Область техники, к которой относится изобретение
Изобретение относится к области медицины, в частности к медицинскому препарату для модуляции, стимуляции роста аксонов нейронов центральной нервной системы после повреждения.
Уровень техники
После периода раннего детства повреждение центральной нервной системы (ЦНС) приводит к функциональным нарушениям, которые в большинстве случаев являются необратимыми. Повреждения в головном мозге или спинном мозге, являющиеся результатом инсульта, травмы или иных случаев, могут привести к пожизненным потерям познавательной, сенсорной и двигательной функций и даже поддержания жизненных функций. Утраченные нервные клетки не восстанавливаются, а резервные клетки обычно не обладают способностью к возобновлению разорванных связей, хотя локальная синаптическая реорганизация в ограниченном количестве может происходить вблизи области повреждения. Утраченные функции в настоящее время являются неизлечимыми.
Невозможность регенерации в ЦНС относили на счет ряда факторов, которые включают присутствие молекул-ингибиторов на поверхности клеток глии, которые супрессируют вырастание аксонов; отсутствие подходящей субстратной молекулы, такой как ламинин, которая способствует росту, и отсутствие подходящих трофических факторов, необходимых для активации программ экспрессии генов, требующихся для выживания и дифференциации клеток.
Напротив, в периферической нервной системе (ПНС) поврежденные нервные волокна могут повторно отрастать на большие расстояния с возможным при соответствующих условиях отменным восстановлением функции. В течение последних 15 лет специалисты в области нейронаук пришли к осознанию того, что данный факт не является следствием внутренних различий между нервными клетками периферической и центральной нервной системы, примечательно, что нейроны ЦНС будут вытягивать свои аксоны на большие расстояния, если получат возможность расти через трансплантированный сегмент ПНС (например, седалищного нерва). Вследствие этого нейроны ЦНС сохраняют способность к росту, если получают соответствующие верные сигналы из внеклеточной среды. Факторы, которые способствуют различию потенциалов роста ЦНС и ПНС включают частично охарактеризованные ингибирующие рост молекулы на поверхности олигодендроцитов, которые окружают нервные волокна в ЦНС, но которые менее распространены в популяции сравнимых клеток ПНС (шванновские клетки); молекулы на базальной пластинке и других поверхностях, которые способствуют росту в ПНС, но отсутствуют в ЦНС (например, ламинин), а также трофические факторы, растворимые полипептиды, которые активируют программы экспрессии генов, лежащие в основе выживания и дифференциации клеток. Хотя данные трофические факторы считают основными для поддержания жизнеспособности и дифференциации нервных клеток, определенные факторы, которые являются ответственными за индукцию аксонной регенерации в ЦНС, остаются точно неизвестными. В результате этого в настоящее время не разработаны эффективные способы лечения повреждений ЦНС.
Соответственно, способы и композиции для модуляции роста нейронов ЦНС являются все еще необходимыми.
Сущность изобретения
Данное изобретение представляет способы и композиции для модуляции вырастания аксонов нейронов центральной нервной системы, в частности нейронов центральной нервной системы млекопитающих. Изобретение основано, по крайней мере, отчасти на обнаружении того факта, что пуриновые нуклеозиды и их аналоги являются способными к модуляции (т.е. либо стимуляции, либо ингибированию) вырастания аксонов нейронов ЦНС, включая нейроны ЦНС млекопитающих, такие как нейроны ганглия сетчатки. Более того, пуриновые нуклеозиды и их аналоги, соответствующие изобретению, являются эффективными при модулировании аксонного роста нейронов ЦНС в отсутствие каких-либо дополнительных модуляторов роста нейронов (таких как фактор роста нервов).
Согласно этому, способы, соответствующие изобретению, в основном включают контактирование нейронов центральной нервной системы с пуриновым нуклеозидом или его аналогом. В одном аспекте изобретение представляет способы стимуляции вырастания предпочтительно с использованием инозинового или гуанозинового нуклеозидов или их аналогов. В другом аспекте изобретение представляет способы ингибирования вырастания предпочтительно с использованием 6-тиогуанина. В особенно предпочтительном варианте осуществления способы, соответствующие изобретению, модулируют рост аксонов клеток ганглия сетчатки.
Соответствующие изобретению способы стимуляции вырастания (восстановления) аксонов нейронов центральной нервной системы могут быть использованы после разрушения или другого повреждения нейронов ЦНС (например, инсульта, черепно-мозговой травмы, церебральной аневризмы, повреждения спинного мозга и т.п.). Соответствующие изобретению способы ингибирования вырастания аксонов нейронов ЦНС могут быть использованы при нейропролиферативных нарушениях, при которых может происходить аберрантный аксонный рост, таких как эпилепсия (например, посттравматическая эпилепсия) и невропатический болевой синдром.
В одном аспекте пуриновый нуклеозид или его аналог вводят субъекту в соответствии с данным изобретением путем интродукции в центральную нервную систему субъекта, например в спинномозговую жидкость субъекта. В ряде аспектов изобретения пуриновый нуклеозид или его аналог вводят подоболочечно, например, в желудочек головного мозга, поясничную область или большую цистерну. В предпочтительном варианте осуществления способ стимуляции, соответствующий изобретению, способствует росту поврежденных клеток ганглия сетчатки. В данных обстоятельствах пуриновый нуклеозид или его аналог могут быть введены локально в клетки ганглия сетчатки для стимуляции вырастания аксонов.
В еще одном варианте осуществления пуриновый нуклеозид или его аналог вводят в фармацевтически приемлемом препарате. Фармацевтически приемлемый препарат может представлять собой дисперсионную систему, например препарат на основе липидов, липосомный препарат или мультивезикулярный липосомный препарат. Фармацевтически приемлемый препарат может также содержать полимерный материал, выбранный, например, из синтетических полимеров, таких как полиэфиры (полилактид, полимер на основе молочной и гликолевой кислот), полиэтиленгликоль, полиоксомеры, полиангидриды и полимеры на основе плуроновой кислоты, или выбранных из естественным путем выделенных полимеров, таких как альбумин, альгинат, производные целлюлозы, коллаген, фибрин, желатин и полисахариды.
В предпочтительном варианте осуществления фармацевтически приемлемый препарат обеспечивает задержанную доставку, например "медленное высвобождение" пуринового нуклеозида в организме субъекта в течение, по меньшей мере, одной недели, более предпочтительно, по меньшей мере, одного месяца после введения фармацевтически приемлемого препарата субъекту. Предпочтительные подходы к получению задержанной доставки препарата, соответствующего изобретению, включают применение полимерных капсул с медленным выходом или инфузионного насоса, который содержит препарат.
Изобретение, кроме того, охватывает применение пуринового нуклеозида или его аналога в приготовлении лекарственного препарата для модуляции вырастания аксонов нейронов центральной нервной системы, предпочтительно нейронов ЦНС млекопитающих. В предпочтительном варианте осуществления данный лекарственный препарат не включает другие модуляторы роста нейронов, отличные от пуринового нуклеотида или его аналогов. Например, в одном варианте осуществления лекарственный препарат не включает фактор роста нервов.
Изобретение представляет также фармацевтические композиции и упакованные препараты, содержащие пуриновый нуклеозид или его аналог, соответствующие изобретению, а также фармацевтически приемлемый носитель.
Другие признаки и преимущества изобретения станут очевидными из последующего подробного описания и формулы изобретения.
На Фигурах 1.1-1.4 представлены графики, показывающие количественное определение пуринергических эффектов на вырастание аксонов.
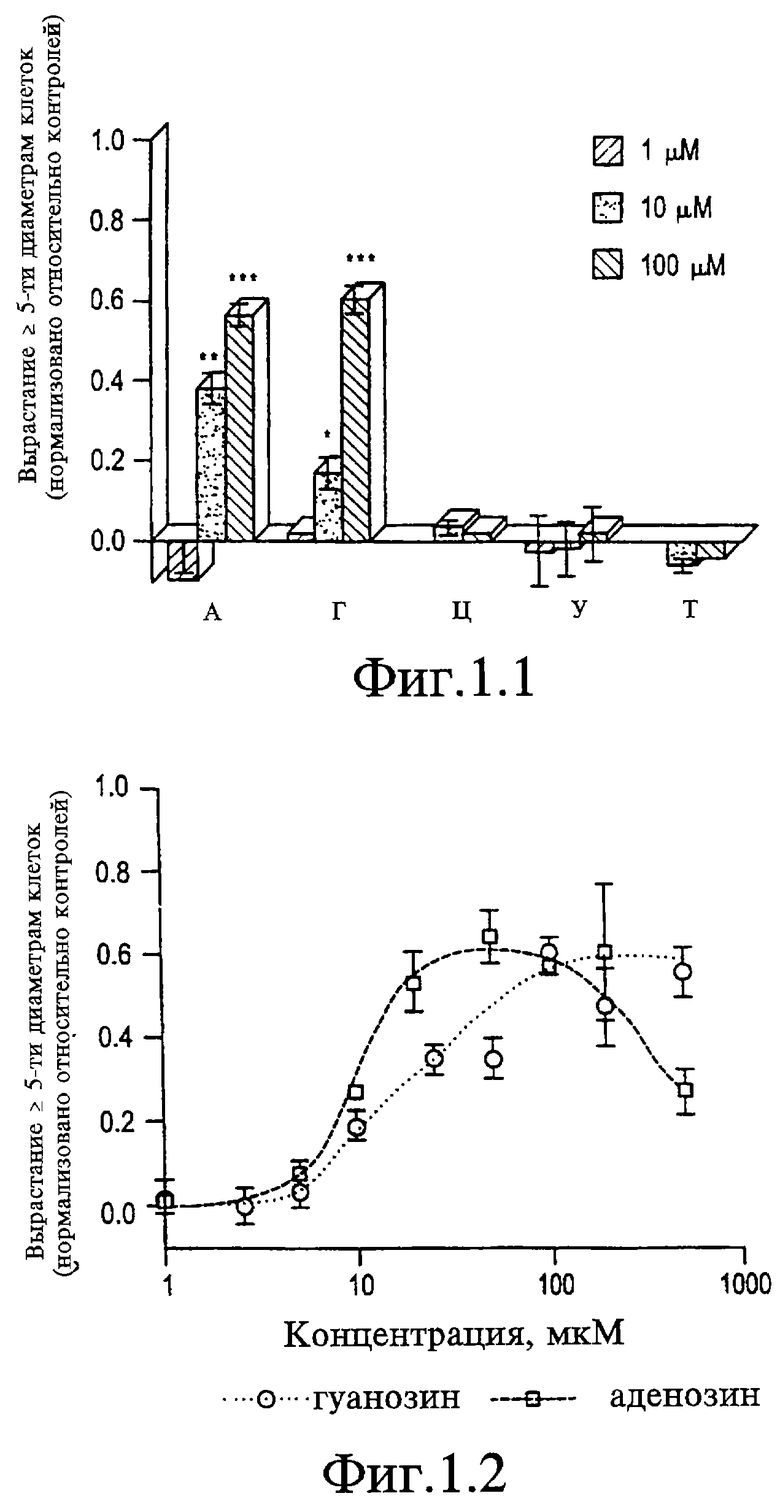
На Фигуре 1.1 представлен график, показывающий рост аксонов в ответ на нуклеозиды аденозин (А), гуанозин (Г), цитидин (Ц), уридин (У) и тимидин (Т) при концентрации 1, 10 и 100 мкМ, как указано. Данные нормализованы вычитанием уровня роста в отрицательных контролях с последующим делением на значение роста сети в положительных контролях, обработанных 20-30% АФ-1 (аксогенезисный фактор - 1).
На Фигуре 1.2 представлен график, показывающий кривые доза-зависимости для аденозина и гуанозина. Значения EC50, полученные на основании данных результатов, составляют 10-15 мкМ для аденозина и 20-30 мкМ для гуанозина.
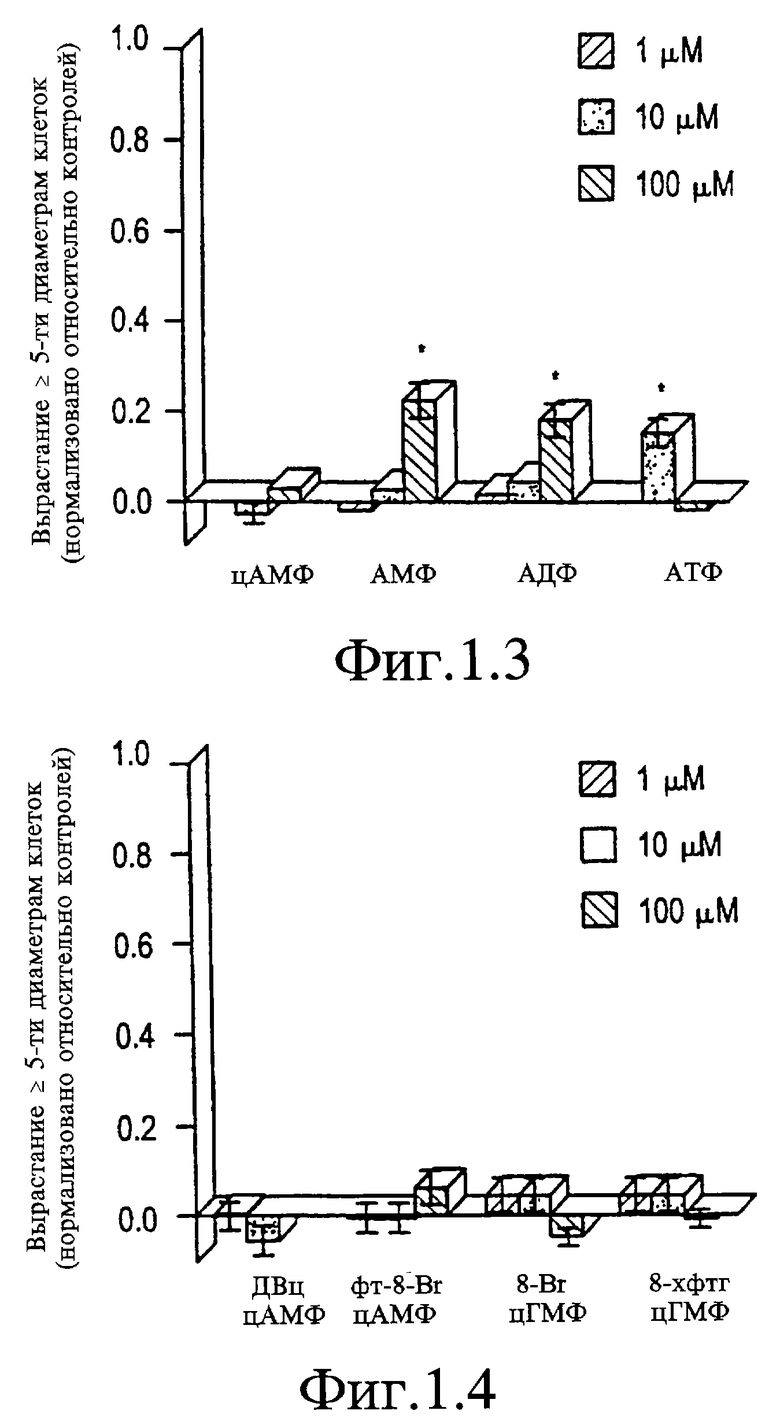
На Фигуре 1.3 представлен график, показывающий эффекты аденозиновых нуклеотидов.
На Фигуре 1.4 представлен график, показывающий эффекты мембранопроницаемых аналогов циклического АМФ (дибутирил циклического АМФ (ДБ-цАМФ)), 8-бромаденозин-3',5'-циклического монофосфортиоата (8-Вг-АцМФТ) или циклического ГМФ (8-бром-циклического ГМФ (8-Вr-цГМФ)); 8-(4-хлорфенилтио)-гуанозин-3', 5'-циклического монофосфата (8-ХФТГ-цМФ)). Данные представляют средние значения + стандартные ошибки среднего значения (SEM, не приведено, если <0,02) и объединены по 2-4 независимым экспериментам. Значения р при сравнении роста с отрицательными контролями основываются на двусторонних t-критериях. *р<0,05; **р<0,01; ***р<0,001.
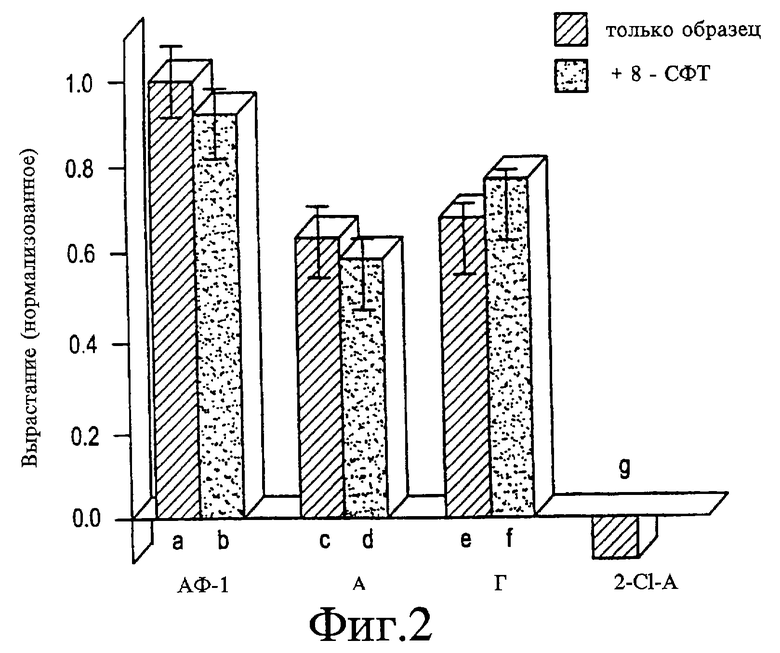
На Фигуре 2 представлен график, показывающий, что аденозин не стимулирует рост посредством экстрацеллюларных рецепторов. На вырастание, стимулируемое АФ-1 (а-b), 100 мкМ аденозина (A) (c-d) или 100 мкМ гуанозина (Г) (e-f), не влияет добавление 20 пМ 8-р-(сульфофенилтеофиллина) (СФТ), ингибитора аденозиновых рецепторов А1 и А2 (ср. рост в а, с и е с ростом в b, d и f). Негидролизующийся аналог аденозина, 2-хлораденозин ((2-Cl-A), 100 мкМ) снижает рост до значений ниже базовых уровней (g) (р<0,001 в 3 экспериментах).
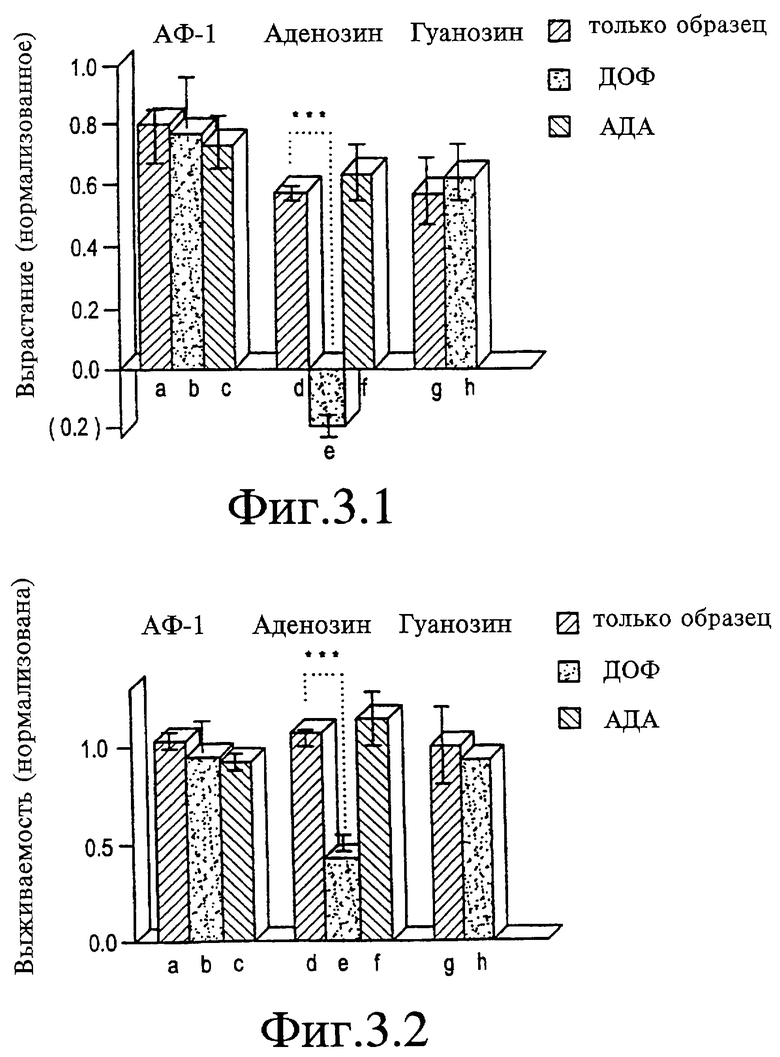
На Фигурах 3.1 и 3.2 представлен график, показывающий, что аденозин должен быть гидролизован для того, чтобы стимулировать рост. На Фигуре 3.1 представлен график, показывающий эффекты дезоксикоформицина (ДОФ) и экзогенной аденозиндеаминазы (АДА) на рост, индуцируемый АФ-1 (а-с), аденозином (d-f) и гуанозином (g, h). На Фигуре 3.2 представлен график, показывающий эффекты дезоксикоформицина (ДОФ) и экзогенной аденозиндеаминазы (АДА) на выживаемость, индуцируемую АФ-1 (а-с), аденозином (d-f) и гуанозином (g, h). В то время как проведение гидролиза аденозина экзогенной АДА сохраняет активность аденозина неизменной (f), блокирующая активность эндогенной АДА с ДОФ вызывает супрессирующее действие аденозина на рост (е, Фигура 3.1) и выживаемость (е, Фигура 3.2). ***р<0,001.
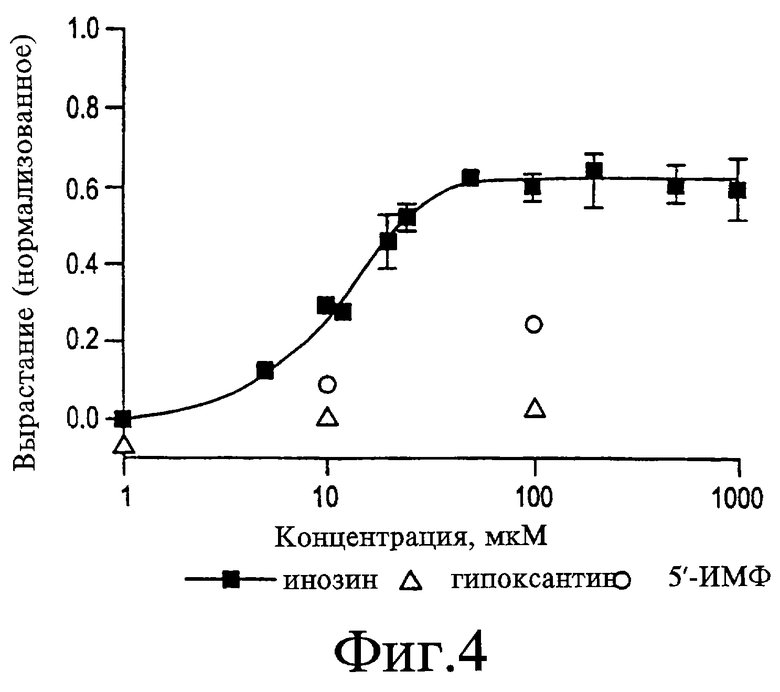
На Фигуре 4 представлен график, показывающий кривые доза-зависимости для инозина. При концентрациях свыше 50 мкМ инозин стимулирует приблизительно 60% от максимального уровня роста, достигаемого при использовании АФ-1. Значение EC50 (EC-эффективная доза) для инозина определяют как 10-15 мкМ. Гипоксантин был неактивным, тогда как 5'-ИМФ (инозинмонофосфат), вероятно, обладает менее чем 1/10 активности инозина. Рост, стимулируемый всеми концентрациями инозина более 10 мкМ, значительно превышает фоновые значения (р<0,001).
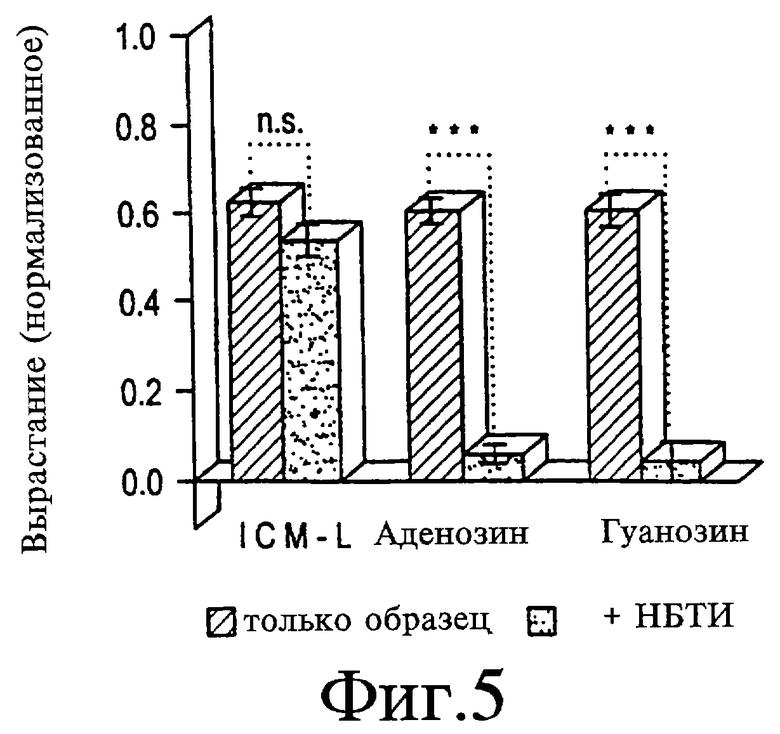
На Фигуре 5 представлен график, показывающий, что инозин и гуанозин стимулируют рост посредством внутриклеточного механизма. При концентрации 20 мкМ NBTI ингибитор транспорта пуринов не влияет на активность АФ-1, но блокирует приблизительно 90% активности инозина (50 мкМ) или гуанозина (100 мкМ). *** различия в росте с добавлением или без добавления лекарств являются значимыми при р<0,001. Объединены данные по 4 независимым экспериментам.
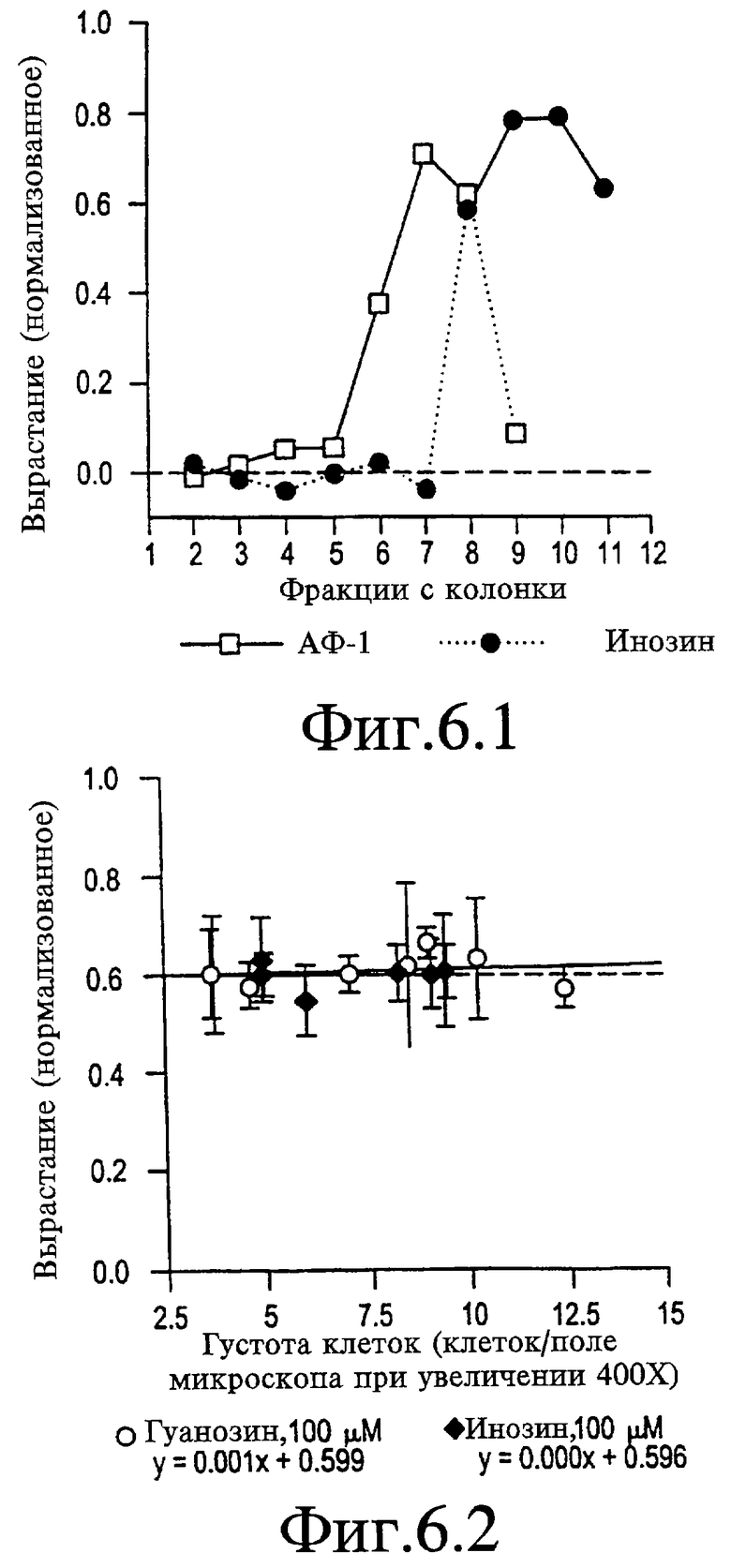
На Фигуре 6-1 представлен график, показывающий, что АФ-1 не обнаруживает активности инозина. На колонке Sephadex G-10 АФ-1 элюируется с пиком 7 минут при отсутствии определяемой активности во время элюции пика инозина (т.е. 9-10 мин).
На Фигуре 6-2 представлен график, показывающий, что эффекты инозина и гуанозина не зависят от густоты клеток. Данные, полученные во множестве независимых экспериментов, каждый из которых указан одной точкой, были проанализированы на эффект наложения густоты на рост клеток. Во всех случаях концентрацию инозина или гуанозина поддерживали на уровне 100 мкМ. Линии регрессии были вычислены выравниванием по методу наименьших квадратов (График Крикета) и представлены ниже обозначений.
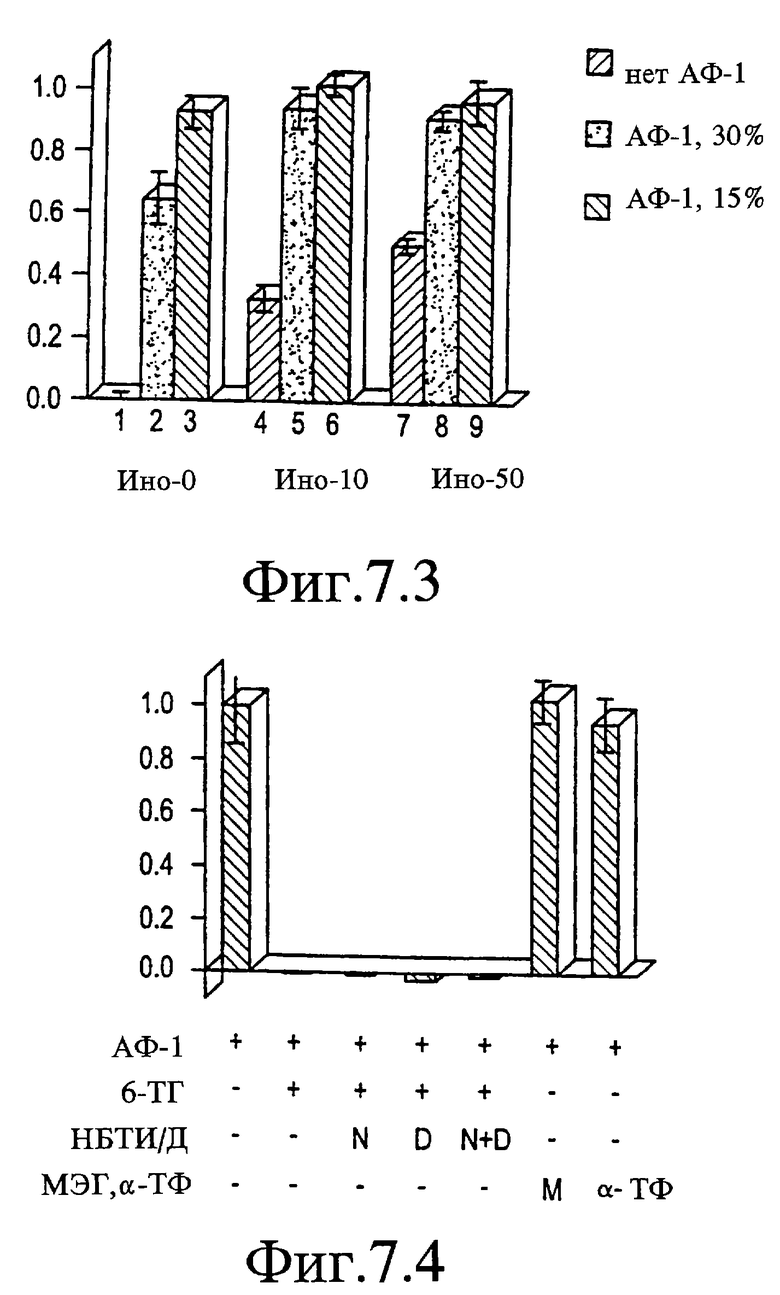
На Фигурах 7.1-7.4 представлены графики, показывающие, что эффекты АФ-1 ингибируются 6-тиогуанином (6-ТГ), но восстанавливаются инозином.
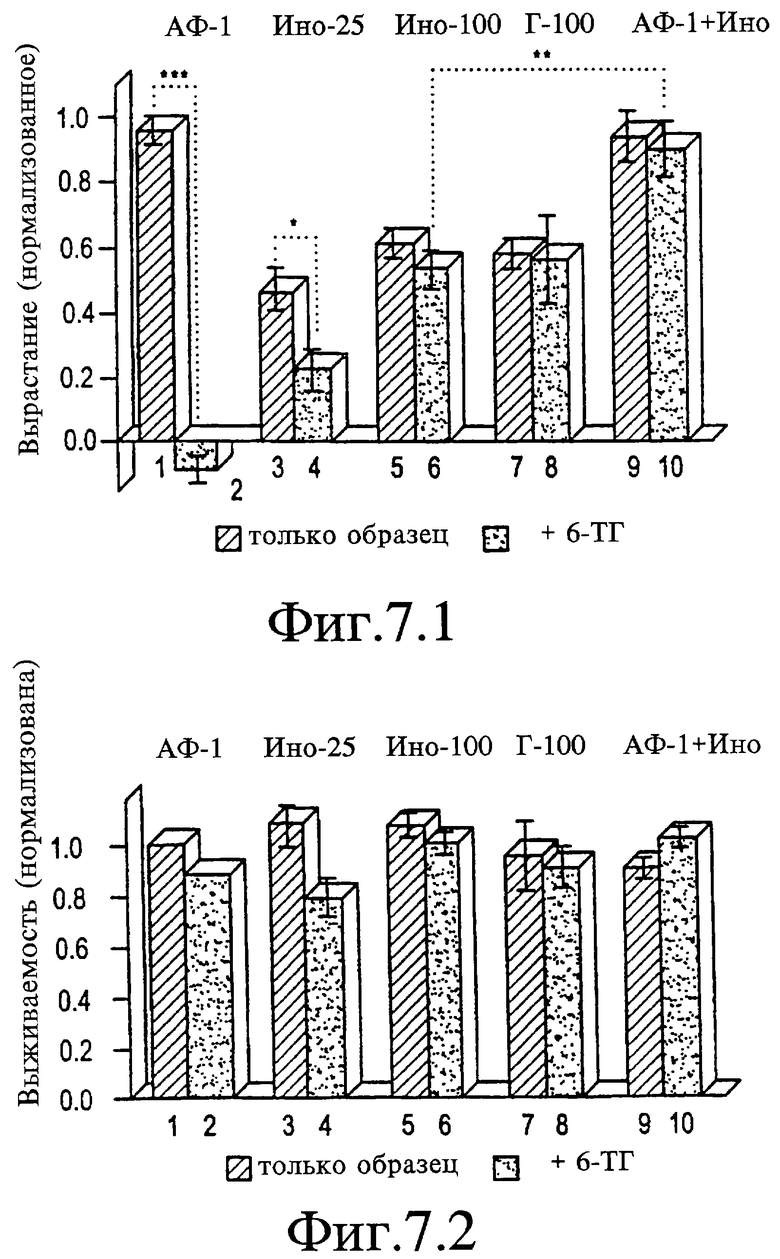
На Фигуре 7.1 показано, что при концентрации 10 мкМ пуриновый аналог 6-ТГ супрессировал рост, индуцированный АФ-1, до уровня ниже базового (дорожка 2 относительно 1, р<0,001) и снижал рост, индуцированный 25 мкМ инозина (ИНО-25) приблизительно на 50% (дорожка 4 относительно 3). Рост, индуцированный более высокими концентрациями инозина или гуанозина (Г-100: дорожки 8 относительно 7), не был подвержен воздействию. Инозин в концентрации 100 мкМ восстанавливал весь рост, индуцированный АФ-1, в присутствии 10 мкМ 6-ТГ (дорожка 10), который является значительно более высоким, чем рост, индуцированный 100 мкМ инозина, как в виде монокомпонента, так и в комбинации с 10 мкМ 6-ТГ (р<0,01).
На Фигуре 7.2 представлен график, показывающий, что концентрация 6-ТГ, используемая в данных экспериментах, не оказывала воздействия на жизнеспособность клеток.
На Фигуре 7.3 представлен график, показывающий, что АФ-1 и инозин обладают частичными аддитивными эффектами. Рост оценивали для АФ-1 и инозина (каждый при концентрации 0, ЕС50 или насыщающей концентрации). Поскольку эффекты полумаксимальных концентраций каждого компонента были аддитивными (дорожка 5), рост достигал уровня плато в присутствии более высоких концентраций каждого компонента (дорожки 6, 8, 9).
На Фигуре 7.4 представлены дальнейшие исследования эффектов 6-тиогуанина. Рост, стимулированный АФ-1, был полностью блокирован 6-ТГ (10 мкМ) и не восстанавливался в присутствии нитробензилтиоинозина (НБТИ или N, 20 мкМ) и/или дипиридамола (D, 10 мкМ), ингибиторов блокаторов транспорта пуринов, которые супрессируют активность инозина. Эффекты ингибирования 6-ТГне имитировались двумя восстанавливающими агентами, α-токоферолом (α-ТФ, 30 мкМ) или α-метиловым сложным эфиром глутатиона (МЭГ, 100 мкМ).
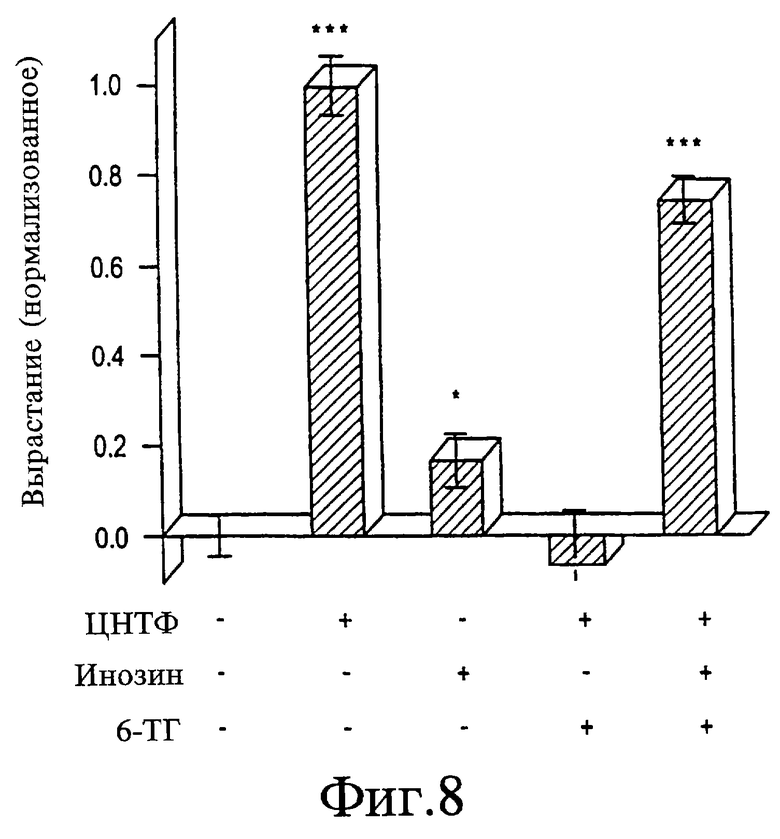
На Фигуре 8 представлен график, показывающий эффекты пуринов на клетки ганглия сетчатки крыс (количественные исследования). Рост, стимулированный цилиарным нейротрофическим фактором (ЦНТФ), ингибируется 6-ТГ (10 мМ), но полностью восстанавливается при добавлении 25 мкМ инозина. Значимость отличий от контроля составляет: *р=0,03; ***р<0,001. Объединены результаты по 3 независимым исследованиям.
Сведения, подтверждающие возможность осуществления изобретения
Данное изобретение представляет способы модуляции вырастания аксонов нейронов центральной нервной системы (ЦНС) и, в частности, нейронов ЦНС млекопитающих. Изобретение основано, по крайней мере, частично на обнаружении того, что ряд пуриновых нуклеозидов (например, инозин и гуанозин), а также их аналоги индуцируют стимуляцию вырастания аксонов из клеток ганглия сетчатки как серебряного карася, так и млекопитающих (см. Примеры 1 и 11 соответственно). Кроме того, изобретение основано, по крайней мере, частично на обнаружении того, что другие пуриновые нуклеозиды, такие как аденозиновые нуклеозиды и их аналоги, индуцируют ингибирование вырастания аксонов из клеток ганглия сетчатки (см. Пример 10). Как показано в Примере 2, пуриновые нуклеозиды являются более активными, чем их нуклеотидные варианты, и они проявляют свое действие с использованием внутриклеточных путей (см. Пример 6). Более того, превращение аденозина в инозин путем деамидирования (например, эндогенной аденозиндеаминазой) приводит к стимуляции вырастания аксонов, тогда как блокада деамидирования аденозина приводит к ингибированию вырастания аксонов (см. Пример 4). В дополнение к этому данный эффект пуриновых нуклеозидов или их аналогов на рост аксонов не требует присутствия других модуляторов роста нейронов (таких как фактор роста нервов).
Согласно этому, соответствующие изобретению способы модуляции вырастания аксонов нейронов ЦНС в основном включают контактирование нейронов центральной нервной системы с пуриновым нуклеозидом или его аналогом таким образом, что происходит модуляции вырастания аксонов.
В предпочтительных вариантах осуществления способы, соответствующие изобретению, используют для стимуляции (например, с использованием инозина или гуанозина) вырастания аксонов нейронов центральной нервной системы после повреждения, такого как, например, инсульт, черепно-мозговая травма, церебральная аневризма или повреждение спинного мозга.
В других предпочтительных вариантах осуществления способы, соответствующие изобретению, используют для ингибирования (например, с использованием 6-тиогуанина) вырастания аксонов нейронов ЦНС при нейропролиферативных нарушениях, например, когда может происходить аберрантный или избыточный рост аксонов, таких как эпилепсия или невропатические болевые нарушения. Было показано, что при эпилепсии, например посттравматической эпилепсии, быстрое вырастание поврежденных аксонов пирамидальных нейронов приводит в формированию повторных возбуждающих синапсов и гипервозбудимой нервной сети (см. статьи Prince D. A. и соавт., Nature Medicine, 3:957-958, (1997) и McKinney R. A. и соавт. , Nature Medicine, 3:990-996, (1997)). Более того, невропатический болевой синдром был связан с нежелательным разрастанием нервных окончаний (описано, например, в статье Woolf C.J. и соавт., Nature, 306:686-688, (1983)).
Как используют в данном контексте, выражение "модуляция вырастания аксонов нейронов центральной нервной системы" предусматривает включение способности стимулировать или ингибировать рост аксонов нейронов центральной нервной системы до различных уровней, например до уровней, которые позволяют лечить целевые повреждения ЦНС.
Как используют в данном контексте, термин "рост" ("вырастание") (т.е. восстановление аксонов) относится к процессу, при котором аксоны вырастают из нейрона ЦНС. Вырастание может приводить к образованию полностью нового аксона или восстановлению частично поврежденного аксона. Рост обычно бывает засвидетельствован процессом вытягивания аксонов на, по меньшей мере, 5 диаметров клеток в длину. Более того, вырастание аксонов может быть подтверждено экспрессией GAP-43 (которая может быть определена, например, иммунным окрашиванием).
Как используют в данном контексте, термин "нейроны ЦНС" предусматривает включение нейронов головного мозга и спинного мозга, которые являются нечувствительными к фактору роста нервов (NGF). Термин не предназначен для включения поддерживающих или защитных клеток, таких как астроциты, олигодендроциты, клетки микроглии, эпендимы и т.п., как и не предназначен для включения нейронов периферической нервной системы (например, соматической, вегетативной, симпатической или парасимпатической нервной системы). Предпочтительными нейронами ЦНС являются нейроны млекопитающих, более предпочтительными - нейроны человека.
Как используют в данном контексте, выражение "контактирование" предусматривает включение способов как in vivo, так и in vitro приведения пуринового нуклеозида или его аналога на близкое расстояние от нейрона ЦНС, так что пуриновый нуклеозид или его аналог могут модулировать процессы вырастания аксонов из указанного нейрона ЦНС.
Как используют в данном контексте, термин "пуриновый нуклеозид" известен в области техники и предусматривает включение любого пуринового основания, связанного с сахаром, или его аналога. Например, пуриновые нуклеозиды включают гуанозин, инозин или аденозин и аналоги включают 6-тиогуанин (6-ТГ) и т. п. Как используют в данном контексте, термин "аналог" пуринового нуклеозида относится к соединению, которое сохраняет химические структуры пуринового нуклеозида, необходимые для функциональной активности, такие как пуриновое кольцо, связанное с сахаром, но которые содержат также некоторые химические структуры, не обнаруженные в существующих в естественных условиях пуриновых нуклеозидах, такие как модификация боковой группы (например, тиогруппа или группа хлора).
В одном варианте осуществления вырастание аксонов нейронов ЦНС стимулируют, предпочтительно используя инозиновый или гуанозиновый нуклеозиды или их аналоги. В другом варианте осуществления вырастание аксонов нейронов ЦНС ингибируют, предпочтительно используя 6-тиогуанин. Аденозин действует как пуриновый нуклеозид - ингибитор, но под действием аденозиндеаминазы превращается в инозин, который является пуриновым нуклеозидом - стимулятором. Соответственно, аденозин может быть использован в качестве пуринового нуклеозида - стимулятора в случаях деамидирования с образованием инозина (например, в присутствии эндогенной активности аденозиндеаминазы). Альтернативно, в ситуациях, когда активность аденозиндеаминазы блокирована, аденозин может быть использован как пуриновый нуклеозид - стимулятор. Аналог аденозина, 2-хлораденозин, также может быть использован в качестве нуклеозида-ингибитора, хотя его другие эффекты, такие как воздействие на А1-, А2- и/или A3-рецепторы может делать его менее предпочтительным для применения in vivo.
Изобретение представляет также способы стимуляции вырастания нейронов центральной нервной системы после повреждений. Способ включает введение субъекту пуринового нуклеозида (например, инозина или гуанозина) или его аналога.
Как используют в данном контексте, термин "субъект" предусматривает включение животных, чувствительных к повреждениям ЦНС, предпочтительно млекопитающих, наиболее предпочтительно человека. В предпочтительном варианте осуществления субъект представлен приматом. В еще более предпочтительном варианте осуществления примат является человеком. Другие примеры субъектов включают собак, кошек, коз и коров.
Как используют в данном контексте, термин "повреждение" предусматривает включение повреждения, которое прямо или косвенно влияет на нормальное функционирование ЦНС. Например, повреждение может быть повреждением клеток ганглия сетчатки, черепно-мозговой травмой, повреждением, связанным с инсультом, повреждением, связанным с церебральной аневризмой, повреждением спинного мозга, включая моноплегию, диплегию, параплегию и квадриплегию; нейропролиферативным нарушением, эпилепсией, например, посттравматическим повреждением головного мозга, или невропатическим болевым синдромом.
Как используют в данном контексте, термин "инсульт" известен в области техники и предусматривает включение внезапного снижения или потери сознания, ощущений и произвольных движений, вызванных закупоркой или обструкцией (например, сгустком крови) артерии головного мозга.
Как используют в данном контексте, термин "черепно-мозговая травма" известен в области техники и предусматривает включение состояния, при котором травматический удар головы вызывает повреждение головного мозга или связанного с ним спинного мозга, часто не затрагивая череп. Обычно исходная травма может приводить к распространению гематомы, субарахноидальному кровотечению, отеку головного мозга, повышенному внутричерепному давлению (ICP) и гипоксии головного мозга, которые в свою очередь приводят к тяжелым вторичным событиям, обусловленным низким кровотоком в головном мозге (CBF).
Фармацевтически приемлемые препараты
В способе, соответствующем изобретению, пуриновый нуклеозид или его аналог могут быть введены в фармацевтически приемлемом препарате. Данный фармацевтически приемлемый препарат обычно включает пуриновый нуклеозид или его аналог, а также фармацевтически приемлемый носитель(и) и/или наполнитель(и). Как используют в данном контексте, выражение "фармацевтически приемлемый носитель" включает любой и все растворители, дисперсионную среду, покрытия, антибактериальные и противогрибковые агенты, изотонические или задерживающие всасывание агенты и т.п., которые являются физиологически совместимыми. Например, носитель может быть пригодным для инъекции в спинномозговую жидкость. Наполнители включают фармацевтически приемлемые стабилизаторы и дезинтегрирующие агенты. Данное изобретение касается всех фармацевтически приемлемых препаратов, включая синтетические или природные полимеры в форме макромолекулярных комплексов, нанокапсул, микросфер или частиц и препаратов на липидной основе, включая эмульсии типа масло в воде, мицеллы, смешанные мицеллы, синтетические мембранные пузырьки и конъюгированные эритроциты.
В одном варианте осуществления фармацевтически приемлемые препараты содержат полимерный матрикс.
Термины "полимер" или "полимерный" известны в области техники и предусматривают включение структурного скелета, состоящего из повторяющихся мономерных структур, способных доставлять пуриновый нуклеозид или его аналог, так что осуществляется лечение целевого состояния, например повреждения ЦНС. Термины включают также сополимеры и гомополимеры, например синтетические или существующие в естественных условиях. Предполагается включение также линейных полимеров, разветвленных полимеров и перекрестно сшитых полимеров.
Например, полимерные материалы, пригодные для образования фармацевтически приемлемого препарата, используемого в данном изобретении, включают выделенные из природных источников полимеры, такие как альбумин, альгинат, производные целлюлозы, коллаген, фибрин, желатин и полисахариды, а также синтетические полимеры, такие как полиэфиры (полилактид, полимер на основе молочной и гликолевой кислот), полиэтиленгликоль, полиоксомеры, полиангидриды и полимеры на основе плуроновой кислоты. Данные полимеры являются биосовместимыми с нервной системой, включая центральную нервную систему, они являются биодеградируемыми в центральной нервной системе без образования токсических побочных продуктов деградации, и они обладают способностью модифицировать способ и продолжительность выхода пуриновых нуклеозидов посредством манипуляции кинетических характеристик полимеров. Как используют в данном контексте, термин "биодеградируемый" означает, что полимер будет разлагаться в течение времени под действием ферментов, посредством гидролитического воздействия и/или воздействия других аналогичных механизмов в теле субъекта. Как используют в данном контексте, термин "биосовместимый" означает, что полимер является совместимым с живой тканью или живым организмом, поскольку он не является токсичным или вредным и не вызывает иммунологического отторжения.
Полимеры могут быть приготовлены с использованием способов, известных в уровне техники (см. работы Sandler S.R., Karo W. "Синтез полимеров" (Polymer Syntheses); Harcourt Brace, Boston, 1994; Shalaby W., Ikada Y., Langer R., Williams J. "Полимеры биологического и биомедицинского значения" (Polymers of Biological and Biomedical Significans), ACS Symposium Series 540; American Chemical Society; Washington, DC, 1994). Возможно конструирование гибких полимеров; расстояние между биоактивными боковыми цепями и длина линкера между полимерным скелетом и группой могут контролироваться. Другие подходящие полимеры и способы их получения описаны в Патентах США NoNo 5455044 и 5576018, содержание которых введено в данный контекст в виде ссылки.
Полимерные препараты могут быть получены диспергированием пуринового нуклеозида в ожиженном полимере, как представлено в Патенте США No 4883666, описание которого введено в данном контексте в виде ссылки, или такими способами, как полимеризация в суспензии, межфазная полимеризация, полимеризация в растворе и циклическая полимеризация, как описано в монографии Odian G. "Основы полимеризации и полимеризация с раскрытием цикла" (Principles of Polymerization and ring opening polymerization), 2-е изд., John Wiley & Sons, New York, 1981, содержание которой введено в данный контекст в виде ссылки. Свойства и характеристики препаратов контролируются изменением таких параметров, как температура реакции, концентрация полимера и пуринового нуклеозида, типы использованных растворителей и время реакции.
Пуриновый нуклеозид или его аналог могут быть инкапсулированы в один или более фармацевтически приемлемых полимеров с образованием микрокапсулы, микросферы или микрочастицы (термины, используемые в данном контексте взаимозаменяемо). Микрокапсулы, микросферы или микрочастицы обычно представляют собой свободно текущие порошки, состоящие из сферических частиц диаметром 2 мм или менее, обычно диаметром 500 мкм или менее. Частицы меньше 1 мкм принято называть нанокапсулами, наночастицами или наносферами. По большей части различие между микрокапсулой и нанокапсулой, микросферой и наносферой или микрочастицей и наночастицей состоит в размере, в общем существует небольшое (если оно имеется) различие между внутренними структурами двух видов частиц. В одном аспекте данного изобретения среднее значение диаметра составляет менее чем 45 мкм, предпочтительно менее чем 20 мкм и более предпочтительно между 0,1 и 10 мкм.
В другом варианте осуществления фармацевтически приемлемые препараты содержат препараты на липидной основе. Любые известные системы доставки лекарств на липидной основе могут быть использованы при практической реализации изобретения. Например, мультивезикулярные липосомы (MVL), многослойные липосомы (известные также как многослойные носители или "MLV"), однослойные липосомы, включая маленькие однослойные липосомы (известные также как однослойные носители или "SUV") или большие однослойные липосомы (известные также как большие однослойные носители или "LUV"), могут быть использованы при условии, что может быть установлена скорость задержанного выхода инкапсулированного пуринового нуклеозида или его аналога. В одном варианте осуществления препарат на основе липидов может быть системой многослойных липосом. Способы получения систем доставки лекарств на основе многослойных липосом с контролируемым выходом описаны в международных заявках РСТ с регистрационными номерами US96/11642, US94/12957 и US94/04490, содержание которых введено в данном контексте в виде ссылки.
Композиция синтетического мембранного носителя обычно представляет собой комбинацию фосфолипидов, обыкновенно в сочетании со стероидами, в частности холестерином. Другие фосфолипиды или другие липиды также могут быть использованы.
Примеры липидов, используемых при получении синтетических мембранных носителей, включают фосфатидилглицерины, фосфатидилхолины, фосфатидилсерины, фосфатидилэтаноламины, сфинголипиды, цереброзиды и ганглиозиды. Предпочтительно используют фосфолипиды, включающие яичный фосфатидилхолин, дипальмитоилфосфатидилхолин, дистеароилфосфатидилхолин, диолеоилфосфатидилхолин, дипальмитоилфосфатидилглицерин и диолеоилфосфатидилглицерин.
При получении носителей на липидной основе, содержащих пуриновый нуклеозид или его аналог, следует принимать во внимание такие изменяющиеся параметры, как эффективность инкапсуляции пуринового нуклеозида, лабильность пуринового нуклеозида, гомогенность и размер полученной популяции носителей, соотношение пуриновый нуклеозид - липид, проницаемость, нестабильность препарата и фармацевтическая приемлемость препарата (см. работы Szoka и соавт., Annual Reviews of Biophysics and Bioengineering, 9:467, 1980; Deamer и соавт. в монографии "Липосомы" (Liposomes), Marcel Dekker, New York, 1983, стр. 27 и Hope и соавт. , Chem. Phys. Lipids, 40:89, 1986, содержание которых включено в данный контекст в виде ссылки).
Введение фармацевтически приемлемого препарата
Фармацевтически приемлемые препараты, соответствующие изобретению, вводят таким образом, чтобы пуриновый нуклеозид или его аналог контактировали с нейронами центральной нервной системы, модулируя таким образом рост их аксонов. Как местное, так и системное введение препаратов рассматриваются в изобретении, хотя местное введение может быть предпочтительным для достижения эффективных локальных концентраций пуринового нуклеозида или аналога, а также для избежания возможных побочных эффектов при системном введении агента. В одном варианте осуществления пуриновый нуклеозид или его аналог вводят путем интродукции в центральную нервную систему субъекта, например в спинномозговую жидкость субъекта. В ряде аспектов изобретения пуриновый нуклеозид или его аналог вводят интратекально, например, в желудочек головного мозга, в область поясницы или в большую цистерну. В другом аспекте пуриновый нуклеозид или его аналог вводят внутриглазно для контакта таким образом с клетками ганглия сетчатки.
Фармацевтически приемлемые препараты могут быть легко суспендированы в водных носителях и введены с помощью обычных игл для подкожных инъекций или с использованием инфузионных насосов. Перед введением препараты могут быть простерилизованы предпочтительно γ-облучением или стерилизацией пучком электронов, которые описаны в Патенте США No 436742, содержание которого введено в данный контекст в виде ссылки.
В одном варианте осуществления препарат пуринового нуклеозида, описанный в данной заявке, вводят субъекту в период от момента повреждения до приблизительно 100 часов после того, как произошло повреждение, например в течение 24, 12 или 6 часов с момента повреждения.
В другом варианте осуществления препарат пуринового нуклеозида вводят субъекту подоболочечно (интратекально). Как используют в данном контексте, термин "подооболочечное введение" предусматривает включение доставки препарата пуринового нуклеозида непосредственно в спинномозговую жидкость субъекта способами, включающими латеральную церебровентрикулярную инъекцию через трепанационное отверстие или посредством цистернальной или поясничной (люмбальной) пункции или подобных мер (см. монографию Lazorthes и соавт. "Успехи в разработке систем доставки лекарств и их применение в нейрохирургии" (Advances in Drug Delivery Systems and Applications in Neurosurgery), стр. 143-192 и Omaya и соавт., Cancer Drug Delivery, 1:169-179, содержание которых введено в данном контексте в виде ссылки). Термин "поясничный участок" предусматривает включение области между третьим и четвертым поясничными позвонками (позвонками нижнего отдела спины). Термин "большая цистерна" предусматривает включение области, в которой заканчивается череп и начинается спинной мозг в нижней части головы. Термин "желудочек головного мозга" предусматривает включение области полостей головного мозга, которые связаны с центральным каналом спинного мозга. Введение пуринового нуклеозида в какой-либо из вышеуказанных участков может достигаться прямой инъекцией препарата пуринового нуклеозида или применением инфузионных насосов. Для инъекции препарат пуринового нуклеозида, соответствующий изобретению, может быть приготовлен в жидких растворах, предпочтительно в физиологически совместимых буферах, таких как раствор Хенка или раствор Рингера. Кроме того, препарат пуринового нуклеозида может быть приготовлен в твердой форме и перерастворен или суспендирован непосредственно перед применением. Лиофилизированные формы также включаются. Инъекция может быть, например, в форме болюсной инъекции или непрерывного введения (например, с использованием инфузионных насосов) препарата пуринового нуклеозида.
В одном варианте осуществления изобретения препарат пуринового нуклеозида вводят путем латеральной церебровентрикулярной инъекции в головной мозг субъекта, предпочтительно в течение 100 часов после того, как произошло повреждение (например в течение 6, 12 или 24 часов с момента повреждения). Инъекция может быть проведена, например, через трепанационное отверстие, сделанное в черепе субъекта. В другом варианте осуществления изобретения препарат вводят через хирургически выполненный шунт в желудочке головного мозга субъекта, предпочтительно в течение 100 часов после того, как произошло повреждение (например, в течение 6, 12 или 24 часов с момента повреждения). Например, инъекция может быть сделана в латеральные желудочки, которые больше по размеру, хотя может быть выполнена также инъекция путем введения в третий и четвертый, меньшие по размеру, желудочки. В еще одном варианте осуществления препарат пуринового нуклеозида вводят путем инъекции в большую цистерну или поясничную область субъекта, предпочтительно в течение 100 часов после того, как произошло повреждение (например, в течение 6, 12 или 24 часов с момента повреждения).
Продолжительность и уровни введения
В предпочтительном варианте осуществления способа, соответствующего изобретению, пуриновый нуклеозид или его аналог контактирует с нейронами ЦНС в течение продолжительного периода времени для получения эффекта модуляции вырастания аксонов. Длительный контакт с пуриновым нуклеозидом или аналогом может быть достигнут, например, повторным введением пуринового нуклеозида или аналога через такой период времени, как одна неделя, несколько недель, один месяц или более. Более предпочтительно, когда фармацевтически приемлемый препарат, используемый для введения пуринового нуклеозида или аналога, обеспечивает задержанную доставку субъекту, например "медленное высвобождение" пуринового нуклеозида или аналога. Например, препарат может доставлять пуриновый нуклеозид или аналог в течение, по меньшей мере, одной, двух, трех или четырех недель после введения фармацевтически приемлемого препарата субъекту. Предпочтительно, чтобы субъекту, которого лечат в соответствии с данным изобретением, вводили пуриновый нуклеозид или аналог в течение, по меньшей мере, 30 дней (либо путем повторного введения или путем использования системы задержанной доставки, либо обоими путями).
Как используют в данном контексте, термин "задержанная доставка" предусматривает включение непрерывной доставки пуринового нуклеозида или его аналога in vivo в течение периода времени после введения, предпочтительно в течение, по крайней мере, нескольких дней, недели, нескольких недель, одного месяца или более. Задержанная доставка пуринового нуклеозида или его аналога может быть продемонстрирована, например, в виде постоянного терапевтического эффекта пуринового нуклеозида или его аналога в течение времени (например, задержанная доставка пуринового нуклеозида или его аналога может быть продемонстрирована в виде постоянного роста или постоянного ингибирования роста нейронов ЦНС в течение времени). Альтернативно задержанная доставка пуринового нуклеозида или его аналога может быть продемонстрирована детекцией присутствия пуринового нуклеозида или его аналога in vivo в течение времени.
Предпочтительные подходы к задержанной доставке включают использование полимерной капсулы или мининасоса для доставки препарата. Полимерные капсулы могут быть приготовлены, как описано выше. Имплантируемые системы инфузионных насосов (например, Infusaid; см., например, статьи Zierski J. и соавт., Acta Neurochem. Suppl., 43:94-99, (1988); Kanoff R.B., J. Am. Osteopath. Assoc. , 94:487-493 (1994)) и осмотические насосы (реализуются фирмой Alza Corporation) имеются в области техники. Другим способом введения является использование имплантируемого программируемого снаружи инфузионного насоса. Подходящие системы инфузионных насосов и системы резервуаров также описаны Blomquist в Патенте США No 5368562 и Doan в Патенте США No 4731058, разработанном в Pharmacia Deltec Inc.
Фармацевтический препарат, использованный в способе, соответствующем изобретению, содержит терапевтически эффективное количество пуринового нуклеозида или его аналога. Термин "терапевтически эффективное количество" означает количество, эффективное при дозах и в течение времени, которые необходимы для достижения желаемого результата. Терапевтически эффективное количество пуринового нуклеозида или его аналога может изменяться в соответствии с такими факторами, как состояние болезни, возраст и масса субъекта и способность пуринового нуклеозида или его аналога (в виде монотерапии или в комбинации с одним или более других агентов) вызывать желаемую реакцию у субъекта. Схемы лечения могут быть подработаны для обеспечения оптимальной терапевтической реакции. Терапевтически эффективное количество также представляет собой количество, при котором какие-либо токсические или вредные эффекты пуринового нуклеозида или его аналога перевешиваются терапевтически благоприятными эффектами. Неограничивающий диапазон доз составляет приблизительно 5-1000 мкМ, хотя особенно оптимальная дозировка будет изменяться в зависимости от (среди прочих факторов) определенного используемого пуринового нуклеозида или его аналога.
Для получения стимуляции вырастания аксонов инозином неограничивающий диапазон для терапевтически эффективной концентрации составляет 5-1000 мкМ, более предпочтительно 10-500 мкМ. Даже более предпочтительная локальная концентрация инозина, контактирующая с нейронами ЦНС, составляет приблизительно 25 мкМ.
Для получения стимуляции вырастания аксонов гуанозином неограничивающий диапазон для терапевтически эффективной концентрации составляет 5-1000 мкМ, более предпочтительно 10-500 мкМ. Даже более предпочтительная локальная концентрация инозина, контактирующая с нейронами ЦНС, составляет приблизительно 100 мкМ.
Для получения ингибирования вырастания аксонов 6-тиогуанином локальная концентрация 6-тиогуанина, контактирующая с нейронами ЦНС, предпочтительно составляет 50 мкМ или менее.
Аденозин может быть использован для ингибирования роста невритов в относительно высоких дозах, например выше 5 мМ (при этом ингибируется его превращение в инозин). Однако при данных концентрациях аденозин может стать токсичным. Таким образом аналоги аденозина, например 6-тиогуанин, являются вследствие этого предпочтительными для введения субъектам-млекопитающим с целью ингибирования вырастания аксонов.
Следует заметить, что значения доз могут отличаться в связи с тяжестью состояния, которое следует облегчить. Кроме того, следует понимать, что для любого определенного субъекта специфические схемы лечения должны подбираться со временем в соответствии с нуждами субъекта и профессиональным мнением медицинского работника, проводящего введение или руководящего введением пуринового нуклеозида или его аналога, и что представленные в данном контексте диапазоны доз являются лишь примерными и не предназначены для ограничения объема или практической реализации заявленного изобретения.
Изобретение в другом варианте осуществления представляет фармацевтическую композицию, состоящую в основном из пуринового нуклеозида или его аналога (например, инозина, гуанозина, 6-тиогуанина) и фармацевтически приемлемого носителя, а также способы ее использования для модуляции вырастания аксонов путем контактирования нейронов ЦНС с композицией. Термин "состоит в основном из" означает, что фармацевтическая композиция не содержит какие-либо другие модуляторы роста нейронов, такие как, например, фактор роста нервов (NGF). В одном варианте осуществления фармацевтическая композиция, соответствующая изобретению, может быть представлена в виде упакованного препарата. Упакованный препарат может включать фармацевтическую композицию, соответствующую изобретению, в контейнере и инструкции в отпечатанном виде по применению композиции для лечения субъекта, имеющего нарушение, связанное с повреждением нейронов центральной нервной системы, например повреждение клеток ганглия сетчатки, повреждение спинного мозга или черепно-мозговую травму. Применение пуриновых нуклеозидов или их аналогов, соответствующих изобретению, в качестве лекарственного средства для модуляции роста нейронов ЦНС (например, нейронов ЦНС млекопитающих) также охватывается изобретением.
Обработка нейронов ЦНС in vitro
Нейроны ЦНС могут далее контактировать с пуриновым нуклеозидом или его аналогом in vitro для модуляции роста аксонов in vitro. Соответственно, клетки нейронов ЦНС могут быть выделены у субъекта и выращены in vitro с использованием технологий, известных в уровне техники, а затем обработаны в соответствии с данным изобретением с целью модуляции вырастания аксонов. Вкратце, культура клеток нейронов ЦНС может быть получена в условиях, позволяющих клеткам нейронов мигрировать из фрагментов нервной ткани, прикрепленной к подходящему субстрату (например, чашке для культивирования), или путем дезагрегации клеток ткани, например, механическом или ферментативном, для получения суспензии клеток нейронов ЦНС. Например, могут быть использованы ферменты трипсин, коллагеназа, эластаза, гиалуронидаза, ДНКаза, проназа, диспаза или их различные комбинации. Трипсин и проназа обеспечивают наиболее полную дезагрегацию, но могут повреждать клетки. Коллагеназа и диспаза приводят к менее полной дезагрегации, но являются менее вредными. Способы выделения ткани (например, нервной ткани) и дезагрегации ткани для получения клеток описаны в монографии Freshney R.L. Культура клеток животных. Руководство по основным методикам (Culture of Animal Cells, A Manual of Basic Technique), 3-е изд. (1994), содержание которой введено в данном контексте в виде ссылки.
Такие клетки могут последовательно контактировать с пуриновым нуклеозидом или его аналогом в количествах и в течение времени, как описано выше. После достижения модуляции вырастания аксонов в нейронах ЦНС данные клетки могут быть введены обратно субъекту, например, путем имплантации.
Изобретение дополнительно иллюстрируют следующими примерами, которые не следует рассматривать как дальнейшее ограничение. Содержание всех ссылок, патентов и опубликованных патентных заявок, приведенных в данной заявке, введено в данный контекст в виде ссылки.
Примеры
В последующих примерах используют следующие методики:
Приготовление образца
Фактор аксогенеза 1 (АФ-1) получают в основном, как описано в статьях Schwalb и соавт. , (1995) и Schwalb и соавт., Neuroscience, 72:901-910, (1996), содержание которых введено в данном контексте в виде ссылки. Глазные нервы препарируют, разрезают на фрагменты длиной 1 мм и инкубируют в соотношении 6 нервов в 3 мл либо среды L-15 (Gibco BRL), либо забуференного фосфатом физиологического раствора (Gibco BRL). Через 3-4 часа фрагменты нервов удаляют фильтрованием через фильтр с низкой связываемостью белка с размером пор 0,22 мкм (Gelman). Низкомолекулярную фракцию кондиционированной среды получают ультрафильтрацией, сначала удалением фракции молекулярной массы 3 кД (Amicon Centriprep-3), затем удалением фракции 1 кД (Filtron). Фильтрат используют в качестве положительного контроля в конечной концентрации 20-30%. Аденозин, аденозин 5'-монофосфат, аденозиндеаминазу, аденозиндифосфат, аденозинтрифосфат, 8-бром-3', 5'-циклический гуанозинмонофосфат, 3',5'-циклический аденозинмонофосфат, 5'-циклический гуанозинмонофосфат, цитидин, гуанозин, гипоксантин, инозин, 5'-инозинмонофосфат, α-токоферол, 6-тиогуанин, тимидин, уридин и ксантин выпускаются фирмой Sigma Chemical Co., St. Louis, МО, 8-р-сульфофенилтеофиллин, дибутирил циклический аденозинмонофосфат и 2-дезоксикоформицин получают от Calbiochem, 2-хлораденозин, эритро-9-(2-гидрокси-3-нонил)аденин и IB-MECA - от Research Biochemicals, Inc. (Natick, MA) и 4-(нитробензил-6-тиоинозин) - от Aldrich Chemicals, Inc. Мембранопроницаемые негидролизующиеся аналоги цАМФ и цГМФ, 8-бромаденозин-3', 5'-циклический монофосфоротиоат и 8-(4-хлорфенилтио)гуанозин-3',5'-циклический монофосфат получают от фирмы Biolog.
Диссоциированные культуры сетчатки
Серебряных карасей (вид Comet, Mt. Parnell Fisheries, Mt. Parnell PA) длиной 6-10 см адаптируют к темноте и удаляют у них сетчатку. Образцы сетчатки инкубируют с папаином (20 мкг/мл), активированным цистеином (2,8 мМ), в течение 30 минут при комнатной температуре, а затем разрушают путем осторожного измельчения. Повторение циклов измельчения и осаждения приводит к получению культур почти гомогенных клеток ганглия, которые легко идентифицируют по овальной форме, внешнему виду при микроскопии в проходящем свете, размеру (диаметр 15 мкм) и удлинению в виде только 1 или 2 невритов одного размера, данные критерии проверяют введением ретроградационной метки (см. статьи Schwartz & Agranoff, Brain Res., 206:331-343, (1981) и Schwalb и соавт. , J. Neuroscience, 15:5514-5625, (1995), содержание которых введено в данном контексте в виде ссылки). Культуры с низкой густотой получают, помещая приблизительно 5•103 клеток/лунку в покрытые поли L-лизином 24-луночные планшеты для культивирования (Costar, Cambridge, MA). Клетки поддерживают при 21oС в бессывороточной среде определенного состава, содержащей инсулин, селен, трансферрин, бычий сывороточный альбумин, каталазу, супероксиддисмутазу, гормоны и витамины в среде Игла L-15, как описано в работе Schwalb и соавт. , содержание которой введено в данном контексте в виде ссылки. Диссоциированные культуры очищенных клеток ганглия сетчатки крыс готовят с помощью иммунопэннинга, как описано в статье Barres и соавт., Neuron, 1:791-803, (1988), содержание которой введено в данном контексте в виде ссылки. Вкратце, сетчатки новорожденных крыс Sparague-Dawley в возрасте 8 дней диссоциируют с использованием папаина, активированного цистеином. Макрофаги удаляют путем инкубирования с антителами против крысиных макрофагов (Accurate) с последующим иммунопэннингом с использованием антител против IgG кролика. Клетки ганглия выделяют иммунопэннингом с использованием антитела против ТПу-1, затем удаляют с помощью трипсина с целью использования культур с низкой густотой. Клетки ганглия сетчатки крыс поддерживают при 37oС в инкубаторе с СО2, используя ту же самую среду, что описана выше, за исключением присутствия 30 мМ бикарбоната.
Схема эксперимента
В типичном эксперименте образцы помещают в четырех повторностях в случайные положения 24-луночного планшета и код скрывают, чтобы гарантировать оценку роста слепым способом. В каждый эксперимент входит 4 лунки отрицательного контроля (только среда и добавки) и 4 лунки положительного контроля (стандартизированный образец АФ-1 известной активности). Рост и выживаемость оценивают через 6 дней для всех клеток ганглия в 25 последовательных полях, просмотренных в каждой лунке с использованием фазово-контрастного микроскопа с увеличением 400Х (подсчитывают приблизительно 150 клеток ганглия/лунку). Критерием роста является вытягивание клеток на 5 диаметров в длину, поскольку это ясно отличает стимулированные клетки от отрицательных контролей (см. работу Schwalb и соавт., 1995). После завершения подсчета распечатывают код, сводят данные в таблицу и вычисляют средние значения и стандартные ошибки для 4 повторов лунок для каждого образца, используя график Крикета (CA Associates, Islandia, NY). Данные нормализуют вычитанием роста в отрицательных контролях (обычно 4-5%) и делением на рост сети в положительных контролях. В наиболее удачных экспериментах в более 50% клеток ганглия сетчатки (КГС), обработанных АФ-1, вырастают аксоны длиной 5 диаметров клетки через 6 дней. Сравнения в группах делают на основе попарных тестов с использованием двустороннего критерия Стьюдента. С большинством образов проводят ряд независимых экспериментов, что отражают в подписях к чертежам. В ряде случаев выживаемость клеток оценивают с помощью красителя 5,6-карбоксифлуоресцеина диацетата. Выживаемость клеток показывают как число жизнеспособных КГС/поле при сильном увеличении.
Пример 1. Индуцированная пурином стимуляции вырастания аксонов из клеток ганглия сетчатки серебряного карася
Низкомолекулярный фактор роста АФ-1, секретируемый глией зрительного нерва, индуцирует сильный рост клеток ганглия сетчатки серебряного карася. Слабый рост происходит в условиях контроля и с использованием только среды определенного состава. Два данных ограничения служат основой для нормализации результатов по другим факторам. При тестировании нуклеозидов (А, Г, Ц, У и Т) в концентрациях между 1-100 мкМ аденозин и гуанозин стимулируют рост клеток ганглия сетчатки серебряного карася почти на таком же уровне, что АФ-1 (см. Фигуру 1.1). Пиримидиновые основания не обладают активностью в данном диапазоне концентраций. Более полная кривая доза-зависимости для пуринов показывает, что аденозин является более активным из этих двух пуринов при EC50 10-15 мкМ (см. Фигуру 1.2). При концентрациях 50-100 мкМ аденозин индуцирует максимальный ответ, равный 60% уровня, индуцируемого АФ-1, но при более высоких концентрациях рост снижается. Гуанозин имеет более высокое значение EC50, чем аденозин (25 мкМ, см. Фигуру 1.2), и при концентрациях 100 мкМ он стимулирует такой же максимальный уровень активности, как аденозин при отсутствии очевидного снижения активности при более высоких концентрациях.
Пример 2. Пуриновые нуклеотиды являются менее активными, чем нуклеозиды
Вне клетки аденозин мог бы стимулировать либо Р1-рецепторы, которые оптимально чувствительны к аденозину как таковые, или Р2-рецепторы, которые максимально реагируют на АТФ или другие нуклеотиды. АМФ и АДФ показывают уровень активности пограничной значимости при 100 мкМ (р 0,05), как АТФ при концентрации 10 мкМ (но не при 100 мкМ) (см. Фигуру 1.3). Поскольку активность пуриновых нуклеозидов значительно ниже активности самих пуринов, маловероятно, что в механизме участвуют Р2-рецепторы. Вероятно, пурины могли бы функционировать внутри клетки как предшественники циклических нуклеотидов, которые могли бы служить вторичными переносчиками при аксогенезе. Вследствие этого исследуют биологическую активность мембранопроницаемых аналогов цАМФ и цГМФ. Ни дибутирил цАМФ (ДБ-цАМФ)), ни 8-Вr-цГМФ не проявляют никакую активность при концентрациях 1-100 мкМ (см. Фигуру 1.4). Недавно полученные негидролизующиеся мембранопроницаемые аналоги цАМФ (8-бромаденозин-3',5'-циклический монофосфоротиоат (8-Вr-АцМФТ)) и цГМП (8-(4-хлорфенилтио)гуанозин-3', 5'-циклический монофосфат (8-ХФТГ-цМФ)), как показывают, также являются неактивными при тестировании в концентрациях до 1 мМ (см. Фигуру 1.4).
Пример 3. Положительные эффекты аденозина не опосредуются внеклеточными аденозиновыми рецепторами
8-р-(сульфофенилтеофиллин)(8-СФТ), описанный в статье Collis и соавт., Brit. J. Pharmacol. , 92:69-75 (1987), содержание которой введено в данном контексте в виде ссылки, представляет собой ингибитор двух наиболее распространенных аденозиновых рецепторов (А1 иА2). При концентрации 20 мкМ дозе, которая почти полностью блокирует рецептор-опосредованные эффекты аденозина у крыс, 8-СФТ не действует на рост, стимулированный аденозином, гуанозином или АФ-1 (см. Фигуру 2). Дальнейшее доказательство того, что положительные эффекты аденозина не опосредуются внеклеточными аденозиновыми рецепторами, происходит из исследований с использованием негидролизующегося аналога 2-хлораденозина (2-Cl-A), который является агонистом относительно А1-, А2- и А3-рецепторов. При концентрациях 10 и 100 мкМ 2-Cl-A вызывает небольшое, но значимое снижение роста ниже базового в 3 из 3 независимых экспериментах (см. Фигуру 2).
Пример 4. Аденозин должен быть гидролизованным до инозина, чтобы стимулировать рост
Для исследования, является ли активность аденозина обусловленной образованием активного метаболита, активность АДФ ингибируют, используя либо дезоксикоформицин(ДОФ) либо эритро-9-(2-гидрокси-3-нонил)аденин (ЭГНА). В присутствии 10 мкМ ДОФ аденозин в концентрации 100 мкМ не только не стимулирует рост, но вызывает его уменьшение ниже базовых уровней (Фигура 3, дорожки е и d). Выживаемость клеток также снижается при блокировании гидролиза аденозина. В присутствии 10 мкМ ДОФ аденозин в концентрации 10 мкМ вызывает снижение выживаемости на 20% (не показано), а аденозин в концентрации 100 мкМ вызывает снижение выживаемости на 57% (Фигура 3-2, дорожка е). Эффекты ДОФ на рост и выживаемость специфически связаны с присутствием негидролизованного аденозина, поскольку они отсутствуют при использовании одного ДОФ, с АФ-1 или гуанозином (Фигура 3, дорожки b и h). Подобно ДОФ, 10 мкМ ЭГНА делает аденозин (100 мкМ) неэффективным для стимуляции вырастания и вызывает снижение выживаемости клеток на приблизительно 30% (данные не приводят). ЭГНА также проявляет неспецифические эффекты, снижая, однако, рост, стимулируемый либо гуанозином, либо АФ-1, приблизительно на 50%, не вызывая при этом изменений выживаемости клеток. Дальнейшее доказательство того, что для положительных эффектов аденозина необходим его гидролиз, вытекает из экспериментов, в которых добавляют экзогенный АДА. При концентрации 0,4 ед./мл фермент не снижает вырастания аксонов, стимулируемого аденозином в концентрации 100 мкМ, или не влияет на выживаемость клеток (см. Фигуру 3, дорожка f).
Пример 5. Инозин является активным метаболитом
Инозин, первичный продукт деамидирования аденозина, как считают, является сильным активатором вырастания аксонов. Как показано на Фигуре 4, EC50 для инозина составляет 10-15 мкМ, и максимальный ответ, равный приблизительно 60% от уровня, достигаемого с использованием АФ-1, получают при концентрациях выше 25 мкМ. При том что ЕС50 и максимальный ответ, индуцируемый инозином, аналогичны данным эффектам аденозина, одно заметное различие состоит в том, что при более высоких концентрациях инозин не вызывает снижение роста в отличие от аденозина. Дальнейший гидролиз инозина приводит к образованию гипоксантина, который совсем не проявляет активность (см. Фигуру 4). Инозин 5'-монофосфат (5'-ИМФ) является неактивным при концентрации 10 мкМ, а при концентрации 100 мкМ он проявляет активность, меньшую, чем у инозина при концентрации 10 мкМ (см. Фигуру 4).
Пример 6. Пурины стимулируют рост через внутриклеточные механизмы
Два ингибитора переносчика пуринов, нитробензилтиоинозин (НБТИ) и дипиридамол, используют для исследования необходимости вхождения инозина и гуанозина в нейроны для стимуляции вырастания. При концентрации 20 мкМ НБТИ блокирует приблизительно 90% роста, индуцированного либо инозином, либо гуанозином (см. Фигуру 5; 86%-ная потеря активности для инозина в концентрации 50 мкМ, р<0,001; 93%-ная потеря активности для гуанозина в концентрации 100 мкМ, р<0,01). Дипиридамол (10 мкМ) также снижает рост, индуцированный инозином (снижение 114%; р<0,01, не показано, гуанозин не исследуют). Напротив, для АФ-1 показывают небольшое ингибирование НБТИ (10%-ное снижение, не представлено) и несколько большее - дипиридамолом (25%-ное снижение, не представлено). Связанная с НБТИ потеря активности для пуринов значительно больше, чем для АФ-1 (р<0,001).
Пример 7. Активность АФ-1 обусловлена не инозином
В плане возможности содержания в препаратах АФ-1 пуринов, которые могли бы обусловливать часть их биологической активности, проводят хроматографирование нативного АФ-1 и инозина на колонке для гель-хроматографии с Sephadex G-10 (Pharmacia Biotech, Uppsala, Sweden) диаметром 1 см, длиной 10 см. Образцы наносят в объеме 0,5 мл и собирают фракциями по 1 мл. Буфер для колонки представляет собой либо 20% метанол в дистиллированной воде, либо 0,14 М NaCl. Биоанализ фракций проводят при концентрации 30%. Как показывают на Фигуре 6-1, пик активности инозина приходится на 9-10 минуту, тогда как для АФ-1 приходится на 7 минуту.
Пример 8. Рост аксонов обусловлен эффектом инозина и гуанозина, а не эффектом вторичного фактора
Используемые культуры содержат 70-90% клеток ганглия, где остальной материал представляет собой другие элементы нервов и не нервов сетчатки (см. работы Schwartz & Agranoff, 1982 и Schwalb и соавт., 1995, содержание которых введено в данном контексте в виде ссылки). Данная гетерогенность обусловливает возможность того, что инозин или гуанозин могут первично воздействовать на другую популяцию клеток, которая секретирует вторичный фактор, стимулирующий рост клеток ганглия сетчатки. В таком случае можно было бы ожидать изменение эффекта пуринов в зависимости от густоты клеток, поскольку концентрация вторичного фактора повышалась бы пропорционально увеличению густоты. Для исследования этого изучают рост в ответ на фиксированную концентрацию инозина или гуанозина при 3-4-кратном диапазоне густоты клеток. Линии регрессии для данных как по инозину, так и по гуанозину демонстрируют, что рост не является функцией густоты клеток (см. Фигуру 6-2), что является аргументом против присутствия зависимого от концентрации вторичного фактора.
Пример 9. Индукция пуринами экспрессии фосфопротеина GAP-43
Одной из характеристик регенерации зрительного нерва in vivo является повышенная экспрессия мембранного фосфопротеина GAP-43. Для исследования, индуцируется ли данная позитивная регуляция пуринами, проводят иммуногистохимический анализ с использованием поликлональной антисыворотки кролика против GAP-43 серебряного карася. Рекомбинантный GAP-43 зубатого карася получают путем трансформации Е. coli кДНК, выделенной д-ром Eva Reinhard Базельского университета, Швейцария (см. статью Reinhard и соавт., Development, 120: 1757-1775, (1994), содержание которой введено в данном контексте в виде ссылки), субклонированнной в прокариотный экспрессионный вектор pTrcHisB(lnvitrogen). Полученный белок очищают Ni2+-NТА-аффинной хроматографией и используют для иммунизации кроликов. Специфичность полученного антитела демонстрируют с помощью вестерн-блоттинга, в котором антитело распознает уникальную полосу 48 кД, которая усилена в клетках ганглия сетчатки, в которых проходит регенерация, или в синаптосомных плазматических мембранах головного мозга серебряного карася.
Все агенты, АФ-1, инозин и гуанозин, вызывают значительное повышение уровней GAP-43 относительно обработанных L-15 контролей. Полуколичественный анализ проводят путем оценки уровня иммунореактивности GAP-43 как 0 (отсутствующего), 1 (среднего) или 2 (высокого) и корреляции интенсивности окрашивания с длиной клеточного аксона для 150-200 клеток, обработанных L-15, инозином или АФ-1. Инозин вызывает увеличение в 5,5 раз числа интенсивно окрашенных клеток относительно L-15, тогда как АФ-1 приводит к 8-кратному увеличению. Во всех 3 случаях интенсивность иммуноокрашивания GAP-43 коррелирует в высокой степени с длиной аксонов.
Пример 10. Блокада вырастания аксонов 6-тиогуанином (6-TГTG)
В КГС серебряного карася 6-ТГ в концентрации 10 мкМ блокирует всякий рост, стимулируемый АФ-1 (см. Фигуру 7-1, дорожку 2), но не оказывает воздействия на выживаемость клеток (см. Фигуру 7-2). Такая же концентрация 6-ТГ снижает рост, стимулируемый инозином в концентрации 25 мкМ только на 50% (см. Фигуру 7-1, дорожки 3 и 4) и не оказывает воздействия на рост, стимулируемый либо инозином в концентрации 100 мкМ, либо гуанозином в концентарции 100 мкМ (см. Фигуру 7-1, дорожки 5-8). Инозин в концентрации 100 мкМ полностью восстанавливает рост, индуцируемый АФ-1, в присутствии 6-ТГ в концентрации 10 мкМ до его исходного уровня, который значительно превышает уровень роста, индуцируемый одним инозином (см. Фигуру 7-1, дорожка 10 относительно 6). Вследствие этого инозин и ТГ-6, вероятно, воздействуют конкурентно на уровень внутриклеточного сигнала, который также используется АФ-1 для стимуляции вырастания. Дальнейшее доказательство того, что инозин может активировать тот же самый путь, который используется сигналом АФ-1, вытекает из наблюдения, что при комбинировании двух данных агентов при уровнях их EC50 они проявляют адцитивные эффекты, тогда как в насыщающих концентрациях рост насыщается на уровне, стимулируемом высокими уровнями одного АФ-1 (см. Фигуру 7-3, дорожка 9). Поскольку 6-ТГ содержит свободный тиол, он мог бы действовать как восстановитель, а не аналог пуринов. Однако два других восстанавливающих агента, α-токоферол (α-ТФ) в концентрации 30 мкМ или α-метиловый эфир глутатиона (МЭГ) в концентрации 100 мкМ не оказывают воздействия на рост, стимулируемый АФ-1 (см. Фигуру 7-4). Другая возможность состоит в том, что инозин мог бы блокировать эффект ингибирования 6-ТГ на вырастание путем нарушения его транспорта в клетки. Однако два ингибитора транспорта, которые блокируют активность инозина, НБТИ и дипиридамол (D), не могут препятствовать блокированию 6-ТГ вырастания, стимулируемого АФ-1 (см. Фигуру 7-4).
Пример 11. Клетки ганглия сетчатки млекопитающих вытягивают аксоны в ответ на инозин
Клетки ганглия сетчатки выделяют у крыс в возрасте 8 дней посредством иммунопэннинга, как описано в статье Barres и соавт., Neuron, 1:791-803, (1988), содержание которой введено в данном контексте в виде ссылки, и выращивают на определенной среде. Инозин в концентрациях 25 или 50 мкМ стимулирует 50%-ное увеличение числа клеток, вытягивающих аксоны на длину 5-ти диаметров клеток (см. Фигуру 8). Цилиарный нейротрофический фактор (ЦНТФ) индуцирует более значительное усиление вырастания (см. Фигуру 8) и повышенную выживаемость клеток. 6-ТГ в концентрации 10 мкМ блокирует ЦНТФ-индуцированный рост. Добавление инозина в концентрации 50 мкМ восстанавливает ЦНТФ-индуцированный рост почти до его исходного уровня (см. Фигуру 8).
Эквивалентные решения
Специалисты узнают или способны установить, используя не более чем рутинные эксперименты, много эквивалентных решений специфических вариантов осуществления описанного изобретения. Предусматривается, что данные эквивалентные решения охватываются следующей формулой изобретения.
Изобретение относится к области медицины, а именно к фармакотерапии. При повреждениях центральной нервной системы, а также больным, перенесшим инсульт, вводят фармацевтическую композицию, которая в качестве активного компонента содержит эффективное количество пуринового нуклеозида или его аналога. Способ позволяет повысить эффективность лечения за счет стимуляции роста нейронов. 5 с. и 12 з.п. ф-лы, 16 ил.
| WO 9400132 А, 31.12.1993 | |||
| RU 94011780 А, 10.07.1996 | |||
| СПОСОБ ПОЛУЧЕНИЯ ИНОЗИНА | 1990 |
|
RU1755583C |

