Перекрестная ссылка на родственную заявку
Данная заявка является частичным продолжением заявки США с регистрационным номером 09/378088, поданной 20 августа 1999 года.
Предпосылки изобретения
Жесткокрылые представляют собой значительную группу сельскохозяйственных вредителей, которые вызывают сильное повреждение посевов каждый год. Примеры жесткокрылых вредителей включают в себя блошку длинноусую и долгоносика люцернового.
Долгоносик люцерновый, Hypera postica, и близкородственный долгоносик египетский люцерновый, Hypera brunneipennis, являются наиболее важными насекомыми-вредителями, обитающими в США, с 2,9 миллиона акров, пораженных в 1984 году. Ежегодная сумма 20 миллионов долларов расходуется для борьбы с этими вредителями. Долгоносик египетский люцерновый является преобладающим видом в юго-западной части США, где он подвергается диапаузе (летованию) (т.е. спячке) во время жарких летних месяцев. Во всех других отношениях он идентичен долгоносику люцерновому, который преобладает во всей остальной части США.
Личиночная стадия является наиболее повреждающей в жизненном цикле долгоносика. Питаясь на растущих кончиках растений люцерны, личинка приводит к скелетированию листьев (оставлению только жилок листьев), задержке роста, уменьшенному росту растений и, в конечном счете, снижениям в урожае. Несколько нашествий вредителей могут погубить всю заготовку сена. Взрослые особи, также питающиеся листьями, вызывают дополнительное, но менее значительное повреждение.
Приблизительно 10 миллионов акров кукурузы США заражаются комплексом видов блошки длинноусой каждый год. Комплекс видов блошки длинноусой включает северную блошку длинноусую, Diabrotica barberi, южную блошку длинноусую, D. undecimpunctata howardi, и западную блошку длинноусую, D. virgifera virgifera. Живущие в почве личинки этих видов Diabrotica питаются на корнях растения кукурузы, вызывая полегание. Полегание в конечном счете снижает урожай кукурузы и часто приводит к гибели растения. Питаясь на «шелке» початков кукурузы, взрослые жуки понижают опыление и, следовательно, вредным образом влияют на урожай кукурузы на одно растение. Кроме того, взрослые особи и личинки рода Diabrotica нападают на посевы тыквенных (огурцов, дынь, тыквы и т.д.) и многочисленные овощные и полевые культуры в коммерческом производстве, а также выращиваемые в приусадебных огородах.
Борьба с блошкой длинноусой ведется частично способами культивации, такими как чередование культур (ротация) и применение высоких уровней азота для стимуляции роста придаточной корневой системы. Однако химические инсектициды являются наиболее надежными для гарантии желательного уровня защиты. Инсектициды либо наносятся на почву, либо включаются в почву. Проблемами, связанными с применением некоторых химических инсектицидов, являются загрязнение окружающей среды и развитие устойчивости среди обрабатываемых популяций насекомых.
Почвенный микроорганизм Bacillus thuringiensis (B.t.) является грамположительной, спорообразующей бактерией, характеризующейся белковыми включениями в параспорах, которые видны под микроскопом в виде кристаллов отчетливой формы. Некоторые штаммы B.t. продуцируют белки, которые являются токсичными для конкретных отрядов вредителей. Некоторые гены токсинов B.t. были выделены и секвенированы и были получены и одобрены для применения продукты на основе рекомбинантной ДНК B.t. Кроме того, с использованием способов генной инженерии разрабатываются новые подходы для доставки эндотоксинов B.t. в сельскохозяйственные среды, в том числе применение генетически сконструированных растений с генами эндотоксинов для устойчивости к насекомым и применение стабилизированных интактных микробных клеток в качестве доставляющих носителей эндотоксинов B.t. (Gaertner, F.H., L.Kim [1988] TIBTECH 6:S4-S7). Таким образом, выделенные гены эндотоксинов B.t. становятся коммерчески ценными.
Коммерческое применение В.t.-пестицидов первоначально было ограничено узким диапазоном чешуекрылых (гусеничных) вредителей. Препараты спор и кристаллов В.thuringiensis подвида kurstaki использовали в течение многих лет в качестве коммерческих инсектицидов для чешуекрылых вредителей. Например, В.thuringiensis сорта kurstaki HD-1 продуцирует кристаллический δ-эндотоксин, который является токсичным для личинок ряда чешуекрылых насекомых.
Однако в недавние годы исследователи открыли B.t.-пестициды со специфичностями для гораздо более широкого круга вредителей. Например, другие виды B.t., а именно israelensis и tenebrionis (a.k.a. B.t. M-7, a.k.a. B.t. san diego) использовали коммерчески для борьбы с насекомыми отрядов Diptera и Coleoptera соответственно (Gaertner, F.H. [1989] "Cellular Delivery Systems for Insecticidal Proteins: Living and Non-Living Microorganisms", in Controlled Delivery of Crop Protection Agents, R.M.Wilkins, ed., Taylor and Francis, New York and London, 1990, pp.245-255). См. также Couch, T.L. (1980) "Mosquito Pathogenicity of Bacillus thuringiensis var. israelensis". Developments in Industrial Microbiology 22:61-76; Beegle, C.C., (1978) "Use of Entomogenous Bacteria in Agroecosystems", Developments in Industrial Microbiology 20:97-104. Krieg, A., A.M. Huger, G.A. Langenbruch, W. Schnetter (1983) Z. ang. Ent. 96:500-508 описывают Bacillus thuringiensis var. tenebrionis, которая, как сообщается, активна против двух жуков отряда Coleoptera. Это колорадские картофельные жуки, Leptinotarsa decemlineata и Agelastica alni.
Недавно были идентифицированы новые подвиды В.t. и были выделены гены, ответственные за активные δ-эндотоксиновые белки (Hofte, H., H.R.Whiteley [1989] Microbiological Reviews 52 (2):242-255). Hofte и Whiteley классифицировали гены кристаллических белков B.t. на четыре основных класса. Эти классы были Cryl (специфические для Lepidoptera), CryII (специфические для Lepidoptera и Diptera), CryIII (специфические для Coleoptera) и CryIV (специфические для Diptera). Сообщалось открытие штаммов, специфически токсичных в отношении других вредителей (Feitelson, J.S., J.Payne, L.Kim [1992] Bio/Technology 10:271-275).
Номенклатура и классификационная схема 1989 года Hofte и Whiteley для кристаллических белков основывалась как на расшифрованной аминокислотной последовательности, так и на круге хозяев токсина. Эта система была приспособлена для охвата четырнадцати различных типов генов токсинов, которые были разделены на пять основных классов. По мере обнаружения дополнительных генов токсинов эта система начала становиться неприменимой для работы, так как было обнаружено, что гены со сходными последовательностями имеют значимо отличающиеся инсектицидные специфичности. Была предложена пересмотренная и исправленная схема номенклатуры, которая основывается только на аминокислотной идентичности (Crickmore et al. [1996] Society for Invertebrate Pathology, 29th Annual Meeting, 3rd International Colloquium on Bacillus thuringiensis, University of Cordoba, Cordoba, Spain, September 1-6, abstract). Мнемоника «cry» была сохранена для всех классов генов токсинов, за исключением cytA и cytB, которые остаются отдельным классом. Римские цифры были заменены на арабские цифры в первичном ранге, а скобки в третичном ранге были удалены. Текущие границы обозначают приблизительно 95%-ную (третичный ранг), 75%-ную (вторичный ранг) и 48%-ную (первичный ранг) идентичность последовательности. Многие из первоначальных названий были сохранены, с упомянутыми исключениями, хотя номер был заново классифицирован. См. также N.Crickmore, D.R.Zeigler, J.Feitelson, E.Schnepf, J.Van Rie, D.Lereclus, J.Baum, and D.H.Dean (1998), "Revisions of the Nomenclature for the Bacillus thuringiensis Pesticidal Crystal Proteins," Microbiology and Molecular Biology Reviews Vol. 62:807-813; и Crickmore, Zeigler, Feitelson, Schnepf, Van Rie, Lereclus, Baum and Dean, "Bacillus thuringiensis toxin nomenclature" (1999) http: //www. biols. susx. ас.uk/Home/Neil_Crickmore/Bt/index.html. Эта система использует свободно доступные приложения программного обеспечения CLUSTAL W и PHYLIP. Приложение NEIGHBOR в пакете PHYLIP использует алгоритм с арифметическими средними (UPGMA).
Клонирование и экспрессия гена кристаллического белка B.t. в Escherichia coli были описаны в опубликованной литературе (Schnepf, H.E., H.R.Whiteley [1981] Proc. Natl. Acad. Sci. USA 78:2893-2897). Оба патента США 4448885 и 4467036 описывают экспрессию кристаллического белка B.t. в Е.coli.
В патентах США 4797276 и 4853331 описан штамм В. thuringiensis tenebrionis (a.k.a. М-7, a.k.a. B.t. san diego), который может быть использован для борьбы с жесткокрылыми вредителями в различных условиях окружающей среды. В патенте США с номером 4918006 описаны токсины B.t., обладающие активностью против двукрылых. В патенте США 4849217 описаны изоляты B.t., которые обладают активностью против долгоносика люцернового. В патенте США 5208077 описаны активные против жесткокрылых изоляты Bacillus thuringiensis. В патенте США 5632987 описан токсин 130 кДа из PS80JJ1 как имеющий активность против блошки длинноусой. В WO 94/40162, которая является родственной данной заявке, описаны новые классы белков, которые являются токсичными для блошки длинноусой. В патентах США 5151363 и 4948734 описаны некоторые изоляты B.t., которые обладают активностью против нематод.
В патенте США 6083499 и WO 97/40162 описаны «бинарные токсины». Данное изобретение отличается от москитоцидных токсинов, продуцируемых Bacillus sphaericus. См. ЕР 454485; Davidson et al. (1990), "Interaction of the Bacillus sphaericus mosquito larvicidal proteins," Can. J. Microbiol. 36 (12):870-8; Baumann et al. (1988), "Sequence analysis of the mosquitocidal toxin genes encoding 51.4- and 41.9-kilodalton proteins from Bacillus sphaericus 2362 and 2297, J.Bacteriol. 170:2045-2050; Oei et al. (1992), "Binding of purified Bacillus sphaericus binary toxin and its deletion derivatives to Culex quinquefasciatus gut: elucidation of functional binding domains," Journal of General Microbiology 138 (7):1515-26.
Краткая сущность изобретения
Данное изобретение относится к новым материалам и способам борьбы с вредителями не млекопитающими. В предпочтительном варианте данное изобретение представляет материалы и способы для борьбы с жесткокрылыми вредителями. В более предпочтительных вариантах описанные здесь материалы и способы используют для борьбы с блошкой длинноусой - наиболее предпочтительно с западной блошкой длинноусой. Чешуекрылые вредители (в том числе мотылек кукурузный и Helicoverpa zea) могут уничтожаться пестицидными белками данного изобретения.
Данное изобретение преимущественно обеспечивает полинуклеотиды и пестицидные белки, кодируемые этими полинуклеотидами. В предпочтительных вариантах белок 40-50 кДа и белок 10-15 кДа используют вместе, причем эти белки в сочетании являются пестицидными. Таким образом, два класса белков данного изобретения могут быть названы «бинарными токсинами». В применении здесь термин «токсин» или «пестицидный белок» включает в себя любой класс этих белков. Применение белка 40-50 кДа с белком 10-15 кДа является предпочтительным, но не обязательно требуемым. Один класс полинуклеотидных последовательностей, описанных здесь, кодирует белки, которые имеют полноразмерную молекулярную массу приблизительно 40-50 кДа. В характерном варианте эти белки имеют молекулярную массу приблизительно 43-47 кДа. Второй класс полинуклеотидов данного изобретения кодирует пестицидные белки приблизительно 10-15 кДа. В конкретном варианте эти белки имеют молекулярную массу приблизительно 13-14 кДа. Должно быть понятно, что каждый тип токсина/гена является аспектом данного изобретения. В особенно предпочтительном варианте белок 40-50 кДа данного изобретения используют в комбинации с белком 10-15 кДа. Таким образом, белки данного изобретения могут быть использованы для изменения и/или облегчения активности других белковых токсинов.
Данное изобретение включает в себя полинуклеотиды, которые кодируют токсины 40-50 кДа или 10-15 кДа, полинуклеотиды, которые кодируют части или фрагменты полноразмерных токсинов, которые сохраняют пестицидную активность (предпочтительно при использовании в комбинации), и полинуклеотиды, которые кодируют оба типа токсинов. Новые примеры слитых (гибридных) белков (белок 40-50 кДа и белок 10-15 кДа, слитые вместе) и кодирующие их полинуклеотиды также описаны здесь.
В некоторых вариантах, В.t.-токсины, применимые в соответствии с данным изобретением, включают в себя токсины, которые могут быть получены из описанных здесь новых изолятов B.t. Должно быть понятно, что при совместном использовании, например, токсинов 40-50 кДа и 10-15 кДа один тип токсина может быть получен из одного изолята, а другой тип токсина может быть получен из другого изолята.
Данное изобретение включает в себя также применение вариантов приводимых в качестве примеров изолятов и токсинов В.t., которые имеют по существу такие же активные против жесткокрылых насекомых свойства, что и приводимые в качестве конкретных примеров изоляты и токсины B.t. Такие вариантные изоляты могут включать, например, мутанты. Процедуры получения мутантов хорошо известны в области микробиологии. Для этой цели интенсивно используются ультрафиолетовый свет и химические мутагены, такие как нитрозогуанидин.
В предпочтительных вариантах данное изобретение относится к растениям и клеткам растений, имеющим по меньшей мере один выделенный полинуклеотид данного изобретения. Предпочтительно клетки трансгенных растений экспрессируют пестицидные токсины в тканях, потребляемых вредителями-мишенями.
Альтернативно, изоляты B.t. данного изобретения или описанные здесь рекомбинантные микробы, экспрессирующие токсины, могут быть использованы для борьбы с вредителями. В этом отношении данное изобретение включает обработку по существу интактными клетками B.t., и/или рекомбинантными клетками, содержащими экспрессируемые токсины данного изобретения, обработанными для пролонгирования пестицидной активности, когда по существу интактные клетки наносят на окружающую среду вредителя-мишени. Обработанная клетка действует в качестве защитного покрытия для пестицидного токсина.
Токсины данного изобретения являются пероральными интоксикантами, которые действуют на клетки средней кишки насекомого при проглатывании насекомым-мишенью. Таким образом, проглатыванием рекомбинантных клеток-хозяев, которые, например, экспрессируют токсины, насекомое-мишень контактирует тем самым с белками данного изобретения, которые являются токсичными для данного вредителя. Это приводит к защите от вредителя-мишени.
Краткое описание чертежей
Фигура 1 показывает три примера пестицидных токсинов 43-47 кДа, а также консенсусную последовательность для этих пестицидных токсинов.
Фигура 2 показывает взаимосвязь последовательностей 14 и 45 кДа PS80JJ1 (SEQ ID NO:31 и SEQ ID NO:10).
Фигура 3 показывает сравнение величин LC50 из исследования по смешиванию примера 23.
Фигура 4 показывает сопоставления токсинов и генов 51 и 42 кДа Bacillus sphaericus и токсина и гена 45 кДа 149В1.
Фигура 5 показывает сопоставления нуклеотидных последовательностей токсинов и генов 51 и 42 кДа Bacillus sphaericus и токсина и гена 45 кДа 149В1.
Краткое описание последовательностей
SEQ ID NO:1 представляет собой N-концевую последовательность из 5 аминокислот токсина приблизительно 45 кДа 80JJ1.
SEQ ID NO:2 представляет собой N-концевую последовательность из 25 аминокислот токсина приблизительно 45 кДа 80JJ1.
SEQ ID NO:3 представляет собой N-концевую последовательность из 24 аминокислот токсина приблизительно 14 кДа 80JJ1.
SEQ ID NO:4 представляет собой N-концевую последовательность токсина приблизительно 47 кДа из 149В1.
SEQ ID NO:5 представляет собой N-концевую аминокислотную последовательность из 50 аминокислот для очищенного белка приблизительно 14 кДа из PS149B1.
SEQ ID NO:6 представляет собой N-концевую последовательность токсина приблизительно 47 кДа из 167Н2.
SEQ ID NO:7 представляет собой N-концевую последовательность из 25 аминокислот для очищенного белка приблизительно 14 кДа из PS167H2.
SEQ ID NO:8 представляет собой олигонуклеотидный зонд для гена, кодирующего токсин 44,3 кДа PS80JJ1 и является прямым праймером для PS149B1 и PS167H2, используемым в соответствии с данным изобретением.
SEQ ID NO:9 представляет собой обратный праймер для PS149B1 и PS167H2, используемый в соответствии с данным изобретением.
SEQ ID NO:10 представляет собой нуклеотидную последовательность гена, кодирующего токсин приблизительно 45 кДа PS80JJ1.
SEQ ID NO:11 представляет собой аминокислотную последовательность для токсина приблизительно 45 кДа PS80JJ1.
SEQ ID NO:12 представляет собой частичную нуклеотидную последовательность гена, кодирующего токсин приблизительно 44 кДа PS149B1.
SEQ ID NO:13 представляет собой частичную аминокислотную последовательность для токсина приблизительно 44 кДа PS149B1.
SEQ ID NO:14 представляет собой частичную нуклеотидную последовательность гена, кодирующего токсин приблизительно 44 кДа PS167H2.
SEQ ID NO:15 представляет собой частичную аминокислотную последовательность для токсина приблизительно 44 кДа PS167H2.
SEQ ID NO:16 представляет собой пептидную последовательность, используемую в конструировании праймера в соответствии с данным изобретением.
SEQ ID NO:17 представляет собой пептидную последовательность, используемую в конструировании праймера в соответствии с данным изобретением.
SEQ ID NO:18 представляет собой пептидную последовательность, используемую в конструировании праймера в соответствии с данным изобретением.
SEQ ID NO:19 представляет собой пептидную последовательность, используемую в конструировании праймера в соответствии с данным изобретением.
SEQ ID NO:20 представляет собой нуклеотидную последовательность, соответствующую пептиду SEQ ID NO:16.
SEQ ID NO:21 представляет собой нуклеотидную последовательность, соответствующую пептиду SEQ ID NO:17.
SEQ ID NO:22 представляет собой нуклеотидную последовательность, соответствующую пептиду SEQ ID NO:18.
SEQ ID NO:23 представляет собой нуклеотидную последовательность, соответствующую пептиду SEQ ID NO:19.
SEQ ID NO:24 представляет собой обратный праймер на основе обращенного комплемента SEQ ID NO:22.
SEQ ID NO:25 представляет собой обратный праймер на основе обращенного комплемента SEQ ID NO:23.
SEQ ID NO:26 представляет собой прямой праймер на основе токсина 44,3 кДа PS80JJ1.
SEQ ID NO:27 представляет собой обратный праймер на основе токсина 44,3 кДа PS80JJ1.
SEQ ID NO:28 представляет собой общую последовательность, представляющую новый класс токсинов в соответствии с данным изобретением.
SEQ ID NO:29 представляет собой олигонуклеотидный зонд, используемый в соответствии с данным изобретением.
SEQ ID NO:30 представляет собой нуклеотидную последовательность всего генетического локуса, содержащую открытые рамки считывания обоих токсинов 14 и 45 кДа PS80JJ1 и фланкирующие нуклеотидные последовательности.
SEQ ID NO:31 представляет собой нуклеотидную последовательность открытой рамки считывания токсина 14 кДа PS80JJ1.
SEQ ID NO:32 представляет собой расшифрованную аминокислотную последовательность токсина 14 кДа PS80JJ1.
SEQ ID NO:33 представляет собой обратный олигонуклеотидный праймер, используемый в соответствии с данным изобретением.
SEQ ID NO:34 представляет собой нуклеотидную последовательность всего генетического локуса, содержащую открытые рамки считывания обоих токсинов 14 и 44 кДа PS167H2 и фланкирующие нуклеотидные последовательности.
SEQ ID NO:35 представляет собой нуклеотидную последовательность гена, кодирующего токсин приблизительно 14 кДа PS167H2.
SEQ ID NO:36 представляет собой аминокислотную последовательность для токсина приблизительно 14 кДа PS167H2.
SEQ ID NO:37 представляет собой нуклеотидную последовательность гена, кодирующего токсин приблизительно 44 кДа PS167H2.
SEQ ID NO:38 представляет собой аминокислотную последовательность для токсина приблизительно 44 кДа PS167H2.
SEQ ID NO:39 представляет собой нуклеотидную последовательность всего генетического локуса, содержащую открытые рамки считывания обоих токсинов 14 и 44 кДа PS149B1 и фланкирующие нуклеотидные последовательности.
SEQ ID NO:40 представляет собой нуклеотидную последовательность гена, кодирующего токсин приблизительно 14 кДа PS149B1.
SEQ ID NO:41 представляет собой аминокислотную последовательность для токсина приблизительно 14 кДа PS149B1.
SEQ ID NO:42 представляет собой нуклеотидную последовательность гена, кодирующего токсин приблизительно 44 кДа PS149B1.
SEQ ID NO:43 представляет собой аминокислотную последовательность для токсина приблизительно 44 кДа PS149B1.
SEQ ID NO:44 представляет собой оптимизированную для кукурузы последовательность гена, кодирующую токсин приблизительно 14 кДа 80JJ1.
SEQ ID NO:45 представляет собой оптимизированную для кукурузы последовательность гена, кодирующую токсин приблизительно 44 кДа 80JJ1.
SEQ ID NO:46 представляет собой последовательность ДНК обратного праймера, используемого в примере 15, ниже.
SEQ ID NO:47 представляет собой последовательность ДНК прямого праймера (см. пример 16).
SEQ ID NO:48 представляет собой последовательность ДНК обратного праймера (см. пример 16).
SEQ ID NO:49 представляет собой последовательность ДНК прямого праймера (см. пример 16).
SEQ ID NO:50 представляет собой последовательность ДНК обратного праймера (см. пример 16).
SEQ ID NO:51 представляет собой последовательность ДНК из PS131W2, которая кодирует белок 14 кДа.
SEQ ID NO:52 представляет собой аминокислотную последовательность белка 14 кДа PS131W2.
SEQ ID NO:53 представляет собой частичную последовательность ДНК из PS131W2 для белка 44 кДа.
SEQ ID NO:54 представляет собой частичную аминокислотную последовательность для белка 44 кДа PS131W2.
SEQ ID NO:55 представляет собой последовательность ДНК из PS158T3, которая кодирует белок 14 кДа.
SEQ ID NO:56 представляет собой аминокислотную последовательность белка 14 кДа PS158T3.
SEQ ID NO:57 представляет собой частичную последовательность ДНК из PS158T3 для белка 44 кДа.
SEQ ID NO:58 представляет собой частичную аминокислотную последовательность для белка 44 кДа PS158T3.
SEQ ID NO:59 представляет собой последовательность ДНК из PS158X10, которая кодирует белок 14 кДа.
SEQ ID NO:60 представляет собой аминокислотную последовательность белка 14 кДа PS158X10.
SEQ ID NO:61 представляет собой последовательность ДНК из PS185FF, которая кодирует белок 14 кДа.
SEQ ID NO:62 представляет собой аминокислотную последовательность белка 14 кДа PS185FF.
SEQ ID NO:63 представляет собой частичную последовательность ДНК из PS185FF для белка 44 кДа.
SEQ ID NO:64 представляет собой частичную аминокислотную последовательность для белка 44 кДа PS185FF.
SEQ ID NO:65 представляет собой последовательность ДНК из PS185GG, которая кодирует белок 14 кДа.
SEQ ID NO:66 представляет собой аминокислотную последовательность белка 14 кДа PS185GG.
SEQ ID NO:67 представляет собой последовательность ДНК из PS185GG для белка 44 кДа.
SEQ ID NO:68 представляет собой аминокислотную последовательность для белка 44 кДа PS185GG.
SEQ ID NO:69 представляет собой последовательность ДНК из PS185L12, которая кодирует белок 14 кДа.
SEQ ID NO:70 представляет собой аминокислотную последовательность белка 14 кДа PS185L12.
SEQ ID NO:71 представляет собой последовательность ДНК из PS185W3, которая кодирует белок 14 кДа.
SEQ ID NO:72 представляет собой аминокислотную последовательность белка 14 кДа PS185W3.
SEQ ID NO:73 представляет собой последовательность ДНК из PS186FF, которая кодирует белок 14 кДа.
SEQ ID NO:74 представляет собой аминокислотную последовательность белка 14 кДа PS186FF.
SEQ ID NO:75 представляет собой последовательность ДНК из PS187F3, которая кодирует белок 14 кДа.
SEQ ID NO:76 представляет собой аминокислотную последовательность белка 14 кДа PS187F3.
SEQ ID NO:77 представляет собой частичную последовательность ДНК из PS187F3 для белка 44 кДа.
SEQ ID NO:78 представляет собой частичную аминокислотную последовательность для белка 44 кДа PS187F3.
SEQ ID NO:79 представляет собой последовательность ДНК из PS187G1, которая кодирует белок 14 кДа.
SEQ ID NO:80 представляет собой аминокислотную последовательность белка 14 кДа PS187G1.
SEQ ID NO:81 представляет собой частичную последовательность ДНК из PS187G1 для белка 44 кДа.
SEQ ID NO:82 представляет собой частичную аминокислотную последовательность для белка 44 кДа PS187G1.
SEQ ID NO:83 представляет собой последовательность ДНК из PS187L14, которая кодирует белок 14 кДа.
SEQ ID NO:84 представляет собой аминокислотную последовательность белка 14 кДа PS187L14.
SEQ ID NO:85 представляет собой частичную последовательность ДНК из PS187L14 для белка 44 кДа.
SEQ ID NO:86 представляет собой частичную аминокислотную последовательность для белка 44 кДа PS187L14.
SEQ ID NO:87 представляет собой последовательность ДНК из PS187Y2, которая кодирует белок 14 кДа.
SEQ ID NO:88 представляет собой частичную аминокислотную последовательность белка 14 кДа PS187Y2.
SEQ ID NO:89 представляет собой частичную последовательность ДНК из PS187Y2 для белка 44 кДа.
SEQ ID NO:90 представляет собой частичную аминокислотную последовательность для белка 44 кДа PS187Y2.
SEQ ID NO:91 представляет собой последовательность ДНК из PS201G, которая кодирует белок 14 кДа.
SEQ ID NO:92 представляет собой аминокислотную последовательность белка 14 кДа PS201G.
SEQ ID NO:93 представляет собой последовательность ДНК из PS201HH, которая кодирует белок 14 кДа.
SEQ ID NO:94 представляет собой аминокислотную последовательность белка 14 кДа PS201HH.
SEQ ID NO:95 представляет собой последовательность ДНК из PS201L3, которая кодирует белок 14 кДа.
SEQ ID NO:96 представляет собой аминокислотную последовательность белка 14 кДа PS201L3.
SEQ ID NO:97 представляет собой последовательность ДНК из PS204C3, которая кодирует белок 14 кДа.
SEQ ID NO:98 представляет собой аминокислотную последовательность белка 14 кДа PS204C3.
SEQ ID NO:99 представляет собой последовательность ДНК из PS204G4, которая кодирует белок 14 кДа.
SEQ ID NO:100 представляет собой аминокислотную последовательность белка 14 кДа PS204G4.
SEQ ID NO:101 представляет собой последовательность ДНК из PS204I11, которая кодирует белок 14 кДа.
SEQ ID NO:102 представляет собой аминокислотную последовательность белка 14 кДа PS204I11.
SEQ ID NO:103 представляет собой последовательность ДНК из PS204J7, которая кодирует белок 14 кДа.
SEQ ID NO:104 представляет собой аминокислотную последовательность белка 14 кДа PS204J7.
SEQ ID NO:105 представляет собой последовательность ДНК из PS236B6, которая кодирует белок 14 кДа.
SEQ ID NO:106 представляет собой аминокислотную последовательность белка 14 кДа PS236B6.
SEQ ID NO:107 представляет собой последовательность ДНК из PS242K10, которая кодирует белок 14 кДа.
SEQ ID NO:108 представляет собой аминокислотную последовательность белка 14 кДа PS242K10.
SEQ ID NO:109 представляет собой частичную последовательность ДНК из PS242K10 для белка 44 кДа.
SEQ ID NO:110 представляет собой частичную аминокислотную последовательность для белка 44 кДа PS242K10.
SEQ ID NO:111 представляет собой последовательность ДНК из PS246P42, которая кодирует белок 14 кДа.
SEQ ID NO:112 представляет собой аминокислотную последовательность белка 14 кДа PS246P42.
SEQ ID NO:113 представляет собой последовательность ДНК из PS69Q, которая кодирует белок 14 кДа.
SEQ ID NO:114 представляет собой аминокислотную последовательность белка 14 кДа PS69Q.
SEQ ID NO:115 представляет собой последовательность ДНК из PS69Q для белка 44 кДа.
SEQ ID NO:116 представляет собой аминокислотную последовательность для белка 44 кДа PS69Q.
SEQ ID NO:117 представляет собой последовательность ДНК из КВ54, которая кодирует белок 14 кДа.
SEQ ID NO:118 представляет собой аминокислотную последовательность белка 14 кДа КВ54.
SEQ ID NO:119 представляет собой последовательность ДНК из KR1209, которая кодирует белок 14 кДа.
SEQ ID NO:120 представляет собой аминокислотную последовательность белка 14 кДа KR1209.
SEQ ID NO:121 представляет собой последовательность ДНК из KR1369, которая кодирует белок 14 кДа.
SEQ ID NO:122 представляет собой аминокислотную последовательность белка 14 кДа KR1369.
SEQ ID NO:123 представляет собой последовательность ДНК из KR589, которая кодирует белок 14 кДа.
SEQ ID NO:124 представляет собой аминокислотную последовательность белка 14 кДа KR589.
SEQ ID NO:125 представляет собой частичную последовательность ДНК из KR589 для белка 44 кДа.
SEQ ID NO:126 представляет собой частичную аминокислотную последовательность для белка 44 кДа KR589.
SEQ ID NO:127 представляет собой полинуклеотидную последовательность для гена, обозначенного как 149В1-15-РО, который оптимизирован для экспрессии в Zea mays. Этот ген кодирует токсин приблизительно 15 кДа, полученный из PS149B1, который описан в WO 97/40162.
SEQ ID NO:128 представляет собой полинуклеотидную последовательность для гена, обозначенного как 149В1-45-РО, который оптимизирован для экспрессии в Zea mays. Этот ген кодирует токсин приблизительно 45 кДа, полученный из PS149B1, который описан в WO 97/40162.
SEQ ID NO:129 представляет собой полинуклеотидную последовательность для гена, обозначенного как 80JJ1-15-PO7, который оптимизирован для экспрессии в кукурузе. Это альтернативный ген, который кодирует токсин приблизительно 15 кДа.
SEQ ID NO:130 представляет собой аминокислотную последовательность для токсина, кодируемого геном, обозначенным как 80JJ1-15-P07.
SEQ ID NO:131 представляет собой олигонуклеотидный праймер (15kfor1), используемый в соответствии с данным изобретением (см. пример 20).
SEQ ID NO:132 представляет собой олигонуклеотидный праймер (45krev6), используемый в соответствии с данным изобретением (см. пример 20).
SEQ ID NO:133 представляет собой последовательность ДНК из PS201L3, которая кодирует белок 14 кДа.
SEQ ID NO:134 представляет собой аминокислотную последовательность белка 14 кДа PS201L3.
SEQ ID NO:135 представляет собой частичную последовательность ДНК из PS201L3 для белка 44 кДа.
SEQ ID NO:136 представляет собой частичную аминокислотную последовательность для белка 44 кДа PS201L3.
SEQ ID NO:137 представляет собой последовательность ДНК из PS187G1, которая кодирует белок 14 кДа.
SEQ ID NO:138 представляет собой аминокислотную последовательность белка 14 кДа PS187G1.
SEQ ID NO:139 представляет собой последовательность ДНК из PS187G1, которая кодирует белок 44 кДа.
SEQ ID NO:140 представляет собой аминокислотную последовательность белка 44 кДа PS187G1.
SEQ ID NO:141 представляет собой последовательность ДНК из PS201HH2, которая кодирует белок 14 кДа.
SEQ ID NO:142 представляет собой аминокислотную последовательность белка 14 кДа PS201HH2.
SEQ ID NO:143 представляет собой частичную последовательность ДНК из PS201HH2 для белка 44 кДа.
SEQ ID NO:144 представляет собой частичную аминокислотную последовательность для белка 44 кДа PS201HH2.
SEQ ID NO:145 представляет собой последовательность ДНК из KR1369, которая кодирует белок 14 кДа.
SEQ ID NO:146 представляет собой аминокислотную последовательность белка 14 кДа KR1369.
SEQ ID NO:147 представляет собой последовательность ДНК из KR1369, которая кодирует белок 44 кДа.
SEQ ID NO:148 представляет собой аминокислотную последовательность белка 44 кДа KR1369.
SEQ ID NO:149 представляет собой последовательность ДНК из PS137A, которая кодирует белок 14 кДа.
SEQ ID NO:150 представляет собой аминокислотную последовательность белка 14 кДа PS137A.
SEQ ID NO:151 представляет собой последовательность ДНК из PS201V2, которая кодирует белок 14 кДа.
SEQ ID NO:152 представляет собой аминокислотную последовательность белка 14 кДа PS201V2.
SEQ ID NO:153 представляет собой последовательность ДНК из PS207C3, которая кодирует белок 14 кДа.
SEQ ID NO:154 представляет собой аминокислотную последовательность белка 14 кДа PS207C3.
SEQ ID NO:155 представляет собой олигонуклеотидный праймер (F1new) для применения в соответствии с данным изобретением (см. пример 22).
SEQ ID NO:156 представляет собой олигонуклеотидный праймер (R1new) для применения в соответствии с данным изобретением (см. пример 22).
SEQ ID NO:157 представляет собой олигонуклеотидный праймер (F2new) для применения в соответствии с данным изобретением (см. пример 22).
SEQ ID NO:158 представляет собой олигонуклеотидный праймер (R2new) для применения в соответствии с данным изобретением (см. пример 22).
SEQ ID NO:159 представляет собой слитый белок приблизительно 58 кДа.
SEQ ID NO:160 представляет собой слитый ген, кодирующий белок SEQ ID NO:159.
SEQ ID NO:161 представляет собой праймер 45kD5' для применения в соответствии с данным изобретением (см. пример 27).
SEQ ID NO:162 представляет собой праймер 45kD3'rc для применения в соответствии с данным изобретением (см. пример 27).
SEQ ID NO:163 представляет собой праймер 45kD5'01 для применения в соответствии с данным изобретением (см. пример 27).
SEQ ID NO:164 представляет собой праймер 45kD5'02 для применения в соответствии с данным изобретением (см. пример 27).
SEQ ID NO:165 представляет собой праймер 45kD3'03 для применения в соответствии с данным изобретением (см. пример 27).
SEQ ID NO:166 представляет собой праймер 45kD3'04 для применения в соответствии с данным изобретением (пример 27).
Подробное описание изобретения
Данное изобретение относится к двум новым классам полинуклеотидных последовательностей, а также новым пестицидным белкам, кодируемым этими полинуклеотидами. В одном варианте эти белки имеют полноразмерную молекулярную массу приблизительно 40-50 кДа. В конкретных вариантах, приведенных в качестве примеров здесь, эти белки имеют молекулярную массу около 43-47 кДа. Во втором варианте пестицидные белки имеют молекулярную массу приблизительно 10-15 кДа. В конкретных вариантах, приведенных в качестве примеров здесь, эти белки имеют молекулярную массу около 13-14 кДа.
В предпочтительных вариантах белок 40-50 кДа и белок 10-15 кДа используют вместе и эти белки являются пестицидными в комбинации. Таким образом, эти два класса белков данного изобретения могут быть названы "бинарными токсинами". В применении здесь термин "токсин" относится к любому классу пестицидных белков. Данное изобретение относится к полинуклеотидам, которые кодируют токсины 40-50 кДа или 10-15 кДа, полинуклеотидам, которые кодируют части или фрагменты полноразмерных токсинов, которые сохраняют пестицидную активность при использовании в комбинации, и полинуклеотидным последовательностям, которые кодируют оба типа токсинов. В предпочтительном варианте эти токсины являются активными против жесткокрылых вредителей, более предпочтительно против блошки длинноусой и наиболее предпочтительно против Западной блошки длинноусой. Мишенями могут быть также чешуекрылые вредители.
Некоторые конкретные токсины приведены здесь в качестве примеров. Для токсинов, имеющих известную аминокислотную последовательность, известна также молекулярная масса. Специалистам в данной области будет понятно, что средняя молекулярная масса белка, определяемая гель-электрофорезом, будет иногда отличаться от истинной молекулярной массы. Таким образом, ссылку, например, на белок 45 кДа или белок 14 кДа следует понимать как ссылку на белки приблизительно такого размера, даже если истинная молекулярная масса является несколько отличающейся.
Данное изобретение относится не только к полинуклеотидам, которые кодируют эти классы токсинов, но также к применению этих полинуклеотидов для получения рекомбинантных хозяев, экспрессирующих эти токсины. В дополнительном аспекте данное изобретение относится к комбинированному применению токсина приблизительно 40-50 кДа данного изобретения вместе с токсином приблизительно 10-15 кДа данного изобретения для достижения высокоэффективной защиты от вредителей, в том числе жесткокрылых, таких как блошка длинноусая. Например, корни одного растения могут экспрессировать оба типа токсинов.
Таким образом, борьба с вредителями с использованием изолятов, токсинов и генов данного изобретения может выполняться различными способами, известными специалистам в данной области. Эти способы включают, например, нанесение изолятов B.t. на вредителей (или место их локализации), нанесение рекомбинантных микробов на вредителей (или места их локализации) и трансформацию растений генами, которые кодируют пестицидные токсины данного изобретения. Микробами для применения в соответствии с данным изобретением могут быть, например, B.t., E.coli и/или Pseudomonas. Рекомбинантные хозяева могут быть получены специалистами в данной области с использованием стандартных способов. Материалы, необходимые для этих трансформаций, описаны здесь или иным образом, легко доступны квалифицированному работнику. Борьба с насекомыми и другими вредителями, такими как нематоды и клещи, может выполняться также специалистами в данной области с использованием стандартных способов в сочетании с обеспеченными здесь указаниями.
Новые классы токсинов и полинуклеотидных последовательностей, обеспеченные здесь, определяются в соответствии с несколькими параметрами. Одной решающей характеристикой описанных здесь токсинов является пестицидная активность. В конкретном варианте эти токсины обладают активностью против жесткокрылых вредителей. Одним из вариантов являются также токсины, активные против чешуекрылых вредителей. Токсины и гены данного изобретения могут быть определены дополнительно их аминокислотными и нуклеотидными последовательностями. Последовательности молекул в каждом новом классе могут быть идентифицированы и определены их сходством или идентичностью с определенными приведенными в качестве примеров последовательностями, а также способностью гибридизоваться с определенными приведенными в качестве примеров зондами или амплифицироваться с определенными приведенными в качестве примеров праймерами. Классы токсинов, обеспеченные здесь, могут быть также идентифицированы на основе их иммунореактивности с определенными антителами и на основе их соответствия общей формуле.
Специалисту в данной области должно быть понятно, что гены, кодирующие пестицидные белки в соответствии с данным изобретением, могут быть получены с использованием нескольких способов. Конкретные гены, приведенные в качестве примеров здесь, могут быть получены из изолятов, депонированных в депозитариях культур, описанных здесь. Гены и токсины данного изобретения могут быть также сконструированы синтетически, например, с использованием синтезатора генов.
Последовательности трех примеров токсинов 45 кДа обеспечены в последовательностях SEQ ID NO:11, 43 и 38. В предпочтительных вариантах токсины этого класса имеют последовательность, которая согласуется с общей последовательностью, представленной в виде SEQ ID NO:28. В предпочтительных вариантах токсины этого класса будут согласоваться с консенсусной последовательностью, показанной на фигуре 1.
С указаниями, обеспеченными здесь, специалист в данной области мог бы легко получить и использовать различные токсины и полинуклеотидные последовательности новых классов, описанные здесь.
Микроорганизмы, применимые в соответствии с данным изобретением, были депонированы в постоянной коллекции Agricultural Research Service Patent Culture Collection (NRRL), Northern Regional Research Center, 1815 North University Street, Peoria, Illinois 61604, USA. Хранилищные номера культур депонированных штаммов являются следующими:
Изолят PS80JJ1 является общедоступным в соответствии с патентом США 5151363 и другими патентами.
Дополнительный аспект данного изобретения относится к новым изолятам и токсинам и генам, полученным из этих изолятов. Новые изоляты были депонированы и они включены в приведенный выше список. Эти изоляты были депонированы при условиях, гарантирующих, что доступ к культурам будет разрешен во время нахождения заявки на рассмотрении лицу, определенному Уполномоченным по Патентам и Товарным знакам как лицо, имеющее право на это, под 37 CFR 1.14 и 35 U.S.С. 122. Эти депозиты являются доступными согласно требованиям иностранных патентных законов в странах, в которых регистрируются копии данной заявки или вытекающие из нее заявки. Однако должно быть понятно, что доступность депозита не является лицензией на применение на практике данного изобретения в частичном аннулировании патентных прав, выданных заключением государственного патентного ведомства.
Далее, данные депозиты культур будут храниться и станут общедоступными в соответствии с условиями Будапештского Договора в отношении депонирования микроорганизмов, т.е. они будут храниться со всеми условиями, необходимыми для сохранения их жизнеспособными и незагрязненными, в течение периода по меньшей мере пяти лет после самого последнего запроса в отношении предоставления пробы депозита, и в любом случае, в течение периода по меньшей мере 30 (тридцати) лет после даты депонирования или в течение осуществимого периода жизни любого патента, который может дать описание этих культур. Депонент подтверждает его обязанность заменять депозит (депозиты) в том случае, если депозитарий будет неспособен обеспечить пробу при ее запросе вследствие состояния депозита (депозитов). Все ограничения в отношении общедоступности данных культуральных депозитов будут окончательно сняты при выдаче патента, раскрывающего их.
Далее следует таблица 1, которая обеспечивает характеристики определенных изолятов B.t., которые применимы в соответствии с данным изобретением.
Описание штаммов B.t., токсичных для жесткокрылых вредителей
Другие изоляты данного изобретения могут быть также охарактеризованы формой и местоположением включений токсина.
Токсины, гены и зонды. Полинуклеотиды данного изобретения могут быть использованы для образования полных «генов» для кодирования белков или пептидов в желаемой клетке-хозяине. Например, как будет вполне понятно квалифицированному специалисту, некоторые из полинуклеотидов в прилагаемом списке последовательностей показаны без стоп-кодонов. Таким образом, данные полинуклеотиды могут быть подходящим образом помещены под контроль промотора, представляющего интерес хозяина, как известно в данной области.
Как понятно квалифицированному специалисту, ДНК обычно существует в двухцепочечной форме. В этом расположении одна цепь комплементарна другой цепи, и наоборот. Когда ДНК, например, реплицируется в растении, продуцируются дополнительные комплементарные цепи ДНК. Термин "кодирующая цепь" часто используют в данной области для обозначения цепи, которая связывается с антисмысловой цепью. мРНК транскрибируется с "антисмысловой" цепи ДНК. "Смысловая", или "кодирующая", цепь имеет ряд кодонов (кодон представляет собой три нуклеотида, которые могут быть считаны в виде трех за один раз, с образованием конкретной аминокислоты), которые могут считываться в виде открытой рамки считывания (ОРС) с образованием представляющего интерес белка или пептида. Для экспрессии белка in vivo цепь ДНК обычно транскрибируется в комплементарную цепь мРНК, которая используется в качестве матрицы для белка. Таким образом, данное изобретение включает применение приведенных в качестве примеров полинуклеотидов, показанных в прилагаемом списке последовательностей, и/или комплементарных цепей. РНК и ПНК (пептид-нуклеиновые кислоты), которые функционально эквивалентны приведенным в качестве примеров ДНК, включены в данное изобретение.
Токсины и гены данного изобретения могут быть идентифицированы и получены с использованием, например, олигонуклеотидных зондов. Эти зонды являются детектируемыми нуклеотидными последовательностями, которые могут быть детектируемыми благодаря подходящей метке или могут быть изготовлены в виде флуоресцентных зондов, как описано в Международной заявке с номером WO 93/16094. Зонды (и полинуклеотиды данного изобретения) могут быть ДНК-, РНК- или ПНК-зондами (и полинуклеотидами). Кроме аденина (А), цитозина (С), гуанина (G), тимина (Т) и урацила (U; для РНК-молекул), синтетические зонды (и полинуклеотиды) данного изобретения могут также иметь инозин (нейтральное основание, способное образовывать пары со всеми четырьмя основаниями; иногда используемое вместо смеси всех четырех оснований в синтетических зондах). Таким образом, при ссылке на синтетический, вырожденный олигонуклеотид и использовании "n" в общем смысле, "n" может быть G, А, Т, С или инозином. Двусмысленные коды, используемые здесь, находятся в соответствии со стандартными номенклатурными конвенциями IUPAC, касающимися подачи данной заявки (например, R обозначает А или G, Y обозначает С или Т, и т.д.).
Как хорошо известно в данной области, если молекула-зонд и проба нуклеиновой кислоты гибридизуются с образованием сильной связи между этими двумя молекулами, то резонно предположить, что эти зонд и проба имеют существенные гомологию/сходство/идентичность. Предпочтительно гибридизацию проводят при жестких условиях способами, хорошо известными в данной области, как описано, например, в Keller, G.H., М.М.Manak (1978) DNA Probes, Stockton Press, New York, NY, pp.169-170. Например, как указано в этой работе, условия высокой степени жесткости могут быть достигнуты промыванием сначала 2х SSC (Стандартным Солевым Цитратом)/0,1% ДСН (додецилсульфатом натрия) в течение 15 мин при комнатной температуре. Обычно выполняют две промывки. Более высокая жесткость может быть достигнута снижением концентрации соли и/или повышением температуры. Например, за описанной выше промывкой могут следовать две промывки 0,1x SSC/0,1% ДСН в течение 15 мин, каждый раз при комнатной температуре, с последующими промывками 0,1x SSC/0,1% ДСН в течение 30 мин каждый раз при 55°С. Эти температуры могут быть использованы с другими протоколами гибридизации и промывок, приведенными здесь, как должно быть известно специалисту в данной области (SSPE может быть использован в качестве соли вместо SSC, например). Смесь 2х SSC/0,1% ДСН может быть получена добавлением 50 мл 20х SSC и 5 мл 10% ДСН к 445 мл воды. 20х SSC может быть получен объединением NaCl (175,3 г/0,150 М), цитрата натрия (88,2 г/0,015 М) и воды до 1 л, с последующим доведением рН до 7,0 10 н. NaOH. 10% ДСН может быть получен растворением 10 г ДСН в 50 мл автоклавированной воды, разбавлением до 100 мл и получением аликвот.
Детектирование зонда обеспечивает средство для определения известным образом, произошла ли гибридизация. Такой анализ с использованием зондов обеспечивает быстрый способ для идентификации кодирующих токсин генов данного изобретения. Нуклеотидные сегменты, которые используют в качестве зондов в соответствии с данным изобретением, могут быть синтезированы с использованием ДНК-синтезатора и стандартных процедур. Эти нуклеотидные последовательности могут быть также использованы в качестве ПЦР-праймеров для амплификации генов данного изобретения.
Характеристики гибридизации молекулы могут быть использованы для определения полинуклеотидов данного изобретения. Таким образом, данное изобретение включает полинуклеотиды (и/или их комплементы, предпочтительно их полные комплементы), которые гибридизуются с приведенным здесь примером полинуклеотида (таким как последовательности ДНК, включенные в последовательности SEQ ID NO:46-166).
В применении здесь термин "жесткие" условия гибридизации относится к условиям, которые дают такую же или примерно такую же степень специфичности гибридизации, что и условия, использованные в данной заявке. Конкретно, гибридизацию иммобилизованной ДНК на Саузерн-блотах с 32Р-мечеными генспецифическими зондами выполняли стандартными способами (Maniatis, Т., E.F.Fritsch, J.Sambrook [1982] Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY). В общем виде, гибридизацию и последующие промывки проводили при жестких условиях, которые позволяли детектирование целевых последовательностей (с гомологией генам токсина PS80JJ1, например). Для двухцепочечных генных ДНК-зондов гибридизацию проводили в течение ночи при температуре на 20-25°С ниже температуры плавления (Tm) ДНК-гибрида в 6Х SSPE, 5X растворе Денхардта, 0,1% ДСН, 0,1 мг/мл денатурированной ДНК. Температура плавления описывается следующей формулой (Beltz, G.A., К.A.Jacobs, Т.Н.Eickbush, P.Т.Cherbas and F.C.Kafatos [1983] Methods of Enzymology, R. Wu, L.Grossman and K.Moldave [eds.] Academic Press, New York 100:266-285):
Tm=81,5°C+16,6Log[Na+]+0,41(%G+C)-0,61(% формамид)-600/ длина дуплекса в п.н.
Промывки обычно проводят следующим образом:
(1) Дважды при комнатной температуре в течение 15 мин в IX SSPE, 0,1% ДСН (промывка с низкой жесткостью).
(2) Один раз при Tm-20°С в течение 15 мин в 0,2Х SSPE, 0,1% ДСН (промывка с умеренной жесткостью).
Для олигонуклеотидных зондов гибридизацию проводили в течение ночи при 10-20°С ниже температуры плавления (Tm) гибрида в 6Х SSPE, 5X растворе Денхардта, 0,1% ДСН, 0,1 мг/мл денатурированной ДНК. Tm для олигонуклеотидных зондов определяли при помощи следующей формулы:
Tm (°С)=2 (число Т/А п.н.)+4 (число G/C п.н.)
(Suggs S.V., Т.Miyake, E.H.Kawashime, M.J.Johnson, К.Itakura, and R.B.Wallace [1981] ICN-UCLA Symp.Dev.Biol. Using Purified Genes, D.D.Brown [ed.], Academic Press, New York, 23:683-693).
Промывки обычно проводили следующим образом:
(1) Дважды при комнатной температуре в течение 15 мин в 1X SSPE, 0,1% ДСН (промывка с низкой жесткостью).
(2) Один раз при температуре гибридизации в течение 15 мин в 1X SSPE, 0,1% ДСН (промывка с умеренной жесткостью).
Токсины, получаемые из изолятов PS149B1, PS167H2 и PS80JJ1, были охарактеризованы как имеющие по меньшей мере одну из следующих характеристик (новые токсины данного изобретения могут быть подобным образом охарактеризованы этой и иной идентифицирующей информацией, приведенной здесь):
(a) указанный токсин кодируется нуклеотидной последовательностью, которая гибридизуется при жестких условиях с нуклеотидной последовательностью, выбранной из группы, состоящей из: ДНК, которая кодирует SEQ ID NO:2, ДНК, которая кодирует SEQ ID NO:4, ДНК, которая кодирует SEQ ID NO:6, SEQ ID NO:8, SEQ ID NO:10, ДНК, которая кодирует SEQ ID NO:11, SEQ ID NO:12, ДНК, которая кодирует SEQ ID NO:13, SEQ ID NO:14, ДНК, которая кодирует SEQ ID NO:15, ДНК, которая кодирует SEQ ID NO:16, ДНК, которая кодирует SEQ ID NO:17, ДНК, которая кодирует SEQ ID NO:18, ДНК, которая кодирует SEQ ID NO:19, SEQ ID NO:20, SEQ ID NO:21, SEQ ID NO:22, SEQ ID NO:23, SEQ ID NO:24, SEQ ID NO:25, SEQ ID NO:26, SEQ ID NO:27, ДНК, которая кодирует пестицидную часть SEQ ID NO:28, SEQ ID NO:37, ДНК, которая кодирует SEQ ID NO:38, SEQ ID NO:42 и ДНК, которая кодирует SEQ ID NO:43;
(b) указанный токсин иммунореагирует с антителом к пестицидному токсину приблизительно 40-50 кДа, или его фрагменту, из изолята Bacillus thuringiensis, выбранного из группы, состоящей из PS80JJ1, имеющего идентифицирующие характеристики NRRL В-18679, PS149B1, имеющего идентифицирующие характеристики NRRL B-21553 и PS167H2, имеющего идентифицирующие характеристики NRRL B-21554;
(c) указанный токсин кодируется нуклеотидной последовательностью, где часть указанной нуклеотидной последовательности может быть амплифицирована при помощи ПЦР с использованием пары праймеров, выбранных из группы, состоящей из SEQ ID NO:20 и 24, с получением фрагмента приблизительно 495 п.н., SEQ ID NO:20 и 25 с получением фрагмента приблизительно 594 п.н., SEQ ID NO:21 и 24 с получением фрагмента приблизительно 471 п.н., и SEQ ID NO:21 и 25 с получением фрагмента приблизительно 580 п.н.;
(d) указанный токсин содержит пестицидную часть аминокислотной последовательности, показанной в SEQ ID NO:28;
(e) указанный токсин содержит аминокислотную последовательность, которая имеет по меньшей мере приблизительно 60%-ную гомологию с пестицидной частью аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO:11, SEQ ID NO:13, SEQ ID NO:15, SEQ ID NO:38 и SEQ ID NO:43;
(f) указанный токсин кодируется нуклеотидной последовательностью, которая гибридизуется при жестких условиях с нуклеотидной последовательностью, выбранной из группы, состоящей из ДНК, которая кодирует SEQ ID NO:3, ДНК, которая кодирует SEQ ID NO:5, ДНК, которая кодирует SEQ ID NO:7, ДНК, которая кодирует SEQ ID NO:32, ДНК, которая кодирует SEQ ID NO:36, и ДНК, которая кодирует SEQ ID NO:41;
(g) указанный токсин иммунореагирует с антителом к пестицидному токсину приблизительно 10-15 кДа, или его фрагменту, из изолята Bacillus thuringiensis, выбранного из группы, состоящей из PS80JJ1, имеющего идентифицирующие характеристики NRRL В-18679, PS149B1, имеющего идентифицирующие характеристики NRRL B-21553 и PS167H2, имеющего идентифицирующие характеристики NRRL B-21554;
(h) указанный токсин кодируется нуклеотидной последовательностью, в которой часть указанной нуклеотидной последовательности может быть амплифицирована при помощи ПЦР с использованием пары праймеров SEQ ID 29 и SEQ ID NO:33; и
(i) указанный токсин содержит аминокислотную последовательность, которая имеет по меньшей мере приблизительно 60%-ную гомологию с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID NO:3, SEQ ID NO:5, SEQ ID NO:7, пестицидных частей SEQ ID NO:32, пестицидных частей SEQ ID NO:36 и пестицидных частей SEQ ID NO:41.
Модификация генов и токсинов. Гены и токсины, применимые в соответствии с данным изобретением, включают не только конкретно приведенные в качестве примеров полноразмерные последовательности, но также части и/или фрагменты (в том числе внутренние и/или концевые делеции, в сравнении с полноразмерными молекулами) этих последовательностей, их варианты, мутанты, химеры и слияния (гибриды). Белки данного изобретения могут иметь замененные аминокислоты до тех пор, пока они сохраняют характерную пестицидную активность белков, конкретно представленных здесь. "Вариантные" гены имеют нуклеотидные последовательности, которые кодируют те же самые токсины или которые кодируют токсины, имеющие пестицидную активность, эквивалентную приведенному в качестве примера белку. В применении здесь термин "эквивалентные токсины" относится к токсинам, имеющим ту же самую или по существу ту же самую биологическую активность против вредителей-мишеней, что и приведенные в качестве примеров токсины. В применении здесь термин "по существу та же самая" последовательность относится к последовательностям, которые имеют аминокислотные замены, делеции, присоединения или инсерции, которые фактически не влияют на пестицидную активность. Фрагменты, сохраняющие пестицидную активность, также включены в это определение. Фрагменты и эквиваленты, сохраняющие пестицидную активность приведенных в качестве примеров токсинов, будут также находиться в объеме данного изобретения.
Эквивалентные токсины и/или гены, кодирующие эквивалентные токсины, могут быть получены из изолятов B.t. дикого типа или изолятов рекомбинантных B.t. и/или из других организмов дикого типа или рекомбинантных организмов с использованием обеспеченных здесь указаний. Другие виды Bacillus, например, могут быть использованы в качестве источников-изолятов.
Вариации генов могут быть легко сконструированы с использованием стандартных способов, например получения точковых мутаций. В патенте США №5605793, например, также описаны способы генерирования дополнительного молекулярного разнообразия с использованием повторной укладки ДНК после случайной фрагментации. Вариантные гены могут быть использованы для получения вариантных белков; рекомбинантные хозяева могут быть использованы для получения вариантных белков. Фрагменты полноразмерных генов могут быть получены с использованием коммерчески доступных экзонуклеаз или эндонуклеаз в соответствии со стандартными процедурами. Например, ферменты, такие как Ваl31, или сайт-направленный мутагенез могут быть использованы для систематического отрезания нуклеотидов от концов этих генов. Гены, кодирующие активные фрагменты, могут быть также получены с использованием разнообразных ре-стрикционных ферментов (рестриктаз). Протеазы могут быть использованы для прямого получения активных фрагментов этих токсинов.
Имеется ряд способов для получения пестицидных токсинов данного изобретения. Например, антитела к пестицидным токсинам, описанным и заявленным здесь, могут быть использованы для идентификации и выделения других токсинов из смеси белков. Конкретно, могут быть продуцированы антитела к частям токсинов, которые являются наиболее константными и наиболее отличающимися от других токсинов B.t. Эти антитела могут быть затем использованы для специфической идентификации эквивалентных токсинов с характерной активностью посредством иммунопреципитации, твердофазного иммуноферментного анализа (ELISA) или Вестерн-блоттинга. Антитела к описанным здесь токсинам, или к эквивалентным токсинам, или к фрагментам этих токсинов могут быть легко получены с использованием стандартных процедур. Гены, кодирующие эти токсины, могут быть затем получены из микроорганизма-источника.
Вследствие избыточности генетического кода множество различных ДНК-последовательностей могут кодировать описанные здесь аминокислотные последовательности. Вполне в рамках квалификации лица, специализирующего в данной области, находится создание этих альтернативных ДНК-последовательностей, кодирующих те же самые или по существу те же самые токсины. Эти вариантные ДНК-последовательности находятся в рамках данного изобретения.
Определенные токсины данного изобретения приводятся здесь конкретно в качестве примеров. Поскольку эти токсины являются лишь примерами токсинов данного изобретения, должно быть вполне очевидным, что данное изобретение включает вариантные или эквивалентные токсины (и нуклеотидные последовательности, кодирующие эквивалентные токсины), имеющие такую же или сходную пестицидную активность в сравнении с активностью приведенного в качестве примера токсина. Эквивалентные токсины будут обладать сходством аминокислотных последовательностей (и/или гомологией) с приведенным в качестве примера токсином. Аминокислотная идентичность будет обычно более 60%, предпочтительно более 75%, более предпочтительно более 80%, даже более предпочтительно более 90% и может быть более 95%. Предпочтительные полинуклеотиды и белки данного изобретения могут также определяться более конкретными параметрами идентичности и/или сходства. Например, идентичность и/или сходство может быть 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99% в сравнении с приведенной здесь в качестве примера последовательностью. Если нет других указаний, в применении здесь процентную идентичность и/или процентное сходство двух нуклеиновых кислот определяют с использованием алгоритма Karlin and Altschul (1990), Proc. Natl. Acad. Sci. USA 87:2264-2268, модифицированного, как описано в Karlin and Altschul (1993), Proc. Natl. Acad. Sci. USA 90:5873-5877. Такой алгоритм включен в программы NBLAST и XBLAST Altschul et al. (1990), J. Mol. Biol. 215:402-410. Нуклеотидный поиск BLAST выполняют с программой NBLAST, оценка =100, длина слова =12, для получения нуклеотидных последовательностей с желаемой процентной идентичностью последовательности. Для получения имеющих гэпы сопоставлений для сравнительных целей используют Gapped BLAST, как описано Altschul et al. (1997), Nucl. Acids Res. 25:3389-3402. При использовании программ BLAST и Gapped BLAST используют параметры по умолчанию соответствующих программ (NBLAST и XBLAST). См. http://www.ncbi.nih.gov. Оценки могут быть также рассчитаны с использованием способов и алгоритмов Crickmore et al., как описано в разделе Предпосылки выше.
Аминокислотная гомология может быть наивысшей в критических районах токсина, которые ответственны за биологическую активность или участвуют в определении трехмерной конфигурации, которая в конечном счете является ответственной за биологическую активность. В этом отношении определенные аминокислотные замены являются приемлемыми и могут ожидаться, если эти замены находятся в районах, которые не являются критическими для активности или являются консервативными заменами аминокислот, которые не влияют на трехмерную конфигурацию молекулы. Например, аминокислоты могут быть помещены в следующие классы: неполярные, незаряженные полярные, основные и кислотные. Консервативные замены, посредством которых аминокислота одного класса заменяется другой кислотой того же самого типа, попадают в рамки данного изобретения, пока эта замена не изменяет существенно биологическую активность соединения. Таблица 2 обеспечивает список примеров аминокислот, принадлежащих к каждому классу.
В некоторых случаях могут быть произведены неконсервативные замены. Критическим фактором является то, что эти замены не должны уменьшать биологическую активность токсина.
В применении здесь термины «выделенные» полинуклеотиды и/или "очищенные" токсины относятся к молекулам, которые не связаны с другими молекулами, с которыми они были бы обнаружены в природе; эти термины включали бы их использование в растениях. Таким образом, ссылка на "выделенные" и/или "очищенные" означает участие "руки человека", как описано здесь.
Синтетические гены, которые функционально эквивалентны токсинам данного изобретения, могут быть также использованы для трансформации хозяев. Способы получения синтетических генов могут быть найдены, например, в патенте США №5380831.
Трансгенные хозяева. Кодирующие токсин гены данного изобретения могут быть введены в большое разнообразие микробных или растительных хозяев. В предпочтительных вариантах экспрессия гена токсина, прямо или опосредованно, приводит к внутриклеточному продуцированию и поддержанию пестицидных белков. При проглатывании трансгенных/рекомбинантных/транс-формированных клеток-хозяев вредителями вредители будут проглатывать токсин. Это предпочтительный способ, в котором обеспечивается контакт вредителя с токсином. Результатом является защита от вредителя (убивание или заболевание вредителя). Альтернативно, подходящие микробные хозяева, например Pseudomonas, такой как Р.fluorescens, могут наноситься на место локализации вредителя, где некоторые из них могут размножаться, и проглатываться вредителями-мишенями. Микро6, несущий ген токсина, может быть обработан в условиях, пролонгирующих активность токсина и стабилизирующих клетку. Затем обработанная клетка, сохраняющая токсическую активность, может быть внесена в окружение вредителя-мишени.
В предпочтительных вариантах используют рекомбинантные клетки растения и растения. Предпочтительными растениями (и клетками растения) являются кукуруза и/или маис.
При введении гена токсина B.t. в подходящем векторе в микробного хозяина и внесении указанного хозяина в окружающую среду в живом состоянии должны использоваться определенные микробы-хозяева. Выбирают микроорганизмы-хозяева, о которых известно, что они заселяют «фитосферу» (поверхность листьев, филлосферу, ризосферу и/или поверхность корней) одной или нескольких представляющих интерес культур. Эти микроорганизмы выбирают таким образом, чтобы они были способны успешно конкурировать в конкретной окружающей среде (посеве или других местах обитания) с микроорганизмами дикого типа, обеспечивать стабильное поддержание и экспрессию гена, экспрессирующего полипептидный пестицид, и, желательно, обеспечивать улучшенную защиту пестицида от разрушения и инактивации условиями окружающей среды.
Известно, что большое число микроорганизмов заселяют филлоповерхность (поверхность листьев растений) и/или ризосферу (почву, окружающую корни растений) большого круга важных культур. Эти микроорганизмы включают бактерии, водоросли и грибы. Особый интерес представляют микроорганизмы, такие как бактерии, например роды Pseudomonas, Erwinia, Serratia, Klebsiella, Xanthomonas, Streptomyces, Rhizobium, Rhodopseudomonas, Methylophilius, Agrobacterium, Acetobacter, Lactobacillus, Arthrobacter, Azotobacter, Leuconostoc и Alcaligenes; грибки, в частности дрожжи, например роды Saccharomyces, Cryptococcus, Kluyveromyces, Sporobolomyces, Rhodotorula и Aureobasidium. Особый интерес представляют такие бактериальные виды фитосферы, как Pseudomonas syringae, Pseudomonas fluorescens, Serratia marcescens, Acetobacter xylinium, Agrobacterium tumefaciens, Rhodopseudomonas spheroides, Xanthomonas campestris, Rhizobium melioti, Alcaligenes entrophus и Azotobacter vinlandii; и виды дрожжей фитосферы, такие как Rhodotorula rubra, R.glutinis, R.marina, R.aurantiaca, Cryptococcus albidus, C.diffluens, C.laurentii, Saccharomyces rosei, S.pretoriensis, S.cerevisiae, Sporobolomyces roseus, S.odorus, Kluyveromyces veronae и Aureobasidium pollulans. Особый интерес представляют пигментированные микроорганизмы.
Большое разнообразие способов доступно для введения гена B.t., кодирующего токсин, в хозяина-мишень в условиях, которые делают возможными стабильное поддержание и экспрессию гена. Эти способы хорошо известны специалистам в данной области и описаны, например, в патенте США 5135867, который включен здесь в качестве ссылки.
Обработка клеток. Как упоминалось выше, B.t. или рекомбинантные клетки, экспрессирующие токсин B.t., могут быть обработаны для пролонгирования активности токсина и стабилизации клетки. Пестицидная микрокапсула, которая образуется, содержит токсин B.t. в клеточной структуре, которая была стабилизирована, и будет защищать этот токсин при нанесении микрокапсулы в среду вредителя-мишени. Подходящие клетки-хозяева могут включать прокариоты или эукариоты, причем они обычно ограничиваются клетками, которые не продуцируют вещества, токсичные для высших организмов, таких как млекопитающие. Однако организмы, которые продуцируют вещества, токсичные для высших организмов, могли бы быть использованы, если эти токсические вещества являются нестабильными или уровень применения является достаточно низким, чтобы избежать какой-либо возможности токсичности для хозяина-млекопитающего. В качестве хозяев особый интерес представляют прокариоты и низшие эукариоты, такие как грибы.
Клетка обычно должна быть интактной и по существу находиться в пролиферативной форме при обработке, а не в форме споры, хотя в некоторых случаях могут быть использованы споры.
Обработка микробной клетки, например микроба, содержащего ген токсина B.t., может быть проведена химическими или физическими способами или комбинацией химических и/или физических способов, пока способ не оказывает неблагоприятного влияния на свойства токсина и не уменьшает клеточной способности защиты токсина. Примерами химических реагентов являются галогенирующие агенты, в частности галогены с атомными номерами 17-80. Более конкретно, иод может быть использован при мягких условиях и в течение достаточного времени для получения желательных результатов. Другие подходящие способы включают обработку альдегидами, такими как глутаровый альдегид; антиинфекционными средствами, такими как зефиранхлорид и цетилпиридинийхлорид; спиртами, такими как изопропиловый спирт и этанол; различными гистологическими фиксаторами, такими как Люголь-иод, фиксатор Bouin, различные кислоты и фиксатор Helly (См.: Humason, Gretchen L., Animal Tissue Techniques, W.H. Freeman and Company, 1967); или комбинацию физических (нагревание) и химических агентов, которые сохраняют и пролонгируют активность токсина, продуцируемого в клетке, при введении в окружение хозяина. Примерами физических средств являются излучение коротких длин волн, таких как гамма-излучение и рентгеновское излучение, замораживание, УФ-излучение, лиофилизация и т.п. Способы обработки микробных клеток описаны в патентах США 4695455 и 4695462, которые включены здесь в качестве ссылки.
Клетки обычно будут иметь повышенную структурную стабильность, которая будет усиливать устойчивость к условиям окружающей среды. Если пестицид является проформой, способ обработки клеток должен быть выбран таким образом, чтобы он не ингибировал процессинг проформы в зрелую форму пестицида патогенным вредителем-мишенью. Например, формальдегид будет сшивать белки и мог бы ингибировать процессинг проформы полипептидного пестицида. Способ обработки должен сохранять по меньшей мере существенную часть биодоступности или биоактивности токсина.
Характеристики, представляющие особенный интерес в выборе клетки-хозяина для целей продуцирования, включают легкость введения гена B.t. в данного хозяина, доступность экспрессионных систем, эффективность экспрессии, стабильность пестицида в данном хозяине и присутствие вспомогательных генетических способностей. Представляющие интерес характеристики для применения в виде пестицидной микрокапсулы включают защитные качества в отношении пестицида, такие как толстые клеточные стенки, пигментация и внутриклеточная упаковка или образование телец включения; выживание в водных средах; отсутствие токсичности для млекопитающих; привлекательность для проглатывания для насекомых; легкость убивания и фиксации без повреждения токсина и т.п. Другие рассмотрения включают легкость приготовления и манипулирования, дешевизну, стабильность при хранении и т.п.
Выращивание клеток. Клеточный хозяин, содержащий инсектицидный ген B.t., может выращиваться в любой подходящей питательной среде, предпочтительно, где ДНК-конструкция обеспечивает селективное преимущество, обеспечивая селективную среду, так что по существу все или все клетки сохраняют ген B.t. Эти клетки могут быть затем собраны согласно общепринятым способам. Альтернативно, клетки могут быть обработаны перед их сбором.
Клетки B.t. данного изобретения могут культивироваться с использованием стандартных в данной области сред и способов ферментации. По завершении ферментационного цикла бактерии могут быть собраны сначала отделением спор и кристаллов B.t. из ферментационного бульона способами, хорошо известными в данной области. Извлеченные споры и кристаллы B.t. могут быть приготовлены в виде смачивающегося порошка, жидкого концентрата, гранул или других препаратов добавлением поверхностно-активных веществ, диспергирующих агентов, инертных носителей и других компонентов для облегчения манипулирования и применения для конкретных вредителей-мишеней. Эти композиции и процедуры нанесения хорошо известны в данной области.
Композиции. Приготовленные гранулы-приманки, содержащие аттрактант и споры и кристаллы изолятов B.t., или рекомбинантные микробы, содержащие гены, полученные из описанных здесь изолятов B.t., могут вноситься в почву. Приготовленный продукт может также наноситься в виде покрытия семян или для обработки корней или обработки целых растений на последних стадиях цикла культуры. Обработки растений и почвы клетками B.t. могут проводиться в виде смачивающихся порошков, гранул или дустов смешиванием различных инертных материалов, таких как неорганические минералы (филлосиликаты, карбонаты, сульфаты, фосфаты и т.п.) или ботанические материалы (порошкообразные стержни кукурузных початков, скорлупа риса, скорлупа лесного ореха и т.п.). Препараты могут включать распределяющие-приклеивающие адъюванты, стабилизирующие агенты, другие пестицидные добавки или поверхностно-активные вещества. Жидкие композиции могут быть на водной основе или безводными и использоваться в виде пен, гелей, суспензий, эмульгируемых концентратов или т.п. Ингредиенты могут включать реологические агенты, поверхностно-активные вещества, эмульгаторы, диспергирующие агенты или полимеры.
Как должно быть понятно специалисту в данной области, концентрация пестицида будет очень широко варьировать в зависимости от природы конкретной композиции, в частности от того, является ли она концентратом или должна использоваться непосредственно. Пестицид будет присутствовать в количестве по меньшей мере 1 мас.% и может составлять 100 мас.%. Сухие композиции будут иметь от приблизительно 1 до 95 мас.% пестицида, тогда как жидкие композиции будут обычно содержать приблизительно 1-60 мас.% твердых веществ в жидкой фазе. Композиции будут обычно содержать от приблизительно 102 до 104 клеток/мг. Эти композиции будут вводиться при приблизительно от 50 мг (жидкости или сухого ингредиента) до 1 кг или более на гектар.
Данные композиции могут вноситься в окружающую среду вредителя, например почву или листву, разбрызгиванием, распылением, дождеванием или т.п.
Мутанты. Мутанты изолятов данного изобретения могут быть получены согласно процедурам, хорошо известным в данной области. Например, аспорогенный мутант может быть получен посредством мутагенеза изолята с использованием этилметансульфоната (ЕМС). Мутанты могут быть получены с использованием ультрафиолетового света и нитрозогуанидина процедурами, хорошо известными в данной области.
Меньший процент аспорогенных мутантов будет оставаться интактным и не лизироваться в течение продолжительных периодов ферментации; эти штаммы названы лизис-минус (-). Лизис-минус-штаммы могут быть идентифицированы скринингом аспорогенных мутантов в среде во встряхиваемых колбах и отбором тех мутантов, которые все еще остаются интактными и содержат кристаллы токсина в конце ферментации. Лизис-минус-штаммы пригодны для способа обработки клеток, который дает защищенный инкапсулированный белок токсина.
Для получения фагоустойчивого варианта указанного аспорогенного мутанта аликвоту фагового лизата распределяют на питательный агар и дают высохнуть. Затем аликвоту фагочувствительного бактериального штамма высевают непосредственно на высушенный лизат и дают высохнуть. Чашки инкубируют при 30°С. Чашки инкубируют в течение 2 дней и в это время можно видеть многочисленные растущие на агаре колонии. Некоторые из этих колоний извлекают и субкультивируют на чашки с питательным агаром. Эти видимые устойчивые культуры испытывают на устойчивость перекрестным посевом штрихом с фаговым лизатом. Линию фагового лизата наносят штрихом на чашку и дают ей высохнуть. Затем предположительно устойчивые культуры "штрихуют" поперек линии фага. Устойчивые бактериальные культуры не обнаруживают лизиса нигде в штрихе, проходящем через линию фага после инкубации в течение ночи при 30°С. Устойчивость к фагу повторно подтверждают затем засевом газона устойчивой культуры на чашку с питательным агаром. Чувствительный штамм также высевают таким же образом в качестве положительного контроля. После высушивания каплю фагового лизата помещают в центр чашки и дают высохнуть. Устойчивые культуры не обнаруживают лизиса в зонах, где был помещен фаговый лизат после инкубирования при 30°С в течение 24 часов.
Далее следуют примеры, которые иллюстрируют процедуры для применения на практике данного изобретения. Эти примеры не должны пониматься как ограничивающие. Все проценты являются мас.% и все соотношения смесей растворителей являются объемными, если нет других указаний.
ПРИМЕР 1
Культивирование изолятов B.t. данного изобретения
Субкультура изолятов B.t. или их мутантов может быть использована для инокуляции следующей среды, среды, содержащей пептон, глюкозу, соли.
Солевой раствор (100 мл)
CaCl2-раствор (100 мл)
Солевой раствор и CaCl2-раствор стерилизуют фильтрацией и добавляют к автоклавированному и сваренному бульону во время инокуляции. Колбы инкубируют при 30°С на ротационном шейкере при 200 об/мин в течение 64 час.
Приведенную выше процедуру можно легко масштабировать до больших ферментеров процедурами, известными в данной области.
Споры и/или кристаллы B.t., полученные в описанной выше ферментации, могут быть выделены процедурами, хорошо известными в данной области. Часто используемой процедурой является применение к собранному ферментационному бульону способов разделения, например центрифугирования.
ПРИМЕР 2
Активность спорулированных культур Bacillus thurinqiensis на блошке длинноусой
Жидкие культуры PS80JJ1, PS149B1 или PS167H2 выращивали до споруляции во встряхиваемых колбах и осаждали центрифугированием. Культуральные осадки ресуспендировали в воде и анализировали на активность против блошки длинноусой в биотестах с верхним нанесением, как описано выше. Количества белков 14 кДа и 44,3 кДа, присутствующих в культуральных осадках, определяли денситометрией и использовали для расчета удельной активности, выражаемой в виде LC50. Активность каждого нативного штамма В. thuringiensis представлена в таблице 3 (биотест WCRW с верхним нанесением штаммов B.t.).
Биотест WCRW с верхним нанесением штаммов B.t.
ПРИМЕР 3
Очистка белка для белка 80JJ1 45 кДа
Один грамм лиофилизированного порошка 80JJ1 суспендировали в 40 мл буфера, содержащего 80 мМ Трис-Cl рН 7,8, 5 мМ ЭДТА, 100 мкМ ПМСФ, 0,5 мкг/мл лейпептина, 0,7 мкг/мл пепстатина и 40 мкг/мл бестатина. Суспензию центрифугировали и полученный супернатант отбрасывали. Осадок промывали пять раз с использованием 35-40 мл вышеуказанного буфера для каждой промывки. Промытый осадок ресуспендировали в 10 мл 40% NaBr, 5 мМ ЭДТА, 100 мкМ ПМСФ, 0,5 мкг/мл лейпептина, 0,7 мкг/мл пепстатина и 40 мкг/мл бестатина и помещали на качающуюся платформу на 75 мин. NaBr-суспензию центрифугировали, супернатант удаляли и осадок обрабатывали второй раз 40% NaBr, 5 мМ ЭДТА, 100 мкМ ПМСФ, 0,5 мкг/мл лейпептина, 0,7 мкг/мл пепстатина и 40 мкг/мл бестатина, как описано выше. Супернатанты (40% NaBr-растворимые) объединяли и диализовали против 10 мМ CAPS pH 10,0, 1 мМ ЭДТА при 4°С. Диализованные экстракты центрифугировали и полученный супернатант удаляли. Осадок (осадок 40% NaBr-диализа) суспендировали в 5 мл H2O и центрифугировали. Полученный супернатант удаляли и отбрасывали. Промытый осадок промывали второй раз в 10 мл Н2O и центрифугировали, как описано выше. Этот промытый осадок суспендировали в 1,5 мл H2O, и он содержал первично три белковые полосы с видимыми подвижностями приблизительно 47 кДа, 45 кДа и 15 кДа при анализе электрофорезом в ДСН-ПААГ. На этой стадии очистки суспендированный 40% NaBr-диализный осадок содержал приблизительно 21 мг/мл белка согласно анализу по Лоури.
Белки в суспензии осадка разделяли с выше. Этот промытый осадок суспендировали в 1,5 использованием электрофореза в ДСН-ПААГ (Laemlli, U.K. [1970] Nature 227:680) в 15% акриламидных гелях. Затем разделенные белки электрофоретически блоттировали на мембрану PVDF (Millipore Corp.) в 10 мМ CAPS pH 11,0, 10% МеОН при 100 В постоянного тока. Спустя один час мембрану PVDF промывали в воде в течение короткого времени и помещали на 3 мин в 0,25% Кумасси синий R-250, 50% метанол, 5% уксусную кислоту. Окрашенную мембрану обесцвечивали в 40% МеОН, 5% уксусной кислоте. Обесцвеченную мембрану сушили на воздухе при комнатной температуре (LeGendre et al., [1989] In A Practical Guide to Protein Purification For Microsequencing, P.Matsudaira, ed., Academic Press, New York, NY). Мембрану секвенировали с использованием автоматизированной деградации Эдмана в газовой фазе (Hunkapillar, M.W., R.M.Hewick, W.L.Dreyer, L.E.Hood [1983] Meth. Enzymol. 91:399).
Аминокислотный анализ выявил, что N-концевая последовательность полосы 45 кДа была следующей: Met-Leu-Asp-Thr-Asn (SEQ ID NO:1).
Полосу 47 кДа также анализировали, и было определено, что N-концевая аминокислотная последовательность является той же самой аминокислотной последовательностью из 5 аминокислот, которая показана в SEQ ID NO:1. Таким образом, N-аминокислотные последовательности пептида 47 кДа и пептида 45 кДа были идентичными.
Эта аминокислотная последовательность соответствует последовательности, полученной из пептида 45 кДа, полученного из порошков спор/кристаллов PS80JJ1, с использованием другого протокола очистки, со следующей N-концевой последовательностью: Met-Leu-Asp-Thr-Asn-Lys-Val-Tyr-Glu-Ile-Ser-Asn-Leu-Ala-Asn-Gly-Leu-Tyr-Thr-Ser-Thr-Tyr-Leu-Ser-Leu (SEQ ID NO:2).
ПРИМЕР 4
Очистка пептида 14 кДа PS80JJ1
0,8 мл белой диализной суспензии (приблизительно 21 мг/мл), содержащей пептиды 47 кДа, 45 кДа и 15 кДа, растворяли в 10 мл 40% NaBr и добавляли 0,5 мл 100 мМ ЭДТА. Спустя приблизительно 18 часов (ночь) получали белую непрозрачную суспензию. Ее собирали центрифугированием и отбрасывали. Супернатант концентрировали в Центриконе-30 (Amicon Corporation) до конечного объема приблизительно 15 мл. Отфильтрованный объем промывали водой посредством фильтр-диализа и инкубировали на льду, что в конечном счете давало молочно-белую суспензию. Суспензию центрифугировали, и осадок, и супернатант разделяли и сохраняли. Затем осадок суспендировали в 1,0 мл воды (приблизительно 6 мг/мл). Осадок содержал по существу чистый белок 15 кДа при анализе электрофорезом в ДСН-ПААГ.
Было определено, что N-концевая аминокислотная последовательность была следующей: Ser-Ala-Arg-Glu-Val-His-Ile-Glu-Ile-Asn-Asn-Thr-Arg-His-Thr-Leu-Gln-Leu-Glu-Ala-Lys-Thr-Lys-Leu (SEQ ID NO:3).
ПРИМЕР 5
Биоанализ белка
Препарат нерастворимой фракции из диализованного NaBr-экстракта 80JJ1, содержащий пептиды 47 кДа, 45 кДа и 15 кДа, подвергали биоанализу против Западной блошки длинноусой, и было обнаружено, что он проявляет значимую токсиновую активность.
ПРИМЕР 6
Очистка белка и характеристика белка 45 кДа PS149B1
Осадок Р1 ресуспендировали с двумя объемами деионизованной воды на единицу сырого веса и к суспензии добавляли девять объемов 40% (мас./мас.) водного бромида натрия. Эту и все последующие операции проводили на льду или при 4-6°С. Спустя 30 мин суспензию разбавляли 36 объемами охлажденной воды и центрифугировали при 25000 х g в течение 30 мин с получением осадка и супернатанта.
Полученный осадок ресуспендировали в 1-2 объемах воды и наслаивали на 20-40% (мас./мас.) градиент бромида натрия и центрифугировали при 8000 х g в течение 100 мин. Слой, дающий полосу при приблизительно 32% (мас./мас.) бромиде натрия («включения», или INC), извлекали и диализовали в течение ночи против воды с использованием диализной мембраны с отсечением MW 6-8 кДа. Состоящий из частиц материал извлекали центрифугированием при 25000 х g, ресуспендировали в воде и делили на аликвоты и анализировали на белок по способу Лоури и с использованием электрофореза в ДСН-ПААГ.
Полученный супернатант концентрировали в 3-4 раза при помощи концентратора Центрикона-10, затем диализовали в течение ночи против воды с использованием диализной мембраны с отсечением MW 6-8 кДа. Состоящий из частиц материал извлекали центрифугированием при 25000 х g, ресуспендировали в воде и делили на аликвоты и анализировали на белок по способу Лоури и с использованием электрофореза в ДСН-ПААГ. Эта фракция была обозначена как Р1.Р2.
Пептиды в суспензии осадка разделяли при помощи электрофореза в ДСН-ПААГ (Laemlli, U.K., supra) в 15% акриламидных гелях. Затем разделенные белки электрофоретически блоттировали на мембрану PVDF (Millipore Corp.) в 10 мМ CAPS pH 11,0, 10% МеОН при 100 В постоянного тока. Спустя один час мембрану PVDF промывали в воде в течение короткого времени и помещали на 3 мин в 0,25% Кумасси синий R-250, 50% метанол, 5% уксусную кислоту. Окрашенную мембрану обесцвечивали в 40% МеОН, 5% уксусной кислоте. Обесцвеченную мембрану сушили на воздухе при комнатной температуре (LeGendre et al., supra). Мембрану секвенировали с использованием автоматизированной деградации Эдмана в газовой фазе (Hunkapillar et al., supra).
Белковый анализ показал присутствие двух основных полипептидов с молекулярными массами 47 кДа и 14 кДа. Молекулярные массы определяли против стандартных полипептидов с известной молекулярной массой. Этот способ обеспечивает только приближенную оценку истинной молекулярной массы. Полоса 47 кДа из PS149B1 мигрировала при электрофорезе в ДСН-ПААГ точно так, как белок 47 кДа из PS80JJ1. Подобным образом, полоса 14 кДа из PS149B1 мигрировала при электрофорезе на ДСН-ПААГ точно так, как полосы 14 кДа из PS167H2 и PS80JJ1. Кроме этих двух полипептидов, которые, как приблизительно определено, составляют 25-35% (47 кДа) и 35-55% (15 кДа) окрашивающегося Кумасси материала соответственно, могут наблюдаться малые полосы, в том числе полосы приблизительно определенных MW при 46 кДа, 130 кДа и 70 кДа.
Белковый анализ показал, что фракция INC содержала единственный полипептид с MW 47 кДа и что фракция Р1. Р2 содержала единственный полипептид с MW 14 кДа. Эти полипептиды извлекали с выходами более 50% из Р1.
N-концевая аминокислотная последовательность для очищенного белка 47 кДа из PS149B1 представляет собой: Met-Leu-Asp-Thr-Asn-Lys-Val-Tyr-Glu-Ile-Ser-Asn-His-Ala-Asn-Gly-Leu-Tyr-Ala-Ala-Thr-Tyr-Leu-Ser-Leu (SEQ ID NO:4).
N-концевая аминокислотная последовательность для очищенного белка 14 кДа из PS149B1 представляет собой: Ser-Ala-Arg-Glu-Val-His-Ile-Asp-Val-Asn-Asn-Lys-Thr-Gly-His-Thr-Leu-GIn-Leu-Glu-Asp-Lys-Thr-Lys-Leu-Asp-Gly-Gly-Arg-Trp-Arg-Thr-Ser-Pro-Xaa-Asn-Val-Ala-Asn-Asp-Gln-Ile-Lys-Thr-Phe-Val-Ala-Glu-Ser-Asn (SEQ ID NO:5).
ПРИМЕР 7
Аминокислотная последовательность для токсинов 45 кДа и 14 кДа PS167H2
N-концевая аминокислотная последовательность для очищенного белка 45 кДа из PS167H2 представляет собой: Met-Leu-Asp-Thr-Asn-Lys-Ile-Tyr-Glu-Ile-Ser-Asn-Tyr-Ala-Asn-Gly-Leu-His-Ala-Ala-Thr-Tyr-Leu-Ser-Leu (SEQ ID NO:6).
N-концевая аминокислотная последовательность для очищенного белка 14 кДа из PS167H2 представляет собой: Ser-Ala-Arg-Glu-Val-His-Ile-Asp-Val-Asn-Asn-Lys-Thr-Gly-His-Thr-Leu-GIn-Leu-Glu-Asp-Lys-Thr-Lys-Leu (SEQ ID NO:7).
Эти аминокислотные последовательности могут быть сравнены с последовательностью, полученной для пептида 47 кДа из порошков спор/кристаллов 80JJ1, с N-концевой последовательностью (SEQ ID NO:1) и с последовательностью для пептида 14 кДа, полученного из порошков спор/кристаллов 80JJ1, с N-концевой последовательностью (SEQ ID NO:3).
Ясно, что белки 45-47 кДа являются высокородственными и белки 14 кДа являются высокородственными.
ПРИМЕР 8
Биоанализ белка
Очищенные белковые фракции из PS149B1 подвергали биоанализу против Западной блошки длинноусой, и было обнаружено, что они проявляют значимую токсиновую активность при комбинировании. В самом деле, комбинирование восстанавливало активность до активности, отмеченной в исходном препарате (Р1). Нижеследующий набор данных биоанализа представляет процент смертности и демонстрирует этот эффект (таблица 4).
ПРИМЕР 9
Молекулярное клонирование, экспрессия и анализ последовательности ДНК нового гена δ-эндотоксина из штамма Bacillus thurinqiensis PS80JJ1
Общую клеточную ДНК получали из клеток Bacillus thuringiensis (B.t.), выращенных до оптической плотности, при 600 нм, 1,0. Клетки осаждали центрифугированием и ресуспендировали в буфере для протопластов (20 мг/мл лизоцима в 0,3 М сахарозе, 25 мМ Трис-Cl [рН 8,0], 25 мМ ЭДТА). После инкубирования при 37°С в течение 1 часа протопласты лизировали двумя циклами замораживания и оттаивания. Девять объемов раствора 0,1 М NaCl, 0,1% ДСН, 0,1 М Трис-Cl добавляли для завершения лизиса. Осветленный лизат экстрагировали дважды смесью фенол-хлороформ (1:1). Нуклеиновые кислоты осаждали двумя объемами этанола и осадок собирали центрифугированием. Осадок ресуспендировали в ТЕ-буфере и добавляли РНКазу до конечной концентрации 50 мкг/мл. После инкубирования при 37°С в течение 1 часа раствор экстрагировали один раз смесью фенол-хлороформ (1:1) и один раз ТЕ-насыщенным хлороформом. ДНК осаждали из водной фазы добавлением одной десятой объема 3 М NaOAc и двух объемов этанола. Осадок ДНК получали центрифугированием, промывали 70% этанолом, сушили и ресуспендировали в ТЕ-буфере.
Олигонуклеотидный зонд для гена, кодирующего токсин 45 кДа PS80JJ1, конструировали на основании данных о N-концевой пептидной последовательности. Последовательность олигонуклеотидного зонда из 29 оснований была следующей: 5'-ATG YTW GAT ACW ААТ ААА GTW TAT GAA АТ-3' (SEQ ID NO:8). Этот олигонуклеотид был смешанным при четырех положениях, как показано. Этот зонд радиоактивно метили 32P и использовали в стандартных условиях гибридизации Саузерн-блотов общей клеточной ДНК PS80JJ1, расщепленной различными рестрикционными эндонуклеазами (рестриктазами). Характерные авторадиографические данные из этих экспериментов, показывающие размеры рестрикционных фрагментов ДНК, содержащих гомологию последовательности в отношении олигонуклеотидного зонда токсина 44,3 кДа SEQ ID NO:8, представлены в таблице 5.
RFLP фрагментов клеточной ДНК PS80JJ1 на Саузерн-блотах, которые гибридизовались при стандартных условиях с олигонуклеотидным зондом гена токсина 44,3 кДа (SEQ ID NO:8)
Эти ДНК-фрагменты, идентифицированные в этих анализах, содержат весь ген токсина 45 кДа PS80JJ1 или сегмент этого гена. Приблизительные размеры гибридизующихся ДНК-фрагментов в таблице 5 находятся в разумном соответствии с размерами субпопуляции фрагментов PS80JJ1, гибридизующихся с зондом субгена токсина 45 кДа PS80JJ1, используемым в отдельных экспериментах, как предсказано (см. таблицу 6 ниже).
Библиотеку генов конструировали из ДНК PS80JJ1, частично расщепленной Sau3AI. Продукты частичной рестрикции фракционировали электрофорезом на агарозном геле. ДНК-фрагменты 9,3-23 т.п.н. вырезали из геля, электроэлюировали из срезов геля, очищали на ионообменной колонке Elutip-D (Schleicher and Schuell, Keene, NH) и извлекали осаждением этанолом. Sau3AI-вставки лигировали в расщепленный BamHI LamblaGem-11 (Promega, Madison, WI). Рекомбинантный фаг упаковывали и высевали на клетки Е.coli KW251. Бляшки подвергали скринингу гибридизацией с олигонуклеотидным зондом, описанным выше. Гибридизующиеся фаги очищали из бляшек и использовали для инфицирования жидкой культуры клеток Е.coli KW251 для выделения ДНК стандартными процедурами (Maniatis et al., supra).
Блот-анализ по Саузерну выявил, что один из рекомбинантных фаговых изолятов содержал XbaI-SacI-полосу приблизительно 4,8 т.п.н., которая гибридизовалась с зондом гена токсина PS80JJ1. Sacl-сайт, фланкирующий ген токсина PS80JJ1, является клонирующим сайтом фагового вектора, тогда как фланкирующий Xbal-сайт локализован в ДНК-вставке PS80JJ1. Этот рестрикционный ДНК-фрагмент субклонировали стандартными способами в pBluescript S/K (Stratagene, San Diego, CA) для анализа последовательности. Полученная плазмида была обозначена как pMYC2421. Эту ДНК-вставку субклонировали также в pHTBlueII (челночный вектор Е.coli/B. thuringiensis, содержащий pBluescript S/K [Stratagene, La Jolla, CA] и начало репликации из плазмиды-резидента B.t. [D. Lereclus et al. (1989) FEMS Microbiology Letters 60:211-218]) с получением pMYC2420.
Олигонуклеотидный зонд для гена, кодирующего токсин 14 кДа PS80JJ1, конструировали на основании данных о N-концевой пептидной последовательности. Последовательность олигонуклеотидного зонда из 28 оснований была следующей: 5'-GW GAA GTW CAT ATW GAA ATW AAT AAT AC-3' (SEQ ID NO:29). Этот олигонуклеотид был смешанным по четырем положениям, как показано. Этот зонд радиоактивно метили 32P и использовали в стандартных условиях гибридизации Саузерн-блотов общей клеточной ДНК PS80JJ1 и ДНК pMYC2421, расщепленных различными рестрикционными эндонуклеазами (рестриктазами). Эксперименты по RFLP-картированию продемонстрировали, что ген, кодирующий токсин 14 кДа, локализован на том же самом геномном EcoRI-фрагменте, который содержит N-концевую кодирующую последовательность для токсина 44,3 кДа.
Для тестирования экспрессии генов токсина PS80JJ1 в B.t., pMYC2420 трансформировали в некристаллизующегося (Cry-) В.t.-хозяина, CryB (A. Aronson, Purdue University, West Lafayette, IN), электропорацией. Экспрессия как токсина приблизительно 14 кДа, так и токсина 44,3 кДа PS80JJ1, кодируемых pMYC2420, была продемонстрирована электрофорезом в ДСН-ПААГ. Кристаллические препараты токсина из рекомбинантного штамма CryB[pMYC2420], MR536, тестировали, и было обнаружено, что они активны против Западной блошки длинноусой.
Гены токсина PS80JJ1, кодируемые pMYC2421, секвенировали с использованием автоматизированной системы секвенирования ABI373 и соответствующего программного обеспечения. Последовательность всего генетического локуса, содержащая как открытые рамки считывания, так и фланкирующие нуклеотидные последовательности, показана в SEQ ID NO:30. Терминирующий кодон гена токсина 14 кДа представляет собой 121 пару оснований выше по ходу транскрипции (5') от инициирующего кодона гена токсина 44,3 кДа (фигура 2). Нуклеотидная последовательность открытой рамки считывания токсина 14 кДа PS80JJ1 (SEQ ID NO:31), нуклеотидная последовательность открытой рамки считывания токсина 44,3 кДа (SEQ ID NO:10) и соответствующие расшифрованные аминокислотные последовательности (SEQ ID NO:32 и SEQ ID NO:11) являются новыми в сравнении с генами других токсинов, кодирующими пестицидные белки.
Таким образом, нуклеотидная последовательность, кодирующая токсин 14 кДа PS80JJ1, показана в SEQ ID NO:31. Расшифрованная аминокислотная последовательность токсина 14 кДа PS80JJ1 показана в SEQ ID NO:32. Нуклеотидные последовательности, кодирующие как токсин 14 кДа, так и токсин 45 кДа PS80JJ1, а также фланкирующие последовательности, показаны в SEQ ID NO:30. Взаимосвязь этих последовательностей показана на фигуре 2.
Субкультура Е.coli NM552, содержащая плазмиду pMYC2421, была депонирована в постоянной коллекции Patent Culture Collection (NRRL), Regional Research Center, 1815 North University Street, Peoria, Illinois 61604, USA 28 марта 1996 года. Номер доступа NRRL B-21555.
ПРИМЕР 10
RFLP- и ПЦР-анализ дополнительных новых генов δ-эндотоксинов из штаммов PS149B1 и PS167H2 Bacillus thuringiensis
Два дополнительных штамма, активных против блошки длинноусой, PS149B1 и PS167H2, также продуцируют параспоральные белковые кристаллы, состоящие частично из полипептидов размером приблизительно 14 и 45 кДа. Гибридизация по Саузерну и частичный анализ последовательности ДНК использовали для исследования родства этих токсинов с токсинами 80JJ1. ДНК экстрагировали из этих штаммов B.t., как описано выше, и стандартные гибридизации по Саузерну проводили с использованием олигонуклеотидного зонда токсина 14 кДа (SEQ ID NO:29) и ПЦР-фрагмента приблизительно 800 пар оснований последовательности, кодирующей ген токсина 44,3 кДа 80JJ1. Данные RFLP из этих экспериментов, показывающие размеры рестрикционных фрагментов ДНК, содержащих гомологию последовательности в отношении токсина 44,3 кДа, представлены в таблице 6. Данные RFLP из этих экспериментов, показывающие размеры рестрикционных фрагментов ДНК, содержащих гомологию последовательности в отношении токсина приблизительно 14 кДа, представлены в таблице 7.
RFLP фрагментов клеточной ДНК PS80JJ1, PS149B1 и PS167H2 на Саузерн-блотах, которые гибридизовались с олигонуклеотидным зондом субгена токсина 44,3 кДа 800 п.н. PS80JJ1 при стандартных условиях
Каждый из трех штаммов обнаружил уникальные параметры RFLP. Гибридизующиеся ДНК-фрагменты из PS149B1 или PS167H2 содержат все гены токсинов или часть генов токсинов с гомологией последовательности относительно токсина 44,3 кДа PS80JJ1.
Полиморфизмы длины рестрикционных фрагментов клеточной ДНК PS80JJ1, PS149B1 и PS167H2 на Саузерн-блотах, которые гибридизовались с олигонуклеотидным зондом токсина 14 кДа PS80JJ1 при стандартных условиях
Каждый из трех штаммов обнаружил уникальные параметры RFLP. Гибридизующиеся ДНК-фрагменты из PS149B1 или PS167H2 содержат все гены токсинов или часть генов токсинов с гомологией последовательности относительно гена токсина 14 кДа PS80JJ1.
Части генов токсинов PS149B1 или PS167H2 амплифицировали при помощи ПЦР с использованием пары прямого и обратного олигонуклеотидных праймеров, сконструированных на основе последовательности гена токсина 44,3 кДа PS80JJ1. Для PS149B1 использовали следующую пару праймеров:
Прямой:
5'-ATG YTW GAT ACW AAT AAA GTW TAT GAA AT-3' (SEQ ID NO:8)
Обратный:
S'-GGA TTA ТСТ АТС ТСТ GAG TGT TCT TG-3' (SEQ ID NO:9) Для PS167H2 использовали ту же самую пару праймеров. Полученные ПЦР фрагменты секвенировали с использованием автоматизированной системы секвенирования ABI373 и связанного с ней программного обеспечения. Полученные частичные генные и пептидные последовательности показаны в SEQ ID NO:12-15. Эти последовательности содержат части нуклеотидных кодирующих последовательностей и пептидных последовательностей новых активных в отношении блошки длинноусой токсинов, присутствующих в штаммах B.t. PS149B1 или PS167H2.
ПРИМЕР 11
Молекулярное клонирование и анализ
последовательности ДНК новых генов δ-эндотоксинов из штаммов Bacillus thurinqiensis PS149B1 и PS167H2
Общую клеточную ДНК экстрагировали из штаммов PS149B1 и PS167H2, как описано для PS80JJ1. Библиотеки генов фракционированных по размеру частичных рестрикционных фрагментов Sau3A конструировали в LambdaGem-11 для каждого соответствующего штамма, как описано ранее. Рекомбинантный фаг упаковывали и высевали на клетки Е.coli KW251. Бляшки подвергали скринингу гибридизацией с олигонуклеотидным зондом, специфическим для гена токсина 44 кДа. Гибридизующиеся фаги очищали из бляшек и использовали для инфицирования жидкой культуры клеток Е.coli KW251 для выделения ДНК стандартными процедурами (Maniatis et al., supra).
Для PS167H2, блот-анализ по Саузерну выявил, что один из рекомбинантных фаговых изолятов содержал HindIII-полосу приблизительно 4,0-4,4 т.п.н., которая гибридизовалась с 5'-олигонуклеотидным зондом гена токсина 44 кДа PS80JJ1 (SEQ ID NO:8). Этот рестрикционный фрагмент ДНК субклонировали стандартными способами в pBluescript S/K (Stratagene, San Diego, CA) для анализа последовательности. Этот фрагмент субклонировали также в высококопийный челночный вектор, рНТ370 (Arantes, О., D.Lereclus [1991] Gene 108:115-119) для анализов экспрессии в Bacillus thuringiensis (см. ниже). Полученная рекомбинантная высококопийная бифункциональная плазмида была обозначена как pMYC2427.
Гены токсина PS167H2, кодируемые pMYC2427, секвенировали с использованием автоматизированной системы секвенирования ABI и связанного с ней программного обеспечения. Последовательность всего генетического локуса, содержащая как открытые рамки считывания, так и фланкирующие нуклеотидные последовательности, показана в SEQ ID NO:34. Терминирующий кодон гена токсина 14 кДа представляет собой 107 пар оснований выше по ходу транскрипции (5') от инициирующего кодона гена токсина 44 кДа. Кодирующая последовательность токсина 14 кДа PS167H2 (SEQ ID NO:35), кодирующая последовательность токсина 44 кДа (SEQ ID NO:37) и соответствующие расшифрованные аминокислотные последовательности (SEQ ID NO:36 и SEQ ID NO:38) являются новыми в сравнении с другими известными генами токсинов, кодирующими пестицидные белки. Эти гены токсинов расположены сходным образом и имеют некоторую гомологию с токсинами 14 и 44 кДа PS80JJ1.
Субкультура Е.coli NM522, содержащая плазмиду pMYC2427, была депонирована в постоянной коллекции Patent Culture Collection (NRRL), Regional Research Center, 1815 North University Street, Peoria, Illinois 61604, USA 26 марта 1997 года. Номер доступа NRRL B-21672.
Для PS149B1, блот-анализ по Саузерну с использованием олигонуклеотидного 5'-зонда 44 кДа PS80JJ1 (SEQ ID NO:8) продемонстрировал гибридизацию ClaI-фрагмента ДНК приблизительно 5,9 т.п.н. Полные продукты расщепления Clal геномной ДНК PS149B1 фракционировали по размеру на агарозных гелях и клонировали в pHTBlueII. Этот фрагмент субклонировали также в высококопийный челночный вектор, рНТ370 (Arantes, О., D.Lereclus [1991] Gene 108:115-119) для анализов экспрессии в Bacillus thuringiensis (см. ниже). Полученная рекомбинантная высококопийная бифункциональная плазмида была обозначена как PMYC2429.
Гены токсина PS149B1, кодируемые pMYC2429, секвенировали с использованием автоматизированной системы секвенирования ABI и связанного с ней программного обеспечения. Последовательность всего генетического локуса, содержащая как открытые рамки считывания, так и фланкирующие нуклеотидные последовательности, показана в SEQ ID NO:39. Терминирующий кодон гена токсина 14 кДа представляет собой 108 пар оснований выше по ходу транскрипции (5') от инициирующего кодона гена токсина 44 кДа. Кодирующая последовательность токсина 14 кДа PS149B1 (SEQ ID NO:40), кодирующая последовательность токсина 44 кДа (SEQ ID NO:42) и соответствующие расшифрованные аминокислотные последовательности (SEQ ID NO:41 и SEQ ID NO:43) являются новыми в сравнении с другими известными генами токсинов, кодирующими пестицидные белки. Эти гены токсинов расположены сходным образом и имеют некоторую гомологию с токсинами 14 и 44 кДа PS80JJ1 и PS167H2. Вместе эти три оперона токсинов составляют новое семейство пестицидных токсинов.
Субкультура Е.coli NM522, содержащая плазмиду pMYC2429, была депонирована в постоянной коллекции Patent Culture Collection (NRRL), Regional Research Center, 1815 North University Street, Peoria, Illinois 61604, USA 26 марта 1997 года. Номер доступа NRRL B-21673.
ПРИМЕР 12
ПЦР-амплификация для идентификации и клонирования нового активного в отношении блошки длинноусой токсина
ДНК- и пептидные последовательности трех новых активных в отношении блошки длинноусой токсинов приблизительно 45 кДа из PS80JJ1, PS149B1 и PS167H2 (SEQ ID NO:12-15) сопоставляли с использованием программы Pileup анализа последовательности Genetics Computer Group с использованием веса гэпа 3,00 и веса длины гэпа 0,10. Сопоставления последовательностей использовали для идентификации консервативных пептидных последовательностей, к которым конструировали олигонуклеотидные праймеры, которые были подходящими для гибридизации с генами, кодирующими членов этого нового семейства токсинов. Такие праймеры могут быть использованы в ПЦР для амплификации диагностических ДНК-фрагментов для этих и родственных генов токсинов. Возможны многочисленные конструкции праймеров к различным последовательностям, четыре из которых описаны здесь в качестве примера. Этими пептидными последовательностями являются:
Asp-Ile-Asp-Asp-Tyr-Asn-Leu (SEQ ID NO:16)
Trp-Phe-Leu-Phe-Pro-Ile-Asp (SEQ ID NO:17)
GIn-Ile-Lys-Thr-Thr-Pro-Tyr-Tyr (SEQ ID NO:18)
Tyr-Glu-Trp-Gly-Thr-Glu (SEQ ID NO:19).
Соответствующими нуклеотидными последовательностями являются:
5'-GATATWGATGAYTAYAAYTTR-3' (SEQ ID NO:20)
5'-TGGTTTTTRTTTCCWATWGAY-3' (SEQ ID NO:21)
5'-CAAATHAAAACWACWCCATATTAT-3' (SEQ ID NO:22)
5'-TAYGARTGGGGHACAGAA-3' (SEQ ID NO:23)
Прямые праймеры для полимеразной амплификации в термоциклической реакции были сконструированы на основе нуклеотидных последовательностей SEQ ID NO:20 и 21.
Обратные праймеры были сконструированы на основе обратного комплемента SEQ ID NO:22 и 23:
5'-ATAATATGGWGTWGTTTTDATTTG-3' (SEQ ID NO:24)
5'-TTCTGTDCCCCAYTCRTA-3'(SEQ ID NO:25)
Эти праймеры могут быть использованы в комбинации для амплификации ДНК-фрагментов следующих размеров (таблица 8), которые идентифицируют гены, кодирующие новые активные против блошки длинноусой токсины.
Предсказанные размеры диагностических ДНК-фрагментов (п.н.), амплифицируемых с праймерами, специфическими для новых активных против блошки длинноусой токсинов
Подобным образом, полные гены новых активных против блошки длинноусой токсинов могут быть выделены полимеразной амплификацией в термоциклических реакциях с использованием праймеров, сконструированных на основе последовательностей ДНК, фланкирующих открытые рамки считывания. Для токсина 44,3 кДа PS80JJ1 одна подобная пара праймеров была сконструирована, синтезирована и использована для амплификации диагностического ДНК-фрагмента 1613 п.н., который включал полную кодирующую последовательность токсина. Этими праймерами были:
Прямой: 5'-CTCAAAGCGGATCAGGAG-3' (SEQ ID NO:26)
Обратный: 5'-GCGTATTCGGATATGCTTGG-3' (SEQ ID NO:27)
Для ПЦР-амплификации токсина 14 кДа PS80JJ1, олигонуклеотид, кодирующий N-концевую пептидную последовательность (SEQ ID NO:29), может быть использован в комбинации с различными обратными олигонуклеотидными праймерами на основе последовательностей в локусе гена токсина PS80JJ1. Один такой обратный праймер имеет следующую последовательность:
5'-CATGAGATTTATCTCCTGATCCGC-3' (SEQ ID NO:33).
При использовании в стандартных ПЦР-реакциях эта пара праймеров амплифицировала диагностический ДНК-фрагмент 1390 п.н., который включает полную кодирующую последовательность токсина 14 кДа и некоторую часть 3'-фланкирующих последовательностей, соответствующих межгенному спейсеру из 121 оснований и части гена токсина 44,3 кДа. При использовании в комбинации с прямым праймером 14 кДа ПЦР будет генерировать диагностический ДНК-фрагмент 322 п.н.
ПРИМЕР 13
Биоанализы клонов на зависимость ответной реакции от дозы
Оперон токсина PS80JJ1 субклонировали из pMYC2421 в рНТ370 для прямого сравнения биоактивности с рекомбинантными токсинами, клонированными из PS149B1 и PS167H2. Полученная рекомбинантная высококопийная бифункциональная плазмида была обозначена как pMYC2426.
Субкультура Е.coli NM522, содержащая плазмиду pMYC2426, была депонирована в постоянной коллекции Patent Culture Collection (NRRL), Regional Research Center, 1815 North University Street, Peoria, Illinois 61604, USA 26 марта 1997 года. Номер доступа NRRL B-21671.
Для тестирования экспрессии генов токсинов PS80JJ1, PS149B1 и PS167H2 в B.t., pMYC2426, pMYC2427 и pMYC2429 no отдельности трансформировали в акристаллического (Cry-) В.t.-хозяина, CryB (A. Aronson, Purdue University, West Lafayette, IN), электропорацией. Рекомбинантные штаммы были обозначены как MR543 (CryB [pMYC2426]), MR544 (СгуВ [pMYC2427]) и MR546 (CryB [pMYC2429]) соответственно. Экспрессию обоих токсинов приблизительно 14 и 44 кДа продемонстрировали электрофорезом в ДСН-ПААГ для каждого рекомбинантного штамма.
Кристаллические препараты токсинов из рекомбинантных штаммов анализировали против западной блошки длинноусой. Рацион насекомых был дополнен сорбиновой кислотой и pen-strep-ampho-B Sigma. Материал наносили сверху при норме 50 мкл суспензии на см2 площади зоны рациона. Биотесты проводили с новорожденными личинками западной блошки длинноусой в течение 4 дней при приблизительно 25°С. Процентную смертность и приблизительные оценки LC50 для верхнего нанесения для этих клонов (осадков) представлены в таблице 9. Контроль дН2О давал 7% смертность.
Количества белков 14 кДа и 44,3 кДа, присутствующих в кристаллических препаратах, определяли денситометрией и использовали для расчета удельной активности, выражаемой в виде LC50. Приблизительные оценки LC50 для этих клонов (осадков) представлены в таблице 10 (биоанализ WCRW с верхним нанесением В.t.-клонов).
Биоанализ WCRW с верхним нанесением В.t.-клонов
**90% CL
ПРИМЕР 14
Мутационный анализ полипептидов 14 и 44 кДа в опероне бинарного токсина PS80JJ1
Гены бинарных токсинов данного изобретения, в их состоянии дикого типа (немутантном состоянии), обычно расположены в опероне, в котором сначала транскрибируется ген белка 14 кДа, а затем транскрибируется ген белка 45 кДа. Эти гены разделены относительно коротким некодирующим участком. Характерные ОРС показаны в SEQ ID NO:30, SEQ ID NO:34 и SEQ ID NO:39.
Для исследования вклада индивидуальных кристаллических белков 14 и 44,3 кДа в активность против блошки длинноусой каждый ген в опероне PS80JJ1 мутировали в отдельных экспериментах для отмены экспрессии одного из этих белков. Затем интактный ген экспрессировали в B.t. и рекомбинантные белки испытывали на активность против блошки длинноусой.
Сначала, ген 44,3 кДа, кодируемый на pMYC2421, мутировали укорочением при сайте EcoRI в положении оснований 387 открытой рамки считывания. Это укорочение и последующее лигирование с векторными последовательностями приводило к открытой рамке считывания, кодирующей гипотетический гибридный (слитый) белок приблизительно 24 кДа. Полученный оперон, кодирующий интактный ген 14 кДа и укороченный ген 45 кДа, субклонировали в высококопийный челночный вектор, рНТ370 (Arantes, О., D.Lereclus [1991] Gene 108:115-119) для анализов экспрессии в Bacillus thuringiensis. Полученную плазмиду, pMYC2424, трансформировали в акристаллического (Cry-) В.t.-хозяина, CryB (A. Aronson, Purdue University, West Lafayette, IN), электропорацией. Полученный рекомбинантный штамм был обозначен как MR541. Только белок 14 кДа PS80JJ1 детектировался анализом с использованием электрофореза в ДСН-ПААГ спорулированных культур MR541. Смертность не наблюдали для препаратов MR541, экспрессирующих только белок 14 кДа PS80JJ1, в биоанализах с верхним нанесением против блошки длинноусой.
Затем ген 14 кДа, кодируемый на pMYC2421, мутировали встраиванием олигонуклеотидного линкера, содержащего терминирующие кодоны во всех возможных рамках считывания при сайте Nrul в положении оснований 11 открытой рамки считывания. Последовательность этого линкера была 5'-TGAGTAACTAGATCTATTCAATTA-3'. Этот линкер вводит сайт BglII для подтверждения встраивания рестрикционным BglII-расщеплением. Плазмидные клоны, содержащие мутагенный линкер, идентифицировали с использованием BglII и секвенировали для проверки. Вставку оперона, кодирующую нонсенс-мутации, субклонировали в рНТ370, получая плазмиду pMYC2425. Эту плазмиду трансформировали в СгуВ электропорацией с получением рекомбинантного В.t.-штамма MR542. Только белок 44,3 кДа PS80JJ1 экспрессировался в спорулированных культурах MR542, как показано анализом, с использованием электрофореза в ДСН-ПААГ. Смертность против блошки длинноусой не наблюдали для препаратов MR542, экспрессирующих только белок 44,3 кДа.
ПРИМЕР 15
Гетерологичная экспрессия отдельных генов, очистка и биоанализ полипептидов 14 и 44,3 кДа из PS149B1 в Pseudomonas fluorescens
Гены полипептидов 14 кДа и 44,3 кДа из PS149B1 конструировали отдельно в плазмидные векторы стандартными способами клонирования ДНК и трансформировали в Pseudomonas fluorescens. Рекомбинантный штамм Pseudomonas fluorescens, экспрессирующий только ген 14 кДа PS149B1, был обозначен как MR1253. Рекомбинантный штамм Pseudomonas fluorescens, экспрессирующий только ген 44,3 кДа PS149B1, был обозначен как MR1256.
MR1253 и MR1256, каждый из которых экспрессирует индивидуально один из двух бинарных белков, выращивали в ферментационных резервуарах на 1 л. Затем часть каждой культуры осаждали центрифугированием, лизировали лизоцимом и обрабатывали ДНКазой I с получением полуочищенных белковых включений. Затем эти белковые включения солюбилизировали в 50 мМ цитрате натрия (рН 3,3) осторожным качанием при 4°С в течение 1 часа. Белок 14 кДа легко растворялся в этом буфере, тогда как белок 44,3 кДа был частично растворимым. Затем солюбилизированные фракции центрифугировали при 15000 х g в течение 20 минут и супернатанты сохраняли.
Белок 14 кДа дополнительно очищали ионообменной хроматографией. Солюбилизированный белок 14 кДа связывали с колонкой Econo-S и элюировали 0-1 М градиентом хлорида натрия.
Затем хроматографически чистый MR1253 (белок 14 кДа) и солюбилизированный цитратом натрия (рН 3,3) препарат MR1256 (белок 45 кДа) испытывали на активность на блошке длинноусой по отдельности или вместе при молярном соотношении 1:10 (белок 45 кДа:белок 14 кДа). Наблюдаемая смертность для каждого из этих белков по отдельности была не выше уровней фона (пробы вода/контроль), но получали 87% смертность при комбинировании их в вышеуказанном соотношении (см. таблицу 11).
ПРИМЕР 16
Идентификация дополнительных генов новых токсинов 14 кДа и 44,3 кДа гибридизацией общей геномной ДНК B.t. и анализом RFLP
Общую геномную ДНК из каждого изолята получали с использованием набора Qiagen DNEasy 96 well tissue. ДНК в 96-луночных планшетах денатурировали перед блоттингом добавлением 10 мкл каждой пробы ДНК и 10 мкл 4 М NaOH к 80 мкл стерильной дистиллированной воды. Пробы инкубировали при 70°С в течение одного часа, после чего в каждую лунку добавляли 100 мкл 20Х SSC. Общую геномную ДНК PS149B1 включали с каждым набором из 94 проб в качестве положительного контроля гибридизации, а общую геномную ДНК CryB включали с каждым набором из 94 проб в качестве отрицательного контроля гибридизации. Каждый набор из 96 проб наносили на найлоновые мембраны Magnacharge с использованием двух 48-луночных слот-блот-распределителей (Hoefer Scientific), с последующими двумя промывками 10 Х SSC. Мембраны отжигали при 80°С в течение одного часа и хранили в сухом виде до использования. Мембраны предгибридизовали и гибридизовали в стандартном формамидном растворе (50% формамид, 5Х SSPE, 5X раствор Денхардта, 2% ДСН, 100 мкг/мл одноцепочечной ДНК) при 42°С. Мембраны промывали при двух типах условий: 2Х SSC/0,1% ДСН при 42°С (низкой жескости) и 0,2Х SSC/0,1% ДСН при 65°С (умеренной-высокой жесткости). Мембраны зондировали ПЦР-фрагментом приблизительно 1,3 т.п.н. гена 44,3 кДа PS149B1, амплифицированным из pMYC2429 с использованием прямого праймера SEQ ID NO:8 и обратного праймера с последовательностью 5'-GTAGAAGCAGAACAAGAAGGTATT-3' (SEQ ID NO:46). Зонд радиоактивно метили с использованием набора Prime-it II (Stratagene) и 32P-dCTP, очищали на колонках Сефадекса, денатурировали при 94°С и добавляли к свежему гибридизационному раствору. Штаммы, содержащие гены с гомологией относительно зонда PS149B1, идентифицировали экспонированием мембран на рентгеновской пленке.
Следующие штаммы были идентифицированы по положительным реакциям гибридизации: PS184M2, PS185GG, PS187G1, PS187Y2, PS201G, PS201HH2, PS242K10, PS69Q, КВ54А1-6, KR136, KR589, PS185L12, PS185W3, PS185Z11, PS186L9, PS187L14, PS186FF, PS131W2, PS147U2, PS158T3, PS158X10, PS185FF, PS187F3, PS198H3, PS201H2, PS201L3, PS203G2, PS203J1, PS204C3, PS204G4, PS204I11, PS204J7, PS210B, PS213E8, PS223L2, PS224F2, PS236B6, PS246P42, PS247C16, KR200, KR331, KR625, KR707, KR959, KR1209, KR1369, КВ2С-4, КВ10Н-5, КВ456, КВ42С17-13, КВ45А43-3, КВ54А33-1, КВ58А10-3, КВ59А54-4, KB59A54-5, КВ53В7-8, КВ53В7-2, KB60F5-7, KB60F5-11, КВ59А58-4, KB60F5-15, КВ61А18-1, КВ65А15-2, КВ65А15-3, КВ65А15-7, КВ65А15-8, КВ65А15-12, КВ65А14-1, KB3F-3, T25, KB53A71-6, KB65A11-2, KB68B57-1, KB63A5-3 и КВ71А118-6.
Дополнительную идентификацию и классификацию новых генов токсинов в препаратах общей геномной ДНК выполняли с использованием 32Р-меченых зондов и условий гибридизации, описанных выше в этом примере. Общую геномную ДНК получали, как описано выше, или с набором Qiagen Genomic-Tip 20/G, и набором буферов геномной ДНК согласно протоколу для грамположительных бактерий (Qiagen Inc.; Valencia, CA) использовали в анализах по Саузерну. Для Саузерн-блотов приблизительно 1-2 мкг общей геномной ДНК из каждого идентифицированного слот-блот-анализом штамма расщепляли ферментами Dral и Ndel, подвергали электрофорезу на 0,8% агарозном геле и иммобилизовали на стандартной найлоновой мембране с использованием стандартных способов (Maniatis et al.). После гибридизации мембраны промывали при низкой жесткости (2Х SSC/0,1% ДСН при 42°С) и экспонировали на пленке. Размеры ДНК-фрагментов определяли с использованием программного обеспечения BioRad Chemidoc system. Полиморфизмы длины рестрикционных фрагментов использовали для (произвольной) классификации генов, кодирующих токсин 44 кДа. Эти классификации приведены в таблице 12.
ПРИМЕР 17
ДНК-секвенирование дополнительных генов бинарных токсинов
Вырожденные олигонуклеотиды были сконструированы для амплификации полных генов 14 и 44,3 кДа или части этих генов из В.t.-штаммов, идентифицированных гибридизацией с описанным выше ПЦР-продуктом 149В1. Эти олигонуклеотиды были сконструированы с консервативными блоками последовательностей, идентифицированными сопоставлением генов 14 кДа или 44,3 кДа из PS149B1, PS167H2 и PS80JJ1. Прямые праймеры для обоих генов были сконструированы таким образом, что они начинаются с инициирующего кодона ATG. Обратные праймеры были сконструированы насколько возможно близко к 3'-концу каждого соответствующего гена.
Праймеры, сконструированные для амплификации гена 14 кДа, были следующими:
149DEG1 (прямой): 5'-ATG TCA GCW CGY GAA GTW CAY ATT G-3' (SEQ ID NO:47)
149DEG2 (обратный): 5'-GTY TGA ATH GTA TAH GTH АСА TG-3' (SEQ ID NO:48)
Эти праймеры амплифицируют продукт приблизительно 340 п.н.
Праймеры, сконструированные для амплификации гена 44,3 кДа, были следующими:
149DEG3 (прямой): 5'-ATG TTA GAT ACW ААТ ААА RTW TAT G-3' (SEQ ID NO:49)
149DEG4 (обратный): 5'-GTW ATT TCT TCW ACT TCT TCA TAH GAA G-3' (SEQ ID NO:50)
Эти праймеры амплифицируют продукт приблизительно 1100 п.н.
Условия ПЦР, используемые для амплификации генных продуктов, являются следующими:
95°С, 1 мин, один цикл
95°С, 1 мин
50°С, 2 мин, повторяли 35 циклов
72°С, 2 мин
72°С, 10 мин, один цикл
ПЦР-продукты фракционировали на 1% агарозных гелях и очищали из матрикса геля с использованием набора Qiaexll (Qiagen). Полученные очищенные фрагменты лигировали в клонирующий вектор pCR-TOPO с использованием набора для клонирования ТОРО ТА (Invitrogen). После лигирования одну половину реакции лигирования трансформировали в XL10 Gold ультракомпетентные клетки (Stratagene). Затем трансформанты подвергали скринингу при помощи ПЦР с векторными праймерами 1212 и 1233. Клоны, содержащие вставки, выращивали на среде LB/карбенициллин для получения плазмид с использованием набора Qiagen plasmid DNA miniprep (Qiagen). Затем клонированные ПЦР-полученные фрагменты секвенировали с использованием автоматической системы секвенирования Applied Biosystems и соответствующего программного обеспечения. Последовательности дополнительных новых генов и полипептидов бинарных токсинов, относящихся к голотипу токсинов 14 и 44,3 кДа из PS80JJ1 и PS149B1, представлены в списке в виде SEQ ID NO:51-126. Приведенный ранее раздел "Краткое описание последовательностей" обеспечивает дополнительное объяснение этих последовательностей.
Токсины и гены 14 кДа-типа из трех дополнительных B.t.-штаммов PS137A, PS201V2 и PS207C3 были также секвенированы с использованием описанных выше процедур (с любыми различиями, указанными ниже). Проводили ПЦР с использованием 149DEG1 (прямого) и 149DEG2 (обратного) праймеров. Эти праймеры ам-плифицируют продукт приблизительно 340 п.н. ПЦР проводили при следующих условиях:
1. 95°С, 3 мин
2. 94°С, 1 мин
3. 42°С, 2 мин
4. 72°С, 3 мин+5 сек/цикл
5. Стадии 2-4 повторяли 29 раз
ПЦР-продукты очищали на геле с использованием набора для экстракции гелей QiaQuick (Qiagen), очищенный фрагмент лигировали в клонирующий вектор pCR-TOPO с использованием набора для клонирования ТОРО ТА (Invitrogen) и затем трансформировали в XL10 Gold ультракомпетентные клетки Е.coli (Stratagene). Получение ДНК трансформантов описано выше. Последовательности гена токсина 14 кДа для каждого из трех новых штаммов получали, как описано выше. Нуклеотидная и полипептидная последовательности приведены в прилагаемом списке последовательностей следующим образом: PS137A (SEQ ID NO:149 и 150), PS201V2 (SEQ ID NO:151 и 152) и PS207C3 (SEQ ID NO:153 и 154).
ПРИМЕР 18
Трансгены токсина PS149B1 и трансформация растений
Отдельные синтетические трансгены, оптимизированные использованием кодонов кукурузы, были сконструированы для обоих компонентов токсинов 14 и 44,3 кДа. Синтетические версии конструировали для модификации неоднозначности кодонов гуанина и цитозина до уровня, более типичного для ДНК растений. Предпочтительные оптимизированные для растений трансгены описаны в SEQ ID NO:127-128. Промоторным участком, используемым для экспрессии обоих трансгенов, был промотор убихитина Zea mays плюс экзон 1 Z. mays и интрон 1 Z.mays (Christensen, A.H. et al. (1992) Plant Mol. Biol. 18:675-689). Терминатором транскрипции, используемым для обоих трансгенов, был терминатор ингибитора II протеиназы картофеля (PinII) (An, G. et al. 1989 Plant Cell 1:115-22).
Фосфинотрицинацетилтрансферазу (PAT) использовали в качестве селектируемого маркера для трансформации растений. Ген фосфинотрицинацетилтрансферазы (pat) выделяли из бактерии Streptomyces viridochromogenes (Eckes P. et al., 1989). Белок PAT ацетилирует фосфинотрицин или его предшественник деметилфосфинотрицин, обеспечивая устойчивость к химически синтезированному фосфинотрицину, такому как гербицид глуфозинат-аммоний. Ацетилирование превращает фосфинотрицин в неактивную форму, которая уже нетоксична для растений кукурузы. Глуфозинат-аммоний является несистемным, неселективным гербицидом широкого спектра действия. Регенерация тканей кукурузы или индивидуальных растений кукурузы, устойчивых к гербициду глуфозинат-аммонию, может быть легко идентифицирована по включению PAT в среду для регенерации или опрыскиванием этим гербицидом листьев.
Получали синтетическую версию pat-гена для модификации неоднозначности кодона гуанина и цитозина до уровня, более типичного для ДНК растений. Промотором pat-гена является промотор CaMV транскрипта 35S из вируса мозаики цветной капусты (Pietrzak et al., 1986). Терминатором транскрипции является терминатор 35S CaMV.
Для трансформации ткани кукурузы линейную часть ДНК, содержащую кодирующие последовательности как 14 кДа и 44,3 кДа PS149B1, так и селектируемого маркера pat и регуляторные компоненты, необходимые для экспрессии, вырезали из полной плазмиды. Эту линейную часть ДНК, называемую вставки, использовали в процессе трансформации.
Растения кукурузы, содержащие трансгены 14 кДа и 44,3 кДа PS149B1, получали бомбардировкой микроснарядами с использованием генного ружья Biolistics®Ò PDS-100He, производимого Bio-Rad, по существу, как описано Klein et al., (1987). Незрелые зародыши, выделенные из «колосьев» кукурузы, собранных спустя приблизительно 15 дней после опыления, культивировали на средах для инициации каллусов в течение трех-восьми дней. В день трансформации микроскопические частицы вольфрама покрывали очищенной ДНК и ускоряли для вхождения их в культивированные зародыши, где ДНК-вставку включали в клеточную хромосому. Спустя шесть дней после бомбардировки подвергнутые бомбардировке зародыши переносили на среду для инициации каллуса, содержащую глуфозинат (Bialaphos) в качестве селектирующего агента. Получали здоровую устойчивую каллусную ткань и повторяемо переносили ее на свежую селективную среду в течение приблизительно 12 недель. Растения регенерировали и переносили в парник. Всего получали 436 регенерированных растений. Пробы листьев брали для молекулярного анализа для подтверждения присутствия трансгенов при помощи ПЦР и для подтверждения экспрессии чужеродного белка с использованием ELISA. Затем растения подвергали биоанализу с целым растением с использованием блошки длинноусой. Положительные растения скрещивали с инбредными линиями для получения семян из исходных трансформированных растений. Было обнаружено, что эти растения были устойчивыми к повреждению блошкой длинноусой как в тепличных, так и в полевых испытаниях.
ПРИМЕР 19
Дополнительные биоанализы
Белковые препараты из штаммов, идентифицированных в примере 16, анализировали на активность против западной блошки длинноусой с использованием основных способов анализа с верхним нанесением, как описано в примере 13. Результаты показаны в таблице 13.
ПРИМЕР 20
Молекулярное клонирование, экспрессия и анализ последовательности ДНК новых генов бинарных эндотоксинов из штамма Bacillus thuringiensis PS201L3
Геномную ДНК из PS201L3 получали из клеток, выращиваемых в культуре во встряхиваемых колбах, с использованием набора Qiagen Genomic-Tip 500/G и набора буферов геномных ДНК согласно протоколу для грамположительных бактерий (Qiagen Inc.; Valencia, CA). Библиотеку генов конструировали из ДНК PS201L3, частично расщепленной Sau3AI. Продукты частичной рестрикции фракционировали электрофорезом в агарозном геле. ДНК-фрагменты размером 9,3-23 т.п.н. вырезали из геля, электроэлюировали из срезов геля, очищали на ионообменной колонке Elutip-D (Schleicher and Schuell, Keene, NH) и выделяли осаждением этанолом. Sau3AI-вставки лигировали в расщепленный BamHI LamblaGem-11 (Promega, Madison, WI). Рекомбинантный фаг упаковывали с использованием пакующего экстракта Gigapack III XL (Stratagene, La Jolla, CA) и высевали на клетки Е.coli KW251. Бляшки поднимали на Nytran Nylon Transfer мембраны (Schleicher and Schuell, Keene, NH) и зондировали 32Р-dCTP-меченым генным зондом для кодирующих бинарный токсин последовательностей. Этот генный зонд был ПЦР-продуктом приблизительно 1,0 т.п.н., амплифицированным с использованием геномной ДНК PS201L3 в качестве матрицы и олигонуклеотидов "15kfor1" и "45krev6".
Последовательности олигонуклеотидов, используемых для ПЦР и секвенирования, были следующими:
15kfor1 (SEQ ID NO:131) ATGTCAGCTCGCGAAGTACAC
45krev6 (SEQ ID NO:132) GTCCATCCCATTAATTGAGGAG
Мембраны гибридизовали с зондом в течение ночи при 65°С и затем промывали три раза 1X SSPE и 0,1% ДСН. Тринадцать бляшек идентифицировали радиоавтографией. Эти бляшки затем извлекали и замачивали в течение ночи в 1 мл SM-буфера+10 мкл CHCl3. Фаг высевали для конфлюэнтного лизиса на клетки-хозяева KW251; 6 конфлюэнтных чашек замачивали в SM и использовали для крупномасштабных получений фаговой ДНК. Очищенную фаговую ДНК расщепляли различными ферментами и подвергали электрофорезу на 0,7% агарозных гелях. Гели переносили на мембраны Nytran блоттингом по Саузерну и зондировали тем же самым ПЦР-амплифицированным ДНК-фрагментом, описанным выше. Идентифицировали гибридизующуюся полосу Xbal приблизительно 6,0 т.п.н. и субклонировали в рНТ370, челночный вектор Е.coli/Bacillus thuringiensis (Arantes, О., D. Lereclus [1991] Gene 108:115-119) для получения pMYC2476. XL10 Gold ультракомпетентные клетки Е.coli (Stratagene), трансформированные pMYC2476, были обозначены как MR1506. Затем pMYC2476 трансформировали в акристаллические клетки CryB электропорацией и отбором на чашках с DM3+эритромицин (20 мкг/мл) при 30°С. Рекомбинантный CryB[pMYC2476] был обозначен как MR561.
Субкультура MR1506 была депонирована в постоянной коллекции Patent Culture Collection (NRRL), Regional Research Center, 1815 North University Street, Peoria, Illinois 61604, USA 1 июня 2000 года. Номер доступа В-30298.
В.t.-штамм MR561 испытывали на экспрессию белков бинарного токсина PS201L3 при помощи иммуноблоттинга. Клетки выращивали в жидкой среде NYS-CAA+эритромицин (10 мкг/мл) в течение ночи при 30°С. Затем культуру осаждали центрифугированием и часть клеточного осадка ресуспендировали и подвергали электрофорезу на гелях ДСН-ПААГ. Как белок 14 кДа, так и белок 44 кДа обнаруживались Вестерн-блот-анализом при зондировании антителами, специфическими для токсинов 14 кДа или 44 кДа PS149B1 соответственно.
ДНК-секвенирование генов токсинов, кодируемых pMYC2476, проводили с использованием автоматического секвенатора ABI377. Последовательность ДНК для гена 14 кДа PS201L3 показана в SEQ ID NO:133. Расшифрованная пептидная последовательность для токсина 14 кДа PS201L3 показана в SEQ ID NO:134. Последовательность ДНК для гена 44 кДа PS201L3 показана в SEQ ID NO:135. Расшифрованная пептидная последовательность для токсина 44 кДа PS201L3 показана в SEQ ID NO:136.
Следующая таблица 14 показывает сходство и идентичность последовательностей бинарных генов и белков из 201L3 и 149В1. Программу BESTFIT (часть пакета программного обеспечения GCG) использовали для этих сравнений. BESTFIT использует алгоритм локальной гомологии Smith and Waterman (Advances in Applied Mathematics 2:482-489 (1981)).
ПРИМЕР 21
Молекулярное клонирование и анализ последовательности ДНК новых генов δ-эндотоксинов из штаммов Bacillus thurinqiensis PS187G1, PS201HH2 и KR1369
Общую клеточную ДНК получали из штаммов Bacillus thuringiensis PS187G1, PS201HH2 и KR1369, выращенных до оптической плотности 0,5-1,0 при видимом свете 600 нм в бульоне Luria Bertani (LB). ДНК экстрагировали с использованием набора Qiagen Genomic-Tip 500/G и набора буферов геномной ДНК согласно протоколу для грамположительных бактерий (Qiagen Inc.; Valencia, CA). Космидные библиотеки PS187G1, PS201HH2 и KR1369 конструировали из вектора SuperCos1 (Stratagene) с использованием вставок общей клеточной ДНК PS187G1, PS201HH2 и KR1369 соответственно, частично расщепленной NdeII. Клетки XL1-Blue MR (Stratagene) трансфицировали упакованными космидами с получением клонов, устойчивых к карбенициллину и канамицину. Для каждого штамма 576 космидных колоний выращивали в 96-луночных блоках в 1 мл LB+карбенициллин (100 мкг/мл)+канамицин (50 мкг/мл) при 37°С в течение 18 часов и переносили методом реплик (отпечатков) на найлоновые фильтры для скрининга посредством гибридизации.
ПЦР-ампликон, содержащий приблизительно 1000 п.н. оперона токсинов 14 кДа и 44 кДа PS187G1, PS201HH2 или KR1369, амплифицировали из геномной ДНК PS187G1, PS201HH2 или KR1369 с использованием праймеров, сконструированных для амплификации бинарных гомологов:
15kfor1: 5'-ATG TCA GCT CGC GAA GTA CAC-3' (SEQ ID NO:131)
45krev6: 5'-GTC CAT CCC ATT AAT TGA GGA G-3' (SEQ ID NO:132)
ДНК-фрагмент очищали на геле с использованием QiaQuick экстракции (Qiagen). Зонд радиоактивно метили 32P-dCTP с использованием набора Prime-It II (Stratagene) и использовали в водном гибридизационном растворе (6Х SSPE, 5X раствор Денхардта, 0,1% ДСН, 0,1 мг/мл денатурированной ДНК) с фильтрами для подъема колоний при 65°С в течение 16 часов. Фильтры для подъема колоний промывали в течение короткого времени 1X в 0,5Х SSC/0,1% ДСН при комнатной температуре с последующими двумя дополнительными промывками в течение 10 мин при 65°С в 0,5Х SSC/0,1% ДСН. Затем фильтры экспонировали на рентгеновской пленке в течение 20 мин (PS187G1 и PS201HH2) или 1 часа (KR1369). Один космидный клон, который гибридизовался сильно с этим зондом, отбирали для дополнительного анализа для каждого штамма. Было подтверждено, что эти космидные клоны содержат приблизительно 1000 п.н. гена-мишени токсина 14 кДа и 44 кДа, согласно ПЦР-амплификации с указанными выше праймерами. Космидный клон PS187G1 был обозначен как pMYC3106; рекомбинантные клетки Е.coli XL1-Blue MR, содержащие pMYC3106, были обозначены как MR1508. Космидный клон PS201HH2 был обозначен как pMYC3107; рекомбинантные клетки Е.coli XL1-Blue MR, содержащие pMYC3107, были обозначены как MR1509. Космидный клон KR1369 был обозначен как pMYC3108; рекомбинантные клетки Е.coli XL1-Blue MR, содержащие pMYC3108, были обозначены как MR1510. Субкультуры MR1509 и MR1510 были депонированы в постоянной коллекции Patent Culture Collection (NRRL), Regional Research Center, 1815 North University Street, Peoria, Illinois 61604, USA 8 августа 2000 года. Номера доступа NRRL В-30330 и NRRL В-30331 соответственно.
Гены токсинов 14 кДа и 44 кДа PS187G1, PS201HH2 и KR1369, кодируемые pMYC3106, pMYC3107 и pMYC3108 соответственно, секвенировали с использованием автоматической системы секвенирования АВ1377 и соответствующего программного обеспечения.
Нуклеотидная и расшифрованная полипептидная последовательности 14 кДа и 44 кДа PS187G1 показаны в виде SEQ ID NO:137-140. Последовательности генов как токсина 14 кДа, так и 44 кДа являются полными открытыми рамками считывания. Нуклеотидная последовательность открытой рамки считывания токсина 14 кДа PS187G1, нуклеотидная последовательность открытой рамки считывания токсина 44 кДа и соответствующие расшифрованные аминокислотные последовательности являются новыми в сравнении с другими генами токсинов, кодирующими пестицидные белки.
Нуклеотидная и расшифрованная полипептидная последовательности 14 кДа и 44 кДа PS201HH2 показаны в виде SEQ ID NO:141-144. Последовательность гена токсина 14 кДа является полной открытой рамкой считывания. Последовательность гена токсина 44 кДа является частичной последовательностью этого гена. Нуклеотидная последовательность открытой рамки считывания токсина 14 кДа PS201HH2, нуклеотидная последовательность частичной открытой рамки считывания токсина 44 кДа и соответствующие расшифрованные аминокислотные последовательности являются новыми в сравнении с другими генами токсинов, кодирующими пестицидные белки.
Нуклеотидная и расшифрованная полипептидная последовательности 14 кДа и 44 кДа KR1369 показаны в виде SEQ ID NO:145-148. Последовательности генов как токсина 14 кДа, так и 44 кДа являются полными открытыми рамками считывания. Нуклеотидная последовательность открытой рамки считывания токсина 14 кДа KR1369, нуклеотидная последовательность открытой рамки считывания токсина 44 кДа и соответствующие расшифрованные аминокислотные последовательности являются новыми в сравнении с другими генами токсинов, кодирующими пестицидные белки.
ПРИМЕР 22
Конструирование и экспрессия гибридного генного слияния, содержащего гены бинарных токсинов PS149B1 14 кДа и 44 кДа
Были сконструированы олигонуклеотидные праймеры к 5'- и 3'-концам как гена 14 кДа, так и гена 44 кДа из PS149B1. Эти олигонуклеотиды сконструированы для создания генного слияния посредством SOE-ПЦР ("Gene Splicing By Overlap Extension: Tailor-made Genes Using PCR," Biotechniques 8:528-535, May 1990). Эти два гена сливали вместе в обратном порядке, обнаруженном в опероне нативного бинарного токсина (т.е. сначала ген 44 кДа, затем ген 14 кДа).
Последовательности олигонуклеотидов, используемых для SOE-ПЦР, были следующими:
F1new: AAATATTATTTTATGTCAGCACGTGAAGTACACATTG (SEQ ID NO:155)
R1new: tctctGGTACCttaTTAtgatttatgcccatatcgtgagg (SEQ ID NO:156)
F2new: agagaACTAGTaaaaaggagataaccATGttagatactaataaag (SEQ ID NO:157)
R2new: CGTGCTGACATAAAATAATATTTTTTTAATTTTTTTAGTGTACTTT (SEQ ID NO:158)
Олиго "F1new" был сконструирован для направления амплификации от 5'-конца гена 14 кДа и гибридизации до 3'-конца гена 44 кДа. Олиго "R1new" был сконструирован для направления амплификации от 3'-конца гена 14 кДа. Этот праймер был сконструирован с двумя стоп-кодонами для обеспечения терминации трансляции. Он был также сконструирован с KpnI-сайтом для направленного клонирования в плазмидный экспрессирующий вектор для Pseudomonas fluorescens. Олиго «F2new" был сконструирован для направления амплификации от 5'-конца гена 44 кДа. Он также включает последовательность связывания рибосом и клонирующий Spel-сайт. Олиго "R2new" был сконструирован для направления амплификации от 3'-конца гена 44 кДа и гибридизации до 5'-конца гена 14 кДа.
Эти два гена сначала независимо амплифицировали из геномной ДНК PS149B1; ген 14 кДа с использованием "F1new" и "R1new", а ген 44 кДа с использованием «F2new» и "R2new". Затем продукты комбинировали в одном цикле ПЦР и вместе амплифицировали с использованием "R1new" и "F2new". В этой точке использовали усиленную геркулазой™ полимеразную смесь (Stratagene, La Jolla, CA) при температуре отжига 48°С для амплификации ДНК-фрагмента ˜ 1,5 т.п.н., содержащего генное слияние. Этот ДНК-фрагмент затем расщепляли Kpnl и Spel, фракционировали на агарозных гелях и очищали электроэлюцией. Этот плазмидный вектор также расщепляли Kpnl и Spel, фракционировали на агарозных гелях, очищали электроэлюцией и обрабатывали фосфатазой. Затем вектор и вставку лигировали вместе в течение ночи при 14°С. Лигированные ДНК-фрагменты трансформировали в клетки Р. f. MB214 электропорацией и отбором в течение ночи на чашках с LB+тетрациклин (30 мкг/мл). Штаммы, содержащие генное слияние, идентифицировали диагностической ПЦР и секвенировали для подтверждения успешного сплайсинга генов. Один характерный штамм, содержащий клонированное генное слияние, был обозначен как MR1607; рекомбинантная плазмида была обозначена как pMYC2475.
Субкультура MR1607 была депонирована в постоянной коллекции Patent Culture Collection (NRRL), Regional Research Center, 1815 North University Street, Peoria, Illinois 61604, USA 8 августа 2000 года. Номер доступа NRRL B-30332. MR1607 выращивали, и продуцирование белка подтверждали электрофорезом в ДСН-ПААГ и иммуноблоттингом. Была идентифицирована белковая полоса при ˜58 кДа, представляющая слитый продукт 44 кДа+14 кДа, при зондировании Вестерн-блотов антителами, специфическими либо для токсина 14 кДа, либо для токсина 44 кДа.
Последовательность слитого (гибридного) белка 58 кДа представлена в SEQ ID NO:159. Последовательность ДНК для этого генного слияния представлена в SEQ ID NO:160.
ПРИМЕР 23
Исследование по смешиванию бинарных гомологов
Выращивание гомологичных штаммов
Были отобраны четыре штамма, по одному из каждого основного семейства бинарных токсинов - 149В1, 80JJ1, 201L3 и 167Н2. Для уменьшения времени, затрачиваемого для очистки индивидуальных белков токсинов, вместо этого выращивали следующие клоны Pseudomonas fluorescens (P.f.): MR1253 (14 кДа 149В1) и MR1256 (44 кДа 149В1). Подобным образом использовали клоны B.t. MR541 (экспрессирующие 14 кДа 80JJ1) и MR542 (44 кДа 80JJ1). Штаммы B.t. выращивали, как описано в примере 1. Осадки промывали ЗХ водой и хранили при -20°С до использования. Штаммы P.f. выращивали в 10-литровых резервуарах в ферментерах Biolaffite с использованием стандартных процедур. Осадки хранили при -80°С до использования.
Экстракция и очистка токсинов
Очистка 167Н2, MR541, MR542, 201L3. Экстракции клеточных осадков выполняли с использованием 100 мМ натрий-цитратного буфера при рН от 3,0 до 5,5. В типичной экстракции осадки экстрагировали объемом буфера 1/10-1/ЗХ исходного культурального объема. Осадки суспендировали в буфере и помещали на качающуюся платформу при 4°С на периоды времени от 2,5 час до ночи. Экстракты центрифугировали и супернатанты сохраняли. Эту процедуру повторяли с каждым штаммом, пока не получали по меньшей мере приблизительно 10 мг каждого белка. Электрофорез в ДСН-ПААГ подтверждал присутствие/отсутствие белковых токсинов в экстрактах посредством использования NuPAGE Bis/Tris-гель-системы (Invitrogen). Пробы получали в соответствии с инструкциями изготовителя и наносили на 4-12% гели и электрофоретограммы проявляли в рабочем буфере MES.
Исключением в отношении этой процедуры было получение всех проб 201L3. Эти пробы получали разбавлением 1/2Х буфера для проб Laemmli BioRad и нагреванием при 95°С в течение 4 минут. Определение количества белка выполняли лазерной сканирующей гель-денситометрией с БСА в качестве стандарта (Molecular Dynamics Personal Densitometer SI). Экстракты осветляли фильтрованием через мембранный фильтр 2 мкм и хранили при 4°С.
Очистка MR1253 и MR1256. Рекомбинантные белки MR1253 и MR1256, соответствующие белкам 14 кДа и 44 кДа 149В1 соответственно, готовили в виде солюбилизированных включений. Тела включения готовили с использованием стандартных процедур. Тела включения солюбилизировали в 1 мМ ЭДТА, 50 мМ цитрате натрия, рН 3,5.
Очистка индивидуальных токсинов, 167Н2 и 201L3. Все экстракты, о которых было известно, что они содержат либо 14 кДа, либо 44 кДа, либо оба белка, объединяли. Этот объединенный экстракт диализовали против 100 мМ цитрата натрия, 150 мМ NaCl, рН 4. Диализная трубка была из Pierce (Snakeskin 10k MWCO). Пробы обычно диализовали в течение приблизительно 6 часов и затем опять в течение ночи в свежем буфере.
Затем экстракты концентрировали с использованием центрифужного фильтрующего устройства Centriprep 10 или Centricon Plus-20 (Biomax -5, 5000 NMWL) (Millipore), определяли количества белков 14 кДа и 44 кДа и подвергали гель-фильтрационной хроматографии.
В подготовке для хроматографии все пробы и буферы фильтровали через фильтр 0,2 мкм и дегазировали. Затем пробы наносили на гель-фильтрационную колонку HiPrep 26/60 Sephacryl S-100, которая была уравновешена двумя колоночными объемами разделительного буфера, содержащего 100 мМ цитрат натрия, 150 мМ NaCI, pH 4,0. Объемы проб были в диапазоне 5-10 мл. Хроматография контролировалась системой жидкостной экспресс-хроматографии белков Akta purifier 100 FPLC (Amersham Pharmacia). Хроматографию проводили при температуре окружающей среды. Ток буфера через колонку во время хроматографии поддерживали при 0,7 мл/мин. Белки детектировали мониторингом УФ-поглощения при 280 нм. Фракции собирали и хранили при 4°С. Фракции, содержащие белок 14 или 44 кДа, объединяли и проверяли на чистоту при помощи электрофореза в ДСН-ПААГ, как описано выше.
Для проб 167Н2 детектировали два больших пика, которые были хорошо отделены друг от друга при линии фона. Электрофорез в ДСН-ПААГ показал, что каждый пик представлял один из белковых токсинов.
В пробе 201L3 детектировали три хорошо разделенных пика и один пик в виде плеча. Электрофорез в ДСН-ПААГ выявил, что первый пик представлял белок 100 кДа плюс белок 80 кДа. Второй пик представлял белок 44 кДа, тогда как пик-плечо был белком 40 кДа. Третий пик был белком 14 кДа. Фракции с 44 кДа из обеих проб объединяли, как и все фракции, содержащие 14 кДа.
Белки 149В1 получали индивидуально из клонов P.f. MR1253 и MR1256 и, следовательно, дополнительная очистка не была необходимой. Подобным образом рекомбинанты 80JJ1, MR541 и MR542 давали индивидуальные белки 14 и 44 кДа, устраняя тем самым необходимость дополнительной очистки.
Получение проб для биоанализа wCRW LC50
Диализ. Пробы индивидуальных белков бинарных токсинов диализовали против 6 л 20 мМ цитрата натрия, рН 4,0. Первый диализ протекал в течение нескольких часов, пробы переносили в свежий буфер и давали им диализоваться в течение ночи. Наконец, пробы переносили в свежий буфер и диализовали еще несколько часов. Источниками белковых проб были либо объединенные гель-фильтрационные фракции (167Н2, 201L3), экстракты осадков (MR541, MR542) или экстракты осадков тел включений (MR1253, Mr1256). Все пробы фильтровали через мембраны 0,2 мкм для стерилизации.
Концентрирование. Пробы концентрировали с использованием центрифужных фильтрующих устройств Centricon Plus-20 (Biomax-5, 5000 NMWL) (Millipore).
Количественные определения. В пробах определяли количества белка, как описано выше. Для удовлетворения требованиям биоанализа LC50 минимально 6,3 мг каждого белка токсина требовалось в диапазоне концентраций 0,316-1,36 мг/мл для различных комбинаций. В случае необходимости пробы концентрировали, как описано выше, или разбавляли буфером (20 мМ цитрат натрия, рН 4,0) и опять определяли количество белка.
Смешивание бинарных токсинов/биоанализ LC50. Для каждого из четырех штаммов 14 кДа объединяли с количеством 44 кДа каждого штамма с получением отношения по массе 1/1. Максимальная доза была 50 мкг/см2 для этих смесей, за исключением смесей с белком 14 кДа 203J1. Максимальные дозы смесей с этим белком были только 44 мкг/см2. Для контролей каждый белок предоставляли индивидуально, так же как буфер для экстракции, 20 мМ цитрат натрия, рН 4,0. Нативные комбинации тоже тестировали (т.е. 14 кДа+44 кДа 149В1). Все комбинации токсинов и буферные контроли оценивали три раза посредством биоанализа против западной блошки длинноусой, тогда как индивидуальные токсины тестировали один раз.
Результаты приведены ниже в таблице 15 (LC50-результаты для комбинаций токсинов) и таблице 16 (сравнение активности штаммов к 149В1).
Сравнение штаммов к 149В1
Эти результаты изображены также графически на фигуре 3.
Нативные комбинации были высокоактивными против западной блошки длинноусой, за исключением 201L3. Однако 44 кДа 201L3 был активным при комбинации с 14 кДа 167Н2 или 149В1. Другими активными комбинациями были 14 кДа 149В1 с 44 кДа 80JJ1 или 167Н2, причем последняя комбинация является более активной, чем нативная смесь 149В1. Не было зависимости реакции от дозы ни для индивидуальных белков, ни для буферных и водных контролей.
ПРИМЕР 24
Борьба с южной блошкой длинноусой с использованием белка 14 кДа PS149B1
Порошок, содержащий приблизительно 50% (мас./мас.) δ-эндотоксина 14 кДа, первоначально обнаруженного в штамме Bacillus thuringiensis PS149B1, выделяли из рекомбинантного штамма Pseudomonas fluorescens (MR1253). Этот порошок оценивали на инсектицидную активность с использованием следующей процедуры.
Искусственный рацион для насекомых (R.I. Rose and J.M. McCabe (1973), "Laboratory rearing techniques for rearing corn rootworm," J. Econ. Entomol. 66(2): 398-400) распределяли при ˜0,5 мл на лунку в 128-луночные планшеты (лотки) для биоанализа (C-D International, Pitman, NJ) для получения поверхностных зон ˜1,5 см2. Суспензии в буфере (10 мМ калий-фосфат, рН 7,5) порошка белка 14 кДа наносили на поверхность искусственного рациона для насекомых при 50 мкл на лунку и поверхности рациона давали высохнуть. Буферные контроли также включали в каждый тест. Отдельных новорожденных личинок блошки длинноусой, Diabrotica undecimpunctata howardi помещали в каждую лунку и лунки герметизировали крышками, которые поставлялись с планшетами (лотками). Биотесты выдерживали в течение 6 дней при 28°С, после чего живых личинок взвешивали в виде группы для каждого варианта обработки. Процент ингибирования роста рассчитывали вычитанием веса живых насекомых из каждого варианта обработки из веса живых контрольных насекомых и затем делением на вес контролей. Этот результат умножали на 100 для превращения этого числа в процент. Ингибирование роста рассчитывали для каждого из 5 тестов, каждый из которых содержал 16 насекомых на вариант обработки, и рассчитывали среднюю величину ингибирования роста по всем тестам.
Результаты показали, что белок 14 кДа ингибировал рост южных блошек длинноусых зависимым от концентрации образом.
Таблица 17 показывает ингибирование роста южной блошки длинноусой белком 14 кДа PS149B1.
ПРИМЕР 25
Борьба с европейским мотыльком кукурузным и совкой кукурузной с использованием бинарного токсина PS149B1
Порошок, содержащей 54% δ-эндотоксина 14 кДа, и другой порошок, содержащий 37% δ-эндотоксина 44 кДа, оба из которых были первоначально обнаружены в штамме Bacillus thuringiensis PS149B1, выделяли из рекомбинантных штаммов Pseudomonas fluorescens MR1253 и MR1256 соответственно. Смеси этих порошков оценивали на инсектицидную активность с использованием следующей процедуры.
Искусственный рацион для насекомых (R.I. Rose and J.M. McCabe (1973), "Laboratory rearing techniques for rearing corn rootworm," J. Econ. Entomol. 66(2): 398-400) распределяли при ˜0,5 мл на лунку в 128-луночные планшеты (лотки) для биоанализа (C-D International, Pitman, NJ) для получения поверхностных зон ˜1,5 см2. Суспензии в буфере (10 мМ калий-фосфат, рН 7,5) порошков белков смешивали и затем наносили на поверхность искусственного рациона для насекомых при 50 мкл на лунку. Поверхности рациона давали высохнуть. Буферные контроли также включали в каждый тест. Отдельных новорожденных личинок помещали в каждую лунку и лунки герметизировали крышками, которые поставлялись с планшетами (лотками). Тесты проводили с европейским мотыльком кукурузным, Ostrinia nubilalis, и совкой кукурузной, Helicoverpa zea (оба насекомых являются чешуекрылыми). Биотесты выдерживали в течение 6 дней при 28°С, после чего живых личинок взвешивали в виде группы для каждого варианта обработки. Процент ингибирования роста рассчитывали вычитанием веса живых насекомых из каждого варианта обработки из веса живых контрольных насекомых и затем делением на вес контролей. Этот результат умножали на 100 для превращения этого числа в процент. Ингибирование роста рассчитывали для каждого из 4 тестов, каждый из которых содержал 14-16 насекомых на вариант обработки, и рассчитывали среднюю величину ингибирования роста по всем тестам.
Результаты показали, что белок 14 кДа ингибировал рост европейского мотылька кукурузного и совки кукурузной зависимым от концентрации образом. Таблица 18 показывает ингибирование роста совки кукурузной (CEW) и европейского мотылька кукурузного (ЕСВ) смесями белков PS149B1.
ПРИМЕР 26
Дополнительная характеристика белков 45 кДа и конструирование праймера для идентификации дополнительных полинуклеотидов и белков
Данное изобретение включает не только конкретные приведенные в качестве примеров последовательности. Части рассматриваемых генов и токсинов могут быть использованы для идентификации других родственных генов и токсинов. Таким образом, данное изобретение включает полинуклеотиды, которые кодируют белки или полипептиды, содержащие по меньшей мере десять смежных аминокислот, например любых белков или полипептидов бинарного типа, включенные в прилагаемый список последовательностей и описанные здесь. Другие варианты включают полинуклеотиды, которые кодируют, например, по меньшей мере 20, 30, 40, 50, 60, 70, 80, 90 и 100 смежных аминокислот приведенного здесь в качестве примера белка; эти цифры относятся также подобным образом к смежным нуклеотидам приведенного в качестве примера полинуклеотида. Белки, кодируемые такими полинуклеотидами, включены в данное изобретение. Подобным образом полинуклеотиды, содержащие смежные нуклеотиды (которые кодируют белки или полипептиды, содержащие пептиды этих приблизительных размеров), включены в данное изобретение.
Считается, что, хотя все еще очень отличающиеся, "ближайшими" токсинами относительно токсинов данного изобретения являются москитоцидные (убивающие комаров) белки 51 и 42 кДа Bacillus sphaericus. В прилагаемых фигурах 4 и 5 показаны сопоставления белков и нуклеотидных последовательностей токсинов и генов 51 и 42 кДа sphaericus и токсина и гена 45 кДа 149В1.
Два блока последовательностей выделены в нуклеотидном сопоставлении, к которым могли бы быть изготовлены праймеры. Пример пары ПЦР-праймеров приведен ниже и в ориентации 5'-3' (45kD3'rc показан в качестве комплемента). Эти праймеры были успешно использованы для идентификации дополнительных членов бинарного семейства 45 кДа. Ниже включены также полностью избыточные последовательности и предсказанная пара.
45kD5': GAT RAT RAT CAA TAT ATT ATT AC (SEQ ID NO:161)
45kD3'rc: CAA GGT ART AAT GTC CAT CC (SEQ ID NO:162)
Эти последовательности могли бы использоваться в том виде, в каком они написаны, и в качестве обратного комплемента (03 и 04 являются комплементарными 45kD3'rc, приведенному в качестве примера обратному праймеру).
45kD5'01: GAT GATGrTmrAk wwATTATTrC A (SEQ ID NO:163).
45kD5'02: GAT GATGrTmrAT ATATTATTrC A (SEQ ID NO:164).
45kD3'03: GGAwG krCdyTwdTm CCwTGTAT (SEQ ID NO:165).
45kD3'04: GGAwG kACryTAdTA CCTTGTAT (SEQ ID NO:166).
Что касается способа, которым были идентифицированы токсины sphaericus, поиск в базе данных BLAST (Altschul et al. (1997), "Gapped BLAST and PSI-BLAST: a new generation of protein database search programs," Nucleic Acids Res. 25:3389-3402) с использованием белка 45 кДа 149В1 обнаружил совпадения с кристаллическим белком включения 42 кДа В. sphaericus (ожидаемая оценка 3·10-14) и кристаллическим белком включения 51 кДа В. sphaericus (ожидаемая оценка 3·10-9).
Сопоставление пептидной последовательности 45 кДа 149В1 с кристаллическим белком включения 42 кДа В. sphaericus привело к сопоставлению, имеющему 26% идентичность на протяжении 325 остатков. Оценка сопоставления имеет 27,2 sd (стандартных отклонений) выше средней оценки 100 случайных сопоставлений. Подобный анализ пептидной последовательности 45 кДа 149В1 с кристаллическим белком включения 42 кДа В. sphaericus привел к сопоставлению, имеющему 29% идентичность на протяжении 229 остатков. Оценка сопоставления имеет 23,4 sd выше средней оценки 100 случайных сопоставлений. Оценки сопоставлений >10 sd выше среднего случайных сопоставлений рассматриваются как значимые (Lipman, D.J. and Pearson, W.R. (1985), "Rapid and sensitive similarity searches," Science 227:1435-1441; Doolittle, R.F. (1987), Of URFs and ORFs: a primer on how to analyze derived amino acid sequences, University Science Books, Mill Valley, CA).
Для ссылки структурно сходные белковые последовательности Cry1Aa, Cry2Aa и Сгу3Аа сравнивали таким же образом. Сry2Аа против Cry1Aa и Cry2Aa против Сry3Аа имеют 29% и 27% идентичность на протяжении 214 и 213 остатков соответственно с оценками сопоставления 32,2 sd и 29,5 sd выше средней оценки 100 случайных сопоставлений. Сопоставление последовательности белка 45 кДа 149В1 и последовательности белка Cry2Aa привело к оценке сопоставления в пределах 1 sd среднего из 100 случайных сопоставлений.
Указываются также следующие сравнения:
Для целей дополнительного сравнения и для дополнительного конструирования праймеров указываются следующие ссылки:
Oei et al. (1992), "Binding of purified Bacillus sphaericus binary toxin and its deletion derivatives to Culex quinquefasciatus gut: elucidation of functional binding domains," Journal of General Microbiology 138(7): 1515-26.
Для 51 кДа: 35-448 является активным; 45-448 является неактивным; 4-396 является активным; 4-392 является неактивным.
Для 42 кДа: 18-370 является активным; 35-370 является неактивным; 4-358 является активным; 4-349 является неактивным.
Эта работа была проведена с GST-слияниями, очищенными и расщепленными тромбином. Все укороченные варианты анализировали с=другой интактной субъединицы. Все делеции имели некоторую потерю активности. Р51дельтаС56 связывается, но не интернализует 42. Р51дельтаN45 не связывается. Только 42 кДа+51 кДа интернализуются. Как N-концевые, так и С-концевые нетоксичные белки 42 кДа не могут связывать белок 51 кДа или комплекс белок 51 кДа-рецептор.
Davidson et al. (1990), "Interaction of the Bacillus sphaericus mosquito larvicidal proteins," Can. J. Microbiol. 36 (12):870-8. N-концы очищенных электрофорезом в ДСН-ПААГ белков, полученные из В. sphaericus. S29 и N31 51 кДа и S9 42 кДа в комплексах 68-74 кДа (невосстановленные). S9 и S29 51 и N31 42 из полосы 51 кДа (невосстановленные). В восстановленных гелях полоса 45 кДа имела S29 и N31 51 кДа и полоса 39 кДа содержала S9 белка 42 кДа.
Baumann et al., (1988), "Sequence analysis of the mosquitocidal toxin genes encoding 51,4- and 41,9-kilodalton proteins from Bacillus sphaericus 2362 and 2297," J. Bacteriol. 17:2045-2050. N-концы 41,9 кДа при D5 из протеазы В. sphaericus и I11 из химотрипсина; С-конец после R349 с трипсином. Были идентифицированы участки повышенного сходства, которые соответствуют многим из вышеуказанных участков. Сходные блоки последовательности A-D между белками 51 и 42 кДа.
В целом, токсины sphaericus, обсуждаемые выше, не включены в рамки данного изобретения (фактически, они особо исключаются). С учетом этого различающиеся непрерывные последовательности, приведенные в качестве примеров в обсуждаемых выше сопоставлениях, могут быть использованы в качестве праймеров для идентификации уникальных токсинов, которые предполагаются, но не приводятся в качестве конкретных примеров здесь. Однако консервативные смежные последовательности, показанные в этих сопоставлениях, могут быть также использованы в соответствии с данным изобретением для идентификации дополнительных новых бинарных токсинов типа 15/45 кДа (активных против блошки длинноусой и других вредителей).
ПРИМЕР 27
Встраивание и экспрессия генов токсинов в растениях
Одним аспектом данного изобретения является трансформация растений полинуклеотидами данного изобретения, которые экспрессируют белки данного изобретения. Трансформированные растения являются устойчивыми к нападению вредителя-мишени.
Новые активные против блошки длинноусой гены, описанные здесь, могут быть оптимизированы для экспрессии в других организмах. Например, оптимизированные для кукурузы последовательности генов, кодирующие токсины 14 и 44 кДа PS80JJ1, представлены в SEQ ID NO:44 и 45 соответственно.
Гены, кодирующие пестицидные токсины, описанные здесь, могут быть встроены в клетки растений с использованием разнообразных способов, которые хорошо известны в данной области. Например, большое число клонирующих векторов, содержащих систему репликации в Е.coli и маркер, позволяющий отбирать трансформированные клетки, являются доступными для подготовки для встраивания чужеродных генов в высшие растения. Эти векторы включают, например, pBR322, серию pUC, серию М13 mp, pACYC184 и т.д. Таким образом, последовательность, кодирующая токсин B.t., может быть встроена в вектор в подходящем сайте рестрикции. Полученную плазмиду используют для трансформации в Е.coli. Клетки Е.coli культивируют в подходящей питательной среде, затем собирают и лизируют. Плазмиду извлекают. Анализ последовательности, рестрикционный анализ, электрофорез и другие биохимические-молекулярно-биологические способы обычно используют в качестве способов анализа. После каждой манипуляции используемая последовательность ДНК может быть расщеплена и присоединена к следующей последовательности ДНК. Каждая плазмидная последовательность может быть клонирована в те же самые или другие плазмиды. В зависимости от способа встраивания желательных генов в растение, могут быть необходимы другие последовательности ДНК. Если, например, для трансформации клетки растения используют Ti- или Ri-плазмиду, то по меньшей мере одна правая граница, но часто правая граница и левая граница Т-ДНК Ti- или Ri-плазмиды должна быть присоединена в качестве фланкирующего участка встраиваемого гена.
Применение Т-ДНК для трансформации клеток растений интенсивно исследовалось и достаточно описано в ЕР 120516; Hoekema (1985) In: The Binary Plant Vector System, Offsetdurkkerij Kanters B.V., Alblasserdam, Chapter 5; Fraley et al., Crit. Rev.Plant Sci. 4:1-46; and An et al. (1985) EMBO J. 4:277-287.
После интеграции встроенной ДНК в геном она является относительно стабильной в нем и, как правило, не выходит снова из него. Обычно она содержит селективный маркер, который придает трансформированному растению устойчивость к биоциду или антибиотику, такому как канамицин, G 418, блеомицин, гигромицин или хлорамфеникол, inter alia. Индивидуально применяемый маркер должен, таким образом, делать возможным отбор трансформированных клеток, а не клеток, которые не содержат встроенной ДНК.
Большое число способов являются доступными для встраивания ДНК в растительную клетку-хозяина. Эти способы включают в себя трансформацию Т-ДНК с использованием Agrobacterium tumefaciens или Agrobacterium rhizogenes в качестве трансформирующего агента, слияние, инъекцию, биолистику (бомбардировку микрочастицами) или электропорацию, а также другие возможные способы. Если Agrobacteria используют для трансформации, то встраиваемая ДНК должна быть клонирована в специальные плазмиды, а именно в промежуточный вектор или в бинарный вектор. Промежуточные векторы могут быть интегрированы в Ti- или Ri-плазмиду гомологичной рекомбинацией благодаря последовательностям, которые являются гомологичными с последовательностями Т-ДНК. Ti- или Ri-плазмида содержит также участок vir, необходимый для переноса Т-ДНК. Промежуточные векторы не могут самореплицироваться в Agrobacteria. Промежуточный вектор может быть перенесен в Agrobacterium tumefaciens посредством хелперной плазмиды (конъюгации). Бинарные векторы могут самореплицироваться как в Е.coli, так и в Agrobacteria. Они содержат ген селективного маркера и линкер или полилинкер, который обрамлен правым и левым пограничными участками Т-ДНК. Они могут быть трансформированы непосредственно в Agrobacteria (Holsters et al. [1978] Mol. Gen. Genet. 163:181-187). Agrobacterium, используемая в качестве клетки-хозяина, должна содержать плазмиду, несущую участок vir. Участок vir необходим для переноса Т-ДНК в клетку растения. Может содержаться дополнительная Т-ДНК. Трансформированную таким образом бактерию используют для трансформации клеток растений. Эксплантаты растений могут предпочтительно культивироваться с Agrobacterium tumefaciens или Agrobacterium rhizogenes для переноса ДНК в клетку растения. Затем могут быть регенерированы целые растения из инфицированного растительного материала (например, кусочков листа, сегментов стебля, корней, но также протопластов или суспензионно-культивируемых клеток) в подходящей среде, которая может содержать антибиотики или биоциды для отбора. Затем полученные таким образом растения могут быть тестированы на присутствие встроенной ДНК. Нет особых требований в отношении плазмид в случае инъекции или электропорации. Возможно использование обычных плазмид, таких как, например, производных pUC.
Трансформированные клетки растут в растениях обычным образом. Они могут образовать зародышевые клетки и передавать трансформированный признак (признаки) потомкам растения. Такие растения могут выращиваться обычным путем и скрещиваться с растениями, которые имеют те же самые трансформированные наследственные факторы или другие наследственные факторы. Полученные гибридные индивидуумы имеют соответствующие фенотипические свойства.
В предпочтительном варианте данного изобретения растения будут трансформироваться генами, в которых использование кодонов было оптимизировано для растений. См., например, патент США №5380831, который включен здесь в качестве ссылки. Также предпочтительно будут использоваться растения, кодирующие укороченный токсин. Укороченный токсин обычно будет кодировать приблизительно 55% - приблизительно 80% полноразмерного токсина. Способы создания синтетических генов B.t. для применения в растениях известны в данной области.
ПРИМЕР 28
Клонирование генов B.t. в вирусы насекомых
Известен ряд вирусов, которые инфицируют насекомых. Эти вирусы включают, например, бакуловирусы и энтомопоксвирусы. В одном варианте данного изобретения гены, кодирующие инсектицидные токсины, описанные здесь, могут быть помещены в геном вируса насекомого с усилением тем самым патогенности вируса. Способы конструирования вирусов насекомых, содержащих гены токсинов B.t., хорошо известны и легко практикуются специалистами в данной области. Эти процедуры описаны, например, в Merryweather et al. (Merryweather, А.Т., U. Weyer, M.P.G.Harris, M.Hirst, T.Booth, R.D.Possee (1990) J.Gen. Virol. 71:1535-1544) и Martens et al. (Martens, J.W.M., G.Honee, D.Zuidema, J.W.M.van Lent, B. Visser, J.M.Vlak (1990) Appl. Environmental Microbiol. 56(9):2764-2770).
Все патенты, патентные заявки, предварительные заявки и публикации, на которые ссылаются или которые цитируются здесь, включены ссылкой во всей полноте до степени, в которой они не являются несовместимыми с определенными указаниями данного описания.
Должно быть понятно, что описанные здесь примеры и варианты приведены только для иллюстративных целей и что различные модификации или изменения в свете этих примеров и вариантов будут предполагаться как очевидные для специалистов в данной области и они должны быть включены в рамки идеи и сферы действия данной заявки и объема прилагаемой формулы изобретения.




























































































| название | год | авторы | номер документа |
|---|---|---|---|
| ИНСЕКТИЦИДНЫЕ БЕЛКИ, ВЫДЕЛЕННЫЕ ИЗ ВИДОВ БАКТЕРИЙ Bacillus, И ИХ ПРИМЕНЕНИЕ | 2004 |
|
RU2382822C2 |
| НОВЫЕ ПЕСТИЦИДНЫЕ ПРОТЕИНЫ И ШТАММЫ | 1995 |
|
RU2196824C2 |
| КОМБИНАЦИИ, ВКЛЮЧАЮЩИЕ БЕЛКИ Cry34Ab/35Ab И Сry3Ba, ДЛЯ ПРЕДОТВРАЩЕНИЯ РАЗВИТИЯ УСТОЙЧИВОСТИ У КУКУРУЗНЫХ КОРНЕВЫХ ЖУКОВ (Diabrotica spp.) | 2011 |
|
RU2591519C2 |
| ИНСЕКТИЦИДНЫЕ CRY-ТОКСИНЫ DIG-3 | 2010 |
|
RU2590708C2 |
| КОМБИНАЦИИ, ВКЛЮЧАЮЩИЕ БЕЛКИ Cry34Ab/35Ab И Cry3Aa, ДЛЯ ПРЕДОТВРАЩЕНИЯ РАЗВИТИЯ УСТОЙЧИВОСТИ У КУКУРУЗНЫХ КОРНЕВЫХ ЖУКОВ (Diabrotica spp.) | 2011 |
|
RU2582249C2 |
| КОМБИНАЦИИ, ВКЛЮЧАЮЩИЕ БЕЛКИ Cry34Ab/35Ab И Cry6Aa, ДЛЯ ПРЕДОТВРАЩЕНИЯ РАЗВИТИЯ УСТОЙЧИВОСТИ У КУКУРУЗНЫХ КОРНЕВЫХ ЖУКОВ (Diabrotica spp. ) | 2011 |
|
RU2582262C2 |
| КОМБИНАЦИИ, ВКЛЮЧАЮЩИЕ БЕЛКИ Cry3Aa И Cry6Aa, ДЛЯ ПРЕДОТВРАЩЕНИЯ РАЗВИТИЯ УСТОЙЧИВОСТИ У КУКУРУЗНЫХ КОРНЕВЫХ ЖУКОВ (Diabrotica spp.) | 2011 |
|
RU2576005C2 |
| СПОСОБЫ БОРЬБЫ С ВРЕДИТЕЛЕМ - СОВКОЙ AGROTIS | 2000 |
|
RU2311767C2 |
| КОМБИНИРОВАННОЕ ИСПОЛЬЗОВАНИЕ БЕЛКОВ CRY1Ca И CRY1Ab ДЛЯ КОНТРОЛЯ УСТОЙЧИВОСТИ НАСЕКОМЫХ | 2010 |
|
RU2596406C2 |
| НОВЫЕ БЕЛКИ, ПРОЯВЛЯЮЩИЕ ИНГИБИРУЮЩУЮ АКТИВНОСТЬ ПО ОТНОШЕНИЮ К НАСЕКОМЫМ | 2016 |
|
RU2765310C2 |


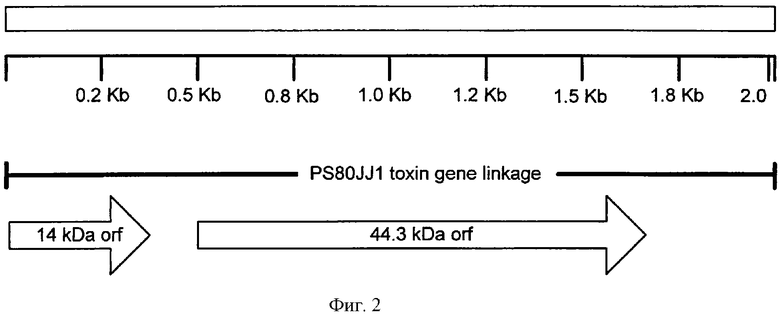
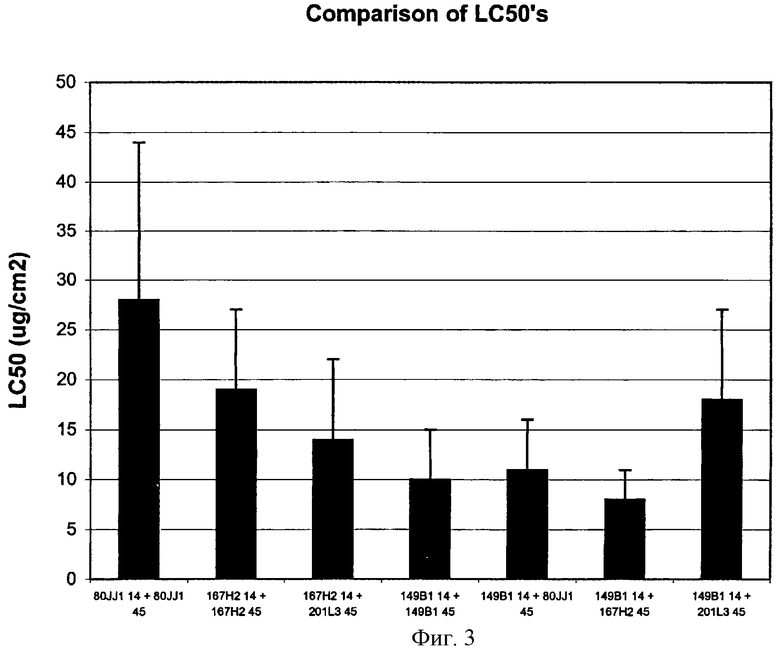
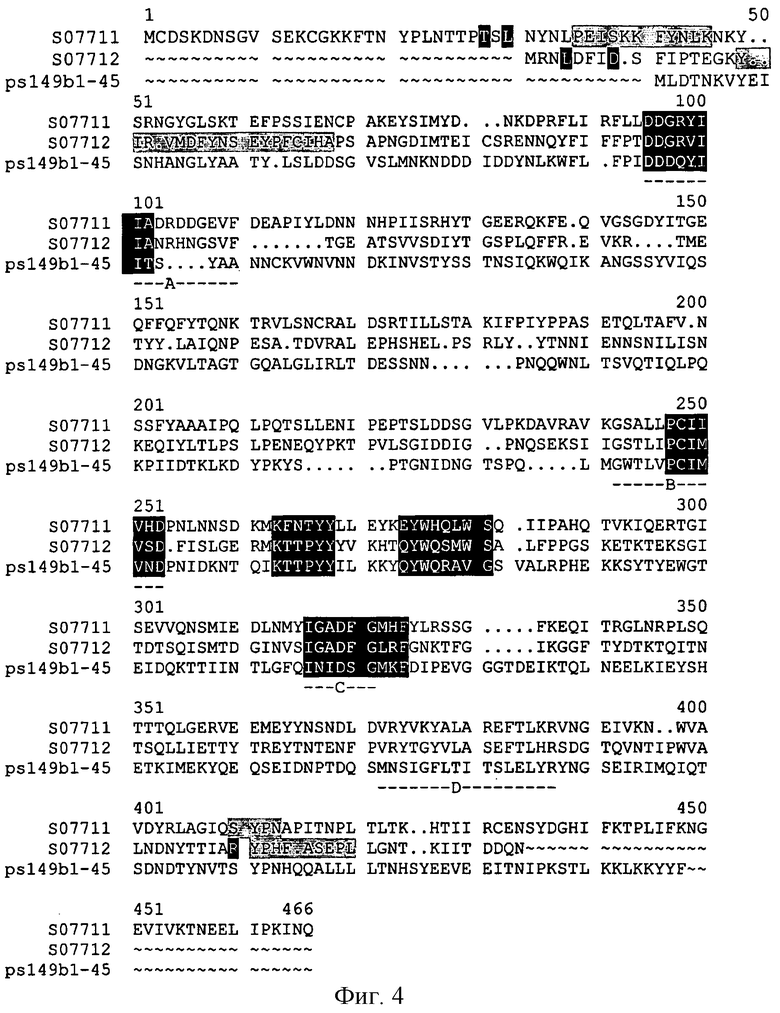



Данное изобретение относится к биотехнологии, в частности к новым классам белков и полинуклеотидным последовательностям, обладающим пестицидной активностью, а также к способу борьбы с вредителями. Выделяют полинуклеотид, который обладает токсической для жесткокрылого насекомого активностью, или сам белок, обладающий данной токсичностью (нуклеотидные и аминокислотные последовательности указаны в материалах заявки). Борьбу с жесткокрылыми насекомыми проводят путем контактирования вредителя с вышеуказанным белком. Изобретение позволяет повысить эффективность борьбы с сельскохозяйственными вредителями группы жесткокрылых и тем самым снизить производственные затраты и повысить сбор сельскохозяйственной продукции. 4 н. и 17 з.п. ф-лы, 5 ил., 20 табл.
| WO 9740162 30.10.1997 | |||
| US 4797276 10.01.1989 | |||
| ШТАММ BACILLUS THURINGIENSIS - ПРОДУЦЕНТ ЭНДОТОКСИНА ПРОТИВ ЖЕСТКОКРЫЛЫХ НАСЕКОМЫХ И СПОСОБ ЕГО КУЛЬТИВИРОВАНИЯ | 1995 |
|
RU2108384C1 |

