ОБЛАСТЬ ТЕХНИКИ
Данное изобретение относится в основном к терапии и диагностике такой злокачественной опухоли, как рак предстательной железы. Изобретение более конкретно относится к полипептидам, содержащим, по меньшей мере, часть опухолевого белка предстательной железы, и к полинуклеотидам, кодирующим такие полипептиды. Такие полипептиды и полинуклеотиды могут быть использованы в вакцинах и фармацевтических композициях для профилактики и лечения рака предстательной железы и для диагностики и мониторинга таких раковых опухолей.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Рак предстательной железы является наиболее распространенной формой рака среди мужчин с оцениваемой встречаемостью 30% у мужчин в возрасте свыше 50 лет. Многочисленные клинические доказательства показывают, что рак предстательной железы человека обладает тенденцией метастазировать в кость, и это заболевание, по-видимому, неуклонно прогрессирует из андроген-зависимого статуса в андроген-резистентный статус, приводя к повышенной смертности пациентов. Это преобладающее заболевание является в настоящее время второй по частоте причиной смерти от рака среди мужчин в Соединенных Штатах.
Несмотря на значительные исследования в терапии указанного заболевания, лечение рака предстательной железы остается трудным. Обычно лечение основывается на хирургии и/или лучевой терапии, но эти способы являются неэффективными в значительном проценте случаев. Два идентифицированных ранее специфических белка предстательной железы - специфический антиген предстательной железы (PSA) и кислая фосфатаза предстательной железы (PAP) - имеют ограниченный терапевтический и диагностический потенциал. Например, содержание PSA не всегда хорошо коррелирует с наличием рака предстательной железы, являясь положительным в некотором проценте случаев, не являющихся раком предстательной железы, в том числе в случае доброкачественной гиперплазии рака предстательной железы (ВРН). Кроме того, содержание PSA коррелирует с объемом предстательной железы и не указывает уровень метастазирования.
Несмотря на значительные исследования в области терапии этих и других раковых опухолей, рак предстательной железы по-прежнему трудно диагностировать и лечить. Соответственно в данной области имеется потребность в усовершенствованных способах обнаружения и лечения подобных раковых опухолей. Данное изобретение удовлетворяет эти потребности и дополнительно обеспечивает другие связанные с ними преимущества.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Вкратце, данное изобретение обеспечивает композиции и способы для диагностики и терапии рака, такого как рак предстательной железы. В одном аспекте данное изобретение обеспечивает полипептиды, содержащие, по меньшей мере, часть опухолевого белка предстательной железы или его вариант. Определенные части и другие варианты являются иммуногенными, так что способность варианта реагировать с антиген-специфическими антисыворотками по существу не является уменьшенной. В некоторых вариантах указанный полипептид содержит, по меньшей мере, иммуногенную часть опухолевого белка предстательной железы или его вариант, причем опухолевый белок содержит аминокислотную последовательность, которая кодируется полинуклеотидной последовательностью, выбранной из группы, состоящей из: (а) последовательностей, представленных в любой из последовательностей SEQ ID NO:1-111, 115-171, 173-175, 177, 179-305, 307-315, 326, 328, 330, 332-335, 340-375, 381, 382 или 384-472; (b) последовательностей, которые гибридизуются с любой из указанных выше последовательностей при умеренно жестких условиях; и (с) комплементов любой из последовательностей (а) или (b). В некоторых характерных вариантах такой полипептид содержит, по меньшей мере, часть или вариант опухолевого белка, которые включают в себя аминокислотную последовательность, выбранную из группы, состоящей из последовательностей, представленных в любой из последовательностей SEQ ID NO:112-114, 172, 176, 178, 327, 329, 331, 336, 339, 376-380 и 383.
Далее, данное изобретение обеспечивает полинуклеотиды, которые кодируют описанный выше полипептид, или его часть (такую как часть, кодирующую, по меньшей мере, 15 аминокислотных остатков опухолевого белка предстательной железы), экспрессирующие векторы, содержащие такие полинуклеотиды, и клетки-хозяева, трансформированные или трансфицированные такими экспрессирующими векторами.
В других аспектах данное изобретение обеспечивает фармацевтические композиции, содержащие полипептид или полинуклеотид, описанные выше, и физиологически приемлемый носитель.
В родственном аспекте данного изобретения обеспечены вакцины. Такие вакцины содержат полипептид или полинуклеотид, описанные выше, и неспецифический усилитель иммунного ответа.
Данное изобретение обеспечивает дополнительно фармацевтические композиции, которые содержат: (а) антитело или его антигенсвязывающий фрагмент, который специфически связывается с опухолевым белком предстательной железы; и (b) физиологически приемлемый носитель.
В следующих аспектах данное изобретение обеспечивает фармацевтические композиции, содержащие: (а) антиген-презентирующую клетку, которая экспрессирует полипептид, описанный выше; и (b) фармацевтически приемлемый носитель или наполнитель. Антиген-презентирующие клетки включают в себя дендритные клетки, макрофаги, моноциты, фибробласты и В-клетки.
В родственных аспектах обеспечены вакцины, которые содержат: (а) антиген-презентирующую клетку, которая экспрессирует полипептид, описанный выше, и (b) неспецифический усилитель иммунного ответа.
Данное изобретение обеспечивает далее, в других аспектах, слитые (гибридные) белки, которые содержат, по меньшей мере, один полипептид, описанный выше, а также полинуклеотиды, кодирующие такие слитые белки.
В родственных аспектах обеспечены фармацевтические композиции, содержащие слитый белок или полинуклеотид, кодирующий слитый белок, в комбинации с физиологически приемлемым носителем.
Далее, обеспечены вакцины в других аспектах, которые содержат слитый белок или полинуклеотид, кодирующий слитый белок, в комбинации с неспецифическим усилителем иммунного ответа.
В следующих аспектах данное изобретение обеспечивает способы подавления развития раковой опухоли у пациента, предусматривающие введение пациенту фармацевтической композиции или вакцины, описанных выше.
Далее, данное изобретение обеспечивает, в других аспектах, способы удаления опухолевых клеток из биологического образца, предусматривающие взаимодействие биологического образца с Т-клетками, которые специфически реагируют с опухолевым белком предстательной железы, причем стадию взаимодействия выполняют в условиях и в течение времени, достаточных для возможности удаления клеток, экспрессирующих указанный белок, из образца.
В родственных вариантах обеспечены способы подавления развития раковой опухоли у пациента, предусматривающие введение пациенту биологического образца, обработанного, как описано выше.
Дополнительно обеспечены способы, в других аспектах, для стимуляции и/или размножения Т-клеток, специфических для опухолевого белка предстательной железы, предусматривающие взаимодействие Т-клеток с одним или более из таких компонентов, как: (i) полипептид, описанный выше; (ii) полинуклеотид, кодирующий такой полипептид; и/или (iii) антиген-презентирующая клетка, которая экспрессирует такой полипептид; в условиях и в течение времени, достаточных для возможности стимуляции и/или размножения Т-клеток. Также обеспечены выделенные популяции Т-клеток, содержащие Т-клетки, полученные, как описано выше.
В следующих аспектах данное изобретение обеспечивает способы подавления развития раковой опухоли у пациента, предусматривающие введение пациенту эффективного количества популяции Т-клеток, описанной выше.
Далее, данное изобретение обеспечивает способы подавления развития раковой опухоли у пациента, предусматривающие стадии; (а) инкубирования CD4+ и/или CD8+ Т-клеток, выделенных из пациента, с одним или более из таких компонентов, как: (i) полипептид, содержащий, по меньшей мере, иммуногенную часть опухолевого белка предстательной железы; (ii) полинуклеотид, кодирующий такой полипептид; и (iii) антиген-презентирующая клетка, которая экспрессирует такой полипептид; и (b) введения пациенту эффективного количества пролиферированных Т-клеток и тем самым подавления развития раковой опухоли у пациента. Пролиферированные клетки могут, но необязательно, быть клонированы перед введением пациенту.
В следующих аспектах данное изобретение обеспечивает способы определения наличия или отсутствия раковой опухоли у пациента, предусматривающие: (а) взаимодействие биологического образца, полученного из пациента, со связывающим агентом, который связывается с полипептидом, описанным выше; (b) определение в указанном образце количества полипептида, который связывается со связывающим агентом; и (с) сравнение указанного количества полипептида с заранее определенной предельной величиной и определение из указанного сравнения наличия или отсутствия раковой опухоли у пациента. В предпочтительных вариантах связывающим агентом является антитело, более предпочтительно моноклональное антитело. Рак может быть раком предстательной железы.
Данное изобретение обеспечивает также, в других аспектах, способы мониторинга прогрессирования раковой опухоли у пациента. Такие способы включают в себя стадии: (а) взаимодействия биологического образца, полученного из пациента, в первой временной точке со связывающим агентом, который связывается с полипептидом, описанным выше; (b) определения в указанном образце количества полипептида, который связывается со связывающим агентом; (с) повторения стадий (а) и (b) с использованием биологического образца, полученного из пациента, в следующей временной точке; и (d) сравнения количества полипептида, определяемого в стадии (с), с количеством, определяемым в стадии (b), и мониторинга на основании указанного сравнения прогрессирования раковой опухоли у пациента.
Данное изобретение обеспечивает, далее, в других аспектах, способы определения наличия или отсутствия раковой опухоли у пациента, предусматривающие стадии: (а) взаимодействия биологического образца, полученного из пациента, с олигонуклеотидом, который гибридизуется с полинуклеотидом, кодирующим опухолевой белок предстательной железы; (b) определения в указанном образце уровня полинуклеотида, предпочтительно мРНК, который гибридизуется с указанным олигонуклеотидом; и (с) сравнения уровня полинуклеотида, который гибридизуется с олигонуклеотидом, с заранее определенной предельной величиной и определения из указанного сравнения наличия или отсутствия раковой опухоли у пациента. В некоторых вариантах количество мРНК определяют через полимеразную цепную реакцию с использованием, например, по меньшей мере, одного олигонуклеотидного праймера, который гибридизуется с полинуклеотидом, кодирующим полипептид, описанный выше, или комплементом такого полинуклеотида. В других вариантах количество мРНК определяют с использованием гибридизационного способа, применяющего олигонуклеотидный зонд, который гибридизуется с полинуклеотидом, кодирующим полипептид, описанный выше, или комплементом такого полинуклеотида.
В родственных аспектах обеспечены способы мониторинга прогрессирования раковой опухоли у пациента, предусматривающие стадии; (а) взаимодействия биологического образца, полученного из пациента, с олигонуклеотидом, который гибридизуется с полинуклеотидом, кодирующим опухолевой белок предстательной железы; (b) определения в указанном образце количества полинуклеотида, который гибридизуется с указанным олигонуклеотидом; (с) повторения стадий (а) и (b) с использованием биологического образца, полученного из пациента, в следующий момент времени; и (d) сравнения количества полинуклеотида, определяемого в стадии (с), с количеством, определяемым в стадии (b), и на основании указанного мониторинга прогрессирования раковой опухоли у пациента.
В следующих аспектах данное изобретение обеспечивает антитела, такие как моноклональные антитела, которые связываются с описанным выше полипептидом, а также диагностические наборы, содержащие такие антитела. Обеспечены также диагностические наборы, содержащие один или несколько олигонуклеотидных зондов или праймеров, описанных выше.
Эти и другие аспекты данного изобретения станут очевидными в связи со следующим подробным описанием и прилагаемыми чертежами. Все приведенные здесь ссылки включены в качестве ссылки во всей их полноте, как если бы каждая из них была включена отдельно.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ И ИДЕНТИФИКАЦИОННЫЕ НОМЕРА (ИДЕНТИФИКАТОРЫ) ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Фигура 1 иллюстрирует способность Т-клеток узнавать клетки, экспрессирующие показательный опухолевый полипептид предстательной железы P502S в сравнении с контрольными фибробластами. Лизис в процентах показан в виде ряда соотношений эффектор : мишень, как указано.
Фигуры 2А и 2В иллюстрируют способность Т-клеток узнавать клетки, экспрессирующие показательный опухолевый полипептид предстательной железы P502S. В каждом случае количество пятен γ-интерферона показано для разных количеств иммунокомпетентных клеток (клеток-респондеров). На фигуре 2А представлены данные для фибробластов, кратковременно обработанных пептидом PDS-12, в сравнении с фибробластами, кратковременно обработанными контрольным пептидом Е75. На фигуре 2В представлены данные для фибробластов, экспрессирующих P502S, в сравнении с фибробластами, экспрессирующими HER-2/neu.
Фигура 3 представляет тест конкурентного связывания пептидов, показывающий, что пептид P1S#10, произведенный из P501S, связывает HLA-A2. Пептид P1S#10 подавляет рестриктированную (ограниченную) HLA-A2 презентацию пептида fluM58 CTL-клону D150M58 в биоанализе высвобождения TNF. CTL D150M58 является специфическим для НLА-А2-связывающего матричного пептида fluM58 вируса гриппа.
Фигура 4 иллюстрирует способность Т-клеточных линий, получаемых из иммунизированных P1S#10 мышей, специфически лизировать клетки-мишени Jurkat А2Кb, кратковременно обработанные P1S#10, и трансдуцированные P501S клетки-мишени Jurkat А2Кb, в сравнении с трансдуцированными EGFP клетками Jurkat A2Kb. Лизис в процентах показан в виде ряда соотношений эффектор : мишень, как указано.
Фигура 5 иллюстрирует способность клона Т-клеток узнавать и специфически лизировать клетки Jurkat A2Kb, экспрессирующие показательный опухолевый полипептид P501S предстательной железы, показывая, что пептид P1S#10 может быть природно процессируемым эпитопом полипептида P501S.
Фигуры 6А и 6В (7) являются графиками, иллюстрирующими специфичность CD8+ клеточной линии (3А-1) в отношении показательного антигена опухоли предстательной железы (P501S). Фигура 6А показывает результаты анализа высвобождения 51Сr. Специфический лизис в процентах показан в виде ряда соотношений эффектор : мишень, как указано. Фигура 6В (7) показывает продуцирование интерферона-гамма клетками 3А-1, стимулированными аутологичными B-LCL, трансдуцированными P501S, при различных соотношениях эффектор : мишень, как указано.
SEQ ID NO:1 является определенной последовательностью-кДНК для F1-13
SEQ ID NO:2 является определенной последовательностью-кДНК для F1-12
SEQ ID NO:3 является определенной 5’-последовательностью-кДНК для F1-12
SEQ ID NO:4 является определенной 3’-последовательностью-кДНК для F1-16
SEQ ID NO:5 является определенной 3’-последовательностью-кДНК для Н1-1
SEQ ID NO:6 является определенной 3’-последовательностью-кДНК для H1-9
SEQ ID NO:7 является определенной 3’-последовательностью-кДНК для H1-4
SEQ ID NO:8 является определенной 3’-последовательностью-кДНК для J1-17
SEQ ID NO:9 является определенной 5’-последовательностью-кДНК для J1-17
SEQ ID NO:10 является определенной 3’-последовательностью-кДНК для L1-12
SEQ ID NO:11 является определенной 5’-последовательностью-кДНК для L1-12
SEQ ID NO:12 является определенной 3’-последовательностью-кДНК для N1-1862
SEQ ID NO:13 является определенной 5’-последовательностью-кДНК для N1-1862
SEQ ID NO:14 является определенной 3’-последовательностью-кДНК для J1-13
SEQ ID NO:15 является определенной 5’-последовательностью-кДНК для J1-13
SEQ ID NO:16 является определенной 3’-последовательностью-кДНК для J1-19
SEQ ID NO:17 является определенной 5’-последовательностью-кДНК для J1-19
SEQ ID NO:18 является определенной 3’-последовательностью-кДНК для J1-25
SEQ ID NO:19 является определенной 5’-последовательностью-кДНК для J1-25
SEQ ID NO:20 является определенной 5’-последовательностью-кДНК для J1-24
SEQ ID NO:21 является определенной 3’-последовательностью-кДНК для J1-24
SEQ ID NO:22 является определенной 5’-последовательностью-кДНК для К1-58
SEQ ID NO:23 является определенной 3’-последовательностью-кДНК для К1-58
SEQ ID NO:24 является определенной 5’-последовательностью-кДНК для К1-63
SEQ ID NO:25 является определенной 3’-последовательностью-кДНК для К1-63
SEQ ID NO:26 является определенной 5’-последовательностью-кДНК для L1-4
SEQ ID NO:27 является определенной 3’-последовательностью-кДНК для L1-4
SEQ ID NO:28 является определенной 5’-последовательностью-кДНК для L1-14
SEQ ID NO:29 является определенной 3’-последовательностыо-кДНК для L1-14
SEQ ID NO:30 является определенной 3’-последовательностью-кДНК для J1-12
SEQ ID NO:31 является определенной 3’-последовательностью-кДНК для J1-16
SEQ ID NO:32 является определенной 3’ -последовательностью-кДНК для J1-21
SEQ ID NO:33 является определенной 3’-последовательностью-кДНК для К1-48
SEQ ID NO:34 является определенной 3’-последовательностью-кДНК для для К1-55
SEQ ID NO:35 является определенной 3’-последовательностыо-кДНК для L1-2
SEQ ID NO:36 является определенной 3’-последовательностью-кДНК для L1-6
SEQ ID NO:37 является определенной 3’-последовательностью-кДНК для N1-1858
SEQ ID NO:38 является определенной 3’-последовательностью-кДНК для N1-1860
SEQ ID NO:39 является определенной 3’-последовательностью-кДНК для N1-1861
SEQ ID NO:40 является определенной 3’-последовательностью-кДНК для N1-1864
SEQ ID NO:41 является определенной последовательностью-кДНК для Р5
SEQ ID NO:42 является определенной последовательностью-кДНК для Р8
SEQ ID NO:43 является определенной последовательностью-кДНК для Р9
SEQ ID NO:44 является определенной последовательностью-кДНК для Р18
SEQ ID NO:45 является определенной последовательностью-кДНК для Р20
SEQ ID NO:46 является определенной последовательностью-кДНК для Р29
SEQ ID NO:47 является определенной последовательностью-кДНК для Р30
SEQ ID NO:48 является определенной последовательностью-кДНК для Р34
SEQ ID NO:49 является определенной последовательностью-кДНК для Р36
SEQ ID NO:50 является определенной последовательностью-кДНК для Р38
SEQ ID NO:51 является определенной последовательностью-кДНК для Р39
SEQ ID NO:52 является определенной последовательностью-кДНК для Р42
SEQ ID NO:53 является определенной последовательностью-кДНК для Р47
SEQ ID NO:54 является определенной последовательностью-кДНК для Р49
SEQ ID NO:55 является определенной последовательностью-кДНК для Р50
SEQ ID NO:56 является определенной последовательностыо-кДНК для Р53
SEQ ID NO:57 является определенной последовательностью-кДНК для Р55
SEQ ID NO:58 является определенной последовательностью-кДНК для Р60
SEQ ID NO:59 является определенной последовательностью-кДНК для Р64
SEQ ID NO:60 является определенной последовательностью-кДНК для Р65
SEQ ID NO:61 является определенной последовательностью-кДНК для Р73
SEQ ID NO:62 является определенной последовательностью-кДНК для Р75
SEQ ID NO:63 является определенной последовательностью-кДНК для Р76
SEQ ID NO:64 является определенной последовательностью-кДНК для Р79
SEQ ID NO:65 является определенной последовательностью-кДНК для Р84
SEQ ID NO:66 является определенной последовательностью-кДНК для Р68
SEQ ID NO:67 является определенной последовательностью-кДНК для Р80
SEQ ID NO:68 является определенной последовательностью-кДНК для Р82
SEQ ID NO:69 является определенной последовательностью-кДНК для U1-3064
SEQ ID NO:70 является определенной последовательностью-кДНК для U1-3065
SEQ ID NO:71 является определенной последовательностью-кДНК для V1-3692
SEQ ID NO:72 является определенной последовательностью-кДНК для 1А-3905
SEQ ID NO:73 является определенной последовательностью-кДНК для V1-3686
SEQ ID NO:74 является определенной последовательностью-кДНК для R1-2330
SEQ ID NO:75 является определенной последовательностью-кДНК для 1В-3976
SEQ ID NO:76 является определенной последовательностью-кДНК для V1-3679
SEQ ID NO:77 является определенной последовательностью-кДНК для 1G-4736
SEQ ID NO:78 является определенной последовательностью-кДНК для 1G-4738
SEQ ID NO:79 является определенной последовательностью-кДНК для 1G-4741
SEQ ID NO:80 является определенной последовательностью-кДНК для 1G-4744
SEQ ID NO:81 является определенной последовательностью-кДНК для 1G-4734
SEQ ID NO:82 является определенной последовательностью-кДНК для 1Н-4774
SEQ ID NO:83 является определенной последовательностью-кДНК для 1Н-4781
SEQ ID NO:84 является определенной последовательностью-кДНК для 1Н-4785
SEQ ID NO:85 является определенной последовательностью-кДНК для 1Н-4787
SEQ ID NO:86 является определенной последовательностью-кДНК для 1Н-4796
SEQ ID NO:87 является определенной последовательностью-кДНК для 1I-4807
SEQ ID NO:88 является определенной последовательностью-кДНК для 1I-4810
SEQ ID NO:89 является определенной последовательностью-кДНК для 1I-4811
SEQ ID NO:90 является определенной последовательностью-кДНК для 1J-4876
SEQ ID NO:91 является определенной последовательностью-кДНК для 1К-4884
SEQ ID NO:92 является определенной последовательностью-кДНК для 1К-4896
SEQ ID NO:93 является определенной последовательностью-кДНК для 1G-4761
SEQ ID NO:94 является определенной последовательностью-кДНК для 1G-4762
SEQ ID NO:95 является определенной последовательностыо-кДНК для 1Н-4766
SEQ ID NO:96 является определенной последовательностью-кДНК для 1Н-4770
SEQ ID NO:97 является определенной последовательностью-кДНК для 1Н-4771
SEQ ID NO:98 является определенной последовательностью-кДНК для 1Н-4772
SEQ ID NO:99 является определенной последовательностью-кДНК для ID-4297
SEQ ID NO:100 является определенной последовательностью-кДНК для ID-4309
SEQ ID NO:101 является определенной последовательностью-кДНК для ID.1-4278
SEQ ID NO:102 является определенной последовательностью-кДНК для ID-4288
SEQ ID NO:103 является определенной последовательностью-кДНК для ID-4283
SEQ ID NO:104 является определенной последовательностью-кДНК для ID-4304
SEQ ID NO:105 является определенной последовательностью-кДНК для ID-4296
SEQ ID NO:106 является определенной последовательностью-кДНК для ID-4280
SEQ ID NO:107 является определенной полноразмерной последовательностью-кДНК для F1-12 (также называемой p504S)
SEQ ID NO:108 является предсказанной аминокислотной последовательностью для F1-12
SEQ ID NO:109 является полноразмерной последовательностью-кДНК для J1-17
SEQ ID NO:110 является определенной полноразмерной последовательностью-кДНК для L1-12
SEQ ID NO:111 является определенной полноразмерной последовательностью-кДНК для N1-1862
SEQ ID NO:112 является предсказанной аминокислотной последовательностью для J1-17
SEQ ID NO:113 является предсказанной аминокислотной последовательностью для L1-12
SEQ ID NO:114 является предсказанной аминокислотной последовательностью для N1-1862
SEQ ID NO:115 является определенной последовательностью-кДНК для Р89
SEQ ID NO:116 является определенной последовательностью-кДНК для Р90
SEQ ID NO:117 является определенной последовательностью-кДНК для Р92
SEQ ID NO:118 является определенной последовательностью-кДНК для Р95
SEQ ID NO:119 является определенной последовательностью-кДНК для Р98
SEQ ID NO:120 является определенной последовательностью-кДНК для Р102
SEQ ID NO:121 является определенной последовательностью-кДНК для Р110
SEQ ID NO:122 является определенной последовательностью-кДНК для Р111
SEQ ID NO:123 является определенной последовательностью-кДНК для Р114
SEQ ID NO:124 является определенной последовательностью-кДНК для Р115
SEQ ID NO:125 является определенной последовательностью-кДНК для Р116
SEQ ID NO:126 является определенной последовательностью-кДНК для Р124
SEQ ID NO:127 является определенной последовательностью-кДНК для Р126
SEQ ID NO:128 является определенной последовательностью-кДНК для Р130
SEQ ID NO:129 является определенной последовательностью-кДНК для Р133
SEQ ID NO:130 является определенной последовательностью-кДНК для Р138
SEQ ID NO:131 является определенной последовательностью-кДНК для Р143
SEQ ID NO:132 является определенной последовательностыо-кДНК для Р151
SEQ ID NO:133 является определенной последовательностью-кДНК для Р156
SEQ ID NO:134 является определенной последовательностью-кДНК для Р157
SEQ ID NO:135 является определенной последовательностью-кДНК для Р166
SEQ ID NO:136 является определенной последовательностью-кДНК для Р176
SEQ ID NO:137 является определенной последовательностью-кДНК для Р178
SEQ ID NO:138 является определенной последовательностью-кДНК для Р179
SEQ ID NO:139 является определенной последовательностью-кДНК для Р185
SEQ ID NO:140 является определенной последовательностью-кДНК для Р192
SEQ ID NO:141 является определенной последовательностью-кДНК для Р201
SEQ ID NO:142 является определенной последовательностью-кДНК для Р204
SEQ ID NO:143 является определенной последовательностью-кДНК для Р208
SEQ ID NO:144 является определенной последовательностью-кДНК для Р211
SEQ ID NO:145 является определенной последовательностыо-кДНК для Р213
SEQ ID NO:146 является определенной последовательностью-кДНК для Р219
SEQ ID NO:147 является определенной последовательностью-кДНК для Р237
SEQ ID NO:148 является определенной последовательностыо-кДНК для Р239
SEQ ID NO:149 является определенной последовательностью-кДНК для Р248
SEQ ID NO:150 является определенной последовательностью-кДНК для Р251
SEQ ID NO:151 является определенной последовательностью-кДНК для Р255
SEQ ID NO:152 является определенной последовательностью-кДНК для Р256
SEQ ID NO:153 является определенной последовательностью-кДНК для Р259
SEQ ID NO:154 является определенной последовательностью-кДНК для Р260
SEQ ID NO:155 является определенной последовательностью-кДНК для Р263
SEQ ID NO:156 является определенной последовательностью-кДНК для Р264
SEQ ID NO:157 является определенной последовательностью-кДНК для Р266
SEQ ID NO:158 является определенной последовательностью-кДНК для Р270
SEQ ID NO:159 является определенной последовательностью-кДНК для Р272
SEQ ID NO:160 является определенной последовательностью-кДНК для Р278
SEQ ID NO:161 является определенной последовательностью-кДНК для Р105
SEQ ID NO:162 является определенной последовательностью-кДНК для Р107
SEQ ID NO:163 является определенной последовательностью-кДНК для Р137
SEQ ID NO:164 является определенной последовательностью-кДНК для Р194
SEQ ID NO:165 является определенной последовательностью-кДНК для Р195
SEQ ID NO:166 является определенной последовательностью-кДНК для Р196
SEQ ID NO:167 является определенной последовательностью-кДНК для Р220
SEQ ID NO:168 является определенной последовательностью-кДНК для Р234
SEQ ID NO:169 является определенной последовательностью-кДНК для Р235
SEQ ID NO:170 является определенной последовательностью-кДНК для Р243
SEQ ID NO:171 является определенной последовательностью-кДНК для P703P-DE1
SEQ ID NO:172 является предсказанной аминокислотной последовательностью для P703P-DE1
SEQ ID NO:173 является определенной последовательностью-кДНК для P703P-DE2
SEQ ID NO:174 является определенной последовательностью-кДНК для P703P-DE6
SEQ ID NO:175 является определенной последовательностью-кДНК для P703P-DE13
SEQ ID NO:176 является предсказанной аминокислотной последовательностью для P703P-DE13
SEQ ID NO:177 является определенной последовательностью-кДНК для P703P-DE14
SEQ ID NO:178 является предсказанной аминокислотной последовательностью для P703P-DE14
SEQ ID NO:179 является определенной удлиненной последовательностью-кДНК для 1G-4736
SEQ ID NO:180 является определенной удлиненной последовательностью-кДНК для 1G-4738
SEQ ID NO:181 является определенной удлиненной последовательностью-кДНК для 1G-4741
SEQ ID NO:182 является определенной удлиненной последовательностью-кДНК для 1G-4744
SEQ ID NO:183 является определенной удлиненной последовательностью-кДНК для 1G-4774
SEQ ID NO:184 является определенной удлиненной последовательностью-кДНК для 1G-4781
SEQ ID NO:185 является определенной удлиненной последовательностью-кДНК для 1G-4785
SEQ ID NO:186 является определенной удлиненной последовательностью-кДНК для 1G-4787
SEQ ID NO:187 является определенной удлиненной последовательностью-кДНК для 1Н-4796
SEQ ID NO:188 является определенной удлиненной последовательностью-кДНК для 11-4807
SEQ ID NO:189 является определенной 3’-последовательностью-кДНК для 11-4810
SEQ ID NO:190 является определенной 3’-последовательностью-кДНК для 11-4811
SEQ ID NO:191 является определенной удлиненной последовательностью-кДНК для 1J-4876
SEQ ID NO:192 является определенной удлиненной последовательностью-кДНК для 1К-4884
SEQ ID NO:193 является определенной удлиненной последовательностью-кДНК для 1К-4896
SEQ ID NO:194 является определенной удлиненной последовательностью-кДНК для 1G-4761
SEQ ID NO:195 является определенной удлиненной последовательностью-кДНК для 1G-4762
SEQ ID NO:196 является определенной удлиненной последовательностью-кДНК для 1Н-4766
SEQ ID NO:197 является определенной 3’-последовательностью-кДНК для 1Н-4770
SEQ ID NO:198 является определенной 3’-последовательностью-кДНК для 1Н-4771
SEQ ID NO:199 является определенной удлиненной последовательностью-кДНК для 1Н-4772
SEQ ID NO:200 является определенной удлиненной последовательностью-кДНК для ID-4309
SEQ ID NO:201 является определенной удлиненной последовательностью-кДНК для 1D.1-4278
SEQ ID NO:202 является определенной удлиненной последовательностью-кДНК для 1D-4288
SEQ ID NO:203 является определенной удлиненной последовательностью-кДНК для 1D-4283
SEQ ID NO:204 является определенной удлиненной последовательностью-кДНК для 1D-4304
SEQ ID NO:205 является определенной удлиненной последовательностью-кДНК для 1D-4296
SEQ ID NO:206 является определенной удлиненной последовательностью-кДНК для 1D-4280
SEQ ID NO:207 является определенной последовательностью-кДНК для 10-d8fwd
SEQ ID NO:208 является определенной последовательностыо-кДНК для 10-Н10cоn
SEQ ID NO:209 является определенной последовательностью-кДНК для 11-C8rev
SEQ ID NO:210 является определенной последовательностью-кДНК для 7.g6fwd
SEQ ID NO:211 является определенной последовательностью-кДНК для 7.g6rev
SEQ ID NO:212 является определенной последовательностью-кДНК для 8-b5fwd
SEQ ID NO:213 является определенной последовательностью-кДНК для 8-b5rev
SEQ ID NO:214 является определенной последовательностью-кДНК для 8-b6fwd
SEQ ID NO:215 является определенной последовательностью-кДНК для 8-b6rev
SEQ ID NO:216 является определенной последовательностью-кДНК для 8-d4fwd
SEQ ID NO:217 является определенной последовательностью-кДНК для 8-d9rev
SEQ ID NO:218 является определенной последовательностью-кДНК для 8-g3fwd
SEQ ID NO:219 является определенной последовательностью-кДНК для 8-g3rev
SEQ ID NO:220 является определенной последовательностью-кДНК для 8-h11rev
SEQ ID NO:221 является определенной последовательностью-кДНК для g-f12fwd
SEQ ID NO:222 является определенной последовательностью-кДНК для g-f3rev
SEQ ID NO:223 является определенной последовательностью-кДНК для P509S
SEQ ID NO:224 является определенной последовательностью-кДНК для P510S
SEQ ID NO:225 является определенной последовательностью-кДНК для P703DE5
SEQ ID NO:226 является определенной последовательностью-кДНК для 9-А11
SEQ ID NO:227 является определенной последовательностью-кДНК для 8-С6
SEQ ID NO:228 является определенной последовательностью-кДНК для 8-Р7
SEQ ID NO:229 является определенной последовательностью-кДНК для JPTPN13
SEQ ID NO:230 является определенной последовательностью-кДНК для JPTPN14
SEQ ID NO:231 является определенной последовательностью-кДНК для JPTPN23
SEQ ID NO:232 является определенной последовательностью-кДНК для JPTPN24
SEQ ID NO:233 является определенной последовательностью-кДНК для JPTPN25
SEQ ID NO:234 является определенной последовательностью-кДНК для JPTPN30
SEQ ID NO:235 является определенной последовательностью-кДНК для JPTPN34
SEQ ID NO:236 является определенной последовательностью-кДНК для PTPN35
SEQ ID NO:237 является определенной последовательностью-кДНК для JPTPN36
SEQ ID NO:238 является определенной последовательностью-кДНК для JPTPN38
SEQ ID NO:239 является определенной последовательностью-кДНК для JPTPN39
SEQ ID NO:240 является определенной последовательностью-кДНК для JPTPN40
SEQ ID NO:241 является определенной последовательностью-кДНК для JPTPN41
SEQ ID NO:242 является определенной последовательностью-кДНК для JPTPN42
SEQ ID NO:243 является определенной последовательностью-кДНК для JPTPN45
SEQ ID NO:244 является определенной последовательностью-кДНК для JPTPN46
SEQ ID NO:245 является определенной последовательностью-кДНК для JPTPN51
SEQ ID NO:246 является определенной последовательностью-кДНК для JPTPN56
SEQ ID NO:247 является определенной последовательностью-кДНК для PTPN64
SEQ ID NO:248 является определенной последовательностью-кДНК для JPTPN65
SEQ ID NO:249 является определенной последовательностью-кДНК для JPTPN67
SEQ ID NO:250 является определенной последовательностью-кДНК для JPTPN76
SEQ ID NO:251 является определенной последовательностью-кДНК для JPTPN84
SEQ ID NO:252 является определенной последовательностью-кДНК для JPTPN85
SEQ ID NO:253 является определенной последовательностью-кДНК для JPTPN86
SEQ ID NO:254 является определенной последовательностью-кДНК для JPTPN87
SEQ ID NO:255 является определенной последовательностью-кДНК для JPTPN88
SEQ ID NO:256 является определенной последовательностью-кДНК для JP1F1
SEQ ID NO:257 является определенной последовательностью-кДНК для JP1F2
SEQ ID NO:258 является определенной последовательностью-кДНК для JP1C2
SEQ ID NO:259 является определенной последовательностью-кДНК для JP1B1
SEQ ID NO:260 является определенной последовательностью-кДНК для JP1B2
SEQ ID NO:261 является определенной последовательностью-кДНК для JP1D3
SEQ ID NO:262 является определенной последовательностью-кДНК для JP1A4
SEQ ID NO:263 является определенной последовательностью-кДНК для JP1F5
SEQ ID NO:264 является определенной последовательностыо-кДНК для JP1E6
SEQ ID NO:265 является определенной последовательностью-кДНК для JP1D6
SEQ ID NO:266 является определенной последовательностью-кДНК для JP1B5
SEQ ID NO:267 является определенной последовательностью-кДНК для JP1A6
SEQ ID NO:268 является определенной последовательностью-кДНК для JP1E8
SEQ ID NO:269 является определенной последовательностью-кДНК для JP1D7
SEQ ID NO:270 является определенной последовательностью-кДНК для JP1D9
SEQ ID NO:271 является определенной последовательностью-кДНК для JP1C10
SEQ ID NO:272 является определенной последовательностью-кДНК для JP1A9
SEQ ID NO:273 является определенной последовательностыо-кДНК для JP1F12
SEQ ID NO:274 является определенной последовательностью-кДНК для JP1E12
SEQ ID NO:275 является определенной последовательностью-кДНК для JP1D11
SEQ ID NO:276 является определенной последовательностью-кДНК для JP1C11
SEQ ID NO:277 является определенной последовательностью-кДНК для JP1C12
SEQ ID NO:278 является определенной последовательностью-кДНК для JP1B12
SEQ ID NO:279 является определенной последовательностью-кДНК для JP1A12
SEQ ID NO:280 является определенной последовательностью-кДНК для JP8G2
SEQ ID NO:281 является определенной последовательностью-кДНК для JP8H1
SEQ ID NO:282 является определенной последовательностью-кДНК для JP8H2
SEQ ID NO:283 является определенной последовательностью-кДНК для JP8A3
SEQ ID NO:284 является определенной последовательностью-кДНК для JP8A4
SEQ ID NO:285 является определенной последовательностью-кДНК для JP8C3
SEQ ID NO:286 является определенной последовательностью-кДНК для JP8G4
SEQ ID NO:287 является определенной последовательностью-кДНК для JP8B6
SEQ ID NO:288 является определенной последовательностью-кДНК для JP8D6
SEQ ID NO:289 является определенной последовательностью-кДНК для JP8F5
SEQ ID NO:290 является определенной последовательностью-кДНК для JP8A8
SEQ ID NO:291 является определенной последовательностью-кДНК для JP8C7
SEQ ID NO:292 является определенной последовательностью-кДНК для JP8D7
SEQ ID NO:293 является определенной последовательностью-кДНК для P8D8
SEQ ID NO:294 является определенной последовательностью-кДНК для JP8E7
SEQ ID NO:295 является определенной последовательностью-кДНК для JP8F8
SEQ ID NO:296 является определенной последовательностью-кДНК для JP8G8
SEQ ID NO:297 является определенной последовательностью-кДНК для JP8B10
SEQ ID NO:298 является определенной последовательностью-кДНК для JP8C10
SEQ ID NO:299 является определенной последовательностью-кдНК для JP8E9
SEQ ID NO:300 является определенной последовательностью-кДНК для JP8E10
SEQ ID NO:301 является определенной последовательностью-кДНК для JP8F9
SEQ ID NO:302 является определенной последовательностью-кДНК для JP8H9
SEQ ID NO:303 является определенной последовательностью-кДНК для JP8C12
SEQ ID NO:304 является определенной последовательностью-кДНК для JP8E11
SEQ ID NO:305 является определенной последовательностью-кДНК для JP8E12
SEQ ID NO:306 является аминокислотной последовательностью для пептида PS2#12
SEQ ID NO:307 является определенной последовательностью-кДНК для Р711Р
SEQ ID NO:308 является определенной последовательностью-кДНК для Р712Р
SEQ ID NO:309 является определенной последовательностью-кДНК для СLONE 23
SEQ ID NO:310 является определенной последовательностью-кДНК для Р774Р
SEQ ID NO:311 является определенной последовательностью-кДНК для Р775Р
SEQ ID NO:312 является определенной последовательностью-кДНК для Р715Р
SEQ ID NO:313 является определенной последовательностью-кДНК для Р710Р
SEQ ID NO:314 является определенной последовательностью-кДНК для Р767Р
SEQ ID NO:315 является определенной последовательностью-кДНК для Р768Р
SEQ ID NO:316-325 являются определенными последовательностями-кДНК ранее выделенных генов
SEQ ID NO:326 является определенной последовательностью-кДНК для P703PDE5
SEQ ID NO:327 является предсказанной аминокислотной последовательностью для P703PDE5
SEQ ID NO:328 является определенной последовательностью-кДНК для Р703Р6.26
SEQ ID NO:329 является предсказанной аминокислотной последовательностью для Р703Р6.26
SEQ ID NO:330 является определенной последовательностью-кДНК для Р703РХ-23
SEQ ID NO:331 является предсказанной аминокислотной последовательностью для Р703РХ-23
SEQ ID NO:332 является определенной полноразмерной последовательностью-кДНК для P509S
SEQ ID NO:333 является определенной удлиненной последовательностью-кДНК для Р707Р (также называемой 11-С9)
SEQ ID NO:334 является определенной последовательностью-кДНК для Р714Р
SEQ ID NO:335 является определенной последовательностью-кДНК для Р705Р (также называемой 9-F3)
SEQ ID NO:336 является предсказанной аминокислотной последовательностью для Р705Р
SEQ ID NO:337 является аминокислотной последовательностью пептида Р1S#10
SEQ ID NO:338 является аминокислотной последовательностью пептида р5
SEQ ID NO:339 является аминокислотной последовательностью P509S
SEQ ID NO:340 является определенной последовательностью-кДНК для Р778Р
SEQ ID NO:341 является определенной последовательностью-кДНК для Р787Р
SEQ ID NO:342 является определенной последовательностью-кДНК для Р789Р
SEQ ID NO:343 является определенной последовательностью-кДНК для клона, обнаруживающего гомологию с мРНК ММ46 Homo sapiens
SEQ ID NO:344 является определенной последовательностью-кДНК для клона, обнаруживающего гомологию с мРНК белка АВС (АВС50), стимулируемой TNF-альфа Homo sapiens
SEQ ID NO:345 является определенной последовательностью-кДНК для клона, обнаруживающего гомологию с мРНК Е-кадхерина Homo sapiens
SEQ ID NO:346 является определенной последовательностью-кДНК для клона, обнаруживающего гомологию с ядерно-кодируемой митохондриальной серингидроксиметилтрансферазой (SHMT) человека
SEQ ID NO:347 является определенной последовательностью-кДНК для клона, обнаруживающего гомологию с природным связанным с устойчивостью белком 2 макрофагов (NRAMP2) Homo sapiens
SEQ ID NO:348 является определенной последовательностью-кДНК для клона, обнаруживающего гомологию с фосфоглюкомутаза-родственным белком (PGMRP) Homo sapiens
SEQ ID NO:349 является определенной последовательностью-кДНК для клона, обнаруживающего гомологию с мРНК для субъединицы протеосомы р40 человека
SEQ ID NO:350 является определенной последовательностью-кДНК для Р777Р
SEQ ID NO:351 является определенной последовательностью-кДНК для Р779Р
SEQ ID NO:352 является определенной последовательностью-кДНК для Р790Р
SEQ ID NO:353 является определенной последовательностью-кДНК для Р784Р
SEQ ID NO:354 является определенной последовательностью-кДНК для Р776Р
SEQ ID NO:355 является определенной последовательностью-кДНК для Р780Р
SEQ ID NO:356 является определенной последовательностью-кДНК для P544S
SEQ ID NO:357 является определенной последовательностью-кДНК для P745S
SEQ ID NO:358 является определенной последовательностью-кДНК для Р782Р
SEQ ID NO:359 является определенной последовательностью-кДНК для Р783Р
SEQ ID NO:360 является определенной последовательностью-кДНК для неизвестного 17984
SEQ ID NO:361 является определенной последовательностью-кДНК для Р787Р
SEQ ID NO:362 является определенной последовательностью-кДНК для Р788Р
SEQ ID NO:363 является определенной последовательностью-кДНК для неизвестного 17994
SEQ ID NO:364 является определенной последовательностью-кДНК для Р781Р
SEQ ID NO:365 является определенной последовательностью-кДНК для Р785Р
SEQ ID NO:366-375 являются определенными последовательностями-кДНК для сплайсинговых вариантов B305D
SEQ ID NO:376 является предсказанной аминокислотной последовательностью, кодируемой последовательностью SEQ ID NO:366
SEQ ID NO:377 является предсказанной аминокислотной последовательностью, кодируемой последовательностью SEQ ID NO:372
SEQ ID NO:378 является предсказанной аминокислотной последовательностью, кодируемой последовательностью SEQ ID NO:373
SEQ ID NO:379 является предсказанной аминокислотной последовательностью, кодируемой последовательностью SEQ ID NO:374
SEQ ID NO:380 является предсказанной аминокислотной последовательностью, кодируемой последовательностью SEQ ID NO:375
SEQ ID NO:381 является определенной последовательностью-кДНК для В716Р
SEQ ID NO:382 является определенной полноразмерной последовательностью-кДНК для Р711Р
SEQ ID NO:383 является предсказанной аминокислотной последовательностью для Р711Р
SEQ ID NO:384 является последовательностью-кДНК для Р1000С
SEQ ID NO:385 является последовательностью-кДНК для CGI-82
SEQ ID NO:386 является последовательностью-кДНК для 23320
SEQ ID NO:387 является последовательностью-кДНК для CGI-69
SEQ ID NO:388 является последовательностью-кДНК для 1-идитол-2-дегидрогеназы
SEQ ID NO:389 является последовательностью-кДНК для 23379
SEQ ID NO:390 является последовательность-кДНК для 23381
SEQ ID NO:391 является последовательностью-кДНК для KIAA0122
SEQ ID NO:392 является последовательность-кДНК для 23399
SEQ ID NO:393 является последовательностью-кДНК для ранее идентифицированного гена
SEQ ID NO:394 является последовательностью-кДНК для HCLBP
SEQ ID NO:395 является последовательностью-кДНК для транс-глутаминазы
SEQ ID NO:396 является последовательностью-кДНК для ранее идентифицированного гена
SEQ ID NO:397 является последовательностью-кДНК для РАР
SEQ ID NO:398 является последовательностью-кДНК для Ets фактора транскрипции PDEF
SEQ ID NO:399 является последовательностью-кДНК для hTGR
SEQ ID NO:400 является последовательностью-кДНК для KIAA0295
SEQ ID NO:401 является последовательностью-кДНК для 22545
SEQ ID NO:402 является последовательностью-кДНК для 22547
SEQ ID NO:403 является последовательностью-кДНК для 22548
SEQ ID NO:404 является последовательностью-кДНК для 22550
SEQ ID NO:405 является последовательностью-кДНК для 22551
SEQ ID NO:406 является последовательностью-кДНК для 22552
SEQ ID NO:407 является последовательностью-кДНК для 22553
SEQ ID NO:408 является последовательностью-кДНК для 22558
SEQ ID NO:409 является последовательностью-кДНК для 22562
SEQ ID NO:410 является последовательностью-кДНК для 22565
SEQ ID NO:411 является последовательностью-кДНК для 22567
SEQ ID NO:412 является последовательностью-кДНК для 22568
SEQ ID NO:413 является последовательностью-кДНК для 22570
SEQ ID NO:414 является последовательностью-кДНК для 22571
SEQ ID NO:415 является последовательностью-кДНК для 22572
SEQ ID NO:416 является последовательностью-кДНК для 22573
SEQ ID NO:417 является последовательностью-кДНК для 22574
SEQ ID NO:418 является последовательностью-кДНК для 22575
SEQ ID NO:419 является последовательностью-кДНК для 22580
SEQ ID NO:420 является последовательностью-кДНК для 22581
SEQ ID NO:421 является последовательностью-кДНК для 22582
SEQ ID NO:422 является последовательностью-кДНК для 22583
SEQ ID NO:423 является последовательностью-кДНК для 22584
SEQ ID NO:424 является последовательностью-кДНК для 22585
SEQ ID NO:425 является последовательностью-кДНК для 22586
SEQ ID NO:426 является последовательностью-кДНК для 22587
SEQ ID NO:427 является последовательностью-кДНК для 22588
SEQ ID NO:428 является последовательностью-кДНК для 22589
SEQ ID NO:429 является последовательностью-кДНК для 22590
SEQ ID NO:430 является последовательностью-кДНК для 22591
SEQ ID NO:431 является последовательностью-кДНК для 22592
SEQ ID NO:432 является последовательностью-кДНК для 22593
SEQ ID NO:433 является последовательностью-кДНК для 22594
SEQ ID NO:434 является последовательностью-кДНК для 22595
SEQ ID NO:435 является последовательностью-кДНК для 22596
SEQ ID NO:436 является последовательностью-кДНК для 22847
SEQ ID NO:437 является последовательностью-кДНК для 22848
SEQ ID NO:438 является последовательностью-кДНК для 22849
SEQ ID NO:439 является последовательностью-кДНК для 22851
SEQ ID NO:440 является последовательностью-кДНК для 22852
SEQ ID NO:441 является последовательностью-кДНК для 22853
SEQ ID NO:442 является последовательностью-кДНК для 22854
SEQ ID NO:443 является последовательностью-кДНК для 22855
SEQ ID NO:444 является последовательностью-кДНК для 22856
SEQ ID NO:445 является последовательностью-кДНК для 22857
SEQ ID NO:446 является последовательностью-кДНК для 23601
SEQ ID NO:447 является последовательностью-кДНК для 23602
SEQ ID NO:448 является последовательностью-кДНК для 23605
SEQ ID NO:449 является последовательностью-кДНК для 23606
SEQ ID NO:450 является последовательностью-кДНК для 23612
SEQ ID NO:451 является последовательностью-кДНК для 23614
SEQ ID NO:452 является последовательностью-кДНК для 23618
SEQ ID NO:453 является последовательностью-кДНК для 23622
SEQ ID NO:454 является последовательностью-кДНК для фолатгидролазы
SEQ ID NO:455 является последовательностью-кДНК для белка LIM
SEQ ID NO:456 является последовательностью-кДНК для известного гена
SEQ ID NO:457 является последовательностью-кДНК для известного гена
SEQ ID NO:458 является последовательностью-кДНК для ранее идентифицированного гена
SEQ ID NO:459 является последовательностью-кДНК для 23045
SEQ ID NO:460 является последовательностыо-кДНК для 23032
SEQ ID NO:461 является последовательностыо-кДНК для 23054
SEQ ID NO:462-467 являются последовательностями-кДНК для известных генов
SEQ ID NO:468-471 являются последовательностями-кДНК для Р710Р
SEQ ID NO:472 является последовательностью-кДНК для Р1001С.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Как отмечалось выше, данное изобретение относится, в общем, к композициям и способам для терапии и диагностики рака, такого как рак предстательной железы. Описанные здесь композиции могут включать в себя опухолевые полипептиды предстательной железы, полинуклеотиды, кодирующие такие полипептиды, связывающие агенты, такие как антитела, антиген-презентирующие клетки (АРС) и/или клетки иммунной системы (например, Т-клетки). Полипептиды данного изобретения обычно содержат, по меньшей мере, часть (такую как иммуногенная часть) опухолевого белка предстательной железы или его вариант. “Опухолевый белок предстательной железы” представляет собой белок, который экспрессируется в клетках опухоли предстательной железы на уровне, который является, по меньшей мере, в 2 раза более высоким и предпочтительно, по меньшей мере, в пять раз более высоким, чем уровень экспрессии в нормальной ткани, как определено с использованием репрезентативного анализа, обеспеченного здесь. Некоторые опухолевые белки предстательной железы являются опухолевыми белками, которые определяемо реагируют (в иммуноанализе, таком как ELISA или Вестерн-блот) с антисыворотками пациента, имеющего рак предстательной железы. Полинуклеотиды данного изобретения обычно содержат последовательность ДНК или РНК, которая кодирует весь такой полипептид или часть такого полипептида или которая является комплементарной такой последовательности. Антитела обычно являются белками иммунной системы или их антигенсвязывающими фрагментами, которые способны связываться с полипептидом, описанным выше. Антиген-презентирующие клетки включают в себя дендритные клетки, макрофаги, моноциты, фибробласты и В-клетки, экспрессирующие полипептид, описанный выше. Т-клетки, которые могут использоваться в таких композициях, обычно являются Т-клетками, которые являются специфическими в отношении полипептида, описанного выше.
Данное изобретение основано на открытии опухолевых белков предстательной железы. Последовательности полинуклеотидов, кодирующих некоторые опухолевые белки, или их части, обеспечены в последовательностях SEQ ID NO:1-111, 115-171, 173-175, 177, 179-305, 307-315, 326, 328, 330, 332-335, 340-375, 382 или 384-472. Последовательности полипептидов, содержащих, по меньшей мере, часть опухолевого белка, обеспечены в последовательностях SEQ ID NO:112-114, 172, 176, 178, 327, 329, 331, 336, 339, 376-380 и 383.
ПОЛИНУКЛЕОТИДЫ ОПУХОЛЕВОГО БЕЛКА ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ
Любой полинуклеотид, кодирующий опухолевый белок предстательной железы или его часть или вариант, описанный здесь, включен в данное изобретение. Предпочтительные полинуклеотиды содержат, по меньшей мере, 15 последовательных нуклеотидов, предпочтительно, по меньшей мере, 30 последовательных нуклеотидов и более предпочтительно, по меньшей мере, 45 последовательных нуклеотидов, которые кодируют часть опухолевого белка предстательной железы. Более предпочтительно, полинуклеотид кодирует иммуногенную часть опухолевого белка предстательной железы. Полинуклеотиды, комплементарные любой такой последовательности, также включены в данное изобретение. Полинуклеотиды могут быть одноцепочечными (кодирующими или антисмысловыми) или двухцепочечными и могут быть ДНК- (геномными, кДНК- или синтетическими) или РНК-молекулами. РНК-молекулы включают в себя молекулы гяRNА, которые содержат интроны и соответствуют ДНК-молекуле, как один к одному, и мРНК-молекулами, которые не содержат интронов. Дополнительные кодирующие или некодирующие последовательности могут, но необязательно, присутствовать в полинуклеотиде данного изобретения, и полинуклеотид может быть, но необязательно, связан с другими молекулами и/или веществами-носителями.
Полинуклеотиды могут содержать нативную последовательность (т.е. эндогенную последовательность, которая кодирует опухолевый белок предстательной железы или его часть), или могут содержать вариант такой последовательности. Полинуклеотидные варианты могут содержать одну или несколько замен, добавлений, делений и/или инсерций (вставок), так чтобы иммногенность кодируемого полипептида не уменьшалась относительно нативного опухолевого белка. Действие на иммуногенность кодируемого полипептида может обычно оцениваться, как описано здесь. Варианты предпочтительно обнаруживают приблизительно 70% идентичность, более предпочтительно, по меньшей мере, приблизительно 80% идентичность и наиболее предпочтительно, по меньшей мере, приблизительно 90% идентичность полинуклеотидной последовательности, кодирующей нативный опухолевый белок предстательной железы или его часть.
Говорят, что две полинуклеотидные или полипептидные последовательности являются “идентичными”, если последовательность нуклеотидов или аминокислот в этих двух последовательностях является одинаковой при сопоставлении для максимального соответствия, как описано ниже. Сравнения между двумя последовательностями обычно выполняют путем сравнения этих последовательностей на протяжении окна сравнения для идентификации и сравнения локальных районов сходства последовательностей. “Окно сравнения” в применении здесь относится к сегменту из, по меньшей мере, 20 смежных (непрерывных) положений, обычно от 30 до приблизительно 75, от 40 до приблизительно 50, в котором последовательность может сравниваться со сравниваемой последовательностью из такого же числа смежных (непрерывных) положений после оптимального выравнивания этих двух последовательностей.
Оптимальное выравнивание последовательностей для сравнения может быть выполнено с использованием программы Megalign в комплекте Lasergene программного обеспечения биоинформатики (DNASTAR, Inc. Madison, WI) с использованием параметров по умолчанию. Эта программа включает в себя несколько схем выравнивания, описанных в следующих ссылках: Dayhoff, М.О. (1978) A model of evolutionary change in proteins - Matrices for detecting distant relationships. В Dayhoff, M.O. (ed.) Atlas of Protein Sequences and Structure, National Biomedical Research Foundation, Washington DC Vol.5, Suppl.3, pp.345-358; Hein J. (1990) Unified Approach to Alignment and Phylogenes pp.626-645 Methods in Enzymology vol.183, Academic Press, Inc., San Diego, CA; Higgins, D.G. and Sharp, P.M. (1989) CABIOS 5:151-153; Myers, E.W. and Muller W. (1988) CABIOC 4:11-17; Robinson, E.D. (1971) Comb. Theor 11:105; Santou, N, Nes, M. (1987) Mol. Biol. Evol. 4:406-425; Sneath, P.H.A. and Sokal, R.R. (1973) Numerical Taxonomy - the Principles and Practice of Numerical Taxonomy. Freeman Press, San Francisco, CA; Wilbur, W.J. and Lipman, D.J. (1983) Proc. Natl. Acad. Sci. USA 80:726-730.
Предпочтительно “процент идентичности последовательности” определяют путем сравнения двух оптимально выравненных последовательностей над окном сравнения из, по меньшей мере, 20 положений, причем доля полинуклеотидной или полипептидной последовательности в окне сравнения может содержать добавления или делеции (т.е. гэпы (промежутки)), составляющие 20% или менее, обычно 5-15% или 10-12%, в сравнении со ссылочной последовательностью (которая не содержит добавлений или делеций) для оптимального выравнивания этих двух последовательностей. Указанный процент рассчитывают путем определения числа положений, при которых идентичные основания нуклеиновых кислот или аминокислотные остатки встречаются в обеих последовательностях, с получением числа совпадающих положений, деления числа совпадающих положений на общее число положений в ссылочной последовательности (например, с размером окна) и умножения этих результатов на 100 с получением процента идентичности последовательностей.
Варианты могут быть также или альтернативно по существу гомологичными нативному гену или его части или комплементу. Такие полинуклеотидные варианты способны гибридизоваться при умеренно жестких условиях с природно встречающейся последовательностью ДНК, кодирующей нативный опухолевый белок предстательной железы (или комплементарной последовательностью). Подходящие умеренно жесткие условия включают в себя предварительное промывание в растворе 5×SSC, 0,5% ДСН, 1,0 мМ ЭДТА (рН 8,0); гибридизацию при 50-65°С, 5×SSC, в течение ночи; с последующим промыванием дважды при 65°С в течение 20 минут каждым из 2х, 0,5х и 0,2х SSC, содержащим 0,1% ДСН.
Средним специалистам в данной области должно быть понятно, что вследствие вырожденности генетического кода имеется много нуклеотидных последовательностей, которые кодируют описанный здесь полипептид. Некоторые из этих полинуклеотидов имеют минимальную гомологию относительно любого нативного гена. Тем не менее полинуклеотиды, которые отличаются вследствие различий в использовании кодонов, особо рассматриваются данным изобретением. Далее, аллели генов, содержащие обеспеченные здесь полинуклеотидные последовательности, находятся в объеме данного изобретения. Аллели являются эндогенными генами, которые изменены в результате одной или нескольких мутаций, таких как делеции, добавления и/или замены нуклеотидов. Полученные мРНК и белок могут, но необязательно, иметь измененные структуру или функцию. Аллели могут быть идентифицированы с использованием стандартных способов (таких как гибридизация, амплификация и/или сравнение последовательностей баз данных).
Полинуклеотиды могут быть получены любым из различных способов. Например, полинуклеотид может быть идентифицирован, как описано более подробно ниже, посредством скрининга микроматрицы кДНК на связанную с опухолями экспрессию (т.е. экспрессию, которая является, по меньшей мере, в пять раз более высокой в опухоли предстательной железы, чем в нормальной ткани, как определено с помощью показательного анализа, описанного здесь). Такие скрининги могут быть выполнены с использованием микроматрицы Synteni (Palo Alto, CA) в соответствии с инструкциями изготовителя (и по существу, как описано Schena et al., Proc. Natl. Acad. Sci. USA 93:10614-10619, 1996 и Heller et al., Proc. Natl. Acad. Sci. USA 94:2150-2155, 1997). Альтернативно, полипептиды могут быть амплифицированы из кДНК, полученной из клеток, экспрессирующих описанные здесь белки, таких как опухолевые клетки предстательной железы. Такие полинуклеотиды могут быть амплифицированы с помощью полимеразной цепной реакции (ПЦР). Для указанного подхода могут быть сконструированы последовательность-специфические праймеры на основе обеспеченных здесь последовательностей или они могут быть куплены или синтезированы.
Амплифицированный белок может быть использован для выделения полноразмерного гена из подходящей библиотеки (например, библиотеки кДНК опухоли предстательной железы) с использованием хорошо известных способов. В таких способах библиотеку (кДНК или геномную) подвергают скринингу с использованием одного или нескольких полинуклеотидных зондов или праймеров, пригодных для амплификации. Предпочтительно библиотека является выбранной по размеру таким образом, чтобы она включала в себя более крупные молекулы. Случайно праймированные библиотеки могут быть предпочтительными для идентификации 5’-районов гена и районов гена, лежащих выше по ходу транскрипции. Геномные библиотеки являются предпочтительными для получения интронов и удлинения 5’-последовательностей.
Для способов гибридизации частичная последовательность может быть меченной (например, ник-трансляцией или концевым мечением 32Р) с использованием хорошо известных способов. Затем бактериальную или бактериофаговую библиотеку подвергают скринингу при помощи фильтров для гибридизации, содержащих денатурированные бактериальные колонии (или “газоны”, содержащие фаговые бляшки), с меченым зондом (Sambrook et al., Molecular Cloning; A Laboratory Manual, Cold Spring Harbor, NY, 1989). Гибридизующиеся колонии или бляшки отбирают и размножают и ДНК выделяют для последующего анализа.
Клоны кДНК могут быть анализированы для определения количества дополнительной последовательности, например, при помощи ПЦР с использованием праймера из частичной последовательности и праймера из вектора. Рестрикционные карты и частичные последовательности могут быть генерированы для идентификации одного или нескольких перекрывающихся клонов. Затем может быть определена полная последовательность с использованием стандартных способов, которые могут включать в себя генерирование ряда делеционных клонов. Затем полученные перекрывающиеся последовательности собирают в единую непрерывную последовательность. Полноразмерная кДНК-молекула может быть генерирована лигированием подходящих фрагментов с использованием хорошо известных способов.
Альтернативно, существуют многочисленные способы амплификации для получения полноразмерной кодирующей последовательности из частичной последовательности кДНК. В таких способах амплификацию обычно выполняют посредством ПЦР. Любой из многочисленных коммерчески доступных наборов может быть использован для выполнения стадии амплификации. Праймеры могут быть сконструированы с использованием, например, программного обеспечения, хорошо известного в данной области. Праймеры предпочтительно имеют 22-30 нуклеотидов в длину, имеют содержание GC, по меньшей мере, 50% и отжигаются с последовательностью-мишенью при температурах приблизительно 68-72°С. Амплифицированный район может быть секвенирован, как описано выше, и перекрывающиеся последовательности могут быть собраны в непрерывную последовательность.
Одним из таких способов амплификации является обратная полимеразная цепная реакция (Triglia et al., Nucl. Acids Res. 16:8186, 1988), в которой используют рестрикционные ферменты (рестриктазы) для генерирования фрагмента в известном районе гена. Затем указанный фрагмент замыкают в кольцо внутримолекулярным лигированием и используют в качестве матрицы для ПЦР с дивергентными праймерами, произведенными из указанного известного района. В альтернативном подходе последовательности, смежные с частичной последовательностью, могут быть исправлены амплификацией с праймером для линкерной последовательности и праймером, специфическим для известного района. Эти амплифицированные последовательности обычно подвергают второму циклу амплификации с тем же самым линкерным праймером и вторым праймером, специфическим для известного района. Вариация этой процедуры, которая использует два праймера, которые инициируют удлинение в противоположных направлениях от известной последовательности, описана в WO 96/38591. Другой подобный способ известен как “быстрая амплификация концов кДНК” или RACE. Указанный способ предусматривает использование внутреннего праймера и наружного праймера, который гибридизуется с полиА-районом или векторной последовательностью, для идентификации последовательностей, которые находятся 5’ и 3’ от известной последовательности. Дополнительные способы включают в себя “улавливающую” ПЦР (Lagerstrom et al., PCR Methods Applic. 1:111-19, 1991) и “прогулочную” ПЦК (Parker et al., Nucl. Acids Res. 19:3055-60, 1991). Другие способы с использованием амплификации также могут применяться для получения полноразмерной последовательности кДНК.
В некоторых случаях можно получить полноразмерную последовательность-кДНК анализом последовательностей, обеспечиваемых в базе данных меток экспрессируемых последовательностей (EST), такой как база данных, доступная из GenBank. Поиски на перекрывающиеся EST могут обычно проводиться с использованием хорошо известных программ (например, программы NCBI BLAST searches), и такие EST могут быть использованы для генерирования непрерывной полноразмерной последовательности.
Некоторые последовательности нуклеиновых кислот молекул кДНК, кодирующие, по меньшей мере, часть опухолевого белка предстательной железы, обеспечены в последовательностях SEQ ID NО:1-111, 115-171, 173-175, 177, 179-305, 307-315, 326, 328, 330, 332-335, 340-375, 381, 382 или 384-472. Выделение этих полинуклеотидов описано ниже. Каждый из этих опухолевых белков предстательной железы сверхэкспрессируется в опухолевой ткани предстательной железы.
Варианты полинуклеотидов могут быть обычно получены при помощи любого способа, известного в данной области, в том числе химического синтеза, например твердофазного фосфорамидитного химического синтеза. Модификации в полинуклеотидной последовательности могут быть также введены с использованием стандартных способов мутагенеза, таких как олигонуклеотид-направляемый сайт-специфический мутагенез (Adelman et al., DNA 2:183, (1983). Альтернативно, молекулы РНК могут быть получены in vitro- или in vivo-транскрипцией последовательностей ДНК, кодирующих опухолевый белок предстательной железы, или их части, при условии, что эта ДНК включена в вектор с подходящим промотором РНК-полимеразы (таким как Т7 или SP6). Некоторые части могут быть использованы для получения кодируемого полипептида, как описано здесь. Кроме того, или альтернативно, такая часть может быть введена пациенту таким образом, что кодируемый ею полипептид генерируется in vivo (например, трансфекцией антиген-презентирующих клеток, таких как дендритные клетки, конструкцией-кДНК, кодирующей опухолевый полипептид предстательной железы, и введением этих трансфицированных клеток пациенту).
Часть последовательности, комплементарной кодирующей последовательности (т.е. антисмысловой полинуклеотид), может быть также использована в качестве зонда или для модуляции экспрессии генов. Конструкции-кДНК, которые могут быть транскрибированы в антисмысловую РНК, также могут быть введены в клетки тканей для облегчения продуцирования антисмысловой РНК. Антисмысловой полинуклеотид может быть использован, как описано здесь, для подавления экспрессии опухолевого белка. Антисмысловая технология может быть использована для регуляции экспрессии генов через образование тройной спирали, которая нарушает способность двойной спирали достаточно открываться для связывания полимераз, транскрипционных факторов или регуляторных молекул (Gee et al., In Huber and Carr, Molecular and Iinmuno logic Approaches, Futura Publishing Co. (Mt. Kisco, NY; 1993)). Альтернативно, может быть сконструирована антисмысловая молекула для гибридизации с регуляторным районом гена (например, промотором, энхансером или сайтом инициации транскрипции) и блокирования транскрипции данного гена; или блокирования трансляции путем подавления связывания транскрипта с рибосомами.
Часть кодирующей последовательности или комплементарной последовательности может быть также сконструирована в виде зонда или праймера для определения экспрессии гена. Зонды могут быть помечены различными репортерными группами, такими как радионуклиды и ферменты, и предпочтительно имеют длину, по меньшей мере, 10 нуклеотидов, более предпочтительно, по меньшей мере, 20 нуклеотидов и еще более предпочтительно, по меньшей мере, 30 нуклеотидов. Праймеры, как отмечалось выше, имеют длину 22-30 нуклеотидов.
Любой полинуклеотид может быть дополнительно модифицирован для увеличения стабильности in vivo. Возможные модификации включают в себя, но не ограничиваются ими, добавление фланкирующих последовательностей при 5’- и/или 3’-концах; предпочтительное использование фосфоротиоатных или 2’-О-метильных, а не фосфодиэстеразных связей в молекулярном скелете и/или включение нетрадиционных оснований, таких как инозин, квеозин и вибутазин, а также ацетил-, метил-, тио- и других модифицированных форм аденина, цитидина, гуанина, тимина и уридина.
Нуклеотидные последовательности, описанные здесь, могут быть соединены с различными другими нуклеотидными последовательностями при помощи стандартных способов рекомбинантных ДНК. Например, полинуклеотид может быть клонирован в любой из многочисленных клонирующих векторов, в том числе в плазмиды, фагмиды, производные фага лямбда и космиды. Представляющие особый интерес векторы включают в себя экспрессирующие векторы, репликационные векторы, генерирующие зонд векторы и секвенирующие векторы. В общем, вектор будет содержать точку начала репликации, функциональное в, по меньшей мере, одном организме, подходящие сайты рестрикционных эндонуклеаз (рестриктаз) и один или более селектируемых маркеров. Другие элементы будут зависеть от желательного применения и будут очевидными средним специалистам в данной области.
В некоторых вариантах полинуклеотиды могут быть приготовлены таким образом, чтобы сделать возможными вхождение в клетку млекопитающего и экспрессию в ней. Такие композиции применимы, в частности, для терапевтических целей, как описано ниже. Специалистам в данной области будет понятно, что существует много способов достижения экспрессии полинуклеотида в клетке-мишени и может быть использован любой подходящий способ. Например, полинуклеотид может быть включен в вирусный вектор, такой как, но не только, аденоассоциированный вирус, ретровирус или вирус коровьей оспы или других поксвирусов (вирусов оспы) (например, птичьего поксвируса). Способы включения ДНК в такие векторы хорошо известны средним специалистам в данной области. Ретровирусный вектор может дополнительно переносить или включать ген для селектируемого маркера (для помощи в идентификации или селекции трансдуцированных клеток) и/или нацеливающую часть молекулы, такую как ген, кодирующий лиганд для рецептора на специфической клетке-мишени, для придания специфичности в отношении мишени указанному вектору. Нацеливание может выполняться с использованием антител способами, известными средним специалистам в данной области.
Другие композиции для терапевтических целей включают в себя коллоидальные диспергированные системы, такие как макромолекулярные комплексы, нанокапсулы, микросферы, гранулы и системы на основе липидов, в том числе эмульсии типа масло-в-воде, мицеллы, смешанные мицеллы и липосомы. Предпочтительной коллоидальной системой для применения в качестве вектора доставки in vitro и in vivo является липосома (т.е. искусственный мембранный пузырек). Получение и применение таких систем хорошо известны в данной области.
ОПУХОЛЕВЫЕ ПОЛИПЕПТИДЫ ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ
В контексте данного изобретения полипептиды могут содержать, по меньшей мере, иммуногенную часть опухолевого белка предстательной железы или его вариант, как описано здесь. Как отмечалось выше, "опухолевый белок предстательной железы" является белком, который экспрессируется опухолевыми клетками предстательной железы. Белки, которые являются опухолевыми белками предстательной железы, также взаимодействуют определяемо в иммуноанализе (таком как ELISA) с антисыворотками из пациента с раком предстательной железы. Описанные здесь полипептиды могут быть любой длины. Могут присутствовать дополнительные последовательности, произведенные из последовательностей нативного белка и/или гетерологичных последовательностей, и такие последовательности могут (но необязательно) обладать дополнительными иммуногенными или антигенными свойствами.
"Иммуногенная часть" в применении здесь является частью белка, которая распознается (т.е. специфически связывается) рецептором антигена поверхности В-клеток и/или Т-клеток.
Такие иммуногенные части обычно содержат, по меньшей мере, 5 аминокислотных остатков, более предпочтительно, по меньшей мере, 10, еще более предпочтительно, по меньшей мере, 20 аминокислотных остатков опухолевого белка предстательной железы или его варианта. Некоторые предпочтительные иммуногенные части включают в себя пептиды, в которых были делегированы N-концевая лидерная последовательность и/или трансмембранный домен. Другие предпочтительные иммуногенные части могут содержать небольшую N- или С-концевую делецию (например, 1-30 аминокислот, предпочтительно 5-15 аминокислот) относительно зрелого белка.
Иммуногенные части могут быть обычно идентифицированы с использованием хорошо известных способов, таких как суммированные в Paul, Fundamental Immunology, 3rd ed., 243-247 (Raven Press, 1993) и цитированных в этой работе ссылках. Такие способы включают в себя скрининг полипептидов на способность реагировать с антиген-специфическими антителами, антисыворотками и/или Т-клеточными линиями или клонами. В применении здесь антисыворотки и антитела являются “антиген-специфическими”, если они специфически связываются с антигеном (т.е. реагируют с белком в ELISA или другом иммуноанализе и не реагируют определяемо с посторонними белками). Такие антисыворотки и антитела могут быть получены, как описано здесь, и с использованием хорошо известных способов. Иммуногенная часть нативного опухолевого белка предстательной железы является частью, которая реагирует с такими антисыворотками и/или Т-клетками на уровне, который по существу не меньше, чем реактивность полноразмерного полипептида (например, в ELISA и/или анализе Т-клеточной реактивности). Такие иммуногенные части могут реагировать в таких анализах на уровне, который является одинаковым или более высоким, чем реактивность полноразмерного полипептида. Такие скрининги могут обычно проводиться с использованием способов, хорошо известных средним специалистам в данной области, таких как описанные в Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, 1988. Например, полипептид может быть иммобилизован на твердом носителе и приведен в контакт с сыворотками пациентов для возможности связывания антител в сыворотках с иммобилизованном полипептидом. Несвязанные сыворотки могут быть затем удалены, а связанные антитела могут быть определены с использованием, например, 125I-меченного белка А.
Как отмечалось выше, композиция может содержать вариант нативного опухолевого белка предстательной железы. “Вариант” полипептида в применении здесь является полипептидом, который отличается от нативного опухолевого белка предстательной железы одной или несколькими заменами, делениями, добавлениями и/или инсерциями (вставками), так что иммуногенность указанного полипептида по существу не уменьшается. Другими словами, способность варианта реагировать с антиген-специфическими антисыворотками может быть повышенной или неизмененной относительно нативного белка или может быть сниженной на менее чем 50%, и предпочтительно на менее чем 20% относительно нативного белка. Такие варианты могут быть идентифицированы модификацией одной из вышеуказанных полипептидных последовательностей и определением реактивности модифицированного полипептида с антиген-специфическими антителами или антисыворотками, как описано здесь. Предпочтительные варианты включают в себя варианты, в которых одна или несколько частей, таких как N-концевая лидерная последовательность или трансмембранный домен, были удалены. Другие предпочтительные варианты включают варианты, в которых небольшая часть (например, 1-30 аминокислот, предпочтительно 5-15 аминокислот) были удалены из N- и/или С-конца зрелого белка. Варианты полипептида предпочтительно обнаруживают, по меньшей мере, приблизительно 70%, более предпочтительно, по меньшей мере, приблизительно 90% и наиболее предпочтительно, по меньшей мере, приблизительно 95% идентичность (определяемую, как описано выше) относительно идентифицированных полипептидов.
Предпочтительно вариант содержит консервативные замены. “Консервативной заменой” является замена, при которой аминокислота заменена другой аминокислотой, которая имеет сходные свойства, так чтобы специалист с квалификацией в данной области мог ожидать, что вторичная структура и гидропатический характер данного полипептида по существу не будут измененными. Аминокислотные замены обычно могут производиться на основе сходства в полярности, заряде, растворимости, гидрофобности, гидрофильности и/или амфипатической природы остатков. Например, отрицательно заряженные аминокислоты включают в себя аспарагиновую кислоту и глутаминовую кислоту; положительно заряженные аминокислоты включают в себя лизин и аргинин; и аминокислоты с незаряженными полярными головными группами, имеющие одинаковые величины гидрофильности, включают в себя лейцин, изолейцин и валин, глицин и аланин, аспарагин и глутамин, серин, треонин, фенилаланин и тирозин. Другие группы аминокислот, которые могут представлять консервативные замены, включают в себя: (1) ala, pro, gly, glu, asp, gln, asn, ser, thr; (2) cys, ser, tyr, thr; (3) val, ile, leu, met, ala, phe; (4) lys, arg, his; (5) phe, tyr, thr, his. Вариант может также, или альтернативно содержать неконсервативные замены. В предпочтительном варианте полипептиды варианта отличаются от нативной последовательности заменой, делецией или добавлением пяти или меньшего количества аминокислот. Варианты могут также (или альтернативно) быть модифицированы, например, делецией или добавлением аминокислот, которые имеют минимальное влияние на иммуногенность, вторичную структуру и гидропатический характер полипептида.
Как отмечалось выше, полипептиды могут содержать сигнальную (или лидерную) последовательность на N-концевой стороне белка, которая котрансляционно или посттрансляционно направляет перенос указанного белка. Полипептид может быть также конъюгирован с линкерной или другой последовательностью для облегчения синтеза, очистки или идентификации указанного полипептида (например, поли-His) или для усиления связывания указанного полипептида с твердым носителем. Например, полипептид может быть конъюгирован с Fc-районом иммуноглобулина.
Полипептиды могут быть получены с использованием любого из разнообразных хорошо известных способов. Рекомбинантные полипептиды, кодируемые описанными выше последовательностями ДНК, могут быть легко получены из этих последовательностей ДНК с использованием различных экспрессирующих векторов, известных средним специалистам в данной области. Экспрессия может быть достигнута в любой подходящей клетке-хозяине, которая была трансформирована или трансфицирована экспрессирующим вектором, содержащим молекулу ДНК, кодирующую рекомбинантный полипептид. Подходящие клетки-хозяева включают в себя прокариоты, дрожжи и высшие эукариотические клетки. Предпочтительно используемыми клетками-хозяевами являются Е.coli, дрожжи или линия клеток млекопитающих, такая как COS или СНО. Супернатанты из подходящих систем хозяин-вектор, которые секретируют рекомбинантный белок или полипептид в культуральные среды, могут быть сначала сконцентрированы при помощи коммерчески доступного фильтра. После концентрирования концентрат может быть нанесен на подходящий очищающий матрикс, такой как аффинный матрикс или ионообменная смола. Наконец, для дополнительной очистки рекомбинантного полипептида могут быть использованы одна или несколько стадий обращеннофазовой ВЭЖХ.
Части или другие варианты, имеющие менее приблизительно 100 аминокислот и обычно менее приблизительно 50 аминокислот, могут быть получены синтетическими способами с использованием способов, хорошо известных средним специалистам в данной области. Например, такие полипептида могут быть синтезированы при помощи любого из коммерчески доступных твердофазных способов, таких как способ твердофазного синтеза Меррифилда, в котором аминокислоты последовательно добавляют к растущей аминокислотной цепи. См. Merrifield, J. Am. Chem. Soc. 85:2149-2146, 1963. Оборудование для автоматизированного синтеза полипептидов является коммерчески доступным от поставщиков, таких как Perkin Elmer/Applied BioSystems Division (Foster City, CA), и оно может эксплуатироваться в соответствии с инструкциями изготовителя.
В некоторых характерных вариантах полипептид может быть слитым белком, который содержит множественные полипептиды, описанные здесь, или который содержит, по меньшей мере, один полипептид, описанный здесь, и постороннюю последовательность, такую как известный опухолевый белок. Слитый партнер может, например, способствовать обеспечению эпитопов Т-клеток-хелперов (иммунологический партнер слияния), предпочтительно эпитопов Т-клеток-хелперов, узнаваемых человеком, или может способствовать экспрессии белка (усилитель экспрессии) с более высокими выходами, чем нативный рекомбинантный белок. Некоторые предпочтительные партнеры слияния являются как иммунологическими, так и усиливающими экспрессию партнерами слияния. Другие партнеры слияния могут быть выбраны таким образом, чтобы они увеличивали растворимость белка или позволяли белку быть нацеленным на желательные внутриклеточные компартменты. Следующие партнеры слияния включают в себя аффинные метки, которые облегчают очистку данного белка.
Слитые белки могут быть обычно получены при помощи стандартных способов, в том числе при помощи химической конъюгации. Предпочтительно слитый белок экспрессируется в виде рекомбинантного белка, позволяя получать увеличенные уровни относительно неслитого белка в системе экспрессии. Вкратце, последовательности-ДНК, кодирующие эти полипептидные компоненты, могут быть собраны отдельно и лигированы в подходящий экспрессирующий вектор. 3’-конец последовательности-ДНК, кодирующей один полипептидный компонент, лигируют с пептидным линкером или без него с 5’-концом последовательности-ДНК, кодирующей второй полипептидный компонент, так что рамки считывания этих последовательностей находятся в фазе. Это делает возможной трансляцию в единый слитый белок, который сохраняет биологическую активность обоих полипептидов-компонентов.
Пептидная линкерная последовательность может быть использована для разделения первого и второго полипептидного компонентов расстоянием, достаточным для гарантии того, что каждый полипептид укладывается в его вторичную и третичную структуры. Такую пептидную линкерную последовательность включают в слитый белок с использованием стандартных способов, хорошо известных в данной области. Подходящие пептидные линкерные последовательности могут быть выбраны на основании следующих факторов: (1) их способности принимать гибкую распрямляющуюся конформацию; (2) их неспособности принимать вторичную структуру, которая могла бы взаимодействовать с функциональными эпитопами на первом и втором полипептидах; и (3) отсутствия гидрофобных или заряженных остатков, которые могут взаимодействовать с функциональными эпитопами полипептида. Предпочтительные пептидные линкерные последовательности содержат остатки Gly, Asn и Ser. Другие почти нейтральные аминокислоты, такие как Thr и А1а, могут быть также использованы в линкерной последовательности. Аминокислотные последовательности, которые могут обычно использоваться в качестве линкеров, включают в себя последовательности, описанные в Maratea et al., Gene 40:39-46, 1985; Murphy et al., Proc. Natl. Acad. Sci. USA 83:8258-8262, 1986; патент Соединенных Штатов № 4935233 и патент Соединенных Штатов № 4751180. Линкерная последовательность обычно может иметь длину от 1 до приблизительно 50 аминокислот. Линкерные последовательности не требуются, когда первый и второй полипептиды имеют районы заменимых N-концевых аминокислот, которые могут быть использованы для отделения функциональных доменов и предотвращения стерических затруднений.
Лигированные последовательности-ДНК оперативно связаны с подходящими регуляторными элементами транскрипции или трансляции. Регуляторные элементы, ответственные за экспрессию ДНК, расположены только 5’ (слева) относительно последовательности-ДНК, кодирующей первый полипептид. Подобным образом стоп-кодоны, требующиеся для конечных сигналов терминации трансляции и транскрипции, присутствуют только 3’ (справа) относительно последовательности-ДНК, кодирующей второй полипептид.
Обеспечены также слитые белки, содержащие полипептид данного изобретения вместе с неродственным иммуногенным белком. Предпочтительно указанный иммуногенный белок способен вызывать вторичный иммунный ответ. Примеры таких белков включают в себя белки столбняка, туберкулеза и гепатита (например, Stoute et al., New Engl. J. Med., 336:86-91, 1997).
В предпочтительных вариантах иммунологический партнер слияния получают из белка D, поверхностного белка грамотрицательной бактерии Haemophilus influenza В (WO 91/18926). Предпочтительно производное белка D содержит приблизительно первую треть белка (например, первые 100-110 N-концевых аминокислот) и производное белка D может быть липидировано. В некоторых предпочтительных вариантах первые 109 остатков партнера слияния Липопротеина D включены на N-конце для обеспечения указанного полипептида дополнительными экзогенными Т-клеточными эпитопами и для увеличения уровня экспрессии в Е.coli (для функционирования, следовательно, в качестве усилителя экспрессии). Липидный хвост обеспечивает оптимальную презентацию (представление) антигена антиген-презентирующим клеткам. Другие партнеры слияния включают в себя неструктурный белок из вируса гриппа, NS1 (гемагглютинин). Обычно используют 81 N-концевую аминокислоту, хотя могут быть использованы различные фрагменты, включающие в себя эпитопы Т-клеток-хелперов.
В другом варианте иммунологическим партнером слияния является белок, известный как LYTA, или его часть (предпочтительно С-концевая часть). LYTA произведен из Streptococcus pneumoniae, который синтезирует N-ацетил-L-аланинамидазу, известную как амидаза LYTA (кодируемую геном LytA; Gene 43:265-292, 1980). LYTA является аутолизином, который специфически разрушает определенные связи в структуре пептидогликана. С-концевой домен белка LYTA является ответственным за аффинность в отношении холина или некоторых аналогов холина, таких как ДЭАЭ. Это свойство было использовано для разработки экспрессирующих C-LYTA Е.coli плазмид, применимых для экспрессии слитых белков. Очистка гибридных белков, содержащих фрагмент C-LYTA на амино-конце, была описана (Biotechnology 10:795-798, 1992). В предпочтительном варианте повторная часть LYTA может быть включена в слитый белок. Повторная часть обнаружена в С-концевом районе, и она начинается при остатке 178. Особенно предпочтительная повторная часть включает в себя остатки 188-305.
Обычно полипептиды (в том числе слитые белки) и полинуклеотиды, описанные здесь, выделяют. “Выделенным” полипептидом или полинуклетидом является полипептид или полинуклеотид, который удален из его исходного окружения. Например, природно встречающийся белок является выделенным, если он отделен от некоторых или всех веществ, сосуществующих с ним в природной системе. Предпочтительно такие полипептиды являются чистыми, по меньшей мере, приблизительно на 90%, более предпочтительно, по меньшей мере, приблизительно на 95% и наиболее предпочтительно, по меньшей мере, приблизительно на 99%. Полипептид считается выделенным, если, например, он клонирован в вектор, который не является частью природного окружения.
СВЯЗЫВАЮЩИЕ АГЕНТЫ
Данное изобретение обеспечивает, далее, агенты, такие как антитела и их антигенсвязывающие фрагменты, которые специфически связываются с опухолевым белком предстательной железы. В применении здесь антитело или его антигенсвязывающий фрагмент считают "специфически связанным" с опухолевым белком предстательной железы, если они реагируют при определяемом уровне (например, в анализе ELISA) с опухолевым белком предстательной железы и не реагируют определяемо с неродственными белками при тех же самых условиях. В применении здесь "связыванием" называют нековалентную связь между двумя отдельными молекулами с образованием комплекса. Способность связывания может быть оценена, например, путем определения константы связывания для образования указанного комплекса. Константа связывания является величиной, получаемой при делении концентрации комплекса на произведение концентраций компонентов. Обычно говорят, что два соединения "связываются", в контексте данного изобретения, когда константа связывания для образования комплекса превышает приблизительно 103 л/моль. Константа связывания может быть определена с использованием способов, хорошо известных в данной области.
Связывающие агенты могут быть дополнительно способными к разделению между пациентами, страдающими или не страдающими раковыми опухолями, такими как рак предстательной железы, при помощи показательных анализов, описанных здесь. Другими словами, антитела или другие связывающие агенты, которые связываются с опухолевым белком предстательной железы, будут генерировать сигнал, указывающий на наличие раковой опухоли, по меньшей мере, приблизительно у 20% пациентов с указанным заболеванием и будут генерировать отрицательный сигнал, указывающий на отсутствие указанного заболевания, по меньшей мере, приблизительно у 90% индивидуумов, не страдающих раковой опухолью. Для определения, удовлетворяет ли связывающий агент указанному требованию, биологические образцы (например, кровь, сыворотки, моча и/или опухолевые биоптаты) пациентов, страдающих и не страдающих раковой опухолью (что определяют с использованием стандартных клинических тестов), могут быть подвергнуты анализу, как описано здесь, на присутствие полипептидов, которые связываются с указанным связывающим агентом. Должно быть понятно, что должно быть подвергнуто анализу статистически значимое число образцов с заболеванием и без заболевания. Каждый связывающий агент должен удовлетворять вышеуказанным критериям, однако среднему специалисту в данной области будет понятно, что связывающие агенты могут быть использованы в комбинации для улучшения чувствительности.
Любой агент, который удовлетворяет вышеуказанным требованиям, может быть связывающим агентом. Например, связывающим агентом может быть рибосома с пептидным компонентом или без пептидного компонента, молекула РНК или полипептид. В предпочтительном варианте связывающий агент представляет собой антитело или его антигенсвязывающий фрагмент. Антитела могут быть получены любым из различных способов, известных средним специалистам в данной области, например, Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, 1988. Обычно антитела могут быть получены способами культуры клеток, в том числе генерированием моноклональных антител, как описано здесь, или через трансфекцию генов антител в подходящие клетки-хозяева бактерий или млекопитающих для продуцирования рекомбинантных антител. В одном способе иммуноген, содержащий данный полипептид, сначала инъецируют в любое млекопитающее из большого ряда (например, мышь, крыса, кролик, овца или коза). В этой стадии полипептиды данного изобретения могут служить в качестве иммуногена без модификации. Альтернативно, в частности, для относительно коротких полипептидов, превосходный иммунный ответ может быть вызван, если полипептид соединен с белком-носителем, таким как бычий сывороточный альбумин или гемоцианин фиссуреллы. Иммуноген инъецируют в животное-хозяин, предпочтительно в соответствии с заданным режимом, включающим в себя одну или несколько бустер-иммунизаций, и периодически из животных берут кровь. Поликлональные антитела, специфические для указанного полипептида, могут быть затем очищены из таких антисывороток, например, аффинной хроматографией с использованием полипептида, связанного с подходящим твердым носителем.
Моноклональные антитела, специфические для представляющего интерес антигенного полипептида, могут быть получены, например, с использованием способа Kohler and Milstein, Eur. J. Immunol. 6:511-519, 1976, и его усовершенствованных вариантов. Вкратце, эти способы предусматривают получение иммортализованных клеточных линий, способных продуцировать антитела, имеющие желаемую специфичность (т.е. реактивность с представляющим интерес полипептидом). Такие клеточные линии могут быть получены, например, из клеток селезенки, полученных из животного, иммунизированного, как описано выше. Затем клетки селезенки иммортализуют, например, слиянием с миеломной клеткой в качестве партнера слияния, предпочтительно клеткой, которая является сингенной с иммунизированным животным. Могут использоваться различные способы слияния. Например, клетки селезенки и миеломные клетки могут быть объединены с неионогенным детергентом на несколько минут, и затем производят посев при низкой плотности на селективную среду, которая поддерживает рост гибридных клеток, но не миеломных клеток. В предпочтительном способе отбора используют отбор с HAT (гипоксантином, аминоптерином, тимидином). После достаточного периода времени, обычно приблизительно 1-2 недель, наблюдают колонии гибридов. Отбирают одиночные колонии и их культуральные супернатанты испытывают на связывающую активность в отношении полипептида. Предпочтительными являются гибридомы, имеющие высокую реактивность и высокую специфичность.
Моноклональные антитела могут быть выделены из супернатантов растущих гибридомных колоний. Кроме того, различные способы могут быть использованы для повышения выхода, такие как инъекция гибридомной клеточной линии в перитонеальную полость подходящего позвоночного хозяина, такого как мышь. Затем моноклональные антитела могут быть собраны из асцитной жидкости или крови. Примеси могут быть удалены из антител общепринятыми способами, такими как хроматография, гель-фильтрация, осаждение и экстракция. Полипептиды данного изобретения могут быть использованы в процессе очистки, например, в стадии аффинной хроматографии.
В некоторых вариантах может быть предпочтительным использование антигенсвязывающих фрагментов антител. Такие фрагменты включают в себя Fab-фрагменты, которые могут быть получены с использованием стандартных способов. Вкратце, иммуноглобулины могут быть очищены из кроличьей сыворотки аффинной хроматографией на колонках с гранулами белка А (Наrlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, 1988) и гидролизованы папаином с образованием Fab- и Fс-фрагментов. Fab- и Fc-фрагменты могут быть разделены аффинной хроматографией на колонках с гранулами белка А.
Моноклональные антитела данного изобретения могут быть связаны с одним или несколькими терапевтическими агентами. Подходящие для указанного агенты включают в себя радионуклиды, индукторы дифференцировки, лекарственные средства, токсины и их производные. Предпочтительные радионуклиды включают в себя 90Y, 123I, 125I, 131I, 188Re, 211At и 212Bi. Предпочтительные лекарственные средства включают в себя метотрексат и аналоги пиримидина и пурина. Предпочтительные индукторы дифференцировки включают в себя форболовые эфиры и масляную кислоту. Предпочтительные токсины включают в себя рицин, абрин, дифтерийный токсин, холерный токсин, гелонин, экзотоксин Pseudomonas, токсин Shigella и антивирусный белок фитолакки американской.
Терапевтический агент может быть связан (например, ковалентно связан) с подходящим моноклональным антителом либо непосредственно, либо опосредованно (например, через линкерную группу). Прямая реакция между агентом и антителом возможна, когда агент и антитело обладают заместителями, способными реагировать друг с другом. Например, нуклеофильная группа, такая как амино или сульфгидрильная группа, на одном из них может быть способна реагировать с карбонилсодержащей группой, такой как ангидрид или галогенангидрид карбоновой кислоты, или с алкильной группой, содержащей подходящую уходящую группу (например, галогенид) на другом.
Альтернативно, может быть желательным связывать терапевтический агент и антитело посредством линкерной группы. Линкерная группа может функционировать в качестве спейсера для помещения антитела на расстоянии от агента во избежание помех в отношении связывающих способностей. Линкерная группа может служить также для увеличения химической активности заместителя на агенте или антителе и, следовательно, увеличения эффективности связывания. Увеличение химической активности может также облегчать использование агентов или функциональных групп на агентах, которое в противном случае не было бы возможным.
Специалистам в данной области будет понятно, что различные бифункциональные или полифункциональные реагенты, как гомо-, так и гетерофункциональные (такие как описанные в каталоге Pierce Chemical Co., Rockford, IL), могут быть использованы в качестве линкерной группы. Связывание может выполняться, например, через аминогруппы, карбоксильные группы, сульгидрильные группы или окисленные углеводные остатки. Имеются многочисленные ссылки, описывающие такую методологию, например патент Соединенных Штатов № 4671958, выданный Rodwell et al.
Если терапевтический агент является более сильнодействующим, когда он не содержит являющейся антителом части иммуноконъюгата данного изобретения, может быть желательным использование линкерной группы, расщепляемой во время или после интернализации в клетку. Был описан ряд различных расщепляемых линкерных групп. Механизмы внутриклеточного высвобождения агента от этих линкерных групп включают в себя расщепление восстановлением дисульфидной связи (например, патент Соединенных Штатов № 4489710, выданный Spitler), облучением фотолабильной связи (например, патент Соединенных Штатов, выданный Senter et al.), гидролизом дериватизованных аминокислотных боковых цепей (например, патент Соединенных Штатов № 4638045, выданный Kohn et al.), опосредованным сывороточным комплементом гидролизом (например, патент Соединенных Штатов № 4671958, выданный Rodwell et al.) и катализируемым кислотой гидролизом (например, патент Соединенных Штатов № 4569789, выданный Blattler et al.).
Может быть желательным связывание более чем одного агента с антителом. В одном варианте множественные молекулы терапевтического агента связывают с одной молекулой антитела. В другом варианте более чем один тип агента может быть связан с одним антителом. Независимо от конкретного варианта иммуноконъюгаты с более чем одним агентом могут быть приготовлены различными способами. Например, более чем один агент может быть связан непосредственно с молекулой антитела или могут быть использованы линкеры, которые обеспечивают множественные сайты для присоединения. Альтернативно, может быть использован носитель. Носитель может нести эти агенты различными способами, в том числе посредством ковалентного связывания либо непосредственно, либо через линкер. Подходящие носители включают в себя белки, такие как альбумины (например, патент Соединенных Штатов № 4507234, выданный Kato et al.), пептиды и полисахариды, такие как аминодекстран (например, патент Соединенных Штатов № 4699784, выданный Shih et al.). Носитель может также нести агент посредством нековалентного связывания или посредством инкапсулирования, такого как в пузырьке липосомы (например, патенты Соединенных Штатов № 4429088 и 4873088). Носители, специфические для радионуклидных агентов, включают в себя радиогалогенированные небольшие молекулы и хелатирующие соединения. Например, патент Соединенных Штатов № 4735792 описывает показательные радиогалогенированные небольшие молекулы и их синтез. Содержащий радионуклид хелат может быть образован из хелатирующих соединений, которые включают в себя соединения, содержащие атомы азота и серы в качестве донорных атомов для связывания металла или оксида металла, радионуклида. Например, патент Соединенных Штатов № 4673562, выданный Davison et al., описывает показательные хелатирующие соединения и их синтез.
Могут быть использованы разнообразные способы введения для антител и иммуноконъюгатов. Обычно введение является внутривенным, внутримышечным, подкожным или в место, где находилась иссеченная опухоль. Понятно, что точная доза антитела/иммуноконъюгата будет зависеть от используемого антитела, антигенной плотности на опухоли и скорости клиренса данного антитела.
Т-клетки
Иммунотерапевтические композиции могут также или альтернативно содержать Т-клетки, специфические для опухолевого белка предстательной железы. Такие клетки могут быть обычно получены in vitro или ex vivo с использованием стандартных процедур. Например, Т-клетки могут быть выделены из костного мозга, периферической крови или фракции костного мозга или периферической крови пациента с использованием коммерчески доступной системы разделения клеток, такой как система CEPRATE™, доступная из CellPro Inc., Bothell WA (патент Соединенных Штатов № 5240856; патент Соединенных Штатов № 5215926; WO 89/06280; WO 91/16116 и WO 92/07243). Альтернативно, Т-клетки могут быть получены из родственников пациента или не являющихся родственниками людей, других млекопитающих, клеточных линий или культур.
Т-клетки могут быть стимулированы опухолевым полипептидом предстательной железы, полинуклеотидом, кодирующим опухолевый полипептид предстательной железы, и/или антиген-презентирующей клеткой (АРС), экспрессирующей такой полипептид. Такую стимуляцию выполняют в условиях и в течение времени, достаточных для возможности генерирования Т-клеток, специфических для указанного полипептида. Предпочтительно опухолевый полипептид или полинуклеотид предстательной железы присутствует в доставляющем носителе, таком как микросфера, для облегчения генерирования специфических Т-клеток.
Т-клетки считаются специфическими для опухолевого полипептида предстательной железы, если эти Т-клетки убивают клетки-мишени, содержащие на своей поверхности полипептид или экспрессирующие ген, кодирующий указанный полипептид. Специфичность Т-клеток может оцениваться с использованием любого из разнообразных стандартных способов. Например, в анализе высвобождения хрома или в анализе пролиферации индекс стимуляции более чем двукратного увеличения лизиса и/или пролиферации в сравнении с отрицательными контролями свидетельствует о специфичности Т-клеток. Такие анализы могут проводиться, например, как описано в Chen et al., Cancer Res. 54:1065-1070, 1994. Альтернативно, определение пролиферации Т-клеток может выполняться различными известными способами. Например, пролиферацию Т-клеток можно определять путем измерения увеличенной скорости синтеза ДНК (например, путем кратковременного (импульсного) мечения культур Т-клеток тритированным тимидином и измерения количества тритированного тимидина, включенного в ДНК). Контакт с опухолевым полипептидом предстательной железы (100 нг/мл - 100 мкг/мл, предпочтительно 200 нг/мл - 25 мкг/мл) в течение 3-7 дней должен приводить к по меньшей мере двукратному увеличению пролиферации Т-клеток. Описанный выше контакт в течение 2-3 часов должен приводить к активации Т-клеток, измеряемой с использованием стандартных анализов на цитокины, в которых двукратное увеличение уровня высвобождения цитокина (например, TNF или IFN-γ) является указанием на активацию Т-клеток (Coligan et al., Current Protocols in Immunology, vol.1, Wiley Interscience (Greent 1998). Т-клетки, которые активировались в ответ на опухолевый полипептид, полинуклеотид предстательной железы или экспрессирующие полипептид АРС, могут быть CD4+ и/или CD8+. Т-клетки, специфические для опухолевого белка предстательной железы, могут быть размножены с использованием стандартных способов. В предпочтительных вариантах Т-клетки получают либо из пациента, либо из родственного или неродственного донора и вводят пациенту после стимуляции и размножения.
Для терапевтических целей CD4+ и/или CD8+ Т-клетки, которые пролиферируют в ответ на опухолевый полипептид, полинуклеотид предстательной железы или экспрессирующие указанный полипептид АРС, могут быть размножены до большего числа клеток in vitro или in vivo. Пролиферацию таких Т-клеток in vitro можно проводить различными путями. Например, эти Т-клетки могут быть повторно экспонированы опухолевому полипептиду предстательной железы или короткому пептиду, соответствующему иммуногенной части такого полипептида, с добавлением или без добавления факторов роста Т-клеток, таких как интерлейкин-2, и/или стимулирующих клеток, которые синтезируют опухолевый полипептид предстательной железы. Альтернативно, одна или несколько Т-клеток, которые пролиферируют в присутствии опухолевого белка предстательной железы, могут быть увеличены в числе клеток посредством клонирования. Способы клонирования клеток хорошо известны в данной области и включают в себя лимитирующее разбавление.
ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ И ВАКЦИНЫ
В некоторых аспектах описанные здесь полипептиды, полинуклеотиды, Т-клетки и/или связывающие агенты могут быть включены в фармацевтические композиции или иммуногенные композиции (т.е. вакцины). Фармацевтические композиции содержат одно или несколько таких соединений и физиологически приемлемый носитель. Вакцины могут содержать одно или несколько таких соединений и неспецифический усилитель иммунного ответа. Неспецифические усилители иммунного ответа включают в себя адъюванты, биодеградируемые микросферы (например, галактид полимолочной кислоты) и липосомы (в которые включено данное соединение; например, Fullerton, патент Соединенных Штатов № 4235877). Получение вакцин описано в общем, например, в M.F.Powell and M.J.Newman, eds., "Vaccine Design (the subunit and adjuvant approach)", Plenum Press (NY, 1995). Фармацевтические композиции и вакцины, находящиеся в объеме данного изобретения, могут также содержать другие соединения, которые могут быть биологически активными или неактивными. Например, одна или несколько иммуногенных частей других опухолевых антигенов могут присутствовать либо включенными в слитый полипептид, либо в виде отдельного соединения в этой композиции или вакцине.
Фармацевтическая композиция или вакцина может содержать ДНК, кодирующую один или несколько вышеописанных полипептидов, так что указанный полипептид генерируется in situ. Как отмечалось выше, эта ДНК может присутствовать в любой из различных систем доставки, известных средним специалистам в данной области, в том числе системах экспрессии нуклеиновых кислот, бактериальных и вирусных системах экспрессии. Многочисленные способы доставки генов хорошо известны в данной области, такие как описанные Rolland, Crit. Rev. Therap. Drug Carrier Systems 15:143-198, 1998 и цитированные в этой работе в ссылках. Подходящие системы экспрессии нуклеиновых кислот содержат необходимые последовательности-ДНК для экспрессии в пациенте (такие как подходящие промотор и сигнал терминации). Бактериальные системы доставки предусматривают введение бактерии (такой как Bacillus-Calmette-Guerrin), которая экспрессирует иммуногенную часть данного полипептида на ее клеточной поверхности или секретирует такой эпитоп. В предпочтительном варианте ДНК может быть введена с использованием вирусной экспрессионной системы (например, вируса коровьей оспы или другого поксвируса, ретровируса или аденовируса), которая может предусматривать применение непатогенного (дефектного) репликационно-компетентного вируса. Подходящие системы описаны, например, в Fisher-Hoch et al., Proc. Natl. Acad. Sci. USA 86:317-321; Flexner et al., Ann. N.Y.Acad. Sci. 569:86-103, 1989; Flexner et al., Vaccine 8:17-21, 1990; патент Соединенных Штатов № 4603112 и патент Соединенных Штатов № 5017487; WO 89/01973; патент Соединенных Штатов № 4777127; GB 2200651; ЕР 0345242; WO 91/02805; Berkner, Biotechniques 6:627, 1988; Rosenfeld et al., Science 252;431-434, 1991; Kolls et al., Proc. Natl. Acad. Sci. USA 91:215-219, 1994; Kass-Eisler et al., Proc. Natl. Acad. Sci. USA 90:11498-11502, 1993; Guzman et al., Circulation 88:2838-2848, 1993 и Guzman et al., Cir. Res. 73:1202-1207, 1993. Способы включения ДНК в такие экспрессионные системы хорошо известны средним специалистам в данной области. ДНК может быть также “оголенной” (депротеинизированной), как описано, например, в Ulmer et al., Science 259:1745-1749, 1993 и в обзоре Cohen, Science 259:1691-1692, 1993. Поглощение депротеинизированной ДНК может быть увеличено нанесением этой ДНК в виде покрытия на биодеградируемые гранулы, которые эффективно транспортируются в клетки.
Хотя любой подходящий носитель, известный средним специалистам в данной области, может быть использован в фармацевтических композициях данного изобретения, тип носителя будет меняться в зависимости от способа введения. Композиции данного изобретения могут быть приготовлены для любого подходящего способа введения, в том числе, например, местного, перорального, назального, внутривенного, внутричерепного, внутрибрюшинного, подкожного или внутримышечного введения. Для парентерального введения, такого как подкожная инъекция, носитель предпочтительно содержит воду, солевой раствор, спирт, жир, воск или буфер. Для перорального введения может быть использован любой из вышеуказанных носителей или твердый носитель, такой как маннит, лактоза, крахмал, стеарат магния, натрий-сахарин, тальк, целлюлоза, глюкоза, сахароза и карбонат магния. Биодеградируемые микросферы (например, полилактат-полигликолат) могут быть также использованы в качестве носителей для фармацевтических композиций данного изобретения. Подходящие биодеградируемые микросферы описаны, например, в патентах Соединенных Штатов № 4897268 и 5075109.
Такие композиции могут также содержать буферы (например, нейтрально забуференный солевой раствор или забуференный фосфатом солевой раствор), углеводы (например, глюкозу, маннозу, сахарозу или декстраны), маннит, белки, полипептиды или аминокислоты, такие как глицин, антиоксиданты, хелатообразователи, такие как ЭДТА или глутатион, адъюванты (например, гидроксид алюминия) и/или консерванты. Альтернативно, композиции данного изобретения могут быть приготовлены в виде лиофилизата. Соединения могут быть также инкапсулированы в липосомах с использованием хорошо известной технологии.
Любой из разнообразных неспецифических усилителей иммунного ответа может быть использован в вакцинах данного изобретения. Например, может быть включен адъювант. Большинство адъювантов содержат вещество, предназначенное для защиты антигена от быстрого катаболизма, такое как гидроксид алюминия или минеральное масло, и стимулятор иммунных ответов, такой как липид А, белки, полученные из Bortadella pertussis или Mycobacterium tuberculosis. Подходящие адъюванты является коммерчески доступными, например, неполный адъювант Фрейнда и полный адъювант Фрейнда (Difco Laboratories, Detroit, MI); адъювант 65 Merck (Merck and Company, Inc., Rahway, NJ); соли алюминия, такие как гель гидроксида алюминия (квасцы) или фосфат алюминия; соли кальция, железа или цинка; нерастворимая суспензия ацилированного тирозина; ацилированные сахара; катионно или анионно дериватизованные полисахариды; полифосфазены; биодеградируемые микросферы; монофосфориллипид А и квил А. Цитокины, такие как GM-CSF или интерлейкин-2, -7 или -12, могут быть также использованы в качестве адъювантов.
В рассматриваемых здесь вакцинах адъювантная композиция предпочтительно предназначена для индукции иммунного ответа типа Th1. Высокие уровни цитокинов Th1-типа (например, IFN-γ, IL-2 и IL-12) имеют тенденцию благоприятствовать индукции клеточно-опосредованных иммунных ответов на введенный антиген. В противоположность этому высокие уровни цитокинов Тh2-типа (например, IL-4, IL-5, IL-6, IL-10 и TNF-β) имеют тенденцию благоприятствовать индукции гуморальных иммунных ответов. После применения рассматриваемой здесь вакцины в пациенте будет поддерживаться иммунная реакция, которая включает в себя ответы Th1- и Тh2-типа. В предпочтительном варианте, в котором иммунный ответ является преимущественно ответом Th1-типа, уровень цитокинов Th1-типа будет увеличиваться в большей степени, чем уровень цитокинов Тh2-типа. Уровни этих цитокинов могут быть легко определены с использованием стандартных анализов. В отношении обзора семейств цитокинов см. Mosmann and Coffman, Ann. Rev. Immunol. 7:145-173, 1989.
Предпочтительные адъюванты для применения в индукции преимущественно ответа Th1-типа включают в себя, например, комбинацию монофосфориллипида А, предпочтительно 3-де-O-ацилированного монофосфориллипида A (3D-MPL), вместе с солью алюминия. MPL-адъюванты доступны из Ribi IiniriunoChem Research Inc. (Hamilton, MT; патенты Соединенных Штатов № 4436727, 4877611, 4866034 и 4912024). CpG-содержащие олигонуклеотиды (в которых динуклеотид CpG является неметилированным) также индуцируют преимущественно Th1-ответ. Такие олигонуклеотиды хорошо известны и описаны, например, в WO 96/02555. Другим предпочтительным адъювантом является сапонин, предпочтительно QS21, который может использоваться отдельно или в комбинации с другими адъювантами. Например, усиленная система предусматривает комбинацию монофосфориллипида А и производного сапонина, например комбинацию QS21 и 3D-MPL, как описано в WO 94/00153, или менее реакционноспособную композицию, в которой QS21 погашен холестерином, как описано в WO 96/33739. Другие предпочтительные композиции содержат эмульсию типа масло-в-воде и токоферол. Особенно сильнодействующая адъювантная композиция, содержащая QS21, 3D-MPL и токоферол в эмульсии типа масло-в-воде, описана в WO 95/17210. Любая рассматриваемая здесь вакцина может быть приготовлена с использованием хорошо известных способов, которые могут приводить к получению комбинации антигена, усилителя иммунного ответа и подходящего носителя или наполнителя.
Описанные здесь композиции могут вводиться в виде части формы продолжительного высвобождения (т.е. композиции, такой как капсула или пористый материал, которая осуществляет медленное высвобождение соединения после введения). Такие формы могут быть обычно приготовлены с использованием хорошо известных способов и могут вводиться, например, перорально, ректально или подкожной имплантацией, или имплантацией в целевом участке-мишени. Формы продолжительного высвобождения могут содержать полипептид, полинуклеотид или антитело, диспергированные в матриксе-носителе и/или содержащиеся в резервуаре, окруженном регулирующей скорость мембраной; предпочтительно эта форма обеспечивает относительно константный уровень высвобождения активного компонента. Количество активного компонента, содержащееся в композиции продолжительного высвобождения, зависит от места имплантации, скорости и ожидаемой продолжительности высвобождения и характера подлежащего лечению или предупреждению состояния.
Любой из разнообразных доставляющих носителей может быть использован в фармацевтических композициях и вакцинах для облегчения получения антиген-специфического иммунного ответа, который нацелен на опухолевые клетки. Носители для доставки включают в себя антиген-презентирующие клетки (АРС), такие как дендритные клетки, макрофаги, В-клетки, моноциты и другие клетки, которые могут быть сконструированы таким образом, чтобы они были эффективными АРС. Такие клетки могут быть, но необязательно, генетически модифицированными для увеличения способности презентации антигена, для улучшения активации и/или поддержания Т-клеточного ответа, чтобы иметь противоопухолевые эффекты per se и/или быть иммунологически совместимыми с реципиентом (т.е. иметь совместимый гаплотип HLA). АРС могут быть обычно выделены из любых различных биологических жидкостей и органов, в том числе опухолевых и околоопухолевых тканей, и могут быть аутологичными, аллогенными, сингенными или ксеногенными клетками.
Некоторые предпочтительные варианты данного изобретения используют дендритные клетки или их клетки-предшественники в качестве антиген-презентирующих клеток. Дендритные клетки являются очень сильными АРС (Banchereau and Steinman, Nature 392:245-251, 1998), и было показано, что они являются эффективными в качестве физиологического адъюванта для индукции профилактического или терапевтического противоопухолевого иммунитета (Timmerman and Levy, Ann. Rev. Med. 50:507-529, 1999). Обычно дендритные клетки могут быть идентифицированы на основании их типичной формы (звездообразной in situ, с заметными цитоплазматическими выступами (дендрита-ми), видимыми in vitro) и на основании отсутствия маркеров дифференцировки В-клеток (CD19 и CD20), Т-клеток (CD3), моноцитов (CD14) и натуральных киллеров (CD56), определяемой с использованием стандартных анализов. Дендритные клетки могут быть, конечно, сконструированы для экспрессии специфических поверхностных рецепторов или лигандов, которые обычно не обнаруживаются на дендритных клетках in vivo или ex vivo, и такие модифицированные дендритные клетки рассматриваются данным изобретением. В качестве альтернативы дендритным клеткам в вакцине могут быть использованы секретируемые пузырьки нагруженных антигеном дендритных клеток (называемые экзосомами) (Zitovogel et al.. Nature Med. 4:594-600, 1998).
Дендритные клетки и их клетки-предшественники могут быть получены из периферической крови, костного мозга, инфильтрирующих опухоль клеток, инфильтрирующих околоопухолевые ткани клеток, лимфатических узлов, селезенки, кожи, крови, пупочного канатика или любой другой подходящей ткани или жидкости. Например, дендритные клетки могут быть дифференцированы ех vivo путем добавления комбинации цитокинов, таких как GM-CSF, IL-4, IL-13 и/или TNFα, к культурам моноцитов, собранных из периферической крови. Альтернативно, CD3 4-положительные клетки, собранные из периферической крови, крови пуповины или костного мозга, могут быть дифференцированы в дендритные клетки путем добавления к культуральной среде комбинаций GM-CSF, IL-3, TNFa, СD40-лиганда, LPS, flt3-лиганда и/или другого соединения (соединений), индуцирующих созревание и пролиферацию дендритных клеток.
Дендритные клетки подразделяют для удобства на “незрелые” и “зрелые” клетки, что делает возможным простой способ различения между двумя хорошо охарактеризованными фенотипами. Однако эта номенклатура не должна предназначаться для исключения всех возможных промежуточных стадий дифференцировки. Незрелые дендритные клетки характеризуются как АРС с высокой способностью поглощения и процессирования антигена, которая коррелирует с высокой экспрессией Fcγ-рецептора, рецептора маннозы и DEC-205-маркера. Зрелый фенотип обычно характеризуется более низкой экспрессией этих маркеров, но отличается высокой экспрессией молекул поверхности клетки, ответственных за активацию Т-клеток, таких как МНС класса I и класса II, молекулы адгезии (например, CD54 и CD11) и костимулирующие молекулы (например, CD40, CD80 и CD86).
Обычно АРС могут быть трансфицированы полинуклеотидом, кодирующим опухолевый белок предстательной железы (или его часть или его другой вариант), так что на клеточной поверхности экспрессируется опухолевый полипептид предстательной железы или его иммуногенная часть. Такая трансфекция может иметь место ex vivo, и затем композиция или вакцина, содержащая такие трансфицированные клетки, могут быть использованы для терапевтических целей, как описано здесь. Альтернативно, носитель для доставки гена, которые нацелен на дендритную или другую антиген-презентирующуго клетку, может быть введен пациенту, что приводит к трансфекции, которая происходит in vivo. Трансфекция in vivo и ex vivo дендритных клеток может обычно выполняться, например, с использованием любых способов, известных в данной области, таких как способы, описанные в WO 97/24447, или подход с использованием генной пушки, описанный Mahvi et al., Immunology and Cell Biology 75:456-460, 1997. Нагрузка антигеном дендритных клеток может быть достигнута инкубированием дендритных клеток или их клеток-предшественников с опухолевым полипептидом предстательной железы, ДНК (депротеинизированной или в плазмидном векторе) или РНК; или с экспрессирующими антиген рекомбинантными бактерией или вирусами (например, векторами, являющимся вирусом коровьей оспы, птичьей оспы, аденовирусом или лентивирусом). Перед нагрузкой полипептид может быть ковалентно конъюгирован с иммунологическим партнером, который обеспечивает помощь Т-клеткам (например, молекулой-носителем). Альтернативно, дендритная клетка может быть кратковременно обработана неконъюгированным иммунологическим партнером, отдельно или в присутствии полипептида.
ТЕРАПИЯ РАКА
В дополнительных аспектах данного изобретения описанные здесь композиции могут быть использованы для иммунотерапии рака, такого как рак предстательной железы. В таких способах фармацевтические композиции и вакцины обычно вводят пациентам. В применении здесь термин "пациент" относится к любому теплокровному животному, предпочтительно человеку.
Пациент может быть или может не быть поражен раком. Вышеописанные композиции и вакцины могут быть использованы для предупреждения развития рака или для лечения пациента, пораженного раком. Рак может быть диагностирован с использованием критериев, обычно принятых в данной области, в том числе по наличию злокачественной опухоли. Фармацевтические композиции и вакцины могут вводиться либо до, либо после хирургического удаления первичных опухолей и/или лечения, такого как проведение лучевой терапии или введение общепринятых химиотерапевтических лекарственных средств.
В некоторых вариантах иммунотерапия может быть активной иммунотерапией, в которой лечение основано на стимуляции in vivo эндогенной иммунной системы человека для реакции против опухолей путем введения модифицирующих иммунный ответ агентов (таких как описанные здесь полипептиды и полинуклеотиды).
В других вариантах иммунотерапия может быть пассивной иммунотерапией, в которой лечение предусматривает доставку агентов с установленной опухолеиммунной реактивностью (таких как эффекторные клетки или антитела), которые могут непосредственно или опосредованно медиировать противоопухолевые эффекты и необязательно зависят от интактной иммунной системы хозяина. Примеры эффекторных клеток включают в себя обсужденные выше Т-клетки, Т-лимфоциты (такие как CD8+ цитотоксические Т-лимфоциты и CD4+ Т-хелперные инфильтрирующие опухоль лимфоциты), клетки-киллеры (такие как природные клетки-киллеры и активируемые лимфокинами клетки-киллеры), В-клетки и антиген-презентирующие клетки (такие как дендритные клетки и макрофаги), экспрессирующие предлагаемый здесь полипептид. Рецепторы Т-клеток и рецепторы антител, специфическиие для описанных здесь полипептидов, могут быть клонированы, экспрессированы и перенесены в другие векторы или эффекторные клетки для адоптивной иммунотерапии. Обеспеченные здесь полипептиды могут быть также использованы для генерирования антител или антиидиотипических антител (как описано выше и в патенте Соединенных Штатов № 4918164) для пассивной иммунотерапии.
Эффекторные клетки могут быть обычно получены в достаточных количествах для адоптивной иммунотерапии путем выращивания in vitro, как описано здесь. Условия культуры для размножения одиночных антиген-специфических эффекторных клеток до нескольких биллионов клеток с сохранением узнавания in vivo хорошо известно в данной области. Такие условия культивирования in vitro обычно предусматривают периодическую стимуляцию антигеном, часто в присутствии цитокинов (таких как IL-2) и неделящихся питающих клеток-фидеров. Как отмечалось выше, иммунореактивные полипептиды, обеспеченные здесь, могут быть использованы для быстрого размножения культур антиген-специфических Т-клеток для генерирования достаточного числа клеток для иммунотерапии. В частности, антиген-презентирующие клетки, такие как дендритные клетки, макрофаги, моноциты, фибробласты или В-клетки, могут быть кратковременно обработаны иммунореактивными полипептидами или трансфицированы одним или несколькими полинуклеогидами с использованием стандартных способов, хорошо известных в данной области. Например, антиген-презентирующие клетки могут быть трансфицированы полинуклеотидом, имеющим промотор, пригодный для увеличения экспрессии в рекомбинантной вирусной или другой экспрессионной системе. Культивируемые эффекторные клетки для применения в терапии должны быть способны расти и широко распространяться и выживать в течение длительного времени in vivo. Исследования показали, что культивируемые эффекторные клетки могут быть индуцированы для роста in vivo и длительного выживания в значительных количествах путем повторяемой стимуляции антигеном, дополненным IL-2 (например, Cheever et al., Immunological Reviews 157:177, 1997).
Альтернативно, вектор, экспрессирующий описанный здесь полипептид, может быть введен в антиген-презентирующие клетки, взятые из пациента, и клонально размножен ех vivo для трансплантации обратно в того же самого пациента. Трансфицированные клетки могут быть повторно введены с использованием любого из известных в данной области способов, предпочтительно в стерильной форме посредством внутривенного, внутриполостного, внутрибрюшинного или внутриопухолевого введения.
Пути и частота введения описанных здесь терапевтических композиций, а также доза, будут варьировать от индивидуума к индивидууму и могут быть легко установлены с использованием стандартных способов. Обычно эти фармацевтические композиции могут вводиться инъекцией (например, внутрикожной, внутримышечной, внутривенной или подкожной), интраназально (например, аспирацией) или перорально. Предпочтительно 1-10 доз могут быть введены на протяжении периода 52 недель. Предпочтительно вводят 6 доз с интервалами 1 месяц, и после этого могут периодически проводиться бустер-вакцинации. Для индивидуальных пациентов могут быть подходящими другие протоколы введения. Подходящей дозой является количество соединения, которое при введении, как описано выше, способно стимулировать противоопухолевый иммунный ответ и находится, по меньшей мере, на 10-50% выше фонового уровня (т.е. без введения этого соединения). Такой ответ может быть подвергнут мониторингу путем измерения противоопухолевых антител в пациенте или по вакцина-зависимому генерированию цитолитических эффекторных клеток, способных убивать опухолевые клетки пациента in vitro. Такие вакцины должны быть способны вызывать иммунный ответ, который приводит к улучшенному клиническому результату (например, более частым ремиссиям, полному или частичному или более продолжительному выживанию без заболевания) в случае вакцинированных пациентов в сравнении с невакцинированными пациентами. Обычно для фармацевтических композиций и вакцин, содержащих один или несколько полипептидов, количество каждого полипептида находится в диапазонах доз от приблизительно 100 мкг до 5 мг/кг веса тела хозяина. Подходящие размеры доз будут варьировать в зависимости от размера пациента, но обычно будут находиться в диапазоне от приблизительно 0,1 мл до приблизительно 5 мл.
Обычно подходящая схема приема и лечения обеспечивает активное соединение (активные соединения) в количестве, достаточном для обеспечения терапевтического и/или профилактического преимущества. Такой ответ может наблюдаться по установлению улучшенного клинического результата (например, более частых ремиссий, полного или частичного или более продолжительного выживания без заболевания) в случае получающих лечение пациентов в сравнении с не получающими лечение пациентами. Увеличения предсуществующих иммунных ответов на опухолевый белок предстательной железы обычно коррелируют с улучшенным клиническим результатом. Такие иммунные ответы обычно оценивают с использованием стандартных анализов пролиферации, цитотоксичности или анализов на цитокины, которые могут выполняться с использованием образцов, полученных из пациента до и после лечения.
СПОСОБЫ ОБНАРУЖЕНИЯ РАКА
Обычно рак может быть обнаружен у пациента на основании наличия одного или нескольких опухолевых белков предстательной железы и/или полинуклеотидов, кодирующих такие белки, в биологическом образце (например, крови, сыворотке, моче и/или биопстате опухоли), полученном из пациента. Другими словами, такие белки могут быть использованы в качестве маркеров для указания на наличие или отсутствие рака, такого как рак предстательной железы. Кроме того, такие белки могут быть применимы для обнаружения других раков. Связывающие агенты, обеспеченные здесь, обычно позволяют определять уровень антигена, который связывается с таким агентом, в биологическом образце. Полинуклеотидные праймеры и зонды могут быть использованы для определения уровня мРНК, кодирующей опухолевый белок, который также может указывать на присутствие или отсутствие рака. Обычно опухолевая последовательность мРНК предстательной железы должна присутствовать при уровне, который, по меньшей мере, в три раза выше в опухолевой ткани, чем в здоровой ткани.
Имеются различные форматы анализов, известные средним специалистам в данной области, для использования связывающего агента для определения полипептидных маркеров в образце. Например, Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, 1988. Обычно наличие или отсутствие раковой опухоли у пациента может быть определено (а) взаимодействием биологического образца, полученного из пациента, со связывающим агентом; (b) определением в указанном образце уровня полипептида, который связывается со связывающим агентом; и (с) сравнением уровня полипептида с заранее определенной предельной величиной.
В предпочтительном варианте анализ предусматривает использование связывающего агента, иммобилизованного на твердом носителе, для связывания и удаления указанного полипептида из остального образца. Затем связанный полипептид может определяться с использованием определяющего реагента, который содержит репортерную группу и специфически связывается с комплексом связывающий агент/полипептид. Такие определяющие агенты могут содержать, например, связывающий агент, который специфически связывается с данным полипептидом, или антитело или другой агент, который специфически связывается со связывающим агентом, такой как анти-иммуноглобулин, белок G, белок А или лектин. Альтернативно, может быть использован конкурентный анализ, в котором полипептид метят репортерной группой и дают ему связываться с иммобилизованным связывающим агентом после инкубации связывающего агента с образцом. Степень, до которой компоненты указанного образца подавляют связывание меченого полипептида со связывающим агентом, указывает на реактивность указанного образца с иммобилизованным связывающим агентом. Подходящие полипептиды для использования в таких анализах, включают в себя полноразмерные опухолевые белки предстательной железы и их части, с которыми связывается связывающий агент, как описано выше.
Твердым носителем может быть любой материал, известный средним специалистам в данной области, к которому может быть присоединен опухолевый белок. Например, твердым носителем может быть тест-лунка в планшете для микротитрования или нитроцеллюлозная или другая подходящая мембрана. Альтернативно, носителем может быть гранула или диск, такой как стеклянный, стекловолоконный, латексный или пластиковый материал, такой как полистирол или поливинилхлорид. Носитель может быть также магнитной частицей или волоконно-оптическим датчиком, таким как описанный, например, в патенте Соединенных Штатов № 5359681. Связывающий агент может быть иммобилизован на твердом носителе при помощи различных способов, известных средним специалистам в данной области, которые вполне достаточно описаны в патентной и научной литературе. В контексте данного изобретения термин "иммобилизация" относится как к нековалентной связи, такой как адсорбция, так и к ковалентному присоединению (которое может быть прямой связью между агентом и функциональными группами на носителе или может быть связью посредством сшивающего агента). Предпочтительной является иммобилизация посредством адсорбции с лункой в планшете для микротитрования или с мембраной. В таких случаях адсорбция может достигаться взаимодействием связывающего агента в подходящем буфере с указанным твердым носителем в течение подходящего периода времени. Время взаимодействия варьируется в зависимости от температуры, но обычно находится в диапазоне между приблизительно 1 часом и приблизительно 1 днем. Обычно взаимодействие лунки пластикового планшета для микротитрования (такого как полистироловый или поливинилхлоридный планшет) с количеством связывающего агента от приблизительно 10 нг до приблизительно 10 мкг и предпочтительно от приблизительно 100 нг до приблизительно 1 мкг является достаточным для иммобилизации адекватного количества связывающего агента.
Ковалентное присоединение связывающего агента к твердому носителю обычно достигается сначала реакцией носителя с бифункциональным реагентом, который будет реагировать как с носителем, так и с функциональной группой, такой как гидроксил или аминогруппа, на связывающем агенте. Например, связывающий агент может быть ковалентно присоединен к носителям, имеющим подходящее полимерное покрытие, при помощи бензохинона или посредством конденсации альдегидной группы на носителе с амином и активным водородом на партнере связывания (например, Pierce Immunotechnology Catalog and Handbook, 1991, at A12-A13).
В некоторых вариантах указанный анализ является сэндвич-анализом с использованием двух антител. Указанный анализ может выполняться взаимодействием сначала антитела, которое было иммобилизовано на твердом носителе, обычно лунке планшета для микротитрования, с образцом, так чтобы полипептиды в образце могли связываться с иммобилизованным антителом. Затем несвязанный образец удаляют из иммобилизованных комплексов полипептид-антитело и добавляют определяющий реагент (предпочтительно второе антитело, способное связываться с отличающимся сайтом на указанном полипептиде), содержащий репортерную группу. Затем определяют количество определяющего реагента, которое остается связанным с твердым носителем, с использованием способа, подходящего для этой специфической репортерной группы.
Более конкретно, после иммобилизации антитела на носителе, как описано выше, оставшиеся сайты связывания белка на носителе обычно блокируют любым подходящим блокирующим агентом, известным средним специалистам в данной области, таким как бычий сывороточный альбумин или Твин 20™ (Sigma Chemical Co., St. Louis, МО). Затем иммобилизованное антитело инкубируют с образцом и полипептиду дают связаться с антителом. Образец может быть разбавлен подходящим растворителем, таким как забуференный фосфатом солевой раствор (ЗФР), перед инкубированием. Обычно подходящим временем взаимодействия (т.е. временем инкубации) является период времени, который является достаточным для определения наличия полипептида в образце, полученного из индивидуума с раком предстательной железы. Предпочтительно это время контакта является достаточным для достижения уровня связывания, которое равно, по меньшей мере, приблизительно 95% времени, достигаемого при равновесии между связавшимся и несвязавшимся полипептидом. Среднему специалисту в данной области будет понятно, что время, необходимое для достижения равновесия, может быть легко определено путем определения уровня связывания, которое имеет место на протяжении некоторого периода времени. При комнатной температуре обычно является достаточным время инкубации приблизительно 30 минут.
Затем несвязавшийся образец может быть удален промыванием твердого носителя подходящим буфером, таким как ЗФР, содержащий 0,1% Твин 20™. Затем к твердому носителю может быть добавлено второе антитело, которое содержит репортерную группу. Предпочтительные репортерные группы включают в себя группы, описанные выше.
Затем определяющий реагент инкубируют с комплексом иммобилизованное антитело/полипептид в течение времени, достаточного для определения связанного полипептида. Подходящее время может быть обычно определено определением уровня связывания, происходящего на протяжении некоторого периода времени. Затем несвязанный определяющий реагент удаляют, а связанный определяющий реагент определяют с использованием репортерной группы. Способ, применяемый для определения репортерной группы, зависит от природы этой репортерной группы. Для радиоактивных групп обычно подходят способы сцинтилляционного счета или ауторадиографии. Спектроскопические способы могут быть использованы для определения красителей, люминесцентных групп и флуоресцентных групп. Биотин может определяться с использованием авидина, связанного с отличающейся репортерной группой (обычно радиоактивной или флуоресцентной группой или ферментом). Ферментная репортерная группа может обычно определяться путем добавления субстрата (обычно в течение определенного периода времени) с последующим спектроскопическим или другим анализом продуктов реакции.
Для определения наличия или отсутствия рака, такого как рак предстательной железы, сигнал, определяемый из репортерной группы, которая остается связанной с твердым носителем, обычно сравнивают с сигналом, соответствующим заранее определенной предельной величине. В одном предпочтительном варианте предельной величиной для определения рака является среднее значение сигнала, полученного при инкубировании иммобилизованного антитела с образцами пациентов, не страдающих раковой опухолью. Обычно образцу, генерирующую сигнал, который на три стандартных отклонения выше заранее определенной предельной величины, считают положительным в отношении рака. В другом предпочтительном варианте предельную величину определяют с использованием кривой Receiver Operator Curve в соответствии со способом Sackett et al., Clinical Epidemiology: A Basic Science for Clinical Medicine, Little Brown and Co., 1985, p.106-7. Вкратце, в указанном варианте предельная величина может быть определена из графика пар истинно положительных уровней (т.е. чувствительности) и ложноположительных уровней (100%-специфичность), которые соответствуют каждой возможной предельной величине для каждого диагностического результата. Предельная величина на указанном графике, т.е. наиболее близкая к верхнему левому углу (т.е. величина, включающая в себя наибольшую площадь), является наиболее точной предельной величиной, и образец, генерирующий сигнал, который является более высоким, чем предельная величина, определенная указанным способом, может считаться положительной. Альтернативно, предельная величина может быть сдвинута влево вдоль графика для минимизации ложноположительного уровня или вправо для минимизации ложноотрицательного уровня. Обычно образец, генерирующий сигнал, который является более высоким, чем предельная величина, определенная указанным способом, считают положительным в отношении рака.
В родственном варианте указанный анализ выполняют в формате проточного теста или теста-полоски, в котором связывающий агент иммобилизуют на мембране, такой как нитроцеллюлозная мембрана. В проточном тесте полипептиды в образце связываются с иммобилизованным связывающим агентом при прохождении образца через эту мембрану. Затем второй меченый связывающий агент связывается с комплексом связывающий агент/полипептид при протекании раствора, содержащего второй связывающий агент, через мембрану. Затем может быть выполнено определение связанного второго связывающего агента, как описано выше. В формате тест-полоски один конец мембраны, с которой связан связывающий агент, погружен в раствор, содержащий образец. Образец мигрирует вдоль мембраны через район, содержащий второй связывающий агент, к зоне иммобилизованного связывающего агента. Концентрация второго связывающего агента в зоне иммобилизованного антитела указывает на присутствие рака. Обычно концентрация второго связывающего агента в указанном участке генерирует рисунок, такой как линия, который может регистрироваться визуально. Отсутствие такого рисунка указывает на отрицательный результат. Обычно количество связывающего агента, иммобилизованного на мембране, выбрано таким образом, чтобы генерировать визуально различимый рисунок, когда биологический образец содержит уровень полипептида, который был бы достаточным для генерирования положительного сигнала в сэндвич-анализе с двумя антителами в обсужденном выше формате. Предпочтительными связывающими агентами для применения в таких анализах являются антитела и их антигенсвязывающие фрагменты. Предпочтительно количество антитела, иммобилизованного на мембране, находится в диапазоне от приблизительно 25 нг до приблизительно 1 мкг и более предпочтительно от приблизительно 50 нг до приблизительно 500 нг. Такие тесты обычно могут проводиться с очень маленьким количеством биологического образца.
Конечно, существуют многочисленные другие протоколы анализов, которые пригодны для использования с опухолевыми белками или связывающими агентами данного изобретения. Приведенные выше описания должны рассматриваться только как примеры. Например, средним специалистам в данной области будет понятно, что вышеописанные протоколы могут быть легко модифицированы для применения опухолевых полипептидов предстательной железы для определения антител, которые связываются с такими полипептидами, в биологическом образце. Определение таких специфических для опухолевого белка предстательной железы антител может коррелировать с присутствием рака.
Рак может быть также или альтенативно определяться на основании наличия Т-клеток, которые специфически реагируют с опухолевым белком предстательной железы, в биологическом образце. В некоторых способах биологический образец, содержащий CD4+ и/или CD8+ Т-клетки, выделенный из пациента, инкубируют с опухолевым полипептидом предстательной железы, полинуклеотидом, кодирующим такой полипептид, и/или АРС, которая экспрессирует, по меньшей мере, иммуногенную часть такого полипептида, и определяют наличие или отсутствие специфической активации этих Т-клеток. Пригодные биологические образцы включают в себя, но не ограничиваются ими, выделенные Т-клетки. Например, Т-клетки могут быть выделены из пациента рутинными способами (такими как центрифугирование в градиенте плотности фиколла/Hypaque лимфоцитов периферической крови). Т-клетки могут быть инкубированы in vitro в течение 2-9 дней (обычно 4 дней) при 37°С с опухолевым полипептидом предстательной железы (например, 5-25 мкг/мл). Может быть желательным инкубирование другой аликвоты образца Т-клеток в отсутствие опухолевого полипептида предстательной железы в качестве контроля. Для CD4+ Т-клеток активацию предпочтительно определяют оценкой пролиферации Т-клеток. Для CD8+ Т-клеток активацию предпочтительно определяют оценкой цитолитической активности. Уровень пролиферации, который по меньшей мере в два раза выше, и/или уровень цитолитической активности, который по меньшей мере на 20% выше, чем в не имеющих заболевания пациентах, указывает на наличие раковой опухоли у пациента.
Как отмечалось выше, рак может быть определен также или альтернативно на основании уровня мРНК, кодирующей опухолевый белок предстательной железы, в биологическом образце. Например, по меньшей мере два олигонуклеотидных праймера могут быть использованы в анализе на основе полимеразной цепной реакции (ПЦР) для амплификации части кДНК опухоли предстательной железы, полученной из биологического образца, в котором по меньшей мере один из олигонуклеотидных праймеров является специфическим для (т.е. гибридизуется с ним) полинуклеотида, кодирующего опухолевый белок предстательной железы. Затем амплифицированную кДНК отделяют и определяют с использованием способов, хорошо известных в данной области, таких как гель-электрофорез. Подобным образом, олигонуклеотидные зонды, которые специфически гибридизуются с полинуклеотидом, кодирующим опухолевый белок предстательной железы, могут быть использованы в гибридизационном анализе для определения наличия полинуклеотида, кодирующего опухолевый белок, в биологическом образце.
Для возможности гибридизации в условиях анализа олигонуклеотидные праймеры и зонды должны содержать олигонуклеотидную последовательность, которая имеет по меньшей мере приблизительно 60%, предпочтительно по меньшей мере приблизительно 75% и более предпочтительно по меньшей мере приблизительно 90% идентичность относительно части полинуклеотида, кодирующего опухолевый белок предстательной железы, которая имеет длину по меньшей мере 10 нуклеотидов и предпочтительно по меньшей мере 20 нуклеотидов. Предпочтительно олигонуклеотидные праймеры и/или зонды будут гибридизоваться с полинуклеотидом, кодирующим описанный здесь полипептид, при умеренно жестких условиях, определенных выше. Олигонуклеотидные праймеры и/или зонды, которые могут быть выгодно использованы в диагностических способах, описанных здесь, предпочтительно имеют длину по меньшей мере 10-40 нуклеотидов. В предпочтительных вариантах олигонуклеотидные праймеры и зонды содержат по меньшей мере 10 непрерывных нуклеотидов, более предпочтительно по меньшей мере 15 непрерывных нуклеотидов молекулы ДНК, имеющей последовательность, представленную в SEQ ID NО:1-111, 115-171, 173-175, 177, 179-305, 307-315, 326, 328, 330, 332-335, 340-375 и 381. Способы для анализов на основе ПЦР и гибридизационных анализов хорошо известны в данной области (например, Mullis et al., Cold Spring Harbor Symp. Quant. Biol., 51:263, 1987; Eriich ed., PCR Technology, Stockton Press, NY, 1989).
В одном предпочтительном анализе используют ОТ-ПЦР, в которой ПЦР применяют в сочетании с обратной транскрипцией. Обычно РНК экстрагируют из биологического образца, такого как ткань биопсии, и обратно транскрибируют с получением кДНК-молекул. ПЦР-амплификация с использованием, по меньшей мере, одного специфического праймера генерирует кДНК-молекулу, которая может быть отделена и визуализирована при помощи, например, гель-электрофореза. Амплификация может выполняться на биологических образцах, взятых из обследуемого пациента и из индивидуума, который не имеет рака. Реакция амплификации может быть проведена на нескольких разведениях кДНК, охватывающих два порядка величин. Двукратное или большее увеличение экспрессии в нескольких разведениях образца обследуемого пациента в сравнении с теми же самыми разведениями неракового образца обычно рассматривается как положительное.
В другом варианте описанные композиции могут быть использованы в качестве маркеров прогрессирования рака. В указанном варианте описанные выше анализы для диагностики рака могут выполняться на протяжении времени и оценивается изменение в уровне реакционноспособного полипептида (полипептидов) или полинуклеотида. Например, эти анализы могут выполняться каждые 24-72 часа в течение периода 6 месяцев-1 года и после указанного выполняться по необходимости. Обычно рак является прогрессирующим в пациентах, в которых уровень определяемого полипептида или полинуклеотида увеличивается со временем. В противоположность указанному рак не является прогрессирующим, когда уровень реакционноспособного полипептида или полинуклеотида либо остается постоянным, либо снижается со временем.
Некоторые диагностические анализы in vivo могут проводиться непосредственно на опухоли. Один из таких анализов предусматривает взаимодействие опухолевых клеток со связывающим агентом. Затем связанный связывающий агент может быть детектирован прямо или косвенно через репортерную группу. Такие связывающие агенты могут также использоваться в гистологических приложениях. Альтернативно, полинуклеотидные зонды могут быть использованы для таких приложений.
Как отмечалось выше, для улучшения чувствительности многочисленные маркеры опухолевого белка предстательной железы могут анализироваться в конкретном образце. Должно быть понятно, что связывающие агенты, специфические для различных обеспеченных здесь белков, могут комбинироваться в едином анализе. Далее, многочисленные праймеры или зонды могут использоваться совместно. Выбор маркеров опухолевого белка может основываться на рутинных экспериментах для определения комбинаций, которые дают оптимальную чувствительность. Кроме того, или альтернативно, анализы на опухолевые белки, обеспеченные здесь, могут комбинироваться с анализами на другие известные опухолевые антигены.
ДИАГНОСТИЧЕСКИЕ НАБОРЫ
Данное изобретение обеспечивает, далее, наборы для применения в любом из вышеописанных диагностических способов. Такие наборы обычно содержат два или более компонентов, необходимых для проведения диагностического анализа. Компоненты могут быть соединениями, реагентами, контейнерами и/или оборудованием. Например, один контейнер в наборе может содержать моноклональное антитело или его фрагмент, который специфически связывается с опухолевым белком предстательной железы. Такие антитела или фрагменты могут быть обеспечены присоединенными к материалу-носителю, как описано выше. Один или несколько дополнительных контейнеров могут содержать элементы, такие как реагенты или буферы для использования в анализе. Такие наборы могут также или альтернативно содержать определяющий реагент, описанный выше, который содержит репортерную группу, пригодную для прямого или опосредованного определения связывания антител.
Альтернативно, набор может быть предназначен для определения уровня мРНК, кодирующей опухолевый белок предстательной железы, в биологическом образце. Такие наборы обычно содержат по меньшей мере один олигонуклеотидный зонд или праймер, описанные выше, который гибридизуется с полинуклеотидом, кодирующим опухолевый белок предстательной железы. Такой олигонуклеотид может быть использован, например, в ПЦР-анализе или гибридизационном анализе. Дополнительные компоненты, которые могут присутствовать в таких наборах, включают в себя второй олигонуклеотид и/или диагностический реагент или контейнер для облегчения определения полинуклеотида, кодирующего опухолевый белок предстательной железы.
Нижеследующие примеры предлагаются в качестве иллюстрации, а не для ограничения изобретения.
ПРИМЕРЫ
ПРИМЕР 1
ВЫДЕЛЕНИЕ И ХАРАКТЕРИСТИКА ОПУХОЛЕВЫХ ПОЛИПЕПТИДОВ ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ
Указанный пример описывает выделение некоторых опухолевых полипептидов предстательной железы из библиотеки кДНК опухоли предстательной железы.
Экспрессионную библиотеку кДНК опухоли предстательной железы человека конструировали из поли А+ РНК с использованием Superscript Plasmid System для синтеза кДНК и набора Plasmid Cloning Kit (BRL Life Technologies, Gaithersburg, MD 20897) в соответствии с протоколом изготовителя. Конкретно, ткани опухоли предстательной железы гомогенизировали с политроном (Kinematica, Switzerland) и общую РНК экстрагировали с использованием реагента Тризола (Life Technologies), как описано изготовителем. Затем поли А+ РНК очищали с использованием набора для очистки мРНК Qiagen oligotex spin column (Qiagen, Santa Clarita, CA 91355) в соответствии с протоколом изготовителя. кДНК первой цепи синтезировали с использованием праймера NotI/Oligo-dT18. Синтезировали двухцепочечную кДНК, лигировали с адаптерами EcoRI/BAXI (Invitrogen, San Diego, CA) и расщепляли NotI. После фракционирования по размеру с использованием колонок Chroma Spin-1000 (Clontech, Palo Alto/ CA) кДНК лигировали в сайт EcoRI/NotI pCDNA3.1 (Invitrogen) и трансформировали в клетки ElectroMax E.coli DH10B (BRL Life Technologies) электропорацией.
С использованием той же самой процедуры экспрессионную библиотеку кДНК поджелудочной железы здорового человека получали из пула шести тканевых образцов (Clontech). Библиотеки кДНК были охарактеризованы путем определения числа независимых колоний, процента клонов, несущих вставку, среднего размера вставки и анализа последовательности. Библиотека опухоли предстательной железы содержала 1,64×107 независимых колоний, причем 70% клонов имели вставку и средний размер вставки был 1745 п.н. Библиотека кДНК здоровой поджелудочной железы содержала 3,3×106 независимых колоний, причем 69% клонов имели вставки и средний размер вставки был 1120 п.н. Для обеих библиотек анализ последовательности показал, что большинство клонов имели полноразмерную последовательность кДНК и были синтезированы из мРНК с минимальной примесью рРНК и митохондриальной ДНК.
Вычитание библиотек кДНК (согласно "субтрактивному методу") выполняли с использованием вышеописанных библиотек кДНК опухоли предстательной железы и нормальной поджелудочной железы, как описано Нага et al. (Blood, 84:189-199, 1994) с некоторыми модификациями. Конкретно, являющуюся результатом вычитания опухолеспецифическую библиотеку кДНК предстательной железы получали следующим образом. Библиотеку кДНК здоровой поджелудочной железы (70 мкг) расщепляли EcoRI, NotI и SfuI с последующей реакцией застраивания концов фрагментом Кленова ДНК-полимеразы. После экстракции смесью фенол-хлороформ и осаждения этанолом ДНК растворяли в 100 мкл Н2О, денатурировали нагреванием и смешивали с 100 мкл (100 мкг) биотина Photoprobe (фотозонда) (Vector Laboratories, Burlingame, CA). Как рекомендовано изготовителем, полученную смесь освещали кинолампой “солнце” 270 Вт на льду в течение 20 минут. Добавляли дополнительное количество биотина Photoprobe (50 мкл) и повторяли реакцию биотинилирования. После экстракции бутанолом пять раз ДНК осаждали этанолом и растворяли в 23 мкл Н2О с получением драйвер-ДНК.
Для образования ДНК-метки 10 мкг библиотеки кДНК опухоли предстательной железы расщепляли BamHI и XhoI, экстрагировали смесью фенол-хороформ и пропускали через колонки Chroma Spin-1000 (Clontech). После осаждения этанолом ДНК-метку растворяли в 5 мкл Н2О. ДНК-метку смешивали с 15 мкл драйвер-ДНК и 20 мкл 2 × гибридизационного буфера (1,5 М NaCl/10 мМ ЭДТА/50 мМ HEPES рН 7,5/0,2% додецилсульфат натрия), покрывали минеральным маслом и полностью денатурировали нагреванием. Пробу сразу же переносили в водяную баню на 68°С и инкубировали в течение 20 часов (длительная гибридизация [LH]). Затем реакционную смесь подвергали обработке стрептавидином с последующей экстракцией смесью фенол/хлороформ. Указанный процесс повторяли еще три раза. Полученную вычитанием ДНК осаждали, растворяли в 12 мкл Н2О, смешивали с 8 мкл драйвер-ДНК и 20 мкл 2× гибридизационного буфера и подвергали гибридизации при 68°С в течение 2 часов (кратковременная гибридизация [SH]). После удаления биотинилированной двухцепочечной ДНК полученную вычитанием кДНК лигировали в сайт BamHI/XhoI хлорамфеникол-резистентной pBCSK+ (Stratagene, La Jolla, CA 92037) и трансформировали в клетки Е.coli DH10B ElectroMax электропорацией для генерирования полученной вычитанием библиотеки кДНК, специфической для опухоли предстательной железы (называемой “вычитание 1 предстательной железы”).
Для анализа полученной вычитанием библиотеки кДНК готовили плазмидную ДНК из 100 независимых клонов, случайно выбранных из полученной вычитанием специфической для опухоли предстательной железы библиотеки и сгруппированных на основе размера вставки. Показательные кДНК-клоны дополнительно характеризовали секвенированием ДНК с использованием автоматического секвенатора модели 373А Perkin Elmer/Applied Biosystems (Foster City, CA). Было показано, что шесть кДНК-клонов, далее называемых F1-13, F1-12, F1-16, H1-1, H1-9 и Н1-4, являются обильно представленными в полученной вычитанием специфической для предстательной железы библиотеке кДНК. Определенные 3’- и 5’-последовательности кДНК для Fl-13 представлены в SEQ ID NО:2 и 3 соответственно, а определенные 3’-последовательности кДНК для F1-13, F1-16, H1-1, H1-9 и Н1-4 представлены в SEQ ID NО:1 и 4-7 соответственно.
Последовательности кДНК для выделенных клонов сравнивали с известными последовательностями в банке генов с использованием баз данных EMBL и GenBank (версии 96). Было определено, что четыре из кДНК-клонов опухоли предстательной железы, F1-13, F1-16, H1-1 и Н1-4, кодируют следующие ранее идентифицированные белки: специфический антиген предстательной железы (PSA), гландулярный калликреин человека, усиленный опухолевой экспрессией ген человека, и субъединицу II цитохром С-оксидазы митохондрий. Было обнаружено, что Р1-9 идентичен ранее идентифицированной автономно реплицирующейся последовательности. Не было найдено значимых гомологии с последовательностью-кДНК для F1-12.
Последующие исследования привели к выделению полноразмерной последовательности-кДНК для F1-12. Эта последовательность изображена в SEQ ID NO:107, с соответствующей предсказанной аминокислотной последовательностью, представленной в SEQ ID NO:108.
Для клонирования менее представленных специфических для опухоли предстательной железы генов вычитание библиотеки кДНК проводили путем вычитания библиотеки кДНК опухоли предстательной железы, описанной выше, с библиотекой кДНК здоровой поджелудочной железы и с тремя наиболее представленными генами в специфической для опухоли предстательной железы библиотеке кДНК, полученной ранее вычитанием: гландулярного калликреина человека, специфического антигена предстательной железы (PSA) и субъединицы II цитохром С-оксидазы митохондрий. Конкретно, 1 мкг каждой из кДНК гландулярного калликреина человека, специфического антигена предстательной железы (PSA) и субъединицы II цитохром С-оксидазы митохондрий в pCDNA3.1 добавляли к драйвер-ДНК и вычитание проводили, как описано выше, с получением второй полученной вычитанием библиотеки кДНК, далее называемой “полученная вычитанием специфическая для опухоли предстательной железы библиотека кДНК со “спайком”.
Двадцать два кДНК-клона выделили из полученной вычитанием специфической для опухоли предстательной железы библиотеки кДНК со “спайком”. Определенные 3’- и 5’-последовательности кДНК для клонов, названных J1-17, L1-12, N1-1862, J1-13, J1-19, J1-25, J1-24, K1-58, K1-63, L1-4 и L1-14, представлены в SEQ ID NO:8-9, 10-11, 12-13, 14-15, 16-17, 18-19, 20-21, 22-23, 24-25, 26-27 и 28-29 соответственно. Определенные 3’-последовательности кДНК для клонов, названных J1-12, J1-16, J1-21, K1-48, K1-55, L1-2, L1-6, N1-1858, N1-1860, N1-1861, N1-1864, представлены в SEQ ID NO:30-40 соответственно. Сравнение этих последовательностей с последовательностями банка генов, как описано выше, не выявило значимых гомологии с тремя из пяти наиболее представленных разновидностей ДНК (J1-17, L1-12 и N-1862; SEQ ID NO:8-9, 10-11 и 12-13 соответственно). Было обнаружено, что из остальных двух наиболее представленных разновидностей, одна (J1-12; SEQ ID NO: 30) является идентичной ранее идентифицированному сурфактант-ассоциированному легочному белку человека, а другая (К1-48; SEQ ID NO:33) имеет некоторую гомологию с мРНК R. norvegicus для 2-арилпропионил-СоА-эпимеразы. Было обнаружено, что из 17 менее представленных кДНК-клонов, выделенных из полученной вычитанием специфической для опухоли предстательной железы библиотеки кДНК со “спайком”, четыре (J1-16, K1-55, L1-6 и N1-1864; SEQ ID NO:31, 34, 36 и 40 соответственно) являются идентичными ранее идентифицированным последовательностям, две (J1-21 и N1-1860; SEQ ID NO:32 и 38 соответственно) обнаруживают некоторую гомологию с последовательностями, не относящимися к человеку, и две (L1-2 и N1-1861; SEQ ID NO:35 и 39 соответственно) обнаруживают некоторую гомологию с известными последовательностями человека. Не было обнаружено значимых гомологий к полипептидам J1-13, J1-19, J1-24, J1-25, K1-58, K1-63, L1-4, L1-14 (SEQ ID NO:14-15, 16-17, 20-21, 18-19, 22-23, 24-25, 26-27, 28-29 соответственно).
Последующие исследования привели к выделению полноразмерных последовательностей-кДНК для J1-17, L1-12 и N1-1862 (SEQ ID NO:109-111 соответственно). Соответствующие аминокислотные последовательности представлены в SEQ ID NO:112-114. L1-12 называют также P501S.
В следующем эксперименте были идентифицированы четыре дополнительных клона путем вычитания библиотеки кДНК опухоли предстательной железы с кДНК здоровой предстательной железы из пула трех поли А+ РНК здоровой предстательной железы (называемого "вычитание 2 предстательной железы"). Определенные последовательности-кДНК для этих клонов, далее называемые U1-3064, U1-3065, V1-3692 и 1А-3905, представлены в SEQ ID NO:69-72 соответственно. Сравнение этих определенных последовательностей с последовательностями в банке генов не выявило значимых гомологий к U1-3065.
Второе вычитание со “спайком” (называемое “вычитанием со спайком 2 предстательной железы”) выполняли путем вычитания специфической для опухоли предстательной железы библиотеки кДНК со “спайком” с библиотекой кДНК здоровой поджелудочной железы и с дополнительным "спайком" в виде генов PSA, J1-17, сурфактант-ассоциированного легочного белка, субъединицы II цитохром С-оксидазы митохондрий, N1-1862, автономно реплицирующейся последовательности, L1-12 и усиленного опухолевой экспрессией гена. Были выделены четыре дополнительных гена, далее называемых V1-3686, R1-2330, 1В-3976 и V1-3679. Определенные последовательности-кДНК для этих клонов представлены в SEQ ID NO:73-76 соответственно. Сравнение этих последовательностей с последовательностями банка генов не выявили значимых гомологий к V1-3686 и R1-2330.
Дополнительный анализ трех вычитании предстательной железы, описанных выше (вычитания 2 предстательной железы, полученной вычитанием специфической для опухоли предстательной железы библиотеки кДНК со спайком и вычитания со спайком 2 предстательной железы), привел к идентификации шестнадцати дополнительных клонов, названных 1G-4736, 1G-4738, 1G-4741, 1G-4744, 1G-4734, 1Н-4774, 1Н-4781, 1H-4785, 1H-4787, 1H-4796, 1I-4810, 1I-4811, 1J-4876, 1K-4884 и 1К-4896. Определенные последовательности-кДНК для этих клонов представлены в SEQ ID NO:77-92 соответственно. Сравнение этих последовательностей с последовательностями банка генов не выявили значимых гомологий к 1G-4741, 1G-4734, 1I-4807, 1J-4876 и 1К-4896 (SEQ ID NO:79, 81, 87, 90 и 92 соответственно). Дополнительный анализ этих выделенных клонов привел к определению удлиненных последовательностей-кДНК для 1G-4736, 1G-4738, 1G-4741, 1G-4744, 1Н-4774, 1Н-4781, 1H-4785, 1Н-4787, 1Н-4796, 1I-4876, 1K-4884 и 1К-4896, представленных в SEQ ID NO:179-188 и 191-193 соответственно, и к определению дополнительных частичных последовательностей-кДНК для 1I-4810 и 1I-4811, представленных в SEQ ID NO:189 и 190 соответственно.
Дополнительные исследования вычитания предстательной железы со спайком 2 привело к выделению еще трех клонов. Их последовательности определяли, как описано выше, и сравнивали с самым последним банком генов. Было обнаружено, что все три клона имеют гомологию с известными генами, которые представляют обогащенный цистеином белок, KIAA0242 и KIAA0280 (SEQ ID NО:317, 319 и 320 соответственно). Дальнейший анализ этих клонов с использованием микроматрицы Synteni (Synteni, Palo Alto, CA) показал, что все три клона сверхэкспрессировались в большинстве опухолей предстательной железы и ВРН предстательной железы, а также в большинстве исследованных тканей здоровой предстательной железы, но имели низкую экспрессию во всех других здоровых тканях.
Дополнительное вычитание проводили путем вычитания библиотеки кДНК здоровой предстательной железы с кДНК здоровой поджелудочной железы (называемое “вычитание 3 предстательной железы). Это привело к идентификации шести дополнительных клонов, названных 1G-4761, 1G-4762, 1Н-4766, 1Н-4770, 1Н-4771 и 1Н-4772 (SEQ ID NO:93-98). Сравнение этих последовательностей с последовательностями банка генов не выявили значимых гомологии с 1G-4761 и 1Н-4771 (SEQ ID NO:93 и 97 соответственно). Дальнейший анализ выделенных клонов привел к определению удлиненных последовательностей-кДНК для 1G-4761, 1G-4762, 1Н-4766 и 1Н-4772, представленных в SEQ ID NO:194-196 и 199 соответственно и к определению дополнительных частичных последовательностей-кДНК для 1Н-4770 и 1Н-4771, представленных в SEQ ID NO:197 и 198 соответственно.
Вычитание библиотеки кДНК опухоли предстательной железы, полученной из пула поли A+ РНК из трех пациентов с раком предстательной железы, с библиотекой кДНК здоровой поджелудочной железы (вычитание 4 предстательной железы) привело к идентификации восьми клонов, названных ID-4297, ID-4309, ID.1-4278, ID-4288, ID-4283, ID-4304, ID-4296 и ID-4280 (SEQ ID NO:99-107). Эти последовательности сравнивали с последовательностями банка генов, как описано выше. Не было обнаружено значимых гомологий к ID-4283 и ID-4304 (SEQ ID NO:103 и 104 соответственно). Дальнейший анализ выделенных клонов привел к определению удлиненных последовательностей-кДНК для ID-4320, ID-4278, ID-4288, ID-4283, ID-4304, ID-4296 и ID-4280, представленных в SEQ ID NO:200-206 соответственно.
Клоны кДНК, выделенные из вычитания 1 предстательной железы и вычитания 2 предстательной железы, описанных выше, амплифицировали при помощи ПЦР колоний и их уровни экспрессии мРНК в опухоли предстательной железы, нормальной предстательной железе и в различных других, здоровых тканях определяли с использованием технологии микроматрицы (Sуnteni, Palo Alto, CA). Вкратце, продукты ПЦР-амплификации наносили в виде пятен на предметные стекла в формате микроматрицы, причем каждый продукт занимал уникальное местоположение в этой матрице. мРНК экстрагировали из испытуемого образца ткани, обратно транскрибировали и генерировали флуоресцентно меченные кДНК-зонды. Микроматрицы зондировали мечеными кДНК-зондами, предметные стекла сканировали и измеряли интенсивность флуоресценции. Эта интенсивность коррелирует с интенсивностью гибридизации. Было обнаружено, что два клона (называемые P509S и P510S) сверхэкспрессировались в опухоли предстательной железы и здоровой предстательной железе и экспрессировались при низких уровнях во всех других испытанных нормальных тканях (печени, поджелудочной железе, коже, костном мозгу, головном мозгу, молочной железе, надпочечнике, мочевом пузыре, яичках, слюнных железах, толстой кишке, почке, яичнике, легком, спинном мозгу, скелетной мышце и ободочной кишке). Определенные последовательности-кДНК для P509S и P510S представлены в SEQ ID NO:223 и 224 соответственно. Сравнение этих последовательностей с последовательностями в банке генов, как описано выше, обнаружило некоторую гомологию с ранее идентифицированными EST.
Кроме того, исследования привели к выделению полноразмерной последовательности кДНК для P509S. Эта последовательность представлена в SEQ ID NO:332, а соответствующая аминокислотная последовательность представлена в SEQ ID NO:339.
ПРИМЕР 2
ОПРЕДЕЛЕНИЕ ТКАНЕВОЙ СПЕЦИФИЧНОСТИ ОПУХОЛЕВЫХ ПОЛИПЕПТИДОВ ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ
С использованием ген-специфических праймеров уровни экспрессии мРНК для показательных опухолевых полипептидов предстательной железы F1-16, H1-1, J1-17 (также называемого P502S), L1-12 (также называемого P501S), F1-12 (также называемого P504S), и N1-1862 (также называемого P503S) исследовали в различных нормальных и опухолевых тканях при помощи ОТ-ПЦР.
Вкратце, общую РНК экстрагировали из различных нормальных и опухолевых тканей с использованием реагента Тризола, как описано выше. Синтез первой цепи проводили с использованием 1-2 мкг общей РНК обратной транскриптазой SuperScript (BRL Life Technologies) при 42°С в течение одного часа. Затем эту кДНК амплифицировали при помощи ПЦР с ген-специфическими праймерами. Для обеспечения полуколичественного характера ОТ-ПЦР β-актин использовали в качестве внутреннего контроля для каждой из испытанных тканей. Сначала готовили серийные разведения кДНК первой цепи и анализы ОТ-ПЦР проводили с использованием β-актин-специфических праймеров. Затем выбирали разведение, которое делало возможной амплификацию линейного диапазона матрицы β-актина и которое было достаточно чувствительным для отражения различий в первоначальных копийностях. С использованием этих условий уровни β-актина определяли для каждой реакции обратной транскрипции из каждой ткани. Загрязнение ДНК минимизировали обработкой ДНКазой и обеспечением результата отрицательной ПЦР, когда использовали кДНК первой цепи, которую получали без добавления обратной транскриптазы.
Уровни экспрессии мРНК определяли в четырех различных типах опухолевой ткани (опухоли предстательной железы из 2 пациентов, опухоли молочной железы из 3 пациентов, опухоли ободочной кишки, опухоли легкого) и шестнадцати различных нормальных тканях, в том числе предстательной железе, ободочной кишке, почке, печени, легком, яичнике, поджелудочной железе, скелетной мышце, коже, желудке, яичках, костном мозгу и головном мозгу. Было обнаружено, что F1-16 экспрессировался при высоких уровнях в ткани опухоли предстательной железы, опухоли ободочной кишки и нормальной предстательной железе и при низких уровнях в нормальной печени, коже и яичках, причем экспрессия была неопределяемой в других исследованных тканях. Было обнаружено, что Н1-1 экспрессировался при высоких уровнях в опухоли предстательной железы, опухоли легкого, опухоли молочной железы, нормальной предстательной железе, нормальной ободочной кишке и нормальном головном мозгу, при гораздо более низких уровнях в нормальном легком, поджелудочной железе, скелетной мышце, коже, тонкой кишке, костном мозгу и не определялся в других исследованных тканях. J1-17 (P502S) и L1-12 (P501S), по-видимому, специфически сверхэкспрессировались в предстательной железе, причем оба гена экспрессировались при высоких уровнях в опухоли предстательной железы и нормальной предстательной железе, но при низких - неопределяемых - уровнях во всех других исследованных тканях. Было обнаружено, что N1-1862 (P503S) сверхэкспрессировался в 60% опухолей предстательной железы и был определяемым в нормальной ободочной кишке и почке. Таким образом, результаты ОТ-ПЦР показывают, что F1-16, H1-1, J1-17 (P502S), N1-1862 (P503S) и L1-12 (P501S) либо являются специфическими для предстательной железы, либо экспрессируются при значительно повышенных уровнях в предстательной железе.
Последующие ОТ-ПЦР-исследования показали, что F1-12 (P504S) сверхэкспрессируется в 60% опухолей предстательной железы, является определяемым в нормальной почке, но неопределяемым во всех остальных исследованных тканях. Подобным образом было показано, что R1-2330 сверхэкспрессировался в 40% опухолей предстательной железы, был определяемым в нормальных почке и печени, но не определялся во всех остальных исследованных тканях. Было обнаружено, что U1-3064 сверхэкспрессировался в 60% опухолей предстательной железы и также экспрессировался в опухолях молочной железы и ободочной кишки, но не определялся в нормальных тканях.
ОТ-ПЦР-характеристика R1-2330, U1-3064 и ID-4279 показала, что эти три антигена сверхэкспрессируются в предстательной железе и/или опухолях предстательной железы.
Нозерн-анализ с четырьмя опухолями предстательной железы, двумя образцами нормальной предстательной железы, двумя ВРН-простатами и нормальными ободочной кишкой, печенью, легким, поджелудочной железой, скелетной мышцой, головным мозгом, желудком, яичками, тонкой кишкой и костным мозгом показал, что L1-12 (P501S) сверхэкспрессируется в опухолях предстательной железы и нормальной предстательной железе, тогда как является неопределяемым в других нормальных исследованных тканях. J1-17 (P502S) определялся в двух опухолях предстательной железы, но не в других испытанных тканях. Было обнаружено, что N1-1862 (P503S) сверхэкспрессировался в трех опухолях предстательной железы и экспрессировался в нормальных предстательной железе, ободочной кишке и почке, но не в других исследованных тканях. Было обнаружено, что F1-12 (P504S) экспрессировался на высоком уровне в двух опухолях предстательной железы и был неопределяемым во всех остальных исследованных тканях.
Технологию микроматрицы, описанную выше, использовали для определения уровней экспрессии показательных антигенов, описанных здесь, в опухоли предстательной железы, опухоли молочной железы и следующих нормальных тканях: предстательной железе, печени, поджелудочной железе, коже, костном мозгу, головном мозгу, молочной железе, надпочечнике, мочевом пузыре, яичках, слюнных железах, толстой кишке, почке, яичнике, легком, спинном мозгу, скелетной мышце и ободочной кишке. Было обнаружено, что L1-12 (P501S) сверхэкспрессируется в нормальной предстательной железе и в опухоли предстательной железы и некоторая экспрессия определяется в нормальной скелетной мышце. Было обнаружено, что как J1-12, так и F1-12 (P504S) сверхэкспрессируются в опухоли предстательной железы, причем экспрессия является более низкой или неопределяемой во всех других исследованных тканях. Было обнаружено, что N1-1862 (P503S) экспрессируется при высоких уровнях в опухоли предстательной железы и в нормальной предстательной железе и при низких уровнях в нормальной толстой кишке и нормальной ободочной кишке, причем экспрессия является неопределяемой во всех других исследованных тканях. Было обнаружено, что R1-2330 сверхэкспрессируется в опухоли предстательной железы и нормальной предстательной железе и экспрессируется при более низких уровнях во всех остальных исследованных тканях.
Было обнаружено, что ID-4279 сверхэкспрессируется в опухоли предстательной железы и нормальной предстательной железе, экспрессируется при более низких уровнях в нормальном спинном мозгу и является неопределяемым во всех остальных исследованных тканях.
Дополнительный анализ с использованием микроматрицы для специального определения степени, с которой P501S (SEQ ID NO:110) экспрессируется в опухоли молочной железы, выявил умеренную сверхэкспрессию не только в опухоли молочной железы, но также в метастатической опухоли молочной железы (2/31) с незначительной - низкой - экспрессией в нормальных тканях. Эти данные предполагают, что P501S может сверхэкспрессироваться в различных опухолях молочной железы так же, как в опухолях предстательной железы.
Уровни экспрессии 32 EST (меток экспрессируемых последовательностей), описанных Vasmatzis et al., Proc. Natl. Acad. Sci. USA 95:300-304, 1998), в различных опухолевых и нормальных тканях исследовали с использованием способа микроматрицы, как описано выше. Было найдено, что два из этих клонов (называемые Р1000С и Р1001С) сверхэкспрессровались в опухоли предстательной железы и нормальной предстательной железе и экспрессировались при низких - неопределяемых - уровнях во всех остальных исследованных тканях (нормальной аорте, вилочковой железе, покоящихся и активированных РВМС (мононуклеарных клетках периферической крови), эпителиальных клетках, спинном мозгу, надпочечнике, фетальных тканях, коже, слюнных железах, толстой кишке, костном мозгу, печени, легком, дендритных клетках, желудке, лимфатических узлах, головном мозгу, сердце, тонкой кишке, скелетной мышце, ободочной кишке и почке. Определенные последовательности-кДНК для Р1000С и Р1001С представлены в SEQ ID NO:384 и 472 соответственно. Было обнаружено, что последовательность Р1001С имеет некоторую гомологию с ранее выделенной мРНК человека для белка JM27. Не были обнаружены значимые гомологии относительно последовательности Р1000С.
Экспрессия полипептида, кодируемого полноразмерной последовательностью-кДНК для F1-12 (также называемого P504S; SEQ ID NO:108), исследовали при помощи иммуногистохимического анализа. Кроличьи поликлональные антитела против P504S генерировали против полноразмерного белка P504S при помощи стандартных способов. Последующее выделение и характеристику поликлональных антител также выполняли способами, хорошо известными в данной области. Иммуногистохимический анализ показал, что полипептид P504S экспрессировался в 100% исследованных образцах рака предстательной железы (n=5).
Кроличьи поликлональные антитела против P504S, по-видимому, не метили доброкачественные клетки предстательной железы с тем же самым цитоплазматическим гранулярным окрашиванием, но скорее с легким ядерным окрашиванием. Анализ нормальных тканей выявил, что указанный кодируемый полипептид, как было обнаружено, экспрессируется в некоторых, но не во всех нормальных тканях человека. Положительное цитоплазматическое окрашивание кроличьими поликлональными антителами против P504S было обнаружено в нормальных почке, печени, головном мозгу, ободочной кишке и легочно-ассоциированных макрофагах человека, тогда как сердце и костный мозг были отрицательными.
Эти результаты показывают, что полипептид P504S присутствует в тканях рака предстательной железы и что имеются качественные и количественные различия в окрашивании между тканями доброкачественной гиперплазии предстательной железы и тканями рака предстательной железы, что предполагает, что указанный полипептид может определяться избирательно в опухолях предстательной железы и, следовательно, быть полезным в диагностике рака предстательной железы,
ПРИМЕР 3
ВЫДЕЛЕНИЕ И ХАРАКТЕРИСТИКА ОПУХОЛЕВЫХ ПОЛИПЕПТИДОВ ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ С ИСПОЛЬЗОВАНИЕМ ВЫЧИТАНИЯ НА ОСНОВЕ ПЦР
Являющуюся результатом вычитания библиотеку кДНК, содержащую кДНК из нормальной предстательной железы, полученную вычитанием с кДНК десяти других нормальных тканей (головного мозга, сердца, почки, печени, легкого, яичника, плаценты, скелетной мышцы, селезенки и тимуса) и затем подвергнутую первому циклу ПЦР-амплификации, покупали в Clontech. Эту библиотеку подвергали второму циклу ПЦР-амплификации в соответствии с протоколом изготовителя. Полученные кДНК-фрагменты субклонировали в вектор рТ7 Blue Т-вектор (Novagen, Madison, WI) и трансформировали в Е.coli XL-1 Blue MRF (Stratagene). ДНК выделяли из независимых клонов и секвенировали с использованием автоматизированного секвенатора модели 373А Perkin Elmer/Applied Biosystems Division.
Секвенировали пятьдесят девять положительных клонов. Сравнение последовательностей-ДНК этих клонов с последовательностями в банке генов, как описано выше, не выявило значимых гомологий к 25 из этих клонов, далее называемых Р5, Р8, Р9, Р18, Р20, Р30, Р34, Р36, Р38, Р42, Р49, Р50, Р53, Р55, Р60, Р64, Р65, Р73, Р75, Р76, Р79 и Р84. Определенные последовательности кДНК для этих клонов представлены в SEQ ID NO:41-45, 47-52 и 54-65 соответственно. Было обнаружено, что Р29, Р47, Р68, Р80 и Р82 (SEQ ID NO:46, 53 и 66-68 соответственно) обнаруживают некоторую степень гомологии с ранее идентифицированными последовательностями ДНК. Насколько известно авторам изобретения, ни одна из этих последовательностей не была описана ранее в качестве присутствующей в предстательной железе.
Дальнейшие исследования с использованием методологии на основе ПЦР, описанной выше, привели к выделению более 180 дополнительных клонов, из которых, как было обнаружено, 23 клона не имели значимых гомологий с известными последовательностями. Определенные последовательности-кДНК для этих клонов представлены в SEQ ID NO:115-123, 127, 131, 137, 145, 147-151, 153, 156-158 и 160. Было обнаружено, что двадцать три клона (SEQ ID NO:124-126, 128-130, 132-136, 138-144, 146, 152, 154, 155 и 159) обнаруживают некоторую гомологию с ранее идентифицированными EST. Было найдено, что дополнительные десять клонов (SEQ ID NO:161-170) имеют некоторую степень гомологии с известными генами. Более крупные кДНК-клоны, содержащие последовательность Р20, представляют сплайсинговые варианты гена, называемого P703S. Определенные последовательности ДНК для этих вариантов, называемых DE1, DE13 и DE14, представлены в SEQ ID NO:171, 175 и 177 соответственно с соответствующими предсказанными аминокислотными последовательностями, представленными в SEQ ID NO:172, 176 и 178 соответственно. Определенная последовательность-кДНК для удлиненной сплайсинговой формы P703S представлена в SEQ ID NO:225. Последовательности-ДНК для сплайсинговых вариантов, называемых DE2 и DE6, представлены в SEQ ID NO:173 и 174 соответственно.
Уровни экспрессии мРНК для показательных клонов в опухолевых тканях (предстательной железы (n=5), молочной железы (n=2), ободочной кишки и легкого), нормальных тканях (предстательной железы (n=5), ободочной кишки, почки, печени, легкого (n=2), яичника (n=2), скелетной мышцы, кожи, желудка, тонкой кишки и головного мозга) и активированных и не активированных РВМС определяли при помощи ОТ-ПЦР, как описано выше. Экспрессию определяли в одной пробе каждого типа ткани, если нет других указаний.
Было обнаружено, что Р9 экспрессируется в высокой степени в нормальной предстательной железе и опухоли предстательной железы в сравнении со всеми испытанными нормальными тканями за исключением нормальной ободочной кишки, показавшей сравнимую экспрессию. Было найдено, что Р20, часть гена P703S, экспрессируется в высокой степени в нормальной предстательной железе и опухоли предстательной железы в сравнении со всеми двенадцатью исследованными нормальными тканями. Умеренное увеличение экспрессии Р20 в опухоли молочной железы (n=2), опухоли ободочной кишки и опухоли легкого наблюдали в сравнении со всеми испытанными нормальными тканями за исключением легкого (1 из двух) Увеличенная экспрессия Р18 была обнаружена в нормальной предстательной железе, опухоли предстательной железы и опухоли молочной железы в сравнении с другими нормальными тканями за исключением легкого и желудка. Умеренное увеличение экспрессии Р5 наблюдали в нормальной предстательной железе в сравнении с большинством других нормальных тканей. Однако некоторое повышение экспрессии наблюдали в нормальном легком и РВМС. Повышенную экспрессию Р5 также наблюдали в опухолях предстательной железы (2 из 5), молочной железе и пробе одной опухоли легкого. Для Р30 наблюдали одинаковые уровни экспрессии в нормальной предстательной железе и опухоли предстательной железы в сравнении с шестью из двенадцати других исследованных нормальных тканей. Увеличенную экспрессию наблюдали в опухолях молочной железы, образце одной опухоли легкого и образце одной опухоли ободочной кишки, а также в нормальных РВМС. Было найдено, что Р29 сверхэкспрессировался в опухоли предстательной железы (5 из 5) и нормальной предстательной железе (5 из 5) в сравнении с большинством нормальных тканей. Однако существенную экспрессию Р29 наблюдали в нормальной ободочной кишке и нормальном легком (2 из 2). Было обнаружено, что Р80 сверхэкспрессировался в опухоли предстательной железы (5 из 5) и нормальной предстательной железе (5 из 5) в сравнении со всеми другими исследованными нормальными тканями, причем увеличенную экспрессию наблюдали также в опухоли ободочной кишки.
Дальнейшие исследования привели к выделению двенадцати дополнительных клонов, далее называемых 10-d8, 10-h10, 11-с8, 7-g6, 8-b5, 8-b6, 8-d4, 8-d9, 8-g3, 8-h11, 9-f12 и 9-f3. Определенные последовательности-ДНК для 10-d8, 10-h10, 11-с8, 8-d4, 8-d9, 8-h11, 9-f12 и 9-f3 представлены в SEQ ID NO:207, 208, 209, 216, 217, 220, 221 и 222 соответственно. Определенные прямые и обратные последовательности-ДНК для 7-g6, 8-b5, 8-b6 и 8-g3 представлены в SEQ ID NO:210 и 211, 212 и 213, 214 и 215 и 218 и 219 соответственно. Сравнение этих последовательностей с последовательностями в банке генов не выявило значимых гомологий к последовательности 9-f3. Было обнаружено, что клоны 10-d8, 11-c8 и 8-g11 обнаруживают некоторую гомологию с ранее выделенными EST, тогда как, как было показано, 10-h10, 8-b5, 8-b6, 8-d4, 8-d9, 8-g3 и 9-f12 обнаруживают некоторую гомологию с ранее идентифицированными генами. Дополнительная характеристика 7-G6 и 8-G3 показала идентичность с известными генами РАР и PSA соответственно.
Уровни экспрессии мРНК для этих клонов определеляли с использованием способа микроматрицы, описанного выше. Было обнаружено, что клоны 7-G6, 8-G3, 8-В5, 8-B6, 8-D4, 8-D9, 9-F3, 9-F12, 9-Н3, 10-А2, 10-А4, 11-С9 и 11-F2 сверхэкспрессировались в опухоли предстательной железы и нормальной предстательной железе, причем экспрессия в других испытанных тканей была низкой или неопределяемой. Увеличенную экспрессию 8-F11 наблюдали в опухоли предстательной железы и нормальной предстательной железе, мочевом пузыре, скелетной мышце и ободочной кишке. Увеличенную экспрессию 10-Н10 наблюдали в опухоли предстательной железы и нормальной предстательной железе, мочевом пузыре, легком, ободочной кишке, головном мозгу и толстой кишке. Увеличенную экспрессию 9-В1 наблюдали в опухоли предстательной железы, опухоли молочной железы и нормальной предстательной железе, слюнных железах, толстой кишке и коже, а увеличенную экспрессию 11-C8 наблюдали в опухоли предстательной железы и нормальной предстательной железе и толстой кишке.
Было обнаружено, что дополнительный кДНК-фрагмент, полученный из вычитания на основе ПЦР нормальной предстательной железы, описанного выше, является специфическим для предстательной железы согласно как способу микроматрицы, так и ОТ-ПЦР. Определенная последовательность-кДНК этого клона (называемого 9-А11) представлена в SEQ ID NO:226. Сравнение этой последовательности с последовательностями в общественных базах данных выявило 99% идентичность с известным геном НОХВ13.
Дальнейшие исследования привели к выделению клонов 8-С 6 и 8-Н7. Определенные последовательности-кДНК для этих клонов представлены в SEQ ID NO:227 и 228 соответственно. Было обнаружено, что эти последовательности имеют некоторую гомологию с ранее выделенными EST.
Методологии на основе ПЦР и на основе гибридизации использовали для получения более длинных последовательностей-кДНК для клона Р20 (также называемого Р703Р), что привело к получению трех дополнительных кДНК-фрагментов, которые прогрессивно удлиняют 5’-конец этого гена. Эти фрагменты, называемые P703PDE5, Р703Р6.26 и Р703РХ-23 (SEQ ID NO:326, 328 и 330 с предсказанными аминокислотными последовательностями, представленными в SEQ ID NO:327, 329 и 331 соответственно), содержат дополнительную 5’-последовательность. P703PDE5 извлекали скринингом библиотеки кДНК (№141-26) с частью Р703Р в качестве зонда. Р703Р6.26 извлекали из смеси трех кДНК опухоли предстательной железы и Р703РХ-23 извлекали из библиотеки кДНК (№438-48). Вместе эти дополнительные последовательности все включают в себя возможную зрелую сериновую протеазу вместе с частью возможной сигнальной последовательности. Дальнейшие исследования с использованием библиотеки, полученной вычитанием на основе ПЦР, пула опухолей предстательной железы, подвергнутого вычитанию против пула нормальных тканей (называемому JP: ПЦР-вычитанием), привели к выделению тринадцати дополнительных клонов, семь из которых не имели какой-либо значимой гомологии с известными последовательностями GenBank. Определенные последовательности-кДНК для этих семи клонов (Р711Р, Р712Р, новый 23, Р774Р, Р775Р, Р710Р и Р768Р) представлены в SEQ ID NO:307-311, 313 и 315 соответственно. Было показано, что остальные шесть клонов имеют некоторую гомологию с известными генами. Согласно анализу с использованием микроматрицы все тринадцать клонов обнаружили трехкратную или более высокую сверхэкспрессию в тканях предстательной железы в том числе в опухолях предстательной железы, ВРН и нормальной предстательной железе в сравнении с нормальными не относящимися к предстательной железе тканями. Клоны Р711Р, Р712Р, новый 23 и Р768Р обнаружили сверхэкспрессию в большинстве испытанных опухолей предстательной железы и тканях ВРН (n=29) и в большинстве нормальных тканей предстательной железы (n=4), но показали уровни фона или уровни низкой экспрессии во всех нормальных тканях.
Клоны Р774Р, Р775Р и Р710Р показали сравнительно более низкую экспрессию и экспрессию в меньшем числе опухолей предстательной железы и проб ВРН, с отрицательной - низкой - экспрессией в нормальной предстательной железе.
Полноразмерную кДНК для Р711Р получали путем применения частичной последовательности SEQ ID NO:307 для скрининга библиотеки кДНК предстательной железы. Конкретно, направленно клонированную библиотеку кДНК предстательной железы получали с использованием стандартных способов. Один миллион колоний этой библиотеки высевали на LB/Аmр-чашки. Найлоновые мембранные фильтры использовали для подъема этих колоний и кДНК, которые извлекались этими фильтрами, денатурировали и сшивали с фильтрами при помощи УФ-света. Р711Р-фрагмент кДНК SEQ ID NO:307 радиоактивно метили и использовали для гибридизации с этими фильтрами. Положительные клоны отбирали и кДНК получали и секвенировали с использованием автоматического секвенатора Perkin Elmer/Applied Biosystems. Определенная полноразмерная последовательность Р711Р представлена в SEQ ID NO:382 с соответствующей предсказанной аминокислотной последовательностью, представленной в SEQ ID NO:383.
С использованием методологий на основе ПЦР и на основе гибридизации дополнительную информацию о последовательностях кДНК получали для двух вышеописанных клонов, 11-С9 и 9-F3, далее называемых Р707Р и Р714Р соответственно (SEQ ID NO:333 и 334). После сравнения с самым последним банком генов было обнаружено, что Р707Р является сплайсинговым вариантом известного гена НохВ13. В противоположность этому не было обнаружено значимых гомологий к Р714Р.
Было найдено, что клоны 8-В3, Р89, Р98, Р130 и Р201 (описанные в патентной заявке Соединенных Штатов № 09020956, поданной 9 февраля 1998 года) содержатся в одной непрерывной последовательностии, называемой Р705Р (SEQ ID NO:335, с предсказанной аминокислотной последовательностью, представленной в SEQ ID NO:336), которая, как было определено, является сплайсинговым вариантом известного гена NKX3.1.
ПРИМЕР 4
СИНТЕЗ ПОЛИПЕПТИДОВ
Полипептиды могут быть синтезированы на пептидном синтезаторе 430А Perkin Elmer/Applied Biosystems с использованием FMOC-химии с HPTU-активацией (активацией гексафторфосфатом О-бензотриазол-N,N,N’,N’-тетраметилурония). Последовательность Gly-Cys-Gly может быть присоединена к амино-концу этого пептида для обеспечения способа конъюгации, связывания с иммобилизованной поверхностью или мечения пептида. Отщепление этих пептидов от твердого носителя может проводиться с использованием следующей отщепляющей смеси: трифторуксусная кислота : этандитиол : тиоанизол : вода : фенол (40:1:2:2:3). После отщепления в течение 2 часов пептиды могут быть осаждены в холодном метил-трет-бутиловом эфире. Затем осадки пептидов могут быть растворены в воде, содержащей 0,1% трифторуксусную кислоту (ТФУ), и лиофилизированы перед очисткой обращенно-фазовой ВЖХ на С18. Градиент 0-60% ацетонитрила (содержащего 0,1% ТФУ) в воде (содержащей 0,1% ТФУ) может быть использован для элюции пептидов. После лиофилизации чистых фракций пептиды могут быть охарактеризованы с использованием электрораспылительной или других типов масс-спектрометрии и аминокислотного анализа.
ПРИМЕР 5
ДОПОЛНИТЕЛЬНОЕ ВЫДЕЛЕНИЕ И ХАРАКТЕРИСТИКА ОПУХОЛЕВЫХ ПОЛИПЕПТИДОВ ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ ПРИ ПОМОЩИ ВЫЧИТАНИЯ НА ОСНОВЕ ПЦР
Библиотеку кДНК, генерированную из мРНК первичной опухоли предстательной железы, как описано выше, вычитали с кДНК из нормальной предстательной железы. Вычитание проводили с использованием протокола на основе ПЦР (Clon-tech), который был модифицирован для генерирования более крупных фрагментов. В этом протоколе тестируемую и драйверную двухцепочечную кДНК отдельно расщепляли пятью рестрикционными ферментами (рестриктазами), которые узнают состоящие из шести нуклеотидов сайты рестрикции (MluI, MscI, PvuII, SalI и StuI). Это расщепление приводило к среднему размеру кДНК 600 п.н., а не к среднему размеру 300 п.н., который достигается при расщеплении RsaI согласно протоколу Clontech. Эта модификация не влияла на эффективность вычитания. Затем создавали две различные тестируемые популяции с различными адаптерами, а драйверную библиотеку оставляли без адаптеров.
Затем тестируемую библиотеку и драйверную библиотеку гибридизовали с использованием избытка драйвер-кДНК. В первой стадии гибридизации драйвер отдельно гибридизовали с каждой из двух тестируемых популяций кДНК. Это приводило к популяциям (а) негибридизованных тестируемых кДНК, (b) тестируемых кДНК, гибридизованных с другими тестируемыми кДНК, (с) тестируемых кДНК, гибридизованных с драйвер-кДНК, и (d) негибридизованных драйвер-кДНК. Затем эти две отдельные реакции гибридизации объединяли и повторно гибридизовали в присутствии дополнительной денатурированной драйвер-кДНК. После этой второй гибридизации кроме популяций (а)-(d), получали пятую популяцию (е), в которой тестируемая кДНК с одним адаптером гибридизовалась с тестируемой ДНК со вторым адаптером. Вследствие этого вторая гибридизация приводила к обогащению дифференциально экспрессируемых последовательностей, которые могли бы использоваться в качестве матриц для ПЦР-амплификации с адаптер-специфическими праймерами.
Затем концы достраивали и ПЦР-амплификацию проводили с использованием адаптер-специфических праймеров. Только популяция (е), которая содержала тестируемую ДНК, которая не гибридизовалась с драйвер-кДНК, амплифицировалась экспоненциально. Затем проводили вторую стадию ПЦР-амплификации для уменьшения фона и дополнительного обогащения дифференциально экспрессируемых последовательностей.
Указанный способ вычитания на основе ПЦР нормализует дифференциально экспрессируемые кДНК таким образом, что могут быть извлечены редкие (необычные) транскрипты, которые сверхэкспрессируются в ткани опухоли предстательной железы. Такие транскрипты было бы трудно извлечь традиционными способами вычитания.
Кроме генов, о которых известно, что они сверхэкспрессируются в опухоли предстательной железы, были идентифицированы семьдесят семь дополнительных клонов. Последовательности этих частичных кДНК представлены в SEQ ID NO:29-305. Большинство этих клонов не имели значимой гомологии с последовательностями баз данных. Исключениями были JPTPN23 (SEQ ID NO:231; сходство с валозин-содержащим белком свиньи), JPTPN30 (SEQ ID NO:234; сходство с мРНК субъединицы протеасомы крысы), JPTPN45 (SEQ ID NO:243; сходство с цитозольной НАДФ-зависимой изоцитратдегидрогеназой крысы norvegicus), JPTPN46 (SEQ ID NO:244; сходство с ДНК-последовательностью субклона человека Н8 4 d4), JP1D6 (SEQ ID NO:265; сходство с легкой цепью А динеина G.gallus), JP8D6 (SEQ ID NO:288; сходство с клоном RG016J04 ВАС человека), JP8F5 (SEQ ID NО:289; сходство с ДНК-последовательностью субклона Н8 3 b5 человека) и JP8E9 (SEQ ID NO:299; сходство с последовательностью Alu человека).
Дополнительные исследования с использованием библиотеки вычитания на основе ПЦР, состоящей из пула опухоли предстательной железы, подвергнутого вычитанию против пула нормальной предстательной железы (называемому ПО-ПН ПЦР-вычитанием) давало три дополнительных клона. Сравнение последовательностей-кДНК этих клонов с самым последним банком генов не выявило значимых гомологий к этим двум клонам, названным Р715Р и Р767Р (SEQ ID NO:312 и 314). Оставшийся клон, как было обнаружено, показал некоторую гомологию с известным геном KIAA0056 (SEQ ID NO:318). С использованием анализа микроматриц для измерения уровней экспрессии мРНК в различных тканях было обнаружено, что все три клона сверхэкспрессируются в опухолях предстательной железы и в тканях ВРН. Конкретно, клон Р715Р сверхэкспрессировался в большинстве опухолей предстательной железы и в тканях ВРН с коэффициентом 3 или более, причем повышенная экспрессия наблюдалась в большинстве проб нормальной предстательной железы и фетальной ткани, но экспрессия была отрицательной - низкой - во всех других нормальных тканях. Клон Р767Р сверхэкспрессировался в нескольких опухолях предстательной железы и тканях ВРН с умеренными уровнями экспрессии в половине проб нормальной предстательной железы, и уровни экспрессии были равны фону или были низкими во всех других исследованных нормальных тканях.
Дополнительный анализ при помощи вышеописанной микроматрицы, библиотеки вычитания ПО-ПН ПЦР и библиотеки вычитания ДНК, содержащей кДНК из опухоли предстательной железы, подвергнутую вычитанию с пулом кДНК нормальной ткани, привел к выделению 27 дополнительных клонов (SEQ ID NO:340-365 и 381), которые, как было определено, сверхэкспрессировались в опухоли предстательной железы. Было обнаружено, что клоны SEQ ID NО:341, 342, 345, 347, 348, 349, 351, 355-359, 361, 362 и 364 сверхэкспрессируются в нормальной предстательной железе. Было найдено, что экспрессия всех 26 клонов в различных нормальных тканях была низкой или неопределяемой за исключением P544S (SEQ ID NО:356), который, как было обнаружено, экспрессировался в тонкой кишке. Было обнаружено, что из этих 26 клонов 10 (SEQ ID NO:340-349) обнаруживают некоторую гомологию с ранее идентифицированными последовательностями. Не были обнаружены значимые гомологии к клонам SEQ ID NO:350-365.
ПРИМЕР 6
ПЕПТИДНОЕ ПРАЙМИРОВАНИЕ МЫШЕЙ И РАЗМНОЖЕНИЕ ЛИНИЙ ЦТЛ
6.1. Указанный пример иллюстрирует получение линии клеток ЦТЛ (цитотоксических Т-лимфоцитов), специфической в отношении клеток, экспрессирующих ген P502S.
Мышей, экспрессирующих трансген для HLA A2.1 человека (предоставленный доктором L.Sherman, The Scripps Research Institute, La Jolla, CA), иммунизировали пептидом P2S#12 (VLGWVAEL; SEQ ID NO:306), который был получен из гена P502S (также называемого здесь J1-17, SEQ ID NO:8), как описано Theobald et al., Proc. Natl. Acad. Sci. USA 92:11993-11997, 1995) со следующими модификациями. Мышей иммунизировали 100 мкг P2S#12 и 120 мкг связывающего пептида I-Ab, произведенного из белка вируса гепатита В, эмульгированного в неполном адъюванте Фрейнда. Спустя три недели мышей умерщвляли и получали суспензии одиночных клеток с использованием найлонового сита. Затем клетки ресуспендировали при 6×106 клеток/мл в полной среде (RPMI-1640; Gibco BRL, Gaithersburg, MD), содержащей 10% ФТС, 2 мМ глутамин (Gibco BRL), пируват натрия (Gibco BRL), ненезаменимые аминокислоты (Gibco BRL), 2×10-5 М 2-меркаптоэтанол, 50 Е/мл пенициллина и стрептомицина, и культивировали в присутствии облученных (3000 рад) кратковременно обработанных Р2S#12 (5 мг/мл P2S#12 и 10 мг/мл β2-микроглобулина) ЛПС-бластов (трансгенных клеток селезенки А2, культивируемых в присутствии 7 мкг/мл сульфата декстрана и 25 мкг/мл ЛПС в течение 3 дней). Спустя 6 дней клетки (5×105 клеток/мл) повторно стимулировали (2/5×106 клеток/мл) облученными (20000 рад) кратковременно обработанными пептидом клетками EL4A2Kb (Sherman et al., Science 258:815-818, 1992) и 3×106 клеток на мл питающими трансгенными клетками селезенки А2 один раз в неделю, как описано, в подготовке для клонирования этой линии.
Линию P2S#12 клонировали с использованием анализа лимитирующего разведения с кратковременно обработанными пептидом опухолевыми клетками EL4 А2Кb (1×104 клеток на лунку) в качестве стимуляторов и трансгенными клетками А2 селезенки в качестве клеток-фидеров (питающих клеток) (5×105 клеток на лунку), выращиваемыми в присутствии 30 Е/мл IL-2. В день 14 клетки повторно стимулировали, как описано ранее. В день 21 клоны, которые росли, выделяли и поддерживали в культуре. Некоторые из этих клонов продемонстрировали значимо более высокую реактивность (лизис) против фибробластов человека (экспрессирующих HLA А2.1), трансдуцированных P502S, в сравнении с контрольными фибробластами. Пример представлен на фигуре 1.
Эти данные показывают, что P2S#l2 представляет природно процессируемый эпитоп белка P502S, который экспрессируется в контексте молекулы HLA A2.1 человека.
6.2. Указанный пример иллюстрирует получение мышиных линий ЦТЛ и ЦТЛ-клонов, специфических в отношении клеток, экспрессирующих ген P501S.
Эту серию экспериментов проводили подобно серии, описанной выше. Мышей иммунизировали пептидом P1S#10 (SEQ ID NO:337), который был получен из гена P501S (также называемого здесь L1-12, SEQ ID NO:110). Пептид P1S#10 был произведен анализом предсказанной полипептидной последовательности для P505S на потенциальные HLA-A2-связывающие последовательности, определяемые при помощи опубликованных мотивов НLА-А2-связывания (Parker, КС, et al., J.Immunol., 152:163, 1994). Пептид P1S#10 синтезировали, как описано в примере 4, и эмпирически тестировали на HLA-A2-связывание с использованием конкурентного анализа на основе Т-клеток. Предсказанные А2-связывающие пептиды испытывали на их способность конкурировать за представление HLA-А2-специфического пептида НLА-А2-рестриктированному ЦТЛ-клону (D150M58), который является специфическим в отношении НLА-А2-связывающего пептида fluM58 матрикса вируса гриппа. D150М58-ЦТЛ секретирует TNF в ответ на самопредставление пептида fluM58. В этом конкурентном анализе тест-пептиды в количестве 100-200 мкг/мл добавляли в культуры 0150М58-ЦТЛ для связывания HLA-A2 на этих ЦТЛ. Спустя тридцать минут ЦТЛ, культивируемые с тест-пептидами или контрольными пептидами, испытывали на их зависимость ответа от дозы антигена в отношении пептида fluM58 в стандартном биотесте на TNF. Как показано на фигуре 3, пептид P1S#10 конкурирует за HLA-A2-рестриктированное представление fluM58, демонстрируя, что пептид Р1S#10 связывает HLA-A2.
Мышей, экспрессирующих трансген для HLA-A2 человека, иммунизировали, как описано Theobald et al., Proc. Natl. Acad. Sci. USA 92:11993-11997, 1995) со следующими модификациями. Мышей иммунизировали 62,5 мкг P1S#10 и 120 мкг связывающего пептида I-Ab, произведенного из белка вируса гепатита В, эмульгированного в неполном адъюванте Фрейнда. Спустя три недели мышей умерщвляли и получали суспензии одиночных клеток с использованием найлонового сита. Затем клетки ресуспендировали при 6×106 клеток/мл в полной среде (описанной выше) и культивировали в присутствии облученных (3000 рад) кратковременно обработанных P1S#10 (2 мг/мл P1S#10 и 10 мг/мл β2-микроглобулина) ЛПС-бластов (трансгенных клеток селезенки А2, культивируемых в присутствии 7 мкг/мл сульфата декстрана и 25 мкг/мл ЛПС в течение 3 дней). Спустя 6 дней клетки (5×105 клеток/мл) повторно стимулировали (2,5×106 клеток/мл) облученными (20000 рад) кратковременно обработанными пептидом клетками EL4A2Kb, как описано выше, и 3×106 клеток на мл питающими трансгенными клетками селезенки А2. Клетки культивировали в присутствии 20 Е/мл IL-2. Клетки повторно стимулировали один раз в неделю в подготовке для клонирования. После трех циклов стимуляций in vitro была получена одна линия, которая узнавала обработанные кратковременно P1S#10 клетки-мишени Jurkat A2Kb и трансдуцированные P501S мишени Jurkat, как показано на фигуре 4.
P1S#10-специфическую ЦТЛ-линию клонировали с использованием анализа лимитирующего разведения с обработанными кратковременно пептидом опухолевыми клетками EL4 A2Kb (1×104 клеток на лунку) в качестве стимуляторов и трансгенными клетками селезенки А2 в качестве фидеров (питающих клеток) (5×105 клеток на лунку), выращенных в присутствии 30 Е/мл IL-2. В день 14 клетки повторно стимулировали, как и прежде. В день 21 жизнеспособные клоны выделяли и поддерживали в культуре. Как показано на фигуре 5, пять из этих клонов продемонстрировали специфическую цитолитическую реактивность против трансдуцированных P501S мишеней Jurkat A2Kb. Эти результаты показывают, что P1S#10 представляет природно процессируемый эпитоп белка P501S, который экспрессируется в контексте молекулы HLA-A2.1 человека.
ПРИМЕР 7
СПОСОБНОСТЬ Т-КЛЕТОК ЧЕЛОВЕКА УЗНАВАТЬ ОПУХОЛЕВЫЕ ПОЛИПЕПТИДЫ ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ
Указанный пример иллюстрирует способность Т-клеток, специфических в отношении опухолевого полипептида предстательной железы, узнавать опухоль человека.
CD8+ Т-клетки человека праймировали in vitro пептидом P2S-12 (SEQ ID NO:306), произведенным из P502S (также называемого J1-17), с использованием дендритных клеток в соответствии с протоколом Van Tsai et al. (Critical Reviews in Immunology 18:65-75, 1998). Полученные микрокультуры CD8+ Т-клеток испытывали на их способность узнавать пептид P2S-12, представляемый аутологичными фибробластами или фибробластами, которые были трансдуцированы для экспрессии гена P502S, в анализе ELISPOT с γ-интерфероном (Lalvani et al., J. Exp. Med. 186:859-865, 1997). Вкратце, титрующиеся количества Т-клеток анализировали в двух повторностях на 104 фибробластах в присутствии 3 мкг/мл β2-микроглобулина человека и 1 мкг/мл пептида P2S-12 или контрольного пептида Е75. Кроме того, Т-клетки одновременно анализировали на аутологичных фибробластах, трансдуцированных геном P502S, или, в качестве контроля, фибробластах, трансдуцированных HER-2/neu. Перед анализом фибробласты обрабатывали 10 нг/мл γ-интерферона в течение 48 часов для положительной регуляции экспрессии МНС класса I. Одна из этих микрокультур (№5) продемонстрировала сильное узнавание обоих обработанных кратковременно пептидом фибробластов, а также трансдуцированных фибробластов в анализе ELISPOT с γ-интерфероном. Фигура 2А демонстрирует, что было сильное увеличение числа пятен γ-интерферона с увеличением количеств Т-клеток на фибробластах, кратковременно обработанных пептидом P2S-12 (сплошные столбцы), но не контрольным пептидом Е75 (белые столбцы). Это показывает способность этих Т-клеток специфически узнавать пептид P2S-12. Как показано на фигуре 2В, эта микрокультура показала также увеличение числа пятен γ-интерферона с увеличением количеств Т-клеток на фибробластах, трансдуцированных для экспрессии гена P502S, но не гена HER-2/neu. Эти результаты обеспечивают дополнительное подтверждающее доказательство, что пептид P2S-12 является природно процессируемым эпитопом белка P502S. Кроме того, это также показывает, что в популяции Т-клеток человека существуют высокоаффинные Т-клетки, которые способны узнавать указанный эпитоп. Эти Т-клетки должны также быть способными узнавать опухоли человека, экспрессирующие ген P502S.
ПРИМЕР 8
ПРАЙМИРОВАНИЕ ЦТЛ in vivo ПУТЕМ ИММУНИЗАЦИИ ДЕПРОТЕИНИЗИ-РОВАННОЙ ДНК С АНТИГЕНОМ ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ
Опухолевый антиген предстательной железы L1-12, описанный выше, называют также P501S. Мышей HLA А2Кb Тg (предоставленных доктором L.Sherman, The Scipps Research Institute, La Jolla; CA) иммунизировали 100 мкг VR10132-P501S либо внутримышечно, либо внутрикожно. Мышей иммунизировали три раза с двухнедельным интервалом между иммунизациями. Через две недели после последней иммунизации иммунные клетки селезенки культивировали с трансдуцированными стимуляторными клетками Jurkat A2Kb-P501S. ЦТЛ-линии стимулировали один раз в неделю. После двух недель стимуляции in vitro оценивали активность ЦТЛ против трансдуцированных P501S мишеней. Две из 8 мышей развили сильные ответы ЦТЛ против P501S. Эти результаты показывают, что P501S содержит по меньшей мере один природно процессируемый А2-рестриктированный эпитоп ЦТЛ.
ПРИМЕР 9
ГЕНЕРИРОВАНИЕ ЦТЛ ЧЕЛОВЕКА in vitro ПОСРЕДСТВОМ ПРАЙМИРОВАНИЯ ЦЕЛЫМ ГЕНОМ И СПОСОБЫ СТИМУЛЯЦИИ ОПУХОЛЕВЫМ АНТИГЕНОМ ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ
Посредством праймирования целым геном in vitro с трансдуцированными P501S (в ретровирусном векторе) аутологичными фибробластами (например, Yee et al., The Journal of Immunology, 157 (9):4079-86, 1996) получали линии ЦТЛ человека, которые специфически узнают аутологичные фибробласты, трансдуцированные P501S (также известным как L1-12), как определено при помощи анализа ELISPOT с γ-интерфероном. С использованием панели с HLA-несовместимыми фибробластными линиями, трансдуцированными P501S, было показано, что эти линии ЦТЛ рестриктированы аллелем HLA-A2 класса I. Конкретно, дендритные клетки (ДК) дифференцировались из культур моноцитов, полученных из мононуклеарных клеток периферической крови (РВМС) нормальных доноров человека путем выращивания в течение пяти дней в среде RPMI, содержащей 10% сыворотку человека, 50 нг/мл GM-CSF человека и 30 нг/мл IL-4 человека. После культивирования ДК инфицировали в течение ночи рекомбинантным содержащим P501S вирусом коровьей оспы при множественности заражения (MOI) пять, и они созревали в течение ночи под действием 3 мкг/мл лиганда CD40. Вирус инактивировали облучением УФ. CD8+ Т-клетки выделяли с использованием системы с магнитными гранулами и культуры для праймирования инициировали с использованием стандартных способов культивирования. Культуры повторно стимулировали каждые 7-10 дней с использованием аутологичных первичных фибробластов, трансдуцированных при помощи ретровирусного вектора P501S. После четырех циклов стимуляции идентифицировали линии CD8+ Т-клеток, которые специфически продуцировали интерферон-γ при стимуляции Р501S-трансдуцированными аутологичными фибробластами. Р501S-специфическая активность могла поддерживаться продолжающейся стимуляцией культур Р501S-трансдуцированными фибробластами в присутствии IL-15. Панель HLA-несовместимых линий фибробластов, трансдуцированных P501S, генерировали для определения аллеля рестрикции (ограничения) этого ответа. Путем измерения интерферона-γ в анализе ELISPOT было показано, что P501S-специфическая ответная реакция была рестриктирована (ограничена) HLA-A2. Эти результаты показывают, что CD8+ ЦТЛ-ответ на P501S может быть индуцирован.
ПРИМЕР 10
ИДЕНТИФИКАЦИЯ ПРИРОДНО ПРОЦЕССИРУЕМОГО ЭПИТОПА ЦТЛ, СОДЕРЖАЩЕГОСЯ В АНТИГЕНЕ ОПУХОЛИ ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ
9-мерный пептид р5 (SEQ ID NO:338) получали из антигена Р703Р (также называемого Р20). Пептид р5 является иммуногенным в донорах HLA-A2 и является природно процессируемым эпитопом. Антиген-специфические CD8+ Т-клетки могут быть праймированы после повторяемых стимуляций in vitro моноцитами, кратковременно обработанными пептидом р5. Эти ЦТЛ специфически узнают обработанные кратковременно пептидом р5 клетки-мишени как в анализе ELISPOT, так и в анализе с высвобождением хрома. Кроме того, иммунизация HLA-A2 трансгенных мышей с P509S приводит к генерированию линий ЦТЛ, которые узнают различные Р703Р-трансдуцированные клетки-мишени, экспрессирующие либо HLA-A2Kb, либо HLA-A2. Конкретно, HLA-A2-трансгенных мышей иммунизировали подкожно в подушечку лапки 100 мкг пептида р5 вместе с 140 мкг корового пептида вируса гепатита В (пептида Th) в неполном адъюванте Фрейнда. Через три недели после иммунизации клетки селезенки из иммунизированных мышей стимулировали in vitro ЛПС-бластами, кратковременно обработанными пептидом. Активность ЦТЛ оценивали анализом высвобождения хрома через пять дней после первичной стимуляции in vitro. Ретровирусно трансдуцированные клетки, экспрессирующие контрольный антиген P703S и HLA-A2, использовали в качестве мишеней. Линии ЦТЛ, которые специфически узнавали как кратковременно обработанные P509S мишени, так и экспрессирующие P703S мишени, были идентифицированы.
Эксперименты по праймированию in vitro человека показали, что пептид р5 является иммуногенным в человеке. Дендритные клетки (ДК) дифференцировали из культур моноцитов, полученных из мононуклеарных клеток периферической крови (РВМС) нормальных доноров человека путем культивирования в течение пяти дней в среде RPMI, содержащей 10% сыворотку человека, 50 нг/мл GM-CSF человека и 30 нг/мл IL-4 человека. После культивирования ДК обрабатывали кратковременно пептидом р5 и культивировали с GM-CSF и IL-4 вместе с обогащенными CD8+ Т-клетками РВМС. Линии ЦТЛ повторно стимулировали один раз в неделю кратковременно обработанными р5 моноцитами. Через пять-шесть недель после инициации культур ЦТЛ было продемонстрировано узнавание ЦТЛ кратковременно обработанных пептидом р5 клеток-мишеней.
ПРИМЕР 11
ЭКСПРЕССИЯ ПОЛУЧЕННОГО ИЗ ОПУХОЛИ МОЛОЧНОЙ ЖЕЛЕЗЫ АНТИГЕНА В ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЕ
Выделение антигена B305D из опухоли молочной железы посредством дифференциального изображения описано в патентной заявке США № 08/700014, поданной 20 августа 1996 года. Несколько различных сплайсинговых форм указанного ангтигена были выделены. Определенные последовательности-ДНК для этих сплайсинговых форм представлены в SEQ ID NO:366-375 с предсказанными аминокислотными последовательностями, соответствующими последовательностям SEQ ID NO:292, 298 и 301-303, представленными в SEQ ID NO:299-306 соответственно.
Уровни экспрессии B305D в различных опухолевых и нормальных тканях исследовали ПЦР реального времени и Нозерн-анализом. Результаты показали, что B305D экспрессируется на высоком уровне в опухоли молочной железы, опухоли предстательной железы, нормальной предстательной железе и нормальных яичках, причем экспрессия является низкой или неопределяемой во всех других испытанных тканях (опухоли ободочной кишки, опухоли легкого, опухоли яичника и нормальном костном мозгу, ободочной кишке, почке, печени, легком, яичнике, коже, тонкой кишке, желудке).
ПРИМЕР 12
ВЫЗЫВАНИЕ СПЕЦИФИЧЕСКИХ ДЛЯ АНТИГЕНА ОПУХОЛИ ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ ЦТЛ-ОТВЕТОВ В КРОВИ ЧЕЛОВЕКА
Указанный пример иллюстрирует способность опухолевого антигена предстательной железы вызывать ЦТЛ-ответ в крови здоровых людей.
Аутологичные дендритные клетки (ДК) дифференцировали из культур моноцитов, полученных из мононуклеарных клеток периферической крови (РВМС) нормальных доноров человека путем выращивания в течение пяти дней в среде RPMI, содержащей 10% сыворотку человека, 50 нг/мл GM-CSF человека и 30 нг/мл IL-4 человека. После культивирования ДК инфицировали в течение ночи рекомбинантным экспрессирующим P501S вирусом коровьей оспы при множественности заражения (MOI) 5 и они созревали в течение 8 часов под действием 2 мкг/мл лиганда CD40. Вирус инактивировали облучением УФ, CD8+ Т-клетки выделяли положительным отбором с использованием магнитных гранул и культуры для праймирования инициировали в 24-луночном планшете. После пяти циклов стимуляции идентифицировали CD8+ линии, которые специфически продуцировали интерферон-гамма при стимуляции аутологичными Р501S-трансдуцированными фибробластами. P501S-специфическая активность клеточной линии 3А-1 могла поддерживаться после дополнительных циклов стимуляции на аутологичных B-LCL, трансдуцированных P501S. Было показано, что линия 3А-1 узнает аутологичные B-LCL, трансдуцированные для экспрессии P501S, но не EGFP-трансдуцированные аутологичные B-LCL, как измерено при помощи анализов на цитотоксичность (высвобождения 51Сr) и анализа с продуцированием интерферона-гамма (ELISPOT с интерфероном-гамма; см. выше и Lalvani et al., J. Exp. Med. 186:859-865, 1997). Результаты этих анализов представлены на фигурах 6А и 6В.
ПРИМЕР 13
ИДЕНТИФИКАЦИЯ АНТИГЕНОВ ОПУХОЛИ ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ ПРИ ПОМОЩИ АНАЛИЗА С МИКРОМАТРИЦЕЙ
Указанный пример описывает выделение некоторых опухолевых полипептидов предстательной железы из библиотеки кДНК опухоли предстательной железы.
Экспрессионную библиотеку кДНК опухоли предстательной железы человека, описанную выше, подвергали скринингу с использованием анализа с микроматрицей для идентификации клонов, которые проявляют по меньшей мере трехкратную сверхэкспрессию в опухоли предстательной железы и/или нормальной предстательной железе в сравнении с не относящимися к предстательной железе нормальными тканями (за исключением яичка). Были идентифицированы 372 клона и 319 были успешно секвенированы. Таблица 1 представляет суммирование этих клонов, которые показаны в SEQ ID NO:385-400. Из этих последовательностей SEQ ID NO:386, 389, 390 и 302 соответствуют новым генам, a SEQ ID NO:393 и 396 соответствуют ранее идентифицированным последовательностям. Другие (SEQ ID NO:385, 387, 388, 391, 294, 395 и 397-400) соответствуют известным последовательностям, как показано в таблице 1.

CGI-82 проявлял 4,06-кратную сверхэкспрессию в тканях предстательной железы в сравнении с другими испытанными нормальными тканями. Он сверхэкспрессировался в 43% опухолей предстательной железы, 25% нормальных предстательных желез, не определялся в других испытанных нормальных тканях. L-идитол-2-дегидрогеназа обнаружила 4,94-кратную сверхэкспрессию в тканях предстательной железы в сравнении с другими испытанными нормальными тканями. Она сверхэкспрессировалась в 90% опухолей предстательной железы, 100% нормальных предстательных желез и не определялась в других испытанных нормальных тканях. Ets фактор транскрипции PDEF проявлял 5,55-кратную сверхэкспрессию в тканях предстательной железы в сравнении с другими испытанными нормальными тканями. Он сверхэкспрессировался в 47% опухолей предстательной железы, 25% нормальных предстательных желез и не определялся в других испытанных нормальных тканях. hTGR1 обнаруживал 9,11-кратную сверхэкспрессию в тканях предстательной железы в сравнении с другими испытанными нормальными тканями. Он сверхэкспрессировался в 63% опухолей предстательной железы и не определялся в испытанных нормальных тканях, в том числе в нормальной предстательной железе. KIAA0295 обнаруживал 5,59-кратную сверхэкспрессию в тканях предстательной железы в сравнении с другими испытанными нормальными тканями. Он сверхэкспрессировался в 47% опухолей предстательной железы, имел низкую - неопределяемую - экспрессию в испытанных нормальных тканях, в том числе в тканях нормальной предстательной железы. Кислая фосфатаза предстательной железы обнаружила 9,14-кратную сверхэкспрессию в тканях предстательной железы в сравнении с другими испытанными нормальными тканями. Она сверхэкспрессировалась в 67% опухолей предстательной железы, 50% нормальных предстательных желез и не определялась в других испытанных нормальных тканях. Трансглутаминаза обнаружила 14,84-кратную сверхэкспрессию в тканях предстательной железы в сравнении с другими испытанными нормальными тканями. Она сверхэкспрессировалась в 30% опухолей предстательной железы, 50% нормальных предстательных желез и не определялась в других испытанных нормальных тканях. Связывающий белок липопротеин высокой плотности (HDLBP) обнаружил 28,06-кратную сверхэкспрессию в тканях предстательной железы в сравнении с другими испытанными нормальными тканями. Он сверхэкспрессировался в 97% опухолей предстательной железы, 75% нормальных предстательных желез и был неопределяемым в других испытанных нормальных тканях. CGI-69 обнаружил 3,56-кратную сверхэкспрессию в тканях предстательной железы в сравнении с другими испытанными нормальными тканями. Он был слабо представленным геном, определялся в более 90% опухолей предстательной железы и в 75% нормальных предстательных желез. Экспрессия указанного гена в нормальных тканях была очень низкой. KIAA0122 обнаружил 4,24-кратную сверхэкспрессию в тканях предстательной железы в сравнении с другими нормальными тканями. Он сверхэкспрессировался в 57% опухолей предстательной железы и был неопределяемым во всех испытанных нормальных тканях, в том числе в тканях нормальной предстательной железы. 19142.2 bangur обнаружил 23,25-кратную сверхэкспрессию в тканях предстательной железы в сравнении с другими испытанными нормальными тканями. Он сверхэкспрессировался в 97% опухолей предстательной железы и 100% нормальных предстательных желез. Он был неопределяемым в других испытанных нормальных тканях. 5566.1 Wang обнаружил 3,31-кратную сверхэкспрессию в тканях предстательной железы в сравнении с другими испытанными нормальными тканями. Он сверхэкспрессировался в 97% опухолей предстательной железы, 75% нормальных предстательных желез и также сверхэкспрессировался в нормальном костном мозгу, поджелудочной железе и активированных РВМС. Новый клон 23379 обнаружил 4,86-кратную сверхэкспрессию в тканях предстательной железы в сравнении с другими испытанными нормальными тканями. Он был определяемым в 97% опухолей предстательной железы и 75% нормальных предстательных желез и является неопределяемым во всех других испытанных нормальных тканях. Новый клон 23399 обнаружил 4,09-кратную сверхэкспрессию в тканях предстательной железы в сравнении с другими испытанными нормальными тканями. Он сверхэкспрессировался в 27% опухолей предстательной железы и был неопределяемым во всех испытанных нормальных тканях, в том числе в тканях нормальной предстательной железы. Новый клон 23320 обнаружил 3,15-кратную сверхэкспрессию в тканях предстательной железы в сравнении с другими испытанными нормальными тканями. Он был определяемым во всех испытанных опухолях предстательной железы и 50% тканей нормальной предстательной железы. Он также экспрессировался в нормальных ободочной кишке и трахее. Другие нормальные ткани не экспрессировали указанный ген на высоком уровне.
ПРИМЕР 14
ИДЕНТИФИКАЦИЯ АНТИГЕНОВ ОПУХОЛИ ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ ПРИ ПОМОЩИ ЭЛЕКТРОННОГО ВЫЧИТАНИЯ
В указанном примере описано использование электронного способа вычитания для идентификации антигенов опухоли предстательной железы.
Потенциальные специфические для предстательной железы гены, присутствующие в базе данных EST человека GenBank, идентифицировали при помощи электронного вычитания (электронной субтракции) (подобно способу, описанному Vasmatizis et al., Proc. Natl. Acad. Sci. USA 95:300-304, 1998). Последовательности EST-клонов (43482), произведенные из различных библиотек предстательной железы, были получены из публичной базы данных EST человека банка генов. Каждую EST-последовательность предстательной железы использовали в качестве запрашиваемой последовательности при поиске в BLASTN (Национальном Центре информации по биотехнологии) в сравнении с базой данных EST человека. Все совпадения, рассматриваемые как идентичные (длина совпадающей последовательности >100 п.н., плотность идентифицированных совпадений на протяжении указанного района >70%), группировали (выравнивали для сравнения) вместе в кластер. Кластеры, содержащие более 200 EST, отбрасывали, так как они, возможно, представляли повторяющиеся элементы или экспрессируемые на высоком уровне гены, такие как гены для рибосомных белков. Если два или более кластеров имели общие EST, то эти кластеры группировали вместе в “суперкластер”, что дало 4345 суперкластеров.
Записи для 479 библиотек кДНК человека, представленные в выданном банком генов списке, загружали для создания базы данных записей этих библиотек кДНК. Эти 479 библиотек кДНК группировали на три группы: Плюс (библиотеки нормальной предстательной железы и опухоли предстательной железы и линии клеток молочной железы, в которых была желательной экспрессия), Минус (библиотеки из других тканей взрослых индивидуумов, в которых экспрессия не была желательной), и Другие (фетальные ткани, ткани новорожденных, ткани, обнаруживаемые только у женщин, опухоли, не относящиеся к предстательной железе, и клеточные линии, не являющиеся клеточными линиями предстательной железы, в которых экспрессия рассматривалась как неуместная). Суммирование этих групп библиотек представлено в таблице II.
Каждый суперкластер анализировали на EST в суперкластере. Отмечали источник ткани для каждого ЕSТ-клона и использовали для классификации суперкластеров на четыре группы: ЕSТ-клоны Типа 1, обнаруживаемые только в библиотеках группы Плюс; отсутствие определяемой экспрессии в библиотеках групп Минус и Другие; ЕSТ-клоны Типа 2, обнаруживаемые только в библиотеках групп Плюс и Другие; отсутствие определяемой экспрессии в группе Минус; ЕSТ-клоны типа 3, обнаруживаемые в библиотеках групп Плюс, Минус и Другие, но экспрессия в группе Плюс является более высокой, чем в группах Минус или Другие; и ЕSТ-клоны Типа 4, обнаруживаемые в библиотеках групп Плюс, Минус и Другие, но экспрессия в группе Плюс является более высокой, чем экспрессия в группе Минус. Указанный анализ идентифицировал 4345 кластеров молочной железы (таблица III). Из этих кластеров 3172 ЕSТ-клона были заказаны из Research Genetics, Inc., и они были получены в виде замороженных исходных растворов в глицерине в 96-луночных планшетах.

Вставки ПЦР-амплифицировали с использованием амино-связанных ПЦР-праймеров для анализа микроматриц Synteni. При получении более одного ПЦР-продукта для конкретного клона указанный ПЦР-продукт не использовали для анализа экспрессии. Всего 2528 клонов из способа электронного вычитания были подвергнуты анализу микроматриц для идентификации полученных электронным вычитанием клонов молочной железы, которые имели высокий уровень мРНК опухолей в сравнении с уровнем мРНК нормальной ткани. Такие скрининги проводили с использованием микроматрицы Synteni (Palo Alto, CA) в соответствии с инструкциями изготовителя (и по существу, как описано Schena et al., Proc. Natl. Acad. Sci. USA 93:10614-10619, 1996 и Heller et al., Proc. Natl. Acad. Sci. USA 94:2150-2155, 1997). В этих анализах клоны выстраивали в виде рядов на микросхеме, которую затем зондировали флуоресцентными зондами, генерируемыми из кДНК нормальной и имеющей опухоль предстательной железы, а также различных других нормальных тканей. Эти слайды сканировали и измеряли интенсивность флуоресценции.
Клоны с коэффициентом экспрессии более 3 (т.е. уровень кДНК в опухоли предстательной железы был по меньшей мере в три раза более высоким, чем уровень кДНК в нормальной предстательной железе) идентифицировали как последовательности, специфические для опухоли предстательной железы (таблица IV). Последовательности этих клонов представлены в SEQ ID NO:401-453, а некоторые новые последовательности представлены в SEQ ID NO:407, 413, 416-419, 422, 426, 427 и 450.


ПРИМЕР 15
ДОПОЛНИТЕЛЬНАЯ ИДЕНТИФИКАЦИЯ АНТИГЕНОВ ОПУХОЛИ ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ ПОСРЕДСТВОМ АНАЛИЗА МИКРОМАТРИЦ
В этом примере описано выделение дополнительных полипептидов опухоли предстательной железы из библиотеки кДНК опухоли предстательной железы.
Экспрессионную библиотеку кДНК предстательной железы человека, описанную выше, подвергали скринингу с использованием анализа микроматриц для идентификации клонов, которые обнаруживают по меньшей мере трехкратную сверхэкспрессию в опухоли предстательной железы и/или в нормальной ткани предстательной железы в сравнении с не относящимися к предстательной железе нормальными тканями (за исключением яичка). Были идентифицированы и секвенированы 142 клона. Некоторые из этих клонов представлены в SEQ ID NO:454-467. Из этих последовательностей SEQ ID NO:459-461 соответствуют новым генам. Другие (SEQ ID NO:454-458 и 461-467) соответствуют известным последовательностям.
ПРИМЕР 16
ДОПОЛНИТЕЛЬНАЯ ХАРАКТЕРИСТИКА АНТИГЕНА Р710Р ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ
В этом примере описано полноразмерное клонирование Р710Р.
Библиотеку кДНК предстательной железы, описанную выше, подвергали скринингу фрагментом Р710Р, описанным выше. Один миллион колоний высевали на чашки с LB/ампициллином. Найлоновые мембранные фильтры использовали для подъема (лифтинга) этих колоний и затем кДНК, извлеченные этими фильтрами, денатурировали и сшивали с фильтрами облучением УФ-светом. Фрагмент Р710Р радиоактивно метили и использовали для гибридизации с фильтрами. Положительные кДНК-клоны отбирали и их кДНК извлекали и секвенировали при помощи автоматического ABI-секвенатора. Были получены четыре последовательности и они представлены в SEQ ID NO:468-471.
На основании вышеописанного должно быть понятно, что хотя с целью иллюстрации здесь были описаны характерные варианты данного изобретения, различные модификации могут быть произведены без отхода от идеи и объема данного изобретения. Таким образом, данное изобретение не ограничивается ничем, кроме прилагаемой формулы изобретения.
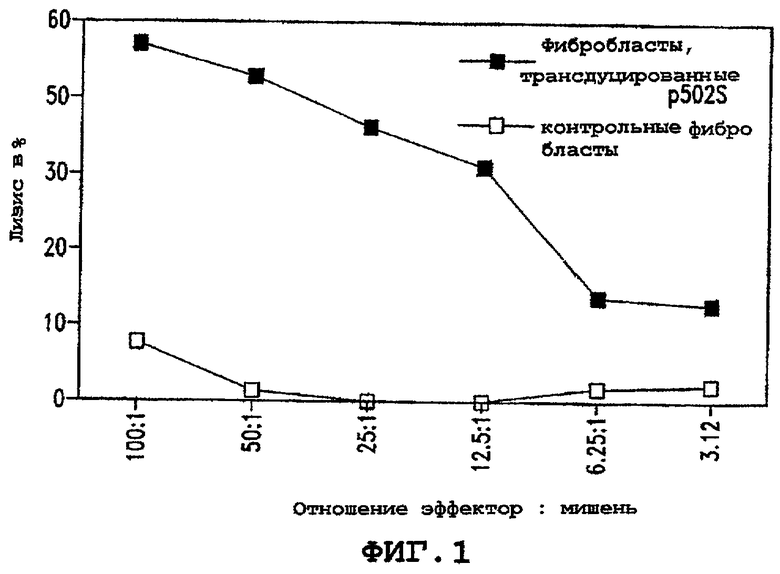















































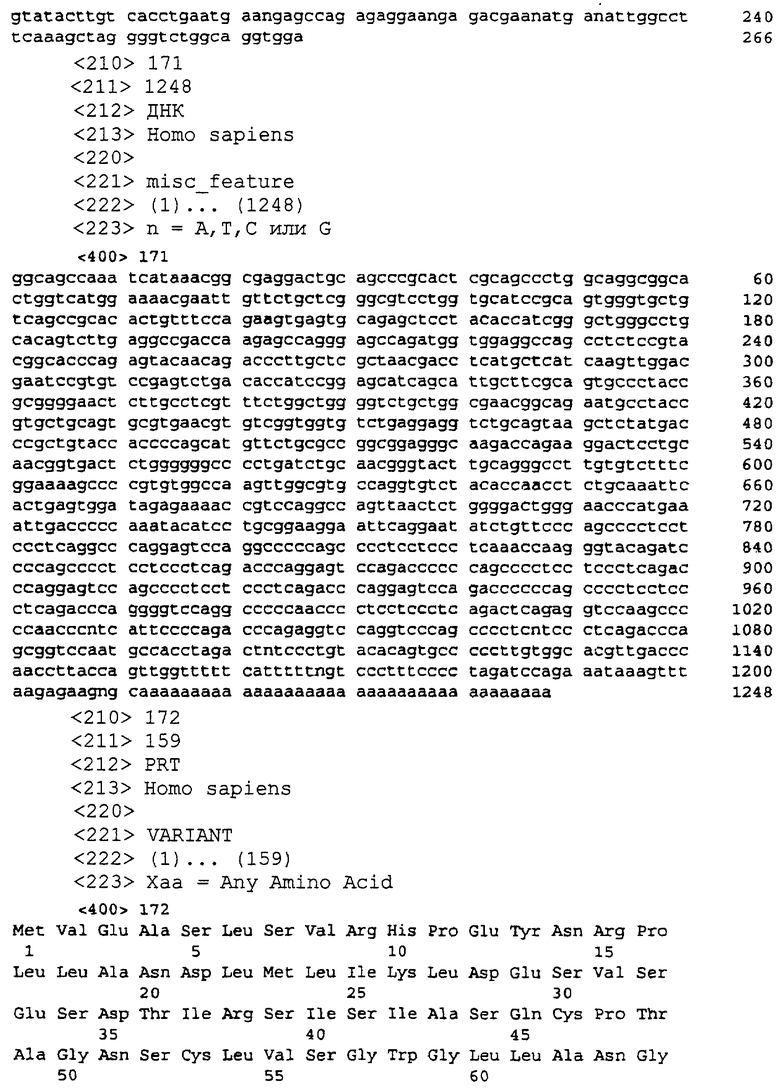
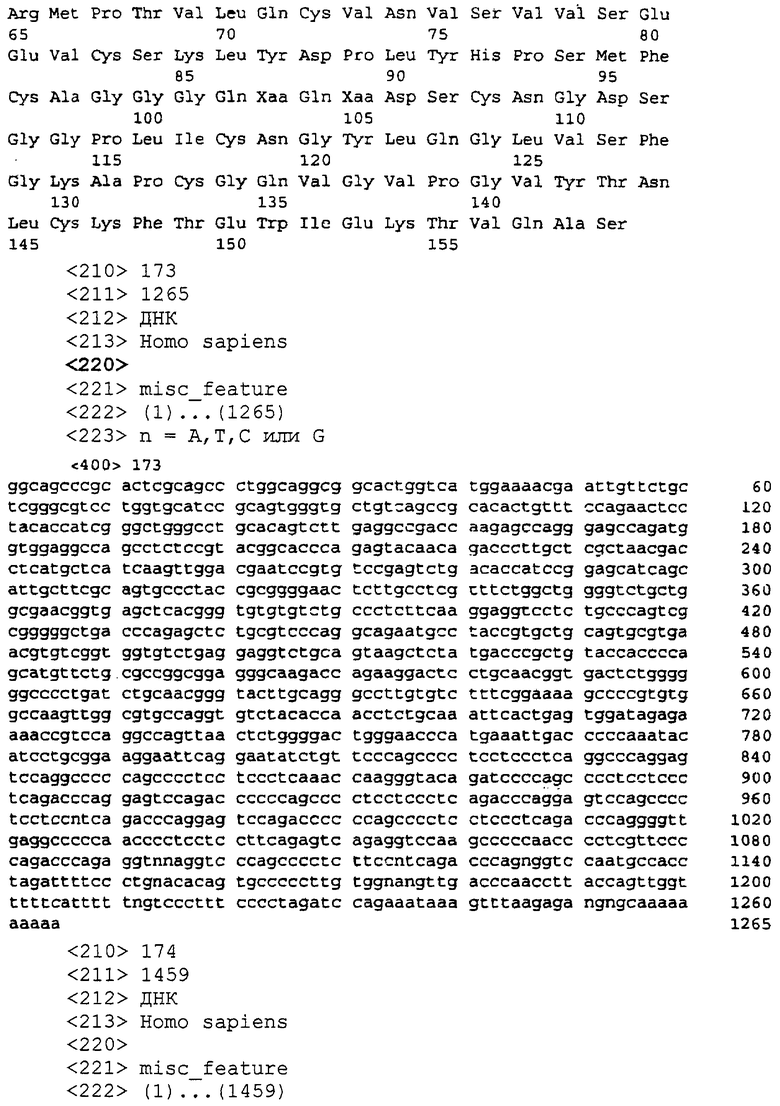









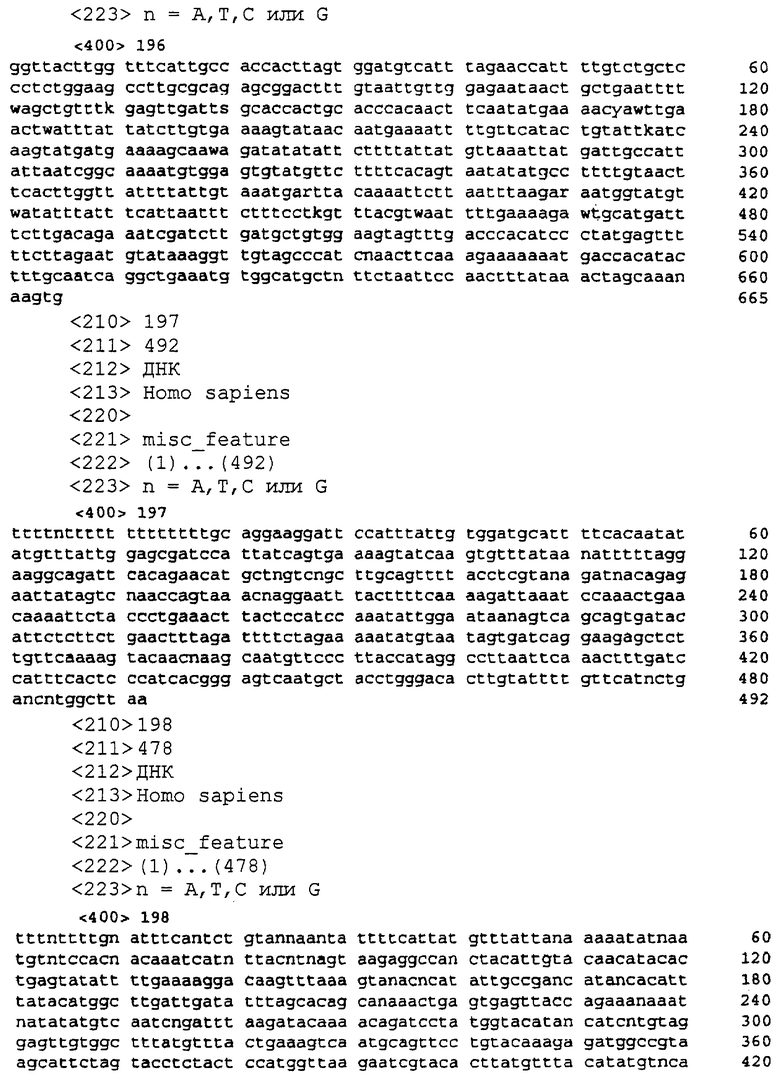




































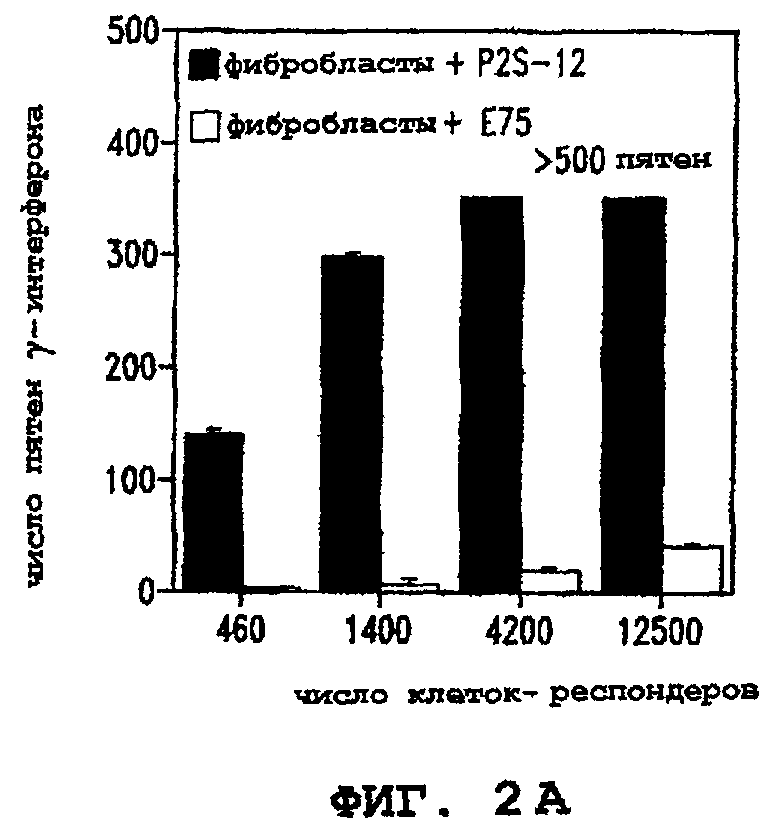
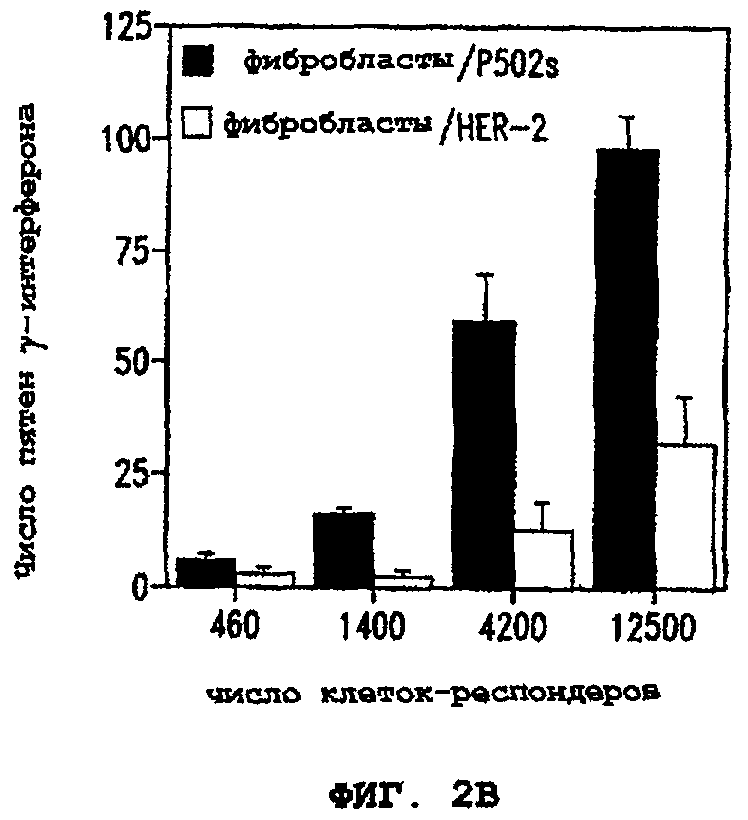
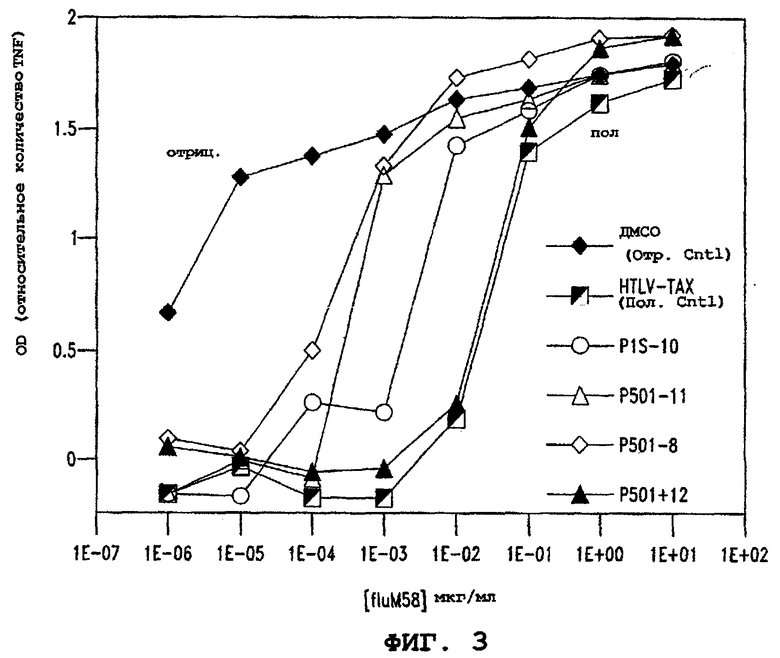
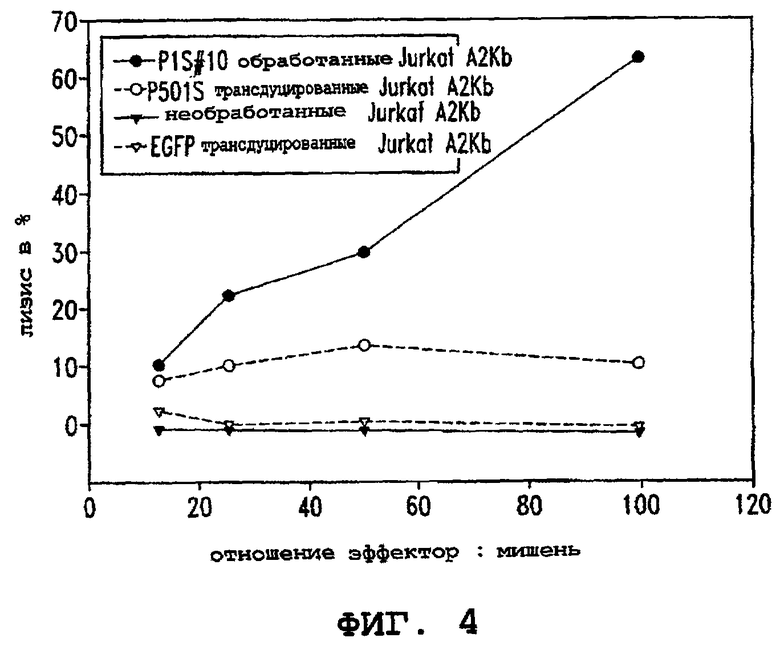
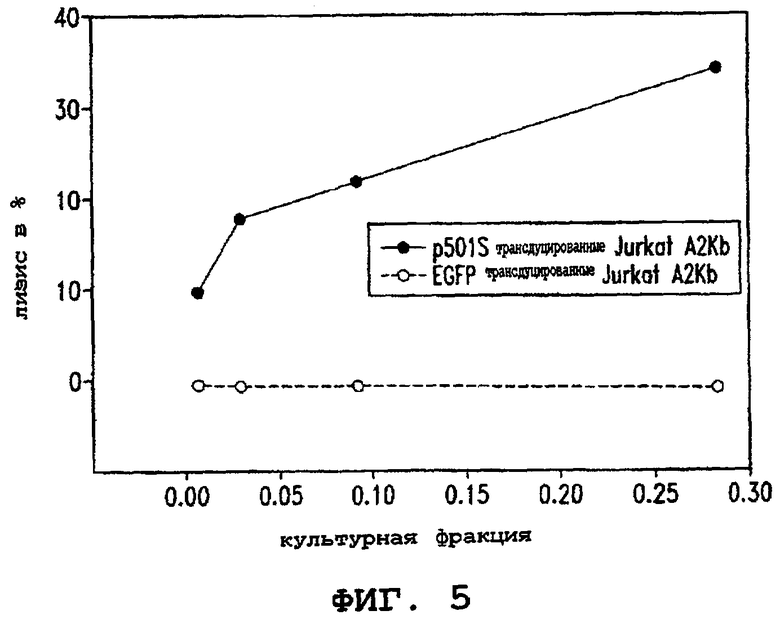
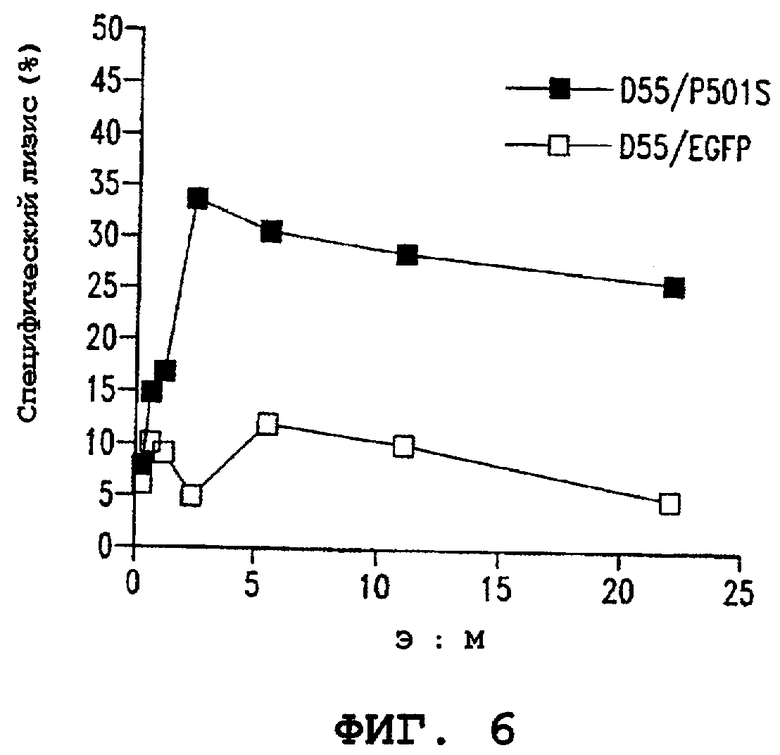
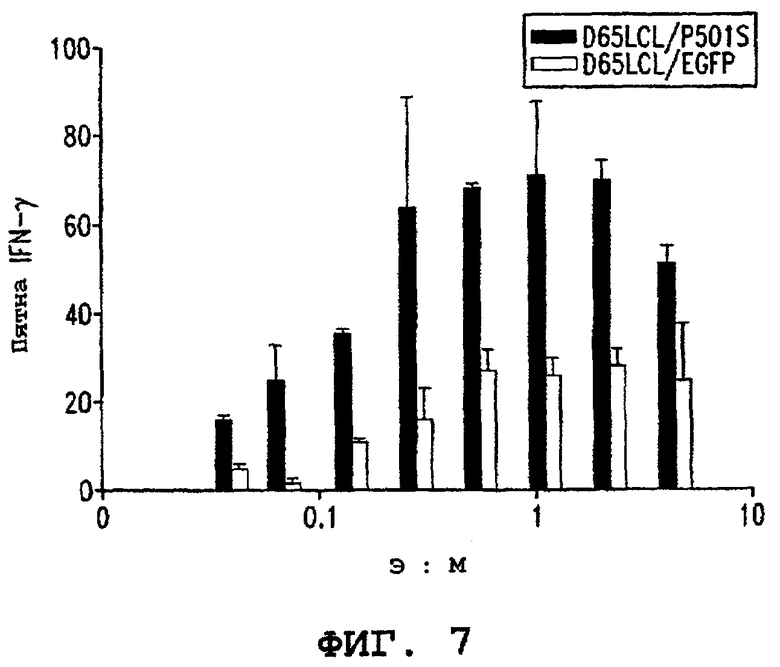
Изобретение относится к области онкологии. Сущность изобретения составляет выделенный опухолевый полипептид предстательной железы, содержащий последовательность SEQ ID NO:327, и полинуклеотид, выделенный из опухолевой ткани предстательной железы, кодирующий опухолевые полипептиды предстательной железы и содержащий последовательность SEQ ID NO:225 или SEQ ID NO:326. Технический результат - такие полипептиды и полинуклеотиды расширяют арсенал генетических средств для использования их в вакцинах и фармацевтических композициях для профилактики и лечения рака предстательной железы и для диагностики и мониторинга таких опухолей. 2 с.п.ф-лы, 4 табл., 7 ил.,
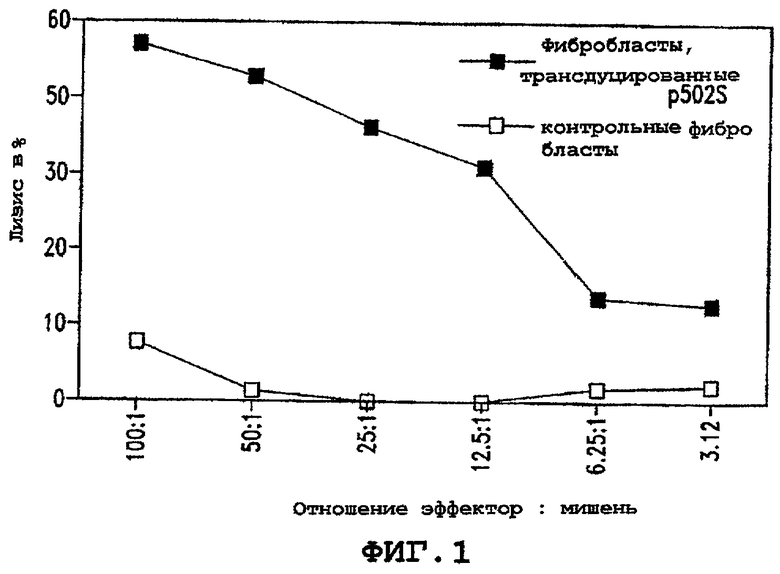
| Домовый номерной фонарь, служащий одновременно для указания названия улицы и номера дома и для освещения прилежащего участка улицы | 1917 |
|
SU93A1 |
| ЦИФРОВОЙ ИЗМЕРИТЕЛЬ ИНДУКТИВНОСТИ | 0 |
|
SU199992A1 |
| Биохимия гормонов и гормональная регуляция | |||
| Под ред | |||
| Н.А.Юдаева | |||
| - М., 1976, с.76-95. | |||

