Изобретение относится к препаративной и технологической биохимии и касается получения биологически активного химического соединения и может быть использовано в клеточной биологии, практической медицине и ветеринарии.
Предшествующий уровень техники
Гликопротеины относятся к конъюгированным белкам или протеидам, содержащим помимо белковой части небелковый органический или неорганический компонент, могущий быть связанным с полипептидной цепью ковалентно, гетерополярно или координационно и вместе с аминокислотами присутствующий в гидролизате. Простетическая группа гликопротеинов может быть представлена нейтральными сахарами (галактоза, манноза, фукоза), аминосахарами (N-ацетилглюкозамин, N-ацетилгалактозамин или кислые производные моносахаридов) (Х. -Д. Якубке, X. Ешкайт "Аминокислоты, пептиды, белки (М.: Мир. 1985, с. 345).
Гликопротеины распространены в природе; к ним относятся важнейшие компоненты сыворотки крови (иммуноглобулины, трансферины и др.), групповые вещества крови, антигены многих вирусов (гриппа, кори и др.), некоторые гормоны, лектины, ферменты и т.д.
Большую группу гликопротеинов представляют собой лектины, белковая часть молекулы которых характеризуется отсутствием остатков серина и треонина, через которые осуществляется связывание углеводного компонента молекулы лектина с полипептидной цепью. Среднее содержание углеводов в лектинах составляет около 5%, их состав, в основном, ограничивается остатками галактозы, маннозы, фукозы, N-ацетилглюкозамина (Х.-Д. Якубке, X. Ешкайт "Аминокислоты, пептиды, белки (М.: Мир, 1985, с.428-429).
Описание известных методов
Из GB 2078229, 1982 известен гликопротеин, обладающий иммуноподавляющей способностью. Гликопротеин получен изоэлектрическим фокусированием сыворотки крови или асцической жидкости человека или теплокровного животного с последующим отбором фракции с изоэлектрической точкой (рI) в области рН 2.6-3.6. Этот гликопротеин применяют для подавления реакции у пациентов, которым проведена трансплантация.
Согласно патенту GB 2095260, 1982 получено биологически активное вещество - гликопротеин с молекулярной массой 3000 5000, которое способно ингибировать размножение токсоплазмы в гомологичных и гетерологичных клетках.
Из патента GB 2117385, 1982 известен гликопротеин, обладающий противораковой активностью, имеющий молекулярную массу 7000 90000, содержание сахаров 8-45%, из них 6-28% гексозы, 1-11% гексозамина и 1-6% сиаловые кислоты; стабилен в водном растворе с рН 2,0; 7,0 или 11,0 при 4oС 24 ч или больше, а в водном растворе с рН 7,0 при 60oС 3 ч и больше; селективно поражает раковые клетки, не затрагивая нормальных.
Из патента СН 634334, 1983 известен гликопротеин иммунологический, содержащий протеиновую часть 89±4%, углеводную часть 11,1±2,2%, гексозу 5,3±1,1%, N-ацетиловый остаток гексозамина 2,8±0,5%, N-ацетиловый остаток нейраминовой кислоты 2,9±0,5%, с молекулярной массой 32000±6000, рI=4,3±0,3, коэффициент седиментации S20n=3,2±0,3.
Из патента RU 2136695, 1999 известен сывороточный гликопротеин с молекулярной массой 12,5 кДа, обладающий биологической активностью в малых дозах, содержащий около 50% остатков углеводов, со значением изоэлектрической точки в области рН 4,6-4,7.
Из патента ЕР 000134, 1978 известен новый гликопротеин, содержащий а-аминокислоту 75±6%, углеводную часть 24,6±5,2%, гексозан 2,9±2%, N-ацетиловый остаток гексозамина 7,1±1,5%, фукоза 0,2±0,2%, N-ацетиловый остаток нейраминовой кислоты 8,4±1,5%, с молекулярной массой 65000±10000, рI=3,4±0,4.
Из FVO и Ceneva P. falciparum выделен и изучен гомогенный гликопротеин с молекулярной массой 56 кДа, значение изоэлектрической точки которого находится в области рН 5,5, содержащий в углеводной части N-ацетилгалактозамин и маннозу (патент US 4835259, кл. С 07 К 15/15, 1989).
Наиболее близким к данному изобретению является гликопротеин с молекулярной массой 5-300 кДа, значением изоэлектрической точки в области рН 2,5-5,0, соотношением белковой и углеводной частей (по массе) 50:50 - 80:20, содержащий в углеводной части остатки фукозы, рибозы, арабинозы, ксилозы, маннозы, галактозы, глюкозы и глюкозамина, а в белковой части остатки аспарагиновой и глутаминовой кислот, треонина, серина, пролина, глицина, аланина, цистеина, валина, метионина, цистатионина, изолейцина, лейцина, тирозина, фенилаланина, триптофана, орнитина, лизина, гистидина, аргинина (US 4683438, кл. С 07 К 15/14, 1987).
Краткое описание сущности изобретения, раскрытие изобретения.
Таким образом, это изобретение касается гликопротеинов, обладающих биологической активностью в сверхмалых дозах, пригодных для использования в медицине, фармацевтической промышленности.
С другой стороны, изобретение относится к получению гликопротеина, обладающего биологической активностью в сверхмалых дозах и применению его в качестве лекарственных препаратов.
Таким образом, технической задачей изобретения является получение гликопротеина, обладающего биологической активностью в сверхмалых дозах (10-12-10-29 моль/л и ниже).
Указанный технический результат достигается новыми гликопротеинами, выделенными изоэлектрическим фокусированием из межклеточного пространства сыворотки крови, желчи, тканей различных органов позвоночных (человека и животных), растворимых в насыщенном (100%-ном) растворе сернокислого аммония, имеющих кажущуюся молекулярную массу 10-45 кДа и обладающих биологической активностью в сверхмалых дозах. Растворы гликопротеинов в концентрации 10-12 10-18 моль/л полностью сохраняют биологическую активность при многократном замораживании-размораживании, а также после нагревания при 100oС в течение 10 мин.
Молекулярный вес гликопротеинов оценивали с помощью метода электрофореза в ПААГ в присутствии додецилсульфата натрия по методу Лемли. В качестве маркеров молекулярной массы использовали набор фирмы LKB, включающий 6 белков с мол. массой от 94 кДа до 14,4 кДа. Кажущаяся мол. масса для гликопротеина из сыворотки крови с рI в области рН 4,65-5,1 (СГ) по данным ПААГ-фореза соответствует 35-37 кДа; по данным гель-хроматографии - 25-27 кДа. Для нейтрального гликопротеина из печени (рI в области рН 6,8-7,2) (НГП) эти значения соответственно были 15 кДа и 22 кДа. Для кислого гликопротеина из печени (КГП) кажущаяся мол. масса была определена только методом гель-фильтрации и соответствовала 17 кДа (методом ПААГ-электрофореза не удалось определить значение мол. массы из-за невозможности окрасить фракцию кислого гликопротеина).
Электрофорез в полиакриламидном геле проводили на приборе для вертикального электрофореза АВГЭ-1 (Himifil, СССР). Применялись денатурирующие условия с добавлением детергента (ДДС-Na) по методу Лэмли. Толщина геля 0.75 мм, размер 115Х115 мм, максимальное количество полос - 13.
Разделяющий гель (12.5%) готовили из следующих компонентов: дистиллированная вода (6.7 мл), 1.5 М Трис-НСl рН 8.8 (5.0 мл), 10% раствор ДДС-Na (0.2 мл), 30% раствор Акриламид/N'N'-бис-метилен-акриламид (8.0 мл), TEMED (10 мкл), 10% раствор персульфата аммония (100 мкл).
Концентрирующий гель (4%) готовили из следующих компонентов: дистиллированная вода (6.1 мл), 0.5 М Трис-НСl рН 6.8 (2.5 мл), 10% раствор ДДС-Na (0.1 мл), 30% раствор Акриламид/N',N'-бис-метилен-акриламид (1.3 мл), TEMED (10 мкл), 10% раствор персульфата аммония (50 мкл).
Электродный буфер: Трис-глицин рН 8.3, 0.1% ДДС-Na. Лиофилизованный образец растворяли (около 0.1 мг/мл) в растворе: дистиллированная вода (4.0 мл), 0.5 М Трис-НСl рН 6.8 (1.0 мл), глицерин (0.8 мл), 10% раствор ДДС-Na (1.6 мл), 2-меркаптоэтанол (0.4 мл), 0.05% раствор бромфенолового синего (0.2 мл). Затем, перед нанесением на гель, образцы инкубировали 3-5 мин при 100oС. На одну полосу наносили 10-15 мкл пробы. Электрофорез проводили при постоянном напряжении 200 В. При прохождении концентрирующего геля прикладывали напряжение 100 В.
В качестве маркеров молекулярной массы использовали набор фирмы LKB, включающий в себя: фосфорилаза b (94 кДа), бычий сывороточный альбумин (67 кДа), альбумин куриного яйца (45 кДа), карбоангидраза (30 кДа), соевый ингибитор трипсина (20 кДа), лизоцим куриный (14.4 кДа).
Прокрашивание гелей осуществляли с помощью коллоидного серебра или красителем Кумасси G-250. Для прокрашивания коллоидным серебром гели предварительно промывали в дистиллированной воде, затем в водном растворе с 5% этанола и 5% уксусной кислоты в течение не менее 3 ч. Далее, гель быстро (5 мин) промывали дистиллированной водой и помещали в 10% раствор глутарового альдегида. Непрореагировавший глутаровый альдегид удаляли в процессе трехпятикратной промывки водой (30 мин каждая промывка). Затем, в течение 30 мин гель инкубировали в растворе дитиотреитола (5 мг/л), и тщательно промывали водой. Промытый гель на 30 мин помещали в раствор AgNO3 (0.1 г/л), снова тщательно промывали водой и помещали в проявитель до появления четких полос. Проявитель представлял собой раствор 37%-го формалина в 3%-ном водном Na2CO3 (50-100 мкл формалина/100 мл раствора). Для остановки проявления добавляли 3-5 мл лимонной кислоты.
При прокрашивании Кумасси G-250 гель фиксировали смесью трихлоруксусная кислота/ метанол/ вода (10:40:50). Прокрашивание проводили 0.04% раствором Кумасси G-250 в 3.5%-ной перхлорной кислоте. Обычно голубые белковые полосы проявляются в течение 5 мин. Чтобы уменьшить фоновую окраску, гель можно поместить в раствор уксусная кислота/ метанол/ вода (10:40:50) на 1-3 ч.
Молекулярный вес гликопротеинов методом гельпроникающей ВЭЖХ был проведен на колонке TSK G3000SW (300х7.5 мм), элюцию проводили 100 мМ NaH2PO4 при рН 7. Для калибровки использовали следующие белки-свидетели: овотрансферрин (78 кДа), бычий сывороточный альбумин (67 кДа), альбумин куриного яйца (45 кДа), карбоангидраза (30 кДа), соевый ингибитор трипсина (20 кДа), миоглобин (17.2 кДа), лизоцим куриный (14.3 кДа) - препараты фирмы Serva (Германия) и Sigma (США).
У выделенных гликопротеинов по изобретению определяли также аминокислотный состав, углеводный состав, наличие гликозилирования.
Определение аминокислотного состава
Определение аминокислотного состава проводили на аминокислотном анализаторе Hitachi 835 в Институте Физико-Химической Биологии им. А.Н. Белозерского. Разделение проводили на хроматографической колонке с сульфополистирольными катионитами марки 2613, детекция - спектрофотометрическая, в нингидриновых производных (длина волны 570 и 440 нм). Перед анализом проводили гидролиз белков смесью 12 н. НСl/конц. трифторуксусная кислота (2:1) с добавлением 0.005% меркаптоэтанола в течение 1 ч при 155oС.
Определение углеводного состава
Определение проводили на углеводном анализаторе Biotronic LC 2000 (ФРГ) на колонке с сорбентом Dun-urn DAX 8-11 (США) при 70oС в 0.4 М боратном буфере рН 8.0. Размер колонки 3.7х75 мм. Детектирование осуществляли при 570 нм с раствором 2,2'-бицинхонината меди. Перед определением проводят гидролиз 1 мл препарата (0.1 мг/мл) в 10 мл 2 М трифторуксусной кислоты 3 ч при 100oС.
Установление наличия гликозилирования
Для определения наличия гликозилирования к 50 мкл раствора белка в воде (0.1 мг/мл) добавляли 5 мкл 0.1 М ацетатного буфера рН 4.6, затем добавляли раствор (5-10 мкл) периодата Na в том же буфере так, чтобы концентрация периодата в реакционной смеси составляла 1-2 мМ, и выдерживали 30 мин при комнатной температуре. Непрореагировавший периодат нейтрализовали 0.5 М раствором тиосульфата натрия в воде до исчезновения окраски. Затем добавляли раствор динитрофенилгидразина (ДНФГ) в диметилсульфоксиде (ДМСО) до концентрации ДНФГ в реакционной смеси 2.5-5 мМ и выдерживали 30 мин при комнатной температуре. Далее, образовавшиеся основания Шиффа восстанавливали с помощью раствора боргидрида натрия в ДМСО (0.1 мг/мл).
Анализ продуктов проводили методом гельпроникающей хроматографии на колонке TSK 2000 PW (5.6х300 мм) с помощью жидкостного хроматографа высокого давления, элюент 50 мМ ацетатный буфер рН 4.5, скорость 0.5 мл/мин, УФ-детекция при длине волны 365 нм. Оценивали время удерживания ДНФГ, интактного гликопротеина (по поглощению на 280 нм) и модифицированного ДНФГ гликопротеина (в случае наличия пика поглощения при 365 нм со временем удерживания, приблизительно равным времени удерживания немодифицированного гликопротеина).
В качестве модельных гликопротеинов были использованы: альбумин куриного яйца, овомукоид, S- карбокси метилированный муцин, родопсин-связывающий гликопротеин из яичного желтка.
Определение гликозилирования гликопротеинов по данному изобретению.
После испытаний на модельных гликопротеинах известной структуры данный метод был применен к выделенным белкам. Результаты свидетельствуют в пользу того, что все эти вещества являются гликопротеинами. На хроматограмме виден пик исследуемого гликопротеина, а также пик, соответствующий непрореагировавшему ДНФГ. Время выхода модифицированного гликопротеина немного отличается от нативного, что можно было ожидать.
Например, для очищенного препарата СГ анализ углеводного состава показал, что данный белок сильно гликозилирован, он содержит 40-55 вес.% углеводов. Из углеводов были детектированы N-ацетилглюкозамин и манноза в соотношении 2: 5. Знание биологических путей синтеза гликопротеинов позволяет сделать предположение о том, что СГ содержит преимущественно маннозобогатые N-связанные олигосахаридные цепи.
Другим аспектом данного изобретения является также применение гликопротеинов, описанных в настоящем изобретении, для приготовления различных фармацевтических композиций (капли, мази, лечебные кремы, гели и т.д.).
Фармацевтическая композиция по настоящему изобретению содержит в эффективном количестве гликопротеин, обладающий биологической активностью в сверхмалых дозах, являющийся одним из аспектов данного изобретения, и фармацевтически приемлемый носитель (примеры 15-17).
Фармацевтически приемлемым носителем может быть органический носитель, полимерный (углеводы, целлюлоза), неорганический носитель, выбор которого определяется прежде всего методом применения (назначения) фармкомпозиции.
Имеющиеся описанные примеры и, в частности, пример 15 описывает как пример фармкомпозицию, которая является биорегулятором репаративных процессов в эпителиальных и соединительных тканях.
Гликопротеины по данному изобретению были выделены из следующих тканей: печени, легкого, тимуса, селезенки, сердца, почек, поджелудочной, молочной и щитовидной желез, кишечника, семенников, яичников, головного мозга, костного мозга, тканей глаза, а также из сыворотки крови и желчи.
Собственно процесс выделения указанных гликопротеинов по данному изобретению включает по крайней мере следующие этапы:
а) получение экстрактов из тканей различных органов человека и животных;
б) высаливание белков тканевого экстракта;
в) собственно выделение (разделение и очистка) гликопротеинов методом изоэлектрической фокусировки;
г) сбор фракций и идентифицирование гликопротеинов, определение их биологической активности, биологического эффекта.
Идентифицированные нами гликопротеины можно разделить условно на три группы:
кислые белки, мигрирующие к аноду;
белки с рI в области рН от 4,6 до 8,5 (слабокислые, нейтральные и слабоосновные);
основные белки, мигрирующие к катоду.
Метод изоэлектрической фокусировки является одним из традиционных методов выделения и очистки (разделения) биополимеров, к числу которых относятся и гликопротеины по настоящему изобретению, используется в физической и аналитической химии.
Он основан на создании под действием внешнего электрического поля стабильного градиента рН, причем значение рН возрастает от анода к катоду. В такой системе каждый белок перемещается в том или ином направлении в соответствии со знаком своего заряда до тех пор, пока не достигнет участка, в котором значение рН совпадает с его изоэлектрической точкой (рI). На этом участке дальнейшее его перемещение под действием электрического поля прекращается, т.к. его заряд становится равным 0.
Приложенное поле, поддерживающее стабильный градиент рН, препятствует диффузионному размыванию зоны. Для создания стабильного градиента рН используют специальные вещества, получившие название амфолинов.
Изоэлектрофокусирование проводили в градиенте сахарозы, используя колонку LKB-440 (LKB, Швеция) и амфолиты (Serva, Швеция) диапазона рН 3,5 10,0, при температуре 4oС в течение 100 ч и при напряжении 500-1500 В. Супернатант в виде раствора вводили в тяжелый градиентный раствор в количестве не более 100 мг общего белка. Детекцию фракций осуществляли спектрофотометрически при 280 нм. Диализ фракций после ИЭФ проводили в мешках из целлюлозной пленки против дистиллированной воды. Собирали следующие фракции:
1. кислые гликопротеины, мигрирующие к положительно заряженному электроду;
2. гликопротеины со значением рI в области рН 4,6-8,5 (слабокислые, нейтральные и слабоосновные);
3. основные гликопротеины, мигрирующие к отрицательно заряженному электроду.
Гликопротеины трех указанных групп были выделены из тканей следующих органов: печени, легкого, тимуса, селезенки, сердца, почек, поджелудочной, молочной и щитовидной желез, кишечника, семенников, яичников, головного мозга, костного мозга, хрусталика глаза, роговицы глаза, пигментного эпителия глаза, сетчатки глаза, а также из сыворотки крови и желчи.
Все идентифицированные гликопротеины проявляли биологическую активность в сверхмалых дозах, соответствующих концентрациям 10-12-10-29 моль/л и ниже.
Для выделенных гликопротеинов по настоящему изобретению определяли биологический эффект (Эa), который расчитывали, например, по формуле ,
,
где Эa - биологический эффект, вызываемый гликопротеином,
Nоn и Nk - количество клеточных ядер, выделившихся из 1 мг ткани соответственно в опыте (органные культуры в присутствии гликопротеина) и в контроле (органные культуры без добавления гликопротеина).
Подсчет клеточных ядер осуществляли в определенном объеме камеры Горяева. Статистическую обработку данных проводили методом вариационной статистики с использованием критерия Стъюдента. Процедура приготовления органной культуры состояла в следующем. После декапитации животного (мыши-гибриды CBA/C57BI, самцы, 16-18 г) печень помещали в 199 среду при комнатной температуре. Из центральной части большой доли печени вырезали фрагменты ткани размером 1-1,5 мм2, при этом края большой доли и области с крупными кровеносными сосудами не использовали. По 5 органных культур помещали в пенициллиновые флаконы с питательной средой состава: 1 мл среды 199+0,2 мл сыворотки крупного рогатого скота или лошадиной сыворотки +0,1 мл исследуемого раствора гликопротеина в определенной концентрации; в контрольные флаконы вместо раствора гликопротеина добавляли 0,1 мл Рингера. Все флаконы инкубировали при 37oС в течение 2 ч.
Для оценки параметра, характеризующего вязкоупругие свойства мембраны гепатоцитов, каждый фрагмент ткани промокали фильтровальной бумагой, помещали в специальный стеклянный гомогенизатор с зазором 50 мкм, добавляли 0,1 мл 0,1% раствора трипанового синего, приготовленного на растворе Рингера, и диспергировали, вращая пестик 25-30 раз. Далее производили подсчет выделившихся при диспергировании одиночных клеточных ядер. Расчет биологического эффекта гликопротеина осуществляли по формуле, указанной выше.
На вязкоупругие свойства мембраны гепатоцитов оказывали влияние гликопротеины трех указанных групп, выделенных из тканей следующих органов: печени, легкого, тимуса, селезенки, сердца, почек, поджелудочной, молочной и щитовидной желез, кишечника, семенников, яичников, головного мозга, костного мозга, хрусталика глаза, роговицы глаза, пигментного эпителия глаза, сетчатки глаза, а также сыворотки крови и желчи.
Для идентифицированных нами гликопротеинов значение Эа составляло от 125 до 150% для концентраций 10-12-10-29 моль/л и ниже (фиг. 1, 2).
Подробное описание осуществления изобретения.
Гликопротеины по настоящему изобретению, проявляющие биологическую активность в сверхмалых дозах, выделяют из межклеточного пространства из сыворотки крови и желчи, из тканей различных органов: печени, легкого, тимуса, селезенки, сердца, почек, поджелудочной, молочной и щитовидной желез, кишечника, семенников, яичников, головного мозга, костного мозга, хрусталика глаза, роговицы глаза, пигментного эпителия глаза, сетчатки глаза с использованием этапов, указанных выше.
Нижеследующие примеры иллюстрируют, но ни в коей мере не ограничивают данное изобретение.
Пример 1. Гликопротеины из печени млекопитающих.
1.1. Получение экстракта печени
Эксперименты проводили на крысах линии Wistar, обоего пола, весом 150-180 г. Животных декапитировали. Через вену Порта в течение 30-40 с печень каждого животного отмывали от крови током перфузирующего раствора (0,15 моль/л NaCl; 0,04 моль/л КСl; 0,001 моль/л CaCl2) со скоростью 5-7 мл/мин, нарезали на фрагменты весом 1,5-2,0 г и помещали в раствор указанного состава при температуре 4oС на 2 ч (15-20 мл раствора на 1 печень).
Полученный экстракт собирали. Ткань заливали свежей порцией физиологического раствора и экстрагировали еще 1 час. Полученные экстракты объединяли. Для удаления клеток крови и поврежденных клеток печени тканевой экстракт центрифугировали при 5000 g в течение 20 мин, затем декантировали и использовали для дальнейшей очистки.
1.2. Высаливание белков тканевого экстракта
В тканевой экстракт при интенсивном перемешивании постепенно добавляли сухой сернокислый аммоний до образования насыщенного раствора соли (4oС, рН 8,0-8,5). Образовавшийся осадок примесных белков осаждали центрифугированием при 35000 g в течение 30 мин, супернатант собирали и длительно диализовали против дистиллированной воды, используя целлюлозную пленку отечественного производства (ГОСТ 7730-89). Во время диализа дистиллированную воду многократно заменяли на свежую. После полного удаления ионов аммония супернатант концентрировали до объема 100 мл, используя метод лиофильного высушивания, и повторяли процедуру высаливания в тех же условиях. Аналогично полученный второй супернатант также диализовали против дистиллированной воды до полного удаления ионов аммония и далее разделяли методом ИЭФ.
Изоэлектрофокусирование супернатанта тканевого экстракта осуществляют по методике, описанной выше.
Пример 2. Гликопротеины из тимуса млекопитающих.
Эксперименты проводили на крысах линии Wistar, обоего пола, весом 150-180 г. Животных декапитировали, вырезали тимусы, промывали тщательно в физиологическом растворе (0,15 моль/л NaCl; 0,04 моль/л КСl; 0,001 моль/л СаСl2) и экстрагировали в растворе указанного состава при температуре 4oС на 2 ч (3-4 мл экстрагирующего раствора на 1 тимус). Полученный экстракт собирали. Тимусы заливали свежей порцией физиологического раствора и экстрагировали еще 1 час. Полученные экстракты объединяли. Для удаления клеток крови и поврежденных клеток печени тканевой экстракт центрифугировали при 5000 g в течение 20 мин, затем декантировали и использовали для дальнейшей очистки.
Высаливание и рН-изоэлектрофокусирование тканевого экстракта проводили по указанной выше методике. После рН-изоэлектрофокусирования так же, как и при разделении экстракта печени, собирали три белковые фракции: кислые, с рI в области рН 4,6-8,5 и основные гликопротеины.
Все идентифицированные гликопротеины тимуса проявляли биологическую активность в сверхмалых дозах, соответствующих концентрациям 10-12-10-29 моль/л и ниже.
Пример 3. Гликопротеины из тканей глаза млекопитающих.
Из свежих глаз быка (30 шт.) выделяли следующие ткани: хрусталик, роговицу, пигментный эпителий, сетчатку. Ткани промывали тщательно в физиологическом растворе (0,15 моль/л NaCl; 0,04 моль/л КСl; 0,001 моль/л СаСl2), разрезали на крупные куски и экстрагировали по отдельности в растворе указанного состава при температуре +4oС в течение 2 ч. Полученные экстракты центрифугировали в течение 15 мин при 3000 g, затем декантировали и использовали для дальнейшей очистки. Высаливание и рН-изоэлектрофокусирование тканевого экстракта проводили по указанной выше методике. После рН-изоэлектрофокусирования собирали по три белковые фракции для каждой из тканей: кислые, с рI в области рН 4,6-8,5 и основные гликопротеины хрусталика глаза, роговицы глаза, пигментного эпителия глаза, сетчатки глаза.
Все идентифицированные гликопротеины из тканей глаза проявляли биологическую активность в сверхмалых дозах, соответствующих концентрациям 10-1-10-29 моль/л и ниже.
Пример 4. Выделение гликопротеина с рI в области рН 4,65-5,1 из сыворотки крови крупного рогатого скота.
К 5 л сыворотки крови крупного рогатого скота постепенно добавляли сухой сернокислый аммоний до образования насыщенного раствора соли (4oС, рН 8,0-8,5). Полученный осадок удаляли фильтрованием, а фильтрат центрифугировали при 10000 g в течение 15 мин. Затем осадок белков удаляли, супернатант собирали и длительно диализовали против дистиллированной воды до полного удаления ионов аммония, после чего лиофильно высушивали и перерастворяли в дистиллированной воде до объема 0,5 л. К полученному раствору добавляли при перемешивании сухой сернокислый аммоний до насыщения, выдерживали смесь в течение 4 суток при 4oС, затем центрифугировали при 10000 g в течение 15 мин. Супернатант диализовали против дистиллированной воды до полного удаления ионов аммония, затем лиофильно высушивали и перерастворяли в 50 мл дистиллированной воды. Дальнейшее разделение проводили методом рН-изоэлекрофокусирования по описанной выше методике. После рН-изоэлектрофокусирования собирали фракцию в области рН 4,65-5,1, диализовали против дистиллированной воды и лиофильно высушивали. Получили 0,55 мг. Полученный препарат при исследовании представлял гомогенный гликопротеин слабо окрашенный в кремовый цвет, мелко-дисперсный порошок. Гликопротеин содержит 40-55% углеводов. Молярное соотношение D-маннозы к N-ацетил-D-глюкозамину составляет от 5:2 до 3: 2, величина биологической активности (влияние на вязкоупругие свойства мембраны гепатоцитов in vitro) при концентрации гликопротеина 10-12-10-29 моль/л и ниже составляет не менее 125% по отношению к контролю.
Аналогичным образом выделяют гликопротеин из сыворотки крови крыс, крови собак, крови лошади и крови человека. Все выделенные гликопротеины из сыворотки крови имеют изоэлектрическую точку в области рН 4,65-5,1, кажущуюся молекулярную массу 35-37 кДа по данным SDS-электрофореза в полиакриламидном геле и 25-27 кДа по данным гель-хроматографии, обладают биологической активностью в сверхмалых дозах при концентрациях 10-12-10-29 моль/л и ниже.
Промышленная применимость
Нижеследующие примеры 5-17, представленные фиг. 1-29 и табл. 1-8 иллюстрируют биологическую активность, биологический эффект гликопротеинов, заявленных в данном изобретении.
Пример 5. Влияние гликопротеинов на вязкоупругие свойства мембраны гепатоцитов in vitro.
Исследование проводили с помощью метода множественного органного культивирования фрагментов ткани печени, весом 1-1,5 мг, в питательной среде, в которую вносили определенный объем раствора изучаемого гликопротеина. Для каждого гликопротеина определяли интервал концентраций, в которой проявляется его биологическое действие. Для этого производили серию последовательного разведения исходного препарата (концентрация 0,1 мг белка/мл) в 10, 100, 1000 и т.д. раз до 10-15 мг белка/мл. В контрольные флаконы вместо исследуемого препарата добавляли аналогичный объем раствора Рингера. Все исследуемые в одном эксперименте эксплантаты были получены из печени одного и того же животного. В каждом отдельном эксперименте на каждую экспериментальную точку (соответствующая концентрация раствора гликопротеина) просчитывали не менее 5 фрагментов ткани. Для определения биологической активности каждого гликопротеина проводили не менее 3-4 опытов.
Биологический эффект рассчитывали по формуле, указанной выше, и проиллюстрирован на фиг. 1-2.
Пример 6. Влияние основного гликопротеина тимуса на протекание реакции гиперчувствительности замедленного типа.
Основной гликопротеин тимуса (ОГТ) вводили четырем группам по 5 мышей C57BL/6 spf статуса весом 20 г внутрибрюшинно трехкратно в дозах 2•10-6, 2•10-12, 2•10-16 и 2•10-18 г на одно животное в течение трех дней, начиная с момента внутрибрюшинной иммунизации мышей эритроцитами барана в дозе 2•108 на одно животное. Через 5 суток после иммунизации реципиентам вводили 1•108 эритроцитов барана в объеме 50 μl подкожно в стопу задней ноги и еще через сутки определяли интенсивность отека по сравнению с контрлатеральной конечностью по методу Kitamura и число антителообразующих клеток селезенки методом Jerne. Результаты представлены в табл. 1.
Как хорошо видно из таблицы, препарат не оказывает существенного влияния на образование антителообразующих клеток в селезенке. Вместе с тем, введение препарата в дозе 2•10-12 г на одно животное приводит к высоко достоверному усилению реакции гиперчувствительности замедленного типа.
Таким образом, нами показана способность препарата ОГТ в дозе 2•10-12 г на одно животное к стимуляции реакции гиперчувствительности замедленного типа, обусловленной функцией Т-лимфоцитов хелперов типа I, от которых зависит развитие реакций специфического противоопухолевого иммунитета.
Jeme N. , Nordin A. (1963) Plaque formation in agar by single antibody production cells. Science, V.140, P.405.
Kitamura K. (1980) A foot pad weight assay to evaluate delayed type hypersensitivity in the mice. J. Immunol. Meth., V.39, P.277-283.
Пример 7. Влияние основного гликопротеина тимуса на выживание мышей с тимомой EL4.
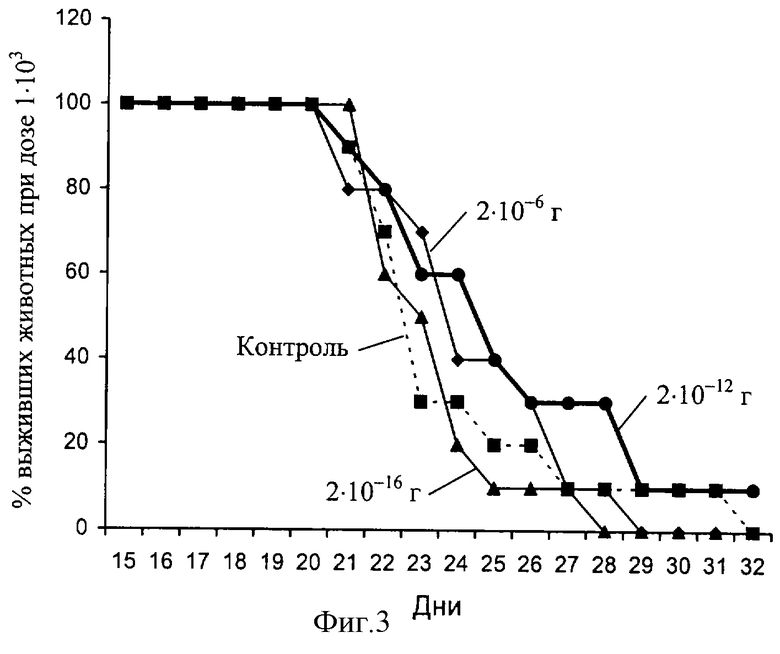
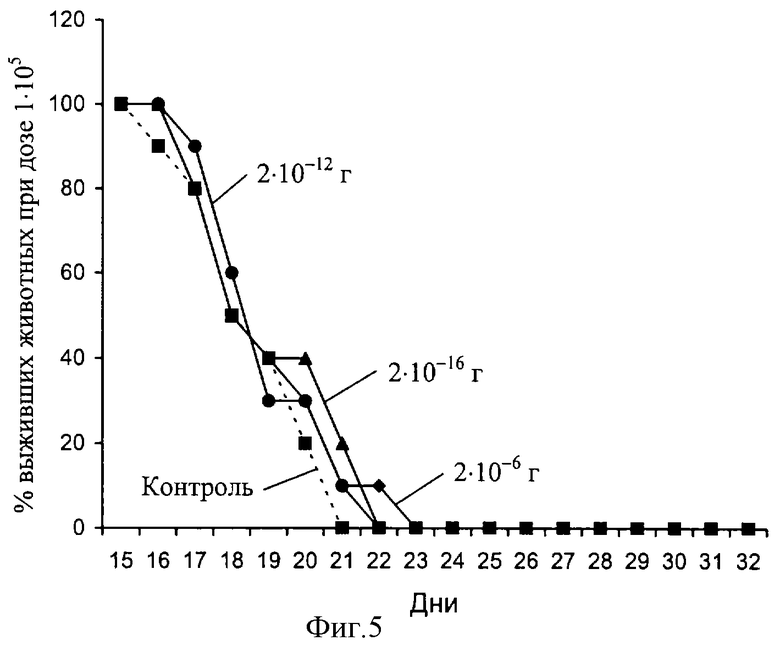
ОГТ был исследован на предмет наличия у него активности, стимулирующей противоопухолевый иммунитет. С этой целью мыши линии C57BL/6 (KbIbDb) получали однократную инъекцию клеток тимомы EL4, происходящей из мышей той же линии, в дозах 1•103, 1•104 и 1•105. Препарат вводили ежедневно в течение недели в дозах 2•10-16, 2•10-12 и 2•10-16 г на одно животное, начиная со дня иммунизации (девять опытных групп и три контрольные группы по 5-7 животных в каждой). Эффект препарата оценивали по продолжительности жизни реципиентов различных доз опухолевых клеток, сравнивая ее с соответствующими группами контрольных реципиентов. Результаты представлены на фиг. 3-5. На фиг. 3 виден протективный эффект препарата в дозе 2•10-6 г на одно животное (жирная линия) по сравнению с контролем (пунктир). В этой экспериментальной группе из 10 реципиентов остался в живых 1, тогда как в контроле все животные погибли в результате роста опухоли в асцитной форме. На фиг. 4 виден незначительный протективный эффект препарата в дозе 2•10-16 г на одно животное (жирная линия) по сравнению с контролем (пунктир). При дозе опухолевых клеток 1•105 увидеть эффект препарата оказалось невозможным (фиг. 5). Из представленных данных следует, что ОГТ в дозе 2•10-12 г на мышь, приводит к статистически достоверному увеличению продолжительности жизни экспериментальных животных - группа, получившая минимальную дозу опухолевых клеток (1•103).
Несмотря на то, что биологический эффект, проявляемый ОГТ невелик, следует учесть тот факт, что время удвоения клеток тимомы в несколько раз превышает время удвоения Т-лимфоцитов с противоопухолевой эффекторной функцией. Кроме того, полное отторжение клеток введенной опухоли наблюдается, как правило, лишь при введении опухолевых вариантов, модифицированных с использованием генетических конструкций для продукции цитокинов, стимулирующих клеточный иммунитет, либо при комбинированной терапии с цитостатиками.
Sumimoto H., Tani К., Nakazaki Y., Tanabe Т., Hibino H., Wu M.S., Izawa K., Hamada H., Asano S. (1998) Superiority of interleukin-12-transduced murine lung cancer cells to GM-CSF or B7-1 (CD 80) transfectants for therapeutic antitumor immunity in syngeneic immunocompetent mice. Cancer Gene Ther V.5, N 1, Р.29-37.
Ehrke M.J., Verstovsek. S., Pocchiari S. K., Krawczyk C. M., Ujhazy P., Zaleskis G. , Maccubbin D.L., Meer J.M., Mihich E. (1998) Thymic anti-tumor effectors in mice cured of lymphoma by cyclophosphamide plus TNF-alpha therapy: phenotypic and functional characterization up to 20 months after initial tumor inoculation. Int. J. Cancer, V.76, N 4, P.579-586.
Пример 8. Влияние кислого гликопротеина тимуса на протекание реакции гиперчувствительности замедленного типа.
Кислый гликопротеин тимуса (КГТ) вводили четырем группам по 5 мышей C57BL/6 spf статуса весом 20 г внутрибрюшинно трехкратно в дозах 2•10-6, 2•10-12, 2•10-16 и 2•10-18 г на одно животное в течение трех дней, начиная с момента внутрибрюшинной иммунизации мышей эритроцитами барана в дозе 2•108 на одно животное. Через 5 суток после иммунизации реципиентам вводили 1•108 эритроцитов барана в объеме 50•1 подкожно в стопу задней ноги и еще через сутки определяли интенсивность отека по сравнению с контрлатеральной конечностью по методу Kitamura и число антителообразующих клеток селезенки методом Jerne. Результаты представлены в табл. 2.
Как хорошо видно из таблицы, препарат не оказывает существенного влияния на образование антителообразующих клеток в селезенке. Вместе с тем, введение препарата в дозе 2-10-12 г на одно животное приводит к высоко достоверному ингибированию реакции гиперчувствительности замедленного типа.
Таким образом, нами показана способность препарата КГТ в дозе 2•10-12 г на одно животное замедлять реакцию гиперчувствительности замедленного типа.
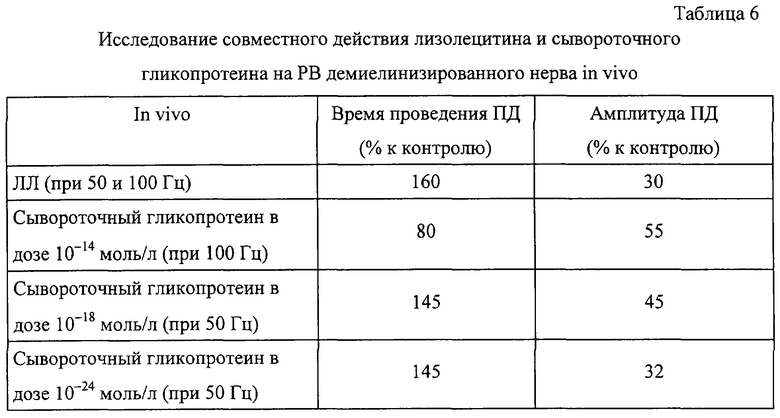
Пример 9. Влияние гликопротеинов из тимуса млекопитающих на проведение возбуждения при демиелинизации нервного волокна.
В качестве объекта исследования использовали миелиновые нервы травяной лягушки (Rana temporaria). Для осуществления фокальной демиелинизации нервов in vivo использовали лизолецитин (ЛЛ), который вводился в оболочку миелинового ischiaticus нерва лягушки при операции (внутриполосная инъекция). Известно, что данное воздействие инициирует демиелинизацию нерва, ряд стадий которой соответствуют автоиммунной демиелинизации. Первые изменения состояния миелина наблюдаются через день после операции, а через 6-12 дней миелин полностью "оголяет" интернодные участки нервного волокна. Через 3-4 недели после операции наблюдается образование нового миелина.
Исследование воздействия основного и кислого гликопротеинов из тимуса на процессы демиелинизации и восстановления при демиелинизации проводили в двух постановках экспериментов: in vitro, на изолированном нерве, а также in vivo, через одну неделю после введения ЛЛ животным. ЛЛ и исследуемый гликопротеин тимуса вводили животным одновременно. При работе in vivo проводили следующие серии экспериментов.
1. Оценивали воздействие на электрофизиологические параметры нерва ЛЛ, гликопротенов тимуса.
2. Определяли эффект совместного действия ЛЛ и соответствующего гликопротеина тимуса.
Контролем служили животные, не подвергавшиеся воздействию ЛЛ и гликопротеинов тимуса, но находящиеся в тех же условиях в течение всего времени эксперимента. При работе in vitro использовали стандартный метод экстраклеточной регистрации мембранного потенциала и потенциала действия (ПД), с помощью которого определяли:
1) динамику изменения амплитуды ПД, порога, скорости проведения ПД и максимальной частоты ритмического ответа изолированного нерва при действии ЛЛ, гликопротеинов тимуса и их совместном действии;
2) динамику изменений амплитуды ПД, порога, скорости проведения ПД и максимальной частоты ритмического ответа через одну неделю после совместного введения ЛЛ и гликопротеинов в животное.
Гликопротеины из тимуса были исследованы в концентрации 10-14 моль/л, 10-18 моль/л и 10-24 моль/л.
Изолированные нервы предварительно выдерживали не менее 30 мин в физиологическом растворе состава (мМ): NaCl - 111,2; КСl - 1,88; CaCl - 1,08; рН - 7,2; 18-20oС. Для приготовления физиологического раствора и растворов гликопротеинов, приготовленных на физиологическом растворе указанного состава, применяли бидистиллят.
Для оценки изменений уровня мембраносвязанного кальция в исследовании применялся метод флуоресцентной спектроскопии с применением локализованного в плазматической мембране Са2+ - связывающего зонда хлортетрациклина, способного образовывать комплекс с ионом кальция. Эксперименты на нервах проводились в специально сконструированной камере, позволяющей одновременно регистрировать уровень флуоресценции нерва и амплитуду ПД. Свечение регистрировали с одного и того же участка нерва или изолированного волокна в продолжение всего эксперимента. Флуоресценцию нервов регистрировали с помощью люминесцентного микроскопа Люмам И-3 (ЛОМО). Возбуждено флуоресценции хлортетрациклина вызывали с помощью галогеновой лампы накаливания КГМ 9•70 и комбинации фильтров ФС-1-6 и СЗС 21-2. Регистрацию проводили с помощью фотометрической насадки и интерференционных светофильтров с длиной волны максимального пропускания светового пучка при 490 и 550 нм. Диаметр фотометрируемого участка нерва составлял 50 мкм, при объективе •10.
При действии ЛЛ и демиелинизации нерва происходят существенные изменения некоторых электрофизиологических параметров ритмического возбуждения (РВ). В ходе настоящего исследования нами было установлено, что in vivo основной и кислый гликопротеины из тимуса в исследуемой дозе компенсируют изменения параметров РВ при демиелинизации нерва (табл. 3, 4 и 5). Отметим, что собственно инъекция растворами гликопротеинов не приводила к достоверным изменениям возбудимости миелинового нерва.
В отдельном эксперименте было исследовано восстановление возбудимости нервов целого животного после инъекции более низких концентраций гликопротеинов тимуса (10-18M и 10-24M). Результаты исследования представлены в табл. 4 и 5.
Известно, что разрушение миелина приводит к снижению скорости распространения ПД. В ходе проведенного нами исследования было установлено, что при действии ЛЛ скорость проведения изолированным нервном ПД снижается, а при совместном действии на нерв ЛЛ и исследуемых гликопротеинов тимуса характер этих изменений уменьшается (фиг. 6, 7). При максимальном разбавлении растворов гликопротеинов тимуса (до 10-24 М) наблюдалась только тенденция (но отсутствовали достоверные различия) восстановления в течение первых 7-15 мин инкубации ЛЛ и растворов гликопротенов. Таким образом, длительная инкубация нервов в растворах гликопротеинов тимуса и ЛЛ приводит к статистически достоверному восстановлению скорости проведения ПД только в случае концентрации 10-14 моль/л.
В следующей серии экспериментов были исследованы изменения амплитуды ПД, при действии ЛЛ и гликопротеинов тимуса (фиг. 8). Установлено, что за время эксперимента (действие ЛЛ) наблюдаются значительные изменения данного параметра РВ (при частоте 100 Гц). При совместном действии ЛЛ и гликопротеинов тимуса (10-14 М) выявленные изменения амплитуды ПД восстанавливаются.
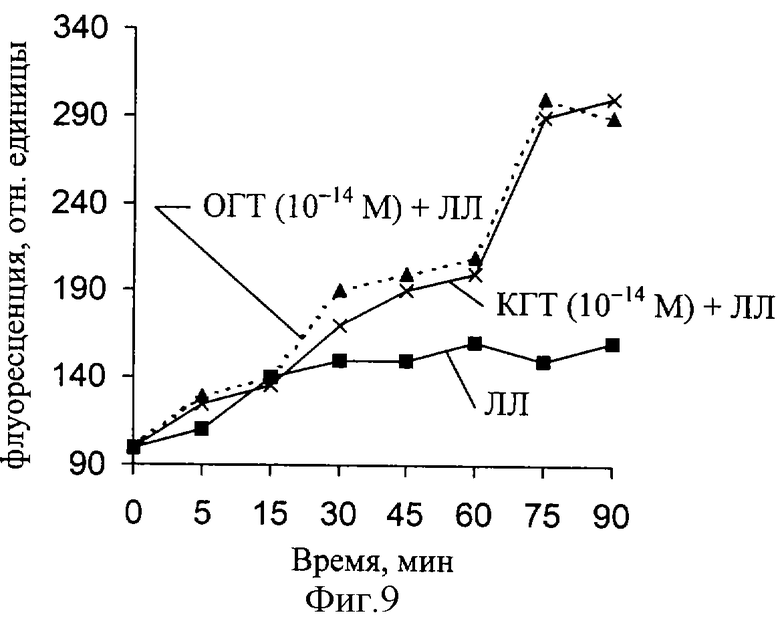
В следующей серии экспериментов изучали перераспределение межклеточного кальция, регистрируя флуоресценцию зонда-хлортетрациклин, что позволило выявить локализацию и изменение уровня мембраносвязанного кальция. Исследовали флуоресценцию нервов, нервных волокон, инкубированных в растворе ЛЛ, и волокон при совместном действии ЛЛ и гликопротеинов тимуса.
Ранее было показано, что при инкубации нерва с хлортетрациклином максимальное значение флуоресценции параметра, пропорционального уровню мембраносвязанного Са2+, регистрируется от мембранных структур нерва (плазматических мембран аксона и шванновской клеткой, а также миелина). При действии ЛЛ уровень мембраносвязанного Са увеличивается, но при совместном действии ЛЛ и гликопротеинов тимуса величина связывания увеличивается еще больше (фиг. 9-11). Как следует из полученных результатов, максимальная величина связывания кальция существенно меняется при действии ЛЛ и гликопротеинов тимуса в концентрации 10-14 М. По мере снижения концентрации гликопротеинов тимуса до 10-18 М максимальное связывание кальция мембранными структурами нерва снижается, но превышает контроль (действие ЛЛ). Снижение концентрации гликопротеинов тимуса до 10-24 М в течение 90 мин приводило к малым изменениям уровня связанного кальция. Полученные данные свидетельствуют о наличии тенденции в способности исследуемых гликопротеинов в "мнимых" растворах изменять уровень мембраносвязанного кальция.
Исследование выявило изменение ряда параметров, характеризующих РВ в условиях демиелинизации нерва, а также наличие характерных перестроек РВ при действии гликопротеинов тимуса как in vivo, так и in vitro.
Кольс О. Р. , Максимов Г.В., Раденович Ч.Н. Биофизика ритмического возбуждения, Москва, МГУ, 206, 1993.
Максимов Г. В., Орлов С.Н. Транспорт ионов кальция при функционировании нервного волокна: механизмы и регуляция. Москва, МГУ, 88, 1994.
Waxman S. G. , Kocsis J.D., Stys P.K. The axon. Structure, function and pathophysiology. Oxford Univ. Press., NY-Oxford, 325, 1995.
Пример 10. Влияние гликопротеина из сыворотки крови млекопитающих (рI в области рН 4,65-5,1) на проведение возбуждения при демиелинизации нервного волокна.
Эксперимент был проведен по методике, приведенной в примере 9. При действии ЛЛ и демиелинизации нерва происходят существенные изменения некоторых электрофизиологических параметров ритмического возбуждения (РВ). Было установлено, что in vivo сывороточный гликопротеин (СГ) в дозе 10-14 моль/л компенсирует изменения параметров РВ при демиелинизации нерва (табл. 6). Отметим, что собственно инъекция растворами гликопротеина не приводила к достоверным изменениям возбудимости миелинового нерва.
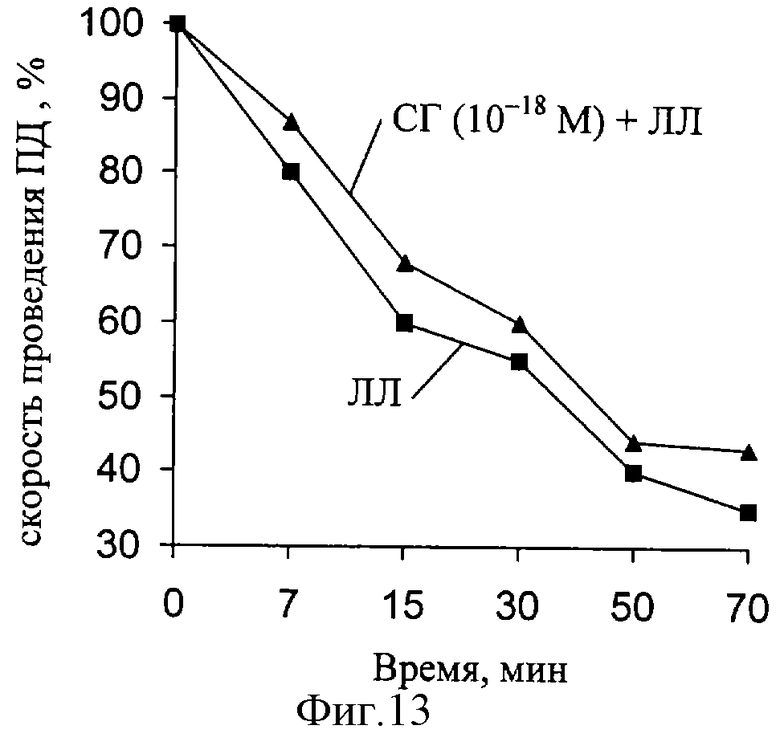
Было установлено, что при совместном действии на нерв ЛЛ и СГ характер изменений, вызываемых введением ЛЛ, уменьшается (фиг. 12, 13). При разбавлении раствора до концентрации 10-18 моль/л величина биологического эффекта была незначительной, но данные имели достоверные различия по сравнению с контрольными. Таким образом, длительная инкубация нервов в растворах СГ и ЛЛ приводит к статистически достоверному восстановлению скорости проведения ПД при концентрациях 10-14 моль/л и 10-18 моль/л.
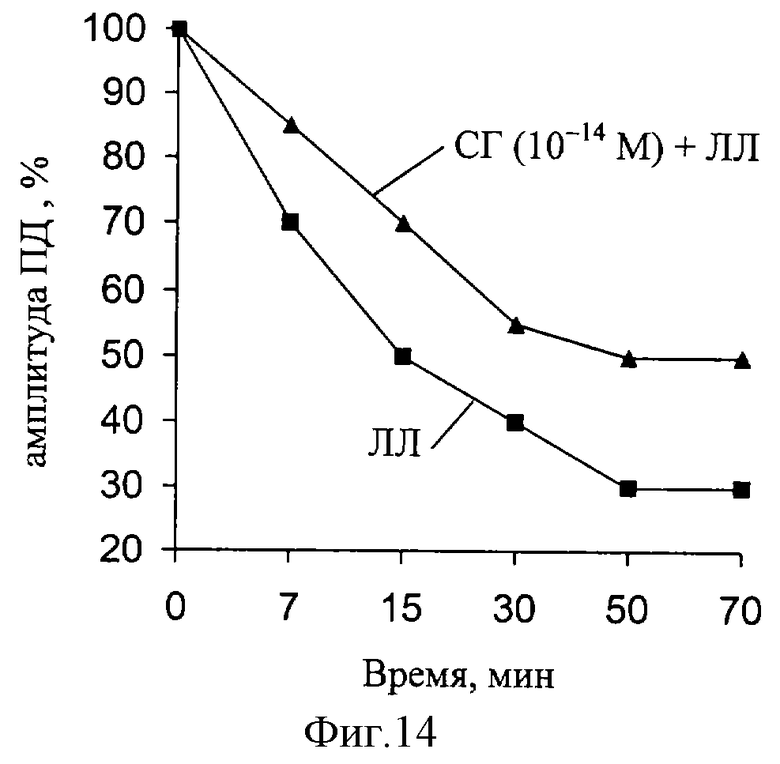
В следующей серии экспериментов были исследованы изменения амплитуды ПД, при действии ЛЛ и СГ (фиг. 14). Установлено, что за время эксперимента (действие ЛЛ) наблюдаются значительные изменения данного параметра РВ (при частоте 100Гц). При совместном действии ЛЛ и СГ (10-14 М) выявленные изменения амплитуды ПД восстанавливаются.
В следующей серии экспериментов изучали перераспределение межклеточного кальция, регистрируя флуоресценцию зондахлортетрациклин, что позволило выявить локализацию и изменения уровня мембраносвязанного кальция. Исследовали флуоресценцию нервов, нервных волокон, инкубированных в растворе ЛЛ, и волокон при совместном действии ЛЛ и СГ.
При действии ЛЛ уровень мембраносвязанного Са увеличивается, но при совместном действии ЛЛ и СГ величина связывания увеличивается еще больше (фиг. 15-17). Как следует из полученных результатов, максимальная величина связывания кальция существенно меняется при действии ЛЛ и СГ в концентрации 10-14 М. По мере снижения концентрации СГ до lO-24 M максимальное связывание кальция мембранными структурами нерва снижается. Полученные данные свидетельствуют о способности СГ в "мнимых" растворах изменять уровень мембраносвязанного кальция.
Исследование выявило изменение ряда параметров, характеризующих РВ в условиях демиелинизации нерва, а также наличие характерных перестроек РВ при действии сывороточного гликопротеина как in vivo, так и in vitro.
Пример 11. Влияние основного гликопротеина из сетчатки глаза быка на проведение возбуждения при демиелинизации нервного волокна.
Эксперимент был проведен по методике, приведенной в примере 9.
При действии ЛЛ и демиелинизации нерва происходят существенные изменения некоторых электрофизиологических параметров ритмического возбуждения (РВ). Было установлено, что in vivo основной гликопротеин из сетчатки глаза быка (ОГС) в дозах 10-14 моль/л и 10-18 моль/л компенсирует изменения параметров РВ при демиелинизации нерва (табл. 7). Отметим, что собственно инъекция раствором гликопротеина не приводила к достоверным изменениям возбудимости миелинового нерва.
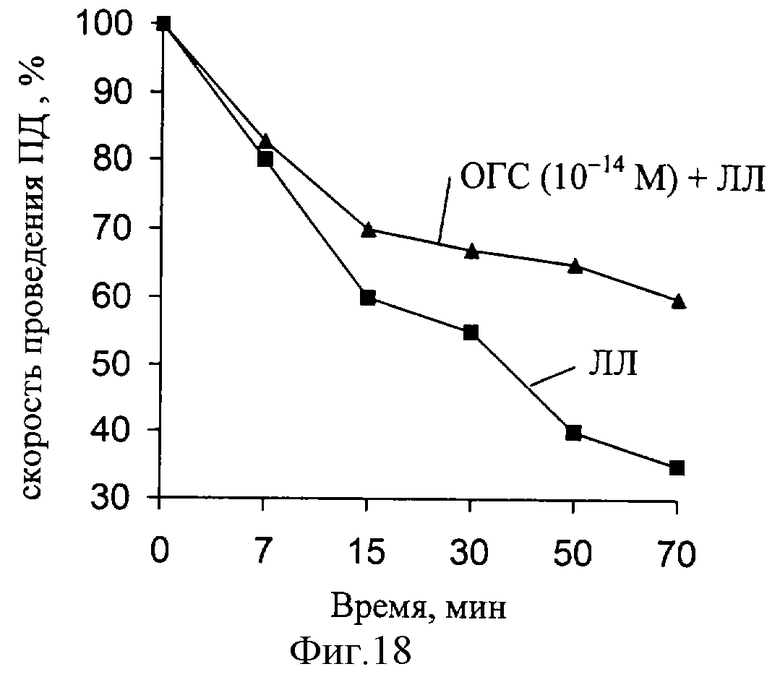
Было установлено, что при совместном действии на нерв ЛЛ и ОГС характер изменений, вызываемых введением ЛЛ, уменьшается (фиг. 18, 19). При разбавлении раствора ОГС до концентрации 10-18 моль/л величина биологического эффекта имела достоверные различия по сравнению с контрольными. Таким образом, длительная инкубация нервов в растворах СГ и ЛЛ приводит к статистически достоверному восстановлению скорости проведения ПД при концентрациях 10-14 моль/л и 10-18 моль/л.
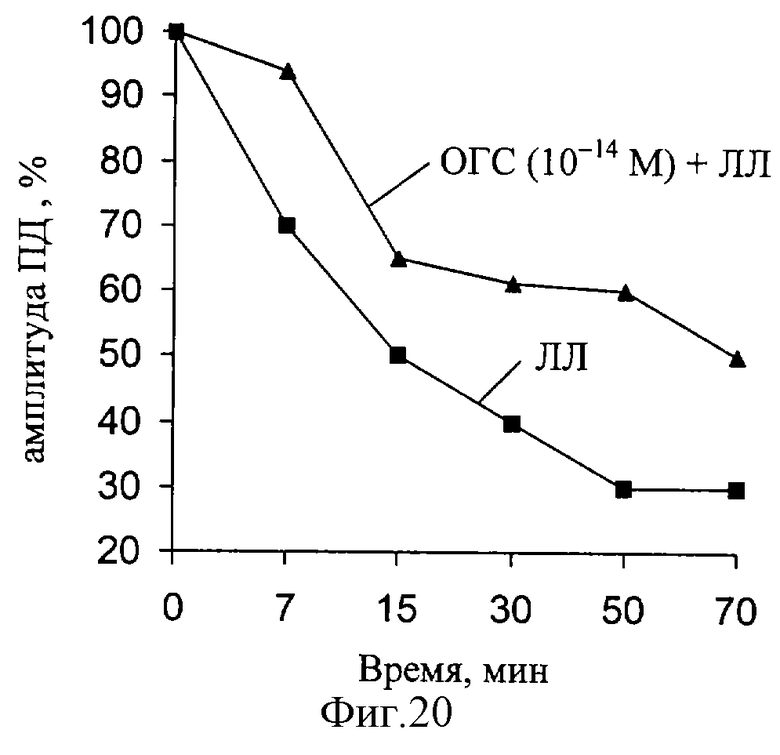
В следующей серии экспериментов были исследованы изменения амплитуды ПД, при действии ЛЛ и ОГС (фиг. 20). Установлено, что за время эксперимента (действие ЛЛ) наблюдаются значительные изменения данного параметра РВ (при частоте 100 Гц). При совместном действии ЛЛ и ОГС (10-14 М) выявленные изменения амплитуды ПД восстанавливаются.
В следующей серии экспериментов изучали перераспределение межклеточного кальция, регистрируя флуоресценцию зонда-хлортетрациклин, что позволило выявить локализацию и изменения уровня мембраносвязанного кальция. Исследовали флуоресценцию нервов, нервных волокон, инкубированных в растворе ЛЛ, и волокон при совместном действии ЛЛ и ОГС.
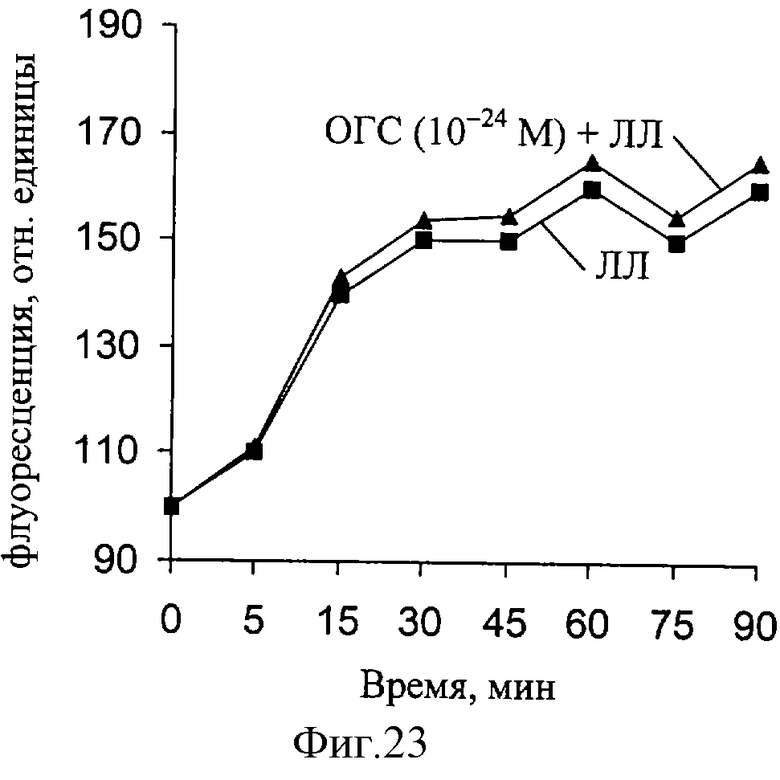
При действии ЛЛ уровень мембраносвязанного Са увеличивается, но при совместном действии ЛЛ и ОГС величина связывания увеличивается еще больше (фиг. 21-23). Как следует из полученных результатов, максимальная величина связывания кальция существенно меняется при действии ЛЛ и ОГС в концентрации 10-14 М. По мере снижения концентрации ОГС до 10-24 M максимальное связывание кальция мембранными структурами нерва снижается. Полученные данные свидетельствуют о способности ОГС в "мнимых" растворах изменять уровень мембраносвязанного кальция.
Исследование выявило изменение ряда параметров, характеризующих РВ в условиях демиелинизации нерва, а также наличие характерных перестроек РВ при действии основного гликопротеина из сетчатки глаза быка как in vivo, так и in vitro.
Пример 12. Влияние основного гликопротеина из пигментного эпителия глаза быка на проведение возбуждения при демиелинизации нервного волокна.
Эксперимент был проведен по методике, приведенной в примере 9.
При действии ЛЛ и демиелинизации нерва происходят существенные изменения некоторых электрофизиологических параметров ритмического возбуждения (РВ). Было установлено, что in vivo основной гликопротеин из пигментного эпителия глаза быка (ОГПЭ) в дозе 10-14 моль/л компенсирует изменения параметров РВ при демиелинизации нерва (табл. 8). Отметим, что собственно инъекция раствором гликопротеина не приводила к достоверным изменениям возбудимости миелинового нерва.
Было установлено, что при совместном действии на нерв ЛЛ и ОГПЭ характер изменений, вызываемых введением ЛЛ, уменьшается (фиг. 24, 25). Длительная инкубация нервов в растворах ОГПЭ и ЛЛ приводит к статистически достоверному восстановлению скорости проведения ПД при концентрации 10-14 моль/л.
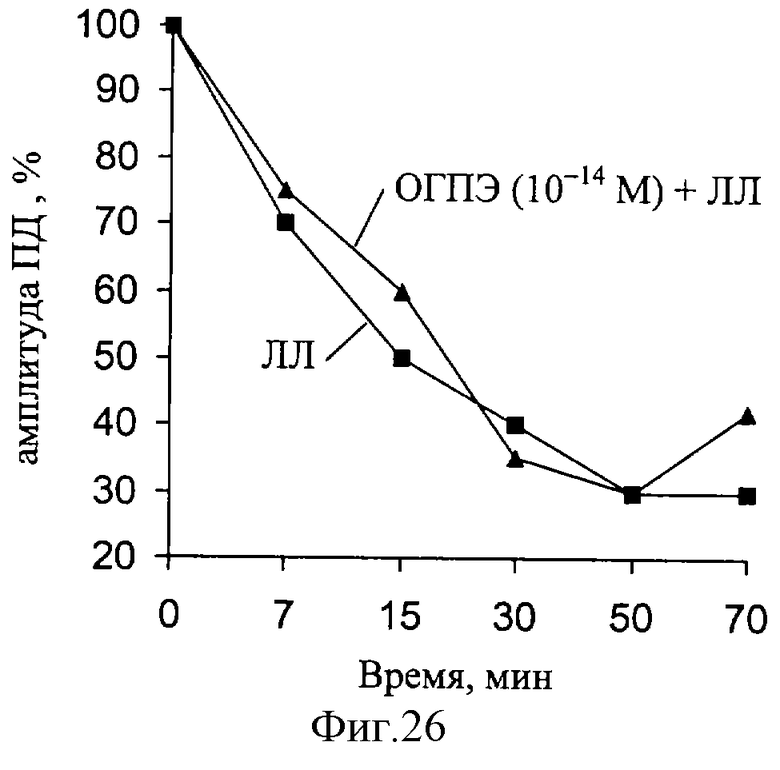
В следующей серии экспериментов были исследованы изменения амплитуды ПД, при действии ЛЛ и ОГПЭ (фиг. 26). Установлено, что за время эксперимента (действие ЛЛ) наблюдаются значительные изменения данного параметра РВ (при частоте 100 Гц). При совместном действии ЛЛ и ОГПЭ (10-14 М) выявленные изменения амплитуды ПД восстанавливаются.
В следующей серии экспериментов изучали перераспределение межклеточного кальция, регистрируя флуоресценцию зонда-хлортетрациклин, что позволило выявить локализацию и изменения уровня мембраносвязанного кальция. Исследовали флуоресценцию нервов, нервных волокон, инкубированных в растворе ЛЛ, и волокон при совместном действии ЛЛ и ОГПЭ.
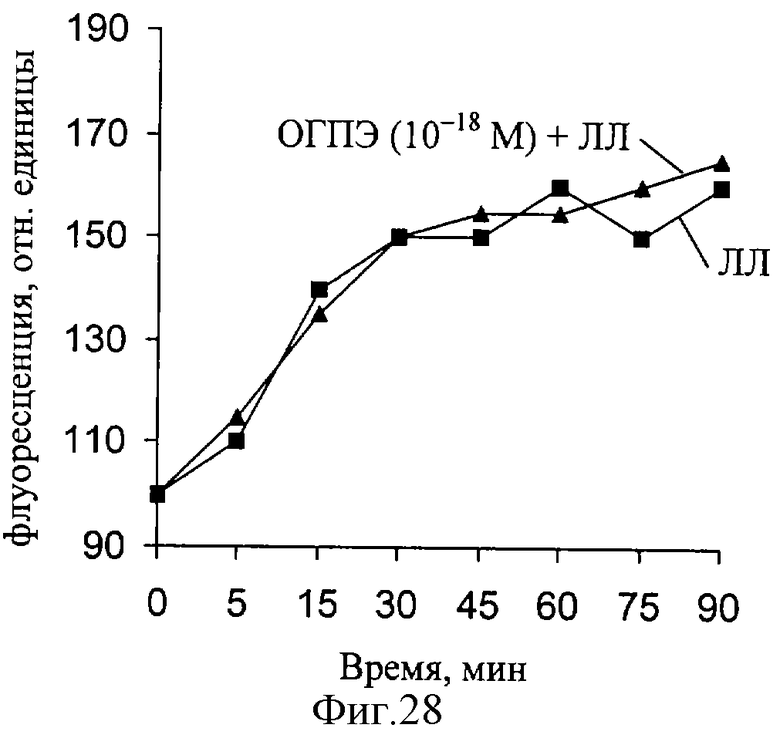
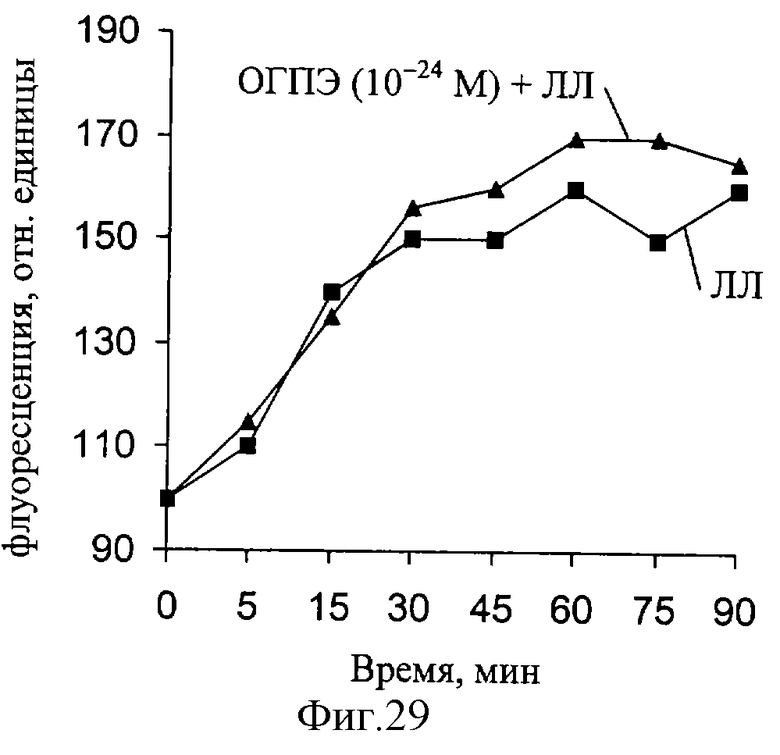
При действии ЛЛ уровень мембраносвязанного Са увеличивается, но при совместном действии ЛЛ и ОГПЭ величина связывания увеличивается еще больше (фиг. 27-29). Как следует из полученных результатов, максимальная величина связывания кальция существенно меняется при действии ЛЛ и ОГПЭ в концентрации 10-14 М. При снижении концентрации ОГПЭ до КГП и КГП максимальное связывание кальция мембранными структурами нерва снижается. Полученные данные свидетельствуют о способности ОГПЭ в "мнимых" (КГ моль/л) растворах изменять уровень мембраносвязанного кальция.
Исследование выявило изменение ряда параметров, характеризующих РВ в условиях демиелинизации нерва, а также наличие характерных перестроек РВ при действии основного гликопротеина из пигментного эпителия глаза быка как in vivo, так и in vitro.
Пример 13. Влияние кислого гликопротеина из печени на проницаемость плазматической мембраны гепатоцитов и интенсивность синтеза белка in vitro.
Эксперимент проводили на множественной органной культуре печени крыс линии Wistar, самцы весом 180 г. Всего было проведено 5 экспериментов. В каждом эксперименте использовали по 25-30 кусочков железы. Все исследуемые в одном эксперименте эксплантаты были получены из печени одного животного. Эксплантаты культивировали на границе жидкой питательной среды и воздуха, используя миллипоровые фильтры (Synpor, диаметр 0,6 мм). Культивирование осуществляли в течение 14-16 ч при 37oС. Группу эксплантатов после культивирования переносили на 10 мин, в бюксы с культуральной средой, содержащей меченый 3H-лейцин в дозе 25 μСu. Интенсивность синтеза белка оценивали по включению лейцина в белки, отнесенному к пулу меченого лейцина в том же образце. Проницаемость клеток для меченого предшественника определяли по пулу свободной меченой аминокислоты. Влияние кислого гликопротеина печени (КГП) определяли для его раствора в концентрации 10-14 моль/л. Раствор КГП добавляли в культуральную среду в опытные флаконы за 2 ч до инкубации с меченым лейцином, а затем на 10 мин переносили в среду, содержащую КГП и меченую аминокислоту в указанных концентрациях.
Было установлено, что интенсивность синтеза белка при воздействии КГП значительно снижалась по сравнению с контролем, приблизительно в два раза (в контроле отношение включения меченого лейцина к пулу этого предшественника составило, например, 0,42±0,09, а в опыте 0,21±0,04). Проницаемость мембраны гепатоцитов увеличивалась весьма значительно в присутствии КГП (в контроле, например, пул свободных аминокислот составлял 1120±50 имп. в 1 мин, а в опытных флаконах соответственно 2180±30).
Морфология эксплантатов опытной органной культуры по сравнению с контрольной не изменялась.
Результаты проведенного исследования показывают, что КГП оказывает влияние на интенсивность синтеза белка в гепатоцитах и проницаемость их плазматических мембран в сверхмалой дозе при условии сохранения структуры ткани печени.
Пример 14. Влияние гликопротеина из сыворотки крови млекопитающих (рI в области рН 4,65-5,1) на проницаемость плазматической мембраны гепатоцитов и интенсивность синтеза белка in vitro.
Эксперимент проводили на множественной органной культуре печени крыс линии Wistar, самцы весом 180 г. Всего было проведено 5 экспериментов. В каждом эксперименте использовали по 25-30 кусочков железы. Все исследуемые в одном эксперименте эксплантаты были получены из печени одного животного. Эксплантаты культивировали на границе жидкой питательной среды и воздуха, используя миллипоровые фильтры (Synpor, диаметр 0,6 мм). Культивирование осуществляли в течение 14-16 ч при 37oС. Группу эксплантатов после культивирования переносили на 10 мин, в бюксы с культуральной средой, содержащей меченый 3H-лейцин в дозе 25 μCu. Интенсивность синтеза белка оценивали по включению лейцина в белки, отнесенному к пулу меченого лейцина в том же образце. Проницаемость клеток для меченого предшественника определяли по пулу свободной меченой аминокислоты. Влияние сывороточного гликопротеина (СГ) определяли для его раствора в концентрации 10-14 моль/л. Раствор СГ добавляли в культуральную среду в опытные флаконы за 2 ч до инкубации с меченым лейцином, а затем на 10 мин переносили в среду, содержащую СГ и меченую аминокислоту в указанных концентрациях.
Было установлено, что интенсивность синтеза белка при воздействии СГ значительно снижалась по сравнению с контролем, приблизительно в два раза (в контроле отношение включения меченого лейцина к пулу этого предшественника составило, например, 0,42±0,09, а в опыте 0,17±0,05). Проницаемость мембраны гепатоцитов увеличивалась весьма значительно в присутствии СГ (в контроле, например, пул свободных аминокислот составлял 1120±50 имп в 1 мин, а в опытных флаконах соответственно 2980±30).
Морфология эксплантатов опытной органной культуры по сравнению с контрольной не изменялась.
Результаты проведенного исследования показывают, что СГ оказывает влияние на интенсивность синтеза белка в гепатоцитах и проницаемость их плазматических мембран в сверхмалой дозе при условии сохранения структуры ткани печени.
Нижеследующие примеры (15-17) описывают фармацевтическую композицию с использованием гликопротеина по данному изобретению.
Пример 15. Композиция на основе гликопротеина из сыворотки крови млекопитающих (рI в области рН 4,65-5,1) в сверхмалых дозах, обладающая фармакологическим действием.
Композиция состава:
Сывороточный гликопротеин - 1•10-10 г
Натрия хлорид - 8,8 г
Кальция хлорид - 0,001 г
Вода - до 1 л
является биорегулятором репаративных процессов в эпителиальных и соединительных тканях.
При применении в качестве глазных капель композиция указанного состава способствует заживлению роговицы глаза после механической травмы или ожога, вызывает образование нежного рубца, ограничивая в то же время излишнее разрастание рубцовой ткани. Особенно эффективна при пересадке роговицы, лечении кератитов и некоторых конъюктивитов. Применение композиции при лечении проникающих ранений роговицы приводит к уменьшению сроков воспалительной реакции, быстрой ликвидации диастаза краев раны, ускорению эпителизации поверхности повреждения, более раннему восстановлению передней камеры, снижению частоты осложнений (выпот фибрина и гипопион), ускорению процессов репаративной регенерации и реконструкции новообразованной рубцовой ткани, в результате чего образуется более компактный и структурированный рубец с преобладанием пролиферативного компонента без активной васкуляризации роговицы. На клеточном уровне отмечается ускорение миграции эпителия, макрофагов, кератобластов и уменьшение лейкоцитарной инфильтрации, раннее формирование непрерывного слоя эндотелиев без признаков десквамации в зоне раневого канала, что в конечном итоге приводит к быстрому заполнению раневого канала эпителиально-фибринозным компонентом, более раннему и точному смыканию краев раны, активной резорбции фибрина и замещению его кератобластическим пролифератом, синтезирующим новые коллагеновые волокна. При применении композиции отмечается более компактное расположение коллагеновых пластин, преобладание волокнистого компонента пролиферата над клеточным, что определяет качество рубцовой ткани. Формирование более нежных, компактных бессосудистых рубцов приводит к меньшему изменению прозрачности и рефракции роговицы. В случае ожогового поражения глаз лечебный эффект композиции указанного состава выражается к 14-м суткам в значительной пролиферации фибробластических элементов, инфильтрующих травмированную роговичную ткань, причем векторная направленность пролиферирующих клеток обусловливает пластинчатое строение роговицы, имитирующее исходную морфологическую структуру ткани.
При применении в инъекционной форме композиция указанного состава стимулирует регенерацию костной ткани при переломах конечностей, в том числе при переломе шейки бедра, эффективна при лечении ряда тяжелых патологий суставов, связанных с нарушением структуры и функции хрящевой ткани, для лечения механических повреждений суставного хряща коленного сустава, при лечении артрозов, синовиитов. Введение инъекций композиции тормозит процесс дегенерации поврежденной хрящевой ткани. При применении композиции для лечения повреждений суставного хряща происходит быстрое накопление массы молодых хрящевых клеток и их дифференцировка, приводящая к более быстрому, чем в контроле образованию замещающей хрящевой ткани и восстановлению ровной суставной поверхности в области повреждения, что в свою очередь играет основную роль в восстановлении подвижности сустава. Происходит заполнение области повреждения молодыми хрящевыми клетками и восстановление хрящевой ткани в дефинитивном виде, так как имеет место ее дифференцировка на слои, присущие исходному гиалиновому хрящу. При внутрисуставном введении композиции отмечено уменьшение болевого синдрома после 2-3-ей инъекции препарата. В сравнении эффективности применения композиции указанного состава с наиболее известными хондропротекторами, например "Zeel", при травматических повреждениях суставного хряща коленного сустава у спортсменов, можно отметить, что болевой синдром и синовиит купировался в среднем на 7-10 сутки в случае применения данной композиции и на 14-17 сутки в случае применения "Zeel". Следовательно, восстановление спортивных результатов на прежнем уровне происходило в 2-2.5 раза быстрее, чем при применении препарата "Zeel".
Композиции различного состава, содержащие в качестве активного компонента сывороточный гликопротеин в концентрации 1•10-10 г гликопротеина/л (или кг) композиции в различной лекарственной форме (гели, мази, свечи, раствор) обладают ранозаживляющим действием и эффективны при восстановлении поврежденных кожных покровов, в том числе при лечении ожоговой болезни, пролежней и предупреждения их образования; стимулируют репаративные процессы в коже после лучевого поражения, возникающего у онкологических больных после радиотерапии. Эффективно применение таких композиций в гастроэнтерологии (язвенная болезнь, гастриты, гастродуодениты), в проктологии (заболевания толстой кишки), в гинекологии (эрозия шейки матки), в кардиологии (реабилитационный период после инфаркта миокарда).
Пример 16. Композиция на основе кислого гликопротеина из сетчатки глаза крупного рогатого скота в сверхмалых дозах, обладающая фармакологическим действием.
Композиция состава:
Кислый гликопротеин из сетчатки глаза - 1•10-10 г
Натрия хлорид - 8,8 г
Кальция хлорид - 0,001 г
Вода - До 1 л
является биорегулятором репаративных процессов, способствующим восстановлению нарушенной функции сетчатки глаза, предотвращает отслоение сетчатки при хирургических вмешательствах.
Композиция указанного состава оказывает стимулирующее действие на функционирование основных ферментных систем в сетчатке глаза, ответственных за осуществление зрительного акта, ингибирует перекисное окисление липидов в мембранах клеток сетчатки. Композиция является эффективным биорегулятором, отвечающим за позиционное расположение клеток в гистоструктуре сетчатки и клеточное деление, способствует восстановлению пространственно-функциональной организации ткани сетчатки после нанесения травмы или развития патологического процесса. Данная композиция оказывает лечебное действие при миопической болезни, дегенерации сетчатки различной этиологии, состояниях после проникающих ранений глаза.
Пример 17. Композиция на основе кислого гликопротеина из пигментного эпителия глаза крупного рогатого скота в сверхмалых дозах, обладающая фармакологическим действием.
Композиция состава:
Кислый гликопротеин из пигментного эпителия глаза - 1•10-10 г
Натрия хлорид - 8,8 г
Кальция хлорид - 0,001 г
Вода - До 1 л
является биорегулятором, способствующим восстановлению нарушенной функции пигментного эпителия глаза.
В настоящее время отсутствуют фармакологические средства, применение которых вызывало бы торможение патологического процесса на начальных стадиях развития ретинитов различной этиологии и макулопатий. Композиция указанного состава эффективна для лечения макулопатий и ретинитов различной этиологии.
























Изобретение относится к медицине, а именно к получению новых биологически активных соединений и созданию лекарственных средств на их основе. Предложены гликопротеины, выделенные изоэлектрическим фокусированием из межклеточного пространства тканей различных органов, сыворотки крови, желчи человека и животных, растворимые в насыщенном (100%-ном) растворе сернокислого аммония и имеющие кажущуюся молекулярную массу 10-45 кДа. Такие гликопротеины обладают биологической активностью в сверхмалых дозах 10-12-10-29 моль/л и ниже. Предложены также фармацевтические композиции, включающие указанные гликопротеины в эффективном количестве и фармацевтически приемлемый носитель. 3 с.п. ф-лы, 29 ил., 8 табл.
| US 4663438 А, 05.05.1987 | |||
| СЫВОРОТОЧНЫЙ ГЛИКОПРОТЕИН, ОБЛАДАЮЩИЙ БИОЛОГИЧЕСКОЙ АКТИВНОСТЬЮ В СВЕРХМАЛЫХ ДОЗАХ | 1998 |
|
RU2136695C1 |
| СПОСОБ ПОЛУЧЕНИЯ СЫВОРОТОЧНОГО БЕЛКОВОГО ПРЕПАРАТА, ВЛИЯЮЩЕГО НА АКТИВНОСТЬ ОКИСЛИТЕЛЬНО-ЭНЕРГЕТИЧЕСКОГО ОБМЕНА | 1992 |
|
RU2078345C1 |
| ЕР 0228357 A3, 08.07.1987 | |||
| ТРУБОПРОВОДНЫЙ ТРАНСПОРТ | 1996 |
|
RU2097647C1 |
| US 4559230 А1, 17.12.1985 | |||
| US 4169139 А1, 25.09.1979. | |||

