Область техники, к которой относится изобретение
Настоящее изобретение относится к новым рекомбинантным производным стафилокиназы. Более конкретно, изобретение относится к рекомбинантным производным стафилокиназы со значительно пониженной полимеризующей способностью по сравнению со стафилокиназой естественного типа, и бифункциональностью тромболитического и антикоагулирующего агента. Кроме того, изобретение относится к получению этих рекомбинантных производных стафилокиназы и к их применению в качестве тромболитического агента.
Уровень техники
Стафилокиназа естественного типа (Sak) представляет собой протеолитический фермент, образовавшийся под действием лизогенного фага Staphylococcus aureus, и состоит из 136 аминокислотных остатков. Разумеется, по своей природе Sak не является ферментом, но он образует 1:1 комплекс с плазминогеном (plg) в человеческой плазме, и затем этот комплекс активируется в Sak*plm под действием следов плазмина (plm) на поверхности сгустков крови. Комплекс Sak*plm является сильным плазминогенным активатором, переводящим свободную форму plg в активную форму plm, которая, в свою очередь, катализирует разложение фибрина, основной матрицы тромба, и таким образом, приводит к лизису тромба. Sak обладает фибриновой специфичностью при активации plg и действует более эффективно, чем другие тромболитические агенты, растворяя старые тромбы и тромбы, обогащенные тромбоцитами. Таким образом, Sak представляет собой эффективный и специфический тромболитический агент (D. Collen et al., Nature Medicine, 4, 279-284 (1998)). В настоящее время рекомбинантные стафилокиназы производятся несколькими фирмами мира, однако они отличаются друг от друга по структуре генов. Тромболитическая терапия острого инфаркта миокарда (ИМ) с помощью рекомбинантных стафилокиназ, изученная D. Collen (Бельгия), завершила II стадию клинических испытаний. В центре Шиксин (Ченгду, Китай) также завершена I стадия клинических испытаний ИМ, причем эффект был вполне удовлетворительным. В 1994 г. В Шанхайском медицинском университете был сконструирован ген Sak, выполнен высокий уровень экспрессии в Е.coli и завершен пилотный способ. Они были использованы для разрешения клинических испытаний, для лечения острого церебрального инфаркта. Однако Sak, как гетеролитический белок, может обладать сильной антигенностью при введении пациентам. Хотя при клинических испытаниях не наблюдается сильная аллергическая реакция, Sak приводит к высокому титру нейтральных антител у большинства пациентов через две недели после введения (P.J. Declerck et al., Thromb. Haemost, 71, 129-133 (1994)). Более того, при исследовании рекомбинантной стафилокиназы обнаружено, что стафилокиназы стремятся образовать димеры и даже полимеры. Образование полимеров увеличивает иммуногенность стафилокиназы.
В ходе тромболитической терапии часто сочетаются тромболитические лекарственные препараты с антитромбиновыми или антитромбоцитными лекарственными препаратами, такими как гепарин и аспирин, чтобы облегчить тромболиз и предотвратить повторный инфаркт. Значительными являются последние исследования тромболитических вспомогательных лекарственных препаратов. Функциональными последовательностями против агрегации тромбоцитов являются Arg-Gly-Asp (RGD) и Lys-Gly-Asp (KGD). Они конкурентно связываются с рецептором IIb/IIIa гликопротеиновой мембраны, ассоциированной с мембранной агрегацией тромбоцитов, и таким образом предотвращается связывание фибриногена с его рецептором и блокируется повторное образование тромба (W.H.Frishman et al., Am. Heart J., 130, 877-892 (1995); A.J.Nichols et al., Trends Pharmacol. Sci., 13, 413-417 (1992)). При введении последовательности RGD/KGD в кДНК тромболитического агента, например, урокиназы, при соответствующем конформационном расщеплении, полученный продукт экспрессии будет обладать бифункциональностью тромболитического агента и антикоагулянта (J. Smith et al., J. Biol. Chem., 270, 30486-30490 (1995)). Однако при клинических испытаниях отмечено, что тромболитический эффект урокиназы значительно ниже, чем стафилокиназы (50% против 75%). Кроме того, были разработаны различные химические имитаторы на основе последовательности RGD/KGD, такие как тирофибан, ламифибан, лефрадафибан, орбофибан, ксемилофибан, интегринлин и т.п., которые могут блокировать рецептор IIb/IIIa. При их введении в сочетании с тромболитическими агентами вероятность повторного инфаркта может быть существенно снижена (W.H.Frishman et al., Am. Heart J., 130, 877-892 (1995); M. Verstraete et. al. 49, 856-884 (1995)).
Целью изобретения является разработка новых производных стафилокиназы, для которых исключено образование димеров и которые обладают бифункциональностью тромболитического и антикоагулирующего агента, и способа их получения.
Сущность изобретения
Настоящее изобретение относится к новым производным стафилокиназы, для которых исключено образование димеров и которые обладают бифункциональностью тромболитического и антикоагулирующего агента, и к способу их получения и применения. В настоящем изобретении разработаны новые молекулярные структуры Sak с биологической структурной, которые получены методом генной инженерии. Кроме эффективного и специфического тромболитического эффекта, полученные продукты обладают новыми свойствами, такими как низкая способность к полимеризации и агрегации тромбоцитов. Способ получения является простым и безопасным. Выход, степень чистоты и активность продуктов являются по существу такими же, как и для Sak дикого типа.
Краткое описание чертежей
Фиг.1 представляет собой SDS-PAGE фотографию различных концентраций стафилокиназы естественного типа и рекомбинантной стафилокиназы настоящего изобретения, на которой: полоса 1 - это молекулярный стандарт; полоса 2, RGD-Sak (3 мг/мл); полоса 3, RGD-Sak (30 мг/мл); полоса 4, Sak (3 мг/мл); полоса 5, Sak (30 мг/мл).
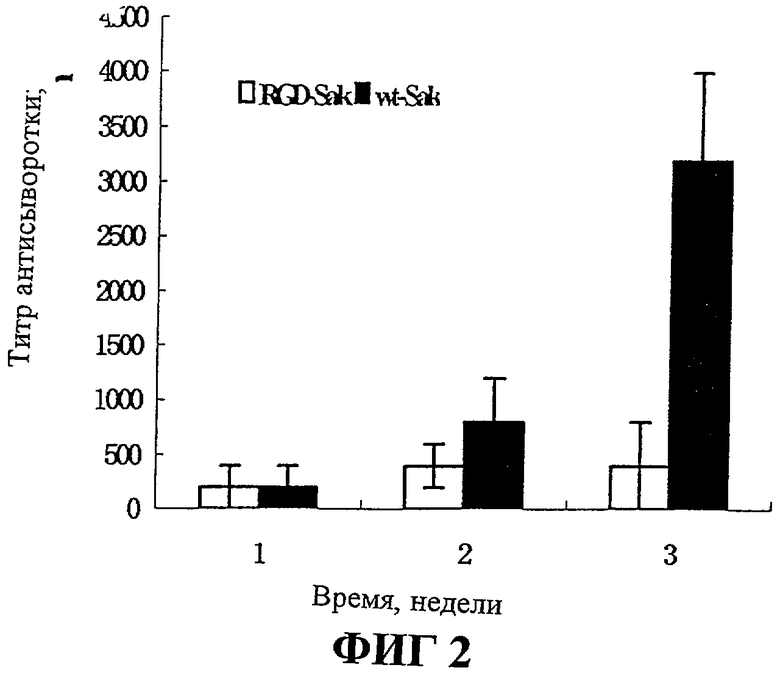
На Фиг.2 представлены результаты испытания чувствительности морских свинок к стафилокиназе естественного типа и к RGD-Sak.
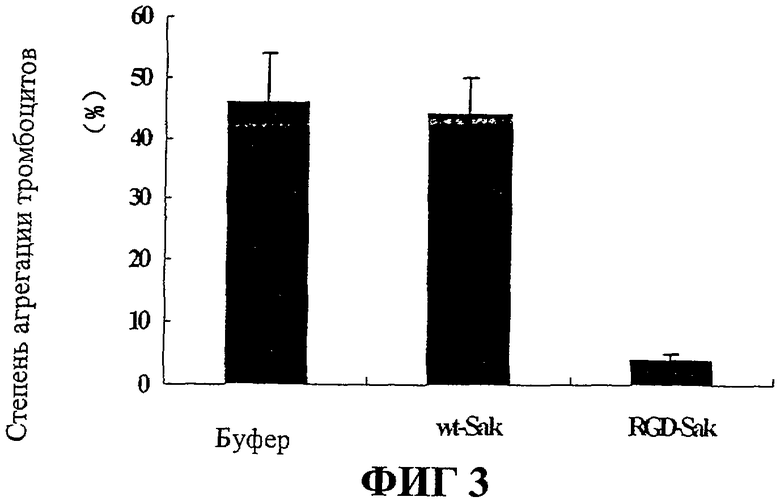
На Фиг.3 представлены результаты испытания активности RGD-Sak против агрегации тромбоцитов.
Подробное описание изобретения
Стафилокиназа естественного типа представляет собой эллипсоидную молекулу, которая включает в себя α-спираль, содержащую 12 остатков аминокислот, перекрытых двумя β-листами, содержащими соответственно 5 и 2 β-витка, простирающихся от аминокислотного остатка 20. Двадцать аминокислот с концевыми аминогруппами свободно выступают наружу, функции которых едва ли можно вывести из кристаллической структуры. Для Sak наблюдается очевидная асимметрия в гидрофобной области и активная область в основном находится в гидрофильной стороне (С.Н.Zhan et al., Acta Cryst., D52, 564-565 (1996); A. Rabijns et al., Nat. Struct. Biol., 5, (1997)). Для определения связывающих областей димера Sak было выполнено моделирование молекулярного объединения с использованием программного обеспечения GRAMM V1.03, разработанного I.A. Vakser (Rockefeller University, USA) на основе кристаллической структуры мономерного Sak по данным рентгеновской дифракции. При использовании одной молекулы Sak в качестве рецептора и другой молекулы Sak в качестве лиганда был выполнен поиск связывающей области в рецепторе Sak относительно лиганда Sak. Были найдены 10 структур комплекса, полученного путем объединения, с общим высоким разрешением параметров объединения, которые рекомендованы автором. Это моделирование осуществлено с использованием графического автоматизированного рабочего стола SGI 02.
Анализ электростатического потенциала и гидрофобности показал, что мономерный Sak является существенно асимметричным в гидрофобной области. Silence и сотр. методом статистического мутагенеза показали, что аминокислоты, определяющие активность, в основном располагаются на гидрофильной стороне (К. Silence et al., J. Biol. Chem., 270, с.27192-27198 (1995)). На гидрофобной стороне Sak имеются две основные гидрофобные области (ГО), которые расположены в области 47-56 (ГО1) остатков и 104-113 (ГО2) соответственно, из которых ГО2 является более гидрофобной. В структурной модели сконструированного димера взаимодействие между гидрофобными областями является очень важным, причем имеются два типа связывания - ГО1-ГО2 и ГО2-ГО2. поскольку ГО1 находится ближе к активной области, эта активная область одной молекулы Sak будет перекрыта, когда две молекулы Sak связываются друг с другом по типу ГО1-ГО2, вероятно, сохраняя активность только одной молекулы Sak. При связывании двух молекул Sak друг с другом по типу ГО2-ГО2, по-видимому, активность не будет сильно изменяться.
Обычно поверхность раздела взаимодействия белковых молекул составляет между 600 и 1300 квадратных Ангстрем (А2), причем каждая молекула предоставляет 10-30 контактирующих остатков. Однако на поверхности раздела имеется так называемое «горячее пятно», на котором лишь 3-5 аминокислот обеспечивают приблизительно 80% энергии связывания. Изменение этих остатков может привести к значительному уменьшению связывающей способности комплекса (В. Li et al.. Science, 270, 1657-1660 (1995)). Таким образом, независимо от конкретного характера связывания димера, образование димеров, вероятно, будет предотвращено, когда изменяются основные связующие остатки ГО2. Фенилаланин (Phe 111), сильно гидрофобная аминокислота, расположен в центральной области ГО2 и находится далеко от активной области. В предпочтительном варианте осуществления изобретения эту кислоту заменяют на сильно полярную аминокислоту, аспарагин (Asp), чтобы нарушить гидрофобный эффект. Ожидается, что мутант сохранит активность. Кроме того, поскольку пептиды последовательности RGD/KGD могут ингибировать агрегацию тромбоцитов, а петлевая область внутри β-листов имеет вполне свободную конформацию, в настоящем изобретении лизин (Lys 109) заменяют на аргинин (Arg), что приводит к последовательности RGD-Sak, или Lys 109 не изменяют, что приводит к последовательности KGD-Sak. Аминокислотная последовательность изобретения RGD-Sak приведена в SEQ ID №1, а аминокислотная последовательность KGD-Sak приведена в SEQ ED №3.
В другом аспекте настоящее изобретение относится к способу получения производных стафилокиназы настоящего изобретения, который включает в себя: приготовление фрагмента ДНК, содержащего, по меньшей мере, часть кодирующей последовательности стафилокиназы, что обеспечивает ее биологическую активность; осуществление in vitro (вне организма) целенаправленного мутагенеза фрагмента ДНК, чтобы заместить один или более кодонов аминокислоты естественного типа на кодон (кодоны) другой аминокислоты (других аминокислот); клонирование мутированного фрагмента ДНК в подходящий вектор; трансформация или трансфекция подходящей клетки хозяина рекомбинантным вектором; культивирование клетки хозяина в условиях, подходящих для экспрессии фрагмента ДНК; и извлечение и очистка желаемых производных стафилокиназы из культуральной среды.
Целенаправленный мутагенез может быть осуществлен путем цепной полимеразной реакции (ЦРП). В предпочтительном варианте осуществления первую амплификацию проводят, используя обратный праймер и мутирующий праймер, с плазмидой pST-Sak в качестве матрицы. После извлечения и очистки методом электрофореза на геле агарозы используют в качестве праймера его амплифицированный двухцепочечный фрагмент, чтобы провести вторую амплификацию с обратным праймером, снова используя плазмиду pST-Sak в качестве матрицы. После очистки полученный фрагмент используется в качестве матрицы в третьей амплификации, вместе с прямым праймером и обратным праймером. Продукт затупляется фрагментом Klenow, расщепляется EcoRI и BamHI, лигируется с плазмидой pUC19, расщепленной EcoRI/BamHI, и трансформируется в штамм JM109 Е.Coli. Положительный клон отбирают путем скрининга в результате анализа расщепления, причем наличие желаемой мутации в ожидаемом положении подтверждается путем анализа нуклеотидной последовательности. Анализ последовательности проводится с помощью анализатора последовательности ABI 377 фирмы Genecore Biotechology Co. Затем ген RGD-Sak удаляется расщеплением EcoRI и BamHI и лигируется в соответствующем сайте вектора экспрессии pLY-4.
прямой праймер: 5'-CGCGAATTCATGTCAAGTTCATTCGAC-3'
обратный праймер: 5'-CGCGGATCCTTATTTCTTTTC-3'
мутирующий праймер (I): 5'-TAAATCTGCGACGACGTCACCACGTTCTGTTATAGG-3' (введен сайт PstI, использованный для конструирования гена RGD-Sak) мутирующий праймер (II): 5'-ATCTGGGACGACGTCACCTTTTTCTG-3' (введен сайт PstI, использованный для конструирования гена KGD-Sak).
Указанным выше способом могут быть получены различные фрагменты ДНК, включающие в себя нуклеотидные последовательности, кодирующие производные стафилокиназы настоящего изобретения. Эти фрагменты включают в себя нуклеотидную последовательность, представленную в SEQ ID №2 или в SEQ ID №4.
Рекомбинантные плазмиды экспрессии получены путем лигатирования фрагментов ДНК изобретения в вектор экспрессии. Настоящее изобретение не ограничивается каким-либо конретным вектором экспрессии при условии, что его можно рекомбинировать с указанными выше фрагментами ДНК с образованием плазмид, подходящих для экспрессии. В предпочтительном варианте осуществления используется прокариотический вектор экспрессии, например, pLY-4, система экспрессии Т7, система экспрессии PL и т.п.
Указанный выше рекомбинантный вектор экспрессии может быть введен в подходящую клетку-хозяина с помощью традиционных методик. Настоящее изобретение не ограничивается какими-либо конретными клетками-хозяина, при условии, что они могут экспрессировать рекомбинантный вектор экспрессии. В предпочтительном варианте осуществления изобретения используется штамм Е.Coli, например, К802, JF1125, серия JM, DH5α и т.п.
Продукт экспрессии настоящего изобретения присутствует в сконструированных клетках в виде тел включения. Желаемый продукт может быть выделен из тел включения и очищен с помощью традиционных методик, например, путем разрушения клеток на прессе French, и сбора тел включения при центрифугировании.
Подробности всех основных лабораторных приемов молекулярной биологии в указанных выше вариантах осуществления описаны в Лабораторном справочнике по молекулярному клонированию.
Кроме того, нуклеиновые кислоты и соответствующие полипептиды настоящего изобретения включают последовательности, которые отличаются от последовательностей, приведенных в SEQ ID №1-4 молчащими мутациями. Эти модификации последовательностей включают, например, нуклеотидные замещения, которые не изменяют аминокислотную последовательность (например, другие кодоны для той же самой аминокислоты или дегенеративные последовательности). Аминокислотные последовательности гомологичных полипептидов могут отличаться от последовательностей SEQ ID №1 или SEQ ID №3 тем, что один или несколько аминокислотных остатков вставляются, удаляются или замещаются другими отличными аминокислотными остатками. Предпочтительно, изменения аминокислот имеют незначительный характер, то есть консервативные замещения аминокислот, которые незначительно влияют на укладку и/или активность белковых молекул; небольшие делеции, обычно длиной приблизительно от 1 до 30 аминокислот; небольшие удлинения на амино- или карбоксильном конце, например, метиониновый остаток на аминоконце; небольшой соединительный пептид, имеющий длину приблизительно 20-25 остатков; или небольшие удлинения, которые могут облегчить очистку путем изменения общего заряда, или которые имеют другие функции, например, поли-гистидиновый тракт, антигенные эпитопы или связывающие домены.
Авторы настоящего изобретения обнаружили, что функции тромболитического агента и антикоагулянта продуктов экспрессии остаются почти неизменными, когда удаляется кодирующая последовательность из 6-10 аминокислот NH2-конца RGD-Sak или KGD-Sak посредством делеционной мутации; продукты экспрессии теряют тромболитическую активность, когда удаляется кодирующая последовательность из 10-15 аминокислот с NH2-конца; указанная бифункциональность продуктов эксперссии сохраняется когда удаляется кодирующая последовательность из 15 иминоксилот и серин (Ser) в положении 16 заменяется на лизин (Lys).
Пример 1. Образец получения и определение свойств RGD-Sak
а. Идентификация r-Sak естественного типа
Стафилокиназа r-Sak естественного типа, полученная в лаборатории заявителя (970923), имеет чистоту выше 98% и сохраняется при -70°С.
Восстановительный и не-восстановительный SDS-PAGE осуществляют по методу Laemmli (см. Лабораторный справочник по молекулярному клонированию).
Загружаемый буферный раствор содержит 0,0625 моль/л Tris-HCl (pH 6,7), 2% додецилсульфата натрия (SDS), 10% глицерина, 5% меркаптоэтанола и 0,001% бромфенолового синего (краситель).
Обработка и загрузка образца: ампулу с лиофилизированным образцом (3 мг/ампула, сохраняется при -70°С в течение более 3 месяцев) растворяют в 3 мл дистиллированной воды. Объем загрузки составляет 10 мкл. Окрашивание геля осуществляют ярко-синим Coomassie R-250 или серебряным красителем.
Сканирование протеиновых полос в геле осуществляют с помощью ImageMaster® VDS (фирма Pharmacia), причем количество протеина, содержащегося в каждой полосе, анализируют, используя прилагаемое программное обеспечение.
После электрофореза гель окрашивают ярко-синим Coomassie, причем появляются плотные полосы в положениях, соответствующих их относительным молекулярным массам приблизительно 15,5 kD, 31 kD, 46 kD и 62 kD (1 kD = 1000 y.e.).
Определение активности с помощью обращенного метода на пластине казеинового геля.
Указанный выше гель последовательно тщательно промывают 2,5% раствором Triton Х-100 и дистиллированной водой, размещают на пластине агарового геля (включает в себя 1% агара), содержащего фибриноген, человеческий плазминоген и тромбин, и выдерживают при 37°С в течение 8 ч. Прозрачные полосы лизиса появляются в положениях, соответствующих указанным выше молекулярным массам, это позволяет предположить, что r-Sak естественного типа стремится образовать при хранении полимеры анти-SDS, которые стабильны и активны.
b. Молекулярное моделирование димера стафилокиназы и образец мутантов
Моделирование осуществляют на автоматизированном графическом рабочем столе SGI 02 с использованием программного обеспечения молекулярного объединения GRAMM V1.03, разработанного I.A.Vakser (Rockefeller University, USA).
Для определения связующей области димера Sak объединение молекул Sak моделируют по программе GRAMM V1.03 на основе кристаллической структуры мономерного Sak по данным рентгеновской дифракции.
В настоящем изобретении замещают фенилаланин (Phe 111) аспарагином, сильно полярной аминокислотой, чтобы нарушить гидрофобное взаимодействие. Предполагается, что мутант сохранит свою активность. Кроме того, поскольку пептиды последовательности RGD могут ингибировать агрегацию тромбоцитов, а петлевая область внутри β-листов имеет вполне свободную конформацию, Lys 109 заменяют на аргинин (Arg), чтобы получить последовательность RGD.
с. Клонирование гена RGD-Sak и конструирование прокариотической плазмиды экспрессии
Используя в качестве матрицы pST-Sak, проводят первую амплификацию с прямым праймером и мутирующим праймером (I), показанными ниже. После выделения и очистки из агарозного геля амплифицированного фрагмента 351bр, последний используют при осуществлении второй амплификации с обращенным праймером, показанными ниже, вновь используя в качестве матрицы pST-Sak. После очистки, используя фрагмент 408bр в качестве матрицы, проводят третью амплификацию с прямым праймером и обращенным праймером. Продукт затупляется фрагментом Klenow, расщепляется EcoRI и BamHI, лигируется с pUC19 и трансформируется. Положительный клон отбирают путем анализа расщепления, причем наличие желаемых мутаций подтверждается путем анализа нуклеотидной последовательности. Анализ последовательности проводится с помощью анализатора последовательности ABI 377 фирмы Genecore Biotechology Co. Затем ген RGD-Sak удаляется путем расщепления EcoRI и BamHI и лигируется в соответствующем сайте вектора экспрессии pLY-4.
прямой праймер: 5'-CGCGAATTCATGTCAAGTTCATTCGAC-3'
обращенный праймер: 5'-CGCGGATCCTTATTTCTTTTC-3'
мутирующий праймер (I): 5'-TAAATCTGCGACGACGTCACCACGTTCTGTTATAGG-3' (введен сайт PstI).
Все ферменты, модифицирующие нуклеофильные кислоты, приобретаются на фирмах GIBCO BRL и Promega. Олигонуклеотиды синтезированы в Группе синтеза ДНК в John Hopkins University (USA).
Штамм JM109 E.Coli и pUC19 имеется в распоряжении в лаборатории заявителя. Штамм JF1125 E.Coli и прокариотический вектор экспрессии pLY-4 любезно предоставлены профессором Цзин-Хуан Лю из Института биохимии Китайской академии наук (Китай). Плазмида pST-Sak сконструирована в лаборатории заявителя (Китайский патент №94 1 12105.4).
Желаемый ген лигируют в pLY-4 и трансформируют в штамм JF1125 E.Coli. Плазмиду получают и идентифицируют путем соответствующего анализа расщепления. Получают определенный фрагмент, что подтверждает положительный клон.
Штамм JF1125 E.Coli, трансформированный с помощью pLY-4-RGD-Sak, культивируют в культуральной среде М9СА при 30°С до достижения значения оптической плотности (ОП 600), равного 0,6. Затем повышают температуру до 42°С и культивирование продолжают еще 3 часа, чтобы вызвать экспрессию. Продукт экспрессии анализируют путем SDS-PAGE. После электрофореза половину продукта окрашивают ярко-синим Coomassie. Наблюдается плотная полоса в положении, соответствующем молекулярной массе около 15,5 kD на трассе продукта лизиса индуцированных бактериальных клеток, которые составляют более чем 50% от суммы протеинов бактериальных клеток, по оценке путем сканирования. Другую половину размещают на пластине агарового геля после удаления додецилсульфата натрия и выдерживают при 37°С в течение нескольких часов. Появляется прозрачная область, соответствующая молекулярной массе 15,5 kD. Другими словами, в этом положении казеин разлагается, что позволяет предположить, RGD-Sak имеет фибринолитическую активность. После разрушения и центрифугирования клеток обнаружено, что полоса 15,5 kD в основном присутствует в осадке и едва наблюдается в супернатанте; это указывает, что продукт экспрессии существует в виде тел включения.
d. Индуцибельная экспрессия в сконструированных штаммах
Сконструированные штаммы подвергают скринингу на высокий уровень экспрессии (например, рекомбинантный протеин экспрессии составляет более чем 50% от суммы протеинов клетки). С выбранным штаммом проводят ферментацию низкой плотности в 10-литровом ферментере. После 3 часов термически индуцированного культивирования, клетки раскручиваются, промываются в фосфатно-солевом буфере и сохраняются при -70°С до использования. Из 10 л культуры получают 80 г влажных клеток. Эти влажные клетки суспендируют в фосфатном буфере, разрушают в гомогенизаторе высокого давления и центрифугируют. Отбирают образцы для SDS-PAGE. Результаты показывают, что желаемый протеин присутствует на трассе осадка, с плотно окрашенной полосой в положении, соответствующем молекулярной массе 15,5 kD, причем какое-либо окрашивание едва наблюдается в положении, соответствующем жидкости над осадком; это позволяет предположить, что RGD-Sak в основном существует в виде тел включения.
е. Выделение, солюбилизация и ренатурация тел включения
После разрушения путем сжатия 80 г клеток сконструированного штамма подвергают центрифугированию при скорости 10000 об/мин и получают 20 г тел включения. После промывки в фосфатном буфере (ФБ, 0,05 моль/л) тела включения растворяют в растворе, содержащем 0,1 моль/л ФБ с рН, равным 5,0, 6 моль/л гидрохлорида гуанидина, 0,5% β-меркаптоэтанола, и выдерживают при комнатной температуре, пока раствор не станет прозрачным. После ультрацентрифугирования при скорости 30000 об./мин осадок отбрасывают, а супернатант разбавляют для ренатурации в ФБ (0,1 моль/л, рН 5,0) и 0,5% β-меркаптоэтанола.
f. Хроматография на колонках Sephadex G-10 и S-Sepharose FF
После концентрирования путем ультрафильтрации (молекулярная масса 1000, Millipore), супернатант фильтруют через колонку Sephadex G-10. Фильтрат подают на колонку S-Sepharose FF, предварительно уравновешенную десятикратным объемом (от объема набивки) фосфатного буфера (0,1 моль/л, рН 5,0). Для контроля скорости потока и детектирования пика протеина используют хроматограф (фирмы Waters). После загрузки продукта колонку промывают до базовой линии (отсутствие сигнала) фосфатным буфером и элюируют продукт раствором хлорида натрия с градиентом 0-1 моль/л. Отбирают элюированные фракции. Распределение желаемого протеина анализируют методом SDS-PAGE и определяют концентрацию по методу Брэдфорда (используемые реагенты закупают на фирме Bio-Rad).
Все операции хроматографического анализа являются обычными для специалистов в этой области техники.
g. Идентификация чистоты и определение молекулярной массы
Образец анализируют методом SDS-PAGE в соответствии с Лабораторным справочником молекулярного клонирования. После окрашивания геля ярко-синим Coomassie R-250 его сканируют на приборе Imagemaster VDS (фирма Pharmacia), чтобы определить степень чистоты и молекулярную массу протеина. Соответственно было найдено, что чистота составляет более 95%, а молекулярная масса равна приблизительно 15,5 kD.
h. Определение биологической активности
Для определения биологической активности проводят анализы числа бляшек на казеиновом геле (A.G. Pipemo et al., J. Exp. Med. 48(1), 223-234 (1978)) и по методу хромогенного субстрата (H.R. Lijnen et al., J. Biol. Chem., 266, 11826-11832 (1991)). Удельная активность составляет около 90000-100000 HU/мг. Для определения единиц см. работу O.Q. Tang et al., Drug Biotechnol, (китайский), 4(1), 1-4 (1997).
i. Определение констант Км и Ккат для комплексов Sak-плазмин и RGD-Sak-плазмин
Выдерживают Sak или RGD-Sak (2 мкмоль/л) с 2 мкмоль/л плазминогена соответственно в 0,1 моль/л фосфатного буфера (рН 5,0) при рН 7,4 и температуре 37°С в течение 30 мин для образования комплексов с плазмином. Затем отбирают каталитическое количество (5 нмоль/л) комплекса для осуществления взаимодействия в течение 0-10 мин в реакционных системах, указанных ниже, в 0,1 моль/л ФБ при рН 7,4 и температуре 37°С, с регистрацией оптической плотности (при 405 нм) каждые 30 с. Приведенные концентрации плазминогена являются усредненными по данным трех анализов.
Активация плазминогена комплексом RGD-Sak-плазмин описывается уравнением Михаэлиса-Ментена (табл.1b).
j. Определение способности к полимеризации
В качестве контроля используют Sak естественного типа. Образцы растворяют в физиологическом растворе. Испытания проводят при двух концентрациях протеина 30 мг/мл (высокая) и 3 мг/мл (низкая). Растворы выдерживают при комнатной температуре. Пробы отбирают каждые 24 ч и анализируют путем электрофореза.
При обеих концентрациях протеина способность к полимеризации RGD-Sak была значительно ниже, чем у Sak естественного типа (фиг.1).
k. Испытание чувствительности на морских свинках
Для испытания чувствительности рекомбинантную стафилокиназу естественного типа, а также мутант RGD-Sak растворяют в стерильном физиологическом растворе при концентрации 2500 ед./мл. Для каждого введения берут интактные ампулы, чтобы приготовить стерильным образом свежие растворы. Здоровых морских свинок (20) случайным образом распределяют на две группы, по 10 животных в каждой. Этим морским свинкам внутрибрюшинно вводят 0,15 мг/кг r-Sak или RGD-Sak через день по три раза. Первое и второе внутривенное воздействие дозой 0,3 мг/кг проводят соответственно на 14 и 21 сутки. Двум здоровым морским свинкам, которым не вводили инъекцию, вводят внутривенно 0,3 мг/кг указанных выше образцов и наблюдают наличие аналогичной реакции, чтобы исключить фармакологические и патологические помехи, связанные с образцами.
В группе с инъекцией r-Sak естественного типа: для 8 морских свинок наблюдалась положительная реакция класса IV и для 2 морских свинок наблюдалась положительная реакция класса II.
В группе с инъекцией RGD-Sak: у 2 морских свинок наблюдалась положительная реакция класса I, а у других животных отсутствовала заметная реакция.
Реакция класса I: мягкий кашель
Реакция класса II: приступы кашля, дрожь
Реакция класса III: сильная дрожь
Реакция класса IV: конвульсия, спазм, недержание фекалий и мочи, шок со смертельным исходом.
Содержание антител в сыворотке морских свинок, иммунизированных в течение 1-3 недель, определяют методом иммуноферментного твердофазного анализа (ELISA), в котором в качестве антигенов используют wt-Sak и RGD-Sak соответственно. В первую неделю количество антител относительно любого антигена было мало. Во вторую неделю количество антител группы wt-Sak (n=10) увеличивается до 1:800, в то время как количество антител группы RGD-Sak (n=10) увеличивается до 1:400. В третью неделю количество антител группы wt-Sak увеличивается до 1:3200, в то время как в группе RGD-Sak остается равным 1:400. Таким образом, иммуногенность RGD-Sak значительно снижается по сравнению с иммуногенностью wt-Sak (фиг.2В).
Приведенные выше результаты указывают, что иммуногенность RGD-Sak значительно снижается по сравнению с иммуногенностью wt-Sak.
l. Анализ ингибирования агрегации тромбоцитов
Медленно центрифугируют (ускорение 150 g, 10 мин) свежую кровь с добавкой 1/10 объема раствора (110 ммоль/л) антикоагулянта - цитрата натрия, чтобы получить плазму, обогащенную тромбоцитами (ПОТ). Добавляют RGD-Sak к ПОТ до окончательной концентрации 2 мкмоль/л, и смесь выдерживают 2 мин при 37°С и непрерывном перемешивании. Затем добавляют аденозиндифосфат (АДФ) до окончательной концентрации 2 мкмоль/л в качестве индуктора. Степень агрегации тромбоцитов определяют в течение 5 минут в двухканальном аппарате агрегации тромбоцитов (CHRONO-LOG560). В качестве контрольных используют образцы r-Sak естественного типа (2 мкмоль/л) и физиологический раствор. Аденозиндифосфат закупают на фирме Sigma, прочие реагенты аналитической чистоты произведены в Китае.
Найдено, что степень агрегации в группе RGD-Sak (5%±2%, n=3) значительно ниже, чем агрегация в группах с r-Sak (58%±3%, n=3) и физиологическим раствором (59%±3%, n=3). Это позволяет предположить, что RGD-Sak обладает сильным потенциалом при ингибировании агрегации тромбоцитов, вызванной АДФ (фиг.3).
m. Тромболитический анализ на животных
Тромболитический анализ на животных проводят с RGD-Sak, полученным, в настоящем изобретении, проверяя, что RGD-Sak сохраняет те же самые тромболитические свойства, что и Sak естественного типа. Для введения RGD-Sak растворяют в стерильном физиологическом растворе, как описано в разделе k этого же примера.
(i) Лечение тромбоза бедренной артерии экспериментальных кроликов RGD-Sak
В каждой группе, обрабатываемой RGD-Sak, обрабатываемой Sak естественного типа и в контрольной группе, без обработки, содержится 6 животных. С помощью артериографии показано, что до лечения средний сегмент бедренной артерии не был видим. При повторном фотографировании через 60 минут после внутривенной инъекции 0,1 мг/кг RGD-Sak бедренная артерия становится полностью заполненной, с восстановлением цикла крови, что согласуется с эффектом в группе Sak естественного типа, в то время как в контрольной группе бедренная артерия не становится заполненной.
(ii) Лечение гифемы экспериментальных кроликов с помощью RGD-Sak
В каждой группе, обрабатываемой RGD-Sak, обрабатываемой Sak естественного типа и в контрольной группе, без обработки, содержится 6 животных. Через 4 ч после внутриглазной инъекции 10-20 мкг RGD-Sak наблюдается лизис тромба гифемы, причем красные кровяные клетки осаждаются и образуют поверхность раздела с водной внутриглазной жидкостью. Внутриглазная кровяная опухоль удаляется через 24 ч. Это согласуется с эффектом в группе Sak естественного типа. Однако в контрольной группе гифема существенно не изменяется.
(iii) Тромболитическая терапия с помощью RGD-Sak является безопасной и эффективной при лечении острого инфаркта миокарда у экспериментальных собак, вызванного тромбозом коронарной артерии. В экспериментальной группе, состоящей из 6 животных, собаки получали дозу RGD-Sak (0,3 мг/кг веса тела) с помощью внутривенной инфузии; в контрольной группе, состоящей из 6 животных, собаки получали физиологический раствор, вместо RGD-Sak, с помощью внутривенной инфузии. До и после ввода дозы проводят коронарную артериографию. До ввода дозы обнаружено, что левое переднее нисходящее ответвление коронарной артерии является незаполненным, или не полностью заполненным у животных обеих групп. При проведении артериографии через 30 минут после лечения обнаружено, что левое переднее нисходящее ответвление вновь заполняется, и животные в экспериментальной группе выживают. Что касается контрольной группы, то в не полностью заполненной артерии отсутствуют заметные изменения, и животные умирают через несколько часов.
(iv) Тромболитическая терапия с помощью RGD-Sak является безопасной и эффективной при лечении острого церебрального инфаркта у экспериментальных свиней, вызванного тромбозом внутричерепной артерии. В экспериментальной группе, состоящей из 6 животных, свиньи получали дозу RGD-Sak (0,2 мг/кг веса тела) с помощью внутривенной инфузии; в контрольной группе, состоящей из 6 животных, свиньи получали физиологический раствор, вместо RGD-Sak, с помощью внутривенной инфузии. До и после ввода дозы проводят внутричерепную DSA артериографию. До ввода дозы обнаружено, что некоторые внутричерепные артерии являются не полностью заполненными у животных обеих групп. При проведении ангиографии через 30 минут после лечения обнаружено, что внутричерепные артерии вновь заполняются, и животные в экспериментальной группе выживают. Что касается контрольной группы, то в не полностью заполненных внутричерепных артериях отсутствуют заметные изменения, и животные умирают спустя несколько дней.
Пример 2. Образец получения и определение свойств KGD-Sak
а. Идентификация r-Sak естественного типа
Стафилокиназа r-Sak естественного типа, полученная в лаборатории заявителя (970923), имеет чистоту выше 98% и сохраняется при -70°С.
Восстановительный и не-восстановительный SDS-PAGE осуществляют по методу Laemmli (см. Лабораторный справочник по молекулярному клонированию).
Загружаемый буферный раствор содержит 0,0625 моль/л Tris-HCl (pH 6,7), 2% додецилсульфата натрия (SDS), 10% глицерина, 5% меркаптоэтанола и 0,001% бромфенолового синего (краситель).
Обработка и загрузка образца: ампулу с лиофилизированным образцом (3 мг/ампула, сохраняется при -70°С в течение более 3 месяцев) растворяют в 3 мл дистиллированной воды. Объем загрузки составляет 10 мкл.
Окрашивание геля осуществляют ярко-синим Coomassie R-250 или серебряным красителем.
Сканирование протеиновых полос в геле осуществляют с помощью ImageMaster® VDS (фирма Pharmacia), причем количество протеина, содержащегося в каждой полосе, анализируют, используя прилагаемое программное обеспечение.
После электрофореза гель окрашивают ярко-синим Coomassie, причем появляются плотные полосы в положениях, соответствующих их относительным молекулярным массам приблизительно 15,5 kD, 31 kD, 46 kD и 62 kD (1 kD = 1000 y.e.).
Определение активности с помощью обращенного метода на пластинах казеинового геля
Указанный выше гель последовательно тщательно промывают 2,5% раствором Triton Х-100 и дистиллированной водой, размещают на пластине агарового геля (включает в себя 1% агара), содержащего фибриноген, человеческий плазминоген и тромбин, и выдерживают при 37°С в течение 8 ч. Прозрачные полосы лизиса появляются в положениях, соответствующих указанным выше молекулярным массам, это позволяет предположить, что r-Sak естественного типа стремится образовать при хранении полимеры анти-SDS, которые стабильны и активны.
b. Молекулярное моделирование димера стафилокиназы и образца мутантов
Моделирование осуществляют на автоматизированном графическом рабочем столе SGI 02 с использованием программного обеспечения молекулярного объединения GRAMM V1.03, разработанного I.A.Vakser (Rockefeller University, USA).
Для определения связывающей области димера Sak объединение молекул Sak моделируют по программе GRAMM V1.03, на основе кристаллической структуры мономерного Sak по данным рентгеновской дифракции.
В настоящем изобретении замещают фенилаланин (Phe 111) аспарагином, сильно полярной аминокислотой, чтобы нарушить гидрофобное взаимодействие. Предполагается, что мутант сохранит свою активность. Кроме того, поскольку пептиды последовательности KGD могут ингибировать агрегацию тромбоцитов, а петлевая область внутри β-листов имеет вполне свободную конформацию, полагают, что тромболитическая активность не будет затронута.
с. Клонирование гена KGD-Sak и конструирование прокариотической плазмиды экспрессии
Используя в качестве матрицы pST-Sak, проводят первую амплификацию с прямым праймером и мутирующим праймером (II), показанными ниже. После выделения и очистки из агарозного геля амплифицированного фрагмента 351bр, последний используют при осуществлении второй амплификации с обращенным праймером, показанными ниже, вновь используя в качестве матрицы pST-Sak. После очистки, используя фрагмент 408bр в качестве матрицы, проводят третью амплификацию с прямым праймером и обращенным праймером. Продукт затупляется фрагментом Klenow, расщепляется EcoRI и BamHI, лигируегся с pUC19 и трансформируется. Положительный клон отбирают путем анализа расщеплением, причем наличие желаемых мутаций подтверждается путем анализа нуклеотидной последовательности. Анализ последовательности проводится с помощью анализатора последовательности ABI 377 фирмы Genecore Biotechology Co. Затем ген KGD-Sak удаляется путем расщепления EcoRI и BamHI и лигируется в соответствующем сайте вектора экспрессии pLY-4.
прямой праймер: 5'-CGCGAATTCATGTCAAGTTCATTCGAC-3'
обращенный праймер: 5'-CGCGGATCCTTATTTCTTTTC-3'
мутирующий праймер (II): 5'-ATCTGGGACGACGTCACCTTTTTCTC-3' (введен сайт PstI).
Все ферменты, модифицирующие нуклеиновые кислоты, приобретаются на фирмах GIBCO BRL и Promega. Олигонуклеотиды синтезированы в Группе синтеза ДНК в John Hopkins University (USA).
Штамм JM109 E.Coli и pUC19 имеется в распоряжении в лаборатории заявителя. Штамм JF1125 E.Coli и прокариотический вектор экспрессии pLY-4 любезно предоставлены профессором Цзин-Хуан Лю из Института биохимии. Китайской академии наук (Китай). Плазмида pST-Sak сконструирована в лаборатории заявителя (см. Китайский патент).
Желаемый ген лигируют в pLY-4 и трансформируется в штамм IF 1125 E.Coli. Плазмиду получают и идентифицируют путем соответствующего анализа расщепления. Получают характерный фрагмент, что подтверждает положительный клон.
Штамм JF1125 E.Coli, трансформированный с помощью pLY-4-KGD-Sak, культивируют в культуральной среде М9СА при 30°С до достижения значения оптической плотности (ОП 600), равного 0,6. Затем повышают температуру до 42°С и культивирование продолжают еще 3 часа, чтобы вызвать экспрессию. Продукт экспрессии анализируют путем SDS-PAGE. После электрофореза половину продукта окрашивают ярко-синим Coomassie. Наблюдается плотная полоса в положении, соответствующем молекулярной массе около 15,5 kD на трассе продукта лизиса индуцированных бактериальных клеток, которые составляют более чем 50% от суммы протеинов бактериальных клеток, по оценке путем сканирования. Другую половину размещают на пластине агарового геля после удаления додецилсульфоната натрия и выдерживают при 37°С в течение нескольких часов. Появляется прозрачная область, соответствующая молекулярной массе 15,5 kD. Другими словами, в этом положении казеин разлагается, что позволяет предположить, KGD-Sak имеет фибринолитическую активность. После разрушения и центрифугирования клеток обнаружено, что полоса 15,5 kD в основном присутствует в осадке и едва наблюдается в супернатанте; это указывает, что продукт экспрессии существует в виде тел включения.
d. Индуцибельная экспрессия в сконструированных штаммах
Сконструированные штаммы подвергают скринингу на высокий уровень экспрессии (например, рекомбинантный протеин экспрессии составляет более чем 50% от суммы протеинов клетки). С выбранным штаммом проводят ферментацию низкой плотности в 10-литровом ферментере. После 3 часов термически индуцированного культивирования, клетки раскручиваются, промываются в фосфатно-солевом буферном растворе и сохраняются при -70°С до использования. Из 10 л культуры получают 80 г влажных клеток. Эти влажные клетки суспендируют в фосфатном буфере, разрушают в гомогенизаторе высокого давления и центрифугируют. Отбирают образцы для SDS-PAGE. Результаты показывают, что желаемый протеин присутствует на трассе осадка, с плотно окрашенной полосой в положении, соответствующем молекулярной массе 15,5 kD, причем какое-либо окрашивание едва наблюдается в положении, соответствующем жидкости над осадком; это позволяет предположить, что KGD-Sak в основном существует в виде тел включения.
е. Выделение, солюбилизация и ренатурация тел включения
После разрушения путем сжатия 80 г клеток сконструированного штамма подвергают центрифугированию при скорости 10000 об./мин и получают 20 г тел включения. После промывки в фосфатном буфере (ФБ, 0,05 моль/л, рН=5,2) тела включения растворяют в растворе, содержащем 0,1 моль/л ФБ с рН, равным 5,2, 6 моль/л гидрохлорида гуанидина, 0,5% β-меркаптоэтанола, и выдерживают при комнатной температуре, пока раствор не станет прозрачным. После ультрацентрифугирования при скорости 30000 об./мин осадок отбрасывают, а жидкость над осадком разбавляют для повторной натурализации в ФБ (0,1 моль/л, рН 5,0) и 0,5% β-меркаптоэтанола.
f. Хроматография на колонках Sephadex G-10 и S-Sepharose FF
После концентрирования путем ультрафильтрации (молекулярная масса 1000, Millipore), жидкость над осадком фильтруют через колонку Sephadex G-10. Фильтрат подают на колонку S-Sepharose FF, предварительно уравновешенную 10 кратным объемом (от объема набивки) фосфатного буфера (0,1 моль/л, рН 5,0). Для контроля скорости потока и детектирования пика протеина используют хроматограф (фирмы Waters). После загрузки продукта колонку промывают до базовой линии (отсутствие сигнала) фосфатным буфером и элюируют продукт раствором хлорида натрия с градиентом 0-1 моль/л. Отбирают элюированные фракции. Распределение желаемого протеина анализируют методом SDS-PAGE и определяют концентрацию по методу Брэдфорда (используемые реагенты закупают на фирме Bio-Rad).
Все операции хроматографического анализа являются обычными для специалистов в этой области техники.
g. Идентификация чистоты и определение молекулярной массы
Образец анализируют методом SDS-PAGE в соответствии с Лабораторным справочником молекулярного клонирования. После окрашивания геля ярко-синим Coomassie R-250 его сканируют на Imagemaster VDS (фирма Pharmacia), чтобы определить степень чистоты и молекулярную массу протеина. Соответственно было найдено, что чистота составляет более 95%, а молекулярная масса равна приблизительно 15,5 kD.
h. Определение биологической активности
Для определения биологической активности проводят анализы числа бляшек на казеиновом геле (A.G. Pipemo et al., J. Exp. Med. 48(1), 223-234 (1978)) и по методу хромогенного субстрата (H.R.Lijnen et al., J. Biol. Chem., 266, 11826-11832 (1991)). Удельная активность составляет около 90000-100000 HU/мг. Для определения единиц см. работу O.Q.Tang et al., Drug Biotechnol (китайский), 4(1), 1-4 (1997).
i. Определение констант Км и Ккат для комплексов Sak-плазмин и KGD-Sak-плазмин
Выдерживают Sak или KGD-Sak (2 мкмоль/л) с 2 мкмоль/л плазминогена соответственно в 0,1 моль/л фосфатного буфера при рН 7,4 и температуре 37°С в течение 30 мин, для образования комплексов с плазмином. Затем отбирают каталитическое количество (5 нмоль/л) комплекса для осуществления взаимодействия в течение 0-10 мин в реакционных системах, указанных ниже, в 0,1 моль/л ФБ при рН 7,4 и температуре 37°С, с регистрацией оптической плотности (при 405 нм) каждые 30 с. Приведенные концентрации плазминогена являются усредненными по данным трех анализов.
Активация плазминогена комплексом KGD-Sak-плазмин описывается уравнением Михаэлиса-Ментена (табл.1c).
j. Определение способности к полимеризации
В качестве контроля используют Sak естественного типа. Образцы растворяют в физиологическом растворе. Испытания проводят при двух концентрациях протеина 30 мг/мл (высокая) и 3 мг/мл (низкая). Растворы выдерживают при комнатной температуре. Пробы отбирают каждые 24 ч и анализируют путем электрофореза.
При обеих концентрациях протеина способность к полимеризации KGD-Sak была значительно ниже, чем у Sak естественного типа.
k. Испытание чувствительности на морских свинках
Для испытания чувствительности рекомбинантную стафилокиназу естественного типа, а также мутант KGD-Sak растворяют в стерильном физиологическом растворе при концентрации 2500 ед./мл. Для каждого введения берут интактные ампулы, чтобы приготовить стерильным образом свежие растворы. Здоровых морских свинок (20) случайным образом распределяют на две группы, по 10 животных в каждой. Этим морским свинкам внутрибрюшинно вводят инъекцию 0,15 мг/кг r-Sak или KGD-Sak через день по три раза. Первое и второе внутривенное воздействие дозой 0,3 мг/кг проводят соответственно на 14 и 21 сутки. Двум здоровым морским свинкам, которым не делали инъекцию, вводят внутривенно 0,3 мг/кг указанных выше образцов и наблюдают наличие аналогичной реакции, чтобы исключить фармакологические и патологические помехи, связанные с образцами.
В группе с инъекцией r-Sak естественного типа: для 8 морских свинок наблюдалась положительная реакция класса IV и для 2 морских свинок наблюдалась положительная реакция класса II.
В группе с инъекцией KGD-Sak: у 2 морских свинок наблюдалась положительная реакция класса I, а у других животных отсутствовала заметная реакция.
Содержание антител в сыворотке морских свинок, иммунизированных в течение 1-3 недель, определяют методом иммуноферментного твердофазного анализа (ELISA), в котором в качестве антигенов используют wt-Sak и KGD-Sak соответственно. В первую неделю количество антител относительно любого антигена было мало. Во вторую неделю количество антител группы wt-Sak (n=10) увеличивается до 1:800, в то время как количество антител группы KGD-Sak (n=10) составляет 1:200. В третью неделю количество антител группы wt-Sak увеличивается до 1:3200, в то время как в группе KGD-Sak увеличивается до 1:400. Таким образом, иммуногенность KGD-Sak значительно снижается по сравнению с иммуногенностью wt-Sak.
Приведенные выше результаты указывают, что иммуногенность KGD-Sak значительно снижается по сравнению с иммуногенностью wt-Sak.
l. Анализ ингибирования агрегации тромбоцитов
Медленно центрифугируют (ускорение 150 g, 10 мин) свежую кровь с добавкой 1/10 объема раствора (110 ммоль/л) антикоагулянта - цитрата натрия, чтобы получить плазму, обогащенную тромбоцитами (ПОТ). Добавляют KGD-Sak к ПОТ до окончательной концентрации 2 мкмоль/л и смесь выдерживают 2 мин при 37°С и непрерывном перемешивании. Затем добавляют аденозиндифосфат (АДФ) до окончательной концентрации 2 мкмоль/л в качестве индуктора. Степень агрегации тромбоцитов определяют в течение 5 минут в двухканальном аппарате агрегации тромбоцитов (CHRONO-LOG560). В качестве контрольных используют образцы r-Sak естественного типа (2 мкмоль/л) и физиологический раствор. Аденозиндифосфат закупают на фирме Sigma, прочие реагенты аналитической чистоты произведены в Китае.
Найдено, что степень агрегации в группе KGD-Sak (3,8%±1,5%, n=3) значительно ниже, чем агрегация в группах с r-Sak (64%±4%, n=3) и физиологическим раствором (60%±3%, n=3). Это позволяет предположить, что KGD-Sak обладает сильным потенциалом при ингибировании агрегации тромбоцитов, вызванной АДФ (фиг.3).
m. Тромболитический анализ на животных
Тромболитический анализ на животных проводят с KGD-Sak, полученным в настоящем изобретении, проверяя, что KGD-Sak сохраняет те же самые тромболитические свойства, что и Sak естественного типа. Для введения KGD-Sak растворяют в стерильном физиологическом растворе, как описано в разделе k этого же примера.
Лечение тромбоза бедренной артерии экспериментальных кроликов KGD-Sak
В каждой группе, обрабатываемой KGD-Sak, обрабатываемой Sak естественного типа и в контрольной группе, без обработки, содержится 6 животных. С помощью артериографии показано, что до лечения средний сегмент бедренной артерии не был видим. При повторном фотографировании через 60 минут после внутривенной инъекции 0,1 мг/кг KGD-Sak бедренная артерия становится полностью заполненной, с восстановлением цикла крови, что согласуется с эффектом в группе Sak естественного типа, в то время как в контрольной группе бедренная артерия не становится заполненной.
Лечение гифемы экспериментальных кроликов с помощью KGD-Sak
В каждой группе, обрабатываемой KGD-Sak, обрабатываемой Sak естественного типа и в контрольной группе, без обработки, содержится 6 животных. Через 4 ч после внутриглазной инъекции 10-20 мкг KGD-Sak наблюдается лизис тромба гифемы, причем красные кровяные клетки осаждаются и образуют поверхность раздела с водной внутриглазной жидкостью. Внутриглазная кровяная опухоль удаляется через 24 ч. Это согласуется с эффектом в группе Sak естественного типа. Однако в контрольной группе гифема существенно не изменяется.
Тромболитическая терапия с помощью KGD-Sak является безопасной и эффективной при лечении острого инфаркта миокарда у экспериментальных собак, вызванного тромбозом коронарной артерии. В экспериментальной группе, состоящей из 6 животных, собаки получали дозу KGD-Sak (0,3 мг/кг веса тела) с помощью внутривенной инфузии; в контрольной группе, состоящей из 6 животных, собаки получали физиологический раствор, вместо KGD-Sak, с помощью внутривенной инфузии. До и после ввода дозы проводят коронарную артериографию. До ввода дозы обнаружено, что левое переднее нисходящее ответвление коронарной артерии является незаполненным или не полностью заполненным у животных обеих групп. При проведении артериографии через 30 минут после лечения обнаружено, что левое переднее нисходящее ответвление вновь заполняется, и животные в экспериментальной группе выживают. Что касается контрольной группы, то в не полностью заполненной артерии отсутствуют заметные изменения, и животные умирают через несколько часов.
Тромболитическая терапия с помощью KGD-Sak является безопасной и эффективной при лечении острого церебрального инфаркта у экспериментальных свиней, вызванного тромбозом внутричерепной артерии. В экспериментальной группе, состоящей из 6 животных, свиньи получали дозу KGD-Sak (0,2 мг/кг веса тела) с помощью внутривенной инфузии; в контрольной группе, состоящей из 6 животных, свиньи получали физиологический раствор, вместо KGD-Sak, с помощью внутривенной инфузии. До и после ввода дозы проводят внутричерепную DSA артериографию. До ввода дозы обнаружено, что некоторые внутричерепные артерии являются не полностью заполненными у животных обеих групп. При проведении ангиографии через 30 минут после лечения обнаружено, что внутричерепные артерии вновь заполняются, и животные в экспериментальной группе выживают. Что касается контрольной группы, то в не полностью заполненных внутричерепных артериях отсутствуют заметные изменения, и животные умирают спустя несколько дней.
Специалисты в этой области техники без дальнейшего подробного описания смогут максимально использовать это изобретение в свете предшествующих рекомендаций. Таким образом, следует понимать, что предпочтительные конкретные варианты осуществления приведены выше с целью иллюстрации и никоим образом не ограничивают объем изобретения.
Все источники, цитированные в описании, введены во всей полноте, со ссылкой. Существенные характеристики настоящего изобретения станут очевидными для специалистов в этой области техники из предшествующего описания. Более того, могут быть осуществлены различные модификации и усовершенствования этого изобретения, без отклонения от сути настоящего изобретения. Предполагается, что такие модификации и усовершенствования входят в объем прилагаемой формулы изобретения.

Изобретение относится к области биотехнологии и может быть использовано в медицине. Предложены новые производные стафилокиназы, которые представляют собой продукты экспрессии в гетерогенной системе мутантных генов, полученных путем локального ПЦР-мутагенеза гена ← фермента дикого типа, и отличаются от соответствующей природной формы стафилокиназы одной или большим числом аминокислотных замен в области между 104 и 113 аминокислотными остатками, приводящих к образованию в названной области последовательности RGD или KGD. Полученные рекомбинантные производные стафилокиназы (RGD/KGD-Sak), а также усеченные с NH2-конца на 1-16 аминокислот варианты этих производных совмещают свойства тромболитического и антикоагулирующего агента и характеризуются пониженными, по сравнению с ферментом дикого типа, способностью к полимеризации и иммуногенностью, что обеспечивает возможность их успешного применения в составе фармкомпозиций и в способах лечения или предупреждения тромбоза. 7 н. и 18 з.п.ф-лы, 3 ил., 3 табл.
a) последовательности, приведенной в SEQ ID NO:1;
b) последовательности, приведенной в SEQ ID NO:3; и
c) NH2-концевого усечения SEQ ID NO:1 или SEQ ID NO:3.
a) получение фрагмента ДНК, содержащего кодирующую последовательность биологически активной стафилокиназы;
b) осуществление in vitro сайт-специфического мутагенеза фрагмента ДНК, чтобы заменить один или более кодонов аминокислотных остатков между аминокислотными остатками 104 и 113 стафилокиназы дикого типа на кодоны других аминокислот для образования в кодируемом производном последовательности RGD или последовательности KGD;
c) клонирование мутированного фрагмента ДНК в подходящем векторе;
d) трансформация или трансфекция подходящей клетки-хозяина рекомбинантным вектором;
e) культивирование клетки-хозяина в условиях, подходящих для экспрессии фрагмента ДНК; и
f) извлечение и очистка желаемого производного стафилокиназы.
| Способ производства широких горячекатаных полос из высокоуглеродистых низколегированных сталей | 1984 |
|
SU1196391A1 |

