Изобретение относится к гликопротеину VI (GPVI), его выделению, очистке и способу получения рекомбинантными методами. В частности, изобретение относится к применению GPVI, предпочтительно - рекомбинантного GPVI, для лечения расстройств и патологий, которые либо непосредственно, либо косвенным образом связаны с такими расстройствами коагуляции крови, как тромбозные и сердечно-сосудистые заболевания. Внеклеточный рекомбинантный протеин можно также применять для скрининга на предмет потенциальных ингибиторов связанного с мембраной GPVI с целью ингибирования взаимодействия тромбоцитов с коллагеном. На протяжении времени жизни тромбоцита в естественных условиях (in vivo) GPVI на его поверхности видоизменяется и может быть использован как маркер для определения возраста тромбоцита.
Гликопротеин VI представляет собой гликопротеин 62/65 кДа (невосстановленный/восстановленный соответственно) мембраны тромбоцита, который образует комплекс с общей субъединицей Fcγ. Субъединица GPVI содержит сайт, связывающий коллаген, а субъединица Fcγ. ответственна за сигнализацию. Этот комплекс является одним из основных рецепторов коллагена на поверхности тромбоцита, ответственных за активирование тромбоцита по отношению к коллагену. Распознавательная последовательность на коллагене состоит из последовательностей (GlyProHyp)n. Известны пациенты из Японии с генетическим дефицитом GPVI. У них проблемы со свертываемостью крови и их тромбоциты весьма слабо реагируют на коллаген, вероятно, через другие рецепторы. Было обнаружено, что сигнальные каскады GPVI нежизнеспособны; эти каскады имеют большое сходство с сигнальными каскадами иммунных рецепторов, в том числе рецепторов Т-клеток, В-клеток и рецепторов природных клеток-убийц. Эти каскады включают группу src тирозинкиназ, таких как Fyn и Lyn, а также p72SYK и многие другие тирозинкиназы, фосфатазы и адаптер типа LAT. Главное назначение этих каскадов - активация фосфолипазы Сγ2, которая расщепляет фосфолипиды на вторые молекулы диацилглицерола и IP3. Предполагается, что GPVI вовлечен в активацию интегрина α2β1 тромбоцита, который играет главную роль в адгезии тромбоцитов с поврежденной стенкой сосуда. Мыши с "выключенной" субъединицей Fcγ. имеют тромбоциты, которые по-прежнему реагируют на коллаген, откуда следует, что состояние покоя α2р1 тоже должно регулироваться комплексом GPVI/Fcγ.
Рецептор коллагена GPVI тромбоцитов тесно связан с активаторными рецепторами семейства p58KAR природных убийц и с FcαR.
Адгезия и активация покоящихся тромбоцитов, циркулирующих на поврежденном участке сосуда, - первый шаг в процессе, который приводит к образованию тромба, который преобразуется в кровоостанавливающую пробку. Коллаген - один из главных компонентов стенки сосуда, ответственный за активацию тромбоцита. Существуют много типов коллагена, и семь из них обнаружены в субэндотелиальных слоях. На тромбоцитах идентифицированы несколько различных рецепторов коллагена, однако в настоящее время главными считаются интегрин α2β1 и неинтегрин GPVI. Притом, что α2β1 всесторонне изучены, и обе субъединицы были клонированы и секвенированы несколько лет назад, структура GPVI остается невыясненной, хотя определены некоторые ее свойства. Около двадцати лет назад было установлено, что GPVI - главный гликопротеин тромбоцита; он имеет массу в интервале 60-65 кДа и кислотное pi. Его роль как предполагаемого рецептора коллагена была установлена после того, как обнаружилось, что тромбоциты у пациента из Японии с плохой свертываемостью крови имеют специфический дефект - они слабо реагируют на коллаген и у них отсутствует названный рецептор. Кроме того, у этого пациента имелись аутоантитела для отсутствующего рецептора, при помощи которых оказалось возможным определить и другие характеристики молекулы. Позднее было установлено, что GPVI образует нековалентную связь с общей субъединицей Fcγ, которая функционирует как сигнальная часть комплекса. Кроме того, было показано, что распознающая GPVI последовательность коллагена представляет собой повторяющийся триплет Gly-Pro-Hyp внутри тройной спирали структуры коллагена, и что основанные на этой структуре синтетические пептиды можно использовать как специфические в отношении GPVI агонисты. Установлено, что комплекс GPVI/Fcγ подает сигнал внутренней части тромбоцита посредством иммунного механизма рецепторного типа, включающего активацию p72SYK, и вызывает каскад взаимодействий киназа/фосфатаза/адаптер протеина, что приводит к активации PCLγ2 и, следовательно, к освобождению гранул и агрегации тромбоцитов. Следующий шаг в выяснении характеристик этой молекулы был сделан, когда обнаружилось, что змеиный лектин С-типа, конвулксин тропической гремучей змеи Crotalus durissus terrificus может активировать тромбоциты путем кластеризации GPVI в ходе многомерного взаимодействия. Было показано, что конвулксин образует специфическую связь с GPVI, что позволило на этой основе разработать методику очистки данного рецептора.
Из всего сказанного следует, что соединение GPVI представляет большой интерес для многих терапевтических сфер, и его особенно важные применения относятся к сфере, которая связана непосредственно или косвенным образом с коагуляцией крови, которая зависит от коллаген-тромбоцитных взаимодействий. В связи с этим цель настоящего изобретения состояла в получении GPVI в рекомбинантной форме и в том, чтобы продемонстрировать его эффективность в прямом терапевтическом назначении, а также - как инструмента для скрининга коротких соединений, в особенности - синтезированных или синтезируемых химически соединений, с возможностью ингибировать или блокировать взаимодействие между природными тромбоцитом и коллагеном. Изобретение относится также к долям или фрагментам протеина GPVI, сохранившим свою биологическую активность, которая выражается в способности образовывать связи с коллагеном.
Изобретение было успешно осуществлено в плане очистки адекватных количеств GPVI для предварительного исследования и секвенирования пептидов. Последовательности были использованы с целью конструирования праймеров для PCR, чтобы идентифицировать положительную последовательность в библиотеке ДНК. Эту последовательность ДНК далее использовали в качестве детектора для выделения из библиотеки почти полной последовательности кДНК, а недостающая 5'-последовательность была получена методом RACE из библиотеки кДНК тромбоцитов.
Изобретение было успешно осуществлено также в плане демонстрации применения рекомбинантных GPVI в качестве терапевтически применимого соединения, которое способно при его назначении, например, пациенту с поврежденными кровеносными сосудами, связывать коллаген и, таким образом, предотвращать появление тромбоцитов со связанными с мембраной GPVI, которые появляются при образовании связей с названным коллагеном. Растворимый рекомбинантный внеклеточный домен GPVI содержит связывающий коллаген сайт и может предотвращать активацию тромбоцита коллагеном. Таким образом, его можно применять для лечения проявлений заболеваний, вызывающих повышенную активацию тромбоцитов коллагеном, таких как атеросклеротическое разрушение тромбоцитов, при таких заболеваниях, как нестабильная стенокардия, или в процессе хирургической операции, такой как подкожная трансиллюминационная коронарная ангиопластика (РТСА), при которой возобновляют проходимость артерий путем введения баллонного катеттера, что приводит к значительному повреждению стенок сосудов и активации большого количества тромбоцитов, что часто обуславливает впоследствии их повторную закупорку. Преимущество применения рекомбинантных фрагментов GPVI по сравнению с существующими методами лечения состоит в том, что они начинают действовать на более ранней стадии путем предотвращения или замедления активации тромбоцитов, а не путем подавления эффектов, возникающих в результате активации тромбоцитов, таких как агрегация антагонистами GPIIb-IIIa. Таким образом, высвобождается меньшее количество содержимого тромбоцитных гранул, включая факторы роста и хемокины, которые участвуют не только в заживлении ран, но и в реконструкции стенки сосуда за счет перемещения гладкой мускулатуры, а также в привлечении фагоцитных клеток - таких, как моноциты, которые стимулируют атеросклероз. В тех же целях и с аналогичным эффектом для блокирования GPVI на поверхности тромбоцита можно применять фрагменты Fab приближенных к человеческим мышиных моноклональных антител против GPVI.
Рекомбинантные GPVI по данному изобретению можно применять также в связывающем анализе коллагена для скринирования маленьких молекул (например, в комбинаторных библиотеках), которые могут ингибировать это взаимодействие и которые можно применять для получения терапевтических соединений, являющихся ингибиторами взаимодействия коллагена с тромбоцитами. Путем соответствующей дериватизации эти соединения можно сделать пригодными для орального приема. Главной целью данного изобретения является получение соединений, уменьшающих взаимодействие GPVI-коллаген и, следовательно, активацию тромбоцита в ситуациях, когда возникает контакт тромбоцитов с коллагеном. Технология скрининга, подобная используемой в данном изобретении, хорошо обоснована в прототипе. При использовании такого скрининга изобретение позволяет обнаруживать и получать новые мишени, которые способны, как антагонисты коллагена, ингибировать образование природных связанных с мембраной GPVI на поверхности тромбоцита. Такие мишени, которые могут представлять собой маленькие химические молекулы, могут создать основу для дальнейших изобретений.
Другое важное применение GPVI и реагентов, которые распознают специфические домены GPVI, - это их применение в качестве маркеров возраста и функциональности тромбоцита. Предполагается, что в общем случае молодые тромбоциты более активны и функциональны, чем они старшие. Конвулксин - пектин С-типа из змеиного яда - связывает и активирует молодые тромбоциты, поскольку он специфичен по отношению к GPVI, однако, по мере старения тромбоцитов это связывание и степень активации снижаются. Это может происходить за счет либо протеолитических, либо конформационных изменений GPVI или его ассоциатов с Fcγ. в результате активации тромбоцита или нарушений циркуляции. Это может оказаться полезным параметром для измерения при медицинском обследовании возрастных и функциональных профилей тромбоцитов у пациентов и у здоровых людей. Возрастные профили тромбоцитов изменяются при многих заболеваниях, поражающих костный мозг или иммунную систему и, если будут разработаны лучшие методы для их определения, могут стать важным критерием в диагностике. Например, у пациентов с заболеваниями, которые сопряжены с ускоренным обновлением тромбоцитов, должно наблюдаться сравнительно большое количество молодых тромбоцитов, в то время как у пациентов, которые проходят курс химиотерапии или облучения, будет преобладать более взрослая популяция. Таким образом, подобные возрастные профили можно использовать для четкого контролирования процесса лечения. В отношении здорового населения о распределении возрастных профилей и их роли в качестве прогнозирующего параметра известно очень мало. Неизвестно, происходят ли изменения в GPVI за счет частичного вовлечения тромбоцитов в события кровоостанавливания, и будут ли изменения более выраженными у пациентов с обширным сердечно-сосудистым заболеванием. В настоящее время для определения молодых сетчатых тромбоцитов, содержащих мРНК, используется тиазоловый оранжевый. Эта мРНК быстро убывает, ограничивая применимость метода только самыми молодыми тромбоцитами. Реагенты, которые следовало бы применять в таком анализе, должны включать специфические по отношению к GPVI протеины змеиного яда, такие как конвулксин, или моноклональные, либо поликлональные антитела, распознающие области N-концов GPVI, или моноклональные антитела, распознающие новые сайты или структуры, обусловленные протеолизом доменов концов N, либо специфические структуры, присутствующие только в неповрежденных молекулах и отсутствующие в старых молекулах или наоборот, или маленькие химические объекты, выбранные для специфической идентификации неповрежденных GPVI, либо его модифицированной формы. Эти реагенты могут быть помечены флюоресцентным маркером или вторым антителом с флюоресцентной меткой, либо реагентом со специфическим сродством; их можно применять проточной цитометрией для измерения профилей связывания тромбоцитов. На последующей стадии, альтернативно можно внедрить менее трудоемкую методику с использованием аппаратуры для автоматического измерения профилей тромбоцитов. При помощи таких методов сортировки клеток, как проточная цитометрия или магнитометрия, окажется возможным выделять молодые и старые тромбоциты для изучения факторов, воздействующих на удаление из системы циркуляции старых тромбоцитов. Для подобных исследований ключевыми являются реагенты, распознающие специфические формы GPVI.
Таким образом, предметом настоящего изобретения является кодирование ДНК для Гликопротеина VI или их биологически активных фрагментов, особенно - последовательности, представленной на Фиг.2.
Другим предметом настоящего изобретения является кодирование ДНК для Гликопротеина VI, включающего аминокислотные последовательности, представленные на Фиг.1a и 1b.
Следующим предметом данного изобретения является фармацевтическая композиция, включающая эффективное рекомбинантный GPVI совместно с разбавителем, приемлемым с точки зрения фармацевтики, носитель или эксципиент, и ее применение для производства лекарств для лечения тромбозных и сердечно-сосудистых заболеваний, а также нарушений, связанных с взаимодействием тромбоцитов с коллагеном. Эта фармацевтическая композиция наряду с указанным протеином может содержать другие известные из уровня техники фармацевтически активные соединения, пригодные для лечения вышеуказанных заболеваний и нарушений
Кроме того, предметом данного изобретения является применение рекомбинантного GPVI в качестве инструмента скринирования - для определения специфических ингибиторов взаимодействий тромбоцитов с коллагеном.
Еще одним предметом данного изобретения является применение GPVI в качестве маркера возраста тромбоцита и для обнаружения сердечно-сосудистых заболеваний.
Возможными медицинскими показаниями и назначениями соответственно являются, например, нестабильная стенокардия, РТСА, введение стентов, операции на сердечных сосудах, общие операции на кровеносных сосудах, операции, которые могут привести к повреждению более крупных кровеносных сосудов, такие как операции на тазобедренных суставах. Кроме того, сюда входят все показания, которые связаны с тромбоэмболическими явлениями, обусловленными расстройствами взаимодействий стенки сосуда с коагуляционной системой, что обуславливает высокую степень риска образования тромбов и блокирования сосудов.
Как показано выше, протеины GPVI и их фрагменты по данному изобретению пригодны для применения в качестве фармацевтически эффективных соединений в фармацевтических композициях и комбинациях.
Фармацевтические рецепты по данному изобретению могут включать дополнительные активные ингредиенты, например антикоагулянты, такие как гирудин или гепарин, либо тромболитические агенты, такие как активатор плазминогена или гементин, либо антагонисты другим рецепторам тромбоцита, например антагонисты GPIIb-IIIa, такие как абциксимаб или эптифибатид, либо антагонисты ADP-рецептора, такие как клопидогрель.
По данному изобретению новый протеин и его биологически активные фрагменты соответственно могут образовывать фармацевтически приемлемые соли с любыми нетоксичными органическими или неорганическими кислотами. В качестве неорганических кислот могут быть использованы, например, соляная, бромистоводородная, серная или фосфорная, а также кислые соли металлов, такие как моногидрогенортофосфат натрия и гидрогенсульфат калия. Примерами органических кислот являются моно-, би- и трикарбоксильные кислоты типа уксусной, гликолевой, молочной, пировиноградной, малоновой, янтарной, глутариловой, фумаровой, яблочной, виннокаменной, лимонной, аскорбиновой, малеиновой, гидроксималеиновой, бензойной, гидроксибензойной, фенилуксусной, коричной, салициловой и сульфоновые кислоты типа метансульфоновой кислоты. Соли карбоксиконцевых компонентов аминокислот включают нетоксичные соли карбоновой кислоты, которые образуются в их реакции с любыми подходящими неорганическими или органическими основаниями. В эти соли могут входить, например, щелочные металлы, такие, как натрий и калий, щелочноземельные металлы, такие как кальций и магний, легкие металлы группы IIIA, включая алюминий, и органические первичные, вторичные и третичные амины, такие как триалкиламины, включая триэтиламин, прокаин, дибензиламин, 1-этенамин, N,N'-дибензилэтилендиамин, дигидроабиэтиламин и N-алкилпиперидин.
В контексте данного изобретения, термин "фармацевтически приемлемый носитель" обозначает инертный, нетоксичный твердый или жидкий наполнитель, растворитель или капсулирующий материал, который не вступает в реакции с действующим соединением или с организмом пациента. Подходящие, предпочтительно жидкие, носители хорошо известны в фармацевтике, к ним относятся, например, стерильные вода, физраствор. жидкая декстроза, сахарные сиропы, этанол, гликоли и масла, включая масла нефтяного, животного, растительного или синтетического происхождения, например арахисовое, соевое и минеральные масла.
Препараты с рецептурой по данному изобретению можно назначать в виде единичных доз, содержащих подходящие нетоксичные фармацевтически приемлемые носители, растворители, вспомогательные лекарственные вещества и связующие, которые обычно используются в фармацевтике для парентеральных назначений.
Термин "парентеральный" охватывает подкожные, внутривенные, внутрисуставные и внутритрахеальные инъекции, а также инфузионные методы. Возможны также оральные назначения и местное применение. Парентеральные составы и комбинации наиболее предпочтительно назначать внутривенно или в виде пилюль, а также в виде постоянной фузии по известной методике. Таблетки и капсулы для оральных назначений содержат подходящие эксципиенты, такие как связующие вещества, наполнители, растворители, таблетирующие вещества, смазочные компоненты, дезинтегрирующие агенты и увлажнители. На таблетках можно формировать покрытия по известным фармацевтическим методикам.
Жидкие оральные препараты можно назначать в виде водных или масляных суспензий, растворов, эмульсий, сиропов и эликсиров; они могут быть представлены также сухим веществом, предназначенным для разведения перед приемом в воде или в другом подходящем носителе. Такие жидкие препараты могут содержать подходящие добавки типа суспендирующих или эмульгирующих агентов, неводных носителей и консервантов. Препараты для местного применения могут быть водными или масляными суспензиями, растворами, эмульсиями, желе, или - что предпочтительно - мазями.
Единичные дозы по настоящему изобретению могут содержать необходимые суточные количества протеина по данному изобретению или их кратные части, из которых можно скомбинировать суточную дозу. Оптимальная терапевтически приемлемая дозировка и дозы, которые требуются конкретному пациенту (млекопитающие, включая человека), зависят от множества факторов, таких как степень активности, связанная с видом деятельности пациента, его возраст, вес, общее состояние здоровья, пол, диета, время и курс назначения препарата, объект лечения, т.е. терапия или профилактика, природа тромбозного заболевания, степень выраженности антитромбоцитной или антикоагуляционной активности.
В составах и комбинациях, показанных для конкретного пациента (in vivo) в качестве антикоагулянтов, фармацевтически эффективная суточная доза пептидов по данному изобретению варьируется в диапазоне от примерно 0,01 до 100 мг/кг веса тела, предпочтительно - от 0,1 до 10 мг/кг веса. В зависимости от способа применения одна доза может содержать от 0,5 до 10 мг ингибитора коллагена. Чтобы достичь антикоагуляционного эффекта в искусственной крови, фармацевтически эффективное количество пептидов по данному изобретению составляет от 0,2 до 150 мг/л искусственной крови, предпочтительно - от 1 до 20 мг/л.
Краткое описание чертежей
Фиг.1. Протеиновая последовательность GPVI (однобуквенный код):
1а: Лидирующая последовательность
1b: Зрелый протеин
Открытая рамка считывания: 339 аминокислот
Фиг.2. Нуклеотидная последовательность GPVI, охватывающая открытую рамку считывания из 1017 п.о. плюс области 3" и 5', в общей сложности - 1249 п.о.
Подробное описание изобретения
Для синтеза праймеров ДНК были выбраны две последовательности из 7 аминокислот с минимальной вырожденностью генетического кода с целью укрупнения при помощи кДНК участка GPVI в реакции PCR. Поскольку, в общем случае, локализация обоих пептидов в протеине не была известна, для каждого из них были приготовлены два вырожденных праймера - один смысловой и один антисмысловой. Эти праймеры были использованы для расширения библиотеки человеческого костного мозга. Комбинация смыслового праймера 5'TYA THC CNG CAN TGA ARMG 3' кодирует последовательность PAMKRSL, а антисмысловой праймер 5'TTR TAN ARN GCR AAY TGR TC 3' соответствует укрупненному DQFALYK фрагменту ДНК из 221 пар основ Дополнительно к выбранным пептидам амплифицированная ДНК кодирует LysC/AspN пептидной DQLELVATGVFAKPSLSAQPGPAVSS, посредством чего связывает последовательность с кДНК для GPVI.
Скрининг 600000 pfu из библиотеки костного мозга при помощи этого фрагмента ДНК из 221 пар основ позволил получить 4 положительных pfu. Три из них имели вставки из 1350 пар основ, обрезанные рестриктазами Sal I или EcoR I и принадлежащие к надсемейству IgG. Четвертая имела вставку 4,6 тысячи гетероциклических оснований нуклеотидов нуклеиновой кислоты, полученную в результате расщепления при помощи Sal I, и после обработки EcoRI предоставила два фрагмента из 2300 пар основ и 1300 пар основ соответственно. Ее ДНК кодировала последовательность для 10 пептидов, полученных в результате аминокислотного секвенирования GPVI, но с оборванными аминоконцами. Не было обнаружено ни исходного метионина, ни лидирующей последовательности, однако присутствовали более 2000 пар основ предварительно секвенированной несчитывающей рамки ДНК, заканчивающейся последовательностью Alu. Эксперимент RACE с концом 5′ был проведен на поли- А РНК тромбоцита с праймерами, локализованными во фрагменте последовательности GPVI, усиленной пептидными последовательностями. До возврата к установленной последовательности GPVI были обнаружены Фрагмент из 348 пар основ, включающий 278 пар основ последовательности четвертого клона и 70 новых пар основ из 1987 пар основ, соответствующих 14 аминокислотам, включая первый метионин. Таким образом, оказалось возможным секвенировать кДНК, содержащую, в общей сложности, 1249 пар основ, последовательность 5' из 25 пар основ, предшествующую стартовому кодону, открытую рамку считывания из 1017 пар основ, кодирующих протеин, включая лидирующую последовательность из 339 аминокислот и области 3' из 207 пар основ, включая стоп-кодон.
С целью восполнить недостающую последовательность 5' была клонирована кДНК, кодирующая GPVI тромбоцита; клонирование и секвенирование осуществляли из библиотеки кДНК человеческого костного мозга при помощи RACE с мРНК тромбоцита. Открытая рамка считывания из 1017 пар основ кодирует 339 аминокислот и нетранслируемую область 3'. При анализе гидрофобности аминокислотных последовательностей полученного протеина было обнаружено присутствие двух предположительно трансмембранных доменов, предположительно сигнальной последовательности из 20 аминокислот и домена между остатками 247 и 265, состоящего из 19 аминокислот. Последовательность и ее аминокислотная трансляция показаны на фиг.2 и 1. Сравнение с аминокислотными последовательностями наиболее похожих молекул из банка генов с очевидностью свидетельствует, что полученная последовательность относится к надсемейству иммуноглобулина, а внеклеточные домены - две спирали Ig домена С2, образованные двумя дисульфидными мостиками. Одна из молекул относится к классу проникающих через мембрану протеинов с концами N на поверхности и проходит через мембрану однократно. Молекулы с наиболее короткими связями относятся к классу рецепторов природных убийц, в котором содержатся как ингибиторы, так и активаторы. Очевидно, что GPVI относятся к подклассу активаторов не только в связи с их функцией, но и потому, что они, в отличие от молекул класса ингибиторов, не содержат последовательность ITIM в своих цитоплазматических доменах. Кроме того, они не содержат каких-либо тирозиновых остатков, которые могли бы быть вовлечены в фосфорилирование. В этом домене присутствует некоторое количество треониновых и сериновых остатков, но они не соответствуют критериям для последовательностей общих типичных элементов структуры киназы. Как и рецепторы NK класса активаторов, GPVI содержит агрининовый остаток в качестве третьей аминокислоты проникающего через мембрану домена, который принимает участие в образовании комплекса с субъединицей Fcγ. Цитоплазматический домен содержит 51 аминокислоту и имеет только весьма небольшое сходство (в области, которая находится непосредственно под мембраной) с цитоплазматическими доменами других членов этого семейства. Соответственно, следует ожидать, что этот домен в GPVI может ассоциироваться скорее с другими типами цитоплазматических молекул, чем с остальными членами семейства. GPVI содержит только единственный предположительно N-гликозилирующий сайт на Asn69. Тем не менее домен, который находится непосредственно над мембраной после бета-слоев конца домена Ig, содержит много тиониновых и сериновых остатков, которые могут предоставлять сайты O-гликозилирования, аналогичные обнаруженным в GPIbα и GPV. Основной функцией этого O-гликозилирования представляется поставка рецепторных структур, распространяющихся далеко от поверхности тромбоцита, что облегчает взаимодействия с их объемистыми лигандами. Поскольку ранее GPVI был идентифицирован как сиалогликопротеин, то различие молекулярных масс между теоретическим значением массы аминокислот (37 кДа) и значением, полученным при помощи гель-электрофореза (65 кДа восстановленного вещества), должно объясняться этим гликозилированием.
При помощи рентгенокристаллографического анализа была установлена структура рецепторов природных убийц двух типов доменов и было показано, что два домена Ig образуют острый угол с рецепторным сайтом для несущих пептид антигенов HLA, расположенным с наружной части угла. Прямое сравнение структуры сайта, связывающего пептиды HLA, со структурой коллагена позволяет сделать вывод об общих истоках этих рецепторов, поскольку повторяющиеся альфа-спиралеобразные структуры сайта, связывающего HLA и пептиды, которые он содержит, чрезвычайно похожи на тройную спиральную структуру коллагена. Постулируется, что рецепторы природных убийц действуют по механизму димеризации, при помощи двух рецепторов, которые распознают два отдельных сайта HLA на клетке, которую исследует клетка-убийца. Возможно, что эта димеризация является частью механизма активации или дезактивации в зависимости от класса рецептора. В случае GPVI должна существовать возможность, что две молекулы GPVI будут объединяться с одной Fcγ., поскольку каждый мономер димера Fcγ. имеет распознающую последовательность. Тем не менее, стехиометрия пока неизвестна, и, основываясь на структуре коллагенов, можно предположить, что сила сигнала коллагеноподобных пептидов, которые действуют через GPVI и конвулксин, зависит от количества объединившихся комплексов GPVI/Fcγ. В число других рецепторов тромбоцита, принадлежащих к этому семейству Ig, входят ICAM-2 (CD102) и РЕСАМ (CD31).
Все микроорганизмы, колонии клеток, происходящие от одного общего предка, системы экспрессии, хозяева экспрессии, плазмиды, промоторы, резистивные маркеры, источники репликации, сайты рестрикции или другие фрагменты и части векторов, которые упомянуты в описании, не имеют непосредственного отношения к предмету данного изобретения они имеются в продаже и общедоступны. Если не оговорено особо, эти материалы используются исключительно как образцы, не имеют значения для сущности изобретения, и могут быть заменены другими инструментами и биологическими материалами.
Методики, которые требуется использовать по данному изобретению, подробно описаны выше и ниже. В отношении других методов, которые широко известны, подробное описание здесь отсутствует, но представлены ссылки на соответствующие патенты и литературные источники, где они описаны достаточно подробно (например, Sambrook и др., 1989, Molecular Cloning: A Laboratory Manual, 2nd Edition, Cold Spring Harbor; Harlow, Lane, 1988, Antibodies: A Laboratory Manual, Cold Spring Harbor).
ПРИМЕРЫ
Материалы - протеин А-сефароза, соединенные с пероксидазой, козьи антимышиные и антикроличьи антитела, альбумин бычьей сыворотки, яд Crotalus durissus temficus, агглютинин зародышей пшеницы (WGA), 4% зерненная арагоза с поперечными связями, активированная N-гидроксисукцинимидилхлорформиатом и Triton Х-114, приобретенные у Sigma Chemical Co. (St Louis, МО), октанол-N-метилглюкамид (ONMG) и нонаноил-N-метилглюкамид (NNMG) от Oxyl Chemie (Bobingen, Germany).
Пример 1: Выделение GPVI из тромбоцитов - мембранные гликопротеины выделяли из тромбоцитов по описанной ранее методике. Вкратце, тромбоциты (из 40 светлых слоев кровяного сгустка) промыли и растворили в 2% Triton X-114 в присутствии ингибиторов протеазы. Отделили Triton X-114 от водной фазы и детергентную фазу загрузили в колонку агглютинина зародышей пшеницы, связанного с сефарозой 4 В. Гликопротеины тромбоцитов элюировали посредством 10 мМ трис HCl, рН 7,4, 30 мМ NaCl, 0,2% октаниол-N-метилглюкамидом (ONMG) и 2% N-ацетилглюкозамином. После диализа и концентрирования раствор гликопротеинов загрузили в колонку конвулксина, связанного с 4% зерненной арагозой с поперечными связями (1 мг/мл), активированной N-гидроксилсукцинамидил-р-нитрофенилхлорформиатом. Колонку промывали 4 объемами 10 мМ трис HCl, рН 7,4, 30 мМ NaCl, 0,2% нонаноил-N-метилглюкамида (NNMG), после этого - 4 объемами 10 мМ трис HCl, рН7,4, 30 мМ NaCl и 2% NNMG. GPVI элюировали 0,08% раствором SDS в 10 мМ трис HCl, рН 7,4. Раствор сконцентрировали и загрузили в препаративный гель 8,5% полиакриламида; использовали установку Model 491 Prep Cell (BioRad, CA). Препаративный электрофорез проводили при условиях, описанных в инструкции производителя установки, которые здесь не приводятся. В результате элюирования GPVI получили единственную полосу при 65 кДа. Фракции отстаивали, концентрировали на установке Centricon-30 (Amicon, Beverly, MA) и повторно суспензировали в 10 мМ трис HCl, рН7,4 и 0,1% ONMG.
Пример 2: Анализ аминокислот GPVI - GPVI переваривали эндопротеиназами LysC и AspN (Boenringer Mannheim, Germany). Полученные 10 пептидов разделили при помощи жидкостной хроматографии высокого разрешения (HPLC) с обращенными фазами и секвенировали на импульсном жидкофазном секвенаторе протеина Applied Biosystem модели 477А, объединенном с фенилтиохидантоиловым анализатором аминокислот модели 120А.
Пример 3: Амплификация фрагмента из 221 п.о., кодирующего участок GPVI из библиотеки кДНК λ gt11 -
Образец (1010 pfu) (бляшкообразующее число) из библиотеки человеческого костного мозга (Clonetech, Palo Alto, CA) был амплифицирован с использованием двух комбинаций из 4 вырожденных праймеров. Конечная концентрация праймеров составляла 2 мкМ, концентрация dNTP была 200 мкМ и использовали 2 U/100 мкл реакционного AmpliTag Gold (Perkin Elmer, Rotkreuz, Switzerland). Условия проведения PCR были следующими: 5 циклов при 37°С, затем 30 циклов при 44°С. Смысловой 19mer 5' TYATHCCNGCNATGAARMG 3' и антисмысловой 20mer 5' TTRTANARNGCRAAYTGRTC 3' амплифицировали фрагментом из 221 пар основ, который был субклонирован в Bluescript KS+ (Stratagene, La Jolla, CA) и секвенировали с использованием секвенатора Т7 Sequenase kit (Amersham, Switzerland).
Пример 4: Скрининг библиотеки кДНК λgИ1 с использованием образца GPVI с 221 пар - Фрагмент из 221 пар основ оторвали от плазмиды, очистили и пометили α32Р-АТР (20 МВк/50 мкл, Hartmann Analytik, Braunschweig, Germany) при использовании High Prime Labelling kit (Boehringer Mannheim, Switzerland). Скрининг библиотеки человеческого костного мозга производили в соответствии с инструкциями производителя установки. Выращивали положительные фаги, выделяли их ДНК и субклонировали в BlueScript с использованием сайтов либо EcoRI, либо Sal I, и затем - секвенировали. Секвенирование производили с использованием системы ABS RACE - Platelet, поли- А РНК получали по описанной ранее методике (Power и др., Cytokine 7, 479-482, 1995). Обратную транскрипцию (30 мкл) осуществляли при использовании 5 мкг поли- А РНК с праймером 5'TGAATGAGACGGTCAGTTCAGC 3' (20 мкм), dNTP (1 мМ), PHKsin (40 U), буфером 1Х AMV и 20 U обратной транскриптазы AMV в течение 20 мин при 45°С, затем в течение 20 мин при 52°С. Далее реакционную смесь обработали при помощи 2 мкл 6N NaOH при 65°С в течение 30 мин, нейтрализовали с использованием 2 мкл 6N уксусной кислоты и сконцентрировали в Centricon 30 (Amicon). Якорь был привязан к первой нити ДНК по системе Aptes и Siebert (BioTechniques 15: 890-893, 1993). Внутригенную PCR провели с использованием праймера, дополнительного к якорю, и праймера 5'TTGTACAGAGCAAATTGGTC 3' (35 циклов, 55°С), затем с праймером 5'GACCAGAGGCTTCCGTTCTG 3' (30 циклов при 53°С). Самая высокая полоса (350 пар основ) была отделена электрофорезом в агарозе от более низких, далее ее субклонировали в BlueScript, затем секвенировали.
Пример 5: Получение анти-GPVI Fab и F(ab')2 - поликлональную антисыворотку против человеческого GPVI вырабатывали в кроликах. IgG из кроличьей антисыворотки анти-GPVI очистили, как описано ранее. Для получения фрагментов Fab произвели расщепление IgG при помощи иммобилизованного папаина (Pierce) в соответствии со стандартной методикой поставщика. Фрагменты Fab отделили от нерасщепленных IgG и фрагментов Fc с использованием колонки с иммобилизованным Протеином Protein A (Sigma). Проточную ячейку преобразовали в диализную трубку, произвели концентрирование при помощи твердого полиэтиленгликоля 20000 и провели диализ до конца на фоне 20 мМ Hepes, 140 мМ NaCl, 4 мМ KCI, pH 7,4 и сохраняли продукт при 4°С до его использования. Фрагменты F(ab')2 получали при помощи расщепления пепсином из IgG при соотношении фермента и основы 1:50 (вес/вес), в 0,5 М ацетатном буфере, pH 4,0, при 37°С в течение 18 часов. Далее довели pH до 7,4 при помощи разбавленной NaOH и подвергли образец диализу на фоне 20 мМ фосфата, pH 7,4. Фрагменты F(ab')2 отделили от нерасщепленных IgG и фрагментов Fc при помощи хроматографии с Протеином А. Проточную ячейку преобразовали в диализную трубку, произвели концентрирование при помощи твердого полиэтиленгликоля 20000 и осуществляли интенсивный диализ на фоне 20 мМ Hepes, 140 мМ NaCl, 4 мМ KCI, pH 7,4; полученный продукт сохраняли в аликвотных количествах при -20°С. Промытые тромбоциты растворили в Triton X-114 и произвели разделение фаз на растворимом материале с последующим выделением мембранных гликопротеинов, ассоциированных с фазой Triton X-114, методом аффинной хроматографии на агглютинин-сефарозе зародышей пшеницы 4В по описанной ранее методике. Поскольку GPVI представляет весьма малую фракцию отстоя гликопротеинов мембраны тромбоцитов, для выделения этого рецептора использовали специфичность змеиного лектина С-типа конвулксина. Аффинная хроматография на конвулксине, связанном с сефарозой 4В, позволила получить в качестве основного продукта протеин 65 кДа. При этом совместно с GPVI были элюированы неисследованные материалы с более высокими или более низкими Mr, которые нельзя было удалить при помощи тщательной промывки колонки. Поэтому в качестве завершающего этапа очистки был произведен препаративный гель-электрофорез на 8,5% полиакриламиде. После отстоя фракции, содержащие GPVI, обнаружили при повторном анализе единственную полосу. Очищенный GPVI подвергли испытаниям на предмет его способности блокировать агрегацию тромбоцитов коллагеном. При добавлении к суспензии тромбоцитов аликвотных растворов GPVI наблюдали слабый ингибирующий эффект. Однако после предварительного инкубирования GPVI с коллагеном последующее добавление его к суспензии тромбоцитов оказывало ингибирующий эффект, степень которого зависела от вводимой дозы GPVI. Агрегация тромбоцитов возобновлялась после введения новой порции коллагена. При невосстанавливающихся условиях выделенный протеин имел Mr 62 кДа со сдвигом в сторону несколько более высоких Мг (65 кДа) при восстановительных условиях. Поскольку было обнаружено, что аминные концы GPVI блокированы, произвели расщепление протеина ферментами LysC и LysC/AspN, в результате было получена 4 и 6 пептидов соответственно, из которых была получена последовательность. Разделение пептидов осуществили при помощи HPLC с обращенными фазами на колонке С4, затем секвенировали по методу Edman. Аминокислотные последовательности подчеркнуты на фиг.1, где представлена транслированная последовательность кДНК.
Фрагмент ДНК, кодирующий первые 187 аминокислот зрелого рецептора GPVI (3 и 4 экзоны), амплифицировали с помощью ПЦР посредством считывания информации полимеразой с кДНК GPVI полной длины с использованием праймеров 5'CTTGGATCCGATTGAGGGACGCCAGAGTGGACCGCTCCC 3'- (смысловой) и
5'TGCGGACCTTAGGTTCCTGTGACCACAAG 3 (антисмысловой). [ДНК фрагмент 687 п.о.]. Последовательность ДНК для сайта расщепления фактором Ха ввели в смысловой праймер для удаления 6 х гистидинового - tag, кодируемого вектором. ПЦР фрагмент ввели в плазмиду pQE31 (Quiagen), полученной конструкцией трансформировали Е.coli M 15 [pREP4]. Для экспрессии трансформированные E.coli выращивали в виде серий в 5-ти литровом ферментере 0,9D600 после введения 0,2% лактозы. После 4 часов E.coli собирали, выделяли тела включения и медленно растворяли в Трис/фосфатном буфере, содержащем 6М гуанидин/HCl (рН 8,0) и супернатант помещали на Ni-NTA колонку (Quiangen). Колонку промывали сразу после элюирования 6 х His-tag протеином в Трис/фосфатном буфере, содержащем 6 М гуанидиниум/HCl при 5,1. Анализы элюированного пика с помощью гель-электрофореза показал основную полосу при 26 кДа и несколько отдельных пиков со значительно меньшей молекулярной массой. Рекомбинантный фрагмент GPVI инкубировали с DTT для снижения количества дисульфидных связей, образующихся в Е.coli. Восстановление складчатости инициировали посредством постепенного добавления буфера для восстановления складчатости (40 мМ NaHPO4, рН 8,0,150 мМ NaCl, 10% глицерин, 0,01% CHAPS и восстановленный глутатион GSH/окисленный глутатион GSSG) с объединением элюированных из колонки Ni-NTA фракций. После восстановления складчатости и диализов рекомбинантный фрагмент GPVI очищали посредством хроматографии на сефарозе Q. Гистидин-tag расщепляли фактором Ха. Очищенный рекомбинантный фрагмент GPVI мигрировал на SDS-PAGE геле при 26 кДа при восстанавливающих условиях и при 24 кДа при невосстанавливающих условиях и обладал чистотой >97%. Прединкубация рекомбинантного протеина GPVI с коллагеном блокирует агрегацию тромбоцитов в зависимости от дозы (IC50 мкг/мл).
Фиг.3. Рекомбинантный фрагмент GPVI, полученный из различных серий E.coli, выделяют с помощью гель-электрофореза SDS-PAGE на 15% полиакриаламиде при восстанавливающих условиях (линии 2, 3, 4) или невосстанавливающих условиях (линии 5 и 6). Четкий сдвиг в миграции при восстановлении показывает правильную складчатость и формирование дисульфидных мостиков. Для тестирования способности рекомбинантного GPVI блокировать коллагеновую активацию тромбоцитов образцы очищали и концентрировали, 4 мкг рекомбинантного GPVI заполняли в случае 1 полосы.
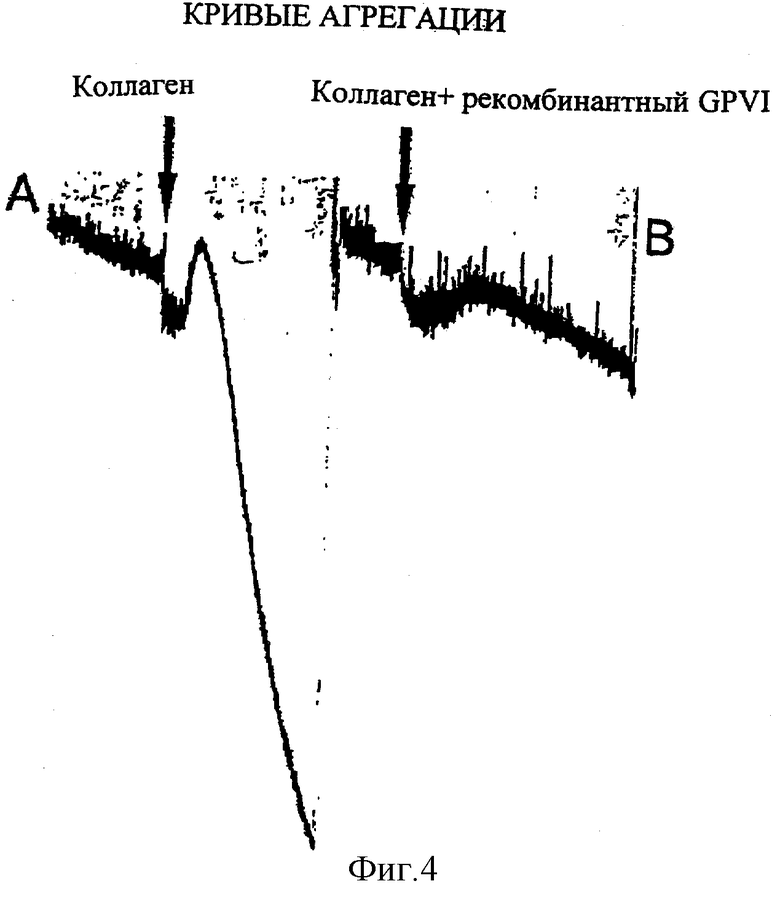
Фиг.4. Агрегацию тромбоцитов ингибировали посредством инкубации коллагена с фрагментом рекомбинантного GPVI до агрегации. Агрегацию осуществляли в агрегометре (Labinhtec, Montpellier, Франция) с растворенными тромбоцитами (5×108 тромбоцитов/мл) рессупендировали в 10 мМ Hepes, 150 мМ NaCl, 5 мМ глюкозного буфера, рН 7,4, в присутствии 2 мМ Ca2+ и 2 мМ Mg2+. В контрольном эксперименте 1 мкм/мл коллагена добавляют в перемешанную суспензию тромбоцитов с индуцированием быстрой агрегации тромбоцитов. В. Агрегация тромбоцитов полностью ингибируется, когда 1 мг/мл коллагена прединкубируют с 5 мг/мл рекомбинатного GPVI.







Изобретение относится к области генной инженерии и биотехнологии и может быть использовано в медицине. Получена ДНК, кодирующая гликопротеин VI (GPVI). На ее основе с помощью методов рекомбинантных ДНК синтезирован рекомбинантный GPVI, который затем использован для создания фармацевтических композиций, обладающих способностью ингибировать или блокировать взаимодействие тромбоцитов и коллагена. Применение изобретения обеспечивает получение GPVI в рекомбинантной форме и применение данного продукта для терапевтического назначения. 5 н. и 1 з.п. ф-лы, 4 ил.
| JANDROT-PERRUS MARTINE ET AL, J | |||
| Biol | |||
| Chem., 1997, vol.272, no.43, стр.27035-27041 | |||
| АБСОРБЕНТ ДЛЯ ОЧИСТКИ ГАЗОВ ОТ HS И CO | 1997 |
|
RU2143942C1 |

