Изобретение относится к микробиологической промышленности, к способу продуцирования L-аргинина и касается использования нового мутантного, устойчивого к ингибированию по типу обратной связи фермента в пути биосинтеза аргинина в штаммах Е.coli - продуцентах L-аргинина.
Описание существующих методов
Биосинтез аргинина из глутамата в клетках Е.coli происходит посредством серии реакций, начинающейся ацетилированием глутамата N-ацетилглутамат синтазой (NAGS), кодируемой геном argA. Этот процесс регулируется путем репрессии транскрипции arg-регулона и ингибированием NAGS аргинином по типу обратной связи (Cunin R. et al, Microbiol. Rev., v. 50, p.314-352, 1986). L-Аргинин подавляет экспрессию гена argA в соотношении более чем 250 раз и ингибирует активность NAGS (Кi=0,02 mM) (Leisinger Т., Haas D., J.Biol.Chem. , v. 250, р. 1690-1693, 1975). Для усиленного биосинтеза аргинина в клетках Е. coli требуются ферменты NAGS, устойчивые к ингибированию по типу обратной связи (также обозначаемые как "fbr").
Мутанты ферментов, устойчивых к ингибированию по типу обратной связи, могут быть получены при спонтанном, химическом или сайт-направленном мутагенезе.
Некоторые из мутантов fbr argA были выделены и изучены. Клетки Serratia marcescens, содержащие fbr argA мутации в хромосоме, были нестабильными и дали начало argA мутантам с пониженной активностью или измененной аффинностью к глутамату (Takagi Т. et al, J. Biochem., v.99, p.357-364, 1986).
Гены fbr argA из пяти штаммов Е.coli, содержащих fbr NAGS, были клонированы, и в каждом из этих штаммов были найдены различные единичные замены оснований в генах argA. Было выявлено, что данные замены вызывают замещение His-15 на Туr, Туr-19 на Cys, Ser-54 на Asn, Arg-58 на His, Gly-287 на Ser, Gln-432 на Arg (Rajagopal B.S. et al, Appl. Environ. Microbiol., 1998, v.64, No. 5, p.1805-1811).
Как правило, fbr фенотип фермента возникает в результате замены одного аминокислотного остатка другим в одном или нескольких местах последовательности белка, и эти замены приводят к уменьшению активности фермента. Например, замена природного Met-256 остальными 19-тью аминокислотными остатками в сериновой ацетилтрансферазе (SAT) (ген cysE) из E.coli в большинстве случаев приводит к появлению fbr фенотипа, но уровень активности мутантных белков SAT не достигает уровня активности природного SAT (Nakamori S. et al, AEM, 64(5): 1607-11, 1998).
Таким образом, недостатком мутантных ферментов, полученных этими методами, является снижение активности мутантных ферментов в сравнении с природными ферментами.
Краткое изложение сути изобретения
Целью настоящего изобретения является получение мутантного, устойчивого к ингибированию по типу обратной связи и высокоактивного фермента, играющего ключевую роль в биосинтезе аргинина бактериями Е.coli.
В настоящем изобретении предложен новый метод синтеза большого набора мутантных генов argA с использованием полной рандомизации фрагмента гена argA. Одновременная замена нескольких аминокислотных остатков в фрагменте последовательности белка, в котором могут быть локализованы мутации fbr, способна привести к образованию мутантных белков с восстановленной активностью, уровень которой близок к природному в виду того, что трехмерная структура фермента становится более правильной. Таким образом описанное ниже настоящее изобретение было осуществлено.
Вот то, что предоставляет настоящее изобретение:
(1) Мутантная N-ацетилглутамат синтаза, в которой последовательность аминокислот, соответствующая положениям с 15 по 19 в природной N-ацетилглутамат синтазе, заменена любой из последовательностей аминокислот под номерами 1-4 (SEQ ID NOS: 1-4) и ингибирование L-аргинином по типу обратной связи утрачено;
(2) Мутантная N-ацетилглутамат синтаза по пункту 1, которая является природной мутантной N-ацетилглутамат синтазой из Escherichia coli;
(3) Мутантная N-ацетилглутамат синтаза по пункту 1, которая содержит делеции, замены, вставки или добавления одной или нескольких аминокислот в одном или множестве положений, кроме положений с 15 по 19, и в которой ингибирование L-аргинином по типу обратной связи утрачено;
(4) ДНК, кодирующая мутантную N-ацетилглутамат синтазу по любому из пунктов 1-3;
(5) Бактерия, принадлежащая к роду Escherichia, трансформированная ДНК по пункту 4 и обладающая активностью по продукции L-аргинина; и
(6) Способ продуцирования L-аргинина, включающий стадии выращивания бактерии по пункту 5 в питательной среде для продукции и накопления L-аргинина в питательной среде и сбора L-аргинина из культуральной жидкости.
Белок NAGS, содержащий любую из fbr мутаций, описанных выше, может упоминаться как "мутантный NAGS", ДНК, кодирующая мутантный NAGS, может упоминаться как "мутантный ген argA", белок NAGS, не содержащий мутаций, может упоминаться как "природный NAGS".
Далее настоящее изобретение будет разъяснено подробнее.
<1> Мутантные белки NAGS и мутантные гены argA.
Мутантные белки NAGS и мутантные гены argA, кодирующие эти белки, были получены с помощью мутагенеза путем рандомизации фрагмента нуклеотидной последовательности гена. Для получения множественных мутаций в гене argA была произведена полная рандомизация фрагмента из 15 нуклеотидов, кодирующих район в последовательности белка с 15-го по 19-й аминокислотный остаток включительно. Полная рандомизация фрагмента из 15 нуклеотидов дает 415 или около 109 различных последовательностей ДНК, кодирующих 205 различных аминокислотных остатков в 5-мерном пептиде. Вероятность невозникновения стоп-кодонов в рамке считывания в этой последовательности равняется 0,955 или около 78%. Таким образом, полная рандомизация фрагмента гена argA, кодирующего пептид с 15-го по 19-й аминокислотный остаток, должна давать приблизительно 2,5 миллиона различных последовательностей белков, отличающихся этим пептидным фрагментом в структуре NAGS. Последовательная селекция и отбор рекомбинантных клонов, несущих мутантные гены argA, клонированные в экспрессирующие векторы, позволяет отобрать fbr варианты мутантных NAGS с различным уровнем их биологической активности вплоть до уровня активности природной NAGS. При селекции авторы изобретения решили, что штаммы, несущие мутантный ген argA, могут быть получены с использованием штаммов argD- и рrоВ- или рrоА-, потому что такие штаммы не могут продуцировать L-пролин вследствие ингибирования NAGS и в связи с этим не могут расти, если в культуральной среде присутствует избыток L-аргинина, но при этом штаммы, несущие fbr NAGS, могут расти на минимальной среде потому, что глутамат полуальдегид, предшественник L-пролина, может получаться с помощью ацетилорнитин деацетилазы (продукт гена argE) из N-ацетилглутамат полуальдегида, предшественника L-аргинина (Eckhardt Т., Leisinger Т., Mol. Gen. Genet., v.138, p. 225-232, 1975). Однако авторы изобретения обнаружили, что описанным выше методом, как это описано в последующих примерах, трудно получить fbr NAGS, обладающую высокой активностью, и что такая fbr NAGS, обладающая высокой активностью, может быть получена путем введения мутантного гена argA в природный штамм и селекцией штаммов, показывающих замедление роста.
В рамках настоящего изобретения были определены последовательности аминокислот мутантных NAGS, соответствующих fbr фенотипу NAGS. Вследствие этого мутантная NAGS может быть получена на основе этих последовательностей введением мутаций в природный ген argA с использованием обычных методов. В качестве примера природного гена argA может быть упомянут ген argA Е.coli (номер Y00492 в GenBank).
Последовательность аминокислот в положении с 15 по 19 в мутантной NAGS согласно настоящему изобретению соответствует любой из последовательностей 1-4 (SEQ ID NOS: 1-4). Соответствующая последовательность известной мутантной NAGS, в которой тирозин в положении 19 заменен на цистеин, а также природная NAGS из Е.coli приведены под номерами 5 и 6 (SEQ ID NOS: 5 и 6). Примеры последовательностей нуклеотидов, кодирующих эти последовательности аминокислот, приведены под номерами 7-12 (SEQ ID NOS: 7-12). Указанные последовательности представлены в таблице 1 (см. в конце описания).
Мутантная NAGS может содержать делеции, замены, вставки или добавки одной или нескольких аминокислот в одном или множестве положений, кроме положений с 15 по 19, при условии, что активность NAGS, которая является активностью по катализу реакции ацетилирования L-глутаминовой кислоты с получением N-ацетилглутамата, не ухудшена.
Значение "несколько" в отношении количества аминокислот различается в зависимости от положения или типа аминокислотных остатков в трехмерной структуре белка по следующей причине. Некоторые аминокислоты обладают высокой гомологией по отношению к другим аминокислотам, и разница при их замене не влияет значительным образом на трехмерную структуру белка. По этой причине мутантной NAGS согласно настоящему изобретению может быть такая NAGS, гомология у которой составляет не менее чем 30-50%, предпочтительно 50-70%, в сравнении с полной последовательностью аминокислот, установленной для NAGS, и которая обладает активностью fbr NAGS.
В настоящем изобретении "последовательность аминокислот, соответствующая последовательности в положении с 15 по 19", означает последовательность аминокислот, соответствующую последовательности аминокислот в положении с 15 по 19 в последовательности природной NAGS из E.coli. Положение аминокислотных остатков может меняться. Например, если остаток какой-либо аминокислоты введен в N-концевой участок, то остаток аминокислоты, расположенный от природы в положении 15, перемещается в положение 16. В таком случае остаток аминокислоты, соответствующий первоначальному положению 15, обозначается в настоящем изобретении как остаток аминокислоты в положении 15.
ДНК, кодирующая по существу такой же белок, как мутантная NAGS, описанная выше, может быть получена, например, путем модификации последовательности нуклеотидов, к примеру, методом сайт-направленного мутагенеза, таким образом, что вместо одного или нескольких аминокислотных остатков в указанном месте появятся делеции, замены, вставки или добавки. Описанная выше модифицированная ДНК может быть создана с использованием традиционных методов обработки для получения мутаций. Обработка для получения мутаций включает в себя метод обработки ДНК, содержащей ген argA, in vitro, например с помощью гидроксиламина, и метод обработки микроорганизма, например бактерии, принадлежащей к роду Escherichia, несущей ген argA, ультрафиолетовым облучением или агентом, вызывающим мутации, таким как N-метил-N'-нитро-N-нитрозогуанидин (NTG) и азотистой кислотой, обычно используемым при обработке для получения мутаций.
Замена, делеция, вставка или добавка нуклеотида, как это описано выше, включает также мутации, которые случаются естественным образом (мутант или вариант), например, на основе индивидуальных различий или различий на уровне вида или рода бактерии, содержащей NAGS.
ДНК, кодирующая по существу такой же белок, как и ген argA, выявляется экспрессией ДНК, содержащей описанную выше мутацию, в подходящей клетке и исследованием активности продукта экспрессии.
Также ДНК, кодирующая по существу такой же белок, как и мутантный NAGS, может быть получена выделением ДНК, которая гибридизуется с ДНК, имеющей известную последовательность гена argA, или зонда, получаемого из нее, в жестких условиях, и которая кодирует белок, обладающий активностью NAGS, из клетки, несущей мутантную NAGS, подвергшейся обработке для получения мутации.
Упомянутый здесь термин "жесткие условия" обозначает условия, при которых образуется так называемый специфический гибрид (дуплекс), а неспецифический - не образуется. Четко описать эти условия с помощью численных значений довольно трудно. Однако, например, жесткими условиями являются такие условия, при которых молекулы ДНК, обладающие высокой гомологией, например ДНК, обладающие гомологией друг с другом не менее 50%, гибридизуются, а ДНК с меньшей гомологией - не гибридизуются. В качестве альтернативы примером жестких условий являются условия, при которых ДНК гибридизуются друг с другом при концентрации солей, соответствующей обычной концентрации при отмывке в ходе гибридизации по Саузерну, т.е. 60oС, 1xSSC, 0,1% SDS, предпочтительно 0,1xSSC, 0,1%SDS.
В число генов, которые гибридизуются в описанных выше условиях, включаются гены, содержащие стоп-кодон внутри кодирующего участка гена, а также те, которые кодируют неактивный белок вследствие мутаций в активном центре. Однако подобные затруднения могут быть легко разрешены путем лигирования гена в коммерчески доступный вектор для экспрессии и изучения активности NAGS.
<2> Бактерия, принадлежащая к роду Escherichia, согласно настоящему изобретению.
Бактерией, принадлежащей к роду Escherichia, согласно настоящему изобретению является бактерия, принадлежащая к роду Escherichia, в которую введен мутантный ген argA, описанный выше. Примером бактерии, принадлежащей к роду Escherichia. является Е.coli. Мутантный ген argA может быть введен, например, путем трансформации бактерии, принадлежащей к роду Escherichia, рекомбинантной плазмидой, содержащей вектор, функционирующий в бактерии, принадлежащей к роду Escherichia, и мутантный ген argA. Мутантный ген argA также может быть введен заменой гена argA на мутантный ген argA в хромосоме.
Примерами векторов, которые можно использовать для введения мутантного гена argA, являются плазмидные векторы, такие как pBR322, pMW118, pUC19 или подобные, фаговые векторы, такие как 11059, 1BF101, M13mp9 или подобные, и транспозоны, такие как Mu, Tn10, Тn5 или подобные.
Введение ДНК в бактерию, принадлежащую к роду Escherichia, может быть осуществлено, например, по методу D.A. Morrison (Methods in Enzymology, 68, 326 (1979)) или методом, в котором бактериальная клетка - реципиент обрабатывается хлоридом кальция для увеличения проницаемости для ДНК (Mandel, M. and Higa, A., J.Mol.Biol. 53, 159 (1970)) или подобньм им методом.
Если мутантный ген argA введен в бактерию, принадлежащую к роду Escherichia, - продуцент L-аргинина, количество продуцируемого L-аргинина может быть увеличено. Кроме того, способность к продукции L-аргинина может быть придана бактерии, в которую мутантный ген argA уже введен.
Примером бактерии, принадлежащей к роду Escherichia, обладающей активностью в продукции L-аргинина, является штамм Е.coli 237 (ВКПМ В-7925). Штамм 237 был депонирован во Всероссийскую коллекцию промышленных микроорганизмов (ВКПМ).
<3> Способ получения L-аргинина.
L-аргинин может быть получен с высокой эффективностью при выращивании бактерии, в которую введен мутантный ген argA, и которая обладает способностью к продукции L-аргинина, в питательной среде, продукции и накоплении L-аргинина в питательной среде и сбора L-аргинина из культуральной жидкости.
В способе согласно настоящему изобретению выращивание бактерии, принадлежащей к роду Escherichia, накопление и сбор L-аргинина из культуральной жидкости может быть осуществлено способом, подобным способу, традиционно используемому для продукции L-аргинина методом ферментации с использованием бактерий. Питательная среда для выращивания может быть как синтетической, так и натуральной, при условии, что она содержит источники углерода и азота, минеральные соединения и, если необходимо, питательные добавки в количестве, необходимом для роста бактерии. Источники углерода включают в себя различные углеводы, такие как глюкоза и сахароза, и различные органические кислоты, в зависимости от способности к их усвоению используемыми бактериями. Могут быть использованы спирты, такие как этанол и глицерин. В качестве источников азота используются аммиак, различные соли аммония, такие как сульфат аммония, другие соединения азота, такие как амины, природные источники азота, такие как пептон, гидролизат соевых бобов и микробный ферментализат. В качестве минеральных соединений используются однозамещенный фосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, карбонат кальция. Выращивание проводят предпочтительно в аэробных условиях, таких как взбалтывание и аэрация с перемешиванием. Температура культуры обычно поддерживается от 20oС до 40oС, предпочтительно от 30oС до 38oС. Значение рН находится в пределах от 5 до 9, предпочтительно в пределах от 6,5 до 7,2. рН среды может быть скорректировано аммиаком, карбонатом кальция, различными кислотами, основаниями и буферами. Обычно выращивание в течение от 1 до 3 дней приводит к накоплению L-аргинина в культуральной жидкости.
Сбор L-аргинина может быть осуществлен путем удаления из питательной среды после выращивания нерастворимых компонент, таких как клетки, методом центрифугирования или фильтрации на мембране, с последующим сбором и очисткой L-аргинина методами ионного обмена, концентрации и фракционной кристаллизации или подобными.
Краткое описание чертежей.
На чертеже показана схема конструирования пула мутантных генов argA.
Наилучший способ осуществления изобретения.
Настоящее изобретение будет детальнее разъяснено со ссылкой на следующие примеры.
Пример 1.
<1> Мутагенез путем рандомизации фрагмента нуклеотидной последовательности гена.
BamHI-SalI фрагмент хромосомной ДНК (2.02 т.п.н.), содержащий ген argA, был клонирован в плазмиду pUC19 (плазмида pUC19-argA). Для амплификации с помощью ПЦР была использована ДНК полимераза PyrobestTM от Takara Shuzo Co. (Япония), она использовалась в условиях, рекомендованных производителем.
Для конструирования пула мутантных генов argA на первой стадии был амплифицирован фрагмент гена argA, кодирующий последовательность с 20-го остатка аминокислоты до конца NAGS (чертеж). В качестве матрицы использовалась плазмида pUC19-argA, смысловая затравка Р1: 5'-CGAGGGATTCCGCNNNNNNNNNNNNNNNATCAATACCCACCGGG-3' (SEQ ID NO:13), была сконструирована на основе последовательности нуклеотидов гена argA, в качестве антисмысловой затравки Р2 использовалась стандартная затравка М13 с обратной последовательностью. Фиксированная последовательность из 16-ти нуклеотидов на 3'-конце затравки Р1 гомологична последовательности гена argA после кодона Туr-19, а фиксированная последовательность из 13-ти нуклеотидов на 5'-конце затравки - перед кодоном His-15. Гомология к последовательности гена argA на 3'-конце затравки Р1 была использована для синтезирования фрагмента ДНК (1,75 т.п.н.) в ходе двадцати циклов ПЦР.
В качестве матрицы 100 нг плазмиды pUC19-argA было добавлено в раствор ПЦР (50 мкл), содержащий обе затравки (40 пмоль каждой). Были выполнены двадцать циклов ПЦР (94oС в течение 0,6 мин, 55oС в течение 0,5 мин, 72oС в течение 2 мин) в термоциклере модели 2400 DNA (Perkin-Elmer Co., Foster City, CA).
На второй стадии проводилось восемь циклов амплификации (94oС в течение 1 мин, 37oС в течение 1 мин, 72oС в течение 0,5 мин), в ходе которых (-) цепь этого фрагмента выступала в качестве затравки для его удлинения для получения полной последовательности гена.
На третьей стадии аликвота 10 мкл реакционной смеси добавлялась в свежую реакционную смесь (40 мкл), содержащую 100 пмоль смысловой затравки Р3: 5'-TGCCATGGTAAAGGAACGTAAAACC-3' (SEQ ID NO:14), гомологичной 5'-концу последовательности гена argA, и антисмысловой затравки Р2, затем проводилось десять циклов амплификации (94oС в течение 0,5 мин, 52oС в течение 0,5 мин, 72oС в течение 2 мин),
ДНК фрагмент (1.78 т.п.н.), кодирующий пул мутантных вариантов полноразмерного гена argA, очищался в ходе электрофореза в агарозном геле, расщеплялся ферментами NcoI (место узнавания включает начальный ATG-кодон гена argA) и HmdIII, затем лигировался в вектор рКК233-2 (Pharmacia, Sweden), расщепленный ферментами NcoI и HindIII.
Около 150 нг дотированной ДНК использовалось в последующих экспериментах для трансформации клеток Е.coli - реципиентов с тем, чтобы получить около 2000 рекомбинантных клонов в каждом случае.
<2> Выделение новых мутантов argA и влияние замен аминокислот на каталитические свойства NAGS.
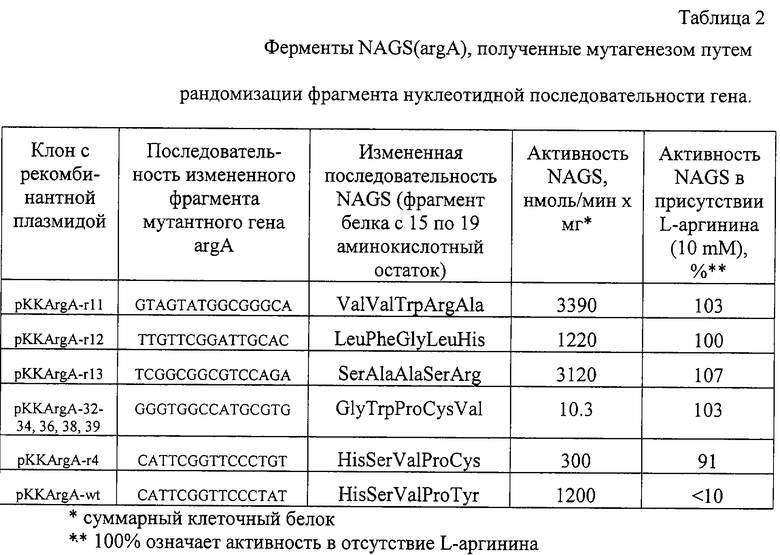
Плазмидный вектор рКК233-2 (Pharmacia, Sweden) использовался для клонирования и экспрессии вариантов гена argA. В качестве штамма Е.coli - реципиента использовался TG1 (supE hsdΔ5 thi Δ(lас-рrоАВ) F'[traD36 proAB+ lacIq lacZΔM15] ) (Sambrook, J. et al. Molecular Cloning. 1989). Отбор клеток TG1, содержащих набор рекомбинантных плазмид pKK-argA-random (с мутантными генами argA)? проводился на чашках с LB агаром. Наблюдалось замедление клеточного роста некоторых мутантных клонов и предполагалось, что этот эффект коррелирует с продукцией активного мутантного fbr фермента NAGS. Из некоторых клонов были выделены плазмиды и методом секвенирования (метод терминации цепи с использованием дидезоксинуклеотидов) была определена последовательность ДНК 5'-концевого фрагмента гена argA (таблица 2).
Для определения активности NAGS штамм Е.coli B3083 (argA-. MetB-) - ауксотроф по аргинину трансформировался полученными плазмидами. Ферменты в растворимых фракциях, полученных из рекомбинантных клеток, разрушенных ультразвуком, были частично очищены осаждением сульфатом аммония и проанализированы, как описано ниже. Активность NAGS из штаммов, содержащих плазмиды pKK-argA-r11 (3390 нмоль/мин х мг), pKK-argA-r12 (1220 нмоль/мин х мг) pKK-argA-r13 (3120 нмоль/мин х мг), значительно выше активности NAGS из штамма, содержащего плазмиду pKK-argA-r4 (300 нмоль/мин х мг). Последняя плазмида содержит мутантный ген argA с такой же заменой (Y19C), как и наиболее активный вариант гена argA с единичной заменой, описанный Rajagopal B. S. et al (Rajagopal B.S. et al. Appl. Environ. Microbiol., 1998, v.64, No.5, p.1805-1811).
Также активность NAGS в штамме, содержащем плазмиду pKK-argA(wt) (природный argA), ниже, чем в случае с pKK-argA-r11, -r12 и -r13. Уровень активности мутантных ферментов в присутствии 10 mM аргинина приблизительно одинаковый, притом что природный фермент в значительной степени ингибировался аргинином (менее одной десятой исходной активности). Эти результаты показывают, что пептидный фрагмент с 15 по 19 аминокислотный остаток отвечает за ингибирование NAGS аргинином по типу обратной связи и за уровень каталитической активности.
(Проверка активности фермента)
Ацетил коэнзим А и все химические реактивы куплены у Sigma Chemical Co., St. Louis, Mo. Для определения активности NAGS клетки Е. coli B3083 (argA-, metB-), содержащие рекомбинантые плазмиды, выращивались в среде М9 (5 мл) до поздней экспоненциальной фазы, отмывались 0,14 М раствором NaCl и ресуспендировались в 2 мл 40 mM калий-фосфатного буфера (рН 7,0) с 100 mM KCl. Клетки разрушались под действием ультразвука и центрифугировались. Фракции, содержащие NAGS, осаждались 5 объемами насыщенного (NH4)2SO4 и осадки растворялись в 2 мл 40 mM калий-фосфатного буфера (рН 7,0) с 100 mM KC1 и 30%(v/v) глицерином. Раствор с NAGS добавлялся к 0,1 мл реакционной смеси (100 mM Tris-HCl (pH 8,5), 35 mM KCl, 20 mM L-глутамата, 1,2 mM ацетил коэнзима А) и реакционная смесь инкубировалась при 37oС в течение 10 мин. Реакция останавливалась добавлением 0,3 мл этанола с последующим центрифугированием реакционной смеси. 0,95 мл 0,24 mM раствора DTNB (5,5-дитио-bis-2-нитробензоата) добавлялись к супернатанту и смесь инкубировалась в течение 15 мин. Активность NAGS определялась измерением поглощения при 412 нм.
<3> Выделение новых мутантов гена argA путем отбора среди клеток В 16-4 (pro- argD-).
Отбор мутантных генов argA из штамма Е.соli В 16-4 (pro- argD-), несущего плазмиды pKK-argA-random, осуществлялся способом, описанным выше. Рекомбинантные клоны с чашек с агаром были суспендированы в среде М9 с L-аргинином и выращивались до стационарной фазы. Аликвота культуры была суспендирована в свежей среде и процедура выращивания повторена четыре раза. После этого аликвота культуры переносилась на агаризованную среду М9, содержащую 5 мг/мл L-аргинина и 100 мкг ампицилина. Из некоторых клонов были выделены плазмиды, определены последовательности 5'-концевых фрагментов мутантных генов argA и проверен, как описано выше, уровень активности мутантных NAGS. 60% мутантов содержали последовательность -Gly-Trp-Pro-Cys-Val- (SEQ ID NO: 4) в подвергавшемся мутагенезу фрагменте фермента и обладали слабой, но fbr активностью NAGS. Очевидно, что такой мутантный белок обладает оптимальным уровнем активности NAGS для роста клеток В16-4 (pro- argD-) в использованных для отбора условиях. Таким образом, условия отбора определяют активность полученной мутантной NAGS.
<4> Продукция L-аргинина с использованием мутантных генов argA.
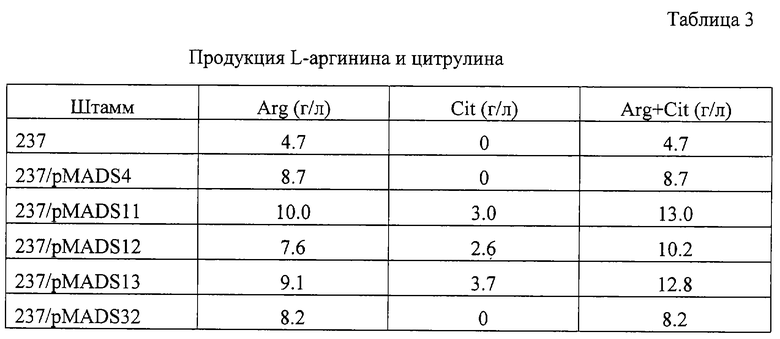
Рекомбинантные плазмиды pKKArgA-r4, 11, 12, 13 и 32 были расщеплены рестриктазами BamHI и SalI, и фрагменты, содержащие мутантные гены argA, находящиеся под контролем промотора trc, были заново клонированы в низкокопийную плазмиду pMW119 (Nippon Gene Co., Tokyo). Полученные плазмиды были названы pMADS4, pMADS11, pMADS12, pMADS13 и pMADS32 соответственно. Эти плазмиды были введены в штамм Е.coli 237 - продуцент L-аргинина (ВКПМ В-7925). Продукция L-аргинина (Arg) и цитрулина (Cit) трансформантами показана в таблице 3. Большинство штаммов - продуцентов с новыми мутантными генами NAGS обладали более высокой продуктивностью Arg и Cit, чем штамм - реципиент или штамм с известной мутантной Tyrl9Cys NAGS (pMADS4).
(Условия выращивания при ферментации в пробирках)
Питательная среда для ферментации содержала 60 г/л глюкозы, 25 г/л сульфата аммония, 2 г/л КН2РO4, 1 г/л MgSO4, 0,1 мг тиамина, 5 г/л дрожжевого экстракта Difco, 25 г/л мела на 1 л воды (рН 7,2). Глюкоза и мел были стерилизованы по отдельности. 2 мл питательной среды помещали в пробирку, высевали одну петлю испытываемого микроорганизма, выращивание продолжали при 32oС в течение 3 дней со встряхиванием.
Перечень последовательностей приведен в конце описания.





Изобретение относится к биотехнологии. L-аргинин получают культивированием бактерии, принадлежащей к роду Escherichia, трансформированной фрагментом ДНК, кодирующим мутантную N-ацетилглутамат синтазу. Мутантная N-ацетилглутамат синтаза представляет собой синтазу, в которой последовательность аминокислот, соответствующая положениям с 15 по 19 в природной N-ацетилглутамат синтазе, заменена любой из последовательностей аминокислот под номерами 1-4 (SEQ ID NOS: 1-4) или синтазу, обладающую указанными свойствами и дополнительно содержащую делеции, замены, вставки или добавления одной или нескольких аминокислот в одной или множестве положений, кроме положений с 15 по 19. В качестве штаммов-продуцентов L-аргинина могут быть использованы штаммы Escherichia coli 237/pMADS11, E.coli 237/pMADS12 или E.coli 237/pMADS13. Данное изобретение позволяет увеличить выход аргинина. 7 с. и 2 з.п. ф-лы, 1 ил., 3 табл.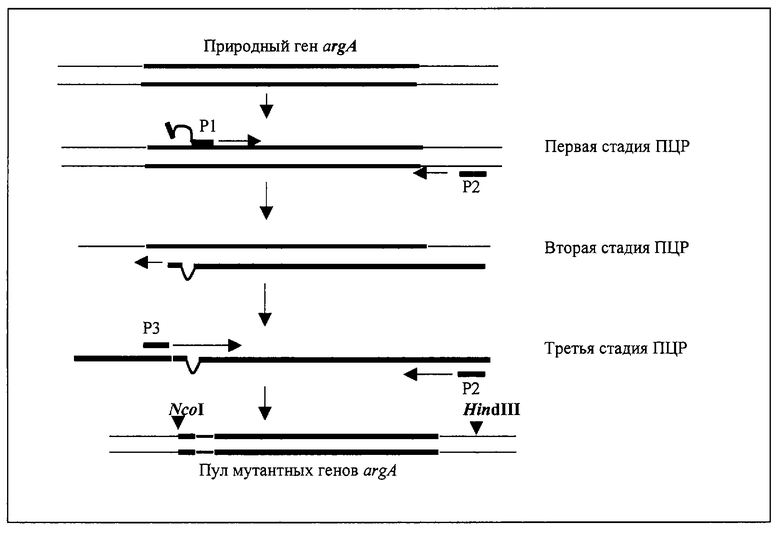
| ФРАГМЕНТ ДНК ИЗ ESCHERICHIA COLI, ОПРЕДЕЛЯЮЩИЙ ПОВЫШЕННУЮ ПРОДУКЦИЮ L-АМИНОКИСЛОТ (ВАРИАНТЫ), И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 1999 |
|
RU2175351C2 |
| US 4430430, 07.02.1984. | |||
