Область техники, к которой относится изобретение
Настоящее изобретение относится к микробиологической промышленности, в частности к способу получения L-аминокислот, таких как L-аргинин, ферментацией бактерии семейства Enterobacteriaceae, которая модифицирована таким образом, что она содержит ген argJ, кодирующий фермент, имеющий, по меньшей мере, орнитинацетилтрансферазную активность, и нарушенный ген argE, кодирующий N-ацетилорнитиндеацетилазу.
Уровень техники
Традиционно L-аминокислоты в промышленности получают ферментационным методом с использованием штаммов микроорганизмов, полученных из природных источников, или их мутантов. Обычно, микроорганизмы модифицируются для увеличения продукции L-аминокислот.
Описано много способов повышения продукции L-аминокислот, например, путем трансформации микроорганизмов рекомбинантной ДНК (см., например, патент США №4,278,765) и изменения регуляторных участков, таких как промотор, лидерная последовательность и/или аттенюаторы или другие, известные для специалиста в данной области (см., например, Патентную заявку США №20060216796 А1 и WO9615246 А1). Другие способы, повышающие выход, включают повышение активности ферментов, вовлеченных в биосинтез аминокислот, и/или снятие ингибирования ключевых ферментов продуцируемой L-аминокислотой по типу обратной связи (см., например, публикацию Патента Японии №56-18596 (1981), WO 95/16042 или Патенты США №5,661,012 и 6,040,160).
Другой способ повышения продукции L-аминокислот заключается в ослаблении экспрессии одного или нескольких генов, вовлеченных в деградацию целевой L-аминокислоты, генов, участвующих в удалении предшественников целевой L-аминокислоты из биосинтетического пути L-аминокислоты, генов, вовлеченных в перераспределение потоков углерода, азота и фосфора, и генов, кодирующих токсины, и т.д.
Принято считать, что, например, в бактерии биосинтез L-аргинина из L-глутамата может проходить по двум путям, линейному или циклическому, в зависимости от конкретного микроорганизма (Cunin R. et al., Microbiol. Rev., 1986, 50:314-352). В бактериях, таких как бактерия, принадлежащая семейству Enterobacteriaceae, в частности, Escherichia coli (E.coli) имеет место линейный путь, который включает 8 стадий до полного образования L-аргинина из L-глутамата (Vogel H.J. и MacLellan W.L., Methods Enzymol., 1970, 17A, 265-269). Биосинтез инициирован ацетилированием L-глутамата аминокислотной N-ацетилтрансферазой (ЕС 2.3.1.1, также называемой как N-ацетил-L-глутаматсинтетаза), кодируемой геном argA. Последующие биосинтетические реакции катализируются ферментами, обычно называемыми как N-ацетилглутаматкиназа, N-ацетил-γ-глутамилфосфатредуктаза, N-ацетилорнитинаминотрансфераза, N-ацетилорнитиндеацетилаза, орнитинкарбамоилтрансфераза, аргининосукцинатсинтетаза и аргининосукцинатлиаза, кодируемые генами argB, argC, argD, argE, argF, argG и argH, соответственно. Ацетильная группа, снимаемая N-ацетилорнитиндеацетилазой (ArgE) с N-ацетилорнитина с образованием орнитина, связывается с коферментом A (HS-CoA, CoA) с образованием ацетилированного кофермента A (AcS-CoA, ацетил-СоА), который выступает как донор ацетильной группы в реакции ацилирования L-глутамина, катализируемой аминокислота-N-ацетилтрансферазой (argA).
Циклический путь биосинтеза L-аргинина был обнаружен в прокариотах, таких как Corynebacterium glutamicum (Udaka S. и Kinoshita S., J. Gen. Appl. Microbiol., 1958, 4:272-282; Sakanyan V. et al., Microbiology, 1996, 142:99-108), Bacillus species (Sakanyan V. et al., J. Gen. Microbiol., 1992, 138:125-130), Thermotoga neapolitana (Marc F. et al., Eur. J. Biochem., 2000, 267(16):5217-5226), и эукариотах (De Deken R.H., Biochim. Biophys. Acta., 1962, 78:606-616). В отличие от линейного пути, в циклическом пути перенос ацетильной группы с N-ацетилорнитина на L-глутамат катализируется бифункциональной орнитинацетилтрансферазой/N-ацетилглутаматсинтазой (ЕС 2.3.1.35/2.3.1.1), кодируемой геном argJ.
Помимо бифункционального фермента, кодируемого геном argJ, известен белок ArgJ, кодируемый геном argJ и проявляющий исключительно орнитинацетилтрансферазную активность (Haas D. et al., Eur. J. Biochem., 1972, 31:290-295; Marc F. et al., Eur. J. Biochem., 2000, 267(16):5217-5226). Монофункциональный и бифункциональный ферменты ArgJ можно различить двумя способами: (i) ферментативным анализом с использованием двух доноров ацетильных групп, таких как N-ацетилорнитин и AcS-CoA; и (ii) комплементационным тестом с использованием мутантов argE и argA Е.coli с клонированием гена argJ. Монофункциональный фермент ArgJ переносит ацетильную группу от N-ацетилорнитина к L-глутамату и комплементирует мутант argE, в то время как бифункциональный фермент ArgJ переносит ацетильную группу от N-ацетилорнитина и AcS-CoA и комплементирует штаммы, мутантные по argE и argA.
Новый биосинтетический путь L-аргинина был недавно обнаружен в Xanthomonas campestris, где N-ацетилорнитин превращается в N-ацетилцитруллин под действием ацетилорнитинкарбамоилтрансферазы, кодируемой геном argF', и цитруллин образуется из N-ацетилцитруллина под действием ArgE (Shi D. et al., J. Biol. Chem., 2005, 280:14366-14369).
Известна бактерия Е.coli, изначально имеющая линейный путь, была модифицирована таким образом, что она содержит ген argJ из Bacillus stearothennophilus или Thermotoga neapolitana (T. neapolitana), который кодирует бифункциональный фермент ArgJ, чтобы инициировать менее энергоемкий циклический путь и, таким образом, увеличить продукцию L-аргинина посредством модифицированной бактерии (патент США №6,897,048 В2).
В микроорганизмах, изначально использующих линейный биосинтетический путь или имеющих модифицированный линейный путь, который функционирует как циклический путь, аминокислота-N-ацетилтрансфераза (N-ацетил-L-глутаматсинтетаза) (ArgA) может быть необходима для инициации и поддержания биосинтеза L-аргинина посредством поставки N-ацетилглутамата. Таким образом, для увеличения продукции L-аргинина с помощью рекомбинантного штамма Е.coli, количество копий гена argA может быть увеличено клонированном гена дикого типа argA на плазмидных векторах и включением их в штамм, имеющий клонированный ген argJ (патент США №6,897,048 В2). С другой стороны, ген argA, кодирующий мутантную аминокислота-N-ацетилтрансферазу с устойчивостью к ингибированию по принципу обратной связи L-аргинином (Eckhardt Т. et al., Mol. Gen. Genet, 1975, 138:225-232), может быть введен в штамм Е.coli для усиления продукции L-аргинина (ЕР 1170361 А2).
Несмотря на то что наличие положительного эффекта на продукцию L-аргинина от использования бифункционального белка ArgJ, дополнительных копий гена argA и/или мутантной N-ацетил-L-глутаматсинтетазы (мутантного ArgA), и т.п., четко установлено, подходы для дальнейшего увеличения продукции L-аргинина с помощью микроорганизмов не очевидны.
В настоящее время нет данных, описывающих влияние нарушенного гена argE так, что активность N-ацетилорнитиндеацетилазы (ArgE) уменьшена или отсутствует, на продукцию L-аргинина модифицированными бактериальными штаммами семейства Enterobacteriaceae, имеющих гетерологичный ген argJ.
Раскрытие сущности изобретения
Авторы настоящего изобретения предположили, что в микроорганизмах, например, в бактериях, имеющих гетерологичный ген argJ, который кодирует монофункциональный фермент орнитинацетилтрансферазу или бифункциональный фермент орнитинацетилтрансферазу/N-ацетилглутаматсинтазу, ослабление экспрессии гена argE или инактивация гена argE может быть источником дополнительной энергии из процесса, связанного с переносом ацетильной группы от N-ацетилорнитина к L-глутамату, таким образом, содействуя продукции L-аргинина с помощью модифицированного штамма по сравнению с родительским штаммом. Авторы настоящего изобретения предположили, что фермент ArgJ, имеющий моно- или двойную активность и кодируемый геном argJ, клонированным из микроорганизма-донора, может восстановить биосинтез L-аргинина в штаммах, мутантных по argE или argA и argE генам. С этой точки зрения, искусственный циклический путь, будучи инициированный путем реализации первой биосинтетической реакции с целью продукции N-ацетилглутамата, может функционировать посредством прямого ацилирования L-глутамата ацетильной группой, полученной из N-ацетилорнитина, нежели из более энергетически богатого AcS-CoA.
Цель настоящего изобретения - предоставление бактерии, принадлежащей к семейству Enterobacteriaceae, которая может принадлежать к роду Escherichia, более точно к виду Escherichia coli, которая имеет активность орнитинацетилтрансферазы или орнитинацетилтрансферазы/N-ацетилглутаматсинтазы (ArgJ) и в которой N-ацетилорнитиндеацетилаза (ArgE) инактивирована или активность ArgE уменьшена.
Другая цель настоящего изобретения - предоставление способа получения L-аминокислот, таких как L-аргинин, с использованием бактерии семейства Enterobacteriaceae как здесь описано.
Эти цели были достигнуты благодаря обнаружению того факта, что инактивация или ослабление экспрессии гена argE в бактерии семейства Enterobacteriaceae, имеющей гетерологичный ген argJ, приводит к повышенной продукции L-аргинина.
Цель настоящего изобретения - предоставление бактерии-продуцента L-аргинина, принадлежащей к семейству Enterobacteriaceae, имеющей рекомбинантную ДНК, которая содержит ген argJ, кодирующий фермент, имеющий, по меньшей мере, орнитинацетилтрансферазную активность, отличающуюся тем, что бактерия модифицирована таким образом, что она содержит N-ацетилорнитиндеацетилазу с нарушенной активностью.
Другая цель настоящего изобретения - предоставление описанной здесь бактерии, отличающейся тем, что ген argJ происходит из микроорганизма, имеющего фермент, обладающий активностью орнитинацетилтрансферазы или орнитинацетилтрансферазы/N-ацетилглутаматсинтазы.
Другая цель настоящего изобретения - предоставление описанной здесь бактерии, отличающейся тем, что ген argJ происходит из микроорганизма, принадлежащего к семейству, выбранному из группы, состоящей из семейств Thermotogaceae, Bacillaceae и Methanocaldococcaceae.
Другая цель настоящего изобретения - предоставление описанной здесь бактерии, отличающейся тем, что ген argJ происходит из вида Thermotoga neapolitana.
Другая цель настоящего изобретения - предоставление описанной здесь бактерии, отличающейся тем, что ген argJ кодирует белок, выбранный из группы, состоящей из:
(A) белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:2, который имеет активность орнитинацетилтрансферазы/N-ацетилглутаматсинтазы;
(B) варианта белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:2, но которая содержит замену, делецию, вставку и/или добавление одного или нескольких аминокислотных остатков и имеет активность орнитинацетилтрансферазы/N-ацетилглутаматсинтазы в соответствии с аминокислотной последовательностью, представленной в SEQ ID NO:2;
и
(C) их комбинации.
Другая цель настоящего изобретения предоставление описанной здесь бактерии, отличающейся тем, что N-ацетилорнитиндеацетилаза является белком, выбранным из группы, состоящей из:
(D) белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:4;
(Е) варианта белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:4, но которая содержит замену, делецию, вставку и/или добавление одного или нескольких аминокислотных остатков и имеет активность N-ацетилорнитиндеацетилазы в соответствии с аминокислотной последовательностью, представленной в SEQ ID NO:4; и
(F) их комбинации.
Другая цель настоящего изобретения - предоставление описанной здесь бактерии, отличающейся тем, что активность N-ацетилорнитиндеацетилазы понижена.
Другая цель настоящего изобретения - предоставление описанной здесь бактерии, отличающейся тем, что активность N-ацетилорнитиндеацетилазы отсутствует.
Другая цель настоящего изобретения - предоставление описанной здесь бактерии, отличающейся тем, что бактерия принадлежит к роду Escherichia.
Другая цель настоящего изобретения - предоставление описанной здесь бактерии, отличающейся тем, что бактерия принадлежит к виду Escherichia coli.
Другая цель настоящего изобретения - предоставление описанной здесь бактерии, отличающейся тем, что бактерия принадлежит к роду Pantoea.
Другая цель настоящего изобретения - предоставление описанной здесь бактерии, отличающейся тем, что бактерия принадлежит к виду Pantoea ananatis.
Цель настоящего изобретения - предоставление способа получения L-аргинина или его соли, включающего:
(i) выращивание описанной выше бактерии в питательной среде;
(ii) накопление L-аргинина или его соли в бактерии или культуральной жидкости, или обоих; и, если необходимо,
(iii) выделение L-аргинина или его соли из бактерии или культуральной жидкости.
Настоящее изобретение детально описано ниже.
Краткое описание фигур
Фигура 1 показывает схему интеграции гена argJ в хромосому штамма Е.coli MG1655ΔargAΔargR.
Фигура 2 показывает схему конструирования штамма Е.coli MG1655ΔargAΔargRΔartP-J::Pnlp8φ10argJargEm24::Cm.
Наилучший способ осуществления изобретения
1. Бактерия
Термин «бактерия-продуцент L-аминокислоты» может означать бактерию семейства Enterobacteriaceae, которая способна продуцировать, экскретировать или секретировать и вызывать накопление L-аминокислоты в культуральной жидкости или бактериальных клетках, когда указанная бактерия выращивается (культивируется) в питательной среде.
Термин «способность бактерии продуцировать L-аминокислоту» может означать способность бактерии продуцировать, экскретировать или секретировать и вызывать накопление L-аминокислоты в культуральной жидкости или бактериальных клетках, что приводит к накоплению L-аминокислоты в количествах, достаточных для ее выделения из культуральной жидкости или бактериальных клеток, когда указанная бактерия выращивается (культивируется) в питательной среде.
Термин «L-аминокислота» может означать L-аланин, L-аргинин, L-аспарагин, L-аспарагиновую кислоту, L-цистеин L-глутаминовую кислоту, L-глутамин, L-гистидин, L-изолейцин, L-лейцин, L-лизин, L-метионин, L-фенилаланин, L-пролин, L-серин, L-треонин, L-триптофан, L-тирозин и L-валин.
Бактерия может обладать способностью к продукции аминокислот изначально, в соответствии со своими природными характеристиками, или может быть модифицирована с помощью мутаций (мутагенеза) или технологий рекомбинантных ДНК таким образом, чтобы она получила способность продуцировать аминокислоты.
Бактерия, принадлежащая к семейству Enterobacteriaceae, может быть выбрана из бактерий, относящихся к родам, входящим в состав этого семейства, таких как Enterobacter, Erwinia, Escherichia, Klebsiella, Morganella, Pantoea, Photorhabdus, Providencia, Salmonella, Yersinia и т.д., и способных продуцировать L-аминокислоту. Более конкретно, могут быть использованы бактерии, классифицируемые как принадлежащие к семейству Enterobacteriaceae в соответствии с таксономией, используемой в базе данных NCBI (National Center for Biotechnology Information) (http://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi?id=543). Предпочтительна для модификаций согласно данному изобретению бактерия, относящаяся к роду Escherichia, Enterobacter или Pantoea.
Выбор штаммов бактерий, принадлежащих к роду Escherichia, которые могут быть модифицированы в настоящем изобретении, не ограничен каким-либо образом, однако, в качестве примеров, бактерии рода Escherichia, описанные в книге Neidhardt et al. (Bachmann, B.J., Derivations и genotypes of some mutant derivatives of E.coli K-12, p.2460-2488. In F.C. Neidhardt et al. (ed.), E.coli и Salmonella: cellular и molecular biology, 2nd ed. ASM Press, Washington, D.C., 1996) могут быть включены в число бактерий согласно настоящему изобретению. В качестве конкретного примера могут быть взяты штаммы E.coli., такие как E.coli W3110 (АТСС 27325), E.coli MG1655 (АТСС 47076) и т.д., которые происходят из исходного штамма дикого типа, т.е. штамма E.coli K-12. Этот штамм может быть получен, в частности, из Американской коллекции типовых культур «American Type Culture Collection, (АТСС)» (P.O. Box 1549, Manassas, VA 20108, United States of America). Каждому штамму присвоен индивидуальный регистрационный номер, и штаммы могут быть заказаны согласно регистрационному номеру (см. ссылку www.atcc.org). Регистрационные номера штаммов находятся в списке каталога Американской коллекции типовых культур «American Type Culture Collection, АТСС».
Примеры бактерий Enterobacter включают Enterobacter agglomerans, Enterobacter aerogenes и т.д. Примеры бактерии Pantoea включают Pantoea ananatis и т.д. Недавно некоторые виды Enterobacter agglomerans были переклассифицированны как Pantoea agglomerans, Pantoea ananatis или Pantoea stewartii на основе анализа нуклеотидной последовательности 16S рРНК и других доказательств. Бактерии, относящиеся к роду Enterobacter или Pantoea, могут быть использованы в соответствии с настоящим изобретением, так как принадлежат семейству Enterobacteriaceae. Полученные с использованием технологий генной инженерии штаммы Pantoea ananatis, такие как штамм Pantoea ananatis AJ13355 (PERM ВР-6614), штамм AJ13356 (PERM BP-6615), штамм AJ13601 (PERM BP-7207) и их производные могут быть использованы в соответствии с настоящим изобретением. Эти штаммы были классифицированы как Enterobacter agglomerans при выделении, и они депонированы как Enterobacter agglomerans. Однако позднее они были классифицированы как Pantoea ananatis на основе анализа нуклеотидной последовательности 16S рРНК и других доказательств.
Термин «бактерия-продуцент L-аргинина» может означать бактерию, которая способна продуцировать, экскретировать или секретировать и вызывать накопление L-аргинина в культуральной жидкости или бактериальных клетках в количествах больших, чем штамм дикого типа или родительский штамм Е.coli, такой как Е.coli K-12, когда бактерия выращена в питательной среде. Способность продуцировать L-аргинин может означать способность бактерии продуцировать, экскретировать или секретировать и вызывать накопление L-аргинина в питательной среде в количестве не менее 0,5 г/л или не менее чем 1,0 г/л, так что L-аргинин может быть выделен из культуральной жидкости и/или бактериальных клеток, когда указанная бактерия выращивается (культивируется) в питательной среде.
Примеры родительских штаммов, которые могут использоваться для получения бактерии, продуцирующей L-аргинин, включают, но не ограничиваются данными примерами, штаммы, принадлежащие к роду Escherichia, такие как штамм Е.coli 237 (ВКПМ В-7925) (патентная заявка США 2002/058315 А1) и его производные штаммы, имеющие мутантную N-ацетилглутаматсинтазу (ЕР 1170361 А2), штамм Е.coli 382 (ВКПМ В-7926) (ЕР1170358 А1) и штамм, продуцирующий L-аргинин, имеющий введенный ген argA (ЕР1170361 А1), и подобные им.
Бактерия-продуцент L-аргинина семейства Enterobacteriaceae, может быть модифицирована далее таким образом, что она содержит гетерологичный ген argJ, который кодирует монофункциональный фермент орнитинацетилтрансферазу или бифункциональный фермент орнитинацетилтрансферазу/N-ацетилглутаматсинтазу (ArgJ). Например, бактерия семейства Enterobacteriaceae может иметь ген argJ, кодирующий моно- или бифункциональный фермент ArgJ, устойчивый к ингибированию по принципу обратной связи L-аргинином. Ген argJ может быть выделен из термофильного микроорганизма, такого как микроорганизм, принадлежащий к семейству Thermotogaceae, и более конкретно - к роду Thermotoga, например, Т. пеароШапа (Т. neapolitana). Ген argJ также может быть выделен из других бактериальных источников, таких как Bacillus stearothermophUus (или Geobacillus stearothermophilus) семейства Bacillaceae или архей, таких как Methanocaldococcus jannaschii (М. jannaschii) (ранее Methanococcus jannaschii) семейства Methanocaldococcaceae, и им подобных. Однако, различные бактериальные источники гена argJ могут использоваться при условии, что ген argJ кодирует фермент имеющий, по меньшей мере, активность орнитинацетилтрансферазы.
Бактерия-продуцент L-аргинина семейства Enterobacteriaceae может быть модифицирована далее таким образом, что она содержит инактивированный ген argE или ген argE с ослабленной экспрессией, так что N-ацетилорнитиндеацетилаза (ArgE) в модифицированной бактерии неактивна или активность N-ацетилорнитиндеацетилазы снижена по сравнению с немодифицированной бактерией.
Термин «бактерия-продуцент L-аргинина» может означать бактерию семейства Enterobacteriaceae, имеющую ген argA, кодирующий дикий тип или мутант аминокислота-N-ацетилтрансферазы (ArgA). Фермент ArgA, устойчивый к ингибированию по принципу обратной связи L-аргинином, может быть назван как мутантная аминокислота-N-ацетилтрансфераза. Например, мутант белка ArgA, устойчивый к ингибированию по принципу обратной связи L-аргинином, может быть получен из белка дикого типа ArgA путем замены аминокислотной последовательности в положении с 15 по 19 на другую аминокислотную последовательность, как указано В ЕР 1170361 А2. Мутантный белок ArgA, содержащий в аминокислотной последовательности замену, делению, вставку и/или добавление одного или нескольких аминокислотных остатков в одном или более положениях, отличных от положений 15-19, также может быть назван как мутантный белок ArgA при условии, что третичная структура мутантной аминокислота-N-ацетилтрансферазы изменена незначительно по сравнению белком дикого типа или активность мутантного фермента не ухудшилась и существует устойчивость к ингибированию по принципу обратной связи L-аргинином.
Термин «бактерия-продуцент L-аргинина» также может означать бактерию семейства Enterobacteriaceae, как описано выше, имеющую усиленный, ослабленный и/или инактивированный ген argA, кодирующий дикий тип или мутант аминокислота-N-ацетилтрансферазы.
Термин «бактерия-продуцент L-аргинина» также может означать бактерию семейства Enterobacteriaceae, модифицированную далее таким образом, что она лишена ArgR-зависимой транскрипционной репрессии. ДНК-связывающий транскрипционный двойной регулятор ArgR, в микроорганизмах участвующий в отрицательном контроле биосинтетического пути L-аргинина, может быть инактивирован, например, инактивацией гена argR, кодирующего ArgR.
Термин «белок дикого типа» может означать нативный белок, естественным образом продуцируемый штаммом дикого типа или родительским бактериальным штаммом семейства Enterobacteriaceae, например, штаммом дикого типа Е.coli MG1655. Белок дикого типа может быть кодирован геном дикого типа, или немодифицированным геном, естественным образом встречающимся в геноме бактерии дикого типа.
Термин «бактерия, модифицированная таким образом, что она содержит ген argJ» может означать, что бактерия, принадлежащая к бактерии первого вида, естественным образом не содержащая ген argJ, и, таким образом, называемая как бактерия-реципиент, модифицирована таким образом, чтобы содержать одну или более рекомбинантных молекул ДНК, имеющих ген argJ, синтезированный и/или происходящий и введенный из бактерии, принадлежащей ко второму виду, называемой как бактерия-донор, которая отлична от бактерии первого вида. Примером модификации по введению рекомбинантной ДНК может быть экспрессия гетерологичного гена. Например, бактерия-реципиент может принадлежать к семейству Enterobacteriaceae, например, вида Е.coli; бактерия-донор может принадлежать к термофильной бактерии, например, вида Т. neapolitana.
Термин «бактерия, модифицированная таким образом, что она содержит рекомбинантную ДНК» может означать, что бактерия модифицирована обычными способами так, что она содержит экзогенную ДНК. К обычным способам относятся, например, трансформация, трансфекция, инфекция, конъюгация и мобилизация. Трансформация, трансфекция, инфекция, конъюгация или мобилизация бактерии молекулой ДНК, кодирующей белок, могут сообщить бактерии способность синтезировать белок, кодируемый молекулой ДНК. Способы трансформации, трансфекции, инфекции, конъюгации и мобилизации включают любые известные ранее описанные способы. Например, известен способ эффективной трансформации и трансфекции ДНК путем обработки клеток-реципиентов Escherichia coli K-12 хлоридом кальция с целью увеличения проницаемости клеток для ДНК (Mandel M. и Higa A., Calcium-dependent bacteriophage DNA infection, J. Mol. Bioi, 1970, 53:159-162). Описаны способы специализированной и/или общей трансдукции (Morse M.L. et al., Transduction in Escherichia coli K-12, Genetics, 1956, 41(1):142-156; Miller J.H., Experiments in Molecular Genetics. Cold Spring Harbor, N.Y.: Cold Spring Harbor La. Press, 1972). Могут быть применены другие способы случайной и/или направленной интеграции ДНК в геном хозяина, например, «Mu-зависимая интеграция/амплификация» (Akhverdyan et al., Appl. Microbiol. Biotechnol., 2011, 91:857-871), «Red/ET-зависимая» или «λRed/ET-зависимая интеграция» (Datsenko K.A. и Wanner B.L., Proc. Natl. Acad. Sci. USA 2000, 97(12):6640-45; Zhang Y., et al., Nature Genet, 1998, 20:123-128). Более того, для многократных вставок желаемых генов в дополнение к Mu-зависимой репликативной транспозиции (Akhverdyan et al., Appl. Microbiol. Biotechnol., 2011, 91:857-871) и химически индуцированной хромосомной эволюции, основанной на recA-зависимой гомологичной рекомбинации, приводящей к амплификации желаемых генов (Туо K.E.J. et al., Nature Biotechnol., 2009, 27:760-765), могут использоваться другие подходы, основанные на различных комбинациях транспозиции, сайт-направленной и/или гомологичной Red/ET-зависимой рекомбинации, и/или P1-зависимой общей трансдукции (см., например, Minaeva N. et al., ВМС Biotechnology, 2008, 8:63; Koma D. et al., Appl. Microbiol. Biotechnol., 2012, 93(2):815-829).
Гетерологичная экспрессия гена argJ в микроорганизмах хозяина может быть осуществлена путем замены редких (мало используемые в организме хозяина) кодонов на синонимичные средне или часто используемые кодоны, при этом термин «использование кодона» может быть определен как число раз (частота) трансляции кодона в клетке организма в единицу времени или как средняя частота встречаемости кодона в сиквенированных белок-кодирующих рамках считывания (Zhang S.P. et al., Gene, 1991, 105(1):61-72). Использование кодонов в организме может быть найдено в базе данных использования кодонов Codon Usage Database, которая является расширенной веб-версией CUTG (Codon Usage Tabulated from GenBank) (http://www.kazusa.or.jp/codon/; Nakamura Y. et al., Codon usage tabulated from the international DNA sequence databases: status for the year 2000, Nucl. Acids Res., 2000, 28(1):292). В Е.coli такие мутации могут включать, не ограничиваясь этим, замену редких Arg-кодонов AGA, AGG, CGG, CGA на CGT или CGC; редкого Ile-кодона АТА на АТС или АТТ; редкого Leu-кодона СТА на CTG, СТС, СТТ, ТТА или TTG; редкого Pro-кодона ССС на CCG или ССА; редкого Ser-кодона TCG на ТСТ, ТСА, ТСС, AGC или AGT; редких Gly-кодонов GGA, GGG на GGT или GGC; и т.д. Замена мало используемых кодонов на синонимичные часто используемые кодоны предпочтительна. Замена редких и/или мало используемых кодонов на синонимичные средне или часто используемые кодоны может комбинироваться с со-экспрессией генов, кодирующих тРНК, распознающие редкие кодоны.
Термин «бактерия, модифицированная таким образом, что она содержит ген argE с нарушенной экспрессией» может означать, что бактерия модифицирована таким образом, что в модифицированной бактерии экспрессия гена argE ослаблена или ген argE инактивирован.
Термин «экспрессия гена argE ослаблена» может означать, что количество белка ArgE в модифицированной бактерии, в которой экспрессия гена argE ослаблена, снижено по сравнению с немодифицированной бактерией, например, штаммом дикого типа или родительским штаммом.
Термин «экспрессия гена argE ослаблена» также может означать, что модифицированная бактерия содержит модифицированный ген argE, который кодирует мутантный белок ArgE, имеющий пониженную активность по сравнению с белком ArgE дикого типа, или функционально связанный с геном участок, включающий последовательности, контролирующие экспрессию гена, такие как промоторы, энхансеры, аттенюаторы, рибосома-связывающие участки (RBS) и т.д., модифицированные с целью снизить уровень экспрессии гена argE, и другие примеры (см., например, WO95/34672; Carrier T.A. и Keasling J.D., Biotechnol. Prog., 1999, 15:58-64).
Подобные определения могут быть даны для терминов «экспрессия гена argR ослаблена» и «экспрессия гена argA ослаблена».
Экспрессия генов argE, argA и/или argR может быть ослаблена путем замены последовательности, контролирующей экспрессию гена, такой как промотор на хромосомной ДНК, на более слабую. Сила промотора определяется частотой актов инициации синтеза РНК. Примеры методов для оценки силы промотора описаны в Goldstein et al., Prokaryotic promoters in biotechnology, Biotechnol. Annu. Rev., 1995, 1:105-128, и т.д. Кроме того, также возможно ввести нуклеотидную замену в промоторный участок целевого гена таким образом, чтобы ослабить силу промотора, как описано в Международной патентной публикации WO00/18935. Кроме того, известно, что замена нескольких нуклеотидов в области между последовательностью Шайна-Дальгарно (Shine-Dalgarno, SD) и стартовым кодоном на рибосома-связывающем участке (RBS), в частности, в последовательности, расположенной выше и сразу за стартовым кодоном, значительно влияет на эффективность трансляции мРНК. Подобная модификация RBS может комбинироваться с уменьшением уровня транскрипции генов argE, argA и/или argR.
Экспрессия генов argE, argA и/или argR также может быть ослаблена путем вставки транспозона или IS-фактора в кодирующий участок гена (патент США №5,175,107), или в участок, контролирующий экспрессию гена, или в близлежащий или удаленный участок по отношению к структурной части гена argE, argA и/или argR, или с помощью обычных методов, таких как мутагенез посредством УФ-излучения или нитрозогуанидина (N-метил-N'-нитро-N-нитрозогуанидин). Кроме того, мутация может быть сайт-направленно введена с помощью известных методов по изменению хромосомы, основанных, например, на λRed/ET-зависимой рекомбинации.
Термины «ферментативная активность ArgE снижена», «ферментативная активность ArgA снижена» и «ферментативная активность ArgR снижена» могут означать, что ферментативная активность N-ацетилорнитиндеацетилазы (ArgE), аминокислота-N-ацетилтрансферазы (ArgA) и/или регулятора ArgR, соответственно, ниже, чем в немодифицированном штамме, например, штамме дикого типа бактерии, принадлежащей к семейству Enterobacteriaceae, более конкретно, к виду Е.coli. Например, ферментативная активность N-ацетилорнитиндеацетилазы (ArgE), аминокислота-N-ацетилтрансферазы и/или регулятора ArgR может быть уменьшена за счет инактивации соответствующего гена.
Ферментативная активность фермента, кодируемого геном(ми) argE, argA и/или argR также может быть уменьшена за счет введения мутации в ген на хромосоме так, чтобы внутриклеточная активность белка, кодируемого геном(ми) argE, argA и/или. argR была уменьшена по сравнению с немодифицированным штаммом. Такой мутацией в гене(ах) или перед геном в структуре оперона может быть замена одного или нескольких оснований, приводящая к замене аминокислотного остатка в белке, кодируемом геном(ами) (миссенс-мутация), введение стоп-кодона (нонсенс-мутация), делеция одного или нескольких оснований, приводящая к сдвигу рамки считывания, вставка гена, сообщающего устойчивость к антибиотику, частичное или полное удаление генов в геноме (Qiu Z. и Goodman M.F., J. Biol. Chem. 1997, 272:8611-8617; Kwon D.H. et al., J. Antimicrob. Chemother. 2000, 46:793-796).
В модифицированной бактерии количество белка ArgE, кодируемого геном argE, может быть понижено до такого уровня, что остаточная активность ArgE меньше или около 3%, или меньше или около 2%, или меньше или около 1% от исходной активности, но выше чем ноль по сравнению с немодифицированной бактерией.
В модифицированной бактерии специфическая активность ArgE, кодируемого геном argE, может быть понижена до такого уровня, что остаточная специфическая активность ArgE не менее чем 1000 нмоль/мг мин, 750 нмоль/мг мин, 500 нмоль/мг мин, 250 нмоль/мг мин, 100 нмоль/мг мин, 50 нмоль/мг мин, 25 нмоль/мг мин, 10 нмоль/мг мин, 5 нмоль/мг мин или около 3 нмоль/мг мин по результатам разброса экспериментальных данных. Специфическая активность бактериального ArgE в пересчете на 1 мг неочищенного белка может быть определена в неочищенном экстракте разрушенных ультразвуком бактериальных клетках описанным способом (Takahara K. et al. FEBS J., 2005, 272:5353-5364). Концентрация неочищенного белка может быть определена методом Бредфорда (Bradford M.M., Anal. Biochem., 1976, 72:248-254), используя в качестве стандарта бычий сывороточный альбумин.
Ферментативная активность N-ацетилорнитиндеацетилазы (ArgE) может быть понижена за счет изменения условий ферментации, таких как кислотность питательной среды (рН), концентрация кофактора(ов), таких как ионы металлов, температура, ионная сила и т.д.
Термин «ген argE инактивирован» может означать, что модифицированный ген кодирует полностью неактивный или нефункциональный белок. Также возможно, что модифицированный участок ДНК не способен к естественной экспрессии гена благодаря делеции части гена или целого гена, сдвигу рамки считывания гена, введению миссенс/нонсенс мутации (ий) или модификации смежного участка гена, включению последовательностей, контролирующих экспрессию гена, таких как промотор(ы), энхансер(ы), аттенуатор(ы), рибосома-связывающий(ие) сайт(ы) и т.д. Инактивация гена также может быть осуществлена обычными способами, такими как мутагенез, путем обработки микроорганизмов, например, УФ-излучением или нитрозогуанидином (N-метил-N'-нитро-N-нитрозогуанидин), сайт-направленный мутагенез, нарушение структуры гена с использованием гомологичной рекомбинации и/или мутагенеза за счет вставки-делеции (Yu D. et al., Proc. Natl. Acad. Sci. USA, 2000, 97(12):5978-83; Datsenko K.A. и Wanner B.L., Proc. Natl. Acad. Sci. USA 2000, 97(12):6640-45), также называемого как «λRed/ET-зависимая интеграция».
Подобные определения могут быть даны для терминов «ген argR инактивирован» и «ген argA инактивирован».
Термин «активность ArgE отсутствует» равнозначен термину «белок ArgE инактивирован» и также может означать, что активность ArgE полностью отсутствует, то есть равна нулю, или активность ArgE ниже определенного уровня, когда активность измеряется с использованием метода, описанного в работе Takahara K. et al. FEBS J., 2005, 272:5353-5364. В качестве контроля сравнения могут служить, например, бактерии дикого типа Enterobacteriaceae, включая штамм Е.coli MG1655, и т.п. В модифицированной бактерии специфическая активность ArgE, кодируемого геном argE, может быть снижена до такого уровня, что остаточная специфичная активность argE равняется нулю или не более чем 0,01 нмоль/мг мин, 0,05 нмоль/мг мин, 0,1 нмоль/мг мин или около 0,5 нмоль/мг мин по результатам разброса экспериментальных данных и при определении уровня активности описанным выше методом.
Как описано выше, активность белка ArgE (N-ацетилорнитиндеацетилазы) может быть снижена и/или отсутствовать. Следовательно, термин «активность ArgE нарушена» может пониматься как «активность ArgE снижена» и/или «активность ArgE отсутствует», как описано выше.
Термин «повышенная экспрессия гена argA» может означать, что общая ферментативная активность соответствующего гену белка, ArgA, увеличена, например, введением и/или увеличением числа копий гена argA в бактериальном геноме или усилением активности в пересчете на молекулу белка (может быть названа как специфическая активность), кодируемого этим геном, по сравнению с немодифицированным штаммом, таким как штамм дикого типа или родительский штамм. Бактерия может быть модифицирована так, что активность белка ArgA в пересчете на клетку увеличена до 150% или более, 200% или более, 300% или более от активности белка в немодифицированном штамме, В качестве примера немодифицированного штамма-сравнения можно привести штаммы дикого типа микроорганизма семейства Enterobacteriaceae, такие как штамм Е.coli MG1655 (АТСС 47076), штамм W3110 (АТСС 27325), штамм Pantoea ananatis AJ13335 (PERM BP-6614) и т.д.
Термин «повышенная экспрессия гена argA» также может означать, что уровень экспрессии гена argA в модифицированном штамме выше, чем уровень экспрессии в немодифицированном штамме, например, штамме дикого типа или родительском штамме.
Способы, которые могут применяться для повышения экспрессии гена argA, включают, но не ограничиваются данными примерами, увеличение числа копий гена argA в бактериальном геноме (на хромосоме и/или автономно реплицирующейся плазмиде) и/или введение гена argA в вектор таким образом, что он становится способным увеличивать число копий и/или уровень экспрессии гена argA в бактерии семейства Enterobacteriaceae в соответствии с методами генной инженерии, известными для специалистов в данной области.
В качестве примеров таких векторов можно привести, но не ограничиться этим, векторы с широким кругом хозяев (broad-host-range vectors), например, pCM110, pRK310, pVK101, pBBR122, pBHR1 и подобные им. Множественные копии argA гена могут быть введены в хромосомную ДНК бактерии посредством, например, гомологичной рекомбинации, Mu-зависимой интеграции или подобными методами. Гомологичная рекомбинация может быть проведена с использованием многокопийной последовательности в хромосомной ДНК. Многокопийные последовательности в хромосомной ДНК включают, но не ограничиваются данными примерами, повторяющие участки ДНК или обращенные повторы на концах перемещающегося генетического элемента. Также, возможно введение argA гена в транспозон с целью введения в хромосомную ДНК нескольких копий гена argA. При использовании Mu-зависимой интеграции более чем 3 копии могут быть введены в хромосомную ДНК за один акт (Akhverdyan V.Z. et al., Biotechnol. (Russian), 2007, 3:3-20).
Повышение уровня экспрессии argA гена также может быть достигнуто путем модификации регуляторного участка, смежного с argA геном, или введением нативных и/или модифицированных чужеродных регуляторных участков. В качестве примеров регуляторных участков или последовательностей можно привести промоторы, энхансеры, аттенюаторы, сигналы терминации и анти-терминации транскрипции, рибосома-связывающие сайты (RBS) и другие элементы контроля экспрессии (например, участки, с которыми связываются репрессоры или индукторы, и/или участки связывания транскрипционных и трансляционных регуляторных белков, например, в составе мРНК). Такие регуляторные последовательности описаны, например, в Sambrook J., Fritsch E.F. и Maniatis Т., «Molecular Cloning: A Laboratory Manual», 2nd ed., Cold Spring Harbor Laboratory Press (1989). Модификации участков, контролирующих экспрессию гена(ов), могут комбинироваться с увеличением числа копий модифицированного гена(ов) в бактериальном геноме с использованием известных методов (см., например, Akhverdyan V.Z. et al., Appl. Microbiol. Biotechnol., 2011, 91:857-871; Tyo K.E.J. et al., Nature Biotechnol., 2009, 27:760-765).
Примером промоторов, усиливающих экспрессию argA гена, могут быть сильные промоторы. Например, lac промотор, trp промотор, trc промотор, tac промотор, PR или PL промоторы фага λ известны как сильные промоторы. Сильные промоторы, обеспечивающие высокий уровень экспрессии гена в бактерии, принадлежащей к семейству Enterobacteriaceae, могут использоваться. Также, влияние промотора может быть усилено, например, введением мутации в участок промотора argA гена с целью получения промотора с более сильной функцией, таким образом, приводя к увеличению уровня транскрипции argA гена, расположенного под промотором. Кроме того, известно, что замена нескольких нуклеотидов в 3D последовательности, и/или в области между SD последовательностью и стартовым кодоном, и/или в последовательности непосредственно перед и/или после стартового кодона в рибосома-связывающем сайте существенно влияет на трансляционную способность мРНК. Например, обнаружен 20-кратный разброс в уровне экспрессии гена в зависимости от природы трех нуклеотидов, предшествующих стартовому кодону (Gold L. et al., Annu. Rev. Microbiol., 1981, 35:365-403; Hui A. etal., EMBO J., 1984, 3:623-629).
Уровень экспрессии гетерологичного гена argJ может быть увеличен с использованием подходов, описанных выше для argA гена.
Количество копий, присутствие или отсутствие гена и/или генов оперона может быть измерено, например, рестрикцией хромосомной ДНК с последующим блоттингом по-Саузерну (Southern blotting), используя зонд, подобранный на основе нуклеотидной последовательности гена, флуоресцентную гибридизацию in situ (fluorescence in situ hybridization, FISH) и подобные им методы. Уровень экспрессии гена может быть определен измерением количества транскрибируемой мРНК с применением хорошо известных методов, включающих, например, Нозерн-блоттинг, количественную ОТ-ПЦР (RT-PCR) и т.п. Количество белков, кодируемых геном, можно определить известными методами, включая ДДС-ПААГ (SDS-PAGE) с последующим иммуноблоттингом (Вестерн-блоттинг) (Western blotting analysis), или масс-спектрометрический анализ образцов белка и т.п.
Методы работы с рекомбинантной молекулой ДНК, расщепления, молекулярного клонирования и гетерологичной экспрессии генов, такие как приготовление плазмидной ДНК, расщепление, лигирование и трансформация ДНК, выбор олигонуклеотидов в качестве праймеров и т.п. могут осуществляться обычными способами, известными специалисту в данной области. Эти методы описаны, например, в Sambrook J., Fritsch E.F. и Maniatis Т., «Molecular Cloning: A Laboratory Manual, 2nd ed.», Cold Spring Harbor Laboratory Press (1989). Методы молекулярного клонирования и экспрессии гетерологичных генов описаны в Bernard R. Glick, Jack J. Pasternak и Cheryl L. Patten, «Molecular Biotechnology: principles и applications of recombinant DNA», 4th ed., Washington, D.C: ASM Press (2009); Evans Jr., T.C. и Xu M.-Q., «Heterologous gene expression in E. сой», 1st ed., Humana Press (2011).
Ген argJ кодирует монофункциональный белок орнитинацетилтрансферазу ArgJ (KEGG, Kyoto Encyclopedia of Genes и Genomes, Киотская энциклопедия генов и геномов, входящий № MJ_0186; UniProtKB/Swiss-Prot, Protein Knowledgebase, входящий № Q57645). Ген argJ (GenBank инвентарный номер NC_000909.1; нуклеотиды, комплементарные нуклеотидам в положении: с 184215 по 185423; Gene ID: 1451033) расположен между геном MJ_0187 на той же молекулярной цепи и геном MJ_0185 на противоположной молекулярной цепи хромосомы М. jannaschii DSM 2661. Нуклеотидная последовательность гена argJ и аминокислотная последовательность монофункционального белка ArgJ, кодируемого геном argJ, приведены в SEQ ID NO:29 и SEQ ID NO:30, соответственно.
Ген argJ кодирует бифункциональный белок орнитинацетилтрансферазу/N-ацетилглутаматсинтазу ArgJ (KEGG, входящий № CTN_1181; UniProtKB/Swiss-Prot, входящий № Q9Z4S1). Ген argJ (GenBank инвентарный № NC_011978.1; нуклеотиды, комплементарные нуклеотидам в положении: с 1146678 по 1147871; Gene ID: 7377498) расположен между генами argC и CTN_1180 на хромосоме Т. neapolitana DSM 4359. Нуклеотидная последовательность гена argJ и аминокислотная последовательность бифункционального белка ArgJ, кодируемого геном argJ, приведены в SEQ ID NO:1 и SEQ ID NO:2, соответственно.
Ген argE кодирует белок N-ацетилорнитиндеацетилазу ArgE (KEGG, входящий № b3957). Ген argE (GenBank инвентарный № NC_000913.2; нуклеотиды, комплементарные нуклеотидам в положении: с 4151719 по 4152870; Gene ID: 948456) расположен между геном ррс на той же молекулярной цепи и геном argC на противоположной молекулярной цепи хромосомы штамма Е.coli K-12. Нуклеотидная последовательность гена argE и аминокислотная последовательность белка ArgE, кодируемого геном argE, приведены в SEQ ID NO:3 и SEQ ID NO:4, соответственно.
Ген argA кодирует белок аминокислота-N-ацетилтрансферазу ArgA (KEGG, входящий № b2818). Ген argA (GenBank инвентарный № NC_000913.2; положения нуклеотидов: с 2947264 по 2948595; Gene ID: 947289) расположены между генами amiC и recD, оба на противоположной цепи хромосомы штамма Е.coli K-12. Нуклеотидная последовательность гена argA и аминокислотная последовательность белка ArgA, кодируемого геном argA, приведены в SEQ ID NO:5 и SEQ ID NO:6, соответственно.
Ген argR кодирует ДНК-связывающий транскрипционный двойной регулятор, связывающий L-аргинин, ArgR (KEGG, входящий № b3237). Ген argR (GenBank инвентарный № NC_000913.2; положения нуклеотидов: с 3382725 по 3383195; Gene ID: 947861) расположен между геном yhcN на той же молекулярной цепи и геном mdh на противоположной молекулярной цепи хромосомы штамма Е.coli K-12. Нуклеотидная последовательность гена argR и аминокислотная последовательность белка ArgR, кодируемого геном argR, приведены в SEQ ID NO:7 и SEQ ID NO:8, соответственно.
Ввиду того, что могут быть некоторые различия в последовательностях ДНК между родами или штаммами семейств Methanocaldococcaceae, Thermotogaceae и Enterobacteriaceae, гены argJ, argE, argA и argR не ограничены генами, представленными в последовательностях SEQ ID NOs: 1, 3, 5, 7 и 29, но могут включать гены, которые являются вариантами нуклеотидных последовательностей или гомологичны нуклеотидным последовательностям SEQ ID NOs: 1, 3, 5, 7 и 29, кодирующие варианты белков ArgJ, ArgE, ArgA и ArgR.
Термин «вариант белка» может означать белок, который содержит одно или несколько изменений в аминокислотной последовательности, приведенной в SEQ ID NOs: 2, 4, 6, 8 или 30, которыми могут быть замены, делеции, вставки и/или добавления одного или нескольких аминокислотных остатков при условии, что активность такого белка такая же, как и у белков ArgJ, ArgE, ArgA и ArgR, соответственно, или третичная структура белков ArgJ, ArgE, ArgA и ArgR не изменилась значительно по сравнению с белком дикого типа или немодифицированным белком. Количество изменений в вариантах белков зависит от вида или положения аминокислотного остатка в третичной структуре белка. Оно может быть, но не ограничено строго, от 1 до 30, в другом примере от 1 до 15, в другом примере от 1 до 10, в другом примере от 1 до 5 в SEQ ID NO:2, 4, 6, 8 и 30.
Примером замены, делеции, вставки и/или добавления одного или нескольких аминокислотных остатков может быть консервативная мутация(и) при условии, что активность и свойства варианта белка сохранены и такие же, как и у белков ArgJ, ArgE, ArgA и ArgR. Примером консервативной мутации является консервативная замена. Консервативными заменами могут быть взаимные замены между Phe, Trp и Tyr, если сайт замещения является ароматической аминокислотой; между Ala, Leu, Ile и Val, если сайт замещения является гидрофобной аминокислотой; между Glu, Asp, Gln, Asn, Ser, His и Thr, если сайт замещения является гидрофильной аминокислотой; между Gln и Asn, если сайт замещения является полярной аминокислотой; между Lys, Arg и His, если сайт замещения является основной аминокислотой; между Asp и Glu, если сайт замещения является кислой аминокислотой; и между Ser и Thr, если сайт замещения является аминокислотой, имеющей гидроксильную группу. Примеры консервативных замен включают замену Ser или Thr на Ala, замену Asn, Glu или Gln на Asp, замену Ser или Ala на Cys, замену Asn, Glu, Lys, His, Asp или Arg на Gln, замену Asn, Gln, Lys или Asp на Glu, замену Pro на Gly, замену Asn, Lys, Gln, Arg или Tyr на His, замену Leu, Met, Val или Phe на Ile, замену Ile, Met, Val или Phe на Leu, замену Asn, Glu, Gln, His или Arg на Lys, замену Ile, Leu, Val или Phe на Met, замену Trp, Tyr, Met, Ile или Leu на Phe, замену Thr или Ala на Ser, замену Ser или Ala на Thr, замену Phe или Tyr на Trp, замену His, Phe или Trp на Tyr и замену Met, Ile или Leu на Val.
Примером замены, делеции, вставки и/или добавления одного или нескольких аминокислотных остатков также может быть неконсервативная(ые) мутация(и) при условии, что такая(ие) мутация(и) компенсируются одной или несколькими мутациями в различных положениях аминокислотной последовательности, так что активность и свойства варианта белка сохранены и такие же, как и у белков ArgJ, ArgE, ArgA и ArgR.
Степень гомологии белка или ДНК, может быть определена с использованием нескольких известных подходов, например, компьютерных алгоритмов BLAST и FASTA и метода ClustalW. Алгоритм BLAST (Basic Local Alignment Search Tool, www.ncbi.nlm.nih.gov/BLAST/), позволяющий проводить иерархический поиск, заложен в программах blastp, blastn, blastx, megablast, tbiastn и tbiastx; эти программы присваювают уровень значимости найденным объектам, используя статистические методы, описанные в Samuel K. и Altschul S.F. («Methods for assessing the statistical significance of molecular sequence features by using general scoring schemes» Proc. Natl. Acad. Sci. USA, 1990, 87:2264-2268; «Applications и statistics for multiple high-scoring segments in molecular sequences». Proc. Natl. Acad. Sci. USA, 1993, 90:5873-5877). Алгоритм BLAST вычисляет три параметра: число аминокислотных остатков, идентичность и сходство. Поисковый алгоритм FASTA описан в Pearson W.R. («Rapid и sensitive sequence comparison with FASTP и FASTA», Methods Enzymol., 1990, 183:63-98). Метод ClustalW описан в Thompson J.D. et al. («CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties и weight matrix choice», Nucleic Acids Res., 1994, 22:4673-4680).
Более того, гены argJ, argE, argA и argR могут быть вариантами нуклеотидных последовательностей. Термин «вариант нуклеотидной последовательности» может означать нуклеотидную последовательность, которая кодирует «вариант белка» с использованием любых синонимичных аминокислотных кодонов в соответствии с таблицей стандарта генетического кода (см., например, Lewin В., Genes VIII, 2004, Pearson Education, Inc., Upper Saddle River, NJ 07458). Термин «вариант нуклеотидной последовательности» также может означать, но не ограничиваться данным примером, нуклеотидную последовательность, которая гибридизуется в жестких условиях с нуклеотидной последовательностью, комплементарной последовательности, представленной в SEQ ID NOs: 1, 3, 5, 7 и 29, или с зондом, который может быть синтезирован на основе указанных нуклеотидных последовательностей, при условии, что он кодирует функциональнуй белок. Под «жесткими условиями» понимаются такие условия, при которых образуется специфический гибрид, например, гибрид, имеющий идентичность не менее чем 70%, не менее чем 80%, не менее чем 90%, не менее чем 95%, не менее чем 96%, не менее чем 97%, не менее чем 98% или не менее чем 99%, и не образуется неспецифический гибрид, например, гибрид, имеющий гомологию меньшую, чем указано выше. Практическим примером жестких условий может быть однократная или многократная отмывка, или, в другом случае, двух- или трехкратная отмывка при концентрации солей 1×SSC (стандарт цитрата натрия или стандарт хлорида натрия) и 0,1% SDS (додецилсульфат натрия) или, в другом случае, 0,1×SSC и 0,1% SDS при 60°С или 65°С. Продолжительность отмывки зависит от типа используемой для блоттинга мембраны и, как правило, такова, как рекомендовано производителем. Например, рекомендуемая продолжительность отмывки для положительно заряженной нейлоновой мембраны Hybond™-N+ (GE Healthcare) при жестких условиях составляет 15 минут. Промывка может быть произведена двух- или трехкратно. В качестве зонда может быть использована часть последовательности, комплементарной последовательности, приведенной в SEQ ID NO:1, 3, 5, 7 и 29. Подобный зонд может быть получен с помощью метода ПЦР (полимеразная цепная реакция, White T.J. et al., Trends Genet, 1989, 5:185-189) с использованием олигонуклеотидных праймеров, приготовленных на основе последовательностей, приведенных в SEQ ID NO:1, 3, 5, 7 и 29 и фрагмента ДНК, содержащего нуклеотидную последовательность в качестве матрицы. Рекомендуемая длина зонда должна быть >50 п.н., она может быть подобрана в зависимости от условий гибридизации и составляет обычно от 100 до 1000 п.н. Например, при использовании в качестве зонда фрагмента ДНК длиной около 300 п.н. условия отмывки могут быть следующие: 2×SSC, 0,1% SDS при 50°С, или при 60°С, или при 65°С.
Гены, кодирующие белки ArgJ видов М. jannaschii и T. neapolitana, и белки ArgE, ArgA и ArgR вида Е.coli, известны (см. описание выше), поэтому гены, кодирующие варианты белков ArgJ, ArgE, ArgA и ArgR, могут быть получены с использованием метода ПЦР (полимеразная цепная реакция, описано в White T.J. et al., Trends Genet, 1989, 5:185-189) с использованием праймеров, синтезированных на основе нуклеотидной последовательности генов argJ, argE, argA и argR, или химически синтезированных как полноразмерные гены. Гены, кодирующие белки ArgJ, ArgE, ArgA и ArgR или их варианты из других микроорганизмов, могут быть получены аналогичным способом.
Активность орнитинацетилтрансферазы (монофункциональный фермент ArgJ) (ЕС: 2.3.1.35; синонимы: глутамат-N-ацетилтрансфераза, N-ацетил-L-глутаматсинтетаза, и т.д.) означает активность в катализе следующей реакции: N2-ацетил-L-орнитин + L-глутамат ↔ L-орнитин + N-ацетил-L-глутамат.
Активность орнитинацетилтрансферазы/N-ацетилглутаматсинтазы (бифункциональный фермент ArgJ) (EC: 2.3.1.35/2.3.1.1) означает активность, в катализе следующей реакции: N2-ацетил-L-орнитин + L-глутамат ↔ L-орнитин + N-ацетил-L-глутамат и ацетил-СоА + L-глутамат ↔ СоА + N-ацетил-L-глутамат.
Активность N-ацетилорнитиндеацетилазы (ArgE) (EC: 3.5.1.16; синонимы: 2-N-ацетил-L-орнитинамидогидролаза, N-ацетилорнитиназа) означает активность в катализе следующей реакции: N2-ацетил-L-орнитин + H2O ↔ ацетат + L-орнитин.
Активность аминокислота-N-ацетилтрансферазы (ArgA) (EC: 2.3.1.1; синонимы: N-ацетилглутаматсинтаза, N-ацетил-L-глутаматсинтетаза) означает активность в катализе следующей реакции: ацетил-СоА + L-глутамат ↔ СоА + N-ацетил-L-глутамат.
Термин «функционально связанный с геном» может означать, что одна или несколько регуляторных последовательностей связаны с нуклеотидной последовательностью молекулы нуклеиновой кислоты или заданного гена таким образом, что возможна экспрессия (например, усиленная, увеличенная, конститутивная, базальная, ослабленная, сниженная или репрессивная экспрессия) нуклеотидной последовательности, предпочтительно экспрессия продукта гена, кодируемого указанной нуклеотидной последовательностью.
Термин «фермент, имеющий, по меньшей мере, орнитинацетилтрансферазную активность» может означать, что фермент имеет активность орнитинацетилтрансферазы (монофункциональный фермент ArgJ) или активность орнитинацетилтрансферазы/N-ацетилглутаматсинтазы (бифункциональный фермент ArgJ), как описано выше.
Бактерия, описанная в настоящем изобретении, может быть получена введением вышеупомянутых ДНК в бактерию, изначально способную продуцировать L-аминокислоту. Кроме того, бактерия, описанная здесь, может быть получена посредством сообщения способности продуцировать L-аминокислоту бактерии, уже содержащей упомянутые ДНК.
Помимо упомянутых выше свойств, бактерия также может обладать другими специфическими свойствами, не выходя за рамки данного изобретения, такими как: нуждаться в различных питательных веществах, обладать чувствительностью, устойчивостью и зависимостью от антибиотиков.
2. Способ получения L-аминокислоты
Способ получения L-аминокислоты может включать стадии выращивания бактерии в питательной среде с целью получения L-аминокислоты, экскрекции и накопления в культуральной жидкости и выделения L-аминокислоты из культуральной жидкости и/или бактериальных клеток.
Выращивание бактерии согласно настоящему изобретению, накопление и очистка L-аминокислоты или ее соли от среды и т.п. может быть проведено способом, подобным традиционным способам ферментации, отличающимся тем, что указанная аминокислота продуцируются с использованием микроорганизма. Питательная среда для получения L-аминокислоты может быть типичной средой, которая содержит источник углерода, источник азота, неорганические ионы и другие необходимые органические компоненты. В качестве источника углерода могут использоваться сахара, такие как глюкоза, лактоза, галактоза, фруктоза, арабиноза, мальтоза, ксилоза, трегалоза, рибоза и гидролизаты крахмала; спирты, такие как глицерин, маннитол и сорбитол; органические кислоты, такие как глюконовая кислота, фумаровая кислота, лимонная кислота, яблочная кислота и янтарная кислота и т.п. В качестве источника азота могут использоваться неорганические аммониевые соли, такие как сульфат аммония, хлорид аммония, фосфат аммония; органические питательные вещества, такие как гидролизаты соевых бобов; аммиачный газ; водный раствор аммиака и т.п. Витамины, такие как витамин В1, необходимые вещества, например, органические питательные вещества, такие как нуклеиновые кислоты, аденин и РНК или экстракт дрожжей и подобные им могут присутствовать в соответствующих количествах или в виде следов. Помимо перечисленных, небольшие количества фосфата кальция, сульфата магния, ионов железа, ионов марганца и подобные им могут быть добавлены при необходимости.
Выращивание осуществляют предпочтительно в аэробных условиях от 16 до 96 часов или от 48 до 72 часов, температура выращивания, в интервале которой выращивание может контролироваться, - от 30 до 45°С, или в интервале от 30 до 37°С; кислотность среды (рН) поддерживают в интервале 5-8, или в интервале 6,5-7,2. Кислотность среды поддерживают с использованием неорганических или органических кислых или щелочных веществ, таких как газообразный аммиак. Обычно, выращивание в течение от 1 до 5 дней приводит к накоплению целевой L-аминокислоты в культуральной жидкости.
После выращивания твердые частицы, такие как клетки и клеточный дебрис, могут быть удалены из жидкой среды центрифугированием или мембранной фильтрацией и затем целевая L-аминокислота может быть выделена из ферментативной смеси сочетанием известных способов, таких как концентрация, ионообменная хроматография, кристаллизация и т.п.
Примеры
Настоящее изобретение более подробно будет описано ниже со ссылкой на следующие, не ограничивающие настоящее изобретение Примеры.
Пример 1.
Получение мутантного штамма Е.coli MG1655, имеющего удаленный ген argE и содержащего плазмиду pJ-T, несущую ген argJ из Т. neapolitana
Сначала, мутантный штамм Е.coli MG1655, имеющий удаленный ген argR, получали с использованием известного метода (Datsenko K.A. и Wanner B.L., Proc. Natl. Acad. Sci. USA, 2000, 97(12):6640-6645) (Пример 1.1). Затем, ген argE удаляли с использованием того же метода (Пример 1.2). Ген устойчивости к хлорамфениколу (cat, CmR-маркер) использовали, чтобы отметить мутацию ΔargR, и ген устойчивости к канамицину (kan, KmR-маркер) использовали, чтобы отметить мутацию ΔargE. Клетки излечивали от CmR-маркера, как описано (Datsenko K.A. и Wanner B.L., Proc. Natl. Acad. Sci. USA, 2000, 97(12):6640-6645). Плазмиду pJ-T, содержащую ген argJ из T. neapolitana (патент США №6897048), вводили в полученные мутантные штаммы Е.coli MG1655ΔargR и ΔargRΔargE::Km методом электропорации. Электропорацию проводили с помощью электропоратора «Bio-Rad» (США) (№165-2098, версия 2-89) в соответствии с инструкциями производителя. Таким образом были получены мутантные штаммы Е.coli MG1655ΔargR/pJ-T и ΔargRΔargE::Km/pJ-T.
Пример 1.1.
Делеция гена argR
Штамм Е.coli MG1655ΔargR получали методом λRed-зависимой интеграции, разработанным Datsenko K.A. и Wanner B.L. [Proc. Natl. Acad. Sci. USA, 2000, 97(12):6640-6645). ДНК-фрагмент, содержащий маркер устойчивости к хлорамфениколу (CmR), получали методом ПЦР с использованием праймеров P1 (SEQ ID NO:9) и Р2 (SEQ ID NO:10) и плазмиды pMW118-attL-Cm-attR в качестве матрицы (WO2005010175 А1). Праймер Р1 содержит участок, комплементарный участку, расположенному на 5'-конце гена argR, и участок, комплементарный участку attR. Праймер Р2 содержит участок, комплементарный участку, расположенному на 3'-конце гена argR, и участок, комплементарный участку attL. Условия для ПЦР были следующие: начальная денатурация - 3 мин при 95°С; профиль для начальных двух циклов: 1 мин при 95°С, 30 сек при 50°С, 40 сек при 72°С; профиль для конечных 25 циклов: 30 сек при 95°С, 30 сек при 54°С, 40 сек при 72°С; финальная элонгация - 5 мин при 72°С. Полученный методом ПЦР продукт (около 1.600 п.н.) очищали методом электрофореза в агарозном геле и использовали для электропорации в штамм Е.coli MG1655, содержащий плазмиду pKD46 с репликоном, обладающим температурной чувствительностью. Плазмида pKD46 (Datsenko K.A. и Wanner B.L., Proc. Natl. Acad. Sci. USA, 2000, 97(12):6640-6б45) содержит ДНК-фрагмент фага λ длиной 2,154 нуклеотидов (положения нуклеотидов от 31088 до 33241, GenBank инвентарный №: J02459), включающий гены λRed гомологичной рекомбинационной системы (γ, β, ехо гены) под контролем арабиноза-индуцируемого промотора ParaB. Плазмида pKD46 необходима для интеграции продукта, полученного методом ПЦР, в хромосому штамма Е.coli МО 1655 (АТСС 47076). Штамм Е.coli MG1655, содержащий рекомбинантную плазмиду pKD46, можно получить из Е.coli Genetic Stock Center, Yale University, New Haven, USA, инвентарный номер CGSC7669.
Электрокомпетентные клетки готовили следующим способом: штамм Е.coli MG1655/pKD46 выращивали при 30°С в течение ночи на среде LB (среда Luria-Bertani, также называемая лизогенная среда, как описано в Sambrook, J. и Russell, D.W. (2001) Molecular Cloning: A Laboratory Manual (3rd ed.). Cold Spring Harbor Laboratory Press), содержащей ампициллин (150 мг/л). Затем клеточную культуру растворяли 100 раз в 5 мл SOB среды (Sambrook J. и Russell D.W., Molecular Cloning: A Laboratory Manual (3rd ed.), Cold Spring Harbor Laboratory Press, 2001), содержащей ампициллин (150 мг/л) и L-арабинозу (1 мМ). Клетки выращивали с аэрацией (250 об/мин) при 30°С до OD600 ≈0.6. Электрокомпетентные клетки готовили, концентрируя в 100 раз и трехкратно промывая деионизированной холодной водой. Электропорацию проводили, используя 70 µл клеток и ≈100 нг продукта ПЦР. После электропорации клетки инкубировали в 1 мл среды SOC (Sambrook J. и Russell D.W., Molecular Cloning: A Laboratory Manual (3rd ed.), Cold Spring Harbor Laboratory Press, 2001) при 37°С в течение 2,5 ч, помещали в L-агар, содержащий хлорамфеникол (25 мг/л), и выращивали при 37°С для отбора CmR-рекомбинантов. Чтобы удалить плазмиду pKD46, клетки высевали два раза на L-агар, содержащий хлорамфеникол (25 мг/л), при 42°С и полученные колонии тестировали на чувствительность к ампициллину. Маркер CmR удаляли с использованием хелперной плазмиды pMW-Int/Xis (WO2005010175 А1), которую электропорировали в выбранные бесплазмидные интегранты с использованием методики, как описано выше для электропорации ПЦР-фрагмента. После электропорации клетки помещали в L-агар, содержащий 0,5% глюкозы и ампициллина (150 мг/л), и инкубировали при 37°С в течение ночи, чтобы индуцировать синтез белков Int/Xis. Рост клонов проводили на L-агаре без хлорамфеникола, чтобы выбрать CmS (чувствительные к хлорамфениколу) варианты. Таким образом получали штамм Е.coli MG1655ΔargR.
Пример 1.2.
Деления гена argE
Штамм Е.coli MG1655ΔargR, имеющий удаленный ген argE, получали методом λRed-зависимой интеграции, изначально разработанным Datsenko K.A. и Wanner B.L. (Proc. Natl. Acad. Sci. USA, 2000, 97(12):6640-6645). ДНК-фрагмент, содержащий маркер KmR, получали методом ПЦР с использованием праймеров Р3 (SEQ ID NO:11) и Р4 (SEQ ID NO:12) и плазмиды pMW118-attL-Km-attR в качестве матрицы (Katashkina Zh.I. et al., Mot Biol. (Mosk.), 2005, 39(5):823-831). Штамм Е.coli MG1655ΔargR/pKD46 использовали для электропорации полученного ДНК-фрагмента (Пример 1.1). Клетки с устойчивостью к антибиотикам отбирали с помощью канамицина (25 мг/л) и полученный KmR-рекомбинант излечивали от плазмиды pKD46, как описано в Примере 1.1. Таким образом получали Е.coli MG1655ΔargRΔargE::Km.
Пример 2.
Получение L-аргинина с помощью модифицированного штамма Е.coli MG1655, имеющего удаленный ген argE и содержащего плазмиду pJ-T, несущую ген argJ из Т. neapolitana
Восемь независимых колоний каждого полученного штамма (Е.coli MG1655ΔargR/pJ-T и ΔargRΔargE::Km/pJ-T) были отобраны для оценки продукции L-аргинина. Полученные штаммы выращивали в чашках с L-агаром при 37°С в течение ночи, затем петлю с каждой из полученных культур вводили в 2 мл ферментационной среды в пробирки 20×200 мм и культивировали при 32°С в течение 72 часов на роторной качалке (220-230 об/мин).
Содержание ферментационной среды (г/л):
Глюкозу и сульфат магния стерилизовали отдельно. СаСО3 стилизовали сухим жаром при 180°С в течение 2 часов. Значение рН поддерживали на уровне 7,0. Антибиотики вводили в среду после стерилизации.
Количество накопленного в среде L-аргинина определяли методом тонкослойной хроматографии (ТСХ) или бумажной хроматографии с использованием подвижной фазы: бутанол: уксусная кислота: вода=4:1:1 (об./об.). Раствор нингидрина (2%) в ацетоне использовали в качестве окрашивающего реагента. Пятно, содержащее L-аргинин, вырезали, L-аргинин элюировали 0,5% водным раствором CdCl2 и количество L-аргинина оценивали спектрофотометрически при 540 нм. Результаты 8 независимых пробирочных ферментации (как среднее значение) представлены в Таблице 1. Как можно видеть из Таблицы 1, модифицированный штамм Е.coli MG1655ΔargR/pJ-T вызывает большее накопление L-аргинина по сравнению с родительским штаммом Е.coli MG1655ΔargRΔargE::Km/pJ-T.
Пример 3.
Получение мутантного штамма Е.coli MG1655, имеющего удаленный ген argE и ген argJ из Т. neapolitana, введенный в хромосому
Ген argJ из Т. neapolitana вставляли в хромосому штамма Е.coli MG1655ΔargR, чтобы избавиться от эффекта неустойчивости плазмиды pJ-T, несущей ген argJ. Чтобы усилить экспрессию гена argJ, получали эффективный искусственный промотор Pnlp8φ10, который был помещен перед геном argJ (Пример 3.1).
С целью исключить естественную продукцию L-аргинина, ген argA удаляли из хромосомы (Пример 3.1).
Штамм Е.coli MG1655ΔargAΔargRΔartP-J::Pnlp8φ10argJ использовали в дельнейших генетических операциях. Мутацию ΔargE помечали геном устойчивости к канамицину (kan) (Пример 1.2) и вводили в хромосому штамма Е.coli MG1655ΔargAΔargRΔartP-J::Pnlp8φ10argJ, используя хорошо известный метод P1-трансдукции (Miller J.H., Experiments in molecular genetics, Cold Spring Harbor Laboratory Press, 1972). Колонии Km-устойчивых трансдуктантов отбирали на L-агаре, содержащем канамицин (25 мг/л) (Пример 1). Таким образом получали штамм Е.coli MG1655ΔargAΔargRΔartP-J::Pnlp8φ10argJΔargE::Km.
Пример 3.1.
Получение штамма Е.coli MG1655ΔargAΔargRΔartP-J::Pnlp8φ10argJ
Штамм Е.coli MG1655, содержащий удаленный ген argA, помеченный cat-геном, получали методом λRed-зависимой интеграции, изначально разработанным Datsenko K.А. и Wanner B.L. (Proc. Natl. Acad. Sci. USA, 2000, 97(12):6640-6645) (Пример 1.1). ДНК-фрагмент, содержащий CmR-маркер, получали методом ПЦР с использованием праймеров Р5 (SEQ ID NO:13) и Р6 (SEQ ID NO:14) и плазмиды pMW118-attL-Cm-attR в качестве матрицы (WO2005010175 А1). Штамм Е.coli MG1655Δarg/pKD46 использовали для электропорации полученным ДНК-фрагментом. Маркер устойчивости к хлорамфениколу (CmR) удаляли с применением плазмиды pMW-Int/Xis (WO2005010175 А1), как описано в Примере 1.1. Таким образом был получен штамм Е.coli MG1655ΔargAΔargR.
ДНК-фрагмент, содержащий промотор гена nlpD из Е.coli, получали методом ПЦР с использованием праймеров Р7 (SEQ ID NO:15) и Р8 (SEQ ID NO:16) и хромосомной ДНК Е.coli MG1655 в качестве матрица. Условия для ПЦР были следующие: начальная денатурация - 3 мин при 95°С; профиль для начальных 2 циклов: 1 мин при 95°С, 30 сек при 50°С, 40 сек при 72°С; профиль для конечных 25 циклов: 20 сек при 94°С, 20 сек при 55°С, 15 сек при 72°С; финальная элонгация - 5 мин при 72°С. Полученный ДНК-фрагмент (около 200 п.н.) очищали методом электрофореза в агарозном геле и обрабатывали эндонуклеазами PaeI и SalI (Fermentas) (Sambrook J., Fritsch E.F. и Maniatis Т., «Molecular Cloning: A Laboratory Manual», 2nd ed., Cold Spring Harbor Laboratory Press (1989); инструкции производителя). Полученный ДНК-фрагмент лигировали с плазмидой pMIV-5JS (Российская патентная заявка №2006132818 и ЕР 1942183), которую предварительно обрабатывали эндонуклеазами Pael и SalI, придерживаясь инструкций производителя. Лигирующую смесь инкубировали при 4°С в течение ночи и затем использовали для трансформации штамма Е.coli MG1655 (АТСС 47076) электропорацией, как описано в Примере 1. Полученные трансформанты помещали на L-агар, содержащий ампициллин (50 мг/л), и чашки инкубировали при 37°С в течение ночи до тех пор, пока индивидуальные колонии не станут заметны. Из полученных трансформантов выделяли плазмиды и анализировали методом рестрикции с использованием нуклеаз Pael и SalI. Таким образом получили плазмиду pMIV-PnlpD, содержащую нативный промотор PnlpD гена nlpD из Е.coli.
Проводили рандомизацию -10 участка промотора PnlpD и отбор промотора Pnlp8. 3'-Конец промотора PnlpD получали методом ПЦР с использованием праймеров Р7 (SEQ ID NO:15) и Р9 (SEQ ID NO:17) и плазмиды pMIV-PnlpD в качестве матрицы. Праймер Р9 содержит случайные нуклеотиды, которые обозначены в последовательности SEQ ID NO:17 как «n», где n это A, G, С или Т. Условия для ПЦР были следующие: начальная денатурация - 3 мин при 95°С; профиль для начальных 2 циклов: 1 мин при 95°С, 30 сек при 50°С, 40 сек при 72°С; профиль для финальных 25 циклов: 20 сек при 94°С, 20 сек при 60°С, 15 сек при 72°С; финальная элонгация - 5 мин при 72°С. 5'-Конец промотора PnlpD получали методом ПЦР с использованием праймеров Р8 (SEQ ID NO:16) и P10 (SEQ ID NO:18) и плазмиды pMIV-PnlpD в качестве матрицы. Праймер Р10 содержит случайные нуклеотиды, которые обозначены в последовательности SEQ ID NO:18 как «n», где n это А, G, С, или Т. Условия для ПЦР были следующие: начальная денатурация - 3 мин при 95°С; профиль для начальных 2 циклов: 1 мин при 95°С, 30 сек при 50°С, 40 сек при 72°С; профиль для финальных 25 циклов: 20 сек при 94°С, 20 сек при 60°С, 15 сек при 72°С; финальная элонгация - 5 мин при 72°С.
Оба полученных ДНК-фрагмента очищали методом электрофореза в агарозном геле, обрабатывали эндонуклеазой BglII (Fermentas) и лигировали в эквимолярном соотношении (Sambrook J., Fritsch E.F. и Maniatis Т., «Molecular Cloning: A Laboratory Manual», 2nd ed., Cold Spring Harbor Laboratory Press, 1989). Лигирующую смесь инкубировали при 4°С в течение ночи. Полученный ДНК-фрагмент получали методом ПЦР с использованием праймеров Р7 (SEQ ID NO:15) и Р8 (SEQ ID NO:16) и полученного ДНК-фрагмента в качестве матрицы. Условия для ПЦР были следующие: начальная денатурация - 3 мин при 95°С; профиль для начальных 2 циклов: 1 мин при 95°С, 30 сек при 50°С, 40 сек при 72°С; профиль для финальных 12 циклов: 20 сек при 94°С, 20 сек при 60°С, 15 сек при 72°С; финальная элонгация - 5 мин при 72°С. Полученный ДНК-фрагмент (около 200 п.н.) очищали методом электрофореза в агарозном геле.
Очищенный ДНК-фрагмент обрабатывали фрагментом Кленова (Fermentas) и лигировали в эквимолярном соотношении с плазмидой pMW118-λattL-KmR-λattR (Российская патентная заявка №2006134574), которую предварительно обрабатывали эндонуклеазой XbaI (Fermentas) и фрагментом Кленова. Лигирующую смесь инкубировали при 4°С в течение ночи и затем использовали для трансформации штамма Е.coli MG1655 электропорацией, как описано в Примере 1. Полученные трансформанты помещали на L-агар, содержащий канамицин (20 мг/л), и чашки инкубировали при 37°С в течение ночи до тех пор, пока не будут заметны индивидуальные колонии. Из полученных трансформантов выделяли плазмиды и анализировали методом рестрикции с использованием нуклеаз PstI и HindIII (Fermentas) (Sambrook J., Fritsch E.F. и Maniatis Т., «Molecular Cloning: A Laboratory Manual», 2nd ed., Cold Spring Harbor Laboratory Press (1989); руководство производителя). Таким образом была получена плазмида pMW-Km-Pnlp8, содержащая промотор Pnlp8.
ДНК-фрагмент, содержащий ген argJ, получали методом ПЦР с использованием праймеров Р11 (SEQ ID NO:19) и Р12 (SEQ ID NO:20) и плазмиды pJ-T (патент США 6897048) в качестве матрицы. Условия для ПЦР были следующие: начальная денатурация - 1 мин при 95°С; профиль для 25 циклов: 1 мин при 95°С, 30 сек при 55°С, 1 мин при 72°С; финальная элонгация - 2 мин при 72°С. Полученный ДНК-фрагмент (около 1.200 п.н.) очищали методом электрофореза в агарозном геле.
ДНК-фрагмент, содержащий кассету λattL-Km-λattR-Pnlp8φ10, получали методом ПЦР с использованием праймеров Р13 (SEQ ID NO:21) и Р14 (SEQ ID NO:22) и плазмиды pMW-Km-Pnlp8 в качестве матрицы. Условия для ПЦР были следующие: начальная денатурация - 1 мин при 95°С; профиль для 25 циклов: 1 мин при 95°С, 30 сек при 55°С, 1 мин 40 сек при 72°С; финальная элонгация 5 мин при 72°С. Полученный ДНК-фрагмент (около 1.700 п.н.) (Фигура 1) очищали методом электрофореза в агарозном геле. Праймеры Р11 (SEQ ID NO:19) и Р14 (SEQ ID NO:22) содержали перекрывающиеся участки.
Полученный ДНК-фрагмент (Фигура 1) с перекрывающимися участками получали методом ПЦР с использованием праймеров Р12 (SEQ ID NO:20) и Р13 (SEQ ID NO:21) и полученного фрагмента ДНК в качестве матрицы. Условия для ПЦР были следующие: начальная денатурация - 2 мин при 95°С; профиль для 30 циклов: 30 сек при 94°С, 30 сек при 55°С, 2 мин при 72°С; финальная элонгация - 5 мин при 72°С. Полученный ДНК-фрагмент (около 2.900 п.н.) очищали методом электрофореза в агарозном геле.
Полученный ДНК-фрагмент содержит KmR-маркер и ген argJ под контролем промотора Pnlp8φ10. Этот фрагмент интегрировали вместо кластера artPIQMJ генов в хромосоме штамма Е.coli MG1655ΔargAΔargR методом λRed-зависимой интеграции, как описано выше. Таким образом получали штамм Е.coli MG1655ΔargAΔargRΔartP-J::Pnlp8φ10argJ.
Пример 4.
Получение L-аргинина с помощью модифицированного штамма Е.coli MG1655, имеющего удаленный ген argE и ген argJ из Т.neapolitana, введенный в хромосому
Шесть независимых колоний каждого из полученных штаммов (Е.coli MG1655ΔargAΔargRΔartP-J::Pnlp8φ10argJ и MG1655ΔargAΔargRΔartP-J::Pnlp8φ10argJΔargE::Km) были отобраны для оценки продукции L-аргинина. Продукцию L-аргинина определяли, как описано в Примере 2. Результаты 6 независимых пробирочных ферментации (как средний результат) приведены в Таблице 2. Как можно видеть из Таблицы 2, модифицированный штамм Е.coli MG1655ΔargAΔargRΔartP-J::Pnlp8φ10argJΔargE::Km вызывает большее накопление L-аргинина по сравнению с родительским штаммом Е.coli MG1655ΔargAΔargRΔartP-J::Pnlp8φ10argJ.
Пример 5.
Получение мутантного штамма Е.coli MG1655, имеющего ослабленный ген argE и ген argJ из Т. neapolitana, введенный в хромосому
Штамм Е.coli MG1655ΔargAΔargRΔartP-J::Pnlp8φ10argJ использовали в качестве родительского штамма для получения штамма, имеющего ген argE с ослабленной экспрессией. Во-первых, вводили CmR-маркер в межгенный участок между генами ррс и argE штамма MG1655 методом λRed-зависимой интеграции (Пример 1.1). ДНК-фрагмент, содержащий CmR-маркер, кодируемый геном cat, получали методом ПЦР с использованием праймеров Р15 (SEQ ID NO:23) и Р16 (SEQ ID NO:24) и плазмиды pMW118-attL-Cm-attR в качестве матрицы (WO2005010175 А1). Праймер Р15 содержит участок, комплементарный участку, расположенному ниже гена argE, и участок, комплементарный attR. Праймер Р16 содержит участок, комплементарный участку, расположенному ниже гена ррс, и участок, комплементарный attL. Условия для ПЦР были следующие: начальная денатурация - 3 мин при 95°С; профиль для начальных 2 циклов: 1 мин при 95°С, 30 сек при 50°С, 40 сек при 72°С; профиль для начальных 25 циклов: 30 сек при 95°С, 30 сек при 54°С, 40 сек при 72°С; финальная элонгация - 5 мин при 72°С. Продукт ПЦР (около 1.600 п.н.) очищали методом электрофореза в агарозном геле и использовали для электропорации штамма Е.coli MG1655/pKD46, как описано в Примере 1.1. После электротрансформации штамма Е.coli MG1655/pKD46 полученным фрагментом ДНК, содержащим ген cat, несколько колоний помещали на чашки с L-агаром, содержащим хлорамфеникол (20 мг/л). Электрокомпетентные клетки готовили, электропорацию проводили и CmR-рекомбинанты отбирали, как описано в Примере 1.1. Таким образом получали штамм Е.coli MG1655-Cm-argE.
Во-вторых, получали замену L76K в ArgE. Первый ДНК-фрагмент, содержащий CmR-маркер и дистальную часть argE, кодирующую мутацию L76K, получали методом ПЦР с использованием праймеров Р17 (SEQ ID NO:25) и Р18 (SEQ ID NO:26), содержащие перекрывающиеся участки, и хромосомную ДНК штамма Е.coli MG1655-Cm-argE в качестве матрицы. Условия для ПЦР были следующие: начальная денатурация - 1 мин при 95°С; профиль для 25 циклов: 1 мин при 95°С, 30 сек при 55°С, 2 мин при 72°С; финальная элонгация - 5 мин при 72°С. Полученный первый ДНК-фрагмент (около 2.800 п.н.) (Фигура 2) очищали методом электрофореза в агарозном геле.
Второй ДНК-фрагмент, содержащий проксимальную часть argE, кодирующую мутацию L76K, получали методом ПЦР с использованием праймеров Р19 (SEQ ID NO:27) и Р20 (SEQ ID NO:28) и хромосомной ДНК штамма Е.coli MG1655-Cm-argE в качестве матрицы. Условия для ПЦР были следующие: начальная денатурация - 1 мин при 95°С; профиль для 25 циклов: 1 мин при 95°С, 30 сек при 55°С, 30 сек при 72°С; финальная элонгация - 5 мин при 72°С. Полученный второй ДНК-фрагмент (около 300 п.н.) (Фигура 2) очищали методом электрофореза в агарозном геле.
Третий ДНК-фрагмент получали методом ПЦР с использованием праймеров Р17 (SEQ ID NO:25) и Р20 (SEQ ID NO:28) и полученных первого и второго ДНК-фрагментов с перекрывающимися участками (Фигура 2) в качестве матрицы. Условия для ПЦР были следующие: начальная денатурация - 2 мин при 95°С; профиль для 30 циклов: 30 сек при 94°С, 30 сек при 55°С, 2 мин 20 сек при 72°С; финальная элонгация - 5 мин при 72°С. Полученный третий ДНК-фрагмент (около 3.100 п.н.) очищали методом электрофореза в агарозном геле. Таким образом получали третий ДНК-фрагмент, содержащий CmR-маркер и ген argE с заменами Т226А и Т227А в нуклеотидной последовательности SEQ ID NO:3, приводящими к мутации L76K. Мутантный ген argE назвали как ген argEm24.
Наконец, третий ДНК-фрагмент интегрировали вместо нативного гена argE в хромосому штамма Е.coli MG1655ΔargAΔargRΔartP-J::Pnlpφ10argJ методом λRed-зависимой интеграции, как описано выше. Таким образом получали штамм Е.coli MG1655ΔargAΔargRΔartP-J::Pnlp8φ10argJargEm24::Cm.
Пример 6.
Продукция L-аргинина с помощью модифицированного штамма Е.coli MG1655, имеющего ослабленный ген argE и ген argJ из Т. neapolitana, введенный в хромосому
Шесть независимых колоний каждого из полученных штаммов (Е.coli MG1655ΔargAΔargRΔartP-J::Pnlp8φ10argJargEm24::Cm и MG1655ΔargAΔargRΔartP-J::Pnlp8φ10argJ) были отобраны для оценки продукции L-аргинина. Продукцию L-аргинина оценивали, как описано в Примере 2. Результаты 6 независимых пробирочных ферментации (как среднее значение) представлены в Таблице 3. Как можно видеть из Таблицы 3, модифицированный штамм Е.coli MG1655ΔargAΔargRΔartP-J::Pnlp8φ10argJΔargEm24::Cm вызывает большее накопление L-аргинина по сравнению с родительским штаммом Е.coli MG1655ΔargAΔargRΔartP-J::Pnlp8φ10argJ.
Пример 7.
Ферментативная активность мутантного белка ArgEL76K
Ферментативную активность контрольного белка и мутантного белка ArgEL76K, кодируемого мутантным геном argEm24, измеряли, как описано в Takahara K. et al. FEBS J., 2005, 272:5353-5364, с использованием экстрактов неочищенных белков, полученных после разрушения клеток ультразвуком. Штаммы Е.coli MG1655, MG1655ΔargE, и MG1655argEm24 использовали в качестве источников белка, чтобы оценить уровень ослабления активности ArgE ввиду мутации L76K. Штаммы Е.coli MG1655ΔargE и MG1655argEm24 получали, как описано в Дополнительном примере 1. Как видно из Таблицы 4, белок ArgEL76K показал специфическую активность менее чем около 1% по сравнению с немодифицированным белком ArgE.
Дополнительный пример 1.
Штамм Е.coli MG1655 ΔargE::Km получали по методике, описанной в Примере 1.2 для штамма Е.coli MG1655ΔargRΔargE::Km. Маркер KmR удаляли с использованием плазмиды pMW-Int/Xis (WO2005010175 A1), как описано в Примере 1.1 для CmR-маркера. Таким образом получали штамм Е.coli MG1655ΔargE.
Штамм Е.coli MG1655argEm24::Cm получали, как описано в Примере 5 для штамма Е.coli MG1655ΔargAΔargRΔartP-J::Pnlp8φ10argJ argEm24::Cm. Маркер CmR удаляли с использованием плазмиды pMW-Int/Xis (WO2005010175 A1), как описано в Примере 1.1. Таким образом получили штамм Е.coli MG1655ΔargEm24.
















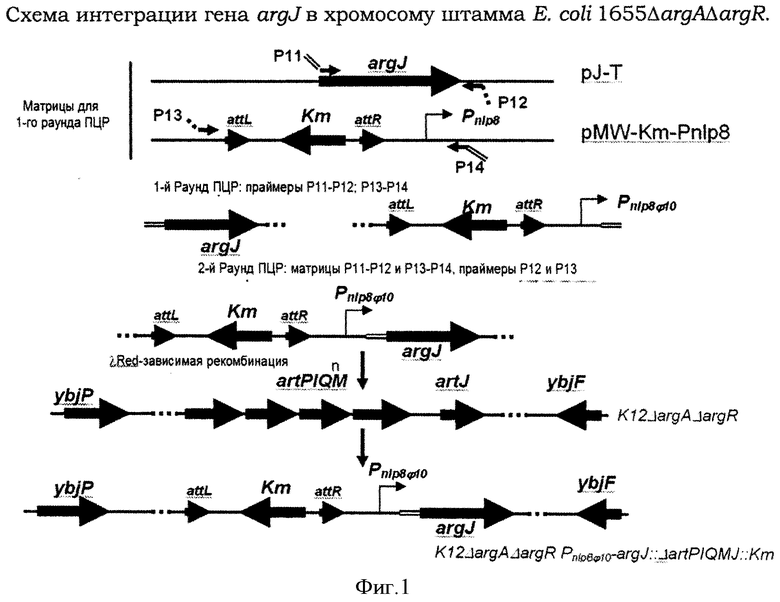
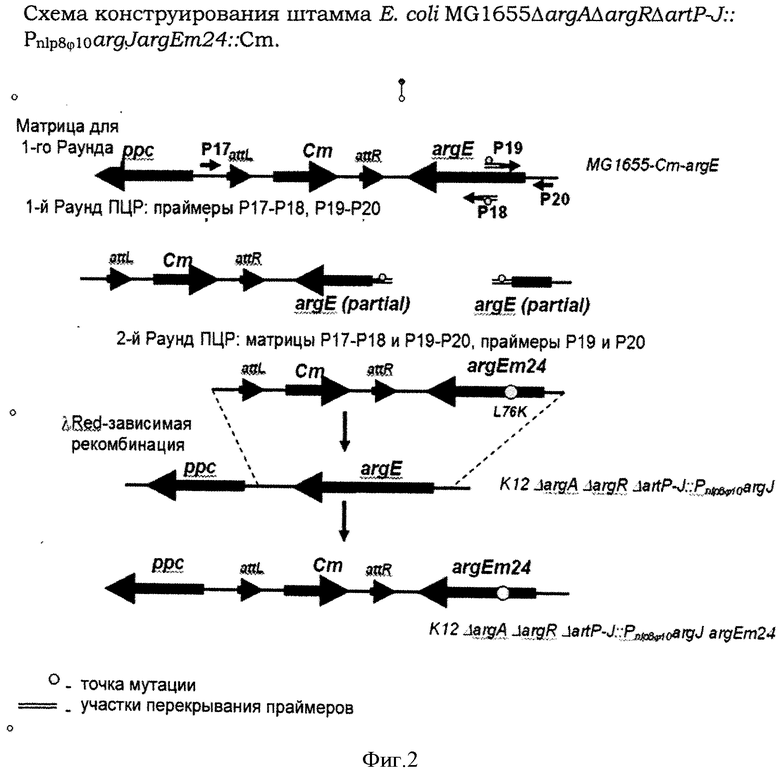
Изобретение относится к биотехнологии и представляет собой способ получения L-аминокислоты, такой как L-аргинин, ферментацией с использованием бактерии рода Escherichia, которая модифицирована таким образом, что содержит ген argJ, который кодирует фермент, имеющий, по меньшей мере, орнитинацетилтрансферазную активность, и при этом бактерия модифицирована таким образом, что содержит N-ацетилорнитиндеацетилазу с нарушенной активностью. Изобретение позволяет получать L-аргинин с высокой степенью эффективности. 2 н. и 8 з.п. ф-лы, 2 ил., 4 табл., 7 пр.
1. Бактерия-продуцент L-аргинина, принадлежащая к роду Escherichia, имеющая рекомбинантную ДНК, которая содержит ген argJ, кодирующий фермент, имеющий, по меньшей мере, орнитинацетилтрансферазную активность, отличающаяся тем, что указанная бактерия модифицирована таким образом, что она содержит N-ацетилорнитиндеацетилазу с нарушенной активностью.
2. Бактерия по п. 1, отличающаяся тем, что указанный ген argJ происходит из микроорганизма, имеющего фермент, обладающий активностью орнитинацетилтрансферазы или орнитинацетилтрансферазы/N-ацетилглутаматсинтазы.
3. Бактерия по п. 1 или 2, отличающаяся тем, что указанный ген argJ происходит из микроорганизма, принадлежащего к семейству, выбранному из группы, состоящей из семейств Thermotogaceae, Bacillaceae и Methanocaldococcaceae.
4. Бактерия по п. 3, отличающаяся тем, что указанный ген argJ происходит из вида Thermotoga neapolitana.
5. Бактерия по п. 1 или 4, отличающаяся тем, что указанный ген argJ кодирует белок, выбранный из группы, состоящей из:
(A) белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:2, который имеет активность орнитинацетилтрансферазы/N-ацетилглутаматсинтазы;
(B) варианта белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO: 2, но которая включает замену, делецию, вставку и/или добавление одного или нескольких аминокислотных остатков и имеет активность орнитинацетилтрансферазы/N-ацетилглутаматсинтазы в соответствии с аминокислотной последовательностью, представленной в SEQ ID NO:2.
6. Бактерия по п. 1, отличающаяся тем, что N-ацетилорнитиндеацетилаза является белком, выбранным из группы, состоящей из:
(C) белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:4;
(D) варианта белка, имеющего аминокислотную последовательность, представленную в SEQ ID NO:4, но которая включает замену, делецию, вставку и/или добавление одного или нескольких аминокислотных остатков и активность N-ацетилорнитиндеацетилазы в соответствии с аминокислотной последовательностью, представленной в SEQ ID NO:4.
7. Бактерия по п. 1, отличающаяся тем, что указанная активность N-ацетилорнитиндеацетилазы понижена.
8. Бактерия по п. 1, отличающаяся тем, что указанная активность N-ацетилорнитиндеацетилазы отсутствует.
9. Бактерия по п. 1, отличающаяся тем, что указанная бактерия принадлежит к виду Escherichia coli.
10. Способ получения L-аргинина или его соли, включающий:
(i) выращивание бактерии по любому из пп. 1-9 в питательной среде;
(ii) накопление L-аргинина или его соли в бактерии или культуральной жидкости, или обеих; и, если необходимо,
(iii) выделение L-аргинина или его соли из бактерии или культуральной жидкости.
| US 0006897048 B2, 24.05.2005 | |||
| БАКТЕРИЯ - ПРОДУЦЕНТ L-АМИНОКИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ (ВАРИАНТЫ) | 2005 |
|
RU2316588C1 |
| Электромагнитно-акустический искатель для контроля ферромагнитных изделий | 1984 |
|
SU1201758A1 |

