Область техники, к которой относится изобретение
Настоящее изобретение, раскрытое здесь, относится к новому способу продукции стафилокиназы (SAK) и ее производных путем регулирования уровня кислорода в среде для ее выращивания, с применением генно-инженерного штамма E. coli. Изобретение относится к последовательностям ДНК, которые кодируют формы стафилокиназы в соответствии с изобретением, и к рекомбинантным плазмидам, в которых присутствуют эти последовательности ДНК, связанные с высокоэффективными сигналами экспрессии, которые контролируются подачей кислорода.
Полный процесс касается нового способа высокоэффективной продукции стафилокиназы и ее аналогов с высоким выходом, с применением особым образом разработанного сигнала экспрессии белка, который не требует применения дорогостоящих экзогенных химических регентов для индукции продукции белка и, следовательно, обеспечивает экономическое преимущество перед известной в настоящее время процедурой продукции стафилокиназы. Стафилокиназа и ее производные, полученные посредством этого процесса, проявляют специфическую тромболитическую активность и способность к активации плазминогена человека, что сопоставимо с нативным аналогом. Таким образом, в принципе, настоящий процесс лежит в основе нового и более экономичного способа продукции стафилокиназы и ее производных, которые могут использоваться в тромболитической терапии.
В связи с этим, изобретение также относится к разработке новой последовательности ДНК, сопоставимой с последовательностью ДНК, кодирующей SAK, которая приводит к накоплению белка стафилокиназы в большом количестве внутри клетки при снижении концентрации кислорода ниже относительно общего содержания атмосферного кислорода на 2% или при достижении клетками стационарной фазы роста. Формы SAK, продуцируемые в соответствии с изобретением, таким образом, могут использоваться в качестве фармацевтических препаратов для лечения тромбоэмболических ангиопатий у человека.
Уровень техники
Стафилокиназа, внеклеточный белок некоторых штаммов Staphylococcus aureus, является перспективным тромболитическим агентом, который обладает множеством желательных тромболитических свойств и может использоваться для лечения пациентов, страдающих тромбоэмболическими нарушениями, и пациентов с инфарктом миокарда [Collen et. al., Fibrinolysis, vol. 6; 232-242, 1992; Collen et. al., vol. 87, 996-1006, 1993]. Стафилокиназа образует бимолекулярный комплекс с белками крови, такими как плазминоген (PG) и плазмин (Pm), и влияет на их фибринолитическую активность посредством преобразования активной неспецифичной сывороточной протеазы плазмина (Pm) в высокоспецифичный протеолитический фермент, который может распознавать субстрат, профермент крови PG, и преобразовывать его в плазмин, способный растворять кровяные тромбы. В плазме SAK может растворять сгустки фибрина без разложения фибриногена [Collen et. al., J. Biol. Chem. 268, 8284-8289, 1993; Lijnen et al., J. Biol. Chem. 266, 11826-11823]. Эта специфичность SAK к фибрину является результатом ингибирования образования комплекса стафилокиназа-плазмин посредством альфа-2-антиплазмина в ходе циркулирования в крови и, в значительной степени, ослабленного взаимодействия с комплексом стафилокиназа-плазмин на поверхности фибрина, что приводит к очень локализованной активации плазминогена на поверхности фибрина. Кроме того, стафилокиназа, обладая слабым сродством к циркулирующему плазминогену, при этом обладает высоким сродством к плазминогену, связанному с фибрином. Последние клинические испытания показали, что при проведении преждевременной перфузии у пациентов с инфарктом миокарда стафилокиназа столь же эффективна, как и t-PA. Таким образом, ее эффективность в тромболитической терапии была определена в нескольких ограниченных клинических испытаниях [Collen et. al., Circulation, 87, 996-1006, 1993; Lijnen and Collen, Fibrinolysis, 10, 119-126, 1996].
Стафилокиназа представляет собой одноцепочечный 16 кД белок, состоящий из 136 аминокислотых остатков. В очень низких количествах ее продуцирует природный хозяин, Staphylococcus aureus [Lack, Nature 161, 558-560, 1948; Robbinson et. al., J. Biol. Chem 242, 2333-2342, 1967]. Продукция интактной, биологически активной SAK в бактериальных системах экспрессии была затруднена из-за микрогетерогенности N-конца, неустойчивости плазмиды или низкого выхода. Учитывая терапевтическую применимость стафилокиназы и клиническую применимость в тромболитической терапии, ранее были предприняты попытки (Gerlach et. al., Zbl. Bakt. Hyg. A269, 1988; Collen et. al., Fibrinolysis 6, 203-213, 1992) поиска альтернативного источника продукции SAK рекомбинантными путями. Ген, кодирующий SAK, был выделен из природного хозяина, Staphylococcus aureus, и клонирован в различных ксеногенных хозяевах, например в E. coli, Bacillus и дрожжах [Sako et. al., Mol. Gen. Genet., 190, 271-277, 1983]. В случае с Bacillus SAK протеолитически разлагалась [Ruiqiong et. al., Biotechnol. Bioeng., 62, 87-96, 1998; Miele, et. al., J. Biol. Chem. 274, 7769-7776, 1999], и в дрожжах было обнаружено, что она глигозилируется, что уменьшало функцию активации плазминогена белка. Поэтому эти гетерологичные системы не слишком подходили для крупномасштабной продукции этого белка. В E. coli внеклеточная продукция SAK приводила к низкому уровню продукции SAK, с последующей деградацией N-конца и образованию двух форм SAK. С целью преодоления этих трудностей ген, кодирующий SAK, экспрессировали с применением сильных сигналов экспрессии, для которых требовались дополнительные экзогенные индукторы, такие как IPTG, триптофан, индолуксусная кислота и тому подобное, что делало эти системы очень дорогостоящими при использовании для крупномасштабной продукции этого белка. В этой системе продукция полноразмерной SAK была сопряжена с получением усеченной формы SAK с отсутствием 10 N-концевых аминокислотных остатков нативной SAK.
Следовательно, настоящее изобретение относится к новому способу высокоэффективной продукции стафилокиназы и ее аналогов с применением нового сигнала экспрессии белка, который не требует использования дорогостоящих экзогенных химических реактивов для индукции продукции белка и, следовательно, обеспечивает экономическое преимущество по сравнению с известным в настоящее время способом продукции стафилокиназы. Кроме того, стафилокиназа и ее производные, полученные с помощью этого способа, проявляют специфическую тромболитическую активность и способность к активации плазминогена человека, что сопоставимо с нативным аналогом. Таким образом, в принципе, настоящий способ лежит в основе нового и более экономичного способа продукции стафилокиназы и ее производных, которые могут использоваться в тромболитической терапии.
Цели настоящего изобретения
Следовательно, главная цель настоящего изобретения состоит в разработке биологически функциональных аналогов SAK, у которых сохраняются N-концевые аминокислотные остатки и которые проявляют полную биологическую активность при крупномасштабной продукции.
Другая цель этого способа состоит в разработке и конструировании эффективной и экономичной системы продукции SAK и ее варианта, для которой не требуется применения какого-либо химического индуктора так, чтобы полная система могла быть экономически рентабельной.
Еще одной целью изобретения является получение части ДНК, несущей полную генетическую информацию для продукции SAK в подходящем хозяине, таком как E. coli, Bacillus, дрожжи или в любой системе микроорганизмов, в которой можно применять характеристики и которая, тем самым, может служить в качестве хозяина.
Еще одна цель изобретения состоит в модификации 1-10 амино-концевых аминокислотных остатков SAK для препятствия ее усечения в ходе крупномасштабной продукции, не влияя на ее суммарную функциональную активность.
Еще одна цель изобретения состоит в конструировании части ДНК и объединении ее с последовательностями ДНК SAK с тем, чтобы экспрессировать SAK и модифицированную на N-конце форму SAK для высокоэффективной экспрессии белков.
Еще одной целью изобретения является получение экспрессионной ДНК плазмиды, несущей генетическую информацию для продукции SAK в рекомбинантной клетке E. coli.
Сущность изобретения
Настоящее изобретение относится к нуклеотидной последовательности кассеты экспрессии OXY-1 с последовательностью SEQ ID No. 1, гену модифицированной стафилокиназы SAK-2 с последовательностью SEQ ID No. 2, пептидной последовательности гена модифицированной стафилокиназы SAK-2 с последовательностью SEQ ID No. 3, трем плазмидам с международными инвентарными номерами BPL-0019, BPL-0020 и BPL-0021 и соответствующим трем рекомбинантным E. Coli; также изобретение относится к способу повышенной продукции стафилокиназы и ее аналогов путем модуляции уровня кислорода в среде для ее выращивания в системе-хозяине и, наконец, к способу растворения кровяного тромба у пациента при необходимости такого растворения.
Подробное описание изобретения
Таким образом, настоящее изобретение относится к нуклеотидной последовательности кассеты экспрессии OXY-1 с последовательностью SEQ ID No. 1, гену модифицированной стафилокиназы SAK-2 с последовательностью SEQ ID No. 2, пептидной последовательности гена модифицированной стафилокиназы SAK-2 с последовательностью SEQ ID No. 3, к трем плазмидам с международными инвентарными номерами BPL-0019, BPL-0020 и BPL-0021 и к соответствующим трем рекомбинантным E. Coli; также изобретение относится к способу повышенной продукции стафилокиназы и ее аналогов путем модуляции уровня кислорода в среде для ее выращивания в системе-хозяине и к способу растворения кровяного тромба у пациента при необходимости такого растворения.
В одном из вариантов осуществления настоящее изобретение относится к нуклеотидной последовательности кассеты экспрессии OXY-1 с последовательностью SEQ ID No. 1.
В другом варианте осуществления настоящее изобретение относится к гену модифицированной стафилокиназы SAK-2 с последовательностью SEQ ID No. 2.
В еще одном варианте осуществления настоящее изобретение относится к пептидной последовательности гена модифицированной стафилокиназы SAK-2 с последовательностью SEQ ID No. 3.
В еще одном варианте осуществления настоящее изобретение относится к плазмиде pRM1 с международным инвентарным номером BPL-0019.
В еще одном варианте осуществления настоящее изобретение относится к плазмиде pOXYSAK-1 с международным инвентарным номером BPL-0020.
В еще одном варианте осуществления настоящее изобретение относится к плазмиде pOXYSAK-2 с международным инвентарным номером BPL-0021.
В еще одном варианте осуществления настоящее изобретение относится к рекомбинантной E. Coli с международным инвентарным номером 5146, международным банком-депозитарием является "Microbial Type Culture Collection" при институте “Institute of Microbial Technology”, Chandigarh, India, содержащей плазмиду pRM1 с международным инвентарным номером BPL-0019.
В еще одном варианте осуществления настоящее изобретение относится к рекомбинантной E. Coli с международным инвентарным номером 5147, международным банком-депозитарием является "Microbial Type Culture Collection" при институте “Institute of Microbial Technology”, Chandigarh, India, содержащей плазмиду pOXYSAK-1 с международным инвентарным номером BPL-0020.
В еще одном варианте осуществления настоящее изобретение относится к рекомбинантной E. Coli с международным инвентарным номером 5148, международным банком-депозитарием является "Microbial Type Culture Collection" при институте “Institute of Microbial Technology”, Chandigarh, India, содержащей плазмиду pOXYSAK-2 с международным инвентарным номером BPL-0021.
В еще одном варианте осуществления настоящее изобретение относится к способу повышенной продукции стафилокиназы и ее аналогов путем модуляции уровня кислорода в среде для ее выращивания в системе-хозяине, который предусматривает стадии:
• получения части ДНК, несущей генетическую информацию для продукции стафилокиназы,
• модифицирования 10 амино-концевых остатков ДНК, кодирующей SAK, где остатки SAK Lys6 и Lys8 заменяют на небольшие нейтральные аминокислотые остатки,
• конструирования кассеты экспрессии ДНК OXY-1,
• интегрирования части ДНК, полученной на стадии (a) или на стадии (b) в кассету экспрессии с OXY-1 с получением pOXYPRO,
• переноса интегрированного продукта стадии (d) в плазмидный вектор с получением плазмидной конструкции pOXYSAK-1 и pOXYSAK-2, соответственно,
• введения плазмидной конструкции стадии (e) в систему-хозяин,
• культивирования клетки-хозяина для повышенной продукции SAK или ее производных при высокой степени аэрации и при изменении уровня кислорода ниже атмосферного уровня кислорода на 5%, когда рост клетки достигает экспоненциальной фазы с получением клеточной массы,
• лизирования клеток стадии (g) для отделения клеточного лизата от клеточного дебриса с получением, таким образом, стафилокиназы и ее аналогов.
В еще одном варианте осуществления настоящего изобретения остатки Lys6 и Lys8 SAK заменяют на небольшие и нейтральные аминокислотные остатки.
В еще одном варианте осуществления настоящего изобретения плазмидный вектор представляет собой плазмиду с высоким или средним числом копий.
В еще одном варианте осуществления настоящего изобретения систему-хозяина выбирают из группы, содержащей E. coli, Bacillus и дрожжи.
В еще одном варианте осуществления настоящего изобретения последовательность OXY-1 модифицируют в зависимости от системы-хозяина.
В еще одном варианте осуществления настоящего изобретения аминокислоты выбирают из группы, содержащей аланин и глицин.
В еще одном варианте осуществления настоящего изобретения среда для выращивания представляет собой питательную среду Luria Broth (LB).
В еще одном из вариантов осуществления настоящего изобретения культивирование клеток-хозяев для повышенной продукции SAK или ее производных осуществляют в культуре встряхиваемой колбы или при ферментации.
В еще одном из вариантов осуществления настоящего изобретения культивирование клеток-хозяев осуществляют до тех пор, пока O.D. 600 не достигнет значения 0,6-0,7.
В еще одном из вариантов осуществления настоящего изобретения ферментация представляет собой двухстадийную периодическую ферментацию с подпиткой.
В еще одном варианте осуществления настоящего изобретения клеточную массу получают путем центрифугирования или фильтрации.
В еще одном из вариантов осуществления настоящего изобретения клетки лизируют способом, выбранным из группы, включающей лизис ультразвуком, химический и механический лизис.
В еще одном из вариантов осуществления настоящего изобретения клеточный лизат отделяют от клеточного дебриса путем центрифугирования.
В еще одном из вариантов осуществления настоящего изобретения способ растворения кровяного тромба у пациента, при необходимости такого растворения, предполагает стадию введения фармацевтически эффективного количества аналога стрептокиназы SAK-2, необязательно, с добавкой (добавками).
В еще одном из вариантов осуществления настоящего изобретения добавку выбирают из группы, включающей питательные вещества, состоящие из белков, углеводов, сахара, талька, стеарата магния, целлюлозы, карбоната кальция, крахмал-желатиновой пасты, и/или фармацевтически приемлемый носитель, эксципиент, разбавитель или растворитель. В еще одном из вариантов осуществления настоящего изобретения соотношение SAK-2 и добавок изменяется в пределах от 1:10 и 10:1.
Таким образом, настоящий способ относится к выделению части ДНК из ДНК Staphylococcus aureus, несущей полную последовательность для продукции SAK и ее модификации, с применением известных методик [Sambrook et al., Molecular Cloning: A laboratory Manual, 1989, Cold Spring Harbor] для получения форм SAK, в которых сохранены N-концевые аминокислоты, что обеспечивает получение единой формы биологически активной SAK. Новая разработанная часть ДНК была сконструирована и связана с ДНК, кодирующей SAK, в соответствующем плазмидном векторе, способном к репликации в подходящем хозяине E. coli с продукцией большого количества SAK и ее аналогов при уровне кислорода в условиях роста ниже атмосферного на 5%. После этих стадий можно получить высокий уровень белков SAK без какой-либо деградации белков.
Описанный здесь способ продукции SAK и ее аналогов не был известен ранее, поэтому он обеспечивает новый эффективный и экономически выгодный способ продукции биологически активных белков SAK путем обычной модуляции уровня атмосферного кислорода в культуральной среде в культуре встряхиваемой колбы или в процессе режима ферментации. SAK или ее аналоги, продуцируемые таким образом, могут быть получены путем лизирования клеток E. coli и отделения загрязняющих клеточных белков после обычных стадий хроматографии. Стафилокиназа и ее модифицированные формы, продуцируемые после этих стадий, проявляют тромболитическую активность и способны активировать плазминоген, что сопоставимо с активностью природного аналога, и могут использоваться в клинической практике для лечения различных нарушений циркуляции.
Соответственно, настоящее изобретение относится к новому способу продукции стафилокиназы и ее производных путем модуляции уровня кислорода в среде для ее выращивания, с применением генно-инженерного штамма E. coli и предусматривает:
(a) получение части ДНК, несущей генетическую информацию для продукции стафилокиназы посредством рекомбинантной технологии или посредством синтетических методов, следуя известным методикам,
(b) модифицирование 10 аминокислотных остатков ДНК, кодирующей SAK, где остатки Lys6 и Lys8 SAK заменяют, например, на аланин, глицин или подобные небольшие и нейтральные аминокислотные остатки,
(c) конструирование синтетической кассеты ДНК, несущей конкретную последовательность ДНК,
(d) интегрирование части ДНК, полученной на стадии (a) или на стадии (b), в последовательность ДНК, полученную на стадии (c) обычным способом рекомбинантной технологии,
(e) перенос последовательностей, кодирующих SAK, связанных со специфичным белковым сигналом экспрессии, полученным на стадии (d), в соответствующий плазмидный вектор репликации известными методиками рекомбинантных ДНК,
(f) подходящий плазмидный вектор в таком случае может представлять собой плазмиду с высоким или средним числом копий, специфичную для системы-хозяина,
(g) введение плазмидных конструкций, содержащих плазмиды, кодирующие SAK, полученных на стадии (e), в подходящий хозяин, такой как E. coli, Bacillus, дрожжи, или любую микробиальную систему, характеристики которой могут быть изменены и которая, таким образом, может служить хозяином,
(h) культивирование клеток-хозяев, содержащих плазмиду экспрессии для продукции SAK или ее модифицированных форм либо в культуре встряхиваемой колбы при высокой степени аэрации, либо на уровне ферментации до тех пор, пока O.D. 600 не достигает значения 0,6-0,7 с последующим понижением уровня кислорода до значения, которое меньше суммарного атмосферного кислорода на 5%,
(i) удаление клеток путем центрифугирования, фильтрации и тому подобное и лизирование клеток обычными методами, такими как лизис ультразвуком, химический и механический лизис, с последующим отделением клеточного лизата от клеточного дебриса после центрифугирования и последующей очисткой стафилокиназы известными методами.
Подробное описание изобретения
Полный способ, разработанный и раскрытый в данном описании, основан на использовании новой последовательности ДНК (фиг.3), связанной с SAK и формами SAK, которая обеспечивает продукцию большого количества стафилокиназы при изменении кислородных параметров условий культивирования и не требует использования внешнего источника для индукции продукции рекомбинантной SAK в E. coli.
Применение E. coli в качестве хозяина для высокоэффективной продукции рекомбинантных белков широко распространено в современной биотехнологической практике. Крупномасштабная продукция эффективных белков обычно достигается посредством двухстадийных периодических ферментаций с подпиткой. На первой стадии такого способа клетки выращивают до высокой клеточной плотности в приемлемо уравновешенных условиях роста. Для этого требуется, чтобы синтез гетерогенных белков был сведен к минимуму. На второй стадии активируют высокоэффективную экспрессию рекомбинантных белков. Также обнаружено, что относительная производительность различных промоторов изменяется от белка к белку. Следовательно, свойства промотора, который регулирует экспрессию рекомбинантного белка, являются критическими для производительности способа. Раньше, ген, кодирующий SAK, клонировали под индуцибельными промоторами, например tac или T7, для которых требовалось использование свободного индуктора или повышение температуры условий роста. Принимая во внимание, что прежний подход способствует наименьшему влиянию на метаболизм клетки-хозяина, в целом, свободные индукторы являются дорогостоящими и, следовательно, негативно сказываются на росте экономических показателей. Кроме того, было показано, что ген нативной SAK при интеграции с этими системами экспрессии процессировал N-концевую область SAK, что приводило к микрогетерогенности белкового продукта. Следовательно, для предотвращения N-концевого процессинга SAK модифицирование Lys6 и Lys8 аминокислотных остатков в N-концевой последовательности не влиял на общие функциональные свойства белка. Если эти формы SAK связывать с разработанными последовательностями экспрессии ДНК, то они могут продуцировать большое количество SAK в клеточной культуре E. coli.
Новая система экспрессии SAK была разработана с применением прототипа системы экспрессии SAK pRM1 (фиг.1, [Rajamohan and Kanak L. Dikshit, FEBS Letters 474, 151-158, 2000]), которая несет ген, кодирующий SAK под IPTG-индуцибельном промоторе T7, и новая кассета экспрессии белка (фиг.3), которая несет последовательность ДНК для связывания кислород-чувствительных регуляторных белков, таких как FNR (ферредоксин-НАДФ+редуктаза), в конкретном положении. Ген, кодирующий SAK (фиг.4), был связан с кассетой экспрессии белка, с применением методик рекомбинантных ДНК, и этот слитый продукт затем клонировали в подходящий реплицирующийся плазмидный вектор (фиг.7 и 8) с получением плазмидного вектора экспрессии SAK, несущего ген SAK, или его модифицированных формы (фиг.6, фиг.9) под новой кассетой экспрессии, названной pOXYPRO (фиг.6), экспрессии белка который при низком уровне кислорода (~менее 5% атмосферного кислорода).
В предпочтительном варианте осуществления настоящее изобретение описывает разработку плазмидного вектора, в котором используют слияние новой последовательности экспрессии белка (фиг.3) с последовательностями ДНК SAK (фиг.4) или его модифицированными на N-конце формами (фиг.5 и фиг.6) для получения большого количества стафилокиназы или ее аналогов, при снижении уровня кислорода в условиях роста на 5% ниже атмосферного уровня кислорода.
В еще одном предпочтительном варианте осуществления способа по настоящему изобретению получали синтетическую кассету, несущую определенную последовательность ДНК (фиг.3), с применением известной методики, соединяли на N-конце ДНК, кодирующей SAK, и клонировали в подходящей плазмиде, которая могла представлять собой pUC или Bluescript.
В другом предпочтительном варианте осуществления для плазмидных векторов экспрессии SAK (фиг.8 и фиг.9), кодирующих SAK или формы SAK, использовали штамм E. coli.
Изобретение проиллюстрировано нижеследующими примерами, которые, тем не менее, не следует принимать как ограничивающие. Способ по настоящему изобретению представлен для иллюстрации настоящего изобретения и, поэтому, не должен рассматриваться как ограничивающий объема настоящего изобретения.
Общие способы, используемые в примерах:
1. Методики рекомбинантных ДНК: Использовали обычные и известные методики рекомбинантных ДНК и молекулярной биологии. Подробное описание этих методик доступно в различных руководствах или справочниках, относящихся к этой области, например в Sambrook et. al., Molecular Cloning: A Laboratory Manual (2nd edition, Cold spring Harbor Press, New York, 1989).
2. Электрофоретический анализ белков: Очищенный препарат SAK или его наличие in vivo исследовали с помощью SDS-PAGE, по существу, в соответствии с методом Laemmli (Nature 227; 680, 1970).
3. Перекрывающийся анализ с казеином-плазминогеном для обнаружения активности SAK in vivo : Бактериальные колонии, продуцирующие стафилокиназу, могут быть обнаружены обычным способом по перекрыванию казеина и плазминогена человека в мягком агаре в соответствии с методом Malke, H. и Ferreti, J. J. (Proc. Natl. Acad. Sc. 81; 3557, 1984). Кратко, 10 мл смеси мягкой агарозы, содержащей 0,8% агарозы, 10% обезжиренного молока, 100 мкг человеческого плазминогена, 150 мМ NaCl и 50 мМ Tris.Cl (pH 8,0) выливали сверху на планшеты с бактериальными колониями, экспрессирующими стафилокиназу. Планшеты инкубировали при 37°C в течение 4-5 ч и колонии, несущие SAK, идентифицировали по наличию прозрачной зоны вокруг колоний.
4. Проба на стафилокиназу с применением субстрата хромогенного пептида. Способность стафилокиназы и ее модифицированных форм к активации плазминогена проверяли с помощью известных методов (Methods of Enzymology; 80; 387). Кратко, один микролитр подходящим образом разбавленного образца SAK смешивали с 25 мкл буфера для образцов (0,15М Tris.Cl, pH 7,5) и 100 мкг плазминогена человека, инкубировали при 37°C в течение 15 минут и затем добавляли 18 мкл NaCl (1,77М в 0,032М Tris.Cl, pH 7,5). Количество плазмина, получаемого таким образом, измеряли после добавления 12 мкл хромогенного субстрата, Chromozyme PL (5 мг/мл в воде, Boehringer Mannheim), и пробирки дополнительно инкубировали при 37°C в течение 10 минут. Активность SAK измеряли при 405 нм посредством высвобождения желтого п-4-нитроанилина.
Проба растворения тромба: Способность SAK или ее модифицированных форм к растворению тромба проверяли в соответствии со стандартными методами (British Pharmacopia, 1980 edition). Растворение фибринового тромба осуществляли в присутствии цитратной плазмы человека или фибриногена человека, содержащей различные концентрации SAK или ее аналогов. Кратко, фибриновый тромб маркировали посредством радиоиодирования (125I), смешивали с подходящими концентрациями SAK и инкубировали при 37°C при медленном вращении. Аликвоту 0,1 мл удаляли через постоянные интервалы времени и высвобождение растворимого фибрина измеряли с помощью количества высвобождаемой радиоактивности с применением гамма-счетчика.
Краткое описание чертежей
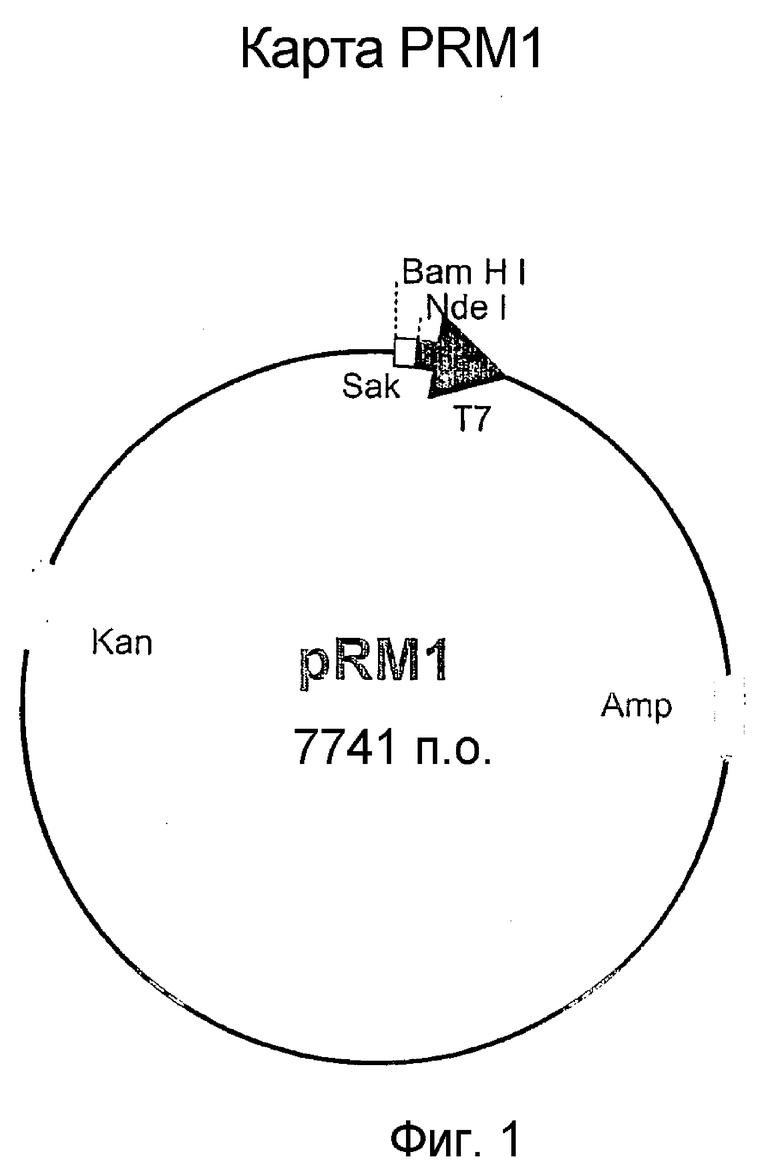
1. Фиг.1: Карта плазмиды pRM1.
2. Фиг.2: Последовательность олигонуклеотида для конструирования кассеты экспрессии белка
3. Фиг.3: Полная нуклеотидная последовательность кассеты экспрессии, OXY-1.
4. Фиг.4: Нуклеотидная последовательность гена SAK.
5. Фиг.5: Нуклеотидная последовательность гена SAK1.
6. Фиг.6: Нуклеотидная последовательность гена SAK2.
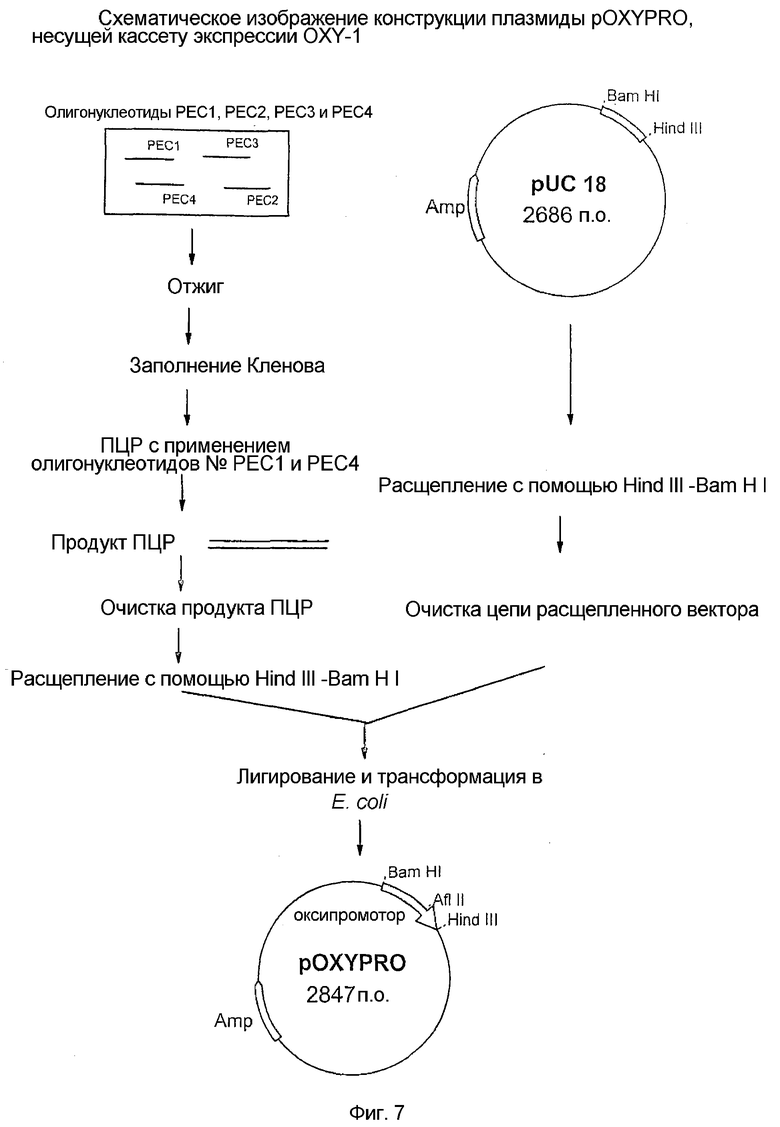
7. Фиг.7: Схематическое изображение конструкции плазмиды pOXYPRO, несущей кассету экспрессии OXY-1.
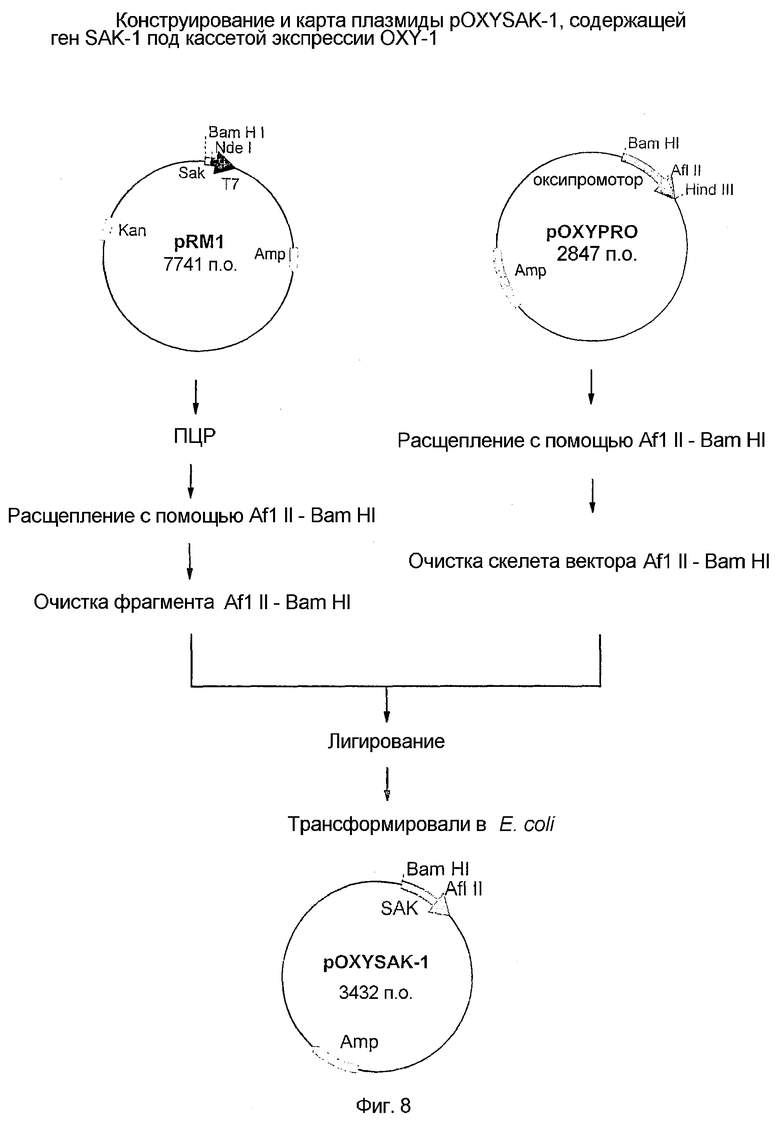
8. Фиг.8: Конструкция и карта плазмиды pOXYSAK-1, содержащей ген SAK-1 под кассетой экспрессии OXY-1.
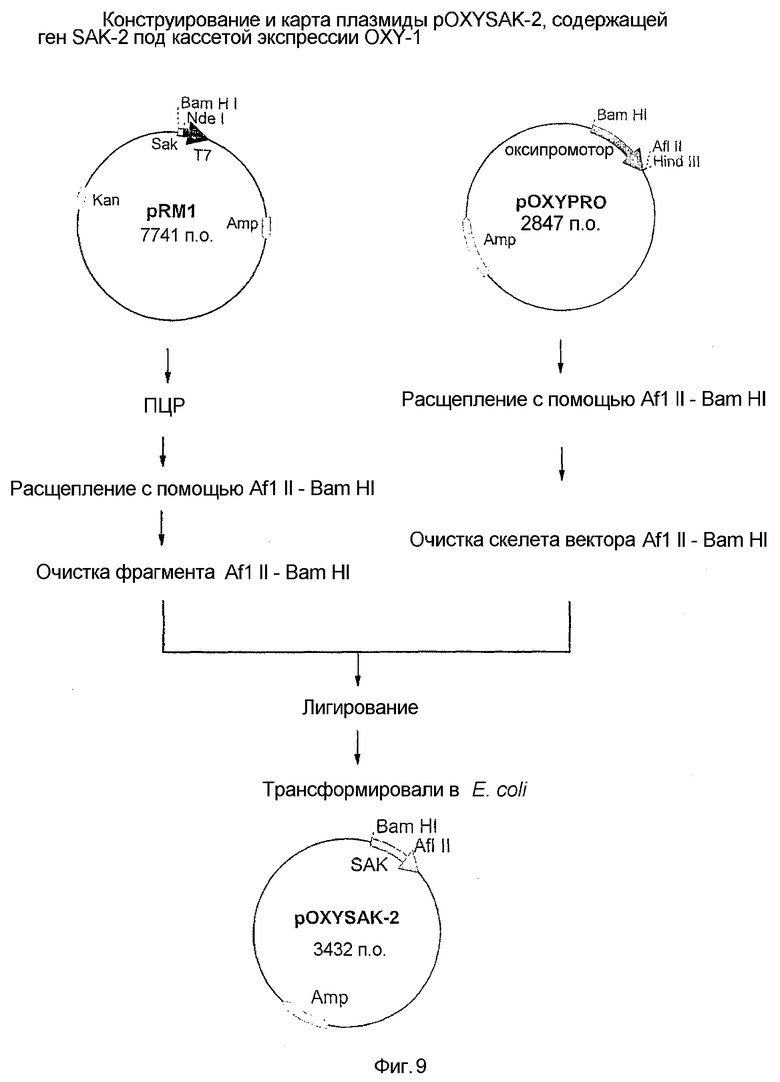
9. Фиг.9: Конструкция и карта плазмиды pOXYSAK-2, содержащей ген SAK-2 под кассетой экспрессии OXY-1.
10. Фиг.10: Последовательности генов SAK, SAK-1 и SAK-2. Терминаторные последовательности в SAK-1 и SAK-2 показаны полужирными буквами и модифицированные остатки в SAK-2 показаны строчными полужирными буквами.
11. Фиг.11: Модификация SAK в SAK-2. Модифицированные остатки показаны полужирными буквами.
Далее изобретение представлено с помощью примеров. Однако примеры не следует рассматривать как ограничивающие объем изобретения.
Пример 1: Конструирование системы экспрессии для продукции внутриклеточной стафилокиназы.
Полный процесс конструирования системы экспрессии для продукции стафилокиназы по существу предусматривал следующие три основные стадии:
Стадия 1: Для конструирования рекомбинантной плазмиды, способной продуцировать рекомбинантную SAK, сходную с нативной, полную геномную ДНК выделяли из определенной выделенной разновидности Staphylococcus aureus (обозначенной как SAK-11) с помощью стандартных методов выделения ДНК. Первую последовательность ДНК, кодирующую SAK, амплифицировали путем ПЦР с применением следующего набора олигонуклеотидных праймеров:
Праймер SAK-1: 5'-GATTGTAGCCATATGTCAAGTTCATTCGACAAAGGAA-3' (37-мер).
Праймер SAK-2: 5'-CGGCTACTGGATCCTCCACTTTTATCCAAACTGATTT-3' (37-мер).
ПЦР осуществляли с помощью матрицы ДНК S. aureus и праймеров SAK-1 и SAK-2 в 100 мкл реакционной смеси, несущей 10 нанограмм матричной ДНК, 20 пикомоль SAK-1 и SAK2, 2,5 единицы Vent ДНК полимеразы (NEB) и 20 мкл каждого дНТФ и 10 мкл стандартного буфера ПЦР (10×буфер; коммерчески доступный от NEB). Для ПЦР общее количество используемых циклов составляло 30 и суммарная продолжительность составляла 10 минут при 72°C. Был получен амплифицированный ДНК продукт с 606 п.о., что было показано на 1% агарозном геле. ДНК продукт расщепляли ферментами рестрикции, клонировали на сайтах NdeI и BamHI плазмидного вектора pET9b и наличие гена SAK на рекомбинантной плазмиде дополнительно проверяли анализом нуклеотидной последовательности (фиг.4) посредством стандартных методик рекомбинантных ДНК. В результате был сконструирован плазмидный вектор pRM1 (депонированный как BPL-0019 в Microbial Type Culture Collection, национальной коллекции культур, расположенной в институте Institute of Microbial Technology, Chandigarh, входящий в состав Council of Scientific and Industrial Research лаборатория. BPL обозначает, что плазмида находится в бактерии, и 0019 обозначает серийный номер депонированного материала).
Основной раствор в глицерине E. coli, несущей рекомбинантную плазмиду, pRM1 (фиг.1) получали в соответствии с описанием в руководстве по методикам рекомбинантной ДНК (Sambrook et. al., Molecular Cloning: A laboratory Manual 1989; Cold Spring Harbor) и поддерживали при -70°C. Для получения биологически активной стафилокиназы культуру для высевания рекомбинантных бактерий, несущих плазмиду экспрессии SAK pRMl, восстанавливали путем инокуляции оттаянного основного раствора в глицерине в 100 мл стандартной среды Luria Broth (LB) (в конической колбе на 500 мл) содержащей 25 мкг/мл канамицина сульфата.
Стадия 2: Вторая стадия включала конструирование синтетической кассеты ДНК OXY-1 для экспрессии SAK и ее производных. Конструирование кассеты ДНК с 150 основаниями OXY-1 проводили с использованием синтетических олигонуклеотидов. Эти олигонуклеотиды синтезировали в коммерческих масштабах, их последовательность приведена на фиг.2. По 1 мкг каждого олигонуклеотида смешивали вместе, нагревали на водяной бане при 100°C в течение 2 минут и после этого медленно охлаждали при комнатной температуре в течение 30 мин. Полученную таким образом смесь для отжига прогоняли на 2% агарозном геле и полосу ДНК с 150 основаниями очищали на геле с применением коммерчески доступных наборов очистки ДНК (набор для очистки ДНК Wizard, Promega Biotech.). Этот очищенный фрагмент ДНК со 150 основаниями расщепляли ферментами рестрикции HindIII и BamHI и связывали на сайте HindIII-BamHI плазмидного вектора pUC18 (New England Bio labs). Получившийся плазмидный вектор обозначали как pOXY-1 (фиг.7). Полную последовательность ДНК со 150 основаниями, вставленную в pUC18, проверяли анализом нуклеотидной последовательности, которая представлена на фиг.3.
Стадия 3. Третья стадия включала конструирование плазмидного вектора экспрессии, pOXYSAK-1 (депонированного как BPL-0020 в Microbial Type Culture Collection, национальной коллекции культур, расположенной в Institute of Microbial Technology, Chandigarh), несущего новый сигнал экспрессии для внутриклеточной продукции SAK в E. coli. Конструирование плазмидного вектора, кодирующего SAK, под контролем кислород-регулируемого белкового сигнала экспрессии осуществляли путем извлечения ДНК, кодирующей SAK, из плазмиды pRM1 (описанной на стадии 1, фиг.l) и объединения ее с новыми белковыми сигналами экспрессии плазмиды pOXYPRO, что приводило к образованию плазмидного вектора pOXYSAK-1.
Схема конструирования плазмиды pOXYSAK-1 изображена на фиг.8. Последовательность ДНК, кодирующую SAK, амплифицировали путем ПЦР из pRM1 с применением следующего набора олигонуклеотидных праймеров:
Праймер SAK-3: 5'-GAACTTAAGGAAGATATACATATGTCAAGTTCATTCGACAA AGGA-3' (45-мер).
Праймер SAK-2: 5'-CGGCTACTGGATCCTCCACTTTTATCCAAACTGATTT-3' (37-мер).
ПЦР осуществляли, используя матрицу ДНК pRM1 и праймеры SAK-3 и SAK-2 в 100 мкл реакционной смеси, несущей 10 нанограмм матричной ДНК, 20 пикомоль SAK-1 и SAK2, 2,5 единицы Vent ДНК полимеразы (NEB) и 20 мкл каждого дНТФ и 10 мкл стандартного буфера для ПЦР (10×буфер; коммерчески доступный от NEB). Для ПЦР общее количество используемых циклов составляло 30 и конечная продолжительность составляла 10 минут при 72°C. Был получен амплифицированный ДНК продукт с 606 п.о., что было показано на 1% агарозном геле. ДНК продукт расщепляли ферментами рестрикции, клонировали на сайтах AflII и BamHI плазмидного вектора OXY-1 в соответствии со стандартными методиками рекомбинантной ДНК.
В отдельной группе 2 мкг плазмидной ДНК pOXYPRO, приготовленной на стадии 2, расщепляли 2 единицами каждого фермента рестрикции AflII и BamHI при 37°C в течение двух часов и смешивали с очищенным на геле фрагментом с 606 основаниями, несущим полный ген SAK. Смесь ДНК осаждали путем добавления 2× объема абсолютного спирта и выдерживания ее при -20°C в течение 1 часа. ДНК восстанавливали после центрифугирования этой смеси в течение 20 минут при 14000 об/мин. Супернатант удаляли и остаток растворяли в 17 мкл воды. Затем смесь ДНК лигировали в присутствии 10 единиц ДНК-лигазы и 1× буфера лигазы (New England Biolabs) при 20°C в течение ночи.
Для стадии трансформации использовали приблизительно 100 нг лигированной ДНК для трансформации электрокомпетентных клеток E. coli JM105 и трансформируемые клетки высевали на планшетах LB-Amp. Собирали 10 трансформированных колоний E. coli и их плазмидные ДНК анализировали на наличие вставки ДНК с 606 основаниями, несущей полный ген, кодирующий SAK, под сигнальной последовательностью OXY-1. Наличие гена SAK на рекомбинантной плазмиде дополнительно проверяли с помощью анализа нуклеотидной последовательности и обозначали как pOXYSAK-1 (фиг.5,9).
Пример 2: Продукция рекомбинантной стафилокиназы из E. coli.
Для продукции рекомбинантной стафилокиназы в E. coli с применением рекомбинантной плазмиды pOXYSAK (фиг.8) использовали основной раствор в глицерине штамма JM109 E. coli, содержащего плазмиду pOXYSAK-1, сохраняемой при -70°C для приготовления культуры для высевания путем инокуляции оттаянного основного раствора в глицерине (приблизительно 100 мкл) в колбу объемом 500 мл со 100 мл питательной среды LB с 100 мкг/мл ампициллина. Колбу инкубировали при 37°C при встряхивании на вращательном шейкере при 200 об/мин в течение 16-18 часов. Клеточную культуру E. coli использовали для засева четырех 2-литровых колб Erlenmyer, содержащих по 500 мл такой же среды (LB, содержащей 100 мкг/мл ампициллина) с применением 5% посевного материала. Колбы инкубировали при 37°C со встряхиванием при 200 об/мин, пока поглощение света культурой при 600 нм не достигало 0,8 (приблизительно 5-6 часов после инокуляции). После этого встряхивание культуры проводили при 50 об/мин и культуры оставляли расти в течение еще 6-8 часов для накопления белка SAK в клетках.
Пример 3. Получение рекомбинантной SAK из E. coli, очистка и характеризация ее биологической активности.
Клетки E. coli, выращенные в течение 6-8 часов, содержащие pOXYSAK (как приведено в примере 2), собирали путем центрифугирования при 6000×g на роторе GS-3 (Sorvall) в течение 30 минут при 4°C. Супернатанты удаляли, остаток клеток ресуспендировали в 50 мл буфере Tris.Cl. и разрушали либо ультразвуком, либо путем химического лизиса, с применением 6M гидрохлорида гуанидина и 20 мМ буфера фосфата натрия, pH 7,2. Лизат клеток центрифугировали при 6000×g при 4°C в течение 15 минут, прозрачные лизаты разбавляли в четыре раза дистиллированной водой и после этого пропускали при комнатной температуре через 10×32 см колонку с SP-сефарозой при скорости потока 1 литр в час. Колонку промывали 0,1M фосфатным буфером, pH 6,2 и элюировали градиентом от 0,1 до 0,5M NaCl. Фракцию, содержащую SAK, проверяли капельным анализом, путем смешивания 1 мкл фракции с 1 мкл плазминогена человека (100 мг/мл) и 1 мл хромозима PL. Фракции, содержащие SAK, продемонстрировали образование желтого цвета. Эти фракции объединяли, доводили до 2,5М с помощью твердого хлорида натрия и подвергали хроматографии гидрофобного взаимодействия на 10×20 см колонке с фенил-сефарозой при комнатной температуре и скорости потока 1 литр в час. Колонку промывали 0,1M фосфатным буфером и элюировали SAK 0,01M фосфатным буфером (pH 6,2). Аликвоты из каждой фракции анализировали на 15% SDS-PAGE для определения относительной чистоты элюируемого белка. SDS-PAGE показал отдельную полосу l6 кД. Специфическую активность очищенного белка SAK оценивали как 1,75 ед/мкг белка, сравнимого с нативной SAK.
Пример 4: Продукция производного SAK, модифицированного на N-конце под сигналом экспрессии OXYPRO.
Создавали производное SAK, которое подвергается модифицированию амино-конца, но сохраняет полную биологическую активность, и подвергали слиянию с последовательностями OXYPRO, для преодоления проблемы N-концевого процессинга SAK, который обычно наблюдается в ходе крупномасштабной продукции SAK.
Удаляли 5 N-концевых остатков и Lys6 и Lys8 заменяли Ala. Выделяли полную геномную ДНК Staphylococcus aureus (SAK-11 как приведено в примере 1 на стадии 1) и использовали ее в качестве матрицы для ПЦР-амплификации с применением следующего набора олигонуклеотидных праймеров:
Праймер SAK 4: 5'-GAACTTAAGCATATGGCTGGAGCTTATAAAAAGGGC-3'.
Праймер SAK 2: 5'-CGGCTACTGGATCCTCCACTTTTATCCAAACTGATTT-3'.
ПЦР осуществляли, используя 10 нанограмм матричной ДНК, 20 пикомоль праймеров SAK-4 и SAK-2, 2,5 единицы Vent ДНК полимеразы и 20 единиц дНТФ и 10 мкл коммерчески доступного стандартного буфера для ПЦР (New England Biolabs). Для ПЦР общее количество используемых циклов составляло 30 и конечная продолжительность составляла 10 минут при 72°C. Был получен амплифицированный ДНК продукт, содержащий около 581 пар оснований, что было показано на 1% агарозном геле. Амплифицированный продукт ДНК расщепляли ферментами рестрикции AflII и BamHI. В параллельной группе 2 мкг плазмидной ДНК pOXYPRO расщепляли 4 единицами ферментов рестрикции AflII и BamHI при 37°C в течение 2 часов в присутствии коммерчески доступного многоцелевого буфера (New England Biolabs), смешивали с амплифицированной в ПЦР ДНК и осаждали этанолом путем добавления двойного объема абсолютного спирта и одной десятой объема ацетата натрия (0,5M) и выдерживания при 4°C в течение 2 часов. Осажденную ДНК восстанавливали после центрифугирования этой смеси в течение 20 минут при 14000 об/мин. Супернатант удаляли и остаток растворяли в 17 мкл воды. Затем смесь лигировали в присутствии 10 единиц ДНК-лигазы и 1 мкл буфер ДНК-лигазы при 20°C в течение ночи. После этого приблизительно 10 нг лигированной ДНК использовали для трансформации электрокомпетентных клеток E. coli JM105 с применением стандартной методики. Трансформированные клетки высевали на планшеты LB, содержащие 100 мкг ампициллина на мл. После ночного инкубирования при 37°C на планшетах появилось несколько колоний, и из них 10 колоний анализировали на наличие вставки ДНК с 430 основаниями, несущей модифицированный ген SAK, то есть SAK-2. Модифицированный ген SAK, кодирующий ДНК с 591 основаниями, секвенировали, последовательность которого приведена на фиг.6. Плазмидную конструкцию обозначали как pOXYSAK-2 (фиг.9). Плазмидный вектор депонирован в Microbial Type Culture Collection как BPL-0021.
E. coli JM105, содержащую pOXYSAK-2, выращивали в колбе объемом 500 мл, содержащей 100 мл питательной среды LB с 100 мкг/мл ампициллина и инкубировали при 37°C со встряхиванием на вращательном шейкере при 200 об/мин в течение 5-6 часов до тех пор, пока поглощение света культурой при 600 нм не достигало 0,8. После этого встряхивание культуры проводили при 50 об/мин и культуры оставляли расти в течение еще 6-8 часов. SAK от этой культуры очищали, следуя стадиям, описанным в примере 3. Анализ SDS-PAGE очищенного белкового препарата продемонстрировал наличие отдельной полосы, соответствующей 16 кД. Специфическую активность очищенного белка SAK оценивали как 1,70000 ед./мг белка, сравнимого с очищенным препаратом стандартной SAK, полученной от нативного хозяина, S. aureus.









Изобретение относится к биотехнологии и представляет собой способ продукции стафилокиназы с использованием кассеты OXY-1 с последовательностью SEQ ID No. 1. Кассета входит в состав двух плазмид с международными инвентарными номерами BPL-0020 и BPL-0021, которыми трансформированы штаммы бактерии Е. coli для продукции стафилокиназы. Изобретение позволяет получать стафилокиназу с высокой степенью эффективности. 6 н. и 10 з.п. ф-лы, 11 ил.
1. Кассета-экспрессии OXY-1, содержащая нуклеотидную последовательность, которая по существу соответствует последовательности SEQ ID No:1.
2. Плазмида pOXYSAK-1, содержащая кассету-экспрессии OXY-1 по п.1, несущую ген SAK, где указанная плазмида задепонирована в международном банке-депозитарии "Microbial Type Culture Collection" при институте "Institute of Microbial Technology", Chandigarh, Индия, под инвентарным номером BPL-0020.
3. Плазмида pOXYSAK-2, содержащая кассету-экспрессии OXY-1 по п.1, несущую ген SAK-2, где указанная плазмида задепонирована в международном банке-депозитарии "Microbial Type Culture Collection" при институте "Institute of Microbial Technology", Chandigarh, Индия, под инвентарным номером BPL-0021.
4. Рекомбинантная E.coli с международным инвентарным номером 5147, использующаяся в качестве хозяина для экспрессии и продукции стафилокиназы, причем международным банком-депозитарием является "Microbial Type Culture Collection" при институте "Institute of Microbial Technology", Chandigarh, Индия, содержащая плазмиду pOXYSAK-1 с международным инвентарным номером BPL-0020.
5. Рекомбинантная E.coli с международным инвентарным номером 5148, использующаяся в качестве хозяина для экспрессии и продукции стафилокиназы, причем международным банком-депозитарием является "Microbial Type Culture Collection" при институте "Institute of Microbial Technology", Chandigarh, Индия, содержащая плазмиду pOXYSAK-2 с международным инвентарным номером BPL-0021.
6. Способ повышенной продукции стафилокиназы SAK-2, где указанный способ включает стадии:
(a) получения ДНК, несущей генетическую информацию для продукции стафилокиназы SAK-2, путем модифицирования 10 амино-концевых остатков ДНК, кодирующей SAK, где остатки SAK Lys6 и Lys8 заменяют на небольшие нейтральные аминокислотые остатки, перед стадией (b);
(b) конструирования ДНК кассеты экспрессии OXY-1 по п.1;
(c) интегрирования полученной ДНК, несущей SAK-2, в кассету-экспрессии OXY-1 с получением плазмидной конструкции pOXYPRO, несущей SAK-2;
(d) переноса pOXYPRO, несущей SAK-2, в плазмидный вектор с получением плазмидной конструкции pOXYSAK-2;
(e) введения плазмидной конструкции pOXYSAK-2 в бактериальную клетку-хозяина;
(f) культивирования клетки-хозяина, полученной на стадии (е), для повышенной продукции стафилокиназы SAK-2 при высокой степени аэрации и при изменении уровня кислорода ниже атмосферного уровня кислорода на 5%, когда рост клетки достигает экспоненциальной фазы с получением клеточной массы, и
(g) лизирования клеток стадии (f) для отделения клеточного лизата от клеточного дебриса с получением, таким образом, стафилокиназы SAK-2.
7. Способ по п.6, где остатки Lys6 и Lys8 SAK заменяют на небольшие и нейтральные аминокислотные остатки.
8. Способ по п.6, где плазмидным вектором является плазмида с высоким или средним числом копий.
9. Способ по п.6, где аминокислоты выбраны из группы, включающей аланин и глицин.
10. Способ по п.6, где средой для выращивания является среда Luria Broth (LB).
11. Способ по п.6, где культивирование клетки-хозяина для повышенной продукции SAK-2 осуществляют в культуре встряхиваемой колбы или при ферментации.
12. Способ по п.11, где культивирование клеток-хозяев осуществляют до тех пор, пока O.D. 600 не достигнет значения 0,6-0,7.
13. Способ по п.11, где ферментация является двухстадийной периодической ферментацией с подпиткой.
14. Способ по п.6, где массу клеток получают путем центрифугирования или фильтрации.
15. Способ по п.6, где лизирование клеток осуществляют способом, выбранным из группы, включающей лизис ультразвуком, химический и механический лизис.
16. Способ по п.6, где отделение клеточного лизата от клеточного дебриса осуществляют путем центрифугирования.
| COLLEN D, et.al | |||
| On the mechanism of the activation of human plasminogen by recombinant staphylokinase | |||
| J Biol Chem | |||
| Способ изготовления фанеры-переклейки | 1921 |
|
SU1993A1 |
| COLLEN D, et.al | |||
| Comparative immunogenicity and thrombolytic properties toward arterial and venous thrombi of streptokinase and recombinant staphylokinase in baboons | |||
| Circulation | |||
| Способ изготовления фанеры-переклейки | 1921 |
|
SU1993A1 |

