ОБЛАСТЬ техники, к которой относится изобретение
Настоящее изобретение относится к области молекулярной биологии и генетической инженерии, в частности к улучшенным ДНК-вакцинам для домашних животных, в особенности собак, кошек и лошадей.
Уровень техники
Использование молекул дезоксирибонуклеиновой кислоты (ДНК) для вакцинации известно с начала 90-х годов (Wolf и др., Science, 247, 1465-1468 (1990)). Этот метод вакцинации индуцирует клеточный и гуморальный иммунитет после трансфекции in vivo клеток вакцинируемого животного молекулами ДНК или РНК, кодирующими и экспрессирующими иммунологически активные белки.
ДНК-вакцина состоит из по меньшей мере одной плазмиды, которая может быть экспрессирована в клетках вакцинируемого животного, и фармацевтически приемлемого наполнителя или эксципиента. Плазмида содержит нуклеотидную последовательность, которая кодирует и экспрессирует один или несколько иммуногенов, таких, как белки или гликопротеины, способные индуцировать у вакцинируемого животного клеточный (мобилизация Т-лимфоцитов) и гуморальный (стимуляция продуцирования антител, специфически направленных против иммуногена) иммунный ответ (Davis H.L., Current Opinion Biotech., 8, 635-640 (1997)).
Поскольку иммуногены, полученные на основе патогена, не всегда могут индуцировать достаточно эффективный защитный иммунный ответ у вакцинируемого животного, в раде случаев бывает необходимо улучшить иммунный ответ.
Каждый путь введения имеет присущие ему ограничения и трудности; так, ДНК-вакцина, эффективная при введении одним путем, может оказаться неэффективной при введении другим путем.
При выборе пути введения нужно учитывать потребности практикующих врачей и животноводов, трудности, связанные с содержанием животных или с природой продукта.
Хотя может быть использован внутримышечный путь введения, подкожный путь введения представляет большой интерес для вакцинации домашних животных, предпочтительно для мелких животных, с которыми трудно обращаться.
ДНК-вакцины, следовательно, должны быть улучшены, чтобы было возможно их эффективное введение различными путями.
ДНК-вакцины уже были использованы экспериментально. В качестве примера можно привести ДНК-вакцину, кодирующую гемагглютинин (НА) вируса кори (Ethart и др., J. Gen. Virol., 78, 1577-1580 (1997)), внутриносовое введение которой в случае мыши оказалось более эффективным, чем пероральное введение. Другим примером является ДНК-вакцина, кодирующая белок оболочки (Env) вируса иммунодефицита человека (ВИЧ), подкожное введение которой оказалось неэффективным по сравнению с внутимышечным введением (Ishii и др., AIDS Res. Hum. Retro, 13, 1421-1428 (1997)).
ДНК-вакцины также были использованы экспериментально против вирусов животных, в частности против вируса чумки собак (CDV). Некоторые иммуногены CDV известны, в частности, нуклеокапсидный белок (N), матриксный белок (М), слитый белок (F) и гемагглютинин (НА) (WO-A-9741236). Однако подкожное введение ДНК-вакцины, кодирующей гемагглютинин и слитый белок CDV, не позволило обнаружить продуцирование антител у мыши, а слабое продуцирование антител удалось детектировать только после внутримышечного введения этой вакцины (Sixt и др., J. Virol., 72, 8472-8476 (1998)).
Инуцирование иммунного ответа и соотношения между различными элементами иммунной системы, принимающими участие в формировании этого ответа, могут слегка различаться у различных видов животных. Многочисленные данные, полученные в результате экспериментов на модели мыши, позволили лучше понять функционирование иммунной системы у мыши, однако эти данные нельзя прямо переносить на другие виды, особенно вследствие того, что индуцировать иммунный ответ у мыши оказалось легче, чем у других видов (van Drunen Little-van den Hurk и др., J. Gen. Virol., 79, 831-839 (1998); Böhm и др., Vaccine, 16, 949-954 (1998)).
Были предложены различные пути введения ДНК-вакцины (интраперитонеальный, внутривенный, внутримышечный, подкожный, внутрикожный, через слизистую оболочку и т.д.). Также были предложены различные средства введения, в частности частицы золота, покрытые ДНК и вносимые через кожу вакцинируемого животного (Tang и др., Nature, 356, 152-154 (1992)), и струйные жидкостные инъекторы, позволяющие осуществлять одновременно трансфекцию клеток кожи и клеток нижележащих тканей (Furth и др., Analytical Bioch., 205, 365-368 (1992)).
Для повышения эффективности трансфекции in vitro ДНК были использованы химические соединения:
А/ - катионные липиды
Катионные липиды разделяют на четыре подгруппы:
1) катионные липиды, содержащие соли четвертичного аммония, как, например, DOTMA (диолеоилоксипропилтриметиламмоний, выпускаемый фирмой Gibco под названием Lipofectine), DOTAP (триметил-2,3-(октадец-9-еноилокси)-1-пропанаммоний; Gregoriadis и др., FEBS Letters, 402, 107-110 (1997)), DMRIE (N-(2-гидроксиэтил)-N,N-диметил-2,3-бис(тетрадецилокси)-1-пропанаммоний; WO-A-9634109), DLRIE (N-(2-гидроксиэтил)-N,N-диметил-2,3-бис(додецилокси)-1-пропанаммоний; Felgner и др., Ann. N.Y. Acad. Sci., 772, 126-139 (1995)).
Катионные липиды, содержащие соли четвертичного аммония, необязательно могут быть получены в виде комплекса с дополнительным нейтральным липидом, таким, как DOPC (диолеоилфосфатидилхолин) или DOPE (диолеоилфосфатидилэтаноламин) (J.P.Behr, Bioconjugate Chemistry, 5, 382-389 (1994));
2) липоамины, как, например, DOGS (диоктадециламидоглицилспермин; продукт, выпускаемый фирмой Promega под названием Transfectam; Abdallah и др., Biol. Cell., 85, 1-7 (1995)), DC-Chol (диметиламиноэтанкарбамоилхолестерин; Gao и Huang, Biochem. Biophys. Res. Commun., 179, 280-285 (1991)), BGSC (бисгуанидинспермидинхолестерин), BGTC (бисгуанидинтренхолестерин) (Vigneron и др., Proc. Natl. Acad. Sci. USA, 93, 9682-9686 (1996));
3) катионные липиды, содержащие соли четвертичного аммония и липоамины, как, например, DOSPA (N,N-диметил-N-(2-(сперминкарбоксамидо)этил)-2,3-бис(диолеоилокси)-1-пропанимидийпентагидрохлорид, выпускаемый фирмой Gibco под названием LipofectAmine®; Hawley-Nelson и др., Focus, 15, 73-79 (1993)), GAP-DLRIE (N-(3-аминопропил)-N,N-диметил-2,3-бис(додецилокси)-1-пропанаминий; Wheeler и др., Proc. Natl. Acad. Sci. USA, 93, 11454-11459 (1996)); Norman и др., Vacine, 15, 801-803 (1997));
4) липиды, содержащие соли амидина, как, например, ADPDE, ADODE (Ruysschaert и др., Biochem. Biophys. Res. Commun., 203, 1622-1628 (1994));В/ - полимеры,
как, например, SuperFect™ (активированные дендримерные молекулы; продукты, выпускаемые фирмой Qiagen; Xu и др., Mol. Genet. Metab., 64, 193-197 (1998)); и
С/ - биохимические агенты,
как, например, токсины, в частности желчные токсины.
Некоторые из этих соединений также были использованы для приготовления ДНК-вакцин с более чем умеренными результатами. Однако результаты, полученные in vitro, нельзя автоматически переносить на вакцинацию ДНК, при которой конечной целью является обеспечение защитной иммунной реакции. Негативное влияние на индуцирование эффективной иммунной защиты также было установлено при использовании соединений, известных для повышения эффективности трансфекции in vitro. Некоторые химические соединения лекарственной формы также являются токсичными в высоких дозах для трансфецируемых клеток.
Согласно уже цитированным работам Etchart (Etchart и др., J. Gen. Virol., 78, 1577-1580 (1997)), использование DOTAP не оказывало адъювантного действия при введении ДНК-вакцины через нос, тогда как оно имело место при введении пероральным путем. DOTAP также был использован в ДНК-вакцинах, кодирующей гемагглютинин (НА) вируса гриппа, на модели мыши при внутриносовом введении (Ban и др., Vaccine, 15, 811-813 (1997)), однако введение DOTAP ингибировало иммунный ответ. Использование DC-Chol или DOTAP/DOPE в ДНК-вакцинах, кодирующей HBs антиген вируса гепатита В, на модели мыши при введении внутримышечным путем позволило усилить ответ в виде антител, тогда как использование липофектина (или DOTMA) не усилило этот ответ (Gregoriadis и др., FEBS Letters, 402, 107-110 (1997)). DC-Chol/DOPE также был использован в ДНК-вакцинах против вируса иммунодефицита человека (ВИЧ, белок Env) на модели мыши, введение которого внутримышечным путем индуцировало более эффективный иммунный ответ, тогда как введение подкожным или внутрикожным путем не усиливало его (Ishii и др., AIDS Res. Hum. Retro, 13, 1421-1428 (1997)).
Также согласно WO-A-98/40499 предлагается реализовать комплексы нуклеиновая кислота + катионные липиды для трансфекции эпителия слизистой оболочки млекопитающих, для генной терапии или экспрессии антигена, предназначенного для индуцирован иммунного ответа. В этом документе предусматривается путь введения через слизистую оболочку за счет ингаляции, например, в случае легочного эпителия. Установлено, что указанные в этом документе результаты отличаются от предшествующих работ. В этой же работе было показано, что для внутримышечного (парентерального) введения ″голая″ ДНК является более эффективной, чем смесь ДНК + липид.
Добавление некоторых цитокинов, в частности интерлейкинов или интерферонов, может усилить иммунный ответ, индуцируемый в особенности ДНК-вакцинами. Каждый цитокин вызывает реакцию, которая свойственна ему, и более или менее ориентирует иммунный ответ в направлении клеточного ответа или в направлении гуморального ответа (Pasquini и др., Immunol. Cell. Biol., 75, 397-401 (1997)); Kim и др., J. Interferon Cytokine Res., 19, 77-84 (1999)). Адъювантные действия цитокина, происходящего от данного вида, не являются обязательно одними и теми же, если изменяется иммунный контекст, особенно если этот цитокин введен другому виду, следовательно, в гетерологичную иммунную систему. Добавление цитокина может также не оказывать никакого адъювантного действия, даже приводить к инверсии искомого эффекта, то есть к уменьшению или ингибированию иммунного ответа. Так, ДНК-вакцина, кодирующая цепь иммуноглобулина, слитого с GM-CSF, не усиливает иммунный ответ, тогда как прямое введение мыши этого слитого белка является эффективным, совершенно также, как введение слитого белка, состоящего из Fv и цитокина IL-1β, или введение ДНК-вакцины, кодирующей вышеуказанный последний слитый белок (Hakim и др., J. Immunol., 157, 5503-5511 (1996)). Использование плазмид, коэкспрессирующих цитокин IL-2 и белок оболочки вируса гепатита В в слитой или неслитой форме, приводит к усилению гуморального и клеточного иммунного ответа (Chow и др., J. Virol., 71, 169-178 (1997)). Однако использование бицистронной плазмиды, кодирующей гликопротеин gp 120 вируса иммунодефицита человека (ВИЧ-1) и цитокин IL-2, индуцирует более слабый специфический анти- gp 120 иммунный ответ, чем ответ, получаемый за счет использования моноцистронной плазмиды, кодирующей только gp 120 (Barouch и др., J. Immunol., 161, 1875-1882 (1998)). Коинъекция мыши двух экспрессирующих векторов, одного, кодирующего гликопротеин G вируса бешенства, другого, кодирующего GM-CSF мыши, стимулирует активность В- и Т-лимфоцитов, тогда как коинъекция плазмиды, кодирующей гамма-интерферон (вместо GM-CSF мыши), приводит к ослаблению иммунного ответа (Xiang и др., Immunity, 2, 129-135 (1995)).
Некоторые модификации на уровне антигенов, такие, как делеции части нуклеотидной последовательности, кодирующей антиген, инсерции фрагмента ДНК в нуклеотидную последовательность, кодирующую антиген, или в нетранскрибируемые области перед и после последовательности, кодирующей антиген, могут также повышать эффективность ДНК-вакцин, особенно повышая уровень экспрессии антигена или содержащей его части.
На практике, однако, манипуляции на уровне нуклеотидной последовательности, кодирующей антиген, могут вызывать уменьшение или потерю первоначальной иммунологической активности. Так, делеция трансмембранного домена в случае гена, кодирующего антиген G вируса бешенства, снижает уровень защиты, индуцированной в случае модели мыши после введения внутримышечно ДНК-вакцины, кодирующей такой модифицированный антиген (Xiang и др., Virol., 209, 569 (1995)). Делеция трансмембранного домена в случае гена, кодирующего гликопротеин gD бычьего герпесвируса (BHV), не позволяет усилить ответ в виде антител и индуцирует только частичную защиту у быков, вакцинированных внутримышечным путем (van Drunen Little-van den Hurk и др., J. Gen. Virol., 79, 831-839 (1998)). Гуморальный и клеточный иммунный ответ и придаваемая защита идентичны у подопытных морских свинок после иммунизации ДНК-вакциной, кодирующей гликопротеин GP вируса Эбола, либо ДНК-вакциной, кодирующей гликопротеин GP в секретированной форме (Xu и др., Nature Medicine, 4, 37-42 (1998)).
Инсерция сигнальной последовательности тканевого плазминогенного активатора человека (human tissue Plasminogen Activator, или человеческий tPA) в ген, кодирующий антиген Pf332 малярии, не позволяет усиливать иммунный ответ в виде антител у мыши, вакцинированной внутримышечно (Haddad и др., FEMS, 18, 193-202 (1997)). Встраивание сигнальной последовательности tPA в ген, кодирующий антиген VP7 ротавируса мыши, также не позволяет усиливать иммунный ответ в виде антител у мыши, вакцинированной внутрикожно, тогда как слитый белок, состоящий из антигена VP4 и сигнальной последовательности tPA, позволяет усилить иммунный ответ, но без индуцирования эффективной защиты (Choi и др., Virology, 250, 230-240 (1998)).
Модификации, осуществленные в случае нуклеотидной последовательности одного антигена, обычно не могут быть прямо перенесены на другой антиген, так как антигены не всегда имеют одну и ту же структурную организацию.
Сущность изобретения
Целью заявителя является повышение эффективности ДНК-вакцин и особенно индуцирование лучшего иммунного ответа и обеспечение эффективной защиты домашних животных, и в частности собак, кошек и лошадей, путем ДНК-вакцинации при различных путях введения и, в частности, при подкожном введении.
Целью заявителя является получение улучшенных ДНК-вакцин, индуцирующих эффективный защитный и иммунный ответ против вируса чумки собак (CDV), вируса респираторного комплекса или лающего кашля (вирус парагриппа-2 или CPI-2), собачьего герпесвируса (CHV-1) у собаки.
Целью изобретателя является получение улучшенных ДНК-вакцин, индуцирующих эффективный защитный и иммунный ответ против кошачьего герпесвируса (FHV-1) у кошки.
Целью изобретателя является получение улучшенных ДНК-вакцин, индуцирующих эффективный защитный и иммунный ответ против лошадей герпесвируса типа 1 (EHV-1), лошадей герпесвируса типа 4 (EHV-4) у лошади.
Целью заявителя также является получение улучшенных ДНК-вакцин, обеспечивающих эффективную защиту собак, включающих по меньшей мере одну валентность, выбираемую из группы, состоящей из вируса CDV, вируса CPI-2, вируса CHV-1, вируса бешенства (рабдовирус), собачьего парвовируса (CPV), собачьего коронавируса (CCV) и Borrelia burgdorferi.
Целью заявителя также является получение улучшенных ДНК-вакцин, обеспечивающих эффективную защиту кошек, включающих по меньшей мере одну валентность, выбираемую из группы, состоящей из кошачьего герпесвируса (FHV-1), кошачьего калицивируса (FCV), вируса бешенства (рабдовирус), кошачьего парвовируса (FPV), кошачьего вируса инфекционного перитонита (FIPV), кошачьего вируса лейкоза (FeLV) и кошачьего вируса синдрома приобретенного иммунодефицита (FTV).
Целью заявителя также является получение улучшенных ДНК-вакцин, обеспечивающих эффективную защиту лошадей, включающих по меньшей мере одну валентность, выбираемую из группы, состоящей из лошадиного герпесвируса типа 1 (EHV-1), лошадиного герпесвируса типа 4 (EHV-4), лошадиного вируса гриппа, лошадиного вируса энцефалита Est, лошадиного вируса энцефалита Quest, лошадиного вируса энцефалита Venezuela, вируса бешенства, Clostridium tetani и Borrelia burgdorferi.
Объектом изобретения являются улучшенные ДНК-вакцины, обеспечивающие эффективную защиту против по меньшей мере одного патогена, инфицирующего домашних животных, в особенности собак, кошек и лошадей. Эффективность ДНК-вакцин улучшают либо оптимизацией способа получения приготовления, либо добавлением GM-CSF, либо оптимизацией антигена или антигенов, либо комбинацией этих подходов.
Предпочтительно ДНК-вакцины улучшают оптимизацией способа получения и необязательно добавлением GM-CSF, либо оптимизацией антигена или антигенов, либо, наконец, одновременным добавлением GM-CSF и оптимизацией антигена или антигенов.
По определению, ДНК-вакцина включает в качестве действующего начала плазмиду, кодирующую и экспрессирующую ген или фрагмент гена. Термин ″плазмида″ означает транскрипционную единицу, включающую полинуклеотидную последовательность, содержащую последовательность экспрессируемого гена и элементы, необходимые для его экспрессии in vivo. Форма плазмиды предпочтительно кольцевая, ″суперскрученная″ или нет. В рамки настоящего изобретения также входит линейная форма.
Каждая плазмида включает промотор, обеспечивающий экспрессию встроенного гена в клетке. Обычно речь идет о сильном эукариотном промоторе и, в особенности, о раннем промоторе цитомегаловируса CMV-IE человека или мыши или другого вида, например, крысы или морской свинки. Как правило, используют промотор либо вирусного происхождения, либо клеточного происхождения. В качестве вирусного промотора, другого, чем CMV-IE, можно назвать ранний или поздний промотор вируса SV40 или промотор области LTR вируса саркомы Рауса. Также можно использовать собственный промотор экспрессируемого гена. В качестве клеточного промотора можно назвать промотор гена питоскелета, такой, как, например, десминовый промотор, или актиновый промотор. Когда в одной и той же плазмиде присутствуют несколько генов, они могут находиться в одной транскрипционной единице или в двух разных транскрипционных единицах.
Согласно первому варианту, в качестве адъюванта для ДНК-вакцин согласно изобретению используют катионные липиды, содержащие соль четвертичного аммония, предпочтительно DMRIE, и более предпочтительно в комплексе с нейтральным липидом, предпочтительно DOPE, для образования DMRIE-DOPE комплекса.
Следовательно, объектом настоящего изобретения является ДНК-вакцина против по меньшей мере одного патогена, поражающего домашних животных, в особенности собак, кошек или лошадей, включающая по меньшей мере одну плазмиду, содержащую по меньшей мере одну нуклеотидную последовательность, кодирующую иммуноген патогена рассматриваемого вида животных, под контролем регуляторных элементов, обеспечивающих экспрессию этой последовательности in vivo, и катионный липид, содержащий соль четвертичного аммония формулы:
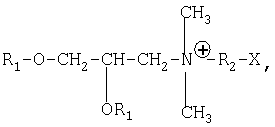
в которой
R1 означает линейный, насыщенный или ненасыщенный алифатический радикал с 12-18 атомами углерода;
R2 означает другой алифатический радикал с 2 или 3 атомами углерода; и Х означает гидроксил или аминогруппу.
Предпочтительно речь идет о DMRIE (N-(2-гидроксиэтил)-N,N-диметил-2,3-бис(тетрадецилокси)-1-пропанаммоний; WO-A-9634109), более предпочтительно в комплексе с нейтральным липидом, предпочтительно с DOPE (диолеоилфосфатидилэтаноламин), с образованием DMRIE-DOPE комплекса.
Смесь рекомбинантного вектора с таким адъювантом предпочтительно готовят перед самым употреблением, предпочтительно перед ее введением животному, инкубируют в течение времени, необходимого для образования комплекса, например, в течение времени от 10 до 60 минут, предпочтительно порядка 30 минут.
При использовании DOPE молярное соотношение DMRIE : DOPE составляет от 95:5 до 5:95, предпочтительно 1:1.
Массовое соотношение плазмида : DMRIE (или DMRIE-DOPE) составляет от 50:1 до 1:10, предпочтительно от 10:1 до 1:5, более предпочтительно от 1:1 до 1:2.
Согласно второму варианту к ДНК-вакцинам согласно изобретению добавляют GM-CSF (Granulocyte macrophage - colony stimulating factor (гранулоцит-макрофаг колониестимулирующий фактор); Clark S.C. и др., Scienct, 230, 1229 (1987); Grant S.M. и др., Drngs, 53, 516 (1992)), что можно осуществлять или путем добавления белка GM-CSF непосредственно в вакцинную композицию или предпочтительно путем встраивания последовательности, кодирующей GM-CSF, в экспрессирующий вектор, обеспечивающий его экспрессию in vivo. В качестве экспрессирующего вектора предпочтительно используют плазмиду, например плазмиду, содержащую нуклеотидную последовательность, кодирующую представляющий интерес антиген или представляющие интерес антигены или другую плазмиду. Видовую принадлежность GM-CSP выбирают в зависимости от вида вакцинируемого животного; так, для собак используют GM-CSF собаки, для кошек используют GM-CSF кошки и для лошадей используют GM-CSF лошадей.
Согласно третьему варианту нуклеотидная последовательность (или нуклеотидные последовательности), кодирующая иммуноген, находится в оптимизированной форме. Под оптимизацией понимают любую модификацию нуклеотидной последовательности, усиливающую экспрессию этой нуклеотидной последовательности путем повышения стабильности мРНК, кодирующей этот антиген, или путем обеспечения секреции этого антигена во внеклеточную среду, приводящую к усилению индуцированного иммунного ответа.
Согласно настоящему изобретению оптимизация представляющего интерес антигена предпочтительно состоит в делеции фрагмента нуклеотидной последовательности, кодирующего трансмембранный домен представляющего интерес антигена (под делецией понимают полную делецию или частичную делецию, достаточную для того, чтобы трансмембранный домен не был более функциональным), и/или встраивание нуклеотидной последовательности, кодирующей сигнальный пептид tPA (tissue plasminogen activator (тканевый плазминогенный активатор); Montgomery и др., Cell. Mol. Biol., 43, 285-292 (1997); Harris и др., Mol. Biol. Med., 3, 279-292 (1986)), и/или встраивание интрона-стабилизатора перед экспрессируемым геном. Делеция фрагмента, кодирующего трансмембранный домен представляющего интерес антигена, благоприятствует секреции во внеклеточную среду укороченных антигенов и, таким образом, повышает их способность контактировать с клетками иммунной системы. Встраивание нуклеотидной последовательности, кодирующей сигнальный пептид tPA, облегчает транслируемость содержащей эту последовательность РНК и, таким образом, увеличивает уровень экспрессии этой мРНК и, следовательно, продуцирование антигенов. Сигнальный пептид tPA также играет роль в секреции синтезированного антигена. Инсерция интрона-стабилизатора в ген, кодирующий представляющий интерес антиген, позволяет избегать аберрантных связывании его информационной РНК и сохраняет физическую целостность последней.
Предпочтительно используют сигнальный пептид tPA человека. Нуклеотидная последовательность сигнального пептида tPA человека доступна из базы данных банка генов под номером NM_000930. Интрон предпочтительно представляет собой интрон II гена бета-глобина кролика (van Ooyen и др., Science, 206, 337-344 (1979)), нуклеотидная последовательность которого доступна из базы данных банка генов под номером V00882 и находится под названием интрон №2.
Объектом настоящего изобретения является улучшенная ДНК-вакцина, способная индуцировать эффективный иммунный и защитный ответ у собаки против чумки собак (по-английски Canine Distemper Virus (вирус собачьей чумы), или CDV).
Болезни Карре вызывает Morbillivirus, относящийся к Paramyxoviridae. Этот вирус вызывает инфекцию не только у собак, но также у диких кошек (Harder и др., J. Gen. Virol., 77, 397-405 (1996)).
Согласно настоящему изобретению можно получать эффективную защищающую собак против болезни Карре ДНК-вакцину, предпочтительно при введении подкожно (Sixt и др., J. Virol., 72, 8472-8476 (1998)).
Согласно изобретению, ДНК-вакцина против CDV получают в виде комплекса с адъювантом согласно изобретению, предпочтительно DMRIE, более предпочтительно DMRIE-DOPE. ДНК-вакцину в комплексе с адъювантом необязательно можно вводить либо в комбинации с GM-CSF собаки (Nash и др., Blood, 78, 50-56 (1991)), либо оптимизировать по меньшей мере один антиген CDV, наконец, либо с добавлять GM-CSF собаки и оптимизировать по меньшей мере один антиген CDV.
Нуклеотидная последовательность, кодирующая GM-CSF собаки, доступна из базы данных банка генов под номером S49738.
GM-CSF собаки можно добавлять либо непосредственно в вакцинную композицию или более предпочтительно путем встраивания нуклеотидной последовательности, кодирующей GM-CSF собаки, в экспрессирующий in vivo вектор, предпочтительно в плазмиду. Нуклеотидную последовательность, кодирующую GM-CSF собаки, предпочтительно встраивают во вторую плазмиду экспрессии, отличную от той (тех), в которую (в которые) встроен ген или гены, кодирующие антиген или антигены CDV.
Оптимизацию происходящих от CDV антигенов осуществляют путем замены сигнальной последовательности антигена (гемагглютинин (НА) и/или слитый белок (F)), предпочтительно сигнальной последовательности tPA человека (банк генов, номер NM 000930), и/или путем делеции фрагмента, кодирующего трансмембранный домен НА и/или F, и/или путем инсерции интрона, предпочтительно интрона II гена бета-глобина кролика (нуклеотидная последовательность которого, обозначаемая как интрон № 2, доступна из базы данных банка генов под номером V00882) перед нуклеотидной последовательностью, кодирующей НА и/или F. ДНК-вакцина против CDV согласно изобретению, следовательно, может кодировать и экспрессировать либо один оптимизированный антиген CDV (НА или F) или оба антигена, то есть оптимизированный НА и оптимизированный F.
Необязательно последовательность, кодирующая природный матриксный белок (М) CDV (без модификации), и/или нуклеотидная последовательность, кодирующая природный нуклеопротеин (N) CDV (без модификации), могут быть также встроены в экспрессирующие плазмиды, причем последние могут быть использованы вместе с плазмидами, содержащими оптимизированный НА и/или оптимизированный F.
Нуклеотидные последовательности, кодирующие антигены CDV, используемые согласно настоящему изобретению, и различные конструкции экспрессирующих векторов проиллюстрированы в прилагаемых примерах и в Международной заявке WO-A-98031199, в особенности в приведенных в ней примерах 8 и 9 и на представленных в ней фигурах 2 и 3.
Согласно изобретению ДНК-вакцину против CDV при введении внутримышечно предпочтительно получают в виде комплекса с DMRIE-DOPE, и вакцина включает две экспрессирующие плазмиды, одна из которых (например, pNS024, фигура 4) содержит антиген НА CDV, оптимизированный путем замены сигнальной последовательности НА сигнальной последовательностью tPA человека, делеции фрагмента, кодирующего трансмембранный домен НА, и инсерции интрона II гена бета-глобина кролика перед геном НА, а другая плазмида (например, pNS021, фигура 3), содержит антиген F CDV, оптимизированный путем делеции фрагмента, кодирующего трансмембранный домен, и инсерции интрона II гена бета-глобина кролика перед геном F.
Согласно изобретению ДНК-вакцину на основе против CDV для введения подкожным путем предпочтительно готовят в виде комплекса с DMRIE-DOPE, и вакцина включает плазмиду, экспрессирующую GM-CSF собаки, и две вышеуказанные плазмиды (например, pNS024 и pNS021).
Объектом настоящего изобретения также является улучшенная ДНК-вакцина, способная индуцировать эффективный защитный и иммунный ответ у собаки против респираторного комплекса или лающего кашля (вирус собачьего парагриппа-2 или CPI-2).
Вирус CPI-2 представляет собой Paramyxovirus и относится к Paramyxoviridae (Bittle и др., J. Am. Vet. Med. Assoc., 156, 1771-1773 (1970); Moloney и др., Aust. Vet. J., 62, 285-286 (1985)).
ДНК-вакцину против CPI-2 предпочтительно готовят в виде комплекса с адъювантом согласно изобретению, предпочтительно DMRIE, более предпочтительно в виде комплекса с DMRIE-DOPE. ДНК-вакцину необязательно можно вводить в комбинации с GM-CSF собаки или использовать хотя бы один оптимизированный антиген CPI-2, или вакцина может содержать по меньшей мере один оптимизированный антиген CPI-2 вместе с GM-CSF собаки.
GM-CSF собаки добавляют также как и при вакцинации против CDV.
Оптимизацию антигенов CPI-2 (гемагглютининнейраминидаза (HN) CPI-2 и/или слитый белок (F) CPI-2) осуществляют путем замены сигнальной последовательности, антигена, предпочтительно сигнальной последовательностью tPA человека и/или путем делеции фрагмента, кодирующего трансмембранный домен HN и/или F, и/или путем инсерции интрона, предпочтительно интрона II гена бета-глобина кролика, перед геном, кодирующим HN и/или F. ДНК-вакцина против CPI-2 согласно изобретению, следовательно, может кодировать и экспрессировать или один (HN или F) или оба (HN и F) оптимизированных антигенов CPI-2.
Нуклеотидные последовательности, кодирующие антигены CPI-2, используемые согласно настоящему изобретению, и различные конструкции экспрессирующих векторов подробно рассмотрены в прилагаемых примерах.
ДНК-вакцину против CPI-2 для введения внутримышечно предпочтительно готовят в виде комплекса с DMRIE-DOPE, и такая вакцина включает две экспрессирующих плазмиды, одна из которых (например, pSB034, фигура 6), содержит антиген HN CPI-2, оптимизированный путем замены сигнальной последовательности HN сигнальной последовательностью tPA человека, делеции фрагмента HN, кодирующего трансмембранный домен, и инсерции интрона II гена бета-глобина кролика перед геном HN, а вторая плазмида (например, pSB032, фигура 5) содержит антиген F CPI-2, оптимизированный путем делеции фрагмента, кодирующего трансмембранный домен F и инсерции интрона II гена бета-глобина кролика перед геном F.
ДНК-вакцину против CPI-2 согласно изобретению, для введения подкожно предпочтительно готовят в виде комплекса с DMRIE-DOPE, и эта вакцина включает плазмиду, экспрессирующую GM-CSF собаки, и две вышеуказанные плазмиды (например, pSB034 и pSB032).
Объектом настоящего изобретения является также улучшенная ДНК-вакцина, способная индуцировать эффективный защитный и иммунный ответ у собаки против собачьего герпесвируса типа 1 (CHV-1).
Вирус CHV-1 относится к Alphaherpesvirinae. Этот вирус вызывает у собак вирусный ринотрахеит. Нуклеотидные последовательности, кодирующие гликопротеины gB, gC и gD, известны (Limbach и др., J. Gen. Virol., 75, 2029-2039 (1994)).
ДНК-вакцину против CHV-1 предпочтительно готовят в виде комплекса с адъювантом согласно изобретению, предпочтительно DMRIE, более предпочтительно DMRIE-DOPE. Вакцину можно необязательно вводить в комбинации с GM-CSF собаки, или использовать по меньшей мере один антиген CHV-1, или можно использовать по меньшей мере один оптимизированный антиген CHV-1 в комбинации с GM-CSF собаки.
GM-CSF собаки добавляют так же, как и при вакцинации против CDV.
Оптимизацию антигенов CHV-1 осуществляют путем делеции фрагмента, кодирующего трансмембранный домен гликопротеина gB, и/или гликопротеина gC, и/или гликопротеина gD CHV-1. Улучшенная ДНК-вакцина против CHV-1 согласно изобретению, следовательно, может кодировать и экспрессировать либо один оптимизированный антиген CHV-1 (gB, gC или gD), либо любые два из них или все три.
Нуклеотидные последовательности, кодирующие антигены CHV-1, используемые согласно настоящему изобретению, и различные конструкции экспрессирующих векторов подробно рассмотрены в прилагаемых далее примерах, а также в Международной заявке WO-A-98/03199 и, в частности, в приведенных в ней примерах 7 и 8 и на представленных в ней фигурах 13 и 14.
ДНК вакцину против CHV-1 для введения внутримышечно предпочтительно готовят в виде комплекса с DMRIE-DOPE. Такая вакцина включает первую экспрессирующую плазмиду (например, pSB016, фигура 7), кодирующую антиген gB CHV-1, оптимизированный путем делеции фрагмента, кодирующего трансмембранный домен, и вторую экспрессирующую плазмиду (например, pSB019, фигура 8), кодирующую антиген gC CHV-1, оптимизированный путем делеции фрагмента кодирующего трансмембранный домен F, и третью экспрессирующую плазмиду (например, pSB017, фигура 9), кодирующую антиген gD CHV-1, оптимизированный путем делеции фрагмента, кодирующего трансмембранный домен.
ДНК-вакцину против CHV-1 для введения подкожно предпочтительно готовят в виде комплекса с DMRIE-DOPE. Такая вакцина включает экспрессирующую плазмиду, кодирующую GM-CSF собаки, и три вышеуказанные плазмиды (например, pSB016, pSB019 и pSB017).
Объектом настоящего изобретения является улучшенная ДНК-вакцина, способная индуцировать эффективный защитный и иммунный ответ у кошки против герпесвируса типа 1 (FHV-1) кошки.
Вирус FHV-1 относится к Alphaherpesvirinae и вызывает вирусный ринотрахеит у кошек (Fargeaud и др., Arch. Virol., 80, 69-82 (1984)).
ДНК-вакцину против FHV-1 предпочтительно готовят в виде комплекса с адъювантом согласно изобретению, предпочтительно с DMRIE, более предпочтительно с DMRIE-DOPE. Вакцину необязательно можно добавлять в комбинации с GM-CSF кошки, либо оптимизировать по меньшей мере один антиген FHV-1, либо, наконец, комбинировать добавление GM-CSF кошки и оптимизацию по меньшей мере одного антигена FHV-1.
GM-CSF кошки вводят либо путем добавления белка GM-CSF непосредственно в вакцинную композицию или путем встраивания нуклеотидной последовательности, кодирующей GM-CSF кошки (например, доступной из базы данных банка генов под номером AF053007), в экспрессирующий in vivo вектор, предпочтительно в плазмиду. Нуклеотидную последовательность, кодирующую GM-CSF кошки, предпочтительно встраивают во вторую экспрессирующую плазмиду, отличную от той (тех), в которую (в которые) встроен ген или гены, кодирующие антиген или антигены FHV-1.
Оптимизацию FHV-1 антигенов осуществляют путем делеции фрагмента, кодирующего трансмембранные домены гликопротеина gB и/или, гликопротеина gC, и/или гликопротеина gD FHV-1. Улучшенная ДНК-вакцина против FHV-1 согласно изобретению, следовательно, может кодировать и экспрессировать один оптимизированный антиген FHV-1 (gB, gC или gD), любые два из них или все три.
Нуклеотидные последовательности, кодирующие антигены FHV-1, используемые согласно настоящему изобретению, и различные конструкции экспрессирующих векторов представлены в прилагаемых примерах и в Международной заявке WO-A-98/03660 и, в частности, в приведенных в ней примерах 14 и 15 и на представленных в ней фигурах 11 и 12.
Согласно изобретению ДНК-вакцину против FHV-1 для введения внутримышечно предпочтительно готовят в виде комплекса с DMRIE-DOPE. Такая вакцина включает первую плазмиду (например, pSB021, фигура 10), экспрессирующую антиген gB FHV-1, оптимизированный путем делеции фрагмента, кодирующего трансмембранный домен, вторую плазмиду (например, pSB023, фигура 11), экспрессирующую антиген gC FHV-1, оптимизированный путем делеции фрагмента, кодирующего трансмембранный домен F, и третью плазмиду (например, pSB024, фигура 12), экспрессирующую антиген gD FHV-1, оптимизированный путем делеции фрагмента, кодирующего трансмембранный домен.
Согласно изобретению ДНК-вакцину против FHV-1 для введения подкожно предпочтительно готовят в виде комплекса с DMRIE-DOPE. Такая вакцина включает плазмиду, экспрессирующую GM-CSF кошки, и три вышеуказанные плазмиды (например, pSB021, pSB023 и pSB024).
Объектом настоящего изобретения является улучшенная ДНК-вакцина, способная индуцировать эффективный защитный и иммунный ответ у лошади против герпесвируса типа 1 (EHV-1) лошадей.
Вирус EHV-1 относится к Alphaherpesvirinae. Вирус EHV-1 вызывает аборт у лошадей (Crabb и др., Adv. Virus Res., 45, 153-190 (1995)). Полный геном этого вируса определен (Telford и др., Virilogy, 189, 304-316 (1992)).
ДНК-вакцину против EHV-1 предпочтительно готовят в виде комплекса с адъвантом согласно изобретению, предпочтительно с DMRIE, более предпочтительно с DMRIE-DOPE. Вакцину необязательно добавляют комбинации с GM-CSF лошади, либо оптимизируют по меньшей мере один антиген EHV-1, либо, наконец, комбинируют добавление GM-CSF лошади и оптимизацию по меньшей мере одного антигена EHV-1.
GM-CSF лошади добавляют либо непосредственно в вакцинную композицию, либо встраивают нуклеотидную последовательность (например, последовательности № 69, фигура 26), кодирующей GM-CSF лошади, в экспрессирующий in vivo вектор, предпочтительно в плазмиду. Нуклеотидную последовательность, кодирующую GM-CSF лошади, предпочтительно встраивают во вторую экспрессирующую плазмиду, отличную от той (или тех), в которую (в которые) встроен ген или гены, кодирующие антиген или антигены EHV-1.
Оптимизацию антигенов EHV-1 осуществляют путем делеции фрагмента, кодирующего трансмембранные домены гликопротеина gB, и/или гликопротеина gC, и/или гликопротеина gD EHV-1. Улучшенная ДНК-вакцина против EHV-1 согласно изобретению, следовательно, может кодировать и экспрессироватъ один оптимизированный антиген EHV-1 (gB, gC или gD) или два из них или все три.
Нуклеотидные последовательности, кодирующие антигены EHV-1, используемые согласно настоящему изобретению, и различные конструкции экспрессирующих векторов представлены в прилагаемых примерах и в Международной заявке WO-A-98/03198, особенно в приведенных в ней примерах 8 и 10 и на представленных в ней фигурах 2 и 4.
Для лошади предпочтительным является внутримышечное введение. Предпочтительно согласно изобретению ДНК-вакцину против EHV-1 для введения внутримышечно готовят в виде комплекса с DMRIE-DOPE. Такая вакцина включает первую плазмиду (например, pSB028, фигура 13), экспрессирующую антиген gB EHV-1, оптимизированный путем делеции фрагмента, кодирующего трансмембранный домен, вторую плазмиду (например, pSB029, фигура 14), экспрессирующую антиген gC EHV-1, оптимизированный путем делеции фрагмента, кодирующего трансмембранный домен, и третью плазмиду (например, pSB030, фигура 15), экспрессирующую антиген gD EHV-1, оптимизированный путем делеции фрагмента, кодирующего трансмембранный домен.
При подкожном введении ДНК-вакцину против EHV-1 предпочтительно готовят в виде комплекса с DMRIE-DOPE, и в этом случае вакцина включает плазмиду, экспрессирующую GM-CSF лошади, и три вышеуказанные плазмиды (например, pSB028, pSB029 и pSB030).
Объектом настоящего изобретения является улучшенная ДНК-вакцина, способная индуцировать эффективный защитный и иммунный ответ у лошади против герпесвируса типа 4 (EHV-4) лошадей.
Вирус EHV-4 относится к Alphaherpesvirinae. Вирус EHV-4 вызывает вирусную ринопневмонию у лошадей (Crabb и др., Adv. Virus Res., 45, 153-190 (1995)). В настоящее время полный геном этого вируса определен (Telford и др., J. Gen. Virol, 79, 1197-1203 (1998)).
ДНК-вакцину против EHV-4 предпочтительно готовят в виде комплекса с адъювантом согласно изобретению, предпочтительно с DMRIE, более предпочтительно с DMRIE-DOPE. ДНК-вакцину необязательно комбинируют либо с добавлением GM-CSF лошадей, либо с оптимизацией по меньшей мере одного антигена EHV-4, либо, наконец, с как добавлением GM-CSF лошади, так и оптимизацией по меньшей мере одного антигена EHV-4.
GM-CSF лошади включают, как это описано для EHV-1.
Оптимизацию антигенов EHV-4 осуществляют путем делеции фрагмента, кодирующего трансмембранный домен гликопротеина gB, и/или гликопротеина gC, и/или гликопротеина gD EHV-4. Улучшенная ДНК-вакцина против EHV-4 согласно изобретению, следовательно, может кодировать и экспрессировать один оптимизированный антиген EHV-4 (gB, gC или gD), два из них или все три.
Нуклеотидные последовательности, кодирующие антигены EHV-4, используемые согласно настоящему изобретению, и различные конструкции экспрессирующих векторов представлены в прилагаемых примерах и в Международной заявке WO-A-98/03198 и, в частности, в приведенных в ней примерах 9 и 11 и на представленных в ней фигурах 3 и 5.
Согласно изобретению ДНК-вакцину против EHV-4 для введения внутримышечно предпочтительно готовят в виде комплекса с DMRIE-DOPE. Такая вакцина включает первую плазмиду (например, pSB025, фигура 16), экспрессирующую антиген gB EHV-4, оптимизированный путем делеции фрагмента, кодирующего трансмембранный домен, вторую плазмиду (например, pSB026, фигура 17), экспрессирующую антиген gC EHV-4, оптимизированный путем делеции фрагмента, кодирующего трансмембранный домен, и третью плазмиду (например, pSB027, фигура 18), экспрессирующую антиген gD EHV-4, оптимизированный путем делеции фрагмента, кодирующего трансмембранный домен.
Для подкожного введения ДНК-вакцину против EHV-4 предпочтительно готовят в виде комплекса с DMRIE-DOPE, и она включает плазмиду, экспрессирующую GM-CSF лошади, и три вышеуказанные плазмиды (например, pSB025, pSB026 и pSB027).
Хотя изобретение описывается в связи с конкретными ДНК-вакцинами, оно относится также к ДНК-вакцинам против других патогенов этих видов животных. Так, различные валентности, описанные в настоящей заявке, могут составлять объект улучшенной вакцины, предпочтительно путем добавления адъюванта согласно изобретению, или GM-CSF, или, в случае необходимости, путем оптимизации гена, или путем сочетаний указанных предложений, как это описывается здесь подробно для некоторых валентностей.
В том же самом порядке представления вакцины согласно изобретению могут быть для одного животного вида комбинированными друг с другом и/или с ДНК-вакцинами против других патогенов того же вида.
Таким образом, объектом настоящего изобретения также являются улучшенные многовалентные ДНК-вакцины, обеспечивающие в случае собак эффективную защиту против по меньшей мере двух патогенов, выбираемых из группы, состоящей из CDV, CPI-2, CHV-1, вируса бешенства (рабдовирус), парвовируса (CPV) собак, коронавируса (CCV) собак или Borrelia burgdorferi.
Объектом настоящего изобретения также являются улучшенные многовалентные ДНК-вакцины, обеспечивающие в случае кошек эффективную защиту против по меньшей мере двух патогенов кошек, выбираемых из группы, состоящей из FHV-1, калицивируса (FCV) кошек, вируса бешенства (рабдовирус), парвовируса (FPV) кошек, вируса инфекционного перитонита (FIPV) кошек, вируса лейкоза (FeLV) кошек, вируса синдрома приобретенного иммунодефицита (FIV) кошек.
Объектом настоящего изобретения также являются улучшенные многовалентные ДНК-вакцины, обеспечивающие в случае лошади эффективную защиту против по меньшей мере двух патогенов лошади, выбираемых из группы, состоящей из EHV-1, EHV-4, вируса гриппа лошадей, вируса энцефалита Est лошадей, вируса энцефалита лошадей Ouest, вируса энцефалита лошадей Venezuela, вируса бешенства, Clostridium tetani, Borrelia burgdorferi.
Многовалентные ДНК-вакцины могут быть улучшены при введении в виде комплекса с адьювантом согласно изобретению, предпочтительно с DMRIE, более предпочтительно с DMRIE-DOPE. Введение таких вакцин необязательно комбинируют либо с добавлением GM-CSF, как это описано выше, либо с оптимизацией по меньшей мере одного представляющего интерес антигена, как это описано выше, либо, наконец, с как добавлением GM-CSF, так и с оптимизацией по меньшей мере одного представляющего интерес антигена.
Улучшенные ДНК-многовалентные вакцины согласно изобретению включают одну или несколько экспрессирующих плазмид, обеспечивающих экспрессию in vivo no меньшей мере одного иммуногена первого патогена и по меньшей мере одного иммуногена по меньшей мере одного другого патогена, инфицирующего тот же вид животных. По меньшей мере один из этих иммуногенов предпочтительно выбирают из группы:
- F CDV, HA CDV, F CPI-2, HN CPI-2, gB CHV-1, gC CHV-1 и gD CHV-1, для собак;
- gB FHV-1, gC FHV-1 и gD FHV-1, для кошек; и
- gB EHV-1, gC EHV-1, gD EHV-1, gB EHV-4, gC EHV-4 и gD EHV-4, для лошадей.
Улучшенные моновалентные или многовалентные ДНК-вакцины согласно изобретению можно также комбинировать по меньшей мере с одной классической (инактивированной, ослабленной живой, субъединичной) вакциной или рекомбинантной вакциной, использующей экспрессирующий in vivo вектор (например, поксвирус, аденовирус, герпесвирус), против по меньшей мере одного патогена, инфицирующего тот же самый вид.
Методы конструирования плазмид приведены в Международной заявке WO-A-98/03198 (вакцины для лошадей), в Международной заявке WO-A-98/03660 (вакцины для кошек) и в Международной заявке WO-A-98/03199 (вакцины для собак).
Объектом настоящего изобретения является также способ вакцинации домашних животных, предпочтительно собак, кошек или лошадей. Этот способ вакцинации включает введение одной из улучшенных, моновалентных или многовалентных, ДНК-вакцин, таких, как описанные выше. Способ вакцинации также включает введение одной или нескольких доз улучшенной ДНК-вакцины.
Количество ДНК, используемое в вакцинах согласно изобретению, составляет от 10 мкг до 1000 мкг, предпочтительно от 50 мкг до 500 мкг для каждой плазмиды. Специалист обладает необходимыми знаниями для точного определения эффективной дозы ДНК, используемой для вакцинации.
Объемы дозы предпочтительно могут составлять от 0,5 мл до 5 мл, предпочтительно от 1 мл до 3 мл.
Улучшенные ДНК-вакцины согласно изобретению могут быть введены в рамках этого способа вакцинации различными путями, предлагаемыми в уровне техники для ДНК-вакцинации, и с помощью известных способов введения.
Согласно двум предпочтительным вариантам осуществления изобретения способы вакцинации включают внутримышечное или подкожное введение улучшенных ДНК-вакцин согласно изобретению.
Перечень фигур
Изобретение теперь более подробно описывается с помощью вариантов осуществления, приводимых в качестве не ограничивающих объема охраны изобретения примеров и со ссылкой на рисунок, на котором представлено:
фигура 1: плазмида рАВ110
фигура 2: плазмида pVR1012
фигура 3: плазмида pNS021
фигура 4: плазмида pNS024
фигура 5: плазмида pSB032
фигура 6: плазмида pSB034
фигура 7: плазмида pSB016
фигура 8: плазмида pSB019
фигура 9: плазмида pSB017
фигура 10: плазмида pSB021
фигура 11: плазмида pSB023
фигура 12: плазмида pSB024
фигура 13: плазмида pSB028
фигура 14: плазмида pSB029
фигура 15: плазмида pSB030
фигура 16: плазмида pSB025
фигура 17: плазмида pSB026
фигура 18: плазмида pSB027
фигура 19: плазмида pJP084
фигура 20: последовательность гена GM-CSF собаки
фигура 21: плазмида pJP089
фигура 22: последовательность гена GM-CSF 3R3 кошки
фигура 23: плазмида pJP090
фигура 24: последовательность гена GM-CSF 3R4 кошки
фигура 25: плазмида pJP097
фигура 26: последовательность гена GM-CSF лошади
фигура 27: последовательность гена CPI-2 F
фигура 28: последовательность гена CPI-2 HN
Список последовательностей:
последовательность №1: олигонуклеотид NS030
последовательность №2: олигонуклеотид NS031
последовательность №3: олигонуклеотид NS034
последовательность №4: олигонуклеотид NS035
последовательность №5: олигонуклеотид NS036
последовательность №6: олигонуклеотид NS037
последовательность №7: олигонуклеотид SB090
последовательность №8: олигонуклеотид SB091
последовательность №9: олигонуклеотид РВ326
последовательность №10: олигонуклеотид РВ329
последовательность №11: олигонуклеотид РВ381
последовательность №12: олигонуклеотид РВ382
последовательность №13: олигонуклеотид РВ383
последовательность №14: олигонуклеотид РВ384
последовательность №15: олигонуклеотид SB101
последовательность №16: олигонуклеотид SB102
последовательность №17: олигонуклеотид SB103
последовательность №18: олигонуклеотид SB104
последовательность №19: олигонуклеотид SB105
последовательность №20: олигонуклеотид SB106
последовательность №21: олигонуклеотид SB107
последовательность №22: олигонуклеотид SB108
последовательность №23: олигонуклеотид SB109
последовательность №24: олигонуклеотид SB110
последовательность №25: олигонуклеотид SB111
последовательность №26: олигонуклеотид SB112
последовательность №27: олигонуклеотид SB113
последовательность №28: олигонуклеотид SB114
последовательность №29: олигонуклеотид SB115
последовательность №30: олигонуклеотид SB116
последовательность №31: олигонуклеотид SB117
последовательность №32: олигонуклеотид SB118
последовательность №33: олигонуклеотид АВ325
последовательность №34: олигонуклеотид АВ326
последовательность №35: олигонуклеотид АВ327
последовательность №36: олигонуклеотид АВ328
последовательность №37: олигонуклеотид АВ329
последовательность №38: олигонуклеотид АВ330
последовательность №39: олигонуклеотид NS003
последовательность №40: олигонуклеотид NS004
последовательность №41: олигонуклеотид NS005
последовательность №42: олигонуклеотид NS006
последовательность №43: олигонуклеотид NS007
последовательность №44: олигонуклеотид NS008
последовательность №45: олигонуклеотид SB119
последовательность №46: олигонуклеотид SB120
последовательность №47: олигонуклеотид SB121
последовательность №48: олигонуклеотид SB122
последовательность №49: олигонуклеотид SB123
последовательность №50: олигонуклеотид SB124
последовательность №51: олигонуклеотид SB125
последовательность №52: олигонуклеотид SB126
последовательность №53: олигонуклеотид SB127
последовательность №54: олигонуклеотид SB128
последовательность №55: олигонуклеотид SB129
последовательность №56: олигонуклеотид SB130
последовательность №57: олигонуклеотид SB131
последовательность №58: олигонуклеотид SB132
последовательность №59: олигонуклеотид SB133
последовательность №60: олигонуклеотид SB134
последовательность №61: олигонуклеотид SB135
последовательность №62: олигонуклеотид SB136
последовательность №63: олигонуклеотид SB137
последовательность №64: олигонуклеотид JP578
последовательность №65: олигонуклеотид JP579
последовательность №66: последовательность гена
GM-CSF собаки (см. фигуру 20)
последовательность №67: последовательность гена
GM-CSF 3R3 кошки (см. фигуру 22)
последовательность №68: последовательность гена
GM-CSF 3R4 кошки (см. фигуру 24)
последовательность №69: последовательность гена
GM-CSF лошади (см. фигуру 26)
последовательность №70: олигонуклеотид JP734
последовательность №71: олигонуклеотид JP735
последовательность №72: последовательность гена CPI-2 F
(см. фигуру 27)
последовательность №73: последовательность гена CPI-2 HN
(см. фигуру 28)
Сведения, подтверждающие возможность осуществления изобретения
Для всех рассматриваемых патогенов каждый ген, как в природной, так и в модифицированной форме, кодирующий главные поверхностные антигены, клонировали в отдельной эукариотической экспрессирующей плазмиде. Секретируемые формы поверхностных антигенов получали путем делеции фрагментов, кодирующих трансмембранные и цитоплазматические домены. Во всех случаях трансмембранные домены гликопротеинов были определены на основе соответствующих аминокислотных последовательностей. Длина белков дикого типа с указанием трансмембранных доменов, укороченных белков, а также названия соответствующих экспрессирующих плазмид приведены в таблице в примере 11.
Пример 1
Базовые плазмидные конструкции
Эукариотическая экспрессирующая плазмида pVR1020 (C.J.Luke и др., J. of Infectious Diseases, 175, 95-97 (1997)), сконструированная на основе pVR1012 (фигура 1, пример 7 WO-A-9803199 или фигура 2 настоящей заявки), содержит область, кодирующую сигнальную последовательность тканевого активатора плазминогена (tPA) человека. Плазмиду pVR1020 модифицировали путем расщепления BamHI-BglII и встраивания полилиннерра, содержащего несколько сайтов клонирования (BamHI, NotI, EcoRi, Xbal, Pmll, PstI, BglII), образующего в результате лигирования следующих олигонуклеотидов в качестве праймеров:
РВ326 (40 нт) (последовательность №9)
5' GATCTGCAGCACGTGTCTAGAGGATATCGAATTCGCGGCC3' и
РВ329 (40 нт) (последовательность №10)
5' GATCCGCGGCCGCGAATTCGATATCCTCTAGACACGTGCT 3'.
Полученный в результате вектор, рАВ110 (фигура 1) использовали для конструкции плазмид, содержащих укороченные формы генов, кодирующих гемагглютинин (НА) вируса чумки собак (CDV) и гемагглютининнейраминидазу (HN) вируса парагриппа типа 2 (CPI-2).
Интрон II гена β-глобина кролика клонировали в векторе pCRII (Invitrogen, Карлсбад, Канада, США) путем полимеразной цепной реакции (PCR) с использованием следующих олигонуклеотидов в качестве примеров:
SB090 (20 нт) (последовательность №7)
5' TTGGGGACCCTTGATTGTTC 3' и
SB091 (21 нт) (последовательность №8)
5' CTGTAGGAAAAAGAAGAAGGC 3',
и тотальную ДНК клеток периферической крови кролика в качестве матрицы. В результате была получена плазмида pNS050.
Пример 2
Плазмиды, кодирующие различные формы антигенов CDV
Гены, кодирующие слитый белок (F) и гемагглютинин (НА) CDV штамма Snyder Hill (SH), получали с помощью полимеразной цепной реакции с обратной транскриптазой (RT-PCR) с использованием вирусной ДНК штамма SH (доступного из библиотеки штаммов Американской коллекции тканевых культур под номером АТСС VR-526) в качестве матрицы.
2.1. Плазмиды, кодирующие различные формы CDV-F
2.1.1. Получение плазмиды рРВ229: клонирование гена F (природная форма) в плазмиде pVR1012
кДНК гена F CDV синтезировали с использованием в качестве праймера РВ383 и амплифицировали с помощью полимеразной цепной реакции с использованием следующей пары олигонуклеотидов в качестве праймеров:
РВ383 (26 нт) (последовательность №13)
5' TTTCTAGACAGCCGAGCCCCATGCAC 3' и
РВ384 (30 нт) (последовательность №14)
5' TTGGATCCGATATATGACCAGAATACTTCA 3'.
Продукт полимеразной цепной реакции рестриктировали с помощью BamHI и XbaI и клонировали в экспрессирующем векторе pVR1012 (пример 1), предварительно рестриктированном BamHI и XbaI. В результате была получена плазмида рРВ229 (6925 пар оснований). Ген F дикого типа CDV кодирует белок из 662 аминокислот.
2.1.2. Получение плазмиды DNS021: клонирование гена F (β-глобиновая форма F ΔТМ) в плазмиде pVR1012
Плазмиду pNS013 (6735 пар оснований), содержащую укороченный ген F (делеция трансмембранного и С-концевого домена), получали лигированием фрагмента Bsu361-BamHI (6593 пары оснований) плазмиды рРВ229 (пример 2.1.1.) и фрагмента (142 пар оснований), полученного с помощью полимеразной цепной реакции с использованием рРВ229 в качестве матрицы и следующих олигонуклеотидов в качестве праймеров:
NS030 (21 нт) (последовательность №1)
5' ATGAGCCCACTCTTACAACAA 3' и
NS031 (35 нт) (последовательность №2)
5' TTTCGCGGATCCATTAAAGGAAGAGCGCCTAACCG 3'
рестриктированного Bsu361-BamHI. Укороченный ген F CDV кодирует белок из 605 аминокислот.
Затем последовательность интрона II гена β-глобина кролика встраивали перед последовательностью, кодирующей укороченный ген F, по сайгу SalI плазмиды pNS013. Соответствующий интрону фрагмент ДНК (573 пары оснований) получали с помощью полимеразной цепной реакции с использование следующих олигонуклеотидов в качестве праймеров:
NS036 (34 нт) (последовательность №5)
5' TTTACGCGTCGACTTGGGGACCCTTGAATTGTTC 3' и
NS037 (36 нт) (последовательность №6)
5' TTTACGCGTCGACCTGTAGGAAAAAGAAGAAGGCAT 3'
pNS050 в качестве матрицы (пример 1) с последующей рестрикцией SalI. Плазмиду pNS013, содержащую интрон II гена β-глобина, назвали pNS021 (7308 пар оснований) (фигура 3).
2.2. Плазмиды, кодирующие различные формы CDV-HA
2.2.1. Получение плазмиды pNS018: клонирование гена НА (природная форма) в плазмиде pVR1012
кДНК гена НА CDV синтезировали с использованием РВ381 в качестве праймера и амплифицировали с помощью полимеразной цепной реакции с использованием следующей пары олигонуклеотидов в качестве праймеров:
РВ381 (30 нт) (последовательность №11)
5' TTCTGCAGATGCTCCTCCTACCAAGAYAAGG 3' и
РВ382 (28 нт) (последовательность №12)
5' TTGTCGACATGTGTATCATCATMCTGTC 3'
Продукт полимеразной цепной реакции клонировали в векторе pCRII (Invitrogen, Карлсбад, Канада, США) и получили плазмиду рРВ235. Фрагмент PstI-SalI (1846 пар оснований) плазмиды рРВ235, содержащий ген НА, клонировали в экспрессирующем векторе pVR1012 (пример 1), рестриктированным PstI-SalI. В результате была получена плазмида pNS018 (6748 пар оснований). Ген НА дикого типа CDV кодирует белок из 607 аминокислот.
2.2.2. Получение плазмиды pNS024: клонировали ген НА (β-глобулиновая форма tPA ΔТМ НА) в плазмиде pVR1012
Укороченную форму гена НА CDV получали путем делеции фрагмента, кодирующего 60 первых аминокислот белка НА. Так как сигнальная и трансмембранная последовательности этого белка перекрываются, секреция укороченного белка обеспечивается включением сигнальной последовательностью тканевого активатора плазминогена человека (tPA). Плазмиду pNS019, полученную на основе pNS018, содержащую укороченный ген НА с сигнальным пептидом tPA, получали лигированием трех фрагментов ДНК:
- фрагмент А получали рестрикцией плазмиды рАВ110 BamHI-EcoRV (пример 1);
- фрагмент В получали с помощью полимеразной цепной реакции с использованием в качестве матрицы pNS018 (пример 2.2.1) и следующих олигонуклеотидов в качестве праймеров:
NS034 (30 нт) (последовательность №3)
5' TTTCGCCGGATCCCACAAAGTATCAACTAGC 3' и
NS035 (23 нт) (последовательность №4)
5' GGGATTTGCTGCCGATGCAATAG 3',
с последующей рестрикцией продукта полимеразнрой цепной реакции BamHI-SapI;
- фрагмент С получали рестрикцией плазмиды pNS018 Sapl-EcoRV.
Гибридный ген tPA ΔТМ НА кодирует белок из 574 аминокислот (1725 пар оснований).
Плазмиду pNS024 получали клонированием Интрона II гена β-глобина кролика (пример 2.1.2) перед геном НА по сайту SalI плазмиды pNS019 (фигура 4).
Пример 3
Плазмиды, кодирующие различные формы антигенов вируса парагриппа типа 2 (CPI-2) собак
Гены F и HN CPI-2 штамма D008 (MERIAL) получали с помощью полимеразной цепной реакции с обратной транскриптазой с использованием вирусной ДНК в качестве матрицы.
3.1. Плазмиды, кодирующие различные формы CPI-2 F
3.1.1. Получение плазмиды рАВ115: клонирование гена F (природная форма), в плазмиде pVR1012
кДНК гена F CPI-2 синтезировали и амплифицировали с помощью полимеразной цепной реакции с обратной транскриптазой с использованием следующей пары олигонуклеотидов в качестве праймеров:
SB 131 (38 нт) (последовательность №57)
5' AAAAACGCGTCGTCGACATGGGTACTATAATTCAATTTCTG 3' и
SB132 (38 нт) (последовательность №58)
5' TTTTCTAGTCTAGATTATTTATGATAAACAAAATTCTC 3'.
Продукт полимеразной цепной реакции рестриктировали SalI и XbaI, с получением фрагмента 1594 пар оснований, которую клонировали в экспрессирующем векторе pVR19012 (пример 1), рестриктированном теми же ферментами и получили плазмиду рАВ115 (6479 пар оснований). Ген F дикого типа CPI-2 (последовательность №72) (фигура 27) кодирует белок из 529 аминокислот.
3.1.2. Получение плазмиды pSB032: клонирование гена F (β-глобиновая форма F ΔTM), в плазмиде pVR1012
Плазмиду pSB031, содержащую укороченный ген F с делецией трансмембранного и цитоплазматического С-концевого доменов, получали с помощью полимеразной цепной реакции с использованием рАВ115 в качестве матрицы (пример 3.1.1.) и следующих олигонуклеотидов в качестве праймеров:
SB 131 (последовательность №57) и
SB 133 (41 нт) (последовательность №59)
5' TTTTCTAGTCTAGATTAGTATGTGTCACTTTGTGCTAAGTG 3',
в результате чего был получен фрагмент длиной 1450 пар оснований. Полученный фрагмент рестриктировали с помощью SalI и XbaI и выделяли фрагмент SalI-XbaI длиной 1436 пар оснований. Этот фрагмент лигировали с вектором pVR1012 (пример 1), предварительном рестриктированном SalI и XbaI, в результате чего получили плазмиду pSB031. Укороченный ген F CPI-2 кодирует белок из 473 аминокислот.
Затем перед последовательностью, кодирующей укороченный ген F CPI-2 по сайту SalI плазмиды pSB031, встраивали интрон II гена β-глобина кролика. Фрагмент ДНК, соответствующий интрону (573 пар оснований), получали с помощью полимеразной цепной реакции с использованием олигонуклеотидов NS036 (последовательность №5) и NS037 (последовательность №6) в качестве праймеров и плазмиды pNS050 (пример 1) в качестве матрицы с последующей рестрикцией SalI. Полученный фрагмент лигировали с плазмидой pSB031, предварительно рестриктированной SalI и дефосфорилированной. В результате была получена плазмида pSB032 (6884 пар оснований) (фигура 5).
3.2. Плазмиды, кодирующие различные формы CPI-2 HN
3.2.1. Получение плазмиды рАВ 114: клонирование гена HN (природная форма), в плазмиде pVR1012
кДНК гена HN CPI-2 синтезировали и амплифицировали с помощью ПЦР с использованием следующих олигонуклеотидов в качестве праймеров:
SB134 (41 нт) (последовательность №60)
5' AAAAACGCGTCGACATGGTTGCAGAAGATGCCCCTGTTAGG 3' и
SB135 (35 нт) (последовательность №61)
5' TTTTGGAAGATCTTTAGGATAGTGTCACCTGACGG 3'
в результате чего был получен фрагмент длиной около 1720 пар оснований. Этот фрагмент рестриктировали SalI и BglII и выделяли фрагмент SalI-BglII длиной 1704 пары оснований. Полученный фрагмент затем лигировали с вектором pVR1012 (пример 1), предварительно рестриктированным SalI и BglII. В результате была получена плазмида рАВ114 (6566 пар оснований). Ген HN дикого типа CPI-2 (последовательность №73) (фигура 28), клонированный в этой плазмиде, кодирует белок из 565 аминокислот.
3.2.2. Получение плазмиды pSB034: клонирование гена HN (β-глобиновая форма HN tPA ΔTM), в плазмиде pVR1012
Укороченную форму гена HN CPI-2 получали путем делеции фрагмента, кодирующего первые 40 аминокислот белка HN. Так как сигнальная и трансмембранная последовательности этого белка перекрываются, секреция укороченного продукта обеспечивается за счет встраивания сигнальной последовательности тканевого активатора плазминогена человека (tPA) перед укороченным геном HN. Плазмиду pSB033, полученную на основе рАВ114 (пример 3.2.1.), получали лигированием фрагмента EcoRI-PmII pAB110 (пример 1), полученной на основе pVR1012 и содержащей открытую рамку считывания, кодирующую сигнальную последовательность tPA, и фрагмента (1599 пар оснований), полученного с помощью полимеразной цепной реакции с использованием следующих олигонуклеотидов в качестве праймеров:
SB136 (37 нт) (последовательность №62)
5' TTAAAAGAATTCGACCCAAAAGCAAATCATGATGAGCCAC 3'
SB137 (33 нт) (последовательность №63)
5' TTAAAAGGCCTTTAGGATAGTGTCACCTGACGG 3',
и pAB114 в качестве матрицы, рестриктированного EcoRI и EcoRV.
Интрон II гена β-глобина кролика встраивали перед геном HN по сайту SalI pBS033, в результате чего была получена плазмида pSB034 (фигура 6). Фрагмент SalI, содержащий интрон, получали с помощью полимеразной цепной реакции с использованием олигонуклеотидов NS036 (последовательность №5) и NS037 (последовательность №6) в качестве праймеров и pNS050 (пример 1) в качестве матрицы.
Пример 4
Плазмиды, кодирующие различные формы гликопротеинов вируса CHV-1
Гены, кодирующие гликопротеины gB, gC и gD штамма Carmichael герпесвируса типа 1 собаки (CHV-1) получали с помощью полимеразной цепной реакции на основе вирусного генома. Клонирование генов, кодирующих gB и gD, в векторе pVR1012 было описано ранее в Международной заявке WO-A-9803199 (плазмиды рАВ037 и рАВ038, фигуры 7 и 8 и примеры 13 и 14 соответственно). Клонирование гена, кодирующего gC, описано в данной заявке.
4.1. Плазмида, кодирующая укороченную форму CHV-gB
4.1.1. Получение плазмиды pSB016: Клонирование гена gB (форма ΔТМ), в плазмиде pVR1012
Согласно профилю гидропатии трансмембранный домен гликопротеина gB CHV-1 (878 амиокислот) расположен между аминокислотными остатками 702 и 769. Плазмиду, содержащую укороченную форму гена, кодирующего gB, получали лигированием следующих трех фрагментов ДНК: (а) вектор pVR1012 (пример 1), расщепленный PstI-XbaI; (b) фрагмент длиной 1997 пар оснований, полученный рестрикцией PstI-NsiI pAB037 (пример 4) и (с) фрагмент длиной 225 пар оснований, полученный с помощью полимеразной цепной реакции с использованием следующих олигонуклеотидов в качестве праймеров:
SB101 (22 нт) (последовательность №15)
5' TATATTGAAGGACAACTTGGGG 3' и
SB102 (36 нт) (последовательность №16)
5' CTAGTCTAGATTAATTATTATCAACTTTTACAACAC 3',
и плазмиды pAB037 в качестве матрицы, рестриктированной NsiI и XbaI. Полученная в результате плазмида, pSB016 (6983 пары оснований) (фигура № 7), содержит укороченный ген gB, кодирующий белок из 701 аминокислот.
4.2. Плазмиды, кодирующие различные формы CHV-gC
4.2.1. Получение плазмиды рSВ018: клонирование гена gC (природная форма), в плазмиде pVR1012
Фрагмент ДНК, содержащий открытую рамку считывания гена gC CHV-1, получали с помощью полимеразной цепной реакции с использованием следующих олигонуклеотидов в качестве праймеров:
SB105 (32 нт) (последовательность №19)
5' AAAACTGCAGATGAGTTTTAAAAATTTTTATC 3' и
SB106 (30 нт) (последовательность №20)
5' CTAGTCTAGATTAGATCTTATTATTTTTG 3',
и вирусной ДНК в качестве матрицы. Продукт полимеразной цепной реакции рестриктировали PstI и XbaI и полученный фрагмент длиной 1400 пар оснований затем лигировали с вектором pVR1012 (пример 1), расщепленный теми же рестриктазами. Полученная в результате плазмида, pSB018 (6253 пар оснований), содержит ген, кодирующий гликопротеин gC из 459 аминокислот.
4.2.2. Получение плазмиды pSB019: клонирование гена gC (форма Δ ТМ), в плазмиде pVR1012
Согласно профилю гидропатии трансмембранный домен гликопротеина gC расположен между аминокислотными остатками 422 и 452. Плазмиду, содержащую укороченную форму гена, кодирующего gC, получали лигированием следующих трех фрагментов ДНК: (а) вектор pVR1012 (пример 1), линеаризированный путем двойной рестрикции PstI-XbaI; (b) фрагмент длиной 934 пары оснований, полученный рестрикцией PstI-StuI pSB018 (предыдущий пример), и (с) фрагмент длиной 335 пар оснований, полученный с помощью полимеразной цепной реакции с использованием следующих олигонуклеотидов в качестве праймеров:
SB107 (24 нт) (последовательность №21)
5' TGGATTGACGGTCTTATAACACG 3' и
SB108 (37 нт) (последовательность №22)
5' CTAGTCTAGATTAATTTTCATCCGATGCATCAAACAC 3',
и плазмиды pSB018 в качестве матрицы, рестриктированной Stul и XbaI. Полученная в результате плазмида, pSB019 (6139 пар оснований) (фигура 8), содержит укороченный ген gC, кодирующий белок из 421 аминокислоты.
4.3. Плазмида, кодирующая укороченную форму CHV-gD
4.3.1. Получение плазмиды pSB017: клонирование гена gD (форма ΔТМ), в плазмиде pVR1012
Трансмембранный домен гликопротеина gD CHV-1 (345 аминокислот) расположен между аминокислотными остатками 310 и 328. Плазмиду, содержащую укороченную форму гена, кодирующего gD, получали дотированием следующих трех фрагментов ДНК: (а) вектор pVR1012 (пример 1), расщепленный PstI-NotI; (b) фрагмент длиной 663 пары оснований, полученный рестрикцией PstI-Avall pAB038 (пример 4), и (с) фрагмент длиной 415 пар оснований, полученный с помощью полимеразной цепной реакции с использованием следующих олигонуклеотидов в качестве праймеров:
SB103 (25 нт) (последовательность №17)
5' CGAGAAACTTGTTATTTTTCTAAAG 3' и
SB104 (51 нт) (последовательность №18)
5' ATAAGAATGCGGCCGCAAAGGCTATATATTTTTGGGGTATTATTTATTGG 3',
и плазмиды pAB038 в качестве матрицы, рестриктированной Avall и NotI. Полученная в результате плазмида, pSB017 (5819 пар оснований) (фигура 9), содержит укороченный ген gD, кодирующий белок из 309 аминокислот.
Пример 5
Плазмиды, кодирующие различные формы гликопротеинов FHV-1
Гены, кодирующие гликопротеины gB, gC и gD штамма СО герпесвируса типа 1 кошки (FHV-1), получали с помощью полимеразной цепной реакции из вирусного генома. В случае гена, кодирующего gD нуклеотидная последовательность идентична таковой у штамма С-27. Заявители использовали плазмиду рАВ029, полученную на основе VR1012, содержащую соответствующий клонированный ген из штамма С-27 и описанную в Международной заявке WO-A-9803660 (плазмида рАВ029, фигура 12 и пример 15).
5.1. Плазмиды, кодирующие различные формы FHV-gB
5.1.1. Получение плазмиды pSB020: клонирование гена gB (природная форма), в плазмиде рVR1012
Фрагмент ДНК, содержащий открытую рамку считывания гена gB FHV-1, получали с помощью полимеразной цепной реакции с использованием следующих олигонуклеотидов в качестве праймеров:
SB113 (34 нт) (последовательность №27)
5' TTTTCTGCAGATGTCCACTCGTGGGGATCTTGGG 3' и
SB114 (40 нт) (последовательность №28)
5' ATAGTTTAGCGGCCGCTTAGACAAGATTTGTTTCAGTATC 3',
и вирусной ДНК в качестве матрицы. Продукт полимеразной цепной реакции рестриктировали PstI и NotI и полученный фрагмент длиной 2849 пар оснований затем лигировали с вектором pVR1012 (пример 1), расщепленной теми же рестриктазами. Полученная в результате плазмида, pSB020 (7728 пар оснований), содержит ген, кодирующий гликопротеин gB из 949 аминокислот.
5.1.2. Получение плазмиды pSB021: клонирование гена gB (форма ΔТМ), в плазмиде pVR1012
Согласно профилю гидропатии трансмембранный домен гликопротеина gB FHV-1 расположен между аминокислотными остатками 761 и 834. Плазмиду, содержащую укороченную форму гена, кодирующего gB, получали лигированием следующих трех фрагментов ДНК: (а) вектор pVR.1012 (пример 1), линеаризированный рестрикцей PstI-NotI; (b) фрагмент длиной 1839 пар оснований, полученный рестрикцией PstI-HindII pSB020 (предыдущий пример), и (с) фрагмент длиной 447 пар оснований, полученный с помощью полимеразной цепной реакции с использованием следующих олигонуклеотидов в качестве праймеров:
SB109 (24 нт) (последовательность №23)
5' CTGTGGACAGAGACCCTAAAACTC 3' и
SB110 (50 нт) (последовательность №24)
5' TTTCCTTTTGCGGCCGCTTATATGCTGTCTATATCATAAAATTTTAAGGC 3',
и плазмиду pSB020 в качестве матрицы, рестриктированную HindII и NotI. Полученная в результате плазмида, pSB021 (7164 пара оснований) (фигура 10), содержит укороченный ген gB, кодирующий белок из 760 аминокислот.
5.2. Плазмиды, кодирующие различные формы FHV-gC
5.2.1. Получение плазмиды pSB022: клонирование гена gC (природная форма), в плазмиде pVR1012
Фрагмент ДНК, содержащий открытую рамку считывания гена gC FHV-1, получали с помощью полимеразной цепной реакции с использованием следующих олигонуклеотидов в качестве праймеров:
SB115 (34 нт) (последовательность №29)
5' TTTTCTGCAGATGAGACGATATAGGATGGGACGC 3' и
SB116 (34 нт) (последовательность №30)
5' AGTTTAGCGGCCGCTTATAATCGCCGGGGATGAG 3',
и вирусную ДНК в качестве матрицы. Продукт полимеразной цепной реакции рестриктировали PstI и NotI, полученный фрагмент длиной 1605 пар оснований затем лигировали с вектором pVR1012 (пример 1), расщепленным теми же рестриктазами. Полученная в результате плазмида, pSB022 (6253 пар оснований), содержит ген, кодирующий гликопротеин gC из 534 аминокислот.
5.2.2. Получение плазмиды рSВ023: клонирование гена gC (форма ΔТМ), в плазмиде PVR1012
Согласно профилю гидропатии, трансмембранный домен гликопротеина gC FHV-1 расположен между аминокислотными остатками 495 и 526. Плазмиду, содержащую укороченную форму гена, кодирующего gC, получали лигированием следующих двух фрагментов ДНК: (а) фрагмент длиной 6198 пар оснований, полученный рестрикцией BclI-NotI pSB022 (предыдущий пример), и (b) фрагмент длиной 168 пар оснований, полученный с помощью полимеразной цепной реакции с использованием следующих олигонуклеотидов в качестве праймеров:
SB117 (24 нт) (последовательность №31)
5' GTTAAATGTGTACCACGGGACGGG 3' и
SB118 (41 нт) (последовательность №32)
5' AGTTTAGCGGCCGCTTATTCAGGGGACGCGTCGTAGACTTG 3',
и плазмиду pSB022 в качестве матрицы, рестриктированный BclI и NotI. Полученная в результате плазмида, pSB023 (6366 пар оснований) (фигура 11), содержит укороченный ген gC, кодирующий белок из 494 аминокислот.
5.3. Плазмида, кодирующая укороченную форму FHV-gD
5.3.1. Получение плазмиды pSB024: клонирование гена gD (форма ΔТМ), в плазмиде pVR1012
Трансмембранный домен гликопротеина gD FHV-1 (374 аминокислоты) расположен между аминокислотными остатками 328 и 353. Плазмиду, содержащую укороченную форму гена, кодирующего gD, получали лигированием следующих двух фрагментов ДНК: (а) фрагмент длиной 5712 пар оснований, полученного рестрикцией XbaI-BglII pAB029 (пример 5), и (b) фрагмент длиной 129 пар оснований, полученный с помощью полимеразной цепной реакции с использованием следующих олигонуклеотидов в качестве праймеров:
SB111 (24 нт) (последовательность №25)
5' GATCGTCCCGCCATACCGTCTGGG 3' и
SB112 (39 нт) (последовательность №26)
5' TTTGGAAGATCTTTACTGATTATTCATGCCCTTGGGAGG 3',
и плазмиду рАВ038 в качестве матрицы, рестриктированный XbaI и BglII. Полученная в результате плазмида, pSB024 (5841 пара оснований) (фигура 12), содержит укороченный ген gD, кодирующий белок из 327 аминокислот.
Пример 6
Плазмиды, кодирующие различные формы гликопротеинов EHV-1
Гены, кодирующие гликопротеины gB, gC и gD штамма 2234/88-2 EHV-1, получали с помощью полимеразной цепной реакции из очищенной вирусной ДНК.
6.1. Плазмиды, кодирующие различные формы EHV-1 gB
6.1.1. Получение плазмиды рАВ127: клонирование гена gB (природная форма), в плазмиде pVR1012
Кодирующую область гена gB EHV-1 амплифицировали с помощью полимеразной цепной реакции с использованием следующих олигонуклеотидов в качестве праймеров:
NS003 (30 нт) (последовательность №39)
5' TTCTGCAGATGTCCTCTGGTTGCCGTTCGT 3' и
NS004 (30 нт) (последовательность №40)
5' TTTCTAGATTAAACCATTTTTTCATTTTCC 3',
полученный фрагмент ДНК рестриктировали PstI и XbatI и лигировали с вектором pVR1012 (пример 1), расщепленным PstI и XbatI, в результате чего была получена плазмида рАВ127 (7818 пар оснований). Ген gB кодирует белок из 980 аминокислот.
6.1.2. Получение плазмиды рSB028: клонирование гена gB (форма ΔТМ), в плазмиде pVR1012
Согласно профилю гидропатии трансмембранный домен гликопротеина gB EHV-1 расположен между аминокислотными остатками 801 и 875. Плазмиду, содержащую укороченную форму гена, кодирующего gB, получали лигированием следующих двух фрагментов ДНК: (а) плазмида рАВ127 (пример 6.1.1), рестриктированная AfeI и XbaI, и (b) фрагмент длиной 276 пар оснований, полученный с помощью полимеразной цепной реакции с использованием следующих олигонуклеотидов в качестве праймеров:
SB125 (24 нт) (последовательность №51)
5' AACAACAGAGGGTCGATAGAAGGC 3' и
SB126 (39 нт) (последовательность №52)
5' AATTTTTCTAGATTACACGTTGACCACGCTGTCGATGTC 3',
и плазмиду рАВ127 в качестве матрицы, рестриктированную AfeI и XbaI. Полученная в результате плазмида, pSB028 (7279 пар оснований), содержит укороченный ген gB EHV-1, кодирующий белок из 800 аминокислот.
6.2. Плазмиды, кодирующие различные формы EHV-1 gC
6.2.1. Получение плазмиды рАВ129: клонирование гена gC (природная форма), в плазмиде pVR1012
Фрагмент ДНК, содержащий открытую рамку считывания гена gC EHV-1, получали с помощью полимеразной цепной реакции с использованием следующих олигонуклеотидов в качестве праймеров:
NS005 (31 нт) (последовательность №41)
5' TTGTCGACATGTGGTTGCCTAATCTCGTGAG 3' и
NS006 (33 нт) (последовательность №42)
5' TTGGATCCCTAAAAGTCAGACTTCTTGTACGGC 3'.
Продукт полимеразной цепной реакции рестриктировали с помощью SalI и BamHI, полученный фрагмент длиной 1412 пар оснований затем лигировали с вектором pVR1012 (пример 1), расщепленным теми же рестриктазами. Полученная в результате плазмида, рАВ129 (6281 пара оснований), содержит ген, кодирующий гликопротеин gC EHV-1, состоящий из 468 аминокислот.
6.2.2. Получение плазмиды рSB029: клонирование гена gC (форма ΔТМ), в плазмиде pVR1012
Согласно профилю гидропатии трансмембранный домен гликопротеина gC EHV-1 расположен между аминокислотными остатками 429 и 455. Плазмиду, содержащую укороченную форму гена, кодирующего gC, получали лигированием следующих двух фрагментов ДНК: (а) плазмида рАВ129 (пример 6.2.1.), расщепленная AspI-BamHI, и (b) фрагмент длиной 287 пар оснований, полученный с помощью полимеразной цепной реакции с использованием следующих олигонуклеотидов в качестве праймеров:
SB127 (24 нт) (последовательность №53)
5' GATCCGGAGGAGGAATACACACCC 3' и
SB128 (39 нт) (последовательность №54)
5' AATTTTGGATCCCTAAACCGGCCTGTCCTCAACAATCGG 3',
и плазмиду рАВ129 в качестве матрицы, рестриктированную AspI и BamHI. Полученная в результате плазмида, pSB029 (6161 пара оснований), содержит укороченный ген gC, кодирующий белок из 428 аминокислот.
6.3. Плазмиды, кодирующие укороченную форму EHV-1 gD
6.3.1. Получение плазмиды рАВ131: клонирование гена gD (природная форма), в плазмиде pVR1012
Фрагмент ДНК, содержащий открытую рамку считывания гена gD EHV-1, получали с помощью полимеразной цепной реакции с помощью следующих олигонуклеотидов:
NS007 (33 нт) (последовательность №43)
5' TTGTCGACATGTCTACCTTCAAGCTTATGATGG 3' и
NS008 (32 нт) (последовательность №44)
5' TTGGATCCTTACGGAAGCTGGGTATATTTAAC 3'.
Продукт полимеразной цепной реакции рестриктировали SalI и BamHI, полученный фрагмент длиной 1214 пар оснований затем лигировали с вектором pVR1012 (пример 1), расщепленной теми же рестриктазами. Полученная в результате плазмида, рАВ131 (6083 пары оснований), содержит ген, кодирующий гликопротеин gD из 402 аминокислот.
6.3.2. Получение плазмиды рSВ030: клонирование гена gD (форма ΔТМ). в плазмиде pVR1012
Согласно профилю гидропатии трансмембранный домен гликопротеина gD EHV-1 расположен между аминокислотными остатками 348 и 371. Плазмиду, содержащую укороченную форму гена, кодирующего gD, получали лигированием следующих трех фрагментов ДНК: (а) плазмида pVR1012 (пример 1), расщепленная SalI-BamHI; (b) фрагмент длиной 825 пар оснований, расщепленный рАВ131 (пример 6.3.1.), SalI и BsmI и (с) фрагмент длиной 239 пар оснований, полученный с помощью полимеразной цепной реакции с использованием следующих олигонуклеотидов в качестве праймеров:
SB129 (24 нт) (последовательность №55)
5'CGGTTTCTTGGTGAATTCAACTTC 3' и
SB130 (42 нт) (последовательность №56)
5' AATTTTGGATCCTTACGTAGAGTTGCTCTTAGACGTTTTTGG 3',
и плазмиду рАВ131 в качестве матрицы, рестриктированную BsmI и BamHI. Полученная в результате плазмида, pSB030 (5921 пара оснований), содержит укороченный ген gD, кодирующий белок из 347 аминокислот.
Пример 7
Плазмиды, кодирующие различные формы гликопротеинов EHV-4
Гены, кодирующие гликопротеины gB, gC и gD штамма KYT 445/2 EHV-4, получали с помощью полимеразной цепной реакции из очищенной вирусной ДНК.
7.1. Плазмиды, кодирующие различные формы EHV-4 gB
7.1.1. Получение плазмиды рАВ136: клонирование гена gB (природная форма), в плазмиде pVR1012
Кодирующую область гена gB EHV-4 амплифицировали с помощью полимеразной цепной реакции с использованием следующих олигонуклеотидов в качестве праймеров:
АВ325 (35 нт) (последовательность №33)
5' TTTCTGCAGATGTCCACTTGTTGCCGTGCTATTTG 3' и
АВ326 (31 нт) (последовательность №34)
5' TTTTCTAGATTAAACCATTTTTTCGCTTTCC 3',
полученный фрагмент ДНК рестриктировали с помощью PstI и XbatI и лигировали с вектором pVR1012 (пример 1), расщепленным PstI и XbatI. Полученная в результате плазмида рАВ136 (7801 пара оснований) содержит ген gB EHV-4, кодирующий белок из 975 аминокислот.
7.1.2. Получение плазмиды pSB025: клонирование гена gB (форма ΔТМ), в плазмиде pVR1012
Согласно профилю гидропатии трансмембранный домен гликопротеина gB EHV-4 расположен между аминокислотными остатками 797 и 867. Плазмиду, содержащую укороченную форму гена, кодирующего gB, получали лигированием следующих двух фрагментов ДНК: (а) плазмида рАВ136 (пример 7.1.1), рестриктированная SplI и XbaI, и (b) фрагмент длиной 231 пары оснований, полученный с помощью полимеразной цепной реакции с использованием следующих олигонуклеотидов в качестве праймеров:
SB119 (36 нт) (последовательность №45)
5' TTTTGGTCTAGATTAGTCCACGTTGACAACGCTGTC 3' и
SB120 (23 нт) (последовательность №46)
5' CGCAAGCTTATCGAGCCGTGCGC 3',
и плазмиды рАВ136 в качестве матрицы, рестриктированной с помощью SplI и Xbal. Полученная в результате плазмида, pSB025 (7264 пары оснований), содержит укороченный ген gB, кодирующий белок из 796 аминокислот.
7.2. Плазмиды, кодирующие различные формы EHV-4 gC
7.2.1. Получение плазмиды рАВ137: клонирование гена gC (природная форма), в плазмиде pVR1012
Фрагмент ДНК, содержащий открытую рамку считывания гена gC EHV-4, получали с помощью полимеразной цепной реакции с использованием следующих олигонуклеотидов в качестве праймеров:
АВ327 (32 нт) (последовательность №35)
5' TTTGTCGACATGGGTTTGGTAAATATAATGCG 3' и
АВ328 (33 нт) (последовательность №36)
5' TTTGGATCCTTAGAAGTCTGCTTTCTTGTAGGG 3'.
Продукт полимеразной цепной реакции рестриктировали SalI и BamHI, полученный фрагмент длиной 1463 пары оснований затем лигировали с вектором pVR1012 (пример 1), расщепленным теми же рестриктазами. Полученная в результате плазмида, рАВ137 (6330 пар оснований), содержит ген, кодирующий гликопротеин gC из 485 аминокислот.
7.2.2. Получение плазмиды pSB026: клонирование гена gC (форма ΔТМ), в плазмиде pVR1012
Согласно профилю гидропатии трансмембранный домен гликопротеина gC EHV-4 расположен между аминокислотными остатками 425 и 472. Плазмиду, содержащую укороченную форму гена, кодирующего gC, получали лигированием следующих двух фрагментов ДНК: (а) плазмида рАВ137 (пример 7.2.1.), расщепленная AclI-BamHI, и (b) фрагмент длиной 237 пар оснований, полученный с помощью полимеразной цепной реакции с использованием следующих олигонуклеотидов в качестве праймеров:
SB121 (24 нт) (последовательность №47)
5' GTATCAATCCCAGCTGACCCCAC 3' и
SB122 (41 нт) (последовательность №48)
5' AATTTTGGATCCTTAGCCGTCCGGGTAACCCTCTATGATGC 3',
и плазмиды рАВ137 в качестве матрицы, рестриктированной с помощью AclI и BamHI. Полученная в результате плазмида, pSB026 (6147 пар оснований), содержит укороченный ген gC, кодирующий белок из 424 аминокислот.
7.3. Плазмиды, кодирующие укороченную форму EHV-4 gD
7.3.1. Получение плазмиды рАВ138: клонирование гена gD (природная форма), в плазмиде pVR1012
Фрагмент ДНК, содержащий открытую рамку считывания гена gD EHV-4, получали с помощью полимеразной цепной реакции с помощью следующих олигонуклеотидов в качестве праймеров:
АВ329 (33 нт) (последовательность №37)
5' TTTGTCGACATGTCTACCTTCAAGCCTATGATG 3' и
АВ330 (33 нт) (последовательность №38)
5' TTTGGATCCTTACGGAAGCTGAGTATATTTGAC 3'.
Продукт полимеразной цепной реакции рестриктировали SalI и BamHI, полученный фрагмент длиной 1214 пар оснований затем лигировали с вектором pVR1012 (пример 1), расщепленным теми же рестриктазами. Полученная в результате плазмида, рАВ138 (6081 пара оснований), содержит ген, кодирующий гликопротеин gD из 402 аминокислот.
7.3.2. Получение плазмиды рSВ027: клонирование гена gD (форма ΔТМ), в плазмиде pVR1012
Согласно профилю гидропатии трансмембранный домен гликопротеина gD EHV-4 расположен между аминокислотными остатками 348 и 371. Плазмиду, содержащую укороченную форму гена, кодирующего gD, получали лигированием следующих двух фрагментов ДНК: (а) плазмида рАВ138 (пример 7.3.1.), расщепленная EcoRI-BamHI, и (b) фрагмент длиной 310 пар оснований, полученный с помощью полимеразной цепной реакции с использованием следующих олигонуклеотидов в качестве праймеров:
SB123 (24 нт) (последовательность №49)
5' TTTTCCGTAACAATTCCGAGCAGC 3' и
SB124 (39 нт) (последовательность №50)
5' AATTTTGGATCCTTACGTAGAGTTGCTATTAGACGCTGG 3',
и плазмиды рАВ138 в качестве матрицы, рестриктированной EcoRI и BamHI. Полученная в результате плазмида, pSB027 (5919 пар оснований), содержит укороченный ген gD, кодирующий белок из 347 аминокислот.
Пример 8
Получение плазмиды, содержащей GM-CSF собаки
8.1. Выделение тотальной РНК из лимфоцитов собаки, стимулированных in vitro митогенами
Кровь собаки (сука породы Бигль) собирали в пробирку, содержащую этилендиамин-тетрауксусную кислоту (EDTA). Мононуклеарные клетки выделяли путем центрифугирования в градиенте плотности Ficoll и культивировали в чашке Петри диаметром 60 мм. Затем мононуклеарные клетки стимулировали конканавалином А (ConA) (конечная концентрация примерно 4 мкг/мл) и фитогемагглютинином (РНА) (конечная концентрация примерно 10 мкг/мл). После стимуляции лимфобласты ″ConA″ и ″РНА″ соскабливали с культуральных чашек и тотальную РНК из стимулированных клеток экстрагировали, используя набор ″набор для выделения мРНК из лейкоцитов″ (Boehringer Mannheim / Roche Cat # 1934325).
8.2. Выделение гена. кодирующего GM-CSF собаки, и конструкция плазмиды pJP074
Тотальную РНК, выделенную из лимфобластов собаки, стимулированных ConA или РНА (пример 8.1.), использовали в качестве матрицы для синтеза первой нити комплементарной ДНК. В качестве праймера для синтеза первой нити комплементарной ДНК использовали олигонуклеотид p(dT)15 (Boehringer Mannheim / Roche Cat # 814270). Полученную однонитевую комплементарную ДНК использовали в качестве матрицы для ПЦР со следующими олигонуклеотидами в качестве праймеров:
JP578 (33 нт) (последовательность №64)
5' TATGCGGCCGCCACCATGTGGCTGCAGAACCTG 3' и
JP579 (36 нт) (последовательность №65)
5' TATGCGGCCGCTACGTATCACTTCTTGACTGGTTTC 3'
для амплификации фрагмента ПЦР длиной примерно 450 пар оснований. Этот фрагмент очищали электрофорезом в агарозном геле, а затем лигировали с вектором pCR2.1 (In Vitrogen, Карлсбад, Канада, США) для получения плазмиды pJP074. Последовательность гена GM-CSF собаки, клонированного в плазмиде pJP074, оказалась эквивалентной последовательности гена GM-CSF собаки из банка генов (номер доступа # S49738).
8.3. Конструкция плазмиды pJP084 и последовательность гена GM-CSF собаки
Плазмиду pJP074 (пример 8.2) рестриктировали NotI и с помощью электрофореза в агарозном геле выделяли фрагмент NotI-NotI длиной примерно 450 пар оснований, содержащий ген GM-CSP собаки. Этот фрагмент лигировали с плазмидой pVR1012 (пример 1). Клон, содержащий последовательность GM-CSP собаки (идентифицированая последовательность №66, фигура 20) в правильной ориентации по отношению к промотору hCMV/IE, назвали pJP084. Эта плазмида имеет длину 5364 пары оснований (фигура 19).
Пример 9
Получение плазмиды, кодирующей GM-CSF кошки
9.1. Выделение тотальной РНК из лимфоцитов кошки, стимулированных w vitro митогенами
Кровь кошки собирали в пробирку, содержащую этилендиаминтетрауксусную кислоту (EDTA). Мононуклеарные клетки вьщеляли центрифугированием в градиенте плотности Ficoll и культивировали в чашке Петри диаметром 60 мм. Затем мононуклеарные клетки кошки стимулировали конканавалином А (ConA) (конечная концентрация примерно 4 мкг/мл) и фитогемагглютинином (РНА) (конечная концентрация примерно 10 мкг/мл). После стимуляции лимфобласты ″ConA″ и ″РНА″ соскабливали с культуральных чашек и тотальную РНК из этих клеток экстрагировали, используя набор ″набор для выделения мРНК из лейкоцитов″ (Boehringer Mannheim / Roche Cat # 1934325).
9.2. Выделение гена, кодирующего GM-CSF кошки, и конструкция плазмид pJP089 и pJP090
Тотальную РНК, выделенную из лимфобластов кошки, стимулированных ConA и РНА (пример 9.1.), использовали в качестве матрицы для слитера первой нити комплементарной ДНК. В качестве праймера для слитера первой нити комплементарной ДНК использовали олигонуклеотид p(dT)15 (Boehringer Mannheim / Roche Cat # 814270). Полученную однонитевую комплементарную ДНК затем использовали в качестве матрицы для ПЦР со следующими олигонуклеотидами в качестве праймеров:
JP578 (33 нт) (последовательность №64)
5' TATGCGGCCGCCACCATGTGGCTGCAGAACCTG 3' и
JP579 (36 нт) (последовательность №65)
5' TATGCGGCCGCTACGTATCACTTCTTGACTGGTTTC 3'
для амплификации фрагмента ПЦР длиной примерно 450 пар оснований. Этот фрагмент рестриктировали NotI и с помощью электрофореза в агарозном геле выделяли фрагмент NotI-NotI длиной 450 пар оснований. Этот фрагмент затем лигировали с плазмидой pVR1012 (пример 1). Два клона, содержащие последовательность гена GM-CSP кошки (идентифицированная последовательность №67 и последовательность №68) в правильной ориентации по отношению к промотору hCMV/IE, назвали pJP089 и pJP090 соответственно. Длина этих плазмид составляет 5364 пары оснований (фигуры №21 и №23).
Последовательность гена GM-CSF кошки, клонированного в плазмиде pJP089, содержит 13 различий на нуклеотидном уровне по сравнению с последовательностью гена GM-CSF кошки, находящейся в банке генов (номер AF053007). Самой существенной заменой является замена С→Т, которая приводит к замене лейцин → фенилаланин (первый нуклеотид кодона аминокислоты 107; фигура 22). Последовательность гена GM-CSF кошки, клонированного в плазмиде pJP090, соответствует последовательности, содержащейся в плазмиде pJP089, однако не содержит замены лейцин → фенилаланин для аминокислоты 107 (фигура 24). Проверка 3' последовательности гена GM-CSF кошки с помощью набора 3' RACE показала, что в положении 107 одной и той же кошки может находиться как лейцин, так и фенилаланин.
Пример 10
Получение плазмиды, кодирующей GM-CSF лошади
10.1. Выделение тотальной РНК из лимфоцитов лошади, стимулированных in vitro митогенами
Кровь лошади, взятую из яремной вены, собирали в пробирку, содержащую этилендиамин-тетрауксусную кислоту (EDTA). Мононуклеарные клетки выделяли центрифугированием в градиенте плотности Ficoll и культивировали в чашке Петри диаметром 60 мм. Затем мононуклеарные клетки лошади стимулировали конканавалином А (ConA) (конечная концентрация примерно 5 мкг/мл), либо фитогемагглютинином (РНА) (конечная концентрация примерно 10 мкг/мл). После стимуляции лимфобласты ″ConA″ и ″РНА″ соскабливали с культуральных чашек и тотальную РНК из клеток выделяли, используя набор ″набор для выделения мРНК из лейкоцитов″ (Boehringer Mannheim / Roche Cat # 1934325).
10.2. Выделение гена. кодирующего GM-CSF лошади
Тотальную РНК, выделенную из лимфобластов лошади, стимулированных ConA или РНА (пример 10.1.), использовали в качестве матрицы для первой нити комплементарной ДНК. В качестве праймера для синтеза первой нити комплементарной ДНК использовали олигонуклеотид p(dT)15 (Boehringer Mannheim / Roche Cat # 814270). Полученную однонитевую комплементарную ДНК затем использовали в качестве матрицы для ПЦР со следующими олигонуклеотидами в качестве праймеров:
JP734 (44 нт) (последовательность №70)
5' CATCATCATGTCGACGCCACCATGTGGCTGCAGAACCTGCTTCT 3' и
JP735 (41 нт) (последовательность №71)
5' CATCATCATGCGGCCGCTACTTCTGGGCTGCTGGCTTCCAG 3'
для амплификации фрагмента ПЦР длиной примерно 500 пар оснований. Этот фрагмент очищали с помощью электрофореза в агарозном геле.
10.3. Конструкция плазмиды pJP097, содержащей последовательность гена GM-CSF лошади
Очищенный фрагмент ПЦР, полученный в примере 10.2, рестриктировали NotI и с помощью электрофореза в агарозном геле выделяли фрагмент NotI-NotI длиной примерно 450 пар оснований, содержащий ген GM-CSF лошади. Этот фрагмент лигировали с плазмидой pVR1012 (пример 1). Клон, содержащий последовательность GM-CSF лошади (последовательность №69, фигура 26) в правильной ориентации по отношению к промотору hCMV/IE называли pJP097. Эта плазмида имеет длину 5334 пары оснований (фигура 25).
Пример 11
Сводная таблица плазмид
** для генов НА CDV и HN CPI-2: трансмембранный домен и сигнальная последовательность перекрываются
Пример 12
Методы молекулярной биологии
Культивирование и выделение вирусов
Вирусы культивировали в соответствующих клеточных системах вплоть до достижения цитопатического эффекта. Используемые для каждого вируса клеточные системы хорошо известны специалисту. Вкратце, соответствующие клетки инфицировали нужным вирусным штаммом с множественностью инфекции 1 и инкубировали при температуре 37°С в течение времени, необходимого для достижения цитопатического эффекта (в среднем 36 часов).
В случае ДНК-содержащих вирусов супернатант и лизированные клетки после культивирования собирали и клеточные остатки удаляли центрифугированием при 1000 g и при температуре 4°С в течение 10 минут. Вирусные частицы осаждали ультрацентрифугированием при 400000 g и при температуре 4°С в течение 1 часа. Осадки после центрифугирования суспендировали в минимальном объеме буфера ТЕ (1 ммоль Трис, 1 ммоль этилендиаминтетрауксусной кислоты (EDTA)).
РНК-содержащие вирусы выделяли согласно стандартным методикам, хорошо известным специалисту.
Выделение вирусной геномной ДНК
Концентрированные вирусные суспензии обрабатывали протеиназой К (конечная концентрация 100 мг/мл) в присутствии додецилсульфата натрия (SDS) (конечная концентрация 0,5%) в течение 2 часов при температуре 37°С. Вирусную ДНК экстрагировали смесью фенол/хлороформ, а затем осаждали двумя объемами абсолютного этанола при температуре -20°С в течение 16 часов и центрифугировали при 10000 g в течение 15 минут при температуре 4°С. Осадки ДНК высушивали и растворяли в минимальном объеме сверхчистой стерильной воды.
Выделение вирусной геномной РНК
Геномную РНК вирусов выделяли ″гуанидинийтиоцианат/фенол-хлороформ″ методом, описанным Р.Chomczynski и N.Sacchi (Anal. Biochem., 162, 156-159 (1987)).
Методы молекулярной биологии
Все плазмиды получали с помощью стандартных методов молекулярной биологии, описанных Sambrook и др. (″Molecular Cloning: A Laboratory Manual″, второе издание. Cold Spring Harbor Laboratory, Cold Spring Harbor, Нью-Йорк, 1989). Все фрагменты рестрикции, используемые согласно настоящему изобретению, получали с помощью набора ″Geneclean″ (BIO101 Inc., La Jolla, Канада). Для всех конструкций клонированные фрагменты ДНК, а также участки соединения с экспрессирующим вектором секвенировали по методу Sanger (Sambrook и др., 1989).
Полимеразная цепная реакция и полимеразная цепная реакция с обратной транскриптазой (PCR и RT-PCR)
Для клонирования необходимых генов или их фрагментов были синтезированы специфические олигонуклеотиды, некоторые из которых включали на 5'-конце сайты рестрикции, облегчающие клонирование амплифицированных фрагментов. Реакции обратной транскрипции (RT) и амплификация с помощью полимеразы (полимеразная цепная реакция, PCR) были осуществлены согласно стандартным методиками (Sambrook и др., 1989).
Получение больших количеств плазмид
Плазмиды в больших количествах (десятки мг), входящие в вакцинные композиции, выделяли в градиенте плотности хлорида цезия с добавлением этидиумбромида (Sambrook и др., 1989).
Пример 13
Приготовление вакцинных композиций плазмид
Раствор ДНК, содержащий одну или несколько плазмид, полученных согласно примерам 2-10, осаждали этанолом, как описано у Sambrook и др. (1989). Осадок ДНК после центрифугирования растворяли 0,9%-ным раствором NaCl до концентрации 1 мг/мл. Модифицированный DMRIE-DOPE растворяли в стерильной воде до получения концентрации 0,75 мМ.
Для получения комплексов плазмидная ДНК-липид смешивали равные объемы 0,75 мМ раствора DMRIE-DOPE и раствора ДНК с концентрацией 1 мг/мл в 0,9%-ном растворе NaCl. Для того чтобы избежать образования пены, раствор ДНК добавляли постепенно с помощью иглы 26G по внутренней стенки флакона, содержащего раствор катионного липида. Полученную смесь медленно перемешивали. В результате получают композицию, включающую 0,375 мМ DMRIE-DOPE и 500 мкг/мл плазмиды.
Все вышеописанные операции проводят при комнатной температуре. Комплексы ДНК/DMRIE-DOPE получают при комнатной температуре в течение 30 минут непосредственно перед проведением иммунизации животных.
Пример 14
Иммунизация собак против CDV
Иммунизацию проводили путем инъекции 2 мл вакцины подкожно или внутримышечно. Иммунизацию повторяли еще раз через 28 дней. Плазмиду при каждой иммунизации вводили в количестве 100, 500, 1000 или 2000 мкг в зависимости от вакцин. Специалист обладает знаниями, необходимыми для установления объема или концентрации в зависимости от требуемой дозы плазмиды.
14.1. Вирулентный тест
Для проведения вирулентного теста использовали штамм в виде измельченного продукта из селезенки, извлеченной у собаки, инфицированной вирусом болезни Карре, штамм Snyder Hill, стократно разведенного PBS. Разведение хранили во льду вплоть до использования.
Стократно разведенный штамм вводили после общей анестезии интракраниально в объеме 0,5 мл спустя 49 дней после первой инъекции.
14.2. Клиническое наблюдение после проведенного теста
Ежедневное клиническое наблюдение осуществляли в течение 21 дня в зависимости от теста. Для каждой собаки проводили клиническое исследование для детектирования возможных клинических симптомов чумки собак и определение ректальной температуры.
Клиническое исследование включает:
- общее состояние животного оценивали по четырехбалльной шкале: "хорошее" - оценка 0; "апатия" - оценка 1; "депрессия" - оценка 2 и "прострация" - оценка 3. Гибель животного соответствовала клинической оценке 10;
- оценка глазоносовых симптомов (установление серозного выделения или гноетечения, ринита и/или конъюнктивита). Конъюнктивит/ринит с серозным выделением из носа эквивалентно оценке 1, конъюнктивит/ринит с гноетечением из носа соответствовала оценке 2;
- оценка пищеварительной системы (установление симптомов гастроэнтерита). Легкий гастроэнтерит соответствовал оценке 1, тяжелый - оценке 2;
- оценка нервной системы (установление миоклоний, конвульсий и/или паралича). Миоклония соответствовала оценке 1, конвульсия - оценке 2 и паралич - оценке 3;
- наблюдение температуры тела животного.
Температура ниже или равная 37,5°С соответствовала оценке 3; температура, составляющая от 37,5°С до 39,5°С, соответствовала оценке 0; температура, равная 39,5°С или от 39,5°С до 40°С, соответствовала оценке 1; температура, равная 40°С или составляющая от 40°С до 40,5°С, соответствовала оценке 2; и, наконец, температура выше 40,5°С, соответствовала оценке 3.
У погибших подопытных животных извлекали селезенку для детекции вируса Карре путем иммунофлуоресценции.
Общую клиническую оценку рассчитывали следующим образом: для каждого клинического симптома вычисляли среднюю оценку на группу животных в период наблюдения, а общую оценку вычисляли как сумму средних оценок для пяти рассмотренных клинических симптомов.
Результаты тестов после иммунизации собак внутримышечно:
* средний титр +/- стандартное отклонение
* средний титр +/- типичное отклонение
Плазмиды pNS016 и pNS017, включающие соответственно природный ген М и природный ген N, были сконструированы так же, как плазмида pNS018.
Неожиданно было обнаружено, что защита, индуцируемая одним оптимизированным НА, выше или равна защите, индуцируемой смесью оптимизированных НА и F или оптимизированных НА и F и нативных М и N.
Должно быть совершенно понятно, что изобретение, точно описанное в прилагаемых пунктах формулы изобретения, не ограничено отдельными вариантами осуществления, указанными в вышеприведенном описании, а охватывает варианты, которые не входят ни в рамки, ни в изобретательский уровень настоящего изобретения.

















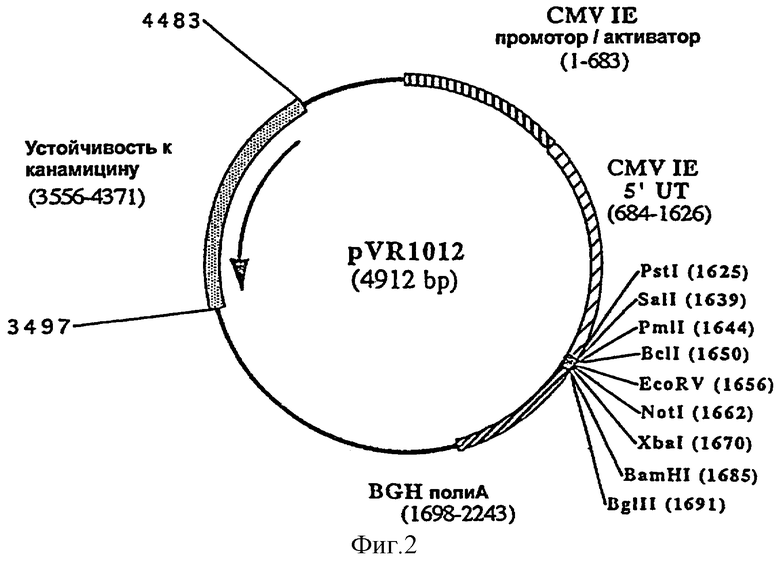

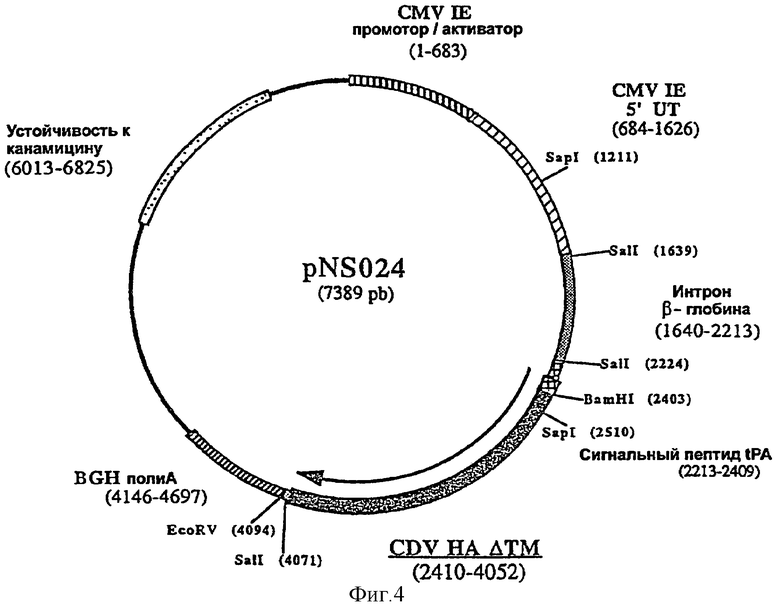
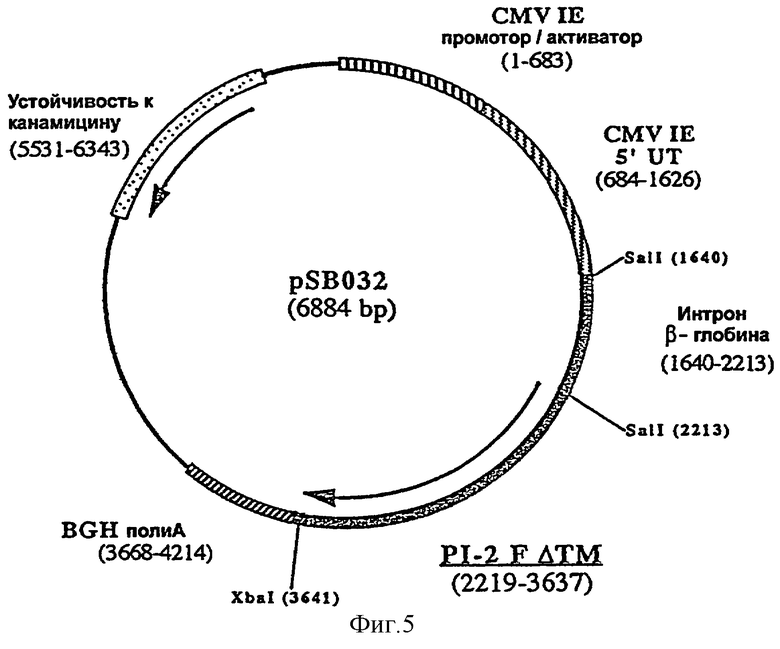
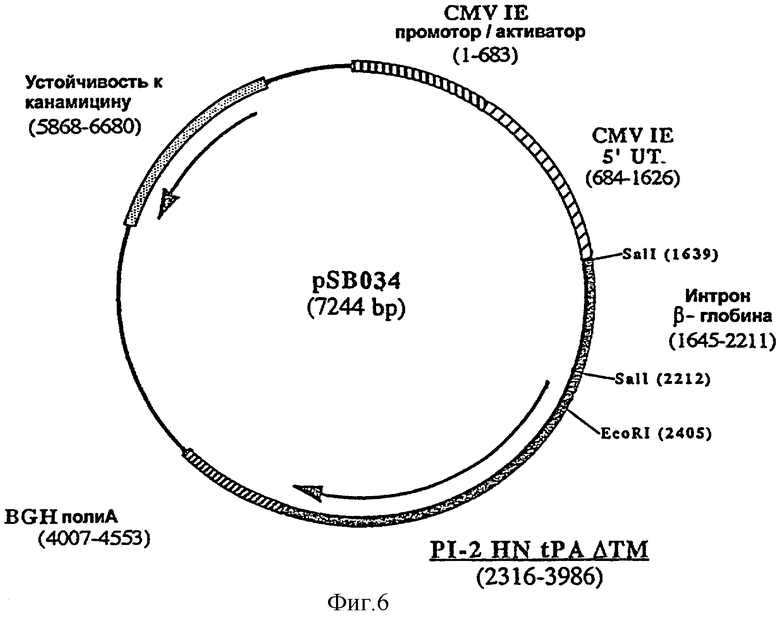
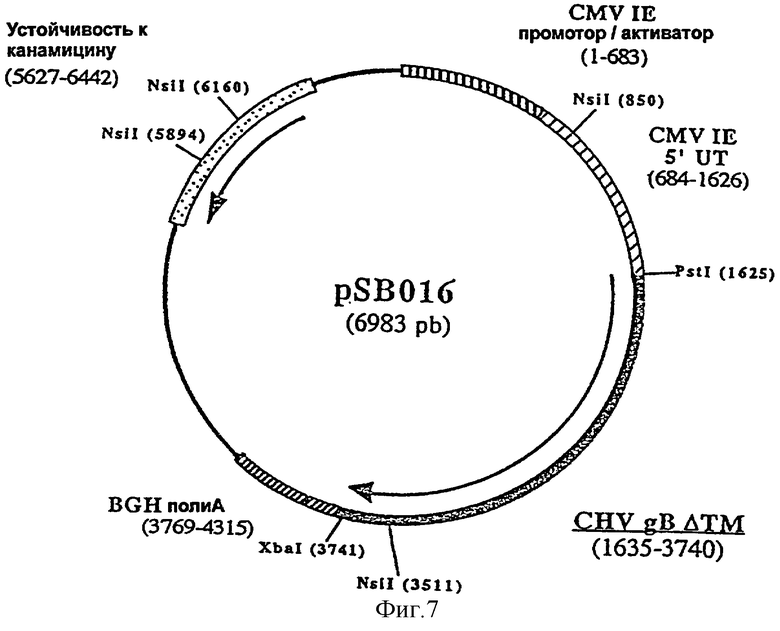
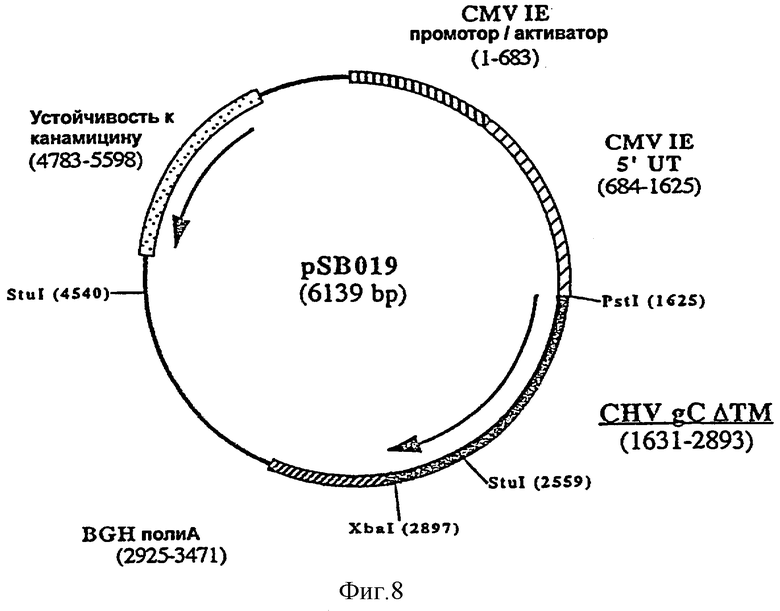
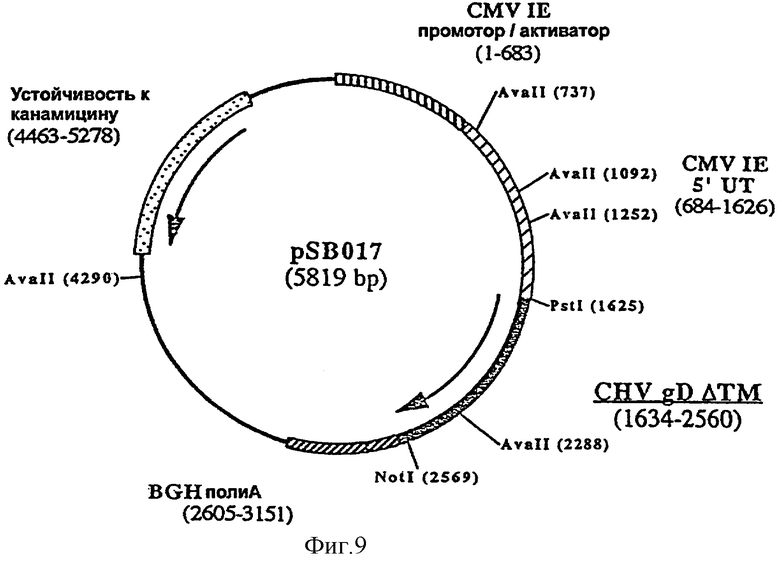
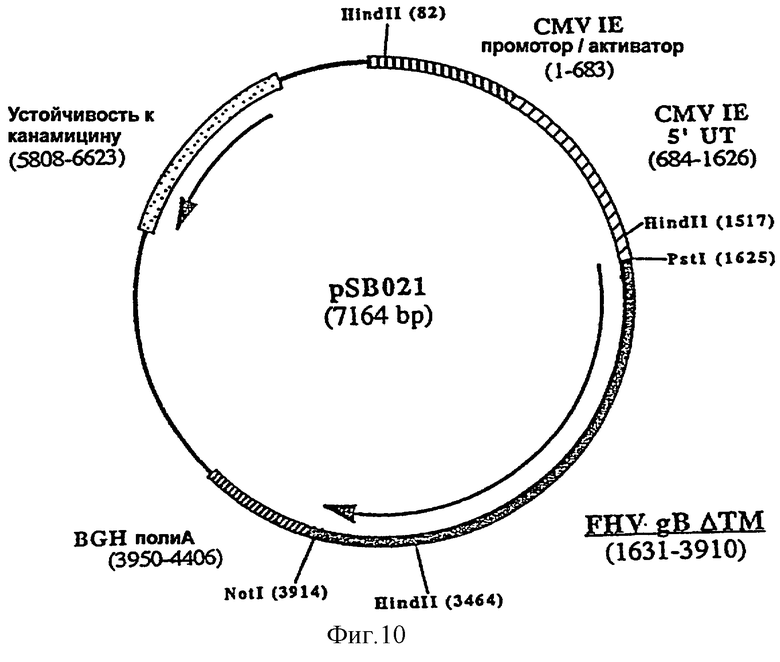
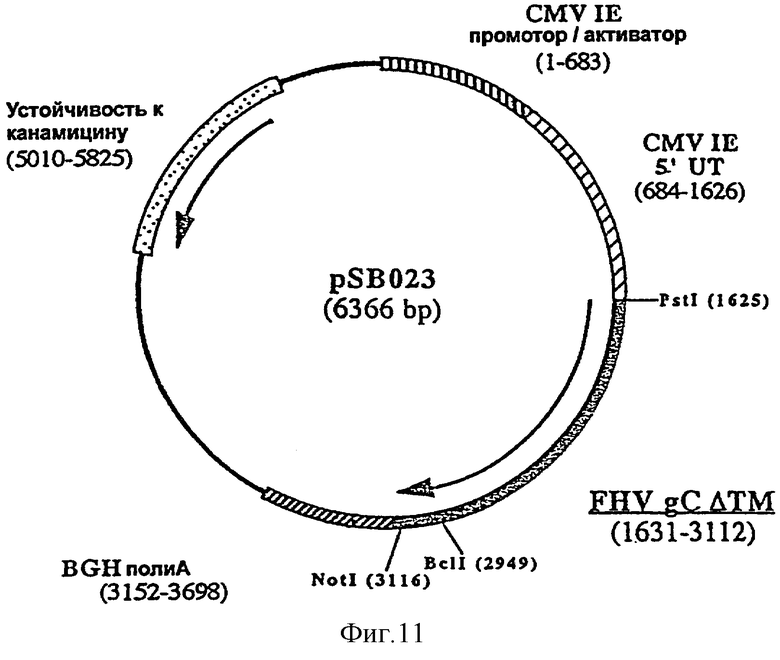
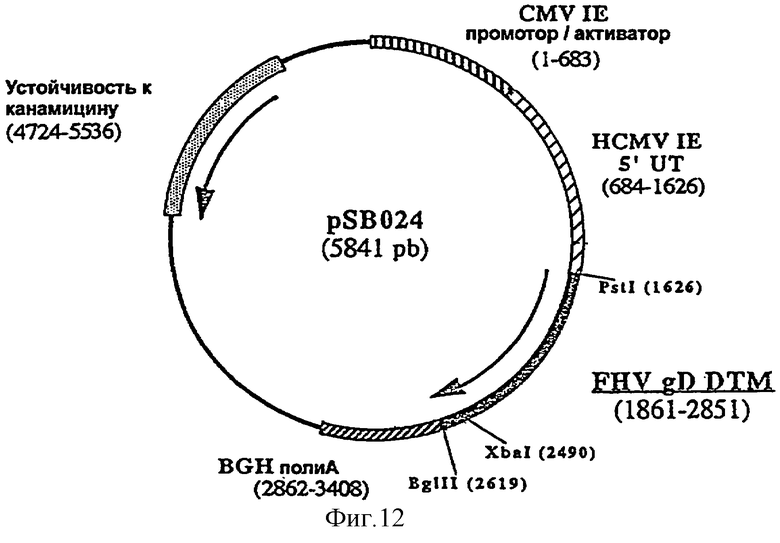
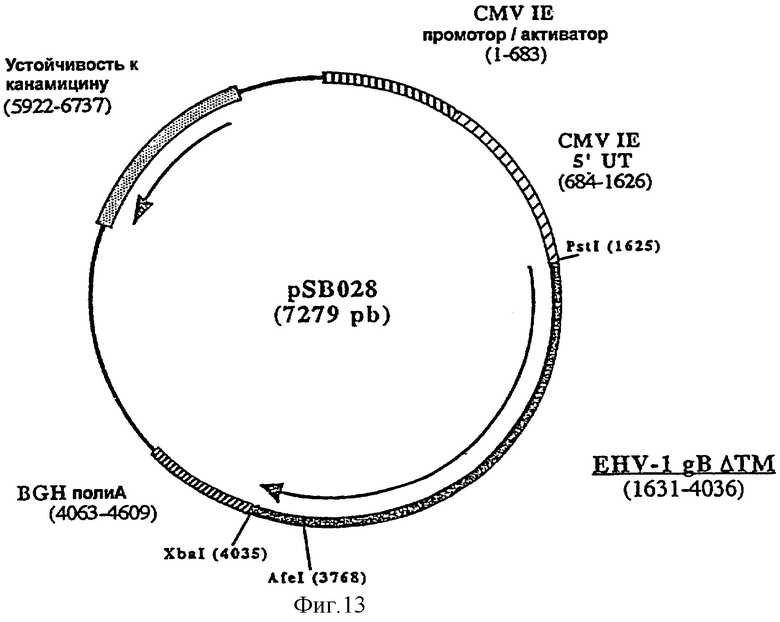
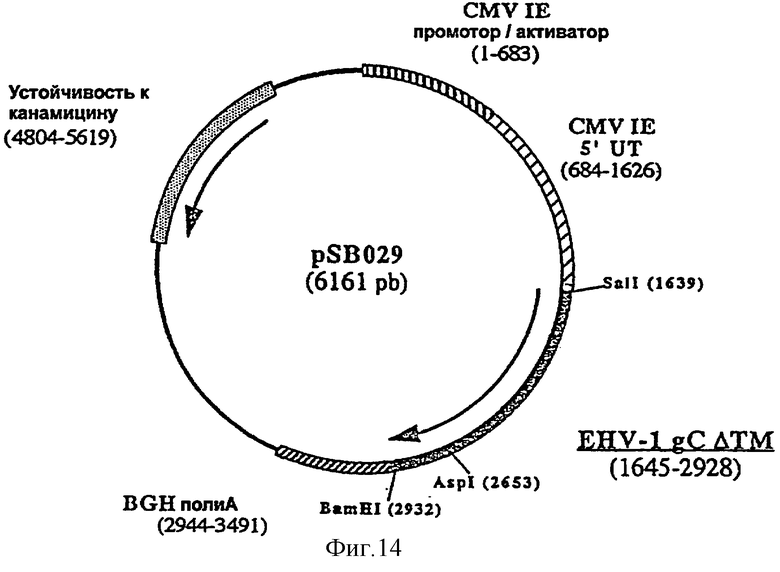
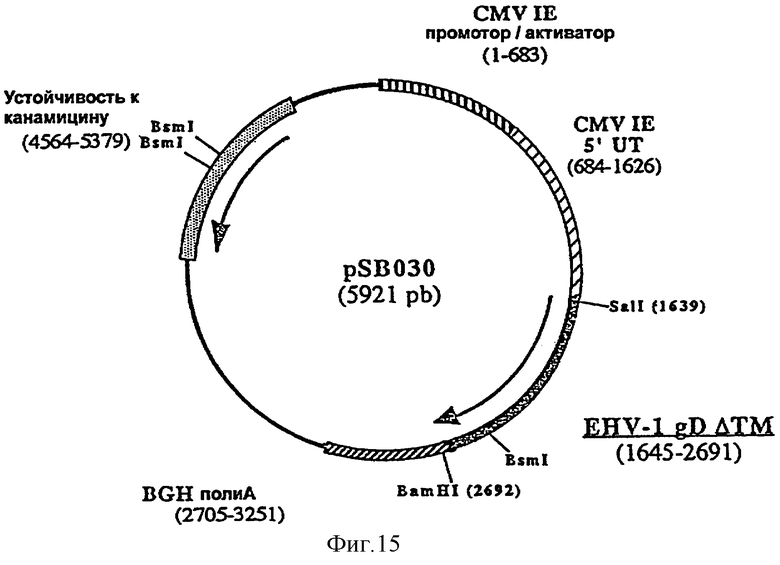
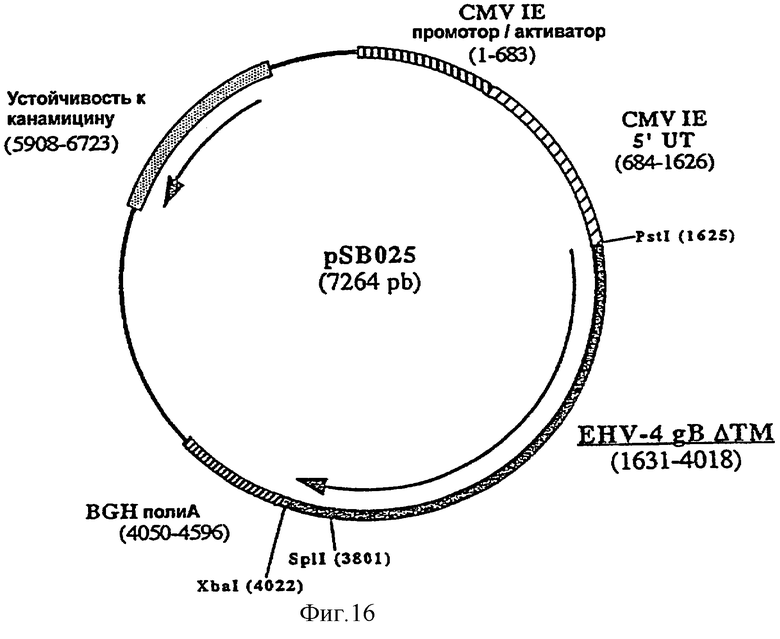
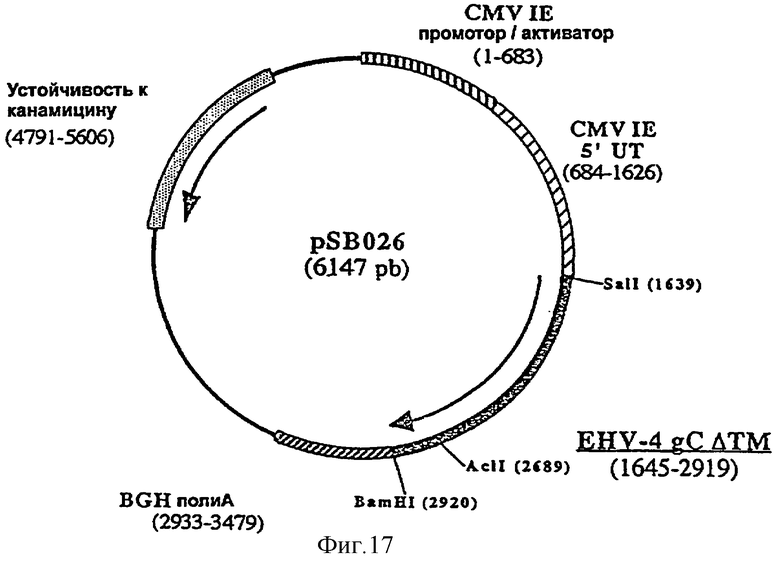
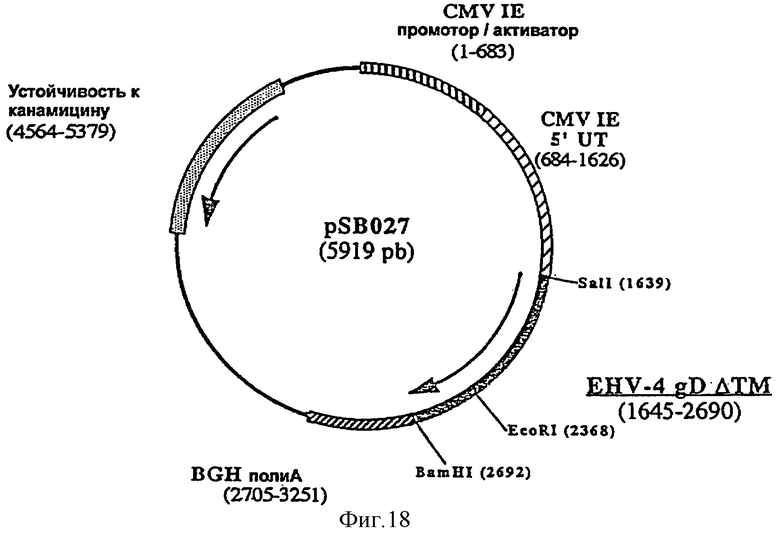
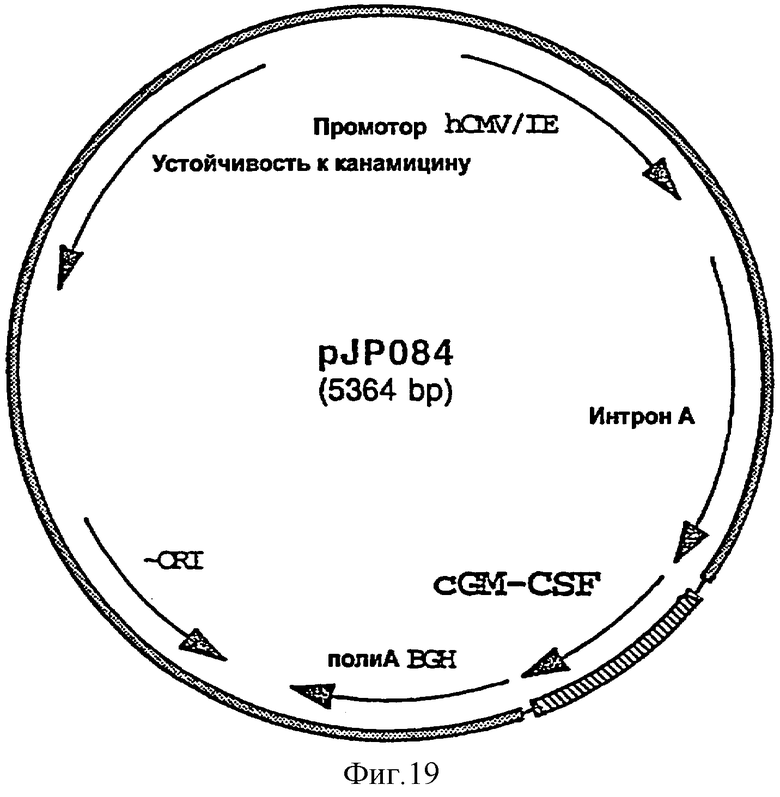

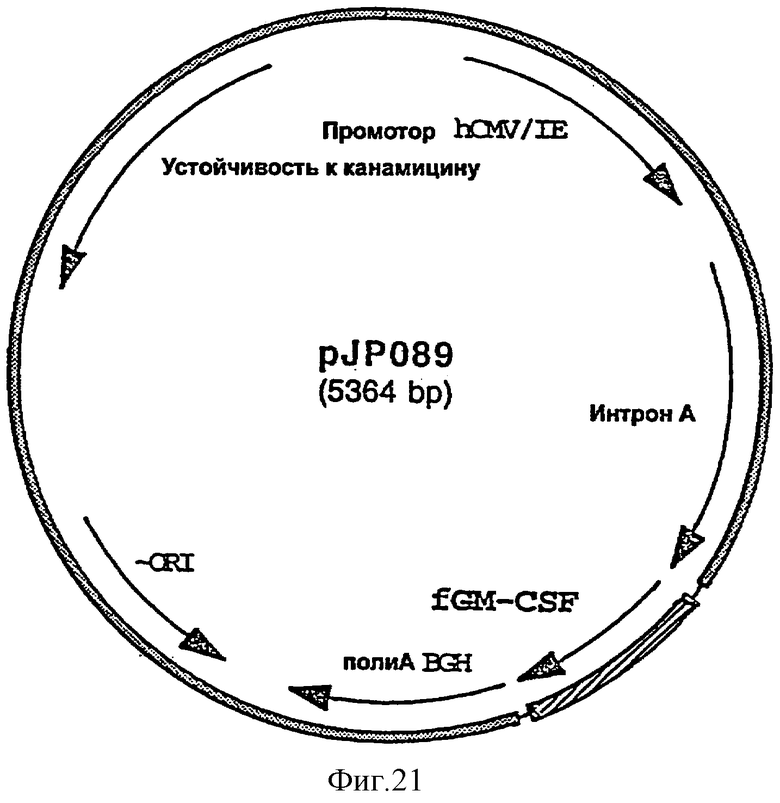

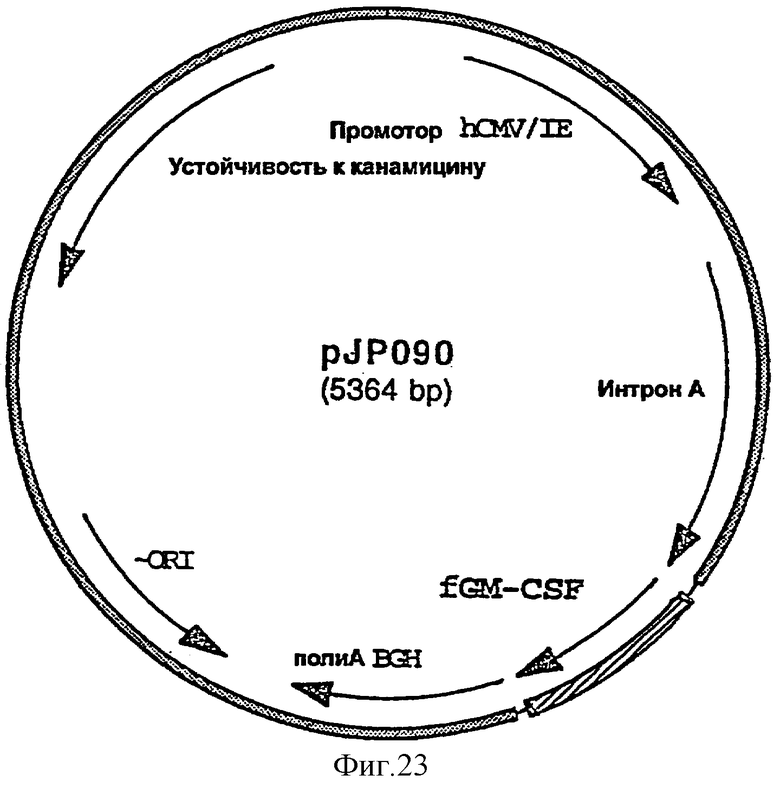

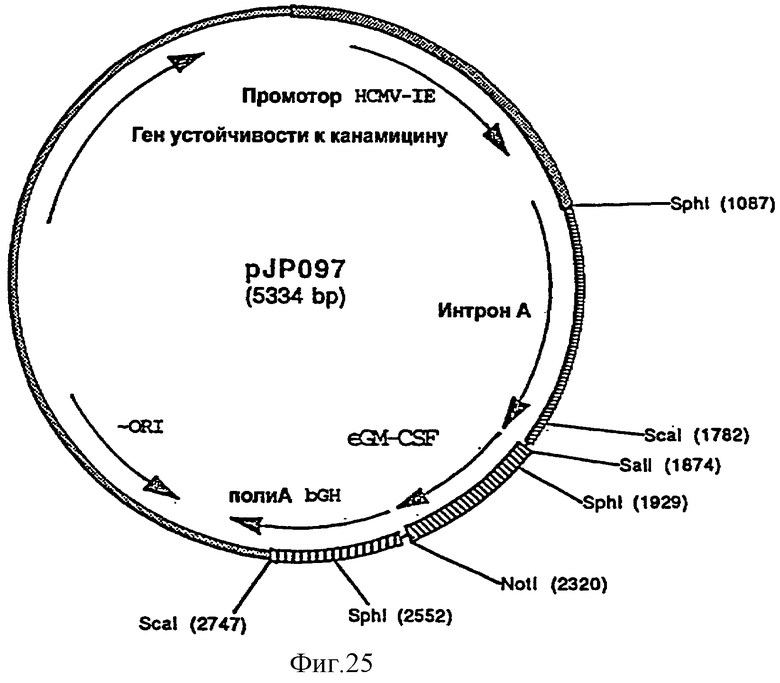

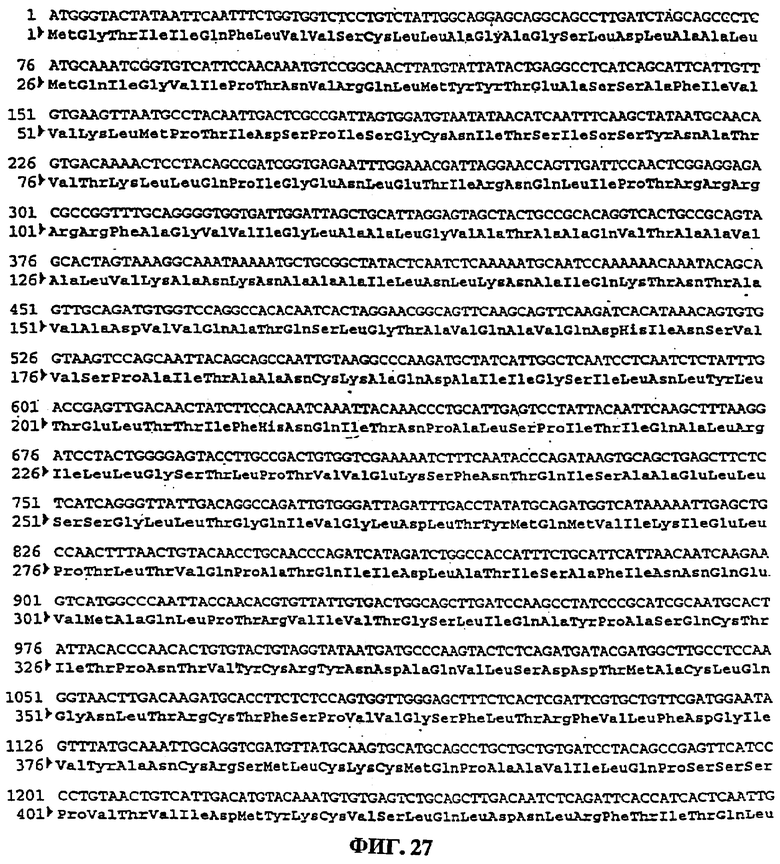



Изобретение относится к генетической инженерии, вирусологии и ветеринарии. Предложена ДНК-вакцина против вируса чумки собак (CDV). Вакцина включает, по меньшей мере, одну ДНК-плазмиду и в качестве адъюванта - катионный липид, содержащий соль аммония согласно формуле, приведенной в п.1 формулы изобретения. При этом ДНК-плазмида содержит и экспрессирует в клетках хозяина нуклеотидную последовательность, кодирующую иммуноген CDV. При этом нуклеотидная последовательность содержит делецию участка, кодирующего трансмембранный домен. Предложена также поливалентная вакцина, содержащая такую ДНК-вакцину, и иммуногенная композиция, содержащая такую ДНК-плазмиду. Изобретение может быть использовано в ветеринарии для вакцинации собак против CDV. 4 н. и 37 з.п. ф-лы, 28 ил., 3 табл.
в которой R1 представляет собой насыщенный или ненасыщенный линейный алифатический радикал, содержащий от 12 до 18 атомов углерода, R2 - другой алифатический радикал, содержащий от 2 до 3 атомов углерода, а Х - гидроксил или аминогруппа.
в которой R1 представляет собой насыщенный или ненасыщенный линейный алифатический радикал, содержащий от 12 до 18 атомов углерода, R2 - другой алифатический радикал, содержащий от 2 до 3 атомов углерода, а Х - гидроксил или аминогруппа.
| SIXT N | |||
| et al | |||
| Canine distemper vims DNA vaccination induces humoral and cellular immunity and protects against a lethal intracerebral challenge | |||
| J | |||
| Virol | |||
| Способ и аппарат для получения гидразобензола или его гомологов | 1922 |
|
SU1998A1 |
| WO 9634109, 31.10.1996 | |||
| US 5679356, 21.10.1997 | |||
| NASH R.A | |||
| et al | |||
| Molecular cloning and in vivo evaluation of canine granulocyte-macrophage colony-stimulating factor | |||
| Blood | |||
| Циркуль-угломер | 1920 |
|
SU1991A1 |
| Aug | |||
| Прибор для нагревания перетягиваемых бандажей подвижного состава | 1917 |
|
SU15A1 |

