Область техники, к которой относится изобретение
Данное изобретение относится к областям молекулярной биологии, вирусологии, иммунологии и медицины. Изобретение представляет композицию, содержащую упорядоченную и содержащую повторы матрицу антигена или антигенной детерминанты. Изобретение представляет также способ получения антигена или антигенной детерминанты в виде упорядоченной и содержащей повторы матрицы. Упорядоченный и содержащий повторы антиген или антигенную детерминанту используют для изготовления вакцин для лечения инфекционных болезней, лечения аллергий и в качестве фармакологического препарата для профилактики или лечения рака и выработки определенных аутоспецифических антител.
Уровень техники
Создание вакцин для профилактики инфекционного заболевания оказало самое большое воздействие на здоровье человека по сравнению с любым изобретением в области медицины. Установлено, что с помощью вакцинации в мире предотвращается три миллиона смертей в год (см. статью Hillemann, Nature Medicine, 4:507 (1998)). Наиболее распространенная стратегия вакцинации, использование аттенуированных (т.е. менее вирулентных) патогенов или близкородственных организмов была впервые продемонстрирована в 1796 г. Эдуардом Дженнером, который проводил вакцинацию против натуральной оспы путем введения менее опасного вируса коровьей оспы. Хотя ряд живых ослабленных вирусов (например, вирусы кори, эпидемического паротита, краснухи, ветряной оспы, аденовирус, полиовирус, вирус гриппа) и бактерий (например, бацилла Кальметта-Герена (Calmette-Guerin) (BCG) против туберкулеза) успешно применяют для вакцинации, существует риск развития серьезных осложнений, связанных с реверсией (возвратом) вирулентности и инфекцией "вакцинным" организмом, в частности, у иммунокомпромиссных субъектов.
Специфическое конструирование ослабленных вирусов в настоящее время достигается технологией рекомбинантной ДНК (т.е. генетической инженерией) путем создания делеционных или мутационных вариантов. Например, было показано, что введение инженерного вируса иммунодефицита обезьян (SIV) с делецией в гене nef защищает макак от последующей инфекции патогенным штаммом SIV (см. статью Daniel и соавт., Science, 258: 1938-1941 (1992)). Однако развитие симптомов, подобных синдрому приобретенного иммунодефицита (СПИДа), у животных, которым вводили аттенуированный SIV, вызывает озабоченность, связанную с проблемами безопасности (см. статью Baba и соавт., Science, 267: 1820-1825 (1995)).
В качестве альтернативного подхода возможно использование аттенуированных (ослабленных) вирусов или бактерий как носителей антиген-кодирующих генов патогена, который считают слишком небезопасным для введения в аттенуированной форме (например, вируса иммунодефицита человека (ВИЧ)). После доставки антиген-кодирующего гена хозяину антиген синтезируется in situ. Вирусы коровьей оспы и подобный поксвирусу птиц были использованы в качестве таких носителей для различных генов в предклинических и клинических исследованиях ряда заболеваний (например, см. статью Shen и соавт., Science, 252:440 (1991)). Одним из недостатков данной стратегии вакцинации является то, что в ней не имитируется поверхность вириона, поскольку рекомбинантный белок экспрессируется на поверхности клетки-хозяина. Кроме того, у иммунокомпромиссных субъектов могут развиваться осложнения, что доказано на примере представляющих угрозу для жизни диссеминированных инфекций коровьей оспы (см. статью Redfield N., Eng. J. Med., 316: 673 ((1998)).
Четвертый подход к вакцинации включает применение выделенных компонентов патогена, которые либо очищены из патогена, выращенного in vitro (например, гемагглютинина или нейраминидазы гриппа), либо получены в результате гетерологичной экспрессии единичного вирусного белка (например, поверхностного антигена гепатита В). Например, рекомбинантные мутантные токсины (детоксифицированные) используют для вакцинации против дифтерийного, столбнячного, холерного и коклюшного токсинов (см. монографию Levine и соавт., Вакцины нового поколения (New generation vaccines), 2-е изд., Marcel Dekker, Inc., New York, 1997), а при оценке рекомбинантных белков ВИЧ (gp120 и gp160 полной длины) в качестве средств для индукции нейтрализующих антител против ВИЧ были получены неудовлетворительные результаты (см. статью Connor и соавт., J. Virol., 72: 1552 (1998)). Недавно были получены перспективные результаты с растворимым олигомерным gp160, который может индуцировать ответ CTL (цитотоксических Т-лимфоцитов) и вызывать образование антител с нейтрализующей активностью в отношении изолятов ВИЧ-1 (см. статью Van Cortt и соавт., J. Virol., 71:4319(1997)). Кроме того, могут быть использованы пептидные вакцины, в которых известные В- и Т-клеточные эпитопы антигена связаны с молекулой-носителем, разработанным для повышения иммуногенности эпитопа путем стимуляции Т-клеток-хелперов. Однако одной существенной проблемой при данном подходе является то, что он дает ограниченный иммунный ответ на белок в целом. Более того, вакцины должны индивидуально разрабатываться для различных гаплотипов МНС (главного комплекса гистосовместимости). Наиболее серьезной проблемой для данного типа вакцин является то, что защитные антивирусные антитела распознают сложные трехмерные структуры, которые не могут имитироваться пептидами.
Более новая стратегия вакцинации представляет собой применение ДНК-вакцин (см. Dоnnеllу и соавт., Апп.Rev. Immunol., 15: 617(1997)), которые могут вызывать ответы CTL с рестрикцией МНС класса I (без использования живого вектора). Это может обеспечить более широкую защиту от различных штаммов вируса посредством направленности на эпитопы из консервативных внутренних белков, характерных для многих штаммов одного вируса. Поскольку антиген образуется с использованием посттрансляционной модификации, конформации и олигомеризации, характерных для млекопитающих, более вероятно, что он является близким или идентичным белку дикого типа, который образуется при вирусной инфекции, чем рекомбинантные или химически модифицированные белки. Однако, данное отличие может стать недостатком для применения бактериальных антигенов, поскольку не природная посттрансляционная модификация может в результате привести к пониженной иммуногенности. Кроме того, вирусные поверхностные белки не имеют высокого уровня организации в отсутствие белков матрикса.
В дополнение к профилактике инфекционной болезни вакцинную технологию используют в настоящее время для решения проблем иммунитета, связанных с аллергиями. У субъектов с аллергией антитела изотипа IgЕ образуются при неадекватном гуморальном иммунном ответе в отношении определенных антигенов (аллергенов). Лечение аллергий путем иммунотерапии аллергий требует еженедельного введения последовательно возрастающих доз определенного антигена в течение периода до 3-5 лет. Вероятно, образуются "блокирующие" антитела IgG, которые задерживают аллергены в выделениях носа и дыхательных путей или в оболочках до того, как они реагируют с антителами IgE или мастоцитами. Однако не существует никакой постоянной связи между титрами IgG и облегчением симптомов. В настоящее время это представляет собой чрезвычайно длительный и дорогостоящий процесс, который предусматривается только для пациентов, у которых каждый год проявляются тяжелые симптомы в течение длительного периода.
Точно установлено, что введение только очищенных белков обычно не является достаточным для получения сильного иммунного ответа; выделенный антиген, как правило, должен вводиться вместе с вспомогательными субстанциями, называемыми адъювантами. Вместе сданными адъювантами введенный антиген является защищенным от быстрой деградации, а адъювант обеспечивает пролонгированное освобождение антигена на низком уровне.
В отличие от выделенных белков вирусы индуцируют быстрые и эффективные иммунные ответы в отсутствие каких-либо адъювантов как с помощью Т-клеток, так и без нее (см. статью Bachmann и Zinkernagel, Ann. Rev. Immunol., 15: 235-270 (1997)). Хотя вирусы часто состоят из небольшого числа белков, они способны включать более сильные иммунные ответы, чем выделенные из них компоненты. Для В-клеточных ответов известно, что одним из решающих факторов иммуногенности вирусов является повторяемость и порядок поверхностных эпитопов. Многие вирусы имеют квазикристаллическую поверхность, на которой представлена упорядоченная последовательность эпитопов, которые эффективно перекрестно сшивают эпитоп-специфические иммуноглобулины на В-клетках (см. статью Bachmann и Zinkernagel, Immunol. Today, 17:553-555 (1996)). Данное перекрестное сшивание поверхностных иммуноглобулинов на В-клетках представляет собой сильный сигнал активации, который непосредственно индуцирует развитие клеточного цикла и продукцию антител IgM. Кроме того, стимулированные В-клетки способны активировать Т-клетки-хелперы, которые в свою очередь индуцируют переключение в В-клетках продукции антител с IgM на IgG и образование длительной В-клеточной памяти - цели любой вакцинации (см. статью Bachmann и Zinkernagel, Ann. Rev. Immunol., 15: 235-270 (1997)). Структура вируса связана даже с генерацией анти-антител при аутоиммунных заболеваниях и как части естественного ответа на патогены (см. Fehr Т. и соавт., J. Exp. Med., 185: 1785-1792 (1997)). Таким образом антигены на вирусных частицах, которые организованы в виде упорядоченной и содержащей повторы матрицы, являются высокоиммуногенными, поскольку они могут непосредственно активировать В-клетки.
Кроме сильных В-клеточных ответов вирусные частицы способны также индуцировать образование ответа цитотоксических Т-клеток, другого решающего компонента иммунной системы. Данные цитотоксические Т-клетки являются особенно важными для элиминации нецитопатических вирусов, таких как ВИЧ или вирус гепатита В, и для борьбы с опухолями. Цитотоксические Т-клетки не распознают нативные антигены, но, однако, распознают продукты их деградации, связанные с молекулами МНС класса 1 (см. статью Townsend и Bodmer, Ann. Rev. Immunol., 7: 601-624 (1989)). Макрофаги и дендритные клетки способны поглощать и процессировать экзогенные вирусные частицы (но не их растворимые выделенные компоненты) и представлять образованные продукты деградации цитотоксическим Т-клеткам, вызывая их активацию и пролиферацию (см. статьи Kovacsovics-Bankowski и соавт., Proc. NatI. Acad. Sci. USA, 90: 4942-4946 (1993); Bachmann и соавт., Eur. J. Immunol., 25: 2595-2600 (1996)).
Вирусные частицы в качестве антигенов обладают двумя преимуществами относительно выделенных из них компонентов: (1) благодаря их поверхностной структуре с высокой степенью повторов они способны непосредственно активировать В-клетки, что приводит к высокому титру антител и длительной памяти В-клеток, и (2) вирусные частицы, но не растворимые белки способны индуцировать ответ цитотоксических Т-клеток даже, если вирусы являются неинфекционными и адъюванты отсутствуют.
В ряде новых вакцинных стратегий используют присущую вирусам иммуногенность. Некоторые из данных подходов сфокусированы на гранулярной природе вирусной частицы, например, см. статьи Harding C.V. и Song R. (J. Immunology, 153:4925 (1994)), в которой описана вакцина, состоящая из частиц латекса и антигена; Kovacsovics-Bankowski М. и соавт. (Рrос. NatI. Acad. Sci. USA, 90:4942-4946 (1993)), в которой описана вакцина, состоящая из частиц оксида железа и антигена; Патент США No 5334394, полученный Kossovsky N., и соавт., в котором раскрывают коровые частицы, покрытые антигеном; Патент США No 5871747, в котором раскрывают синтетические частицы, несущие на поверхности один или более белков, ковалентно связанных с ними, а также коровую частицу с нековалентно связанным покрытием и по меньшей мере одним биологически активным агентом, находящимся в контакте с указанной покрытой частицей кора (см., например, заявку WO /94/15585).
Однако недостаток данных имитирующих вирусы систем состоит в том, что они неспособны к воссозданию упорядоченной презентации антигенов, обнаруженной на поверхности вирусов. Антигены, связанные с поверхностью в произвольной ориентации, как обнаружено, индуцируют ответ CTL и не индуцируют или индуцируют слабый ответ В-клеток. Для эффективной вакцины оба компонента иммунной системы должны быть сильно активированы, как описано выше и в статье Bachmann и Zinkernagel, Ann. Rev. Immunol., 15: 235(1997).
В другом примере рекомбинантные вирусы используют для доставки антигена. Было показано, что вирус, представленный нитчатым фагом, содержащий антиген, слитый с капсидным белком, обладает высокой иммуногенностью (см. статьи Perham R.N., и соавт., FEMS Microbiol. Rev., 17: 25-31 (1995); Willis и соавт., Gene, 128: 85-88 (1993); Minenkova и соавт., Gene, 128: 85-88 (1993)). Однако данная система ограничена очень маленькими пептидами (из 5 и 6 остатков аминокислот) при экспрессии слитого белка на высоком уровне (см. статью lannolo и соавт., J. Mol. Biol., 248: 835-844 (1995)) или ограничена низким уровнем экспрессии более крупных белков (см. статью de la Cruz и соавт., J. Biol. Chem., 263: 4318-4322 (1988)). При этом для маленьких пептидов отмечается только ответ CTL и отсутствует или имеется только слабый ответ В-клеток.
В еще одной системе предложены альфавирусы в качестве средства для доставки антигена (см. Патенты США NoNo 5766602; 5792462; 5739026; 5789245 и 5814482). Проблемы с использованием рекомбинантных вирусных систем, описанных до настоящего времени, включают низкую плотность экспрессии гетерологичного белка на поверхности вируса и/или сложность успешного и воспроизводимого получения новых и различных рекомбинантных вирусов для различных областей применения.
В дальнейших разработках вирусоподобные частицы (VLP) будут использоваться в области получения вакцин вследствие как их структурных свойств, так и их неинфекционной природы. VLP представляют собой супермолекулярные структуры, построенные симметричным образом из множества белковых молекул одного или более типов. В них отсутствует вирусный геном, вследствие чего они являются неинфекционными. Часто VLP можно получить в больших количествах путем гетерологичной экспрессии и их можно легко очистить.
Примеры VLP включают капсидные белки вируса гепатита В (см. статью Ulrich и соавт., Virus Res., 50: 141-182 (1998)), вируса кори (см. статью Warnes и соавт., Gene, 160: 173-178(1995)); вируса Sindbis, ротавируса (Патенты США NoNo 5071651 и 5374426), вируса ящура (см. статьи Twomey и соавт., Vaccine, 13: 1603-1610, (1995)), вируса Norwalk (см. статьи Jiang X. и соавт., Science 250: 1580-1583 (1990); MatsuiSM. и соавт., J. Clin. Invest. 87: 1456-1461 (1991)), ретровирусный белок GAG (см. Патентную заявку РСТ No. WO 96/30523), белок р1 ретротранспозона Ту, поверхностный белок вируса гепатита В (см. WO 92/11291) и вируса папилломы человека (см. WO 98/15631). В ряде случаев возможно применение технологии рекомбинантной ДНК для слияния гетерологичного белка с белком VLP (см. статью Kratz Р.А. и соавт., Proc. NatI. Acad. Sci. USA, 96:1915-1920 (1999)).
Таким образом в данной области техники существует необходимость разработки новых и усовершенствованных вакцин, которые стимулируют сильный иммунный ответ CTL и В-клеток так же эффективно, как природные патогены.
Сущность изобретения
Изобретение представляет собой многофункциональную новую технологию, которая позволяет получать частицы, покрытые любым желательным антигеном. Технология позволяет создавать высокоэффективные вакцины против инфекционных болезней и разрабатывать вакцины для лечения аллергий и злокачественных новообразований.
В первом варианте осуществления изобретение представляет новую композицию, содержащую (А) не природный молекулярный носитель и (Б) антиген или антигенную детерминанту.
Не природный молекулярный носитель включает коровую частицу, выбранную из группы, состоящей из коровой частицы не природного происхождения и коровой частицы природного происхождения, а также организатор, содержащий по меньшей мере один первый центр связывания, причем указанный организатор соединен с указанной коровой частицей по меньшей мере одной ковалентной связью.
Антиген или антигенная детерминанта имеет по меньшей мере один второй центр связывания, который выбран из группы, состоящей из центра связывания, не встречающегося в природе с указанным антигеном или антигенной детерминантой, и центра связывания, существующего в природе с указанным антигеном или антигенной детерминантой.
Изобретение представляет упорядоченную и содержащую повторы антигенную матрицу, полученную путем ассоциации второго центра связывания с первым центром связывания посредством по меньшей мере одной непептидной связи. Таким образом антиген или антигенная детерминанта и не природный молекулярный носитель соединены вместе за счет данной ассоциации первого и второго центров связывания с образованием упорядоченной и содержащей повторы антигенной матрицы.
В другом варианте осуществления коровая частица вышеупомянутой композиции содержит вирус, вирусоподобную частицу, бактериофаг, вирусную капсидную частицу или их рекомбинантную форму. Альтернативно коровая частица может быть представлена синтетическим полимером или металлом.
В конкретном варианте осуществления организатор может содержать по меньшей мере один первый центр связывания. Первый и второй центры связывания являются особенно важными элементами композиции, соответствующей изобретению. В различных вариантах осуществления изобретения первый и/или второй центр связывания могут быть представлены антигеном и антителом или фрагментом антитела против антигена; биотином и авидином; стрептавидином и биотином; рецептором и его лигандом; лиганд-связывающим белком и его лигандом; полипептидами, взаимодействующими по типу лейциновой молнии; аминогруппой и реагирующей с ней химической группой; карбоксильной группой и реагирующей с ней химической группой; сульфгидрильной группой и реагирующей с ней химической группой; или их комбинацией.
В более предпочтительном варианте осуществления в изобретении представлено связывание практически любого выбранного антигена с поверхностью вируса, бактериофага, вирусоподобной частицы или вирусной капсидной частицы. В изобретении разработана сильная антивирусная иммунная реакция хозяина за счет введения антигена в квазикристаллическую "вирусоподобную" структуру с получением высокоэффективного иммунного ответа, т.е. вакцинации против представленного антигена.
В одном предпочтительном варианте осуществления коровая частица может быть выбрана из группы, состоящей из рекомбинантных белков ротавируса, рекомбинантных белков вируса Norwalk, рекомбинантных белков альфавируса, рекомбинантных белков вируса ящура, рекомбинантных белков ретровируса, рекомбинантных белков вируса гепатита В, рекомбинантных белков вируса табачной мозаики, рекомбинантных белков вируса помещений для животных и рекомбинантных белков вируса папилломы человека.
В другом предпочтительном варианте осуществления антиген может быть выбран из группы, состоящей из: (1) белка, пригодного для индукции иммунного ответа против раковых клеток; (2) белка, пригодного для индукции иммунного ответа против инфекционных болезней, (3) белка, пригодного для индукции иммунного ответа против аллергенов, и (4) белка, пригодного для индукции иммунного ответа у сельскохозяйственных животных.
В особенно предпочтительном варианте осуществления изобретения первый центр связывания и/или второй центр связывания содержат полипептид, взаимодействующий по типу лейциновой молнии. В наиболее предпочтительном варианте осуществления первый центр связывания и/или второй центр связывания выбраны из группы, содержащей (1) домен белка лейциновой молнии JUN и (2) домен белка лейциновой молнии FOS.
В другом предпочтительном варианте осуществления первый центр связывания и/или второй центр связывания выбраны из группы, содержащей (1) генно-инженерный остаток лизина и (2) генно-инженерный остаток цистеина, т.е. два остатка, которые могут химически связываться друг с другом.
Другие варианты осуществления изобретения включают способы получения композиций, соответствующих изобретению, и способы терапевтического лечения с использованием указанных композиций.
Следует понимать, что как предшествующее общее описание, так и последующее подробное описание представлены только в качестве примеров и разъяснений и предназначены для дальнейшего объяснения изобретения в том виде, как оно заявлено.
Перечень чертежей
На Фигуре 1 представлен результат вестерн-блоттинга, демонстрирующий образование вирусных частиц, содержащих слитый белок E2-JUN, с помощью экспрессионного вектора pCYTts::E2JUN.
На Фигуре 2 представлен результат вестерн-блоттинга, демонстрирующий образование вирусных частиц, содержащих слитый белок E2-JUN, экспрессированный с помощью экспрессионного вектора pTE5’2J::E2JUN.
На Фигуре 3 представлен результат вестерн-дот-блоттинга, демонстрирующий экспрессию антигена FOS-ngh в бактериях и эукариотах.
На Фигуре 4 представлена экспрессия HBcAg-JUN в клетках Е. coli.
На Фигуре 5 представлен результат вестерн-блоттинга, демонстрирующий, что HBcAg-JUN является растворимым в лизатах Е coli.
На Фигуре 6 представлен анализ с помощью SDS-PAGE (электрофорез в полиакриламидном геле с использованием додецилсульфата натрия) обогащения капсидных частиц HBcAg-JUN в градиенте плотности сахарозы.
На Фигуре 7 представлен анализ с помощью SDS-PAGE в невосстанавливающих условиях связывания частиц hGH-FOS и HBcAg-JUN.
Сведения, подтверждающие возможность осуществления изобретения
1. Определения
Следующие определения представлены для разъяснения сущности изобретения, которая, как полагают изобретатели, составляет данное изобретение.
Альфавирус: Как используют в данном контексте, термин "альфавирус" относится к любому из РНК-вирусов, включенных в род Alphavirus. Описания представителей данного рода содержатся в работе Strauss и Strauss, Microbiol. Rev., 58: 491-562 (1994)). Примеры альфавирусов включают вирусы Aura, Bebaru, Cabassou, Chikungunya, восточного энцефаломиелита лошадей, Fortmorgan, Getah, Kyzylagach, Mayoaro, Middleburg, Mucambo, Ndumu, Pixuna, Tonate, Triniti, Una, западного энцефаломиелита лошадей, Whataroa, Sindbis (SIN), Semliki forest (SFV), венесуэльского энцефаломиелита лошадей (VEE) и Ross River.
Антиген: Как используют в данном контексте, термин "антиген" представляет молекулу, способную быть связанной антителом. Антиген дополнительно способен индуцировать гуморальный иммунный ответ и/или клеточный иммунный ответ, приводящие к образованию В- и/или Т-лимфоцитов. Антиген может иметь один или более эпитопов (В- и Т-эпитопы). Специфическая реакция, относящаяся к вышеизложенному, предназначается для обозначения того, что антиген будет реагировать с высокой селективностью с соответствующим ему антителом и не будет реагировать с множеством других антител, которые могут стимулироваться другими антигенами.
Антигенная детерминанта: Как используют в данном контексте, термин "антигенная детерминанта" обозначает ту часть антигена, которая специфически распознается В- или Т-лимфоцитами. В-лимфоциты отвечают на чужеродные антигенные детерминанты путем выработки антител, тогда как Т-лимфоциты являются медиатором клеточного иммунитета. Таким образом, антигенные детерминанты или эпитопы представляют собой те части антигена, которые распознаются антителами или в контексте МНС Т-клеточными рецепторами.
Ассоциация: Как используют в данном контексте, термин "ассоциация" применительно к первому и второму центрам связывания используют по отношению к по меньшей мере одной непептидной связи. Природа ассоциации может быть ковалентной, ионной, гидрофобной, полярной или любой их комбинацией.
Центр связывания, первый: Как используют в данном контексте, выражение "первый центр связывания" относится к элементу "организатора", который сам связан с коровой частицей упорядоченным образом и с которым может ассоциироваться (связываться) второй центр связывания, находящийся на антигене или антигенной детерминанте. Первый центр связывания может быть белком, полипептидом, пептидом, сахаром, полинуклеотидом, природным или синтетическим полимером, вторичным метаболитом или соединением (биотином, флуоресцеином, ретинолом, дигоксигенином, ионами металлов, фенилметилсульфонилфторидом) или их комбинацией или их химически реактивной группой. Множество первых центров связывания присутствует на поверхности не природного молекулярного носителя в содержащей повторы конфигурации.
Центр связывания, второй: Как используют в данном контексте, выражение "второй центр связывания" относится к элементу, присоединенному к антигену или антигенной детерминанте, с которым может ассоциироваться первый центр связывания "организатора", находящийся на поверхности не природного молекулярного носителя. Второй центр связывания антигена или антигенной детерминанты может быть белком, полипептидом, пептидом, сахаром, полинуклеотидом, природным или синтетическим полимером, вторичным метаболитом или соединением (биотином, флуоресцеином, ретинолом, дигоксигенином, ионами металлов, фенилметилсульфонилфторидом) или их комбинацией или их химически реактивной группой. На антигене или антигенной детерминанте присутствует по меньшей мере один второй центр связывания.
Коровая частица: Как используют в данном контексте, термин "коровая частица" относится к жесткой структуре с присущей ей организацией с повторами, которая обеспечивает основу для присоединения "организатора". Коровая частица, как используют в данном контексте, может быть продуктом процесса синтеза или продуктом биологического процесса.
Цис-действующий: Как используют в данном контексте, выражение "цис-действующая" последовательность относится к последовательностям нуклеиновых кислот, которые связывает репликаза для того, чтобы катализировать РНК-зависимую репликацию молекул РНК. Данные репликационные события приводят к репликации молекул РНК полной длины или частичных молекул РНК, и, таким образом, субгеномный промотор альфавирусов также представляет собой "цис-действующую" последовательность. Цис-действующие последовательности могут находиться на 5’-конце, 3’-конце или обоих концах молекулы нуклеиновой кислоты или около них, а также внутри последовательности.
Слияние: Как используют в данном контексте, термин "слияние" относится к комбинации последовательностей аминокислот различной природы в полипептидной цепи посредством комбинации в рамке считывания кодирующих их нуклеотидных последовательностей. Термин "слияние" однозначно охватывает внутренние слияния, т.е. инсерцию последовательностей различной природы в полипептидную цепь, в дополнение к слиянию с одним из ее концов.
Гетерологичная последовательность: Как используют в данном контексте, термин "гетерологичная последовательность" относится к нуклеотидной последовательности, которая присутствует в векторе, соответствующем изобретению. Термин "гетерологичная последовательность" относится также к любой последовательности аминокислот или РНК, кодируемой гетерологичной последовательностью ДНК, содержащейся в векторе, соответствующем изобретению. Гетерологичные нуклеотидные последовательности могут кодировать молекулы белков или РНК, которые нормально экспрессируются в типе клеток, в которых они присутствуют, или молекулы, которые нормально не экспрессируются в них (например, структурные белки Sindbis).
Выделенный: Как используют в данном контексте, где термин "выделенный" используют по отношению к молекуле, данный термин означает, что молекула была удалена из естественной для нее среды. Например, полинуклеотид или полипептид, присутствующий в естественных условиях в организме живого животного, не является "выделенным", но тот же самый полинуклеотид или полипептид, отделенный от материалов, совместно с ним существующих в его природном состоянии, является "выделенным". Кроме того, рекомбинантные молекулы ДНК, содержащиеся в векторе, считают выделенными с учетом целей данного изобретения. Выделенные молекулы РНК включают представленные РНК продукты репликации молекул ДНК и РНК in vivo или in vitro. Выделенные молекулы нуклеиновых кислот далее включают молекулы, полученные синтетическим путем. Кроме того, векторные молекулы, содержащиеся в рекомбинантных клетках-хозяевах, также являются выделенными. Таким образом, не все "выделенные" молекулы должны быть "очищенными".
Иммунотерапевтический: Как используют в данном контексте, термин "иммунотерапевтический" представляет композицию для лечения заболеваний или нарушений. В частности, термин обычно касается способа лечения аллергий или способа лечения рака.
Субъект: Как используют в данном контексте, термин "субъект" относится к многоклеточным организмам и включает как растения, так и животных. Предпочтительно, когда многоклеточные организмы являются животными, более предпочтительно позвоночными, даже более предпочтительно представлены млекопитающими и наиболее предпочтительно человеком.
Низкий или неопределяемый: Как используют в данном контексте, выражения "низкий или неопределяемый" при использовании касательно уровня экспрессии гена относится к уровню экспрессии, который является либо значительно более низким, чем определяемый при максимальной индукции гена (например, по меньшей мере в пять раз ниже), либо является трудно определяемым способами, используемыми в следующем разделе примеров.
Лектин: Как используют в данном контексте, белки, полученные в основном из семян бобовых растений, но также из многих других растительных и животных источников, которые имеют центры связывания для специфических моно- или олигосахаридов. Примеры включают конканавалин А и агглютинин зародышей пшеницы, которые широко используют в качестве аналитических и препаративных агентов в исследования гликопротеинов.
Естественной природы (происхождения): Как используют в данном контексте, термин "естественной природы (происхождения)" означает, что целое или его части не являются синтетическими и существуют или образуются в природе (т.е. в естественных условиях).
Не природный: Как используют в данном контексте, термин в основном означает полученный не из природы, в частности, термин означает "сделанный руками человека".
Не природного происхождения: Как используют в данном контексте, термин "не природного происхождения" означает синтетический или полученный не из природы, в частности, термин означает сделанный руками человека.
Не природный молекулярный носитель: Как используют в данном контексте, выражение "не природный молекулярный носитель" относится к любому продукту, сделанному руками человека, который может служить для получения жесткой и содержащей повторы последовательности первых центров связывания. В идеальном случае данные первые центры связывания расположены в геометрическом порядке, но это не является необходимым. Не природный молекулярный носитель может быть органическим или неорганическим и может быть синтезирован химическим путем или с помощью биологического процесса частично или полностью. Не природный молекулярный носитель состоит из (а) коровой частицы природного или не природного происхождения и (b) организатора, который сам содержит по меньшей мере один первый центр связывания и соединен с частицей кора по меньшей мере одной ковалентной связью. В определенном варианте осуществления не природный молекулярный носитель может быть представлен вирусом, вирусоподобной частицей, вирусной капсидной частицей, фагом, их рекомбинантными формами или синтетической частицей.
Упорядоченная и содержащая повторы матрица антигенов или антигенных детерминант: Как используют в данном контексте, термин "упорядоченная и содержащая повторы матрица антигенов или антигенных детерминант" в основном касается повторов структуры антигена или антигенной детерминанты, характеризующихся одинаковой пространственной организацией антигенов или антигенных детерминант относительно основы. В одном варианте осуществления изобретения образец повтора может иметь геометрическую форму. Идеальная упорядоченная и содержащая повторы матрица антигенов или антигенных детерминант будет представлять собой имеющую четкие повторы паракристаллическую структуру антигена или антигенной детерминанты с расстояниями 5-15 нанометров.
Организатор: Как используют в данном контексте, термин "организатор" применяют для обозначения элемента, связанного с частицей кора непроизвольным образом, который обеспечивает участок образования ядра для создания упорядоченной и содержащей повторы антигенной матрицы. Организатор представляет собой любой элемент, содержащий по меньшей мере один первый центр связывания, который соединен с частицей кора с помощью по меньшей мере одной ковалентной связи. Организатор может быть белком, полипептидом, пептидом, аминокислотой (т.е. остатком белка, полипептида или пептида), сахаром, полинуклеотидом, природным или синтетическим полимером, вторичным метаболитом или соединением (биотином, флуоресцеином, ретинолом, дигоксигенином, ионами металлов, фенилметилсульфонилфторидом) или их комбинацией или их химически реактивной группой.
Пермиссивная температура: Как используют в данном контексте, выражения "пермиссивная температура" относится к температурам, при которых фермент имеет относительно высокий уровень каталитической активности.
Очищенный: Как используют в данном контексте, когда термин "очищенный" используют в отношении молекулы, он означает, что концентрация очищаемой молекулы повысилась относительно молекул, ассоциированных с ней в ее естественной среде. Ассоциированные естественным образом молекулы включают белки, нуклеиновые кислоты, липиды и сахара, но обычно не включают воду, буферы и реагенты, добавляемые для поддержания целостности или облегчения очистки молекулы, которую очищают. Например, даже если мРНК разбавить водным растворителем во время колоночной хроматографии с использованием олиго dT, молекулы мРНК очищают посредством данной хроматографии, если естественным образом ассоциированные нуклеиновые кислоты и другие биологические молекулы не связываются с колонкой и отделяются от данных молекул РНК.
Рецептор: Как используют в данном контексте, термин "рецептор" относится к белкам или гликопротеинам или их фрагментам, способным к взаимодействию с другой молекулой, называемой лигандом. Лиганд может принадлежать к любому классу биохимических или химических соединений. Рецептор не обязательно должен быть связанным с мембраной белком. Растворимый белок, такой как, например, белок, связывающий мальтозу, или белок, связывающий ретинол, также является рецептором.
Остаток: Как используют в данном контексте, термин "остаток" предназначен для обозначения определенной аминокислоты в скелете или боковой цепи полипептида.
Температурочувствительный: Как используют в данном контексте, выражение "Температурочувствительный" относится к ферменту, который легко катализирует реакцию при одной температуре, но медленно катализирует ту же реакцию или совсем не катализирует ее при другой температуре. Примером температурочувствительного фермента является белок репликазы, кодируемый вектором pCYTts, который обладает легко определяемой активностью репликазы при температурах, не превышающих 34°С, и имеет низкую или неопределяемую активность при 37°С.
Транскрипция: Как используют в данном контексте, термин "транскрипция" касается образования молекул РНК с ДНК-матриц, которое катализируется РНК-полимеразами.
Рекомбинантная клетка-хозяин: Как используют в данном контексте, термин "рекомбинантная клетка-хозяин" касается клетки-хозяина, в которую была интродуцирована одна или более молекул нуклеиновой кислоты, соответствующих изобретению.
Рекомбинантный вирус: Как используют в данном контексте, выражение "рекомбинантный вирус" относится к вирусу, который является генетически модифицированным вмешательством человека. Выражение покрывает любой вирус, известный в области техники. В частности, выражение касается альфавируса, генетически модифицированного человеком, и более специально выражение относится к вирусу Sinbis, генетически модифицированному человеком.
Ограничивающая температура: Как используют в данном контексте, выражение "ограничивающая температура" относится к температурам, при которых фермент имеет низкие или неопределяемые уровни каталитической активности. Известны оба, как "тепло-", так и "холодо"чувствительные мутанты, и таким образом ограничивающая температура может быть выше или ниже пермиссивной температуры.
Событие РНК-зависимой репликации РНК: Как используют в данном контексте, выражение "событие РНК-зависимой репликации РНК" относится к процессам, которые приводят к образованию молекулы РНК с использованием молекулы РНК в качестве матрицы.
РНК-зависимая РНК-полимераза: Как используют в данном контексте, выражение " РНК-зависимая РНК-полимераза" относится к полимеразе, которая катализирует образование молекулы РНК с помощью другой молекулы РНК. Данный термин используют в данном контексте как синоним термина "репликаза".
Нетранслируемая РНК: Как используют в данном контексте, выражение "нетранслируемая РНК" относится к последовательности РНК или молекуле, которая не кодирует открытую рамку считывания или кодирует открытую рамку считывания или ее фрагмент, но в формате, в котором не образуется последовательность аминокислот (например, не присутствует никакой кодон инициации). Примерами данных молекул являются молекулы тРНК, молекулы рРНК и рибозимы.
Вектор: Как используют в данном контексте, термин "вектор" относится к агенту (например, плазмиде или вирусу), используемому для переноса генетического материала в клетку-хозяин. Вектор может состоять либо из ДНК, либо из РНК.
Один или какой-либо: При использовании терминов "один или какой-либо" в данном описании они означают "по меньшей мере один" или "один или несколько", если не указано иначе.
2. Композиции упорядоченных и содержащих повторы матриц антигена или антигенной детерминанты и способы их получения
Описанное изобретение представляет композиции, содержащие упорядоченный и содержащий повторы антиген или антигенную детерминанту. Более того, изобретение представляет специалисту-практику удобную возможность конструировать упорядоченные и содержащие повторы матрицы антигена или антигенной детерминанты для различных лечебных целей, которые включают профилактику инфекционных болезней, лечение аллергий и лечение злокачественных новообразований.
Композиции, соответствующие изобретению, в основном содержат два элемента: (1) не природный молекулярный носитель и (2) антиген или антигенную детерминанту с по меньшей мере одним вторым центром связывания, способным образовывать ассоциацию посредством по меньшей мере одной непептидной связи с указанным первым центром связывания.
Не природный молекулярный носитель включает (а) коровую частицу, выбранную из группы, состоящей из (1) коровой частицы не природного происхождения и (2) коровой частицы природного происхождения, и (b) организатор, содержащий по меньшей мере один первый центр связывания, где указанный организатор связан с указанной коровой частицей по меньшей мере одной ковалентной связью.
Антиген или антигенная детерминанта имеет по меньшей мере один второй центр связывания, который выбран из группы, состоящей из (а) центра связывания, не встречающегося в природе в данном антигене или антигенной детерминанте, и (b) центра связывания, который имеется в естественных условиях в данном антигене или антигенной детерминанте.
Изобретение представляет упорядоченную и содержащую повторы антигенную матрицу, полученную за счет ассоциации второго центра связывания с первым центром связывания с помощью по меньшей мере одной непептидной связи. Таким образом, антиген или антигенная детерминанта и не природный молекулярный носитель соединены вместе за счет данной ассоциации первого и второго центров связывания с образованием упорядоченной и содержащей повторы антигенной матрицы.
Практикующий специалист может специально сконструировать антиген или антигенную детерминанту и второй центр связывания так, что расположение (размещение) всех антигенов или антигенных детерминант, связанных с не природным молекулярным носителем (основой), будет равномерным. Например, можно поместить единственный второй центр связывания на карбоксильном или аминоконце антигена или антигенной детерминанты, и такая конструкция обеспечит единообразное расположение всех молекул антигена или антигенной детерминанты, связанных с не природным молекулярным носителем. Таким образом, изобретение представляет удобное средство для размещения любого антигена или антигенной детерминанты на не природном молекулярном носителе в определенном порядке и повторах.
Для компетентных специалистов в данной области будет ясно, что некоторые варианты осуществления изобретения включают применение технологий рекомбинантных нуклеиновых кислот, таких как клонирование, полимеразная цепная реакция, очистка ДНК и РНК, экспрессия рекомбинантных белков в прокариотических и эукариотических клетках и т.п. Данные методологии хорошо известны компетентным специалистам в данной области и их можно легко найти в опубликованных руководствах по лабораторным методам (например, см. монографию под ред. Sambrook J. и соавт., Лабораторное руководство по молекулярному клонированию (Molecular Cloning, A Laboratory Manual), 2-е изд., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989); монографию под ред. Ausubel F. и соавт. Современные методы молекулярной биологии (Current Protocols in Molecular Biology), John H. Wiley & Sons, Inc. (1997)). Фундаментальные лабораторные методики для работы с культурами тканей и клеточными линиями (см. монографию Биология клетки (Cell Biology) под ред. Celis J., Academic Press, 2-е изд. (1998)) и технологии на основе антител (см. монографию Harlow E. и Lane D., "Лабораторное руководство по антителам" ("Antibodies: A Laboratory Manual,") Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. (1988); работу Deutscher M.P. "Руководство по очистке белков" ("Guide to Protein Purification,") Meth. Enzymol., 128, Academic Press, San Diego (1990); монографию Scopes R.K. "Принципы и практические аспекты очистки белков" ("Protein Purification Principles and Practice") 3-е изд., Springer-Verlag, New York (1994)) также адекватно описаны в литературе, которая включена в данном контексте в виде ссылки.
А. Конструирование не природного молекулярного носителя
Одним из элементов композиции, соответствующей изобретению, является не природный молекулярный носитель, содержащий коровую частицу и организатор. Как используют в данном контексте, выражение "не природный молекулярный носитель" относится к любому продукту, сделанному человеком, который может служить для получения жесткой и содержащей повторы последовательности первых центров связывания. В частности, не природный молекулярный носитель включает (а) коровую частицу, выбранную из группы, состоящей из (1) коровой частицы не природного происхождения и (2) коровой частицы природного происхождения, и (b) организатор, содержащий по меньшей мере один первый центр связывания, причем указанный организатор связан с указанной коровой частицей по меньшей мере одной ковалентной связью.
Как будет легко понять компетентным в данной области специалистам, коровая частица не природного молекулярного носителя, соответствующего изобретению, не ограничена каким-либо конкретным видом. Коровая частица может быть органической или неорганической и может быть синтезирована химическим путем или с помощью биологического процесса.
В одном из вариантов осуществления изобретения не природная коровая частица может быть синтетическим полимером, липидной мицеллой или металлом. Данные коровые частицы известны в уровне техники и представляют собой основу для создания нового не природного молекулярного носителя (скелета), соответствующего изобретению. Например, коровые частицы из синтетического полимера или металла описаны в Патенте США No 5770380, в котором раскрыто применение каликсаренового органического носителя, с которым связано множество пептидных петель, для создания "имитатора антитела", а в Патенте США No 5334394 описывают нанокристаллические частицы, используемые в качестве вирусной ловушки, которые состоят из широкого круга неорганических материалов, включая металлы или керамику. Предпочтительные металлы в данном варианте осуществления включают хром, рубидий, железо, цинк, селен, никель, золото, серебро, платину. Предпочтительные керамические материалы в данном варианте осуществления включают диоксид кремния, диоксид титана, оксид алюминия, оксид рутения и оксид олова. Частицы кора, соответствующие данному варианту осуществления, могут быть сделаны из органических материалов, включая углерод (алмаз). Предпочтительные полимеры включают полистирол, нейлон и нитроцеллюлозу. Для нанокристаллической частицы данного типа особенно предпочтительными являются частицы, сделанные из оксида олова, диоксида титана или углерода (алмаза). Липидные мицеллы готовят с помощью любых средств, известных в области техники. Например, мицеллы могут быть получены в соответствии с процедурой, описанной Baiselle и Millar (см. статью Baiselle С.J. и Millar D.B., Biophys. Chem., 4: 355-361 (1975)) или Corti и соавт. (см. статью Corti М., DegriorgioV., Sonnino S., Ghidoni R., Masserini М. и Tettamanti, G., Chem. Phys. Lipids, 38: 197-214 (1981)) или Lopez и соавт. (Lopez O., de la Maza A., Coderch L, Lopez-lglesias C., Wehrii E. и Parra J.L, FEBS Lett., 426: 314-318 (1998)) или Topchieva и Karezin (Topchieva I. и Karaezin K., J. Colloid Interface Sci., 213:29-35 (1999)) или Morein и соавт. (Morein В., Sundquist В., Hoglund S., Dalsgaard К. и Osterhaus A., Nature 308: 457-60 (1984)), которые введены в данном контексте в виде ссылки.
Частица кора может быть также получена с помощью биологического процесса, который может быть природным или не природным. Например, данный тип варианта осуществления может включать коровую частицу, содержащую вирус, вирусоподобную частицу, фаг, частицу вирусного капсида или их рекомбинантные формы. В более специфическом варианте осуществления коровая частица может содержать рекомбинантные белки ротавируса, рекомбинантные белки вируса Norwalk, рекомбинантные белки альфавируса, рекомбинантные белки вируса ящура, рекомбинантные белки ретровируса, рекомбинантные белки вируса гепатита В, рекомбинантные белки вируса табачной мозаики, рекомбинантные белки вируса помещений для животных и рекомбинантные белки вируса папилломы человека.
Будучи природного или не природного происхождения, коровая частица, соответствующая изобретению, характеризуется тем, что содержит организатор, который присоединен к природной или не природной коровой частице по меньшей мере одной ковалентной связью. Организатор представляет собой элемент, связанный с коровой частицей не случайным образом, который обеспечивает участок ядра для создания упорядоченной и содержащей повторы антигенной матрицы. В идеальном случае, организатор связан с коровой частицей в геометрическом порядке, но это не является необходимым. Минимальным условием является то, что организатор содержит первый центр связывания.
Как определено ранее, организатор может быть любым элементом, содержащим по меньшей мере один первый центр связывания, который присоединен к коровой частице по меньшей мере одной ковалентной связью. Организатор может быть представлен белком, полипептидом, пептидом, аминокислотой (т.е. остатком белка, полипептида или пептида), сахаром, полинуклеотидом, природным или синтетическим полимером, вторичным метаболитом или соединением ((биотином, флуоресцеином, ретинолом, дигоксигенином, ионами металлов, фенилметилсульфонилфторидом) или их комбинацией или их химически реактивной группой. В более специфическом варианте осуществления организатор может включать по меньшей мере один первый центр связывания, содержащий антиген, антитело или фрагмент антитела, биотин, авидин, стрептавидин, рецептор, лиганд рецептора, лиганд, лиганд-связывающий белок, полипептид, взаимодействующий по типу лейциновой молнии, аминогруппу, химическую группу, реагирующую с аминогруппой, карбоксильную группу, химическую группу, реагирующую с карбоксильной группой, сульфгидрильную группу, химическую группу, реагирующую с сульфгидрильной группой, или их комбинацию.
В предпочтительном варианте осуществления коровая частица не природного молекулярного носителя содержит вирус, бактериофаг, вирусоподобную частицу, частицу вирусного капсида или их рекомбинантную форму. Любой вирус, известный в области техники, как имеющий упорядоченную содержащую повторы оболочку и/или структуру коревого белка, может быть выбран в качестве не природного молекулярного носителя, соответствующего изобретению; примеры подходящих вирусов включают sindbis и другие альфавирусы; вирус везикулярного стоматита, рабдо- (например, вирус везикулярного стоматита), пикорна-, тога-, ортомиксо-, вирусы, вирус полиомы, парвовирус, ротавирус, вирус ящура, ретровирус, вирус гепатита В, вирус табачной мозаики, вирус помещений для животных, вирус папилломы (например, см. Таблицу 1 в статье Bachman M.F. и Zinkernagel P.M., Immunol. Today, 77: 553-558 (1996)).
В одном из вариантов осуществления в изобретении используют генетическую инженерию вируса для создания слияния между упорядоченным и содержащим повторы белком вирусной оболочки и организатором, содержащим выбранные гетерологичный белок, пептид, антигенную детерминанту или реакционный остаток аминокислоты. Другие генетические манипуляции, известные специалистам в данной области, могут быть использованы при конструировании не природного молекулярного носителя, например, может быть желательным ограничение репликационной способности рекомбинантного вируса посредством генетической мутации. Вирусный белок, выбранный для слияния с белком организатора (т.е. первым центром связывания), должен иметь упорядоченную и содержащую повторы структуру, более предпочтительно паракристаллическую организацию, в оптимальном случае с расстояниями 5-15 нм на поверхности вируса. Создание слитого белка данного типа будет в результате приводить к образованию множества упорядоченных и содержащих повторы организаторов на поверхности вируса. Таким образом, полученная в результате упорядоченная и содержащая повторы организация первых центров связывания будет отражать нормальную организацию природного вирусного белка.
Как будет более подробно обсуждаться в данном контексте, в предпочтительном варианте осуществления изобретения носитель представляет собой рекомбинантный альфавирус и, в более частном случае, рекомбинантный вирус Sinbis. Альфавирусы являются вирусами с положительной цепью РНК, которые реплицируют свою геномную РНК полностью в цитоплазме инфицированной клетки и без участия промежуточной ДНК (см. статью Strauss J. и Strauss E., Microbiol. Rev., 58: 491-562 (1994)). Некоторые представители семейства альфавирусов, в частности, Sindbis (см. статьи Xiong С. и соавт., Science, 243: 1188-1191 (1989); Schlesinger S., Trends Biotechnol., 11: 18-22 (1993)), вирус Semliki Forest (SFV) (см. статью  P. и Garoff H., BioТechnology, 9: 1356-1361 (1991)) и другие (см. статью Davis N.L и соавт., Virology, 171: 189-204(1989)) привлекли большое внимание в плане использования в качестве векторов для экспрессии ряда различных белков на вирусной основе (см. статьи Lundstrom К., Curr. Opin. Biotechnol., 8: 578-582(1997); LiljestrЦm P., Curr. Opin. Biotechnol. 5: 495-500 (1994)) и в качестве кандидатов для разработки вакцин. Недавно был выдан ряд Патентов, направленных на применение альфавирусов для экспрессии гетерологичных белков и разработки вакцин (см. Патенты США NoNo 5766602; 5792462; 5739026; 5789245 и 5814482). Конструкция альфавирусного носителя, соответствующего изобретению, может быть сделана с помощью средств, в основном известных в области технологии рекомбинантной ДНК, как описано в вышеупомянутых статьях, которые включены в данном контексте в виде ссылки.
P. и Garoff H., BioТechnology, 9: 1356-1361 (1991)) и другие (см. статью Davis N.L и соавт., Virology, 171: 189-204(1989)) привлекли большое внимание в плане использования в качестве векторов для экспрессии ряда различных белков на вирусной основе (см. статьи Lundstrom К., Curr. Opin. Biotechnol., 8: 578-582(1997); LiljestrЦm P., Curr. Opin. Biotechnol. 5: 495-500 (1994)) и в качестве кандидатов для разработки вакцин. Недавно был выдан ряд Патентов, направленных на применение альфавирусов для экспрессии гетерологичных белков и разработки вакцин (см. Патенты США NoNo 5766602; 5792462; 5739026; 5789245 и 5814482). Конструкция альфавирусного носителя, соответствующего изобретению, может быть сделана с помощью средств, в основном известных в области технологии рекомбинантной ДНК, как описано в вышеупомянутых статьях, которые включены в данном контексте в виде ссылки.
Для получения коровые частицы на основе вируса для присоединения антигена или антигенной детерминанты может быть использовано множество различных рекомбинантных клеток-хозяев. Например, известно, что альфавирусы имеют широкий круг хозяев, вирус Sindbis инфицирует культивируемые клетки млекопитающих, рептилий и амфибий, а также некоторые клетки насекомых (см. статьи Clark H., J. Natl. Cancer Inst. 51: 645 (1973); Leake С., J. Gen. Virol., 35: 335 (1977); раздел, написанный Stollar V. в монографии "Тогавирусы" (The Togaviruses), под ред. R. W. Schlesinger, Academic Press, (1980), стр.583-621). Таким образом, для практической реализации изобретения может быть использовано множество рекомбинантных клеток-хозяев. Клетки ВНК, COS, Vero, HeLa и СНО являются особенно подходящими для получения гетерологичных белков, поскольку они обладают активностью гликозилирования гетерологичных белков, близкой активности клеток человека (см. статью Watson E. и соавт., Glycobiology 4: 227, (1994)), и могут быть отселектированы (см. статью Zang М. и соавт., Bio/Technology, 13: 389 (1995)) или получены генной инженерией (см. статьи Renner W. и соавт., Biotech. Bioeng., 4: 476 (1995); Lee К. и соавт., Biotech. Bioeng. 50: 336 (1996)), с целью выращивания в бессывороточной среде, а также в виде суспензии.
Интродукция полинуклеотидных векторов в клетки-хозяева может быть осуществлена способами, описанными в стандартных лабораторных руководствах (см., например, монографию под ред. Sambrook J. и соавт., Лабораторное руководство по молекулярному клонированию (Molecular Cloning, A Laboratory Manual), 2-е изд., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989), Глава 9; монографию под ред. Ausubel F. и соавт. Современные методы молекулярной биологии (Current Protocols in Molecular Biology), John H. Wiley & Sons, Inc. (1997), Глава 16), включающих такие способы, как электропорация, DEAE [(диэтиламино)этилцеллюлоза)]-декстран-опосредованная трансфекция, трансфекция, микроинъекция, опосредованная катионными липидами трансфекция, трансдукция, нагрузка с помощью соскоба, баллистическая интродукция (бомбардировка) и инфекция. Способы интродукции экзогенных последовательностей ДНК в клетки-хозяева обсуждают в Патенте No 5580859, выданном Felgner P. и соавт.
Упакованные последовательности РНК также могут быть использованы для инфицирования клеток-хозяев. Данные упакованные последовательности РНК можно интродуцировать в клетки-хозяева путем введения их в среду для культивирования. Например, в ряде источников, включая работу "Экспрессионная система Sindbis" ("Sindbis Expression System"), вариант С, (Invitrogen, каталожный No. K750-1), описан препарат неинфекционных альфавирусных частиц.
При использовании клеток млекопитающих в качестве рекомбинантных клеток-хозяев для получения коровых частиц на основе вируса данные клетки, как правило, выращивают в тканевой культуре. Способы выращивания клеток в культуре хорошо известны в области техники (см. монографию под ред. Celis J. Биология клетки (Cell Biology), Academic Press, 2-е изд. (1998); монографию под ред. Sambrook J. и соавт., Лабораторное руководство по молекулярному клонированию (Molecular Cloning, A Laboratory Manual), 2-е изд., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989); монографию под ред. Ausubel F. и соавт. Современные методы молекулярной биологии (Current Protocols in Molecular Biology), John H. Wiley & Sons, Inc. (1977); монографию Freshney R. Культура клеток животных (Culture of Animal Cells), Alan R. Liss, Inc. (1983)).
Специалистам в данной области будет понятно, что первый центр связывания может являться любым подходящим белком, полипептидом, сахаром, полинуклеотидом, пептидом (аминокислотой), природным или синтетическим полимером, вторичным метаболитом или быть их фрагментом или их комбинацией, которая может служить для специфического присоединения выбранного антигена или антигенной детерминаты к носителю. В одном из вариантов осуществления центр присоединения представлен белком или пептидом, который может быть выбран из известных в области техники. Например, первый центр связывания может быть выбран из следующей группы: лиганд, рецептор, лектин, авидин, стрептавидин, биотин, такой эпитоп, как НА или Т7 tag, Мус, Мах, домены иммуноглобулина и любая другая аминокислотная последовательность, о которой в области техники известно, что она была бы приемлемой в качестве первого центра связывания.
Далее специалистам в данной области следует понимать, что в другом варианте осуществления изобретения первый центр связывания может быть создан вторично по отношению к организатору (т.е. белку или полипептиду), использованному при конструировании в рамке слияния с капсидным белком. Например, белок может быть использован для слияния с белком оболочки с последовательностью аминокислот, о которой известно, что она гликозилируется специфическим образом, и введенная в результате этого молекула сахара затем служит первым центром связывания вирусного носителя посредством связывания лектина, который служит вторичным центром связывания антигена. Альтернативно последовательность организатора может быть биотинилирована in vivo и биотиновая часть может служить первым центром связывания, соответствующим изобретению, или последовательность организатора может быть подвергнута химической модификации различных остатков аминокислот in vitro, при этом данная модификация служит первым центром связывания.
В одном из специфических вариантов осуществления изобретения используют вирус Sinbis. Геномная РНК вируса Sinbis упакована в капсидный белок, который окружен двойным липидным слоем, содержащим три белка, названные Е1, Е2 и ЕЗ. Данные так называемые белки оболочки являются гликопротеинами, и их гликозилированные участки расположены вне двойного липидного слоя, где комплексы данных белков образуют "пики", которые можно видеть на электронных микрофотографиях выступающими из поверхности вируса. В предпочтительном варианте осуществления изобретения первый центр связывания выбран из домена белка лейциновой молнии JUN или FOS, который слит в рамке считывания с белком оболочки Е2. Однако для всех специалистов в данной области будет ясно, что можно использовать другие белки оболочки в конструкции слитого белка для помещения первого центра связывания в носитель, соответствующий изобретению.
В наиболее предпочтительном варианте осуществления изобретения первым центром связывания выбран домен белка лейциновой молнии JUN-FOS, который слит в рамке считывания с белком капсида (кора) вируса гепатита В. Однако для всех специалистов в данной области будет ясно, что можно использовать другие белки вирусного капсида в конструкции слитого белка для помещения первого центра связывания в носитель, соответствующий изобретению.
В другом предпочтительном варианте осуществления изобретения первым центром связывания выбран остаток лизина или цистеина, который слит в рамке считывания с белком капсида (кора) вируса гепатита. Однако для всех специалистов в данной области будет ясно, что можно использовать другие вирусный капсид или вирусоподобные частицы в конструкции слитого белка для помещения первого центра связывания в носитель, соответствующий изобретению.
Пример 1 представлен для демонстрации конструирования белка, полученного при слиянии в рамке считывания белка оболочки Е2 вируса Sinbis и белкового домена лейциновой молнии JUN с использованием вектора pTE5’2J, описанного Hahn и соавт. (см. Proc. NatI. Acad. Sci. USA, 89: 2679-2683(1992)). Последовательность аминокислот JUN, использованная для первого центра связывания, является следующей:
CGGRIARLEEKVKTLKAQNSELASTANMLREQVAQLKQKVMNHVGC (SEQ ID NO: 59). В данном случае ожидаемым вторым участком связывания на антигене был бы домен белка лейциновой молнии FOS и последовательность аминокислот была бы следующей:
CGGLTDTLQAETDQVEDEKSALQTEIANLLKEKEKLEFILAAHGGC (SEQ ID NO: 60).
Данные последовательности выделены из факторов транскрипции JUN и FOS, и каждая из них фланкирована короткой последовательностью, содержащей остаток цистеина на обеих сторонах. Известно, что данные последовательности взаимодействуют друг с другом. В исходной гипотетической структуре, предложенной для димера JUN-FOS, предполагается, что гидрофобные боковые цепи мономера переплетаются с соответствующими боковыми цепями другого мономера как в молнии (см. статью Landschuiz и соавт., Science, 240:1759-1764 (1988)). Однако, как доказано, эта гипотеза является неверной, и данные белки, как известно, образуют α-закрученную спираль (см. статьи O’Shea и соавт., Science, 243: 538-542 (1989); O’Shea и соавт., CelI, 68: 699-708 (1992); Cohen и Parry, Trends Biochem. Sci., 11: 245-248 (1986)). Таким образом, термин "лейциновая молния" часто используют для обозначения данных белковых доменов больше по историческим причинам, чем с точки зрения структуры. В данном патенте термин "лейциновая молния" используют для обозначения последовательностей, представленных выше, или последовательностей, в основном аналогичных вышеописанным последовательностям. Термины JUN и FOS используют для обозначения соответственных доменов лейциновой молнии, а не для целых белков JUN и FOS.
В одном варианте осуществления изобретение представляет получение носителя на основе Е2 вируса Sinbis E2JUN с использованием экспрессионной системы pCYTts (см. Патентную заявку США No. 60/079562, поданную 27 марта 1998 г.). Экспрессионная система pCYTts представляет собой новые экспрессионные векторы, которые обеспечивают жесткую регуляцию экспрессии генов в эукариотических клетках. ДНК-векторы данной системы транскрибируются с образованием молекул РНК, которые затем реплицируются с помощью температурочувствительной репликазы с образованием дополнительных молекул РНК. Полученные при репликации молекулы РНК содержат нуклеотидную последовательность, которая может быть транслирована с целью образования представляющего интерес белка или которая кодирует одну или более нетранслируемых молекул РНК. Таким образом экспрессионная система обеспечивает продукцию рекомбинантных частиц вируса Sinbis.
В Примере 2 представлены детали получения не природного молекулярного носителя Sinbis E2-JUN, соответствующего изобретению. В Примере 3 дополнительно представлен другой способ получения носителя на основе рекомбинантного вируса Sinbis E2-JUN с использованием вектора pTE5’2JE2:JUN, полученного в Примере 1. Таким образом, в изобретении представляют два средства - экспрессионную систему pCYTts (см. Пример 2) и векторную систему pTE5’2J (см. Пример 3), с помощью которых можно получить не природный молекулярный носитель на основе рекомбинантного вируса Sinbis E2-JUN. Анализ вирусных частиц, полученных в каждой системе, представлен на Фигуре 1 и Фигуре 2.
Как ранее показано, изобретение включает коровые частицы вирусной природы, которые содержат вирус, вирусоподобную частицу, фаг, частицу вирусного капсида или их рекомбинантные формы. Компетентные исследователи знают, как получить данные коровые частицы и присоединить к ним организаторы. Посредством представления других примеров изобретение предлагает в данном контексте получение частиц, подобных вирусу гепатита В, и частиц капсида вируса кори в качестве коровых частиц (см. Примеры 17-22). В данном варианте осуществления домен белка лейциновой молнии JUN или домен белка лейциновой молнии FOS могут быть использованы в качестве организатора и, следовательно, в качестве первого центра связывания для не природного молекулярного носителя, соответствующего изобретению.
В Примерах 23-29 приведены детали получения коровых частиц вируса гепатита В, несущих пептид, слитый в рамке считывания с реактивным остатком лизина, и антигены, несущие генетически слитый остаток цистеина, в качестве первого и второго центров связывания соответственно.
В. Конструирование антигена или антигенной детерминанты со вторым центром связывания
Вторым элементом композиции, соответствующей изобретению, является антиген или антигенная детерминанта, обладающая по меньшей мере одним вторым центром связывания, способным ассоциироваться с помощью по меньшей мере одной непептидной связи с первым центром связывания не природного молекулярного носителя. В изобретении представлены композиции, которые различаются в соответствии с антигеном или антигенной детерминантой, выбранными из соображений желательного терапевтического эффекта. Другие композиции получены путем изменения молекулы, выбранной для второго центра связывания.
Антигены, соответствующие изобретению, могут быть выбраны из группы, состоящей из следующих компонентов: (а) белки, пригодные для индукции иммунного ответа в отношении раковых клеток; (b) белки, пригодные для индукции иммунного ответа в отношении инфекционных болезней; (с) белки, пригодные для индукции иммунного ответа в отношении аллергенов, и (а) белки, пригодные для индукции иммунного ответа у сельскохозяйственных животных.
В одном специфическом варианте осуществления изобретения антиген или антигенная детерминанта представляет собой структуру, используемую для профилактики инфекционной болезни. Данный способ будет эффективным для лечения широкого ряда инфекционных болезней, поражающих широкий круг хозяев, например человека, корову, овцу, свинью, собаку, кошку, другие виды млекопитающих, а также немлекопитающих. Инфекционные болезни, которые лечат, хорошо известны компетентным специалистам в данной области, их примеры включают инфекции вирусной этиологии, такие как ВИЧ, грипп, герпес, вирусный гепатит, болезни, вызываемые вирусами Эпштейн-Барра и полиовирусами, вирусный энцефалит, корь, птичья оспа и т.п. или инфекции бактериальной этиологии, такие как пневмония, туберкулез, сифилис и т.п., или инфекции паразитарной этиологии, такие как малярия, трипаносомиоз, лейшманиоз, трохомониоз, амебиоз и т.п. Таким образом, антигены или антигенные детерминанты, выбранные для композиций, соответствующих изобретению, хорошо известны в области медицины; примеры антигенов или антигенных детерминант включают следующее: антигены ВИЧ gр 140 и gр 160, антигены гриппа гемагглютинин и нейраминидаза, поверхностный антиген гепатита В, белок циркумспорозоита возбудителя малярии.
В другом специфическом варианте осуществления композиции, соответствующие изобретению, являются иммунотерапевтическими, которые могут быть использованы для лечения аллергий и рака.
Выбор антигенов или антигенных детерминант для композиции и способ лечения аллергий должны быть известны компетентным специалистам в области медицины, касающейся лечения данных нарушений; репрезентативные примеры данного типа антигена или антигенной детерминанты включают следующее: фосфолипаза пчелиного яда A2, Bet v I (аллерген пыльцы березы), 5 Dol m V (аллерген яда осы пятнистой), Der p I (аллерген клещей домашней пыли).
Выбор антигенов или антигенных детерминант для композиции и способ лечения рака должны быть известны компетентным специалистам в области медицины, касающейся лечения данных нарушений; репрезентативные примеры данного типа антигена или антигенной детерминанты включают следующее: Неr2 (рак молочной железы), GD2 (нейробластома), EGF-R (злокачественная глиобластома), СЕА(медуллярный рак щитовидной железы), CD52 (лейкоз).
В определенном варианте осуществления изобретения антиген или антигенная детерминанта выбраны из группы, состоящей из: (а) рекомбинантного белка ВИЧ, (b) рекомбинантного белка вируса гриппа, (с) рекомбинантного белка вируса гепатита С, (d) рекомбинантного белка Toxoplasma, (e) рекомбинантного белка Plasmodium falciparum, (f) рекомбинантного белка Plasmodium vivax, (g) рекомбинантного белка Plasmodium ovale, (h) рекомбинантного белка Plasmodium malariae, (i) рекомбинантного белка клеток рака молочной железы, (j) рекомбинантного белка клеток рака почки, (k) рекомбинантного белка клеток рака простаты, (1) рекомбинантного белка клеток рака кожи, (m) рекомбинантного белка клеток рака головного мозга, (n) рекомбинантного белка лейкозных клеток, (о) рекомбинантного профилирования, (р) рекомбинантного белка аллергии на укус пчелы, (q) рекомбинантных белков аллергии на орехи, (r) рекомбинантных белков аллергий на пищевые продукты, рекомбинантных белков астмы и рекомбинантного белка Chlamydia.
После того как антиген или антигенная детерминанта композиции выбраны, по меньшей мере один второй центр связывания может быть введен в молекулу для конструирования организованной и содержащей повторы последовательности, связанной с не природным молекулярным носителем, соответствующим изобретению. Репрезентативные примеры вторых центров связывания включают без ограничения перечисленным антиген, антитело или фрагмент антитела, биотин, авидин, стрептавидин, рецептор, лиганд рецептора, лиганд, лиганд-связывающий белок, полипептид, взаимодействующий по типу лейциновой молнии, аминогруппу, химическую группу, реагирующую с аминогруппой, карбоксильную группу, химическую группу, реагирующую с карбоксильной группой, сульфгидрильную группу, химическую группу, реагирующую с сульфгидрильной группой, или их комбинацию.
Ассоциация между первым и вторым центрами связывания будет определяться характеристиками соответствующих выбранных молекул, но будет включать по меньшей мере одну непептидную связь. В зависимости от комбинации первого и второго центров связывания природа ассоциации может быть ковалентой, ионной, гидрофобной, полярной или их комбинацией.
В одном варианте изобретения второй центр связывания может быть представлен доменом белка лейциновой молнии FOS или доменом белка лейциновой молнии JUN.
В наиболее специфическом варианте осуществления изобретения выбранный второй центр связывания представляет собой домен белка лейциновой молнии FOS, который специфически ассоциирован с доменом белка лейциновой молнии JUN не природного молекулярного носителя, соответствующего изобретению. Ассоциация доменов белка лейциновой молнии FOS и JUN обеспечивает основу для образования организованной и содержащей повторы матрицы антигена или антигенной детерминанты на поверхности носителя. Домен белка лейциновой молнии FOS может быть слит в рамке считывания с выбранным антигеном или антигенной детерминантой по аминоконцу, карбоксильному концу, либо быть локализованным внутри белка, если это желательно.
Некоторые слитые конструкции FOS приведены в качестве примеров. Гормон роста человека (см. Пример 4), фосфолипаза A2 пчелиного яда (PLA) (см. Пример 9), овальбумин (см. Пример 10) и gр 140 ВИЧ (см. Пример 12).
Для упрощения создания слитых конструкций FOS описан ряд векторов, которые обеспечивают возможность выбора плана создания и конструкции антигенов и антигенных детерминант (см. Пример 6). Векторы pAV1-4 были разработаны для экспрессии слитых форм FOS в Е.соli; векторы pAV5 и pAV6 были разработаны для экспрессии слитых белков FOS в эукариотических клетках. Краткое описание свойств данных векторов приведено ниже:
1. pAVI: Данный вектор был разработан для секреции слитых белков, содержащих FOS на С-конце, в периплазматическое пространство Е.соli. Представляющий интерес ген (g.o.i.) может быть лигирован в сайты StuI/NotI вектора
2. рАV2: Данный вектор был разработан для секреции слитых белков, содержащих FOS на N-конце, в периплазматическое пространство Е. соli. Представляющий интерес ген (g.o.i.) может быть лигирован в сайты NotI/EcoRV (или Notl/Hindlll) вектора.
3. рАV3: Данный вектор был разработан для цитоплазматической продукции слитых белков, содержащих FOS на С-конце, в Е.соli. Представляющий интерес ген (g.o.i.) может быть лигирован в сайты EcoRV/NotI вектора.
4. рАV4: Данный вектор был разработан для цитоплазматической продукции слитых белков, содержащих FOS на N-конце, в Е.соli. Представляющий интерес ген (g.o.i.) может быть лигирован в сайты NotI/EcoRV (или NotI/Hindlll) вектора. N-концевой остаток метионина протеолитически удаляют при синтезе белка (см. статью HireI и соавт., Рсос. NatI. Acad. Sci. USA, 86: 8247-8251 (1989)).
5. рАV5: Данный вектор был разработан для эукариотической продукции слитых белков, содержащих FOS на С-конце. Представляющий интерес ген (g.o.i.) может быть введен между последовательностями, кодирующими сигнальную последовательность hGH и доменом FOS путем лигирования в сайты Eco47III/NotI вектора. Альтернативно ген, содержащий свою собственную сигнальную последовательность, может быть слит с участком, кодирующим FOS путем лигирования в сайты StuI/NotI.
6. DAV6: Данный вектор был разработан для эукариотической продукции слитых белков, содержащих FOS на N-конце. Представляющий интерес ген (g.o.i.) NotI/StuI (или NotI/HindIII) вектора.
Компетентным в данной области специалистам будет понятно, что конструкция слитого белка FOS-антиген или -антигенная детерминанта может включать дополнение в виде определенных генетических элементов для облегчения продукции рекомбинантного белка. Пример 4 представляет собой руководство по введению ряда элементов Е.соli для регуляции трансляции, а Пример 7 представляет собой руководство по введению эукариотической сигнальной последовательности. Могут быть выбраны другие генетические элементы в зависимости от специфических требований практикующего специалиста.
Изобретение, что также показано, включает продукцию слитого белка FOS-антиген или FOS-антигенная детерминанта либо в бактериальной (см. Пример 5), либо в эукариотической клетках (см. Пример 8). Выбор типа клетки для экспрессии слитого белка находится в сфере знания компетентного специалиста и зависит от таких факторов, как то, являются ли посттрансляционные модификации важным аспектом в создании композиции.
Как ранее замечено, изобретение раскрывает различные способы конструирования слитого белка FOS-антиген или FOS-антигенная детерминанта посредством использования векторов pAV. Кроме обеспечения экспрессии в прокариотах и эукариотах данные векторы позволяют практикующему специалисту выбрать между N- и С-концевым введением в антиген домена белка лейциновой молнии FOS. Представлены специфические примеры, в которых FOS сливают по N- и С-концам с PLA (см. Пример 9) и овальбумином (см. Пример 10). Пример 11 демонстрирует очистку слитых белков PLA- и овальбумин-FOS.
В наиболее специфическом варианте осуществления изобретение касается антигена или антигенной детерминанты, кодируемыми геномом ВИЧ. В частности, антиген ВИЧ представлен gр140. Как показано в Примерах 11-15, может быть получен gр140 с доменом белка лейциновой молнии FOS и синтезирован и очищен слитый белок для присоединения к не природному молекулярному носителю, соответствующему изобретению. Как известно компетентному специалисту в данной области, другие антигены и антигенные детерминанты ВИЧ могут быть использованы для создания композиции, соответствующей изобретению.
В наиболее специфическом варианте осуществления изобретения выбранный второй центр связывания является остатком цистеина, который специфически связывается с остатком лизина не природного молекулярного носителя, соответствующего изобретению. Химическая связь остатка лизина (Lys) и остатка цистеина (Cys) обеспечивает основу для образования организованной и содержащей повторы последовательности антигена или антигенной детерминанты на поверхности носителя. Остаток цистеина может быть инженерным способом введен в рамку считывания выбранного антигена или антигенной детерминанты по аминоконцу, карбоксильному концу либо быть локализованным внутри последовательности, если это желательно. В качестве примера приведены PLA и gр 140 ВИЧ с остатком цистеина для связывания с остатком лизина первого центра связывания.
С. Получение частиц альфа-вакцины
В изобретении представлены новые композиции и способы конструирования упорядоченных и содержащих повторы антигенных матриц. Как должен знать компетентный в данной области специалист, условия для сборки упорядоченной и содержащей повторы антигенной матрицы в значительной степени зависят от специфического выбора первого центра связывания не природного носителя и специфического выбора второго центра связывания антигена или антигенной детерминанты. Таким образом, выбор практикующего исследователя при создании композиции (т.е. выбор первого и второго центров связывания, антигена и не природного носителя) будет определять конкретные условия сборки частицы альфавакцины (комбинации упорядоченной и содержащей повторы антигенной матрицы и не природного молекулярного носителя). Информация, касающаяся сборки частицы альфавакцины, входит в профессиональные знания практикующего исследователя, и существует множество ссылок для помощи практикующему исследователю (например, см. монографию под ред. Sambrook J. и соавт., Лабораторное руководство по молекулярному (клонированию (Molecular Cloning, A Laboratory Manual), 2-е изд., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989); монографию под ред. Ausubel F. и соавт. Современные методы молекулярной биологии (Current Protocols in Molecular Biology), John H. Wiley & Sons, Inc. (1997); монографию под ред. Celis J. Биология клетки (Cell Biology), Academic Press, 2-е изд. (1998); монографию Harlow E. и Lane D. Лабораторное руководство по антителам (Antibodies: A Laboratory Manual,") Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. (1988), которые все включены в данном контексте в виде ссылки.
В конкретном варианте осуществления изобретения домены белка лейциновой молнии JUN и FOS используют соответственно как первый и второй центры связывания, соответствующие изобретению. При приготовлении частиц альфавакцины антиген следует получать и очищать в условиях, способствующих сборке упорядоченной и содержащей повторы антигенной матрицы на не природном носителе. В особом варианте домена белка лейциновой молнии JUN/FOS FOS-антиген или FOS-антигенную детерминанту следует обработать восстанавливающим агентом (например, дитиотреитолом (DTT)) для снижения или устранения возможности образования дисульфидных связей (см. Пример 15).
Для приготовления не природного носителя (т.е. рекомбинантного вируса Sinbis) в варианте осуществления с доменом белка лейциновой молнии JUN/FOS рекомбинантные вирусные частицы E2-JUN должны быть сконцентрированы, нейтрализованы и обработаны восстанавливающим агентом (см. Пример 16).
Сборку упорядоченной и содержащей повторы антигенной матрицы в варианте JUN/FOS осуществляют в присутствии окислительно-восстановительной пары. Вирусные частицы E2-JUN смешивают с 240-кратным молярным избытком FOS-антигена или FOS-антигенной детерминанты в течение 10 часов при 4°С. Затем частицы альфа-вакцины концентрируют и очищают с помощью хроматографии (см. Пример 16).
В другом варианте осуществления изобретения связывание не природного молекулярного носителя с антигеном или антигенной детерминантой может быть выполнено путем химического перекрестного сшивания. В наиболее предпочтительном варианте осуществления химический агент представлен гетеробифункциональным перекрестно-сшивающим агентом, таким как N-гидроксисукцинимидный сложный эфир ∈-малеимидкапроновой кислоты (см. статьи Tanimori и соавт., J. Pharm. Dyn., 4: 812 (1981); Fujiwara и соавт., J. Immunol. Meth., 45: 195 (1981)), который содержит сукцинимидную группу, реагирующую с аминогруппами, и (2) малеимидную группу, реагирующую с SH-группами. Гетерологичный белок или полипептид первого центра связывания можно сконструировать так, чтобы он содержал один или более остатков лизина, которые будут служить в качестве реакционной части для сукцинимидной группы гетеробифункционального перекрестно-сшивающего агента. После завершения химического связывания первых центров связывания не природного молекулярного носителя малеимидная группа гетеробифункционального перекрестно-сшивающего агента будет доступной для реакции с SH-группой остатка цистеина на антигене или антигенной детерминанте. Получение антигена или антигенной детерминанты в данном случае может требовать конструирования остатка цистеина, введенного в белок или полипептид, выбранный в качестве второго центра связывания так, чтобы он мог вступать в реакцию со свободной малеимидной группой на перекрестно-сшивающем агенте, который связан с первыми центрами связывания не природного молекулярного носителя.
3. Композиции, вакцины и их применение, а также способы лечения
В одном из вариантов осуществления в изобретении представлены вакцины для профилактики инфекционных болезней у широкого круга видов, в частности видов млекопитающих, таких как человек, обезьяна, корова, собака, кошка, лошадь, свинья и т.п. Могут быть разработаны вакцины для лечения инфекций вирусной этиологии, таких как ВИЧ, грипп, герпес, вирусный гепатит, болезни, вызываемые вирусами Эпштейн-Барра и полиовирусами, вирусный энцефалит, корь, птичья оспа и т.п., или инфекций бактериальной этиологии, таких как пневмония, туберкулез, сифилис и т.п., или инфекций паразитарной этиологии, таких как малярия, трипаносомиоз, лейшманиоз, трихомониоз, амебиоз и т.п.
В другом варианте осуществления в изобретении представлены вакцины для профилактики рака у широкого круга видов, в частности видов млекопитающих, таких как человек, обезьяна, корова, собака, кошка, лошадь, свинья и т.п. Могут быть разработаны вакцины для лечения всех типов злокачественных новообразований: лимфом, карцином, сарком, меланом и т.п.
В другом варианте осуществления изобретения композиции, соответствующие изобретению, могут быть использованы для разработки вакцин для лечения аллергий. Антитела изотипа IgE являются важными компонентами аллергических реакций. Мастоциты связывают антитела IgE на своей поверхности и высвобождают гистамины и другие медиаторы аллергической реакции при связывании специфического антигена с молекулами IgE, которые связаны с поверхностью мастоцитов. Ингибирование образования антител IgE вследствие этого является перспективной целью в плане защиты от аллергий. Это было бы возможным при получении ответа Т-клеток-хелперов. Ответы Т-клеток-хелперов можно разделить на ответы Т-клеток-хелперов типа 1 (ТH1) и типа 2 (ТH2) (см. статью Romagnani, Immunol. Today, 18:263-266 (1997)). Клетки ТH1 секретируют γ-интерферон и другие цитокины, которые запускают продукцию антител lgGI-3 в В-клетках.
Напротив, критический цитокин, образуемый клетками ТH2, представлен ИЛ-4 (интерлейкином-4), который направляет продукцию lgG4 и IgE в В-клетках. Во многих экспериментальных системах появление ответов ТH1 и ТH2 является взаимоисключающим, поскольку клетки ТH1 супрессируют индукцию клеток ТH2 и наоборот. Таким образом антигены, которые запускают сильный ответ ТH1, одновременно супрессируют развитие ответов ТH2 и, следовательно, продукцию антител IgE. Интересно, что в действительности все вирусы индуцируют ответ ТH1 в хозяине и не вызывают продукцию антител IgE (см. статью Coutelier и соавт., J. Exp. Med., 165: 64-69 (1987)). Данный образец изотипа не ограничен живыми вирусами, но был отмечен для инактивированных или рекомбинантных вирусных частиц (см. статью Lo-Man и соавт., Eur. J. Immunol., 28: 1401-1407 (1998)). Таким образом, используя процессы, соответствующие изобретению (например, технологию альфа-вакцин), на вирусные частицы можно поместить различные аллергены и использовать для иммунизации. Благодаря полученной "вирусной структуре" аллергена будет появляться ответ ТH1, будут образовываться "защитные" антитела lgG1-3, что будет препятствовать продукции антител IgE, которые вызывают аллергические реакции. Поскольку аллерген презентируется вирусными частицами, которые распознаются другим набором Т-клеток-хелперов, чем сам аллерген, вероятно, что аллерген-специфические антитела lgG1-3 будут индуцироваться даже у больных аллергией субъектов, несущих предварительно существовавшие клетки ТH2, специфические в отношении аллергена. Присутствие высоких концентраций антител IgG может препятствовать связыванию аллергенов с IgE, связанных с мастоцитами, ингибируя таким образом выход гистамина.
Вследствие этого присутствие антител IgG может защищать от lgE-опосредованных аллергических реакций. Типичные субстанции, вызывающие аллергию, включают пыльцу злаков, амброзии, березы или можжевельника мексиканского, домашнюю пыль, клещей, перхоть животных, плесень, яд насекомых или лекарственные вещества (например, пенициллин). Таким образом, иммунизация субъектов вирусными частицами, на которые помещены аллергены, должна быть эффективной не только перед, но и после появления аллергий.
Специалистам в данной области следует понимать, что, когда композиции, соответствующие изобретению, вводят субъекту, они могут находиться в препарате, который содержит соли, буферы, адъюванты или другие субстанции, которые являются желательными для повышения эффективности композиции. Примеры материалов, пригодных для применения при изготовлении фармацевтических композиций, приведены в многочисленных источниках, включая Справочник по фармацевтическим наукам Ремингтона (Remington’s Pharmaceutical Sciences (под ред. Osol А., Mack Publishing Co., (1980)).
Считают, что композиции, соответствующие изобретению, являются "фармакологически приемлемыми", если их введение может переноситься субъектом-реципиентом. Кроме того, композиции, соответствующие изобретению, будут вводиться в терапевтически эффективном количестве (т.е. в количестве, которое создает желательный физиологический эффект).
Композиции, соответствующие данному изобретению, могут вводиться различными способами, известными в уровне техники, но в норме будут вводиться путем инъекции, инфузии, ингаляции, перорально или другими подходящими физическими способами. Композиции могут альтернативно вводиться внутримышечно, внутривенно или подкожно. Компоненты композиций для введения включают стерильные водные (например, физиологический раствор) или неводные растворы и суспензии. Примерами неводных растворителей являются пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло, и инъекционные органические сложные эфиры, такие как этилолеат. Носители или окклюзионные повязки можно использовать для повышения проницаемости кожи и повышения всасывания антигена.
В дополнение к вакцинным технологиям в других вариантах осуществления изобретения представлены способы медицинского лечения рака и аллергий.
Все патенты и публикации, представленные в данном контексте, специально включены в виде ссылки.
Примеры
Ферменты и реагенты, используемые в последующих примерах, включают: ДНК-лигазу Т4, приобретаемую в фирме New England Biolabs; ДНК-полимеразуТао, набор плазмид QlАргер Spin Plasmid Kit, набор средних плазмид QIAGEN plasmid Midi Kit, набор для гель-экстракции QiaExll Gel Extraction Kit, набор для очистки при ПЦР (полимеразная цепная реакция) QIAquick PCR Purification Kit, приобретаемые в фирме QIAGEN; набор для очистки MPHK QuickPrep Micro mRNA Purification Kit, приобретаемый в фирме Pharmacia; набор для одностадийной ОТ-ПЦР (полимеразная цепная реакция с использованием обратной транскриптазы) Superscript One-step RT PCR Kit, сыворотка телячьих эмбрионов (PCS), бактотриптон и дрожжевой экстракт, приобретаемые в фирме Gibco BRL; олигонуклеотиды, приобретаемые в фирме Microsynth (Швейцария); рестриктазы, приобретаемые в фирме Boehringer Mannheim, New England Biolabs или МВI Fermentas; полимераза Pwo и dNTP (дезоксинуклеозидтрифосфаты), приобретаемые в фирме Boehringer Mannheim. Среду HP-1 приобретают в фирме CeII culture technologies (Glattbrugg, Швейцария). Все стандартные химические реактивы приобретают в фирме Fluka-Sigma-Aldrich, а все материалы, связанные с культурой клеток, приобретают в фирме ТРР.
Манипуляции с ДНК проводят, используя стандартные технологии. ДНК готовят в соответствии с инструкциями изготовителя либо из 2 мл бактериальной культуры, используя набор QIAprep Spin Plasmid Kit или из 50 мл культуры, используя набор QIAGEN Plasmid Midi Kit. Для разрезания рестриктазой ДНК инкубируют в течение по меньшей мере 2 часов с соответствующей рестриктазой в концентрации 5-10 единиц (U) фермента/мг ДНК в рекомендуемых изготовителем условиях (буфер и температура). Реакции разрезания более чем одним ферментом проводят одновременно, если условия реакции являются подходящими для всех ферментов, в другом случае реакции проводят последовательно. Фрагменты ДНК, выделенные для дальнейших манипуляций, разделяют путем электрофореза в 0,7-1,5% агарозном геле, вырезают из геля и очищают, используя набор для экстракции из геля QiaExll Gel Extraction Kit в соответствии с инструкциями, представленными производителем. Для лигирования фрагментов ДНК 100-200 пг очищенной векторной ДНК инкубируют в течение ночи с трехкратным молярным избытком фрагмента инсерции при 16°С в присутствии 1 ед. ДНК-лигазы Т4 в буфере, приобретенном у изготовителя (общий объем составляет 10-20 мкл). Аликвоту (0,1-0,5 мкл) реакции лигирования используют для трансформации Е.соli XL1-Blue (Stratagene). Трансформацию выполняют путем электропорации, используя пульсирующее устройство для введения гена Gene Pulser (BioRAD) и кюветы для Gene Pulser 0,1 см (BioRAD) при 200 ом, 25 мкф, 1,7 кВ. После электропорации клетки инкубируют при встряхивании в течение 1 час в 1 мл среды S.O.B. (см. работу Miller, 1972) перед тем, как поместить их на селективный агар S.O.B.
Пример 1:
Инсерция амфипатического спирального домена JUN в Е2
В вектор pTE5’2J (см. статью Hahn и соавт., Proc. NatI. Acad. Sci. USA, 89: 2679-2683, (1992)), интродуцируют сайты рестриктаз MluI и BstEII между кодонами 71 (Gln) и 74 (Thr) последовательности, кодирующей структурный белок Е2, что приводит к образованию вектора pTE5’2JBM. Интродукцию данных сайтов рестриктаз осуществляют с помощью ПЦР, используя следующие олигонуклеотиды:
Олигонуклеотид 1:
E2insBstEII/BssHII:
5’-ggggACGCGTGCAGCAggtaaccaccgTTAAAGAAGGCACC-3’ (SEQ ID NO:1) Олигонуклеотид 2:
E2insMlulStul:
5’-cggtggttaccTGCTGCACGCGTTGCTTAAGCGACATGTAGCGG-3’ (SEQ ID NO:2) Олигонуклеотид 3:
E2insStuI:
5’-CCATGAGGCCTACGATACCC-3’ (SEQ ID NO:3)
Олигонуклеотид 4:
E2insBssHII:
5’-GGCACTCACGGCGCGCTTTACAGGC-3’ (SEQ ID NO:4)
Для ПЦР используют 100 пмоль каждого олигонуклеотида с 5 нг матричной ДНК в 100 мкл реакционной смеси, содержащей 4 единицы полимеразы Taq или Pwo, 0,1 мМ dNTP и 1,5 мМ MgSO4. Все концентрации ДНК определяют фотометрически, используя устройство GeneQuant (Pharmacia). Полимеразу добавляют непосредственно перед началом ПЦР (исходная точка - 95°С). Температурный циклинг осуществляют следующим образом и в следующем порядке: 95°С в течение 2 минут; 5 циклов, состоящих из стадий: 95°С (45 сек), 53°С (60 сек), 72°С (80 сек); и 25 циклов, состоящих из стадий: 95°С (45 сек), 57°С (60 сек), 72°С (80 сек).
Два фрагмента, полученные при ПЦР, анализируют и очищают гель-электрофорезом с использованием агарозы. Для получения конечной конструкции проводят сборку с помощью ПЦР двух полученных при ПЦР фрагментов, используя олигонуклеотиды 3 и 4 для амплификации.
Для сборки с помощью ПЦР используют 100 пмоль каждого олигонуклеотида с 2 нг очищенных фрагментов, полученных при ПЦР, в 200 мкл реакционной смеси, содержащей 4 ед. полимеразы Taq или Pwo, 0,1 мМ dNTP и 1,5 мМ MgSO4. Все концентрации ДНК определяют фотометрически, используя устройство GeneQuant (Pharmacia). Полимеразу добавляют непосредственно перед началом ПЦР (исходная точка - 95°С). Температурный циклинг осуществляют следующим образом и в следующем порядке: 95°С в течение 2 минут; 5 циклов, состоящих из следующих стадий: 95°С (45 сек), 57°С (60 сек), 72°С (90 сек); и 25 циклов, состоящих из следующих стадий: 95°С (45 сек), 59°С (60 сек), 72°С (90 сек).
Конечный продукт ПЦР очищают, используя колонки для ПЦР Qia spin (Qiagen), и разрезают в подходящем буфере, используя по 10 ед. рестриктаз BssHII и StuI, в течение 12 часов при 37°С. Фрагменты ДНК очищают гель-электрофорезом и лигируют в разрезанный BssHII/StuI и очищенный гель-электрофорезом вектор pTE5’2J (см. статью Hahn и соавт., Proc. Natl. Acad. Sci. USA, 89:2679-2683 (1992)). Правильность инсерции продукта ПЦР сначала анализируют с помощью рестрикционного анализа с использованием BstEII and MIuI, а затем путем секвенирования ДНК фрагмента, полученного при ПЦР.
Последовательность ДНК, кодирующую домен амфипатической спирали JUN, амплифицируют с помощью ПЦР из вектора pJuFo (см. статью Crameri и Suter, Gene, 137:69 (1993)), используя следующие олигонуклеотиды:
Олигонуклеотид 5:
JUNBstEII:
5’-CCTTCTTTAAcggtggttaccTGCTGGCAACCAACGTGGTTCATGAC-3’ (SEQ ID NO:5) Олигонуклеотид 6:
MlulJUN:
5’-AAGCATGCTGCacgcgtgTGCGGTGGTCGGATCGCCCGGC-3’ (SEQ ID NO:6)
Для ПЦР используют 100 пмоль каждого олигонуклеотида с 5 нг матричной ДНК в 100 мкл реакционной смеси, содержащей 4 единицы полимеразы Taq или Pwo, 0,1 мМ dNTP и 1,5 мМ MgSO4. Все концентрации ДНК определяют фотометрически, используя устройство GeneQuant (Pharmacia). Полимеразу добавляют непосредственно перед началом ПЦР (исходная точка - 95°С). Температурный циклинг осуществляют следующим образом и в следующем порядке: 95°С в течение 2 минут; 5 циклов, состоящих из следующих стадий: 95°С (45 сек), 60°С (30 сек), 72°С (25 сек.); и 25 циклов, состоящих из следующих стадий: 95°С (45 сек), 68°С (30 сек), 72°С (20 сек).
Конечный продукт ПЦР очищают гель-электрофорезом и лигируют в разрезанный и очищенный гель-электрофорезом pBluescript II(KS’). Из полученного вектора выделяют последовательность JUN путем разрезания MluI/BstEII, очищают с помощью QiaExII и лигируют в вектор pTE5’2JBM (предварительно разрезанный теми же самыми рестриктазами) для получения вектора pTE5’2J:E2JUN.
Пример 2:
Получение вирусных частиц, содержащих E2-JUN с использованием системы pCYTts
Структурные белки амплифицируют с помощью ПЦР, используя pTE5’2J:E2JUN в качестве матрицы и олигонуклеотиды XbaIStruct (ctatcaTCTAGAATGAATAGAGGATTCTTTAAC) (SEQ ID NO: 12) и StructBsp-1201 (tcgaatGGGCCCTCATCTTCGTGTGCTAGTCAG) (SEQ ID NO: 87). Для ПЦР используют 100 пмоль каждого олигонуклеотида и 5 нг матричной ДНК в 100 мкл реакционной смеси, содержащей 4 единицы полимеразы Taq или Pwo, 0,1 мМ dNTP и 1,5 мМ MgSO4. Все концентрации ДНК определяют фотометрически, используя устройство GeneQuant (Pharmacia). Полимеразу добавляют непосредственно перед началом ПЦР (исходная точка - 95°С). Температурный циклинг осуществляют следующим образом: 95°С в течение 3 минут c последующими 5 циклами, состоящими из следующих стадий: 93°С (35 сек), 54°С (35 сек), 72°С (270 сек), и последующими 25 циклами, состоящими из следующих стадий: 92°С (30 сек), 63°С (35 сек), 72°С (270 сек). Продукт ПЦР очищают гель-электрофорезом и разрезают рестриктазами XbaI/Bsp1201 и лигируют в вектор pCYTts, предварительно разрезанный теми же ферментами (см. Патентную заявку США No.60/079562, поданную 27 марта 1998 г.).
Двадцать мкг pCYTtsE2:JUN инкубируют с 20 ед. ScaI в подходящем буфере в течение по меньшей мере 4 час при 37°С. Реакцию прекращают экстракцией фенолом/хлороформом с последующим осаждением изопропанолом линеаризованной ДНК. Реакцию рестрикции проверяют гель-элетрофорезом в агарозе. Для трансфекции 5,4 мкг линеаризованного pCYTtsE2:JUN смешивают с 0,6 мкг линеаризованного pSV2Neo в 30 мкл H2O и добавляют 30 мкл 1 М раствора СаСl2. После добавления 60 мкл фосфатного буфера (50 мМ HEPES, 280 мМ NaCl, 1,5 мМ Na2HPO4, pH 7,05) раствор взбалтывают в течение 5 секунд, а затем инкубируют при комнатной температуре в течение 25 секунд. Раствор немедленно добавляют к 2 мл среды НР-1, содержащей 2% FCS (2% среда FCS). Среду конфлюэнтной на 80% культуры клеток ВНК21 в 6-луночной плате затем замещают ДНК-содержащей средой. После инкубирования в течение 5 часов при 37°С в инкубаторе с СО2 ДНК-содержащую среду удаляют и замещают 2 мл 15% глицерина в 2% среде FCS. Глицерин-содержащую среду удаляют после 30-секундной фазы инкубирования и промывают клетки 5 мл среды НР-1, содержащей 10% FCS. Наконец, добавляют 2 мл свежей среды НР-1, содержащей 10% FCS. Отбирают стабильно трансфицированные клетки и выращивают на селективной среде (НР-1 с добавлением G418) при 37°С в инкубаторе с СО2. Когда смешанная популяция вырастает до конфлюэнтности, культуру делят на две чашки с последующим периодом роста в течение 12 час при 37°С. Одну чашку с клетками переносят в условия культивирования при 30°С для индукции экспрессии вирусных частиц, другую чашку сохраняют при 37°С.
Экспрессию вирусных частиц определяют с помощью вестерн-блоттинга (см. Фигуру 1). Культуральную среду (0,5 мл) осаждают метанолом/хлороформом и ресуспендируют осадок в буфере для образцов для SDS-PAGE. Образцы нагревают в течение 5 минут при 95°С перед внесением в 15% акриламидный гель. После SDS-PAGE белки переносят на нитроцеллюлозные мембраны Protan (Schleicher & Schuell, Germany), как предложено в разделе, написанном Bass и Yang, в монографии под ред. Creighton Т.Е. Практические подходы к функции белка (Protein Function: A Practical Approach), 2-е изд., IRL Press, Oxford (1997), стр.29-55. Мембрану блокируют 1% бычьим альбумином (Sigma) в TBS (10 × TBS/л: 87,7 г NaCl, 66,1 r Trizma гидрохлорида (Sigma) и 9,7 r Trizma основания (Sigma), рН 7,4) в течение 1 часа при комнатной температуре с последующим инкубированием с антителом против EI/E2 (поликлональная сыворотка) в течение 1 часа. Блот промывают 3 раза по 10 минут с помощью TBS-T (TBS с добавлением 0,05% Tween 20) и инкубируют в течение 1 часа с конъюгатом щелочной фосфатазы с IgG против кролика (0,1 мкг/мл, Amersham Life Science, England). После промывания 2 раза в течение 10 минут в TBS-T и 2 раза по 10 минут в TBS проводят проявляющую реакцию с использованием реагентов для детекции щелочной фосфатазы (10 мл буфера АР (100 мМ Трис/HCl, 100 мМ NaCl, pH 9,5) с 50 мкл раствора NBT (7,7% нитро синего тетразолия (Sigma) в 70% диметилформамиде) и 37 мкл раствора Х-фосфата (5% 5-бром-4-хлор-3-индолилфосфата в диметилформамиде).
Получение вирусных частиц представлено на Фигуре 1. Образец вестерн-блоттинга показывает, что E2-JUN (дорожка 1) мигрирует в направлении более высокой молекулярной массы при SDS-PAGE по сравнению с Е2 дикого типа (дорожка 2) и линия клеток-хозяев ВНК21 не дает никакого фона.
Пример 3:
Получение вирусных частиц, содержащих E2-JUN, с использованием вектора pTE5’2JE2:JUN
Не содержащий РНКазу вектор (1,0 мкг) линеаризуют разрезанием с помощью Pvul. Затем проводят транскрипцию in vitro, используя набор для транскрипции in vitro SP6 (InvitroscripCAP, изготавливаемый InvitroGen, Invitrogen BV, NVLeek, Netherlands). Полученную мРНКс5’-кэпом анализируют в восстанавливающем агарозном геле.
Транскрибированную in vitro мРНК (5 мкг) электропорацией вводят в клетки ВНК21 (АТСС: CCL10) в соответствии с инструкцией Invitrogen (Экспрессионная система Sindbis (Sindbis Expression system), Invitrogen BV, Netherlands). После инкубирования в течение 10 час при 37°С среду, содержащую FCS, заменяют средой НР-1, не содержащей FCS, с последующим дополнительным инкубированием при 37°С в течение 10 час. Супернатант собирают и анализируют с помощью вестерн-блоттинга на образование вирусных частиц точно так же, как описано в Примере 2.
Полученный результат идентичен результату, полученному с использованием pCYTtsE2:JUN, как представлено на Фигуре 2.
Пример 4:
Слияние гормона роста человека (hGH) с доменом лейциновой молнии FOS (сигнальная последовательность ОmрА)
Ген hGH без человеческой лидерной последовательности амплифицируют из оригинальной плазмиды (АТСС 31389) с помощью ПЦР. Олигонуклеотид 7 с внутренним сайтом XbaI создают для отжига гена hGH по 5’-концу и олигонуклеотид 9 с внутренним сайтом EcoRI примируют по 3’-концу гена hGH. Для ПЦР используют 100 пмоль каждого олигонуклеотида и 5 нг матричной ДНК в 75 мкл реакционной смеси (4 единицы полимеразы Taq или Pwo, 0,1 мМ dNTP и 1,5 мМ MgSO4).
ПЦР-циклинг осуществляют следующим образом: 30 циклов при температуре отжига 60°С и время элонгации 1 мин при 72°С.
Очищенный гель-электрофорезом и выделенный продукт ПЦР используют в качестве матрицы для второй ПЦР с целью интродукции сигнальной последовательности оmрА и последовательности Шайна-Дальгарно (Shine-Dalgarno). Для ПЦР используют 100 пмоль олигонуклеотидов 8 и 9 и 1 нг полученного при ПЦР фрагмента-матрицы в 75 мкл реакционной смеси (4 единицы полимеразы Taq или Pwo, 0,1 мМ dNTP и 1,5 мМ MgSO4). Температура отжига для первых пяти циклов составляет 55°С при времени элонгации 60 сек при 72°С, другие 25 циклов проводят при температуре отжига 65°С и времени элонгации 60 сек при 72°С.
Олигонуклеотид 7:
gggtctagattcccaaccattcccttatccaggctttttgacaacgctatgctccgcgcccatcgtctgcaccag
ctggcctttgacacc (SEQ ID NO:7)
Олигонуклеотид 8:
gggtctagaaggaggtaaaaaacgatgaaaaagacagctatcgcgattgcagtggcactggctggtttcgct accgtagcgcaggccttcccaaccattcccttatcc (SEQ ID NO:8)
Олигонуклеотид 9:
cccgaattcctagaagccacagctgccctcc (SEQ ID NO:9)
Полученный рекомбинантный ген hGH субклонируют в pBluescript с помощью XbaI/EcoRI. Правильность последовательностей обеих цепей подтверждают секвенированием ДНК.
Последовательность ДНК, кодирующую домен амфипатической спирали FOS, амплифицируют с помощью ПЦР из вектора pJuFo (см. статью Crameri & Suter, Gene, 137:69 (1993)), используя олигонуклеотиды:
omp-FOS:
5’- ccTGCGGTGGTCTGACCGACACCC-3’ (SEQ ID NO: 10)
FOS-hgh:
5’- ccgcggaagagccaccGCAACCACCGTGTGCCGCCAGGATG-3’ (SEQ ID NO:11).
Для ПЦР используют 100 пмоль каждого олигонуклеотида и 5 нг матричной ДНК в 75 мкл реакционной смеси (4 единицы полимеразы Taq или Pwo, 0,1 мМ dNTP и 1,5 мМ MgSO4). Выполняют следующие температурные циклы: 95°С в течение 2 минут с последующими 5 циклами, состоящими из следующих стадий: 95°С (45 сек), 60°С (30 сек), 72°С (25 сек), и последующими 25 циклами, состоящими из следующих стадий: 95°С (45 сек), 68°С (30 сек), 72°С (20 сек).
Продукт ПЦР очищают, выделяют и клонируют в разрезанный с помощью StuI pBluescript-ompA-hGH. Гибридный ген затем клонируют в плазмиду pKK223-3 (Pharmacia).
Пример 5:
Бактериальная экспрессия FOS-hGH
ompA-FOS-hGH в pkk223-3 экпрессируют под контролем индуцируемого промотора, используя JM101 в качестве штамма Е. соli-хозяина. Экспрессию осуществляют в колбе для встряхивания. Клетки индуцируют 1 мМ IPTG (изопропилтиогалактозид) (конечная концентрация) при OD (оптическая плотность) 600 при 0,5. Экспрессию продолжают в течение 10 часов при 37°С. Клетки собирают центрифугированием при 3600×g при 10°С в течение 15 мин Осадок клеток замораживают (-20°С или жидким N2) и хранят в течение 16 час. Затем осадок оттаивают при 4°С и ресуспендируют в 10 мл 10 мМ Трис- HCl, рН 7,4, содержащем 600 мМ сахарозы. После перемешивания в течение 15 мин при 4°С периплазматические белки освобождают с помощью процедуры осмотического шока. Добавляют охлажденную (4°С) деионизированную воду и перемешивают суспензию в течение 30 мин при 4°С. Осадок разводят, ресуспендируют и добавляют лизоцим для разложения клеточной стенки бактерий. Клетки и сферопласты периплазматической фракции отделяют центрифугированием в течение 20 мин при 11000×g при 4°С. Содержащий FOS-hGH супернатант анализируют с помощью SDS-PAGE в восстанавливающих и невосстанавливающих условиях и дот-блоттинга. Дот-блоттинг проводят, как описано в Примере 8, используя антитело против hGH (Sigma) в качестве первого антитела и конъюгат щелочная фосфатаза (АР)-антитело против мыши в качестве второго антитела.
Правильно процессированный FOS-hGH полной длины можно определять в восстанавливающих и невосстанавливающих условиях. Часть FOS-hGH связывается с другими неидентифицированными белками с помощью свободных цистеинов, присутствующих на амфипатической спирали FOS. Однако более 50% экспрессированного FOS-hGH находится в своей нативной мономерной конформации (см. Фигуру 3).
Очищенный FOS-hGH будет использован для проведения первых экспериментов по введению JUN-содержащих вирусных частиц.
Пример 6:
Конструирование серий векторов pAV для экспрессии слитых белков FOS
Конструируют многофункциональную векторную систему, которая обеспечивает либо цитоплазматическую продукцию, либо секрецию белков, слитых с FOS по N- или С-концам, в Е.соli, либо продукцию белков, слитых с FOS по N- или С-концам, в эукариотических клетках. Векторы pAV1 - pAV4, созданные для продукции слитых с FOS белков в Е.соli, охватывают нижеперечисленные кассеты ДНК, которые содержат следующие генетические элементы в следующем порядке: (а) сильный сайт связывания рибосом и 5’-нетранслируемый участок, выделенные из гена Е.соli оmрА (aggaggtaaaaaacg) (SEQ ID NO: 13); (b) последовательность, кодирующую сигнальный пептид белка наружной мембраны Е.соli оmрА (MKKTAIAIAVALAGFATVAQA) (SEQ ID NO: 14); (с) последовательность, кодирующую домен димеризации FOS, фланкированную с обеих сторон остатками глицина и остатком цистеина (CGGLTDTLQAETDQVEDEKSALQTEIANLLKEKEKLEFILAAHGGC) (SEQ ID NO: 15), и (d) участок, кодирующий короткий пептидный линкер (AAASGG (SEQ ID NO: 16) или GGSAAA (SEQ ID NO: 17)), связывающий представляющий интерес белок с доменом димеризации FOS. Соответствующие кодирующие участки даны прописными буквами. Организация сайтов рестрикции обеспечивает легкость конструирования слитых с FOS генов, содержащих или не содержащих сигнальную последовательность. Кассеты клонируют в сайт рестрикции EcoRI/HindIII экспрессионного вектора рКК.223-3 (Pharmacia) с целью экспрессии слитых генов под контролем сильного промотора tac.
pAVI
Данный вектор разрабатывают для секреции слитых белков, содержащих FOS на С-конце, в периплазматическое пространство Е.соli. Представляющий интерес ген (g.o.i.) можно лигировать в сайты StuI/NotI вектора.

pAV2
Данный вектор разрабатывают для секреции слитых белков, содержащих FOS на N-конце, в периплазматическое пространство Е.соli. Представляющий интерес ген (g.o.i.) можно лигировать в сайты NotI/EcoRV (или Notl/Hindlll) вектора.

рАV3
Данный вектор разрабатывают для цитоплазматической продукции слитых белков, содержащих FOS на С-конце, в Е.соli. Представляющий интерес ген (g.o.i.) можно лигировать в сайты EcoRV/NotI вектора.
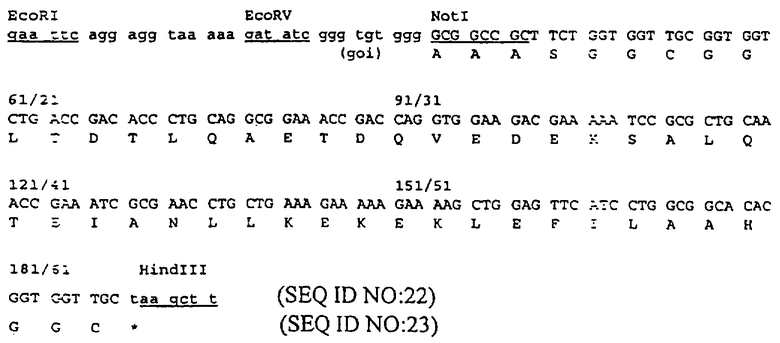
рАV4
Данный вектор разрабатывают для цитоплазматической продукции слитых белков, содержащих FOS на N-конце, в Е.coil. Представляющий интерес ген (g.o.i.) можно лигировать в сайты NotI/EcoRV (или NotI/HindIII) вектоpa. N-концевой остаток метионина удаляют протеолитически при синтезе белка (см. статью Hirel и соавт. Proc. NatI. Acad. Sci. USA 86: 8247-8251 (1989)).
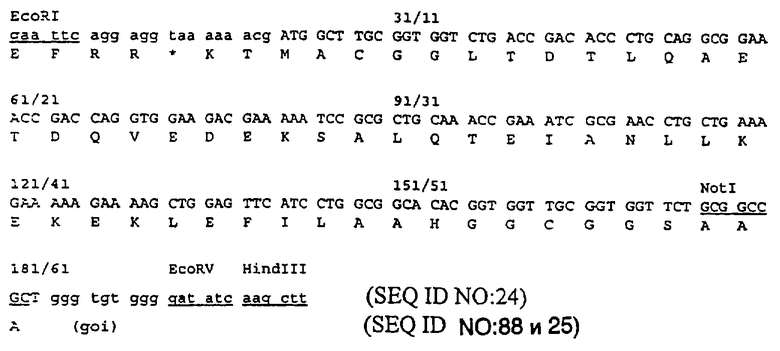
Векторы pAV5 и pAV6, которые разрабатывают для эукариотической продукции слитых с FOS белков, охватывают следующие генетические элементы, расположенные в различном порядке: (а) участок, кодирующий лидерный пептид человеческого гормона роста (MATGSRTSLLLAFGLLCLPWLQEGSA) (SEQ ID NO: 26); (b) последовательность, кодирующую домен димеризации FOS, фланкированный с обеих сторон двумя остатками глицина и остатком цистеина (CGGLTDTLQAETDQVEDEKSALQTEIANLLKEKEKLEFILAAHGGC) (SEQ ID NO: 15); и (с) участок, кодирующий короткий пептидный линкер (AAASGG (SEQ ID NO: 16) или GGSAAA (SEQ ID NO: 17)), связывающий представляющий интерес белок с доменом димеризации FOS. Соответствующие кодирующие участки даны прописными буквами. Организация сайтов рестрикции обеспечивает легкость конструирования слитых с FOS генов. Кассеты клонируют в сайт рестрикции EcoRI/HindIII экспрессионного вектора pMPSVEH (см. статью Artelt и соавт., Gene, 68:213-219 (1988)).
pAV5
Данный вектор разрабатывают для эукариотической продукции слитых белков, содержащих FOS на С-конце. Представляющий интерес ген (g.o.i.) можно ввести между последовательностями, кодирующими сигнальную последовательность hGH и домен FOS, путем лигирования в сайты Eco47III/NotI вектора. Альтернативно ген, содержащий свою собственную сигнальную последовательность, можно слить с участком, кодирующим FOS, путем лигирования в сайты StuI/NotI.

рАV6
Данный вектор разрабатывают для эукариотической продукции слитых белков, содержащих FOS на N-конце. Представляющий интерес ген (g.o.i.) можно лигировать в сайты NotI/StuI (или NotI/HindIII) вектора.
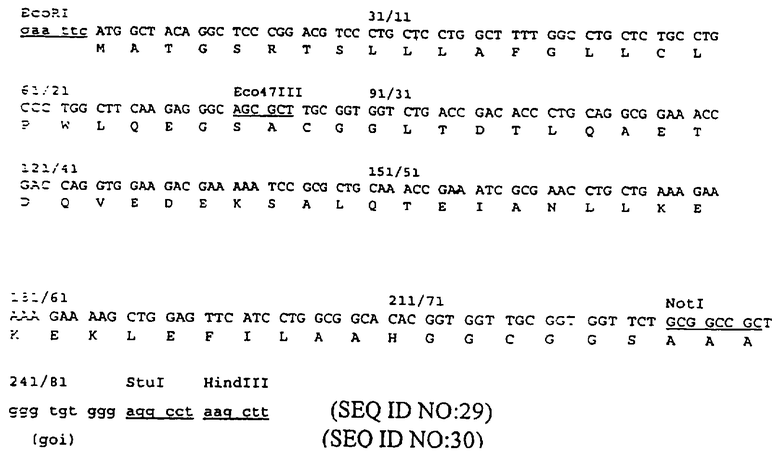
Конструирование экспрессионных векторов pAVI-pAV6
Для конструирования экспрессионных векторов pAVI-pAV6 синтезируют следующие олигонуклеотиды:
FOS-FOR1:
CCTGGGTGGGGGCGGCCGCTTCTGGTGGTTGCGGTGGTCTGACC(SEQ IDNO: 31);
FOS-FOR2:
GGTGGGAATTCAGGAGGTAAAAAGATATCGGGTGTGGGGCGGCC (SEQ ID NO: 32);
FOS-FOR3:
GGTGGGAATTCAGGAGGTAAAAAACGATGGCTTGCGGTGGTCTGACC (SEQ ID NO: 33);
FOS-FOR4:
GCTTGCGGTGGTCTGACC (SEQ ID NO: 34);
FOS-REV1:
CCACCAAGCTTAGCAACCACCGTGTGC (SEQ ID NO: 35);
FOS-REV2:
CCACCAAGCTTGATATCCCCACACCCAGCGGCCGCAGAACCACCGCAACCACCG (SEQ ID NO: 36);
FOS-REV3:
CCACCAAGCTTAGGCCTCCCACACCCAGCGGC (SEQ ID NO: 37);
OmpA-FOR-1:
GGTGGGAATTCAGGAGGTAAAAAACGATG (SEQ ID NO: 38);
hGH-FOR1:
GGTGGGAATTCAGGCCTATGGCTACAGGCTCC (SEQ ID NO: 39) и
hGH-FOR2:
GGTGGGAATTCATGGCTACAGGCTCCC (SEQ ID NO: 40).
Для конструирования вектора рАV2 амплифицируют участки, кодирующие сигнальную последовательность ОmрА и домен FOS, из слитого гена ompA-FOS-hGH в векторе рКК223-3 (см. Пример 5), используя пару праймеров OmpA-FORI/FOS-REV2. Продукт ПЦР разрезают с помощью EcoRI/HindIII и лигируют в те же сайты вектора рКК223-3 (Pharmacia).
Для конструирования вектора pAV1 амплифицируют участок, кодирующий FOS, из слитого гена ompA-FOS-hGH в векторе рКК223-3 (см. Пример 5), используя пару праймеров FOS-FORI/FOS-REV1. Продукт ПЦР разрезают с помощью HindIII и лигируют в разрезанный StuI/HindIII вектор pAV2.
Для конструирования вектора pAV3 амплифицируют участок, кодирующий домен FOS, из вектора pAV1, используя пару праймеров FOS-FOR2/FOS-REV1. Продукт ПЦР разрезают с помощью EcoRI/Hindlll и лигируют в такие же сайты вектора рКК223-3 (Pharmacia).
Для конструирования вектора рАV4 амплифицируют участок, кодирующий домен FOS, из слитого гена ompA-FOS-hGR в векторе рКК223-3 (см. Пример 5), используя пару праймеров FOS-FOR3/ FOS-REV2. Продукт ПЦР разрезают с помощью EcoRI/Hindlll и лигируют в те же сайты вектора рКК223-3 (Pharmacia).
Для конструирования вектора рАV5 амплифицируют участок, кодирующий сигнальную последовательность hGH из слитого гена hGH-FQS-hGH в векторе pSINrepS (см. Пример 7), используя пару праймеров hGH-FOR1/hGHREV1. Продукт ПЦР разрезают с помощью EcoRI/NotI и лигируют в те же сайты вектора pAV1. Затем полученную кассету, кодирующую сигнальную последовательность hGH и домен FOS, выделяют и клонируют в вектор pMPSVEH (см. статью Artelt и соавт., Gene, 68: 213-219 (1988)), разрезанную теми же ферментами.
Для конструирования вектора рАV6 амплифицируют участок, кодирующий FOS, из вектора pAV2, используя пару праймеров FOS-FOR4/FOS-REV3. Продукт ПЦР разрезают с помощью HindIII и клонируют в расщепленный Eco47III/HindIII вектор pAV5. Целую кассету, кодирующую сигнальную последовательность hGH и домен FOS, затем повторно амплифицируют из полученного вектора, используя пару праймеров hGH-FOR2/FOS-REV3, расщепляют с помощью EcoRI/HindIII и лигируют в вектор pMPSVEH (см. статью Artelt и соавт., Gene, 68:213-219 (1988)), расщепленный теми же ферментами.
Пример 7:
Конструирование FOS-hGH с человеческой (hGH) сигнальной последовательностью
Для эукариотической экспрессии слитого белка FOS-hGH выделяют слитый ген OmpA-FOS-hGH из pBluescript::OmpA-FOS-hGH (см. Пример 4) путем разрезания с помощью XbaI/Bsp1201 и клонируют в вектор pSINrepS (Invitrogen), расщепленный теми же ферментами. Сигнальную последовательность hGH синтезируют с помощью ПЦР (реакционная смесь содержит 50 пмоль каждого праймера, dATP, dGTP, dTTP, dCTP (no 200 мкМ каждого), 2,5 ед. ДНК-полимеразы Taq (Qiagen), общий объем 50 мкл в буфере, представляемом изготовителем, при следующих условиях амплификации: 92°С в течение 30 сек, 55°С в течение 30 сек, 72°С в течение 30 сек, 30 циклов) с использованием перекрывающихся олигонуклеотидов Sig-hGH-FOR
(GGGTCTAGAATGGCTACAGGCTCCCGGACGTCCCTGCTCCTGGCTTTTGGCCTG CTCTG) (SEQ ID NO: 41) и Sig-hGH-REV (CGCAGGCCTCGGCACTGCCCTCTTGAAGCCAGGGCAGGCAGAGCA GGCCAAAAGCCAG) (SEQ ID NO: 42). Продукт ПЦР очищают, используя набор QiaExII, разрезают с помощью StuI/XbaI и лигируют в вектор pSINrep5::OmpA-FOS-hGH, расщепленный теми же ферментами.
Пример 8:
Эукариотическая экспрессия FOS-hGH
Не содержащий РНКазу вектор (1,0 мкг) (pSINrep5::OmpA-FOS-hGH) и 1,0 мкг DHEB (см. статью Bredenbeek и соавт., J. Virol, 67:6439-6446 (1993)) линеаризуют, разрезая рестриктазой Seal. Затем проводят транскрипцию in vitro с использованием набора для транскрипции in vitro SP6 (InvitroscripCAP, изготавливаемый InvitroGen, Invitrogen BV, NVLeek, Нидерланды). Полученную мРНК с 5’-кэпом анализируют в восстанавливающих условиях в агарозном геле.
Транскрибированную in vitro мРНК (5 мкг) электропорацией вводят в клетки ВНК21 (АТСС: CCL10) в соответствии с инструкцией Invitrogen (Экспрессионная система Sindbis (Sindbis Expression system), Invitrogen BV, Netherlands). После инкубирования в течение 10 час при 37°С среду, содержащую FCS, заменяют средой НР-1, не содержащей FCS, с последующим дополнительным инкубированием при 37°С в течение 10 час. Супернатант собирают и анализируют с помощью дот-блоттинга на образование FOS-hGH.
Культуральную среду (2,5 мкл) наносят в виде пятна на нитроцеллюлозную мембрану и сушат в течение 10 мин при комнатной температуре. Мембрану блокируют 1% бычьим альбумином (Sigma) в TBS (10 × TBS/л: 87,7 г NaCl, 66,1 г Trizma гидрохлорида (Sigma) и 9,7 г Trizma основания (Sigma), рН 7,4) в течение 1 часа при комнатной температуре с последующим инкубированием с 2 мкг антитела кролика против hGH человека в течение 1 часа. Блот промывают 3 раза по 10 минут в TBS-T и инкубируют в течение 1 часа с щелочной фосфатазой, конъюгированной с IgG против кролика (Jackson ImmunoResearch Laboratories, Inc.), разбавленной в соотношении 1:5000 в TBS-T. После промывания 2 раза в течение 10 минут в TBS-T блот проявляют окрашиванием АР, как описано в Примере 2. Результаты представляют на Фигуре 3.
Пример 9:
Конструирование FOS-PLA (N- и С-концевого)
Следующий ген конструируют путем химического синтеза гена, кодирующего каталитически неактивный вариант (см. статью 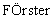 и соавт., J. Allergy Clin. Immunol. 95: 1229-1235 (1995)) фосфолипазы A2 пчелиного яда (PLA).
и соавт., J. Allergy Clin. Immunol. 95: 1229-1235 (1995)) фосфолипазы A2 пчелиного яда (PLA).

Для слияния PLA с М-концом домена димеризации FOS участок амплифицируют, используя олигонуклеотиды PLA-FOR1 (CCATCATCTACCCAGGTAC) (SEQ ID NO: 45) и PLA-REV1 (CCCACACCCAGCGGCCGCGTATTTGCGCAGGTCG) (SEQ ID NO: 46). Продукт ПЦР разрезают NotI и лигируют в вектор pAV1, предварительно расщепленный рестриктазами StuI/NotI. Для слияния PLA с С-концом домена димеризации FOS участок амплифицируют, используя олигонуклеотиды PLA-FOR2 (CGGTGGTTCTGCGGCCGCTATCATCTACCCAGGTAC) (SEQ ID N0:47) и PLA-REV2 (TTAGTATTTGCGCAGGTCG) (SEQ ID NO: 48). Продукт, полученный при ПЦР, разрезают с помощью NotI и лигируют в вектор pAV2, предварительно расщепленный рестриктазами NotI/EcoRV.
Пример 10:
Конструирование слитого гена FOS-овальбумина (N- и С-концевого)
Для клонирования последовательности, кодирующей овальбумин, получают мРНК из ткани куриных яйцеводов, используя набор для очистки MPHKQuickPrep™ Micro mRNA Purification Kit (Pharmacia) согласно инструкциям изготовителя. С помощью набора для одностадийной ОТ-ПЦР Superscript™ One-step RT PCR Kit (Gibco BRL) синтезируют кДНК, кодирующую зрелую часть овальбумина (соответствующую нуклеотидам 68-1222 мРНК (см. статью McReynolds и соавт., Nature, 273: 723-728 (1978)) с использованием праймеров Ova-FORI (CCGGCTCCATCGGTGCAG) (SEQ ID NO: 49) и Ova-REVI (ACCACCAGAAGCGGCCGCAGGGGAAACACATCTGCC) (SEQ ID NO: 50). Продукт ПЦР разрезают с помощью NotI и клонируют в разрезанный StuI/NotI вектор pAVI с целью экспрессии слитого белка с доменом димеризации FOS на С-конце. Для получения слитого белка с доменом димеризации FOS на N-конце участок, кодирующий овальбумин, амплифицируют из сконструированного вектора (pAV1::Ova), используя праймеры Ova-FOR2 (CGGTGGTTCTGCGGCCGCTGGCTCCATCGGTGCAG) (SEQ ID NO: 51) и Ova-REV2 (TTAAGGGGAAACACATCTGCC) (SEQ ID NO: 52). Продукт ПЦР разрезают с помощью NotI и клонируют в разрезанный с помощью Notl/EcoRV вектор pAV2. Клонированные фрагменты проверяют с помощью анализа последовательности ДНК.
Пример 11:
Продукция и очистка слитых белков FOS-PLA и FOS-овальбумин
Для цитоплазматической продукции слитых белков FOS подходящий штамм Е.соli трансформируют векторами pAV3::PLA, pAV4::PLA, pAV4::Ova, pAV3::Ova или pAV4::Ova. Культуру инкубируют в обогащенной среде в присутствии ампициллина при встряхивании при 37°С. По достижении оптической плотности (550 нм) добавляют 1,1 мМ IPTG и продолжают инкубирование в течение последующих 5 час. Клетки собирают с помощью центрифугирования, ресуспендируют в подходящем буфере (например, Трис-HCl, рН 7,2, 150 мМ NaCl), содержащем ДНКазу, РНКазу и лизоцим, и разрушают, проводя через камеру под давлением Френча (French). После центрифугирования (Sorvall RC-5C, ротор SS34, 15000 об/мин, 10 мин, 4°С) осадок ресуспендируют в буфере для промывания телец включений (20 мМ Трис-HCl, 23% сахарозы, 0,5% Triton X-100, 1 мМ EDTA (этилендиаминтетрауксуная кислота), рН 8) при 4°С и повторно центрифугируют, как описано выше. Данную процедуру повторяют, пока супернатант, полученный после центрифугирования, не становится практически прозрачным. Тельца включений ресуспендируют в 20 мл буфера для солюбилизации (5,5 М гидрохлорида гуанидина, 25 мМ Трис-HCl, рН 7,5) при комнатной температуре и удаляют нерастворимый материал центрифугированием с последующим пропусканием супернатанта через стерильный фильтр (0,45 мкм). Раствор белка хранят при 4°С в течение по меньшей мере 10 час в присутствии 10 мМ EDTA и 100 мМ DTT, а затем трижды диализуют против 10 объемов 5,5 М гидрохлорида гуанидина, 25 мМ Трис-HCl, 10 мМ EDTA, рН 6. Раствор дважды диализуют против 5 л 2 М мочевины, 4 мМ EDTA, 0,1 M NH4Cl, 20 мМ бората натрия (рН 8,3) в присутствии подходящей окислительно-восстановительной пары (окисленный глутатион/восстановленный глутатион; цистин/цистеин). Переуложенный белок подвергают ионообменной хроматографии. Белок хранят в подходящем буфере с рН выше 7 в присутствии 2-10 мМ DTT для того, чтобы сохранить цистеиновые остатки, фланкирующие домен FOS, в восстановленной форме. Перед связыванием белка с частицами альфавируса DTT удаляют, пропуская раствор белка через колонку для гель-фильтрации Sephadex G-25.
Пример 12:
Конструкции gp140-FOS
Ген gp140 (Swiss-Prot:P03375), не содержащий внутренний сайт протеазного расщепления, амплифицируют с помощью ПЦР из оригинальной плазмиды рАbТ4674 (АТСС 40829), содержащей ген gр 160 полной длины, с использованием следующих олигонуклеотидов:
ВИЧ-1:
5’-ACTAGTCTAGAatgagagtgaaggagaaatatc-3’ (SEQ ID NO: 53);
ВИЧ-end:
5’-TAGCATGCTAGCACCGAAtttatctaattccaataattcttg-3’ (SEQ ID NO: 54);
ВИЧ-Cleav:
5’-gtagcacccaccaaggcaaagCTGAAAGCTACCCAGCTCGAGAAACTGgca-3’ (SEQ ID NO: 55) и
BИЧ-Cleav2:
5'-caaagctcctattcccactgcCAGTTTCTCGAGCTGGGTAGCTTTCAG-3' (SEQ ID 10 N0:56).
Для ПЦР I используют 100 пмоль олигонуклеотида HIV-1 и HIV-Cleav2 и 5 нг ДНК-матрицы в 75 мкл реакционной смеси (4 ед. полимеразы Taq или Pwo, 0,1 мМ dNTP и 1,5 мМ МgSO4). Циклинг ПЦР осуществляют следующим образом: 30 циклов при температуре отжига 60°С и времени элонгации 2 мин при 72°С.
Для ПЦР II используют 100 пмоль олигонуклеотида HIV-end и HIV-Cleav и 5 нг ДНК-матрицы в 75 мкл реакционной смеси (4 ед. полимеразы Taq или Pwo, 0,1 мМ dNTP и 1,5 мМ MgSO4). Циклинг ПЦР осуществляют следующим образом: 30 циклов при температуре отжига 60°С и времени элонгации 50 сек при 72°С.
Оба полученные при ПЦР фрагменты очищают, выделяют и используют в реакции сборки с помощью ПЦР. Для сборки с помощью ПЦР используют 100 пмоль олигонуклеотидов HIV-1 и HIV-end и 2 нг каждого из фрагментов, полученных при ПЦР (ПЦРI и ПЦРII), в 75 мкл смеси (4 ед. полимеразы Taq или Pwo, 0,1 мМ dNTP и 1,5 мМ MgSO4). ПЦР-циклинг осуществляют следующим образом: 30 циклов при температуре отжига 60°С и времени элонгации 2,5 мин при 72°С. Продукт сборки с помощью ПЦР разрезают с помощью XbaI и NheI. Амфипатическую спираль FOS сливают в рамке считывания с С-концом gр-140.
Последовательность ДНК, кодирующую домен амфипатической спирали FOS, амплифицируют с помощью ПЦР из вектора pJuFo (см. статью Crameri и Suter, Gene, 137:69 (1993)), используя олигонуклеотиды:
FOS-ВИЧ:
5’-ttcggtgctagcggtggcTGCGGTGGTCTGACCGAC-3’ (SEQ ID NO: 57) и
FOS-Apa:
5’-gatgctgggcccttaaccGCAACCACCGTGTGCCGCC-3’ (SEQ ID NO: 58).
Для ПЦР используют 100 пмоль каждого олигонуклеотида и 5 нг ДНК-матрицы в 75 мкл реакционной смеси (4 ед. полимеразы Taq или Pwo, 0,1 мМ dNTP и 1,5 мМ MgSO4). Температурный циклинг осуществляют следующим образом: 95°С в течение 2 минут с последующими 5 циклами, состоящими из следующих стадий: 95°С (45 сек), 60°С (30 сек), 72°С (25 сек), и последующими 25 циклами, состоящими из следующих стадий: 95°С (45 сек), 68°С (30 сек), 72°С (20 сек). Полученный продукт ПЦР разрезают с помощью NheI и Bsp120L.
Конечный экспрессионный вектор для GP140-FOS получают при лигировании обоих фрагментов, полученных при ПЦР, в реакции лигирования трех фрагментов в pSinRep5. Полученный в результате вектор pSinRep5-GP140-FOS определяют с помощью рестрикционного анализа и секвенирования ДНК.
GP140-FOS клонируют также в pCYTts с помощью XbaI и Bsp120L для получения стабильной индуцируемой клеточной линии, экспрессирующей GP140-FOS.
Пример 13:
Экспрессия GP140FOS с использованием pSinRep5-GP140-FOS
Не содержащий РНКазу вектор (1,0 мкг) (pSinRep5-GP140-FOS) и 1,0 мкг DHEB (см. статью Bredenbeek и соавт., J. Virol, 67: 6439-6446 (1993)) линеаризуют, разрезая рестриктазой. Затем проводят транскрипцию in vitro с использованием набора для транскрипции in vitro SP6 (InvitroscripCAP, изготавливаемый InvitroGen, Invitrogen BV, NVLeek, Нидерланды). Полученную мРНК с 5’-кэпом анализируют в восстанавливающих условиях в агарозном геле.
Транскрибированную in vitro мРНК (5 мкг) электропорацией вводят в клетки ВНК21 (АТСС: CCL10) в соответствии с инструкцией Invitrogen (Экспрессионная система Sindbis (Sindbis Expression system), Invitrogen BV, Netherlands). После инкубирования в течение 10 час при 37°С среду, содержащую FCS, заменяют средой НР-1, не содержащей FCS, с последующим дополнительным инкубированием при 37°С в течение 10 час. Супернатант собирают и анализируют с помощью вестерн-блоттинга на образование растворимого GP140-FOS точно, как описано в Примере 2.
Пример 14:
Экспрессия GP140-FOS с использованием pCYTts-GP140-FOS
Двадцать мкг pCYT-GP140-FOS линеаризуют, разрезая рестриктазой. Реакцию прекращают экстракцией фенолом/хлороформом с последующим осаждением изопропанолом линеаризованной ДНК. Реакцию рестрикции проверяют гель-элетрофорезом в агарозе. Для трансфекции 5,4 мкг линеаризованного pCYT-GP140-FOS смешивают с 0,6 мкг линеаризованного pSV2Neo в 30 мкл Н2О и добавляют 30 мкл 1 М раствора СаСl2. После добавления 60 мкл фосфатного буфера (50 мМ HEPES, 280 мМ NaCl, 1,5 мМ Na2HPO4, pH 7,05) раствор взбалтывают в течение 5 секунд, а затем инкубируют при комнатной температуре в течение 25 секунд. Раствор немедленно добавляют к 2 мл среды НР-1, содержащей 2% FCS (2% среда FCS). Среду конфлюэнтной на 80% культуры клеток ВНК21 (в 6-луночной плате) затем замещают ДНК-содержащей средой. После инкубирования в течение 5 часов при 37°С в инкубаторе с CO2 ДНК-содержащую среду удаляют и замещают 2 мл 15% глицерина в 2% среде FCS.
Глицерин-содержащую среду удаляют после 30-секундной фазы инкубирования и промывают клетки 5 мл среды НР-1, содержащей 10% FCS. Наконец, добавляют 2 мл свежей среды НР-1, содержащей 10% FCS.
Отбирают стабильно трансфицированные клетки и выращивают на селективной среде (среда НР-1 с добавлением G418) при 37°С в инкубаторе с СО2. Когда смешанная популяция вырастает до конфлюэнтности, культуру делят на две чашки с последующим периодом роста в течение 12 час при 37°С. Одну чашку с клетками переносят в условия культивирования при 30°С для индукции экспрессии растворимого GP140-FOS. Другую чашку сохраняют при 37°С.
Экспрессию растворимого GP140-FOS определяют с помощью вестерн-блоттинга. Культуральную среду (0,5 мл) осаждают метанолом/хлороформом и ресуспендируют осадок в буфере для образцов для SDS-PAGE. Образцы нагревают в течение 5 минут при 95°С перед внесением в 15% акриламидный гель. После SDS-PAGE белки переносят на нитроцеллюлозные мембраны Protan (Schleicher & Schuell, Germany), как предложено в разделе, написанном Bass и Yang, в монографии под ред. Creighton Т.Е. Практические подходы к функции белка (Protein Function: A Practical Approach), 2-е изд., IRL Press, Oxford (1997), стр.29-55. Мембрану блокируют 1% бычьим альбумином (Sigma) в TBS (10 × TBS/л: 87,7 г NaCl, 66,1 rTrizma гидрохлорида (Sigma) и 9,7 г Trizma основания (Sigma), pH 7,4) в течение 1 часа при комнатной температуре с последующим инкубированием с антителом против GP140 или GP160 в течение 1 часа. Блот промывают 3 раза по 10 минут с помощью TBS-T (TBS с добавлением 0,05% Tween 20) и инкубируют в течение 1 часа с конъюгатом щелочной фосфатазы с IgG против мыши/кролика/обезьяны/человека. После промывания 2 раза в течение 10 минут в TBS-T и 2 раза в течение 10 минут в TBS проводят проявляющую реакцию с использованием реагентов для детекции щелочной фосфатазы (10 мл буфера АР (100 мМ Трис/HCl, 100 мМ NaCl, pH 9,5) с 50 мкл раствора NBT (7,7% нитро синего тетразолия (Sigma) в 70% диметилформамиде) и 37 мкл раствора Х-фосфата (5% 5-бром-4-хлор-3-индолилфосфата в диметилформамиде).
Пример 15:
Продукция и очистка GP140-FOS
Антитело против gр120 ковалентно связывают с NHS/EDC-активированным декстраном и упаковывают в хроматографическую колонку. Супернатант, содержащий GP140-FOS, помещают на колонку и после достаточного промывания элюируют GP140-FOS, используя 0,1 М HCl. Элюат нейтрализуют непосредственно во время сбора, используя 1 М Трис pH 7,2, находящийся в пробирках для сбора.
Поскольку во время очистки могло бы произойти образование дисульфидной связи, собранный образец обрабатывают 10 мМ DTT в 10 мМ Трис, pH 7,5 в течение 2 час при 25°С.
DTT удаляют при последующем диализе против 10 мМ Mes, 80 мМ NaCl, pH 6,0. Наконец, GP140-FOS смешивают с альфавирусными частицами, содержащими лейциновую молнию JUN в Е2, как описано в Примере 16.
Пример 16:
Получение частиц альфа-вакцины
Вирусные частицы (см. Примеры 2 и 3) концентрируют, используя устройства для фильтрации Millipore Ultrafree Centrifugal Filter Devices, удаляющие соединения молекулярной массы 100 кД, в соответствии с инструкций, представляемой изготовителем. Альтернативно вирусные частицы концентрируют путем центрифугирования в градиенте сахарозы, как описано в руководстве для Экспрессионной системы Sindbis (Sindbis Expression System) (Invitrogen, San Diego, California). pH суспензии вируса подводят до 7,5 и инкубируют вирусные частицы в присутствии 2-10 мМ DTT в течение нескольких часов. Вирусные частицы очищают от белковых примесей на колонке Sephacryl S-300 (Pharmacia) (вирусные частицы элюируют свободным объемом) в подходящем буфере.
Очищенные вирусные частицы инкубируют с по меньшей мере 240-кратным молярным избытком слитого белка FOS-антиген в подходящем буфере (pH 7,5-8,5) в присутствии окислительно-восстановительной пары (окисленный глутатион/восстановленный глутатион; цистин/цистеин) в течение по меньшей мере 10 час при 4°С. После концентрирования частиц с использованием устройства Millipore Ultrafree Centrifugal Filter Device, удаляющего соединения с молекулярной массой 100 кД, смесь пропускают через колонку для гель-фильтрации Sephacryl S-300 (Pharmacia). Вирусные частицы элюируют свободным объемом.
Пример 17:
Слияние амфипатической спирали JUN с аминоконцом НВсАg(1-144)
Спираль JUN сливают с аминоконцом последовательности аминокислот 1-144 HBcAg (конструкция JUN-HBcAg). Для конструирования последовательности ДНК JUN-HBcAg последовательности, кодирующие спираль JUN и НВсАg(1-144), раздельно амплифицируют с помощью ПЦР. Последовательность JUN амплифицируют из плазмиды pJuFo, используя праймеры EcoRI-JUN(s) и JUN-SacII(as). Праймер EcoRI-JUN(s) интродуцирует сайт EcoRI, следующий за стартовым кодоном ATG. Праймер JUN-SacII(as) интродуцирует линкер, кодирующий последовательность аминокислот GAAGS. Последовательность НВсАg(1-144) амплифицируют из плазмиды рЕсо63 (получена в АТСС No. 31518), используя праймеры JUN-HBcAg(s) и HBcAg(1-144)Hind(as). JUN-HBcAg(s) содержит последовательность, соответствующую 3’-концу последовательности, кодирующей спираль JUN, за которой находится последовательность, кодирующая линкер GAAGS и 5’-конец последовательности HBcAg. HBcAg(1-144)Hind(as) интродуцирует стоп-кодон и сайт HindIII после кодона 144 гена HBcAg. Для проведения ПЦР используют 100 пмоль каждого олигонуклеотида и 50 нг матричных ДНК в 50 мкл реакционных смесей, содержащих 2 ед. полимеразы Pwo, 0,1 мМ dNTP и 2 мМ MgSO4. Для обеих реакций температурный циклинг проводят следующим образом: 94°С в течение 2 мин с последующими 30 циклами, состоящими из следующих стадий: 94°С (1 мин), 50°С (1 мин), 72°С (2 мин).
Последовательности праймеров:
EcoRI-JUN(s):
(5’-CCGGAATTCATGTGCGGTGGTCGGATCGCCCGG-3’) (SEQ ID NO: 61);
JUN-SacII(as):
(5’-GTCGCTACCCGCGGCTCCGCAACCAACGTGGTTCATGAC-3’) (SEQ ID NO: 62);
JUN-HBcAg(s):
(5’-GTTGGTTGCGGAGCCGCOGGTAGCGACATTGACCCTTATAAAGAATTTGG-3’) (SEQ ID NO: 63);
HBcAg(1-144)Hind(as):
(5’-CGCGTCCCAAGCTTCTACGGAAGCGTTGATAGGATAGG-3’) (SEQ ID NO: 64).
Слияние двух фрагментов, полученных при ПЦР, осуществляют с помощью ПЦР, используя праймеры ЕсоRI-JUN(s) и HBcAg(1-144)Hind(as). 100 пмоль каждого олигонуклеотида используют со 100 нг очищенных полученных при ПЦР фрагментов в 50 мкл реакционной смеси, содержащей 2 ед. полимеразы Pwo, 0,1 мМ dNTP и 2 мМ MgSO4. Используют следующие условия ПЦР-циклинга: 94°С в течение 2 мин с последующими 35 циклами, состоящими из стадий: 94°С (1 мин), 50°С (1 мин), 72°С (2 мин). Конечный продукт ПЦР анализируют с помощью гель-электрофореза с использованием агарозы, очищают и разрезают в течение 16 час в подходящем буфере рестриктазами EcoRI and HindIII. Фрагмент разрезанной ДНК лигируют в разрезанный с помощью EcoRI/HindIII вектор рКК для создания экспрессионного вектора pKK-JUN-HBcAg. Инсерцию продукта ПЦР определяют рестрикционным анализом с помощью EcoRI/HindIII и путем секвенирования ДНК инсерции.
Пример 18:
Слияние амфипатической спирали JUN с карбоксиконцом НВсАg(1-144)
Спираль JUN сливают с карбоксиконцом последовательности аминокислот 1-144 HBcAg (конструкция HBcAg-JUN). Для конструирования последовательности ДНК HBcAg-JUN последовательности, кодирующие спираль JUN и НВсАg(1-144), раздельно амплифицируют с помощью ПЦР. Последовательность JUN амплифицируют из плазмиды pJuFo, используя праймеры SacII-JUN(s) и JUN-HindIII(as). SacII-JUN(s) интродуцирует линкер, кодирующий последовательность аминокислот LAAG. Данная последовательность содержит также сайтSасII. JUN-HindIII(as) интpoдyциpyeт стоп-кодон (ТАА), за которым следует сайт HindIII. Последовательность ДНК НВсАg(1-144) амплифицируют из плазмиды рЕсо63, используя праймеры EcoRI-HBcAg(s) и HBcAg(1-144)-JUN(as). EcoRI-HBcAg(s) интродуцирует сайт EcoRI перед стартовым кодоном ATG последовательности, кодирующей HBcAg. HBcAg(1-144)-JUN(as) интродуцирует последовательность, кодирующую пептидный линкер (LAAG), который содержит также сайт SacII. Для проведения ПЦР используют 100 пмоль каждого олигонуклеотида и 50 нг матричных ДНК в 50 мкл реакционных смесей, содержащих 2 ед. полимеразы Pwo, 0,1 мМ dNTP и 2 мМ MgSO4. Температурный циклинг проводят следующим образом: 94°С в течение 2 мин с последующими 30 циклами, состоящими из следующих стадий: 94°С (1 мин), 50°С (1 мин), 72°С (2 мин). Последовательности праймеров:
SacII-JUN(s):
(5’-CTAGCCGCGGGTTGCGGTGGTCGGATCGCCCGG-3’) (SEQ ID NO: 65);
JUN-HindIII(as):
(5’-CGCGTCCCAAGCTTTTAGCAACCAACGTGGTTCATGAC-3’) (SEQ ID NO: 66);
EcoRI-HBcAg(s):
(5’-CCGGAATTCATGGACATTGACCCTTATAAAG-3’) (SEQ ID NO: 67) и HBcAg-JUN(as):
(5’-CCGACCACCGCAACCCGCGGCTAGCGGAAGCGTTGATAGGATAGG-3’)(SEQID NO: 68).
Слияние двух фрагментов, полученных при ПЦР, осуществляют с помощью ПЦР, используя праймеры EcoRI-HBcAg(s) и JUN-HindIII(as). Для слияния с помощью ПЦР 100 пмоль каждого олигонуклеотида используют со 100 нг очищенных полученных при ПЦР фрагментов в 50 мкл реакционной смеси, содержащей 2 ед. полимеразы Pwo, 0,1 мМ dNTP и 2 мМ MgSO4. Используют следующие условия ПЦР-циклинга: 94°С в течение 2 мин с последующими 35 циклами, состоящими из стадий: 94°С (1 мин), 50°С (1 мин), 72°C (2 мин). Конечный продукт ПЦР анализируют с помощью гель-электрофореза с использованием агарозы и разрезают в течение 16 час в подходящем буфере рестриктазами EcoRI and HindIII. Фрагмент разрезанной ДНК очищают гель-хроматографией и лигируют в разрезанный с помощью EcoRI/HindIII вектор рКК для создания экспрессионного вектора pKK-HBcAg-JUN. Инсерцию продукта ПЦР определяют рестрикционным анализом с помощью EcoRI/HindIII и путем секвенирования ДНК инсерции.
Пример 19:
Инсерция амфипатической спирали JUN в эпитоп с/е1 НВсАд(1-144)
Известно, что эпитоп с/е1 (остатки 72-88) НВсАд расположен в концевом участке на поверхности капсида вируса гепатита В. Фрагмент данного участка (остатки 76-82) белка генетически замещают спиралью JUN для получения центра связывания для антигенов (конструкция HBcAg-JUNIns). Последовательность ДНК HBcAg-JUNIns получают с помощью ПЦР, для чего последовательность спирали JUN и две последовательности, кодирующие фрагменты НВсАg (остатки аминокислот 1-75 и 83-144), амплифицируют раздельно с помощью ПЦР. Последовательность JUN амплифицируют из плазмиды pJuFo с использованием праймеров BamHI-JUN(s) и JUN-SacII(as). BamHI-JUN(s) интродуцирует линкерную последовательность, кодирующую линкерную последовательность пептида GSGGG, который содержит также сайт BamHI. JUN-SacII(as) интродуцирует последовательность, кодирующую пептидный линкер GMGS, за которым следует последовательность, комплементарная 3’-концу последовательности, кодирующей JUN. Последовательность ДНК НВсАg(1-75) амплифицируют из плазмиды рЕсо63, используя праймеры EcoRIHBcAg(s) и HBcAg75-JUN(as). EcoRIHBcAg(s) интродуцирует сайт EcoRI, за которым следует последовательность, соответствующая 5’-концу последовательности HbcAg. HBcAg75-JUN(as) интродуцирует линкер, кодирующий пептид GSGGG, после аминокислоты 75 HbcAg, за которым следует последовательность, комплементарная 5’-концу последовательности, кодирующей спираль JUN. Фрагмент НВсАg (83-144) амплифицируют, используя праймеры JUN-HBcAg83(s) и HBcAg(1-144)Hind(as). JUN-HBcAg83(s) содержит последовательность, соответствующую 3’-концу последовательности, кодирующей JUN, за которой следует линкер, кодирующий пептид GAAGS, и последовательность, соответствующая 5’-концу последовательности, кодирующей НВсАg(83-144). HBcAg(1-144)Hind(as) интродуцирует стоп-кодон и сайт HindIII после кодона 144 гена НВсАg. Для ПЦР используют 100 пмоль каждого олигонуклеотида и 50 нг матричных ДНК в 50 мкл реакционных смесей (2 ед. полимеразы Pwo, 0,1 мМ dNTP и 2 мМ MgSO4). Температурный циклинг проводят следующим образом: 94°С в течение 2 мин с последующими 35 циклами, состоящими из стадий: 94°С (1 мин), 50°С (1 мин), 72°С (2 мин).
Последовательности праймеров:
BamHI-JUN(s):
(5’-CTAATGGATCCGGTGGGGGCTGCGGTGGTCGGATCGCCCGGCTCGAG-3’) (SEQ ID NO:69);
JUN-SacII(as):
(5’-GTCGCTACCCGCGGCTCCGCAACCAACGTGGTTCATGAC-3’) (SEQ ID NO: 70);
EcoRIHBcAg(s):
(5’-CCGGAATTCATGGACATTGACCCTTATAAAG-3’) (SEQ ID NO: 71);
HBcAg75-JUN (as):
(5’-CCGACCACCGCAGCCCCCACCGGATCCATTAGTACCCACCCAGGTAGC-3’) (SEQ ID NO: 72);
JUN-HBcAg83(s):
(5’-GTTGGTTGCGGAGCCGCGGGTAGCGACCTAGTAGTCAGTTATGTC-3’) (SEQ ID NO: 73) и
HBcAg(1-144)Hind(as):
(5’-CGCGTCCCAAGCTTCTACGGAAGCGTTGATAGGATAGG-3’) (SEQ ID NO: 74).
Слияние трех полученных при ПЦР фрагментов осуществляют следующим образом. Во-первых, фрагмент, кодирующий НВсАg (1-75), сливают с последовательностью, кодирующей JUN, с помощью ПЦР, используя праймеры EcoRIHBcAg(s) и JUN-SacII(as). Во-вторых, полученный продукт сливают с фрагментом HbcAg (83-144) с помощью ПЦР, используя праймеры EcoRI HBcAg(s) и HBcAg HindIII(as). Для слияний с помощью ПЦР 100 пмоль каждого олигонуклеотида используют со 100 нг очищенных полученных при ПЦР фрагментов в 50 мкл реакционной смеси, содержащей 2 ед. полимеразы Pwo, 0,1 мМ dNTP и 2 мМ MgSO4. Такие же циклы ПЦР используют для получения отдельных фрагментов. Конечный продукт ПЦР разрезают в течение 16 час в подходящем буфере с помощью рестриктаз EcoRI и HindIII. Фрагмент ДНК лигируют в разрезанный EcoRI/HindIII вектор рКК, получая вектор pKK-HBcAg-JUNIns. Инсерцию продукта ПЦР определяют с помощью рестрикционного анализа с использованием EcoRI/HindIII и путем секвенирования ДНК инсерции.
Пример 20:
Слияние амфипатической спирали JUN с карбоксиконцом белка нуклеокапсида (N) вируса кори
Спираль JUN сливают с карбоксиконцом усеченного фрагмента белка N вируса кори, содержащего остатки аминокислот 1-473 (конструкция N473-JUN). Для конструирования последовательности ДНК, кодирующей N473-JUN, последовательность, кодирующую спираль JUN, и последовательность, кодирующую N473-JUN, амплифицируют раздельно с помощью ПЦР. Последовательность JUN амплифицируют из плазмиды pJuFo с помощью праймеров SacII-JUN(s) и JUN-HindIII(as). SacII-JUN(s) интродуцирует последовательность, кодирующую пептидный линкер LAAG. Данная последовательность содержит также сайт Sacli. Антисмысловой праймер JUN-HindIII(as) интродуцирует стоп-кодон (ТАА), за которым следует сайт HindIII. Последовательность N (1-473) амплифицируют из плазмиды pSC-N, содержащей последовательность, кодирующую полный белок N вируса кори (предоставленный М. Billeter, Zurich), с использованием праймеров EcoRI-Nmea(s) и Nmea-JUN(as). EcoRI-N(mea)(s) интродуцирует сайт EcoRJ перед стартовым кодоном ATG последовательности, кодирующей N. N(mea)-JUN(as) является комплементарным 3’-концу последовательности, кодирующей N(1-473), за которой следует последовательность, комплементарная последовательности, кодирующей пептидный линкер (LAAG). Для ПЦР используют 100 пмоль каждого олигонуклеотида и 50 нг матричных ДНКв 50 мкл реакционных смесей, содержащих 2 ед. полимеразы Pwo, 0,1 мМ dNTP и 2 мМ MgSO4. Температурный циклинг проводят следующим образом: 94°С в течение 2 мин с последующими 35 циклами, состоящими из стадий: 94°С (1 мин), 50°С (1 мин), 72°С (2 мин).
Последовательности праймеров:
SacII-JUN(s):
(5’-CTAGCCGCGGGTTGCGGTGGTCGGATCGCCCGG-3’) (SEQ ID NO: 75);
JUN-HindIII(as):
(5’-CGCGTCCCAAGCTTTTAGCAACCAACGTGGTTCATGAC-3’) (SEQ IDNO:76);
EcoRI-Nmea(s):
(5’-CCGGAATTCATGGCCACACTTTTAAGGAGC-3’) (SEQ ID NO: 77) и
Nmea-JUN(as):
(5’-CGCGTCCCAAGCTTTTAGCAACCAACGTGGTTCATGAC-3’) (SEQ ID NO: 78).
Слияние двух полученных при ПЦР фрагментов осуществляют в следующей реакции ПЦР, используя праймеры EcoRI-Nmea(s) и Nmea-JUN(as). Для слияния с помощью ПЦР 100 пмоль каждого олигонуклеотида используют со 100 нг очищенных полученных при ПЦР фрагментов в 50 мкл реакционной смеси, содержащей 2 ед. полимеразы Pwo, 0,1 мМ dNTP и 2 мМ MgSO4. Температурный циклинг проводят следующим образом: 94°С в течение 2 мин с последующими 35 циклами, состоящими из стадий: 94°С (1 мин), 50°С (1 мин), 72°С (2 мин). Продукт ПЦР разрезают в течение 16 час в подходящем буфере с помощью рестриктаз EcoRI и HindIII. Фрагмент ДНК очищают и лигируют в разрезанный EcoRI/HindIII вектор рКК, получая плазмиду pKK-N473-JUN. Инсерцию продукта ПЦР определяют с помощью рестрикционного анализа с использованием EcoRI/HindIII и путем секвенирования ДНК инсерции.
Пример 21:
Экспрессия и частичная очистка HBcAg-JUN
Штамм Е.coli xl-1 blue трансформируют pKK-HBcAg-JUN. 1 мл суточной культуры бактерий используют для инокуляции 100 мл среды LB, содержащей 100 мкг/мл ампициллина. Данную культуру выращивают в течение 4 час. При 37°С до того, как OD при 600 нм не достигнет значения приблизительно 0,8. Индукцию синтеза HBcAg-JUN осуществляют путем добавления IPTG до конечной концентрации 1 мМ. После индукции бактерии продолжают встряхивать при 37°С в течение 16 час. Бактерии собирают центрифугированием при 5000×g в течение 15 мин. Осадок замораживают при -20°С. Осадок оттаивают и ресуспендируют в буфере для лизиса бактерий (10 мМ Na2PO4, рН 7,0, 30 мМ NaCl, 0,25% Tween-20, 10 мМ EDTA, 10 мМ DTT) с добавлением 200 мкг/мл лизоцима и 10 мкл бензоназы (Merck). Клетки инкубируют в течение 30 мин при комнатной температуре и разрушают в камере под давлением Френча (French). Triton X-100 добавляют к лизату до конечной концентрации 0,2% и инкубируют лизат в течение 30 мин на льду при периодическом встряхивании. На Фигуре 4 представляют экспрессию белка HBcAg-JUN в Е.соli при индукции с помощью IPTG. Клетки Е.соli, несущие экспрессионную плазмиду pKK-HBcAg-JUN или контрольную плазмиду, используют для индукции экспрессии HBcAg-JUN с помощью IPTG. Перед добавлением IPTG берут образец бактериальной культуры, несущей плазмиду pKK-HBcAg-JUN (дорожка 3), и культуры, несущей контрольную плазмиду (дорожка 1). Через 16 час после добавления IPTG снова берут образцы культуры, содержащей pKK-HBcAg-JUN (дорожка 4), и контрольной культуры (дорожка 2). Экспрессию белка мониторируют с помощью SDS-PAGE с последующим окрашиванием кумасси.
Затем лизат центрифугируют в течение 30 мин при 12,000×g с целью удаления нерастворимого клеточного дебриса. Супернатант и осадок анализируют с помощью вестерн-блоттинга, используя моноклональные антитела против НВсАg (YVS1841, приобретаемые в фирме Accurate Chemical and Scientific Corp., Westbury, NY, USA), показывая, что значительное количество белка НВсАg-JUN является растворимым (см. Фигуру 5). Вкратце, лизаты, полученные из клеток Е.соli, экспрессирующих НВсАg-JUN, и из контрольных клеток центрифугируют при 14,000×g в течение 30 мин. Супернатант (=растворимую фракцию) и осадок (=нерастворимую фракцию) разделяют и разбавляют буфером для образцов SDS до равных объемов. Образцы анализируют, используя SDS-PAGE с последующим вестерн-блоттингом с использованием моноклонального антитела против НВсАд YVS 1841. На дорожке 1 представлены растворимая фракция, контрольные клетки; на дорожке 2 представлены нерастворимая фракция, контрольные клетки; на дорожке 3 представлены растворимая фракция, клетки, экспрессирующие НВсАg-JUN; на дорожке 4 представлены нерастворимая фракция, клетки, экспрессирующие HBcAg-JUN.
Осветленный лизат клеток используют для центрифугирования в ступенчатом градиенте с использованием ступенчатого градиента сахарозы, состоящего из 4 мл 65% раствора сахарозы, на который накладывают 3 мл 15% раствора сахарозы, на который наносят 4 мл лизата бактерий. Образец центрифугируют в течение 3 час при 100,000×g при 4°С. После центрифугирования 1 мл фракций из верхней части градиента собирают и анализируют с помощью SDS-PAGE с последующим окрашиванием кумасси (см. Фигуру 6). На дорожке 1 представлен общий лизат Е.coli перед центрифугированием. На дорожках 1 и 2 представлены фракции 1 и 2 из верхней части градиента. На дорожках 4-7 представлены фракции 5-8 (15% сахароза). Белок HBcAg-JUN определяют окрашиванием кумасси.
Белок HBcAg-JUN накапливается на границе раздела между 15 и 65% сахарозы, что указывает на то, что он формирует капсидную частицу. Большая часть бактериальных белков остается в не содержащем сахарозу верхнем слое градиента, вследствие чего центрифугирование частиц HBcAg-JUN в ступенчатом градиенте приводит как к обогащению, так и частичной очистке частиц.
Пример 22:
Ковалентное связывание hGH-FOS с HBcAg-JUN
Для того чтобы продемонстрировать связывание белка с частицами HBcAg-JUN, выбирают гормон роста человека (hGH), слитый по карбоксиконцу со спиралью FOS, в качестве модели белка (hGH-FOS). Частицы HBcAg-JUN смешивают с частично очищенным hGH-FOS и инкубируют в течение 4 час при 4°С для связывания белков. Затем смесь диализуют в течение ночи против 3000-кратного объема буфера для диализа (150 мМ NaCl, 10 мМ раствора Трис-HCl, рН 8,0) с целью удаления DTT, присутствующего как в растворе HBcAg-JUN, так и в растворе hGH-FOS, обеспечивая таким образом ковалентное связывание белков путем образования дисульфидных связей. В качестве контролей растворы HBcAg-JUN и hGH-FOS также диализуют против буфера для диализа. Образцы всех трех диализуемых растворов белков анализируют с помощью SDS-PAGE в невосстанавливающих условиях.
Связывание hGH-FOS c HBcAg-JUN определяют путем иммуноблоттинга с антителом против hGH (см. Фигуру 7). hGH-FOS, связанный с HBcAg-JUN, должен мигрировать с предполагаемой молекулярной массой приблизительно 53 кД, тогда как несвязанный hGH-FOS мигрирует с предполагаемой молекулярной массой 31 кД. Продукт диализа анализируют с помощью SDS-PAGE в отсутствие восстанавливающего агента (дорожка 3) и в присутствии восстанавливающего агента (дорожка 2) и определяют окрашиванием кумасси. В качестве контроля hGH-FOS, который не смешивают с капсидными частицами, также вносят в гель в присутствии восстанавливающего агента (дорожка 1).
Сдвиг hGH-FOS до молекулярной массы приблизительно 53 кД отмечают в присутствии капсидного белка HBcAg-JUN, что позволяет предположить, что имеет место эффективное связывание hGH-FOS с HBcAg-JUN.
Пример 23:
Инсерция пептида, содержащего остаток лизина, в эпитоп с/е1 НВсАg(1-149)
Эпитоп с/е1 (остатки 72-88) НВсАg расположен в концевом участке на поверхности капсида вируса гепатита В (НВсАg). Фрагмент данного участка (Пролин 79 и Аланин 80) генетическим путем замещают пептидом Gly-Gly-Lys-Gly-Gly (конструкция HBcAg-Lys). Интродуцированный остаток лизина содержит в боковой цепи реакционную аминогруппу, которая может быть использована для межмолекулярного химического перекрестного связывания частиц НВсАg с любым антигеном, содержащим свободную группу цистеина.
Последовательность ДНК HBcAg-Lys получают с помощью ПЦР: Два фрагмента, кодирующих фрагменты НВсАg (остатки аминокислот 1-78 и 81-149), раздельно амплифицируют с помощью ПЦР. Праймеры, используемые для данных ПЦР, интродуцируют также последовательность ДНК, кодирующую пептид Gly-Gly-Lys-Gly-Gly. Фрагмент НВсАg (1-78) амплифицируют из рЕсо63, используя праймеры EcoRIHBcAg(s) и Lys-HBcAg(as). Фрагмент НВсАg (81-149) амплифицируют из рЕсо63, используя праймеры Lys-HBcAg(s) и HBcAg(1-149)Hind(as). Праймеры Lys-HBcAg(as) и Lys-HBcAg(s) интродуцируют комплементарные последовательности ДНК на концах двух продуктов ПЦР, обеспечивая слияние двух продуктов ПЦР в последующей ПЦР для сборки. Собранные фрагменты амплифицируют с помощью ПЦР, используя праймеры EcoRIHBcAg(s) и HbcAg(1-149)Hind(as).
Для ПЦР используют 100 пмоль каждого олигонуклеотида и 50 нг матричных ДНК в 50 мкл реакционных смесей, содержащих 2 ед. полимеразы Pwo, 0,1 мМ dNTP и 2 мМ MgSO4. Для обеих реакций температурный циклинг проводят следующим образом: 94°С в течение 2 мин с последующими 30 циклами, состоящими из стадий: 94°С (1 мин), 50°С (1 мин), 72°С (2 мин).
Последовательности праймеров:
EcoRIHBcAg(s):
(5’-CCGGAATTCATGGACATTGACCCTTATAAAG-3’) (SEQ ID NO: 79);
Lys-HBcAg(as):
(5’-CCTAGAGCCACCnTGCCACCATCTTCTAAATTAGTACCCACCCAGGTAGC-3’) (SEQ ID NO: 80);
Lys-HBcAg(s):
(5’-GAAGATGGTGGCAAAGGTGGCTCTAGGGACCTAGTAGTCAGTTATGTC-3’)
(SEQ ID NO: 81);
HBcAg(1-149)Hind(as):
(5’-CGCGTCCCAAGCTTCTAAACAACAGTAGTCTCCGGAAG-3’) (SEQ ID NO: 82).
Для слияния с помощью ПЦР двух полученных с использованием ПЦР фрагментов 100 пмоль праймеров EcoRIHBcAg(s) и HBcAg(1-149)Hind(as) используют со 100 нг двух очищенных полученных при ПЦР фрагментов в 50 мкл реакционной смеси, содержащей 2 ед. полимеразы Pwo, 0,1 мМ dNTP и 2 мМ МgSO4. Используют следующие условия ПЦР-циклинга: 94°С в течение 2 мин с последующими 30 циклами, состоящими из стадий: 94°С (1 мин), 50°С (1 мин), 72°С (2 мин). Собранный продукт ПЦР анализируют с помощью гель-электрофореза с использованием агарозы, очищают и разрезают в течение 19 час в подходящем буфере рестриктазами EcoRI and HindIII. Фрагмент разрезанной ДНК лигируют в разрезанный с помощью EcoRI/HindIII вектор рКК для создания экспрессионного вектора pKK-HBcAg-Lys. Инсерцию продукта ПЦР определяют рестрикционным анализом с помощью EcoRI/HindIII и путем секвенирования ДНК инсерции.
Пример 24:
Экспрессия и частичная очистка HBcAg-Lys
Штамм Е.coli XL-1 blue трансформируют рКК-HBcAg-Lys. 1 мл суточной культуры бактерий используют для инокуляции 100 мл среды LB, содержащей 100 мкг/мл ампициллина. Данную культуру выращивают в течение 4 час при 37°С до того, как OD при 600 нм не достигнет значения приблизительно 0,8. Индукцию синтеза HBcAg-Lys осуществляют путем добавления IPTG до конечной концентрации 1 мМ. После индукции бактерии продолжают встряхивать при 37°С в течение 16 час Бактерии собирают центрифугированием при 5000×g в течение 15 мин. Осадок замораживают при -20°С. Осадок оттаивают и ресуспендируют в буфере для лизиса бактерий (10 мМ Na2HPO4, рН 7,0, 30 мМ NaCl, 0,25% Tween 20, 10 мМ EDTA, 10 мМ DTT) с добавлением 200 мкг/мл лизоцима и 10 мкл бензоназы (Merck). Клетки инкубируют в течение 30 мин при комнатной температуре и разрушают в камере под давлением Френча (French). Triton X-100 добавляют к лизату до конечной концентрации 0,2% и инкубируют лизат в течение 30 мин на льду при периодическом встряхивании. Клетки Е.соli, несущие экспрессионную плазмиду pKK-HBcAg-Lys или контрольную плазмиду, используют для индукции экспрессии HBcAg-Lys с помощью IPTG. Перед добавлением IPTG берут образец бактериальной культуры, несущей плазмиду рКК-HBcAg-Lys, и культуры, несущей контрольную плазмиду. Через 16 час после добавления IPTG снова берут образцы культуры, содержащей рКК-HBcAg-Lys, и контрольной культуры. Экспрессию белка мониторируют с помощью SDS-PAGE с последующим окрашиванием кумасси.
Затем лизат центрифугируют в течение 30 мин при 12,000×g с целью удаления нерастворимого клеточного дебриса. Супернатант и осадок анализируют с помощью вестерн-блоттинга, используя моноклональные антитела против HBcAg (YVS1841, приобретаемые в фирме Accurate Chemical and Scientific Corp., Westbury, NY, USA), показывая, что значительное количество белка HBcAg-Lys является растворимым. Вкратце, лизаты, полученные из клеток E. соli, экспрессирующих HBcAg-Lys, и из контрольных клеток, центрифугируют при 14,000×g в течение 30 мин. Супернатант (=растворимую фракцию) и осадок (=нерастворимую фракцию) разделяют и разбавляют буфером для образцов SDS до равных объемов. Образцы анализируют, используя SDS-PAGE с последующим вестерн-блоттингом с использованием моноклонального антитела против НВсАg YVS1841.
Осветленный лизат клеток используют для центрифугирования в ступенчатом градиенте с использованием ступенчатого градиента сахарозы, состоящего из 4 мл 65% раствора сахарозы, на который накладывают 3 мл 15% раствора сахарозы, на который наносят 4 мл лизата бактерий. Образец центрифугируют в течение 3 час при 100000×g при 4°С. После центрифугирования 1 мл фракций из верхней части градиента собирают и анализируют с помощью SDS-PAGE с последующим окрашиванием кумасси. Белок HBcAg-Lys накапливается на границе раздела между 15 и 65% сахарозы, что указывает на то, что он формирует капсидную частицу. Большая часть бактериальных белков остается в не содержащем сахарозу верхнем слое градиента, вследствие чего центрифугирование частиц HBcAg-Lys в ступенчатом градиенте приводит как к обогащению, так и частичной очистке частиц.
Пример 25:
Химическое связывание пептида FLAG с HBcAg-Lys с использованием гетеробифункционального перекрестно-сшивающего линкера SPDP
Синтетический пептид FLAG с остатком цистеина на аминоконце (последовательность аминокислот CGGDYKDDDDK) химически связывают с очищенными частицами HBcAg-Lys с целью получения иммунного ответа против пептида FLAG. 600 мкл 95% очищенного раствора частиц HBcAg-Lys (2 мг/мл) инкубируют в течение 30 мин при комнатной температуре с гетеробифункциональным перекрестно-сшивающим линкером М-сукцинимидил 3-(2-пиридилдитио)пропионатом (SPDP) (0,5 мМ). После завершения реакции смесь диализуют в течение ночи против 1 л 50 мМ фосфатного буфера (рН 7,2) со 150 мМ NaCl с целью удаления свободного SPDP. Затем 500 мкл дериватизированного капсида HBcAg-Lys (2 мг/мл) смешивают с 0,1 мМ пептидом FLAG (содержащим аминоконцевой цистеин) в присутствии 10 мМ ЕDТА для предупреждения катализируемого металлом окисления сульфгидрила. Реакцию мониторируют по повышению оптической плотности раствора при 343 нм, что обусловлено выходом пиридин-2-тиона из SPDP при реакции со свободным цистеином пептида. Реакция дериватизированных остатков Lys с пептидом заканчивается приблизительно через 30 мин.
Частицы, несущие FLAG, инъецируют мышам.
Пример 26:
Конструирование pMPSV-gp140Cys
Ген др140 амплифицируют с помощью ПЦР из pCytTSgp140FOS, используя олигонуклеотиды gp140 CysEcoRI и SaIIgp140. Для проведения ПЦР используют 100 пмоль каждого олигонуклеотида и 50 нг матричных ДНК в 50 мкл реакционных смесей, содержащих 2 ед. полимеразы Pwo, 0,1 мМ dNTP и 2 мМ MgSO4. Для обеих реакций температурный циклинг проводят следующим образом: 94°С в течение 2 мин с последующими 30 циклами, состоящими из стадий: 94°С (0,5 мин), 55°С (0,5 мин), 72°С (2 мин).
Продукт ПЦР очищают, используя набор QiaEXII, разрезанный с помощью SaII/EcoRI, и лигируют в вектор pMPSVHE, разрезанный теми же ферментами.
Последовательности олигонуклеотидов:
Gp140CysEcoRI:
5’-GCCGAATTCCTAGCAGCTAGCACCGAATTTATCTAA-3’ (SEQ ID NO: 83);
SaIIgp140
5’-GGTTAAGTCGACATGAGAGTGAAGGAGAAATAT-3’ (SEQ ID NO: 84).
Пример 27:
Экспрессия pMPSVgp140Cys
pMPSVgp140Cys (20 мкг) линеаризуют, разрезая рестриктазой. Реакцию прекращают экстракцией фенолом/хлороформом с последующим осаждением изопропанолом линеаризованной ДНК. Реакцию рестрикции проверяют гель-электрофорезом в агарозе. Для трансфекции 5,4 мкг линеаризованного pMPSVgp140-Cys смешивают с 0,6 мкг линеаризованного pSV2Neo в 30 мкл Н2О и добавляют 30 мкл 1 М раствора СаСl2. После добавления 60 мкл фосфатного буфера (50 мМ HEPES, 280 мМ NaCl, 1,5 мМ Nа2РО4, рН 7,05) раствор взбалтывают в течение 5 секунд, а затем инкубируют при комнатной температуре в течение 25 секунд. Раствор немедленно добавляют к 2 мл среды НР-1, содержащей 2% FCS (2% среда FCS). Среду конфлюэнтной на 80% культуры клеток ВНК21 (в 6-луночной плате) затем замещают ДНК-содержащей средой. После инкубирования в течение 5 часов при 37°С в инкубаторе с CO2 ДНК-содержащую среду удаляют и замещают 2 мл 15% глицерина в 2% среде FCS. Глицерин-содержащую среду удаляют после 30-секундной фазы инкубирования и промывают клетки 5 мл среды НР-1, содержащей 10% FCS. Наконец, добавляют 2 мл свежей среды НР-1, содержащей 10% FCS.
Отбирают стабильно трансфицированные клетки и выращивают на селективной среде (среда НР-1 с добавлением G418) при 37°С в инкубаторе с CO2. Когда смешанная популяция вырастает до конфлюэнтности, культуру делят на две чашки с последующим периодом роста в течение 12 час при 37°С. Одну чашку с клетками переносят в условия культивирования при 30°С для индукции экспрессии растворимого GP140-Cys. Другую чашку сохраняют при 37°С.
Экспрессию растворимого GP140-Cys определяют с помощью вестерн-блоттинга. Культуральную среду (0,5 мл) осаждают метанолом/хлороформом и ресуспендируют осадок в буфере для образцов для SDS-PAGE. Образцы нагревают в течение 5 минут при 95°С перед внесением в 15% акриламидный гель. После SDS-PAGE белки переносят на нитроцеллюлозные мембраны Protan (Schleicher & Schuell, Germany), как предложено в разделе, написанном Bass и Yang, в монографии под ред. Creighton Т.Е. Практические подходы к функции белка (Protein Function: A Practical Approach), 2-е изд., IRL Press, Oxford (1997), стр.29-55. Мембрану блокируют 1% бычьим альбумином (Sigma) в TBS (10 × TBS/л: 87,7 г NaCl, 66,1 г Trizma гидрохлорида (Sigma) и 9,7 г Trizma основания (Sigma), pH 7,4) в течение 1 часа при комнатной температуре с последующим инкубированием с антителом против GP-140 или GP-160 в течение 1 часа. Блот промывают 3 раза по 10 минут с помощью TBS-T (TBS с добавлением 0,05% Tween 20) и инкубируют в течение 1 часа с конъюгатом щелочной фосфатазы с IgG против мыши/кролика/обезьяны/человека. После промывания 2 раза в течение 10 минут в TBS-T и 2 раза в течение 10 минут в TBS проводят проявляющую реакцию с использованием реагентов для детекции щелочной фосфатазы (10 мл буфера АР (100 мМ Трис/HCl, 100 мМ NaCl, pH 9,5) с 50 мкл раствора NBT (7,7% нитро синего тетразолия (Sigma) в 70% диметилформамиде) и 37 мкл раствора Х-фосфата (5% 5-бром-4-хлор-3-индолилфосфата в диметилформамиде).
Пример 28
Очистка GP140Cys
Антитело против gр120 ковалентно связывают с NHS/EDC-актвированным декстраном и упаковывают в хроматографическую колонку. Супернатант, содержащий GP140Cys, помещают на колонку и после достаточного промывания элюируют GP140Cys, используя 0,1 М HCl. Элюат нейтрализуют непосредственно во время сбора, используя 1 М Трис рН 7,2, находящийся в пробирках для сбора.
Поскольку во время очистки могло бы произойти образование дисульфидной связи, собранный образец обрабатывают 10 мМ DTT в 10 мМ Трис, рН 7,5 в течение 2 час при 25°С.
DTT удаляют при последующем диализе против 10 мМ Mes, 80 мМ NaCI, рН 6,0. Наконец, GP140Cys смешивают с альфавирусными частицами, содержащими остаток JUN в Е2, как описано в Примере 16.
Пример 29
Конструирование PLA2-Cys
Ген PLA2 амплифицируют с помощью ПЦР из pAVSPLAfos, используя олигонуклеотиды EcoRIPLA и PLA-Cys-hind. Для проведения ПЦР используют 100 пмоль каждого олигонуклеотида и 50 нг матричных ДНК в 50 мкл реакционных смесей, содержащих 2 ед. полимеразы Pwo, 0,1 мМ dNTP и 2 мМ МgSO4. Для обеих реакций температурный циклинг проводят следующим образом: 94°С в течение 2 мин с последующими 30 циклами, состоящими из стадий: 94°С (0,5 мин), 55°С (0,5 мин), 72°С (2 мин).
Продукт ПЦР очищают, используя набор QiaEXII, разрезанный с помощью EcoRI/HindIII, и лигируют в вектор pAV3, разрезанный теми же ферментами.
Последовательности олигонуклеотидов:
EcoRIPLA:
5’-TAACCGAATTCAGGAGGTAAAMGATATGG-3’ (SEQ ID NO: 85)
PLACys-hind:
5’-GAAGTAAAGCTTTTAACCACCGCAACCACCAGAAG-3’ (SEQ ID NO: 86).
Пример 30
Экспрессия и очистка PLA-cys
Для цитоплазматической продукции меченых Cys Е.соli XL-1-Blue трансформируют векторами pAV3::PLA и pPLA-Cys. Культуру инкубируют в обогащенной среде в присутствии ампициллина при 37°С, встряхивая. По достижении оптической плотности (550 нм) добавляют 1 мМ IPTG и продолжают инкубирование в течение последующих 5 час. Клетки собирают с помощью центрифугирования, ресуспендируют в подходящем буфере (например, Трис-HCl, рН 7,2, 150 мМ NaCl), содержащем ДНКазу, РНКазу и лизоцим, и разрушают, проводя через камеру под давлением Френча (French). После центрифугирования (SorvaII RC-5C, ротор SS34, 15000 об/мин, 10 мин, 4°С) осадок ресуспендируют в буфере для промывания телец включений (20 мМ Трис-HCl, 23% сахарозы, 0,5% Triton Х-100, 1 мМ EDTA, рН 8) при 4°С и повторно центрифугируют, как описано выше. Данную процедуру повторяют, пока супернатант, полученный после центрифугирования, не становится практически прозрачным. Тельца включений ресуспендируют в 20 мл буфера для солюбилизации (5,5 М гидрохлорида гуанидина, 25 мМ Трис-HCl, рН 7,5) при комнатной температуре и удаляют нерастворимый материал центрифугированием с последующим пропусканием супернатанта через стерильный фильтр (0,45 мкм). Раствор белка хранят при 4°С в течение по меньшей мере 10 час в присутствии 10 мМ EDTA и 100 мМ DTT, а затем трижды диализуют против 10 объемов 5,5 М гидрохлорида гуанидина, 25 мМ Трис-HCl, 10 мМ EDTA, рН 6. Раствор дважды диализуют против 5 л 2 М мочевины, 4 мМ EDTA, 0,1 М NH4Cl, 20 мМ бората натрия (рН 8,3) в присутствии подходящей окислительно-восстановительной пары (окисленный глутатион/восстановленный глутатион; цистин/цистеин). Переуложенный белок подвергают ионообменной хроматографии. Белок хранят в подходящем буфере с рН выше 7 в присутствии 2-10 мМ DTT для того, чтобы сохранить цистеиновые остатки в восстановленной форме. Перед связыванием белка с частицами альфавируса DTT удаляют, пропуская раствор белка через колонку для гель-фильтрации Sephadex G-25.




































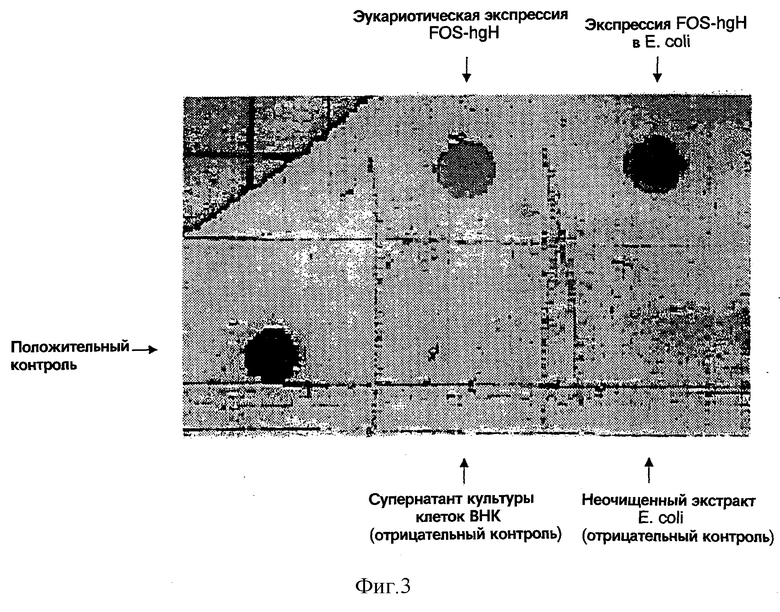

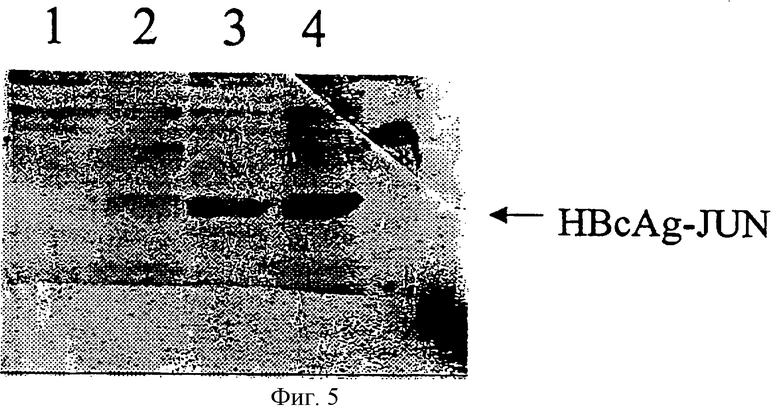
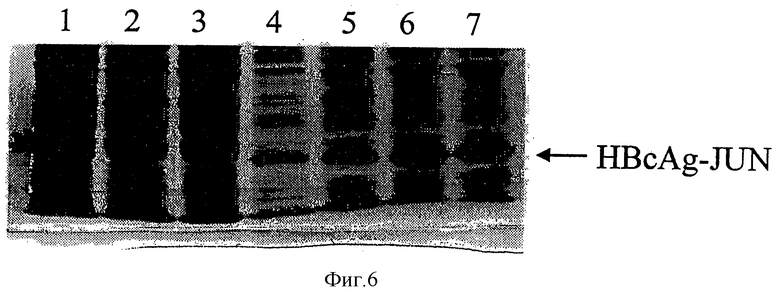
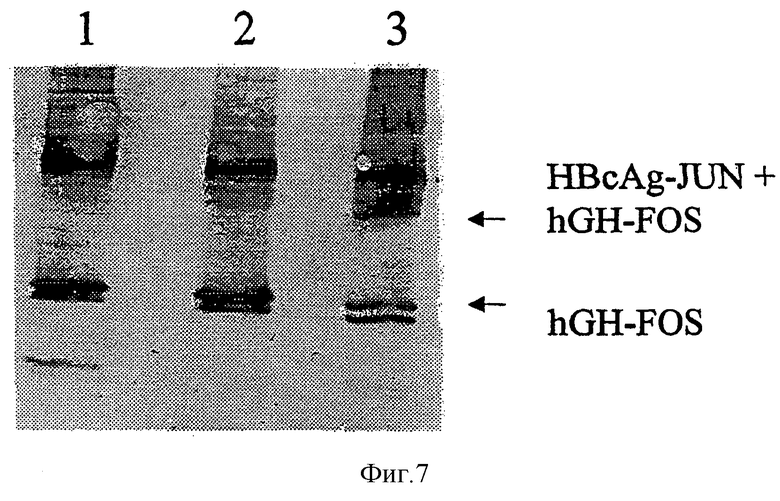
Изобретение относится к медицине и касается композиции, способа получения не природной упорядоченной и содержащей повторы антигенной матрицы, способа терапевтического лечения и способа иммунизации. Сущность изобретения включает композиции, которые используют для приготовления вакцин для профилактики инфекционных болезней, лечения аллергий и лечения злокачественных новообразований. В различных вариантах осуществления изобретения представлены вирус, вирусоподобная частица, вирусная капсидная частица, фаг или их рекомбинантные формы, покрытые в высокой степени упорядоченно и с повторами любым желаемым антигеном, что достигается специфическими взаимодействиями. В одном из специфических вариантов осуществления многофункциональная новая технология, основанная на системе кассетного типа (Технология альфавакцины), позволяет получить вирусные частицы, покрытые антигеном. Другие специфические варианты осуществления позволяют получить покрытые антигеном частицы, подобные вирусу гепатита В, или покрытые антигеном частицы, подобные вирусу кори. Преимущество изобретения заключается в разработке усовершенствованных вакцин, которые стимулируют сильный иммунный ответ. 6 н. и 38 з.п. ф-лы, 7 ил.
Приоритет по пунктам:
| АНТИМИКРОБНАЯ КОМПОЗИЦИЯ | 1989 |
|
RU2092180C1 |
| Экономайзер | 0 |
|
SU94A1 |
| FROLOV I | |||
| et al., Alphavirus – based expression vectors: strategies and applications, Proc | |||
| Natl., Acad | |||
| Sci USA,1996, oct.15, 93(21), pp.11371-11377 | |||
| ESCPOSITO G | |||
| et al., Comformational study of a short pertussis toxin T cell epitope incorporated in a multiple antigen peptide template by CD and two-dimensional NMP | |||
| Analysis of the structural effect on the activity of synthetic immunogens, Eur | |||
| J | |||
| Biochem., 1993, oct 1, 217(1), рр.171-187. | |||

