Уровень изобретения
Люди выращивают кукурузу для использования в качестве пищи и источника энергии. Люди также выращивают хлопок и множество других зерновых культур, включающих в себя сою. Насекомые поедают и повреждают растения и таким образом подрывают усилия человека. Ежегодно затрачиваются миллиарды долларов для контроля насекомых-вредителей, где ущерб, который они наносят, исчисляется дополнительными миллиардами долларов. Для контроля насекомых-вредителей применяются в первую очередь синтетические органические химические инсектициды, вместе с тем, в некоторых сферах важную роль играют биологические инсектициды, такие как инсектицидные белки, полученные из Bacillus thuringiensis (Bt). Способность производить устойчивые к насекомым растения посредством их трансформации генами инсектицидного белка Bt представляет собой революционное изменение в современном сельском хозяйстве и повышает важность и значение инсектицидных белков и их генов.
Для создания устойчивых к насекомым трансгенных растений использовались несколько Bt-белков, которые в настоящее время успешно зарегистрированы и введены в коммерческое обращение. Они включают в себя белки кукурузы Cry1Ab, Cry1Ac, Cry1Fa и Cry3Bb, белки хлопка Cry1Ac и Cry2Ab и картофельный белок Cry3A.
Экспрессирующие указанные белки коммерческие продукты экспрессируют единственный белок, кроме тех случаев, когда желательно объединить инсектицидный спектр 2 белков (например, Cry1Ab и Cry3Bb в кукурузе объединяют для обеспечения устойчивости к чешуекрылым вредителям и к личинкам, повреждающим корни, соответственно), или если независимое действие белков делает их полезными в качестве инструмента для задержки развития устойчивости у восприимчивых популяций насекомых (например, объединяют белки хлопка Cry1Ac и Cry2Ab для обеспечения контроля устойчивости к листовертке-почкоеду табака).
В этой связи, некоторые из качеств устойчивых к насекомым трансгенных растений, которые привели к быстрому и широко распространенному принятию этой технологии, также вызывают обеспокоенность возможностью развития у популяций вредителей устойчивости к инсектицидным белкам, которые продуцируются этими растениями.
Был предложен ряд стратегий для сохранения полезности свойств устойчивости к насекомым, обусловленной Bt белками, которые включают в себя применение белков в высоких дозах, применение в комбинации с «убежищами», и совместное применение с различными токсинами или повреждение этими токсинами (McGaughey et al. (1998), "B.t. Resistance Management", Nature Biotechnol. 16: 144-146).
Белки, выбранные для использования с целью управления устойчивостью насекомых (IRM), в совокупности должны проявлять свой инсектицидный эффект независимо таким образом, чтобы устойчивость, развивающаяся к одному белку, не вызывала развитие устойчивости ко второму белку (то есть, чтобы отсутствовала перекрестная устойчивость к этим белкам). Если, например, популяция вредителя, выбранная по устойчивости к "белку А", восприимчива к "белку B", можно сделать вывод об отсутствии перекрестной устойчивости, и что комбинация белка A и белка B будет эффективно задерживать развитие устойчивости к белку А единственному.
При отсутствии устойчивых популяций насекомых можно проводить оценку на основе других характеристик, которые, вероятно, связаны с механизмом действия и интенсивностью перекрестной устойчивости. Было предложено применение рецептор-опосредованного связывания для идентификации инсектицидных белков с вероятным отсутствием перекрестной устойчивости (van Mellaert et al. 1999). Ключевым фактом для прогноза отсутствия перекрестной устойчивости, который лежит в основе упомянутого подхода, является то, что инсектицидные белки не конкурируют за рецепторы у восприимчивых видов насекомых.
В случае, если два токсина B.t. Cry конкурируют за один и тот же рецептор, и впоследствии у насекомого происходит определенная мутация рецептора, в результате которой один из токсинов больше не связывается с этим рецептором и поэтому теряет инсектицидное действие против этого насекомого, то у этого насекомого может также возникать устойчивость ко второму токсину (который конкурентно связывается с этим рецептором). Вместе с тем, если два токсина связываются с двумя разными рецепторами, это может служить признаком отсутствия одновременной устойчивости насекомого к двум упомянутым токсинам.
Для защиты растений от множества насекомых-вредителей в настоящее время применяется Cry1Ab, представляющий собой инсектицидный белок, используемый в трансгенной кукурузе. Белок Cry1Ab обеспечивает защиту от основного вредителя кукурузы, которым является европейский кукурузный мотылек.
Дополнительные токсины Cry можно найти в перечне на вебсайте комитета по официальной номенклатуре B.t (Crickmore et al.; lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/). См. Приложение A к указанному источнику. В настоящее время существует около 60 основных групп токсинов "Cry" (Cry1-Cry59), с дополнительными токсинами Cyt, токсинами VIP и подобными токсинами. Многие из этих многочисленных групп имеют подгруппы, обозначаемые прописными буквами, и в подгруппах с прописными буквами выделяют подподгруппы, обозначаемые строчными буквами. (Например, Cry1 имеет подгруппы A-L, и в подгруппе Cry1A выделяют подподгруппы а-i).
Краткая сущность изобретения
Настоящее изобретение частично относится к неожиданному открытию, что Cry1Ca обладает высокой активностью против популяции огневки сахарного тростника, включающей в себя популяцию устойчивой к Cry1Ab огневки сахарного тростника. Специалистам в данной области техники будет очевидно, в плане преимущества настоящего раскрытия, что растения, продуцирующие Cry1Ca и Cry1Ab (включающие в себя инсектицидные участки указанных белков), будут полезными для замедления или предотвращения развития устойчивости к любому одному из указанных инсектицидных белков. Например, ген cry1Fa также может состоять из этих генов/белков из двух оснований.
Настоящее изобретение также относится к открытию, что Cry1Ca и Cry1Ab не конкурируют друг с другом за связывание с рецепторами кишечника кукурузной листовой совки (Spodoptera frugiperda; FAW).
КРАТКОЕ ОПИСАНИЕ ФИГУР
Фиг.1 показывает конкурентное связывание корового токсина Cry1Ab, корового токсина Cry1Ca и корового токсина Cry1Ab, меченного 125I, с мембранными везикулами щеточной каймы (BBMV) Spodoptera frugiperda.
Фиг.2 показывает конкурентное связывание корового токсина Cry1Ca, корового токсина Cry1Ab и корового токсина Cry1Ab, меченного 125I, с BBMV Spodoptera frugiperda.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO:1 показывает коровый Cry1Ca/протоксин Cry1Ab химерный белок 1164 aa (DIG-152),
SEQ ID NO:2 показывает коровый токсин Cry1Ca,
SEQ ID NO:3 показывает коровый токсин Cry1Ab.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение частично относится к неожиданному открытию, что Cry1Ca обладает высокой активностью против популяции огневки сахарного тростника (SCB; Diatraea saccharalis), которая устойчива к Cry1Ab. Соответственно, настоящее изобретение частично относится к неожиданному открытию, что Cry1Ca можно использовать в комбинации или в "комплекте" с Cry1Ab для борьбы с развитием устойчивости к любому одному из упомянутых инсектицидных белков. Иначе говоря, настоящее изобретение частично относится к неожиданному открытию, что популяция огневки сахарного тростника, выбранная по устойчивости к Cry1Ab, не обладает устойчивостью к Cry1Ca; популяция огневки сахарного тростника с устойчивостью к токсину Cry1Ab является восприимчивой (то есть, не проявляет перекрестной устойчивости) к Cry1Ca. Таким образом, настоящее изобретение включает в себя использование токсина Cry1Ca для контроля популяций огневки сахарного тростника, которые являются устойчивыми к Cry1Ab.
Специалистам в данной области техники будет очевидно, в плане преимущества настоящего раскрытия, что растения, продуцирующие Cry1Ca и Cry1Ab (включающие в себя инсектицидные участки указанных белков), будут полезными для замедления или предотвращения развития устойчивости к любому одному из указанных инсектицидных белков.
Настоящее изобретение включает в себя использование Cry1Ca для защиты сахарного тростника и других экономически важных видов растений от повреждения и потери урожая, вызванных огневкой сахарного тростника или популяциями огневки сахарного тростника, которые стали устойчивыми к Cry1Ab. Огневка сахарного тростника может также быть вредителем кукурузы. Это особенно актуально для некоторых стран Центральной и Южной Америки, например, Бразилии и Аргентины. Таким образом, согласно настоящему изобретению также можно защищать, например, кукурузу.
Настоящее изобретение, таким образом, описывает совокупность мероприятий по управлению устойчивостью насекомых (IRM) для предотвращения или уменьшения развития устойчивости к Cry1Ab и/или Cry1Ca у огневки сахарного тростника.
Дополнительно, исследования связывания рецептора с помощью радиомеченого Cry1Ca и ткани насекомого Spodoptera frugipera; кукурузной листовой совки (FAW), показали, что Cry1Ab не конкурирует за участок высокоаффинного связывания, с которым связывается Cry1Ca. Эти результаты указывают, что комбинацию Cry1Ab и Cry1Ca можно использовать как эффективный способ снижения развития устойчивости в популяциях насекомого (таких как FAW и SCB) к белкам Cry1Ab и/или Cry1Ca для растений (таких как кукуруза и сахарный тростник), продуцирующих оба белка. Совместные исследования токсина показали, что белок Cry1Ca связывается с двумя белками в BBMV у S.frugiperda, один из которых имеет молекулярную массу 40 кДа и другой 44 кДа, где белок Cry1Ab связывается с единственным белком 150 кДа (Aranda et al., 1996) и его не задействовали в исследованиях неконкурентного связывания.
Таким образом, настоящее изобретение также включает в себя комбинацию Cry1Ca и Cry1Ab в качестве совокупности мероприятий IRM по снижению развития устойчивости кукурузной листовой совки и/или огневки сахарного тростника к какому-либо белку, или по борьбе с устойчивостью популяций огневки сахарного тростника, проявляемой к Cry1Ab.
Настоящее изобретение относится к следующему: композициям для контроля чешуекрылых вредителей, где указанные композиции содержат клетки, которые экспрессируют коровый токсин-содержащий белок Cry1Ca и коровый токсин-содержащий белок Cry1Ab;
к клеткам-хозяевам, трансформированным для экспрессии обоих белков: корового токсин-содержащего белка Cry1Ab и корового токсин-содержащего белка Cry1C, где указанный хозяин представляет собой клетку микроорганизма или растения (полунуклеотидный субъект (субъекты) предпочтительно находятся в генетической конструкции под контролем промотора, не происходящего из Bacillus thuringiensis (функционально связанного с ним/содержащими его); рассматриваемые полинуклеотиды могут содержать кодон, используемый для усиления экспрессии в растении);
к способу контроля чешуекрылых вредителей, содержащему контакт указанных вредителей или окружающей среды указанных вредителей с эффективным количеством композиции, которая продуцирует коровый токсин-содержащий белок Cry1Ab, и к клетке, экспрессирующей коровый токсин-содержащий белок Cry1C;
к растению (такому как, например, кукуруза, или соя, или хлопок, или сахарный тростник), которое содержит ДНК, кодирующую коровый токсин-содержащий белок Cry1Ca, и ДНК, кодирующую коровый токсин-содержащий белок Cry1Ab; и к семенам такого растения;
к растению (такому как, например, кукуруза, или соя, или хлопок, или сахарный тростник), где в указанное растение кукурузу была вставлена ДНК, кодирующая коровый токсин-содержащий белок Cry1Ca, и ДНК, кодирующая коровый токсин-содержащий белок Cry1Ab; и к семенам такого растения.
Авторы изобретения в биотестах с искусственной питательной средой продемонстрировали, например, что Cry1Ca (белок из рекомбинантного штамма Pseudomonas fluorescens MR1206/DC639; плазмида pMYC2547), обладает высокой эффективностью для контроля популяций огневки сахарного тростника (SCB; Diatraea saccharalis), которые были выбраны по устойчивости к Cry1Ab. Это является показателем полезности Cry1Ca для контроля популяций SCB, которые устойчивы к Cry1Ab, или для снижения развития устойчивости к Cry1Ab в популяциях SCB.
Частично исходя из описанных в изобретении данных, считается, что совместная экспрессия Cry1Ca и Cry1Ab может создать высокоэффективный комплекс IRM для контроля SCB. Для расширения спектра действия к указанной комбинации можно добавлять другие белки. Например, для кукурузы, добавление Cry1Fa сможет создать комплекс IRM против европейского кукурузного мотылька (ECB), Ostrinia nubilalis (Hübner), при добавлении еще одного МОА создается комплекс для контроля огневки сахарного тростника SCB.
Информацию о белке Cry1C как потенциальном биоинсектициде для растений см. в публикации (Avisar et al. 2009). Avisar D, Eilenberg H, Keller M, Reznik N, Segal M, Sneh B, Zilberstein A (2009) The Bacillus thuringiensis delta-endotoxin Cry1C as a potential bioinsecticide in plants. Plant Science 176:315-324.
Рецепторы насекомых. Как описано в разделе примеров, исследования конкурентного рецепторного связывания с использованием радиомеченого корового белка-токсина Cry1Ca показали, что коровый белок-токсин Cry1Ab не конкурирует за присутствующий в тканях насекомых FAW высокоаффинный сайт связывания, с которыми связывается Cry1Ca. Эти результаты показывают, что комбинация белков Cry1Ab и Cry1Ca может быть эффективным средством для снижения развития устойчивости в популяциях FAW к Cry1Ab (и аналогично, для снижения развития устойчивости к Cry1Ca), и, вероятно, будет повышать уровень устойчивости к этому вредителю кукурузных растений, экспрессирующих оба белка.
Исходя из этих данных, также предполагается, что Cry1Ca будет эффективен для контроля популяций SCB, которые обладают устойчивостью к Cry1Ab. Один вариант осуществления состоит в использовании упомянутых белков Cry в регионах, где Cry1Ab перестал быть эффективным для контроля SCB по причине развития устойчивости. Другой вариант осуществления состоит в использовании Cry1Ca в комбинации с Cry1Ab для снижения развития устойчивости у SCB к белку Cry1Ab.
Комбинации токсинов, описанных в изобретении, можно использовать для контроля чешуекрылых вредителей. Взрослые чешуекрылые, то есть, бабочки и моли, питаются в основном цветочным нектаром. Почти все личинки, то есть, гусеницы, питаются растениями, и многие из личинок являются серьезными вредителями. Гусеницы питаются на листьях или внутри листьев, или поедают корни или стебли растения, где они лишают растение питательных веществ и часто разрушают конструкцию физической опоры растения. Дополнительно, гусеницы питаются фруктами, тканями и хранящимся зерном и мукой, разрушая указанные продукты для продажи или в значительной степени уменьшая их стоимость. Упоминаемые в изобретении чешуекрылые вредители относятся к разным стадиям жизни вредителя, включая в себя личиночные стадии.
Химерные токсины настоящего изобретения содержат полный N-концевой токсический участок корового токсина B.t. и, в некоторой точке после конца участка токсина белок несет транзицию в гетерологичной последовательности протоксина. N-концевой токсический участок токсина B.t. называется согласно изобретению "коровым" токсином. Транзиция в гетерологичном сегменте протоксина может происходить примерно около соединения токсина/протоксина или, как альтернатива, может сохраняться участок нативного протоксина (расположенный после участка токсина) с транзицией в гетерологичном протоксине, расположенной в 5'-3' направлении.
В качестве примера, один химерный токсин настоящего изобретения имеет полный коровый токсический участок Cry1Ab (аминокислоты от 1 до 601) и гетерологичный протоксин (аминокислоты от 602 до C-конца). В одном предпочтительном варианте осуществления участок химерного токсина, содержащий протоксин, происходит из белка-токсина Cry1Ab. В качестве второго примера, второй химерный токсин настоящего изобретения, показанный в SEQ ID NO:1 (DIG-152), имеет полный коровый токсический участок Cry1Ca (аминокислоты от 1 до 619) и гетерологичный протоксин (аминокислоты от 620 до C-конца). В предпочтительном варианте осуществления участок химерного токсина, содержащий протоксин, происходит из белка-токсина Cry1Ab.
Специалистам в данной области техники будет очевидно, что токсины B.t., даже относящиеся к конкретному классу, например, к Cry1Ca, до некоторой степени могут варьировать по длине и точной локализации транзиции от участка токсина до участка протоксина. Обычно длина токсинов Cry1Ca составляет от около 1150 до около 1200 аминокислот. Транзиция от участка токсина до участка протоксина будет обычно занимать участок в диапазоне от около 50% до около 60% от общей длины токсина. Химерный токсин настоящего изобретения будет полностью включать в себя всю протяженность этого корового N-концевого участка токсина. Таким образом, химерный токсин будет содержать по меньшей мере около 50% полноразмерных B.t. токсинов Cry1Ca или Cry1Ab. Обычно они будут составлять по меньшей мере около 590 аминокислот. В отношении участка протоксина, полноразмерный участок Cry1(b) протоксина расположен от конца участка токсина до C-конца молекулы. Конец этого участка длиной примерно от 100 до 150 аминокислот является наиболее важным для включения в него химерного токсина настоящего изобретения.
Гены и токсины. Гены и токсины, полезные согласно настоящему изобретению, включают в себя не только полноразмерные раскрытые последовательности, но также и фрагменты этих последовательностей, варианты, мутанты и слитые белки, которые сохраняют свойства пестицидной активности токсинов, конкретно описанных в изобретении в качестве примеров. Используемые в изобретении термины "варианты" или "изменения" генов относятся к нуклеотидным последовательностям, которые кодируют одинаковые токсины, или которые кодируют эквивалентные токсины, обладающие пестицидным действием. Используемый в изобретении термин "эквивалентные токсины" относится к токсинам, имеющим одинаковое или по существу одинаковое биологическое действие против целевых вредителей, как и токсины, указанные в формуле изобретения.
Используемые в настоящем изобретении пределы идентичности последовательностей составляют около 95% (белки Cry1Ab и 1Са), 78% (Cry1A и Cry1C) и 45% (Cry1), см. публикации "Revision of the Nomenclature for the Bacillus thuringiensis Pesticidal Crystal Proteins", N. Crickmore, D.R. Zeigler, J. Feitelson, E. Schnepf, J. Van Rie, D. Lereclus, J. Baum, and D.H. Dean. Microbiology and Molecular Biology Reviews (1998) Vol 62: 807-813. Эти пределы также можно применять только для коровых токсинов (для токсинов Cry1Ab и Cry1C). Перечень номеров GENBANK, приведенный ниже в Приложении A, также можно использовать для получения последовательности для любого из генов и белков, раскрытых или упомянутых в настоящем изобретении.
Специалистам в данной области техники будет очевидно, что существует ряд способов идентификации и получения генов, кодирующих активные токсины. Конкретные гены или участки генов, упомянутые в изобретении в качестве примера, можно выделять из коллекций депозитария культур, как описано выше. Эти гены, или их участки или варианты, также можно конструировать искусственно, например, при помощи синтезатора генов. Вариации генов можно легко конструировать с помощью стандартных технологий создания точечных мутаций. Также можно создавать фрагменты этих генов согласно стандартным методикам с использованием коммерчески доступных экзонуклеаз или эндонуклеаз. Например, можно использовать ферменты, такие как Bal31, или сайт-направленный мутагенез для систематического отсечения нуклеотидов от концов этих генов. Также можно получать гены, кодирующие активные фрагменты, с помощью ряда ферментов рестрикции. Можно использовать протеазы для прямого создания активных фрагментов упомянутых токсинов.
Фрагменты и эквиваленты, которые сохраняют пестицидное действие рассматриваемых токсинов, входят в объем настоящего изобретения. Кроме того, благодаря избыточности генного кода множество разных последовательностей ДНК может кодировать раскрытые в изобретении аминокислотные последовательности. Специалистам в данной области техники будут очевидны способы создания таких альтернативных последовательностей ДНК, кодирующих одинаковые, или по существу аналогичные токсины. Эти различные последовательности ДНК входят в объем настоящего изобретения. Используемое в изобретении понятие "по существу аналогичная" последовательность относится к последовательностям, имеющим аминокислотные замены, делеции, добавления или инсерции, которые фактически не влияют на пестицидное действие. В это определение также включены фрагменты, сохраняющие пестицидное действие.
Дополнительным способом идентификации генов и генных участков, кодирующих токсины и полезных согласно настоящему изобретению, является использование олигонуклеотидных зондов. Эти зонды представляют собой обнаружимые нуклеотидные последовательности. Детекцию таких последовательностей можно проводить с помощью подходящей метки, или можно задействовать флуоресцентные свойства, как описано в международной патентной заявке № WO 93/16094. Как известно в данной области техники, если при гибридизации молекулы зонда и образца нуклеиновой кислоты образуется прочная связь между этими двумя молекулами, то обоснованно предполагается, что зонд и образец имеют значительную степень гомологии. Предпочтительно, гибридизацию проводят при строгих условиях способами, известными в данной области техники, которые описаны, например, авторами Keller, G.H., M.M. Manak (1987) DNA Probes, Stockton Press, New York, N.Y., pp. 169-170. Некоторые примеры концентраций солей и температурных комбинаций представляют собой (в порядке увеличения степени строгости): 2 X SSPE или SSC (раствор хлорида и цитрата натрия) при комнатной температуре; 1 X SSPE или SSC при 42°C; 0,1 X SSPE или SSC при 42°C; 0,1 X SSPE или SSC при 65°C. Детекция зонда представляет собой общепринятый способ для определения факта проведения гибридизации. Такой анализ зондов обеспечивает быстрый способ идентификации кодирующих токсины генов настоящего изобретения. Нуклеотидные сегменты, которые используются в качестве зондов согласно изобретению, можно синтезировать с помощью синтезатора ДНК и стандартных методик. Указанные нуклеотидные последовательности можно также использовать в качестве ПЦР-праймеров для амплификации генов настоящего изобретения.
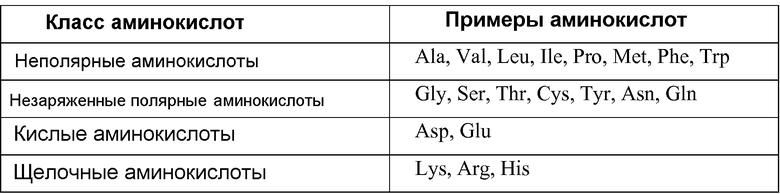
Конкретные токсины настоящего изобретения приведены в изобретении в качестве конкретных примеров. Поскольку упомянутые токсины являются просто примерами токсинов настоящего изобретения, будет очевидно, что настоящее изобретение содержит варианты или эквиваленты токсинов (и нуклеотидные последовательности, кодирующие эквивалентные токсины), обладающие пестицидным действием, которое одинаково или сходно с действием рассматриваемого токсина. Эквивалентные токсины и рассматриваемый токсин имеют аминокислотную гомологию. Эта аминокислотная гомология обычно превышает 75%, предпочтительно составляет более 90%, и наиболее предпочтительно более 95%. Наиболее выраженную аминокислотную гомологию выявляют в самых важных областях токсина, которые обуславливают биологическое действие или вовлечены в предопределение трехмерной конфигурации, которая в конечном счете отвечает за биологическое действие. В этом отношении подходят конкретные замены аминокислоты, и можно предполагать, что эти замены находятся в областях, не имеющих решающего значения для действия, или они представляют собой консервативные аминокислотные замены, которые не затрагивают трехмерную конфигурацию молекулы. Например, аминокислоты могут относиться к следующим классам: неполярные, незаряженные полярные, щелочные и кислые. Консервативные замены, посредством которых аминокислота одного класса заменяется на другую аминокислоту того же типа, входят в объем настоящего изобретения, если эта замена в значительной степени не изменяет биологическое действие соединения. В таблице 1 приведен перечень примеров аминокислот, принадлежащих к каждому классу.
В некоторых случаях также можно делать неконсервативные замены. Решающим фактором является то, что эти замены не должны в значительной степени снижать биологическое действие токсина.
Рекомбинантные хозяева. Гены, кодирующие токсины настоящего изобретения, можно вставлять в широкий спектр клеток-хозяев микроорганизмов или растений. Экспрессия генов токсина прямо или опосредованно приводит к внутриклеточной продукции и сохранению пестицида. Можно использовать конъюгационный перенос и рекомбинантный перенос для создания штамма B.t., который экспрессирует оба токсина настоящего изобретения. Другие организмы-хозяева также можно трансформировать одним или двумя генами токсина, используемых в этом случае для достижения синергетического эффекта. Можно применять микроорганизмы с подходящими микробными хозяевами, например, Pseudomonas, у вредителя в месте их размножения и поглощения. Результатом является контроль над вредителем. Альтернативно, микроорганизм, являющийся хозяином гена токсина, можно обрабатывать при условиях, которые пролонгируют действие токсина и стабилизируют клетку. Затем обработанную клетку, которая сохраняет токсическую активность, можно применять в окружающей среде целевого вредителя.
Если ген токсина B.t. внедряют в микроорганизм-хозяин посредством подходящего вектора, и указанный хозяин используется в окружающей среде в живом состоянии, имеет значение использование конкретных микроорганизмов-хозяев. Хозяев выбирают из микроорганизмов, которые обладают известной способностью в одной или более рассматриваемых сельскохозяйственных культур заселять "фитосферу" (филлоплан, филлосферу, ризосферу и/или ризоплан). Выбирают такие микроорганизмы, которые способны успешно конкурировать в конкретной среде (сельскохозяйственные культуры и другие среды обитания насекомых) с микроорганизмами дикого типа, обеспечивать устойчивое сохранение и экспрессию гена, экспрессирующего полипептидный пестицид, и обеспечивать улучшенную защиту пестицида от разложения и инактивации в окружающей среде.
Известно, что большое количество микроорганизмов населяет филлоплан (поверхность листьев растения) и/или ризосферу (почву, окружающую корни растения) у широкого спектра важных сельскохозяйственных культур. Эти микроорганизмы включают в себя бактерии, морские водоросли и грибы. Особый интерес вызывают такие микроорганизмы, как, например, бактерии рода Pseudomonas, Erwinia, Serratia, Klebsiella, Xanthomonas, Streptomyces, Rhizobium, Rhodopseudomonas, Methylophilius, Agrobactenum, Acetobacter, Lactobacillus, Arthrobacter, Azotobacter, Leuconostoc и Alcaligenes; грибы, особенно дрожжи, например, рода Saccharomyces, Cryptococcus, Kluyveromyces, Sporobolomyces, Rhodotorula и Aureobasidium. Особый интерес вызывают такие фитосферные виды бактерий как Pseudomonas syringae, Pseudomonas fluorescens, Serratia marcescens, Acetobacter xylinum, Agrobactenium tumefaciens, Rhodopseudomonas spheroides, Xanthomonas campestris, Rhizobium melioti, Alcaligenes entrophus и Azotobacter vinlandii; и фитосферные виды дрожжей, такие как Rhodotorula rubra, R. glutinis, R. marina, R. aurantiaca, Cryptococcus albidus, C. diffluens, C. laurentii, Saccharomyces rosei, S. pretoriensis, S. cerevisiae, Sporobolomyces roseus, S. odorus, Kluyveromyces veronae и Aureobasidium pollulans. Особый интерес вызывают пигментированные микроорганизмы.
Широкое разнообразие путей доступно для внедрения в микроорганизм-хозяин гена B.t., кодирующего токсин при условиях, которые позволяют устойчиво сохранять и экспрессировать ген. Эти способы известны специалистам в данной области техники и описаны, например, в патенте США № 5135867, который включен в изобретение путем ссылки.
Обработка клеток. Bacillus thuringiensis или рекомбинантные клетки, экспрессирующие токсины B.t., можно обрабатывать для пролонгирования действия токсина и стабилизации клетки. Образуемая микрокапсула пестицида содержит токсин B.t. или токсины в клеточной структуре, которая была стабилизирована, и будет защищать токсин в ходе применения микрокапсулы в среде вредителя-мишени. Подходящие клетки-хозяева могут включать в себя или прокариоты или эукариоты, и обычно ограничены клетками, которые не продуцируют веществ, являющихся токсичными для высших организмов, например, млекопитающих. Вместе с тем, можно использовать организмы, которые продуцируют токсичные вещества для высших организмов, если эти токсичные вещества нестабильны или уровень применения является достаточно низким, чтобы избежать любой вероятности токсичного воздействия на хозяина-млекопитающего. В качестве хозяев особый интерес представляют прокариоты и низшие эукариоты, такие как грибы.
Обычно для обработки задействуют интактные клетки, в основном в пролиферативной форме, а не в виде споры, хотя в некоторых случаях можно использовать споры.
Обработку микробной клетки, например, микроорганизма, содержащего ген или гены токсина B.t., можно осуществлять химическими или физическими способами, или комбинацией химических и/или физических способов при условии, что отсутствует и вредное воздействие технологии на свойства токсина, и уменьшение способности клеток к защите токсина. Примерами химических реагентов являются галогеновые агенты, в частности, галогены с атомными номерами 17-80. Более конкретно, можно использовать иод при умеренных условиях и в течение достаточного времени для достижения желательных результатов. Другие подходящие способы включают в себя обработку альдегидами, такими как глутаральдегид; противоинфекционными агентами, например, зефиран хлоридом и цетилпиридиния хлоридом; спиртами, такими как изопропил и этанол; различными гистологическими фиксаторами, такими как иодный раствор Люголя, фиксатор Боуэна, различные кислоты и фиксатор Хэлли (См.: Humason, Gretchen L., Animal Tissue Techniques, W. H. Freeman and Company, 1967); или комбинацию физических воздействий (нагревание) и химических агентов, которые сохраняют и пролонгируют действие токсина, продуцируемого в клетке при введении клетки в среду хозяина. Примерами физических способов является коротковолновое излучение, например, гамма-излучение и рентгеновское излучение, замораживание, ультрафиолетовое облучение, лиофилизация и т.п. Способы обработки микробных клеток раскрыты в патентах США № 4 695455 и 4695462, включенных в изобретение путем ссылки.
Обычно клетки имеют повышенную структурную устойчивость, которая увеличивает их стабильность в условиях окружающей среды. Если пестицид находится в предварительной форме, необходимо выбирать способ обработки клеток, который не ингибирует превращение пре-формы в зрелую форму пестицида посредством патогена целевого вредителя. Например, формальдегид сшивает белки и способен ингибировать превращение пред-формы полипептида пестицида. Способ обработки должен сохранять по меньшей мере значительную часть свойств биодоступности или биоактивности токсина.
Особенно важные свойства для отбора клеток-хозяев в целях продукции включают в себя простоту внедрения гена или генов B.t. в клетку-хозяин, доступность систем экспрессии, эффективность экспрессии, стабильность пестицида в хозяине и присутствие дополнительных генетических способностей. Рассматриваемые свойства для применения в качестве пестицидной микрокапсулы включают в себя защитные характеристики для пестицида, такие как толстые клеточные стенки, пигментация и внутриклеточная упаковка или образование телец включения; выживание в водной среде; отсутствие токсичности для млекопитающих; привлекательность для поглощения вредителями; способность легко вызывать гибель и фиксироваться без повреждения токсина; и т.п. Другие важные условия включают в себя простоту изготовления и использования, экономичность, стабильность при хранении и тому подобное.
Рост клеток. Клетку-хозяин, содержащую инсектицидные ген или гены B.t., можно выращивать в любой удобной питательной среде, в которой конструкция ДНК дает селективное преимущество, и во всей среде или по существу во всех клетках в селективной среде обеспечивается сохранение гена B.t. Затем эти клетки можно собирать общепринятыми способами. Альтернативно, клетки можно обрабатывать перед их сбором.
Клетки B.t., продуцирующие токсины по изобретению, можно культивировать с использованием стандартных в данной области техники сред и способов ферментации. После завершения цикла ферментации бактерии можно собирать из ферментационного бульона первой сепарацией спор B.t. и кристаллов с помощью способов, известных в данной области. Восстановленные споры B.t. и кристаллы можно объединять в смачивающийся порошок, жидкий концентрат, гранулы или другие рецептуры с помощью добавления сурфактантов, диспергирующих агентов, инертных носителей и других компонентов, чтобы облегчить обработку и применение для конкретных целевых вредителей. Эти рецептуры и методики применения широко известны в данной области техники.
Рецептуры. Рецептуры гранулированных приманок, содержащие аттрактант и споры, кристаллы и выделенные B.t. токсины, или рекомбинантные микроорганизмы, которые содержат гены, получаемые из изолятов B.t., раскрытые в настоящем изобретении, можно применять на почве. Рецептуру продукта также можно применять в качестве покрытия семян, или для обработки корней, или для полной обработки растения на более поздних этапах цикла злакового растения. Для обработки растений и почвы можно использовать B.t.-клетки в виде смачивающихся порошков, гранул или порошковых препаратов путем смешивания с различными инертными материалами, такими как неорганические минералы (филлосиликаты, карбонаты, сульфаты, фосфаты и т.п.) или растительными материалами (порошок из кукурузных початков, рисовая шелуха, скорлупа грецкого ореха и т.п.). Рецептуры могут включать в себя адгезивные адъюванты, стабилизирующие агенты, другие пестицидные добавки или сурфактанты. Жидкие рецептуры могут иметь водную или неводную основу и использоваться в виде пены, гелей, суспензий, эмульгируемых концентратов или подобных форм. Компоненты могут включать в себя реологические агенты, сурфактанты, эмульгаторы, диспергирующие агенты или полимеры.
Специалистам в данной области техники будет очевидно, что концентрация пестицида может широко варьировать в зависимости от природы конкретной рецептуры, в частности, является ли она концентратом или изготовлена для непосредственного применения. Пестицид может присутствовать в количестве по меньшей мере 1% веса и может составлять 100% веса. В сухих рецептурах пестицид может составлять примерно от 1 до 95% веса, тогда как жидкие рецептуры обычно могут иметь примерно от 1 до 60% веса твердых частиц в жидкой фазе. В рецептурах обычно может находиться от около 102 около до 104 клеток/мг. Указанные рецептуры можно применять в количестве примерно от 50 мг (в жидком или сухом виде) до 1 кг или больше на гектар.
Рецептуры можно применять в среде обитания чешуекрылых вредителей, например, на листве или на почве, путем распыления, опудривания, опрыскивания или подобными способами.
Трансформация растений. Предпочтительным рекомбинантным хозяином, продуцирующим инсектицидные белки настоящего изобретения, является трансформированное растение. Гены, кодирующие Bt белки-токсины, раскрытые в изобретении, можно вставлять в клетки растения, используя множество способов, известных в данной области техники. Например, для подготовки к вставке чужих генов в высшие растения доступно большое количество векторов клонирования, содержащих систему репликации в Escherichia coli и маркер, позволяющий проводить селекцию трансформированных клеток. Векторы содержат, например, среди прочего, pBR322, серии pUC, серии M13mp, pACYC184. Соответственно, фрагмент ДНК, несущий последовательность, кодирующую Bt белок-токсин, можно вставлять в вектор на подходящем участке рестрикции. Полученную плазмиду используют для трансформации в E. coli. Клетки E. coli выращивают в подходящей питательной среде, затем собирают и лизируют. Восстанавливают плазмиду. В качестве способов анализа обычно проводят анализ последовательностей, анализ рестрикции, электрофорез и другие биохимические и молекулярно-биологические тесты. После каждой манипуляции используемую последовательность ДНК можно расщеплять и соединять со следующей последовательностью ДНК. Каждую последовательность плазмиды можно клонировать в той же плазмиде или в других плазмидах. В зависимости от способа вставки желательных генов в растение могут потребоваться другие последовательности ДНК. Например, если для трансформации растительной клетки использована плазмида Ti или Ri, то по меньшей мере правая граница, но часто и правая и левая граница Ti или Ri плазмиды T-ДНК должны соединяться в качестве фланкирующей области генов, предназначенных для вставки. Проведены интенсивные исследования использования T-ДНК для трансформации растительных клеток, что в достаточном объеме описано в патенте EP 120516, Lee and Gelvin (2008), Hoekema (1985), Fraley et al., (1986) и An et al., (1985), и хорошо известно в данной области техники.
После включения в геном растения вставленная ДНК является относительно стабильной. Вектор трансформации обычно содержит выбираемый маркер, который придает трансформированным растительным клеткам резистентность к биоциду или антибиотику, такому как биалафос, канамицин, G418, блеомицин или гигромицин, среди прочего. Конкретный используемый маркер должен соответственно позволять отбор трансформированных клеток, а не клеток, не содержащих ДНК-вставку.
Для вставки ДНК в растительную клетку-хозяин доступно большое количество способов. Эти способы включают в себя трансформацию с T-ДНК, с использованием в качестве агента трансформации Agrobacterium tumefaciens или Agrobacterium rhizogenes, слияние, инъекцию, биолистику (бомбардировку микрочастицами) или электропорацию, а также другие возможные способы. Если для трансформации используют Agrobacteria, вставляемую ДНК необходимо клонировать в специальные плазмиды, а именно, или в промежуточный вектор или в бинарный вектор. Промежуточные векторы можно интегрировать в плазмиды Ti или Ri путем гомологичной рекомбинации обусловленной последовательностями, которые являются гомологичными последовательностям в T-ДНК. Плазмиды Ti или Ri также содержат vir-область, необходимую для переноса T-ДНК. Промежуточные векторы не могут самостоятельно реплицироваться в Agrobacteria. Промежуточный вектор можно переносить в Agrobacterium tumefaciens посредством хелперной плазмиды (конъюгация). Бинарные векторы могут реплицироваться и в E. coli и в Agrobacteria. Они содержат ген маркера селекции и линкер или полилинкер, которые ограничены правыми и левыми пограничными областями T-ДНК. Они могут быть трансформированы непосредственно в Agrobacteria (Holsters et al., 1978). Используемая в качестве клетки-хозяина Agrobacterium должна содержать плазмиду, несущую vir-область. Эта vir-область необходима для переноса T-ДНК в клетку растения. Может присутствовать дополнительная T-ДНК. Трансформированная указанным путем бактерия используется для трансформации клеток растения. Растительные экспланты предпочтительно можно культивировать с Agrobacterium tumefaciens или Agrobacterium rhizogenes для переноса ДНК в клетку растения. Затем можно восстанавливать целые растения из инфицированного растительного материала (например, из частей листьев, сегментов стеблей, корней, но также из протопластов или суспензии культивированных клеток) в подходящей среде, которая может содержать антибиотики или биоциды для селекции. Полученные указанным путем растения затем можно тестировать на присутствие вставки ДНК. Специальных требований к плазмидам в случае инъекции и электропорации не существует. Можно использовать обычные плазмиды, например, такие как производные pUC.
Трансформированные клетки растут внутри растений обычным путем. Они могут формировать зародышевые клетки и передавать трансформированный признак (признаки) растениям-потомкам. Такие растения можно выращивать обычным путем и скрещивать с растениями, которые имеют те же самые трансформированные наследственные факторы или другие наследственные факторы. Получаемые гибридные субъекты имеют соответствующие фенотипические свойства.
В предпочтительном варианте осуществления настоящего изобретения растения трансформируют генами, в которых используемый кодон был оптимизирован для растений. См., например, патент США № 5380831, включенный в изобретение путем ссылки. Некоторые усеченные токсины рассмотрены в изобретении в качестве примера, вместе с тем в области техники Bt-белков известно, что токсины 130 кДа-типа (полноразмерные) имеют N-концевую группу, которая представляет собой коровый токсин, и C-концевой остаток, который является "хвостом" протоксина. Таким образом, соответствующие "хвосты" можно использовать с усеченными/коровыми токсинами настоящего изобретения. См. например, патент США № 6218188 и патент США № 6673990. Дополнительно, в данной области техники известны способы создания синтетических Bt-генов для использования в растениях (Stewart and Burgin, 2007). Одним из неограничивающих примеров предпочтительного трансформированного растения является фертильное растение кукурузы, содержащее растительный экспрессируемый ген, кодирующий белок Cry1Fa, и дополнительно содержащее второй растительный экспрессируемый ген, кодирующий белок Cry1Ca.
Перенос (или вставка) признака (признаков) Cry1Ab и Cry1C в инбредные линии кукурузы может осуществляться размножением с рекуррентной селекцией, например, путем обратного скрещивания. В этом случае, желательный рекуррентный родитель вначале скрещивают с инбредным донором (нерекуррентным родителем), который несет соответствующий ген (гены) для признаков Cry1Ab и Cry1C. Потомство от этого скрещивания затем обратно спаривают с рекуррентным родителем, с последующей селекцией получаемого потомства на желательный признак (признаки), переносимый от нерекуррентного родителя. Через три, предпочтительно четыре, более предпочтительно, через пять или больше поколений обратных скрещиваний с рекуррентным родителем с селекцией на желательный признак (признаки), потомство будет гетерозиготным по локусам, отвечающим за переносимый признак (признаки), но будет похожим на рекуррентного родителя по большинству или почти по всем остальным генам (см., например, Poehlman & Sleper (1995) Breeding Field Crops, 4th Ed., 172-175; Fehr (1987) Principles of Cultivar Development, Vol. 1: Theory and Technique, 360-376).
Стратегии управления устойчивостью насекомых (IRM). Авторы Roush et al., например, выделяют стратегию двух токсинов, также называемый "пирамидинг" или "комплект", для управления инсектицидными трансгенными сельскохозяйственными культурами. (The Royal Society. Phil. Trans. R. Soc. Lond. B. (1998) 353, 1777-1786). На вебсайте Управления по охране окружающей среды США (the United States Environmental Protection Agency: epa.gov/oppbppdl/biopesticides/pips/bt_corn_refuge_2006.htm) опубликованы следующие требования для обеспечения посевов нетрансгенных (то есть не-B.t) «убежищ» (блок не-Bt сельскохозяйственных культур/кукурузы), высеваемый вместе с трансгенными зерновыми культурами, продуцирующими единственный Bt белок, активный против целевых вредителей.
Существуют следующие конкретные структурированные требования для кукурузной продукции, Bt-защищенной от кукурузного мотылька (Cry1Ab или Cry1F):
Структурированные «убежища»:
20% площади в кукурузной зоне отводят под «убежище» для Bt-незащищенной от чешуекрылых вредителей кукурузы
50% площади в хлопковой зоне отводят для «убежища» культур, Bt-незащищенных от чешуекрылых вредителей
Блоки
1. Внутренние (то есть в пределах Bt-поля)
2. Внешние (то есть отдельные поля в пределах ½ мили (¼ мили, по возможности) от Bt-поля, для максимального увеличения частоты случайного спаривания)
Полосы внутри поля
Ширина полос должна составлять по меньшей мере 4 ряда (предпочтительно 6 рядов), для уменьшения эффекта перемещения личинок.
На вебсайте Национальной ассоциации производителей кукурузы (ncga.com/insect-resistance-management-fact-sheet-bt-com), также опубликовано подобное руководство в отношении требований. Например:
Требования IRM по кукурузному мотыльку:
- По меньшей мере 20% площадей отводят под выращивание «убежищ» гибридной кукурузы
- В зонах выращивания хлопка «убежища» должны составлять 50%
- Выращивание должно быть на расстоянии в пределах 1/2 мили от гибридов «убежища»
- «Убежища» можно выращивать в виде полос внутри Bt-поля; полосы «убежища» должны иметь ширину по меньшей мере 4 ряда
- «Убежище» можно обрабатывать обычными пестицидами, только в случае достижения экономического порога для целевого насекомого
- В «убежище» кукурузы нельзя применять распыляемые инсектициды на основе Bt
- Соответствующее «убежище» должно выращиваться на каждой ферме с Bt-кукурузой
Авторы Roush et al. утверждают (например, правая колонка, страницы 1780 и 1784) что комплект или «пирамидинг» из двух разных белков, каждый из которых эффективен против целевых вредителей и не имеет какой-либо перекрестной устойчивости или слабую перекрестную устойчивость, может дать возможность применять убежища меньшего размера. Roush предполагает, что при успешном комплекте размер убежища менее 10% убежища может дать сопоставимый контроль устойчивости до уровня 50% убежища для единственного (не-пирамидного) признака. Для доступных в настоящее время пирамидных Bt-кукурузных продуктов согласно требованиям Управления по охране окружающей среды США необходимо структурированное убежище не-Bt кукурузы значительно меньшей площади (обычно 5%), чем для продуктов с единственным признаком (обычно 20%).
Любое из вышеупомянутых процентных соотношений (таких, как например, для 1F/Ab), или подобных соотношений убежищ можно применять для рассматриваемых двойных или тройных комплектов или пирамид. Настоящее изобретение включает в себя коммерческую площадь земли в акрах, например, более 10 акров, с выращиванием с упомянутым убежищем (или без него), и с растениями согласно настоящему изобретению.
Существуют разные способы выращивания убежищ, включающие в себя выращивание в полях с различной геометрической планировкой (как упомянуто выше), и упаковки с семенной смесью, как дополнительно рассмотрено Roush и, например, в патенте США № 6551962.
Все патенты, патентные заявки, предварительные заявки и публикации, упомянутые или цитируемые в изобретении, включены в изобретение путем ссылки во всей полноте при условии, что они не противоречат идее настоящей заявки. Если конкретно не обозначено или подразумевается, используемые в изобретении термины в единственном числе обозначают "по меньшей мере один".
Далее в изобретении приведены следующие иллюстративные примеры. Примеры не должны рассматриваться как ограничение объема изобретения.
ПРИМЕР 1
Дизайн химерных токсинов, содержащих коровые токсины Cry1 и гетерологичные протоксины, и инсектицидное действие белка DIG-152, продуцируемого в Pseudomonas fluorescens
Химерные токсины. Химерные белки, несущие домен корового токсина из одного токсина Cry, которые слиты с сегментом протоксина из другого токсина Cry, описаны ранее, например, в патенте США № 5593881 и патенте США № 5932209.
Химерные варианты белка Cry1Ca настоящего изобретения включают в себя химерные токсины, содержащие N-концевой сегмент корового токсина, происходящий из инсектицидного токсина Cry1Ca3, который слит с сегментом гетерологичного дельта-эндотоксинового протоксина в некоторой точке после конца сегмента корового токсина. Транзиция от корового токсина до сегмента гетерологичного протоксина может происходить примерно около сочленения нативного корового токсина/протоксина, может сохраняться ora-участок нативного протоксина (расположенный после сегмента корового токсина), с транзицией гетерологичного протоксина, возникающей в 5'-3' направлении. Разными механизмами сегменты корового токсина и протоксина могут содержать точную аминокислотную последовательность нативных токсинов, из которой они происходят, или могут включать в себя аминокислотные добавления, делеции или замены, которые не уменьшают биологическую функцию сегментов при слиянии их друг с другом и способны усилить указанную функцию.
Например, химерный токсин настоящего изобретения содержит сегмент корового токсина, происходящий из Cry1Ca3 и гетерологичного протоксина. В предпочтительном варианте осуществления изобретения сегмент корового токсина, происходящий из Cry1Ca3 (619 аминокислот), является слитым с гетерологичным сегментом, содержащим сегмент протоксина, происходящий из дельта-эндотоксина Cry1Ab (545 аминокислот). Состоящая из 1164 аминокислот аминокислотная последовательность химерного белка, именуемого в изобретении DIG-152, раскрыта в SEQ ID NO:1. Подразумевается, что другие химерные слияния, содержащие варианты корового токсина Cry1Ca3 и протоксины, происходящие из Cry1Ab, входят в объем настоящего изобретения.
Инсектицидное действие белка DIG-152 против чешуекрылых было продемонстрировано на личинках новорожденной огневки сахарного тростника (SCB; Diatraea saccharalis) и Cry1Ab-резистентной SCB (rSCB) в экспериментах зависимости реакции от дозы, с помощью методик введения рациона. Тельца включения DIG-152 солюбилизировали при аккуратном покачивании при 4º в течение 4 часов в 7,5 мл 100 мМ CAPS, pH 11, 1 мМ ЭДТА, к которому добавляли 200 мкл ингибитора бактериальной протеазы (Sigma P4865; приготовленной согласно инструкции поставщика). После центрифугирования до получения осадка нерастворимого материала стоковую концентрацию белка доводили до 4,0 мг/мл в 100 мМ CAPS, pH 11. Для биотеста с насекомыми приготовляли корм с концентрацией белка DIG-152 в диапазоне от 0,030 мкг до 102 мкг/г, путем смешивания подходящих объемов с рационом меридик (meridic) (Bio-Serv, Frenchtown, NJ) непосредственно перед распределением около 0,7 мл рациона в отдельные ячейки в кюветах с 128 ячейками (Bio-Ba-128, C-D International).
Трипсин-активированный белок Cry1Ab (используемый в качестве положительного контроля инсектицидной активности) тестировали в диапазоне концентрации от 0,03125 мкг до 32 мкг/г рациона (приготовленного путем смешивания лиофилизированного порошка с соответствующим количеством дистиллированной воды перед приготовлением рациона).
В качестве контрольных введений использовали рационы, приготовленные с дистиллированной водой (чистый контроль для тестов Cry1Ab) или с буфером единственным (100 мМ CAPS pH 11 для DIG-152 теста). Из каждой ячейки с поверхности рациона забирали одну новорожденную личинку D. saccharalis (< через 24 часа после вылупления). После инокуляции личинки ячейки накрывали крышками с отверстиями (C-D International), и кюветы с биопробами помещали в камеру с искусственным климатом, где поддерживали условия 28ºС, относительную влажность RH 50% и цикл свет/темнота 16 часов:8 часов. На седьмой день после инокуляции регистрировали смертность личинок, вес личинок и число выживших личинок, которые не показывали увеличение веса (<0,1 мг на личинку). Тест с каждой комбинацией штамм насекомого/концентрация белка Cry повторяли четыре раза, и в каждом повторе задействовали от 16 до 32 личинок.
Критерии смертности личинок измеряли как "фактическую" смертность, которая учитывала и мертвые (смертность) личинки и выжившие (чахлые, не питающиеся) личинки, которые не показывали значительный прирост массы тела (то есть <0,1 мг на личинку). Фактическую смертность личинок при обработке вычисляли с помощью уравнения:
Фактическая смертность (%) = [TDS/TNIT]×100
где TDS обозначает общее количество мертвых личинок плюс количество чахлых личинок,
и TNIT обозначает общее количество насекомых при обработке.
"Фактическую" смертность (далее для упрощения называемую "смертностью") каждого штамма D. saccharalis корректировали по относительной личиночной смертности, наблюдаемой с чистым водным контрольным рационом для анализа результатов после обработки Cry1Ab, или с рационом единственного буфера при обработке DIG-152.
Результаты экспериментов зависимости реакции от дозы дополнительно анализировали для установления значения GI50, (то есть, концентрации B.t. белка в рационе, при котором показатели замедления роста личинок (%GI) составляли 50). Процентный показатель %GI личинок при рационе, содержащем белок Cry1Ab, вычисляли с помощью формулы:
%GI = [TWC-TWT]/TWC×100
где TWC обозначает общую массу тела личинок, питающихся контрольным водным рационом, и
TWT обозначает общую массу тела личинок, питающихся вводимым рационом Cry1Ab,
где для анализа показателя %GI личинок в результате потребления белка DIG-152 расчеты проводили по формуле:
%GI = [TWB-TWT]/TWB×100
где TWB обозначает общую массу тела личинок, потреблявших контрольный рацион с единственным буфером, и
TWT обозначает общую массу тела личинок, потреблявших рацион с белком DIG-152.
100% замедление роста личинок объяснялось репликацией, если отсутствовали какие-либо личинки с выраженным увеличением веса (< 0,1 мг на личинку). Анализ показателей замедления роста проводили с помощью двухпоточной ANOVA со штаммом насекомых и концентрацией белка Cry в качестве основных двух факторов. Для определения разницы в тестах на уровне = 0,05 применяли тесты LSMEANS.
Результаты биотестов поглощения рационов личинками Diatraea saccharalis приведены в таблице 2.

bмкг белка/г рациона
сИзмерение смертности личинок проводили согласно описанию в тексте изобретения
dЭти процентные показатели рассчитывали по формуле, приведенной в тексте
еЭти процентные показатели рассчитывали по формуле, приведенной в тексте
fNT = тест не проводили
Анализ данных. Скорректированные данные доза/смертность затем подвергали пробит-анализу для определения концентраций используемого белка, которые вызывали смертность в 50% случаев (значение LC50), и соответствующие 95%-ые доверительные интервалы (CI). Используемые в пробит-анализе введения белков показатели включали в себя самую высокую концентрацию, которая вызывала нулевую смертность, самую низкую концентрацию, которая вызывала 100% смертность, и все результаты между этими крайними значениями. Соотношения устойчивости вычисляли путем деления значения LC50 от штамма rSCB на этот показатель от насекомых SCB. Тест соотношения летальной дозы использовали для определения выраженности соотношений устойчивости на уровне = 0,05. Также использовали двухпоточный тест ANOVA для анализа данных смертности, с последующим тестом LSMEANS на уровне = 0,05, с целью определения разницы при введении белков. Результаты исследований представлены в таблице 3.
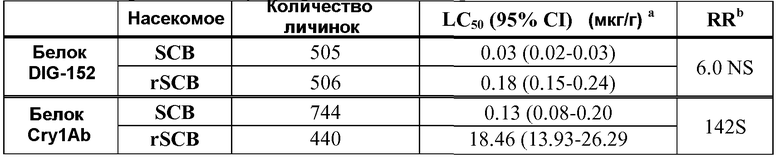
bСоотношения устойчивости, обозначенные буквой S, были значительными, NS обозначает незначительный уровень в 5% на основе тестов с летальной дозой.
Свойством белка DIG-152 настоящего изобретения является замедление роста новорожденных личинок огневки сахарного тростника (Diatraea saccharalis) или смерть личинок после поглощения белка DIG-152 в количестве, подобном количеству активированного белка Cry1Ab, который вызывает аналогичную биологическую реакцию. Дополнительное свойство белка DIG-152 состоит в том, что личинки Diatraea saccharalis, которые являются устойчивыми к токсическому действию белка Cry1Ab, тем не менее восприимчивы к токсическому действию белка DIG-152.
ПРИМЕР 2
Конструирование плазмид экспрессии, кодирующих химерные белки, и экспрессия в Pseudomonas
Использовали стандартные способы клонирования (описанные, например, авторами Sambrook et al, (1989) и Ausubel et al, (1995), с обновлениями] для дизайна конструкции экспрессии Pseudomonas fluorescens (Pf) pMYC2547, которую создавали генноинженерным способом для продукции полноразмерных химерных белков DIG-152. Продукцию белка осуществляли в штаммах MB214 Pseudomonas fluorescens (производное штамма MB101; биовар I P. fluorescens), которые несли вставку модифицированного lac-оперона, согласно раскрытию в патенте США № 5169760. Основная стратегия клонирования подразумевала субклонирование в плазмидные векторы фрагмента ДНК, кодирующего DIG-152, посредством чего его помещали под контроль экспрессии промотором Ptac и терминатором rrnBTlT2 из плазмиды pKK223-3 (PL Pharmacia, Milwaukee, WI). Одна из таких плазмид называется pMYC2547, и изолят MB214, несущий указанную плазмиду, называется Dpf108.
Анализ роста и экспрессии во встряхиваемых колбах. Продукцию белка DIG-152 для изучения свойств и биотестов с насекомыми осуществляли путем выращивания во встряхиваемых колбах штамма Dpf108 P.fluorescens. Продукцию белка DIG-152 с помощью промотора Ptac проводили, согласно описанию из ранее опубликованного патента США № 5527883. Подробности микробиологических манипуляций доступны в публикациях Squires et al., (2004), патентной заявке США № 20060008877, патентной заявке США 20080193974 и патентной заявке США, которые включены в изобретение путем ссылки. Экспрессию индуцировали добавлением изопропил-β-D-1-тиогалактопиранозида (IPTG) после начальной инкубации в течение 24 часов при 30º со встряхиванием. Культуры собирали в момент индукции и в разные точки времени после индукции. Плотность клеток измеряли по оптической плотности при 600 нм (OD600).
Фракционирование клеток и анализ SDS-PAGE образцов встряхиваемых колб. В каждый момент сбора образцов плотность клеток в образцах регулировали до OD600=20, и аликвотные количества объемом 1 мл центрифугировали при 14000×g в течение пяти минут. Клеточный осадок замораживали при -80°С. Растворимые и нерастворимые фракции из замороженных образцов клеточного осадка из встряхиваемых колб получали с помощью экстракционного раствора EasyLyse™ для бактериального белка (EPICENTRE® Biotechnologies, Madison, WI). Каждый клеточный осадок ресуспендировали в 1 мл раствора EasyLyse™, дополнительно разводили 1:4 в лизирующем буфере и инкубировали со встряхиванием при комнатной температуре в течение 30 минут. Лизат центрифугировали при 14000 оборотов в минуту в течение 20 минут при 4°С, и супернатант восстанавливали в виде растворимой фракции. Осадок (нерастворимую фракцию) затем ресуспендировали в равном объеме фосфатно-буферного раствора (ФБР; 11,9 мМ Na2HPO4, 137 мМ NaCl, 2,7 мМ KC1, pH 7,4).
Образцы смешивали 1:1 с 2 X буфера для образцов Laemmli, содержащего β-меркаптоэтанол (Sambrook et al., выше), и кипятили в течение 5 минут до загрузки на гели Criterion XT 12% Bis-Tris (Bio-Rad Inc., Hercules, CA). Электрофорез проводили в рекомендуемом буфере XT MOPS. Гели окрашивали биобезопасным красителем Кумасси (Bio-Safe Coomassie) согласно протоколу изготовителя (Bio-Rad) и визуализировали с использованием системы отображения Alpha Innotech (San Leandro, CA).
Приготовление телец включения. Приготовление телец включения (IB) белка DIG-152 проводили на клетках после ферментации P. fluorescens, при которой продуцировался нерастворимый инсектицидный Bt-белок, что выявляли с помощью электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия SDS-PAGE и MALDI-MS (лазерной десорбцией/ионизацией с использованием матрицы - масс-спектрометрией). Ферментационный осадок P. fluorescens оттаивали при 37°С на водяной бане. Клетки ресуспендировали до 25% вес/объем в лизирующем буфере [50 мм Трис, pH 7,5, 200 мМ NaCl, 20 мМ динатриевой соли ЭДТА (этилендиаминтетрауксусная кислота), 1% Тритона X-100 и 5 мМ дитиотретиола (ДТТ); непосредственно перед использованием добавляли 5 мл/л ингибирующего коктейля бактериальной протеазы (№ в каталоге P8465; Sigma-Aldrich, St.Louis, MO)]. Клетки суспендировали с помощью переносного гомогенизатора с установками на самом низком уровне (Tissue Tearor, BioSpec Products, Inc., Bartlesville, OK). К суспензии клеток добавляли лизозим (25 мг Sigma L7651, из белка куриного яйца) путем смешивания металлической лопаточкой, и суспензию инкубировали при комнатной температуре в течение одного часа. Суспензию охлаждали на льду в течение 15 минут, затем обрабатывали ультразвуком с помощью Branson Sonifier 250 (две сессии по 1 минуте, рабочий цикл 50%, выход 30%). Лизис клеток проверяли микроскопией. При необходимости добавляли дополнительные 25 мг лизозима, и повторяли инкубацию и обработку ультразвуком. После подтверждения лизиса клеток с помощью микроскопии лизат центрифугировали при 11500×g в течение 25 минут (4°С) для получения осадка IB, затем удаляли супернатант. Осадок IB ресуспендировали с 100 мл лизирующего буфера, гомогенизировали в переносном миксере и центрифугировали, как указано выше. Осадок IB повторно промывали путем ресуспендирования (в 50 мл лизирующего буфера), проводили гомогенизацию, обработку ультразвуком и центрифугирование, пока супернатант не становился бесцветным, и получали твердый осадок IB грязно-белого цвета. Для заключительного промывания осадок IB ресуспендировали в фильтрованной в стерильных условиях (0,22 мкМ) дистиллированной воде, содержащей 2 мМ ЭДТА и центрифугировали. Конечный осадок ресуспендировали в стерильной фильтрованной дистиллированной воде, содержащей 2 мМ ЭДТА, и сохраняли в аликвотных количествах 1 мл при -80°С.
Анализ SDS-PAGE и количественное определение белка в препаратах IB проводили путем оттаивания аликвотного количества 1 мл осадка IB и разведения его 1:20 в стерильной фильтрованной дистиллированной воде. Затем разведенный образец кипятили с 4 X редуцирующим буфером для образцов [250 мМ Трис, pH 6,8, 40% глицерин (объем/объем), бромфенол синий 0,4% (вес/объем), 8% додецилсульфат натрия SDS (вес/объем) и 8% β-меркаптоэтанол (об/об)] и загружали на гель 4-20% Novex® Трис-глицин, 12+2 лунок (Invitrogen), который пропускали с 1X буфером Трис/глицин/SDS (BioRad). Гель пропускали в течение 60 минут при 200 вольт, затем окрашивали Кумасси синим (50% G-250/50% R-250 в 45% метаноле, 10% уксусная кислота) и обесцвечивали 7% уксусной кислотой, 5% метанолом в дистиллированной воде. Количественное определение целевых полос проводили путем сравнения денситометрических показателей для полос со стандартными образцами бычьего сывороточного альбумина (BSA), которые прогоняли на том же геле для получения стандартной кривой.
Солюбилизация телец включения. Шесть мл суспензии телец включения DIG-152 из Pf клона DPf108 центрифугировали с самыми высокими установками на микроцентрифуге модели 5415C Eppendorf (около 14000×g) до осаждения включений. Супернатант буфера для хранения удаляли и заменяли на 25 мл 100 мМ буфера карбоната натрия, pH 11, в конической пробирке 50 мл. Включения ресуспендировали, используя пипетку и перемешивание на вортексе для полного смешивания. для экстракции целевого белка пробирку оставляли на плавно качающейся платформе при 4°С в течение ночи. Экстракт центрифугировали при 30000×g в течение 30 минут при 4°С, и полученный супернатант концентрировали до 5-кратного изменения объема с помощью Amicon Ultra-15 центрифужного фильтра из восстановленной целлюлозы (с усеченной молекулярной массой 30000; Millipore). Затем буфер для образцов заменяли на 10 мМ CAPS [3-(циклогексамино)1-пропансульфоновая кислота], pH 10, с использованием одноразовых колонок PD-10 (GE Healthcare, Piscataway, NJ).
Солюбилизация и активация трипсином белка телец включения. В некоторых случаях суспензии телец включения DIG-152 из Pf клона DPf108 центрифугировали с самыми высокими установками на микроцентрифуге модели 5415C Eppendorf (около 14000×g) до осаждения включений. Супернатант буфера для хранения удаляли и заменяли на 100 мМ CAPS, pH 11, чтобы получить концентрацию белка около 50 мг/мл. Пробирку оставляли на качалке при комнатной температуре в течение трех часов до полной солюбилизации белка. Добавляли трипсин в количестве, равном от 5% до 10% (вес/вес, в пересчете на исходный вес порошка IB), и в ходе инкубации доводили до расщепления, оставляя на качалке в течение ночи при 4°С или 90-120 минут на качалке при комнатной температуре. Нерастворимый материал удаляли центрифугированием при 10000×g в течение 15 минут, и супернатант загружали в анионобменную колонку MonoQ (10 мм на 10 см). Активированный белок DIG-152 элюировали (как указано в условиях SDS-PAGE, см. ниже) с градиентом 1М NaCl от 0% до 100% в более чем 25 колоночных объемах. Объединяли фракции, содержащие активированный белок, и при необходимости концентрировали до менее, чем 10 мл, с помощью оборудования - центрифужного фильтра Amicon Ultra-15 из восстановленной целлюлозы, как указано выше. Затем материал пропускали через колонку Superdex 200 (16 мм на 60 см) в буфере, содержащем 100 мМ NaCl, 10% глицерина, 0,5% Твин-20 и 1 мМ ЭДТА. Проводили анализ SDS-PAGE чтобы определить элюирование от 65 до 70 мл активированного (ферментативно усеченного) белка. Фракции, содержащие активированный белок, объединяли и концентрировали с использованием центрифужного концентратора, как описано выше.
Электрофорез в геле. Концентрированные белковые препараты были подготовлены для электрофореза путем разведения 1:50 в буфере для образцов NuPAGE® LDS (Invitrogen), содержащем 5 мМ DTT в качестве редуцирующего агента, и нагревали при 95°С в течение 4 минут. Образец загружали в две полосы геля NuPAGE® 4-12% рядом с пятью стандартными полосами BSA в диапазоне от 0,2 мкг до 2 мкг/полосу (для создания стандартной кривой). Применяли напряжение 200 V с использованием подвижного буфера MOPS-SDS (Invitrogen), пока краска-свидетель не достигала основания геля. Гель окрашивали 0,2% Кумасси синим G 250 в 45% метаноле, 10% уксусной кислоте, и обесцвечивали вначале коротким воздействием 45% метанола, 10% уксусной кислотой и затем более тщательно с 7% уксусной кислотой, 5% метанол до очищения фона. После обесцвечивания гель сканировали на визуализаторе BioRad Fluor-S Multilmager. В качестве программного обеспечения применяли Quantity One Software v.4.5.2, чтобы получить вычтенные фоновые значения объема окрашенных полос белка и создать стандартную кривую BSA, которую использовали для расчета концентрации химерного белка DIG-152 в стоковом растворе.
ПРИМЕР 3
Приготовление коровых белков-токсина Cry1Ca и Cry1Ab и выделение мембранных везикул щеточной каймы Spodoptera frugiperda для использования в экспериментах конкурентного связывания
Следующие примеры оценивают конкурентное связывание коровых белков-токсинов Cry1 с предполагаемыми рецепторами в тканях кишечника насекомого. Показано, что 125I меченный коровый белок-токсин Cry1Ca связывается с высокой аффинностью с препаратом мембранных везикул щеточной каймы (BBMV), изготовленным из Spodoptera frugiperda (кукурузной листовой совки), и указанный коровый белок-токсин Cry1Ab не конкурирует за это связывание. В качестве альтернативы показано, что 125I меченный коровый белок-токсин Cry1Ab связывается с высокой аффинностью с BBMV, изготовленными из S. frugiperda, и что коровый белок-токсин Cry1Ca не конкурирует за это связывание.
Очистка белков Cry. Осуществляли экспрессию гена, кодирующего химерный белок DIG-152, который содержит коровый токсин Cry1Ca3 и протоксин Cry1Ab, в экспрессирующем штамме Pseudomonas fluorescens, как описано в Примере 2. Аналогично, экспрессировали ген, кодирующий белок Cry1Ab, в системе экспрессии Pf. Штамм P. fluorescens, который экспрессирует белок Cry1Ab, называется DPf88.
Белки очищали способами из примера 2, и затем расщепляли трипсином для получения активированных коровых токсинов из полноразмерных белков, затем продукты очищали способами, описанными в примере 2. Белковые препараты после обработки трипсином (активированный коровый токсин) имели чистоту >95% и молекулярную массу около 65 кДа, что экспериментально определяли анализом SDS-PAGE. Используемый в изобретении активированный коровый токсин, приготовленный из белка DIG-152, называют коровым белком-токсином Cry1Ca, и активированный коровый токсин, приготовленный из белка Cry1Ab, называют коровым белком-токсином Cry1Ab.
Приготовление и фракционирование солюбилизированных BBMV. Стандартные способы количественного определения белка и электрофорез в SDS-полиакриламидном геле использовали согласно, например, авторам Sambrook et al. (1989) и Ausubel et al. (1995), с обновлениями указанных способов.
Личинок S. frugiperda в последней возрастной стадии не кормили в течение ночи и затем вскрывали после охлаждения на льду в течение 15 минут. Ткань средней кишки удаляли из полости, оставляя после удаления заднюю кишку, соединенную с наружным покровом. Среднюю кишку помещали в 9 X объем ледяного буфера для гомогенизации (300 мМ маннит, 5 мМ этиленгликоль-тетрауксусная кислота (ЭГТА), 17 мМ основания Триса, pH 7,5), с добавлением протеазного ингибирующего коктейля (Sigma-Aldrich P-2714), разведенного согласно рекомендации изготовителя. Ткань гомогенизировали 15 толчками стеклянного тканевого гомогенизатора. Приготовляли BBMV способом преципитации с MgCl2 по Wolfersberger (1993). Коротко, равные объемы 24 мМ раствора MgCl2 в 300 мМ манните смешивали с гомогенатом средней кишки, перемешивали в течение 5 минут и отставляли на льду на 15 мин. Раствор центрифугировали при 2500×g в течение 15 минут при 4°С. Супернатант сохраняли и осадок ресуспендировали в исходный объем 0,5 X разведенного буфера для гомогенизации, и затем снова центрифугировали. Два супернатанта объединяли и центрифугировали при 27000×g в течение 30 минут при 4°С для получения фракции BBMV. Осадок ресуспендировали в буфер для хранения BBMV (10 мМ HEPES, 130 мМ KCl, 10% глицерин, pH 7,4) до концентрации белка около 3 мг/мл. Концентрацию белка определяли с использованием в качестве стандарта бычьего сывороточного альбумина (BSA). Перед замораживанием образцов проводили определение щелочной фосфатазы (маркерный фермент для фракции BBMV), с использованием комплекта для анализа на щелочную фосфатазу QuantiChrom™ DALP-250 (Gentaur Molecular Products, Kampenhout, BE) согласно инструкциям изготовителя. Обычно специфичная активность указанного фермента повышалась в 7 раз по сравнению с аналогичными показателями, выявляемыми в исходном гомогенате фракции средней кишки. Аликвотные количества BBMV разделяли на 250 образцов, мгновенно замораживали в жидком азоте и сохраняли при -80°С.
Электрофорез. Проводили анализ белков SDS-PAGE при редуцирующих условиях (а именно, в 5% β-меркаптоэтанол, BME) и денатурирующих условиях (а именно, с нагреванием в течение 5 минут при 90°С в присутствии 2% SDS). Белки загружали в ячейки с Трис-глицин полиакриламидным гелем от 4% до 20% (BioRad; Hercules, CA) и сепарировали при 200 вт в течение 60 минут. Детекцию полос белка проводили с помощью окрашивания Кумасси бриллиантовым синим R-250 (BioRad) в течение одного часа, и обесцвечивали раствором 5% метанола в 7% уксусной кислоте. Гели визуализировали и анализировали с помощью BioRad Fluro-S MultiImager™. Относительные молекулярные массы полос белка определяли по сравнению с подвижностью белков с известной молекулярной массой, наблюдаемых в образце BenchMark™ Protein Ladder (Life Technologies, Rockville, MD), который загружали в одну ячейку геля.
Иодирование коровых белков-токсинов Cry1Ca или Cry1Ab. Очищенные коровый белок-токсин Cry1 или коровый белок-токсин Cry1Ab подвергали иодированию с помощью шариков для иодирования Pierce (Thermo Fisher Scientific, Rockford, IL). Коротко, два шарика иодирования дважды промывали 500 мкл ФБР (20 мМ фосфат натрия, 0,15М NaCl, pH 7,5), и помещали в центрифужную пробирку объемом 1,5 мл с 100 ФБР. Затем добавляли 0,5 mCi 125I меченного иодида натрия, компоненты оставляли для реакции в течение 5 минут при комнатной температуре, после этого к раствору добавляли 1 мкг корового белка-токсина Cry1Ca (или 1 мкг корового белка-токсина Cry1Ab) и оставляли для проведения реакции в течение дополнительных 3-5 минут. Реакцию завершали добавлением с помощью пипетки раствора из шариков иодирования и вносили раствор на спиновую колонку Zeba™ (Invitrogen), уравновешивали добавлением 50 мМ CAPS, pH 10,0, 1 мМ ДТТ (дитиотретиол), 1 мМ ЭДТА и 5% глицерина. Шарики для иодирования дважды промывали 10 мкл ФБР и раствор для мытья также вносили на обессоливающую колонку Zeba™. Радиоактивный раствор элюировали через спиновую колонку путем центрифугирования при 1000×g в течение 2 мин. Затем 125I-радиомеченый коровый белок-токсин Cry1Ca (или коровый белок-токсин Cry1Ab) диализировали против 50 мМ CAPS, pH 10,0, 1 мМ DTT, 1 мМ ЭДТА и 5% глицерина.
Визуализация. Чистоту в отношении радиоактивности иодированных коровых белков-токсинов Cry1Ca или Cry1Ab определяли с помощью SDS-PAGE и отображения на фосфорном покрытии. Коротко, гели SDS-PAGE высушивали с использованием устройства для сушки геля BioRad согласно инструкциям изготовителя. Высушенные гели визуализировали путем заворачивания их в пленку Майлара (толщиной 12 мкм) и в течение 1 часа оставляли их под воздействием молекулярного люминесцентного экрана с фосфором с длительным послесвечением Molecular Dynamics (35×43 см). Планшеты обрабатывали с использованием Molecular Dynamics Storm, визуализатора с люминесцентным фосфорным покрытием 820 и изображения анализировали с программным обеспечением ImageQuant™.
ПРИМЕР 4
Связывание 125 I меченного корового белка-токсина Cry1 с BBMV из Spodoptera frugiperda
Для определения оптимального количества белка BBMV создавали кривую насыщения, чтобы применять ее анализе связывания с коровыми белками-токсинами Cry1Ca и Cry1Ab. Инкубировали 0,5 нм 125I-радиомеченого корового белка токсина Cry1 в течение 1 часа при 28°С в связывающем буфера (8 мМ NaHPO4, 2 мМ KH2PO4, 150 мМ NaCl, 0,1% BSA pH 7,4) с количеством белка BBMV в диапазоне от 0 мкг/мл к 500 мкг/мл (полный объем 0,5 мл). Затем сепарировали 125I-меченный коровый белок-токсин Cry1, связанный с белками BBMV, от несвязанной фракции, путем отбора в трехкратном исполнении 150 мкл образца реакционной смеси в отдельные центрифужные пробирки 1,5 мл и центрифугированием образцов при 14000×g в течение 8 минут при комнатной температуре. Супернатант аккуратно удаляли и осадок промывали три раза с ледяным связывающим буфером. Дно центрифужной пробирки, содержащей осадок, отсекали, помещали в стеклянную культуральную пробирку 13×75 мм, и каждый из образцов подсчитывали в течение 5 минут в гамма-счетчике. Графически изображали полученные показатели CPM (число импульсов в минуту) минус исходные показатели CPM (реакция без белка BBMV) в зависимости от концентрации белка BBMV. Согласно результатам других исследователей (Luo et al. 1999), была определена оптимальная концентрация белка BBMV для использования в анализе связывания, которая составляла 150 мкг/мл.
ПРИМЕР 5
Анализы конкурентного связывания BBMV из S. frugiperda с коровыми белками-токсинами Cry1Ab и Cry1Ca
Анализы гомологичного и гетерологичного конкурентного связывания проводили с использованием 150 мкг/мл белка BBMV из S. frugiperda и 0,5 нМ 125I-радиомеченого корового белка-токсина Cry1Ca. Концентрации конкурентного не-радиомеченого корового белка-токсина Cry1Ab, добавленного к реакционной смеси, варьировали от 0,045 нм до 300 нМ, и этот белок добавляли в те же точки времени, что и радиоактивный коровый белок-токсин Cry1Ca, чтобы гарантировать действительно конкурентное связывание. Инкубацию проводили в течение 1 часа при 28°С, и количественное измерение 125I-меченного корового белка-токсина Cry1Ca, связанного с BBMV (специфичное связывание), выполняли согласно вышеприведенному описанию. Неспецифичное связывание было отражено при подсчете, полученном в присутствии 1000 нМ не-радиомеченого корового белка-токсина Cry1Ca. Стопроцентным суммарным связыванием считалось количество связывания при отсутствии какого-либо конкурента корового белка-токсина Cry1Ab.
В анализе связывания рецептора с использованием 125I-меченного корового белка-токсина Cry1Ca определяли способность корового белка-токсина Cry1Ab перемещать этот радиомеченый лиганд из его сайта связывания на BBMV из S. frugiperda. Результаты (фиг.1) показывают, что коровый белок-токсин Cry1Ab не перемещает связанный 125I-меченный коровый белок-токсин Cry1Ca из своего рецепторного белка (белков) при таких высоких концентрациях, как 300 нМ (в 600 раз выше концентрации радиоактивного связывающего лиганда). Предполагается, что немеченный коровый белок-токсин Cry1Ca способен перемещать радиомеченый коровый белок-токсин Cry1Ca из своего связывающего белка (белков), демонстрируя сигмоидальную кривую зависимости реакции от дозы с 50% смещением, происходящим при концентрации 5 нМ.
Таким образом показано, что коровый белок-токсин Cry1Ca взаимодействует со связывающим участком BBMV из S. frugiperda, который не связывается с коровым белком-токсином Cry1Ab.
ПРИМЕР 6
Анализ конкурентного связывания BBMV из S. frugiperda с коровыми белками-токсинами Cry1Ca и Cry1Ab
Анализы гомологичного и гетерологичного конкурентного связывания проводили с использованием 150 мкг/мл белка BBMV из S. frugiperda и 0,5 нМ 125I-радиомеченого корового белка-токсина Cry1Ab. Концентрации конкурентного не-радиомеченого корового белка-токсина Cry1Ca, добавленного к реакционной смеси, варьировали от 0,045 нм до 300 нМ, и этот белок добавляли в те же точки времени, что и радиоактивный коровый белок-токсин Cry1Ab, чтобы гарантировать действительно конкурентное связывание. Инкубацию проводили в течение 1 часа при 28°С, и количественное измерение 125I-меченного корового белка-токсина Cry1Ab, связанного с BBMV (специфичное связывание), выполняли согласно вышеприведенному описанию. Неспецифичное связывание было отражено при подсчете, полученном в присутствии 1000 нМ не-радиомеченого корового белка-токсина Cry1Ab. Стопроцентным суммарным связыванием считалось количество связывания при отсутствии какого-либо конкурента корового белка-токсина Cry1Ca.
В анализе связывания рецептора с использованием 125I-меченного корового белка-токсина Cry1Ab определяли способность корового белка-токсина Cry1Ca перемещать этот радиомеченый лиганд из его сайта связывания на BBMV из S. frugiperda. Результаты (фигура 2) показывают, что коровый белок-токсин Cry1Ca не перемещает связанный 125I-меченный коровый белок-токсин Cry1Ab из своего рецепторного белка (белков) при таких высоких концентрациях, как 300 нМ (в 600 раз выше концентрации радиоактивного связывающего лиганда). Предполагается, что немеченный коровый белок-токсин Cry1Ab способен перемещать радиомеченый коровый белок-токсин Cry1Ab из своего связывающего белка (белков), демонстрируя сигмоидальную кривую зависимости реакции от дозы с 50% смещением, происходящим при концентрации 5 нМ.
Таким образом показано, что коровый белок-токсин Cry1Ab взаимодействует со связывающим участком BBMV из S. frugiperda, который не связывается с коровым белком-токсином Cry1Ca.
Ссылки
Finney, D.J. 1971. Probit analysis. Cambridge University Press, England.
Hua, G., L. Masson, J. L. Jurat-Fuentes, G. Schwab, and M.J. Adang. Binding analyses of Bacillus thuringiensis Cry d-endotoxins using brush border membrane vesicles of Ostrinia nubilalis. Applied and Environmental Microbiology 67[2], 872-879. 2001.
LeOra Software. 1987. POLO-PC. A user's guide to probit and logit analysis. Berkeley, CA.
McGaughey, W.H., F. Gould, and W. Gelernter. Bt resistance management. Nature Biotechnology 16[2], 144-146. 1998.
Marcon, P.R.G.C, L.J. Young, K. Steffey, and B.D. Siegfried. 1999. Baseline susceptibility of the European corn borer, Ostrinia nubilalis (Hubner) (Lepidoptera: Pyralidae) to Bacillus thuringiensis toxins. J. Econ. Entomol. 92 (2): 280-285.
Robertson, L.J. and H.K. Preisler. 1992. Pesticide bioassays with arthropods. CRC Press, Boca Ranton, FL.
SAS Institute Inc. 1988. SAS procedures guide, Release 6.03 edition. SAS Institute Inc, Cary, NC.
Stone, B.F. 1968. A formula for determining degree of dominance in cases of monofactorial inheritance of resistance to chemicals. Bull. WHO 38: 325-329.
Van Mellaert, FL, J. Botterman, J. Van Rie, and H. Joos. Transgenic plants for the prevention of development of insects resistant to Bacillus thuringiensis toxins. (Plant Genetic Systems N.V., Belg. 89-401499[400246], 57-19901205. EP. 5-31-1989.
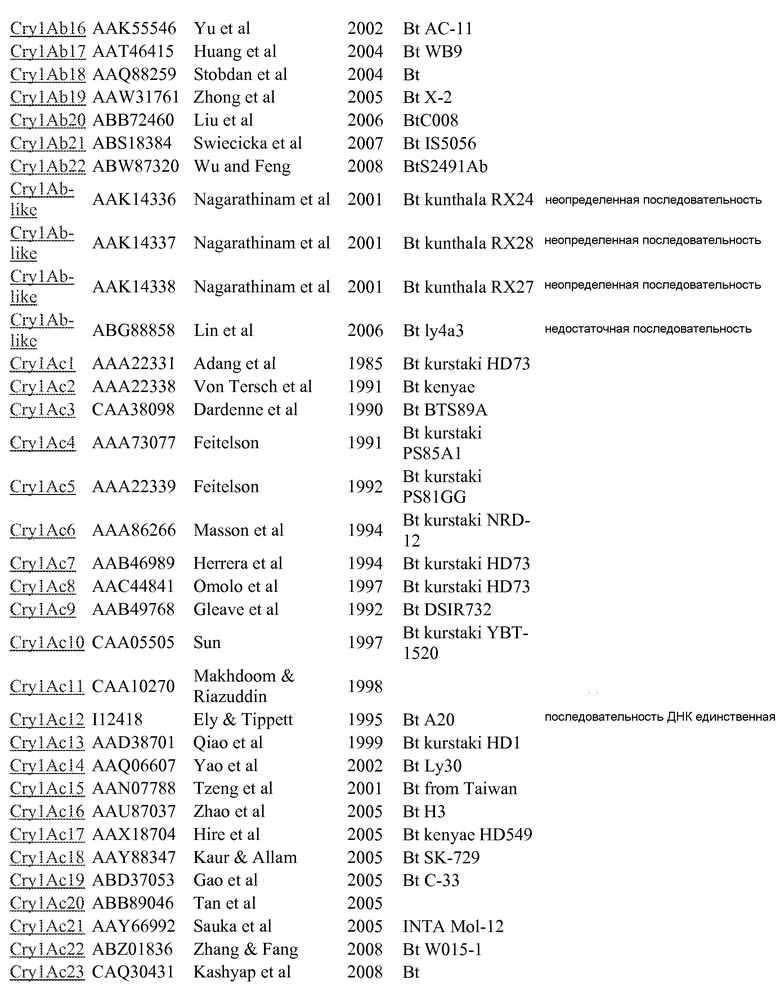

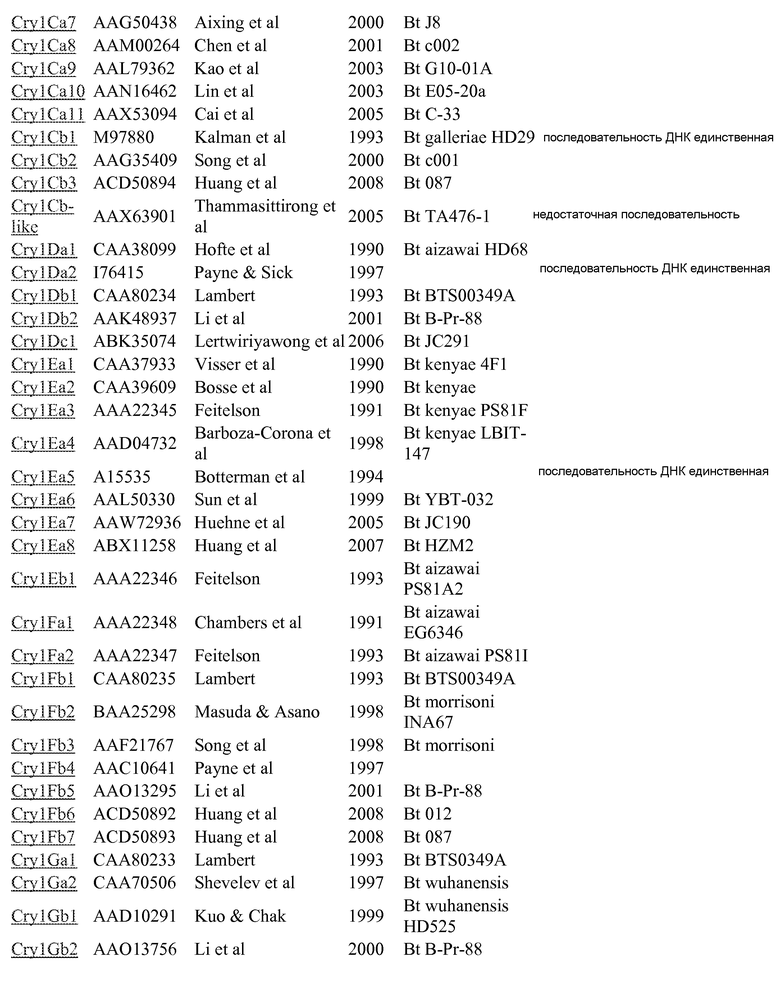








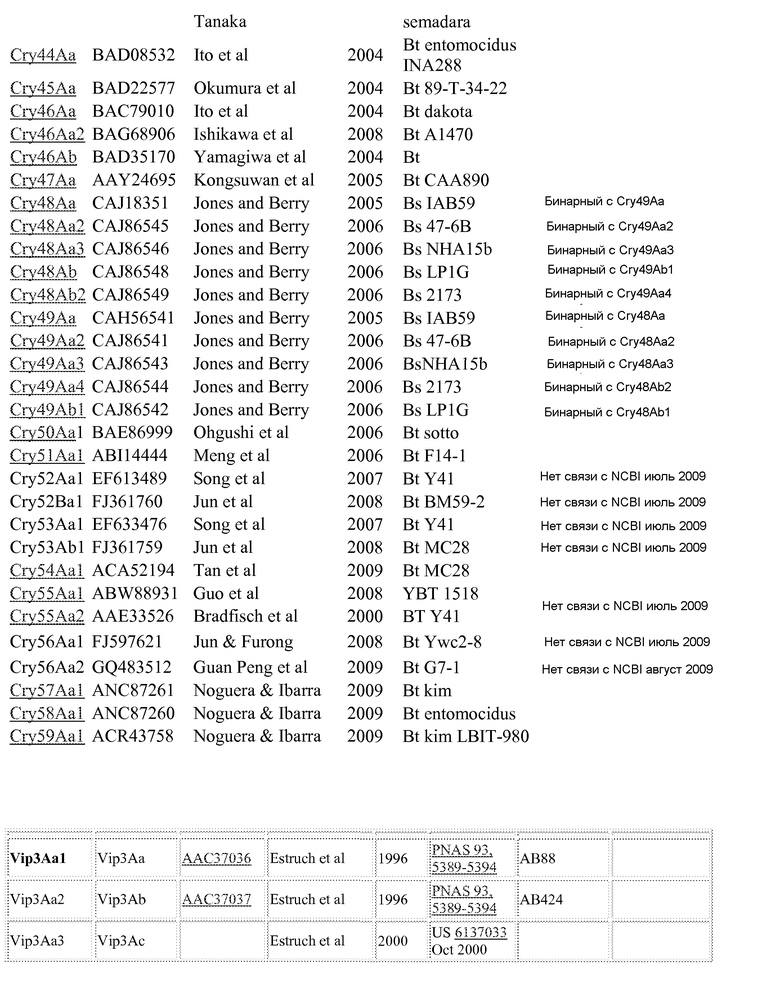
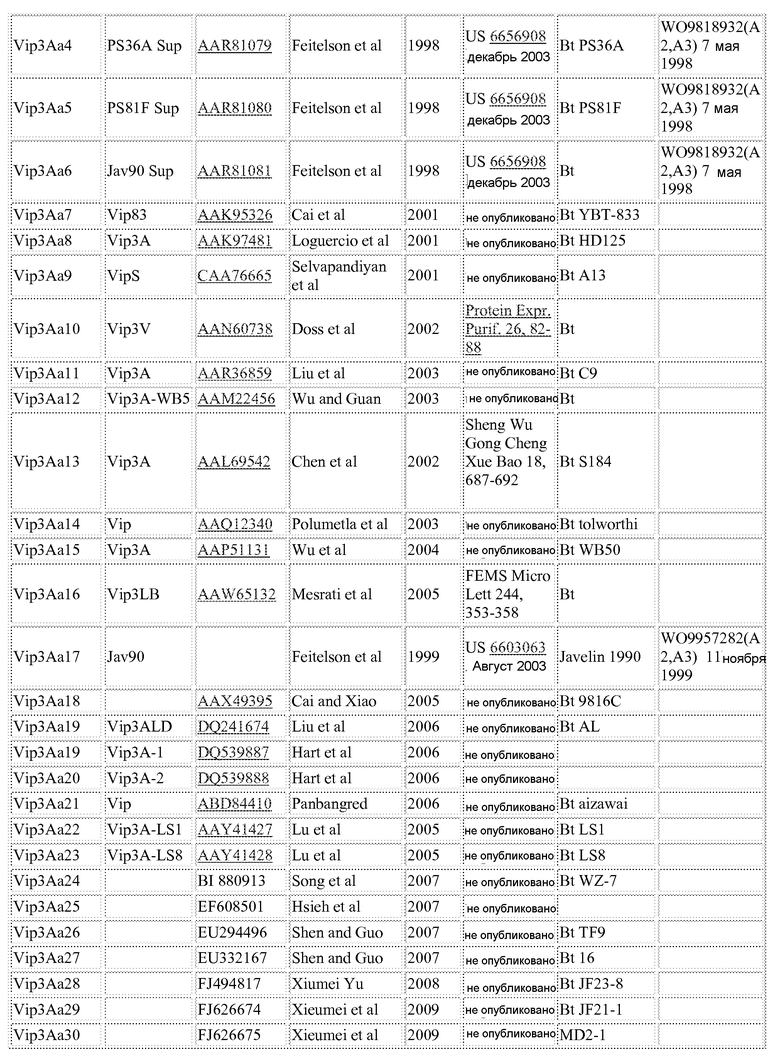
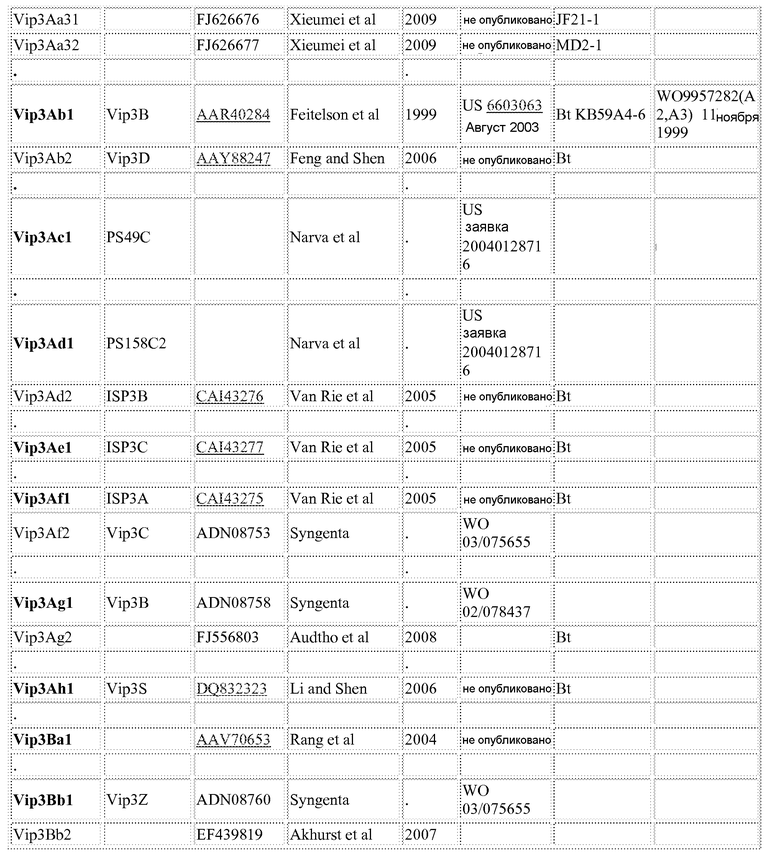
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМБИНИРОВАННОЕ ПРИМЕНЕНИЕ БЕЛКОВ CRY1Ca И CRY1Fa ДЛЯ БОРЬБЫ С РЕЗИСТЕНТНОСТЬЮ У НАСЕКОМЫХ | 2010 |
|
RU2593961C2 |
| ПРИМЕНЕНИЕ Cry1Da В СОЧЕТАНИИ С Cry1Ca ДЛЯ УПРАВЛЕНИЯ УСТОЙЧИВЫМИ НАСЕКОМЫМИ | 2010 |
|
RU2569108C2 |
| ПРИМЕНЕНИЕ Vip3Ab В СОЧЕТАНИИ С Cry1Ca ДЛЯ УПРАВЛЕНИЯ УСТОЙЧИВЫМИ НАСЕКОМЫМИ | 2010 |
|
RU2575084C2 |
| ПРИМЕНЕНИЕ ИНСЕКТИЦИДНОГО КРИСТАЛЛИЧЕСКОГО БЕЛКА DIG3 В КОМБИНАЦИИ С CRY1AB ДЛЯ РЕГУЛИРОВАНИЯ УСТОЙЧИВОСТИ К КУКУРУЗНОМУ МОТЫЛЬКУ | 2012 |
|
RU2624031C2 |
| ПРИМЕНЕНИЕ Cry1Da В СОЧЕТАНИИ С Cry1Be ДЛЯ УПРАВЛЕНИЯ УСТОЙЧИВЫМИ НАСЕКОМЫМИ | 2010 |
|
RU2590592C2 |
| ПРИМЕНЕНИЕ CRY1AB В КОМБИНАЦИИ С CRY1BE ДЛЯ УПРАВЛЕНИЯ РЕЗИСТЕНТНОСТЬЮ НАСЕКОМЫХ | 2010 |
|
RU2583288C2 |
| КОМБИНИРОВАННОЕ ПРИМЕНЕНИЕ БЕЛКОВ Cry1Fa И Cry1Ab ДЛЯ БОРЬБЫ С ОГНЕВКОЙ САХАРНОГО ТРОСТНИКА, РЕЗИСТЕНТНОЙ К Cry-БЕЛКАМ, И ДЛЯ УПРАВЛЕНИЯ РЕЗИСТЕНТНОСТЬЮ НАСЕКОМЫХ НА САХАРНОМ ТРОСТНИКЕ | 2010 |
|
RU2604790C2 |
| КОМБИНИРОВАННОЕ ПРИМЕНЕНИЕ БЕЛКОВ Vip3Ab И Cry1Fa ДЛЯ ВЫРАБАТЫВАНИЯ РЕЗИСТЕНТНОСТИ К НАСЕКОМЫМ | 2010 |
|
RU2607666C2 |
| КОМБИНИРОВАННОЕ ПРИМЕНЕНИЕ БЕЛКОВ Cry1Da И Cry1Fa ДЛЯ ВЫРАБАТЫВАНИЯ РЕЗИСТЕНТНОСТИ К НАСЕКОМЫМ | 2010 |
|
RU2603257C2 |
| КОМБИНИРОВАННОЕ ПРИМЕНЕНИЕ VIP3AB И CRY1AB ДЛЯ РЕГУЛИРОВАНИЯ УСТОЙЧИВЫХ НАСЕКОМЫХ | 2011 |
|
RU2608500C2 |











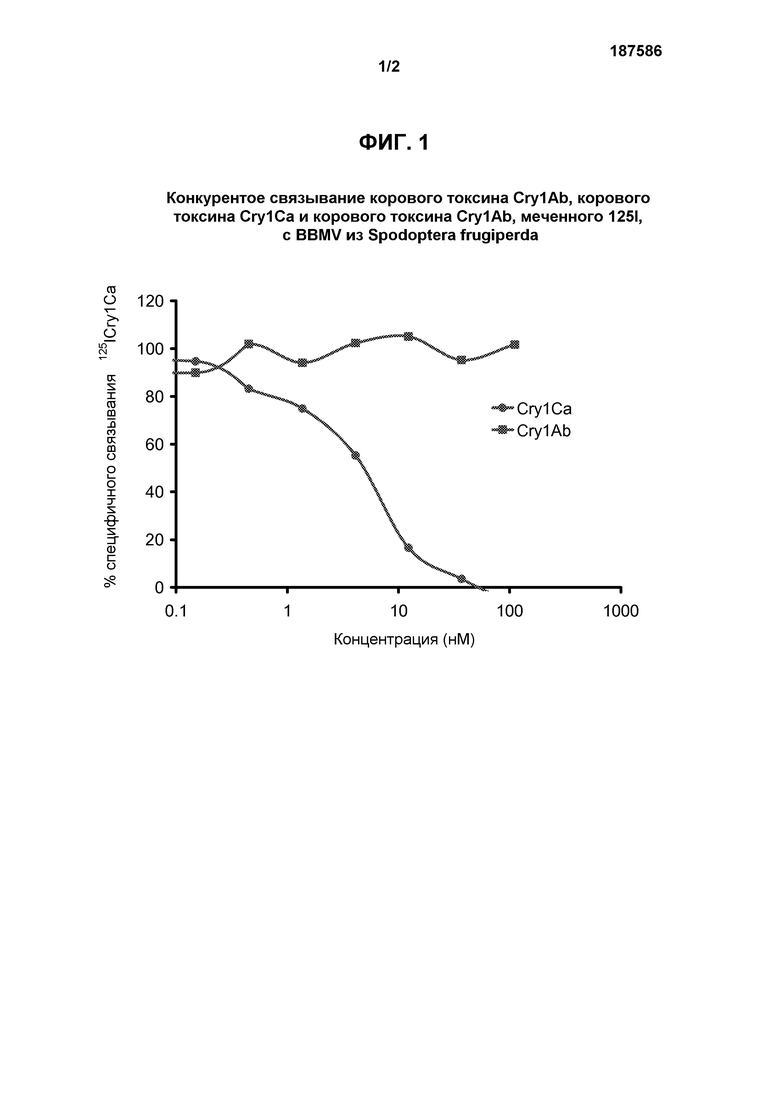
Изобретение относится к области биохимии, в частности к трансгенному растению, обладающему устойчивостью к насекомым-вредителям кукурузной листовой совке и огневке сахарного тростника, содержащему ДНК, которая кодирует Cry1Ca с SEQ ID NO: 2, и ДНК, которая кодирует Cry1Ab с SEQ ID NO: 3, его семени, а также к способу понижения развития устойчивости насекомых огневки сахарного тростника и кукурузной листовой совки к белкам Cry1Ca и Cry1Ab с его использованием. Также раскрыта совокупность растений в поле, содержащая множество вышеуказанных трансгенных растений и не являющихся Bacillus thuringiensis (не-Bt) растения, и смесь семян, содержащая не-Bt семена от не Bt-растений и совокупность вышеуказанных семян. Изобретение также относится к композиции для понижения количества чешуекрылых вредителей, содержащей клетки, экспрессирующие белки Cry1Ab и Cry1Ca. Изобретение позволяет эффективно бороться с кукурузной листовой совкой и огневкой сахарного тростника. 11 н. и 7 з.п. ф-лы, 2 ил., 3 табл., 6 пр.
1. Трансгенное растение, обладающее устойчивостью к насекомым-вредителям кукурузной листовой совке (FAW; Spodoptera frugiperda) и огневке сахарного тростника (SCB; Diatraea saccharalis), содержащее ДНК, которая кодирует инсектицидный белок Cry1Ca с последовательностью SEQ ID NO: 2, и ДНК, которая кодирует инсектицидный белок Cry1Ab с последовательностью SEQ ID NO: 3; где указанное растение выбирают из группы, состоящей из кукурузы, сои и сахарного тростника.
2. Семя растения, обладающего устойчивостью к кукурузной листовой совке и огневке сахарного тростника, по п. 1, содержащее ДНК, которая кодирует инсектицидный белок Cry1Ca с последовательностью SEQ ID NO: 2, и ДНК, которая кодирует инсектицидный белок Cry1Ab с последовательностью SEQ ID NO: 3, указанное семя сеют с целью понижения развития устойчивости кукурузной листовой совки и огневки сахарного тростника к белкам Cry1Ca и Cry1Ab.
3. Трансгенное растение по п. 1, где ДНК, кодирующая инсектицидный белок Cry1Ca, и ДНК, кодирующая инсектицидный белок Cry1Ab, были внедрены в указанное растение.
4. Семя растения, обладающего устойчивостью к кукурузной листовой совке и огневке сахарного тростника, по п. 3, содержащее ДНК, которая кодирует инсектицидный белок Cry1Ca с последовательностью SEQ ID NO: 2, и ДНК, которая кодирует инсектицидный белок Cry1Ab с последовательностью SEQ ID NO: 3, указанное семя сеют с целью понижения развития устойчивости кукурузной листовой совки и огневки сахарного тростника к белкам Cry1Ca и Cry1Ab.
5. Совокупность растений в поле, содержащая растения, не являющиеся Bacillus thuringiensis (не-Bt), не экспрессирующие трансгенный инсектицидный белок, и совокупность трансгенных растений по п. 1, обладающих инсектицидной активностью в отношении кукурузной листовой совки и огневки сахарного тростника, где указанные трансгенные растения содержат ДНК, которая кодирует инсектицидный белок Cry1Ca с последовательностью SEQ ID NO: 2, и ДНК, которая кодирует инсектицидный белок Cry1Ab с последовательностью SEQ ID NO: 3, где указанные не Bt-растения составляют 5-50% всех злаковых растений в указанной совокупности растений, где указанная совокупность растений задерживает развитие устойчивости кукурузной листовой совки и огневки сахарного тростника к белкам Cry1Ca и Cry1Ab, где указанная совокупность растений содержит по меньшей мере одно не Bt-растение.
6. Совокупность растений в поле по п. 5, где указанные не Bt-растения высажены блоками или полосами.
7. Смесь семян, содержащая не-Bt семена от не Bt-растений, не экспрессирующих трансгенные инсектицидные белки, и совокупность семян по п. 4, содержащих ДНК, которая кодирует инсектицидный белок Cry1Ca с последовательностью SEQ ID NO: 2, и ДНК, которая кодирует инсектицидный белок Cry1Ab с последовательностью SEQ ID NO: 3, где указанные не-Bt семена составляют 5-50% всех семян в смеси, где указанную смесь семян сажают для контроля развития устойчивости кукурузной листовой совки и огневки сахарного тростника к белкам Cry1Ca и Cry1Ab, и где указанная смесь семян содержит по меньшей мере одно не-Bt семя.
8. Способ понижения развития устойчивости насекомых огневки сахарного тростника и кукурузной листовой совки к белкам Cry1Ca и Cry1Ab, где указанный способ включает посев семян с получением совокупности трансгенных растений, содержащих ДНК, которая кодирует инсектицидный белок Cry1Ca с последовательностью SEQ ID NO: 2, и ДНК, которая кодирует инсектицидный белок Cry1Ab с последовательностью SEQ ID NO: 3; и приведение указанных насекомых в контакт с указанной совокупностью трансгенных растений.
9. Растение по п. 1 и указанное растение дополнительно содержит ДНК, кодирующую коровый Cry1Fa токсин-содержащий белок.
10. Совокупность растений в поле, содержащая не Bt-растения, не экспрессирующие трансгенный инсектицидный белок, и совокупность трансгенных растений кукурузы по п. 9, обладающих инсектицидной активностью в отношении кукурузной листовой совки и огневки сахарного тростника, где указанные не Bt-растения составляют менее 20% всех злаковых растений в указанной совокупности растений; где указанная совокупность растений задерживает развитие устойчивости кукурузной листовой совки и огневки сахарного тростника к белкам Cry1Ca и Cry1Ab, где указанная совокупность растений содержит по меньшей мере одно не Bt-растение.
11. Совокупность растений в поле, содержащая не Bt-растений, не экспрессирующих трансгенный инсектицидный белок, и совокупность трансгенных растений по п. 9, обладающих инсектицидной активностью в отношении кукурузной листовой совки и огневки сахарного тростника, где указанные не Bt-растения составляют менее 10% всех злаковых растений в указанной совокупности растений; где указанная совокупность растений задерживает развитие устойчивости кукурузной листовой совки и огневки сахарного тростника к белкам Cry1Ca и Cry1Ab, где указанная совокупность растений содержит по меньшей мере одно не Bt-растение.
12. Способ понижения развития устойчивости насекомых огневки сахарного тростника и кукурузной листовой совки к белками Cry1Ca и Cry1Ab, где указанный способ включает посев семян с получением совокупности растений по п. 11 и приведение указанных насекомых в контакт с указанной совокупностью трансгенных растений.
13. Композиция для понижения количества чешуекрылых вредителей, которая содержит клетки, экспрессирующие инсектицидно активное количество как корового токсин-содержащего белка Cry1Ab с последовательностью SEQ ID NO:3, так и корового токсин-содержащего белка Cry1Ca с последовательностью SEQ ID NO:2; где указанные чешуекрылые вредители представляют собой кукурузную листовую совку и огневку сахарного тростника, устойчивых к Cry1Ab и Cry1Ca.
14. Композиция по п. 13, содержащая хозяина, трансформированного для экспрессии как корового токсин-содержащего белка Cry1Ab с последовательностью SEQ ID NO:3, так и корового токсин-содержащего белка Cry1Ca с последовательностью SEQ ID NO:2, где указанный хозяин представляет собой клетку микроорганизма или клетку растения.
15. Способ понижения количества чешуекрылых вредителей, содержащий контакт указанных вредителей или окружающей среды указанных вредителей с инсектицидно активным количеством композиции по п. 13; где указанные чешуекрылые вредители представляют собой кукурузную листовую совку и огневку сахарного тростника, устойчивых к Cry1Ab и Cry1Ca.
16. Совокупность растений в поле по п.5 или 10, где указанные растения занимают площадь более 10 акров.
17. Растение по любому из пп.1, 3 и 9, где указанное растение представляет собой растение кукурузы.
18. Растение по любому из пп.1, 3, 9 или 17, где указанный инсектицидный белок Cry1Ca состоит по существу из SEQ ID NO:2, и указанный инсектицидный белок Cry1Ab состоит по существу из SEQ ID NO:3.
| WO2008145406 A1, 04.12.2008 | |||
| BRAVO A.et al., How to cope with insect resistance to Bt toxins?, 2008, Vol | |||
| Прибор для получения стереоскопических впечатлений от двух изображений различного масштаба | 1917 |
|
SU26A1 |
| Котел для водяного отопления с внутренним перегревателем воды для побуждения циркуляции в сети и с регулятором наружной температуры котла | 1924 |
|
SU573A1 |
| PARDO-LOPEZ L | |||
| et al., Strategies to improve the insecticidal activity of Cry toxins from Bacillus thuringiensis, Peptides | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| Прибор для переработки спирта в газовую смесь для двигателей внутреннего сгорания | 1920 |
|
SU589A1 |
| ФРАГМЕНТ ГИБРИДНОГО ТОКСИНА Bacillus thuringiensis, ОБЛАДАЮЩИЙ ИНСЕКТИЦИДНОЙ АКТИВНОСТЬЮ, КОДИРУЮЩАЯ ЕГО РЕКОМБИНАНТНАЯ ДНК, СПОСОБ БОРЬБЫ С НАСЕКОМЫМИ | 1994 |
|
RU2210593C2 |

