих компонентов: сигнальную последовательность, ориджин репликации, один или несколько маркерных генов, энхансерный элемент, промотор и последовательность терминации транскрипции.
(i) Компонент сигнальной последовательности
Антитела по настоящему изобретению могут быть получены в рекомбинантной системе, не только в непосредственном виде, но также в виде антител слияния с гетерологичными антителами. В одном варианте гетерологичное антитело представляет собой сигнальную последовательность или другое антитело, содержащее специфический сайт расщепления на N-конце зрелого белка или антитела. Выбранная гетерологичная сигнальная последовательность представляет собой такую последовательность, которая распознается и процессируется (например, расщепляется сигнальной пептидазой) клеткой-хозяином. В случае прокариотических клеток-хозяев, которые не распознают и не процессируют нативную сигнальную последовательность антитела, сигнальную последовательность замещают прокариотической сигнальной последовательностью, выбранной, например, из группы, включающей щелочную фосфатазу, пенициллиназу, lpp или лидерные последовательности термостабильного энтеротоксина II. В случае дрожжевой секреции, нативная сигнальная последовательность может быть замещена, например, лидерной последовательностью для дрожжевой инвертазы, лидерной последовательностью α-фактора (включая лидерные последовательности α-фактора из Saccharomyces и Kluyveromyces) или лидерной последовательностью кислой фосфатазы, лидерной последовательностью глюкоамилазы C. albicans или сигнальной последовательностью, описанной в WO90/13646. В системе экспрессии в клетках млекопитающих доступны сигнальные последовательности млекопитающих, а также секреторные лидерные последовательности вирусов, например сигнальная последовательность gD из вируса простого герпеса (herpes simples). ДНК для такого участка-предшественника лигируют в рамке считывания с ДНК, кодирующей антитело.
В другом варианте продукция антител может быть осуществлена в цитоплазме клетки-хозяина, и в этом случае не требуется наличие секретируемых сигнальных последовательностей внутри каждого цистрона. Соответственно, легкие и тяжелые цепи иммуноглобулина экспрессируются, подвергаются образованию складчатой структуры и сборке с образованием функциональных иммуноглобулинов внутри цитоплазмы. Определенные штаммы-хозяева (например, штамм E. coli trxB) создают такие условия в цитоплазме, которые способствуют образованию дисульфидных связей, что позволяет создавать соответствующую складчатую структуру и осуществлять сборку экспрессированных белковых субъединиц (Proba and Plukthun, 1995, Gene, 149:203).
(ii) Компонент ориджина репликации
Как векторы экспрессии, так и векторы клонирования содержат последовательность нуклеиновой кислоты, которая позволяет вектору реплицироваться в одной или нескольких выбранных клетках-хозяевах. В основном в векторах клонирования данная последовательность представляет собой такую последовательность, которая позволяет вектору реплицироваться независимо от хромосомальной ДНК клетки-хозяина, и включает ориджины репликации или автономно реплицирующиеся последовательности. Такие последовательности хорошо известны для множества бактерий, дрожжей, вирусов. Ориджин репликации из плазмиды pBR322 приемлем для большинства Грам-отрицательных бактерий, ориджин репликации плазмиды 2μ приемлем для дрожжей, и различные вирусные ориджины репликации (SV40, полиомы, аденовируса, VSV или BPV) используются для векторов клонирования в клетках млекопитающих. В основном компонент ориджина репликации не является обязательным для векторов экспрессии в клетках млекопитающих (ориджин SV40 в типичном случае используется только в том случае, если он содержит ранний промотор).
(iii) Компонент, определяемый селекцией гена
Векторы экспрессии и клонирования могут содержать ген селекции, также называемый селектируемым маркером. Типичные гены селекции включают белки, которые (а) придают резистентность к антибиотикам или другим токсинам, например ампициллину, неомицину, метотрексату или тетрациклину, (b) комплементауксотрофную недостаточность или (с) восполняют важнейшие питательные компоненты, недоступные из комплексных средств, например ген, кодирующий D-аланин-рацемазу для Bacilli.
Один пример схемы селекции включает использование препарата, останавливающего рост клетки-хозяина. Те клетки, которые были успешно трансформированы гетерологичным геном, продуцируют белок, придающий лекарственную резистентность, и таким образом выживают в режиме селекции. Примеры такой доминантной селекции включают использование таких препаратов, как неомицин, микофеноловая кислота и гигромицин.
Другой пример подходящих селектируемых маркеров для клеток млекопитающих включает такие маркеры, которые позволяют идентифицировать клетки, компетентные по поглощению нуклеиновой кислоты для антитела, такие как гены DHFR, тимиидинкиназы, металлотионеина-I и -II, предпочтительно гены металлотионеина приматов, аденозиндезаминазы, орнитиндекарбоксилазы и т.п.
Например, клетки, трансформированные геном селекции DHFR, вначале идентифицируют путем культивирования всех трансформантов в культуральной среде, которые содержат метотрексат (Mtx), конкурентный антагонист DHFR. Подходящей клеткой-хозяином в том случае, когда используют DHFR дикого типа, является клеточная линия яичника китайского хомяка (CHO), дефицитная по DHFR активности.
Альтернативно, клетки-хозяева (в особенности клетки-хозяева дикого типа, которые содержат эндогенную DHFR), трансформированные или сотрансформированные последовательностями ДНК, кодирующими антитело, белок DHFR дикого типа и другой селектируемый маркер, такой как аминогликозид-3'-фосфотрансфераза (APH), могут быть отобраны путем выращивания клеток в среде, содержащей агент селекции, соответствующий селектируемому маркеру, такой как аминогликозидный антибиотик, например канамицин, неомицин или G418. См. патент США № 4965199.
Подходящим геном селекции, используемым в случае дрожжей, является ген trp1, присутствующий в дрожжевой плазмиде YRp7 (Stinchcomb et al., 1979, Nature, 282:39). Ген trp1 обеспечивает наличие маркера селекции для мутантного штамма дрожжей, не способных расти при наличии триптофана, например мутанта, зарегистрированного в ATCC № 44076 или PEP4-1 (Jones, 1977, Genetics, 85: 12). Наличие trp1-повреждения в геноме дрожжевых клеток-хозяев обеспечивает эффективное окружение для выявления трансформации при росте в отсутствие триптофана. Аналогично, Leu2-дефицитные дрожжевые штаммы (например, штаммы с номером доступа ATCC 20622 или 38626) компенсируются известными плазмидами, содержащими ген Leu2.
Кроме того, векторы, полученные из кольцевой плазмиды pKD1 размером 1,6 мкм, могут использоваться для трансформации дрожжей Kluyveromyces. Альтернативно, система экспрессии для целей крупномасштабного получения рекомбинантного химозина теленка описана применительно к K. lactis (см. Van der Berg, 1990, Bio/Technology, 8:135). Стабильные многокопийные векторы экспрессии для секреции зрелого рекомбинантного сывороточного альбумина человека промышленными штаммами Kluyveromyces также описаны в литературе (см. Fleer et al., 1991, Bio/Technology, 9: 968-975).
(iv) Промоторный компонент
Вектор экспрессии и клонирования обычно содержит промотор, который распознается организмом-хозяином и который оперативно соединен с нуклеиновой кислотой для антитела. Промоторы, подходящие для использования в прокариотических клетках-хозяевах, включают промотор phoA, β-лактамазную и лактозную промоторную системы, щелочную фосфатазу, промоторную систему триптофана (trp) и гибридные промоторы, такие как промотор tac. Однако могут использоваться и другие известные бактериальные промоторы. Промоторы, которые могут использоваться в бактериальных системах, также будут содержать последовательность Шайна-Далгарно (Shine-Dalgarno (S.D.)), оперативно связанную с ДНК, кодирующей антитело.
Промоторные последовательности известны также и для эукариотов. Фактически, все эукариотические гены содержат АТ-обогащенную область, расположенную примерно между 25 и 30 основаниями против направления считывания информации относительно сайта инициации транскрипции. Другая последовательность выявлена в положении 70-80 оснований против направления считывания информации относительно старта транскрипции многих генов на участке CNCAAT, где N может представлять собой любой нуклеотид. На 3'-конце большинства эукариотических генов находится последовательность AATAAA, которая может быть сигналом для добавления фрагмента поли-A к 3'-концу кодирующей последовательности. Все указанные последовательности соответствующим образом встраиваются в эукариотические векторы экспрессии.
Примеры подходящих промоторных последовательностей, которые могут использоваться с дрожжевыми клетками-хозяевами, включают промоторы для 3-фосфоглицераткиназы или других гликолитических ферментов, таких как энолаза, глицеральдегид-3-фосфатдегидрогеназа, гексокиназа, пируватдекарбоксилаза, фосфофруктокиназа, глюкозо-6-фосфатизомераза, 3-фосфоглицератмутаза, пируваткиназа, триозофосфатизомераза, фосфоглюкозоизомераза и глюкокиназа.
Другие дрожжевые промоторы, которые представляют собой индуцибельные промоторы, обладающие дополнительным плюсом наличия транскрипции, контролируемой ростовыми условиями, включают промоторные участки для алкогольдегидрогеназы 2, изоцитохрома С, кислой фосфатазы, ферментов деградации, связанных с азотным метаболизмом, металлотионеина, глицеральдегид-3-фосфатдегидрогеназы и ферментов, отвечающих за утилизацию мальтозы и галактозы. Подходящие векторы и промоторы, которые могут использоваться для экспрессии в дрожжах, также описаны в EP 73657. Энхансеры дрожжей также удобно использовать с дрожжевыми промоторами.
Транскрипция антитела на основе векторов в клетках-хозяевах млекопитающих контролируется, например, промоторами, полученными из геномов вирусов, таких как вирус полиомы, вирус осповакцины, аденовирус (такой как Аденовирус 2), вирус бычьей папилломы, вирус птичьей саркомы, цитомегаловирус, ретровирус, вирус гепатита В и наиболее предпочтительно обезьяньего вируса-40 (SV40), из гетерологичных промоторов млекопитающих, например актинового промотора или иммуноглобулинового промотора, из промоторов белков теплового шока, при условии, что такие промоторы совместимы с системами, функционирующими в клетках-хозяевах.
Ранние и поздние промоторы вируса SV40 обычно получают в виде рестрикционного фрагмента SV40, который также содержит ориджин репликации вируса SV40. Очень ранний промотор человеческого цитомегаловируса может быть получен в виде фрагмента рестрикции при использовании HindIII E. Система экспрессии ДНК в клетках-хозяевах млекопитающих с использованием вируса бычьей папилломы в качестве вектора описана в патенте США № 4419446. Модификация данной системы описана в патенте США № 4601978 (см. также Reyes et al., 1982, Nature, 297: 598-601) применительно к экспрессии кДНК β-интерферона человека в клетках мыши под контролем тимидинкиназного промотора из вируса простого герпеса (hepes simplex). Альтернативно, в качестве промотора может использоваться длинный концевой повтор из вируса саркомы Рауса.
(v) Компонент энхансерного элемента
Транскрипция ДНК, кодирующей антитело по настоящему изобретению, высшими эукариотами зачастую повышается при встраивании энхансерной последовательности в вектор. В настоящее время известно множество энхарсерных последовательностей из генов млекопитающих (глобина, эластазы, альбумина, α-фетопротеина и инсулина). Однако в типичном случае используют энхансер из вируса эукариотической клетки. Примеры их включают энхансер SV40, локализованный на поздней стороне ориджина репликации (100-270 н.п.), энхансер раннего промотора цитомегаловируса, энхансер полиомы на поздней стороне ориджина репликации и энхансеры аденовируса. Энхансерные элементы, влияющие на активацию эукариотических промоторов, описаны также Yaniv, 1982, Nature, 297: 17-18. Энхансер может также подвергаться сплайсингу при встраивании в вектор в положении 5' или 3' относительно последовательности, кодирующей антитело, и предпочтительно он расположен в 5'-положении относительно промотора.
(vi) Компонент терминации транскрипции
Векторы экспрессии, используемые в эукариотических клетках-хозяевах (дрожжи, грибы, насекомые, растения, животные, человек или ядросодержащие клетки из других многоклеточных организмов), также содержат последовательности, необходимые для терминации транскрипции и для стабилизации мРНК. Такие последовательности в основном доступны от 5'- и, иногда, от 3'-нетранслируемых участков эукариотической или вирусной ДНК или кДНК. Указанные участки содержат нуклеотидные сегменты, транскрибируемые в виде полиаденилированных фрагментов в нетранслируемой части мРНК, кодирующей антитело. Один полезный компонент терминации транскрипции представляет собой участок полиаденилирования бычьего гормона роста. См. WO 94/11026 и описанный в данном документе вектор экспрессии.
(vii) Модуляция силы трансляции
Иммуноглобулины по настоящему изобретению могут также экспрессироваться на основе системы экспрессии, в которой количественное соотношение экспрессируемых легкой и тяжелой цепей может модулироваться для максимизации выхода секретированных и соответствующим образом собранных антител полной длины. Такая модуляция осуществляется при одновременной модуляции сил трансляции для легкой и тяжелой цепей.
Одна методика модуляции силы трансляции описана Simmons et al., патент США № 5840523 и Simmons et al., 2002, J. Immunol. Methods, 263: 133-147. В данной методике используются варианты участка инициации трансляции (TIR) внутри цистрона. Для данного TIR может быть создана серия вариантов аминокислотных последовательностей или последовательностей нуклеиновой кислоты с диапазоном трансляционной силы, что обеспечивает подходящие механизмы корректировки данного фактора для достижения желательного уровня экспрессии конкретной цепи. Варианты TIR могут быть созданы с помощью традиционных методик мутагенеза, которые приводят к изменениям в кодонах и которые в свою очередь могут изменить аминокислотную последовательность, хотя предпочтительны молчащие изменения в нуклеотидной последовательности. Изменения в TIR могут включать, например, изменения в количестве или локализации последовательности Шайна-Далгамо (Shine-Dalgamo), вместе с изменениями в сигнальной последовательности. Одним предпочтительным методом создания мутантных сигнальных последовательностей является создание «кодонового банка» в начале кодирующей последовательности, который не меняет аминокислотную последовательность в сигнальной последовательности (то есть изменения являются молчащими). Этого можно достичь путем изменения третьего нуклеотидного положения в каждом кодоне; некоторые аминокислоты, такие как лейцин, серин и аргинин, характеризуются неоднозначностью первого и второго положений, что вносит дополнительную сложность в процесс создания банка. Такой метод мутагенеза подробно описан Yansura et al., 1992, METHODS: A Companion to Methods in Enzymol., 4: 151-158.
Предпочтительно создается множество векторов, включающих некоторый диапазон силы TIR для каждого цистрона. Такой ограниченный набор обеспечивает возможность сравнения уровней экспрессии каждой цепи, а также выхода продуктов полной длины при сочетании TIR разной силы. Сила TIR может быть определена путем количественной оценки уровня экспрессии репортерного гена, как описано подробно Simmons et al., патент США № 5840523 и Simmons et al., 2002, J. Immunol. Methods, 263: 133-147. В контексте настоящего изобретения сочетание силы трансляции для конкретной пары TIR в векторе описывают выражением (N - легкая цепь, М - тяжелая цепь), где N обозначает относительную силу TIR для легкой цепи и М обозначает относительную силу TIR для тяжелой цепи. Например, выражение (3 - легкая цепь, 7 - тяжелая цепь) означает, что вектор обеспечивает относительную силу TIR, равную примерно 3, для экспрессии легкой цепи и относительную силу TIR, равную примерно 7, для экспрессии тяжелой цепи. На основе результатов сравнения силы трансляции можно выбрать желательные отдельные TIR, которые могут быть далее объединены в конструкциях вектора экспрессии по настоящему изобретению.
(vii) Отбор и трансформация клеток-хозяев
Подходящие клетки-хозяева для клонирования или экспрессии ДНК в векторах по настоящему изобретению включают клетки прокариотов, дрожжей или высших эукариотов, описанные выше. Подходящие прокариоты для данной цели включают Archaebacteria и Eubacteria, такие как Грам-отрицателные или Грам-положительные организмы, например Enterobacteriaceae, такие как Escherichia, например E. coli, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, например Salmonella typhimurium, Serratia, например Serratia marcescans, и Shigella, а также Bacilli, такие как B. subtilis и B. licheniformis (например, B. licheniformis 41P, описанные в документе DD266710, опубликванном 12 апреля 1989 года), Pseudomonas, такие как P. aeruginosa, и Streptomyces. Одним предпочтительным для клонирования организмом-хозяином E. coli является E. coli 294 (ATCC 31446), хотя могут быть применимы и другие штаммы, такие как E. coli B, E. coli X1776 (ATCC 31537) и E. coli W3110 (ATCC 27325). Данные примеры следует рассматривать как иллюстративные и неограничивающие. Предпочтительно также для клетки-хозяина, чтобы она секретировала минимальное количество протеолитических ферментов, и желательно, чтобы в клеточную культуру были включены дополнительные ингибиторы протеазы. Прокариотические клетки-хозяева могут также содержат мутацию(и) в системах, функционирующих на пути тиоредоксина и/или глютатиона.
Кроме прокариотов, в качестве клеток-хозяев для клонирования или экспрессии антителокодирующих векторов также приемлемы эукариотические микроорганизмы, такие как нитчатые грибы или дрожжи. Saccharomyces cerevisiae, или обычные пекарские дрожжи, представляют собой чаще всего используемые микроорганизмы из низших эукариотических микроорганизмов-хозяев. Однако широко доступно и может быть применимо в контексте настоящего описания множество других родов и штаммов, таких как клетки-хозяева Schizosaccharomyces pombe; Kluyveromyces, в частности K. lactis, K. fragilis (ATCC 12424), K. bulgaricus (ATCC 16045), K. wickeramii (ATCC 24178), K. waltii (ATCC 56500), K. drosophilarum (ATCC 36906), K. thermotolerans и K. marxianus; yarrowia (EP 402226); Pichia pastoris (EP 183070); Candida; Trichoderma reesia (EP 244234); Neurospora crassa; Schwanniomyces, такие как Schwanniomyces occidentalis; и нитчатые грибы, такие как, например, клетки-хозяева Neurospora, Penicillium, Tolypocladium и Aspergillus, в частности A. nidulans и A. niger.
Подходящие клетки-хозяева, применимые для экспрессии гликозилированного антитела, получают из многоклеточных организмов. Примеры беспозвоночных включают клетки растений и клетки насекомых. Были также идентифицированы различные штаммы бакуловирусов и их варианты и соответствующие пермиссивные клетки-хозяева насекомых из таких организмов-хозяев, как Spodoptera frugiperda (гусеница), Aedes aegypti (комар), Aedes albopictus (комар), Drosophila melanogaster (плодовая мушка) и Bombyx mori. Для трансфекции доступно множество штаммов вирусов, например L-1 вариант Autographa californica NPV и Bm-5 штамм Bombyx mori NPV, и все такие вирусы могут использоваться в качестве вируса по настоящему изобретению, в особенности для трансфекции клеток Spodoptera frugiperda. Культуры растительных клеток хлопка, кукурузы, картофеля, соевых бобов, петуньи, томатов и табака также могут использоваться в качестве клеток-хозяев.
Широко используются в качестве хозяев также клетки позвоночных организмов, а размножение клеток позвоночных в культуре (культура ткани) стало обычной процедурой. Примеры клеточных линий млекопитающих, используемых в качестве хозяев, включают линию почки обезьяны CV1, трансформированную SV40 (COS-7, ARCC CRL 1651); клеточную линию почек эмбриона человека (клетки 293 или клетки 293, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen. Virol. 36: 59 (1977)); клетки почки детеныша хомяка (BHK, ATCC CCL 10); клетки яичника китайского хомяка /-DHFR (CHO, Urlaub et al., 1980, Proc. Natl. Acad. Sci. USA 77:4216); клетки Сертоли мыши (TM4, Mather 1980, Biol. Reprod. 23:243-251); клетки почки обезьяны (CV1 ATCC CCL 70); клетки почки африканской зеленой мартышки (VERO-76, ATCC CRL-1587); клетки цервикальной карциномы человека (HELA, ATCC CCL 2); клетки почки собаки (MDCK, ATCC CCL 34); клетки печени крысы буффало (BRL 3A, ATCC CRL 1442); клетки легкого человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); клетки опухоли молочной железы мыши (MMT 060562, ATCC CCL 51); клетки TRI (Mather et al., 1982, Annals N.Y. Acad. Sci. 383: 44-68); клетки MRC-5; клетки FS4; клетки миеломы мыши, такие как NSO (например, RCB0213, 1992, Bio/Technology 10:169) и клетки SP2/0 (например, клетки SP2/0-Ag14, ATCC CRL 1581); клетки миеломы крысы, такие как клетки YB2/0 (например, клетки YB2/3HL.P2.G11.16Ag.20, ATCC CRL 1662); и клеточную линию гепатомы человека (Hep G2). Клетки CHO являются предпочтительной клеточной линией для практического осуществления настоящего изобретения, а клетки CHO-K1, DUK-B11, CHO-DP12, CHO-DG44 (Somatic Cell and Molecular Genetics 12:555 (1986)) и Lec13 представляют собой репрезентативные примеры линии клеток-хозяев. В случае клеток-хозяев CHO-K1, DUK-B11, DG44 или CHO-DP12, данные клетки могут быть изменены таким образом, чтобы сделать их дефицитными по способности осуществлять фукозилирование экспрессированных белков.
Настоящее изобретение также применимо к гибридомным клеткам. Термин «гибридома» относится к гибридной клеточной линии, получаемой при слиянии иммортализованной клеточной линии иммунологического происхождения и антитело-продуцирующей клетки. Термин охватывает потомство гетерогибридных миеломных белков слияния, которое является результатом слияния клеток человека и клеточной линии миеломы мыши с последующим их слиянием с плазматической клеткой, известное как триомная клеточная линия. Кроме того, термин включают любую иммортализованную гибридную клеточную линию, которая продуцирует антитела, такие как, например, квадромы (см., например, Milstein et al., 1983, Nature, 537: 3053). Гибридные клеточные линии могут быть получены на основе любого вида, включая человека и мышь.
В наиболее предпочтительном варианте осуществления настоящего изобретения клетка млекопитающего представляет собой клетку млекопитающего, отличную от гибридомы, которая была трансформирована экзогенной выделенной нуклеиновой кислотой, кодирующей интересующее антитело. Термин «экзогенная нуклеиновая кислота» или «гетерологичная нуклеиновая кислота» обозначает последовательность нуклеиновой кислоты, которая является чужеродной для клетки или является гомологичной для клетки, но в таком положении в составе нуклеиновой кислоты клетки-хозяина, в котором нуклеиновая кислота обычно не встречается.
(viii) Культивирование клеток-хозяев
Клетки-хозяева трансформируют указанными выше векторами экспрессии или векторами клонирования с целью получения антитела и культивируют в обычных питательных средах, модифицированных соответствующим образом для индукции промоторов, селекции трансформантов или амплификации генов, кодирующих желательные последовательности.
Клетки-хозяева, используемые для продукции антитела по настоящему изобретению, могут культивироваться в различных средах. Для культивирования клеток-хозяев приемлемы коммерчески доступные среды, такие как среда Хэма F10 (Ham's F10) (Sigma), минимальная эссенциальная среда ((MEM) (Sigma), RPMI-1640 (Sigma)) и среда Игла в модификации Дульбекко (DMEM) (Sigma). Кроме того, любая среда, описанная Ham et al., 1979, Meth. Enz. 58:44, Barnes et al., 1980, Anal. Biochem. 102: 255, а также в патентах США №№ 4767704, 4657866, 4927762, 4560655 или 5122469; в документах WO 90/03430; WO 87/00195 или в патенте США Re. 30985 может использоваться в качестве культуральной среды для выращивания клеток-хозяев. В любую из указанных сред могут быть внесены добавки, при необходимости, такие как гормоны и/или другие ростовые факторы (такие как инсулин, трансферрин или эпидермальный фактор роста), соли (такие как хлорид натрия, соли кальция, магния и фосфат), буферы (такие как HEPES), нуклеотиды (такие как аденозин и тимидин), антибиотики (такие как терапевтическое средство гентамицин (GENTAMYCINTM)), минорные элементы (определяемые как неорганические соединения, обычно присутствующие в конечных концентрациях в микромолярном диапазоне) и глюкоза или другой эквивалентный источник энергии. Любые другие необходимые добавки могут также включаться в соответствующих концентрациях, которые известны специалистам в данной области. Условия культивирования, такие как температура, pH и т.п., соответствуют значениям, обычно используемым в случае данной клетки-хозяина, выбранной для экспрессии, которые известны специалистам в данной области.
Все культуральные среды в типичном случае обеспечивают по меньшей мере один компонент, выбранный из одной или нескольких приведенных ниже категорий:
1) источник энергии, обычно в форме углевода, такого как глюкоза;
2) незаменимые аминокислоты и обычно основной набор из двадцати аминокислот плюс цистин;
3) витамины и/или другие органические соединения, необходимые в низких концентрациях;
4) свободные жирные кислоты; и
5) минорные элементы, где минорные элементы определяются как неорганические соединения или природные элементы, которые в типичном случае необходимы в очень низких концентрациях, обычно в микромолярном диапазоне.
Культуральная среда предпочтительно не содержит сыворотку, то есть содержит меньше примерно 5%, предпочтительно меньше примерно 1% и более предпочтительно меньше примерно 0-0,1% сыворотки и других белков животного происхождения. Однако они могут использоваться при желании. В предпочтительном варианте осуществления настоящего изобретения культуральная среда для выращивания клеток включает избыток аминокислот. Аминокислоты, которые вносятся в избытке, могут быть выбраны, например, из Asn, Asp, Gly, Ile, Leu, Lys, Met, Ser, Thr, Trp, Tyr и Val. Предпочтительно вносят в избытке Asn, Asp, Lys, Met, Ser и Trp. В частности, например, могут использоваться аминокислоты, витамины, минорные элементы и другие компоненты среды в однократном или двукратном диапазоне, указанные в европейском патенте EP 307247 или в патенте США № 6180401. Указанные два документа включены в настоящее описание в качестве ссылки.
В случае культуры клеток млекопитающих, экспрессирующих желательный белок и способных добавлять желательные углеводы в конкретных позициях, могут использоваться различные культуральные условия, в которых уделяется конкретное внимание культивируемой клетке-хозяину. Подходящие условия для культивирования клеток млекопитающих известны специалистам в данной области (W. Louis Clevelend et al., 1983, J. Immunol. Methods 56: 221-234) или могут быть определены на основе доступной информации (см., например, Animal Cell Culture: A Practical Approach 2nd Ed., Rickwood, D. and Hames, B. D., eds. Oxford University Press, New York (1992)) и которые варьируют в зависимости от выбранной для работы конкретной клетки-хозяина.
(ix) Очистка антитела
В соответствии с рекомбинантными методиками антитело может быть получено внутри клетки, в периплазматическом пространстве или может быть непосредственно секретировано в среду. Если антитело получают внутриклеточно, то на первой стадии удаляют, например, центрифугированием или ультрафильтрацией остатки частиц клеток-хозяев или лизированных фрагментов. Carter et al., 1992, Bio/Technology 10: 163-167, описывает процедуру выделения антител, которые были секретированы в периплазматическое пространство E. coli. Вкратце, процедура состоит в том, что клеточную пасту оттаивают в присутствии ацетата натрия (pH 3,5), ЭДТА и фенилметилсульфонилфторида (PMSF) в течение примерно 30 минут. Клеточные осколки удаляют центрифугированием. В том случае, когда антитело секретируется в среду, супернатанты, отобранные из таких систем экспрессии, в основном вначале концентрируют с использованием коммерчески доступного фильтра для концентрирования белка, например фильтра Амикон (Amicon) или устройства для ультрафильтрации Миллипор (Millipore Pellicon). На любой последующей стадии очистки может использоваться ингибитор протеазы, такой как PMSF, для ингибирования протеолиза, а также антибиотики - для предотвращения роста нежелательных контаминирующих организмов.
Композиции антител, получаемые из клеток, могут быть очищены с использованием, например, хроматографии на гидроксилапатите, гель-электрофореза, диализа, аффинной хроматографии, при этом предпочтительной методикой очистки является аффинная хроматография. Пригодность белка А в качестве лиганда для аффинной хроматографии определяется видом и изотипом Fc-участка иммуноглобулина, имеющегося в антителе. Белок А может использоваться для очистки антител, которые основаны на тяжелых цепях γ1, γ2 или γ4 антител человека (Lindmark et al., 1983 J. Immunol. Meth. 62: 1-13). Белок G рекомендован для очистки всех изотипов мыши и γ3 человека (Guss et al., 1986, EMBO J. 5: 1567-1575). Матрица, к которой обычно присоединяют аффинный лиганд, чаще всего представляет собой агарозу, но приемлемы также и другие матрицы. Механически стабильные матрицы, такие как стекло с контролируемым размером пор или поли(стиролдивинил)бензол, позволяют достичь больших скоростей и сократить время процессинга, в сравнении с вариантом использования агарозы. В том случае, когда антитело включает домен CH3, для очистки может использоваться смола Bakerbond ABXTM (J. T. Baker, Phillipsburg, NJ). Приемлемы также другие методики для очистки белка, такие как фракционирование на ионообменной колонке, осаждение этанолом, обращенно-фазовая ВЭЖХ, хроматография на силикагеле, хроматография на гепаринсефарозе (SEPHAROSETM), хроматография на катионообменной смоле (такая как хроматография на колонке с полиаспарагиновой кислотой), хроматофокусирование, электрофорез в SDS-PAGE и осаждение сульфатом аммония, выбор которых зависит от выделяемого антитела.
В одном варианте гликопротеин может быть очищен с использованием метода адсорбции на лектиновом субстрате (например, с использованием аффинной хроматографии с лектином) для удаления из препарата фукозосодержащего гликопротеина и тем самым для обогащения его бесфукозным гликопротеином.
(x) Тесты для определения активности антител
Иммуноглобулины по настоящему изобретению могут быть охарактеризованы по их физическим/химическим свойствам и биологическим функциям с использованием различных известных в данной области методик тестирования. В одном аспекте изобретения важно сравнить селективность антитела по настоящему изобретению по связыванию с иммуногеном в сравнении с другими связываемыми мишенями. В частности, антитело, с которым селективно связывается FcγRIIB, предпочтительно не будет связываться или будет демонстрировать слабую аффинность по связыванию с другими FcγR, в частности FcγRIIA.
В некоторых вариантах осуществления настоящего изобретения иммуноглобулины, получаемые в соответствии с описанными в нем методиками, анализируют на биологическую активность. В некоторых вариантах иммуноглобулины по настоящему изобретению тестируют на их антигенсвязывающую активность. Тесты на связывание антигена, которые известны в данной области и которые могут использоваться в контексте настоящего изобретения, включают, но ими не ограничиваются, любой непосредственный или конкурентный тест на связывание с использованием таких методик, как вестерн-блоттинг, радиоиммуноанализ, ELISA (иммуноферментный твердофазный анализ), иммунологический анализ по методу «сэндвича», анализ в рамках процедуры иммунопреципитации, флуоресцентный иммунологический анализ и иммунологический анализ с использованием белка А. Ниже в разделе «Примеры» приведены репрезентативные тесты на связывание антигена.
Очищенные иммуноглобулины могут быть далее охарактеризованы с использованием серии тестов, включающих, но ими не ограничивающихся, N-концевое секвенирование, аминокислотный анализ, высокоэффективную жидкостную хроматографию в неденатурирующих условиях (ВЭЖХ), масс-спектрометрию, ионообменную хроматографию и расщепление папаином. Методы количественного определения белка известны в данной области. Например, образцы экспрессированных белков сравнивают количественно по интенсивности окрашивания на содержащем кумасси SDS-PAGE. Альтернативно, конкретная интересующая полоса(ы) (например, полоса полной длины) может быть выявлена, например, при проведении вестерн-блоттинга и/или анализа с использованием AME5-RP.
С. Фармацевтические композиции
Терапевтические композиции антитела могут быть получены путем смешивания антитела, имеющего желательную степень чистоты, с необязательными физиологически приемлемыми носителями, эксципиентами или стабилизаторами (Remingron's Pharmaceutical Science 16th edition, Osol, A. Ed. (1980)) с получением лиофилизированных композиций или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципиентов в используемых дозировках и концентрациях и включают буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включающие аскорбиновую кислоту и метионин; консерванты (такие как октадецилдиметилбензилхлорид аммония; хлорид гексаметония; хлорид бензалкония; хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил или пропилпарабен; катехол; резорцинол; циклогексанол; 3-пентанол; и м-крезол); низкомолекулярные (например, содержащие менее примерно 10 остатков) антитела; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глютамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включающие глюкозу, маннозу или декстрины; хелатирующие агенты, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексы металлов (например, Zn-белковые комплексы); и/или неионные сурфактанты, такие как TweenTM (TWEENTM), плуроники (PLURONICSTM) или полиэтиленгликоль (ПЭГ).
Описываемая в настоящем изобретении композиция может также содержать более одного активного соединения, необходимого для конкретного подлежащего лечению состояния, предпочтительно с комплементарными активностями, которые не оказывают друг на друга неблагоприятного воздействия. Например, композиция может также включать другое антитело или химиотерапевтический агент. Такие молекулы присутствуют в соответствующем сочетании в количествах, эффективных для целей лечения.
Активные ингредиенты могут быть также захвачены получаемыми микрокапсулами и встроены в их состав, например, путем использования методик коацервации или путем межпоточной полимеризации, например, в состав гидрометилцеллюлозных или желатиновых микрокапсул или поли(метилметакрилатных)микрокапсул, соответственно, в состав коллоидных систем доставки лекарственных средств (например, в состав липосом, альбуминовых микросфер, микроэмульсий, наночастиц и нанокапсул) или в состав макроэмульсий. Такие методики описаны Remingron's Pharmaceutical Science 16th edition, Osol, A. Ed. (1980).
Подлежащие введению in vivo композиции должны быть стерильными. Стерилизация может быть осуществлена путем фильтрования через стерильные мембраны для фильтрации.
Могут быть также получены препараты с пролонгированным высвобождением. Подходящие примеры препаратов с пролонгированным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащих антитело, где указанные матрицы имеют определенную форму, например, пленок или микрокапсул. Примеры матриц с пролонгированным высвобождением включают сложные полиэфиры, гидрогели (например, поли-(2-гидроксиэтилметакрилат) или поли(виниловый спирт)), полилактиды (патент США № 3773919), сополимеры L-глютаминовой кислоты и γ-этил-L-глютамата, неразлагаемый этиленвинилацетат, разлагаемые сополимеры молочной кислоты-гликолевой кислоты, такие как LUPRON DEPOTTM (инъецируемые микросферы, состоящие из сополимера молочной кислоты-гликолевой кислоты и ацетата леупролида) и поли-D-(-)-3-гидроксимасляную кислоту. Поскольку полимеры, такие как этиленвинилацетат и сополимер молочной кислоты-гликолевой кислоты, позволяют осуществлять высвобождение молекул в течение периода времени, превышающего 100 дней, некоторые гидрогели высвобождают белки в течение более коротких периодов времени. В том случае, когда инкапсулированные антитела остаются в организме в течение длительного периода времени, они могут денатурировать или агрегировать в результате воздействия влаги при 37°С, что приводит к потере биологической активности и возможным изменениям в иммуногенности. В этой связи, должны быть разработаны рациональные стратегии стабилизации, в зависимости от конкретного вовлеченного механизма. Например, если механизм агрегации имеет отношение к образованию межмолекулярных связей S-S через тиодисульфидный взаимообмен, то стабилизация может быть достигнута путем модификации сульфгидрильных остатков, лиофилизации из кислых растворов, контролирования содержания влаги, при использовании соответствующих добавок и путем разработки специфических полимерных матричных композиций.
D. Нетерапевтические направления использования антитела
Антитело по настоящему изобретению может использоваться в качестве агента для аффинной очистки. В рамках данного способа антитело иммобилизуют на твердой фазе, такой как смола Сефадекс (SephadexTM) или фильтровальная бумага, с использованием известных в данной области методик. Иммобилизованное антитело приводят в контакт с образцом, содержащим подлежащий очистке антиген, и далее подложку промывают подходящим растворителем, который удаляет по существу весь материал в образце, за исключением антигена, который подлежит очистке и который связывается иммобилизованным антителом. В конце процедуры подложку промывают другим подходящим растворителем, таким как глициновый буфер, pH 5,0, который будет высвобождать антиген из антитела.
Антитело может также использоваться в диагностических тестах, например, для выявления экспрессии интересующего антигена в конкретных клетках, тканях или сыворотке. Для целей диагностического применения антитело в типичном случае метят выявляемым фрагментом. Доступно множество различных меток, которые могут быть сгруппированы в основном по следующим категориям:
(а) Радиоактивные изотопы, такие как 35S, 14C, 125I, 3H и 131I. Антитело может быть помечено радиоактивным изотопом с использованием методик, описанных, например, в соответствующем руководстве (Current Protocols on Immunology, Volumes 1 and 2, Coligen et al., Ed. Wiley-Interscience, New York, New York, Pub. (1991)), и радиоактивность может быть далее определена с использованием сцинтилляционного счетчика.
(b) Флуоресцентные метки, такие как хелаты щелочно-земельных металлов (хелаты европия) или флуоресцеин и его производные, родамин и его производные, дансил, лиссамин, фикоэритрин и техасский красный также доступны. Флуоресцентные метки могут быть конъюгированы с антителом с использованием методик, описанных в соответствующем руководстве (см., например, Current Protocols on Immunology, выше). Количественное содержание флуоресцентной метки может быть определено с помощью флуориметра.
(с) Различные метки, содержащие энзиматические субстраты, также доступны, и в патенте США № 4275149 приводится обзор некоторых таких меток. В основном фермент катализирует химическое изменение в хромогенном субстрате, которое может быть определено с помощью разных методик. Например, фермент может катализировать изменение окраски субстрата, что можно количественно оценить спектрофотометрически. Альтернативно, фермент может изменять флуоресценцию или хемилюминесценцию субстрата. Методики количественной оценки изменений флуоресценции были описаны выше. Хемилюминесцентный субстрат становится электронно возбужденным в ходе химической реакции и затем может испускать свет, показатели которого могут быть количественно измерены (с использованием, например, хемилюминометра), или может отдавать энергию флуоресцентному акцептору. Примеры энзиматических меток включают люциферазы (например, люциферазу светлячков и бактериальную люциферазу; патент США № 4737456), люциферин, 2,3-дигидрофталазиндионы, малатдегидрогеназу, уреазу, пероксидазу, такую как пероксидаза хрена (HRPO), щелочную фосфатазу, β-галактозидазу, глюкоамилазу, лизоцим, оксидазы сахаридов (например, оксидазу глюкозы, оксидазу галактозы и глюкоза-6-фосфатдегидрогеназу), гетероциклические оксидазы (такие как уриказа и ксантиноксидаза), лактопероксидазу, микропероксидазу и т.п. Методики конъюгирования ферментов с антителами описаны в ряде работ (O'Sullivan et al., Methods for the Preperation of Enzyme-Antibody Conjugates for use in Enzyme Immunoassay, in Methods in Enzym. (ed. J. Langone and H. Van Vunakis), Academic Press, New York, 73: 147-166 (1981)).
Примеры фермент-субстратных сочетаний включают, например:
1) пероксидазу хрена (HRPO), которая использует перекись водорода для окисления предшественника красителя (например, ортофенилендиамин (OPD) или гидрохлорид 3,3',5,5'-тетраметилбензидина (TMB));
2) щелочную фосфатазу (AP) с пара-нитрофенилфосфатом в качестве хромогенного субстрата; и
3) β-D-галактозидазу (β-D-Gal) с хромогенным субстратом (например, п-нитрофенил-β-D-галактозидазой) или флуорогенным субстратом, 4-метилумбелиферил-β-D-галактозидазой.
Для специалиста в данной области известно и доступно множество других фермент-субстратных сочетаний. Один из таких обзоров приведен, например, в патентах США №№ 4275149 и 4318980.
Иногда метку подвергают опосредованной конъюгации с антителом. Специалистам в данной области известно множество методик, используемых для достижения этой цели. Например, антитело может быть конъюгировано биотином, и любая из указанных выше меток, входящих в три основные категории, может быть конъюгирована с авидином, или наоборот. Биотин селективно связывается с авидином и, следовательно, метка может быть конъюгирована с антителом таким опосредованным образом. Альтернативно, для достижения опосредованной конъюгации метки с антителом антитело конъюгируют с небольшим гаптеном (например, дигоксином) и одну из указанных выше меток любого типа конъюгируют с антигаптеновым антителом (например, антидигоксиновым антителом). Таким образом, может быть достигнута опосредованная конъюгация метки с антителом.
В другом варианте осуществления настоящего изобретения нет необходимости метить антитело, а присутствие антитела может быть выявлено с использованием меченого антитела, которое связывается с антителом.
Антитело по настоящему изобретению может использоваться в любом известном методе тестирования, таком как конкурентные методы тестирования связывание, опосредованные и прямые тесты по методу «сэндвич» и тесты на основе иммунопреципитации (Zola, Monoclonal Antibodies: A Manual of Techniques, pp. 147-158 (CRC Press, Inc. 1987)).
Антитело может также использоваться для проведения диагностических тестов in vivo. В основном антитело метят радионуклидом (таким как 111In, 99Tc, 14C, 131I, 125I, 3H, 32P или 35S), так что антиген или клетка, экспрессирующая его, могут быть выявлены с использованием иммуносцинтиографии.
Е. Направления использования антитела in vivo
(i) Сниженная ингибирующая активность FcγRIIB (CD32B): помехи связыванию с Fc антитела
В другом варианте анти-FcγRIIB антитело вводят вместе с терапевтическим агентом для усиления функции терапевтического агента. Например, анти-FcγRIIB вводят в организм млекопитающего для блокирования связывания IgG c FcγRIIB, препятствуя тем самым FcγRIIB-опосредованному ингибированию иммунного ответа. Это приводит к повышению цитотоксичности IgG терапевтического антитела. Например, в том случае, когда терапевтическое антитело специфично для опухолевого антигена, совместное введение анти-FcγRIIB по настоящему изобретению с антителом против опухолевого антигена повышает цитотоксичность антитела против опухолевого антигена.
Терапевтические антитела, многие из которых описаны выше, были разработаны и разрешены для лечения множества заболеваний, включая рак. Например, RITUXAN® (ритуксимаб (IDEC Pharm/Genentech, Inc.)) используют для лечения B-клеточных лимфом, AVASTINTM (бевацизумаб (Genentech, Inc.)) используют для лечения метастазирующего колоректального рака и HERCEPTIN® (трастумаб), представляющий собой гуманизированное анти-HER-2 моноклональное антитело, используют для лечения метастазирующего рака молочной железы. Несмотря на то, что механизм лечения рака всеми моноклональными антителами, разработанными для целей такого лечения, не совсем понятен, по меньшей мере в ряде случаев, часть достигаемой при антительной терапии эффективности можно объяснить рекрутментом иммунной эффекторной функции (Houghton et al., 2000, Nature Medicine, 6: 373-374; Clynes et al., 2000, Nature Medicine, 6: 433-446). XOLAIR® (омализумаб (Genentech, Inc.)) представляет собой анти-IgE антитело, используемое для лечения аллергий.
FcγRIIB экспрессируется на клетках лимфоидной и миелоидной линий, но не на природных клетках-киллерах и представляет собой ингибирующий рецептор. В случае активации, FcγRIIB может, например, ингибировать сигнальную функцию FcγRIII, который, в ином случае активировал бы макрофаги, природные клетки-киллеры и тучные клетки. Ингибирование FcγRIIB (например, блокирование связывания Fc с FcγRIIB) ослабляет его ингибирующий эффект на иммунную эффекторную функцию, что способствует терапии MAb. Ravetch, J. (WO 01/79299) описывает способ повышения цитотоксичности противоопухолевого антитела за счет снижения аффинности Fc-участка для FcγRIIB и, таким образом, ограничения SHIP-опосредованного ингибирования клеточной активации.
В одном варианте антитело, которое селективно связывается с FcγRIIB, вводят вместе с противоопухолевым антителом млекопитающему при необходимости такого лечения. Селективность в отношении FcγRIIB является желательным компонентом с тем, чтобы активация иммунного эффекторного ответа другими FcγR, включая FcγRIIA, не затрагивалась. При неудаче достичь перекрестной реакции с FcγRIIA ингибирующая функция FcγRIIB будет блокироваться более эффективно, что дополнительно повышает эффект совместно вводимого терапевтического агента.
В одном варианте анти-FcγRIIB антитело по настоящему изобретению вводят в организм млекопитающего для блокирования связывания IgG антител, что, в свою очередь, блокирует ингибирующий эффект FcγRIIB и, например, повышает В-клеточную пролиферацию.
(ii) Повышение ингибирующей активности FCγRIIB: совместная агрегация с активирующим рецептором
In vivo FcγRIIB может быть подвергнут совместной агрегации с большим числом активирующих рецепторов, включающих, в качестве неограничивающих примеров, рецептор В-клеточного антигена (BCR), высокоаффинный рецептор для IgE (IgER или FcεRI), FcγRIIA и рецепторный с-набор (с-kit) рецепторов (FcγRIII). Активирующие рецепторы включают, в качестве неограничивающих примеров, трансмембранные белки с активирующей активностью для иммунного эффекторного ответа, а также мотив активации ITAM. FcγRIIB активируется под действием совместной агрегации FcγRIIB с активирующим рецептором и ослабляет сигналы, доставляемые через активирующие рецепторы. К настоящему времени не было показано, что FcγRIIB фосфорилируется путем самоагрегации или гомодимеризации. Рецептор FcγRIIB был подвергнут в экспериментальном режиме гетеродимеризации или совместной агрегации (или совместному лигированию) с другими рецепторами, экспрессирующими фосфорилированный ITAM (мотив активации), и путем опосредованной ассоциации с белковыми тирозинкиназами (PTK) FcγRIIB ITIM может быть фосфорилирован. Фосфорилированный FcγRIIB ITIM мобилизует домен SH2, содержащий фосфатазу SHIP (инозитполифосфат-5'-фосфатазу), и ингибирует ITAM-инициированную мобилизацию кальция и клеточную пролиферацию (Daeron et al., 1995, Immunity 3, 635; Malbec et al., 1998, J. Immunol. 169, 1647; Ono et al., 1996, Nature, 383, 263), Итоговый эффект представляет собой блокирование кальциевого потока с созданием ограничений длительному сигнальному пути через кальциевый поток, что препятствует прохождению кальцийзависимых процессов, таких как дегрануляция, фагоцитоз, ADCC и высвобождение цитокинов (Ravetch et al., 2000, Science, 290: 84-89), хотя некоторые FcγRIIB-опосредованные механизмы блокирования сигнальной функции могут также не зависеть от кальция. Остановка пролиферации В-клеток также зависит от механизма ITIM.
Активация FcγRIIB-ингибирующей активности может сопровождаться опосредованным сшиванием моноклональных антител, специфичных для FcγRIIB и связанного активирующего рецептора. Опосредованные сшивающие реагенты включают авидин для биотинилированных моноклональных антител, поликлональные антитела, специфичные для Fc-части моноклонального IgG мыши, и поливалентный антиген, который образует иммунный комплекс, связывающий как ингибирующие, так и активирующие рецепторы. В большинстве описанных экспериментальных моделей используют В-клеток мыши или тучные клетки и моноклональное антитело (G4.2 крысы), которые вступают в перекрестную реакцию с рецепторами FcγRII и FcγRIII мыши.
В рамках настоящего изобретения гетеробифункциональное антитело, включающее моноклональный FcγRIIB Fab против человеческого компонента и моноклональный Fab, специфичный для активирующего рецептора, получают в соответствии с методами химического или генетического конструирования, известными в данной области.
Терапевтический потенциал такого бифункционального антитела включает ослабление сигналов, вовлекаемых в воспаление и аллергию. При активации и под действием IgE и аллергена (через FcεR) тучные клетки и базофилы секретируют воспалительные медиаторы и цитокины, которые активируются на сосудистых и мышечных клетках и мобилизуют воспалительные клетки. Воспалительные клетки, в свою очередь, секретируют воспалительные медиаторы и осуществляют рекрутмент воспалительных клеток в длительном процессе, который приводит к длительному воспалению. В этой связи, средства контроля IgE-индуцированной активации тучных клеток обеспечивают новый терапевтический подход для лечения аллергических заболеваний путем прерывания инициации воспалительного ответа. Как было указано выше, бифункциональное антитело также включает антитело или его фрагмент, который селективно связывается с FcγRIIB, и включает антитело или его фрагмент, который связывается, например, с FcεRI или FcεRI, связанным с IgE, ослабляя IgE-опосредованую активацию через ингибирование активности FcγRIIB.
Дополнительные примеры бифункциональных антител (например, биспецифических антител) включают сочетания антитела или его фрагмента, который селективно связываются с FcγRIIB, и второго антитела или его фрагмента, который связывается с активирующим рецептором, вовлекаемым в патологию астмы (моноклональный Fab FcγRIIB против молекулы человека и моноклональный Fab, специфичный для FcεRI, связанный с IgE или CD23), ревматоидного артрита и системной красной волчанки (моноклональный Fab FcγRIIB против молекулы человека и моноклональный Fab, специфичный для FcγRI), псориаза (моноклональный Fab FcγRIIB против молекулы человека и моноклональный Fab, специфичный для CD11a), иммунноопосредованной тромбоцитопении, ревматоидного артрита и системной красной волчанки (моноклональный Fab FcγRIIB против молекулы человека и моноклональный Fab, специфичный для FcγRIII (CD16) или CD4), болезни Крона и язвенного колита (моноклональный Fab FcγRIIB против молекулы человека и моноклональный Fab, специфичный для интегрина альфа4бета7, субъединицы интегрина бета7 или субъединицы интегрина альфа4, или связываемая часть указанных моноклональных антител) и другие аутоиммунные расстройства, в которые активно вовлекаются такие клетки, как тучные клетки, базофилы, В-клетки, моноциты, природные клетки-киллеры, нейтрофилы и дендритные клетки. Различные аутоиммунные заболевания были описаны выше в разделе «Определения» данного описания. Антитело может также использоваться для лечения аутоиммунных заболеваний, в случае которых имеется выраженная связанность с комплексным иммунным компонентом.
В некоторых вариантах антитело по настоящему изобретению используют для активации ингибирующих FcγRIIB рецепторов у млекопитающего, подлежащего лечению антителом, с целью ингибирования провоспалительных сигналов и/или В-клеточной активации, опосредованных активирующими рецепторами. В этой связи антитело используют для лечения воспалительных расстройств и/или аутоиммунных заболеваний, таких как указаны выше. Активация ингибирующей функции FcγRIIB осуществляется биспецифическим антителом по настоящему изобретению, которое непосредственно сшивает FcγRIIB и активирующий рецептор, и/или антителом, которое опосредованно сшивает FcγRIIB и активирующий рецептор.
В некоторых вариантах антитело по настоящему изобретению ингибирует дегрануляцию, связанную с активацией. Ингибирование дегрануляции, связанной с активацией, связано с супрессией высвобождения гистамина, которая может быть измерена. В некоторых вариантах антитело по настоящему изобретению ингибирует высвобождение гистамина по меньшей мере до 70% относительно общего высвобождения гистамина. В других вариантах ингибирование высвобождения гистамина достигает по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, включая промежуточные целые значения в диапазоне от 70 до 100%, где 100% снижение высвобождения гистамина принимается как эквивалентное фоновому высвобождению гистамина.
Для предотвращения или лечения заболевания соответствующая для применения дозировка антитела будет зависеть от типа заболевания, подлежащего лечению, тяжести и течения заболевания, от того, с какой целью, профилактической или терапевтической, вводят данное антитело, от предшествующей терапии, от истории болезни пациента и его отзывчивости на антитело, и определяется лечащим врачом. Антитело вводят пациенту соответствующим образом однократно или в виде курсового лечения.
В зависимости от типа и тяжести заболевания дозировка для пациента будет составлять примерно 1 мкг/кг - 15 мг/кг (например, 0,1-20 мг/кг) антитела, вводимого, например, однократно или в виде нескольких отдельных введений или путем непрерывной инфузии. Типичная дневная дозировка может варьировать от примерно 1 мкг/кг до примерно 100 мг/кг или более, в зависимости от указанных выше факторов. Для повторных введений в течение нескольких дней или более длительного периода, в зависимости от состояния, лечение может быть длительным, до достижения желательной супрессии имеющихся симптомов заболевания. Однако другие режимы дозирования также могут использоваться. Прогресс данной терапии легко оценивается традиционными методиками и тестами.
Антительная композиция может быть изготовлена, представлена в дозированной форме и введена в режиме, соответствующем медицинской практике. Факторы, которые должны приниматься при этом в расчет, включают природу конкретного заболевания, подлежащего лечению, конкретное млекопитающее, подлежащее лечению, состояние здоровья данного пациента, причина расстройства, место доставки агента, способ введения, режим введения и другие факторы, известные специалистам в данной области. «Терапевтически эффективное количество» подлежащего введению антитела будет определяться с учетом указанных выше соображений и представляет собой минимальное количество, необходимое для предотвращения, ослабления или лечения заболевания или расстройства. Антитело необязательно может быть включено в состав композиции с использованием одного или нескольких агентов, обычно используемых для профилактики или лечения рассматриваемого расстройства. Эффективное количество таких других агентов зависит от количества антитела, имеющегося в композиции, от типа расстройства или вида лечения, а также от других обсуждавшихся выше факторов. В основном они используются в одних и тех же дозах и режимах дозирования, которые были указанных выше, и составляют примерно от 1 до 99% от указанных выше используемых дозировок.
Композиции терапевтических антител вводят в контейнер, имеющий стерильный порт доступа, например в пакет с раствором для внутривенного введения или во флакон с пробкой, которую прокалывают гиподермической иглой для инъекций.
Изобретение также относится к продукту, получаемому в процессе производства, и к набору, содержащему материалы, применяемые для лечения, например, рака. Производимый продукт включает контейнер с меткой. Подходящие контейнеры включают, например, бутыли, флаконы и опытные пробирки. Контейнеры могут быть изготовлены с использованием множества материалов, таких как стекло или пластик. Контейнер содержит композицию, включающую антитело, приведенное в настоящем описании. Активный агент композиции представляет собой конкретное антитело. Метка на контейнере указывает, что композицию используют для лечения или профилактики конкретного заболевания или расстройства, и может также включать инструкции по применению in vivo, например, указанные выше.
Набор по настоящему изобретению включает контейнер, описанный выше, и второй контейнер, содержащий буфер. Он может также включать другие материалы, желательные с точки зрения коммерческого применения и с точки зрения пользователя, в том числе другие буферы, разбавители, наполнители, иглы, шприцы и вкладыши с инструкциями по применению.
Например, для лечения аутоиммунных заболеваний, в патогенез которых вовлекается адгезия воспалительных клеток (например, лейкоцитов), миграция и активация, таких как ревматоидный артрит и волчанка, антитело может вводиться совместно, например, с анти-LFA-1 антителом (таким как анти-CD11a или анти-CD18 антитело) или анти-ICAM антителом, таким как ICAM-1, -2 или -3. Дополнительные агенты для лечения ревматоидного артрита, сочетаемые с антителом по настоящему изобретению, включают EnbrelTM, DMARDS, например метотрексат, и НСПВС (нестероидные противовоспалительные средства). Может также использоваться более одного из таких других активных агентов, отличных от антитела по настоящему изобретению. Дополнительно может использоваться инсулин для лечения диабета, анти-IgE для лечения астмы, анти-CD11a для лечения псориаза, анти-альфа4бета7 и гормон роста (GH) для лечения воспалительного заболевания кишечника.
Кроме того, композиция может соответствующим образом вводиться вместе с эффективным количеством гипогликемического агента. В контексте настоящего описания термин «гипогликемический агент» относится к соединениям, которые используются для регуляции метаболизма глюкозы, предпочтительно пероральные агенты. В контексте настоящего описания более предпочтительными для применения человеком являются инсулин и пероральные гипогликемические агенты из класса сульфонимочевин, которые вызывают секрецию инсулина поджелудочной железой. Соответствующие примеры включают глибурид, глипизид и гликлазид. Кроме того, агенты, которые повышают чувствительность к инсулину или являются сенсибилизаторами инсулина, такие как бигуаниды (включая метформин и фенформин), и тиазолидендионы, такие как инсулиновый сенсибилизатор REZULINTMTM (троглитазон) и другие соединения, которые связываются с PPAR-гамма ядерным рецептором, охватываются данным определением и также являются предпочтительными.
Гипогликемический агент вводят млекопитающему с использованием любого подходящего способа, включая парентеральный, интраназальный, пероральный и любой другой эффективный режим введения. Более предпочтительно, введение осуществляют перорально. Например, таблетки MICRONASETM (глибурид), поставляемые на фармацевтический рынок компанией Upjohn, в концентрациях 1,25, 2,5, и 5 мг на таблетку, приемлемы для перорального введения. Обычная поддерживающая доза в составе такой терапии в случае диабета типа II составляет от примерно 1,25 до 20 мг в день, которая может вводиться в виде однократной дозы или при разделении на несколько приемов в течение дня, какой вариант будет наиболее подходящим (Physician's Desk Reference, 2563-2565 (1995)). Другие примеры основанных на глибуриде таблеток, доступные для приема, включают GLYNASETM (Upjohn) и DIABETATM (Hoechst-Roussel). GLUCOTROLTM (Pratt), представляющий торговое наименование глипизида (1-циклогексил-3-(п-(2-(5-метилпиразинкарбоксамид)этил)фенил)сульфонил)мочевины) в виде таблеток, доступных в дозе 5 и 10 мг, также рекомендуется для приема при диабете типа II пациентам, нуждающимся в гипогликемической терапии после диетической корректировки, или тем пациентам, которые перестали отвечать на другие сульфонилмочевины (Physician's Desk Reference, 1902-1903 (1995)). Могут также использоваться другие, отличные от сульфинилмочевины гипогликемические агенты, такие как бигуаниды (например, метформин и фенформин) или тиазолидиндионы (например, троглитозон) или другие лекарственные средства, воздействующие на действие инсулина. Если тиазолидиндион используют вместе с пептидом, его берут в обычных, принятых количествах, или даже в несколько сниженных количествах, которые можно откорректироваться с учетом эффектов, выявляемых при использовании одного пептида или в сочетании с дионом. Типичная доза троглитозона (REZULINTMTM) при использовании его одного составляет примерно 100-1000 мг в день, более предпочтительно 200-800 мг/день, и этот диапазон также приемлем в настоящем контексте (см., например, Ghazzi et al., Diabets, 46: 433-439 (1997)). Другие тиазолидиндионы, которые представляют собой более мощные инсулинсенсибилизирующие агенты, чем троглитазон, также используют в сниженных дозах.
F. Депонирование материалов
Приведенная ниже гибридомная клеточная линия была депонирована в Американской Коллекции типовых культур (American Type Culture Collection, 10801 University Blvd., Manassas, VA 20110-2209 USA (ATCC)):
Депонирование было проведено на условиях Будапештского Договора, включаещего условие международного признания депонированных материалов для целей патентований и их регламентирования (Будапештский Договор). Гарантируется поддержание жизнеспособности клеточной культуры в течение 30 лет от даты депонирования. Клеточная линия, доступная от ATCC на условиях Будапештского Договора, является субъектом договора между Genentech, Inc. и ATCC, который гарантирует, что (а) доступ к культуре в ходе рассмотрения патентной заявки будет определен официальным лицом в соответствии с 37 CFR § 1.14 и 35 USC §122 и (b) что все ограничения по доступности к депонированной культуре будут необратимо удалены после выдачи патента.
Владелец настоящей заявки согласился, что если культура, подлежащая депонированию, погибнет, или будет потеряна или разрушена при культивировании в соответствующих условиях, она будет сразу же заменена жизнеспособным образцом той же культуры. Доступность депонированной клеточной линии не должна истолковываться в контексте лицензии на практическое осуществление изобретения, противоречащее правам, предоставляемым официальным ведомством любого правительства, в соответствии с его патентными законами.
Приведенное выше описание считается достаточным для того, чтобы специалист в данной области мог осуществить настоящее изобретение. Настоящее изобретение не ограничивается депонированной культурой, поскольку вариант, относящийся к депонированию, представляют собой лишь одну иллюстрацию одного аспекта изобретения, и любая культура, которая функционально эквивалентна ей, охватывается областью настоящего изобретения. Информацию о депонировании материала не следует рассматривать как указание на то, что данное описание не является адекватным для практического осуществления любого аспекта изобретения, включая наилучший способ его осуществления, ее не следует также трактовать в плане ограничения области прилагаемой формулы изобретения какими-то конкретными иллюстративными вариантами. Фактически, различные модификации изобретения, в дополнение к показанным и описанным вариантам, будут очевидны специалистам в данной области на основе приведенного выше описания, которое охватывается областью прилагаемой формулы изобретения.
Ниже настоящее изобретение для целей лучшего пояснения описывается со ссылками на соответствующие примеры. Однако их не следует трактовать в плане ограничения области настоящего изобретения. Все литературные и патентные ссылки включены в настоящее описание в соответствующем объеме.
III. ПРИМЕРЫ
Несмотря на противоположный характер функциональных свойств, FcγRIIA человека (активирующий рецептор) и FcγRIIB человека (ингибирующий рецептор) представляют собой высоко гомологичные белки (участок гомологии выделен в виде прямоугольника на фиг.2А), которые различаются примерно по девяти аминокислотам в доменах связывания IgG1 и 3. Коммерчески доступные моноклональные антитела связываются и с FcγRIIA человека, и с FcγRIIB. Моноклональное антитело, которое специфически связывается с FcγRIIB, также будет полезно, тогда как дополнительная способность блокировать связывание IgG рассматривается как желательная.
В приведенных примерах и прилагаемых чертежах FcγRIIB обозначает FcγRIIB человека и в основном относится к FcγRIIB1 человека, если особо не указано иное. FcγRIIB взаимозаменяемо может рассматриваться как FcgRIIB, FcGRIIb, huFcγRIIB, huFcGRIIb, hFcRIIB, Fcγ-RIIb, FcγR2B, FcγR2b или IgGR. Специфические аллельные варианты обозначаются путем добавления нумерических показателей 1, 2 или 3, например huFcGRIIb1. FcεRI обозначает FcεRI человека и относится к FcεRIα человека. FcεRI может взаимозаменяемо относиться к FceRI, FceRIa, FcERI, IgER, IgE-R, FcεRIα, Fcε-RI или FcεRIa.
Антитела для любого из указанных выше белков указываются либо по их названию, либо в основном путем добавления приставки «анти-» к соответствующему белковому антигену, например анти-FcγRIIB, анти-IgER и т.п. Внеклеточные домены белка обозначаются путем добавления символа ECD к названию белка, например, FcγRIIB ECD. Клетки, экспрессирующие интересующий(ие) белок(ки), могут быть обозначены соответствующим образом так, чтобы в эти наименования включались вариации в названии белка в названии клеточной линии, и обозначаются добавлением термина «клетки».
Пример 1.0 Материалы и методы
1.1 Материалы
ПЦР с обратной транскриптазой проводят с использованием GeneAmpTM от Perkin Elmer Life Science. Плазмиду pGEX-4T2, колонки с белком А и реактивы, а также белок G FcγRIII: колонки и реагенты получают от Amersham Pharmacia Biotech. Колонки Ni-NTA и соответствующие реактивы приобретают от Qiagen, Valencia, CA. Концентраторы Centriprep-30 получают от Millipore, Bedford, MA. SDS-полиакриламидные гели и поливинилидендифторидные мембраны получают от NOVEX, San Diego, CA. FuGENE® 6 получают от Roche.
кДНК, кодирующие внеклеточные трансмембранные домены FcγRIIA человека (CD32A; His131 аллотип), FcγRIIB (CD32B) и FcγRIIIA (CD16A; Val158 аллотип); и изоформы глюкозо-6-фосфатизомеразы (GPI) FcγRIIB и FcγRIIA были получены от доктора J. Ravetch (Rockefeller University, New York). Аллотипы FcγRIIA-Arg131 и FcγRIIIA-Phe158 создают методом направленного мутагенеза (31). Информация по последовательностям: FcγRIIB1 (SEQ ID NO:11) также доступна под номером NP_003992; FcγRIIB2 (SEQ ID NO:10) c номером доступа NP_001002273; FcγRIIA (SEQ ID NO:9) c номером доступа NP_067674 и FcγRIII (две изоформы) c номерами доступа NP_000560 и NP_000561.
Антитело AT10 получают от Biosource International, Camirillo, CA. Антитело mopc21 получают от BD Pharmagen. Мышиные моноклональные антитела получают из следующих источников: 32.2 (анти-FcγRI), IV.3 (анти-FcγRII) и 3G8 (анти-FcγRIII) - от Medarex, Annandale, NJ; и B1G6 (анти-b2-микроглобулин) получают от Beckman Coulter, Palo Alto, CA. Анти-GST антитело получают от Zymed Laboratories Inc. Анти-GST-биотин представляет собой получаемый получают от Genentech клон 15H4.1.1. JW8.5.13 получают от Serotec Inc., Raleigh, NC.
Планшеты для ELISA, например планшеты Nunc® maxisorb, получают от Nalge-Nunc, Naperville, IL. Планшеты для культуры ткани могут быть получены, например, от Linbro или Fisher. Бычий сывороточный альбумин (BSA), Tween 20®, Triton Х-100, EMEM (минимальная эссенциальная среда Игла), иономицин, протаминсульфат, дигидрохлорид о-фениленлдиамина (OPD) и иодид пропидия получают от Sigma (St. Louis, MO). Стрептавидин и казеиновый блокатор (Prod # 37528) получают от Pierce (Rockford, IL). Конъюгат противомышиного IgG антитела кролика и пероксидазы хрена и конъюгированный с пероксидазой F(ab')2 фрагмент козьего противочеловеческого F(ab')2-специфического IgG получают от Jackson ImmunoResearch Laboratoreis (West Grove, PA). Конъюгированный с пероксидазой белок G получают от Bio-Rad. Стрептавидин-HRP получают от Boehringer Mannheim или Zymed. Субстрат TMB (Prod # TMBW-0100-01) и останавливающий раствор (Prod # BSTP-0100-01) получают от BioFx Laboratory. Противомышиный козий IgG-флуоресцеин получают от American Qualex Labs. NP-(11)-OVA и TNP-(11)-OVA получают от Biosearch Technologies, Inc. (Novado, Ca). Стрептавидин-PE и противомышиный крысиный IgG-PE или флуоресцеиновые конъюгаты получают от BD Pharmagen (Franklin, Lakes, NJ).
Проточную цитометрию проводят на проточном цитрометре FACScanTM или FACSCaliburTM от BD Pharmagen (Franklin, Lakes, NJ). Поглощение измеряют с использованием счетчика для планшетов Vmax от Molecular Devices (Mountain View, CA). Метод ELISA по гистамину проводят с использованием гистаминового набора для ELISA, получаемого от IBL Immunobiological Labs (Hamburg, Germany), поставляемого от RDI, Inc (NJ).
1.2 Получение белков слияния GST-Fc рецептор
кДНК для FcγRI (CD64) выделяют путем ПЦР с обратной транскриптазой олиго(dT)-праймированной РНК из U937 клеток с использованием праймеров, которые образуют фрагмент, кодирущий внеклеточный домен α-цепи. Кодирующие участки всех рецепторов субклонируют в описанных ранее векторах экспрессии для клеток млекопитающих pRK (Eaton, D. et al., 1986, Biochemistry 25: 8343-8347). В случае всех плазмид FcγR pRK, трансмембранный и внутриклеточный домены заменяют ДНК, кодирующей метку Gly-His6 и глютатион-S-трансферазу человека (GST). Последовательность GST из 234 аминокислот получают методом ПЦР из плазмиды pGEX-4T2 с сайтами рестрикции для NheI и XbaI на 5'- и 3'-концах соответственно. Таким образом, экспрессированные белки содержат внеклеточные домены α-цепи, слитые по их карбоксильным концам с Gly/His6/GST в следующих положениях аминокислот: FcγRI, His292; FcγRIIA, Met216; FcγRIIB, Met195; FcγRIIIA, Gln191 (номера остатков включают сигнальные пептиды).
Плазмиду трансфицируют в трансформированные аденовирусом клетки линии эмбриональных клеток почки человека 293 по методу осаждения фосфатом кальция (Gorman et al., 1990, DNA Prot. Eng. Tech. 2: 3-10). Супернатанты собирают через 72 часа после превращения в бессывороточную PSO4-содержащую среду с добавкой 10 мг/л рекомбинантного бычьего инсулина, 1 мг/л трансферина человека и следовых количеств элементов. Белки очищают хроматогрфией с использованием никельнитрилотриуксусной кислоты (Ni-NTA) и заменяют буфер на фосфатно-буферный раствор (PBS) с использованием концентраторов Centriprer-30. Белки анализируют на 4-20% SDS-полиакриламидных гелях, переносят на поливинилидендифторидные мембраны и проводят секвенирование по аминоконцам последовательности для подтверждения корректности расщепления сигнальной последовательности. Оценивают конформацию рецептора методом ELISA с использованием мышиных моноклональных антител 32.2 (анти-FcγRI), IV.3 (анти-FcγRII), 3G8 (анти-FcγRIII) и B1G6 (анти-b2-микроглобулин). Концентрации рецепторов определяют по поглощению при длине волны 280 нм с использованием коэффициентов возбуждения, полученных при анализе аминокислотного состава.
1.3 Получение FcγRIIB антител
FcγRIIB-специфичные антитела человека, которые блокируют IgG Fc-связывание с рецептором, получают против белков слияния FcγRIIB-His6-GST. Мышей BALB/c иммунизируют, вводя в подушечки лап 2 мкг huFcγRIIB-His6-GST. Спленоциты от иммунизированных мышей сливают с миеломными клетками P3X63Ag8U1 (клетки, описанные в Oi VT, Herzenberg LA., 1981, Immunoglobulin producting hybrid cell lines. In: Selecred methods in cellular immunology (Mishell BB, Csiigi SM, eds), pp 351-372. San Francisco: Freeman) с получением примерно 900 гибридом.
Метод ELISA осуществляют следующим образом: белок слияния с рецептором в количестве примерно 1,5 мг/мл в PBS, pH 7,4, наслаивают на ячейки планшетов для ELISA и оставляют на 18 часов при 4°С. Планшеты блокируют добавлением буфера для тестирования при 25°С на 1 час. Подвергаемые скринингу серийные 3-кратные разбавления антител и контрольные антитела (10,0-0,0045 мг/мл) добавляют к планшетам и инкубируют в течение 2 часов. После промывки планшетов буфером для тестирования IgG, связанный с рецептором, выявляют с использованием конъюгированного с пероксидазой фрагмента F(ab')2 козьего противочеловеческого F(ab')2-специфичного IgG или белка G, конъюгированного с пероксидазой. Используемый субстрат представляет собой дигидрохлорид о-фенилендиамина. Поглощение при длине волны 490 нм определяют с использованием счетчика для планшетов Vmax.
1.4 Первичный скрининг FcγRIIB-специфичных антител
При проведении первичного скрининга супернатант, содержащий антитела, экспрессированные из гибридомных субклонов, тестируют на наличие позитивного связывания с FcγRIIB-His6-GST. Антитела, реактивные с FcγRIIB-His6-GST, по данным метода ELISA, подвергают повторному скринингу для определения способности связываться с FcγRIIB-His6-GST и для подтверждения негативного результата по связыванию с FcγRIIA(R131 вариант)-His6-GST и FcγRIII(F158 вариант)-His6-GST, по данным метода ELISA.
Отбирают примерно 50 антител в ходе первичного скрининга для дальнейшего анализа.
1.5 Вторичный скрининг FcγRIIB-специфичных антител
При проведении вторичного скрининга антитела подвергают повторному скринингу для определения специфичности к рецептору по методу ELISA и для анализа связывания с клетками с использованием клеточных линий CHO, экспрессирующих глюкозо-6-фосфатизомеразу (GPI), связанных FcγRIIB и FcγRIIA. ELISA проводят, как было описано выше, и результаты анализов показаны на фиг.4. Приведенная на фиг.4 диаграмма показывает относительные уровни связывания антител с GST-huFcγRIIB относительно GST-huFcγRIIA и белков слияния GST-huFcγRIII. Антитела 1D1, 5A6,
5H11 и 6А5 селективно связываются с GST-huFcγRIIB относительно GST-huFcγRIIA и белков слияния GST-huFcγRIII. Антитело 5В9 селективно связывается и с GST-huFcγRIIB, и с GST-huFcγRIIA относительно GST-huFcγRIII.
На фиг.5 приведены данные по специфичности связывания, по результатам определения методом иммунофлуоресценции связывания с клетками CHO, экспрессирующими GPI-huFcγRIIB, относительно клеток CHO, экспрессирующих GPI-huFcγRIIA. Отдельные аликвоты клеток CHO окрашивают контрольным изотипом mIgG1 (mopc21) или (противоFcγRIIB человека) моноклональными антителами 1D1, 5A6, 5B9, 5D11 и 6А5. Связывание выявляют опосредованно при вторичной инкубации с конъюгированным с флуоресцеином F(ab')2 противомышиного козьего IgG (F(ab')2-специфическим антителом) и анализируют методом проточной цитометрии. Антитело 5А6 предпочтительно связывается с клетками CHO, экспрессирующими GPI-huFcγRIIB относительно клеток CHO, экспрессирующих GPI-huFcγRIIA. Результаты аналогичны связыванию с конструкциями GST.
Дополнительные данные по связыванию, полученные методом ELISA, проиллюстрированы на фиг.6-9. На фиг.6-9 показан график, демонстрирующий аффинность по связыванию различных анти-FcγRII (CD32) МАт с GST-huFcγRIIB, GST-huFcγRIIA(H131) или GST-huFcγRIIA (R131). AT10 представляет собой mIgG, специфичный для FcγRIIBA, и mopc21 представляет собой контрольный изотип mIgG. 5А6 mIgG1 характеризуется значением EC50, равным 0,06 нМ, для связывания с GST-huFcγRIIB, как показано на фиг.6. Тогда как EC50 для 5А6 mIgG1 в случае связывания GST-huFcγRIIA (Н131) выше, чем 50 мкг/мл (фиг.9), и для связывания GST-huFcγRIIA (R131) составляет 2,5 мкг/мл (фиг.8).
1.6 Экспрессия антитела и его очистка
Антитело 5А6.2.1 (в контексте настоящего описания называемое взаимозаменяемо как 5А6.2.1 или 5А6) отбирают для асцитов и очищают хроматографией на белке G (Amersham Pharmacia Biotech). Выделяют ДНК, кодирующую 5А6.2.1, и проводят секвенирование с использованием традиционных методик. Аминокислотные последовательности и CDR тяжелой цепи (SEQ ID NO: 7) и легкой цепи (SEQ ID NO: 8) показаны на фиг.10. CDR для тяжелых цепей включают: DAWMD (SEQ ID NO: 1), EIRSKPNNHATYYAESVKG (SEQ ID NO: 2) и FDY (SEQ ID NO: 3). CDR для легких цепей включают: RASQEISGYLS (SEQ ID NO: 4), AASALDS (SEQ ID NO: 5) и LQYVSYPL (SEQ ID NO: 6).
Предположительные связывающие эпитопы для 5А6 моноклонального антитела включают аминокислотные остатки K158-V161 и F174-N180, где нумерация показана для FcγRIIB2 на фиг.2А (FcγRIIB2, SEQ ID NO: 10). Рецепторы FcγRIIB1 и FcγRIIB2 содержат структурные домены, показанные на фиг.2А и 2В (проиллюстрированы на примере FcγRIIB2), в виде IgG-подобного домена 1 на участке остатков T43-P123 и IgG-подобного домена 3 на участке остатков W132-P217. ITIM мотив показан на фиг.2А для FcγRIIB2 и включает остатки N269-M277. В последнее время появилось сообщение о том, что аминокислотная последовательность FcγRIIA F165-T171, показанная в виде FSRLDPT (SEQ ID NO: 39) на фиг.2А, может представлять собой FSHLDPT (SEQ ID NO: 40), что указывает на большее различие по последовательностям между FcγRIIA и FcγRIIB в предположительном эпитопе связывания FcγRIIB для антитела 5А6 (см. фиг.2 и номера доступа NP_067674, SEQ ID NO:30, где указанная аминокислотная последовательность также включает изменения аминокислотных остатков на N-концевом участке FcγRIIA).
1.7 Конкуренция с комплексами Е27:IgE
Данный тест используется для скрининга способности 5A6 MAb препятствовать связыванию IgG1 с FcγRIIA и FcγRIIB. FcγRII обладают слабой аффинностью к мономерным IgG1 и соответственно IgG1-связывание тестируют с использованием стабильного гексамерного комплекса из трех IgE и трех молекул анти-IgE, например E27, гуманизированного IgG1 антитела, которое связывается с IgE (Shields, R. L. et al., J. Biol. Chem., 276: 6591-6604 (2001)). MAb 5А6 подвергают скринингу на способность нейтрализовать связывание с IgG при оценке способности антитела конкурировать с FcγRIIA и FcγRIIB человека за связывание с гексамерными комплексами E27-IgE. Данный конкурентный тест проводят по указанной ниже процедуре, и результаты его показаны на фиг.11 и 12.
Белки слияния FcγRIIB и FcγRIIA в концентрации 1 мг/мл в PBS, pH 7,4, наслаивают на ячейки планшетов для ELISA и оставляют на 48 часов при 4°С. К планшетам добавляют для блокирования трис-буферный солевой раствор, 0,5% бычьий сывороточный альбумин, 0,05% полисорбат-20, 2 мМ ЭДТА, pH 7,45 (буфер для тестирования), при 25°С в течение 1 часа. Гексамерный комплекс E27-IgE получают в буфере для тестирования путем смешивания эквимолярных количеств Е27 и миеломного IgE человека (Nilsson, K., Bennich, H., Johansson, S.G.O. and Ponten, J., (1970) Clin. Exp. Immunol. 7: 477-489) при 25°С в течение 1 часа. E27-IgE (10,0 мг/мл в буфере для тестирования) добавляют к ячейкам планшета и инкубируют в течение 2 часов. Планшеты промывают для удаления несвязанного E27-IgE. 5А6 MAb, 5А6 F(ab)2, 5A6 Fab, mIgG1 (контроль) и 5B9 (анти-FcγRIIA/B) готовят в буфере для тестирования в различных концентрациях от 0,01 нМ до 100 нМ. Антитела добавляют в отдельные ячейки и инкубируют в течение 1 часа. После промывания планшетов буфером для тестирования выявляют гексамерные комплексы E27-IgE, которые остались связанными с FcγRIIA или FcγRIIB в присутствии конкурентного антитела. Выявление включает связывание с IgG1 частью E27 конъюгированного с пероксидазой F(ab')2 фрагмента козьего противочеловеческого F(ab')2-специфичного IgG. Выявляемый субстрат пероксидазы, используемый в данном случае, представляет дигидрохлорид о-фенилендиамина. Определяют поглощение при длине волны 490 нм с использованием счетчика для планшетов Vmax. На фиг.11 показано, что 5А6 не блокирует связывание гексамера E27-IgE с huFcγRIIA, как видно из сохраняющегося связывания гексамера E27-IgE с FcγRIIA при повышении концентрации конкурентного антитела (5А6 MAb, 5А6 F(ab)2, 5A6 Fab, mIgG1 и 5В9). Только антитело 5В9, в отношении которого известно, что оно связывается и с FcγRIIA, и с FcγRIIB (см. фиг.4 и 5), оказалось способным конкурировать за связывание с гексамером E27-IgE. На фиг.12 показано, что 5А6 конкурирует с FcγRIIB за связывание с гексамером E27-IgE, что следует из снижения связывания гексамера E27-IgE при повышении 5А6 антитела, Fab или F(ab)2. Как и ожидалось, контрольное IgG1 антитело не конкурирует. Связывание антител с huFcγRIIB, (5А6, 5А5, 5Н11.1 и 5А6 Fab'2) и IgG1 (гексамер E27-IgE) с FcγRIIB, FcγRIIA (R131) или FcγRIIA (H131) проиллюстрировано также на фиг.13-16. На фиг.14 показано, что IgG не связывается с FcγRIIB в присутствии антител 5А6.2.1 и 6А5, тогда как связывание IgG с FcγRIIA (R131), показанное на фиг.13, и связывание IgG с FcγRIIA (H131), показанное на фиг.15, не блокируется.
1.8 Анализ связывания с использованием метода иммунофлуоресценции
На фиг.16 показан опосредованный анализ связывания с использованием иммунофлуоресценции 5А6 MAb с нативным FcγRIIA, экспрессированным на эритролейкемических клетках К562 (ATCC № CCL-243). Отдельные аликвоты клеток K562 окрашивают контрольным изотипом mIgG1 (mopc21), моноклональным антителом 5A6 (противочеловеческий FcγRIIB) или моноклональным антителом Medarex 4.3 MAb (противочеловеческие FcγRIIA/B). Связывание выявляют опосредованно при проведении вторичной инкубации с конъюгированным с флуоресцеином F(ab')2 козьего противомышиного IgG (F(ab')2-специфическим антителом) и анализируют методом проточной цитометрии. Medarex 4.3 MAb связывается с huFcγRIIA (CD32A), как показано на фиг.16. 5A6, анти-huFcγRIIB (анти-CD32B) антитело не связывается с huFcγRIIA (CD32A), что соответствует контрольному изотипу mopc21 антитела, которое также не связывается с huFcγRIIA (CD32A), как показано прерывистой линией на фиг.4.
ПРИМЕР 2.0 Свойства анти-FcγRIIB антитела
2.1 Материалы
Анти-FcεRI MAb, 22Е7 MAb связывается с FcεRI при наличии или в отсутствие 22Е7, связанного с рецептором. MAb 22Е7 выделяют и очищают из клеточной линии, полученной от Hoffman-LaRoche, IGE4R: 22E7.2D2.1D11 (Risek, F., et al., 1991, J. Biol. Chem. 266: 11245-11251). Клетки от Hoffman-LaRoche, экспрессирующие 22E7 MAb, растят в средах Дульбекко в модификации Искова с добавлением 10× PBS, 1× Pen-Strep и 1× глутамина. MAb 22E7 очищают хроматографией с белком А и белком G. Объединяют содержащие 22E7 экстракты и подтверждают наличие аффинности для FcεRI.
2.2 Клеточные линии RBL
Клеточная линия RBL48, полученная из родительской линии тучных клеток крысы RBL-2H3 (ATCC# CRL-2256), экспрессирует α-субъединицу высокоаффинного рецептора IgE человека (FcεRI) (Gilfillian A. M. et al., 1992, Immunology, 149: 2445-2451). Клеточную линию RBL48 трансфицируют по методу электропорации клоном кДНК для α-субъединицы полной длины FcγRIIB1 человека (Muta, T., et al., 1994, Nature 368: 70-73), который был субклонирован в векторе экспрессии с селектируемым пуромициновым маркером (Morgenstern, J.P., et al., 1990, Nucleic Acid Research, 18: 3587-3596). Клоны отбирают с использованием 1 мкМ пуромицина и анализируют на наличие экспрессии FcγRIIB на клеточной поверхности путем иммунофлуоресцентного окрашивания с использованием против FcγRIIB человека моноклонального антитела 5A6.2.1. Выбранный субклон был обозначен как RBL48.C.4.
2.3 Высвобождение гистамина
Эффекты сшивки FcγRIIB (которые также могут взаимозаменяемо обозначаться как совместная перекрестная сшивка, коагрегация или колигирование) на активирующих рецепторах определяют количественно на основании определения способности антитела блокировать высвобождение гистамина из сенсибилизированных аллергеном клеток RBL48.C.4. Ниже описаны методы тестирования, а их результаты показаны на фиг.17.
Клон RBL48.C.4 инкубируют в ячейках 96-ячеечного микротитрационного планшета с плоским дном в буфере для тестирования (EMEM (минимальная эссенциальная среда Игла с добавлением Earle's BSS) с 2 мМ L-глютамина, 1 мМ пирувата натрия, 0,1 мМ необязательных аминокислот, 1,5 г/л бикарбоната натрия, пенициллина, стрептомицина, 15% фетальной сыворотки теленка) с добавлением 2 мкг/мл анти-FcεRI MAb 22Е7 и контрольного изотипа mIgG1 (mopc21) или 5А6 MAb в концентрациях, варьирующих от 0,002 до 2 мкг/мл, при 37°С в течение 30 минут в инкубаторе в атмосфере CO2. Клетки промывают два раза буфером для тестирования и инкубируют с F(ab)'2 фрагментом козьего противомышиного Fc-специфичного сшивающего антитела при 37°С в течение 30 минут. Супернатанты отбирают и исследуют на содержание гистамина методом ELISA, как было описано выше, с использованием гистаминового набора для ELISA.
Показатели высвобождения гистамина выражают в виде среднего значения и СКО по результатам измерения в трех ячейках и представляют графически на фиг.5. Для ингибирования высвобождения гистамина требуется и 5A6, и 22E7 со сшивающим антителом. Высвобождение гистамина подавляется связыванием 5A6 с FcγRIIB и связыванием 22E7 с FcεRI, где 5A6 и 22E7 также сшиваются козьим противомышиным Fc-специфичным сшивающим антителом. Наиболее эффективным для ингибирования высвобождения гистамина было соотношение 5A6 к 22E7, равное 1:1, хотя заметная супрессия наблюдалась уже в соотношении 1:10, 1:100 и 1:1000.
ПРИМЕР 3.0 Получение биспецифических антител
В данном примере описывается конструирование и очистка биспецифических антител, содержащих вариант шарнирного участка без цистеиновых остатков, образующих дисульфидную связь («бесшарнирных»). Описано также конструирование биспецифических антител, содержащих шарнирную последовательность дикого типа; указанные антитела могут использоваться для оценки эффективности получаемых комплексов антител различного вида.
3.1 Конструирование векторов экспрессии
Все плазмиды, используемые для экспрессии антител полной длины, основаны на отдельной цистронной системе (Simmons et al., 2002, J. Immunol. Methods, 263:133-147; Simmons, et al., патент США № 5840523), которая основана на отдельно локализованных промоторах phoA (AP) (Kikuchu et al., 1981, Nucleic Acids Res., 9: 5671-5678) для транскрипции тяжелой и легкой цепей, за которыми следует trp последовательности Shine-Dalgarno для инициации трансляции (Yanovsky et al., 1981, Nucleic Acids Res., 9: 6647-6668, и Chang et al., 1987, Gene, 55: 189-196). Дополнительно используют термостабильную сигнальную последовательность энтеротоксина II (STII) (Picken et al., 1983, Infect. Immun. 42: 269-275 и Lee et al., 1983, Infect. Immun., 42:264-268) для периплазматической секреции тяжелой и легкой цепей. Тонкий контроль трансляции обеих цепей достигается с использованием описанных ранее вариантов STII сигнальной последовательности с определенной относительной силой трансляции, которые содержат молчащие изменения в кодонах на участке инициации трансляции (TIR) (Simmons and Yansura, 1996, Nature Biotechnol., 14: 629-634, и Simmons et al., выше). В контексте настоящего описания сочетание силы трансляции для конкретной пары TIR в составе вектора обозначается выражением (N - легкая цепь, M - тяжелая цепь), где N обозначает относительную силу TIR для легкой цепи и M обозначает относительную силу TIR для тяжелой цепи. И, наконец, терминатор транскрипции λt0 (Schlosstissek and Grosse, 1997, Nucleic Acids Res., 15: 3185) помещают в направлении транскрипции кодирующих последовательностей обеих цепей. Для всех плазмид используют рамки считывания pBR322-векторной системы (Sutcliff, 1978, Cold Spring Harbor Symp. Quant. Biol.,43: 77-90).
Для усиления ассоциации биспецифических полипептидных цепей вводят мутации по типу «выступ и впадина» (“knob-and-hole”) в участки димеризации. Следует понимать, что любая цепь может включать мутацию по типу «выступа» («knob»), тогда как другая цепь будет включать комплементарную мутацию по типу «впадина» («hole»). Настоящее изобретение включает оба варианта. В приведенном репрезентативном примере плечо 5А6 биспецифического антитела конструируют таким образом, чтобы оно включало мутацию по типу «выступа» («knob»), а плечо 22E7 биспецифического антитела конструируют таким образом, чтобы оно включало комплементарную мутацию по типу «впадина» («hole»).
(i) Плазмида p5A6.11.Knob.Hg-
Для создания желательной плазмиды p5A6.11.Knob.Hg требуются две промежуточных плазмиды. Вначале переносят вариабельный домен из 5A6 (анти-FcγRIIB) химерной легкой цепи в плазмиду pVG11.VNERK.Knob для создания промежуточной плазмиды p5A6.1.L.VG.1.H.Knob. Затем вариабельный домен 5A6 из химерной тяжелой цепи переносят на плазмиду p5A6.1.L.VG.1.H.Knob для создания промежуточной плазмиды p5A6.11.Knob. Ниже описана методика получения указанных промежуточных плазмид p5A6.1.LC.VG.1.HC.Knob и p5A6.11.Knob с последующим конструированием p5A6.11.Knob.Hg- и p5A6.1.L.VG.1.H.Knob.
Данную плазмиду конструируют для целей переноса вариабельного домена мышиной легкой цепи антитела 5A6 в плазмиду, совместимую для целей создания антитела полной длины, несущего тяжелую-легкую цепь (H/L) мономерного антитела. Конструирование данной плазмиды включает лигирование двух фрагментов ДНК. Первый из них представляет вектор pVG11.VNERK.Knob, из которого удален фрагмент EcoRI-PacI. Плазмида pVG11.VNERK.Knob представляет собой производное отдельного цистронного вектора с относительной силой TIR 1 для легкой, и 1 - для тяжелой цепей (Simmons et al., 2002, выше), где вариабельные домены легкой и тяжелой цепей были изменены на соответствующий вариант анти-VEGF антитела (VNERK) за счет мутации по типу «выступа» («knob») (T366W) (Merchant et al., 1998, Nature Biotechnology 16: 677-681) с обеспечением всех описанных выше контрольных элементов. Вторая часть процесса лигирования включает лигирование последовательности, изображенной на фиг.25 (SEQ ID NO: 35), в описанном выше векторе pVG11.VNERK.Knob, расщепленном EcoRI-PacI. Данная последовательность кодирует промотор щелочной фосфатазы (phoA), сигнальную последовательности STII и полную (включая вариабельный и константный домены) легкую цепь антитела 5A6.
p5A6.11.Knob
Данную плазмиду конструируют для целей введения вариабельного домена мышиной тяжелой цепи 5A6 антитела в каркасную область тяжелой цепи человеческого антитела для создания химерного мономерного антитела полной длины, включающего тяжелую цепь/легкую цепь (H/L). Конструирование p5A6.11.Knob включает лигирование двух фрагментов ДНК. Первый фрагмент представляет собой описанный выше вектор p5A6.1.L.VG.1.H.Knob, из которого был удален малый фрагмент MluI-PspOMI. Второй фрагмент включает лигирование последовательности, изображенной на фиг.27 (SEQ ID NO: 37), в вектор p5A6.1.L.VG.1.H.Knob, расщепленный MluI-PspOMI. Данная последовательность кодирует последние 3 аминокислоты в сигнальной последовательности STII и примерно 119 аминокислот из вариабельного домена тяжелой цепи мышиного антитела 5A6.
p5A6.11.Knob.Hg-
Плазмиду p5A6.11.Knob.Hg- конструируют для целей экспрессии химерного бесшарнирного Knob антитела 5A6 полной длины, включающего тяжелую/легкую (H/L) цепь мономерного антитела. Конструирование плазмиды вклчюает лигирование двух фрагментов ДНК. Первый фрагмент представляет собой вектор p5A6.11.Knob, указанный выше, из которого был удален фрагмент PspOMI-SacII. Второй фрагмент представляет собой фрагмент PspOMI-SacII, включающий примерно 514 пар оснований из p4D5.22.Hg-, кодирующего примерно 171 аминокислоту из тяжелой цепи человека, в которой два цистеина из шарнирной области были преобразованы в серины (C226S, C229S, EU схема нумерации приведена согласно Kabat, E.A., et al., (eds.), 1991, page 672 in Seguences of proteins of Immunological interest, 5th ed. Vol. 1. NIH, Bethesda MD). Плазмида p4D5.22.Hg- представляет собой производное отдельного цистронного вектора с относительной силой TIR, равной 2 для легкой и 2 - для тяжелой цепей (Simmons et al., J. Immunol. Methods, 263:133-147 (2002)), в которой вариабельные домены легкой и тяжелой цепей изменены на анти-HER2 антитело и два цистеина в шарнирной области преобразованы в серины (C226S, C229S).
(ii) Плазмида p5A6.22.Knob.Hg-
Для создания желательной плазмиды p5A6.22.Knob.Hg- требуется одна промежуточная плазмида. Промотор phoA и сигнальную последовательность STII (с относительной силой TIR, равной 2 для легкой цепи) переносят вначале в плазмиду p5A6.22.Knob.Hg- для создания промежуточной плазмиды p5A6.21.Knob.Hg-. Ниже описана методика получения промежуточной плазмиды p5A6.21.Knob.Hg- с последующим конструированием p5A6.22.Knob.Hg-.
p5A6.21.Knob.Hg-
Данную плазмиду конструируют для введения STII сигнальной последовательности (с относительной силой TIR, равной 2) в легкую цепь. Конструирование p5A6.21.Knob.Hg- включает лигирование трех фрагментов ДНК. Первый фрагмент представляет собой вектор p5A6.11.Knob.Hg-, из которого был удален малый EcoRI-PacI фрагмент. Второй фрагмент представляет собой фрагмент Nsil-PacI, включающий примерно 658 пар оснований из плазмиды p5A6.11.Knob.Hg-, кодирующей легкую цепь химерного антитела 5А6. Третья часть системы лигирования представляет собой фрагмент ПЦР EcoRI-NsiI, включающий примерно 489 пар оснований, полученный из плазмиды p1H1.22.Hg- с использованием следующих праймеров:
5'-AAAGGGAAAGAATTCAACTTCTCCAGACTTTGGATAAGG
(SEQ ID NO: 27)
5'-AAAGGGAAAATGCATTTGTAGCAATAGAAAAAACGAA
(SEQ ID NO: 28)
Плазмида p1H1.22.Hg- представляет собой производное отдельного цистронного вектора с относительной силой TIR, равной 2 для легкой цепи и 2 - для тяжелой цепи (Simmons et al., J. Immunol. Methods, 263:133-147 (2002)), в котором вариабельные домены легкой и тяжелой цепей изменены на антитело крысы против тканевого фактора и два цистеина в шарнирной области преобразованы в серины (C226S, C229S).
p5A6.22.Knob.Hg-
Данную плазмиду конструируют для введения STII сигнальной последовательности с относительной силой TIR, равной 2 для тяжелой цепи. Конструирование p5A6.22.Knob.Hg- включает лигирование двух фрагментов ДНК. Первый фрагмент представляет собой вектор p5A6.21.Knob.Hg-, из которого был удален малый фрагмент PacI-MluI. Вторая часть системы лигирования представляет собой фрагмент PacI-MluI, включающий примерно 503 пары оснований из плазмиды p1H1.22.Hg-, кодирующий терминатор транскрипции λt0 для легкой цепи, промотор phoA и STII сигнальную последовательность (с относительной силой TIR, равной 2 для тяжелой цепи).
(iii) Плазмида p22E7.11.Hole.Hg-
Требуются две промежуточных плазмиды для создания желательной плазмиды p22E7.11.Hole.Hg-. Вначале переносят вариабельный домен химерной легкой цепи 22E7 (анти-FcεRI) в плазмиду pVG11.VNERK.Hole с получением промежуточной плазмиды p22E7.1.L.VG.1.H.Hole. Вариабельный домен химерной тяжелой цепи 22E7 затем переносят на плазмиду p22E7.1.L.VG.1.H.Hole с получением промежуточной плазмиды p22E7.11.Hole. Ниже описана методика получения указанных промежуточных плазмид p22E7.1.L.VG.1.H.Hole и p22E7.11.Hole с последующим конструированием p22E7.11.Hole.Hg-.
p22E7.1.L.VG.1.H.Hole
Данную плазмиду конструируют для переноса вариабельного домена легкой цепи антитела 22E7 мыши в плазмиду, совместимую для целей создания мономерного антитела полной длины, включающего тяжелую/легкую цепь (H/L). Конструирование данной плазмиды включает лигирование двух фрагментов ДНК. Первый фрагмент представляет собой вектор pVG11.VNERK.Hole, из которого был удален малый фрагмент EcoRI-PacI. Плазмида pVG11.VNERK.Hole представляет собой производное отдельного цистронного вектора с относительной силой TIR 1 - для легкой цепи и 1 - для тяжелой цепи (Simmons et al., 2002, J. Immunol. Methods, 263:133-147), в котором вариабельные домены легкой и тяжелой цепей изменены на анти-VEGF антитело (VNERK), содержащего мутации по типу «впадины» (“hole”) (T36S, L368A, Y407V) (Merchant et al., 1998, Nature Biotechnology 16: 677-681), а также все контрольные элементы, описанные выше. Вторая часть процесса лигирования включает лигирование последовательности, изображенной на фиг.26 (SEQ ID NO: 36), в векторе pVG11.VNERK.Hole, расщепленном EcoRI-PacI, как было описано выше. Данная последовательность кодирует промотор щелочной фосфатазы (phoA), сигнальную последовательности STII и полностью (включая вариабельный и константный домены) легкую цепь антитела 22E7.
p22E7.11.Hole
Данную плазмиду конструируют для целей введения вариабельного домена тяжелой цепи мышиного антитела 22E7 в каркасную область тяжелой цепи антитела человека для создания химерного антитела полной длины, включающего тяжелую цепь/легкую цепь (H/L) мономерного антитела. Конструирование p22E7.11.Knob включает лигирование двух фрагментов ДНК. Первый фрагмент представляет собой вектор p22E7.1.L.VG.1.H.Hole, описанный выше, из которого был удален малый фрагмент MluI-PspOMI. Вторая часть системы лигирования включает лигирование последовательности, изображенной на фиг.28 (SEQ ID NO: 38), в векторе p22E7.1.L.VG.1.H.Hole, расщепленном MluI-PspOMI. Данная последовательность кодирует последние 3 аминокислоты в сигнальной последовательности STII и примерно включает 123 аминокислоты из вариабельного домена тяжелой цепи мышиного антитела 22E7.
p22E7.11.Hole.Hg-
Плазмиду p22E7.11.Hole.Hg- конструируют для целей экспрессии химерного бесшарнирного антитела полной длины 22E7 Hole, включающего тяжелую/легкую цепь (H/L) мономерного антитела. Конструирование плазмиды включает лигирование двух фрагментов ДНК. Первый фрагмент представляет собой вектор p22E7.11.Hole, из которого был удален небольшой фрагмент PspOMI-SacII. Вторая часть системы лигирования включает фрагмент PspOMI-SacII, состоящий примерно из 514 пар оснований, из p4D5.22.Hg-, кодирующий тяжелую цепь человека, включающую примерно 171 аминокислоту, в которой два цистеиновых остатка в шарнирной области были преобразованы в серины (C226S, C229S).
(iv) Плазмида p22E7.22.Hole.Hg-
Для создания желательной плазмиды p22E7.22.Hole.Hg- требуется одна промежуточная плазмида. Промотор phoA и сигнальную последовательность STII (с относительной силой TIR, равной 2) для легкой цепи переносят вначале в плазмиду p22E7.11.Hole.Hg- для создания промежуточной плазмиды p22E7.21.Hole.Hg-. Ниже описана процедура получения промежуточной плазмиды p22E7.21.Hole.Hg- с последующим конструированием p22E7.22.Hole.Hg-.
p22E7.22.Hole.Hg-
Данную плазмиду конструируют для введения STII сигнальной последовательности (с относительной силой TIR, равной 2) для легкой цепи. Конструирование p22E7.21.Hole.Hg- ключает лигирование трех фрагментов ДНК. Первый фрагмент представляет собой вектор p22E7.11.Hole.Hg-, из которого был удален малый фрагмент EcoRI-PacI. Второй фрагмент представляет собой фрагмент EcoRV-PacI с длиной примерно в 647 пар оснований из плазмиды p22E7.11.Hole.Hg-, кодирующий легкую цепь химерного антитела 22E7. Третий фрагмент представляет собой фрагмент EcoRI-EcoRV размером примерно 500 пар оснований из плазмиды p1H1.22.Hg-, кодирующий промотор щелочной фосфатазы (phoA) и сигнальную последовательность STII.
p22E7.22.Hole.Hg-
Данную плазмиду конструируют для введения STII сигнальной последовательности (с относительной силой TIR, равной 2) для тяжелой цепи. Конструирование p22E7.22.Hole.Hg- включает лигирование трех фрагментов ДНК. Первый фрагмент представляет собой вектор p22E7.21.Hole.Hg-, из которого был удален малый фрагмент EcoRI-MluI. Второй фрагмент представляет собой фрагмент EcoRI-PacI размером примерно 1141 пар оснований из плазмиды p22E7.21.Hole.Hg-, кодирующий промотор щелочной фосфатазы, STII сигнальную последовательность и легкую цепь химерного антитела 22E7. Третий фрагмент представляет собой фрагмент PacI-MluI, включающий примерно 503 пары оснований из плазмиды p1H1.22.Hg-, кодирующий терминатор транскрипции λt0 для легкой цепи и сигнальную последовательность для легкой цепи STII (с относительной силой TIR 2) для тяжелой цепи.
3.2 Экспрессия антитела -5А6 Knob и 22E7 Hole
Биспецифическое полноразмерное антитело создают в соответствии со стратегией создания «выступов во впадинах» («knobs into holes») для усиления гетеродимеризации при создании анти-FcγRIIB (5А6)/анти-FcεRI (22Е7) антитела. Мутации по типу «выступов во впадинах» в CH3 домене Fc-последовательности, как было показано ранее, существенно снижают образование гомодимеров (Merchant et al., Nature Biotechnology 16: 677-681 (1998)). Получают конструкции для компонента анти-FcγRIIB (p5A6.11.Knob) путем встраивания мутации по типу «выступа» («knob») (T366W) в Fc-участок и для компонента анти-FcεRI (p22E7.11.Hole) путем встраивания мутаций по типу «впадина» («hole») (T366S, L368A, Y407V) (Merchant et al., 1998, выше).
Синтезируют небольшую партию антител с использованием плазмид p5A6.11.Knob для получения knob анти-FcγRIIB мономерного антитела (с мутацией по типу выступа) и p22E7.11.Hole для hole анти-FcεRI мономерного антитела (с мутацией по типу впадина). Каждая плазмида обладает относительной силой TIR 1 как для легкой, так и для тяжелой цепей. Для экспрессии данной конструкции в рамках небольшой партии штамм E. coli 33D3 (W3110 ∆fhuA (∆tonA) ptr3 lac Iq lacL8 ∆ompT ∆(nmpc-fepE) degP41 kanR) используют в качестве клеток-хозяев. После проведения трансформации полученные трансформанты отбирают и инокулируют в 5-мл среду Luria-Bertani с добавкой карбенициллина (50 мкг/мл) и растят при 30°С на качалке для культуры тканей в течение ночи. Далее каждую культуру разбавляют (1:100) в лимитированных по фосфату средах C.R.A.P. (Simmons et al., J. Immunol. Methods, 263:133-147, 2002). Затем к индуцируемой культуре добавляют карбенициллин в концентрации 50 мкг/мл и культуру растят примерно в течение 24 часов при 30°С на качалке. Если особо не указано иное, индукцию культуры проводят в качалочных колбах объемом 5 мл.
Невосстановленные лизаты целых клеток из индуцированных культур получают следующим образом: (1) 1 мл образцов индуцированной культуры с OD600 центрифугируют в микроцентрифужной пробирке; (2) каждый полученный осадок ресуспендируют в 90 мкл ТЕ (10 мМ Трис, pH 7,6, 1 мМ ЭДТА); (3) 10 мкл 100 мМ йодуксусной кислоты (Sigma I-2512) добавляют к каждому образцу для блокирования свободных цистеинов и для предотвращения перегруппировки дисульфидных связей; (4) к каждому образцу добавляют 20 мкл 10% SDS. Образцы подвергают обработке в вихревом смесителе, нагревают до примерно 90°С в течение 3 минут и затем снова перемешивают в вихревом смесителе. После охлаждения образцов до комнатной температуры добавляют 750 мкл ацетона для осаждения белка. Образцы перемешивают в вихревом смесителе и оставляют при комнатной температуре примерно на 15 минут. После центрифугирования в течение 5 минут в микроцентрифуге супернатант из каждого образца отбирают отсасыванием и белковые осадки ресуспендируют в 50 мкл dH2O плюс 50 мкл 2× NOVEX SDS буфера для образцов. Затем образцы нагревают в течение четырех минут при 90°С, перемешивают в вихревом смесителе и оставляют охлаждаться до комнатной температуры. Проводят окончательное пятиминутное центрифугирование и супернатанты переносят в чистые пробирки.
Восстановленные лизаты целых клеток из индуцированных культур получают следующим образом: (1) 1 мл образцов индуцированной культуры с OD600 центрифугируют в микроцентрифужной пробирке; (2) каждый полученный осадок ресуспендируют в 90 мкл ТЕ (10 мМ Трис, pH 7,6, 1 мМ ЭДТА); (3) к каждому образцу для восстановления дисульфидных связей добавляют 10 мкл 1 М дитиотрейтола (Sigma D-5545); (4) к каждому образцу добавляют 20 мкл 10% SDS. Образцы подвергают обработке в вихревом смесителе, нагревают до примерно 90°С в течение 3 минут и затем снова перемешивают в вихревом смесителе. После охлаждения образцов до комнатной температуры добавляют 750 мкл ацетона для осаждения белка. Образцы перемешивают в вихревом смесителе и оставляют при комнатной температуре примерно на 15 минут. После центрифугирования в течение 5 минут в микроцентрифуге супернатант из каждого образца отбирают отсасыванием, и белковые осадки ресуспендируют в 10 мкл 1 М дитиотрейтола, плюс 40 мкл dH2O, плюс 50 мкл 2× NOVEX SDS буфера для образца. Затем образцы нагревают в течение 4 минут при приблизительно 90°С, перемешивают в вихревом смесителе и оставляют охлаждаться до комнатной температуры. Проводят окончательное пятиминутное центрифугирование, и супернатанты переносят в чистые пробирки.
После получения образцов по 5-8 мкл каждого из них вносят в 10 ячеек, по 1,0 мм, в 12% Трис-глициновом SDS-PAGE (NOVEX) и проводят электрофорез при напряжении ~120 В в течение 1,5-2 часов. Полученные гели либо окрашивают кумасси синим, либо используют для вестерн-блоттинг-анализа.
При проведении вестерн-блоттинга SDS-PAGE гели переносят на нитроцеллюлозную мембрану (NOVEX) в 10 мМ буфере CAPS, pH 11, +3% метанола. Мембрану блокируют раствором 1× NET (150 мМ NaCl, 5 мМ ЭДТА, 50 мМ Трис, pH 7,4, 0,05% Triton Х-100) плюс 0,5% желатина и оставляют на приблизительно 30 минут - 1 час при комнатной температуре на качалке. По завершению стадии блокирования мембрану помещают в раствор 1× NET/0,5% желатина/анти-Fab антитело (фракция конъюгированного с пероксидазой козьего IgG против IgG Fab человека; CAPPEL #55223) для вестерн-блоттинга анти-Fab. Разбавляют анти-Fab антитело в диапазоне от 1:50000 до 1:1000000, в зависимости от лота антитела. Альтернативно, мембрану помещают в раствор 1× NET/0,5% желатина/анти-Fc антитело (фракция конъюгированного с пероксидазой козьего IgG против фрагмента Fc человека; BETHYL #A80-104P-41) для проведения вестерн-блот-анализа анти-Fc. Разбавляют анти-Fc антитело в диапазоне от 1:50000 до 1:250000, в зависимости от лота антитела. В каждом случае мембрану оставляют в антительном растворе антитела в течение ночи при комнатной температуре на качалке. На следующее утро мембрану промывают минимум в течение трех раз по 10 минут в 1× NET/0,5% желатина и затем 1 раз в течение 15 минут в TBS (20 мМ Трис, pH 7,5, 500 мМ NaCl). Визуализируют полосы белка, связанные анти-Fab антителом и анти-Fc антителом, с использованием набора для их обнаружения (Amersham Pharmacia Biotech ECL) с последующей экспозицией мембраны с рентгеновской пленкой.
На фиг.18 показаны результаты анти-Fab вестерн-блоттинга для оценки экспрессии p5A6.11.Knob (knob анти-FcγRIIB) и p22E7.11.Hole (hole анти-FcεRI) антитела. Они выявляют экспрессию полностью уложенного и собранного вида, включающего тяжелую-легкую (HL) цепи для knob анти-FcγRIIB антитела на линии 1 и для hole анти-FcεRI антитела на линии 2. Анти-Fab антитело характеризуется разной аффинностью для разных вариабельных доменов легкой цепи. Анти-Fab антитело в основном имеет сниженную аффинность для тяжелой цепи. В случае невосстановленных образцов, экспрессия каждого антитела приводит к выявлению вида, включающего тяжелую-легкую цепи. А именно: обнаруживается гомодимерный вид антитела полной длины для hole анти-FcεRI антитела, однако это лишь небольшая часть полностью уложенного и собранного вида антитела. Складчатость и сборка гомодимерного вида антитела полной длины скорее всего не является результатом включения мутации knob в анти-FcγRIIB антитело и мутаций по типу hole в анти-FcεRI антитела. В случае восстановленных образцов, легкая цепь обнаруживается для knob анти-FcγRIIB антитела и для hole анти-FcεRI антитела.
Аналогично, вестерн-блоттинг анти-Fc дает результаты, показанные на фиг.19, которые также выявляют экспрессию полностью уложенного и собранного вида, включающего тяжелую-легкую цепи (HL), для knob анти-FcγRIIB антитела в линии 1 и для hole анти-FcεRI антитела в линии 2. Анти-Fc антитело не способно связывать легкую цепь, так что легкая цепь не выявляется. В случае невосстановленных образцов экспрессия каждого антитела снова приводит к выявлению вида, включающего тяжелую-легкую цепи, но не гомодимерного вида антитела полной длины. В случае восстановленных образцов отмечаются аналогичные количества тяжелой цепи, выявляемые для knob анти-FcγRIIB антитела и для hole анти-FcεRI антитела.
3.3 Экспрессия антитела 5A6 Knob с вариантом по шарнирной области и антитела 22E7 Hole с вариантом по шарнирной области
Вид первичного антитела, получаемый при экспрессии конструкции p5A6.11.Knob и p22E7.11.Hole, представляет собой полностью уложенный и собранный вид, включающий тяжелую-легкую цепи (HL). Однако для упрощения методики получения приведенного в описании биспецифического анти-FcγRIIB/анти-FcεRI (5A6/22E7) антитела, последовательность шарнирной области двух тяжелых цепей модифицируют путем замещения двух цистеиновых остатков в шарнирной области сериновыми остатками (C226S, C229S, схема нумерации EU, соответствующая номенклатуре Kabat, E.A. et al., выше). Варианты по шарнирной области также называются как «бесшарнирные».
Получают плазмидные конструкции для knob анти-FcγRIIb (5A6) антитела и для hole анти-FcεRI (22E7) антитела, включающих варианты по шарнирной области с замещениями в C226S, C229S. Для каждого антитела получают две плазмидные конструкции. Одна конструкция характеризуется TIR силой 1 для легкой и тяжелой цепей, а вторая конструкции имеет относительную TIR силу 2, также и для легкой, и для тяжелой цепей.
Knob анти-FcγRIIB антитело (из плазмиды p5A6.11.Knob), hole анти-FcεRI антитело (p22E7.11.Hole), knob бесшарнирные анти-Fcγ-RIIb антитела
(p5A6.11.Knob.Hg- и p5A6.22.Knob.Hg-) и hole бесшарнирные анти-FcεRI антитела (p22E7.11.Hole.Hg- и p22E7.22.Hole.Hg-) далее подвергают экспрессии на основе соответствующих плазмид, как было описано выше. Получают лизаты целых клеток, проводят разделение в SDS-PAGE, переносят на нитроцеллюлозу и идентифицируют с помощью козьего противочеловеческого Fab-конъюгированного антитела и козьего противочеловеческого Fc-конъюгированного антитела, как было описано выше.
Результаты анти-Fab вестерн-блоттинга проиллюстрированы на фиг.20, которые указывают на существенное повышение уровня складчатости и сборки вида, включающего тяжелую-легкую (HL) цепи, для варианта knob бесшарнирного анти-FcγRIIB мономерного антитела (с относительной силой TIR 1 - для легкой цепи и 1 - для тяжелой цепи) в линии 2 и для варианта hole бесшарнирного анти-FcεRI мономерного антитела (с относительной силой TIR 1 - для легкой цепи и 1 - для тяжелой цепи) в линии 5. Кроме того, результаты анти-Fab вестерн-блоттинга обнаруживают повышение уровня складчатости и сборки вида, включающего тяжелую-легкую (HL) цепи, для мономерного HL knob бесшарнирного анти-FcγRIIB антитела (линия 3) и для мономерного HL hole бесшарнирного анти-FcεRI антитела (линия 6), при повышении относительной силы TIR для легкой цепи и тяжелой цепи от 1 до 2. Анти-Fab антитело имеет отличающуюся аффинность для разных вариабельных доменов легкой цепи и в основном характеризуется сниженной аффинностью для тяжелой цепи. В случае невосстановленных образцов экспрессия каждого антитела приводит к выявлению вида, включающего тяжелую-легкую цепи, но не вида антитела полной длины, возникающего в результате превращения цистеинов шарнирной области в серины. Отмечается существенное повышение уровня складчатости и сборки вида, включающего тяжелую-легкую (HL) цепи, для knob бесшарнирного анти-FcγRIIb антитела и hole бесшарнирного анти-FcεRI антитела, в том случае, когда два цистеина шарнирной области преобразуются в серины и когда относительная сила TIR для легкой цепи и тяжелой цепи также повышается от 1 до 2. В случае восстановленных образцов обнаруживаются и тяжелая цепь, и легкая цепь для разных анти-Fcγ-RIIb и анти-FcεRI антител. Повышение количества тяжелой и легкой цепей выявляется в том случае, когда относительная сила TIR возрастает от 1 до 2.
Аналогично, анти-Fc вестерн-блоттинг дает результаты, показанные на фиг.21, которые указывают на существенное повышение уровня складчатости и сборки мономерного вида, включающего тяжелую-легкую (HL) цепи, и для knob бесшарнирного анти-Fcγ-RIIB антитела, и для hole бесшарнирного анти-FcεRI антитела, в том случае, когда два цистеина в шарнирной области тяжелой цепи (HC) превращают в серины и когда относительная сила TIR для легкой цепи и тяжелой цепи возрастает от 1 до 2. Анти-Fc антитело не способно связываться с легкой цепью, так что легкая цепь не выявляется. В случае восстановленных образцов тяжелая цепь выявляется для разных анти-FcγRIIB и анти-FcεRI антител. Повышение количества тяжелых цепей выявляется в том случае, когда относительная сила TIR возрастает от 1 до 2.
3.4 Очистка компонентов биспецифических антител
Простота и эффективность получения очищенных и функциональных биспецифических антител оценивается далее применительно к антителам, содержащим вариантный шарнирный участок, описанный выше.
1. Экстракция из пасты E. coli
Замороженную пасту E. coli оттаивают и суспендируют в 5 объемах (объем/вес) дистиллированной воды, доводят pH до 5 с помощью HCl, центрифугируют и отбрасывают супернатант. Нерастворимый остаток ресуспендируют в 5-10 объемах буфера, pH 9, с использованием политрона (Brinkman) и супернатант оставляют после центрифугирования. Данную стадию повторяют еще один раз.
Нерастворимый остаток далее ресуспендируют в 5-10 объемах того же буфера и клетки разрушают пропусканием через микрофлюидизатор (Microfluidics). Супернатант оставляют после центрифугирования.
Супернатанты анализируют при проведении электрофореза в SDS-полиакриламидном геле (SDS-PAGE) и по методу вестерн-блоттинга, и собирают фракции, содержащие антитело с одним плечом (то есть полосу, соответствующую молекулярному весу одной тяжелой цепи плюс легкая цепь).
2. Аффинная хроматография с белком А
В объединенных супернатантах корректируют значение рН до 8 и добавляют шарики ProSepTM-А (Millipore) (примерно 250 мл шариков на 10 литров). Смесь перемешивают в течение 24-72 часов при 4°С, шарикам дают осесть и супернатант сливают. Шарики переносят на хроматографическую колонку (Amersham Biosciences XK50TM) и промывают 10 мМ трис-буфером, pH 7,5. Затем проводят элюцию с колонки в градиенте pH в буфере, содержащем 50 мМ цитрата и 0,1 М NaCl. Исходный буфер доводят до pH 6 и получают градиент путем линейного разбавления буфером с pH 2.
Фракции доводят до pH 5 и до 2 М уровня мочевины при добавлении 8 М мочевины и трис-основания, после чего анализируют в SDS-PAGE и объединяют.
3. Катионообменная хроматография
Колонку с S-сефарозой (S-Sepharose Fast FlowTM) (Amersham Biosciences) уравновешивают 25 мМ MES, pH 5,5, с 2 М мочевиной. Объединенный элюат ProSepTM-А разбавляют равным объемом уравновешивающего буфера и вносят на колонку. После промывки вначале уравновешивающим буфером и затем 25 мМ MES, pH 5,5 через колонку пропускают линейный градиент 0-1 М NaCl в 25 мМ MES, pH 5,5. Фракции объединяют с учетом результатов анализа в SDS-PAGE.
4. Хроматография, основанная на гидрофобном взаимодействии
Колонку HI-PropylTM (J.T. Baker) уравновешивают 25 мМ MES, pH 6, с 0,5 М сульфатом натрия. Элюат S-FastFlowTM доводят до уровня 0,5 М сульфата натрия, pH 6 вносят на колонку и пропускают через колонку градиент 0,5-0 M сульфата натрия в 25 мМ MES, pH 6. Фракции объединяют, с учетом результатов анализа в SDS-PAGE.
5. Гель-хроматография
Объединенный элюат HI-PropylTM концентрируют с использованием концентратора CentriPrerTM YM10 (Amicon) и вносят на колонку SuperdexTM SX200 (Amersham Biosciences), уравновешенную 10 мМ сукцинатом или 10 мМ гистидином в 0,1 М NaCl, pH 6, и колонку элюируют со скоростью 2,5 мл/мин. Фракции объединяют, с учетом результатов анализа в SDS-PAGE.
3.5 Отжиг компонентов антитела для получения биспецифических антител
Ниже описаны два аналогичных (но не идентичных) метода отжига, и оба приводят к хорошим выходам биспецифических антител. Тяжелая цепь антител и компоненты антител, описанные ниже, включают варианты по шарнирной области, описанные выше.
Вариант 5A6Knob по шарнирной области и вариант 22E7Hole по шарнирной области, получаемые отжигом по методу 1
Очищенные мономерные антитела 5A6Knob и 22E7Hole, содержащие тяжелую цепь/легкую цепь, в 25 мМ MES, pH 5,5, 0,5 М NaCl, смешивают в равных молярных соотношениях, определяемых соответствующими концентрациями. Далее смесь нагревают при 50°С в течение 5 минут - 1 часа. Температуру отжига устанавливают по результатам анализа кривых плавления, описанных ранее для данных вариантов CH3 (Atwell, S., et al., 1997, J. Mol. Biol., 270: 26-35). Полученные при отжиге антитела далее анализируют для определения их биспецифичности.
Анализ биспецифичности
1) Изоэлектрофокусирование
В полученных при отжиге антителах подтверждают их биспецифичность путем изоэлектрофокусирования образцов. Антитело 5A6Knob имеет pI 7,13, тогда как антитело 22E7Hole имеет pI 9,14. Биспецифическое антитело 5A6Knob/22E7Hole имеет значение pI 8,67. На фиг.22 показано движение 5A6Knob, 22E7Hole и биспецифического 5A6Knob/22E7Hole (до и после нагревания) антител в геле для изоэлектрофокусирования (Invitrogen, Novex pH 3-10 IEF) после окрашивания кумасси синим. Хотя и происходит некоторый отжиг при смешивании в условиях комнатной температуры, нагревание до 50°С, по всей видимости, ускоряет завершение процесса. Появление новой белковой полосы со значением pI в интервале значений для 5A6Knob и 22E7Hole подтверждает образование биспецифического антитела.
2) Анализ на аффинной колонке
Поведение 5A6Knob, 22E7Hole и биспецифического 5A6Knob/22E7Hole антител оценивают на аффинных колонках с FcγRIIB. Белок слияния FcγRIIB человека (внеклеточный домен)-GST связывают с твердой подложкой в маленькой колонке, в соответствии с инструкциями производителя (Pierce, набор для иммобилизации UltraLinkTM № 46500). 5A6Knob, 22E7Hole и биспецифическое 5A6Knob/22E7Hole антитела в PBS (137 мМ NaCl, 2,7 мМ KCl, 8 мМ Na2HPO4, 1,5 мМ KH2PO4, pH 7,2) вносят на три отдельные аффинные колонки с FcγRIIB примерно в количестве, составляющем 10-20% от теоретической связывающей способности каждой колонки. Затем колонки промывают с использованием 16 колоночных объемов PBS. Собирают протекающие фракции после загрузки и промывки колонки, объединяют и концентрируют примерно в 10 раз на микроконцентраторе CentriconTM (Amicon). Затем каждый концентрат в одном и том же буфере разбавляют в 2 раза буфером для образца 2× SDS и анализируют в SDS-PAGE (Invitrogen, Novex Трис-глицин). Белковую полосу далее переносят на нитроцеллюлозу путем электроблоттинга в 20 мМ Na2HPO4, pH 6,5, и проводят зондирование противочеловеческим IgG-Fab антителом, конъюгированным с пероксидазой (CAPPELL#55223). Полосы антител выявляют затем с использованием набора от Amersham Pharmacia Biotech ECLTM в соответствии с инструкциями производителя.
Результаты данного анализа показаны на фиг.23. Аффинная колонка с FcγRIIB должна удерживать 5A6Knob антитело и биспецифическое антитело 5A6Knob/22E7Hole. Антитело 22E7Hole должно протекать через колонку, как показано на фиг.23. Отсутствие антитела в полосе, соответствующей биспецифическому 5A6Knob/22E7Hole антителу, указывает на биспецифичность.
Поведение 5A6Knob, 22E7Hole и биспецифического 5A6Knob/22E7Hole антител также можно проследить на колонках для аффинной хроматографии с FcεRI. Аффинная колонка со слитым IgE может быть получена и использована, как было описано выше, применительно к аффинным колонкам с FcγRIIB. Аффинная колонка с FcεRI должна удерживать 22E7Hole антитело и 5A6Knob/22E7Hole антитело. 5A6Knob антитело должно проходить через колонку. Отсутствие антитела в полосе, соответствующей 5A6Knob/22E7Hole антителу, указывает на биспецифичность.
Вариант 5A6Knob по шарнирной области и вариант 22E7Hole по шарнирной области, получаемые отжигом по методу 2
Проводят очистку компонентов антител (5A6Knob и 22E7Hole с одним плечом), как было описано выше.
«Гетеродимер» образуется в результате отжига при 50°С с использованием небольшого молярного избытка 5A6, который затем очищают на катионообменной колонке.
5 мг 5A6(Knob) и 4,5 мг 22E7(Hole) H/L мономерных антител объединяют в общем объеме 10 мл, содержащем 8 мМ сукцината, 80 мМ NaCl буфера, и доводят до уровня 20 мМ трис, pH 7,5.
Мономерные антитела подвергают отжигу при нагревании смеси до 50°С в водяной бане в течение 10 минут, после чего охлаждают до 4°С с получением биспецифического антитела.
Анализ биспецифичности
1. Изоэлектрофокусирование
Анализ в геле для изоэлектрофокусирования (Cambrex, pH 7-11) указывает на образование одной полосы со значением pI ~ 8,5, в смеси для отжига, что соответствует наличию биспецифического антитела (которое имеет расчетное значение pI 8,67). См. фиг.24.
2. Очистка на катионообменной колонке
Колонку CM-FastFlow объемом 5 мл (HiTrap, Amersham Biosciences) уравновешивают буфером с pH 5,5 (30 мМ MES, 20 мМ hepes, 20 мМ имидазола, 20 мМ трис, 25 мМ NaCl). Объединенные продукты отжига разбавляют равным объемом уравновешивающего буфера и доводят рН до значения 5,5, наносят на колонку и промывают колонку буфером для уравновешивания. Через колонку пропускают тот же буфер со скоростью 1 мл/мин в градиенте pH от 5,5 до 9,0 в течение 30 минут.
Фракции анализируют по методу IEF, результаты которого указывают на то, что 5A6 элюируется раньше гетеродимера. Анализ светорассеяния объединенных фракций, содержащих гетеродимер, не выявляет мономера.
ПРИМЕР 4.0 Характеристика биспецифического антитела 5А6/22E7 Knob-Hole
Цель данного примера продемонстрировать тот факт, что 5А6/22Е7, а не 5А6 и 22Е7 по одиночке представляет собой биспецифическое антитело. 5А6/22Е7 имеет двойную связывающую специфичность для FcγRIIB-His6-GST и FcεRI-ECD-Fc человека по данным теста ELISA в модификации «сэндвич». Результаты представлены на фиг.29 и 30. 5А6(а) и 5А6(b) обозначают два разных белковых препарата 5А6. Описанные ниже биспецифические антитела 5А6/22Е7 представляют собой гетеродимеры по типу «выступ во впадинах» (knobe in holes) с шарниром дикого типа или без шарнира. Биспецифическое антитело взаимозаменяемо обозначается как BsAb.
Двойная специфичность по связыванию 5А6/22Е7 бесшарнирного биспецифического антитела в отношении huFcγRIIB-His6-GST и huFcεRI-ECD-Fc (слияние по типу IgE-рецептор) была продемонстрирована по методу ELISA и результаты данного испытания представлены на фиг.29. На планшеты для ELISA наслаивают на ночь при 4°С 100 мкл раствора с концентрацией 1 мкг/мл FcγRIIB-His6-GST в PBS, pH 7,4. Планшеты промывают PBS и блокируют добавлением 1% казеинового блокатора в PBS. Ячейки промывают три раза PBS/0,05% Tween®. Готовят раствор CD4-IgG с концентрацией 10 мкг/мл в разбавляющем буфере для ELISA (50 мМ Трис-HCl, pH 7,5, 150 мМ NaCl, 0,05% Tween-20, 0,5% BSA, 2 мМ ЭДТА) и добавляют к ячейкам в количестве 100 мкл/ячейку для блокирования связывания FcγRIIB-His6-GST с Fc-частью каждого из исследуемых антител: 5А6(А)/22Е7 по типу «выступ во впадинах» (knob in holes) с шарнирной областью дикого типа, биспецифическое антитело; 5А6(B)/22Е7 по типу «выступ во впадинах» (knob in holes), с шарнирной областью дикого типа, BsAb; 5А6/22Е7 по типу «выступ во впадинах» (knob in holes), бесшарнирное BsAb; 5А6 MAb и 22Е7 MAb. После промывки планшетов три раза PBS/0,05% Tween® делают серийные разведения трех антител: 5А6/22Е7 BsAb, 5А6 MAb и 22Е7 MAb в разбавляющем буфере для ELISA и добавляют к ячейкам по 100 мкл/ячейку каждого из разведений. Планшеты инкубируют в течение 1 часа при комнатной температуре. После промывки планшетов три раза PBS/0,05% Tween® к каждой ячейке добавляют 100 мкл huFcεRI-ECD-Fc с концентрацией 1 мкг/мл и планшеты инкубируют в течение 1 часа при комнатной температуре. После промывки планшетов три раза PBS/0,05% Tween® к каждой ячейке добавляют 100 мкл IgE-биотина с концентрацией 1 мкг/мл и инкубируют в течение 1 часа при комнатной температуре. Планшет промывают PBS/0,05% Tween® и инкубируют 30 минут с добавлением по 100 мкл/ячейку стрептавидина-HRP (1:2000) в разбавляющем буфере для ELISA. После промывки PBS/0,05% Tween® планшет инкубируют в течение 5 минут со 100 мкл субстрата TMB. Реакцию гасят добавлением 100 мкл/ячейку останавливающего раствора и планшет анализируют в денситометре для 96-ячеечных планшетов при длине волны 630 нм (Molecular Devices). Результаты показывают, что IgE связывается в ячейках, содержащих биспецифические антитела 5А6/22Е7. Биспецифические антитела: 5А6(A) + 22Е7 BsAb, 5А6(B) + 22Е7 BsAb и 5А6 + 22Е7 бесшарнирное BsAb по типу «выступ во впадинах» (knob in holes) успешно связываются с FcγRIIB-GST и IgE-биотином. См. фиг.29.
Проводят эксперимент в рамках комплементарного ELISA по указанной ниже процедуре, результаты данного эксперимента представлены на фиг.30. На планшеты для ELISA наслаивают на ночь при 4°С 100 мкл 1 мкг/мл раствора huFcεRI-ECD-Fc в PBS, pH 7,4. Планшет промывают PBS и блокируют добавлением 1% казеинового блокатора в PBS. Ячейки промывают три раза PBS/0,05% Tween®. Разбавляющим буфером для ELISA делают серийные разведения 5А6/22Е7 биспецифических антител, 5А6 антитела или 22Е7 антитела и каждое из разведений добавляют к ячейкам в количестве 100 мкл/ячейку. Планшеты инкубируют в течение 1 часа при комнатной температуре. После промывки планшетов три раза PBS/0,05% Tween® к каждой ячейке добавляют FcγRIIB-His6-GST в количестве 100 мкл с концентрацией 1 мкг/мл, в присутствии 10 мкг/мл CD4-IgG для блокирования связывания FcγRIIB-His6-GST с Fc-частью исследуемого антитела, huFcεRIIB-ECD-Fc и вторичное антитело (анти-GST-биотин) и инкубируют в течение 1 часа при комнатной температуре. После промывки планшетов три раза PBS/0,05% Tween® к каждой ячейке добавляют по 100 мкл анти-GST-биотина в концентрации 1 мкг/мл и инкубируют в течение 1 часа при комнатной температуре. Планшет промывают PBS/0,05% Tween® и инкубируют в течение 30 минут со стрептавидином-HRP (1:2000) в количестве 100 мкл/ячейку в разбавляющем буфере для ELISA. После промывки PBS/0,05% Tween® планшет инкубируют в течение 5 минут со 100 мкл субстрата TMB. Реакцию гасят добавлением 100 мкл/ячейку останавливающего буфера и планшет анализируют при длине волны 630 нм. Результаты показывают, что анти-GST-биотин связывается в ячейках, содержащих биспецифические антитела 5А6/22Е7. Биспецифические антитела: 5А6(А) + 22Е7 и 5А6(В) + 22Е7, бесшарнирные, а также 5А6(В) + 22Е7, бесширнирные, и биспецифические антитела 5А6/22Е7, сконструированные по типу «выступ во впадинах» (knob in holes), успешно связываются с huFcεRI-ECD-Fc и FcγRIIB-GST. См. фиг.30.
На фиг.29 и 30 показаны графики кривых, описывающих оба эксперимента. Успешное связывание с FcγRIIB-GST и huFcεRI-ECD-Fc было продемонстрировано только для 5А6(А) + 22Е7 и для 5А6(В) + 22Е7 бесширнирных биспецифических антител. В таблице 1 приведены значения IC50, которые также показаны на фиг.29 и 30.
5А6(А)+22Е7: 55,2
5А6(В)+22Е7: 76,0
MAb
5А6(А): 3,3е+06
5А6(В): 1,4е+07
22Е7: 1,0е+05
BsAb «выступ во впадинах» (knob in holes), бесшарнирное
5А6+22Е7, бесшарнирное «выступ во впадинах»: 23
5А6(А)+22Е7: 490
5А6(В)+22Е7: 291,5
MAb
5А6(А): 5,3е+06
5А6(В): 1,0е+07
22Е7: 2,8е+06
BsAb «выступ во впадинах» (knob in holes), бесшарнирное
5А6+22Е7, бесшарнирное «выступ во впадинах»: 76,5
ПРИМЕР 5.0 Свойства бесшарнирного биспецифического антитела 5A6/22E7 по типу «выступ во впадинах» (Knob in Holes)
5.1 Материалы
В предыдущих примерах FcγRIIB рассматривался применительно к huFcγRIIB1, представляющему один из трех сплайсинговых FcγRIIB человека. В оставшихся примерах использованы и соответствующим образом обозначены FcγRIIB1 и дополнительный сплайсинговый вариант FcγRIIB2.
JW8.5.13 представляет собой химерное антитело, состоящее из вариабельного участка мыши, специфичного для NP (нитрофенол, антиген) и Fc-участка IgE человека. Вариабельный участок JW8.5.13 IgE специфичен для NP и не вступает в перекрестную реакцию с TNP. Взятая от человека IgE часть JW8.5.13 связывается специфически с huFcεRI и не связывается с эндогенным FcεRI крысы в клеточных линиях, полученных из RBL. Связывание JW8.5.13 с huFcεRI оказывает позитивную регуляцию его экспрессии и нагружает его антиген-специфическим IgE.
Клетки RBL-2Н3 (ATCC# CRL-2256), экспрессирующие FcεRI, α-субъединицу высокоаффинного IgE рецептора человека (FcεRI) (Gilfillan et al., (1995) Int Arch Allergy Immunol. 107(1-3): 66-68), трансфицируют сочетаниями (то есть при наличии или в отсутствие) huFcεRI и/или huFcγRIIB для получения клеточных линий, производных от RBL. Варианты клеточной линии RBL 2H3 получают путем трансдукции ретровирусом RBL 2H3 клеток FcγRIIB1 человека или FcγRIIB2 с использованием ретровирусных векторов экспрессии, получаемых из Вашинтонского Университета (Washington University, MO), которые аналогичны pQCXIR (Retro-X Q векторы), доступным от BD-Clontech. кДНК генов полной длины человека субклонируют в ретровирусном векторе по одиночке или в сочетании с IRES (Internal Ribosomal Entry Sequence (внутренняя рибосомальная входная последовательность)) для бисцистронной совместной трансфекции и совместной экспрессии двух генов. Ниже приводится подробное описание метода ретровирусной трансдукции.
PG13 упакованные клетки (ATCC CRL-10686) высевают на культуральные чашки для среды диаметром 10 см с плотностью 2×106 клеток на чашку (среда DMEM с высоким содержанием глюкозы, 10% FCS, пенициллин, стрептомицин, 2 мМ L-глютамин) на 24 часа. Клетки трансфицируют конструкциями ДНК pMSCV с использованием FuGENE 6 и затем культивируют в течение двух дней при 37°С в атмосфере 5% CO2. Супернатант клеточной культуры, содержащий ретровирусные частицы, собирают и фильтруют через фильтр с размером 0,4 микрона. Добавляют стерильный протаминсульфат до конечной концентрации 10 мкг/мл и используют 4 мл супернатанта для инфицирования примерно 1×106 клеток RBL путем спиновой инфекции при 32°С в течение 90 минут с последующим продолжением культивирования с ретровирусным супернатантом в течение 3-4 часов при 37°С в атмосфере 5% CO2. Инфицированные клетки RBL собирают, переносят в среду RBL и размножат для последующей сортировки. Позитивно трансфицированные клетки идентифицируют по методу, включающему флуоресцентную сортировку (FACS), с использованием 22E7 и/или 5A6 для антител для выявления FcεRIA человека и FcγRIIB человека соответственно.
Полученные клеточные линии обозначают следующим образом: клетки RBL huFcεRI с экспрессированным на поверхности FcεRIα человека; клетки RBL huFcγRIIB с экспрессированным на поверхности FcγRIIB1 человека, клетки RBL huFcεRI+huFcγRIIB1 с экспрессированным на поверхности FcεRIα человека и FcγRIIB1 человека; и клетки RBL huFcεRI+huFcγRIIB2 с экспрессированным на поверхности FcεRIα человека и FcγRIIB2 человека.
Биотинилированное биспецифическое антитело 5А6/22Е7, сконструированное по принципу «выступ во впадинах» (knob in holes, бесшарнирное), получают при объединении 20× молярный избыток EZ-linkTM NHS-PEO4-биотин (Pierce, Rockford, IL) с биспецифическим антителом в PBS.
Внеклеточный домен huFcεRIα (huFcεRIα ECD) получают при субклонировании в системе экспрессии бакуловируса и очищают на колонке со сшитой CNBr-сефарозой и на колонке для гель-фильтрации на сефадексе. Внеклеточный домен huFcγRIIB (huFcγRIIB ECD) получают при субклонировании в рамке считывания с С-концевым фрагментом His6 с последующей экспрессией в системе экспрессии бакуловируса. huFcγRIIB ECD очищают на смоле NiNTA.
5.2 Тест на высвобождение гистамина
Способность биспецифического антитела 5А6/22Е7 сшивать huFcγRIIB1 или huFcγRIIB2 с huFcεRI на поверхности клеток была показана при селективном блокировании высвобождения гистамина в рамках следующего теста. Приведенное ниже описание дополнительно иллюстрируется фиг.31-33.
Трансфицированные клетки RBL 48 (показанные выше) выращивают (EMEM (минимальная эссенциальная среда Игла с добавкой BSS Эрла) с содержанием 2 мМ L-глютамина, 1 мМ пирувата натрия, 0,1 мМ неэссенциальных аминокислот, 1,5 г/л бикарбоната натрия, пенициллина, стрептомицина, 15% бычьей сыворотки теленка в стандартных колбах для культуры ткани при 37°С в увлажненной атмосфере c 5% содержанием CO2 в инкубаторе. Клетки собирают и обрабатывают 4 мл раствора PBS/0,05% трипсина/0,53 мМ ЭДТА в течение 2 минут при 37°С, после чего центрифугируют (400×g, 10 минут) и ресуспендируют в свежей среде EMEM. Клетки в суспензии просчитывают на гемоцитометре (Reichert-Jung) и плотность корректируют до уровня примерно 105-106 клеток/мл.
Описанные выше трансфицированные клетки RBL, клетки RBL huFcεRI, клетки RBL huFcεRI+huFcγRIIB1 и клетки RBL huFcεRI+FcγRIIB2 высевают в 96-ячеечный планшет для культуры ткани с плоским дном с плотностью 105 клеток/ячейку в 200 мкл среды EMEM. Клетки инкубируют в течение 24 часов при 37°С с добавлением или без добавления 1 мкг/мл JW8.5.13 («NP-специфичный IgE человека»). Далее клетки промывают три раза свежей средой для удаления несвязанного NP-специфичного IgE человека. Некоторые клетки обрабатывают 1-5 мкг/мл биспецифического антитела в условиях насыщения и инкубируют в течение 1 часа при 37°С до активации антигеном.
Клетки инкубируют с овальбумином, конъюгированным с нитрофенолом (NP) (NP (11)-OVA), антигеном, который связывает JW8.5.13, IgE, или TNP(11)-OVA, нерелевантным антигеном, в течение 1 часа при 37°С. Дегрануляцию, связанную с активацией (высвобождение гистамина) клеток RBL huFcεRI, RBL huFcεRI+huFcγRIIB1 и клеток RBL huFcεRI+huFcγRIIB2, при наличии или в отсутствие биспецифического антитела, тестируют по NP-(11)-OVA и TNP в диапазоне концентраций антигена от 0,0001 до 10 мкг/мл. После инкубации измеряют уровень гистамина в клеточных супернатантах (культуральная среда от клеток) методом ELISA, как описано выше. Общий уровень гистамина в клетках, который служит в качестве позитивных контролей, независимых от активации, также получают либо путем лизирования клеток Triton Х-100, либо путем запуска общего высвобождения гистамина через стимуляцию иономицином. Определяют также фоновый уровень высвобождения гистамина клетками RBL. Уровни высвобождения гистамина определяют количественно по процедуре ELISA с использованием гистаминового набора для ELISA (KMI, Diagnostics Minneapolis, MN).
Результаты теста на высвобождение гистамина представлены на фиг.31-33. Ожидается, что высвобождение гистамина будет увеличиваться в присутствии hIgE (JW8.5.13) и NP(11)-OVA антигена («NP»), если не будет происходить специфического ингибирования. На фиг.31 приведены данные по уровню высвобождения гистамина в клетках RBL huFcεRI при использовании варьирующих концентраций TNP или NP (11)-OVA. В клетках RBL huFcεRI высвобождение гистамина запускается NP и hIgE. Как и ожидалось, биспецифическое антитело не воздействует (не подавляет, или не ингибурует) высвобождение гистамина в отсутствие huFcγRIIB (см. вариант «+hIgE+NP+биспецифическое антитело», темно-серые колонки в дальнем правом углу каждого образца на фиг.31, график А).
На фиг.32 представлены данные по высвобождению гистамина в клетках RBL huFcεRI+huFcγRIIB1 и на фиг.33 представлены данные по высвобождению гистамина в клетках RBL huFcεRI+FcγRIIB2. В клетках RBL huFcεRI+huFcγRIIB1 и RBL huFcεRI+FcγRIIB2 биспецифическое антитело ингибирует высвобождение гистамина (показаны для целей сравнения: «+hIgE+NP» светло-серые прямоугольники и темно-серые прямоугольники «+hIgE+NP+биспецифическое антитело» на графике А, фиг.32 и на графике А, фиг.33).
Активация высвобождения гистамина в клеточных линиях RBL специфична для антигена и происходит зависимым от дозы образом через IgE человека, связанный с FcεRI человека. Клетки не активируются в отсутствие IgE человека, а также не активируются при стимуляции нерелевантным антигеном (например, TNP). Добавление биспецифического антитела 5А6/22Е7 ингибирует высвобождение гистамина (до фонового уровня) в клетках RBL huFcεRI+huFcγRIIB1 и RBL huFcεRI+huFcγRIIB2, но не в клетках RBL huFcεRI, указывая на то, что присутствие FcγRIIB необходимо для осуществления ингибирующей функции. Аналогичные результаты отмечаются для huFcγRIIB1 и huFcγRIIB2 в присутствии huFcεRI.
Биспецифическое антитело по настоящему изобретению также ингибирует анти-IgE-индуцированное высвобождение гистамина в первичных базофилах человека. Первичные базофилы выделяют от шести нормальных доноров людей, от которых было получено информированное согласие. Образец крови человека обогащают базофилами с использованием процедуры осаждения декстраном. Вкратце, процедура состоит в том, что каждые 40 мл донорской крови, подлежащей осаждению, смешивают в конической пробирке на 50 мл, содержащей 375 мг декстрозы, 5,0 мл 0,1 М ЭДТА и 12,5 мл 6% декстрана клинической чистоты. Смесь разделяют на две конических пробирки по 50 мл и добавляют к каждой пробирке по 20 мл крови. Кровь оставляют для осаждения на 60-90 минут, после чего плазменный слой отбирают и центрифугируют при 110×g в течение 8 минут при 4°С и оставляют осевшие клетки, которые затем ресуспендируют, промывают PAG (декстроза, 1 г/л: 1× PIPES, pH 7,3: 0,003% сывороточный альбумин человека) и ресуспендируют в PAG. Клетки стимулируют анти-IgE антителом либо в виде обогащенного декстраном препарата, либо после очистки в процедуре разделения с использованием магнитных шариков Miltenyi (Miltenyi Biotec, Auburn, CA; см., например, Kepley С. et al., J. Allergy Clin. Immunol. 102: 304-315 (1998)) в ходе инкубации при 37°С в течение одного часа с последующим центрифугированием для осаждения клеток. Супернатант оставляют для анализа. Базофилы могут быть выделены стандартными процедурами, таким как описаны Kepley, С. L. et al., J. Allergy Clin. Immunol. 106(2): 337-348 (2000). Препарат, обогащенный базофилами, может быть далее очищен в процедуре разделения с использованием магнитных шариков Miltenyi (Miltenyi Biotec, Auburn, CA; см., например Kepley, С. et al., J. Allergy Clin. Immunol. 102: 304-315 (1998)), и/или путем сортировки по методу проточной цитометрии (Kepley, С. et al., (1994) выше). Козий противочеловеческий IgE получают от Caltag Laboratories, Burlingame, CA, США. Выделенные базофилы, осуществляющие совместную экспрессию huFcγRIIB и huFcεRI, инкубируют с анти-IgE (козий противочеловеческий IgE (Caltag Laboratories)) или при добавлении биспецифического антитела 5А6/22Е7 в течение одного часа при 37°С. Для стимуляции базофилов используют козий анти-IgE в разведении 1:100 (по объему) в присутствии биспецифического антитела 5А6/22Е7 в диапазоне от 0 до 20000 нг/мл в исследуемом растворе. Высвобождение гистамина оценивают по описанной выше методике. Диаграмма на фиг.62 показывает, что высвобождение гистамина индуцируется в присутствии противочеловеческого IgE. Добавление биспецифического антитела 5А6/22Е7 ингибирует высвобождение гистамина в режиме, который достаточно приближенно можно назвать зависимым от дозы. В отсутствие любого антитела или в присутствии одного биспецифического антитела 5А6/22Е7 отмечается ограниченное фоновое высвобождение гистамина. На основе анализа базофильных образцов от шести человек - нормальных доноров крови, получено среднее значение показателя ингибирования высвобождения гистамина под действием биспецифического антитела 5А6/22Е7, равное 67±9%. Ранее сообщалось, что среднее значение высвобождения гистамина из базофилов у пациентов Xolair® составляет примерно 50% через 90 дней (MacGlashan, D. W. et al., J. Immunol. 158: 1438-1445 (1997)) на основании данных по отрицательной регуляции экспрессии FcεRI. Эти результаты показывают, что анти-huFcγRIIB/анти-huFcεRI биспецифическое антитело может быть полезно в качестве терапевтической молекулы для быстрого ингибирования иммунной реакции (такой как высвобождение гистаминов в базофилах) у пациента-человека, путем ингибирования активности FcεRI через сшивку с FcγRIIB. Анти-huFcγRIIB/анти-huFcεRI биспецифическое антитело также полезно для целей применения в комбинированой терапии при сочетании с анти-IgE антителом. При использовании комбинированной терапии анти-huFcγRIIB/анти-huFcεRI биспецифическое антитело действует в направлении быстрого ингибирования высвобождения гистамина путем сшивания с FcγRIIB с последующей отрицательной регуляцией экспрессии FcεRI анти-IgE антителом (таким как Xolair® анти-IgE антитело, Genentech, Inc.).
5.3 Сшивание huFcεRI и huFcγRIIB биспецифическим антителом
Данный пример приведен с целью продемонстрировать зависимость ингибирования гистамина и совместного сшивания FcεRI человека и FcγRIIB человека на поверхности клеток биспецифическим антителом 5А6/22Е7. Метод тестирования описан ниже, а его результаты показаны на фиг.34-41.
Клетки RBL huFcεRI+huFcγRIIB1 и RBL huFcεRI+huFcγRIIB2 инкубируют в течение 24 часов при 37°С с 5 мкг/мл NP-специфического IgE человека и затем промывают три раза свежей средой EMEM для удаления несвязанного NP-специфичного IgE человека. Перед добавлением к клеткам RBL биспецифическое антитело 5А6/22Е7 подвергают предварительной инкубации в течение 30 минут с очищенным huFcεRIα ECD и huFcγRIIB ECD в разных молярных соотношениях. После предварительной инкубации биспецифическое антитело 5А6/22Е7 добавляют к клеточной культуре RBL до достижения конечной концентрации 5 мкг/мл биспецифического антитела 5А6/22Е7 и далее инкубируют в течение одного часа при 37°С. Клетки активируют путем инкубации с NP-конъюгированным овальбумином в течение 1 часа при 37°С. Дегрануляцию, связанную с активацией, измеряют путем количественного определения уровня высвобождение гистамина в клеточной культуральной среде с использованием процедур ELISA, в основном описанных выше. Зависимость ингибирования высвобождения гистамина от сшивки FcεRI человека и FcγRIIB человека биспецифическим антителом по настоящему изобретению, продемонстрировано на фиг.34 (для клеток RBL huFcεRI+huFcγRIIB1) и на фиг.36 (для клеток RBL huFcεRI+huFcγRIIB2).
Связывание биспецифического антитела с клетками RBL-линии также оценивают в присутствии huFcεRIα ECD и huFcγRIIB ECD с использованием проточной цитометрии. Клетки и материалы для данной процедуры были описаны выше. Клетки собирают и сортируют на аликвоты, содержащие 105-106 клеток. Далее клетки промывают и ресуспендируют в FACS буфере (PBS с 2% FCS). Клетки промывают второй раз и суспендируют в FACS буфере с добавкой 10% крысиной сыворотки, 2 мкг/мл IgG человека и 1 мкг/мл биотинилированного биспецифического антитела. Клетки инкубируют в течение 30 минут на льду, промывают и ресуспендируют в FACS буфере со стрептавидином-PE. После инкубации в течение 30 минут на льду смесь промывают холодным FACS буфером, откручивают и ресуспендируют в FACS буфере с 0,1% йодидом пропидия. Образцы анализируют методом проточной цитометрии и результаты выражают в виде относительных единиц флуоресценции (RFU). Результаты указанных тестов на связывание показаны на фиг.35 и 37-41, где указаны соотношения ECD к биспецифическому антителу. На фиг.35 и 37 показаны графики, иллюстрирующие результаты исследования методом проточной цитометрии связывания биспецифического антитела 5А6/22Е7 с клетками RBL huFcεRI + huFcγRIIB1 (фиг.35) или с клетками RBL huFcεRI + huFcγRIIB2 (фиг.37) в присутствии huFcεRI ECD и huFcγRIIB ECD. Как и ожидалось, более высокие соотношения ECD к биспецифическому антителу приводят к снижению связывания биспецифического антитела с клетками. Результаты представлены в сравнительном аспекте: светлый пик (клетки, связанные BsAb в присутствии ECD) против темного пика (позитивный контроль - клетки, связанные BsAb в отсутствие ECD).
На фиг.38-41 проиллюстрировано использование метода проточной цитометрии для анализа уровня связывания биспецифического антитела 5А6/22Е7 с различными клетками, полученными из RBL, в присутствии huFcεRI ECD, huFcγRIIB ECD или их обоих: huFcεRI ECD и huFcγRIIB ECD. На фиг.38-41 темный пик относится к связыванию рецептора на клеточной поверхности с 5А6/22Е7 в присутствии ECD. Результаты показаны на сравнительном аспекте: светло-серый пик (клетки, не связанные BsAb) и темно-серый пик (клетки, связанные BsAb в отсутствие ECD). Как и ожидалось, связывание 5А6/22Е7 с клетками RBL huFcεRI (см. фиг.38) блокируется возрастающими концентрациями huFcεRI ECD, но не huFcγRIIB ECD, при этом блокирование обоих ECD дает результаты, аналогичные для huFcεRI ECD. Связывание 5А6/22Е7 с клетками RBL huFcγRIIB (см. фиг.39) не зависит от huFcεRI ECD, но блокируется huFcγRIIB ECD. Аналогичные результаты по связыванию отмечаются в клетках RBL huFcεRI+huFcγRIIB1 (см. фиг.40) и в клетках RBL huFcεRI+huFcγRIIB2 (см. фиг.41). Как и ожидалось, уровень связывания 5А6/22Е7 снижается при соотношении 10:1 для huFcεRI ECD или huFcγRIIB ECD, при этом полное блокирование 5А6/22Е7 клеток huFcεRI+huFcγRIIB (1 или 2) наблюдается только при наличии обоих ECD в соотношении 10:1 (насыщающая концентрация).
Приведенные эксперименты показывают, что ингибирование высвобождения гистамина зависит от сшивания RBL FcεRI и FcγRIIB на поверхности клеток, поскольку не наблюдается ингибирования высвобождения гистамина при предварительной инкубации биспецифического антитела 5А6/22Е7 с 10-кратным молярным избытком внеклеточных доменов huFcεRIα и huFcγRIIB. В этих условиях связывание биспецифического антитела 5А6/22Е7 с клеточной поверхностью полностью блокируется, как следует из данных оценки методом проточной цитометрии. Предварительная инкубация со сниженными молярными соотношениями huFcεRI ECD и huFcγRIIB ECD (2:2:1, 1:1:1 или 0,1:0,1:1 huFcεRI:huFcγRIIB:биспецифическое антитело) сказывается на неполном блокировании связывания биспецифического антитела 5А6/22Е7 с клетками RBL и на неполном ингибировании высвобождения гистамина. В этой связи подавление высвобождения гистамина в тучных клетках требует сшивки на клеточной поверхности FcεRIα и FcγRIIB.
Ингибирование высвобождения гистамина биспецифическим антителом 5А6/22Е7 в приведенных концентрациях, которые ниже насыщающих, позволяет полагать, что полное занятие рецепторов не является необходимым условием для достижения желательного уровня ингибирования.
5.4 Ингибирование биспецифическим антителом в субнасыщающих концентрациях
Ингибирование биспецифическим антителом 5А6/22Е7 высвобождения гистамина и связывания клеток RBL huFcεRI+huFcγRIIB1 измеряют в концентрациях, которые ниже насыщающих по связыванию, по приведенному ниже методу, и результаты теста представлены на фиг.42-46.
Клетки RBL huFcεRI+huFcγRIIB1 или клетки RBL huFcεRI+huFcγRIIB2 инкубируют в течение 24 часов при 37°С с 5 мкг/мл NP-специфического IgE человека и затем промывают три раза свежей средой для удаления несвязанного NP-специфичного IgE человека. Перед активацией антигеном клетки дополнительно инкубируют в течение 1 часа при 37°С с варьирующими концентрациями биспецифического антитела 5А6/22Е7. Затем клетки разделяют на два варианта: для анализа методом проточной цитометрии или для оценки экспрессии гистамина.
Уровень связывания биспецифического антитела оценивают методом проточной цитометрии, как было описано выше. Проводят проточную цитометрию с использованием сравнимых концентраций биотинилированного биспецифического антитела, выявляемого с использованием стрептавидина-PE.
Клетки, подвергнутые предварительной инкубации, как было указано выше, активируют путем инкубации с 0,1 мкг/мл или 1 мкг/мл NP-конъюгированного овальбумина в течение 1 часа при 37°С. Грануляцию, связанную с активацией, измеряют путем количественной оценки уровней гистамина, высвобождаемых в клеточную культуральную среду, как было описано выше.
Данные по высвобождению гистамина и по связыванию биспецифического антитела 5А6/22Е7 с клетками RBL huFcεRI+huFcγRIIB1 представлены на фиг.42 и 43 соответственно, а данные по высвобождению гистамина и по связыванию биспецифического антитела 5А6/22Е7 с клетками RBL huFcεRI+huFcγRIIB2 представлены на фиг.44 и 45 соответственно. Подавление высвобождения гистамина до фоновых уровней показано для концентраций биспецифического антитела выше 0,0025 мкг/мл в клетках RBL huFcεRI+huFcγRIIB1 и в клетках RBL huFcεRI+huFcγRIIB2.
Оценка методом проточной цитометрии связывания биспецифического антитела с клетками RBL huFcεRI+huFcγRIIB1 и с клетками RBL huFcεRI+huFcγRIIB2 указывает на то, что насыщение по связыванию достигается в концентрации примерно 2,5 мкг/мл биспецифического антитела. На фиг.46 представлены результаты титрования по методу проточной цитометрии биспецифического антитела в диапазоне концентраций от 0,1 мкг/мл до 2,5 мкг/мл для четырех RBL-производных клеточных линий, клеток RBL huFcεRI, клеток RBL huFcγRIIB, клеток RBL huFcεRI+huFcγRIIB1 и клеток RBL huFcεRI+huFcγRIIB2. Выделенный жирным шрифтом пик соответствует клеткам, связанным с биотинилированным биспецифическим антителом. Результаты титрования для выявления связывания биспецифического антитела с RBL-производными клеточными линиями указывает на то, что связывание биспецифического антитела с клетками RBL huFcεRI+huFcγRIIB1 и RBL huFcεRI+huFcγRIIB2 снижается при снижении концентрации биспецифического антитела и не обнаруживается при значениях менее 0,0025 мкг/мл. Ингибирование биспецифическим антителом высвобождения гистамина из RBL, показанное на фиг.42 и 44, сохраняется в концентрациях биспецифического антитела ниже насыщающей концентрации для связывания, в случае использования двух разных концентраций NP-антигенного стимула.
5.5 Эффекты биспецифического антитела на уровни поверхностной экспрессии FcεRIα
Отрицательная регуляция экспрессии FcεRI на тучных клетках и базофилах представляет собой способ снижения чувствительности тучных клеток и базофилов в отношении антиген-индуцированной активации и представляет механизм, посредством которого терапевтический агент может оказывать благоприятный эффект при астме или аллергии.
Способность биспецифического антитела модулировать уровень поверхностной экспрессии FcεRI оценивают при проведении эксперимента по позитивной или отрицательной регуляции IgE-индуцированного FcεRI в присутствии и в отсутствие биспецифического антитела с использованием приведенных ниже процедур.
Клетки RBL huFcεRI+huFcγRIIB1 и RBL huFcεRI+huFcγRIIB2 инкубируют с 1 мкг/мл U266 IgE (ATCC TIB196) в присутствии или в отсутствие 2 мкг/мл биспецифического антитела в течение 1, 2, 3 или 7 дней. На фиг.47 и 48 показано, что концентрации биспецифического антитела 5А6/22Е7 и IgE остаются неизменными, по данным анализа методом ELISA с использованием IgG1 и IgE человека для выявления, в течение 7-дневного курса, указывая на то, что клеточная культуральная среда не истощается по данным реагентам. Общий уровень FcεRI человека на поверхности клеток определяют методом проточной цитометрии с использованием антитела против IgE человека (Caltag, Laboratories) после насыщения всех рецепторов FcεRI на льду в присутствии U266 IgE.
Результаты анализа позитивной регуляции FcεRI методом проточной цитометрии показаны на фиг.49-54. Биспецифическое антитело не оказывает эффекта на IgE-индуцированную позитивную регуляцию уровней поверхностной экспрессии FcεRI в 2 образцах клеток RBL FcεRI, как показано на фиг.49 и 50, и в 2 образцах клеток RBL huFcεRI+huFcγRIIB1, как показано на фиг.51 и 52. Однако биспецифическое антитело снижает выраженность позитивной регуляции FcεRI при сшивке huFcεRI и huFcγRIIB2 в 2 образцах клеток RBL huFcεRI+huFcγRIIB2, как показано на фиг.53 и 54.
Был также оценен эффект биспецифического антитела на отрицательную регуляцию FcεRIα после удаления IgE, и результаты определения приведены на фиг.55-57. FcεRIα на клетках RBL подвергают позитивной регуляции в течение 7 дней, воздействуя 1 мкг/мл U266 IgE. Затем IgE отмывают из среды для клеточной культуры и выявляют отрицательную регуляцию FcεRIα методом проточной цитометрии в присутствии и в отсутствие биспецифического антитела через 1, 2, 3 и 7 дней после удаления IgE. Биспецифическое антитело не оказывает эффекта на отрицательную регуляцию FcεRIα в клетках RBL huFcεRI и RBL huFcεRI+huFcγRIIB1, как показано на фиг.55 и 56. Однако уровень отрицательной регуляции FcεRIα повышается при воздействии биспецифического антитела в клетках RBL huFcεRI+huFcγRIIB2, как показано на фиг.57. Повторяют эксперимент с использованием клеток RBL huFcεRI+huFcγRIIB2, но биспецифическое антитело 5А6/22Е7 добавляют в присутствии IgE в дни ноль, три и четыре (см. фиг.63). Приведенные данные показывают, что биспецифическое антитело снижает IgE-индуцированную экспрессию FcεRI в данных клетках. В этих исследованиях было также показано, что изоформа huFcγRIIB1 не демонстрирует отрицательной регуляции в отношении huFcεRI.
Приведенные исследования показывают, что биспецифическое антитело может снижать уровень поверхностной экспрессии FcεRI на тучных клетках и базофилах за счет поперечного сшивания FcεRIα с В2 изоформой FcγRIIB. Приведены результаты анализа методом RT-ПЦР экспрессии мРНК для huFcεRIα, FcγRIIB1, FcγRIIB2, huRPL 19 (контроль) и крысиного FcεRIα в тучных клетках: клетках RBL huFcεRI (обозначены как huFcERIa), клетках RBL huFcεRI+huFcγRIIB1 (обозначены как huFcGRIIb1) и клетках RBL huFcε+huFcγRIIB2 (обозначены как huFcGRIIb2); и на базофилах человека от трех разных доноров. Идентификацию в масштабе реального времени результатов RT-ПЦР изоформ FcγRIIB1 и FcγRIIB2 проводят на основе мРНК, полученной из очищенных базофилов периферической крови от трех разных людей-доноров. Базофилы крови человека выделяют из 100 мл крови путем очистки на магнитных шариках (набор MAC для выделения базофилов человека, Miltenyi). мРНК из 106 базофилов получают с использованием мининабора RNeasyTM (Qiagen). В таблице 2 приведен перечень праймеров/зондов, используемых для анализа по методу RT-ПЦР в масштабе реального времени.
Таблица 2


РНК анализируют в системе определения последовательности ABI PRISM® 7700 с использованием большого набора для проведения одностадийной RT-ПЦР TaqMan® One-Step RT-PCR Master Mix (Applied Biosystems) в соответствии с рекомендуемым производителем протоколом. И В1, и В2 изоформы FcγRIIB экспрессируются на базофилах человека, как показано на фиг.58-61, выявленная при этом способность биспецифического антитела осуществлять модуляцию по типу отрицательной регуляции уровня поверхностной экспрессии FcεRI путем сшивки с FcγRIIB2 в клетках делает методы использования анти-FcγRIIB-анти-FcεRI биспецифического антитела по настоящему изобретению особенно полезным для лечения пациентов, имеющих расстройства, в случае которых ингибирование и/или отрицательная регуляция FcεRI обеспечивает ослабление симптомов расстройства.
5.6 Биспецифическое антитело ингибирует высвобождение цитокинов в клеточной линии RBL
Высвобождение цитокинов MCP-1 (моноцитарный хемотаксический протеин-1), IL-4 (интерлейкин-4) и TNFα (фактор некроза опухолевой ткани α) ингибируется в присутствии анти-FcγRIIB-анти-FcεRI биспецифического антитела 5А6/22Е7, как показано в приведенном ниже тесте. Клетки RBL трансфицируют кДНК, кодирующей huFcγRIIB2 или huFcγRIIB1 и huFcεRI, и культивируют по процедурам, описанным выше в тексте данного примера 5. Клетки стимулируют для высвобождения цитокинов путем воздействия конъюгированного с нитрофенолом (NP) овальбумином (NP(11)-OVA) и IgE (анти-NP IgE человека), как было описано в примере 5, в разделе, относящемся к анализу высвобождения гистамина. К исследуемым образцам добавляют биспецифическое антитело 5А6/22Е7 в концентрации 5 мкг/мл. Выявление и количественное определение каждого из интересующих цитокинов проводят по методике, описанной ниже применительно к интересующим цитокинам. MCP-1 и IL-4 выявляют с использованием набора для цитокинов Beablyte Rat Multi-cytokine Beadmaster kit (номер по каталогу 38-200, Upstate, Charlottesville, VA, USA). TNF-альфа крысы выявляют с использованием набора с противокрысиным TNF-альфа для ELISA, в соответствии с инструкцией производителя. Тесты проводят по инструкциям от производителя. На фиг.64 показаны результаты определения высвобождения цитокинов в клетках RBL, трансфицированных huFcγRIIB2 и huFcεRI, однако, результаты были такие же, как и в случае клеток RBL, трансфицированных huFcγRIIB1 и FcεRI. Высвобождение цитокинов из тучных клеток крысы ингибируется в присутствии биспецифического антитела 5А6/22Е7 (5 мкг/мл, светлые прямоугольники), тогда как в клеточной культуре высвобождение цитокинов не ингибируется и повышается в течение периода времени, равного пяти часам (темные прямоугольники).
5.7 Биспецифическое антитело ингибирует синтез и высвобождение арахидоновой кислоты в клеточной линии RBL
Присутствие аллергена инициирует множественные иммунные реакции, включающие высвобождение так называемых «предсформированных» воспалительных медиаторов, таких как гистамин, из тучных клеток, продукцию арахидоновой кислоты и ее превращение в так называемые «эйкозаноидные» медиаторы, такие как простагландины, и продукцию и высвобождение цитокинов хемокинов. Предсформированные медиаторы высвобождаются сразу при воздействии, тогда как высвобождение эйкозаноидных медиаторов задерживается примерно на 30 минут - 2 часа, а высвобождение цитокинов и хемокинов задерживается еще примерно на 5-24 часа. Будучи одним из способов защиты организма, получившим название «каскад арахидоновой кислоты», данный механизм продуцирует три заново образуемых воспалительных медиатора: простагландины, тромбоксаны и лейкотриены, которые в целом известны как эйкозаноиды. Отслеживают высвобождение метаболитов арахидоновой кислоты для выявления способности биспецифического антитела 5А6/22Е7 ингибировать эффект воздействия аллергена в направлении считывания информации. Клетки RBL трансфицируют кДНК, кодирующей huFcγRIIB1 или huFcγRIIB2 и FcεRI, и культивируют, как было описано выше в тексте данного примера 5. Каскад арахидоновой кислоты стимулируется под действием овальбумина, конъюгированного с нитрофенолом (NP) (NP (11)-OVA), в качестве антигена, в сочетании с IgE (анти-NP IgE человека), как было описано в данном примере 5 в разделе, относящемся к анализу высвобождения гистамина. Количественное определение метаболита лейкотриена C4 (LTC4) проводят с использованием набора для EIA (номер по каталогу 520211, Cayman Chemical Company, Ann Arbor, MI, USA) в соответствии с инструкциями производителя. Количественное определение метаболита простагландина D2 (PGD2) проводят с использованием набора MOX для EIA (номер по каталогу 212011, Cayman Chemical Company, выше) в соответствии с инструкциями производителя. Приведенные на фиг.65 результаты показывают, что в клетках RBL, экспрессирующих huFcγRIIB1 и FcεRI, метаболизм арахидоновой кислоты повышается со временем в присутствии IgE плюс антиген, но не в присутствии нерелевантного антигена (TNP(11)-OVA), как следует из данных по продукции LTC4 и PGD2. В присутствии 5 мкг/мл биспецифического антитела 5А6/22Е7 метаболизм арахидоновой кислоты ингибируется. Такие же результаты были получены с использованием клеток RBL, экспрессирующих huFcγRIIB2 и FcεRI (данные не показаны). Полученные результаты показывают, что важный иммунный путь ингибируется анти-FcγRIIB-анти-FcεRI биспецифическим антителом.
5.8 Биспецифическое антитело ингибирует IgE-индуцированное выживание тучных клеток
Выживание тучных клеток, полученных из костного мозга человека (huBMMC), индуцируется IgE мыши. Для определения того, ингибирует ли биспецифическое антитело 5А6/22Е7 такое выживание, проводят следующий тест. Стволовые клетки-предшественники гематопоэтических клеток человека (CD34+) получают от Allcells (номер по каталогу ABM012, Allcells, LLC, Berkeley, CA, USA). Клетки от каждого из трех доноров культивируют две недели в бессывороточной среде StemPro-34® (Gibco Cell Culture Systems, Invitrogen, Carlsbad, CA, USA), содержащей IL-3 (в концентрации 30 нг/мл), IL-6 (в концентрации 200 нг/мл) и фактор стволовых клеток (SCF, в концентрации 100 нг/мл). Выживание тучных клеток оценивают с использованием процедуры окрашивания аннексином (Annexin/7-AAD (7-амино-актиномицин D)) (набор для проточной цитометрии, BD/Pharmingen; Becton Dickenson & Company, Franklin Lakes, NJ, USA) в следующих условиях: (1) одна среда StemPro®, (2) среда StemPro® + 30 нг/мл IL-3, 200 нг/мл IL-6 и 100 нг/мл SCF, (3) среда StemPro® + 5 мкг/мл SPE-7 (мышиное IgE анти-DNP моноклональное антитело) (SPE-7, Sigma, St. Louis, MO, USA), (4) среда StemPro® + 5 мкг/мл кипяченого денатурированного SPE-7, и (5) среда StemPro® + 5 мкг/мл SPE-7+5 мкг/мл биспецифического антитела 5А6/22Е7. Выживание клеток отслеживают в течение 10 дней после исходного двухнедельного периода культивирования. Клетки поддерживают при 37°С в атмосфере 5% CO2 в ходе обеих фаз. В период времени от 4 до 7 дней после начала культивирования клеток определяют уровень выживания клеток. Средний процент ингибирования выживания клеток для трех образцов донорских клеток составляет 65±9%. Приведенные результаты показывают, что ингибирование активности рецептора FcεRI за счет сшивания с рецептором FcγRIIB с использованием анти-FcγRIIB-анти-FcεRI биспецифического антитела подавляет индуцированное IgE мыши выживание тучных клеток, полученных из костного мозга человека.
Приведенное выше описание, примеры и другие данные представляют полное описание получения и использования композиции по настоящему изобретению. Несмотря на то, что настоящее изобретение может быть реализовано в виде множества вариантов, все они осуществляются в рамках его принципов и тематики, которые зафиксированы в прилагаемой ниже формуле изобретения.

























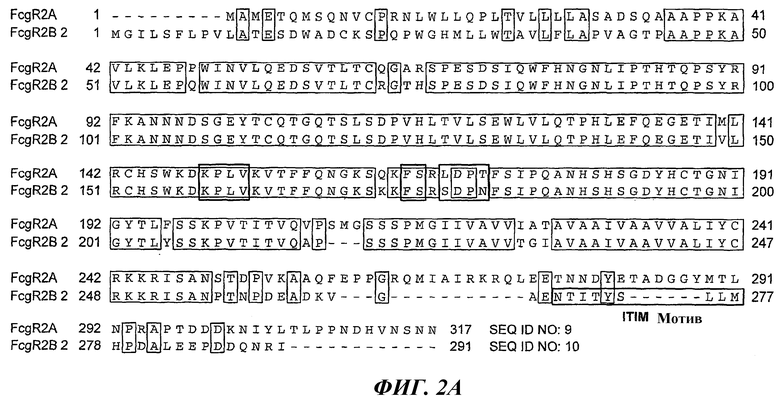


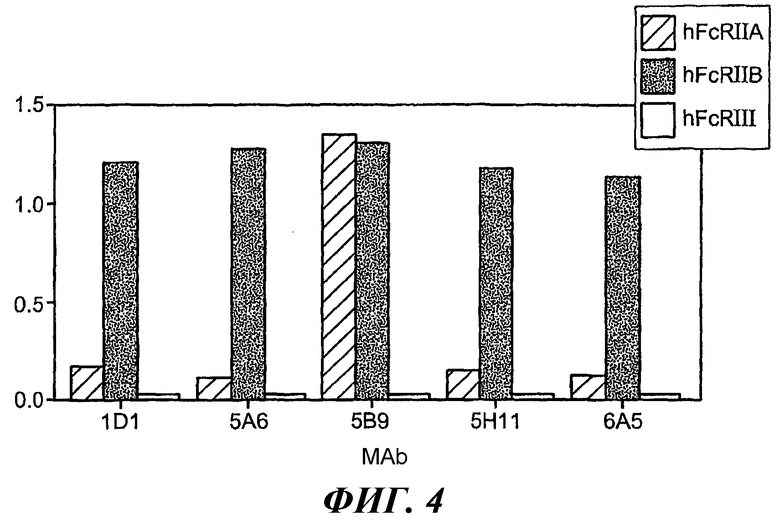
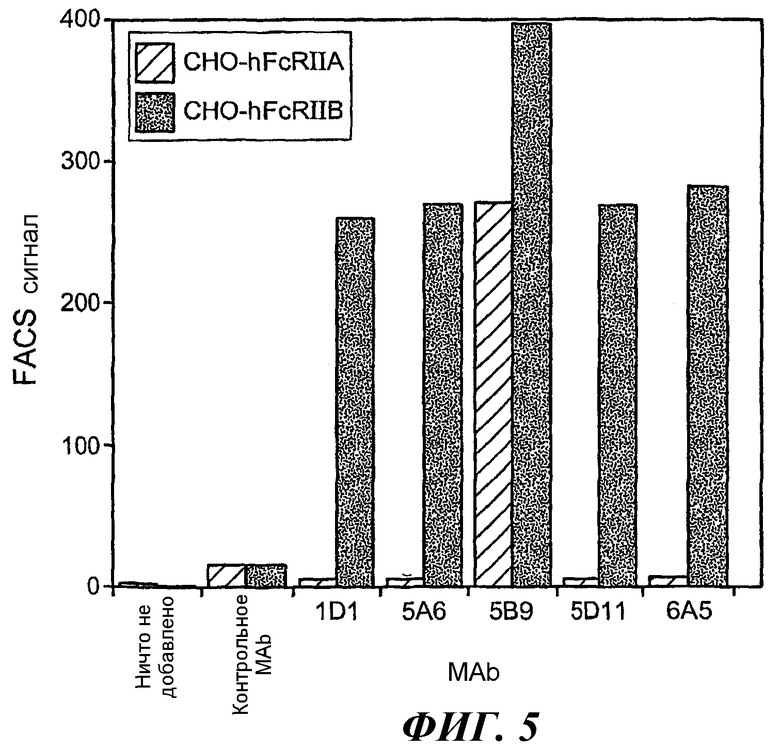

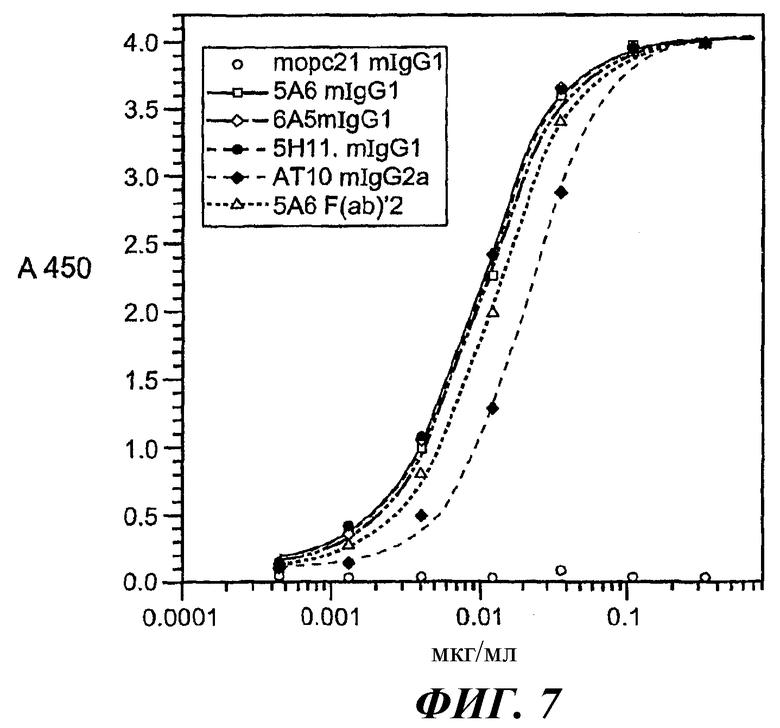
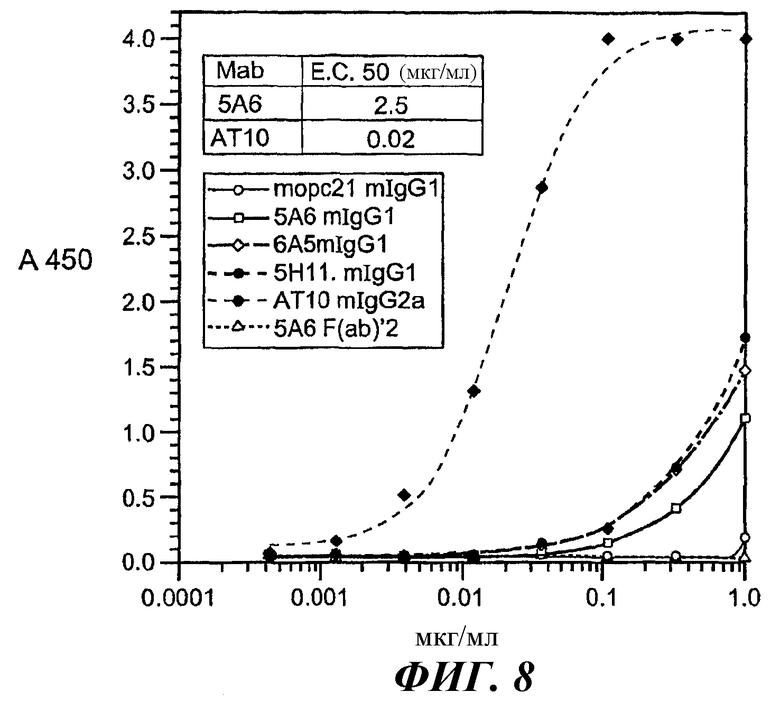
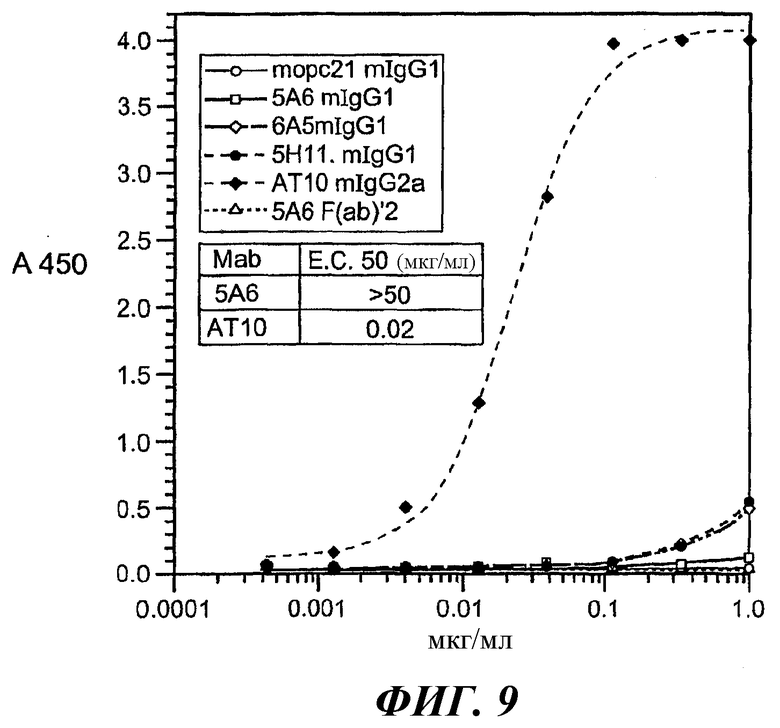
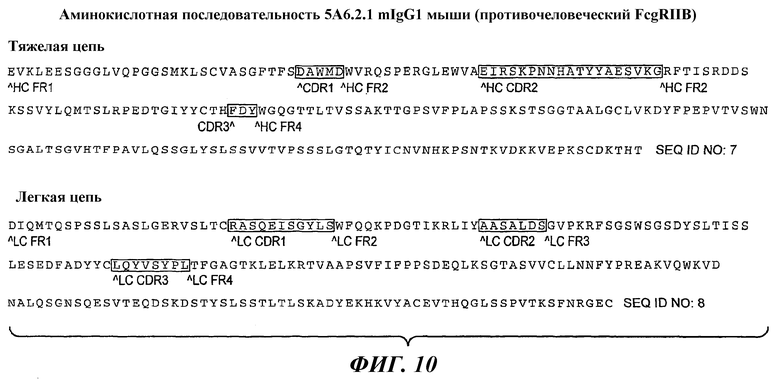
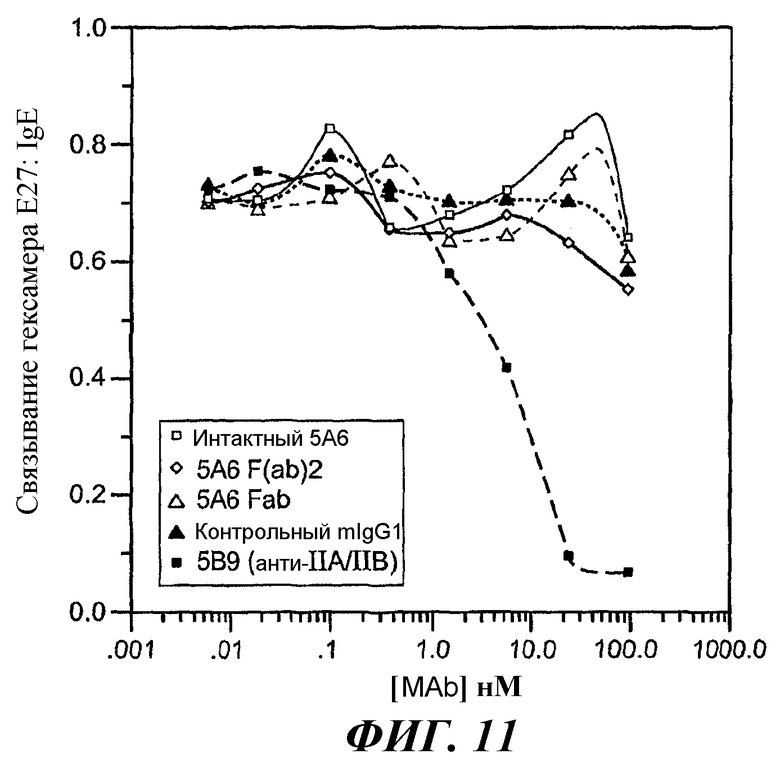
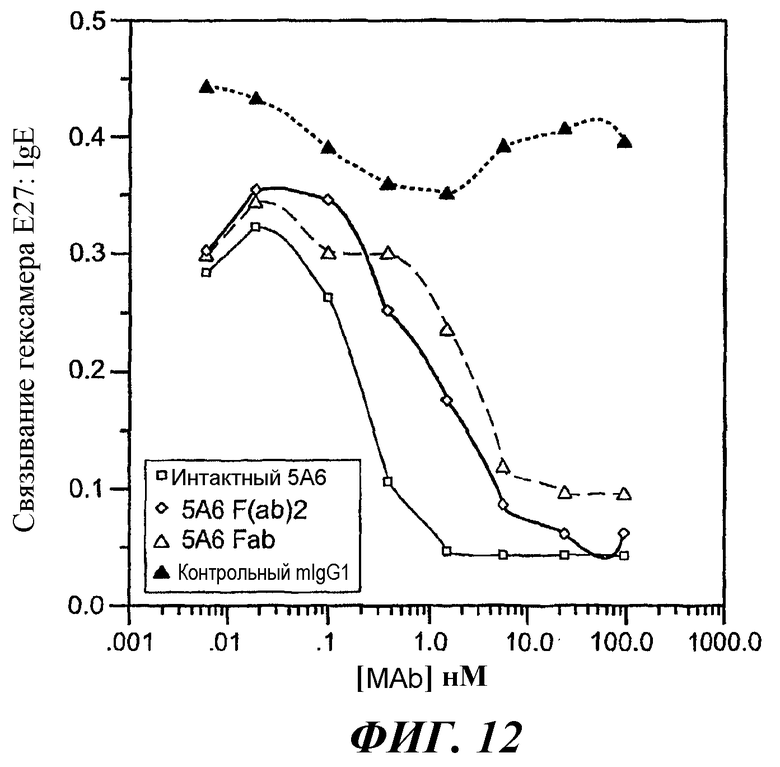
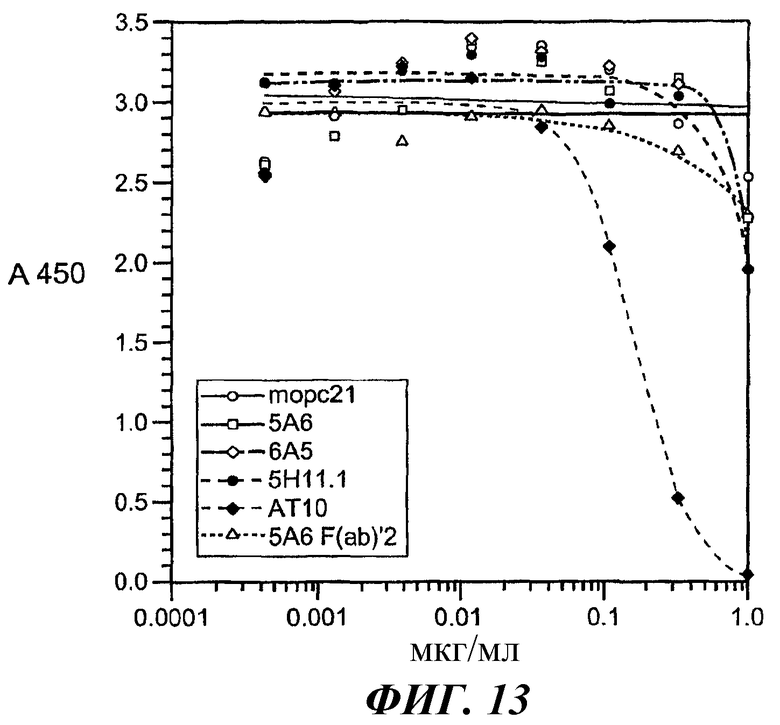
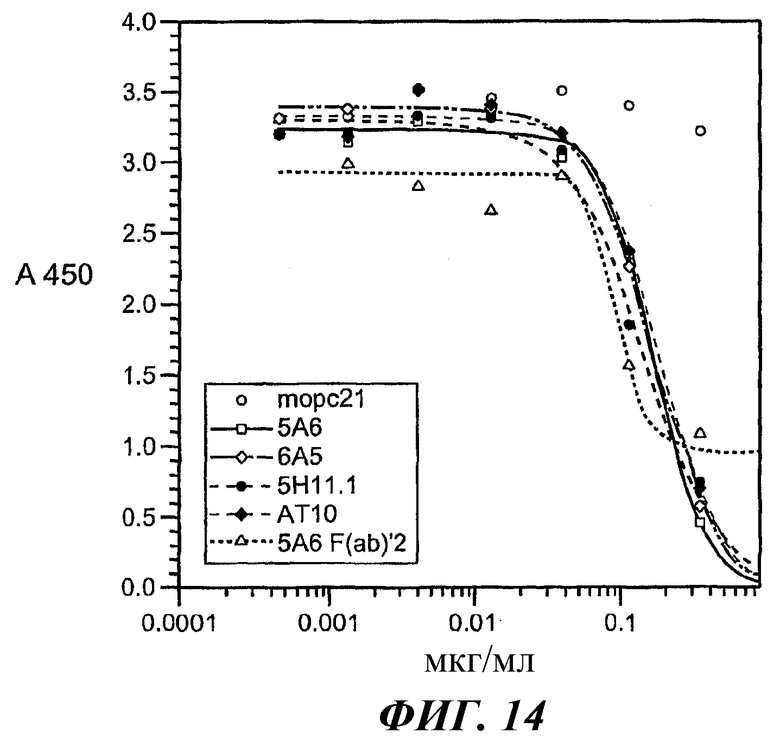
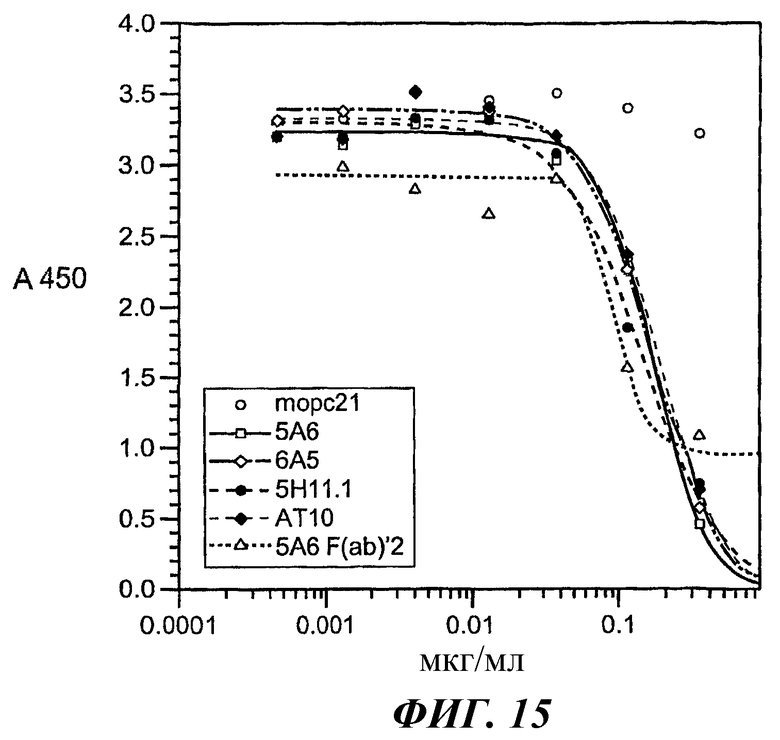
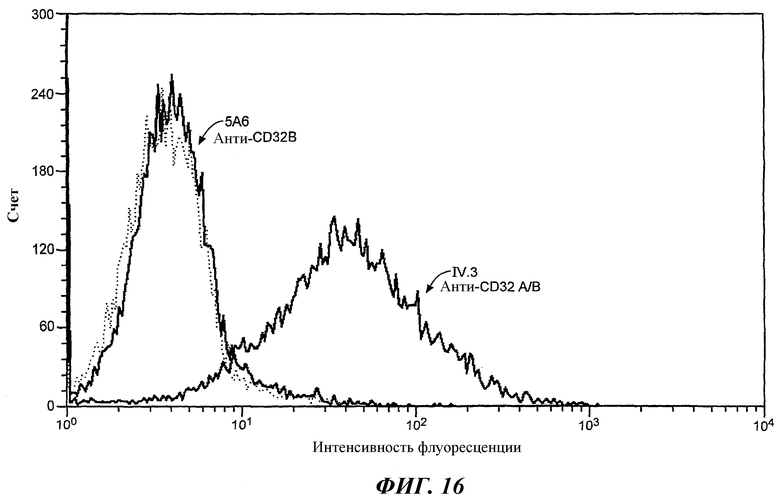
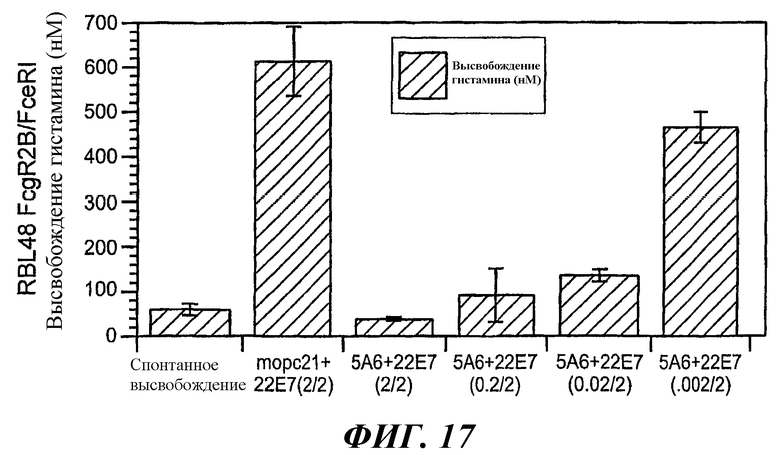
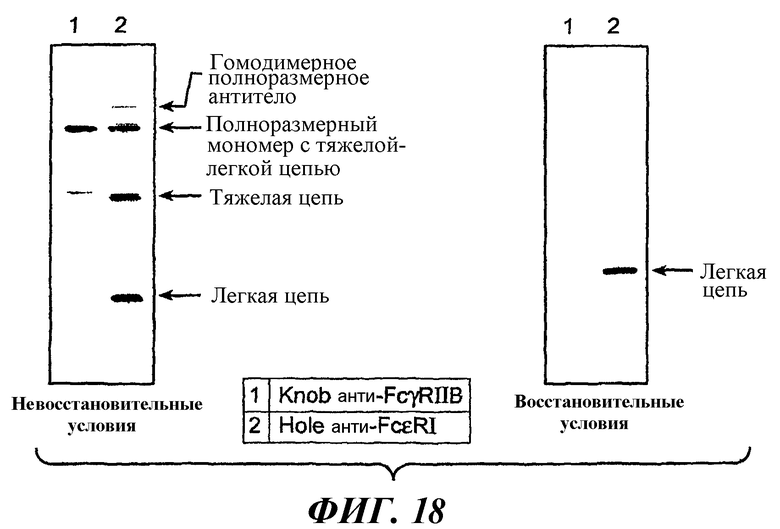
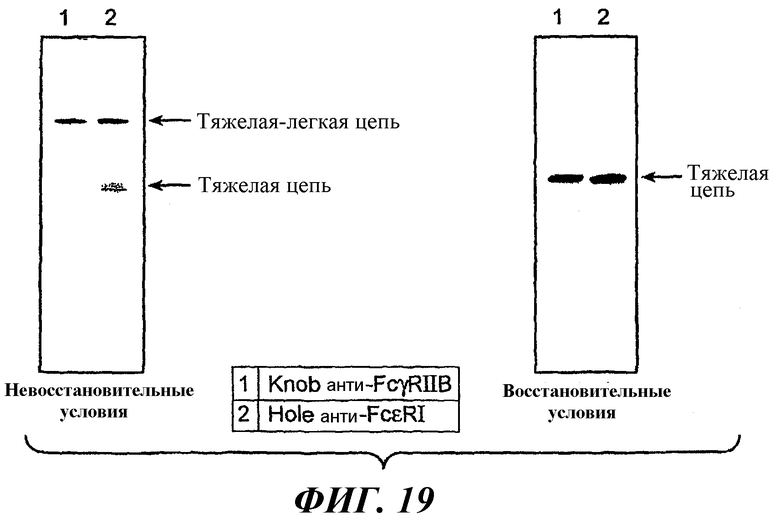
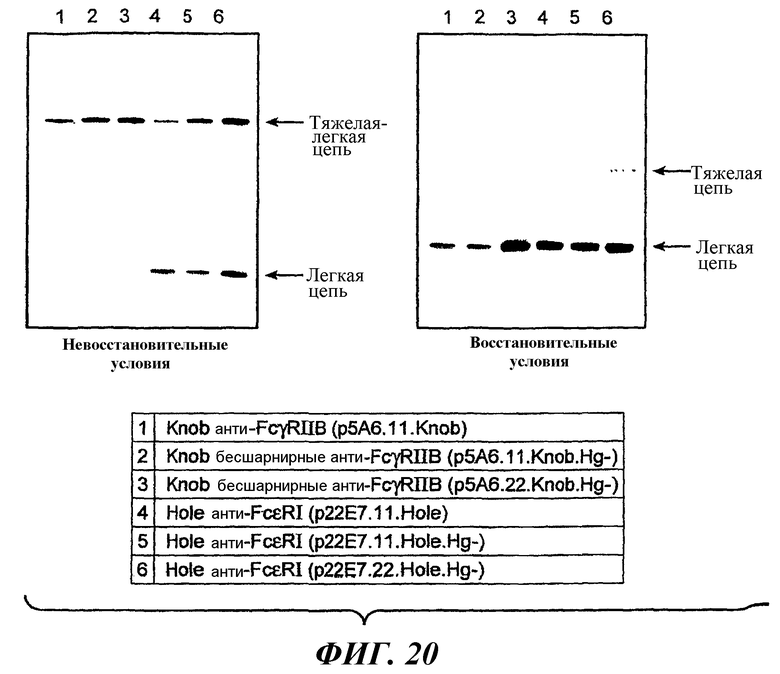
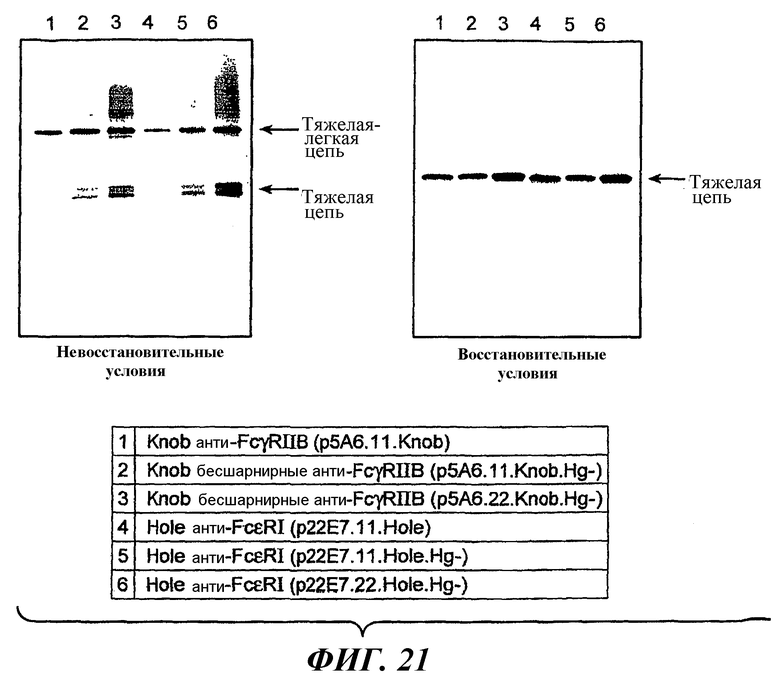
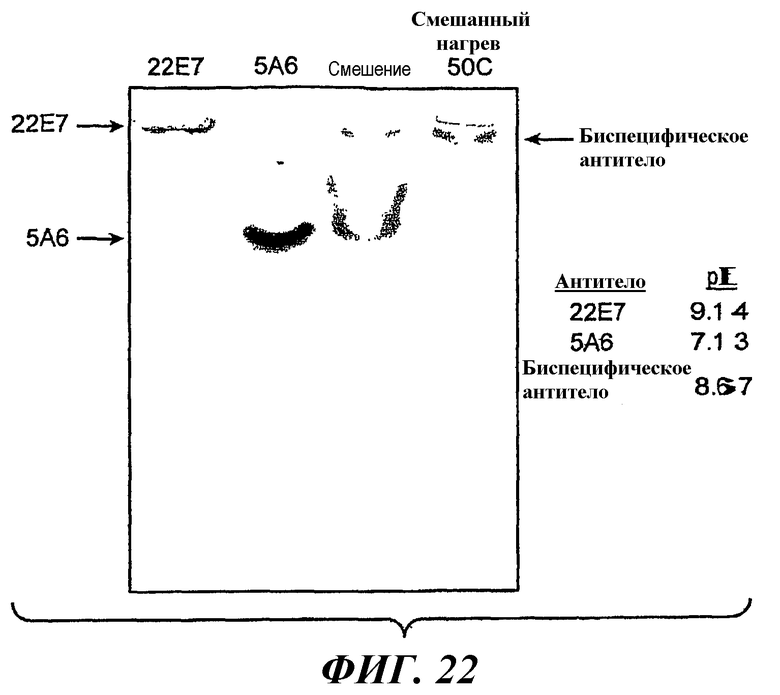
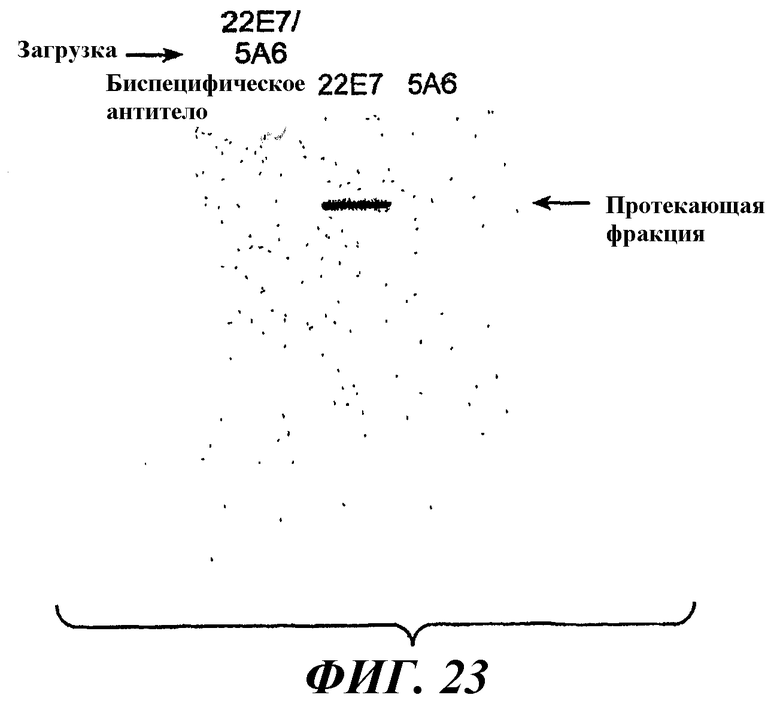
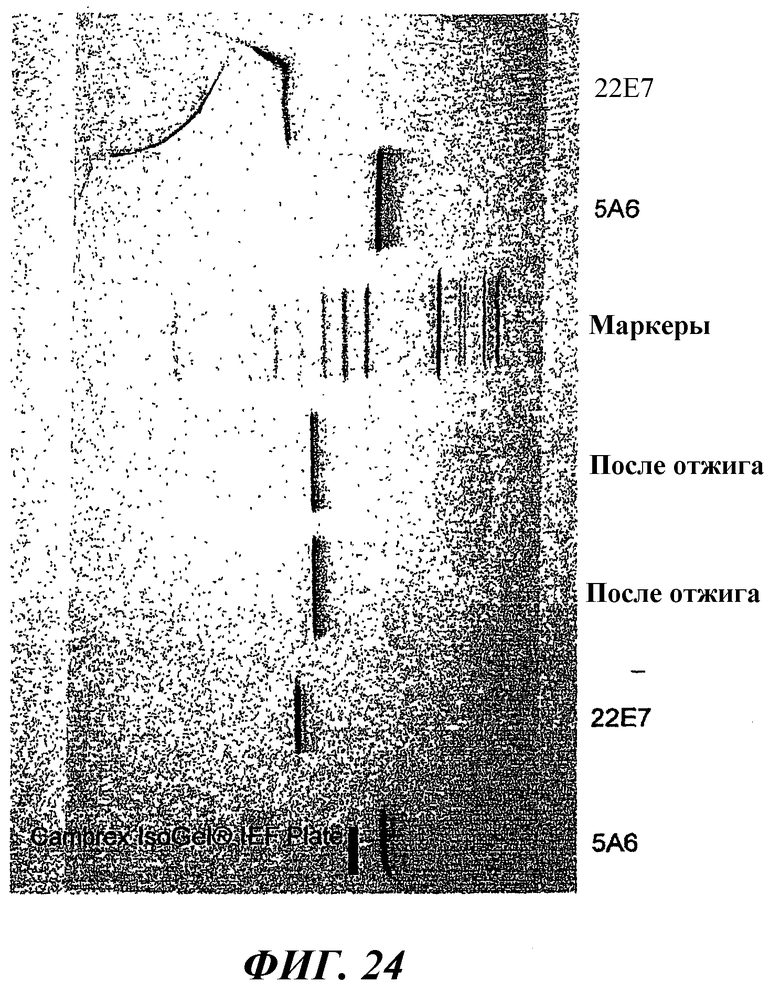





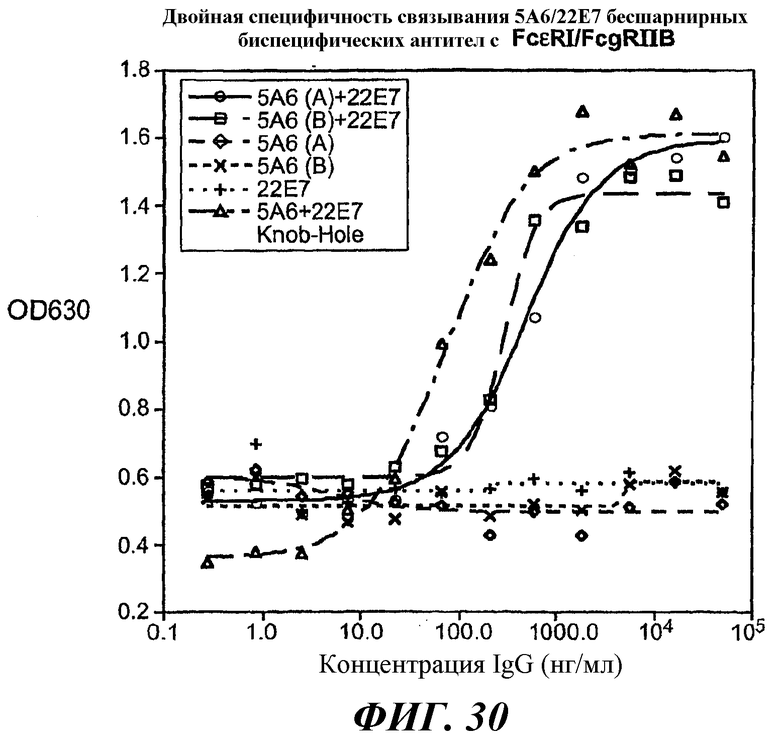
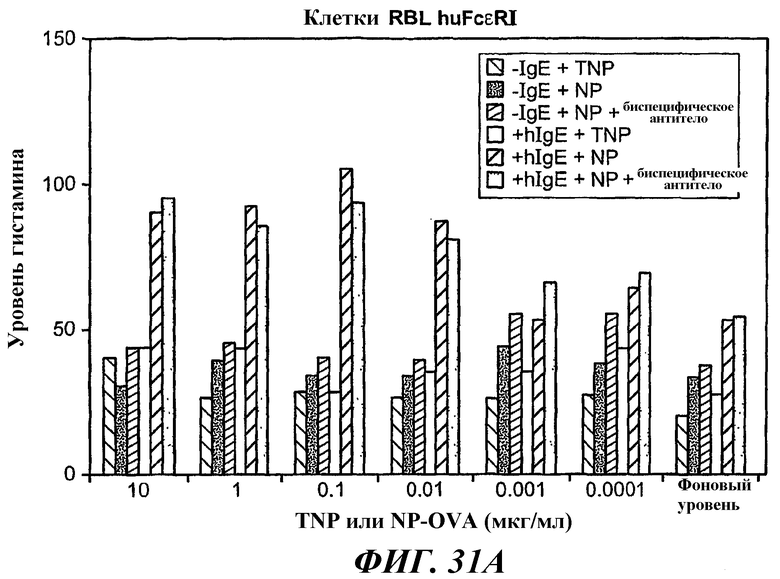


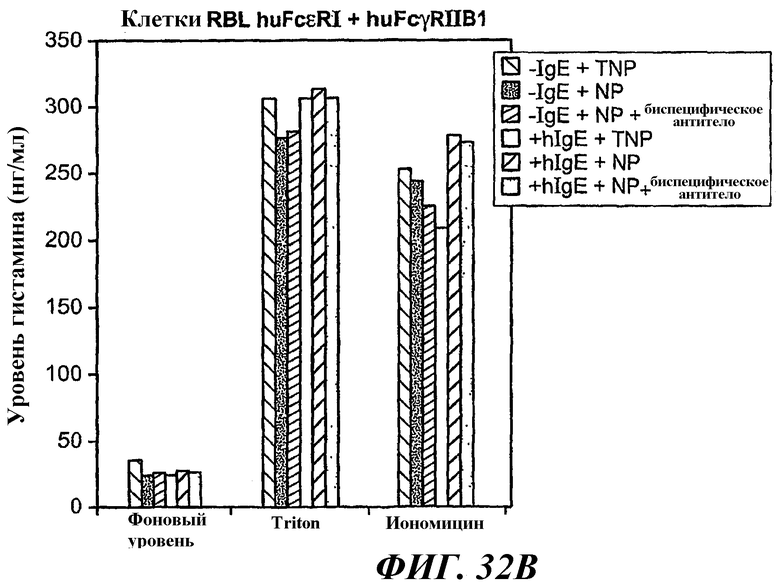
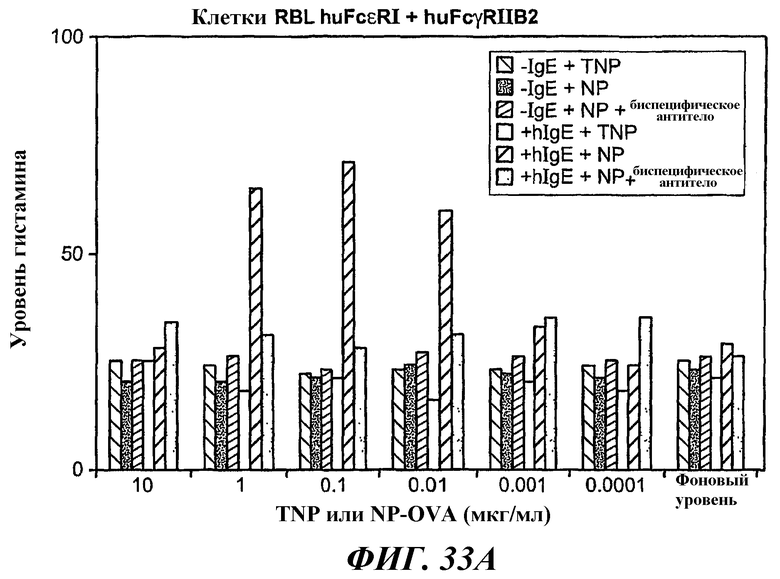
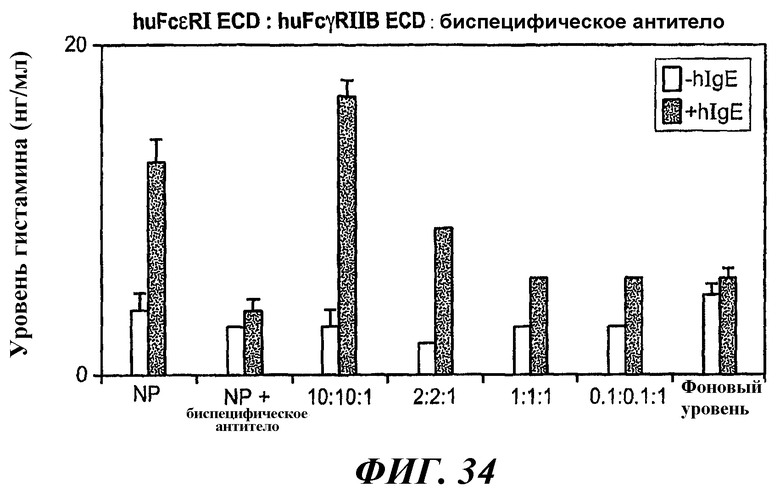

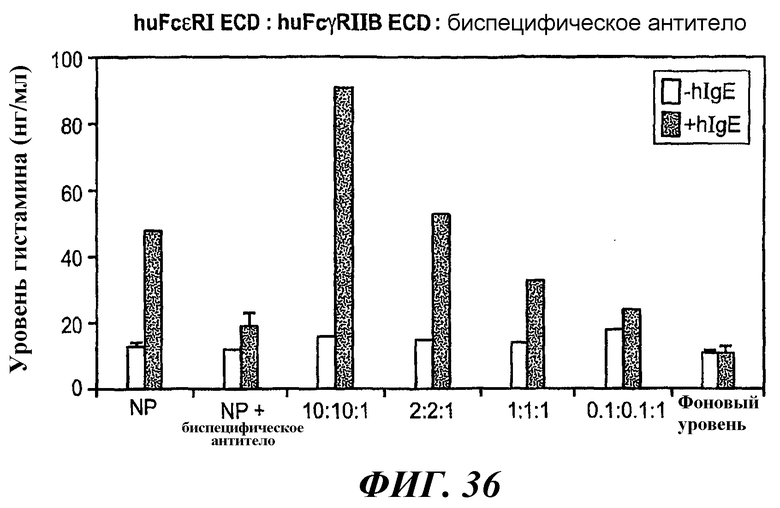
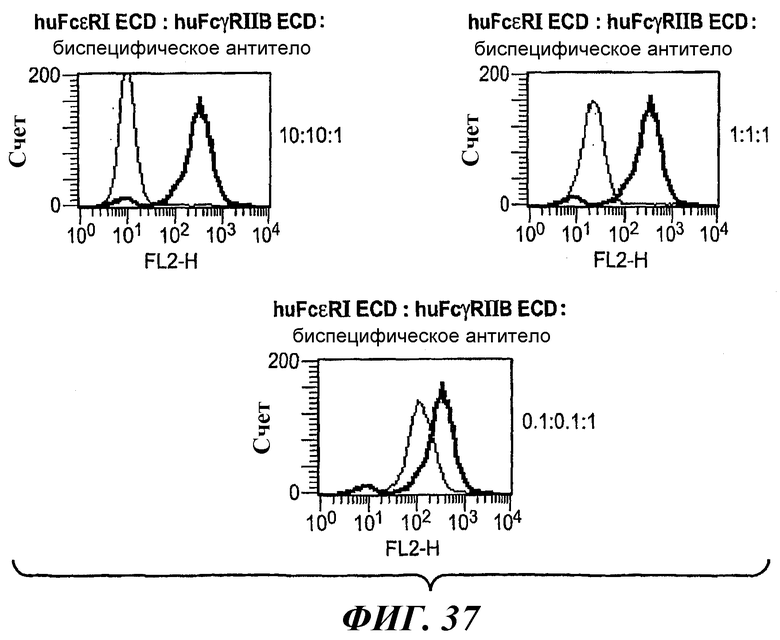

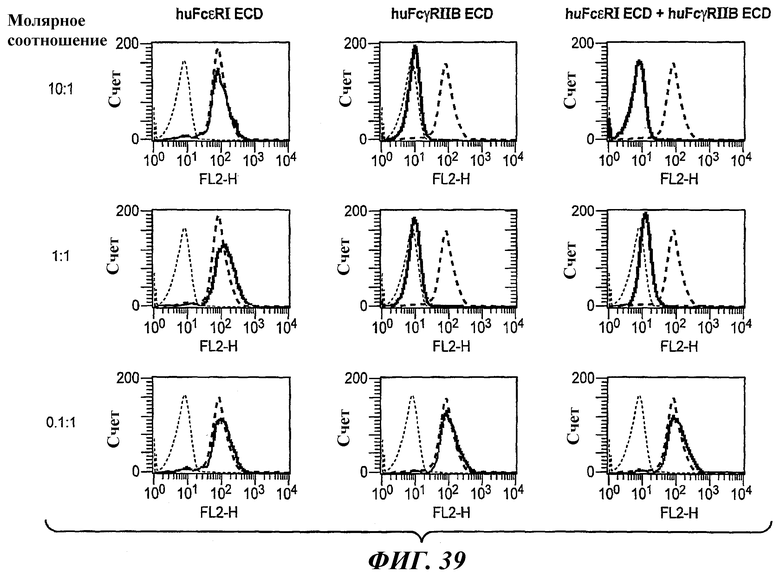
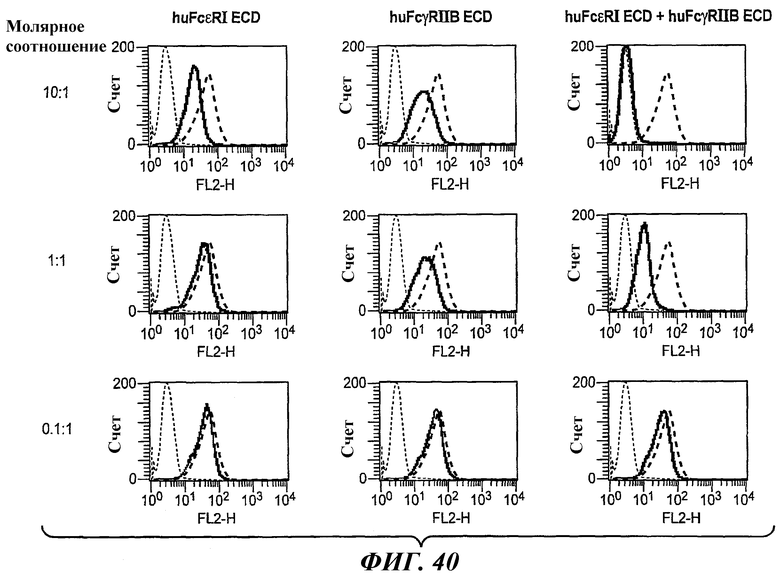
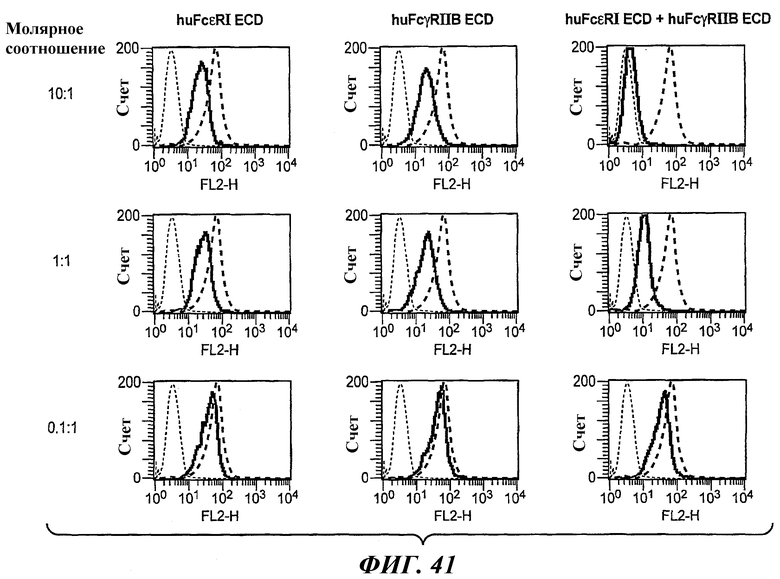
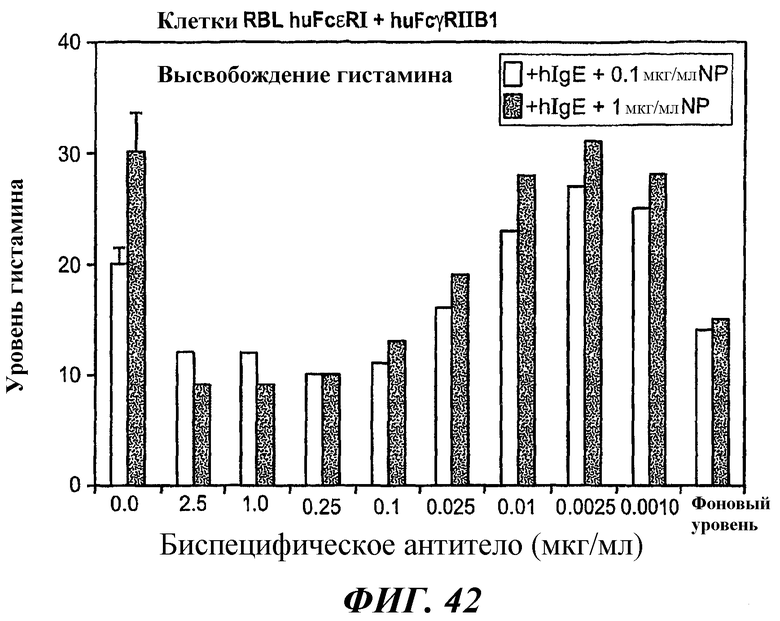
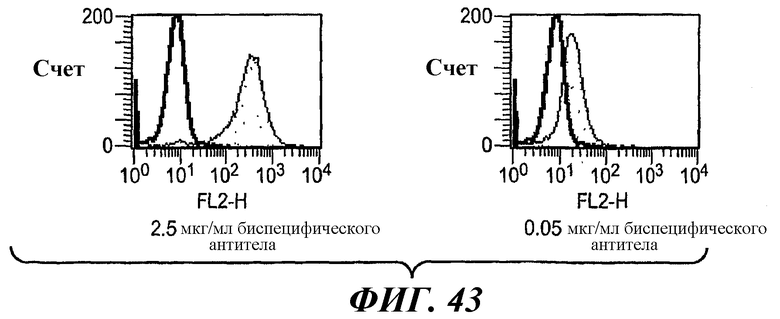
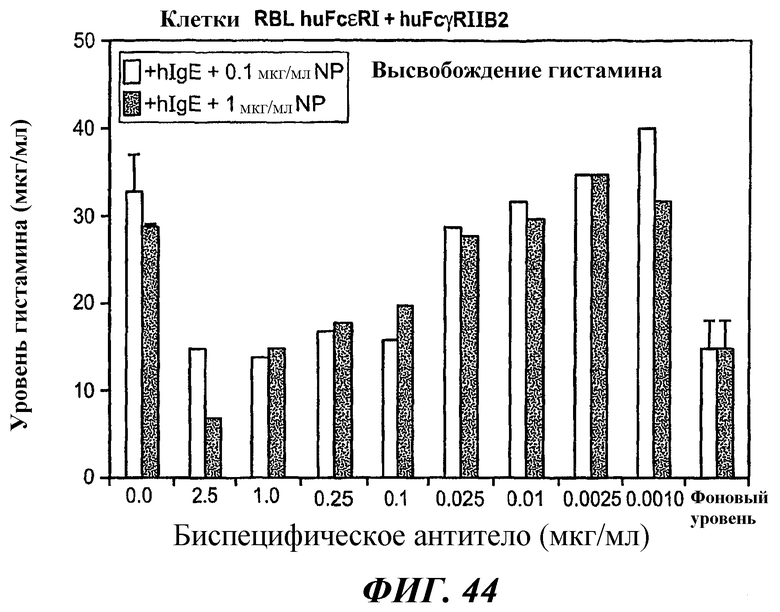
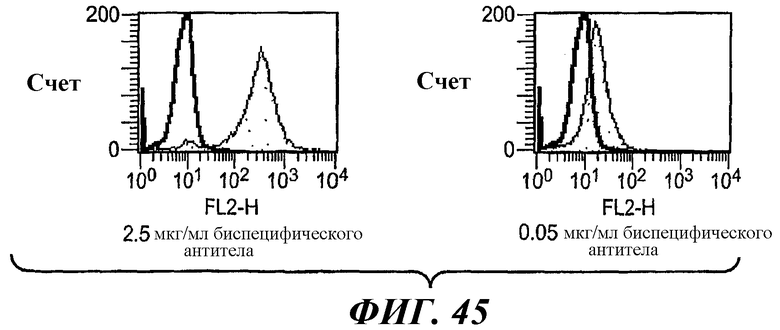
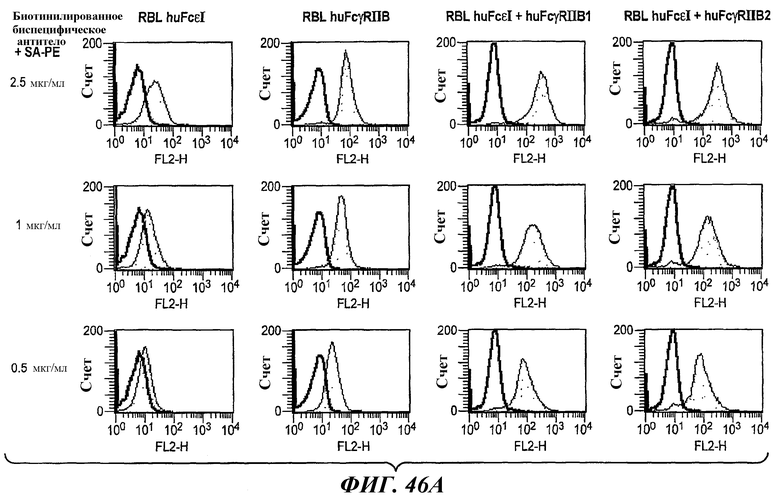
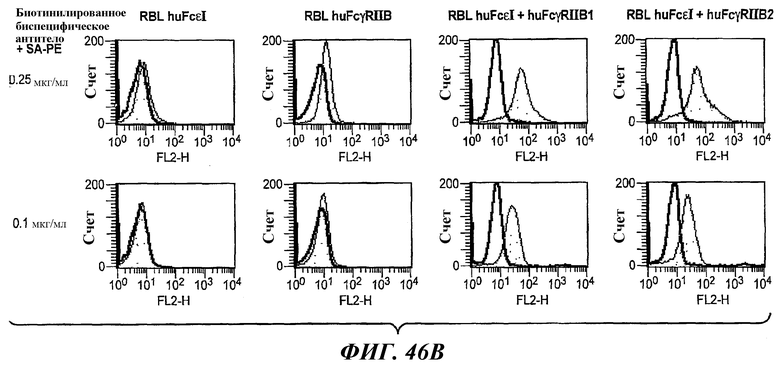
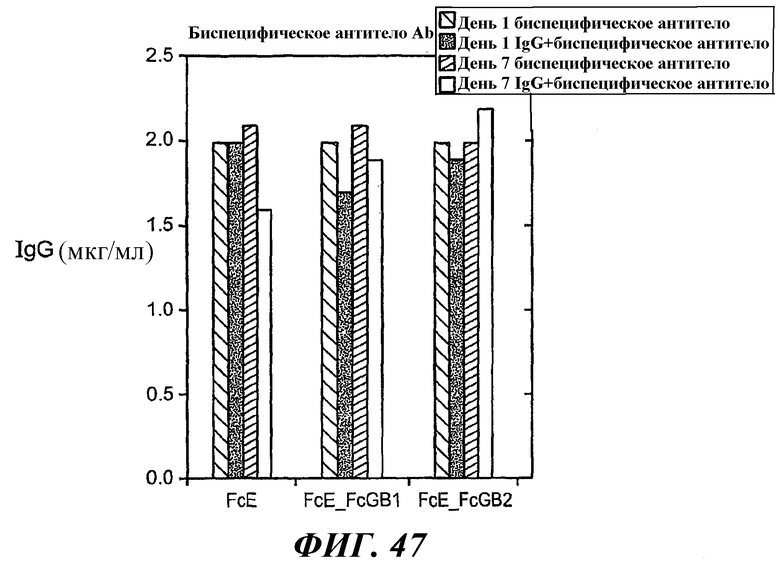

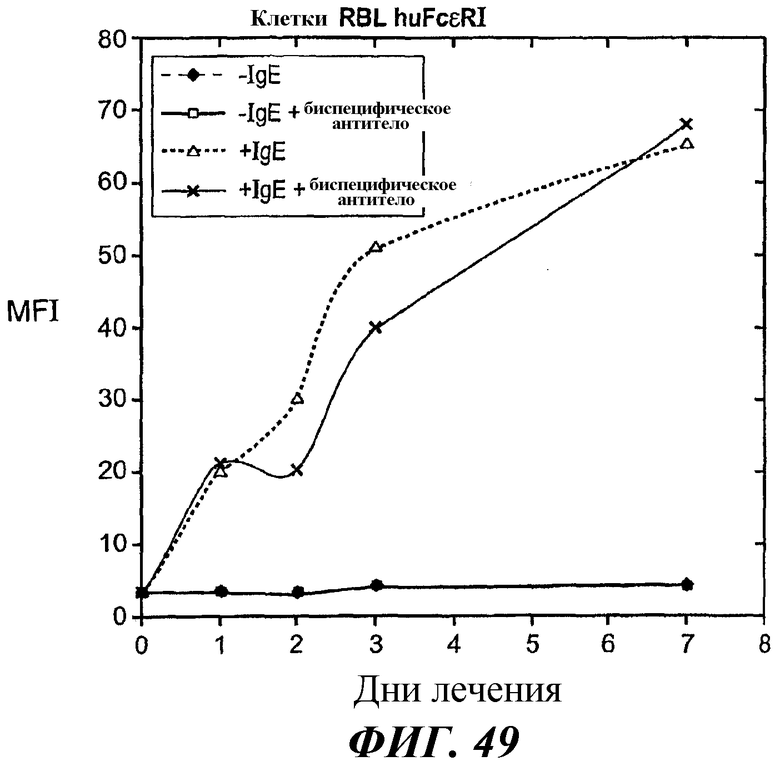
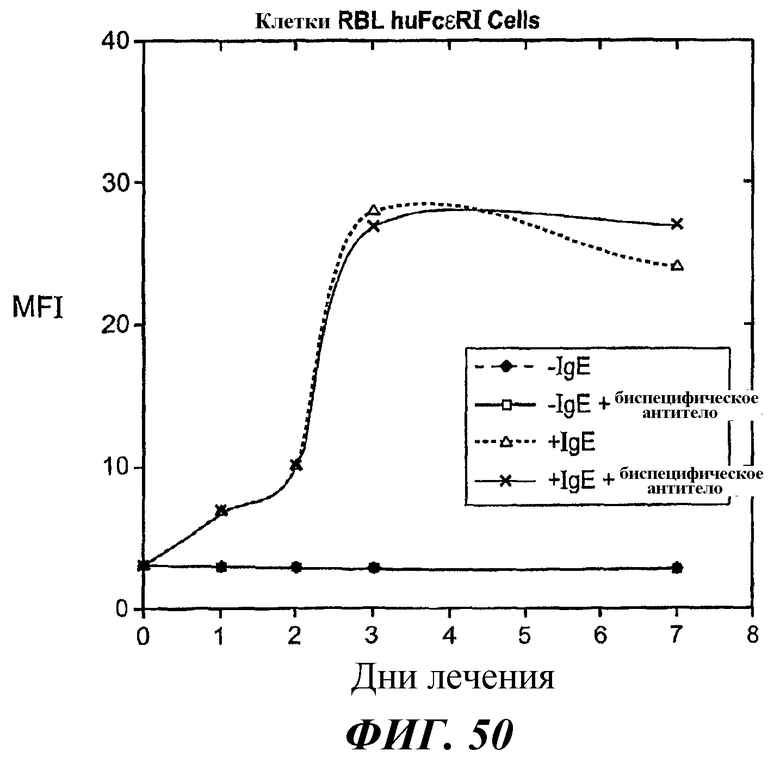
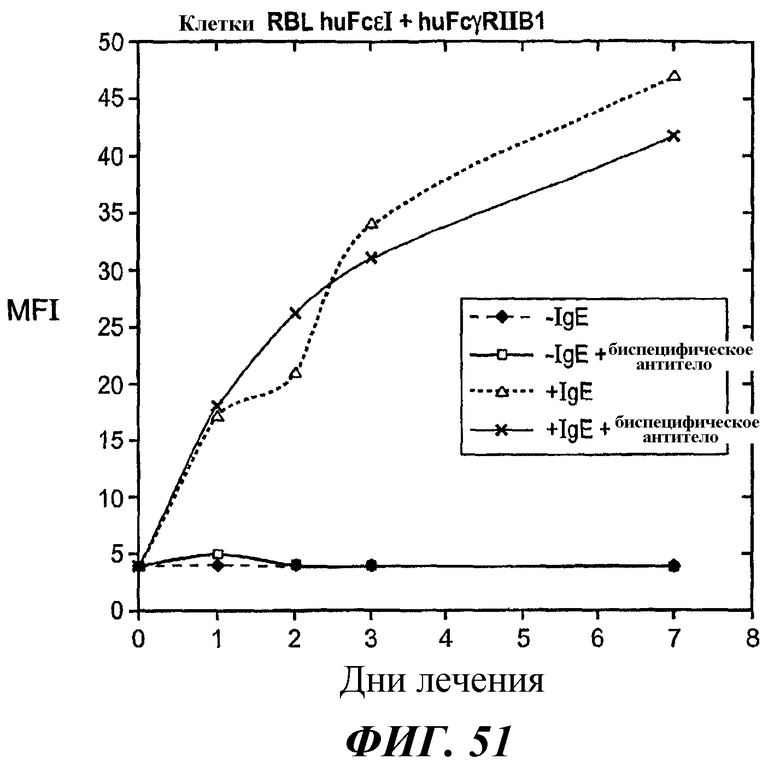

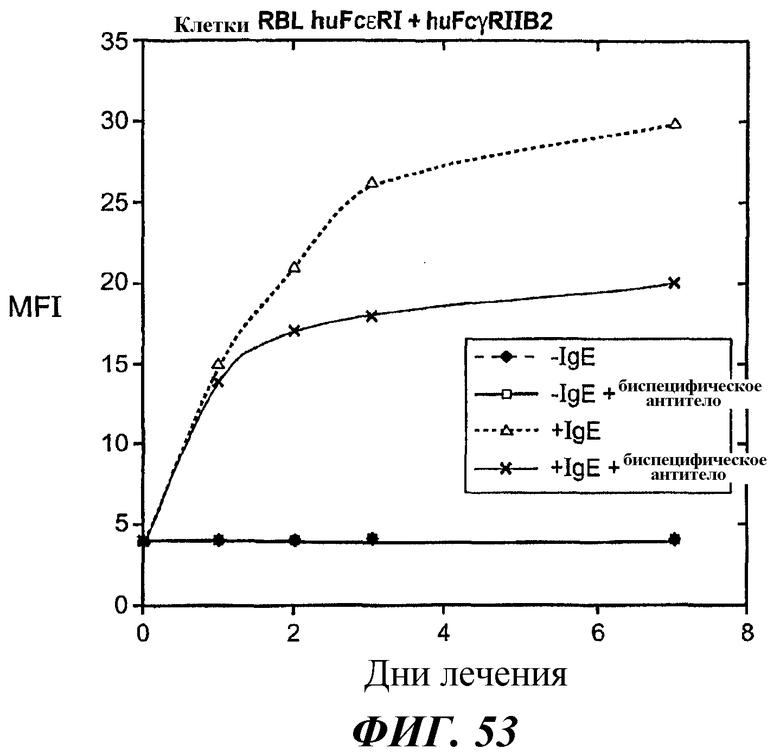
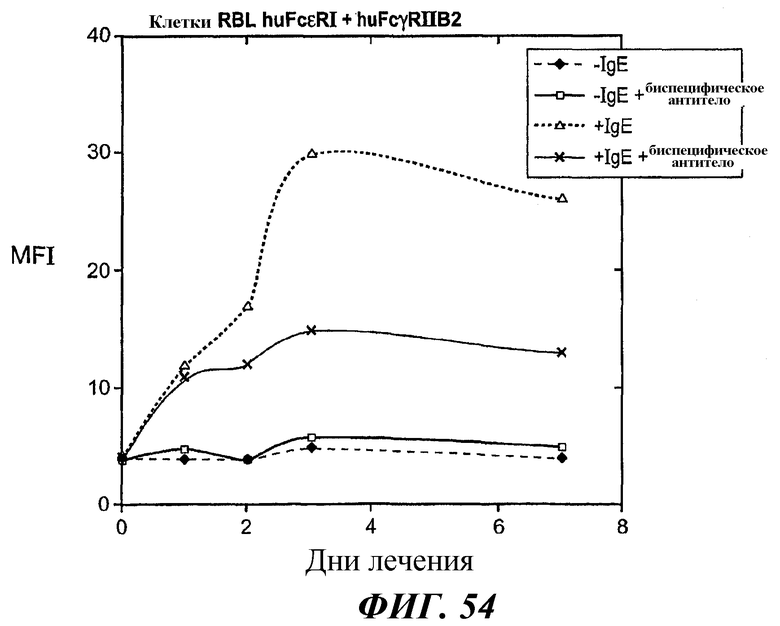
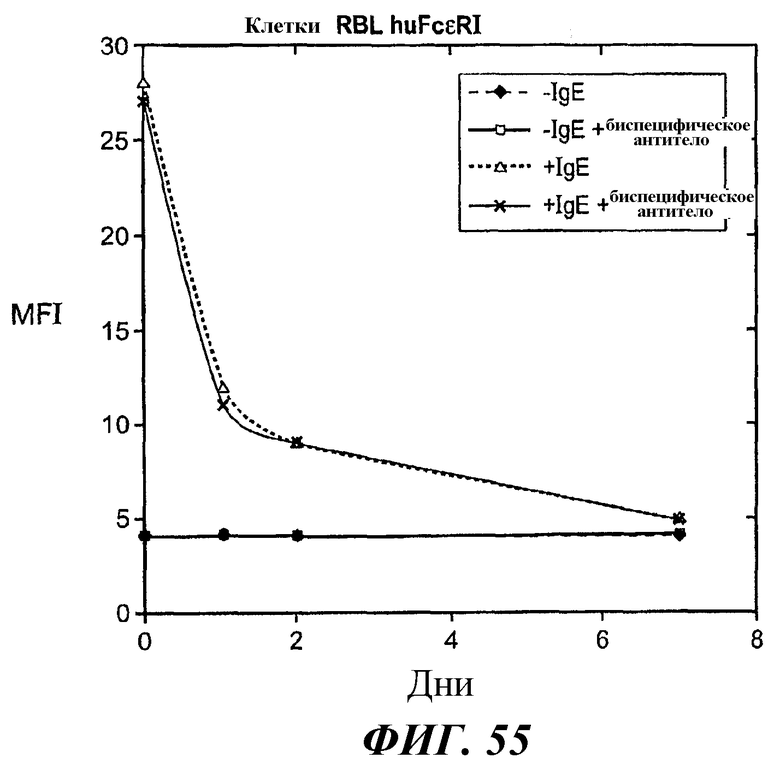

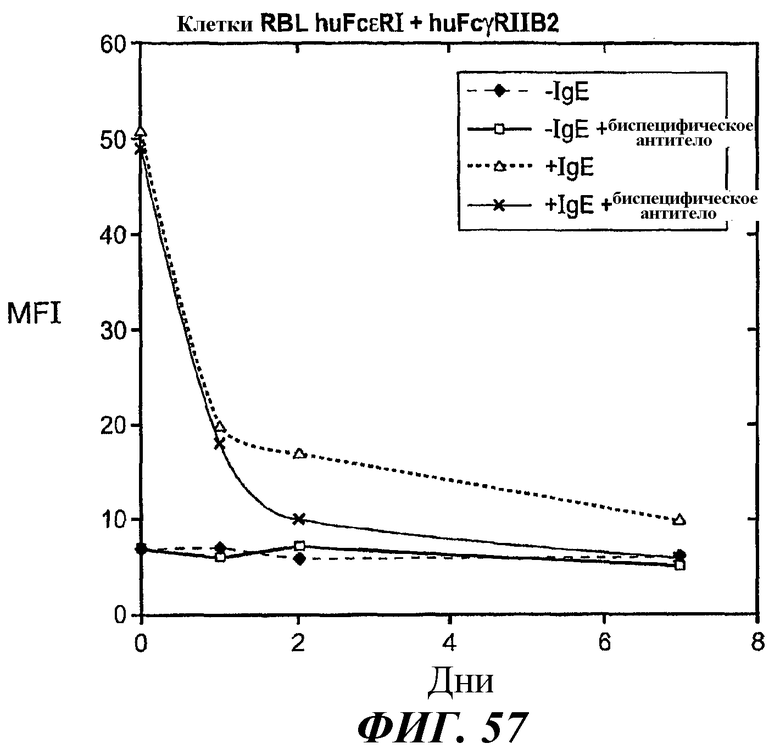
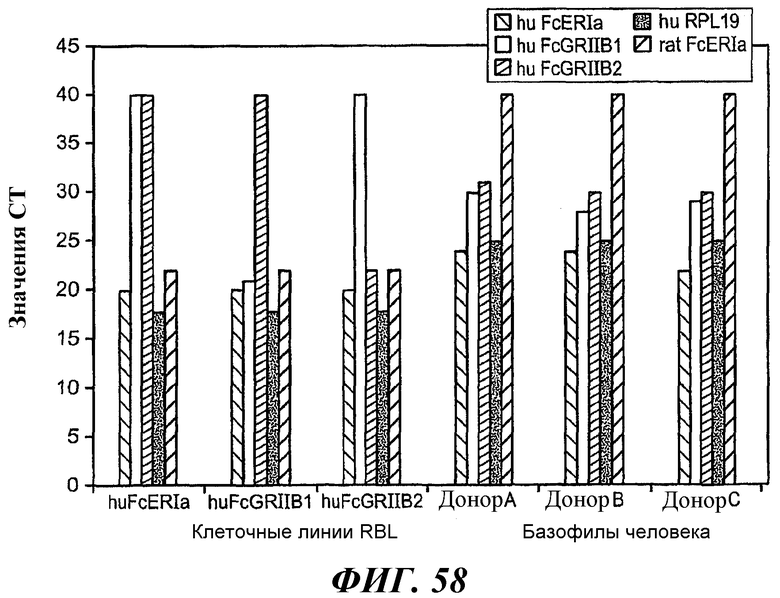
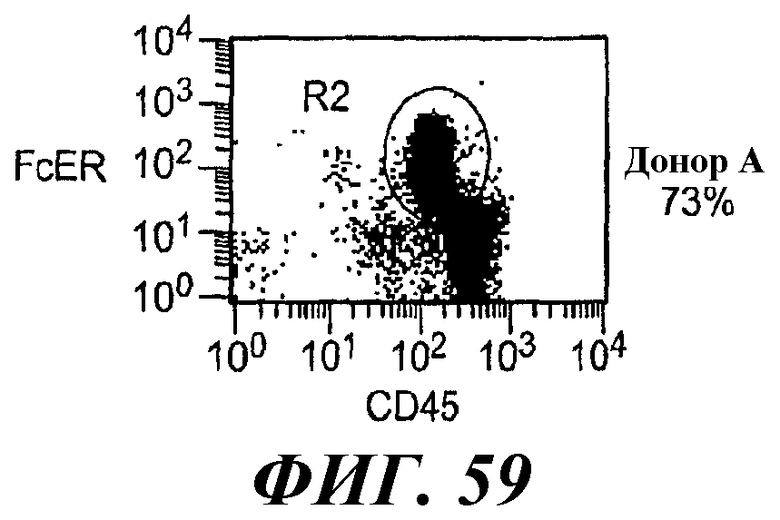
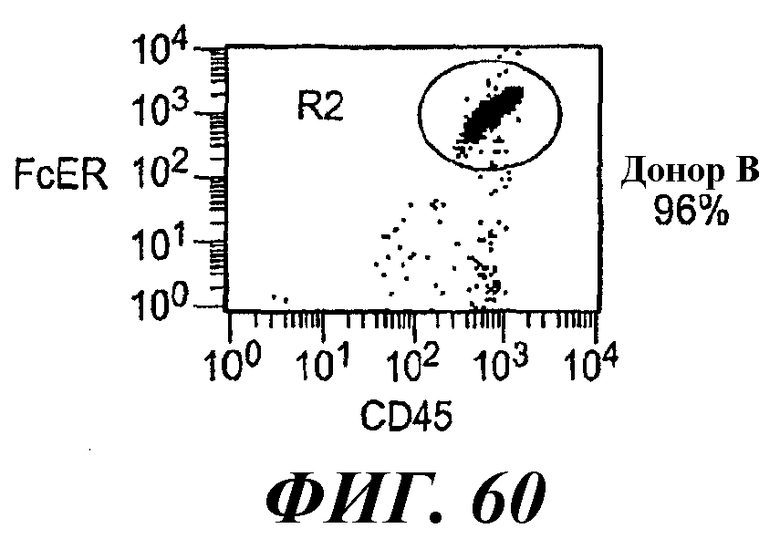
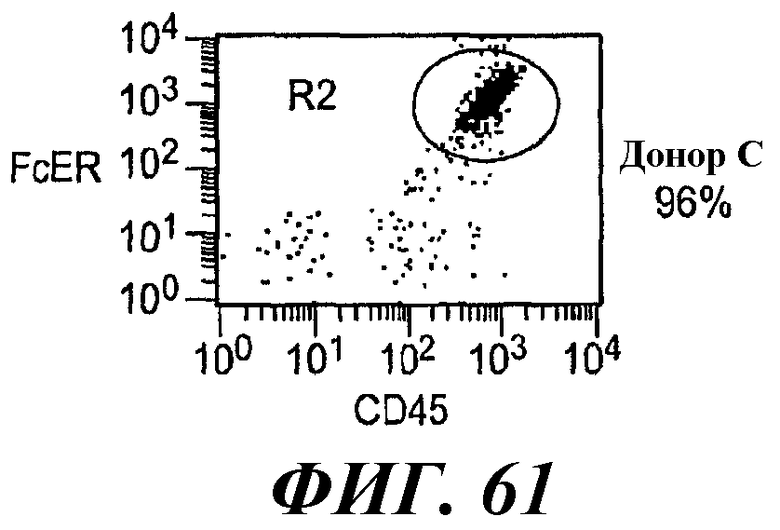
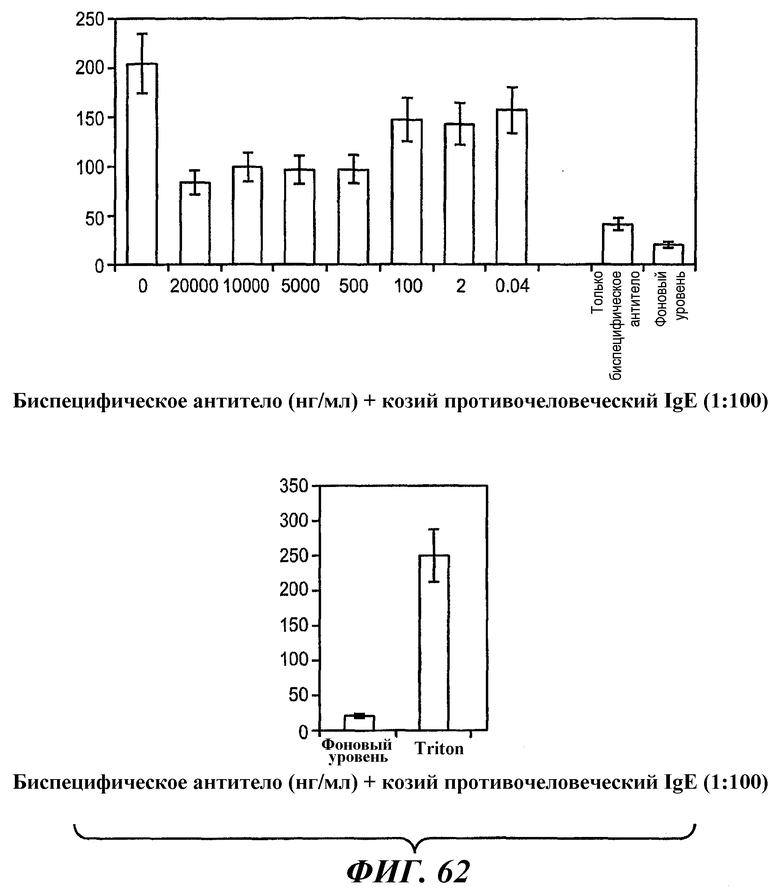

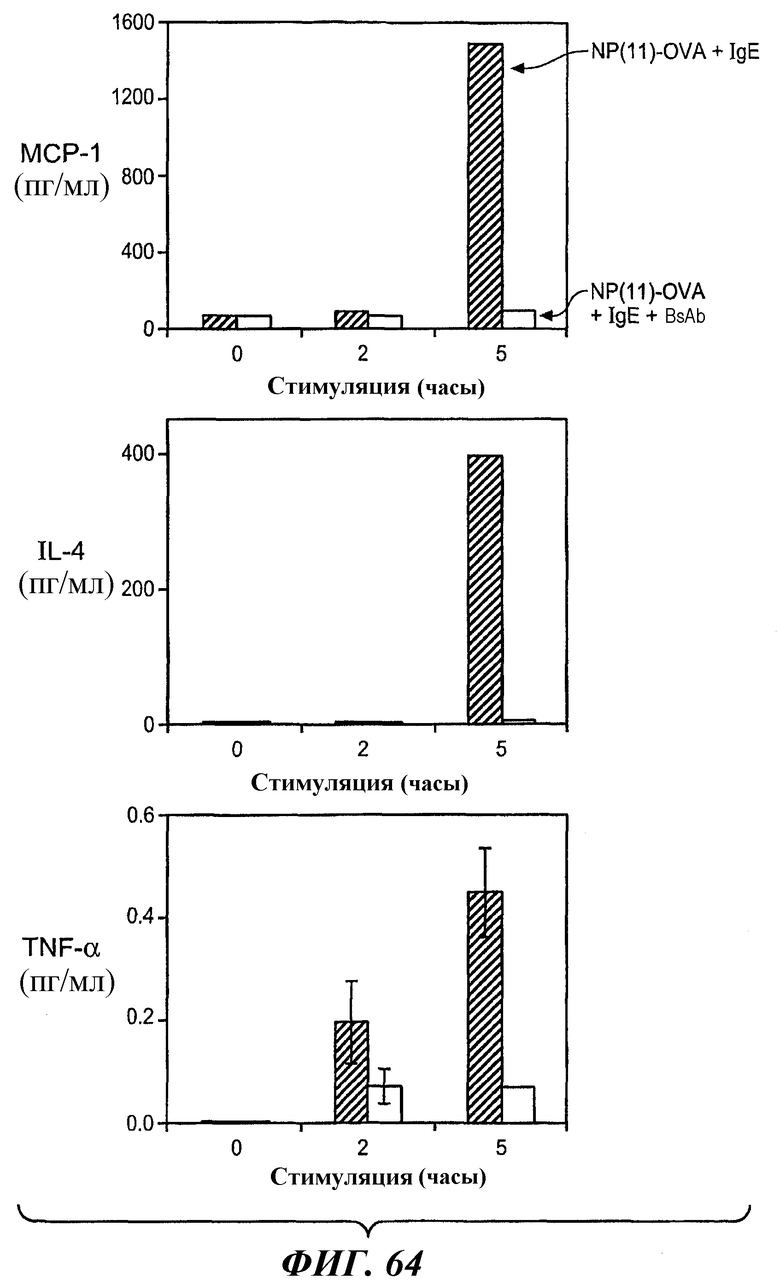

Изобретение относится к биотехнологии. Описано выделенное анти-FcγRIIB/анти-FcεRIα биспецифическое антитело, неспособное к перекрестному связыванию с FcγRIIA человека. Предложены способы ингибирования иммунного ответа у млекопитающего, подавления выброса гистамина, связанного с иммунным ответом у млекопитающего, и лечения, включающие введение описанного биспецифического антитела. Представлены композиция для подавления активности FcεRI в выделенных клетках млекопитающих, композиция, используемая для активации FcγRIIB в выделенных клетках млекопитающих, экспрессирующих FcγRIIB, путем совместной агрегации FcγRIIB с FcεRIα и композиция, использующаяся для ингибирования экспрессии рецептора FcεRI в выделенных клетках, содержащие в качестве активного вещества описанное антитело. Изобретение позволяет получать биспецифические анти-FcγRIIB/анти-FcεRIα антитела. 14 н. и 35 з.п. ф-лы, 65 ил., 2 табл.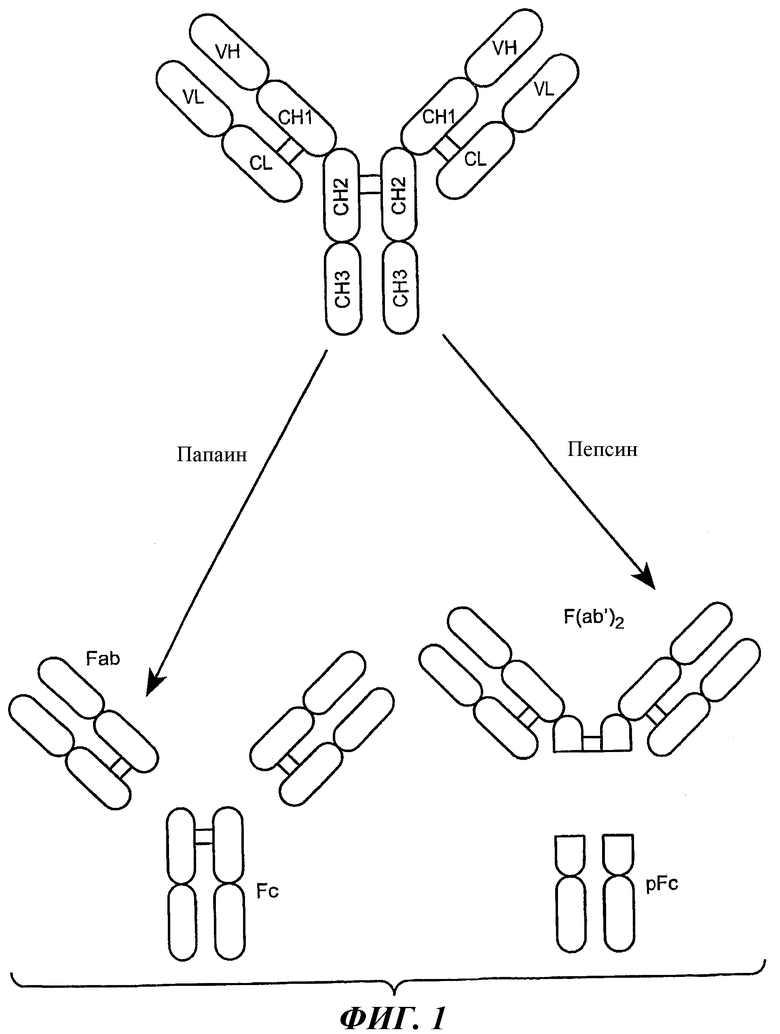
1. Выделенное анти-FcγRIIB/анти-FcεRIα биспецифическое антитело, неспособное к перекрестному связыванию с FcγRIIA человека, содержащее:
a) первый антигенсвязывающий полипептид, который селективно связывается с FcγRIIB и, по существу, неспособен перекрестно связываться с FcγRIIA, причем указанный антигенсвязывающий полипептид получен следующим образом:
(i) получают антитела FcγRIIB против белка экстрацеллюлярного домена FcγRIIB;
(ii) проводят первичный скрининг на антитела, специфичные в отношении FcγRIIB, путем положительного связывания с FcγRIIB,
(iii) проводят вторичный скрининг на специфические антитела против FcγRIIB путем связывания с FcγRIIB и отсутствия связывания с FcγIIA, и
b) второй антигенсвязывающий полипептид, который специфически связывается с FcεRIα.
2. Выделенное биспецифическое антитело по п.1, где второй антигенсвязывающий полипептид представляет собой моноклональное антитело, химерное антитело или гуманизированное антитело, или их фрагмент.
3. Выделенное биспецифическое антитело по п.1 или 2, где CDR 1, 2 и 3 тяжелой цепи первого антигенсвязывающего полипептида содержит SEQ ID NO: 1, 2 и 3 соответственно.
4. Выделенное биспецифическое антитело по п.1 или 2, где CDR 1, 2 и 3 легкой цепи первого антигенсвязывающего полипептида содержит SEQ ID NO: 4, 5 и 6 соответственно.
5. Выделенное биспецифическое антитело по п.1 или 2, где первый антигенсвязывающий полипептид обладает характеристиками антитела, продуцируемого клетками гибридомы с номером доступа в АТСС РТА-4614.
6. Выделенное анти-FcγRIIB/анти-FcεRIα биспецифическое антитело, содержащее первый антигенсвязывающий полипептид и второй антигенсвязывающий полипептид, где:
a) первый антигенсвязывающий полипептид, продуцируемый гибридомной клеточной линией с номером доступа в АТСС РТА-4614, или его функциональный фрагмент, или химерное антитело, или гуманизированное антитело, содержащее такой антигенсвязывающий полипептид или его антигенсвяызывающий фрагмент, селективно связывается с FcγRIIB; и
b) второй антигенсвязывающий полипептид специфически связывается с FcεRIα.
7. Выделенное биспецифическое антитело по п.6, где второй антигенсвязывающий полипептид представляет собой моноклональное антитело, химерное антитело, гуманизированное антитело или их фрагмент.
8. Выделенное биспецифическое антитело по п.6 или 7, где первый антигенсвязывающий полипептид или его фрагмент содержит CDR тяжелой или легкой цепи антитела, продуцируемого гибридомной клеточной линией с номером доступа в АТСС РТА-4614.
9. Выделенное биспецифическое антитело по п.6 или 7, где первый антигенсвязывающий полипептид, или его фрагмент, или второй антигенсвязывающий полипептид представляет собой фрагмент антитела, выбранный из группы, состоящей из Fab, Fab', Fab2, F(ab')2, Fd, Fd', scFv, scFv2, dAb.
10. Биспецифическое антитело по п.1 или 6, где первый антигенсвязывающий полипептид ковалентно связан со вторым антигенсвязывающим полипептидом.
11. Биспецифическое антитело по п.9, где первый и второй антигенсвязывающий полипептид ковалентно связан посредством линкера, содержащего по меньшей мере пять аминокислот.
12. Биспецифическое антитело по п.1 или 6, содержащее вариант шарнирной области тяжелой цепи, неспособный образовывать внутренние дисульфидные связи между тяжелыми цепями.
13. Способ ингибирования иммунного ответа у млекопитающего, включающий введение биспецифического антитела по любому из пп.1-12.
14. Способ подавления выброса гистамина, связанного с иммунным ответом у млекопитающего, включающий введение биспецифического антитела по любому из пп.1-12.
15. Способ по п.14, отличающийся тем, что выброс гистамина связан с аллергией, астмой или воспалением.
16. Применение биспецифического антитела по любому из пп.1-12 для получения лекарственного средства для ингибирования иммунного ответа у млекопитающего.
17. Применение биспецифического антитела по любому из пп.1-12 для получения лекарственного средства для подавления выброса гистамина, связанного с иммунным ответом у млекопитающих.
18. Применение по п.17, где выброс гистамина связан с аллергией, астмой или воспалением.
19. Композиция для подавления активности FcεRI в выделенных клетках млекопитающих, содержащая в качестве активного вещества антитело по любому из пп.1-12 в эффективном количестве.
20. Композиция по п.13, где клетки млекопитающих являются клетками человека.
21. Композиция, используемая для активации FcγRIIB в выделенных клетках млекопитающих, экспрессирующих FcγRIIB, путем совместной агрегации FcγRIIB с FcεRIα, содержащая эффективное количество биспецифического антитела по любому из пп.1-12.
22. Композиция по п.21, где совместная агрегация FcγRIIB и FcεRIα подавляет экспрессию FcεRI.
23. Композиция по п.21, где клетки представляют собой В-клетки или тучные клетки.
24. Композиция по п.21, где клетки млекопитающих являются клетками человека.
25. Композиция, использующаяся для ингибирования экспрессии рецептора FcεRI в выделенных клетках, содержащая эффективное количество биспецифического антитела по любому из пп.1-12.
26. Композиция по п.25, где клетка представляет собой клетку млекопитающего, имеющего расстройство, которое ослабляется при ингибировании экспрессии FcεRI в клетке.
27. Композиция по п.26, где указанное расстройство представляет собой хроническое расстройство.
28. Композиция по п.27, где указанное млекопитающее представляет собой человека.
29. Композиция по п.27, где указанное расстройство представляет собой атеросклероз, недостаточность лейкоцитарной адгезии, ревматоидный артрит, системную красную волчанку (SLE), сахарный диабет, рассеянный склероз, синдром Рейно, аутоиммунный тиреоидит, аллергический энцефаломиелит, синдром Шегрена и ювениальный диабет.
30. Композиция по п.25, которая улучшает лечение хронического расстройства, связанного с активностью FcεRI.
31. Композиция, содержащая анти-FcγRIIВ/анти-FcεRIα биспецифическое антитело по любому из пп.1-12 и фармацевтический носитель, для терапевтического применения в сочетании с анти-IgE антителом или анти-IgE-связывающим полипептидом для лечения аллергии, астмы и/или воспаления у млекопитающих.
32. Композиция по п.31, где анти-IgE антитело представляет собой омализумаб.
33. Набор, содержащий композицию по п.31, дополнительно содержащий этикетку с указанием, что биспецифическое антитело вводят в сочетании с анти-IgE антителом или анти-IgE-связывающим полипептидом для лечения аллергии, астмы и/или воспаления у млекопитающих.
34. Набор по п.33, где введение биспецифического антитела и анти-IgE антитела или анти-IgE связывающего полипептида проводят независимо друг от друга.
35. Набор по п.33, где введение биспецифического антитела и анти-IgE антитела или анти-IgE связывающего полипептида проводят одновременно.
36. Набор по п.33, где млекопитающим является человек.
37. Набор по п.33, где анти-IgE антитело представляет собой омализумаб.
38. Набор, содержащий композицию по п.31, дополнительно содержащий этикетку с указанием, что биспецифическое антитело вводят в сочетании с анти-IgE антителом или анти-IgE-связывающим полипептидом для лечения аллергии, астмы и/или воспаления у млекопитающих.
39. Набор по п.38, где млекопитающим является человек.
40. Набор по п.38, где анти-IgE антитело представляет собой омализумаб.
41. Способ лечения, включающий введение анти-FcγRIIB/анти-FcεRIα биспецифического антитела по любому из пп.1-12 в сочетании с анти-IgE антителом или анти-IgE-связывающим полипептидом млекопитающему, имеющему расстройство, выбранное из группы, состоящей из аллергии, астмы и воспаления.
42. Способ по п.41, отличающийся тем, что введение биспецифического антитела и анти-IgE антитела или анти-IgE-связывающего полипептида проводят отдельно друг от друга.
43. Способ по п.41, отличающийся тем, что введение биспецифического антитела и анти-IgE антитела или анти-IgE-связывающего полипептида проводят одновременно.
44. Способ по п.41, отличающийся тем, что указанным млекопитающим является человек.
45. Способ по п.41, отличающийся тем, что анти-IgE антитело представляет собой омализумаб.
46. Применение анти-FcγRIIB/анти-FcεRIα биспецифического антитела по любому из пп.1-12 в сочетании с анти-IgE антителом или анти-IgE-связывающим полипептидом для получения лекарственного средства для лечения млекопитающего, имеющего расстройство, выбранное из группы, состоящей из аллергии, астмы и воспаления.
47. Применение по п.46, где анти-FcγRIIB/анти-FcεRIα биспецифическое антитело не способно к перекрестному взаимодействию с рецептором FcγRIIA.
48. Применение по п.46, где млекопитающее является человеком.
49. Применение по п.46 или 48, где анти-IgE антитело представляет собой омализумаб.
| ТАМ S ЕТ AL: "A bispecific antibody against human IgE and human FcgammaRII that inhibits antigen-induced histamine release by human mast cells and basophils", ALLERGY (OXFORD), vol.59, no.7, July 2004, p.772-780 | |||
| ZHU DAOCHENG ЕТ AL: "A novel human immunoglobulin Fcgamma-Fcepsilon bifunctional fusion protein inhibits FcepsilonRI-mediated |

