Область изобретения
Изобретение относится к области молекулярной биологии, биологии рака и терапевтической медицины. Изобретение главным образом направлено на идентификацию генов, которые ингибируют развитие клеток рака и вызывают в них апоптоз, и на полипептиды, кодируемые такими генами. В частности, изобретение касается нуклеотидной последовательности одного из таких ингибирующих опухоли генов, tag7, аминокислотной последовательности полипептида, кодируемого tag7, антител, которые специфически связывают генный продукт tag7 и способов ингибирования развития раковых клеток и терапии рака с применением гена tag7 и генного продукта.
Уровень техники
Метастазирование опухоли
Метастазирование опухолевых клеток представляет собой комплексный процесс, включающий сложный каскад процессов. Среди этих процессов пролиферация опухолевых клеток и блокирование механизма апоптоза (см. статью Harrington Е.А. и др. в журнале Embo J. 73:3286-3295 (1994)), распространение опухолевых клеток в окружающих тканях, проникновение опухолевых клеток в циркуляционные контуры крови и лимфы и закрепление и размножение опухолевых клеток на новом месте (см. статьи Schirmacher А.Е. в журнале Adv.Cancer Res. 43: и Liotta L.A. и др. в журнале Lab.Invest.49:636-649 (1983)). Пока молекулярные механизмы индуцирования фенотипа метастазов остаются слабо исследованными; вероятно, что когда опухоль становится метастазирующей, происходит активация и/или инактивакция различных регуляторных и структурных генов. Описаны несколько генов, чья экспрессия коррелируется с потенциальной способностью опухолей к метастазированию (см. статьи Yasushi Т. и др. в журнале J.Biol.Chem. 269(37): 22958-22963 (1994); Ebralidze А.К. и др. в журнале Genetika (USSR),25(5): 932-936 (1989); Wolf С. и др. в журнале Proc.Natl.Acad.Sci. USA 90:1843-1847 (1993); Bernbard E.J. и др. в журнале Proc.Natl.Acad.Sci. USA 91;61-65; Sato Н. и др. в журнале Nature 370:61-65 (1994)).
Предполагалось, что популяция клеток в пределах определенной опухоли может быть гетерогенной в отношении их потенциальной способности к метастазированию (см. статью Fidler I.J. и Hart I.R. в журнале Science 217: 998-1001(1982). Это предположение указывало на возможность получения родственных опухолей, которые по потенциальной способности к метастазированию заметно отличаются от одной родительской опухоли. Например, получили опухоли, которые можно трансплантировать мыши и которые имеют переменную повторяемость и специфичность метастазирования по отношению к органу, как результат выбора характера метастазирования (см. статью Senin V.M. в журнале Vestn.Akad.Med.Nauk.SSSR 0(5): 85-91 (1984)).
Полипептиды, ингибирующие опухоли
Ряд полипептидов, естественньм путем продуцируемых клетками млекопитающих, проявляют антиопухолевую активность (то есть индуцируют задержку роста, апоптоз и/или дифференциацию раковых клеток). Например, сообщается, что комбинации определенных цитокинов, таких как интерлейкины и колониестимулирующие факторы, индуцируют конечную дифференциацию и сопутствующую задержку роста некоторых типов опухолевых клеток и клеточных линий (см. обзор, данный Pimental E. в Handbook of Growth Factors, Vol.1, Roca Raton, Florida: CRC Press, c.28-34 (1994)). К тому же, трансформирующий (β-фактор роста (TGF-β) и интерфероны известны в качестве потенциальных ингибиторов развития опухолевых клеток при определенных условиях in vivo и in vitro (см. статью Keski-Oja J. и Moses H.L. в журнале Med-Biol. 66:13-20 (1987); статью Ohta M. и др. в журнале Nature 329:539-541 (1987)). Другие естественные полипептиды млекопитающих проявляют разнообразную противоопухолевую активность, например индуцируют апоптоз в некоторых опухолевых клетках, содержат факторы некроза опухолей (TNFs) (см. обзор, данный Pimental E. в Handbook of Growth Factors, Vol.3, Roca Raton, Florida: CRC Press, c.241-278 (1994)).
Недавно было идентифицировано семейство TNF-цитокинов и их рецепторов, чьи структурные свойства до некоторой степени являются обычными. Это семейство включает системы рецептор-лиганд из TNF, LT-α, LT-β, Fas, CD27, CD40, ОХ-40 и фактора роста нерва (NGF) (см. статьи Smith С. и др. в журнале Cell 76:959-962(1994); Armitage R.J. в журнале Curr.Opin.Immunol. 6:407-413 (1994)). За исключением NGF, все эти TNF-цитокины, как предполагается, вовлечены в регуляцию иммунной системы. TNF и лимфотоксин-альфа (LT-α или TNF-β) представляют собой цитокины, вовлеченные во многие регуляторные процессы (см. статьи Vassalli P. в журнале Ann. Rev. Immunol. 10: 411-452 (1992); Paul N. и Ruddle N. в журнале Ann.Rev.Immunol. 6:407-438 (1988)), но их роль в иммунной системе, хотя и является, без сомнения, решающей, остается загадочной (см. статью Kossodo S. и др. в журнале Exp.Med. 176:1259-1264 (1994)). TNF синтезируется разными типами клеток как ответ на различные неблагоприятные воздействия; это обычно является одним из первичных явлений в каскаде воспалительных процессов, включающем сильный противоопухолевый эффект у мыши (см. статью Blankenstein Т. и др. в журнале J.Exp.Med.173:1047-1052 (1991)). Активированные макрофаги являются основным источником мембраносвязанного TNF, хотя он также продуцируется активированньми лимфоцитами и несколькими клетками других типов. Наоборот, LT-α продуцируется только лимфоцитами и существует в мембраносвязанной форме только через гримерный комплекс с LT-β (см. статью Browning J. и др. в журнале Cell 72:847-855 (1993)). LT-β обнаруживает спектр действия, аналогичный спектру действия TNF в системах in vitro, но слабее (см. статью Browning J. и Ribolini А. в журнале J.Immunol. 743:1859-1867 (1989)). И TNF, и LT-α индуцируют апоптоз в различных системах (см. статьи Cohen J.J. и др. в журнале Ann. Rev.Immunol. 10:267-293 (1992); Golstein Р. и др. в журнале Immunol.Rev.121:29-65 (1991); Sarin А. и др. в журнале J.Immunol. l55:3716-3718 (1995)). Недавно появилось сообщение об ингибировании роста опухолей, опосредованном LT-α (см. статью Qin Z. и Blankenstein Т. в журнале Cancer Res. 55-A747-4751 (1995)).
TNF и LT-α также вырабатываются некоторыми опухолевыми клетками различного происхождения, такими как фибросаркомы мыши, линии эпителиальных клеток человека, а также клетки и клеточные линии лейкемии Т-клеток (см. статьи Rubin B.Y. и др. в журнале J.Exp.Med.164:1350-1255 (1986); Spriggs D.R. и др. в журнале J.Clin.Invest. 81:455-460 (1988); Ishibashi К. и др. в журнале Blood77: 2451-2455 (1991)). Гены, кодирующие TNF, LT-α и LT-P, лежат на близком расстоянии один от другого внутри области класса III основного комплекса гистосовместимости (МНС) (см. статьи Spies и др. в журнале Proc.Natl.Acad. USA 53:8699-8702 (1986); Nedospasov S.A. и др. в журнале Nucl.Acids Res. 14:7713-7725 (1986); Gardner S.М.и др. в журнале J.Immunol. 139:476-483 (1987)). Представляется, что эти гены являются эволюционно родственными и формируют локус за счет тандемных дупликаций гена, хотя противоположная ориентация транскрипции LT-β означает, что могут иметь место более сложные эволюционные процессы. В последнее время был клонирован и определен в качестве индуцирующего апоптоз TNF-лиганда (TRAIL), еще один новый член семейства TNF (см. статью Wiley S.R. и др. в журнале Immunity 3: 673-682 (1995)).
КРАТКОЕ ИЗЛОЖЕНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение преимущественно относится к генам и полипептидам, ингибирующим развитие опухолей, и способам лечения рака с использованием таких генов и полипептидов. Конкретно, изобретение предлагает выделенные молекулы нуклеиновой кислоты tag7, содержащие нуклеотидную последовательность по меньшей мере на 65% (более предпочтительно по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 99%) идентичную используемой в качестве ссылки последовательности, выбранной из группы, включающей:
(a) нуклеотидную последовательность, представленную в SEQ ID NO:1;
(b) нуклеотидную последовательность, кодирующую полипептид tag7 имеющий полноразмерную аминокислотную последовательность, представленную в SEQ ID NO:2;
(c) нуклеотидную последовательность, кодирующую зрелый полипептид tag7, имеющий аминокислотную последовательность в положениях 20-182 в SEQ ID NO:2;
(d) нуклеотидную последовательность полинуклеотида, который гибридизуется при жестких условиях с полинуклеотид, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:1;
(e) нуклеотидную последовательность полинуклеотида, который гибридизуется при определенных условиях с полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:1;
(f) нуклеотидную последовательность, комплементарную к любой из последовательностей по (а), (b), (с), (d) или (е), или их фрагмент.
Изобретение также относится к выделенной молекуле нуклеиновой кислоты, содержащей полинуклеотид, который гибридизуется при жестких условиях с полинуклеотидом, имеющим нуклеотидную последовательность, идентичную той, которая содержится в выделенных молекулах нуклеиновой кислоты, описанных выше, и которая может или не может кодировать полипептид с tag7-активностью.
Изобретение также относится к выделенной молекуле нуклеиновой кислоты, содержащей полинуклеотид, который гибридизуется при определенных условиях с полинуклеотидом, имеющим нуклеотидную последовательность, идентичную той, которая содержится в выделенных молекулах нуклеиновой кислоты, описанных выше, и которая может или не может кодировать полипептид с tag7-активностью.
В предпочтительном воплощении изобретение относится к выделенным молекулам нуклеиновой кислоты tag7 содержащим полинуклеотид, имеющий:
(a) нуклеотидную последовательность, представленную в SEQ ID NO:3;
(b) нуклеотидную последовательность, кодирующую человеческий полипептид tag7, имеющий полноразмерную аминокислотную последовательность, представленную в SEQ ID NO:4;
(c) нуклеотидную последовательность полинуклеотида, который гибридизуется при жестких условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:3;
(d) нуклеотидную последовательность полинуклеотида, который гибридизуется при определенных условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:3;
(e) нуклеотидную последовательность, комплементарную к любой из последовательностей по (а), (b), (с), или (d), или их фрагмент.
Изобретение также относится к векторам, в частности векторам экспрессии, содержащим такие выделенные молекулы нуклеиновой кислоты, и клеткам-хозяевам, которые содержат эти выделенные молекулы или векторы. Предпочтительные клетки-хозяева согласно изобретению включают, не ограничиваясь этим, бактериальные клетки, дрожжевые клетки, клетки животных (в особенности клетки млекопитающих или насекомых) и клетки растений.
Изобретение также касается способов продуцирования выделенного полипептида tag7, предусматривающих культивирование вышеописанных клеток-хозяев при условиях, обеспечивающих экспрессию полипептида tag7 и выделение его. Изобретение направлено также на выделенные полипептиды tag7, продуцированные согласно этим способам.
Изобретение относится также к выделенным полипептидам tag7, имеющим аминокислотную последовательность по меньшей мере на 65% (более предпочтительно на по меньшей мере 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 99%), идентичную используемой в качестве ссылки последовательности, выбранной из группы, включающей:
(a) аминокислотную последовательность, кодируемую выделенной молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность, представленную в SEQ ID NO:1;
(b) полноразмерную аминокислотную последовательность полипептида tag7, представленную в SEQ ID NO:2;
(c) аминокислотную последовательность полипептида tag7, кодируемую полинуклеотидом, который гибридизуется при жестких условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:1;
(d) аминокислотную последовательность полипептида tag7, кодируемую полинуклеотидом, который гибридизуется при определенных условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:1; или их фрагмент.
В предпочтительном воплощении изобретение относится к человеческому полипептиду tag7, имеющему:
(a) аминокислотную последовательность, кодируемую выделенной молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность, представленную в SEQ ID NO:3;
(b) аминокислотную последовательность полипептида tag7, представленную в SEQ ID NO:4; или их фрагмент.
Изобретение относится также к фармацевтическим композициям, содержащим один или более из вышеописанных выделенных полипептидов tag7, или их фрагментов и фармацевтически приемлемый носитель, или среду для них.
Изобретение касается также способов продуцирования выделенного tag7-специфичного антитела, предусматривающих иммунизацию животного вышеописанными выделенными полипептидами tag7 и выделение из животного tag7-специфичного антитела. Изобретение направлено также на tac7-специфичные антитела, продуцируемые этими способами. Антитела согласно настоящему изобретению могут быть моноклональными или поликлональными и могут быть мечены для их обнаружения или иммобилизованы на твердой опоре.
Изобретение касается также способов ингибирования развития опухолей у млекопитающих, таких как опухоли у человека. В одном из предпочтительных воплощений, такие способы согласно изобретению могут предусматривать воздействие на клетку млекопитающего композиции, содержащей один или более выделенных полипептидов tag7, причем выделенный полипептид tag7 имеет аминокислотную последовательность по меньшей мере на 65% (более предпочтительно по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 99%) идентичную используемой в качестве ссылки последовательности, выбранной из группы, включающей:
(a) аминокислотную последовательность, кодируемую выделенной молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность, представленную в SEQ ID NO:1;
(b) полноразмерную аминокислотную последовательность полипептида tag7, представленную в SEQ ID NO:2;
(c) аминокислотную последовательность зрелого полипептида tag7, имеющего аминокислотную последовательность, представленную в положениях 20-182 в SEQ ID NO:2;
(d) аминокислотную последовательность, кодируемую полинуклеотидом, который гибридизуется при жестких условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:1;
(e) аминокислотную последовательность, кодируемую полинуклеотидом, который гибридизуется при определенных условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:1, при этом воздействие на клетку млекопитающего полипептидом tag7 ингибирует рост клетки.
В предпочтительном варианте осуществления этого способа опухолью млекопитающего является опухоль человека, и на человеческие клетки воздействуют композицией, содержащей человеческий полипептид tag7, имеющий аминокислоту, кодируемую выделенной молекулой нуклеиновой кислоты, который имеет нуклеотидную последовательность, представленную в SEQ ID NO:3, или ее фрагмент.
В другом предпочтительном варианте осуществления данное изобретение относится к способам ингибирования развития опухоли у млекопитающего, предусматривающим введение в клетку млекопитающего молекулы нуклеиновой кислоты, содержащей полинуклеотид, имеющий полинуклеотидную последовательность, по меньшей мере на 65% (более предпочтительно, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 99%), идентичную используемой в качестве ссылки последовательности, выбранной из группы, включающей:
(a) нуклеотидную последовательность, представленную в SEQ ID NO:l;
(b) нуклеотидную последовательность, кодирующую полипептид tag 7, имеющий полноразмерную аминокислотную последовательность, представленную в SEQ ID NO:2;
(c) нуклеотидную последовательность, кодирующую зрелый полипептид tag7, имеющий аминокислотную последовательность в положениях 20-182 в SEQ ID NO:2;
(d) нуклеотидную последовательность молекулы нуклеиновой кислоты, кодирующей полипептид tag7, содержащий полинуклеотид, который гибридизуется при жестких условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:l;
(е) нуклеотидную последовательность молекулы нуклеиновой кислоты, кодирующей полипептид tag7, содержащий полинуклеотид, который гибридизуется при определенных условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:1, при этом введение выделенной молекулы нуклеиновой кислоты в клетку млекопитающего ингибирует развитие опухоли.
В предпочтительном варианте осуществления данное изобретение относится к способам ингибирования развития опухоли человека, предусматривающим введение в клетку человека молекулы нуклеиновой кислоты, содержащей человеческий полинуклеотид tag7, имеющий нуклеотидную последовательность, представленную в SEQ ID NO:3, или его фрагмент.
В соответствии с данным изобретением в результате осуществления настоящих способов можно индуцировать в вышеописанных опухолях апоптоз. Опухоли млекопитающих, развитие которых ингибируется способами по изобретению, преимущественно включают опухолевые клетки человека, в частности клетки карциномы (включая, но не ограничиваясь этим, клетки карциномы печени, клетки карциномы яичника, клетки карциномы груди, клетки карциномы затылочной части шеи, клетки карциномы легкого, клетки карциномы простаты, клетки карциномы желудка, клетки карциномы мочевого пузыря, клетки карциномы яичка, клетки карциномы прямой кишки, клетки кардиномы поджелудочной железы, клетки карциномы полости рта, клетки карциномы чешуйчатых клеток, клетки карциномы головы и шеи и клетки тератокарциномы), клетки саркомы (включая, но не ограничиваясь этим, клетки саркомы Kaposi, клетки фибросаркомы и клетки остеосаркомы), клетки меланомы и клетки лейкемии.
Изобретение также относится к способам лечения рака у страдающего им животного (в частности, млекопитающего, такого как человек). В одном из предпочтительных воплощений такие способы могут предусматривать воздействие на указанное животное композиции, содержащей один или более выделенных полипептидов tag7, причем указанный выделенный полипептид tag7 имеет аминокислотную последовательность, по меньшей мере на 65% (более предпочтительно по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 99%) идентичную используемой в качестве ссылки последовательности, выбранной из группы, включающей:
(a) аминокислотную последовательность, кодируемую выделенной молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность, представленную в SEQ ID NO:1;
(b) полноразмерную аминокислотную последовательность полипептида tag 7, представленную в SEQ ID NO:2;
(c) аминокислотную последовательность зрелого полипептида tag 7, имеющего аминокислотную последовательность, представленную в положениях от 20 до 182 в SEQ ID NO:2;
(d) аминокислотную последовательность, кодируемую молекулой выделенной нуклеиновой кислоты, содержащей полинуклеотид, который гибридизуется при жестких условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:1;
(е) аминокислотную последовательность, кодируемую молекулой выделенной нуклеиновой кислоты, содержащей полинуклеотид, который гибридизуется при определенных условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID: 1, причем указанное лечение ингибирует развитие рака или индуцирует ремиссию рака.
В другом предпочтительном варианте осуществления данного изобретения такие способы согласно настоящему изобретению могут предусматривать введение в организм животного молекулы нуклеиновой кислоты, содержащей полинуклеотид, имеющий нуклеотидную последовательность по меньшей мере на 65% (более предпочтительно по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 99%), идентичную используемой в качестве ссылки последовательности, выбранной из группы, включающей:
(a) нуклеотидную последовательность, представленную в SEQ ID NO:1;
(b) нуклеотидную последовательность, кодирующую полипептид tag7 имеющий полноразмерную аминокислотную последовательность, представленную в SEQ ID NO:2;
(c) нуклеотидную последовательность, кодирующую зрелый полипептид tag7 имеющий аминокислотную последовательность в положениях от 20 до 182 в SEQ ID NO:2;
(d) нуклеотидную последовательность полинуклеотида, который гибридизуется, при жестких условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:1;
(е) нуклеотидную последовательность полинуклеотида, который гибридизуется при определенных условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:1, причем указанное лечение ингибирует развитие рака или индуцирует ремиссию.
В предпочтительном варианте осуществления такие способы могут предусматривать введение человеку композиции, содержащей выделенный человеческий полинуклеотид tag7, имеющий полинуклеотидную последовательность, представленную в SEQ ID NO:3, или по меньшей мере один из фрагментов.
В другом предпочтительном варианте осуществления данное изобретение относится к способам лечения рака у страдающего им животного, предусматривающих введение животному по меньшей мере одной вышеописанной композиции, содержащей один или несколько из выделенных полипептидов tag7 по настоящему изобретению.
В соответствии с изобретением выделенные полипептиды tag7, используемые в вышеописанных способах, преимущественно имеют аминокислотную последовательность по меньшей мере на 95% идентичную вышеописанным последовательностям. Композиции, содержащие tag7 и используемые в вышеописанных способах, могут далее содержать фармацевтически приемлемые носитель или среду для выделенного полипептида tag7.
Также в соответствии с настоящим изобретением полинуклеотиды, используемые в вышеописанных способах, имеют последовательности нуклеиновых кислот по меньшей мере на 65% (более предпочтительно по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% или по меньшей мере на 99%), идентичные вышеописанным последовательностям, и могут содержаться в векторе или вирионе, которые можно произвести из аденовируса или аденоассоциированного вируса.
Животным, страдающим от рака и подвергаемьм лечению способами согласно настоящему изобретению, может быть млекопитающее, например человек. Рак, излечиваемый этими способами, может включать, не ограничиваясь этим, карциному (такую как карцинома печени, карцинома яичника, карцинома груди, карцинома затылочной части шеи, карцинома легкого, карцинома простаты, карцинома желудка, карцинома мочевого пузыря, карцинома яичка, карцинома прямой кишки, карцинома поджелудочной железы, карцинома полости рта, карцинома чешуйчатых клеток, карцинома головы и шеи или тератокарцинома), саркому (например, саркому Kaposi, фибросаркому и остеосаркому), меланому или лейкемию.
Другие предпочтительные варианты осуществления настоящего изобретения будут ясны обычному специалисту с учетом прилагаемых чертежей и описания изобретения, а также формулы изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фиг.1 изображает нуклеотидную последовательность кодирующего сегмента клонированной к ДНК из tag7 (SEQ ID NO:1) и аминокислотную последовательность (SEQ ID NO: 2) полипептида tag7, кодируемого этой кодирующей последовательностью.
Фиг.2 представляет сложный график различных структурных характеристик полипептида tag7, показывающий (на основе первичной аминокислотной структуры полипептида): вероятные участки альфа-спирали, бета-складчатой структуры, бета-изгиба и альфа-закрученной структуры; вероятные альфа- и бета-амфипатические участки; гидрофильность полипептида; показатель антигенности полипептида; возможную локализацию полипептида на клеточной поверхности.
Фиг.3 представляет радиоавтографию, сравнивающую результаты анализа, выявляющего различия в мРНК у неметастазирующих опухолевых клеток VMR-0 (линии 1, 3) и опухолевых клеток VMR-L, дающих метастазы в печени (линии 2, 4), кДНК были получены из 0,2 мг РНК путем обратной транскрипции с помощью обратной транскриптазы вируса лейкемии из мыши Moloney (M-MuLV) в присутствии праймера Т12АС (5'-ТТТТТТТТТТТТАС-3') (SEQ ID NO: 5) и двух различных 10-основных олигонуклеотидных праймеров: 5'-AATCGGGCTG-3' (SEQ ID NO:6; линии 1, 2) и 5'AGT-CAGCCAC-3' (SEQ ID NO:7; линии 3, 4). Различия в популяциях мРНК между двумя разными типами клеток показаны стрелками.
Фиг.4 представляет радиоавтографию Нозерн-блоттинг гибридизации реамплифицированных кДНК проб tag7 с помощью тотальной РНК, полученной из опухолевых клеток VMR-0 (линия 1) и опухолевых клеток VMR-L (линия 2). Относительное количество материала в каждой линии определяется по интенсивности сигнала гибридизации от кДНК гена глицеральдегид-3-фосфатдегидрогеназы (GAPDH).
Фиг.5 представляет радиоавтографию Нозерн-блоттинг гибридизации тотальной РНК, выделенной из различных органов здоровой мыши, с обработкой пробой, представляющей собой 32Р-меченную ДНК клона tag7. Линия 1: клетки VMR-0; линия 2: клетки VMR-L; линия 3: исходная печень; линия 4: исходная зобная железа; линия 5: исходный яичник; линия 6: исходное сердце; линия 7: исходный головной мозг; линия 8: исходная почка; линия 9: исходная селезенка; линия 10: исходное легкое; линия 11: исходная кожа. GAPDH является меченной пробой, используемой в качестве стандарта, как на фиг.2.
Фиг.6 представляет радиоавтографию Нозерн-блоттинг-гибридизации тотальной РНК, выделенной из различных клеточных линий мыши, с обработкой пробой, представляющей собой 32Р-меченную ДНК из клона tag7. Линия 1: клетки CSML-0; линия 1:клетки CSML-100; линия 3: клетки VMR-0; линия 4: клетки VMR-L; линия 5: клетки VMR-Ov.
Фиг.7 представляет радиоавтографию Нозерн-блоттинг-гибридизации тотальной РНК, выделенной из различных первичных опухолей мыши и соответствующих опухолевых клеточных линий мыши, с обработкой пробой, представляющей собой 32Р-меченную ДНК из клона tag7.
Фиг.8 представляет набор фотографий, отображающих распределение транскриптов tag7 в тканях мыши.
(А): Радиоавтография Нозерн-блоттинга тотальной РНК, выделенной из указанных тканей здоровой мыши.
(В-Е): Гибридизация in situ участков ткани взрослой мыши с помощью проб 35S-меченной ДНК из клона tag 7; (В): гиппокампус и зубчатая извилина головного мозга; (С): мозжечок; (D): кишки; (Е): кишки, расщепленные РНК-азой перед гибридизацией.
Фиг.9 представляет радиоавтографию Нозерн-блоттинга транскрипции tag7 в лимфоидных и кроветворных клетках мыши, культивированных в средах без добавок или в средах, содержащих липополисахарид (LPS).
Фиг.10 представляет фотографию Вестерн-блоттинга растворимых и связанных с клеткой форм tag7 в различных мышиных клетках.
(A): tag7 связанный с клеткой. Общие клеточные лизаты и кондиционированные среды ("supe") LPS-индуцированных и неиндуцированных опухолевых мышиных клеток VMR-L и VMR-0 были иммуноосаждены с антителами анти-tag7 и проанализированы методом Вестерн-блоттинга с использованием в качестве контроля рекомбинантного tag7 (rtagT).
(B): Растворимый tag7. Спленоциты мыши были стимулированы липополисахаридом в течение различных отрезков времени, и лизаты или супернатанты были иммуноосаждены и проанализированы методом блоттинга с антителами анти-tag7, как описано выше. Величины маркеров молекулярной массы указаны стрелкой.
Фиг.11 представляет в сочетании линейный график (А) и фотографию геля агарозы, окрашенного бромидом этидия (В), показывающие, что растворимый белок tag7 индуцирует гибель и фрагментацию ДНК в клетках L929.
(А): Клетки L929 были культивированы в присутствии tag7 (среды, кондиционированные клетками VMR-L, стимулированными с помощью LPS) или фактора некроза опухоли (TNF), с антителами анти-tag7 или без них и тестированы на гибель клеток путем окрашивания трипановым синим (кондиционированная среда и TNF) или расщепления лактатдегидрогеназой (TNF). Результаты показаны как среднее от пяти отдельных опытов, а ошибки представляют собой средние квадратические отклонения.
(В): Фотография электрофореза в 1.8% геле агарозы, окрашенном бромидом этидия, ДНК из клеток L929, которые были обработаны только TNF ("контроль") или супернатантами клеток VMR-L, стимулированных с помощью LPS ("Tag7 supe"). Маркеры калибрования положения ДНК представлены слева.
Фиг.12 относится к ингибированию развития опухолевых клеток "in vivo" с помощью генетически модифицированных клеток VMR-0.
(A): Нозерн-блоттинг гибридизация тотальной РНК, выделенной из трансфецированных и родительских клеток с помощью проб 32Р-меченной ДНК из tag7.
(B): График темпа развития опухоли, показывающий рост клеток (незакрашенные квадратики - родительские опухолевые клетки VMR-0, закрашенные треугольники - псевдо-трансфецированные VMR-0, незакрашенные кружочки - подавление роста 4SX путем инъекции поликлонального антитела анти tag7 в место расположения клеток, закрашенные квадратики - наблюдаемый эффект совместной инъекции родительских (VMR-0) и tag7 - модифицированных опухолевых клеток (4SX), незакрашенные треугольники - скорость роста клона 12SX, закрашенные кружочки - скорость роста клона 4SX.C - родительских и модифицированных (клон 4SX) опухолей, полученных путем рассечения из сингенных животных).
(C): Фотографии, показывающие размер опухолей, образованных родительскими клетками и tag7модифицированными опухолевыми клетками.
(D): Определение белка tag7 с помощью Вестерн-блоттинга (верхняя полоса) и цитотоксичности культуральной среды для клеток (нижняя полоса).
Фиг.13 показывает анализ трансформированных опухолевых клеток, анализируемых в голой мыши (недостаток Т-клеток) и SCID -мыши (недостаток Т- и В-клеток). Размер опухоли контролировали в течение 40 дней.
Фиг.14 представляет фотографию голой мыши, которой вводили клетки, экспрессирующие tag7 (4SX, верхняя полоса) и родительские клетки VMR-0 (нижняя полоса).
Фиг.15: Гистологический анализ и электронная микроскопия
(A) показывает гистологический срез опухоли, образуемой родительскими клетками (клетками VMR-0), окрашенный эозин/гематоксилином.
(B) показывает изображения, полученные электронной микроскопией митотических клеток в опухоли, образуемой родительскими клетками (клетками VMR-0).
(C) показывает изображения, полученные электронной микроскопией апоптических клеток в опухоли, образуемой клетками, модифицированными tag7 (клетки 4SX).
Фиг.16: Иммуногистохимический анализ. Распознавание эффекторных клеток.
(A) Опухоли, продуцирующие tag7 (4SX).
(B) Родительские опухоли (VMR-0).
Фиг.17: Индуцирование защитного иммунитета путем временной экспрессии tag7 в М3-клетках меланомы мыши. Выживаемость DBA/2 мыши.
Фиг.18 представляет радиоавтографию Нозерн-блоттинг-гибридизации тотальной РНК, выделенной из различных человеческих органов с помощью проб 32P-меченной ДНК из мышиного клона tag 7, демонстрирующий экспрессию гомологии tag7 в различных человеческих тканях.
Фиг.19 показывает определение tag7, экспрессируемого как фьюжн-белок GST-tag7.
Фиг.20 показывает иммунногистохимический анализ аденокарциномы легкого с помощью антител анти- tac7.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ Обзор
Для распознавания генов, обнаруживающих разный уровень экспрессии в опухолях на различных стадиях развития, согласно настоящему изобретению использовали две опухолевых линии аденокарциномы грудной железы мыши (VMR-0, имеющих низкую способность к метастазированию, и VMR-L, метастазирующих в печень с высокой частотой). С использованием технологии "выявления различий в РНК" (см. статью Liang Р. и Pardee A.B. в журнале Science 257:967-971(1992)) был получен ранее не описанный ген, экспрессированный в опухолевой линии VMR-L, который был назван tag7 и структурно охарактеризован. Хотя ген tag7 первоначально был выделен из мышиных тканей, настоящее изобретение относится также к человеческому гомологу tag7, который экспрессируется в человеческих клетках и тканях. Поэтому обычному специалисту в данной области будет понятно, что термин "tag7", как он здесь применяется, означает выделенные tac7-молекулы нуклеиновой кислоты, полинуклеотиды, полипептиды и антитела, которые могут быть мышиного или человеческого происхождения, и настоящее изобретение таким образом охватывает tog-7-молекулы нуклеиновой кислоты, полинуклеотиды, полипептиды и антитела мышиного и человеческого происхождения.
Молекулы нуклеиновой кислоты
Все нуклеотидные последовательности, кроме особо оговоренных случаев, определяемые здесь путем секвенирования молекулы ДНК, определяются секвенированием вручную, например, дидезокси-секвенированием, в соответствии с методами, которые являются обычными для специалиста в данной области (см. статьи Sanger F. и Coulson A.R. в журнале J.Mol.Biol. 94:444-448 (1975); Sanger F. и др. в журнале Proc.Natl.Acad.Sci.USA 74:5463-5467 (1977)), или автоматическим секвенированием, например с использованием с Автоматического Секвенатора для биосистем (Applied Biosystems Automated Sequenator), в соответствии с инструкциями изготовителя. Все аминокислотные последовательности полипептидов, кодированные определенными здесь молекулами ДНК, были предсказаны путем умозрительной трансляции ДНК-последовательности, определенной выше. Поэтому, как известно в данной области знаний в отношении любой ДНК-последовательности, определяемой в соответствии с такими подходами, всякая ДНК-последовательность, определяемая здесь, может содержать некоторые ошибки. Нуклеотидные последовательности, определяемые такими методами, обычно являются идентичными по меньшей мере на 90%, более типично - по меньшей мере от 95% до 99,9%, действительной нуклеотидной последовательности секвенированной молекулы ДНК. Как известно также специалисту в данной области, единичная вставка или делеция в определенной нуклеотидной последовательности по сравнению с действительной последовательностью приводит к такому сдвигу рамки в трансляции нуклеотидной последовательности, что предсказанная аминокислотная последовательность, кодируемая определенной нуклеотидной последовательностью, будет совершенно иной, чем аминокислотная последовательность, в действительности кодируемая молекулой ДНК, начиная от точки расположения такой вставки или делеции.
Если не оговорено иного, каждая "нуклеотидная последовательность", изображенная здесь, представлена как последовательность дезоксирибонуклеотидов (аббревиатуры A, G, С и Т). Однако под "нуклеотидной последовательностью" молекулы нуклеиновой кислоты или полинуклеотида подразумевается для молекулы ДНК или полинуклеотида последовательность дезоксирибонуклеотидов, а для молекулы РНК или полинуклеотида соответствующая последовательность рибонуклеотидов (A,G,C и U), где каждый дезоксирибонуклеотид тимидин (Т) в специфической дезоксирибонуклеазной последовательности замещен рибонуклеотидом уридином (U). Например, упоминание о tag7-молекуле РНК, имеющей последовательность SEQ ID NO:1, представленную с использованием дезоксирибонуклеотидных аббревиатур, как предполагается, указывает на молекулу РНК, имеющую последовательность, в которой каждый дезоксирибонуклеотид A,G или С последовательности SEQ ID NO:1 был замещен соответствующим рибонуклеотидом А, G или С и каждый дезоксирибонуклеотид Т был замещен рибонуклеотидом U.
Используя данную здесь информацию, например, о нуклеотидной последовательности на фиг.1, можно получить молекулу нуклеиновой кислоты согласно настоящему изобретению, кодирующую полипептид tag7 с применением стандартных методик клонирования и скрининга, таких, какие применяются для клонирования кДНК, когда в качестве исходного материала используются мРНК. Термин "полипептид tag7", применяемый здесь, означает полипептид или его фрагмент, которые кодируются полинуклеотидом, содержащим нуклеотидную последовательность, показанную на фиг.1 (SEQ ID NO:1), или которые имеют аминокислотную последовательность, как показано на фиг.1 (SEQ ID NO:2). Предпочтительные технологии клонирования и скрининга, используемые в настоящем изобретении, включают методы клонирования на основе PCR (полимеразной цепной реакции), таких как PCR с обратной транскриптазой (RT-PCR), с использованием праймеров, например тех, которые будут описаны ниже в примерах. На фиг.1 в качестве иллюстрации к изобретению изображена определенная нуклеотидная последовательность кодирующего сегмента (549 пар оснований) кДНК tag7 (SEQ ID NO:1). Предсказанный полипептид tag7 из 182 аминокислот, кодируемый этой кодирующей последовательностью, имеет аминокислотную последовательность, которая показана на фиг.1 (SEQ ID NO:2) и выведенный молекулярный вес около 20 кД.
Настоящее изобретение также относится к зрелой форме (формам) полипептида tag7 согласно изобретению. Полипептиды, секретируемые клетками млекопитающих, имеют сигнальную или секреторную лидерную последовательность, которая отщепляется от зрелого белка как только инициируется перенос растущей белковой цепи через шероховатый эндоплазматический ретикулум. Большинство клеток млекопитающих и даже клетки насекомых расщепляют секретируемые белки с одинаковой специфичностью. Однако в некоторых случаях расщепление секретируемого белка не совсем единообразно, что выражается в наличии двух или более зрелых семейств белков. Кроме того, давно известно, что специфичность расщепления секретируемого белка в конечном счете определяется первичной структурой полного белка, это означает, что характер расщепления определяется аминокислотной последовательностью полипептида. Как будет детально изложено в нижеследующих примерах, предсказанный полипептид tag7 имеет N-концевой гидрофобный участок примерно между 10 и 30 аминокислотами, который гомологичен сигнальным последовательностям некоторых белков, что позволяет думать, что полипептид tag7 может являться трансмембранным или секреторным белком. Поэтому настоящее изобретение относится к молекулам нуклеиновой кислоты, кодирующим зрелый полипептид tag7, имеющий аминокислотную последовательность, кодируемую полинуклеотидом, имеющим последовательность нуклеиновых кислот, показанную на фиг.1 (SEQ ID NO:1). Под зрелым полипептидом tag7, имеющим аминокислотную последовательность, кодируемую полинуклеотидом, имеющим последовательность нуклеиновых кислот, показанную на фиг.1 (SEQ ID NO:1), понимается зрелая форма (формы) полипептид a tag7, продуцируемого путем экспрессии в клетке млекопитающего (например, COS-клетке, как описано ниже) полинуклеотида, имеющего последовательность нуклеиновых кислот, показанную на фиг.1 (SEQ ID NO:1). Как показано ниже, зрелый полипептид tag7 может отличаться или не отличаться от предсказанного "зрелого" полипептида tag7, представленного на фиг.1 (SEQ ID NO:2; аминокислоты примерно от 20 до 182), в зависимости от точности предсказания сайта расщепления, основанного на компьютерном анализе.
В случае секреторного белка предсказанный сайт расщепления может быть предварительно определен по ранее описанньм правилам (см. статью Heijne и др. в журнале Eur.J.Biochem. 133:17-21 (1992)) и/или с помощью компьютерного анализа. В соответствии с диаграммой гидрофильности, полученной для полипептида tag7 (фиг.2), сайт расщепления для tag7 расположен приблизительно в аминокислотном остатке 20, хотя в зависимости от точности этого анализа сайт расщепления можно ожидать в любом месте от аминокислоты 10 до аминокислоты 30. Как будет понятно специалисту в данной области, расположение сайта расщепления, которое определяет длину сигнального пептида (в секретированном белке) или участок закрепления на мембране (в трансмембранном белке), можно подтвердить путем N-концевого секвенирования белка tag7 (натурального или полученного с помощью рекомбинантных технологий).
Как будет понятно обычному специалисту, поскольку при секвенировании возможны погрешности, а также изменение сайтов расщепления сигнальных последовательностей у различных известных белков, действительный полипетид tag7, кодируемый полинуклеотидом, изображенным на фиг.1 (SEQ ID NO:1), содержит около 182 аминокислот, но может быть где угодно в пределах примерно 150 - 190 аминокислот; и действительная N-концевая гидрофобная сигнальная последовательность этого белка содержит около 20-22 аминокислот, но может быть где угодно в пределах примерно 10-30 аминокислот.
Молекулы нуклеиновой кислоты согласно настоящему изобретению могут находиться в форме РНК, такой как мРНК, или в форме ДНК, включая, например, кДНК и геномную ДНК, полученную клонированием или произведенную синтетически. ДНК может быть двухцепочечной или одноцепочечной. Одноцепочечная ДНК или РНК может представлять собой кодирующую цепочку, также известную как смысловая цепочка, или она может быть некодирующей цепочкой, также упоминаемой как антисмысловая цепочка.
Под "выделенной" молекулой (молекулами) понимается молекула нуклеиновой кислоты, ДНК или РНК, которая была удалена из ее нативной среды. Например, молекулы рекомбинантной ДНК, содержащиеся в векторе, предполагаются выделенными для целей настоящего изобретения. Другие примеры выделенных ДНК-молекул включают молекулы рекомбинантной ДНК, содержащиеся в гетерологичных клетках-хозяевах, причем эти ДНК-молекулы очищены (частично или в значительной степени) из раствора, независимо от того, произведены ли они с помощью рекомбинантной ДНК или химическим синтезом. Выделенные молекулы РНК включают РНК-транскрипты молекул ДНК in vitro и in vivo согласно настоящему изобретению. Однако предполагается, что использованный здесь термин "выделенный" не предусматривает наличия кДНК tag7 в библиотеке кДНК или в препарате очищенной или выделенной геномной ДНК, содержащей ген tag7 или его участок в смеси с одной или более других молекул ДНК или фрагментов ДНК.
Молекулы нуклеиновой кислоты согласно настоящему изобретению далее включают генетические конструкции, содержащие, по меньшей мере, одну последовательность ДНК tag7, оперативно связанную с регуляторными последовательностями ДНК (которые могут представлять собой гетерологические регуляторные последовательности), например, промоторы или энхансеры, как будет описано ниже, причем из экспрессии этих последовательностей ДНК в клетках-хозяевах, преимущественно в клетках бактериальных, грибковых (включая дрожжевые), растительных или животных (включая клетки насекомых или млекопитающих), продуцируются один или более полипептидов tag7. В таких конструкциях регуляторные последовательности могут быть оперативно связаны с полинуклеотидом tag7, кодирующим зрелый полипептид tag7 или любые его варианты, предшественники, фрагменты или производные, которые могут включать один или более нуклеотидов, имеющих последовательность нуклеиновых кислот, комплементарную к почти всей или к участку молекулы нуклеиновой кислоты, имеющей последовательность, показанную на фиг.1 (SEQ ID NO:1) или показанную в SEQ ID NO:3. Примененный здесь термин "почти вся" по отношению к молекуле нуклеиновой кислоты или полипептиду означает участок молекулы нуклеиновой кислоты или полипептида, который содержит более чем примерно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% нуклеотидной последовательности, показанной в SEQ ID NO:l или в SEQ ID NO:3, или полипептидной последовательности, показанной в SEQ ID NO:2 или SEQ ID NO:4. Примененный здесь термин "участок" или "фрагмент" молекулы нуклеиновой кислоты или пептида означает сегмент полинуклеотида или полипептида, содержащий по меньшей мере 15, а предпочтительно по меньшей мере 20 смежных полинуклеотидов или аминокислот референс-полинуклеотида или полипептида (SEQ ID NO: 1,2,3 или 4)), если не будет специально оговорено что-нибудь иное.
Выделенные молекулы согласно настоящему изобретению включают:
(а) молекулы ДНК, кодирующие полипептид tag7 молекулы ДНК, имеющие нуклеотидную последовательность, соответствующую той, которая изображена на фиг.1 (SEQ ID NO:1) или в SEQ ID NO:3; (b) молекулы ДНК, содержащие кодирующую последовательность для полипептида tag7 показанную на фиг.1 (SEQ ID NO:2) или в SEQ ID NO:4; (с) молекулы ДНК, содержащие последовательность, существенно отличную от той, которая описана выше, но которая вследствие вырожденности генетического кода еще кодирует полипептид tag7. Поскольку генетический код является хорошо известным в данной области знаний, обычному специалисту нетрудно получить описанные выше вырожденные варианты без специальной экспериментальной работы.
Согласно другому аспекту изобретение относится к выделенной молекуле нуклеиновой кислоты, имеющей нуклеотидную последовательность, показанную на фиг.1 (SEQ ID NO:1) или в SEQ ID NO:3, или молекуле нуклеиновой кислоты, имеющей последовательность, комплементарную к почти всей такой молекуле или к ее участку. Такие выделенные молекулы, в особенности молекулы ДНК, используются в качестве проб для картирования гена путем гибридизации in situ с хромосомами и для детектирования экспрессии гена tag7 в ткани животного (главным образом млекопитающего, включая человека), особенно в опухолевых тканях и клетках, например, с помощью Нозерн-блоттинга.
Молекулы нуклеиновой кислоты согласно настоящему изобретению, которые кодируют полипептид tag7, могут включать, но не ограничиваясь этим, молекулы, сами по себе кодирующие аминокислотную последовательность зрелого полипептида; кодирующую последовательность для зрелого полипептида и дополнительные кодирующие последовательности, например, кодирующие лидерную или секреторную последовательность из примерно 20 аминокислот, такие как пре-последовательность, или про-последовательность, или пре-про-последовательность белка; кодирующую последовательность зрелого полипептида, содержащую или не содержащую вышеупомянутые дополнительные кодирующие последовательности, вместе с дополнительными некодирующими последовательностями, включая, например, интроны и некодирующие 5' и 3'-последовательности, такие как транскрибированные, нетранслированные участки (URTs) или другие 5'-фланкирующие последовательности, которые могут играть роль в транскрипции (например, через сайты, связывающие рибосому или фактор транскрипции), процессинге мРНК (например, сигналы сплайсинга и полиаденилирования) и стабилизации мРНК; кодирующую последовательность для зрелого полипептида tag7 оперативно связанную с регуляторной последовательностью ДНК, в особенности с гетерологической последовательностью ДНК, такой как промотер или энхансер; и кодирующую последовательность для зрелого полипептида tag7, связанную по меньшей мере с одной кодирующей последовательностью, которая кодирует аминокислоты, обладающие дополнительными функциями. Таким образом, последовательность, кодирующая полипептид, может быть слита с маркерной последовательностью, например, последовательностью, кодирующей пептид, который способствует очистке слитого полипептида. В некоторых вариантах осуществления этого аспекта изобретения маркерная аминокислотная последовательность может являться гекса-гистидиновым пептидом, таким как tag, представленный в векторе pQE (Qiagen, Inc.), a также находиться в ряду других, многие из которых коммерчески доступны. Как описано, например, в статье Gentz и др. в журнале Proc.Natl.Acad.Sci. USA 86:821-824 (1989), гекса-гистидин используется как подходящее средство для очистки фьюжн-белка. "НА" tag представляет собой другой пептид, применяемый для очистки и соответствующий эпитопу, полученному из белка гемагглютинина вируса гриппа, который описан у Wilson и др. в Cell 37:767 (1984). Еще одним маркерньм пептидом, используемым для очистки tag7, является глютатион S-трансфераза (GST), кодируемая фьюжн-вектором pGEX (смотри, например, Winnacker, From Genes to Clones, New York: VCH Publishers, (1987)). Как будет обсуждено ниже, и другие такие же фьюжн-белки включают tag7 соединенные с иммуноглобулином Fc на Nили С-конце.
Далее, настоящее изобретение относится к вариантам молекул нуклеиновой кислоты, которые кодируют области, аналоги или производные полипептида tag7. Варианты могут иметь природное происхождение, например природный аллельный вариант. Под "аллельным вариантом" понимается одна из нескольких альтернативных форм гена, занимающих определенное место на хромосоме в организме (см., например, у Lewin В., изд.. Genes II, John Wiley & Sons, New York (1985)). Варианты неприродного происхождения могут быть получены с использованием известных специалисту технологий мутагенеза.
Такие варианты включают те, которые получены с помощью нуклеотидных замещений, делений или добавлений. В эти замещения, делеции или добавления могут быть вовлечены один или более нуклеотидов. Варианты можно менять в кодирующих областях, некодирующих областях или в тех и других. Изменения в кодирующих областях могут продуцировать консервативные или неконсервативные аминокислотные замещения, делеции или добавления. Особенно предпочтительными среди них являются молчащие замещения, добавления и делеции, которые не изменяют свойства и активность белка или его участков. Также наиболее предпочтительными в этом отношении являются консервативные замещения.
Другие варианты осуществления изобретения охватывают выделенную молекулу нуклеиновой кислоты, содержащую полинуклеотид, имеющий нуклеотидную последовательность, по меньшей мере на 65% идентичную, а более предпочтительно по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, или по меньшей мере на 99% идентичную:
(a) нуклеотидной последовательности, представленной на фиг.1 (SEQ ID NO:l);
(b) нуклеотидной последовательности, кодирующей полноразмерный полипептид tag7 имеющий полную аминокислотную последовательность, представленную на фиг.1 (в SEQ ID NO:2), включая предсказанную сигнальную последовательность на N-конце; (с) нуклеотидной последовательности, кодирующей зрелый полипептид tag7 (полноразмерный полипептид с удаленной сигнальной последовательностью), который может, например, иметь аминокислотную последовательность, указанную в положениях приблизительно 20-182 на фиг.1 (SEQ IDNO:2);
(d) нуклеотидной последовательности полинуклеотида, кодирующего tag7, который гибридизуется при жестких условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, идентичную той, которая имеется в описанных выше выделенных молекулах нуклеиновой кислоты;
(e) нуклеотидной последовательности полинуклеотида, кодирующего tag7 который гибридизуется при определенных условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, идентичную той, которая имеется в описанных выше выделенных молекулах нуклеиновой кислоты; или
(е) нуклеотидной последовательности, комплементарной к почти всей или к участку любой из последовательностей по вышеуказанным пунктам (а), (b), (с), (d) или (е),
или их фрагмент.
Под используемым здесь термином "жесткие условия гибридизации" имеется в виду инкубирование в течение ночи при 42°С в растворе, содержащем: 50% формамида, 5xSSC (1X SSC=150 мМ NaCL, 15 мМ тринатрийцитрата), 50 мМ фосфата натрия (рН 7,6), 5х раствора Denhardt'a, 10% декстрансульфата и 20 мкг/мл денатурированной, фрагментированной ДНК спермы лосося, вслед за чем проводят промывку фильтров в 0.lx SSC при 65°С.
Под используемым здесь термином "определенные условия гибридизации" понимается предгибридизация при 65°С в течение 2 часов в Church-буфере (0,5 М фосфата натрия (рН 7,2), 7% SDS, 1мМ EDTA), денатурация при 95°С в течение пяти минут, добавление свежего Church-буфера для гибридизации при 55°С и трехкратная промывка при 50°С в течение 15 минут каждый раз промывочным Church-буфером (40 мМ фосфата натрия (рН 7,2), 1% SDS) или эквивалентные условия гибридизации в SSC или SSPE, как описано в стандартных протоколах (см., например. Molecular Cloning. A Laboratory Manual, изд.2, Sambrook J. и др., Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press (1989), которые во всей полноте приводятся в качестве ссылки).
Под полинуклеотидом, имеющим нуклеотидную последовательность, по меньшей мере на 65% "идентичную" используемой в качестве ссылки нуклеотидной последовательности, кодирующей полипептид tag7 подразумевается, что нуклеотидная последовательность полинуклеотида идентична используемой в качестве ссылки последовательности, за исключением того, что последовательность полинуклеотида может включать до 35 точковых мутаций на каждые 100 нуклеотидов используемой в качестве ссылки нуклеотидной последовательности, кодирующей полипептид tag7. Другими словами, чтобы получить полинуклеотид, имеющий нуклеотидную последовательность, по меньшей мере на 65% идентичную используемой в качестве ссылки нуклеотидной последовательности, до 5% нуклеотидов в используемой в качестве ссылки последовательности могут быть делегированы, или замещены другим нуклеотидом, или некоторое количество нуклеотидов до 35% всего их количества в используемой в качестве ссылки последовательности может быть встроено в используемой в качестве ссылки последовательность. Эти мутации используемой в качестве ссылки последовательности могут иметь место на 5' или 3'-концевых положениях используемой в качестве ссылки нуклеотидной последовательности или в любом месте между этими концевыми положениями, распределяясь либо поодиночке среди нуклеотидов используемой в качестве ссылки последовательности, либо в одной или более смежных групп в пределах используемой в качестве ссылки последовательности.
На практике, чтобы по стандартам определить, любая ли из отдельных молекул нуклеиновой кислоты является по меньшей мере на 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичной нуклеотидной последовательности, показанной на фиг.1 (SEQ ID NO:l) или в SEQ ID NO:3, можно применять известные компьютерные программы, такие как FASTA (Гейдельберг, Германия), BLAST (Вашингтон, Федеральный округ Колумбия) или BESTFIT (Wisconsin Sequence Analysis Package, версия 8 для Unix, Genetics Computer Group, University Research Park, 575 Science Drive, Madison, WI 53711), которая использует алгоритм локальной гомологии (см. Smith и Waterman, Advances in Applied Mathematics 2:482-489 (1981)) для того, чтобы найти сегмент наилучшей гомологии между последовательностями. При использовании FASTA, BLAST, BESTFIT или любой другой программы выравнивания последовательностей для определения того, является ли конкретная последовательность, например, на 65% идентичной используемой в качестве ссылки последовательности в соответствии с настоящим изобретением, устанавливают такие параметры, чтобы процентное значение идентичности подсчитывалось по всей длине используемой в качестве ссылки нуклеотидной последовательности и чтобы позволялись пропуски в гомологии до 35% общего количества нуклеотидов в используемой в качестве ссылки последовательности.
Настоящее изобретение касается молекул нуклеиновой кислоты, по меньшей мере на 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичных последовательности нуклеиновых кислот, показанной на фиг.1 (SEQ ID NO:1) или в SEQ ID NO:3, и их фрагментов, независимо от того, кодируют ли они полипептид, обладающий tag7-активностью. Поэтому, даже если конкретная молекула нуклеиновой кислоты не кодирует полипептид, обладающий tag7-активностью, специалист в данной области знает, как использовать молекулу нуклеиновой кислоты, например, в качестве пробы при гибридизации или праймера цепной полимеразной реакции (PCR). Возможности применения молекул нуклеиновой кислоты согласно настоящему изобретению, которые не кодируют полипептид, проявляющий tag7-активность, включают, среди прочих, (1) выделение гена tag7 или его аллельных вариантов в библиотеке геномных ДНК; (2) гибридизацию in situ (например "FISH") с метафазным хромосомным веретеном для определения точного расположения гена tag7 на хромосоме, как описано для локализации человеческого гена у Verma и др., Human Chromosomes: A Manual of Basic Techniques, Pergamon Press, New York (1988); и (3) Нозерн-блоттинг для регистрации экспрессии мРНК tag7 в отдельных тканях.
Конечно, любой обычный специалист в данной области сразу поймет, что, вследствие вырожденности генетического кода, большое количество молекул нуклеиновой кислоты, имеющих последовательность, по меньшей мере на 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичную последовательности нуклеиновых кислот, показанной на фиг.1 (SEQ ID NO:1) или в SEQ ID NO:3, и их фрагментов будет кодировать полипептид, обладающий структурой и/или активностью полипептида tag7. Действительно, поскольку все вырожденные варианты этих нуклеотидных последовательностей кодируют одинаковый полипептид, это специалисту в данной области будет ясно и без выполнения вышеописанных сравнительных анализов. Кроме того, любой специалист поймет, что из тех нуклеиновых кислот, которые не являются вырожденными вариантами, приемлемое количество также будет кодировать полипептид, обладающий tag7-структурой и/или активностью. Учитывается также, что опытный специалист полностью осведомлен о заместителях аминокислот, от которых менее вероятно или совсем невероятно ожидать значительного влияния на функции белка (в частности, замены одной алифатической аминокислоты другой алифатической аминокислотой). Например, указания в отношении того, как выполнить фенотипически молчащие замещения аминокислот, даются в статье Bowie J.U. и др.. в журнале Science 247:1306-1310 (1990) и в приведенных там ссылочных материалах.
Как указано выше, изобретение относится также к фрагментам описанных молекул нуклеиновой кислоты. Предпочтительные фрагменты нуклеиновых кислот согласно настоящему изобретению включают выделенные молекулы нуклеиновой кислоты, кодирующие участки полипептида tag7, несущие эпитоп. В частности, такие фрагменты нуклеиновой кислоты согласно настоящему изобретению кодируют: полипептид, содержащий аминокислотные остатки примерно от 20 до 40 на фиг.1 (SEQ ID NO:2); полипептид, содержащий аминокислотные остатки примерно от 55 до 75 на фиг.1 (SEQ ID NO:2); полипептид, содержащий аминокислотные остатки примерно от 90 до 110 на фиг.1 (SEQ ID NO:2); полипептид, имеющий аминокислотную последовательность, состоящую в основном из аминокислотных остатков примерно от 145 до 160 на фиг.1 (SEQ ID NO:2). Изобретатели также установили, что указанные полипептидные фрагменты являются антигенными областями предсказанного полипептида tag7. Способы определения других таких участков полипептида tag7, несущих эпитоп, которые можно также использовать для определения несущих эпитоп участков человеческого полипептида tag7, подробно описаны ниже.
В другом аспекте изобретение касается выделенной молекулы нуклеиновой кислоты, содержащей полинуклеотид, который гибридизуется при строгих условиях гибридизации с практически целым полинуклеотидом или с участком полинуклеотида в описанной выше молекуле нуклеиновой кислоты, например молекуле нуклеиновой кислоты, имеющей нуклеотидную последовательность, представленную на фиг.1 (SEQ ID NO:1) или в SEQ ID NO:3.
Еще в одном аспекте изобретение касается выделенной молекулы нуклеиновой кислоты, содержащей полинуклеотид, который гибридизуется при определенных условиях гибридизации с практически целым полинуклеотидом или с участком полинуклеотида в описанной выше молекуле нуклеиновой кислоты, например, молекуле нуклеиновой кислоты, имеющей нуклеотидную последовательность, представленную на фиг.1 (SEQ ID NO:1) или предпочтительно представленную в SEQ ID NO:3, и который кодирует полипептид с активностью tag7, предпочтительно человеческий полипептид tag 7.
Под нуклеотидом, который гибридизуется с "участком" полинуклеотида, имеется в виду полинуклеотид (как ДНК, так и РНК), гибридизующийся по меньшей мере с примерно 15 нуклеотидами, более предпочтительно по меньшей мере с примерно 20 нуклеотидами, еще более предпочтительно по меньшей мере примерно с 30 нуклеотидами, а еще более предпочтительно по меньшей мере примерно с 30-70 нуклеотидами, используемого в качестве ссылки полинуклеотида. Эти гибридизующиеся полинуклеотиды используются в качестве диагностических проб и праймеров, как было указано выше и более подробно будет рассмотрено ниже.
Конечно, полинуклеотиды, гибридизующиеся с большим участком используемого в качестве ссылки полинуклеотида (например, молекулой нуклеиновой кислоты, содержащей последовательность, кодирующую tag7 и имеющую нуклеотидную последовательность, представленную на фиг.1 (SEQ ID NO:1) или в SEQ ID NO:3), в частности с участком длиной примерно в 50-500 нуклеотидов или даже со всей длиной используемого в качестве ссылки нуклеотида, также используются в качестве проб согласно настоящему изобретению, так же как полинуклеотиды, соответствующие большей части, но не всей нуклеотидной последовательности из нуклеотидной последовательности, представленной на фиг.1 (SEQ ID NO:1) или в SEQ ID NO:3. Под участком полинуклеотида, например, "длиной по меньшей мере 20 нуклеотидов" подразумеваются 20 или более смежных нуклеотидов из нуклеотидной последовательности используемого в качестве ссылки полинуклеотида (например, нуклеотидной последовательности, представленной на фиг.1 (SEQ ID NO:1) или в SEQ ID NO:3). Как уже отмечено, такие участки имеют диагностическое применение либо в качестве проб в соответствии с известными технологиями гибридизации ДНК или в качестве праймеров для амплификации последовательности-мишени путем полимеразной цепной реакции (PCR), как описано, например, в Molecular Cloning. A Laboratory Manual, изд.2, Sambrook J., Fritish E.F. и Маniatis Т., изд. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press (1989), которые во всей полноте приводятся в качестве ссылки.
Поскольку определенная нуклеотидная последовательность, кодирующая полипептид tag7, приводится на фиг.1 (SEQ ID NO:1) или в SEQ ID NO:3, производные полинуклеотиды, которые гибридизуются с участком молекулы кДНК tag7, являются понятными для специалиста в данной области. Например, расщепление с помощью рестрикционной эндонуклеазы или разрушение путем ультразвуковой обработки клона кДНК tag7 можно без труда применить для получения участков ДНК различного размера, которые и являются полинуклеотидами, гибридизующимися с участком молекулы кДНК tag7. С другой стороны, гибридизующиеся полинуклеотиды по настоящему изобретению можно получить с помощью синтеза по известным технологиям. Следовательно, хотя настоящее изобретение относится к молекулам нуклеиновой кислоты tag7 и полипептидам из мыши и человека, любой из обычных специалистов в данной области легко может получить и/или выделить гомологи молекул нуклеиновой кислоты tag7 и полипептидов по настоящему изобретению из других организмов (в особенности других млекопитающих), используя нуклеотидную последовательность tag7 (SEQ ID NO:l) и последовательность аминокислот (SEQ ID NO:2), описанные здесь, и обычные методы молекулярной биологии, например скрининг библиотек кДНК, которые хорошо известны в данной области и описаны в стандартных протоколах (см., например, Molecular Cloning. A Laboratory Manual, изд. 2, Sambrook J. и др., изд. Соld Spring Harbor, New York: Cold Spring Harbor Laboratory Press (1989)). Например, подходящей библиотекой кДНК для получения молекулы нуклеиновой кислоты, кодирующей человеческий гомолог tag7 (далее обозначаемый как человеческий tag7), является библиотека кДНК костного мозга. Библиотеки кДНК костного мозга и кДНК других тканей человека, а также геномные библиотеки, коммерчески доступны, например, от Clontech (Palo Alto, Калифорния).
Векторы и клетки-хозяева
Настоящее изобретение также относится к генетическим конструкциям, содержащим выделенные молекулы нуклеиновой кислоты согласно изобретению или их фрагменты, оперативно связанные с регуляторной последовательностью ДНК, как будет подробно описано ниже, векторам, которые содержат эти генетические конструкции или выделенные молекулы ДНК согласно изобретению, и клеток-хозяев, содержащих такие векторы. Кроме того, изобретение касается способов получения полипептидов tag7 или их фрагментов с помощью рекомбинантных технологий с применением этих векторов и клеток-хозяев.
Векторы, содержащие генетические конструкции или выделенные молекулы ДНК согласно настоящему изобретению могут быть введены в клетки-хозяева с использованием хорошо известных методов, таких как инфекция, трансдукция, трансфекция, электропорация и трансформация. Вектор может являться, например, фагом, плазмидой, вирусом или ретровирусом и предпочтительно является вектором экспрессии, как описано ниже. Ретровирусные векторы могут быть компетентные или дефектные по репликации. В последнем случае размножение вируса может иметь место только в комплементирующих клетках-хозяевах.
Полинуклеотиды можно присоединить к вектору, содержащему выбранный маркер для размножения в хозяине. Плазмидный вектор главным образом вводят внутрь клеток млекопитающих или птиц в виде осадка, такого как кальций-фосфатный осадок, или в комплексе с заряженным липидом (например, LIPOFESTAMINETM; Life Technologies, Inc.; Rockville, Мэриленд), или в комплексе с вирусом (таким как аденовирус; см. патенты US №5547932 и №5521291), или с компонентами вируса (такими как вирусные капсидные пептиды). Если вектор представляет собой вирус, он может быть упакован in vitro с использованием подходящей упаковочной клеточной линии и затем трансдуцирован в клетки-хозяева.
Предпочтительными являются векторы, содержащие контролирующие области с cis-действием на нужные полинуклеотиды. Соответствующие факторы с trans-действием могут быть обеспечены клеткой-хозяином, комплементирующим вектором или самим вектором при введении в клетку-хозяин.
В некоторых предпочтительных вариантах осуществления этого аспекта векторы обеспечивают специфическую экспрессию, которая может быть индуцированной и/или быть спецификой типа клетки. Особенно предпочтительными среди таких векторов экспрессии являются те, которые индуцируются факторами окружающей среды, которыми можно легко управлять, такими как температура и питательные добавки.
Векторы экспрессии, используемые в настоящем изобретении, включают векторы хромосомного, эписомального или вирусного происхождения, например, векторы, происходящие из бактериальных плазмид, бактериофагов, эписомов дрожжей, хромосомных элементов дрожжей, вирусов, таких как бакуловирусы, паповавирусы, вирусы вакцин, аденовирусы, вирусы птичьего сифилиса, вирусы псевдобешенства и ретровирусы, и векторы, происходящие из их комбинаций, таких как космиды и фагмиды,
В одном из вариантов осуществления настоящего изобретения выделенная молекула нуклеиновой кислоты согласно изобретению или ее фрагмент могут быть оперативно связаны с соответствующей регуляторной последовательностью, предпочтительно промотором, например промотором лямбда фага PL, промоторами из фагов ТЗ, Т7 и SP6, lac, trp и tac промоторов E.coli, ранних и поздних промоторов SV40 и промоторов ретровирусных LTR и их производных, причем названы лишь немногие как пример. Другие подходящие промоторы известны опытному специалисту. Конструкции экспрессии содержат далее сайты инициации транскрипции, терминации и в транскрибированной области сайт связывания рибосомы для трансляции. Кодирующий участок зрелых транскриптов, экспрессируемых конструкциями, предпочтительно включает кодон инициации трансляции (AUG) в начале полипептида, подлежащего трансляции, и кодон терминации (UAA, UGA или UAG), соответственно расположенный на его конце.
Как отмечено выше, векторы экспрессии предпочтительно включают по меньшей мере один выбранный маркер. Такие маркеры содержат гены устойчивости к дигидрофолатредуктазе (dhfr) или неомицину (nео) для культур эукариотических клеток и к тетрациклину (tef) или ампициллину (amp) для культивируемых клеток E.coli и других бактерий. Типичными примерами подходящих хозяев являются, не ограничиваясь этим, бактериальные клетки, такие как клетки Escherichia spp. (в особенности E.coli), клетки Bacillus spp. (в особенности В. cereus, В. subtilis и В. megaterwm), клетки Streptomyces spp., клетки Salmonella spp. (в особенности S. typhimirium) и клетки Xanthomonas spp.; грибковые клетки, включая дрожжевые клетки, такие как клетки Saccharomyces spp.; клетки насекомых, например клетки Drosophila S2, Spodoptera Sf9 или Sf21 и клетки Trichoplusa High-five; клетки других животных (в особенности клетки млекопитающих и наиболее предпочтительно человеческие клетки), такие как СНО, COS, VERO, HeLa, клетки меланомы Bowes, а также HepG2 и другие клеточные линии печени; клетки высших растений. Подходящие культуральные среды и условия для вышеописанных клеток-хозяев хорошо известны в данной области знаний.
Предпочтительно используемые векторы в бактериях включают pQE70, pQE60 и рQЕ-9, поставляемые Qiagen, векторы pBS, векторы Phabescript, векторы Bluescript, pNH8A, pNH16a, pNH18A и pNH46A, поставляемые Stratagene; pcDNA3, поставляемые Invitrogen; pGEX, pTrxfus, pTrc99a, pET-9, pKK223-3, pKK233-3, pDR540 и pRIT5, поставляемые Pharmacia. Среди предпочтительных эукариотических веторов находятся pWLNEO, pSV2CAT. pOG44, pXTl, pBK и pSG, поставляемые Stratagene; pSVK3, pBPV, pMSG и pSVL, поставляемые Pharmacia. Другие приемлемые векторы могут быть легко понятны опытному специалисту.
Известные бактериальные промоторы, подходящие для использования в настоящем изобретении, включают lacI и lacZ промоторы E.coli, фаговые промоторы Т3,Т7 и SP6, gpt-промотор, промоторы лямбда PR и PL trp-промотор. Приемлемые эукариотические промоторы включают немедленно ранний промотор CMV, промотор тимидинкиназы HSV (вируса простого герпеса), ранний и поздний промоторы SV40, промоторы ретровирусных LTR, например вируса саркомы Rous (RSV), и металлотионеиновые промоторы, такие как мышиный металлотионеин-I-промотор.
Введение конструкции в клетку-хозяин может быть осуществлено путем трансфекции фосфатом кальция, трансфекции, опосредованной DEAE-декстраном, трансфекции, опосредованной катионным липидом, электропорации, трансдукции, инфекции, бомбардировки частицами с покрытием из нуклеиновой кислоты или другими методами. Такие методы описаны во многих стандартных руководствах по лабораторным работам, например, Davis и др. Basic Methods In Molecular Biology (1986).
В некоторых вариантах осуществления настоящего изобретения выделенные полинуклеотиды согласно настоящему изобретению могут быть оперативно связаны с регуляторной последовательностью гена, которая может быть гомологичной или гетерологичной регуляторной генетической последовательностью (такой как энхансер, промотор или репрессор), для образования генетической конструкции. Под генетическими конструкциями в соответствии с этим аспектом настоящего изобретения понимаются не только те, которые содержат полинуклеотид, кодирующий зрелый белок tag7, оперативно связанный с регуляторной последовательностью ДНК, но также и те конструкции, которые содержат одну или более регуляторных последовательностей, оперативно связанных с фрагментом полинуклеотида tag7, не кодирующего зрелый белок tag7, но содержащего достаточной величины фрагмент нуклеотидной последовательности tag7 ("нацеливающий фрагмент"), для того чтобы нацеливать генетическую конструкцию на локус нативного tag7 при введении в клетку-хозяина, где tag7 ген может инактивироваться репрессией или мутацией. Эти конструкции могут быть вставлены в вектор, как описано выше, и вектор вводится в клетку-хозяина, геном которой содержит ген-мишень, любым из описанных выше методов. Полинуклеотид tag7 затем интегрирует в геном клетки-хозяина путем гомологичной рекомбинации. В случае конструкции, содержащей гомологичную или гетерологичную регуляторную последовательность, связанную с нацеливающим фрагментом полинуклеотида tag7, регуляторная последовательность будет нацелена на локус нативного tag7 в клетке-хозяине и будет амплифицировать или дерепрессировать (если регуляторная последовательность содержит, например, промотор или энхансер) или будет ингибировать или подавлять (если регуляторная последовательность содержит, например, репрессор или иным способом интегрируется в нативную регуляторную последовательность для ингибирования или репрессирования (например, "ударным действием")) экспрессию нативного гена tag7 в клетке-хозяине, благодаря чему уровень экспрессии гена tag7 повышается или понижается. В свою очередь такое генетическое нацеливание может быть выполнено с использованием генетических конструкций, содержащих вышеописанный нацеливающий фрагмент tag7 и в отсутствие регуляторной последовательности; такой подход может быть применен, например, для корректировки или введения точковых мутаций в гене tag7 (см. статью Steeg С.М.и др. в журнале Proc. Natl. Acad. Sci. USA 87(12): 4680-4684 (1990), где дано описание применения таких подходов для корректировки точковых мутаций в других генах млекопитающих). Эти методы продуцирования генетических конструкций, введения нужных генов в клетки-хозяева посредством гомологичной рекомбинации и получения кодированных полипептидов в целом описаны в патентах US №5578461; WO 94/12650 (патентная заявка US №07/985586); WO 93/09222 (патентная заявка US №07/911535); WO 90/14092 (патентная заявка US №07/353909), содержание которых во всей его полноте приводится в виде ссылки).
Транскрипция ДНК, кодирующей полипептиды согласно настоящему изобретению, у высших эукариотов может быть увеличена путем встраивания в вектор энхансерной последовательности. Энхансеры представляют собой элементы ДНК с cis-действием, обычно от 10 до 300 пар оснований, действие которых заключается в увеличении транскрипционной активности промотора в данном типе клеток-хозяев. Примерами энхансера являются энхансер SV40, который локализован в поздней области от начала репликации на участке 100-270 нп, энхансер цитомегаловирусного раннего промотора, полиомный энхансер, расположенный в поздней области от начала репликации и аденовирусные энхансеры. В другом варианте осуществления изобретения транскрипционная активация гена tag7 может быть усилена с помощью встраивания в вектор по меньшей мере одного конкатемеризованного элемента из нативного человеческого или tag7-промотора.
Для секреции транслированного полипептида в просвет эндоплазматического ретикулума в периплазматическое пространство или во внеклеточную окружающую среду можно в экспрессируемый полипептид включить соответствующие сигналы секреции. Сигналы могут быть эндогенными по отношению к полипептидам или они могут представлять собой гетерологические сигналы.
Полипептид tag7 может быть экспрессирован в модифицированной форме, такой как фьюжн-белок, и может включать не только сигналы секреции, но также дополнительные гетерологические функциональные области. Например, с N-конца полипептида можно добавить область дополнительных аминокислот, в частности заряженных аминокислот, для улучшения стабильности и сохранения эффективности в клетке-хозяине в процессе очистки или при последующих использовании или хранении. Также для облегчения очистки в полипептид можно добавить части пептида. Такие участки могут быть удалены перед конечным получением полипептида. Добавление частей пептида в полипептид для возбуждения секреции или выделения с целью улучшить стабильность или облегчить очистку, а также для других целей, является хорошо знакомой и обычной практикой для специалиста. Предпочтительный фьюжн-белок содержит гетерологическую область из иммуноглобулина, которая используется для солюбилизации белков. Например, в ЕР 0464533 описаны фьюжн-белки, содержащие различные участки константной области (Fc) молекул иммуноглобулина вместе с другим человеческим белком или его частью. Во многих случаях Fc -участок фьюжн-белка вполне пригоден для использования его в терапии и диагностике и обеспечивает таким образом, например, улучшенные фармакокинетические качественные показатели (ЕР 0232262). С другой стороны, для такого использования было бы желательно обеспечить удаление Fc-части фьюжн-белка после того, как произошли экспрессия фьюжн-белка, его регистрация и очистка ранее описанными выгодными способами. Это тот случай, когда Fc-участок, как доказано, является препятствием для применения фьюжн-белка в терапии, диагностике или других производствах, например, когда он должен быть использован в качестве антигена для иммунизации при получении антител.
Полипептид tag7 можно выделить и очистить из рекомбинантных клеточных культур с помощью хорошо известных методов, включая осаждение сульфатом аммония или этанолом, кислотную экстракцию, анион- или катионообменную хромотографию, лектинхромотографию, гель-фильтрацию, хромотографию на основе гидрофобных взаимодействий, аффинную хромотографию, хромотографию на основе гидроксилапатитовых смол. Наиболее предпочтительно использовать для очистки высокоэффективную жидкостную хроматографию (ВЭЖХ). Полипептиды согласно настоящему изобретению включают натурально очищенные продукты, продукты, полученные в результате процессов химического синтеза и продукты, произведенные рекомбинантными методами из прокариотических или эукариотических хозяев, включая, например, бактериальные и дрожжевые клетки, клетки насекомых, млекопитающих, птиц и высших растений. В зависимости от типа используемого хозяина в технологии рекомбинантного получения полипептиды согласно настоящему изобретению могут быть гликозилированы или не гликозилированы. Кроме того, полипептиды tag7 по изобретению могут также включать исходный модифицированный остаток метионина, в некоторых случаях как результат процессов, опосредованных клеткой-хозяином.
Полипептиды tag7 и фрагменты
Изобретение далее относится к выделенному полипептиду tag7, имеющему аминокислотную последовательность, кодируемую полинуклеотидом, имеющим нуклеотидную последовательность, представленную на фиг.1 (SEQ ID NO:1), к выделенному полипептиду tag7, имеющему аминокислотную последовательность, кодируемую полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:3, полноразмерную аминокислотную последовательность на фиг.1 (SEQ ID NO:2), аминокислотную последовательность зрелого полипептида tag7, имеющего аминокислотную последовательность, представленную в положениях от 20 до 182 на фиг.1 (SEQ ID NO: 2), аминокислотную последовательность, кодируемую полинуклеотидом, который гибридизуется при жестких условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:1, аминокислотную последовательность, кодируемую полинуклеотидом, который гибридизуется при определенных условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:l, или фрагмент вышеупомянутых полипептидов. Используемые здесь термины "пептид" и "олигопептид" являются синонимами (как это обычно признается), и эти термины могут быть использованы как взаимозаменяемые, когда контекст требует обозначить цепочку из по меньшей мере двух аминокислот, связанных посредством (а) пептидильной связи (связей). Термин "полипептид" применяется здесь для обозначения цепочек, содержащих по меньшей мере десять аминокислотных остатков. Как обычно принято в данной области, все формулы или последовательности олигопептидов и полипептидов пишутся слева направо и в направлении от амино-конца к карбокси-концу.
Обычному специалисту в данной области будет понятно, что некоторые аминокислотные последовательности полипептида tag7 могут варьироваться без заметного влияния на структуру или функцию полипептида. Если такие различия в последовательности являются намеренными, следует помнить, что на белке имеются критические области, которые определяют структуру и активность. В основном, возможна замена остатков, которые формируют третичную структуру, при условии, что используются остатки, выполняющие подобную функцию. В других примерах тип остатка может быть совершенно неважен, если изменение имеет место в некритической области полипептида.
Поэтому изобретение далее относятся к вариантам полипептида tag7, включая аллельные варианты, которые обнаруживают существенную гомологию с полипептидом tag7 в структуре или активности или которые содержат области полипептида tag7, например участки, обсуждаемые ниже. Такие мутанты могут включать делеции, вставки, инверсии, повторы и типовые замещения (например, гидрофильный остаток, замещающий другой, но, как правило, без замены в высокой степени гидрофильньм остатком в высокой степени гидрофобного). Небольшие изменения или такие "нейтральные" аминокислотные замещения оказывают, в общем, малое влияние на активность.
Типичными консервативными замещениями являются взаимозамены среди алифатических аминокислот Ala, Val, Leu и Ile; взаимозамена гидроксилированных остатков Ser и Thr; замена кислотных остатков Asp и Glu; замещение один другим амидированных остатков Asn и Gin; замена основных остатков Lys и Arg; замещение один другим ароматических остатков Phe и Туг.
Таким образом, фрагмент, производное или аналог полипептида на фиг.1 (SEQ ID NO:2), или полипептид по SEQ ID NO:4, или тот, который кодируется полинуклеотидом, имеющим последовательность нуклеиновых кислот, как показано на фиг.1 (SEQ ID NO: 1) или в SEQ ID NO:3, могут быть такими (i), у которых по меньшей мере один из аминокислотных остатков замещен консервативным или не консервативным аминокислотным остатком (предпочтительно консервативным аминокислотным остатком) и такой замещенный аминокислотный остаток может быть кодирован генетическим кодом или может являться аминокислотой (например, десмозином, цитруллином, орнитином и т.д.), которая не кодирована генетическим кодом; такими (ii), у которых по меньшей мере один из аминокислотных остатков содержит замещающую группу (например, фосфатную, гидроксильную, сульфатную или другую группу) в дополнение к нормальной группе "R" аминокислоты; такими (iii), у которых зрелый полипептид соединен с другим соединением, например, соединением, увеличивающим период полужизни полипептида (например, полиэтиленглйколем), или (iv) тот, у которого дополнительные аминокислоты слиты со зрелым полипептидом, таким как пептид, являющийся Fc-областью иммуноглобулина, лидерной или секреторной последовательностями, последовательностью, которая используется для очистки зрелого полипептида (такого как GST) или последовательностью пробелка. Имеется в виду, что такие фрагменты, производные или аналоги охватываются настоящим изобретением и находятся в пределах опыта специалиста в данной области, почерпнутого из руководств и состояния знаний на момент создания изобретения.
Полипептиды согласно настоящему изобретению предпочтительно представлены в выделенной форме и предпочтительно в значительной степени очищены. Полученный методами рекомбинантных технологий вариант полипептида tag7 может быть в основном очищен одноступенчатым методом, описанным в статье Smith и Johnson в журнале Gene 67: 31-40 (1998). Термин "в основном очищенный", использованный здесь, означает препарат полипептида tag7, в котором по меньшей мере 50%, предпочтительно по меньшей мере 70%, а более предпочтительно 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% загрязняющих белков (т.е. тех, которые не являются белком tag7), удалены из препарата.
Полипептиды по настоящему изобретению включают те, которые по меньшей мере на 65% идентичны, а предпочтительно по меньшей мере на 70%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, 98% или по меньшей мере на 99% идентичны полипептиду, кодируемому полинуклеотидом, имеющим последовательность нуклеиновых кислот, представленную на фиг.1 (SEQ ID NO:l) или в SEQ ID NO:3, полипептиду, имеющему полноразмерную аминокислотную последовательность, представленную на фиг.1 (SEQ ID NO:2) или в SEQ ID NO:4, зрелому полипептиду tag7, имеющему аминокислотную последовательность, представленную в положениях от 20 до 182 на фиг.1 (SEQ ID NO:2), полипептиду, кодируемому полинуклеотидом, который гибридизуется при жестких условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:1, или полипептиду, кодируемому полинуклеотидом, который гибридизуется при определенных условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:1. Данные полипептиды также включают участки или фрагменты из по меньшей мере 30 аминокислот, а более предпочтительно из по меньшей мере 50 аминокислот.
Под полипептидом, имеющим аминокислотную последовательность, по меньшей мере на 65% "идентичную" используемой в качестве ссылки последовательности полипептида tag7, подразумевается, что аминокислотная последовательность полипептида идентична референс-последовательности, за исключением того, что полипептидная последовательность может включать до 35 аминокислотных изменений на каждые 100 аминокислот используемой в качестве ссылки последовательности полипептида tag7. Другими словами, чтобы получить полипептид, имеющий аминокислотную последовательность, по меньшей мере на 65% идентичную используемой в качестве ссылки последовательности, до 35% аминокислотных остатков в используемой в качестве ссылки последовательности могут быть делегированы или замещены другой аминокислотой или до 35% общего числа аминокислотных остатков в используемой в качестве ссылки последовательности можно встроить в используемую в качестве ссылки последовательность. Эти изменения в используемой в качестве ссылки последовательности могут быть в амино-(N-) или карбокси-(С-) концевых положениях используемой в качестве ссылки аминокислотной последовательности или могут находиться где-нибудь между этими концевыми положениями, распределяясь либо поодиночке среди остатков в используемой в качестве ссылки последовательности, либо в по меньшей мере одной из смежных групп внутри используемой в качестве ссылки последовательности.
Полипептиды согласно настоящему изобретению можно применять в качестве маркеров молекулярного веса на гелях при электрофорезе в полиакриламидном геле с добавлением додецилсульфата натрия (SDS-PAGE) или на молекулярных ситах для гель-фильтрационных колонок с использованием методов, хорошо известных в этой области. Кроме того, как будет подробно изложено ниже, полипептиды согласно настоящему изобретению можно использовать для взаимодействия с поликлональными или моноклональными антителами, которые применяются в анализах для регистрации экспрессии белка tag7 в качестве антагонистов, способных ингибировать функцию белка tag7, в терапевтических способах для ингибирования или задержки роста раковых метастазов или для выделения белка tag7.
В другом аспекте настоящее изобретение относится к пептиду или полипептиду, содержащим несущий эпитоп участок полипептида по изобретению, которые могут быть использованы для взаимодействия с антителами, в особенности моноклональными антителами, которые специфически связывают полипептид tag7 согласно изобретению. Эпитоп участка этого полипептида является иммуногенным или антигенным эпитопом полипептида согласно изобретению. "Иммуногенный эпитоп" означает часть белка, которая вызывает реакцию антитела, когда весь белок является иммуногенным. Эти иммуногенные эпитопы, как полагают, ограничены немногими местами на молекуле. С другой стороны, область белковой молекулы, с которой может быть связано антитело, определена как "антигенный эпитоп". Количество иммуногенных эпитопов белка в основном меньше, чем количество антигенных эпитопов (см., например, статью Geysen и др. в журнале Proc.Natl.Acad.Sci. USA 81:3998-4002 (1983)).
Что касается отбора пептидов или полипептидов, несущих антигенный эпитоп (т.е. тех, которые содержат в белковой молекуле область, с которой может связываться антитело), то хорошо известно в данной области знаний, что относительно короткие синтетические пептиды, которые имитируют часть последовательности белка, обычно способны приводить к получению антисыворотки, которая реагирует с частично имитированным белком (см. статью Sutcliffe J.G. и др. в журнале Science 219:660-666 (1983)). Пептиды, способные приводить к получению белок-реактивных сывороток, часто представлены в праймерной последовательности белка, могут быть охарактеризованы набором простых химических норм и не ограничиваются иммунодоминантными областями неповрежденных белков (т.е. иммуногенными эпитопами) или амино- или карбоксиконцами. Пептиды, которые чрезвычайно гидрофобны и те из них, которые состоят из шести или менее остатков, не являются эффективными при индуцировании антител, которые связываются с имитированным белком; более длинные пептиды, особенно те, которые содержат пролиновые остатки, более эффективны (см. статью Sutcliffe J.G. и др. в журнале Science 219:660-666 (1983)).
Несущие эпитоп пептиды и полипептиды согласно изобретению, составленные в соответствии с вышеуказанными указаниями, предпочтительно содержат последовательность из по меньшей мере семи, более предпочтительно из девяти, а наиболее предпочтительно примерно из 15-30 аминокислот, содержащихся в аминокислотной последовательности полипептида согласно изобретению. Однако пептиды и полипептиды, содержащие более длинные участки аминокислотной последовательности полипептида согласно настоящему изобретению, содержащие примерно от 30 до 50 аминокислот, или участки любой длины до полной аминокислотной последовательности полипептида согласно изобретению и включая ее, также являются желательньми эпитоп-несущими пептидами и полипептидами согласно изобретению и также могут быть использованы для выработки антител, которые реагируют с имитированньм белком. Предпочтительно аминокислотная последовательность эпитоп-несущего пептида выбирается так, чтобы обеспечить существенную растворимость в водных растворителях (т.е. последовательность включает относительно гидрофильные остатки, а высокогидрофобных последовательностей избегают); последовательности, содержащие пролиновые остатки, особенно предпочтительны.
Не ограничивающие объем изобретения примеры эпитоп-несущих пептидов или полипептидов, которые можно использовать для продуцирования tac7-специфичных антител, включают полипептид, имеющий аминокислотную последовательность, состоящую в основном из примерно 20-40 аминокислотных остатков на фиг.1 (SEQ ID NO:2), полипептид, имеющий аминокислотную последовательность, состоящую в основном из примерно 55-75 аминокислотных остатков на фиг.1 (SEQ ID NO:2), полипептид, имеющий аминокислотную последовательность, состоящую в основном из примерно 90-110 аминокислотных остатков на фиг.1 (SEQ ID NO:2), и полипептид, имеющий аминокислотную последовательность, состоящую в основном из примерно 145-165 аминокислотных остатков на фиг.1 (SEQ ID NO:2). Другие эпитоп-несущие полипептиды и пептиды, которые можно использовать для продуцирования tag7-специфичных антител, будут очевидны для обычного специалиста в данной области на основе диаграммы гидрофильности полипептида tag7, показанного на фиг.2.
Эпитоп-несущие пептиды и полипептиды по изобретению могут быть получены любыми подходящими способами производства пептидов или полипептидов, включая рекомбинантные способы, использующие молекулы нуклеиновой кислоты согласно изобретению. Например, короткая эпитоп-несущая аминокислотная последовательность может быть соединена с большим по величине полипептидом, который действует как носитель в процессе рекомбинантного продуцирования и очистки, а также в процессе иммунизации, для получения антипептидных антител. Эпитоп-несущие пептиды также можно синтезировать, используя известные методы химического синтеза (см., например, патент US №4 631 211; статью Houghten R.A. в журнале Proc.Natl.Acad. Sci. USA 52:5131-5135(1985)).
Как будет понятно специалисту в данной области, полипептиды tag7 согласно настоящему изобретению и их эпитоп-несущие фрагменты могут быть иммобилизованы на твердой подложке с помощью технологий, которые являются хорошо известными и обычными. Под "твердой подложкой" подразумевается любая подложка, на которой может быть иммобилизован пептид. Такие твердые подложки включают, не ограничиваясь этим, нитроцеллюлозу, диазоцеллюлозу, стекло, полистирол, поливинилхлорид, полиэтилен, декстран, сефарозу, агар, найлон, шарики и планшеты для микротитрования. Связь пептидов согласно изобретению с твердой подложкой может быть достигнута путем прикрепления одного или обоих концов полипептида к подложке. Прикрепление также может быть сделано и на одном или нескольких внутренних участках пептида. В соответствии с настоящим изобретением можно также применить множественное крепление (как на средних участках, так и на концах пептида). Прикрепление может осуществляться через связующую группу аминокислоты, такую как первичная аминогруппа, карбоксигруппа или сульфгидрильная группа (SH), или с помощью групп химической связи, такой как бромциановая связь (CNBr), а также через спейсер. Для нековалентных присоединений может быть использовано добавление к пептиду аффинной концевой последовательности, например, GST (см. статью Smith D.B. и Johnson K.S. в журнале Gene 67:31 (1988)), полигистидина (см. статью Hochuli E. J. в журнале Chromatog. 41:11 (1987)) или биотина. Такие аффинные концы могут быть использованы для обратимого крепления пептида к подложке. Подобные иммобилизованные полипептиды или фрагменты могут быть использованы, например, при выделении антител, направленных против tag7, как будет описано ниже.
Как будет также понятно специалисту в данной области, вышеописанные полипептиды tag7 согласно настоящему изобретению и эпитопнесущие фрагменты можно комбинировать с частями константного домена иммуноглобулина (Ig), получая в результате химерные полипептиды или полипептиды слияния. Эти полипептиды слияния облегчают очистку и показывают увеличенное время полужизни in vivo (см. патент ЕР 0394827; статью Traunecker и др. в журнале Nature 331:84-86 (1988)).
Антитела анти-tag7
Эпитопнесущие пептиды и tag7 полипептиды согласно изобретению могут быть использованы для продуцирования антител, направленных против tag7, в соответствии с хорошо известными в данной области методами (смотри, например, статьи Sutcliffe J.G. и др. в журнале Science 219:660-666 (1983); Wilson и др. в журнале Cell 37:767 (1984) и Bittle F.J. и др. в журнале J.Gen. Virol 66:2347-2354 (1985). Tag7-специфичные антитела могут быть вызваны против целого полипептида tag7 или по меньшей мере одного из его антигенных полипептидных фрагментов, таких как эпитопнесущие фрагменты вышеописанного tag7. Эти полипептиды или фрагменты могут быть представлены вместе с белком-носителем (например, альбумином) в организме животного (такого как кролик или мышь) или, если они достаточно длинны (по меньшей мере около 25 аминокислот), без носителя.
Термин "антитело" (Аb), используемый здесь, может быть заменен термином "поликлональное антитело" или "моноклональное антитело", за исключением особых контекстов, как будет описано ниже. Эти термины, как они здесь использованы, подразумевают как целые молекулы, так и фрагменты антител (таких, например, как Fab и Р(аb')2-фрагменты), которые способны специфично связывать полипептид tag7 или его участок. Fab и Р(аb')2-фрагменты в отсутствие Fc-фрагмента целого антитела быстрее выводятся из кровообращения и могут обладать меньшим тканенеспецифичным связыванием, чем целое антитело (см. статью Wahl и др. в журнале J.Nucl.Med. 24:316-325 (1983)).
Антитела согласно настоящему изобретению могут быть поликлональными или моноклональными и могут быть получены любым из различных методов. Например, поликлональные антитела могут быть получены путем иммунизации животного одним или более полипептидов tag7 или его участков (включая по меньшей мере один эпитопнесущий фрагмент tag7) согласно настоящему изобретению по стандартным технологиям (см., например, Harlow Е. и Lane D. Antibodies: A Laboratory Manual, Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press (1988); Kaufman P.B. и др. в: Handbook of Molecular and Cellular Methods in Biology and Medicine, Roca Raton, Florida: CRC Press, стр.468-469 (1995). В наиболее предпочтительном методе антитела согласно настоящему изобретению являются моноклональными антителами (или их фрагментами, связывающими белок tag 7). Такие моноклональные антитела могут быть получены с применением гибридомной технологии, которая хорошо известна в данной области (см. статьи  и др. в журнале Nature 256:495 (1975),
и др. в журнале Nature 256:495 (1975),  и др. в журнале Eur.J.Immunol. 6:511 (1976) и
и др. в журнале Eur.J.Immunol. 6:511 (1976) и  и др. в журнале Eur.J.Immunol. 6:292 (1976), а также Hammerling и др. в Monoclonal Antibodies and T-Cell Hybridomas, New York:Elsevier, стр.563-682 (1981); Kaufman P.B. и др. в: Handbook of Molecular and Cellular Methods in Biology and Medicine, Roca Raton, Florida: CRC Press, стр.444-467 (1995).
и др. в журнале Eur.J.Immunol. 6:292 (1976), а также Hammerling и др. в Monoclonal Antibodies and T-Cell Hybridomas, New York:Elsevier, стр.563-682 (1981); Kaufman P.B. и др. в: Handbook of Molecular and Cellular Methods in Biology and Medicine, Roca Raton, Florida: CRC Press, стр.444-467 (1995).
Напротив, антитела, способные связывать полипептиды tag7 или их фрагменты, можно получить в двухстадийном процессе посредством использования антиидиотипических антител. Такой метод основан на том факте, что антитела сами по себе являются антигенами и поэтому можно получить антитело, которое связывает другое антитело. В соответствии с этим методом tag7-специфичные антитела используются для иммунизации животного, предпочтительно мыши. Спленоциты такого животного затем используются для получения гибридомных клеток, а гибридомные клетки подвергают скринингу для распознавания клонов, которые продуцируют антитело, чья способность связывать полипептид tag7 может быть блокирована антигеном полипептида tag7. Такие антитела содержат антиидиотипические антитела к антителу, специфичному по отношению к полипептиду tag7 и могут применяться для иммунизации животного с целью формирования других антител, специфичных по отношению к полипептиду tag7.
В другом предпочтительном варианте осуществления изобретения, антитела могут быть получены как химерные антитела. В соответствии с изобретением такие химерные антитела могут содержать антигенсвязывающий домен (то есть область антитела, связывающую tag7), из первого вида и по меньшей мере одну константную область из второго вида, см. патент US №4816567, который относится к методам получения химерных антител и содержание которого привлекается здесь во всей полноте в качестве ссылки.
Понятно, что Fab, F(ab')2-фрагменты и другие фрагменты антител согласно настоящему изобретению также могут быть использованы в описанных методах. Такие фрагменты обычно получают путем протеолитического расщепления с помощью ферментов, таких как папаин (чтобы получить Fab-фрагменты) или пепсин (чтобы получить F(ab')2-фрагменты. Альтернативно, фрагменты, связывающие белок tag 7, могут быть получены с применением методов получения рекомбинантных ДНК или методами химического синтеза.
Антитела, специфичные по отношению к белку tag7, согласно настоящему изобретению могут быть мечены для их обнаружения, наиболее предпочтительно ферментной, радиоизотопной, нерадиоактивной изотопной, флюоресцентной, хемилюминесцентной меткой, токсином или контрастным агентом, применяемым при анализе с помощью ядерно-магнитного резонанса (NMR). Подходящие типы меток хорошо известны специалисту в данной области. Стандартные способы связывания метки с антителом приведены, например, у Kennedy и др. в Clin.Acta 70:1-31 (1976) и у Schurs и др. в Clin.Acta, 81:1-40 (1977), причем все они привлечены здесь во всей своей полноте в качестве ссылки.
В дополнительном предпочтительном воплощении изобретения антитела, полученные вышеописанными способами, могут быть ковалентно или нековалентно иммобилизованы на твердой подложке. Под "твердой подложкой" подразумевается любая твердая подложка, на которой может быть иммобилизовано антитело, включая, но не ограничиваясь этим, нитроцеллюлозу, диазоцеллюлозу, стекло, полистирол, поливинилхлорид, полипропилен, полиэтилен, декстран, сефарозу, агар, крахмал, найлон, шарики, включая стеклянные, из латекса, магнитные (в том числе парамагнитные и суперпарамагнитные), и планшеты для микротитрования. Связь антител по изобретению с твердой подложкой может быть достигнута путем прикрепления одного или обоих концов антитела к опоре. Прикрепление также может быть сделано и на одном или нескольких внутренних участках антитела. В соответствии с настоящим изобретением можно также применить множественное крепление (как на средних участках, так и на концах пептида). Прикрепление может быть через связующую группу аминокислоты, такую как первичная аминогруппа, карбоксигруппа или сульфгидрильная группа (SH), или с помощью химических связывающих групп, например, с помощью бромциана (CNBr), а также через спайсер. Для нековалентных креплений можно применить добавление к антителу аффинной концевой последовательности, например, GST (см. статью Smith D.B. и Johnson K.S. в журнале Gene 67:31 (1988)), полигистидина (см. статью Hochuli Е. J. в журнале Chromatog. 41:11 (1987)) или биотина. Альтернативно, агент для непрямой связи, такой как Протеин А или Протеин G (коммерчески доступный, например от Sigma Chemical Co., St.Louis, Missouri), который связывается с областью Fc антител, может прикрепляться к твердой подложке, и антитела согласно настоящему изобретению присоединяются посредством простого инкубирования антител с твердой подложкой, содержащей иммобилизованные Протеин А или Протеин G. Такие аффинные концы можно использовать также для обратимого крепления антител по настоящему изобретению к опоре.
Возможности применения
Выделенные tag7 полинуклеотиды, полипептиды и антитела согласно настоящему изобретению могут использоваться в различных методах, например, в промышленных, клинических или исследовательских технологиях. Среди этих возможностей имеет место и применение данных антител анти-tag7 для определения tag7 экспрессии или продукции выделенными клетками, или тканями, или клетками и тканями в организме животного по стандартным иммунологическим методикам, которые хорошо знакомы обычному специалисту. Кроме того, полинуклеотиды и полипептиды согласно настоящему изобретению можно применять в терапевтических методах ингибирования развития или прогрессирования опухоли у животного. Аналогичным образом данные полинуклеотиды и полипептиды могут использоваться для лечения рака у животного (предпочтительно у млекопитающего, например человека), страдающего от рака или опухоли.
Ингибирование развития раковых и опухолевых клеток
Данные полинуклеотиды и полипептиды tag7 можно использовать в способах, предусматриваемых изобретением и направленных на ингибирование развития опухолевых или раковых клеток. В одном из предпочтительных аспектов полинуклеотиды и полипептиды настоящего изобретения могут использоваться для вызывания апоптоза в опухолевых или раковых клетках. Обычному специалисту будет, однако, понятно, что ингибирование развития опухолей или клеток может обуславливаться цитостатическими механизмами (то есть индуцированием задержки развития или задержки клеточного цикла, включая индуцирование дифференциации, без уничтожения опухолевых или раковых клеток) или цитоцидальными механизмами (т.е. индуцированием гибели, как прямым, так и непрямым, в том числе посредством индуцирования апоптоза, цитотоксичности, цитолиза и т.д.).
Способы согласно этому аспекту изобретения могут состоять из одной или нескольких стадий, которые подобраны так, чтобы ингибировать развитие опухолевых или раковых клеток in vitro, in vivo или ex vivo (т.е. в кусочке ткани, удаленном из тела животного, который может быть помещен или нет обратно в животное для дальнейшего лечения животного для ингибирования развития опухолевых клеток). В одном из таких предпочтительных вариантов осуществления данного изобретения для ингибирования развития опухоли в опухоль или, что более предпочтительно, в клетку млекопитающего, можно ввести по меньшей мере одну молекулу нуклеиновой кислоты, содержащую полинуклеотид, кодирующий tag7. Предпочтительные для использования в такой технологии молекулы нуклеиновой кислоты включают молекулу нуклеиновой кислоты, содержащую полинуклеотид, имеющий последовательность нуклеиновых кислот, которая по меньшей мере на 65% идентична, а предпочтительно по меньшей мере на 70%, на 80%, на 85%, на 90%, на 95%, или на 99% идентична используемой в качестве ссылки последовательности, такой как (а) нуклеотидная последовательность, представленная в SEQ ID NO:l; (b) нуклеотидная последовательность, кодирующая полипептид tag7, имеющий полноразмерную аминокислотную последовательность, представленную в SEQ ID NO:2; (с) нуклеотидная последовательность, кодирующая зрелый полипептид tag7, имеющий аминокислотную последовательность в положениях от 20 до 182 в SEQ ID NO:2; (d) нуклеотидная последовательность полинуклеотида, который гибридизуется при жестких условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:1; или (е) нуклеотидная последовательность полинуклеотида, который гибридизуется при определенных условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:1. В особенности предпочтительньми являются такие молекулы нуклеиновых кислот, введение которых в клетку млекопитающих ингибирует развитие опухоли. В особенно предпочтительном варианте осуществления настоящего изобретения такой подход предусматривает использование молекулы нуклеиновой кислоты, содержащей полинуклеотид, имеющий последовательность нуклеиновых кислот, представленную в SEQ ID NO:3; или (b) нуклеотидную последовательность, кодирующую полипептид tag 7, имеющий аминокислотную последовательность, представленную в SEQ ID NO:4.
При таком подходе по меньшей мере одну из вышеописанных молекул нуклеиновой кислоты можно ввести в вектор или вирион, пригодный для введения молекулы нуклеиновой кислоты в клетки млекопитающих, нуждающихся в лечении, для того чтобы сформировать вектор трансфекпии. Подходящие для этой цели векторы или вирионы включают те, которые происходят из ретровирусов, аденовирусов и аденоассоциированных вирусов. Технологии формирования вектора трансфекции, содержащего молекулу нуклеиновой кислоты, кодирующую tag7, широко известны в данной области и в основном описаны в "Working Toward Human Gene Therapy", Chapter 28 в Recombinant DNA, изд.2-е, Watson J.D. и др. изд. New York: Scientific American Books, стр.567-581 (1992), и приведены здесь в качестве ссылки. Другие пригодные технологии введения молекул нуклеиновой кислоты, кодирующих tag7, представляют собой хорошо известные в данной области технологии передачи с применением невирусного гена, такие как методы, основанные на эндоцитозе, опосредованном рецептором, как описано в W093/07293, липосомах, катионных липидах и т.д. При альтернативном подходе выделенные молекулы нуклеиновой кислоты можно вводить в клетку другими методами, хорошо известными опытному специалисту, включая, например, электропорацию, трансдукцию, трансформацию, обработку фосфатом кальция, "генную пушку", гипотоническую порацию и повторное запечатывание и т.п. При выборе вышеописанных подходов уровень экспрессии белка tag7 в клетках, испытывающих воздействие, увеличивается, благодаря чему ингибируется развитие опухоли или индуцируется апоптоз в опухолевых клетках.
В другом предпочтительном способе согласно настоящему изобретению развитие опухоли у млекопитающих может быть ингибировано при воздействии на нее композицией, содержащей по меньшей мере один вышеописанный выделенный полипептид tag7 по изобретению. Выделенные полипептиды tag7, подходящие для использования в этом аспекте изобретения, включают те, которые имеют аминокислотную последовательность, по меньшей мере на 65% идентичную, а предпочтительно по меньшей мере на 70%, на 80%, на 85%, на 90%, на 95% или на 99% идентичную используемой в виде ссылки последовательности, такой как (а) аминокислотная последовательность, кодируемая выделенной молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность, представленную в SEQ ID NO:1; (b) полноразмерная аминокислотная последовательность полипептида tag7 представленная в SEQ ID NO:2; (с) аминокислотная последовательность зрелого полипептида tag7, имеющего аминокислотную последовательность, расположенную от 20 до 182 в SEQ ID NO:2; (d) аминокислотная последовательность полипептида, кодируемая полинуклеотидом, который гибридизуется при жестких условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:1; или (е) аминокислотная последовательность полипептида, кодируемая полинуклеотидом, который гибридизуется при определенных условиях гибридизации с полинуклеотидом, имеющим нуклеотидную последовательность, представленную в SEQ ID NO:1. С другой стороны, в этих методах согласно настоящему изобретению можно с успехом использовать фрагмент по меньшей мере одного из полипептидов, описанных выше в (a), (b),(c), (d) или (е). В особенности предпочтительными для применения в данных методах являются такие полипептиды tag7 или их фрагменты, которые обеспечивают ингибирование развития опухоли при контакте клетки млекопитающего с полипептидом или фрагментом.
В предпочтительном варианте осуществления выделенные полипептиды tag7, предназначенные для использования в этом аспекте изобретения, включают те, которые имеют аминокислотную последовательность, кодируемую выделенной молекулой нуклеиновой кислоты, имеющей нуклеотидную последовательность, представленную в SEQ ID NO:3 или (b) аминокислотную последовательность полипептида tag7, представленную в SEQ ID NO:4, или их фрагменты.
Композиции, применяемые в этом аспекте изобретения, могут необязательно содержать также одно или несколько дополнительных соединений, таких как фармацевтически приемлемые носители или среды, подходящие для использования с выделенным полипептидом tag7, входящим в композицию. Под "фармацевтически приемлемым носителем или средой" подразумевается нетоксичный, твердый, полужидкий или жидкий наполнитель, разбавитель, инкапсулирующий материал или вспомогательные вещества любого типа. Носители могут также содержать малые количества добавок, таких как вещества, усиливающие изотоничность и химическую стабильность, которые хорошо известны специалистам. Таким образом, изобретение относится к фармакологическим композициям, содержащим по меньшей мере один выделенный полипептид tag7 по изобретению и фармацевтически приемлемые носитель или среду для него.
При выборе вышеописанных подходов полипептид tag7 действует напрямую (например, через клеточную поверхность или внутриклеточный рецептор) или косвенно (например, посредством трансмембранной передачи или путем индуцирования действия по меньшей мере одного вторичного мессенджера) для ингибирования роста клетки млекопитающего или индуцирования в ней апоптоза.
Клетки млекопитающих, которых можно лечить данными способами для того, чтобы ингибировать развитие опухоли или индуцировать апоптоз опухолевой клетки, включают те, которые получены из любых млекопитающих, в частности, но не ограничиваясь этим, из мышей, крыс, обезьян, овец, коров, лошадей, собак, кошек, морских свинок, кроликов и наиболее предпочтительно людей. Хотя можно ингибировать рост любой клетки млекопитающего с помощью вышеописанных способов, эти способы особенно эффективны для ингибирования развития опухолевых или других раковых клеток или индуцирования апоптоза в них. Типы опухолевых или раковых клеток, развитие которых можно с успехом ингибировать с помощью способов согласно изобретению, включают клетки карциномы (в частности, клетки карциномы печени, клетки карциномы яичника, клетки карциномы груди, клетки карциномы затылочной части шеи, клетки карциномы легкого, клетки карциномы простаты, клетки карциномы желудка, клетки карциномы мочевого пузыря, клетки карциномы яичка, клетки карциномы прямой кишки, клетки карциномы поджелудочной железы, клетки карциномы полости рта, клетки карциномы чешуйчатых клеток, клетки карциномы головы и шеи и клетк тератокарциномы), клетки саркомы (в частности, клетки саркомы Kaposi, клетки фибросаркомы и клетки остеосаркомы), клетки меланомы и клетки лейкемии.
Возможности терапевтического применения
Выделенные молекулы нуклеиновой кислоты и полипептиды согласно изобретению могут быть также использованы в терапевтических целях, например, при лечении рака у страдающего им животного. В этих случаях задачей терапевтического воздействия является задержка или ингибирование развития или роста опухоли или рака для того, чтобы задержать или ингибировать метастазирование опухоли или рака и/или вызвать их ремиссию.
Генетическая терапия. В соответствии с первым из таких аспектов изобретения больное раком животное можно лечить введением ему одной или более выделенных молекул нуклеиновой кислоты согласно изобретению, содержащей полинуклеотид, кодирующий полипептид tag7, или его фрагмент, преимущественно полинуклеотид, который на 90% или 95% идентичен по меньшей мере одной используемой в виде ссылки нуклеотидной последовательности, описанной выше. Этот подход, известный в общем как "генетическая терапия", направлен на увеличение уровня экспрессии гена tag7 в клетках, из которых состоит рак или опухоль, благодаря чему ингибируется или задерживается рост, развитие или метастазирование рака или опухоли или индуцируется ремиссия. Доказано, что аналогичные методы генетической терапии эффективны или обещают быть таковыми при лечении других заболеваний у млекопитающих, например кистозного фиброза (см. статьи Drumm M.L. и др. в журнале Cell 62:1227-1233 (1990); Gregory R.J. и др. в журнале Nature 347:358-363 (1990); Rich D.R. и др. в журнале Nature 347:358-363 (1990)); болезни Gaucher'a (см. статьи Sorge J. и др. в журнале Рrос.Natl.Acad.Sci. USA 84:906-909 (1987); Fink J.K. и др. в журнале Рrос.Natl.Acad.Sci. USA 87:2334-2338 (1990)), некоторых форм гемофилии (см. статью Bontempo F.A. и др. в журнале Blood 69:1721-1724 (1987); Palmer T.D. и др. в журнале Blood 73:438-445 (1989); Axelrod J.H. и др. в журнале Ргос.Natl.Acad.Sci. USA 87:5173-5177 (1990); Armentano D. и др. в журнале Рrос.Natl.Acad.Sci. USA 87:6141-6145 (1990)) и мышечной дистрофии (см. статьи Partridge T.A. и др. в журнале Nature 337:176-179(1989); Law Р.К. и др. в журнале Lancet 336:114-115 (1990); Morgan J.E. и др. в журнале J. Cell.Biol.111:2437-2449 (1990)), а также в других способах лечения некоторых видов рака, например метастазирующей меланомы (см. статьи Rosenberg S.A. и др. в журнале Science 233:1318-1321 (1986); Rosenberg S.A. и др. в журнале N.Eng.J.Med., 319:1676-1680 (1980); Rosenberg S.A. и др. в журнале N.Eng.J.Med., 323:m-m (1990)).
В предпочтительном из этих способов животному, больному раком, вводят одну или более выделенных молекул нуклеиновой кислоты согласно изобретению, содержащую полинуклеотид, имеющий нуклеотидную последовательность, которая по меньшей мере на 90% или по меньшей мере на 95% идентична одной из используемых в качестве ссылок нуклеотидных последовательностей, описанных выше. Такие выделенные молекулы нуклеиновой кислоты могут быть включены в вектор или вирион, подходящие для введения молекул нуклеиновой кислоты в клетки или ткани подлежащего лечению животного, чтобы сформировать вектор трансфекции. Приемлемые для этой цели векторы и вирионы включают те, которые получены из ретровирусов, аденовирусов и аденоассоциированных вирусов. С другой стороны, молекулы нуклеиновой кислоты по изобретению можно использовать в комплексе с молекулярным коньюгатом вместе с вирусом (например, аденовирусом и аденоассоциированным вирусом) или с вирусными компонентами (например, вирусными капсидными белками).
Технологии формирования векторов и вирионов, содержащих молекулы нуклеиновой кислоты, кодирующие tag7, хорошо известны в данной области и в основном описаны в "Working Toward Human Gene Therapy", Chapter 28 в Recombinant DNA, изд.2-е, Watson J.D. и др., изд. New York: Scientific American Books, стр.567-581 (1992). Кроме того, основные способы конструирования векторов для генетической терапии и введения их в пораженное болезнью животное для терапевтических целей могут быть почерпнуты из вышеуказанных публикаций, содержание которых приводится здесь во всей полноте в качестве ссылки. В одном из таких основных способов векторы, содержащие выделенные полинуклеотиды согласно настоящему изобретению, вводят непосредственно в клетки и ткани пораженного животного, преимущественно путем инъекции, ингаляции, проглатывания или введения в слизистую оболочку через раствор; такие способы в общем называются генетической терапией "in vivo". С другой стороны, клетки, ткани и органы, содержащие раковые или опухолевые клетки, можно удалить из организма больного животного и поместить в культуру способами, хорошо известным обычному специалисту в данной области; векторы, содержащие полинуклеотиды tag7, можно затем ввести в клетки или ткани любым из вышеописанных способ с целью подачи выделенных полинуклеотидов tag7 в клетки или ткани и, после того как пройдет достаточное время для внедрения полинуклеотидов tag7, клетки или ткани можно потом вновь ввести больному животному. Поскольку подача гена tag7 происходит вне тела пораженного болезнью животного, этот способ в общем называется генетическая терапия "ех vivo". Один вид генетической терапии "ех vivo" можно также обозначить как "иммунотерапия" или, более точно, "противораковая вакцина". Она вызывает иммунную реакцию у подвергаемого лечению животного и вследствие этого уничтожение опухолевых клеток. В опытах, выполненных при проверке настоящего изобретения, было показано, что вакцинация животных опухолевыми клетками, экспрессирующими tag7, защищает животных от последующего заражения немодифицированными опухолевыми клетками. В предпочтительном варианте осуществления настоящее изобретение касается противоопухолевой вакцины на основе опухолевых клеток, экспрессирующих tag7. Такие клетки могут быть, как описано выше, аутологичными, т.е. происходить из подлежащего лечению организма, или они могут быть аллогенными, т.е. из одного или нескольких организмов, например, клетки от различных устоявшихся клеточных линий.
При проведении генетической терапии как "w vivo", так и "ex vivo" выделенные полинуклеотиды tag7 согласно изобретению могут по выбору быть оперативно связаны с регуляторной последовательностью ДНК, которая может представлять собой промотор или энхансер tag7, или с гетерологической регуляторной последовательностью ДНК, такой как промотор или энхансер, полученные из другого гена, клетки или организма, для формирования генетической конструкции, как описано выше. Эта генетическая конструкция может быть затем встроена в вектор, который затем напрямую вводят больному животному способом генетической терапии in vivo, например, внутриопухо-левым введением (то есть введением молекулы нуклеиновой кислоты или вектора непосредственно в опухоль животного, например, инъекцией), или в клетки или ткани пораженного животного ех vivo. В другом предпочтительном варианте осуществления генетическую конструкцию согласно изобретению можно ввести в клетки или ткани животного, или in vivo, или ех vivo, в виде молекулярного коньюгата с вирусом (например, аденовирусом и аденоассоциированным вирусом) или с вирусными компонентами (например, вирусными капсидными белками; смотри WO 93/07283). С другой стороны, трансфецированную клетку-хозяин, которая может быть гомологичной или гетерологичной, можно инкапсулировать в полупроницаемый носитель и имплантировать в больное животное, обеспечивая проникновение полипептидов tag7 в ткани и циркулирование в нем, но предотвращая контакт между иммунной системой животного и трансфецированными клетками (смотри WO 93/09222). Такие способы приводят к увеличенной продукции tag7 у подвергнутого лечению животного посредством (а) случайного встраивания гена tag7 в геном клетки-хозяина; (b) введения гена tag7 в ядра клеток, где он может существовать как экстрахромосомный генетический элемент. Общее описание таких методов и подходов можно найти, например, в патенте US №5578461 и WO 93/09222.
Независимо от применяемых подходов, однако, использование этих методов по изобретению приводит к увеличению продуцирования tag7 клетками и тканями больного животного, так что развитие, рост или метастазирование рака и опухолей замедляется или ингибируется или рак и опухоли переходят в стадию ремиссии или излечиваются.
Белковая терапия. При другом предпочтительном терапевтическом подходе, обеспечиваемом с помощью настоящего изобретения, животное, страдающее раком, можно лечить введением ему композиции, содержащей по меньшей мере один выделенный полипептид tag7 по изобретению, причем такое введение ингибирует развитие, рост или метастазирование опухоли или рака или индуцирует их ремиссию. По одному из таких способов выделенные полипептиды tag7 вводят животному в форме вышеописанных фармацевтических композиций. По другому такому способу вводимые животному композиции могут содержать один или несколько выделенных полипептидов tag7, имеющих аминокислотную последовательность, по меньшей мере на 65% или по меньшей мере на 70%, на 75%, на 80%, на 85%, на 90%, на 95% или на 99% идентичную по меньшей мере одной вышеописанной используемой в качестве ссылки последовательности. Следовательно, данное изобретение относится к способам лечения страдающих раком животных, предусматривающим введение таким животным фармацевтической композиции, содержащей терапевтически эффективное количество одного или нескольких выделенных полипептидов tag7 согласно изобретению и необязательно фармацевтически приемлемый носитель или среду для него.
Композиции, содержащие полипептиды tag7, должны быть приготовлены и дозированы так, как это принято в медицинской практике, принимая во внимание клиническое состояние индивидуального пациента (в особенности побочные эффекты, возникающие при лечении с применением только полипептида tag7), место введения композиции, содержащей полипептид tag7, метод введения, график введения и другие факторы, известные специалистам. "Терапевтически эффективное количество" полипептидов tag7 для намеченных здесь целей будет определено следующими соображениями.
В качестве основного предложения общее терапевтически эффективное количество полипептидов tag 7, вводимое парентерально с одной дозой, должно находиться в пределах примерно от 0,01 мкг/кг/день до 1000 мкг/кг/день по отношению к весу тела пациента, хотя, как отмечено выше, это должно быть благоразумно с терапевтической точки зрения. Более предпочтительно, чтобы эта доза составляла по меньшей мере 0,1 мкг/кг/день и наиболее предпочтительно для человека примерно между 0,1 и 100 мкг/кг/день, считая на полипептид. При непрерывном приеме полипептид tag7 можно вводить либо путем 1-10 инъекций в день или путем непрерывной подкожной инфузии, например с помощью мининасоса. Можно также использовать раствор во внутривенной капсуле. Ключевым фактором при выборе подходящей дозировки является получаемый результат, измеряемый, например, через увеличение уровня циркулирующего tag7 или путем определения уменьшения роста опухоли или ее метастазирования, а также ремиссии рака. Обычному специалисту в данной области ясны другие стандартные измерительные методы определения терапевтической эффективности. Продолжительность лечения, в течение которого нужно наблюдать за изменениями, и интервал после лечения, чтобы проявилась реакция на него, могут варьироваться в зависимости от желаемого эффекта.
Фармацевтические композиции, применяемые в таких методах, содержат один или несколько выделенных полипептидов tag7 согласно изобретению и необязательно могут содержать фармацевтически приемлемый носитель или среду для них. Полипептиды tag7 и фармацевтическая композиция согласно изобретению могут быть введены с помощью любых средств, которые обеспечивают достижение намеченных целей - задержки или ингибирования развития и роста опухолей, или рака, или их метастазирования у больного животного. Можно использовать, например, внутриопухолевое введение, оральное введение, введение в глаз, в ухо, ректальное, парентеральное, подкожное, внутривенное, внутримышечное, перитональное, вагинальное, топическое (с помощью порошков, мазей, капель или трансдермальных пластырей), буккальный, внутриоболочечный и внутрикраниальный пути, а также применять средства, распыляемые в полость рта или в нос или глазные и ушные капли. Термин "парентеральное", используемый здесь, указывает на способы введения, включающие внутривенные, внутримышечные, перитональные, внутригрудинные, подкожные и внутрисуставные инъекции и инфузии. Вводимые дозы зависят от возраста, состояния здоровья и веса реципиента, вида сопутствующего лечения, если оно имеет место, повторяемости лечения или природы ожидаемого эффекта. Композиции в рамках настоящего изобретения включают все те, где содержатся tag7 полинуклеотиды или полипептиды в количестве, которое является эффективным для достижения ингибирования или задержки развития и роста опухолей или рака, или их метастазирования, или для индуцирования ремиссии опухоли или рака, или излечения от них. Хотя индивидуальные потребности каждого животного могут быть разными, выбор оптимальных диапазонов эффективного количества каждого компонента находится в пределах компетенции обычного клинициста.
Композиции, содержащие полипептид tag7, можно также вводить в системах замедленного высвобождения. Примерами таких композиций с замедленным высвобождением являются полупроницаемые полимерные матрицы в виде формованных изделий, в частности пленок или микрокапсул. Матрицы с замедленным высвобождением включают полилактиды (патент US №3779919; ЕР 0058481), сополимеры L-глутаминовой кислоты и гамма-этил-b-глутамата (см. статью Sidman U. и др. в журнале Byopolimers 22:547-556 (1983)), поли(2-гидроксиэтилметакрилат) (см. статью Langer R. и др. в журнале J.Biomed. Mat. Res. 15:167-277 (1981); Langer R. в журнале Chem.Tech. 12:98-105 (1982)), этиленвинилацетат (см. Langer R. и др. в Id) и поли-В(-)-3-гидроксибутировую кислоту (ЕР 0133988). Композиции с замедленным высвобождением полипептида tag7 могут также включать полипептиды tag7, заключенные в липосомы или абсорбированные ими, и их можно изготовить любым из известных в данной области методов (см. патенты US №4485045 и US 4544545; статьи Epstein и др. в журнале Proc.Natl.Acad.Sci. (USA) 82:3.688-3692 (1985); Hwang и др. в журнале Proc.Natl.Acad.Sci. (USA) 77:4030-4034 (1980); патенты ЕР 0036676; ЕР 0052322; ЕР 0088046; ЕР 0102324; ЕР 0142641; ЕР 0143949; DE 3218121; и JP 83-118008).
Для парентерального введения, в одном из воплощений, полипептиды tag7 готовят в основном путем смешивания их при желаемой степени чистоты в инъецируемую форму, содержащую одну дозу (раствор, суспензию или эмульсию), с фармацевтически приемлемым носителем, т.е. тем, который не токсичен для реципиентов в используемых дозах и концентрациях и совместим с другими ингредиентами формулы. Например, желательно, чтобы формула не включала окислителей и других соединений, которые известны как повреждающие полипептиды.
В основном, фармацевтические формы получают путем равномерного распределения полипептидов tag7 согласно настоящему изобретению в жидких носителях, или тонко измельченных твердых носителях, или в тех и других. Затем, если необходимо, продукт формуют в желаемый препарат. Предпочтительно носитель представляет собой носитель для парентерального введения, более предпочтительно раствор, который является изотоническим по отношению к крови реципиента. Примеры таких носителей включают воду, солевой раствор, раствор Ringer'a и раствор декстрозы. Приемлемы также неводные носители, такие как фиксирующие масла и этилолеат, а также липосомы.
Полипептид tag7 обычно содержится в таком носителе в концентрации примерно от 0,01 мг/мл до 100 мг/мл, преимущественно примерно от 0,1 до 10 мг/мл, при рН около 3-8. Понятно, что применение некоторых из упомянутых ранее сред, носителей или стабилизаторов может привести к образованию солей полипептида tag 7.
Полипептиды tag 7, используемые для терапевтического введения, должны быть стерильны. Стерильность легко обеспечивается путем фильтрации через стерильную фильтрующую мембрану (например, 0,2-микронные мембраны) или облучения гамма-лучами или ультрафиолетом по известным специалистам технологиям. Терапевтические композиции, содержащие полипептид tag7, в основном помещают в контейнер, имеющий стерильное входное отверстие, например капсулу или пузырек для внутривенного раствора, с пробкой, которую можно проткнуть иглой для гиподермической инъекции.
Терапевтические композиции, содержащие полипептид tag7 согласно настоящему изобретению, обычно можно хранить в контейнере на одну или множество доз, например, в герметично закрытых ампулах или пузырьках, как в виде водного раствора, так и в виде лиофилизированного восстанавливаемого препарата. Как пример приготовления лиофилизированного препарата, пузырьки емкостью в 10 мл наполняют 5 мл отфильтрованным в стерильных условиях 1%-ным водным раствором полипептида tag7, и полученную смесь лиофилизируют. Раствор для инфузии готовят восстановлением лиофилизированного полипептида tag7, используя воду U.S.P., которая годится для инъекции.
Различные виды рака у животных можно лечить терапевтическими способами согласно изобретению. Виды рака, которые поддаются лечению этими способами, включают, не ограничиваясь этим, карциномы, саркомы, меланомы и лейкемии, в частности те, которые упомянуты выше. Способы согласно настоящему изобретению, в частности, хорошо проявляют себя при лечении любых животных, предпочтительно млекопитающих и наиболее предпочтительно человека.
Совершенно очевидно для обычного специалиста в данной области, что любые подходящие модификации и переделки в методах и способах применения, описанных здесь, ясны и могут быть выполнены без выхода за рамки изобретения или любых его воплощений.
Имея теперь детальное описание изобретения, все это может быть более понятно из ссылки на последующие примеры, которые включаются только с целью иллюстрации и не предполагают ограничения изобретения.
Основное обсуждение
Для того чтобы исследовать гены, которые селективно экспрессируются в опухолях с высоким потенциалом метастазирования, была выбрана пара опухолей одинакового происхождения, различающихся по своим метастазирующим свойствам. Два субштамма рака VMR с высокой метастатической активностью (который имеет широкий спектр поражаемых метастазами органов и частоту метастазирования в печени не выше 5%) были выделены в соответствии с предыдущими исследованиями (см. у Senin V.M. в Vestn.Acad.Med.Nauk. SSSR 0(5): 55-91 (1984)): клеточная линия VMR-0 (частота метастазирования в печени около 0%) и линия VMR-L (частота метастазирования в печени около 100%). В отличие от опухолевых линий CSML-0 и CSML-100 ни одна из этих VMR-опухолей не экспрессирует предварительно клонированный ген mts 1, который транскрибируется в большинстве опухолей (см. статью Grigorian M.S. и др. в журнале Genetika (USSR) 25(6): 993-1000 (1989)), вследствие чего целесообразно искать другие гены, которые участвуют в процессе метастазирования. Если рассматривать опухоли, полученные в соответствии с ранее предложенными моделями (см. статью Filder I.J. и Hart I.R. в журнале Science 277:998-1001 (1982)), очевидно, что опухоль VMR-L может происходить из метастазной субпопуляции клеток, которые представлены в исходной опухоли VMR на очень низком уровне и которые имеют очень короткий срок жизни (см. статью Kerbell K.S. и др. в журнале Adv. Cancer Res. 55:87-131 (1990)).
Это предполагаемое общее происхождение пары опухолей, исследуемой здесь, предлагает возможность использования методики "выявления различий в РНК" для идентификации генов, связанных с метастазированием. В общем, генетическая разница между опухолями VMR-0 и VMR-L незаметна, до определенной степени подтверждая их общее происхождение. Тем не менее, с помощью такого подхода можно выделить несколько фрагментов ДНК, соответствующих различно экспрессируемым РНК в опухолях VMR-0 и VMR-L. Анализ выделенных фрагментов гибридизацией посредством Нозерн-блоттинга с РНК из этих опухолей дал возможность отнести большинство фрагментов к классу повторяющихся последовательностей (элемент IAR и т.д.). Однако несколько полученных фрагментов не могли быть так определены и поэтому представляют несомненный интерес.
Одним из таких фрагментов, выделенных и охарактеризованных здесь, является ген tag7, который демонстрирует очень высокий уровень транскрипции в опухоли VMR-L, метастазирующей в печень (см. фиг.2). Анализ последовательности показал, что PCR-фрагмент фланкирован на обеих сторонах посредством 5'-концевого праймера 5'-AATCGGGCTG-3' (SEQ ID NO:4). Когда была проанализирована нуклеотидная последовательность клона кДНК из тотальной библиотеки кДНК VMR-L, последовательность, которая оказалась гомологичной этому праймеру длиной восемь нп, была обнаружена на расстоянии в 52 нуклеотида от 3' полиА+хвоста клонированной ДНК. Вероятно, что традиционный олиго-dT праймер мог бы быть использован в методике "выявления различий в РНК" вместо 3'-концевого праймера и только 5'-концевые праймеры могли бы меняться; такой подход, вероятно, уменьшил бы количество получаемых фрагментов, но одновременно увеличил бы разрешающую способность геля.
Ошибки, возникающие в многочисленных циклах PCR, можно классифицировать как недостатки методики. При сравнении нуклеотидных последовательностей фрагмента, полученного в процессе PCR, и кДНК клона tag7 было обнаружено, что 44 нуклеотида на 5'-конце амплифицированного фрагмента не совпадают с аналогичным сегментом последовательности клона кДНК.
Компьютерный анализ полноразмерной последовательности кДНК мышиного tag7 показал две открытых рамки считывания (ORFs), соответствующие полипептидам длиной приблизительно 182 и 91 аминокислот. Оба места начала трансляции в различной степени соответствовали усредненной последовательности Козака (см. статью Kozak M.J. в журнале J.Cell.Biol. 108:229-241 (1989)). Однако в случае более длинной открытой рамки считывания (ORF) число нуклеотидов, одинаковых по совпадению трансляции, больше, чем при более короткой ORF, и это, вероятно, представляет собой первый метионин, в особенности тот, который является сайтом инициации трансляции (см. статью Kozak M.J. в журнале J.Cell.Biol. 108:229-241 (1989)). Но трансляция функционального полипептида tag7 может быть также инициирована от второго метионина.
Молекулярный вес предсказанного полипептида tag7 (182 аминокислотного остатка; SEQ ID NO:l) составляет около 20 кД. Для того чтобы проверить гомологию с ранее описанньми последовательностями, нуклеотидную последовательность tag7 и предсказанную аминокислотную последовательность его продукта анализировали в банках данных GENE и SWISSPROT с помощью программ поиска гомологии FASTA (Гейдельберг, Германия) и BLAST (Вашингтон, DC, США). Не было найдено существенной гомологии с ранее известными последовательностями, за исключением незначительной гомологии гидрофобного сегмента (между 2 и 12 аминокислотными остатками) на N-конце предсказанного полипептида tag7 с β-цепью рецептора Т-клетки человека. Эта гидрофобная область также является гомологичной сигнальным последовательностям некоторых белков (например, предшественников кальретикулина, остеокальцина и внеклеточного глобина III). Наличие гидрофобной области на N-конце предсказанного полипептида tag7 наводит на мысль, что этот полипептид может иметь трансмембранную локализацию или может быть секретирован.
Не обнаружено корреляции экспрессии tag7 с органоспецифичностью метастазирования; ген tag7 транскрибируется не только в клетках опухоли VMR-L, метастазирующей в печени, но также и в клетках опухоли VMR-0v, метастазирующей в яичниках. Среди обычных органов максимальный уровень экспрессии гена tag7 наблюдался в клетках тех органов, у которых относительно высокое содержание клеток базальной мембраны. Эта корреляция может совпадать не полностью, но отражает роль, которую базальная мембрана играет в процессе метастазирования (см. статью Stetler-Stevenson W.G. и др. в журнале Ann. Rev. Cell Biol. 9:541-573 (1993)). Интересно, что уровень экспрессии гена tag7 в метастазирующих опухолях гораздо выше, чем у тех же клеток в культуре. Этот результат является обратным по отношению к недавно сообщенным результатам исследований экспрессии гена в первичных и культивированных раковых клетках (см. статью Zhang L. и др. в журнале Science 276:1268-1271 (1997)), и может представлять собой ответ на взаимодействие опухолевых клеток с клетками-хозяевами базальной мембраны, например, в процессе поражения базальной мембраны метастатическими опухолевыми клетками. Также нельзя исключить участия клеток стромы в регуляции экспрессии tag7 как происходит, например, при экспрессии гена стромелизина-3 (см. статью Basset P. и др. в журнале Nature 348: 699-704 (1990)).
Также представляет интерес наблюдение, показывающее, что уровень транскрипции гена tag7 в клетках опухоли CSML-0 в несколько раз ниже, чем в клетках VMR-L, тогда как, если взять штаммы тех же опухолей в виде культур, картина заметно меняется. В частности, после культивирования этих клеток уровень транскрипции гена tag7 в клетках VMR-L уменьшается, а в клетках CSML-0 увеличивается и превышает в несколько раз уровень экспрессии tag7 в культивированных клетках VMR-L. В культивированных клетках МТ1 ТС1 наблюдался несколько более низкий уровень транскрипции tag7 по сравнению с культивированными клетками CSML-0. Такое изменение в уровне транскрипции гена tag7 в культивированных клетках VMR-L может быть объяснено тем фактом, что эта культура не является моноклональной и поэтому доля субпопуляции метастазирующих клеток в культуре может быть довольно малой (см. статьи Filder I.J. и Hart I.R. в журнале Science 217:998-1001 (1982) и Senin V.M. в Vestn. Acad.Med.Nauk. SSSR 0(5): 85-91 (1984)). С другой стороны, гетерогенность культуры клеток CSML-0 установлена D.A.Kramer'ом (личная информация). Особенный интерес представляет упоминание о том, что транскрипция гена tag7 наблюдалась в культурах клеток тех опухолей, которые имели эпителиальное происхождение и низкий (5-10%) метастатический потенциал. Однако из 13 клеточных линий с различными метастатическими свойствами, которые были изучены, только в клетках линии VMR-L транскрипция гена tag7 коррелировалась с маркированным метастатическим потенциалом этой опухоли.
На протяжении последнего десятилетия было исследовано и клонировано некоторое количество цитокинов, относящихся к TNF. Протеолитически получаемая форма TNF-α является хорошо известным растворимым цитокином. Кроме того, сообщалось, что растворимая форма Fas-лиганда продуцируется активированными периферийными Т-лимфоцитами (см. статью Tanaka и др. в журнале EMBO J. 14:1129-1135 (1995), а клеточная линия мышиной тимомы, как показано, секретирует растворимый СD40-лиганд (смотри статью Armitage G. и др. в журнале Eur. J. Immunol. 22:2071-2076 (1992). Благодаря своему структурному сходству с другими цитокинами, родственными TNF, tag7 может быть признан новым членом этого развивающегося семейства.
Расположение tag7 на хромосоме позволяет понять роль tag7 in vivo. Цитогенетическая полоса 7А3 генетически связана с нефритом, подобным волчанке в MRL и новозеландских гибридных моделях системного эритематоза волчанки (SLE) (см. статьи Morel L. и др. в журнале Immunity 1:219-229 (1994); Kono D.H. и др. в журнале Proc.Natl.Acad.Sci. USA 97:10168-10172 (1994)). Хотя не существует вероятности того, что SLE вовлечен в мутации с сильными функциональными изменениями, опыты позволяют проникнуть в возможную роль tag7 в этом патогенном процессе.
Локализация РНК tag7 в основном в лимфоидных органах представляет интерес, учитывая цитотоксическое действие растворимого белка. Очевидно, что главная ступень в каскаде регуляции происходит на посттранскрипционном уровне. Даже незначительные изменения в продуцировании РНК tag7 приводят к изменениям общего количества синтезируемого белка и его секреции. Поэтому является возможным, что могут быть задействованы различия в способности передавать сигнал между растворимой и связанной с мембраной формами лиганда.
Одним из биологических свойств tag7, установленных путем настоящих исследований, является его вовлеченность в индуцирование in vitro апоптоза в некоторых клеточных линиях. Апоптоз, или процесс запрограмированной гибели клеток, необходим для нормального развития и гомеостаза организма. Фрагментация ДНК на мультимеры примерно в 180 нп одно из тех изменений, которые возникают при апоптозе.
Интересно было обнаружить, что опухоли VMR-0, трансфецированные tag7, растут значительно медленнее, чем родительские опухоли VMR-0. Было показано, что некоторое количество цитокинов способно индуцировать отторжение опухоли, если они продуцируются локально опухолевыми клетками после передачи гена (см. статью Hock Н. и др. в журнале Proc.Natl.Acad.Sci. USA 90:2774-2778 (1993)). Известны различные цитокины, активирующие гетерогенные транзисторные опухолеподавляющие механизмы, но для полного отторжения опухоли всегда требуются клетки CD8+. Недавно был описан сильный противоопухолевый эффект человеческого LT-α в мышиной экспериментальной модели (см. статью Qin Z. и Blankenstein Т. в журнале Cancer Res. 55:4747-4751 (1995)). В отторжении опухоли задействован комплексный иммунологический механизм, в который вовлечены как Т-клетки, так и В-клетки. Показано, что LT-α менее токсичен для животных, чем TNF, и по меньшей мере столь же эффективен, как TNF (см. статью Qin Z. и др. в журнале Blood 55:2779-2785 (1995)). Не наблюдалось никакого токсического действия клеток, трансфецированных tag7, в мыши, а также какой-либо непосредственной цитоксической активности клонов, трансфецированных конструкцией tag 7, не было обнаружено в L929 цитотоксическом анализе. Хотя активность tag7 в кондиционированной среде клонов ниже границы определения, клон, который производил более низкие количества tag7 РНК, развивался быстрее в сингенной мыши. Ингибиторный эффект локально выделенного tag7 по отношению к развитию опухоли, вероятнее всего, непрямой и может быть аннулирован инъекцией антител анти-tag7. Это доказано также опытом, в котором был подвергнут мониторингу рост опухоли в родительской линии VMR-0 и совместно инъецированных клетках 4SX. Даже когда инъецировали двойное по отношению к нормальному количество клеток, скорость развития опухоли значительно снижается. Наконец, полное уменьшение опухоли или ремиссия могут быть достигнуты посредством секреции более высоких количеств tag7.
В ингибирование развития опухолевой клетки, опосредованное tag7, как представляется, не вовлечены Т-лимфоциты, т.к. в отсутствие Т-клеток (например, в голой мыши) клетки, продуцирующие tag7, растут медленнее, чем в сингенной мыши (E.V.Korobko, неопубликованные наблюдения). Однако несмотря на эти обнаруженные данные, участие других типов клеток в антиопухолевой реакции, индуцированной tag7, не может быть исключено.
Поскольку передача гена в опухолевые клетки является полезньм средством для оценки противоопухолевого действия разных цитокинов и противоопухолевых агентов, было исследовано противоопухолевое действие tag7 путем введения его кДНК в tag-7-негативные опухолевые клетки. Было показано, что ряд цитокинов способен вызывать отторжение опухоли, если они продуцируются локально опухолевыми клетками после передачи гена. Различные цитокины активируют механизмы гетерогенного транзисторного подавления опухолей, но для полного отторжения опухоли всегда требуются клетки CD8+. Недавно был описан сильный противоопухолевый эффект человеческого LT-α, в мышиной экспериментальной модели. В отторжении опухоли задействован комплексный иммунологический механизм, в который вовлечены как Т-клетки, так и В-клетки. Показано, что LT-α менее токсичен для животных, чем TNF, и по меньшей мере столь же эффективен, как TNF. Tag7 в был недавно описан как белок, секретируемый лимфоцитами, цитотоксичный по отношению к ограниченному количеству клеток-мишеней. В настоящем изобретении не наблюдалось никакого токсического действия клеток, трансфецированных tag7, в мыши. Более того, изобретатель не смог обнаружить прямой цитоксической активности нескольких клонов, трансфецированных tag7 конструкцией, в цитотоксическом анализе L929 и должен был переключиться на уровень мРНК tag7. Хотя активность tag7 в кондиционированной среде клонов ниже границы определения, клон, который продуцировал более низкие количества РНК tag7, развивался быстрее в сингенной мыши. Ингибиторное воздействие на развитие опухоли локально выделенного tag7 вероятнее всего, непрямое и может быть аннулировано инъекцией антител анти-tag7. Это доказано также опытом, в котором был подвергнут мониторингу рост опухоли в родительской линии VMR-0 и совместно инъецированных клетках 4SX. Даже когда инъецируют двойное по отношению к нормальному количество клеток, скорость развития опухоли значительно снижается. Наконец, полное уменьшение опухоли или ремиссия могут быть достигнуты посредством секреции более высоких количеств tag7. Эффект отторжения после совместной имплантации родительских и модифицированных клеток в один и тот же сайт приводит к заключению, что действие tag7 по меньшей мере локально ограничено и индуцируется некоторой неспецифичной реакцией клетки-хозяина, которая также направлена на отторжение наблюдаемых опухолевых клеток. Однако временная экспрессия tag7 в клетках меланомы М3 создавала защитный иммунитет в отдаленном участке. Эти результаты означают, что антиопухолевое действие tag7 не имеет локального ограничения и индуцирует некоторую специфичную реакцию хозяина, которая отторгает также и отдаленные клетки.
Хотя изобретатель не обнаружил какой-либо разницы в пролиферации родительских VMR-0 и устойчиво трансфецированных клонов т vitro, результаты гистологической и электронной микроскопии показали значительное уменьшение пролиферации модифицированных опухолей, главным образом вследствие апоптоза модифицированных клеток. Для этого не требуется чувствительности опухолевых клеток к tag7 in vitro, т.к. опухолевые клетки устойчивы к секретированному tag7 в культуре. Однако имеющиеся данные не допускают возможности чувствительности опухолевых клеток к секретируемому tag7 при развитии in vivo.
В подавлении опухолей на основе tag7 по-видимому, не участвуют Т-лимфоциты, поскольку в отсутствие Т-клеток (например, у голой мыши) клетки, продуцирующие tag7, растут даже с меньшей скоростью, чем в сингенной мыши; таким образом, наиболее вероятно, что Т-клетки не являются важными для ингибиторного эффекта. Это ясно показывает, что задействованный механизм может эффективно действовать на опухоли, которым недостает антигенов отторжения, специфичных по отношению к опухолям, и поэтому он может использоваться для лечения неантигенных опухолей.
Иммуногистохимический анализ показал, что модифицированные опухолевые клетки распознавались СD45К-позитивными клетками, предположительно В-лимфоцитами, хотя скорость роста в мыши SCID (в отсутствие В-клеток) показала значительную разницу только в сравнении с мышью, негативной по Т-клеткам (голой). Эти результаты означают, что, очевидно, tag7 восстанавливает В-клетки и активизирует их цитотоксическое действие, приводя к апоптозу опухолевых клеток. Индуцирование защитного иммунитета посредством tag7 позволяет придти к выводу, что перенос tag7 возможно является эффективной терапевтической стратегией.
Имея описание изобретения, для ясности понимания дополненное некоторыми деталями с помощью иллюстраций и примеров, любой обычный специалист в данной области с очевидностью поймет, что то же самое можно провести путем модификаций и изменений изобретения в широких пределах равнозначных условий, формул и других параметров без ущерба для объема изобретения или любых его конкретных воплощений и что такие модификации или изменения, как предполагается, охватываются объемом прилагаемой формулы изобретения.
Все публикации, патенты и патентные заявки, упомянутые в этом описании, характеризуют уровень знаний тех, кто является специалистом в той области, к которой относится данное изобретение, и приведены здесь лишь в качестве ссылки в такой же степени, как если бы каждая отдельная публикация, патент или патентная заявка были специально и индивидуально приведены как ссылка.
Примеры
Материалы и методы
Во всех примерах в основном использованы следующие материалы и методы.
Опухоли и клеточные линии
Метастазирующие (VMR-L) и неметастазирующие (VMR-0) линии рака мышиной молочной железы были получены как трансплантаты в мышь A/Sn. В случае VMR-0 трансплантация была осуществлена с использованием опухолевого материала, в случае VMR-L-линии - с использованием клеток из метастазов в печени. Опухоль имела 1см в диаметре, развивалась в месте инъекции 2-3 недели после внутримышечной инъекции клеток в количестве 5×105. Клетки L929 были получены из American Type Culture Collection (номер ATCC:CRL-2148; Rockwill, Maryland). Клетки из VMR-0, VMR-L, VMR-Lym (см. статью SeninV.M. в журнале Vestn.Akad.Med.Nauk. USSR 0(5):85-91 (1984)), CSML-0, CMSL-100 (см. статью Senin V.M. и др. в журнале Experim. Oncol. (USSR) 5(63):35-39 (1983)), Line 1, LL Met, 10 Т1/2, МТ1 TC1, MT1 ТС3 (см. статью Barnett S.C. и Eccless S.A. в журнале Exp.Metast. 2(1):15-36 (1984)), RAC 5Е, RAC 34E, RAC 10Р, RAC 311C (см. статью Sonnenberg А. и др. в журнале Cancer Res. 46:5913-5922 (1986)) и клеточные линии L929 были выращены в атмосфере 5% СО2 в среде DMEM, содержащей 10% фетальной бычьей сыворотки (FBS) и пенициллин и стрептомицин по 100 единиц/мл.
Клеточная линия М3-мышиной метастатической меланомы (АТСС CCL 53.1): Клетки культивировали в среде RPMI 1640, содержащей 5% FBS, 100 ед/мл стрептомицина, 10 мМ HEPES.
Для исследований in vivo клетки VMR-0 были трансфецированы с использованием трансфекционного реагента DOTAP (Boehringer Mannheim), в соответствии с рекомендациями изготовителя. Клетки выращивали в среде, содержащей G418, в течение двух недель перед отбором индивидуальных клонов.
Мышиные спленоциты, тимоциты, моноциты и перитонеальные макрофаги выделяли, как было опубликовано ранее (см. Klaus G.G.B. Lymphocytes: A Practical Approach, Klaus G.G.B., изд. Oxford U.K.:IRL Press, Chapter 1 (1988)). Клетки культивировали в среде RPMI-1640, содержащей 5% FBS, по 100 ед/мл пенициллина и стрептомицина и 10 мМ HEPES. В некоторых исследованиях клетки активировали с помощью бактериального липополисахарида (LPS) в среде, свободной от сыворотки, в течение различных периодов времени.
Клетки VMR-0 и VMR-L, стимулированные LPS, и кондиционированные среды были получены, как уже было раскрыто ранее (см. статью Sheehan K.C. и др. в журнале J.Immunol. 142:3884-3893 (1989.)). Говоря кратко, клетки VMR-L обрабатывали LPS (6 мкг/мл) в течение 18 часов при 37°С в среде, свободной от сыворотки, не содержащие клеток супернатанты очищали центрифугированием и использовали для проведения цитотоксических анализов или иммунологических исследований.
Выделение РНК
Тотальную клеточную РНК выделяли с помощью гуанидин-изотиоцианатного метода (см. статью Chomczynski P. и Sacchi N. в журнале Anal. Biochem. 162:156-159 (1987)). Полиаденилированную мРНК извлекли путем двух циклов хромотографии на олиго(dТ)целлюлозе (см. Maniatis Т. и др., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press, стр.545 (1982)).
Удаление хромосомальной ДНК из препаратов РНК
50 мкг тотальной РНК инкубировали в течение 30 мин при 37°С с 10 единицами ДНК-азы I и буфером, содержащим 10 мМ TRIS-HCl, рН 8,3, 50 мМ КСl и 1,5 мМ MgCl2 в присутствии 10 единиц ингибитбра человеческой плацентарной рибонуклеазы.
Выявление различий в РНК
Выявление различий в мРНК проводили по стандартньм методикам (см. статью Liang Р. и Pardee A.B. в журнале Science 257:967-971 (1992)). Коротко говоря, кДНК получили из 0,2 мкг РНК с помощью реакции обратной транскрипции, используя праймер Т12 АС (5'-ТТТТТТТТТТТТАС-3') (SEQ ID NO:3) и обратную транскриптазу вируса лейкемии мыши Moloney (M-Mul V). T12 AC и два коротких случайных 5'- олигонулеотидных праймера (праймер 1:5'-AATCGGGCTG-3'(SEQ ID NO:4); праймер 2:5-AGTCAGCCAC-3' (SEQ ID NO:5) были использованы как 3' -праймеры в двух различных комбинациях в процессе полимеразной цепной реакции (PCR). Амплифицированные кДНК разделили электрофорезом в 6% полиакриламидных гелях, содержащих 7М мочевину.
Электрофорез и гибридизация
Электрофорез РНК проводили в 1,2% агарозном геле в присутствии 6% формальдегида и 50 мМ фосфатного буфера (см. Maniatis Т. и др., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press, стр.545 (1982)). На каждую дорожку было нанесено до 20 мкг тотальной РНК. Вслед за электрофорезом РНК перенесли из гелей на найлоновые фильтры (Hybond-N, Amersham, England) и гибридизировали с 32Р-меченными кДНК (см. статью Southern E.M. в журнале J.Mol.Biol. 98:503-517 (1975)). Фрагменты EcoRI/Xhol из клона самой длинной кДНК помечали случайным праймированием с помощью набора Random Primed Labeling kit (Boehringer Mannheim, Austria) по инструкциям производителя и использовали в качестве гибридизационных проб. Для выравнивания количества РНК, нанесенного на каждую линию, осуществляли гибридизацию с пробой ДНК GADPH. Гибридизацию выполняли, как уже известно (см. Maniatis Т. и др., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press, стр.545 (1982)).
С целью гибридизации tag7 с человеческими тканями, блот Human Immune System Multiple Tissue Northern™ Blot II (Clontech), обработали пробой - 32Р-меченной кДНК tag7 приготовленной, как описано выше. Блот содержал примерно 2 мкг полиА+РНК на дорожку из следующих человеческих тканей: селезенки, лимфатических узлов, зобной железы, лейкоцитов периферической крови, костного мозга и печени эмбриона. Затем блот гибридизовали при определенных условиях гибридизации следующим образом: провели предгибридизацию блота при 65°С в течение двух часов в Church-буфере (0,5 М фосфата натрия (рН 7,2), 7% SDS, 1мМ EDTA); пробу денатурировали при 95°С в течение 5 мин и добавили к 7 мл свежеприготовленного Church-буфера для гибридизации; гибридизацию провели при 55°С, и блот трижды промыли при 50°С каждый раз в течение 15 минут в Church-промывочном буфере (40 мМ фосфата натрия (рН 7.2), 1% SDS). Затем блоты экспонировали на рентгеновской пленке в течение 24 часов.
Клонирование и определение нуклеотидной последовательности фрагментов кЦНК
Амплифицированные продукты полимеразной цепной реакции (PCR) клонировали в вектор pGEM-T (Promega, USA). Нуклеотидную последовательность определили по методике Sanger (см. статью Sanger F. и др. в журнале Proc.Natl.Acad.Sci. USA 74:5463-5467 (1977)), а также автоматически, используя автоматический секвенатор (Automated Sequenator (Applied Biosystems)) в соответствии с предлагаемыми изготовителем протоколами.
Конструирование библиотеки кДНК
Двухцепочечную кДНК синтезировали на полиА+РНК, выделенной из опухолей VMR-L, используя набор ZAP-cDNA Gigapack Cloning (Stratagene) в соответствии с рекомендациями изготовителя и клонировали в λZAP-вектор в EcoRI и XhoI участки. Фрагмент кДНК из 390 нп использовали в качестве пробы для Нозерн-гибридизации, и скрининг библиотеки провели по стандартным методикам (см. Maniatis Т. и др., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press, стр.545 (1982)). Положительные клоны очистили и удалили вставки как клоны pBluescript, используя вспомогательный фаг, как описано изготовителем. Эти клоны секвенировали, применив набор для секвенирования Sequenase Version 2.0 (USB-American) и синтетические олигонуклеотидные праймеры (Applied Biosystem 391 DNA Synthesizer). Геномную библиотеку сконструировали в λFIX II -векторе (Stratagene) и провели скрининг в соответствии со стандартной процедурой (см. Sambrook J. и др., Molecular Cloning: A Laboratory Manual, изд.2. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press, (1989)). Вставки субклонировали в вектор pGEM7Z и частично секвенировали, как описано выше.
Были синтезированы олигонуклеотиды, соответствующие 5' и 3'-концам кодирующих областей мышиного гена tag7 с рестрикционными сайтами ВаmHI и HindIII, добавленными на концах олигонуклеотидов. Кодирующую область гена амплифицировали стандартными PCR-методиками, вырезали по BamHI и HindIII и встраивали в рамку считывания по BamHI и НindIII участкам рестрикции вектора экспрессии pQES30 (Qiagen).
Полноразмерный фрагмент кДНК tag7 вырезали из клона pBluescript рестрикционными ферментами ХbаI и XhoI и субклонировали в векторе экспрессии у млекопитающих pBK-CMV (Qiagen) в сайты NheI и XhoI.
Хромосомное картирование гена tag7
Флюоресцентную гибридизацию in situ на мышиных метафазных хромосомах проводили с помощью коммерческого набора Genome Systems, Ink. (St. Louis, Missouri).
Иммунологические методы
Продуцированный Е. coli рекомбинантный белок tag7 (rtag7) был экспрессирован в M15[pREP4] (Qiagen) и очищен нa Ni-NTA-агарозе (Qiagen), соблюдая рекомендации изготовителя. Антитела из кролика, вызванные против rtag7, были аффинно очищены на колонке с Сефарозой (Pharmacia; Piscataway, New Jersey), содержащей rtag7, иммобилизованный в соответствии с рекомендациями изготовителя. Электрофорез в полиакриламидном геле с додецилсульфатом натрия, иммуноосаждение и иммуноблоттинг проводили по стандартным методикам (см. Sambrook J. и др., Molecular Cloning: A Laboratory Manual, изд.2. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press, (1989)).
Цитотоксический анализ tag7 и нейтрализация цитотоксической активности
В качестве источника нативной формы белка tag7 использовали культуральную среду, кондиционированную клетками VMR-L, стимулированными LPS. Клетки L929 культивировали в 96-луночных планшетах при густоте 3×104 на лунку. После инкубирования в течение ночи клетки обработали актиномицином D (1 мкг/мл) в течение двух часов при 37°С в среде, не содержащей сыворотки. Вслед за этим добавили среду, кондиционированную VMR-L, стимулированными LPS, или среду, содержащую LPS, в количестве по 100 мкл на лунку. Где было указано, добавили рекомбинантный фактор-α некроза человеческой опухоли (rhTNF; Sigma) с концентрацией 100 нг/мл и в объеме 100 мкг на лунку. Чтобы определить гибель клеток, использовали набор для анализа Сую Тох 96 Assay kit (Promega) или окрашивали клетки трипановым синим, и закодированные образцы подсчитывали с помощью микроскопа слепым методом при минимальном количестве 100 клеток в каждой группе.
Нейтрализацию цитотоксического эффекта белка tag7 или rhTNF провели с использованием афинно-очищенных поликлонального антитела антиtag7 из кролика и поли-клонального антитела анти-hTNF (Sigma). Поликлональные антитела добавили в среду, кондиционированную LPS-индуцированными VMR-L, с конечной концентрацией 2 мкг/мл, и определили цитотоксический эффект, как было описано выше.
Анализ фрагментации ДНК
Анализ фрагментации ДНК провели, как уже описано (см. статью Tian Q. и др. в журнале Cell 67:629-639 (1991)), с модификациями. Кратко говоря, 2×106 клеток L929 подвергли прединкубированию с актиномицином D (1 мкг/мл) в течение 2 часов при 37°С в среде, не содержащей сыворотки, и затем в течение 5 часов инкубировали со средой, кондиционированной LPS-индуцированными VMR-L, или с rhTNF. Клетки собирали и лизировали в 20 мМ Tris-HCL (pH 8.0) и 10 мМ EDTA (ТЕ-буфер) при содержании 0,8% Triton Х-100. После центрифугирования из супернатантов была осаждена ДНК при -20°С изопропанолом в присутствии NaCl. Осажденную ДНК повторно суспендировали в ТЕ-буфере, обработали РНК-азой А и фрагменты разделили электрофорезом в 1,8% агарозном геле в ТЕ-буфере.
Исследования на животных
Для изучения влияния tag7 на прогрессирование опухоли in vivo были использованы мыши A/Sn в возрасте от 6 до 10 недель. Экспоненциально растущие клетки собрали, промыли в PBS и подкожно инъецировали в спинную область мыши в объемной дозе PBS 0,2 мл. Размер опухоли измерили в миллиметрах с помощью калибра и регистрировали, как уже описано (см. статью Hock H. и др. в журнале Proc.Natl.Acad.Sci. USA 90:2774-2778 (1993)). Для нейтрализации активности tag7 in vivo 100 мкг аффинно-очищенных антител анти-tag7 инъецировали подкожно, вместе с 106 опухолевых клеток в нулевой день и затем такое же количество антител инъецировали в место расположения опухоли дозой PBS объемом 0,2 мл на 2-й, 4-й, 6-й и 8-й дни. Контрольных животных в те же дни инъецировали только PBS.
Пример 1: Распознавание генов, различно экспрессируемых в опухолях, имеющих разную способность к метастазированию
В этих исследованиях для распознавания генов с различным уровнем экспрессии в двух родственных линиях опухолей, различающихся по их метастатическим характеристикам, была использована методика "выявления различий в РНК" (см. статью Liang P. и Pardee A.B. в журнале Science 257:967-971 (1992)).
Полученный с помощью методики "выявления различий в РНК" спектр фрагментов кДНК, экспрессируемых в мышиных опухолях, которые метастазируют в печень (VMR-0) и не метастазируют в печень (VMR-L), показан на фиг.3. Было обнаружено, что спектр фрагментов кДНК, амплифицированных в присутствии одинаковых пар праймеров, в основном идентичен для обеих опухолевых линий. Однако, когда проанализировали бальшое количество комбинаций праймеров, появилась возможность идентифицировать различия в экспрессии некоторых последовательностей РНК (стрелки на фиг.3). Фрагменты ДНК, которые не имели аналогов на соседних дорожках, были элюированы из геля, амплифицированы в присутствии одинаковых пар праймеров и клонированы. Различие в уровнях экспрессии последовательностей РНК в двух проанализированных опухолевых линиях проверили с помощью Нозерн-гибридизации клонированных фрагментов с тотальной РНК из двух опухолей. В результате этого анализа были получены два фрагмента, которые различным образом гибридизировались с РНК из клеток VMR-0 и из клеток VMR-Liv; эти фрагменты были обозначены как tag7 (фиг.4) и tag6 (данные не показаны). Было найдено, что транскрипт tag7 хорошо представлен в мРНК, выделенной из опухолевой линии VMR-L с высокой способностью к метастастазированию (фиг.4, дорожка 2), но не определялся в мРНК, полученной из опухолевой линии VMR-0, не обладающей способностью к метастазированию (фиг.4, дорожка 1).
Основываясь на результатах Нозерн-гибридизации, длину транскрипта tag7 определили приблизительно в 750 нуклеотидов. Нуклеотидная последовательность мышиного гена tag7 (SEQ ID NO:l) была определена секвенированием по Sanger'y и автоматическим секвенированием и показана на фиг.1. Было обнаружено, что ген tag7 фланкируется на обеих сторонах одним и тем же праймером 5'-AATCGGGCTG-3' (SEQ ID NO:6), используемым в полимеразной цепной реакции (PCR). Поскольку эта последовательность не имеет гомологии с известными генами (определено изучением генетических баз данных Genbank и EMBL), была клонирована полноразмерная кДНК этого гена. Библиотека кДНК, полученная обратной транскрипцией тотальной мРНК из опухолевой линии VMR-Liv, содержала приблизительно 800,000 клонов. Были отобраны 52 клона, содержащие кДНК гена tag7, путем гибридизации с фрагментом гена tag7, меченным 32P. Нуклеотидную последовательность определили для нескольких клонов секвенированием по Sanger'y и автоматическим секвенированием, и кодирующая последовательность tag7 наиболее протяженного клона представлена на фиг.1 (SEQ ID NO:l). Последовательность этого клона и фрагмент, полученный при выявлении различий, как было найдено, практически идентичны. Было обнаружено, что нуклеотидная последовательность кодирующего сегмента клона tag7 составляет 549 нп (Фиг.1; SEQ ID NO:1) и кодирует полипептид из 182 аминокислотных остатков (Фиг.1; SЕQ ID NO:2).
Анализ предсказанной аминокислотной последовательности мышиного полипептида tag7 (Фиг.1; SEQ ID NO:2) показывает, что имеет место протяженный участок высоко гидрофобных аминокислот на N-конце полипептида, который предположительно действует как домен, заякоривающийся в мембране, а затем следует потенциальная секреторная сигнальная последовательность (между 20 и 22 аминокислотными остатками). Изучение базы данных по белкам (программа FASTA от IntelliGenetics и BLAST от NCBI) не выявило сколь-нибудь существенного подобия между полипептидом tag7 и теми генными продуктами, о которых сообщалось ранее. Мышиная геномная библиотека была сконструирована в векторе λ FIX II, подвергнута скринингу, и была установлена геномная структура tag7. Было найдено, что мышиный ген tag7 измеряется 8 Kb, состоит из трех экзонов и содержит неконсенсусный ТАТА-бокс (не показано). Стартовый участок РНК был картирован с использованием праймера удлинения (данные не показаны), показав два участка инициации, разделенных четырьмя нуклеотидами на 22 нп ниже по цепи от ТАТА-бокса, вероятно, вследствие неконсенсусной структуры последнего. Компьютерный анализ приблизительно 200 пар оснований последовательности выше по цепи (Signal Scan) показал некоторое количество потенциальных сайтов связывания для некоторых известных транскрипционных факторов, например Ets-1, NF-kb Sp-1 и myoD. Порядок этих сайтов связывания и расстояние от стартового сайта мРНК были очень близки контрольной области мышиного гена LT-β (см. статью Pokholok D.K. и др. в журнале Proc.Natl.Acad.Sci. USA 92:674-678 (1994)). Более того, 5'-нетранслированная область мРНК tag7, как было обнаружено, является относительно короткой, что типично для генов лимфотоксина.
Эти подобия между геномными структурами tag7 и мышиного LT-β подтвердили более детальный анализ аминокислотной последовательности, который показал существование четырех областей во внеклеточных доменах полипептида tag7, структурно подобных внеклеточным доменам других членов семейства TNF. Было найдено, что в первых двух областях уровень идентичности с другими лигандами, относящимися к TNF, высок (данные не приведены), в особенности в остатках, заключенных внутри β-складки (см. статью Smith С. и др. в журнале Cell 76:959-963 (1994)). Четвертый домен обнаружил меньше гомологии, но расположенные внутри остатки все же были консервативными. Было также найдено, что tag7 изобилует цистеиновыми остатками, так же как LT-α (см. статью Aggarwal B.B. и др. в журнале J.Biol.Chem. 269:2334-2339 (1985)), содержит четыре метиониновых остатка. Расстояние между предположительным трансмембранным доменом и доменом в tag7, связывающим рецептор, было приблизительно таким же, как у LT-α (25 аминокислот) и у TNF-α (31 аминокислота).
Для дальнейшего пояснения трехмерной структуры полипептида tag7 и определения ее связи с клеточной мембраной и потенциальными антигенными областями полипептида, были построены диаграммы гидрофильности по Kyte-Doolittle и индекса антигенности Jаmeson-Wolf для полипептида tag7 с использованием компьютерной программы PROTEAN (DNASTAR, Ink.; Madison, Wisconsin). Как показано на фиг.2, полипептид tag7 содержит последовательность из высоко гидрофобных аминокислотных остатков на N-конце (приблизительно от 1 до 20 остатков), наиболее вероятно представляющую N-концевую сигнальную последовательность длиной примерно в 20 аминокислот). Кроме того, график индекса антигенности (фиг.2) выявляет по меньшей мере четыре потенциальных эпитопнесущих областей полипептида tag7, - на аминокислотах 20-40, аминокислотах 55-75, аминокислотах 90-110 и аминокислотах 145-160, а также пятую область с потенциально слабой антигенностью на аминокислотах 120-130.
Пример 2: Анализ транскрипции мышиного гена tag7 в различных нормальных и опухолевых клетках и тканях у мышей
Для проверки уровней транскрипции tag7 в различных нормальных и опухолевых клетках и тканях из различных органов здоровой мыши выделили тотальную РНК и исследовали путем гибридизации с 32Р-меченной пробой ДНК tag7. Как показано на фиг.5, наивысший уровень транскрипции tag7 в здоровой мыши наблюдали в селезенке и легких, и более низкие уровни - в головном мозгу и зобной железе. Было найдено, что транскрипт tag7 во всех органах имел одинаковую длину - около 750 нуклеотидов, включающих кодирующий сегмент 549 нп плюс дополнительные 5' и 3'-нетранслированные области, включающие 3' полиА + хвост (не показано). В печени или любых других органах не наблюдалось экспрессии tag7 даже после продолжительного экспонирования радиавтограмм. Вместе эти результаты показывают, что экспрессия клонированного гена tag7 в мыши очень тканеспецифична.
Однако при проверке различных опухолевых клеточных линий, как было обнаружено, транскрипция гена tag7 не ограничивалась метастатической линией VMR-L. Экспрессия tag7 наблюдалась также в опухолевых линиях семейства VMR, обладающих наиболее высокой способностью к метастазированию, особенно в опухоли VMR-Ov (фиг.6). Ген tag7 также транскрибировался на низком уровне в линии CSML-О, в которой повторяемость метастазирования в легкие составляет около 5%.
Для пояснения связи гена tag7 с процессом метастазирования, РНК из разных культур клеток из опухолей, обладающих различной способностью к метастазированию и специфичностью метастазирования по органам, а также имеющим разное происхождение, были гибридизованы с фрагментом кДНК гена tag7; результаты показаны в таблице 1.
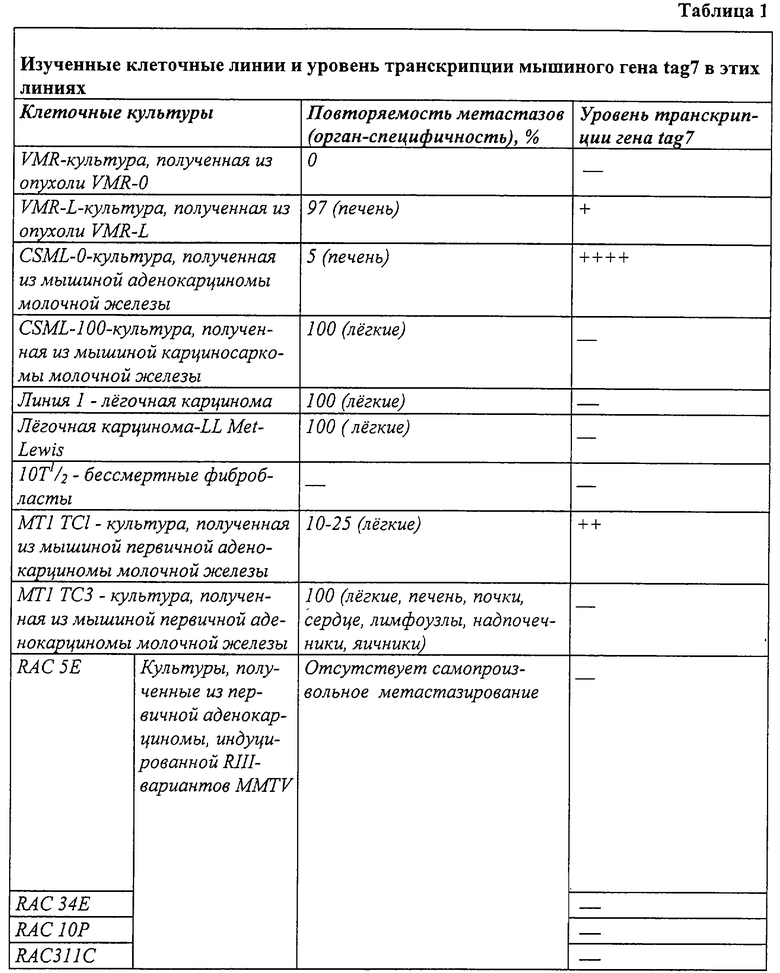
Интересно, что эти результаты демонстрируют то, что, когда опухолевые штаммы превращаются в линии трансплантируемых культур, наблюдались изменения в уровне транскрипции гена tag7 in vitro: транскрипция гена tag7 значительно понижается в клеточной линии VMR-L, в то время как в клеточной линии CSML-0 наблюдался умеренный уровень экспрессии tag 7. Ген tag7 транскрибировался на довольно высоком уровне в линии МТ1 ТС1 - культуре, полученной из мышиной первичной аденокарциномы молочной железы, которая метастазирует в легкие в 10-25% случаев.
Для более тщательной проверки корреляции между экспрессией мРНК tag7 и способностью опухолей к метастазированию применили Нозерн-гибридизацию. Однако при гибридизации тотальной РНК опухолей CSML-0 и CSML-100 не наблюдалось никакой корреляции (фиг.7). Более того, выявилось, что экспрессия гена tag7 в этой паре является специфичной для опухоли с низкой способностью к метастазированию, CSML-0. Более интригующим является подтверждение вышеупомянутого наблюдения, что уровень мРНК tag7 разительно изменился после образования из первичной опухоли клеточной культуры. В других исследованных клеточных линиях из мышиных опухолей не имелось никакой корреляции между способностью к метастазированию и экспрессией мРНК tag7 (данные не показаны).
Для того чтобы определить распределение по ткани транскрипции tag7 провели Нозерн-блоттинг и гибридизацию in situ. Нозерн-гибридизация нескольких видов тканей взрослой мыши (фиг.8А) показала наивысший уровень транскрипции tag7 в легком и селезенке, регистрируемый уровень в головном мозгу и зобной железе и полное отсутствие или только низкий уровень экспрессии мРНК в других исследуемых тканях. Однако гибридизация in situ на срезах отобранных органов дала регистрацию транскриптов tag7 в определенных областях головного мозга и кишечника (фиг.8В-Е).
Был также проверен уровень транскрипции tag7 в кроветворных и лимфоидных клетках, которые, как известно, являются главным источником цитокинов из семейства TNF. Мышиные спленоциты и резидентные перитонеальные макрофаги продемонстрировали существенно высокий уровень мРНК tag7 (фиг.9). Стимуляция IL-2 или фитогемагглютином не привела к значительному увеличению содержания мРНК tag7 в тимоцитах и макрофагах за время проведения исследования (данные не показаны). В культивированных с LPS мышиных спленоцитах уровень транскрипции tag7 увеличивался в течение первых часов активации (максимум 24 часа постстимуляции) (фиг.9). Нозерн-блоттинг клеточной линии WEHI-231 лимфомы мышиных В-клеток и линии LBRM-33 лимфомы Т-клеток не показал регистрируемого уровня транскрипции tag7 в присутствии или отсутствии РМА, LPS или ионофора кальция в течение 20 часов (данные не показаны).
Пример 3: Хромосомное картирование мышиного tag7
Для определения хромосомной локализации tag7 в мышином геноме метафазные хромосомы анализировали путем флюоресцентной гибридизации in situ. Общее количество проанализированных метафазных клеток составляло 80, причем из них 62 обнаружили tag7-специфичное мечение. В этих опытах наблюдалось специфичное мечение центромерной области хромосомы 7 (данные не показаны). Измерения десяти специфически меченных хромосом 7 показало, что сигнал tag7 локализован в положении, которое составляет примерно 9% расстояния от центромерной области, области, которая соответствует полосе 7А3.
Пример 4: Экспрессия растворимого мышиного tag7 в клеточных линиях и лимфоидных клетках
Для изучения экспрессии tag7 на белковом уровне в Вестерн-блоттинге использовали поликлональные антитела кролика, полученные против rtag7. В то время как белок tag7 не определялся в культивированных клетках VMR-L и в культуральных средах, кондиционированных этими клетками, в среде, кондиционированной LPS-стимулированными клетками VMR-L, был зарегистрирован высокий уровень белка tag7 (фиг.10А). Эти результаты позволяют предположить, что, так же как TNF и LT-α, белок tag7 может существовать в двух формах: растворимой и связанной с клеткой. Предполагаемая форма белка клеточной поверхности была определена в клеточных экстрактах, но больше всего белка секретировалось в культуральной среде после обработки LPS. Хотя увеличение уровней мРНК tag7 в процессе активации незначительно (данные не показаны), степень синтеза и секреции белка tag7 относительно высока. Такой же характер экспрессии гена наблюдался также у линии клеток CSML-0, экспрессирующих tag7 (данные не показаны). Свежевыделенные мышиные спленоциты содержали значительное количество связанного с клетками белка tag7 (фиг.10В), которое было увеличено примерно в 5 -7 раз посредством LPS-стимуляции с максимальной постстимуляцией около 24 часов. Интересно, что эти увеличенные уровни экспрессии белка tag7 временно совпадают с изменениями в локализации белка: в то время как больше всего белка tag7 было обнаружено в форме, связанной с клетками в первое время после стимуляции LPS, продолжительное воздействие LPS на клетки приводит не только к увеличению общего количества белка, но также к его секреции. Различия в количестве белка tag7 в неактивированных клетках VMR-L и спленоцитах (фиг.10А,В) могут отражать тот факт, что для этих двух клеточных типов задействованы разные механизмы активации гена, и/или то, что спленоциты уже были активированы in vivo перед их выделением из селезенки.
Пример 5: Фрагментация ДНК, вызванная растворимым мышиным tag7
Было показано, что фактор некроза опухоли (TNF) вызывает апоптоз in vitro в различных клеточных линиях, в некоторых случаях через синергическое взаимодействие с актиномицином D (см. статьи Browning J. и Ribolini А. в журнале J.Immunol. 143:1859-1867; Larrick и др. (1990). Подобия между структурой гена tag7 и цитокинов, относящихся к TNF, и существование растворимой формы белка позволили придти к выводу, что полипептид tag7 может также запускать запрограмированную гибель клетки.
Чтобы исследовать эту возможность для растворимой формы tag7, клетки L929 обработали кондиционированньми супернатантами из как нестимулированных, так и из LPS-стимулированных клеток VMR-L. Уничтожение клеток-мишеней после инкубации со 100 мкг кондиционированных супернатантов подвергли мониторингу путем окрашивания трипановым синим. Результаты, представленные на фиг.11А, показали, что и TNF, и tag7 убивали клетки L929 в присутствии актиномицина D, но гибель клетки, индуцированная tag7, ингибировалась добавлением антител-анти tag7.
Поскольку процесс апоптоза быстро вызывает фрагментацию ДНК, была проверена способность секретированной формы tag7 запускать апоптоз с использованием стандартных способов анализа фрагментации ДНК. L929 клетки-мишени были обработаны или кондиционированными супернатантами VMR-L клеток, стимулированных LPS, или LPS-содержащей средой и рекомбинантными TNF в качестве контролей. После пяти часов инкубации фрагментированную ДНК в клеточной цитоплазме повторно выделили и разделили с помощью электрофореза в агарозном геле. Как показано на фиг.11В, клетки L929 начинали испытывать апоптоз после обработки растворимой формой белка tag7, как доказывается внутринуклеосомальной фрагментацией ДНК. В противоположность этому, контрольные клетки не показали фрагментации ДНК. Эти результаты означают, что обработка tag7 может индуцировать апоптоз в некоторых опухолевых клетках, и приводят к выводу о том, что, как и TNF, tag7 может вызвать запрограмированную гибель клеток также и в других клетках млекопитающих.
Пример 6: Ингибирование развития опухоли, индуцируемое tag7
а) Получение клеточных линий, продуцирующих Tag 7
Продемонстрировав, что tag7 может вызывать апоптоз in vitro, далее проверили влияние tag7 на развитие опухолей in vitro. Для облегчения этих биологических исследований полноразмерные копии кДНК мышиного гена tag7 были субклонированы в векторы экспрессии млекопитающих. Клетки VMR-0, не экспрессирующие сколь-нибудь существенно мРНК tag7 генетически трансформировали этими векторами для получения мРНК tag7; трансфектанты отбирали по устойчивости к G418 и клонировали. Некоторое количество клонов анализировали на уровень транскрипции tag7, и два клона (4SX и 12SX), различающихся по уровню мРНК tag7, отобрали для дальнейшего анализа (фиг.12А). кДНК tag7 клонировали в вектор pM5Gneo и провели устойчивую трансфекцию клеток VMR-0.
Модифицированные клетки отбрали по G418-устойчивости, клонировали и анализировали (клон 8SX). Этот вектор позволил получить некоторое количество белка, определяемое Вестерн-блоттингом (фиг.12 D, верхняя полоса), и цитоксичность культуральной среды (нижняя полоса), в то время как при использовании промотора CMV регистрируются только мРНК tag7 (клоны 12SX, 4SX). Для этого опыта кондиционированную среду клона 8SX отфильтровали через мембрану с размером отверстий 0,2 мкм и добавили к клеткам-мишеням (клетки L929; 100 мкл на лунку, содержащую примерно 104 клеток). После четырех часов инкубации измеряли цитотоксичность путем окрашивания Трипановым Синим, используя набор Cyto Tox Kit (Promega). Однако вектор pM5Gneo, несущий часть gag-гена, вируса миелопролиферативной саркомы, и клетки, псевдотрансфецированные этим вектором, были существенно более иммуногенными в иммунодефицитных животных, чем контрольные клетки, трансфецированные плазмидой pBK/CMV. Родительские клетки VMR-0 и псевдотрансфецированные клетки VMR-0 Neo не продуцировали регистрируемого уровня мРНК tag7. Генетически модифицированные клетки VMR-0 сколько-нибудь существенно не отличаются от родительских клеток как в морфологии, так и в способности пролиферации клеток in vitro.
b) Экспрессия гена tag7 подавляет развитие клеток VMR-0 in vivo в сингенной мыши
Для того чтобы определить, могут ли опухолевые клетки, будучи трансфецированы tag7, изменить свою способность к образованию опухолей, 106 клеток, трансфецированных tag7, и контрольных клеток были подкожно (s.c.) имплантированы в сингенную мышь A/Sn. Клетки, экспрессирующие tag7, росли медленнее, чем контрольные клетки (фиг.12В: незакрашенные квадратики - родительские опухолевые клетки VMR-0, закрашенные треугольники - псевдотрансфецированные VMR-0, незакрашенные кружочки - подавление роста 4SX путем инъекции поликлонального антитела анти tag7 в место расположения клеток, закрашенные квадратики - наблюдаемый эффект совместной инъекции родительских (VMR-0) и tag7-модифицированных опухолевых клеток (4SX), незакрашенные треугольники - скорость роста клона 12SX, закрашенные кружочки - скорость роста клона 4SX.C - родительских и модифицированных (клон 4SX) опухолей, полученных путем иссечения из сингенных животных), и скорость подавления развития опухоли коррелируется с уровнем мРНК tag7 и содержанием белка. В контрольных животных наблюдается прогрессивный рост родительских и псевдотрансфецированных клеток.
Фиг.12С показывает опухоли, образованные родительскими клетками VMR-0, иссеченные через 4 недели после инъекции, и опухоли, образованные клетками 4SX, иссеченные через 5 недель после инъекции.
Самым интригующим наблюдением явилось полное отсутствие геморрагического некроза, даже когда опухоли достигли довольно большого размера через 90 дней после инъекции, тогда как родительские и псевдотрансфецированные опухоли начали отмирать менее чем через две недели после инъекции. Это наблюдение далее подтвердилось гистологическим анализом опухолей, образование которых вызвано инъекцией родительских клеток (клеток VMR-0) и tag7-модифицированных клеток (клеток 4SX). Антипролиферирующее влияние на развитие опухоли является непрямым, так как опухолевые клетки не показали изменений в кинетике развития in vitro. Непрямое влияние на подавление развития опухоли in vivo подтверждается совместной имплантацией клеток 4SX и родительских клеток VMR-0 (по 106, всего 2×106) подкожно в то же самое место. Хотя развитие опухоли ингибировалось не так сильно, как только с трансфецированными клетками, оно показало явно наблюдаемый эффект немодифицированных опухолевых клеток. Чтобы проверить, что уменьшение способности к образованию опухоли явилось результатом секреции tag7 инъецировали подкожно модифицированные опухолевые клетки в сингенную мышь вместе с поликлональными антителами анти-tag7. Уменьшение способности модифицированных опухолевых клеток к образованию опухолей практически блокировалась инъекцией антител анти-tag7 с интервалом 2 дня, и отмена их введения к 8-му дню привела к немедленному восстановлению антипролиферирующего эффекта.
Инъекция tag7-несущих опухолевых клеток не вызвала каких-либо аномалий (например, потери веса) у мышей-хозяев в период опытов. Это наблюдение означает, что секретированный tag7 может индуцировать противоопухолевый эффект по отношению к родительским опухолевым клеткам без возникновения системной токсичности.
Пример 7: Скорость развития модифицированных опухолевых клеток зависит от иммунного состояния организма
Чтобы проанализировать возможные клеточные механизмы, участвующие в антиопухолевой реакции, индуцированной tag7, трансформированные опухолевые клетки анализировали также в голой мыши (дефектной по Т-клеткам) и мыши SCID (дефектной по Т- и В-клеткам) (фиг.13). Для этого опыта использовали только клетки с высоким уровнем tag7 (4SX). Размер опухоли отслеживали в течение 40 дней. Родительские и псевдотрансфецированные клетки имеют совершенно одинаковую кинетику развития в голой мыши и мыши SCID. Скорость развития модифицированных опухолей подавлялась в обеих моделях, однако размер опухоли в конце испытательного срока в SCID-модели был на 50% больше, чем у голой мыши. Это означает, что отсутствие В-лимфоцитов может играть роль в частичном восстановлении скорости роста опухоли.
Фиг.14 показывает голую мышь, зараженную tag7-модифицированными клетками VMR-0 (верхняя фотография, клон 4SX) и родительскими клетками (нижняя фотография).
Пример 8: Отторжение опухоли является следствием процесса апоптоза в модифицированных опухолях
Отторжение tog-7-трансфектантов не зависело от цитотоксических Т-лимфоцитов, потому что оно наблюдалось в голой мыши, дефектной по Т-клеткам. Чтобы понять лежащие в основе этого клеточные механизмы, провели гистологический анализ и электронную микроскопию опухолевой ткани сингенных животных через 3 недели после инъекции. Контрольная опухоль VMR-0 показала непрерывный рост и центральные некротические области. Количество митотических клеток было относительно высоким, примерно 6-8% клеток находились на стадии митоза и апоптозные клетки в родительской опухоли были редки (таблица 2 показывает, что общая разница в пролиферативной активности между родительскими и модифицированными клетками была более чем 50 раз).
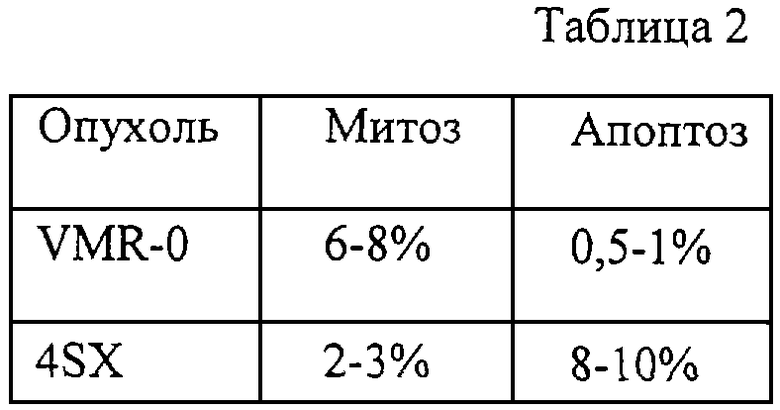
Модифицированные опухоли были разрушены и освобождены от некротических очагов. Электронная микроскопия показала, что более 8% опухолевых клеток обладают свойствами, характерными для апоптоза, начиная с аггрегации конденсированного хроматина, ядерной фрагментации и т.д. Верхняя фотография на фиг.15А изображает гистологический срез опухоли, образованной родительскими клетками (клетками VMR-0), окрашенной эозин/гематоксилином. Нижняя фотография на фиг.15А изображает гистологический срез опухоли, образованной tag7-модифицированными клетками (клетки 4SX), окрашенной эозин/гематоксилином. Апоптозные клетки обозначены стрелками. Фиг.15В показывает полученные электронной микроскопией изображения митотических клеток в опухоли, образованной родительскими клетками (клетки VMR-0); фиг.15С показывает полученные электронной микроскопией изображения апоптозных клеток в опухоли, образованной tag7-модифицированными клетками (клетки 4SX).
Пример 9: В-клетки опосредуют отторжение первичной опухоли
Хотя в мыши SCID рост модифицированной опухоли также подавлялся, средний размер опухолей был больше, чем в голой мыши, что подразумевает роль В-клеток в апоптозе модифицированных опухолевых клеток. Для распознавания эффекторных клеток мы провели иммуногистологический анализ опухолевых тканей. Родительские и tag7-модифицированные опухолевые клетки (4SX) инъецировали в сингенную мыши подкожно, через 3 недели опухолевые ткани выделили и срезы окрасили антителами анти-CD45R/B220, -Gr-1, -CD8a, -CD4, -CD11b(Mac-l). Мы не наблюдали значительной разницы в опухолевых инфильтруемых гранулоцитах, макрофагах или клетках CD4+,CD8+ (данные не показаны) в родительских и модифицированных опухолях. Однако опухоли, продуцирующие tag7, были в большой степени инфильтрованы клетками CD45R+ (фиг.16А), т.е. предпочтительно В-лимфоцитами. Напротив, в родительских опухолях CD45R+ положительные клетки практически отсутствуют (фиг.16В). Наиболее интенсивная инфильтрация наблюдается на границах опухоли и в областях, где сконцентрированы кровеносные сосуды.
Пример 10: Индуцирование защитного иммунитета путем временной экспрессии tag7 в клетках М3 мышиной меланомы (профилактическая модель)
М3-клетки мышиной меланомы были временно трансфецированы конструкцией, экспрессирующей tag7 при промоторе CMV. Через день после трансфекции клетки собрали и имплантировали подкожно в сингенную мышь DBA/2. Через неделю после имплантации трансфецированных опухолевых клеток животных заразили родительскими М3-клетками в местах, не совпадающих с местами подкожной инъекции. У тех животных, которым имплантировали опухолевые клетки, трансфецированные конструкцией, экспрессирующей tag7, скорость роста родительских М3-клеток уменьшалась. Таблица 3 показывает размер опухоли у животных, которые были вакцинированы М3-клетками, трансфецированными как GM-CSF, так и tag7 (при нетрансфецированных М3-клетках в качестве контроля; количество клеток, использованных для вакцинации, дано в таблице), и затем заражены 106 родительских М3-меланомных клеток. Размер опухолей, образуемых родительскими М3-меланомньми клетками, отслеживали; он приведен в таблице, в мм3. Опыт показал, что временная экспрессия tag7 ингибирует развитие этих опухолей по меньшей мере столь же эффективно, как GM-CSF.
Фиг.17 показывает степень выживания мыши DBA/2 в этом опыте (незакрашенные кружочки: 500,000 М3-клеток на мышь: незакрашенные квадраты: 500,000 GM-CSF-трансфецированных М3-клеток на мышь; закрашенные квадраты: 500,000 tag7-трансфецированных М3-клеток на мышь; закрашенные кружочки: 106 родительских М3-клеток на мышь).
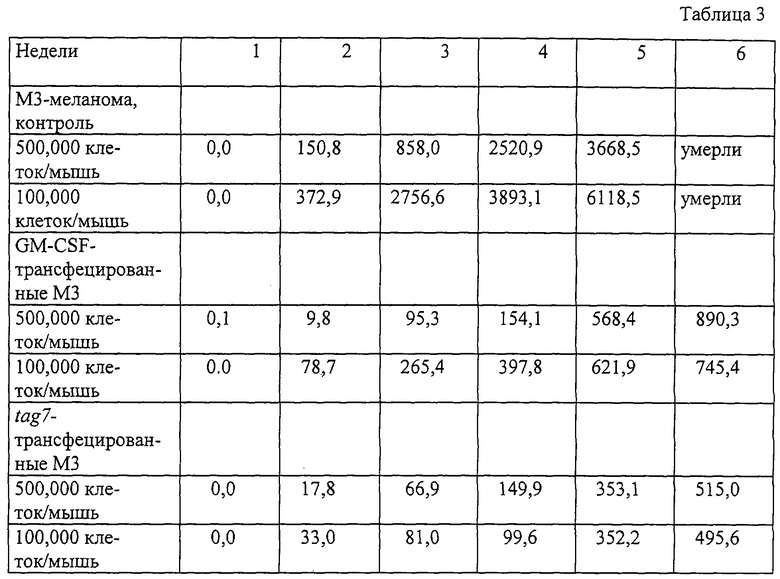
Пример 11: Распознавание гомолога tag7 в человеческих тканях
Чтобы определить, экспрессируется ли в человеке гомолог мышиного гена tag7, коммерчески доступный блот, содержащий полиА + РНК из различных человеческих тканей обработали 32Р-меченными пробами кДНК мышиного tag7. Как показано на фиг.18, меченая проба tag7 гибридизовалась с РНК из нескольких человеческих тканей. Наиболее заметная, сильная гибридизация наблюдалась с полосой 1 т.п.н. из человеческого костного мозга, полосой 2,00 т.п.н. из человеческой эмбриональной печени и полосой 2,1 т.п.н. из человеческих лимфоузлов. Несколько более слабая гибридизация наблюдалась с полосой 2,1 т.п.н. из человеческой селезенки, полосами 0,9 т.п.н. из человеческой эмбриональной печени и полосой 1,0 т.п.н. из периферийных кровяных лейкоцитов. РНК, выделенная из тимуса человека, не гибридизовалась с помощью tag7-меченной пробы, используемой в текущих опытах. Все вместе взятые, эти результаты означают, что человеческий гомолог tag7 существует и экспрессируется в разных человеческих тканях.
Пример 12: Выделение и секвенирование человеческого tag7
Нозерн-блоттинг человеческих РНК с использованием мышиного tag7 в качестве пробы показал высокую экспрессию человеческого гомолога в костном мозге (ВМ) и низкую экспрессию в клетках периферийной крови (PBLs) (Пример 11).
Человеческую ВМ-библиотеку (Clontech, Human Bone Marrow 5'-STRETCH-PLUS cDNA catno. HL5005b) подвергли скринингу в соответствии с протоколом Contech, использовав в качестве пробы кДНК мышиного tag7. Проба была вырезана из плазмиды рХ как фрагмент Xhol-EcoRI, длиной приблизительно в 700 нп. Очищенную полосу пометили Р32 с помощью набора для эффективного мечения ("high prime labeling kit", Boehringer Mannheim, cat.no. 1585584).
1×10 фаговых бляшек были подвергнуты скринингу. 25 позитивных клонов можно было зарегистрировать в первом скрининге, и соответствующие бляшки очистили и проанализировали.
ДНК из указанных позитивных клонов получили с помощью набора Qiagen lambda kit#12549. Вставки вырезали путем рестрикционного ферментного расщепления по NotI и клонировали в вектор pGEM11f+. 3 клона содержали человеческий tag7 (клоны #6, #12 и #13) и один из них содержал полную кДНК (# 13). Полный клон #13 содержал вторую кДНК выше кДНК tag 7, кодирующую MRP-8, белок, родственный фактору ингибирования миграции макрофага (MIF), Асс.по.Х 006234. Этот белок является белком, связывающим кальций в макрофагах. Все три клона были секвенированы. Четвертый клон (#10) был секвенирован прямо из лямбда ДНК с помощью амплифицирования вставки путем PCR перед секвенированием.
В полученной последовательности имелась открытая рамка считывания, как представлено в SEQ ID NO:3, кодирующая tag7 согласно аминокислотной последовательности SEQ ID NO:4.
Человеческий полипептид tag7 имеет на своем N-конце, подобно мышиному tag7 гидрофобный участок из 20 аминокислот, который представляет собой предполагаемую сигнальную последовательность. Для опытного в данной области специалиста обычным делом является определение наличия или длины такой лидерной последовательности, соответственно путем секвенирования N-конца зрелого (натурального или рекомбинантного) полипептида tag7.
Выравнивание последовательности человеческой и мышиной кДНК (с применением кластерного метода с использованием таблицы весовых вкладов остатков) подтвердило 77,2% подобия. На белковом уровне было определено 66,1% подобия с применением кластерного метода с использованием таблицы весовых вкладов остатков.
Пример 13: Экспрессия рекомбинантного мышиного и человеческого tag7
(а) Экспрессия рекомбинантного мышиного tag7 в E.coli
Для первой серии опытов использовали плазмиду экспрессии, обозначенную tag7pDH13/12, которая содержит лидерную последовательность STII и phoA-промотор (получена из pDH13), и плазмиду экспрессии, обозначенную tag7pDH15/l с лидерной последовательностью STII и Тгр-промотором (получена из pDH15).
Чтобы получить плазмиды, лидерная последовательность STII была слита с мышиным tag7 ("mutag 7") в процессе реакции SOE (удлинения с перекрыванием, "splice overlap extension"; см. статью Norton R.M., Hunt H.D., No S.N., Pullen J.K. и Pease L.R. в журнале Gene 77:61-68 (1989)), получили XhoI-PstI-фрагмент и клонировали в векторы pDH13 (патент US 5710027) и pDH15 соответственно. pDH15 идентичен pDH13, только pho-промотор заменен на синтетический Тгр-промотор. Тгр-промотор индуцируется индолакриловой кислотой. STII-лидерная последовательность заменила предполагаемую лидерную последовательность mutag7, как первые аминокислотные остатки mutag7 были выбраны SEWR (начиная с аминокислоты 24 в SEQ ID NO:2). ДНК-фрагменты, которые подлежали слиянию, были получены в двух отдельных PCR-реакциях:
PCR1:
прямой праймер EBI5141: 5'GAAAGGGCCTCGTGATAC3'(SEQ ID NO:8)
обратный праймер АС1=ЕВI8024:
5'CAGGGCCCTCCACTCACTTGCATAGGCATTTGTAGC3'(SEQ ID NO:9)
ДНК-матрица была pDH 13.
PCR2:
прямой праймер: АС2=ЕВI8025
5'GCTACAAATGCCTATGCAAGTGAGTGGAGGGCCCTG3'
(SEQ ID NO: 10),
обратный праймер: ЕВI8203:
5'GGCCGCTAGCCTGCAGTTATCACTCTCGGTAGTGTTCCCAG3'
(SEQ ID NO:11)
ДНК-матрица была рХ-плазмида, несущая mutag7 (SEQ ID NO:l).
PCR-фрагменты разогнали в 1% агарозном геле и очистили с помощью Qiagen Qiaex II. Очищенные PCR-фрагменты были слиты в следующей SOE-реакции:
прямой праймер: EBI5141
обратный праймер: EBI8203
ДНК-матрицы: очищенные PCR1 и РСР2-фрагменты.
Полученный в результате PCR-фрагмент был затем расщеплен Xhol и PstI и клонирован в pDH13 и pDH15, которые были расщеплены такими же фрагментами. pDH13 и pDH15 несут альфа-фрагмент человеческого интерферона в качестве вставки, которую делегировали посредством расщепления XhoI-PstI и заместили SOE-фрагментом mutag7. E.coli DHα5 был трансформирован плазмидами экспрессии. Экстракты E.coli трансформированных бактерий проанализировали на экспрессию tag7 в Вестерн-блотах.
Для следующего опыта использовали плазмиду экспрессии, обозначенную как pGEX-2TmuTag7/клон3, для того чтобы экспрессировать мышиный tag7 как GST-tag7 фьюжн-белок.
Для того чтобы добавить сайты рестрикции BamHl к 5'-концу и EcoRII к 3'-концу кДНК mutag7 в процессе PCR-реакции были использованы синтетические олигонуклеотиды. PCR-реакцию провели с рХ-плазмидой в качестве ДНК-матрицы и праймерами:
EBI8471
(содержит BamHI-сайт) и EBI8472 (содержит EcoRI-сайт)
EBI8471: 5'GGCGGATCCGAGTGGAGGGCCCTGCCATCC3'
(SEQ ID NO:12)
EBI8472: 5'GGCGAATTCTTATCACTCTCGGTAGTGTTCS'
(SEQ ID NO:13)
PCR-фрагмент очистили и расщепили по BamHI и EcoRI и клонировали в те же сайты pGEX-2T (Pharmacia). Структуру праймеров выбрали так, чтобы первыми остатками mutag7, слитого с GST-белком, были Glu Trp Arg Ala (начиная с аминокислоты в положении 25), таким образом исключая предполагаемую лидерную последовательность. E.coli w3110 был трансформирован плазмидой pGEX-2TmuTag7/клон3, и экспрессию CST-мышиного tag7 проверили с помощью Вестерн-блоттинга (фиг.19). Вестерн-блоттинг показывает полосу приблизительно 44.5 кД (26 кД GST плюс 18,5 кД tag7) в линии а; линия b в качестве позитивного контроля показывает мышиный tag7 из супернатантов CSML-0-клеток при молекулярной массе 18,5 кД.
b) Экспрессия рекомбинантного мышиного tag7 в СНО-клетках
Для этих опытов использовали плазмиду экспрессии, обозначенную tag7+LpAD-CMVl, с эндогенной лидерной последовательностью mutag7 (полученной из pAD-CMVl). Для того чтобы добавить сайты рестрикции BglII к 5'-концу и NheI к 3'-концу кДНК mutag7 в процессе PCR-реакции были использованы синтетические олигонуклеотиды. PCR-реакцию провели с рХ-плазмидой в качестве ДНК-матрицы и праймерами:
EBI8233
(содержит BglII-сайт) и EBI8203 (содержит NheI-сайт)
EBI8233: 5'GGCCAGATCTCGTCCAGCATGTTGTTTGCCTGTGCT3'
(SEQ ID NO:14)
PCR-фрагмент очистили и расщепили BglII и NheI и клонировали в pAD-CMVl (ЕР393.438), который расщепили BglII-XbaI.
Были получены устойчивые В16- и С26-клеточные линии, экспрессирующие мышиный tag7. Плазмиду tag7+L pAD-CMVl линеаризовали с помощью FspI и котрансфецировали с плазмидой pCI-neo (Promega) в СНО-клетки. Клоны, экспрессирующие мышиный tag7, распознали с помощью Вестерн-блоттинга клеточных культуральных супернатантов.
Для следующих опытов использовали плазмиду экспрессии, обозначенную Tag7+IgL pAD-CMVl/2, с синтетической лидерной последовательностью легкой цепи иммуноглобулина (получена из pAD-CMVl).
Синтетическая лидерная последовательность легкой цепи иммуноглобулина INРЕР4+лидерная плазмида была слита с mutag7 в SOE-реакции ("splice overlapping extension"), был получен BglII-NheI-фрагмент и клонирован в вектор pAD-CMVl (WO 93/11257). Лидерная последовательность иммуноглобулина заместила предполагаемую лидерную последовательность mutag7, и как первые аминокислотные остатки mutag7 были выбраны EWRA. ДНК-фрагменты, подлежащие слиянию, были получены в двух отдельных PCR-реакциях:
PCR1:
прямой праймер ЕВI8143: 5'GGCACCAAAATCAACGGGAC3'
(SEQ ID NO:15)
обратный праймер
EBI8201: 5'CAGGGCCCTCCACTCACCATCGTGCACCTGGGC3'
(SEQ ID NO:16)
ДНК-матрица была INPEP4+лидерная последовательность.
PCR2:
прямой праймер:
EBI8202:5'CCCAGGTGCACGATGTGAGTGGAGGGCCCTG3'
(SEQ ID NO:17),
обратный праймер: EBI8203:
5'GGCCGCTAGCCTGCAGTTATCACTCTCGGTAGTGTTCCCAG3'
(SEQ ID NO:18)
ДНК-матрица была рХ-плазмида, несущая mutag7.
PCR-фрагменты разогнали в 1% агарозном геле и очистили с помощью Qiagen Qiaex II.
Очищенные PCR-фрагменты были слиты в следующей SOE-реакции:
прямой праймер: EBI8143
обратный праймер: EBI8203
ДНК-матрицы: очищенные PCR1и РСР2-фрагменты.
Полученный в результате SOE-фрагмент был затем расщеплен BglII и NheI и клонирован в pAD-CMVl, который был расщеплен BamHI и NheI.
с) Экспрессия рекомбинантного мышиного tag7 в СНО-клетках
Для этих опытов использовали плазмиду экспрессии, обозначенную hutag7pAD-CMVl, с эндогенной лидерной последовательностью hutag7 (полученной из pAD-CMVl).
Для того чтобы добавить сайты рестрикции HindIII к 5'-концу и EcoRI к 3'-концу кДНК hutag7 в процессе PCR-реакции были использованы синтетические олигонуклеотиды. PCR-реакцию провели с pGem hu Bone Marrow Tag7#13/l-плaзмидoй в качестве ДНК-матрицы и праймерами: EBI8748 (содержит HindIII-сайт) и EBI8749 (содержит EcoRI-сайт)
EBI8748: 5'GGCCAAGCTTCCACCATGTCCCGCCGCTCTATGCT3'
(SEQ ID NO:19)
EBI8749: 5'GGCCGAATTCTTATCAGGGGGAGCGGTAGTGTGGT3'
(SEQ ID NO:20)
PCR-фрагмент очистили и расщепили HindIII и EcoRI и клонировали в те же сайты INPEP4 и pAD-CMVl соответственно.
Пример 14: Получение антисывороток анти-mutag7 и anmu-hutag7
Были синтезированы два разных пептида, полученных из последовательности mutag7 (мышиного tag7), и два разных пептида, полученных из последовательности hutag7 (человеческого tag7):
mutag7-пeптиды
пептид nemu-A: RSEWRALPSECSSRLGHPY (SEQ ID NO:21)
пептид nemu-B: GEDGHVYEGRGWNIKGDHTGCY (SEQ ID NO:22) Для иммунизации пептиды смешали (1:1).
hutag-пептиды
пептид nehu-A: AQETEDPACCSPIVPRNEWKALASEY (SEQ ID NO:23)
пептид nehu-B: GEDGLVYEGRGWNFTGCY (SEQ ID NO:24)
Для иммунизации пептиды смешали (1:1).
Кролики были иммунизированы подкожным введением в 1-й и 21-й дни 500 мкг пептидной смеси. На 42-й день внутримышечно им ввели 250 и на 49-й день 200 мкг пептидной смеси. На 50-й и 51-й дни внутривенно им ввели по 250 мкг пептидной смеси. Кровь брали на 63-й, 70-й, 77-й и 84-й дни.
Аффинная очистка антисывороток анти-mutag и анти-hutag.
Аффинные колонки приготовили путем связывания пептидных смесей mutag7 и hutag7 с сепаративньми колонками Affi-Gel 10 (Biorad) в условиях безводного связывания. По 20 мл каждой поликлональной антисыворотки очистили на соответствующей колонке, причем промывочный буфер представлял собой 0,02 М натрий-фосфат, а элюционный буфер - 0,1 М глицин, рН 2,5. Аффинно-очищенные антисыворотки были подвергнуты диализу в PBS, отфильтрованы в стерильных условиях и хранились при -20°С.
Пример 15: Экспрессия Tag7 в нормальных и опухолевых человеческих тканях.
Провели иммуногистохимический анализ замороженных срезов ткани в соответствии с недавней публикацией (см. Karl-Heinz Heider, Jobst  Petra Skroch-Angel,
Petra Skroch-Angel,  Hans Peter Vollmers, Peter Herrlich и Helmut Ponta "Differential expression of CD44 splice variants in intestinal and diffuse type human gastric adenocarcinomas and normal gastric mucosa", Cancer Res. 53, 4197-4203, 1993). Кратко говоря, замороженные срезы инкубируют с первичным антителом (антисывороткой анти-Tag-7, приготовленной, как указано в примере 14, в количестве 10 мкг/мл, или контрольной сыворотки Ig кролика в количестве 10 мкг/мл) в течение 1 часа при комнатной температуре. Связанные антитела определяли, используя комплексную систему авидин-биотин-пероксидаза (DAKO, Дания). Оценку окрашивания осуществили с помощью Zeiss Axioscop, подразделив интенсивность окрашивания на - (негативную), + (слабо позитивную) и ++(от умеренной до позитивной в высокой степени).
Hans Peter Vollmers, Peter Herrlich и Helmut Ponta "Differential expression of CD44 splice variants in intestinal and diffuse type human gastric adenocarcinomas and normal gastric mucosa", Cancer Res. 53, 4197-4203, 1993). Кратко говоря, замороженные срезы инкубируют с первичным антителом (антисывороткой анти-Tag-7, приготовленной, как указано в примере 14, в количестве 10 мкг/мл, или контрольной сыворотки Ig кролика в количестве 10 мкг/мл) в течение 1 часа при комнатной температуре. Связанные антитела определяли, используя комплексную систему авидин-биотин-пероксидаза (DAKO, Дания). Оценку окрашивания осуществили с помощью Zeiss Axioscop, подразделив интенсивность окрашивания на - (негативную), + (слабо позитивную) и ++(от умеренной до позитивной в высокой степени).
Экспрессию Tag-7-антигена в нормальных и опухолевых человеческих тканях изучили с помощью иммуногистохомических методов. В нормальных тканях позитивное окрашивание было зарегистрировано во всех исследуемых образцах. В мозжечке и коре головного мозга отдельные клетки слабо реагировали по отношению к антителу анти-Tag-7. Лимфатические клетки в двенадцатиперстной кишке, селезенке и лимфоузлах были позитивны от умеренной до сильной степени. Окрашивание главным образом ограничивалось клеточной мембраной и внутриклеточным пространством. В легком как альвеолярный эпителий, так и эндотелий сосудов сильно реагировали по отношению к антителу анти-Tag-7. Результаты суммированы в таблице 4.

Во всех изученных типах опухолей наблюдалось позитивное окрашивание с антителом анти-Tag-7 (таблица 5). В большинстве случаев окрашивание было умеренным до сильного и главным образом ограничивалось опухолевой стромой при диффузном характере окрашивания (см. фиг.1). В карциноме почечных клеток также в высокой степени реагировали с антителом анти-Tag-7 инфильтрующие лимфоциты и сосудистый эндотелий. Опухолевые клетки в основном не обнаруживают реактивности по отношению к анти-Tag-7, за исключением аденокарциномы толстой кишки, где часть опухолевых клеток были позитивны (не показано).

На фиг.20 показаны данные иммуногистохимического анализа аденокарциномы легкого с антителами анти-Tag-7. Стрелки обозначают позитивное окрашивание опухолевой стромы.
Общее обсуждение
Чтобы изучить гены, которые селективно экспрессируются в опухолях с высокой способностью к метастазированию, были выбраны две опухоли одинакового происхождения, но различающиеся по их метастатическим свойствам. В предварительных исследования были выделены два субштамма рака VMR с высокой способностью к метастазированию (который имеет широкий спектр метастазов по органам и частоту метастазов в печень не более 5%) (см. статью Senin V.M., в журнале Vestnik. Akad. Med. Nauk. SSSR 0(5):85-9l (1984)): линия VMR-0 (частота метастазирования в печень около 0%) и линия VMR-L (частота метастазирования в печень около 100%). В отличие от опухолевых линий CSML-0 и CSML-100 ни одна из опухолей VMR не экспрессирует предварительно клонированного mts-1-гена, который транскрибируется большинством опухолей (см. статью Grigorian M.S. и др. в журнале Genetika (USSR) 25(6):993-1000 (1989)), что делает полезным поиск других генов, которые участвуют в процессе метастазирования.
Если рассматривать опухоли, полученные в соответствии с ранее предложенными моделями (см. статью Fidler I.J. и Hart I.R. в журнале Science 217:998-1001 (1982)), очевидно, что опухоль VMR-L можно получить из метастатической субпопуляции клеток, которые имеются в первичной опухоли VMR в очень малом количестве и которые имеют очень короткий срок жизни (см. у Kerbell K.S. в Adv. Cancer Res. 55:87-131 (1990)).
Этот вероятное одинаковое происхождение двух опухолей, исследованных здесь, предлагает возможность использовать методику "выявления различий в РНК" для распознавания генов, связанных с процессом метастазирования. В общем, генетические различия между опухолями VMR-0 и VMR-L незначительны, до определенной степени подтверждая их общее происхождение. Однако с помощью такого подхода возможно выделить несколько фрагментов ДНК соответственно различно экспрессируемым РНК в опухолях VMR-0 и VMR-L. Анализ выделенных фрагментов с помощью Нозерн-гибридизации с тотальной РНК из этих опухолей позволяет отнести большинство фрагментов к классу известных повторяющихся последовательностей (IAR-элемент и т.д.). Но несколько полученных фрагментов не могут быть так определены и поэтому представляют безусловный интерес.
Один из этих фрагментов, выделенный и охарактеризованный здесь, представляет собой ген tag7, который показывает очень высокий уровень транскрипции в опухоли VMR-L, метастазирующей в печень (см. фиг.2). Анализ последовательности обнаружил, что PCR-фрагмент был фланкирован на обеих сторонах с помощью 5'-концевого праймера 5'-AATCGGGCTG-3' (SEQ ID NO:4). Когда проанализировали нуклеотидную последовательность клона кДНК из тотальной библиотеки кДНК VMR-L, последовательность, аналогичная этому праймеру по длине в восемь пар оснований, была найдена на расстоянии в 52 нуклеотида от 3'полиА + хвоста клонированной кДНК. Вероятно, что в методике "выявления различий в РНК" можно использовать стандартный олиго-dT-праймер вместо 3'-концевого праймера и только 5'-концевые праймеры могут варьироваться; такой подход, возможно, приведет к уменьшению количества получаемых фрагментов, но одновременно увеличит разрешающую способность геля.
Те ошибки, которые возникают при проведении многочисленных циклов PCR, могут быть отнесены к недостаткам методики. При сравнении нуклеотидной последовательности фрагмента, полученного в реакции PCR, и кДНК клона tag7 было обнаружено, что 44 нуклеотида на 5'-конце амплифицированного фрагмента не совпадают с аналогичным сегментом последовательности клона кДНК.
Компьютерный анализ полноразмерной последовательности кДНК мышиного tag7 показал две открытые рамки считывания (OFR), соответствующие полипептидам длиной примерно 182 аминокислоты и 91 аминокислота. Оба старта трансляции в разной степени соответствовали усредненной последовательности Козака (см. статью Kozak M. в журнале J. Cell. Biol. 108:229-241 (1989)). Однако в случае более длинной ORF количество нуклеотидов, совпадающих с трансляционной консенсусной последовательностью, больше, чем при более короткой ORF, и, по-видимому, наличие первого метионина свидетельствовало о присутствии сайта инициации трансляции. Кроме того, метионин в позции от +37 до +39 нп в полноразмерной кДНК находится ближе к 5' -концу и поэтому является более благоприятным для инициации трансляции (см. статью Kozak M. в журнале J.Cell.Biol. 108:229-241 (1989)). Тем не менее, трансляция функционального полипептида tag7 может быть также инициирована со второго метионина.
Молекулярный вес предсказанного полипептида tag7 (182 аминокислотных остатка; SEQ ID NO:1) составляет около 20 кД. Чтобы найти гомологию с ранее описанными последовательностями, нуклеотидную последовательность tag7 и предсказанную аминокислотную последовательность ее продукта проанализировали в базах данных GENE и SWISSPROT с помощью программ поиска гомологии FASTA (Heidelberg, Германия) и BLAST (Washington, DC, США). Не было найдено существенных гомологий с ранее известными последовательностями, за исключением незначительной гомологии гидрофобного сегмента (между 2 и 12 аминокислотными остатками) на N-конце предсказанной последовательности полипептида tag7 с β-цепочкой человеческого Т-клеточного рецептора. Эта гидрофобная область также гомологична сигнальным последовательностям некоторых белков (например, предшественников кальретикулина, остеокальцина и внеклеточного глобина III). Наличие гидрофобной области на N-конце предсказанного полипептида tag7 позволяет предположить, что этот полипептид может иметь трансмембранную локализацию или может быть секретирован.
Экспрессия tag7, как было обнаружено, не коррелируется со органоспецифичностью метастазирования; ген tag7 транскрибировался не только в клетках VMR-L-опухоли, метастазирующей в печень, но также и в клетках VMR-Ov-опухоли, метастазирующей в яичник. Среди нормальных органов максимальный уровень экспрессии гена tag7 наблюдался в клетках тех органов, в которых в относительно высокой пропорции имелись клетки базальной мембраны. Эта корреляция может не быть полным соответствием, но может отражать роль, которую играет базальная мембрана в процессе метастазирования (см. статью Stetler-Stevenson W.G. и др. в журнале Ann.Rev.Cell Biol. 9:541-573 (1997)). Интересно, что уровень экспрессии гена tag7 в метастазирующих опухолях много выше, чем в таких же клетках в культуре. Этот результат противоречит недавно опубликованным результатам исследования экспрессии генов в превичных и культивированных раковых клетках (см. статью Zhang L. и др. в журнале Science 276:1268-1271 (1997)) и может представлять реакцию на взаимодействие опухолевых клеток с клетками-хозяевами базальной мембраны, например, в процессе инвазии базальной мембраны метастазными опухолевыми клетками. Участие клеток стромы в регулировании экспрессии tag7, которое имеет место, например, в экспрессии гена стромезилина-3 (см. статью Basset P. и др. Nature 348:699-704 (1990)), также не может быть исключено.
Также интересным является наблюдение, что уровень транскрипции гена tag7 в несколько раз ниже в клетках опухоли CSML-0, чем в клетках VMR-L, тогда как при переводе штаммов тех же опухолей в культуру картина заметно меняется. В частности, при культивировании этих клеток уровень транскрипции гена tag7 в клетках VMR-L уменьшается, в то время как он увеличивается в клетках CSML-0 и в несколько раз превышает уровень экспрессии tag7 в культивированных клетках VMR-L. Несколько более низкий уровень транскрипции tag7 наблюдался в культивированных клетках МТ1 ТС1 в сравнениии с культивированными клетками CSML-0. Такое изменение уровня транскрипции гена tag7 в культивированных клетках VMR-L может быть объяснено тем обстоятельством, что эта культура не является моноклональной, и поэтому пропорция субпопуляции метастазирующих клеток в культуре может быть довольно малой (см. статьи Fidler I.J. и Hart I.R. в журнале Science 21:998-1001 (1982); Senin V.M. в журнале Akad.Med.Nauk. SSSR 0(5):85-9l (1984)). С другой стороны, гетерогенность культуры клеток CMSL-0 установлена D.A.Kramer'ом (выяснено при личном контакте). В частности, интересно заметить, что транскрипция гена tag7 наблюдалась в культурах клеток опухолей, имеющих эпителиальное происхождение, и имеет очень низкую (5-10%) способность к метастазированию. Однако из 13 исследованных клеточных линий с различной способностью к метастазированию только в клетках линии VMR-L транскрипция гена tag7 коррелировалась с замеченной способностью этих опухолей метастазировать.
В течение последнего десятилетия были распознаны и клонированы цитокины, относящиеся к TNF. Хорошо известным растворимым цитокином является протеолитически полученная форма TNF-α. Кроме того, были сообщения, что растворимая форма Fas-лиганда продуцировалась активированными периферическими Т-лимфоцитами (см. статью Tanaka M. и др. в журнале EMBOJ. 14: 129-1135 (1995), а клеточная линия мышиной тимомы, как показано, секретирует расворимый лиганд CD40 (см. статью Armitage G. и др. в журнале Eur.J.Immunol. 22:2071-2076 (1992)). Вследствие его структурного подобия с цитокинами, относящимися к TNF, tag7 может быть новьм членом этого растущего семейства.
Хромосомная локализация tag7 открывает возможность рассмотреть роль tag7 in vivo. Цитогенетическая полоса 7А3 генетически связана с нефритом типа волчанки в гибридных моделях -MRL и Ново-зеландской (New Zealand) системного эритоматоза волчанки (SLE) (см. статьи Morel L. и др. в журнале Immunity 1:219-229 (1994); Kono D.H. и др. в журнале Proc. Natl.Acad.Sci. USA 91:10168-10172 (1994)). Хотя нет вероятности, что SLE вовлечен в мутацию с сильными функциональными изменениями, сенсационные опыты с геном могут пролить свет на возможную роль tag7 в этом патогенном процессе.
Интересной является локализация РНК tag7 в основном в лимфоидных органах, принимая во внимание цитотоксический эффект растворимого белка. Очевидно, что главная стадия в каскаде регуляции имеет место на посттранскрипционном уровне. Даже незначительные изменения в продуцировании мРНК tag7 приводят к изменениям общего количества синтезированного белка и его секреции. Поэтому представляется возможным, что могут быть задействованы различия в способности передавать сигнал между растворимой и связанной с мембраной формами лиганда.
Одним из биологических свойств tag7, установленных путем настоящих исследований, является его вовлеченность в вызывание in vitro апоптоза в некоторых клеточных линиях. Апоптоз, или процесс запрограмированной гибели клеток, необходим для нормального развития и гомеостаза организма. Фрагментация ДНК на мультимеры примерно в 180 нп - одно из тех изменений, которые возникают при апоптозе.
Интересно было обнаружить, что опухоли VMR-0, трансфецированные tag7, растут значительно медленнее, чем родительские опухоли VMR-0. Было показано, что некоторое количество цитокинов способно индуцировать отторжение опухоли, если они продуцируются локально опухолевыми клетками после переноса гена (см. статью Hock Н. и др. в журнале Proc.Natl.Acad.Sci. USA 90:2774-2778 (1993)). Известны различные цитокины, активизирующие гетерогенные временные опухолеподавляющие механизмы, но для полного отторжения опухоли всегда требуются клетки CD8+. Недавно был описан сильный противоопухолевый эффект человеческого LT-α, в мышиной экспериментальной модели (см. статью Qin Z. и Blankenstein Т. в журнале Cancer Res. 55:4747-4751 (1995)). В отторжении опухоли задействован комплексный иммунологический механизм, в который вовлечены как Т-клетки, так и В-клетки. Показано, что LT-α менее токсичен для животных, чем TNF, и по меньшей мере столь же эффективен, как TNF (см. статью Qin Z. и др. в журнале Blood 85:2779-2785 (1995)). Не наблюдалось никакого токсического действия клеток, трансфецированных tag7, в мыши, а также какой-либо цитоксической активности клонов, трансфецированного tag7 конструкцией не обнаружено в L929 цитотоксическом анализе. Хотя активность tag7 в кондиционированной среде клонов ниже границы определения, клон, который продуцируется меньшими количествами РНК tag7 развивался быстрее в сингенной мыши. Ингибиторный эффект локально отщепленного tag7 по отношению к развитию опухоли вероятнее всего непрямой и может быть аннулирован инъекцией антител анти-tag7. Это доказано также опытом, в котором был подвергнут мониторингу рост опухоли в родительской линии VMR-0 и совместно инъецированных клетках 4SX. Даже когда инъецируют двойное по отношению к нормальному количество клеток, скорость развития опухоли значительно снижается. Наконец, полное уменьшение опухоли или ремиссия могут быть достигнуты посредством секреции более высоких количеств tag7.
В ингибирование развития опухолевой клетки, опосредованное tag7, как представляется, не вовлечены Т-лимфоциты, т.к. в отсутствие Т-клеток (например, в голой мыши) клетки, продуцирующие tag7, растут медленнее, чем в сингенной мыши (E.V. Korobko, неопубликованные наблюдения). Однако несмотря на эти обнаруженные данные участие других типов клеток в антиопухолевой реакции, индуцированной tag7, не может быть исключено.
Имея полное описание настоящего изобретения, детали которого представлены с помощью иллюстраций и примеров для ясности понимания, обычный специалист в данной области поймет, что можно выполнить модификации или изменения изобретения в широких и равнозначных пределах условий, форм и иных параметров, не влияя на объем изобретения или его специфические воплощения, и имеется в виду, что эти модификации или изменения охватываются объемом зависимых пунктов формулы.
Все публикации, патенты и патентные заявки, упомянутые в данном описании, характеризуют уровень техники в той области, к которой относится изобретение, и привлечены как ссылка в таком объеме, как если бы каждая из этих публикаций, патентов и патентных заявок отдельно была предназначена для включения посредством ссылки на нее.

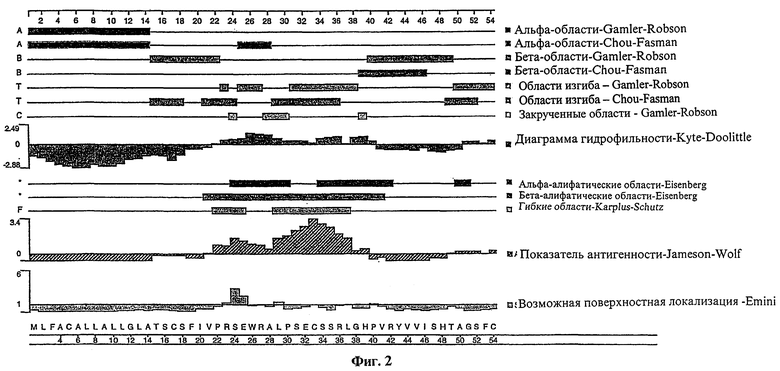
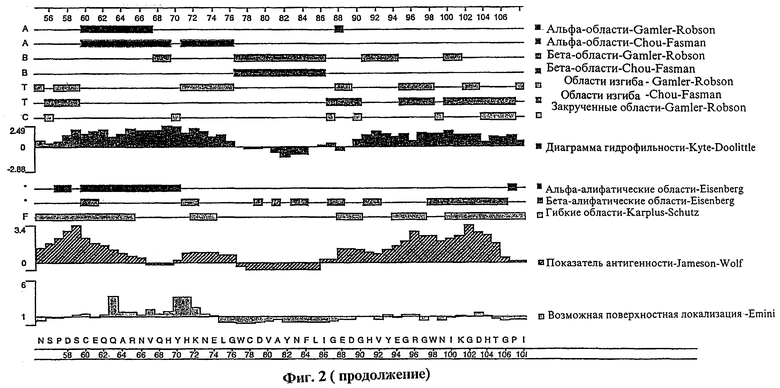
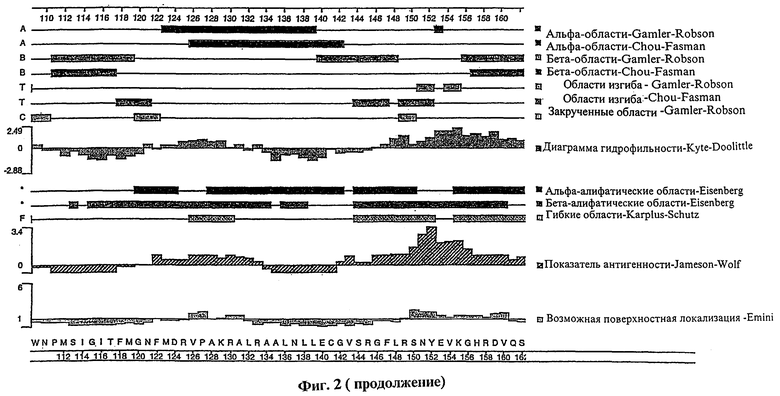
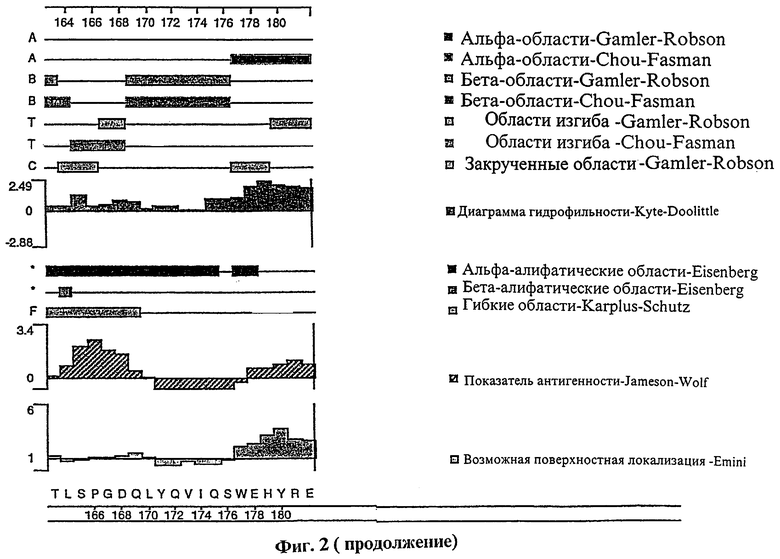




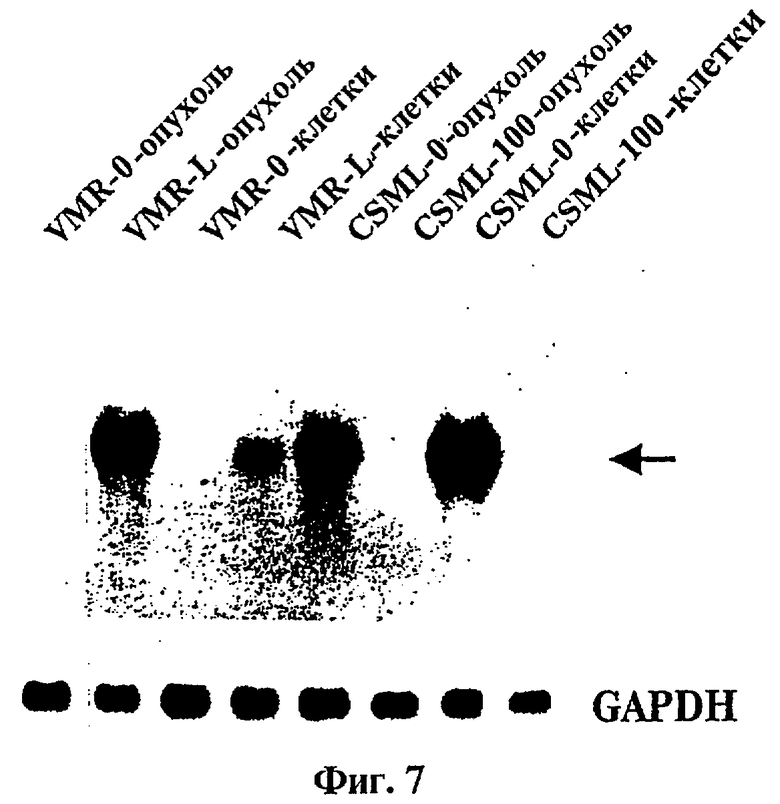
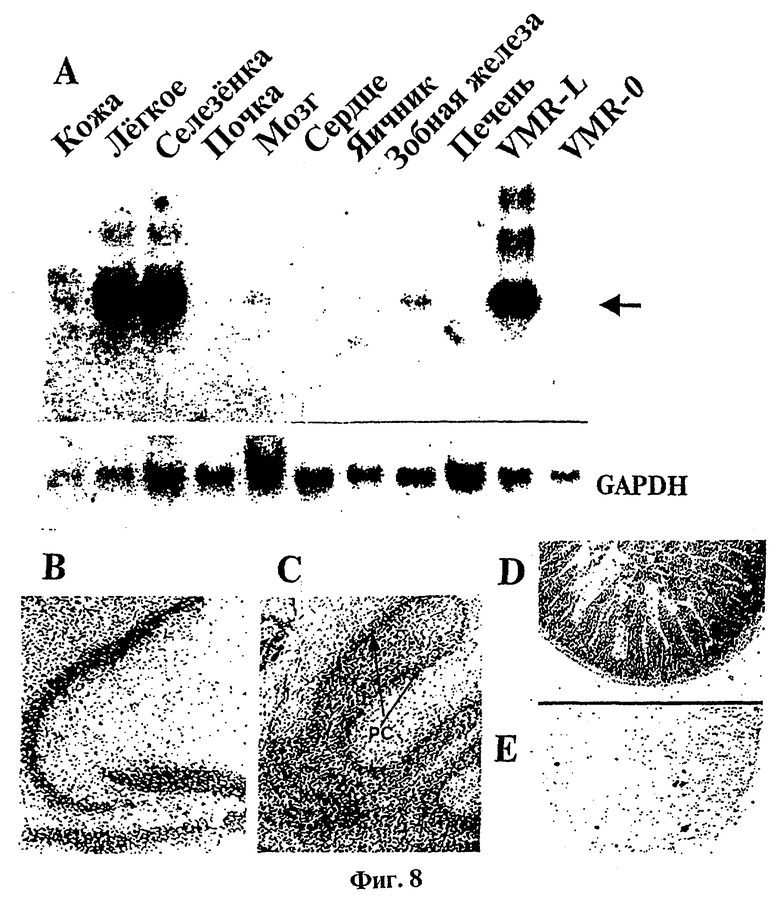

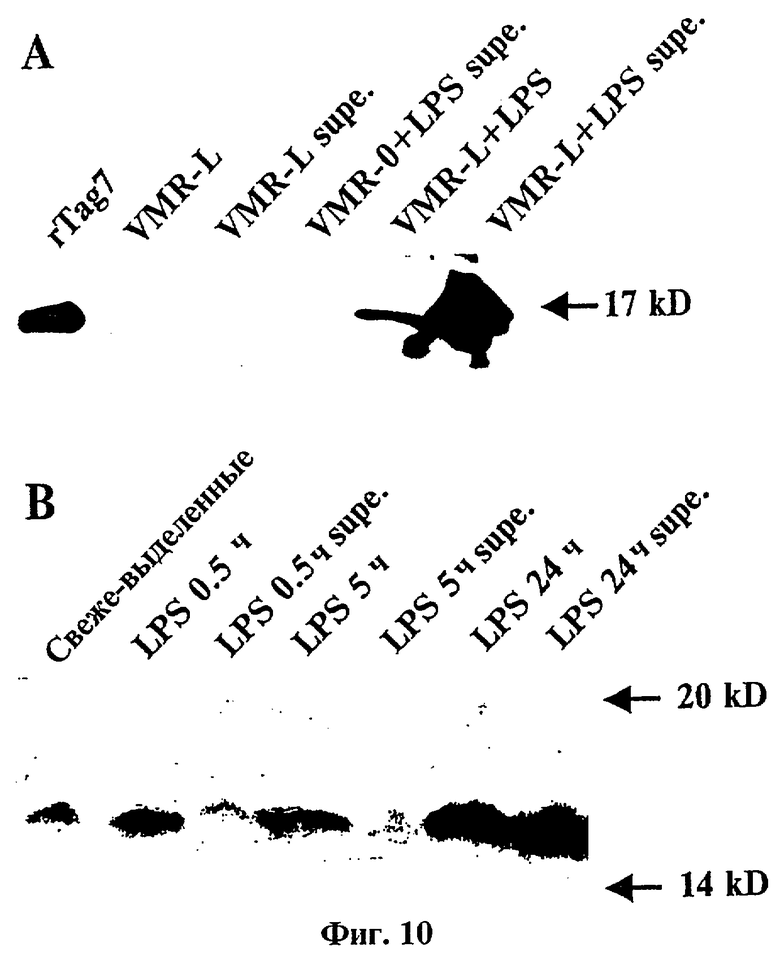
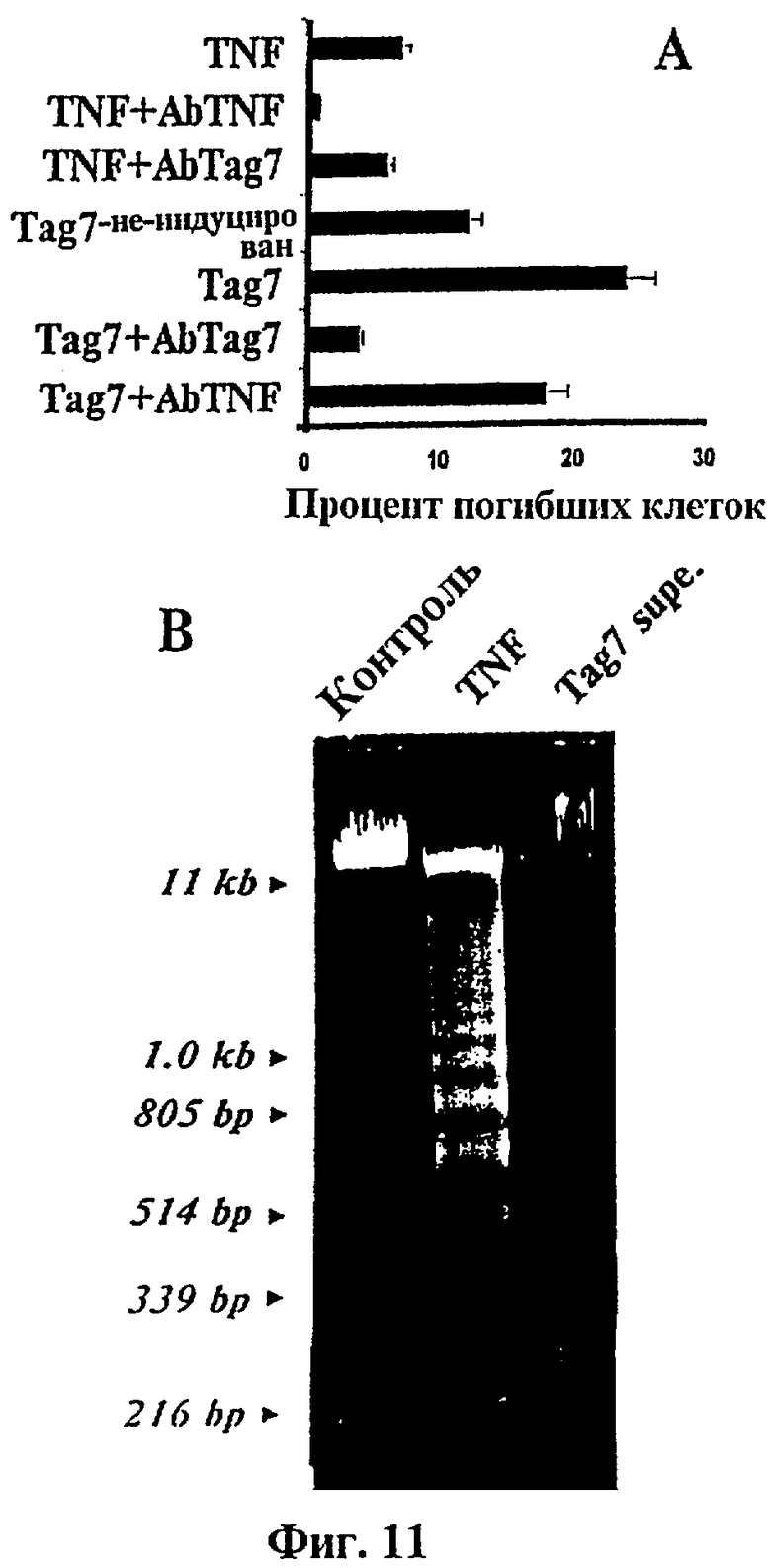
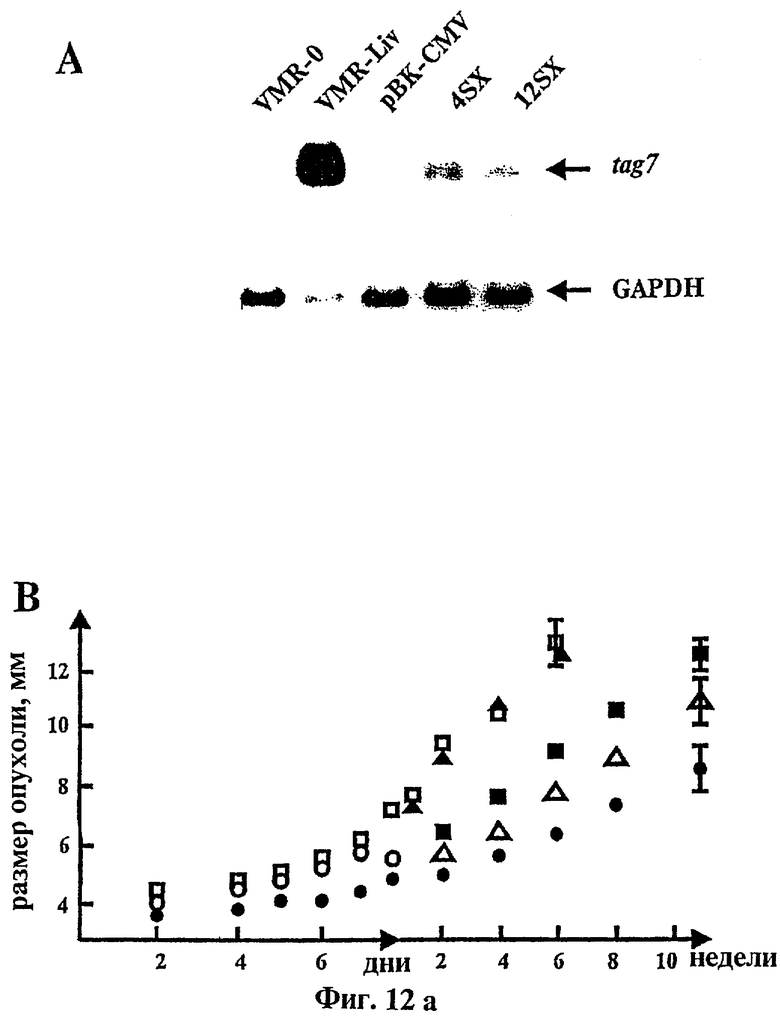
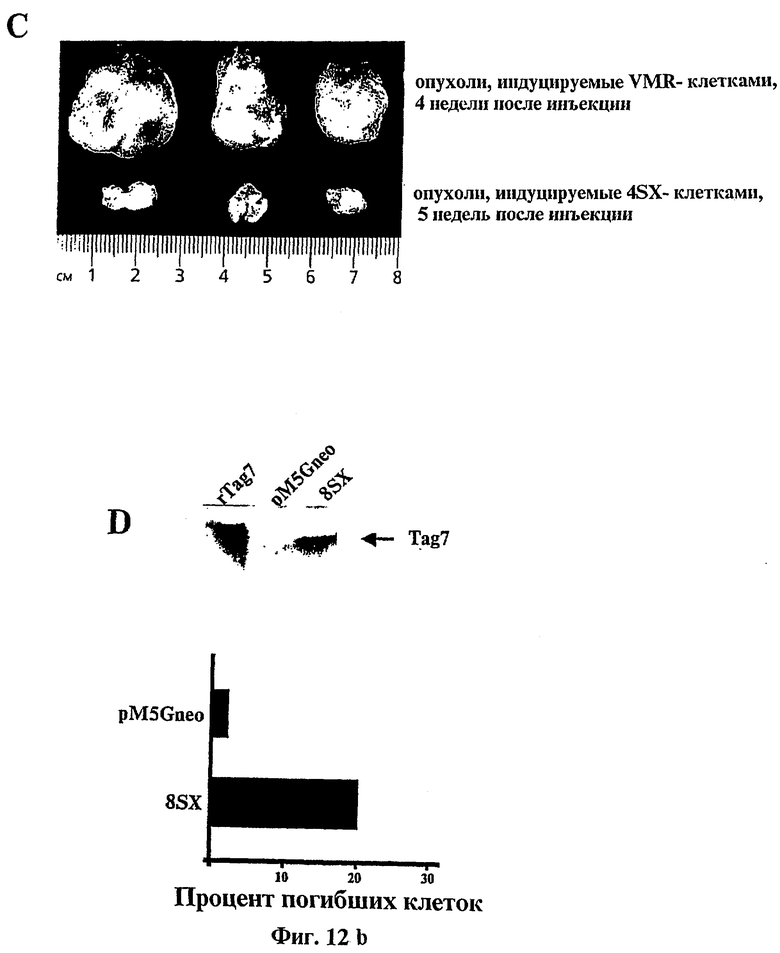
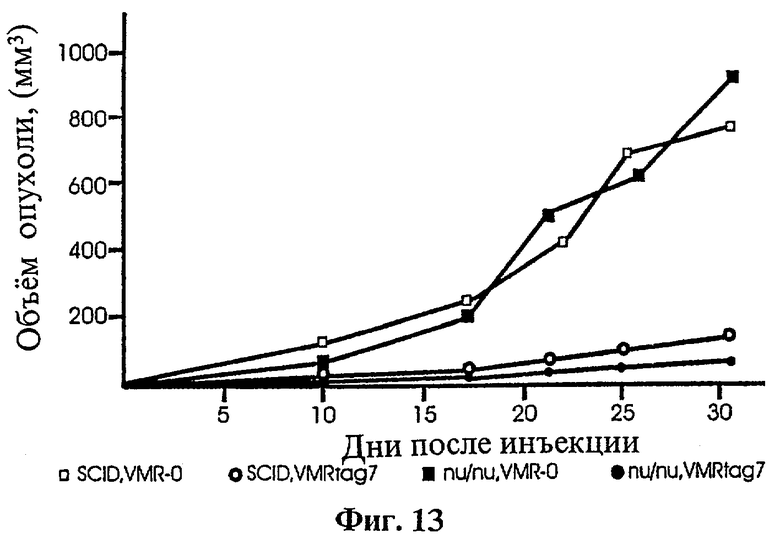
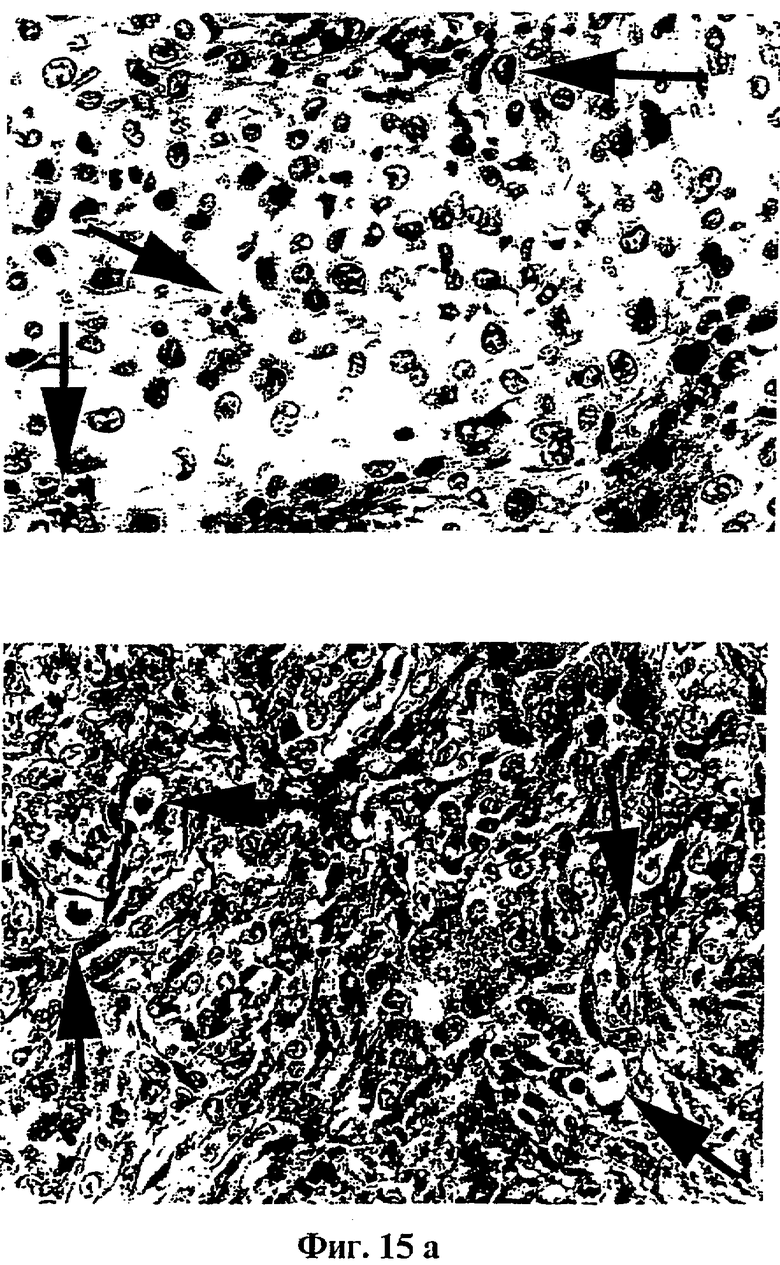
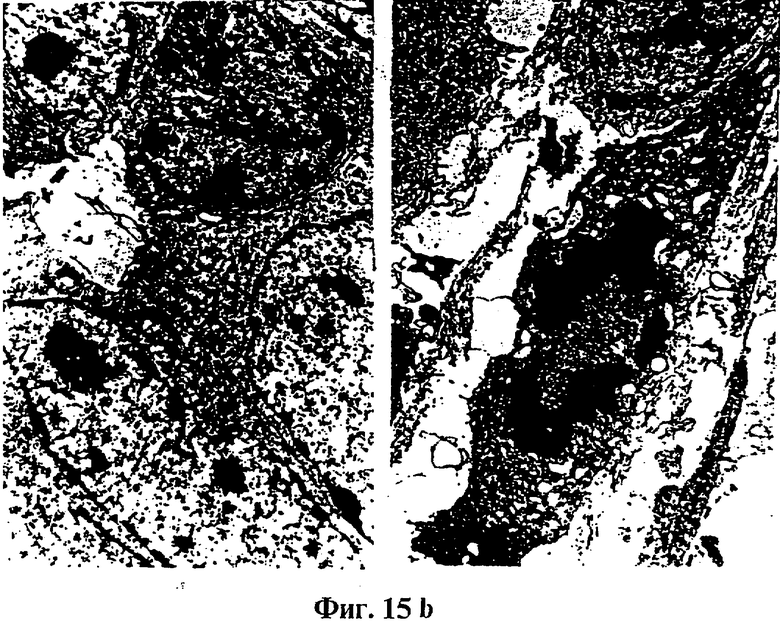
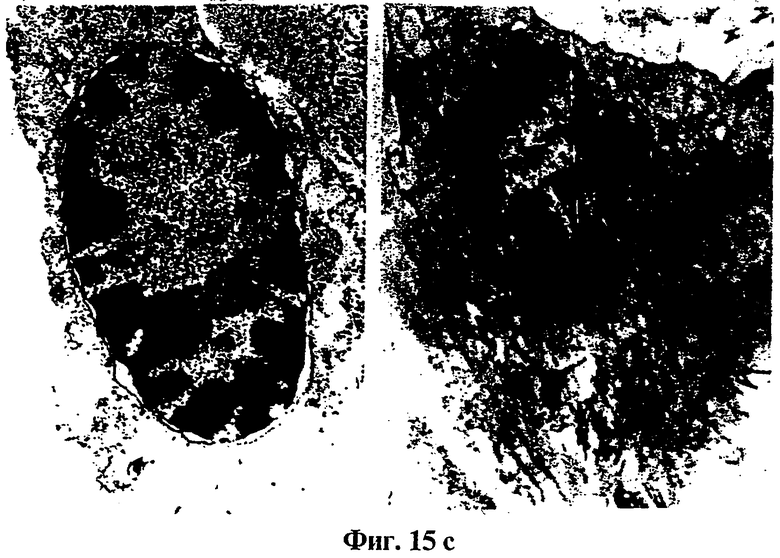
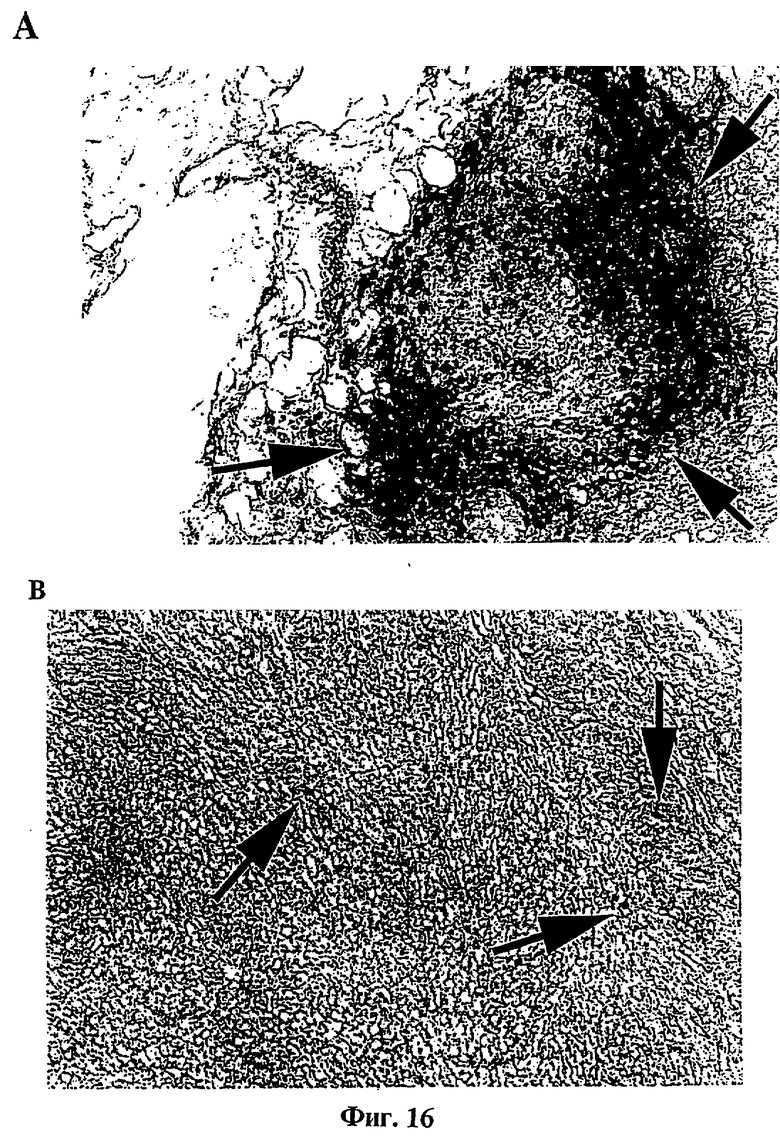
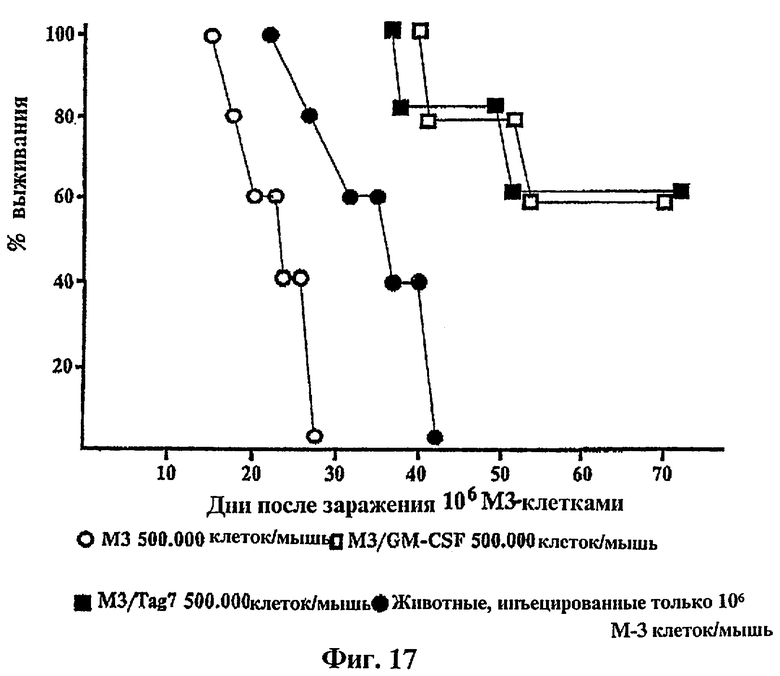
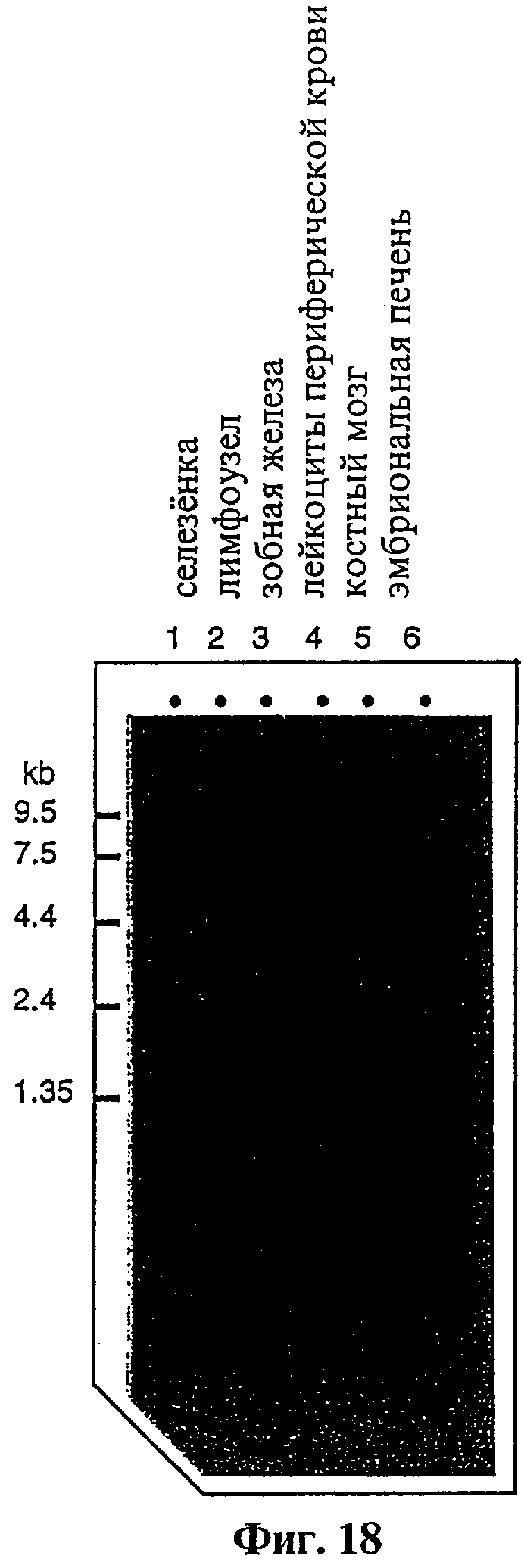
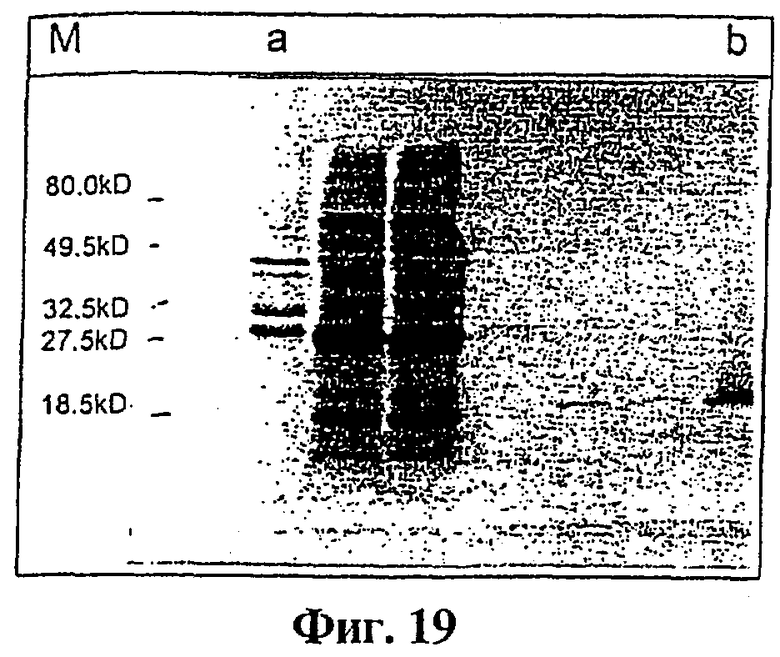

Изобретение относится к биотехнологии и медицине и может быть использовано для получения tag7 полипептида и ингибирования развития опухоли. Полипептид tag7 или изолированную нуклеиновую кислоту, кодирующую полипептид tag7, используют для ингибирования развития опухолей у млекопитающего и лечения рака путем воздействия на клетку. Воздействие нуклеиновой кислотой, в частности, осуществляют с помощью вектора или вириона. Изобретение позволяет разрабатывать лекарственные средства для лечения карциномы, меланомы, саркомы, лейкемии. 6 н. и 29 з.п. ф-лы, 36 ил., 5 табл.
| KUSTIKOVA O.S | |||
| et al | |||
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Способ образования коричневых окрасок на волокне из кашу кубической и подобных производных кашевого ряда | 1922 |
|
SU32A1 |
| ПОРТСИГАР С ПРИСПОСОБЛЕНИЕМ ДЛЯ СКРУЧИВАНИЯ ПАПИРОС | 1922 |
|
SU621A1 |
| MARRA M | |||
| et al | |||
| Прибор для нагревания перетягиваемых бандажей подвижного состава | 1917 |
|
SU15A1 |
| Бесколесный шариковый ход для железнодорожных вагонов | 1917 |
|
SU97A1 |
| SMITH C.A | |||
| et al | |||
| The TNF receptor superfamily of cellular and viral proteins: activation, costimulation, and death, Cell, 1994, v | |||
| Аппарат, предназначенный для летания | 0 |
|
SU76A1 |
| Приспособление для сбрасывания на парашюте почтовых отправлений с летательных аппаратов | 1923 |
|
SU959A1 |

