Перекрестная ссылка на родственные заявки
По настоящей заявке испрашивается приоритет предварительной заявки США № 61/183847, поданной 3 июня 2009 года, и предварительной заявки США № 61/300951, поданной 3 февраля 2010 года, содержание которых, таким образом, в полном объеме включено в качестве ссылки.
Область техники
В основном настоящее изобретение относится к антитромбоцитарной фармакологической системе, содержащей лиганд, представляющий собой нуклеиновую кислоту, который связывается с тромбоцит-специфическим белком гликопротеином VI (GPVI) и регулирует его активность. Также эти лиганды, представляющие собой нуклеиновые кислоты, являются активно обратимыми с применением модулятора, ингибирующего активность лиганда, представляющего собой нуклеиновую кислоту, с нейтрализацией его фармакологического действия и, таким образом, восстановления функционирования GPVI. Изобретение дополнительно относится к композициям, содержащим лиганд, представляющий собой нуклеиновую кислоту, и/или модулятор, а также к способам использования этих средств и композиций при лечении опосредованных тромбоцитами заболеваний и нарушений.
Предшествующий уровень техники
Тромбоциты представляют собой небольшие, безъядерные клетки крови, которые фактически неактивны в нормальных условиях, но которые сразу же отвечают на повреждение сосуда посредством адгезии, активации, агрегации и формирования тромба. Первичной функцией тромбоцитов является остановка потери крови после травмы ткани и экспонирования субэндотелиального матрикса. Хорошо известно, что повреждение кровеносного сосуда может подвергать воздействию крови компоненты внеклеточного матрикса, в частности фактор фон Виллебранда (VWF), коллаген, фибронектин, тромбоспондин и ламинин. Взаимодействие тромбоцитов с этими экспонированными молекулы приводит к активации тромбоцитов.
Хотя длительное время признавалась доминирующая роль тромбоцитов в гемостазе и тромбозе, становится все более очевидным, что тромбоциты также могут играть значимую роль в ряде других нарушений, таких как воспаление, рост и метастазирование опухолей и иммунитет. Таким образом, белки рецепторов тромбоцитов являются привлекательными мишенями для регуляции функции тромбоцитов в качестве способов лечения или предотвращения опосредованных тромбоцитами нарушений.
Гликопротеин VI (GPVI) является особенно предпочтительной мишенью, так как он представляют собой трансмембранный белок, специфически экспрессируемый на тромбоцитах. Клеточно-специфичные молекулы с внеклеточной экспозицией обеспечивают доступность для терапевтической молекулы и значительную вероятность минимальных, если вообще каких-либо, побочных эффектов вследствие ограниченного профиля экспрессии.
Гликопротеин VI (GPVI) представляет собой основной сигнальный рецептор тромбоцитов для коллагена, для которого показано, что он играет роль в образовании внутриклеточных сигналов, опосредующих активацию тромбоцитов (Watson et al., Platelets, 2000, 11:252-258). GPVI представляет собой гликопротеин массой 62-65 кДа из семейства Ig, состоящий из двух петель C2 Ig, содержащих коллагенсвязывающий домен, представляющих собой потенциальные участки-мишени для лекарственных средств. Белок in vivo является очень высоко гликозилированным и сиалированным, и один участок гликозилирования находится на внешнем Ig-C2-подобном домене. Как представитель иммуноглобулинового суперсемейства, GPVI является родственным с рецепторами естественных киллеров. Его сигнал может передаваться опосредованно через γ-цепь FcR или прямо через цитоплазматический домен GPVI. γ-цепь FcR содержит иммунорецепторный активационный мотив на основе тирозина, и в сочетании с нетирозиновой киназой SYK и адаптерами LAT и LCP2 он приводит к активации фосфолипазы C2 и ассоциированных внутриклеточных путей передачи сигнала (Lankhof et al., Thromb. Haemost., 1996, 75:950-958).
Важность GPVI в физиологии и в клинических проявлениях, ассоциированных с формированием сгустков, является общепризнанной, и она подкрепляется эпидемиологическими ассоциациями уровней GPVI с возникновением острого коронарного синдрома (ACS) и случаями инсульта (Bigalke et al., American Heart Journal, 2008, 156:193-200; Bigalke et al., European Journal of Neurology, 2009, 101:911-915) и устойчивостью к тромбозу, продемонстрированной у мышей с дефицитом GPVI (Denis et al., Arteriosclerosis, Thrombosis and Vascular Biology, 2007, 27:728-739). Кроме того, Gawaz et al показали, что радиоактивно меченный GPVI можно использовать в сцинтиллографическом представлении повреждений сосудов у мышей, так как он специфически связывается с поврежденной областью, указывая на то, что в этих участках на поверхности находится коллаген (Thromb. Haemost., 2005, 93:910-913). Penz et al., (FASEB J. 2005, 19:898-909) показали, что атеросклеротические бляшки у человека, полученные у пациентов со стенозом сонных артерий, содержали структуры коллагена типа I и типа III, способные активировать тромбоциты. Кроме того, блокада или отсутствие GPVI могла предотвращать формирование тромба, тогда как блокада α2β1 оказывала небольшое действие. Подобным образом, блокирование коллагена антителами к коллагену или его разрушение коллагеназой предотвращало формирование тромба.
Недавно GPVI связали с более значимой ролью в нарушениях, ассоциированных с нарушениями функционирования тромбоцитов и аномальной экспрессией коллагена. Это частично происходит вследствие того факта, что тромбоциты действуют при адгезии после повреждения сосудов, как в случае тромбоза, и при высвобождении ряда воспалительных медиаторов и цитокинов. Кроме того, после реакции тромбоцитов с участком повреждения сосуда последующая активация тромбоцитов приводит к высвобождению цитокинов и других регуляторных молекул. Таким образом, хотя может показаться, что нарушения, ассоциированные с аномальной активацией тромбоцитов, являются различными, эти нарушения связаны с зависимостью от функции тромбоцитов. Таким образом, ингибирование или предотвращение активации тромбоцитов может обеспечивать значимые терапевтические эффекты.
Опосредованные тромбоцитами нарушения включают сосудистые заболевания, а также ряд нарушений, ассоциированных с высоким риском диабета. Воспалительные нарушения, для которых показано, что они опосредованы тромбоцитами, включают воспалительные артриты и склеродермию. При воспалительных заболеваниях суставов давно известна роль воспаления и лейкоцитарной активности. Недавно в синовиальной жидкости воспаленных суставов выявлено присутствие тромбоцитов (Boilard et al., Science, 2010, 327:580-583). Кроме того, показано, что тромбоцитарные микрочастицы в суставной жидкости пациентов, страдающих воспалительным артритом, являются провоспалительными, вызывая ответ цитокинами у синовиальных фибробластов посредством IL-1. И фармакологический, и генетический подходы продемонстрировали, что GPVI играет ключевую роль в этой провоспалительной природе тромбоцитов при артрите.
Другие нарушения, для которых показано, что они ассоциированы с экспрессией GPVI на поверхности тромбоцитов, включают экспериментальное метастазирование опухолей (Jain et al., J. Thromb. Haemostasis, 2009, 7:1713-1717), диабет (Cabeza et al., 2004, 53:2117-2121) и инфекцию вирусом гепатита C (Zahn et al., Diabetes, 2004, 53:2117-2121).
Несмотря на продемонстрированное расширение роли GPVI, которую он играет в функционировании тромбоцитов и связанных физиологических нарушениях, попытки поиска и разработки антагонистов этого рецептора ограничены. Активный контроль или модуляция интенсивности и длительности антитромбоцитарной терапии могут обеспечить значительное клиническое преимущество. Таким образом, в данной области остается необходимость в регулируемых средствах, предназначенных для специфической направленности на белок GPVI и регуляции его функции.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
В настоящем документе описаны композиции, относящиеся к лигандам, представляющим собой нуклеиновые кислоты, которые специфически связывают гликопротеин VI (GPVI), способы применения и подходы к применению лигандов, представляющих собой нуклеиновые кислоты, специфически связывающие гликопротеин VI (GPVI), и их модуляторы.
В одном из аспектов предоставлен лиганд GPVI, где лиганд содержит выделенную последовательность нуклеиновой кислоты. В другом варианте осуществления по меньшей мере один нуклеотид представляет собой рибонуклеотид. В другом варианте осуществления по меньшей мере один нуклеотид представляет собой дезоксирибонуклеиновую кислоту. В еще одном варианте осуществления выделенная последовательность нуклеиновой кислоты лиганда GPVI содержит комбинацию рибонуклеотидов и дезоксирибонуклеотидов.
В одном из вариантов осуществления лиганд GPVI, представляющий собой нуклеиновую кислоту, содержит вторичную структуру, содержащую один, два или три стебля и одну, две, три или четыре петли.
В одном из вариантов осуществления лиганд GPVI содержит вторичную структуру, где вторичная структура содержит, в направлении от 5' к 3', область первого стебля, область первой петли, область второго стебля, область второй петли, область третьей петли, область третьего стебля и область четвертой петли. В другом варианте осуществления лиганд GPVI по существу состоит, в направлении от 5' к 3', из области первого стебля, области первой петли, области второго стебля, области второй петли, области третьей петли, области третьего стебля и области четвертой петли.
В одном из вариантов осуществления вторичная структура лиганда GPVI, представляющего собой нуклеиновую кислоту, сконструирована в направлении от 5' к 3' в виде первого стебля, первой петли, второго стебля, соединенного со второй петлей и с третьей петлей, третьего стебля, соединенного с третьей петлей и с четвертой петлей, и четвертой петли. В одном из вариантов осуществления второй стебель соединен с первой, второй и третьей петлями.
В одном из вариантов осуществления четвертая петля лиганда GPVI, представляющего собой нуклеиновую кислоту, содержит первую консенсусную последовательность, состоящую из UAA. В другом варианте осуществления четвертая петля содержит последовательность, состоящую из (G/A)UAA.
В одном из вариантов осуществления третий стебель содержит последовательность, состоящую из пары нуклеотидов G-C с последующей парой нуклеотидов C-G.
В одном из вариантов осуществления лиганд GPVI, представляющий собой нуклеиновую кислоту, содержит GC(G/A)UAAGC. В другом варианте осуществления третий стебель и четвертая петля лиганда, представляющего собой нуклеиновую кислоту, содержат GC(G/A)UAAGC. В еще одном варианте осуществления третий стебель и четвертая петля лиганда, представляющего собой нуклеиновую кислоту, содержат последовательность, выбранную из группы, состоящей из GCAUAAGC и GCGUAAGC.
В одном из вариантов осуществления третья петля содержит последовательность YD, где Y представляет собой пиримидин, а D не является цитозином. В другом варианте осуществления третья петля содержит последовательность YU, YG или YA. В еще одном варианте осуществления третья петля содержит последовательность UU, UG, UA, CU, CG или CA. В другом варианте осуществления третья петля состоит из последовательности YD.
В одном из вариантов осуществления первая петля содержит последовательность GAC.
В одном из вариантов осуществления длина выделенной последовательности лиганда GPVI, представляющего собой нуклеиновую кислоту, составляет приблизительно от 20 нуклеотидов (н.) до приблизительно 50 н., приблизительно от 20 н. до приблизительно 45 н., приблизительно от 20 н. до приблизительно 40 н., приблизительно от 20 н. до приблизительно 35 н., приблизительно от 20 н. до приблизительно 30 н. или приблизительно от 30 н. до приблизительно 35 н.
В одном из вариантов осуществления лиганд GPVI содержит выделенную последовательность нуклеиновой кислоты, выбранную из группы, состоящей из SEQ ID NO:7, SEQ ID NO:9, SEQ ID NO:11, SEQ ID NO:12, SEQ ID NO:14 и SEQ ID NO:15. В другом варианте осуществления лиганд GPVI содержит выделенную последовательность нуклеиновой кислоты, выбранную из группы, состоящей из последовательностей от SEQ ID NO:28 до SEQ ID NO:33, включительно. В еще одном варианте осуществления лиганд GPVI содержит выделенную последовательность нуклеиновой кислоты РНК, выбранную из группы, состоящей из РНК EF-1, РНК EF-2, РНК EF-3, РНК E2-6, РНК EF-22 и РНК EF-31. В другом варианте осуществления лиганд GPVI содержит выделенную последовательность нуклеиновой кислоты РНК, выбранную из группы, состоящей из модифицированной EF-1, модифицированной EF-2, модифицированной EF-3, модифицированной E2-6, модифицированной EF-22 и модифицированной EF-31. В другом варианте осуществления лиганд GPVI содержит выделенную последовательность нуклеиновой кислоты, которая по меньшей мере на 80% идентична последовательности, выбранной из группы, состоящей из последовательностей от SEQ ID NO:28 до SEQ ID NO:33, включительно.
В одном из вариантов осуществления лиганд GPVI выбран из группы, состоящей из RB424, RB426, RB427, RB428, RB429, RB430, RB445, RB446, RB447, RB431, RB432, RB433, RB434, RB435, RB436, RB439, RB440, RB441, RB442, RB443 и RB444.
В одном из вариантов осуществления лиганд GPVI выбран из группы, состоящей из RB448, RB452, RB453, RB455, RB460, RB462, RB466, RB478, RB480, RB488, EB490, RB491, RB492, RB493, RB495, RB496, RB497, RB498, RB499, RB500, RB502, RB503, RB504, RB505, RB506, RB507, RB508, RB517, RB518, RB519, RB520, RB521, RB522, RB523, RB524, RB525, RB526, RB527, RB528, RB531, RB532, RB533, RB534, RB535, RB536, RB537, RB538, RB540, RB541, RB542, RB546, RB547, RB548, RB549, RB550, RB551, RB552, RB553, RB554, RB555, RB556, RB560, RB561, RB566, RB567, RB569, RB570, RB571.
В одном из вариантов осуществления выделенная последовательность нуклеиновой кислоты лиганда GPVI содержит один или несколько рибонуклеотидов, дезоксирибонуклеотидов, или комбинацию рибонуклеотидов и дезоксирибонуклеотидов.
В одном из вариантов осуществления один или несколько нуклеотидов выделенной последовательности лиганда GPVI, представляющего собой нуклеиновую кислоту, являются модифицированными. В другом варианте осуществления один или несколько нуклеотидов содержат модификацию в положении 2'-гидроксила. В другом варианте осуществления модификация выбрана из группы, состоящей из 2'-O-метила и 2'-фтора. В еще одном варианте осуществления один или несколько нуклеотидов представляют собой 2'-O-метилцитозин, 2'-O-метилуридин, 2'-O-метиладенозин или 2'-O-метилгуанозин. В еще одном варианте осуществления один или несколько нуклеотидов представляют собой 2'-фторцитидин или 2'-фторуридин.
В одном из вариантов осуществления один или несколько нуклеотидов, содержащих модификацию, выбраны из группы, состоящей из 5-фторурацила, 5-фторцитозина, 5-бромурацила, 5-бромцитозина, 5-хлорурацила, 5-хлорцитозина, 5-йодурацила, 5-йодцитозина, 5-метилцитозина, 5-метилурацила, гипоксантина, ксантина, 4-ацетилцитозина, 5-(карбоксигидроксилметил)урацила, 5-карбоксиметиламин-O-метилтиоуридина, 5-карбоксиметиламин-O-метилурацила, дигидроурацила, бета-D-галактозилквеозина, инозина, N6-изопентениладенина, 1-метилгуанина, 1-метилинозина, 2,2-диметилгуанина, 2-метиладенина, 2-метилгуанина, 3-метилцитозина, 6-метилцитозина, N6-аденина, 7-метилгуанина, 5-метиламин-O-метилурацила, 5-метоксиамин-O-метил-2-тиоурацила, бета-D-маннозилквеозина, 5'-метоксикарбоксиметилурацила, 5-метоксиурацила, 5-метоксицитозина, 2-метилтио-N6-изопентениладенина, урацилоксиуксусной кислоты (v), бутоксозина, псевдоурацила, квеозина, 2-тиоцитозина, 5-метилтиоурацила, 2-тиоурацила, 4-тиоурацила, 5-метилурацила, метилового эфира урацил-5-оксиуксусной кислоты, урацилоксиуксусной кислоты (v), 5-метилтиоурацила, 3-(3-амино-3-N-карбоксипропил)уридина, (acp3U) и 2,6-диаминопурина.
В одном из вариантов осуществления лиганд GPVI содержит по меньшей мере одну модифицированную молекулу сахара.
В одном из вариантов осуществления лиганд GPVI содержит по меньшей мере один модифицированный фосфатный остов.
В одном из вариантов осуществления выделенная последовательность лиганда GPVI, представляющего собой нуклеиновую кислоту, на своем 3'-конце содержит инвертированный тимин.
В одном из вариантов осуществления лиганд GPVI, представляющий собой нуклеиновую кислоту, содержит спейсер. В другом варианте осуществления спейсер представляет собой гликолевый спейсер. В другом варианте осуществления вторая петля лиганда GPVI, представляющего собой нуклеиновую кислоту, содержит гликолевый спейсер. В еще одном варианте осуществления вторая петля лиганда GPVI, представляющего собой нуклеиновую кислоту, состоит из гликолевого спейсера. В еще одном варианте осуществления гликолевый спейсер обеспечивают посредством введения 9-O-диметокситритилтриэтиленгликоль,1-[(2-цианоэтил)-(N,N-диизопропил)]фосфорамидита. В еще одном варианте осуществления гликолевый спейсер присоединяют к 3'-концу первого внутреннего нуклеотида выделенной последовательности нуклеотидного лиганда GPVI и присоединяют к 5'-концу второго внутреннего нуклеотида, расположенного рядом с первым внутренним нуклеотидом выделенной последовательности нуклеотидного лиганда GPVI.
В одном из вариантов осуществления лиганд GPVI, представляющий собой нуклеиновую кислоту, содержит алифатический аминолинкер. В другом варианте осуществления алифатический аминолинкер присоединен к 5'-концу выделенной последовательности лиганда GPVI, представляющего собой нуклеиновую кислоту. В еще одном варианте осуществления алифатический аминолинкер присоединен к 3'-концу выделенной последовательности лиганда GPVI, представляющего собой нуклеиновую кислоту. В еще одном варианте осуществления алифатический аминолинкер обеспечивают посредством введения (2-цианоэтил-N,N-диизопропил)фосфорамидита 6-(трифторацетамино)гексанола.
В одном из вариантов осуществления выделенный лиганд GPVI, представляющий собой нуклеиновую кислоту, связаную по меньшей мере с одной гидрофильной молекулой. В другом варианте осуществления по меньшей мере одна гидрофильная молекула представляет собой полиалкиленгликоль.
В одном из вариантов осуществления лиганд GPVI содержит полиалкиленовую молекулу, присоединенную к 5'-концу и/или к 3'-концу выделенной последовательности нуклеиновой кислоты. В другом варианте осуществления полиалкиленовая молекула присоединена посредством линкера. В еще одном варианте осуществления линкер представляет собой алифатический аминолинкер.
В одном из вариантов осуществления лиганд GPVI связан с молекулой полиэтиленгликоля (PEG) массой 40 кДа с использованием аминолинкера с шестью атомами углерода. В другом варианте осуществления аминолинкер с шестью атомами углерода присоединен к молекуле PEG посредством амидной связи. В еще одном варианте осуществления, молекула PEG представляет собой две молекулы PEG массой двенадцать кДа, присоединенные к одной или нескольким аминокислотам, таким как лизин, которые присоединены посредством амидной связи к аминолинкеру с шестью атомами углерода.
В одном из вариантов осуществления первый лиганд GPVI, представляющий собой нуклеиновую кислоту, содержит тиофосфатную связь.
В одном из вариантов осуществления лиганд GPVI, представляющий собой нуклеиновую кислоту, специфически связывается с GPVI (SEQ ID NO:1). В другом варианте осуществления лиганд GPVI, представляющий собой нуклеиновую кислоту, специфически связывается с внеклеточным доменом GPVI (SEQ ID NO:2).
В одном из вариантов осуществления константа диссоциации лиганда GPVI составляет приблизительно 20 наномоль/л (нМ) или менее.
В одном из вариантов осуществления константа диссоциации лиганда GPVI находится в диапазоне приблизительно от 400 пикомоль/л (пМ) до приблизительно 10 нМ.
В одном из вариантов осуществления константа диссоциации лиганда GPVI находится в диапазоне приблизительно от 100 пМ до приблизительно 10 нМ.
В одном из вариантов осуществления лиганд GPVI, представляющий собой нуклеиновую кислоту, ингибирует связывание GPVI с коллагеном. В другом варианте осуществления лиганд GPVI, представляющий собой нуклеиновую кислоту, ингибирует внутриклеточную передачу сигнала через GPVI. В другом варианте осуществления ингибирование внутриклеточной передачи сигнала через GPVI с использованием лиганда GPVI включает уменьшение образования инозитолтрифосфата или ингибирование колебаний внутриклеточных уровней кальция.
В одном из вариантов осуществления лиганд GPVI, представляющий собой нуклеиновую кислоту, ингибирует связывание GPVI с родственным коллагену пептидом (CRP) и/или с конвулксином.
В одном из вариантов осуществления лиганд GPVI, представляющий собой нуклеиновую кислоту, ингибирует связывание GPVI и с коллагеном, и с CRP.
В одном из вариантов осуществления лиганд GPVI, представляющий собой нуклеиновую кислоту, ингибирует связывание GPVI с коллагеном, но не ингибирует связывание GPVI с CRP. В другом варианте осуществления лиганд GPVI, представляющий собой нуклеиновую кислоту, ингибирует связывание GPVI и с коллагеном, и CRP, но не ингибирует связывание GPVI с конвулксином.
В одном из вариантов осуществления связывание лиганда GPVI, представляющего собой нуклеиновую кислоту, с GPVI стабилизирует активную конформацию GPVI. В другом варианте осуществления связывание лиганда GPVI, представляющего собой нуклеиновую кислоту, с GPVI стабилизирует неактивную конформацию GPVI. В еще одном варианте осуществления связывание лиганда GPVI, представляющего собой нуклеиновую кислоту, с GPVI ингибирует взаимодействие между GPVI и субъединицей γ FcR.
В другом варианте осуществления связывание лиганда GPVI с GPVI приводит к ингибированию или снижению активности GPVI. В еще одном варианте осуществления связывание лиганда GPVI с GPVI приводит к неспособности или к снижению способности GPVI взаимодействовать с γ-цепью FcR. В еще одном варианте осуществления связывание лиганда GPVI с GPVI, экспрессированном на поверхности тромбоцита, приводит к ингибированию или снижению адгезии тромбоцита. В еще одном варианте осуществления связывание лиганда GPVI с GPVI, экспрессированным на поверхности тромбоцита, приводит к ингибированию или снижению активации тромбоцитов. В еще одном варианте осуществления связывание лиганда GPVI с GPVI, экспрессированном на поверхности тромбоцита, приводит к ингибированию или снижению агрегации тромбоцитов.
В одном из вариантов осуществления лиганд GPVI связывается с вариантом GPVI и снижает или ингибирует его функцию, где указанный вариант GPVI по меньшей мере на 80%, 85%, 90%, 91%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичен SEQ ID NO:1.
В одном из вариантов осуществления константа диссоциации ("Kd") лиганда GPVI для GPVI составляет менее чем приблизительно 100 микромоль/л (мкМ), менее чем приблизительно 1 мкМ, менее чем приблизительно 500 наномоль/л (нМ), менее чем приблизительно 100 нМ, менее чем приблизительно 50 нМ, менее чем приблизительно 1 нМ, менее чем приблизительно 500 пикомоль/л (пМ), менее чем приблизительно 300 пМ, менее чем приблизительно 250 пМ или менее чем приблизительно 200 приблизительно пМ.
Во втором аспекте предоставлен модулятор для лиганда GPVI, где модулятор частично или полностью обращает активность лиганда GPVI.
В одном из вариантов осуществления модулятор содержит выделенную последовательность нуклеиновой кислоты. В другом варианте осуществления модулятор содержит последовательность ДНК, последовательность РНК, полипептидную последовательность или любое их сочетание.
В одном из вариантов осуществления модулятор лиганда GPVI, представляющего собой нуклеиновую кислоту, выбран из группы, состоящей из рибозима, ДНКзима, пептидной нуклеиновой кислоты (ПНК), морфолинонуклеиновой кислоты (МНК) и замкнутой нуклеиновой кислоты (ЗНК).
В одном из вариантов осуществления модулятор лиганда GPVI, представляющего собой нуклеиновую кислоту, выбран из группы, состоящей из рибозима, ДНКзима, пептидной нуклеиновой кислоты (ПНК), морфолинонуклеиновой кислоты (МНК) и замкнутой нуклеиновой кислоты (ЗНК), где модулятор специфически связывается или взаимодействует по меньшей мере с частью лиганда GPVI, представляющего собой нуклеиновую кислоту.
В одном из вариантов осуществления модулятор выбран из группы, состоящей из связывающих нуклеиновую кислоту белка или пептида, низкомолекулярного соединения, олигосахарида, связывающих нуклеиновую кислоту липида, полимера, наночастицы и микросферы, где модулятор связывается или взаимодействует по меньшей мере с частью лиганда GPVI, представляющего собой нуклеиновую кислоту.
В одном из вариантов осуществления модулятор представляет собой модулятор, представляющий собой нуклеиновую кислоту, содержащий дезоксирибонуклеотиды, рибонуклеотиды или комбинацию дезоксирибонуклеотидов и рибонуклеотидов. В другом варианте осуществления модулятор, представляющий собой нуклеиновую кислоту, содержит по меньшей мере один модифицированный дезоксирибонуклеотид и/или по меньшей мере один модифицированный рибонуклеотид.
В одном из вариантов осуществления модулятор представляет собой олигонуклеотид, комплементарный по меньшей мере части лиганда GPVI, представляющего собой нуклеиновую кислоту. В другом варианте осуществления модулятор представляет собой олигонуклеотид, комплементарный по меньшей мере части петли в лиганде GPVI. В другом варианте осуществления модулятор представляет собой олигонуклеотид, комплементарный по меньшей мере первой петле, второму стеблю и второй петле лиганда GPVI, представляющего собой нуклеиновую кислоту. В другом варианте осуществления модулятор представляет собой олигонуклеотид, комплементарный третьей петле, третьему стеблю, четвертой петле, и первому стеблю лиганда GPVI.
В одном из вариантов осуществления модулятор содержит выделенную последовательность нуклеиновой кислоты, где длина последовательности составляет приблизительно от 10 н. до приблизительно 30 н., приблизительно от 10 н. до приблизительно 25 н., приблизительно от 10 н. до приблизительно 20 н., приблизительно от 10 н. до приблизительно 15 н. или приблизительно от 15 н. до приблизительно 20 н.
В одном из вариантов осуществления один или несколько нуклеотидов последовательности модулятора, представляющего собой нуклеиновую кислоту, являются модифицированными. В другом варианте осуществления один или несколько нуклеотидов содержат модификацию в положении 2'-гидроксила. В другом варианте осуществления модификация выбрана из группы, состоящей из 2'-O-метила и 2'-фтора. В еще одном варианте осуществления один или несколько нуклеотидов представляют собой 2'-O-метилцитозин, 2'-O-метилуридин, 2'-O-метиладенозин, 2'-O-метилгуанозин 2'-O-метилтимидин. В еще одном варианте осуществления один или несколько нуклеотидов представляют собой 2'-фторцитидин, 2'-фторуридин, 2'-фтораденозин или 2'-фторгуанозин.
В одном из вариантов осуществления модификация одного или нескольких нуклеотидов модулятора, представляющего собой нуклеиновую кислоту, включает модификацию, выбранную из группы, состоящей из 5-фторурацила, 5-фторцитозина, 5-бромурацила, 5-бромцитозина, 5-хлорурацила, 5-хлорцитозина, 5-йодурацила, 5-йодцитозина, 5-метилцитозина, 5-метилурацила, гипоксантина, ксантина, 4-ацетилцитозина, 5-(карбоксигидроксилметил)урацила, 5-карбоксиметиламин-O-метилтиоуридина, 5-карбоксиметиламин-O-метилурацила, дигидроурацила, бета-D-галактозилквеозина, инозина, N6-изопентениладенина, 1-метилгуанина, 1-метилинозина, 2,2-диметилгуанина, 2-метиладенина, 2-метилгуанина, 3-метилцитозина, 6-метилцитозина, N6-аденина, 7-метилгуанина, 5-метиламин-O-метилурацила, 5-метоксиамин-O-метил-2-тиоурацила, бета-D-маннозилквеозина, 5'-метоксикарбоксиметилурацила, 5-метоксиурацила, 5-метоксицитозина, 2-метилтио-N6-изопентениладенина, урацилоксиуксусной кислоты (v), бутоксозина, псевдоурацила, квеозина, 2-тиоцитозина, 5-метилтиоурацила, 2-тиоурацила, 4-тиоурацила, 5-метилурацила, метилового эфира урацил-5-оксиуксусной кислоты, урацилоксиуксусной кислоты (v), 5-метилтиоурацила, 3-(3-амино-3-N-карбоксипропил)уридина (acp3U) и 2,6-диаминопурина.
В одном из вариантов осуществления модулятор содержит по меньшей мере одну модифицированную молекулу сахара.
В одном из вариантов осуществления модулятор содержит по меньшей мере один модифицированный фосфатный остов.
В одном из вариантов осуществления модулятор содержит олигонуклеотид, который в физиологических условиях гибридизуется с четвертой петлей лиганда GPVI. В другом варианте осуществления олигонуклеотид модулятор содержит последовательность 3'-AUU-5'.
В одном из вариантов осуществления модулятор содержит олигонуклеотид, который в физиологических условиях гибридизуется с первой петлей лиганда GPVI. В другом варианте осуществления олигонуклеотид модулятор содержит последовательность 3'-CUG-5'.
В одном из вариантов осуществления модулятор содержит последовательность, выбранную из группы, состоящей из SEQ ID NO:74-88, включительно. В другом варианте осуществления модулятор выбран из группы, состоящей из RB416, RB417, RB418, RB419, RB420, RB421, RB422, RB423, RB513, RB514, RB515, RB516, RB543, RB544 и RB545.
В одном из вариантов осуществления модулятор нарушает вторичную структуру лиганда GPVI, представляющего собой нуклеиновую кислоту. В другом варианте осуществления модулятор стабилизирует вторичную структуру лиганда GPVI.
В одном из вариантов осуществления модулятор нарушает третичную структуру лиганда GPVI, представляющего собой нуклеиновую кислоту. В другом варианте осуществления модулятор стабилизирует вторичную структуру лиганда GPVI.
В одном из вариантов осуществления связывание модулятора с лигандом GPVI экспонирует суицидальное положение в лиганде GPVI, таким образом, нарушая вторичную структуру лиганда GPVI и приводя к увеличенному разрушению лиганда GPVI, представляющего собой нуклеиновую кислоту, нуклеазами.
В одном из вариантов осуществления связывание модулятора с комплексом лиганд GPVI-GPVI уменьшает или устраняет связывание лиганда GPVI с GPVI.
В другом аспекте предоставлен способ модуляции активности лиганда GPVI.
В одном из вариантов осуществления предоставлен способ модуляции активности лиганда GPVI, представляющего собой нуклеиновую кислоту, посредством введения модулятора лиганда GPVI индивидууму, которому вводили лиганд GPVI, представляющий собой нуклеиновую кислоту. В одном из вариантов осуществления модулятор может представлять собой олигонуклеотидный модулятор или его производное, а в определенных вариантах осуществления он комплементарен части лиганда GPVI, представляющего собой нуклеиновую кислоту.
В дополнительном аспекте предоставлен способ регуляции функции GPVI с использованием лиганда GPVI.
В одном из вариантов осуществления способ регуляции функции GPVI включает введение индивидууму терапевтически эффективного количества лиганда GPVI. В другом варианте осуществления способ дополнительно включает введение индивидууму модулятора лиганда GPVI.
В другом аспекте предоставлен способ лечения или облегчения опосредованного тромбоцитами заболевания или нарушения.
В одном из вариантов осуществления способ включает введение нуждающемуся в этом индивидууму терапевтически эффективной дозы лиганда GPVI, который связывается с GPVI. В другом варианте осуществления у индивидуума диагностирован высокий риск диабета. В еще одном варианте осуществления у индивидуума диагностирована злокачественная опухоль с высоким риском метастазирования.
В одном из вариантов осуществления опосредованное тромбоцитами заболевание или нарушение выбраны из группы, состоящей из цереброваскулярных нарушений, острых коронарных синдромов, связанных с диабетом нарушений, аутоиммунных воспалительных нарушений и злокачественной опухоли.
В одном из вариантов осуществления цереброваскулярное нарушение представляет собой тромбоз, тромбоэмболию или транзиторную ишемическую атаку (TIA). В другом варианте осуществления острый коронарный синдром возникает вследствие тромбоза коронарных артерий, нестабильной стенокардии или инфаркта миокарда. В еще одном варианте осуществления связанное с диабетом нарушение представляет собой диабетическую ретинопатию, диабетическую васкулопатию, атеросклероз, ишемический инсульт, заболевание периферических сосудов, острое повреждение почек или хроническую почечную недостаточность. В другом варианте осуществления аутоиммунное воспалительное нарушение представляет собой склеродермию, ревматоидный артрит или воспалительное аутоиммунное нарушение, выбранное из группы, состоящей из псориатического артрита, реактивного артрита, воспалительного заболевания кишечника и анкилозирующего спондилита. В одном из вариантов осуществления злокачественная опухоль выбрана из рака легких, рака молочной железы, рака предстательной железы, рака поджелудочной железы, злокачественной опухоли головного мозга, злокачественной опухоли кости и рака печени.
В одном из вариантов осуществления лиганд GPVI вводят посредством парентерального введения, внутривенной инъекции, интрадермальной доставки, внутрисуставной доставки, интрасиновиальной доставки, интратекально, посредством внутриартериальной доставки, внутрисердечной доставки, внутримышечной доставки, подкожной доставки, интраорбитальной доставки, внутрикапсулярной доставки, интраспинальной доставки, интрастернальной доставки, местной доставки, доставки посредством трансдермального пластыря, ректальной доставки, доставки посредством вагинального или уретрального суппозитория, перитонеальной доставки, чрескожной доставки, доставки посредством назального спрея, доставки посредством хирургического имплантата, доставки посредством внутреннего хирургического нанесения, доставки посредством инфузионного насоса или доставки посредством катетера.
В другом аспекте предоставлен способ лечения нуждающегося в этом индивидуума посредством введения лиганда GPVI, где лиганд GPVI регулирует функцию тромбоцитов.
В одном из вариантов осуществления вводят терапевтически эффективную дозу GPVI.
В одном из вариантов осуществления терапевтически эффективная доза снижает или ингибирует адгезию и/или агрегацию тромбоцитов.
В одном из аспектов предоставлена фармацевтическая композиция, содержащая терапевтически эффективное количество лиганда GPVI, представляющего собой нуклеиновую кислоту, связывающегося с GPVI.
В одном из аспектов предоставлена фармацевтическая композиция, содержащая терапевтически эффективное количество модулятора, где модулятор регулирует активность лиганда GPVI, представляющего собой нуклеиновую кислоту, связывающегося с GPVI.
В одном из вариантов осуществления фармацевтическая композиция содержит лиганд GPVI и фармацевтически приемлемые эксципиенты. В другом варианте осуществления фармацевтическая композиция представляет собой жидкость, подходящую для внутривенной инъекции. В еще одном варианте осуществления фармацевтическая композиция представляет собой жидкость или дисперсию, подходящую для подкожной инъекции.
В одном из аспектов предоставлен набор, содержащий терапевтически эффективное количество лиганда GPVI, представляющего собой нуклеиновую кислоту, и/или модулятора, регулирующего активность лиганда GPVI, представляющего собой нуклеиновую кислоту.
Краткое описание рисунков
Фиг. 1 представляет собой диаграмму способа отбора SELEX лиганда, представляющего собой нуклеиновую кислоту.
На фиг. 2 представлены условия отбора для стадий SELEX, проводимого для идентификации лигандов GPVI, представляющих собой нуклеиновые кислоты.
На фиг. 3 представлены кривые связывания библиотек с концевым мечением маркерным 32P, обогащенных лигандами GPVI, представляющими собой нуклеиновые кислоты.
На фиг. 4A-B представлена первичная последовательность, полученная из случайной области, идентифицированная для индивидуальных клонов в серии отбора "E/F" после 10 стадии отбора и для индивидуальных клонов в серии отбора "E2" после 10 стадии отбора.
На фиг. 5 представлена минимальная предсказанная первичная последовательность, необходимая для связывания GPVI и предсказанная консервативная вторичная структура для последовательностей, идентифицированных в ходе отбора. "L" означает область петли, "S" означает область стебля.
Фиг. 6 представляет собой диаграмму связывания лиганда GPVI EF-3 с GPVI, с отжигом 3'-праймера с EF-3 для маскировки 3'-фиксированной области EF-3 при взаимодействии с белком или без отжига.
Фиг. 7 представляет собой диаграмму с кривыми связывания укороченных лигандов с GPVI в сравнении с исходными лигандами GPVI EF-2 и EF-3.
На фиг. 8 представлена последовательность и предсказанная вторичная структура нескольких укорочений укороченных вариантов лигандов GPVI EF-3 и EF-31.
На фиг. 9 представлена предсказанная вторичная структура укороченных последовательностей лигандов GPVI EF-3 T2 и EF-2 T2 и расположение инактивирующих точковых мутаций.
На фиг. 10 представлены результаты анализов связывания GPVI с вариантами лигандов GPVI, представляющих собой нуклеиновую кислоту.
На фиг. 11 представлены предсказанная вторичная структура и замены сахаров различных лигандов GPVI, представляющих собой нуклеиновые кислоты.
На фиг. 12 представлены результаты анализов связывания GPVI с вариантами лигандов GPVI, представляющих собой нуклеиновую кислоту.
На фиг. 13A-B представлены гексаэтиленгликолевый спейсерный фосфорамидит и спейсерный фосфорамидит, встроенный между двумя нуклеотидами в последовательности нуклеиновой кислоты.
На фиг. 14A-B представлены молекулы PEG, которые можно конъюгировать с лигандом GPVI посредством линкера, и конфигурация конъюгированных молекул.
Фиг. 15 представляет собой диаграмму индуцированной коллагеном агрегации тромбоцитов, выраженной в виде процента от контроля, для укороченных вариантов лигандов GPVI EF-2 и EF-3 по сравнению с вариантами, содержащими инактивирующие точечные мутации.
Фиг. 16 представляет собой диаграмму индуцированной коллагеном агрегации тромбоцитов, выраженной в виде процента от контроля, для различных концентраций выбранных лигандов GPVI, представляющих собой нуклеиновые кислоты.
На фиг. 17 представлены диаграммы индуцированной CRP агрегации тромбоцитов, выраженной в виде процента от контроля, для различных концентраций выбранных лигандов GPVI, представляющих собой нуклеиновые кислоты.
На фиг. 18 представлены диаграммы индуцированной коллагеном агрегации тромбоцитов, выраженной в виде процента от контроля, для различных концентраций выбранных лигандов GPVI, представляющих собой нуклеиновые кислоты.
На фиг. 19 представлены диаграммы индуцированной CRP агрегации тромбоцитов, выраженной в виде процента от контроля, для различных концентраций выбранных лигандов GPVI, представляющих собой нуклеиновые кислоты.
На фиг. 20 представлены диаграммы индуцированной коллагеном и CRP агрегации тромбоцитов, выраженной в виде процента от контроля, при различных концентрациях лиганда GPVI RB571 в обогащенной тромбоцитами плазме.
Фиг. 21 представляет собой диаграмму, демонстрирующую эффекты лигандов GPVI, представляющих собой нуклеиновые кислоты, на накопление тромбоцитов на покрытой коллагеном поверхности, подвергающейся воздействию текущей цельной крови, выраженное в виде % максимального ответа неактивного контрольного лиганда.
На фиг. 22 представлены диаграммы, на которых отображена специфичность связывания лиганда GPVI RB571 с GPVI по сравнению с тромбоцитарными рецепторами P2Y1, P2Y12 и PAR-1.
На фиг. 23 представлены диаграммы, на которых отображена специфичность связывания лиганда GPVI RB571 с GPVI по сравнению со взаимодействием с тромбоцитарным рецептором GP1bα-vWF или тромбоцитарным рецептором тромбоксана A2.
На фиг. 24 представлена предсказанная вторичная структура лигандов GPVI EF2-T2 и EF3-T2 с укороченными последовательностями, а также области комплементарности лигандов и модуляторов лигандов GPVI RB416-423.
На фиг. 25 представлены диаграммы индуцированной коллагеном агрегации тромбоцитов, выраженной в виде процента от контроля, для лиганда GPVI EF2-T2 отдельно или в комбинации с различными концентрациями различных модуляторов лигандов GPVI.
На фиг. 26 представлены диаграммы индуцированной коллагеном агрегации тромбоцитов, выраженной в виде процента от контроля, для лиганда GPVI EF3-T2 отдельно или в комбинации с различными концентрациями различных модуляторов лигандов GPVI.
На фиг. 27 представлены диаграммы индуцированной коллагеном агрегации тромбоцитов, выраженной в виде процента от контроля, для лиганда GPVI RB490 отдельно или в комбинации с различными концентрациями различных модуляторов лигандов GPVI.
На фиг. 28 представлена диаграмма, на которой отображена продолжительность обращения активности лиганда GPVI RB490, представляющего собой нуклеиновую кислоту, направленной против GPVI с использованием модулятора лиганда GPVI RB5I5.
На фиг. 29 представлена предсказанная вторичная структура лиганда GPVI RB571 и его предсказанное взаимодействие с модулятором лиганда GPVI RB515.
На фиг. 30 представлены диаграммы индуцированной коллагеном и индуцированной CRP агрегации тромбоцитов, выраженной в виде процента от контроля, для лиганда GPVI RB571 отдельно или в комбинации с различными концентрациями модулятора лиганда GPVI RB515.
На фиг. 31 представлены диаграммы индуцированной коллагеном и индуцированной CRP агрегации тромбоцитов, выраженной в виде процента от контроля, для лиганда GPVI RB571 отдельно или в комбинации с модулятором лиганда GPVI RB515 в обогащенной тромбоцитами плазме.
Подробное описание
Настоящее изобретение относится к фармацевтическим композициям лигандов, представляющих собой нуклеиновые кислоты, которые связываются с гликопротеином VI (GPVI) на мембранах тромбоцитов, модуляторам лигандов и к способам их применения для лечения опосредованных тромбоцитами заболеваний и нарушений. Кроме того, предоставлены фармацевтические составы, содержащие лиганд GPVI, представляющий собой нуклеиновую кислоту, и/или модулятор лиганда GPVI.
A. Определения
Как применяют в настоящем документе термин "приблизительно" при указании измеряемого значения, такого как количество массы, времени, дозы и т.д., предназначен для указания на отклонения на ±20% или ±10%, ±5%, ±1% или ±0,1% от указанного количества, так как такие отклонения являются подходящими для осуществления описанного способа.
"Лиганд, представляющий собой нуклеиновую кислоту", который в настоящем документе может быть указан как "лиганд" или "аптамер", представляет собой нуклеиновую кислоту, которая может формировать третичную структуру, обеспечивающую взаимодействие с молекулой-мишенью. "Лиганд GPVI, представляющий собой нуклеиновую кислоту" или "лиганд GPVI" или "лиганд против GPVI" "лиганда GPVI, представляющего собой нуклеиновую кислоту" относится к лиганду или аптамеру, специфически связывающемуся с GPVI. Термины относятся к олигонуклеотидам со специфическими связывающими областями, способными к формированию комплексов с заданной молекулой-мишенью в физиологическом окружении. Аффинность связывания лиганда с молекулой-мишенью определяют по величине константы диссоциации (Kd) взаимодействия между лигандом и молекулой-мишенью. Как правило, Kd лиганда и его мишени составляет приблизительно от 1 нМ до приблизительно 100 нМ. Специфичность связывания, как правило, определяют на основе сравнительной константы диссоциации лиганда и мишени по сравнению с константой диссоциации относительно лиганда и других веществ в окружении или посторонних молекул. Как правило, Kd лиганда относительно мишени в 10 раз, 50 раз, 100 раз или 200 раз меньше, чем Kd относительно постороннего вещества или сопутствующего вещества в окружении.
"Пара лиганд-модулятор" означает включение конкретного лиганда для молекулы-мишени и модулятора лиганда, который изменяет вторичную и/или третичную структуру лиганда так, что взаимодействие лиганда с его мишенью изменяется. Модулятор может представлять собой олигонуклеотид, комплементарный части лиганда. Модулятор может изменять конформацию лиганда с уменьшением способности лиганда связывать мишень в физиологических условиях на величину от 10% до 100%, от 20% до 100%, на 25%, 40%, 50%, 60%, 70%, 80%, 90% или 100% или на любую величину в процентах в диапазоне от 10% до 100%.
"Модулятор" "антидот", "регулятор" или "регулирующее средство" относится к любому фармацевтическому средству, которое может связывать лиганд или аптамер, как описано в настоящем документе, и желаемым образом модифицировать взаимодействие между этим лигандом и его молекулой-мишенью (например, изменяя структуру лиганда).
Как применяют в настоящем документе, "модулировать" означает снижение, увеличение или какое-либо другое измеримое изменение активности.
Как применяют в настоящем документе, "фармацевтически приемлемый" означает одобренный регулирующим органом федерального правительства или правительства штата или включенный в фармакопею США или другую общепризнанную фармакопею для применения у людей.
Фармацевтически эффективная доза представляет собой дозу, необходимую для предотвращения, ингибирования возникновения или лечения (облегчения симптома до некоторой степени) болезненного состояния. Фармацевтически эффективная доза зависит от типа заболевания, используемой композиции, маршрута введения, вида млекопитающего, подвергаемого лечению, физических характеристик конкретного рассматриваемого млекопитающего, сопутствующего лечения и других факторов, которые известны специалистам в области медицины. В основном, в зависимости от эффективности лиганда, представляющего собой нуклеиновую кислоту, и модулятора, вводят количество от 0,1 мг/кг до 100 мг/кг массы тела/сутки активных ингредиентов.
"Стабилизированная молекула нуклеиновой кислоты" относится к молекуле нуклеиновой кислоты, которая менее быстро разрушается in vivo (например, посредством экзонуклеазы или эндонуклеазы) по сравнению с нестабилизированной молекулой нуклеиновой кислоты. Стабилизация может являться функцией длины, и/или вторичной структуры, и/или включения химических заместителей в сахар фосфатных частей олигонуклеотидного остова. Стабилизации можно добиваться, управляя, например, вторичной структурой, которая может стабилизировать молекулу. Например, если 3'-конец молекулы нуклеиновой кислоты комплементарен предшествующей области, эта часть может загибаться и формировать структуру "стебель-петля", которая стабилизирует молекулу.
Термины "аффинность связывания" и "активность связывания" предназначены для обозначения стремления молекулы лиганда связываться или не связываться с мишенью. Для "активности связывания" и "аффинности связывания" важна энергетика указанных взаимодействий так как она определяет необходимые концентрации взаимодействующих партнеров, скорости, при которых эти партнеры способны к ассоциации, и относительные концентрации связанных и свободных молекул в растворе. Энергетику, в числе прочих способов, можно охарактеризовать посредством определения константы диссоциации, Kd.
Как применяют в настоящем документе "лечение" означает любое лечение заболевания у млекопитающего, включая: (a) защиту от заболевания, то есть, обуславливая отсутствие развития клинических симптомов; (b) ингибирование заболевания, то есть, задерживая, облегчая, уменьшая или подавляя развитие клинических симптомов; и/или (c) облегчение заболевания, то есть, вызывая регрессию клинических симптомов. Специалисты в данной области должны понимать, что в области медицины человека, не всегда возможно различить "предотвращение" и "подавление", так как первичное ведущее нарушение или нарушения могут оставаться неизвестными, скрытыми или пациента не выявляют до момента, значительно позже возникновения нарушения или нарушений. Таким образом, как применяют в настоящем документе, термин "профилактика" подразумевают как элемент "лечения", включающий и "предотвращение" и "подавление", как определено в настоящем документе. Как применяют в настоящем документе, подразумевается, что термин "предохранение" включает "профилактику".
Термин "эффективное количество" означает дозу, достаточную для обеспечения лечения нарушения или состояния болезни, подвергаемого лечению. Оно варьирует в зависимости от пациента, заболевания и проводимого лечения.
Как применяют в настоящем документе, "вариант" лиганда GPVI, представляющего собой нуклеиновую кислоту, включает варианты, по существу выполняющие те же функции, что и лиганд GPVI, представляющий собой нуклеиновую кислоту, и по существу обладающие той же структурой.
B. Гликопротеин VI
Гликопротеин VI (GPVI) специфически экспрессируется на поверхности тромбоцитов. В многочисленных исследованиях показано, что опосредованная коллагеном активация GPVI играет ключевую роль в адгезии и агрегации тромбоцитов. Таким образом, GPVI представляет собой терапевтическую мишень с возрастающим интересом в отношении лечения опосредованных тромбоцитами и коллагеном заболеваний.
Так как граница между физиологическим гемостазом и патологическим тромбозом является очень узкой, важно иметь возможность обеспечить тонко регулируемый контроль активности тромбоцитов для опосредованной коллагеном активации GPVI. Таким образом, в настоящем документе также предоставлены модуляторные компоненты, способные модулировать или регулировать активность описываемых лигандов GPVI.
GPVI представляет собой гликопротеин длиной 339 аминокислотных остатков (номер доступа GeneBank Q9HCN6; описываемый в настоящем документе как SEQ ID NO:1). Аминокислотные остатки 1-20 представляют собой сигнальную последовательность, которая отщепляется с получением зрелого белка, содержащего 319 аминокислот. GPVI имеет два внеклеточных иммуноглобулиновых домена, муциноподобный кор, короткую пептидную линкерную последовательность, трансмембранный домен и короткий цитоплазматический конец, который связывает киназы семейства Src Fyn и Lyn. GPVI также конститутивно образует комплекс с γ-цепью FcR, обеспечивая сборку и активацию Syk и инициацию активации нисходящего пути передачи сигнала, который имеет большое сходство с путем, используемым рецепторами иммунной системы. Ген, кодирующий GPVI находится в лейкоцитарном рецепторном кластере (LRC) на хромосоме человека 19. У мышей, у которых отсутствует GPVI или γ-цепь FcR обнаруживают значительно ослабленный ответ тромбоцитов на коллаген и уменьшенное формирование тромба. Кроме того, в модели тромбоза у крыс in vivo Fab-фрагмент нового моноклонального антитела к GPVI человека, OM4, ингибирует тромбоз без удлинения времени кровотечения, наблюдаемого при использовании антител к GP IIb/IIIa.
Показано, что внеклеточный домен GPVI (SEQ ID NO:2) специфически связывает коллаген типов I-IV (Jung et al. Platelets, 2008, 19:32-42). Кроме того, известно, что коллаген типов I-IV поддерживает активацию, агрегацию и адгезию тромбоцитов, тогда как нефибриллярные коллагены типов VI, VII и VIII индуцируют только слабую адгезию без агрегации тромбоцитов. Таким образом, проведены исследования для идентификации лигандов GPVI, которые связываются с внеклеточным доменом GPVI, для получения фармацевтического средства, которое может являться пригодным при лечении опосредованных тромбоцитами нарушений или заболеваний.
C. Получение лигандов GPVI, представляющих собой нуклеиновые кислоты
Лиганды, представляющие собой нуклеиновые кислоты, которые специфически связывают белок GPVI идентифицировали способом SELEX. Лиганды, которые исходно получали посредством SELEX, затем целиком охарактеризовывали для понимания свойств лигандов GPVI. Такая характеристика включала секвенирование, выравнивание последовательностей для определения консервативных последовательностей, предсказания вторичной структуры и анализ укорочений и мутаций для идентификации областей лигандов, наиболее критичных для желаемой функции специфического связывания и ингибирования GPVI. После идентификации оптимальных последовательностей лигандов и вторичных структур, проводили модификации для оптимизации лигандов для фармацевтического применения. Примеры этих модификаций включают пегилирование, использование спейсера в лиганде, представляющем собой нуклеиновую кислоту, и выбранные модификации сахарной и фосфатной части лиганда, представляющего собой нуклеиновую кислоту. Для контроля функции лиганда, как результат различных применяемых модификаций, проводили анализы связывания.
SELEX относится к систематической эволюции лигандов при экспоненциальном обогащении. Этот способ обеспечивает эволюцию молекул нуклеиновых кислот in vitro с высокоспецифичным связыванием молекул-мишеней. Способ SELEX описан, например, в патентах США №№ 7087735; 5475096 и 5270163 (также см. WO 91/19813).
Способ SELEX включает отбор олигонуклеотидов-кандидатов из смеси и пошаговые циклические повторения связывания, разделения и амплификации с использованием одной и той же общей схемы отбора с достижением практически любого желаемого показателя аффинности связывания и селективности. Начиная от смеси нуклеиновых кислот, такой как смеси, содержащие участок случайной последовательности, способ SELEX включает стадии приведения смеси в контакт с мишенью в условиях, подходящих для связывания, разделения несвязавшихся нуклеиновых кислот и тех нуклеиновых кислот, которые специфически связались с молекулами-мишенями, диссоциации комплексов нуклеиновая кислота-мишень, амплификации нуклеиновых кислот, диссоциированных из комплексов нуклеиновая кислота-мишень с получением обогащенной лигандами смеси нуклеиновых кислот, затем повторения стадий связывания, разделения, диссоциации и амплификации на протяжении множества циклов насколько желательно для выхода высокоспецифичных, высокоаффинных лигандов для молекулы-мишени.
Основной способ SELEX модифицирован для достижения ряда конкретных задач. Например, в патенте США № 5707796 описано применение SELEX в сочетании с электрофорезом в геле для отбора молекул нуклеиновой кислоты с конкретными структурными характеристиками, такими как изгиб ДНК. В патенте США № 5763177 описан основанный на SELEX способ отбора лигандов, содержащих фотореакционноспособные группы, способные к связыванию и/или к фотосшиванию и/или фотоинактивации молекулы-мишени. В патенте США № 5580737 описан способ идентификации высокоспецифичных лигандов, способных к различению близкородственных молекул, названный Counter-SELEX. В патентах США №№ 5567588 и 5861254 описаны основанные на SELEX способы, позволяющие добиваться высокоэффективного разделения олигонуклеотидов с высокой и низкой аффинностью к молекуле-мишени. В патенте США № 5496938 описаны способы получения улучшенных лигандов после проведения способа SELEX. В патенте США № 5705337 описаны способы ковалентного связывания лиганда с его мишенью.
В патенте США № 5648214 продемонстрирована возможность идентификации лигандов, представляющих собой нуклеиновые кислоты, к небольшим пептидам в растворе. В патенте США № 5780228, относящемся к получению высокоаффинных лигандов, связывающихся с определенными лектинами, проиллюстрирована возможность использовать аффинную элюцию с использованием лигандов с получением лигандов, направленных к конкретному участку на молекуле-мишени. В патенте США № 6127119 описаны способы получения лигандов, представляющих собой нуклеиновые кислоты, к определенным тканям, которые содержат группы типов клеток. В патенте США № 6673553 описано получение определенных модифицированных высокоаффинных лигандов к кишечной фосфатазе теленка. В патенте США № 6716580 описан автоматизированный способ идентификации лигандов, представляющих собой нуклеиновые кислоты, включающий использование роботизированных манипуляторов.
В его основной форме способ SELEX можно определить следующим рядом стадий:
1) Получают смесь-кандидат нуклеиновых кислот с различными последовательностями. Смесь-кандидат в основном содержит области фиксированных последовательностей (т.е., каждый из представителей смеси-кандидата содержит одинаковые последовательности в одинаковом положении) и области случайных последовательностей. Области фиксированных последовательностей выбраны: (a) для помощи на стадиях амплификации, описанных ниже, (b) для имитации последовательности, для которой известно, что она связывается с мишенью или (c) для увеличения концентрации данной структурной конфигурации нуклеиновых кислот в смеси-кандидате. Случайные последовательности могут являться полностью случайными (т.е., вероятность нахождения основания в любом положении равна одной четвертой) или только частично случайной (например, вероятность нахождения основания в любом положении можно выбирать на любом уровне от 0 до 100%).
2) Смесь-кандидат приводят в контакт с выбранной мишенью в условиях, подходящих для связывания мишени и представителей смеси-кандидата. В этих условиях взаимодействие между мишенью и нуклеиновыми кислотами смеси-кандидата можно рассматривать как формирующее комплексы нуклеиновая кислота-мишень между мишенью и нуклеиновыми кислотами с наиболее сильной аффинностью к мишени.
3) Нуклеиновые кислоты с наиболее высокой аффинностью к мишени разделяют с нуклеиновыми кислотами с меньшей аффинностью к мишени. Так как в смеси-кандидате присутствуют только крайне малое количество последовательностей (а возможно только одна молекула нуклеиновой кислоты), соответствующих нуклеиновым кислотам с наиболее высокой аффинностью, как правило, желательно определять условия разделения так, чтобы при разделении сохранялось значительное количество нуклеиновых кислот в смеси-кандидате (приблизительно от 5 до 50%).
4) Затем нуклеиновые кислоты, отобранные при разделении, как обладающие относительно высокой аффинностью к мишени, амплифицируют с получением новой смеси-кандидата, обогащенной нуклеиновыми кислотами с относительно более высокой аффинностью к мишени.
5) При повторении стадий разделения и амплификации, указанных выше, вновь сформированная смесь-кандидат содержит все меньше и меньше слабо связывающихся последовательности, и средняя степень аффинности нуклеиновых кислот к мишени, как правило, возрастает. Если брать в предельном случае, способ SELEX позволяет получать смесь-кандидат, содержащую одну или небольшое количество уникальных нуклеиновых кислот, представляющих собой последовательности нуклеиновых кислот из исходной смеси-кандидата, которые укладываются во вторичную и третичную структуры, обеспечивающие наиболее высокое аффинное взаимодействие с молекулой-мишенью.
SELEX можно использовать для получения бивалентного связывания с двумя или более связывающими доменами с аффинностью к двум или более эпитопам белка, включая рецептор. Конкретно, в одном из вариантов осуществления способ можно использовать для отбора лигандов, представляющих собой нуклеиновые кислоты, с аффинностью к двум или более областям рецептора GPVI. Например, в определенных вариантах осуществления лиганд может связываться по меньшей мере с двумя частями областей C2. В определенных вариантах осуществления лиганд влияет на димеризацию рецептора GPVI, нарушая или стабилизируя конформацию димера. В этих вариантах осуществления можно конструировать модуляторы для снижения связывания только с одним, несколькими или всеми эпитопами, с которыми связывается лиганд, представляющий собой нуклеиновую кислоту. Например, модулятор может препятствовать связыванию только с одним эпитопом, таким как область C2-1 или C2-2 рецептора.
Лиганды, представляющие собой нуклеиновые кислоты, специфичные к GPVI, можно получать проводя SELEX с короткими пептидами, представляющими внеклеточный домен молекулы, способами SELEX, как, например, описано в патенте США № 7087735. Альтернативно лиганды, представляющие собой нуклеиновые кислоты, специфичные к GPVI, можно выделять, проводя SELEX с интактными тромбоцитами, с фракциями мембран тромбоцитов, обогащенных белком, с очищенным GPVI или с линиями клеток, специфически сверхэкспрессирующими рецептор GPVI, способами SELEX, как, например, описано в патенте США № 6730482.
Кроме того, способ SELEX можно проводить для выделения специфических лигандов GPVI, представляющих собой нуклеиновые кислоты, с использованием схем конкурентной аффинной элюции, таких как схемы, описанные в патенте США № 5780228. Например, для выделения лигандов, представляющих собой нуклеиновые кислоты, специфичные к GPVI, элюцию лигандов, связанных с белком можно проводить посредством добавления достаточных количеств активатора или связывающего средства GPVI, такого как коллаген, конвулксин или CRP или родственные соединения.
Можно рекомбинантно экспрессировать рецептор GPVI и очищать белок служащий для процедуры SELEX. В определенных вариантах осуществления лиганд GPVI, представляющий собой нуклеиновую кислоту, связывается с рецептором GPVI в физиологических условиях. Физиологические условия, как правило, относятся к уровню солей и pH раствора. In vitro, физиологические условия, как правило, воспроизводят в буфере, содержащем 150 мМ NaCl, 2 мМ CaCl2 20 мМ HEPES, при pH приблизительно 7,4. В определенных вариантах осуществления для скрининга множества лигандов, представляющих собой нуклеиновые кислоты, как описано выше, и получения обогащенного множества, содержащего лиганды, направленные к белкам, находящимся на тромбоцитах используют нативные, как правило, неактивированные, тромбоциты. Затем обогащенное множество используют со стабильной клеточной линией, сверхэкспрессирующей желаемый рецептор GPVI, или клеточной линией, в которую транзиторно ввели белок. Повторный скрининг можно проводить модифицированным способом SELEX с выделенными из этих клеток рецепторами или с целыми клетками, исследуя конкуренцию лигандов или определяя действие на внутриклеточные пути передачи сигнала.
В определенных вариантах осуществления лиганды, представляющие собой нуклеиновые кислоты, к конкретным мишеням GPVI можно идентифицировать с использованием иммобилизованного белка. В некоторых из этих вариантов осуществления очищенный белок можно связывать с твердым матриксом посредством химического линкера. В других вариантах осуществления можно выделять мембраны клеток, сверхэкспрессирующих конкретный белок, с использованием детергента, такого как анионный детергент (например, холат), с выделением определенной фракции белков, и смесь связывали с иммобилизованной искусственной мембраной. В основном, полагают, что перегруппировку можно проводить, удаляя детергент, в ходе чего липиды и белки перегруппировываются и формируют слой с углеводородными цепями иммобилизованной искусственной мембраны, как правило, находящейся на подлежащем матриксе, таком как гранулы.
Лиганды, представляющие собой нуклеиновые кислоты, специфичные для GPVI, выделенные этими способами SELEX, которые также обладают желаемой функциональной активностью, можно идентифицировать посредством скрининга лигандов, представляющих собой нуклеиновые кислоты, на их способность ингибировать специфическую индуцированную агонистами функцию тромбоцитов и/или события внутриклеточной передачи сигнала, вызываемые GPVI. Так как желаемые лиганды, представляющие собой нуклеиновые кислоты, являются не только связывающими партнерами, но также являются ингибиторами передачи сигнала рецепторами, можно идентифицировать лиганды с желаемой функцией, оценивая действие лиганда на активность тромбоцитов. Это может включать характеристику действий лиганда на различные сигнальные пути, для которых известно, что они регулируются GPVI. Например, передача сигнала GPVI может приводить к кластеризации белка GPVI и активации киназ с запуском локальной сигнальной цепи событий, активирующих фосфолипазу Cγ2, высвобождая вторичные мессенджеры 1,4,5-инозитолтрифосфаты и диацилглицерин, ответственные за повышение уровней Ca2+ и активацию протеинкиназы C. Любые из этих систем или сигналов вторичных мессенджеров можно измерять хорошо известными специалистам в данной области способами.
Кроме того, в этих системах можно измерять коллаген, связывающий GPVI в присутствии или отсутствие лиганда GPVI, представляющего собой нуклеиновую кислоту.
Лиганды также можно подвергать скринингу на ингибирование агрегации тромбоцитов в анализах функции тромбоцитов, таких как оптическая агрегометрия, проводимая в обогащенной тромбоцитами плазме и препаратах отмытых тромбоцитов, или импедансная агрегометрия, проводимая в цельной крови. Кроме того, лиганды также можно подвергать скринингу на ингибирование взаимодействия тромбоцитов с покрытыми коллагеном поверхностями в статических условиях или в потоке цельной крови. Специфичность данного лиганда, представляющего собой нуклеиновую кислоту, к GPVI можно дополнительно определять по способности лиганда блокировать события внутриклеточной передачи сигнала, запускаемые известными агонистами данного рецептора. Например, можно ожидать, что лиганд-ингибитор GPVI, представляющий собой нуклеиновую кислоту, будет блокировать передачу сигнала, запускаемую, например, конвулксином змеи, представляющим собой лектин C-типа, коллагеном или родственным коллагену пептидом (CRP). Специфичность данного лиганда, представляющего собой нуклеиновую кислоту, к GPVI также можно дополнительно определять по отсутствию действия на агрегацию тромбоцитов, когда агрегацию запускает агонист, активирующий рецептор, отличный от GPVI.
Использование любого из описанных выше способов, отдельно или в комбинации, позволяет получать множество лигандов, представляющих собой нуклеиновые кислоты, специфичных к GPVI. После идентификации лиганда, представляющего собой нуклеиновую кислоту, с желаемыми ингибирующими свойствами, можно идентифицировать модуляторы этого лиганда, как описано ниже.
D. Лиганды GPVI, представляющие собой нуклеиновые кислоты.
Лиганды GPVI, описываемые в настоящем документе, предпочтительно представляют собой лиганды, представляющие собой нуклеиновые кислоты, такие как лиганды. Лиганды GPVI, специфически связывающие внеклеточный домен (ECD) GPVI (аминокислотные остатки Gln21-Lys267; SEQ ID NO:2) выбирали с использованием способа SELEX, более подробно описанного в примере 1, затем модифицировали для увеличения стабильности, аффинности к GPVI и/или способности регулировать активность GPVI.
Лиганд GPVI, представляющий собой нуклеиновую кислоту, по настоящему изобретению состоит из выделенной последовательности нуклеиновой кислоты, которая может представлять собой ДНК или РНК и которую можно синтезировать с использованием модифицированных рибо- или дезоксирибонуклеиновых кислот. Как описано в настоящем документе, если используют структуру оснований РНК, структура в последовательности оснований вместо тимидина (T) включает уридин (U). В определенных вариантах осуществления, описываемого в настоящем документе, последовательность нуклеиновых кислот записана в виде последовательности РНК. Подобным образом, в определенных вариантах осуществления, описываемого в настоящем документе, где лиганд, представляющий собой нуклеиновую кислоту, исходно идентифицирован в виде молекулы ДНК, последовательность нуклеиновых кислот записана в виде последовательности ДНК. Следует понимать, что последовательность нуклеотидов, представленная в текстовой форме в виде последовательности ДНК по сути представляет описание соответствующей последовательности РНК, где остатки тимина (T) в последовательности ДНК заменены остатками уридина (U) с получением соответствующей последовательности нуклеотидов РНК. Подобным образом, следует понимать, что последовательность, представленная в текстовой форме в виде последовательности РНК по сути представляет описание соответствующей последовательности ДНК, где остатки уридина (U) в последовательности РНК заменены остатками тимина (T) с получением соответствующей последовательности ДНК.
Некоторые лиганды GPVI, представляющие собой нуклеиновые кислоты, полученные способом SELEX, секвенировали и их последовательности выравнивали. Выравнивание последовательностей, представленное на фиг. 4, привело к идентификации 6 уникальных последовательностей, полученных в результате обогащения посредством процесса отбора. Выравнивание 6 последовательностей, обозначенных как "A"-"F", продемонстрировало присутствие последовательности UAA. Кроме того, эта абсолютно консервативная последовательность содержится в последовательности: (G/A)UAA. Последовательность (G/A)UAA, с каждой стороны фланкированная GC, приводит к консервативной последовательности GC(G/A)UAAGC.
Затем для уникальных лигандов GPVI проводили анализ предсказания вторичной структуры. Вторичная структура вносит вклад в особенности функционирования лиганда. Как хорошо понимают специалисты в данной области, вторичную структуру можно описать в терминах структур стеблей и петель, как они встречаются в молекуле в направлении от 5' к 3'. На основе предсказания вторичной структуры, как описано в примере 2 ниже, лиганды ECD GPVI обладают вторичной структурой, содержащей три стебля и четыре петли. Конфигурация от 5' к 3' содержит первый стебель (стебель 1 или S1), первую петлю (петля 1 или L1), второй стебель (стебель 2 или S2), соединенный со второй и третьей петлей (петля 3 или L3), третью петлю, третий стебель (стебель 3 или S3) и четвертую петлю (петля 4 или L4) (см. фиг. 8A-8C). В определенных вариантах осуществления второй стебель связан с первой, второй и третьей петлями, а третий стебель связан с третьей и четвертой петлями. В одном из вариантов осуществления третий стебель находится рядом с первым стеблем в направлении 5'-3'. Последовательность GC(G/A)UAAGC формирует стебель 3 (пара оснований GC) и петлю 4 ((G/A)UAA). Последовательность GAC формирует петлю 1.
Мутационный анализ лигандов GPVI, идентифицированных SELEX, демонстрирует, что последовательность петли 3 UA, UU или UG обеспечивает высокоаффинное связывание лигандов с GPVI. Таким образом, в некоторых вариантах осуществления петля 3 лиганда GPVI, представляющего собой нуклеиновую кислоту, содержит последовательность 5'-YD-3', где Y представляет собой пиримидин, а D представляет собой U, G или A.
В определенных вариантах осуществления петлю 2 лиганда GPVI, представляющего собой нуклеиновую кислоту, можно известными специалистам в данной области способами замещать спейсером. Спейсер может представлять собой ненуклеотидный спейсер, который обеспечивает структуру, аналогичную петле 2 так, что лиганд GPVI сохраняет свою структуру и функцию, когда петля 2 замещена спейсером. Замещение петли 2 гексаэтиленгликолевым спейсером, обеспечиваемое встраиванием (9-O-диметокситритилтриэтиленгликоль,1-[(2-цианоэтил)-(N,N-диизопропил)]фосфорамидита (см. фиг. 13 A-13B) в лиганд GPVI, представляющий собой нуклеиновую кислоту, не приводит к потере аффинности к GPVI. Таким образом, специалист в данной области может понять, что петлю 2 можно заменить рядом ненуклеотидных спейсеров, которые являются коммерчески доступными. Примеры таких спейсеры в качестве неограничивающих примеров включают спейсеры, получаемые встраиванием в лиганд GPVI, представляющий собой нуклеиновую кислоту, 5'-O-диметокситритил-1'2'-дидезоксирибоза-3'-[(2-цианоэтил)-(N,N-диизопропил)]фосфорамидита; 18-O-диметокситритилгексаэтиленгликоль,1-[(2-цианоэтил)-(N,N-диизопропил)]фосфорамидита и 12-(4,4'-диметокситритилокси)додецил-1-[(2-цианоэтил)-(N,N-диизопропил)]фосфорамидита.
Эффективность лиганда GPVI в регуляции функции GPVI или лечения опосредованного тромбоцитами заболевания во многом зависит от способности лиганда с достаточной аффинностью связываться с белком GPVI. Таким образом, после получения лигандов GPVI способом SELEX, каждый лиганд секвенируют, а затем его можно охарактеризовать в отношении связывания с молекулой-мишенью. Аффинность связывания лигандов по настоящему документу к мишени (GPVI) можно определять посредством Kd. Величину этой константы диссоциации можно определять непосредственно хорошо известными способами, такими как способы связывания радиоактивных лигандов, описанные в примере 1. Однако замечено, что для некоторых низкомолекулярных олигонуклеотидов прямое определение Kd иногда является трудным и может приводить к неоправданно высоким результатам. В этих условиях можно проводить анализ конкурентного связывания молекулы-мишени или другого вещества-кандидата относительно веществ, для которых известно, что они связывают мишень или кандидата. Значение концентрации, при которой происходит 50% ингибирование (Ki), в идеальных условиях равна Kd. Однако ни при каких условиях Ki не будет меньше Kd. Таким образом, определение Ki в качестве альтернативы дает максимальное значение для величины Kd. В тех условиях, где технические трудности препятствуют точному измерению Kd, его можно подходящим образом заменить измерением Ki с получением верхнего предела Kd. Значение Ki также можно использовать для подтверждения того, что лиганд по настоящему изобретению связывает мишень. При характеристике связывающих свойств лиганда GPVI, с использованием конкурентного связывания или функциональных анализов с известными молекулами, связывающими GPVI, такими как коллаген, CRP (родственный коллагену пептид) или конвулксин (Cvx), можно анализировать специфичность.
В определенных вариантах осуществления Kd связывания лиганда с GPVI может находиться в диапазоне приблизительно от 1 нМ до приблизительно 100 нМ, приблизительно от 10 нМ до приблизительно 50 нМ или приблизительно от 20 нМ до приблизительно 0,1 нМ. В других вариантах осуществления Kd связывания лиганда с GPVI по меньшей мере в 2 раза, 3 раза, 4 раза, 5 раз или 10 раз меньше, чем Kd связывания лиганда с посторонним белком или другим сопутствующим веществом в окружении. Посторонний белок также может представлять собой белок с мотивами, родственными мотивам, находящимся в GPVI, такими как другой представитель суперсемейства Ig или другой белок, содержащий коллаген-связывающий домен или другой рецептор активации или адгезии тромбоцитов.
Как более подробно будет описано ниже, активность связывания лиганда, полученного и идентифицированного способом SELEX, можно дополнительно модифицировать или усиливать рядом технологических способов.
В определенных вариантах осуществления лиганд взаимодействует с внеклеточным доменом GPVI. Лиганд может препятствовать связыванию коллагена с рецептором GPVI. В определенных вариантах осуществления лиганд может ингибировать внутриклеточную передачу сигнала через рецептор GPVI, включая снижение образования инозитолтрифосфата или колебания внутриклеточных уровней кальция. Лиганд также может стабилизировать или нарушать конформацию рецептора, такую как димерная конформация так, что у рецептора снижается способность взаимодействовать с коллагеном или FcRγ. Лиганд может воздействовать на активацию тромбоцитов коллагеном или другими агонистами GPVI. Лиганд также может воздействовать на адгезию тромбоцитов к коллагену или к родственным коллагену пептидам. Лиганд может воздействовать на агрегацию тромбоцитов, индуцированную коллагеном или другими агонистами GPVI.
Лиганды, представляющие собой нуклеиновые кислоты, описываемые в настоящем документе, могут функционировать в качестве активно обратимых средств. Они представляют собой средства или фармацевтически активные молекулы, которые после введения пациенту можно напрямую регулировать введением второго средства. Как более подробно описано ниже, второе средство, обозначаемое в настоящем документе как модулятор, может выключать или тонко регулировать фармакологическую активность лиганда. В результате фармакологическую активность лиганда можно обращать средствами, отличными, например, от выведения лекарственного средства.
E. Модуляторы
В определенных вариантах осуществления лиганды GPVI, представляющие собой нуклеиновые кислоты, являются обратимыми. В одном из аспектов изобретение относится к способу модуляции активности лиганда GPVI, представляющего собой нуклеиновую кислоту, посредством введения индивидууму, которому вводили лиганд, представляющий собой нуклеиновую кислоту, модулятора лиганда GPVI.
Модуляторы по настоящему изобретению включают любое фармацевтическое средство, которое может связываться с лигандом, представляющим собой нуклеиновую кислоту, и модифицировать взаимодействие между этим лигандом и его молекулой-мишенью (например, модифицируя структуру лиганда, представляющего собой нуклеиновую кислоту) желаемым образом, или которое разрушает, метаболизирует, разрушает или иным образом химически изменяет лиганд, представляющий собой нуклеиновую кислоту, модифицируя его биологическое действие. Примеры модуляторов по настоящему изобретению включают: олигонуклеотиды или их аналоги, которые комплементарны по меньшей мере части последовательности лиганда, представляющего собой нуклеиновую кислоту (включая рибозимы или ДНКзимы). Другие примеры включают пептидные нуклеиновые кислоты (ПНК), морфолинонуклеиновые кислоты (МНК) или замкнутые нуклеиновые кислоты (ЗНК); связывающие нуклеиновую кислоту белки или пептиды; олигосахариды; низкомолекулярные соединения или связывающие нуклеиновую кислоту полимеры, липиды, наночастицы или основанные на микросферах модуляторы.
Модуляторы можно разрабатывать так, чтобы они связывали конкретный лиганд, представляющий собой нуклеиновую кислоту, с высокой степенью специфичности и желаемой степенью аффинности. Модуляторы также можно разрабатывать так, чтобы после связывания структура лиганда модифицировалась в более или менее активную форму. Например, модулятор можно конструировать так, чтобы после связывания с лигандом, представляющим собой нуклеиновую кислоту, к которому он направлен, изменялась вторичная и/или третичная структура этого лиганда, в результате чего лиганд больше не мог бы связываться с его молекулой-мишенью или связывался с его молекулой-мишенью с меньшей аффинностью. Альтернативно, модулятор можно конструировать так, чтобы после связывания изменялась трехмерная структура лиганда так, чтобы увеличивалась аффинность лиганда к его молекуле-мишени. Т.е., модулятор можно конструировать так, чтобы после связывания структурный мотив модифицировался так, чтобы возрастала аффинность лиганда. В другом варианте осуществления пару лиганд/модулятор конструируют так, чтобы связывание модулятора с молекулой лиганда, представляющего собой нуклеиновую кислоту, которая не может связываться с представляющей интерес мишенью, могла приводить к образованию в лиганде структурного мотива, который, таким образом, позволяет лиганду связываться с его молекулой-мишенью.
Модуляторы также можно конструировать для неспецифического связывания конкретного лиганда, представляющего собой нуклеиновую кислоту, или группы лигандов, представляющих собой нуклеиновые кислоты, с аффинностью, достаточной для формирования комплекса. Такие модуляторы в основном могут связываться с нуклеиновыми кислотами посредством межзарядовых взаимодействий. Также такие модуляторы могут одновременно связываться более чем с одним лигандом, представляющим собой нуклеиновую кислоту. Модулятор можно конструировать так, чтобы после связывания с одним или несколькими лигандами, представляющими собой нуклеиновые кислоты, структура лиганда, представляющего собой нуклеиновую кислоту, изменялась относительно его активной формы незначительно, но вместо этого, модулятор маскировал связывание лиганда, представляющего собой нуклеиновую кислоту, с его молекулой-мишенью или стерически препятствовал ему.
Нуклеотидные модуляторы могут быть любой длины, которая обеспечивает эффективное связывание с молекулой лиганда. Например, длина олигонуклеотидных модуляторов может находиться в диапазоне приблизительно от 10 нуклеотидов (н.) до приблизительно 30 н., приблизительно от 10 н. до приблизительно 20 н. или приблизительно от 15 н. Длина нуклеотидных модуляторов может составлять 8 н., 9 н., 10 н., 11 н., 12 н., 13 н., 14 н., 15 н., 16 н., 17 н., 18 н., 19 н., 20 н., 21 н., 22 н., 23 н., 24 н., 25 н., 26 н., 27 н., 28 н., 29 н. или 30 н. Специалист в данной области также может представить себе нуклеотидные модуляторы с длиной более 30 н.
Как описано в настоящем документе, лиганд, представляющий собой нуклеиновую кислоту, обладает активной третичной структурой, на которую можно влиять формированием соответствующей стабильной вторичной структуры. Таким образом, хотя механизм формирования дуплекса между комплементарным олигонуклеотидным модулятором по изобретению и лигандом, представляющим собой нуклеиновую кислоту, сходен с формированием дуплекса между двумя короткими линейными олигорибонуклеотидами, на правила проектирования таких взаимодействий и кинетику формирования такого продукта можно воздействовать посредством внутримолекулярной структуры лиганда.
В формировании конечного стабильного дуплекса значительную роль играет скорость нуклеации при формировании исходной пары нуклеотидов между лигандом, представляющим собой нуклеиновую кислоту, и олигонуклеотидным модулятором, и скорость этой стадии значительно увеличивается при нацеливании олигонуклеотидного модулятора на одноцепочечные петли и/или одноцепочечные 3'- или 5'- концы, присутствующие в лиганде, представляющем собой нуклеиновую кислоту. Для оптимального формирования межмолекулярного дуплекс, свободная энергия должна идеально благоприятствовать формированию межмолекулярного дуплекса по отношению к формированию существующих внутримолекулярных дуплексов в лиганде-мишени, представляющем собой нуклеиновую кислоту.
Как правило, описанные в настоящем документе модуляторы по изобретению представляют собой олигонуклеотиды, содержащие последовательность, комплементарную по меньшей мере части последовательности лиганда-мишени, представляющего собой нуклеиновую кислоту. Например, модуляторный олигонуклеотид может содержать последовательность, комплементарную приблизительно от 6 н. до 25 н., от 8 н. до 20 н. или от 10 н. до 15 н. лиганда-мишени. Длину модуляторного олигонуклеотида можно легко оптимизировать способами, описанными в настоящем документе и известными специалистам в данной области, учитывая лиганд-мишень и желаемое действие. Олигонуклеотид можно получать с нуклеотидами в стереохимической конфигурации D или L или с их комбинацией. Природные нуклеозиды находятся в D-конфигурации.
Хотя олигонуклеотидные модуляторы по изобретению содержат последовательность, комплементарную по меньшей мере части лиганда, представляющего собой нуклеиновую кислоту, абсолютной комплементарности не требуется. Последовательность, "комплементарная по меньшей мере части лиганда, представляющего собой нуклеиновую кислоту", указываемая в настоящем документе, представляет собой последовательность с достаточной комплементарностью, чтобы обладать способностью гибридизоваться с лигандом, представляющим собой нуклеиновую кислоту. Способность гибридизоваться может зависеть от степени комплементарности и длины нуклеиновой кислоты. Как правило, чем более длинным является гибридизующийся олигонуклеотид, чем большее количество несоответствующих оснований с лигандом-мишенью он может содержать и все же формировать стабильный дуплекс (или триплекс, в зависимости от ситуации). Специалист в данной области может устанавливать допустимую степень несоответствия с использованием стандартных способов определения температуры плавления гибридизующегося комплекса. Олигонуклеотиды по изобретению могут представлять собой одноцепочечные ДНК или РНК или гибридные смеси или их производные или модифицированные варианты.
Модуляторы могут содержать модификации остова нуклеиновой кислоты и структуры отдельных нуклеиновых кислот. В определенных вариантах осуществления модулятор представляет собой нуклеиновую кислоту, комплементарную по меньшей мере одной области петли в лиганде. В определенных вариантах осуществления модулятор представляет собой олигонуклеотид по меньшей мере с последовательностью, в физиологических условиях гибридизующейся с четвертой петлей в двумерную структуру, а особенно олигонуклеотид, содержащий последовательность 3'-AUU-5'. В других вариантах осуществления модулятор представляет собой олигонуклеотид, в физиологических условиях гибридизующийся с первой петлей во вторичной структуре лиганда, и, в частности, олигонуклеотид, по меньшей мере содержащий последовательность 3'-CUG-5' В зависимости от желаемой функции модулятора, модулятор можно конструировать так, чтобы нарушать или стабилизировать вторичную и/или третичную структуру лиганда, представляющего собой нуклеиновую кислоту.
В определенных вариантах осуществления модулятор конструируют так, чтобы он связывал "суицидальное положение" в лиганде и, таким образом, разрушал последовательность лиганда. Суицидальное положение представляет собой одноцепочечную часть лиганда, чувствительную к ферментативному расщеплению. В одном иллюстративном варианте осуществления суицидальное положение становится одноцепочечным и неустойчивым при связывании модулятора с лигандом и может усиливать расщепление лиганда ферментами в циркуляции, такими как эндонуклеазы крови или печени. В определенных вариантах осуществления модулятор связывается с лигандом, после чего лиганд больше не может взаимодействовать с его мишенью.
В одном иллюстративном варианте осуществления модулятор содержит последовательность нуклеиновой кислоты, выбранную из SEQ ID NO:74-88, включительно.
В определенных вариантах осуществления последовательность модулятора содержит по меньшей мере один модифицированный нуклеотид. Например, модификации 2'-O-метил и 2'-фтор, которые могут включать 2'-O-метилцитозин, 2'-O-метилуридин, 2'-O-метиладенозин, 2'-O-метилгуанозин, 2'-фторцитидин или 2'-фторуридин.
Для определения оптимального участка в лиганде, представляющем собой нуклеиновую кислоту, для связывания олигонуклеотидным модулятором можно использовать различные стратегии. Можно использовать эмпирическую стратегию, при которой комплементарные олигонуклеотиды "гуляют" по лиганду, представляющему собой нуклеиновую кислоту. В соответствии с этим подходом можно использовать олигонуклеотиды (например, 2'-O-метил- или 2'-фторолигонуклеотиды) длиной приблизительно 15 нуклеотидов, ступенчато располагающихся на лиганде приблизительно через 5 нуклеотидов (например, олигонуклеотиды, комплементарные 1-15, 6-20, 11-25 и т.д. лиганда). Эмпирическая стратегия может являться особенно эффективной, так как влияние третичной структуры лиганда, представляющего собой нуклеиновую кислоту, на эффективность гибридизации может являться трудным для предсказания.
Для оценки способности различных олигонуклеотидов гибридизоваться с конкретным лигандом, представляющим собой нуклеиновую кислоту, можно использовать анализы, описанные в примерах, приведенных ниже, с особым вниманием на молярном избытке олигонуклеотида, необходимом для достижения полного связывания лиганда, представляющего собой нуклеиновую кислоту. Также можно определять способность различных олигонуклеотидных модуляторов увеличивать скорость диссоциации лиганда, представляющего собой нуклеиновую кислоту, с его молекулой-мишенью или скорость ассоциации лиганда с ней, проводя стандартные кинетические исследования, например, с применением анализов BIACORE. Олигонуклеотидные модуляторы можно выбирать так, что для модификации взаимодействия лиганда и его молекулы-мишени желаемым образом необходим 5-50-кратный молярный избыток олигонуклеотида или менее.
Альтернативно, лиганд-мишень, представляющий собой нуклеиновую кислоту, можно модифицировать так, чтобы он для содействия ассоциации с олигонуклеотидным модулятором содержал одноцепочечный конец (3' или 5'). Подходящие концы могут содержать от 1 до 20 нуклеотидов, от 1 до 10 нуклеотидов, от 1 до 5 нуклеотидов или от 3 до 5 нуклеотидов. Концы также можно модифицировать (например, модификация 2'-O-метил и 2'-фтор, которая может включать 2'-O-метилцитозин, 2'-O-метилуридин, 2'-O-метиладенозин, 2'-O-метилгуанозин, 2'-фторцитидин или 2'-фторуридин). Лиганды, содержащие одноцепочечные концы, можно тестировать в анализах связывания и биологических анализах (например, как описано в примерах ниже) для подтверждения того, что добавление одноцепочечного конца не нарушает активную структуру лиганда, представляющего собой нуклеиновую кислоту. Можно конструировать ряд олигонуклеотидов (например, 2'-O-метилолигонуклеотиды), которые могут формировать, например, 1, 2, 3, 4 или 5 пар оснований с концевой последовательностью и тестировать на их способность связываться с содержащим одноцепочечный конец лигандом отдельно, а также на их способность увеличивать скорость диссоциации лиганда с его молекулой мишенью или скорость ассоциации лиганда с ней. Для подтверждения того, что действие является результатом формирования дуплекса, а не неспецифических эффектов, можно использовать контроли с последовательностями со случайными перестановками.
В другом варианте осуществления модулятор представляет собой рибозим или ДНКзим. Ферментативные нуклеиновые кислоты действуют посредством первичного связывания с РНК- или ДНК-мишенью. Такое связывание осуществляется посредством связывающей мишень части ферментативной нуклеиновой кислоты, которая находится в непосредственной близости с ферментативной частью молекулы, которая действует, расщепляя ДНК-мишень. Таким образом, ферментативная нуклеиновая кислота сначала распознает, а затем связывает РНК- или ДНК- мишень посредством спаривания комплементарных оснований, после связывания в соответствующем участке, ферментативно действует, расщепляя РНК-мишень, таким образом, обеспечивая инактивацию РНК-лигандов. Существуют по меньшей мере пять классов рибозимов, каждый из которых демонстрирует отличающийся тип специфичности. Например, интроны группы 1 представляют собой последовательности длиной приблизительно от 300 до >1000 нуклеотидов, и им в последовательности мишени непосредственно с 5'-конца от участка расщепления необходим U, и они связывают 4-6 нуклеотидов с 5'-конца от участка расщепления. Другим классом является РНК РНКаза P (РНК M1), длина которой составляет приблизительно от 290 до 400 нуклеотидов. Третьим классом являются рибозимы типа головки молотка, длина которых составляет приблизительно от 30 до 40 нуклеотидов. Им необходима последовательность-мишень UH (где H не представляет собой G) непосредственно с 5'-конца от участка расщепления, и они связывают переменное количество нуклеотидов на обоих концах участка расщепления. Четвертым классом являются рибозимы-шпильки, длина которых составляет приблизительно 50 нуклеотидов. Им необходима последовательность-мишень GUC непосредственно с 3'-конца от участка расщепления, и они связывают 4 нуклеотиды на 5'-конце участка расщепления и переменное количество на 3'-конце участка расщепления. Пятой группой является рибозимы вируса гепатита дельта (HDV), длина которых составляет приблизительно 60 нуклеотидов. ДНКзимы являются одноцепочечными и расщепляют и РНК, и ДНК. Общая модель ДНКзима является предположительной и известна как модель "10-23". ДНКзимы по модели "10-23" содержат каталитический домен из 15 дезоксирибонуклеотидов, фланкированный двумя распознающими субстрат доменами каждый из дезоксирибонуклеотидов в количестве от семи до девяти.
В другом варианте осуществления модулятор сам является лигандом, представляющим собой нуклеиновую кислоту. В этом варианте осуществления получают первый лиганд, который связывается с желаемой терапевтической мишенью. На второй стадии способом SELEX, описываемым в настоящем документе, или другим способом получают второй лиганд, который связывается с первым лигандом и модулирует взаимодействие терапевтического лиганда и мишени. В одном из вариантов осуществления второй лиганд прекращает действие первого лиганда.
В другом иллюстративном варианте осуществления модулятор представляет собой основанный на ПНК, МНК, ЗНК или PCO. Нуклеиновые основания олигонуклеотидных модуляторов по изобретению могут соединяться посредством связей между нуклеиновыми основаниями, например, пептидными связями (как в случае пептидных нуклеиновых кислот (ПНК); Nielsen et al. (1991) Science 254, 1497 и патент США № 5539082) и морфолиносвязей (Qin et al., Antisense Nucleic Acid Drug Dev. 10, 11 (2000); Summerton, Antisense Nucleic Acid Drug Dev. 7, 187 (1997); Summerton et al., Antisense Nucleic Acid Drug Dev. 7, 63 (1997); Taylor et al., J. Biol. Chem. 271, 17445 (1996); Partridge et al., Antisense Nucleic Acid Drug Dev. 6, 169 (1996)), или посредством другой природной или модифицированной связи. Также олигонуклеиновые основания могут представлять собой замкнутые нуклеиновые кислоты (LNAs). Nielsen et al., J. Biomol. Struct. Dyn. 17, 175 (1999); Petersen et al., J. Mol. Recognit. 13, 44 (2000); Nielsen et al., Bioconjug. Chem. 11, 228 (2000).
ПНК представляют собой соединения, аналогичные олигонуклеотидам, но отличающиеся по строению. В ПНК, дезоксирибозный остов олигонуклеотида заменен пептидным остовом. Каждая субъединица пептидного остова присоединена к природному или неприродному нуклеиновому основанию. ПНК часто содержат ахиральный полиамидный остов, состоящий из N-(2-аминоэтил)глициновых единиц. Пуриновые или пиримидиновые основания присоединены к каждой единице метиленовым карбонильным линкером (1-3) с направлением на комплементарную нуклеиновую кислоту. ПНК связывается с комплементарной РНК или ДНК в параллельной или антипараллельной ориентации по правилам спаривания Уотсона-Крика. Незаряженный характер олигомеров ПНК увеличивает стабильность гибридных дуплексов ПНК/ДНК(РНК) по сравнению с природными гомодуплексами.
Морфолинонуклеиновые кислоты называют так потому, что они составлены из морфолиновых субъединиц, каждая из которых содержит одно из четырех генетических оснований (аденин, цитозин, гуанин и тимин), присоединенных к 6-членному морфолиновому кольцу. От восемнадцати до двадцати пяти субъединиц из этих четырех типов субъединиц связаны в конкретном порядке неионными фосфородиамидатными межсубъединичными связями с получением морфолиноолиго.
ЗНК представляет собой класс аналогов ДНК, обладают рядом характеристик, которые делают его основным кандидатом в модуляторы по изобретению. Мономеры ЗНК представляют собой бициклические соединения, структурно сходные с мономерами РНК. ЗНК обладает большинством химических свойств ДНК и РНК, она является водорастворимой, ее можно разделять гель-электрофорезом, ее осаждает этанол и т.д. (Tetrahedron, 54, 3607-3630 (1998)). Однако введение мономеров ЗНК в олигонуклеотиды ДНК или РНК приводит к высокой термостабильности дуплексов с комплементарной ДНК или РНК, при соблюдении в то же время правил спаривания оснований Уотсона-Крика.
Также в качестве модулятора по настоящему изобретению можно использовать псевдоциклические олигонуклеиновые основания (PCO) (см. патент США № 6383752). PCO содержат два олигонуклеотидных участка, соединенных их 3'-3' или 5'-5' концами. Один из участков ("функциональный участок") PCO обладает такой же функциональностью (например, комплементарностью к РНК-мишени). Другой участок ("защитный участок") комплементарен 3'- или 5'-концу функционального участка (в зависимости от того конца, которым он присоединен к функциональному участку). В результате комплементарности между функциональным и защитным участками, PCO в отсутствии нуклеиновых кислот-мишеней (например, РНК) формируют внутримолекулярные псевдоциклические структуры. PCO являются более стабильными, чем обычные олигонуклеотиды вследствие присутствия 3'-3' или 5'-5' связей и формирования внутримолекулярных псевдоциклических структур. Исследования фармакокинетики, тканевого распределения и стабильности на мышах позволяет предположить, что, как правило, PCO обладают более высокой стабильностью in vivo, чем PS-олигонуклеотиды и сходными с ними фармакокинетикой и профилями тканевого распределения, но более быстрым выведением из выбранных тканей. Когда с PCO по настоящему изобретению соответствующим образом связаны молекулы флуорофора и гасителя, когда молекула находится в линейной конфигурации, она флуоресцирует, но в циклической конформации флуоресценция гасится. Это свойство можно использовать для скрининга PCO в качестве потенциальных модуляторов.
В другом иллюстративном варианте осуществления модуляторы представляют собой модуляторы на основе пептидов. Основанные на пептидах модуляторы лигандов, представляющих собой нуклеиновые кислоты, представляют собой альтернативный класс молекул модуляторов олигонуклеотидов или их аналогов. Этот класс модуляторов особенно пригоден, если вследствие отсутствия удовлетворительной одноцепочечной области для обеспечения нуклеации между мишенью и олигонуклеотидным модулятором невозможно выделить достаточно активные олигонуклеотидные модуляторы лиганда-мишени, представляющего собой нуклеиновую кислоту. Кроме того, пептидные модуляторы обеспечивают отличную от олигонуклеотидных модуляторов биологическую доступность и фармакокинетику. В одном иллюстративном варианте осуществления модулятор представляет собой протамин (Oney et al., 2009, Nat. Med. 15:1224-1228). Протамины растворимы в воде, не сворачиваются при нагревании и содержат аргинин, аланин и серин (большинство также содержат пролин и валин, и многие содержат глицин и изолейцин). Модуляторы также включают варианты протамина (см. например, Wakefield et al., J. Surg. Res. 63:280 (1996)) и модифицированные формы протамина, включая формы, описанные в публикации США № 20040121443. Другие модуляторы включают фрагменты протамина, такие как фрагменты, описанные в патенте США № 6624141 и публикации США № 20050101532. Как правило, модуляторы также включают пептиды, модулирующие активность гепарина, другие гликозаминогликаны или протеогликаны (см., например, патент США № 5919761). В одном иллюстративном варианте осуществления модуляторы представляют собой пептиды, содержащие катионные NH-группы, обеспечивающие стабилизацию межзарядовых взаимодействий, такие как поли-1-лизин и поли-1-орнитин.
Доступно несколько стратегий выделения пептидов, способных связываться с лигандом-мишенью, представляющим собой нуклеиновую кислоту и, таким образом, модулировать его активность. Например, описаны кодируемые пептидные комбинаторные библиотеки, иммобилизованные на гранулах, и показано, что они содержат пептиды, способные связывать вирусные последовательности РНК и нарушать взаимодействие вирусной РНК и вирусного регуляторного белка, специфически связывающего указанную РНК (Hwang et al. Proc. Natl. Acad. Sci USA, 1999, 96:12997). С использованием таких библиотек, модуляторы лигандов, представляющих собой нуклеиновые кислоты, можно выделять, добавляя метку в лиганд-мишень, представляющий собой нуклеиновую кислоту, и инкубируя вместе меченую мишень и иммобилизованную на гранулах пептидную библиотеку в условиях, благоприятных для связывания некоторых членов библиотеки и нуклеиновой кислоты. Связывание лиганда, представляющего собой нуклеиновую кислоту, с конкретным пептидом на конкретной грануле вызывает "окрашивание" гранулы меткой на лиганде, представляющем собой нуклеиновую кислоту, и, таким образом, обеспечивает идентификацию пептидов, способных связывать мишень, посредством простого выделения гранулы. Прямое взаимодействие пептидов, выделенных такими способами скрининга, и лиганда-мишени, представляющего собой нуклеиновую кислоту, можно подтверждать и определять количественно с использованием любого ряда анализов связывания, описанных для идентификации модуляторов лигандов, представляющих собой нуклеиновые кислоты. Способность указанных пептидов модулировать активность лиганда-мишени, представляющего собой нуклеиновую кислоту, можно подтверждать подходящими биологическими анализами.
В дополнительном варианте осуществления модуляторы представляют собой модуляторы на основе олигосахаридов. Олигосахариды могут взаимодействовать с нуклеиновыми кислотами. Например, антибиотические аминогликозиды являются продуктами видов Streptomyces и специфически взаимодействуют с разнообразным множеством молекул РНК, таким как различные рибозимы, РНК компоненты рибосом и последовательности TAR и RRE ВИЧ-1. Таким образом, олигосахариды могут связываться с нуклеиновыми кислотами, и их можно использовать для модуляции активности лигандов, представляющих собой нуклеиновые кислоты.
В другом варианте осуществления модулятор представляет собой модулятор на основе низкомолекулярного соединения. Низкомолекулярное соединение, встраивающееся между лигандом и мишенью или иным образом нарушающее или модифицирующее связывание лиганда и мишени также можно использовать в качестве терапевтического регулятора. Такие низкомолекулярные соединения можно идентифицировать посредством скрининга кандидатов в анализе, в котором определяют изменения связывания лиганда и мишени в присутствии низкомолекулярного соединения и без, или с применением анализа in vivo или in vitro, в котором определяют отличия биологического действия лиганда на мишень в присутствии низкомолекулярного соединения и без. После идентификации низкомолекулярного соединения, проявляющего желаемое действие, для оптимизации химической структуры для желаемого регуляторного действия можно использовать способы, такие как комбинаторные подходы.
В дополнительном иллюстративном варианте осуществления модулятор представляет собой связывающие нуклеиновые кислоты полимер, липид, наночастицу или микросферу. В дополнительных неограничивающих примерах модулятор можно выбирать из группы, состоящей из: 1,2-диолеоил-sn-глицеро-3-этилфосфохолина (EDOPC); дилауроилэтилфосфатидилхолина (EDLPC); EDLPC/EDOPC; пиридиниевых поверхностно-активных веществ; диолеоилфосфатидилэтаноламина (DOPE); бромида (±)-N-(3-аминопропил)-N,N-диметил-2,3-бис(додецилокси)-1-пропанаминия (GAP-DLRIE) c нейтральным колипидом диолеоилфосфатидилэтаноламином (DOPE) (GAP-DLRIE/DOPE); петагидрохлорида (±)-N,N-диметил-N-[2-(сперминкарбоксамидо)этил]-2,3-бис(диоеилокси-1-пропанаминия) (DOSPA); дилауроилэтилфосфатидилхолина (EDLPC); этилдимиристоилфосфатидилхолина (EDMPC); хлорида (±)-N,N,N-триметил-2,3-бис(z-октадец-9-еноилокси)-1-пропанаминия (DOTAP); бромида (±)-N-2-(2-гидроксиэтил)-N,N-диметил-2,3-бис(тетрадецилокси)-1-пропанаминия (DMRIE); хлорида (±)-N,N,N-триметил-2,3-бис(z-октадец-9-енилокси)-1-пропанаминия (DOTMA); 5-карбоксиспермилглициндиоктадециламида (DOGS); 5-карбоксиспермиламида дипальмитоилфосфатидилэтаноламина (DPPES); 1,3-диолеоилокси-2-(6-карбоксиспермил)пропиламида (DOSPER); тетраметилтетрапальмитоилспермина (TMTPS); (тетраметилтетраолеилспермина (TMTOS); тетраметилтетралаурилспермина (TMTLS); тетраметилтетрамиристилспермина (TMTMS); тетраметилдиолеилспермина (TMDOS); дифитаноилфосфатидилэтаноламина (DPhPE) и бромида (±)-N-(3-аминопропил)-N,N-диметил-2,3-бис(додецилокси)-1-пропанаминия (GAP-DLRIE).
В других вариантах осуществления модулятор выбран из группы, состоящей из: хитозана; производного хитозана; 1,5-диметил-1,5-диазаундекаметиленполиметобромида; блок-сополимеров полиоксиэтилена/полиоксипропилена; поли-L-лизина; полиамидоамина (PAMAM); содержащего β-циклодекстрин поликатиона (CDP); содержащего β-циклодекстрин поликатиона (содержащий имидазол вариант) (CDP-Im); полифосфорамидатного полимера (8кДа, 30кДа) (PPA-DPA 8k, PPA-DPA 30k); полибрена; спермина; PEG-блок-PLL-дендримеров; полиэтиленимина (PEI); манноза-PEI; трансферрин-PEI; линера-PEI (lPEI); желатина; метакрилата/метакриламида; поли(бета-амино сложных эфиров); полиэлектролитных комплексов (PEC); поли(виниламина) (PVA); коллагена; полипропиленимина (PPI): полиаллиламина; поливинилпиридина; аминоацеталированного поли(винилового спирта); акрилового или метакрилового полимера; дендримера Ньюкома; полифенилена; бромида диметилдиоктадециламмония (DAB); бромида цетилтриметиламмония (CTAB); альбумина; обработанного кислотой желатина; полилизина; полиорнитина; полиаргинина; DEAE-целлюлозы; DEAE-декстрана и поли(N,N-диметиламиноэтилметакрилата) и полипропиламина (POPAM).
В одном из вариантов осуществления модулятор выбран из хитозана и производных хитозана. Производные хитозана включают водорастворимые наночастицы хитозана (такие, как описано в патенте США № 6475995; патентной заявке США № 2006/0013885; Limpeanchob et al., (2006) Efficacy and Toxicity of Amphotericin B-Chitosan Nanoparticles; Nareusan University Journal 14(2):27-34). Учитывая поликатионный характер хитозанового полимера (по существу очень большого полиаминового полимера, состоящего из повторяющихся мономеров глюкозамина), хитозан можно использовать для агрегации и/или инкапсуляции лигандов в полиэлектролитный комплекс in vivo после инъекции индивидууму. Это частично основано на взаимодействиях первичных аминов, находящихся в хитозане, и фосфодиэфирного остова лиганда.
В определенных вариантах осуществления первичные амины в хитозановом полимере по существу могут являться модифицированными для изменения водорастворимости и состояния заряда. Производные хитозана вкючают триметилхитозанхлорид (TMC), который можно синтезировать с различными степенями кватернизации; монокарбоксиметилированный хитозан (MCC), представляющий собой полиамфолитический полимер; поперечно сшитое производное глутаральдегида (CSGA): тиолированный хитозан (Lee, et al. (2007) Pharm. Res, 24:157-67); гликольхитозан (GC), производное хитозана, конъюгированное с этиленгликолем (Lee, et al. (2007) Int. J. Pharm.); [N-(2-карбоксибензил)хитозан (CBCS) (Lin, et al. (2007) Carbohydr. Res. 342(1):87-95); бета-циклодекстринхитозановый полимер (Venter, et al. (2006) Int. J. Pharm. 313(l-2):36-42); O-карбоксиметилхитозан; N,O-карбоксиметилхитозан или хитозан, химически модифицированный введением в его остов ксантатной группы.
В одном из вариантов осуществления получают пустые хитозановые наночастицы и используют их в качестве модуляторов. Можно использовать хитозан или производные хитозана в диапазоне молекулярной массы от 10000 Да до >1000000 Да. В определенных вариантах осуществления масса хитозана представляет собой 500000 Да или менее. В определенных вариантах осуществления масса хитозана представляет собой 100000 Да или менее. В определенных вариантах осуществления масса соединения составляет от 10000 до 100000 Да, от 10000 до 90000, от 10000 до 80000, от 20000 до 700000, от 30000 до 70000, приблизительно 30000, приблизительно 40000, приблизительно 50000 или приблизительно 60000 Да.
В определенных вариантах осуществления используют хитозановые полимеры, содержащие различные доли деацетилированных первичных аминов. В этих вариантах осуществления различная степень диацетилирования изменяет состояние заряда полимера и, таким образом, связывающие свойства полимера. После контакта хитозановой наночастицы с лигандами у индивидуума, лиганды могут связываться с поверхностью наночастицы и поверхность наночастицы может их захватывать, или они могут проникать в наночастицу и наночастица может инкапсулировать их посредством ионных взаимодействий.
В другом варианте осуществления модулятор представляет собой микросферу из полифосфатного полимера. В определенных вариантах осуществления модулятор представляет собой производное такой микросферы, такие как поли(сополимер L-лактида с этилфосфитом) или P(LAEG-EOP) и другие, как описано в патенте США № 6548302. Можно получать такие полимеры, содержащие ряд функциональных групп как часть полимерного остова. В одном из примеров, полимеры могут содержать четвертичные амины с положительным зарядом при физиологическом pH так, что они при контакте могут образовывать комплекс с одной или несколькими нуклеиновыми кислотами или инкапсулировать их. В определенных вариантах осуществления полимеры не содержат положительных зарядов.
Настоящее изобретение также относится к способам идентификации модуляторов лигандов GPVI, представляющих собой нуклеиновую кислоту. В основном модуляторы можно идентифицировать посредством анализов связывания, молекулярного моделирования или анализов in vivo или in vitro, в которых определяют модификацию биологической функции. В одном из вариантов осуществления связывание модулятора с нуклеиновой кислотой определяют посредством анализа сдвига в геле. В другом варианте осуществления связывание модулятора с лигандом, представляющим собой нуклеиновую кислоту, определяют посредством анализа BIACORE.
Для идентификации и отбора модуляторов по изобретению можно использовать стандартные анализы связывания. Неограничивающие примеры представляют собой анализ сдвига в геле и анализы BIACORE. Т.е. тестируемые модуляторы можно приводить в контакт с лигандами-мишенями, представляющими собой нуклеиновые кислоты, в условиях тестирования или типичных физиологических условиях и проводить определение того, действительно ли тестируемый модулятор связывается с лигандом. Затем тестируемые модуляторы, для которых выявлено, что они связывают лиганд, представляющий собой нуклеиновую кислоту, можно анализировать в подходящем биологическом анализе (который варьирует в зависимости от лиганда и его молекулы-мишени, например коагуляционные пробы) для определения того, может ли тестируемый модулятор воздействовать на биологическое действие, оказываемое лигандом на его молекулу-мишень.
Анализ сдвига в геле представляет собой хорошо известный способ, применяемый для оценки связывающей способности. Например, фрагмент ДНК, содержащий тестируемую последовательность, сначала инкубируют с тестируемым белком или смесью, содержащей предполагаемые связывающие белки, а затем разделяют на геле посредством электрофореза. Если фрагмент ДНК связывается с белком, он становится большим по размеру и, таким образом, его миграция относительно миграции свободного фрагмента замедляется. Например, один из способов анализа сдвига электрофоретической подвижности в геле может представлять собой (a) приведение связывающего нуклеиновую кислоту белка в контакт с нерадиоактивно или радиоактивно меченой молекулой нуклеиновой кислоты, содержащей молекулярный зонд, в смеси в подходящих условиях с обеспечением специфических связывающих взаимодействий между белком и нуклеиновой кислотой для формирования комплекса, где указанный зонд выбран из группы, состоящей из дцДНК, оцДНК и РНК; (b) электрофорез смеси и (c) детекцию комплекса, связанного с мембраной, посредством детекции нерадиоактивной или радиоактивной метки в комплексе.
Посредством технологии BIACORE определяют события связывания на поверхности сенсорного чипа так, что взаимодействующее вещество, присоединенное к поверхности, определяет специфичность анализа. Тестирование специфичности взаимодействия включает простой анализ того, могут ли различные молекулы связываться с иммобилизованным взаимодействующим веществом. Связывание приводит к немедленному изменению сигнала поверхностного плазмонного резонанса (SPR) так, что непосредственно очевидно происходит ли взаимодействие или нет. Биосенсоры на основе SPR измеряют взаимодействия посредством измерения массовой концентрации биомолекул рядом с поверхностью. Поверхность делают специфичной, прикрепляя один из взаимодействующих партнеров. Образец, содержащий другого партнера(ов), омывает поверхность: когда молекулы из образца связываются с взаимодействующим веществом, присоединенным к поверхности, локальная концентрация изменяется и измеряют ответную SPR. Ответ прямо пропорционален массе молекул, связывающихся с поверхностью.
SPR возникает, когда свет в определенных условиях отражается от проводящего слоя на границе двух сред с различным показателем преломления. В технологии BIACORE среды представляют собой образец и стекло на сенсорном чипе, а проводящий слой представляет собой тонкий слой золота на поверхности чипа. SPR вызывает снижение интенсивности отраженного света при конкретном угле отражения. Этот угол варьирует в зависимости от показателя преломления рядом с поверхностью на стороне, противоположной отраженному свету. Когда молекулы в образце связываются с поверхностью сенсора, концентрация и, таким образом, показатель преломления на поверхности изменяются и детектируют ответ SPR. Нанесение на график зависимости ответа от времени при взаимодействии обеспечивает количественную меру развития взаимодействия. Посредством технологии BIACORE определяют угол минимальной интенсивности отраженного света. Свет не поглощается образцом: вместо этого световая энергия посредством SPR рассеивается в слое золота. Значения ответа SPR выражают в резонансных единицах (РЕ). Одна РЕ представляет собой изменение угла минимума интенсивности на 0,0001°, для большинства белков это приблизительно равно изменению концентрации на поверхности сенсора приблизительно на 1 пг/мм2. Точный коэффициент пересчета между РЕ и поверхностной концентрацией зависит от свойств поверхности сенсора и природы молекулы, ответственной за изменение концентрации.
Существует ряд других анализов, которыми можно определять может ли олигонуклеотид или его аналог, пептид, полипептид, олигосахарид или низкомолекулярное соединение связываться с лигандом таким образом, что модифицируется взаимодействие с мишенью. Например, для оценки способности средства связывать лиганд, представляющий собой нуклеиновую кислоту, можно использовать анализы сдвига электрофоретической подвижности (EMSA), титрационную калориметрию, сцинтилляционные анализы сближения, анализы седиментационного равновесия с применением аналитического ультрацентрифугирования (например, см. www.cores.utah.edu/interaction), анализы поляризации флуоресценции, анализы анизотропии флуоресценции, анализы интенсивности флуоресценции, анализы резонансного переноса энергии флуоресценции (FRET), анализы связывания на нитроцеллюлозном фильтре, ELISA, ELONA (см., например, патент США № 5789163), RIA или анализы равновесного диализа. Можно проводить прямые анализы, в которых непосредственно определяют взаимодействие средства и лиганда, представляющего собой нуклеиновую кислоту, или можно проводить анализы конкуренции или вытеснения, в которых определяют способность средства вытеснять лиганд из его мишени (например, см. Green, Bell and Janjic, Biotechniques 30(5), 2001, p 1094 и патент США № 6306598). После идентификации модулирующего средства-кандидата, его способность модулировать активность лиганда, представляющего собой нуклеиновую кислоту, в отношении его мишени можно подтверждать в биологическом анализе. Альтернативно, если идентифицировано средство, которое может модулировать взаимодействие лиганда с его мишенью, такие анализы связывания можно использовать для подтверждения того, что средство непосредственно взаимодействует с лигандом и можно измерять аффинность указанного взаимодействия.
В другом варианте осуществления для идентификации модулятора, который связывает лиганд, представляющий собой нуклеиновую кислоту, участка(ов) взаимодействия модулятора и лиганда, представляющего собой нуклеиновую кислоту и относительной аффинности связывания средств для лиганда можно использовать масс-спектрометрию (например, см. патент США № 6329146). Такие масс-спектрометрические способы также можно использовать для скрининга химических смесей или библиотек, особенно комбинаторных библиотек, на отдельные соединения, которые связываются с выбранным лигандом-мишенью, которые можно использовать в качестве модуляторов лиганда. Кроме того, масс-спектрометрические способы можно использовать для скрининга нескольких лигандов-мишеней, представляющих собой нуклеиновые кислоты, одновременно, например, с комбинаторной библиотекой соединений. Кроме того, масс-спектрометрические способы можно использовать для идентификации взаимодействия множества видов молекул, особенно "малых" молекул, и молекулярного участка взаимодействия на лиганде-мишени.
Анализы in vivo или in vitro, посредством которых оценивают эффективность модулятора в модификации взаимодействия лиганда, представляющего собой нуклеиновую кислоту, и мишени специфичны для нарушения, подлежащего лечению. Существует множество стандартных анализов биологических свойств, которые хорошо известны и которые можно использовать. Примеры биологических анализов предоставлены в патентах, цитируемых в настоящей заявке, описывающих определенные лиганды, представляющие собой нуклеиновые кислоты, для конкретного применения.
В определенных вариантах осуществления модулятор представляет собой низкомолекулярное соединение. Например, в определенных вариантах осуществления лиганд, представляющий собой нуклеиновую кислоту, связан с молекулой биотина. В таких вариантах для связывания и обращения действия лиганда вводят стрептавидин или авидин (см. Savi et. al. J. Thrombosis and Haemostasis, 6: 1697-1706). Авидин представляет собой тетрамерный белок, продуцируемый в яйцеводах птиц, пресмыкающихся и амфибий, откладывающийся в белках их яиц. Стрептавидин представляет собой тетрамерный белок, выделяемый у бактерии Streptomyces avidinii. Тетрамерный белок содержит четыре идентичные субъединицы (гомотетрамер), каждая из которых с высокой степенью аффинности и специфичности может связываться с биотином (витамин B7, витамин H).
В определенных вариантах осуществления модулятор представляет собой катионную молекулу. В определенных вариантах осуществления лиганд формирует структуру гуанинового квартета (G-квартета или G-квадруплекса). Эти структуры связаны катионными молекулами. В определенных вариантах осуществления молекулы представляют собой хелатирующие молекулы металлов. В определенных вариантах осуществления модулятор представляет собой порфирин. В определенных вариантах осуществления соединение представляет собой TMPyP4. См. Joachimi, et.al. JACS 2007, 129, 3036-3037 и Toro, et.al. Analytical Biochemistry 2008, Aug 1, 379 (1) 8-15.
В одном из вариантов осуществления модулятор обладает способностью значительного связывания с лигандом, представляющим собой нуклеиновую кислоту, в растворе при концентрациях модулятора менее десяти (10,0) микромоль/л (мкМ), одного (1,0) микромоль/л (мкМ), предпочтительно менее 0,1 мкМ, а более предпочтительно - менее 0,01 мкМ. Под "значительным" подразумевают то, что при модуляции в присутствии мишени наблюдают по меньшей мере 50 процентное уменьшение заданной биологической активности, и 50% уменьшение в настоящем документе обозначают как значение IC50.
F. Оптимизация лигандов и модуляторов
Для того, чтобы лиганд был пригоден для использования в качестве терапевтического средства, лиганд предпочтительно должен быть недорог при синтезе, безопасен для применения у индивидуума и стабилен in vivo. Как правило, олигонуклеотиды РНК и ДНК дикого типа нестабильны in vivo вследствие их чувствительности к разрушению нуклеазами. Устойчивость к разрушению нуклеазами можно значительно увеличить введением модифицирующих групп в 2'-положение.
В смеси олигонуклеотидов, из которых затем выбирают лиганды, можно вводить 2'-фтор- или аминогруппы. По настоящему описанию для получения исходной смеси олигонуклеотидов для отбора лигандов в реакции транскрипции in vitro использовали 2'-фторпиримидины (см. пример 1). Однако полученные лиганды, выбранные из таких библиотек, содержащие сахара с 2'-гидроксилом в каждом положении пурина, хотя и являясь более стабильными in vivo, чем сравнимые лиганды, представляющие собой РНК или ДНК, требуют дополнительной оптимизации. Таким образом, лиганды, идентифицированные способами, описываемыми в настоящем документе, затем рядом способов модифицируют с получением лигандов с улучшенной функцией и стабильностью, а также с увеличенной пригодностью для способов крупномасштабного производства.
После исходной идентификации лигандов (например, посредством SELEX) и модуляторов (например, конструирование на основе комплементарности последовательности), лиганды и модуляторы можно рядом способов модифицировать или конструировать для улучшения их желаемой структуры, функции и/или стабильности. Эти способы в качестве неограничивающих примеров включают замену конкретных остатков сахара, изменения состава и размера конкретных областей и/или структур в лиганде и конструирование лигандов, которые можно более эффективно регулировать модулятором.
Конструирование и оптимизация лиганда, представляющего собой нуклеиновую кислоту, включает анализ вторичной структуры лиганда, а также связей между вторичной структурой и контролем модулятора. В отличие от общепринятых способов модулирования нуклеиновых кислот, конструирование лигандов белка GPVI может включать рассмотрение воздействия изменений лиганда на конструирование потенциальных модуляторов. Если лиганд модифицируют, например, посредством укорочения, необходимо сконструировать соответствующий модулятор для контроля укороченного лиганда.
Вторичную структуру лигандов, идентифицированных способом SELEX, можно предсказывать различными способами, известными специалистам в данной области. Например, каждую последовательность можно анализировать с применением такого программного обеспечения, как Mfold (mfold.bioinfo.rpi.edu; также см. Zuker, 2003, Nucleic Acids Res. 31:3406-3415 и Mathews, et al., 1999, J. Mol. Biol. 288:911-940). Затем, можно использовать сравнительный анализ последовательностей для различных отобранных последовательностей с выравниванием последовательностей в зависимости от консервативных консенсусных вторичных структурных элементов для получения по предсказанной вторичной консенсусной структуре лигандов GPVI (см. пример 2). Такой анализ, как описанный выше, позволяет конструировать и тестировать варианты последовательностей, полученных посредством SELEX, с получением лигандов с улучшенной функцией и стабильностью.
Лиганды GPVI по настоящему изобретению, представляющие собой нуклеиновые кислоты, можно модифицировать, варьируя общую длину лиганда, а также длины стеблевых и петлевых структур. Например, можно получать укорочения лигандов, в которых в лиганде, выбранном способом SELEX, удалена часть с 5'- и/или 3'-конца лиганда. Для определения размера укорочений, которые выдерживает лиганд, одним из используемых способов может являться отжиг олигонуклеотида (например, олигонуклеотида ДНК), комплементарного 5'- или 3'-концевой области лиганда, при нагревании с последующим сравнением связывания лиганда с отожженным олигонуклеотидом и без него. Если значительных отличий в связывании лиганда с отожженным олигонуклеотидом и без него не наблюдают, это позволяет предположить, что отожженная часть лиганда необязательна для связывания лиганда белком-мишенью. Этот способ можно проводить с использованием олигонуклеотидов, отжигающихся с 5'- или 3'-концами лиганда различной длины, для определения 5'- и 3'-границ, обеспечивающих полнофункциональный лиганд.
В другом варианте осуществления конструирование включает уменьшение размера лиганда. В другом варианте осуществления в зависимости от размера лиганда изменяют размер модулятора. В еще одном варианте осуществления участки, содержащие остатки гуанина уменьшают до участков, содержащих менее четырех остатков гуанина, или менее трех остатков гуанина, или менее двух остатков гуанина или без остатков гуанина. Однако сочетанное действие этих изменений должно удовлетворять требованию получения лиганда, обеспечивающего достаточную активность, но который легко нейтрализовать модулятором.
Для нацеливания модулятора, улучшенный лиганд также можно модифицировать так, чтобы он включал одноцепочечный конец (3' или 5') для содействия связывания с олигонуклеотидным модулятором. Подходящие концы могут содержать от 1 н. до 20 н., предпочтительно от 1 н. до 10 н., от 1 н. до 5 н. или от 3 н. до 5 н. Легко понять, что такие концы могут включать модифицированные нуклеотиды, как более подробно описано ниже.
Содержащие одноцепочечные концы лиганды можно тестировать в анализах связывания и биологических анализах (например, как описано ниже) для подтверждения того, что добавление одноцепочечного конца не нарушает активную структуру лиганда. Можно конструировать ряд олигонуклеотидов (например, 2'-O-метилолигонуклеотиды), которые могут формировать, например, 1, 2, 3, 4 или 5 пар оснований с концевой последовательностью и тестировать на их способность связываться с содержащим одноцепочечный конец лигандом отдельно, а также на их способность увеличивать скорость диссоциации лиганда с его молекулой мишенью или скорость ассоциации лиганда с ней. Для подтверждения того, что действие является результатом формирования дуплекса, а не неспецифических эффектов, можно использовать контроли с последовательностями со случайными перестановками.
Конструирование лигандов также облегчает определение консенсусной структуры с идентификацией одного или нескольких нуклеотидов, которые могут улучшать или ухудшать структуру и функцию лиганда. Например, можно более эффективно идентифицировать и тестировать добавления, делеции и замены нуклеотидов в конкретные стеблевые и петлевые структуры (см. пример 3).
Знание консенсусной вторичной структуры также позволяет избежать модификаций, которые могут вредить структуре и функции лиганда. Например, определенные модификации, такие как 2'-фтор в области стебля или петли, могут являться консервативными в консенсусной вторичной структуре. В этих случаях, удаление 2'-фтора из стебля или петли лиганда может приводить к потере активности.
В определенных вариантах осуществления лиганды представляют собой молекулы нуклеиновой кислоты, выбранные из таблиц 1-7, включая укороченные варианты и в значительной степени гомологичные им последовательности. Как применяют в настоящем документе, в отношении гомологичных областей, "в значительной степени гомологичная" последовательность представляет собой последовательность, которая в конкретной молекуле формирует такую же вторичную структуру при спаривании оснований по Уотсону-Крику. В определенных вариантах осуществления последовательности являются "в значительной степени гомологичными", если идентичность их последовательности с конкретным лигандом составляет по меньшей мере 80%, 85% или более, например, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%. Относительно лиганда, представляющего собой нуклеиновую кислоту, указанной длины, такой как 50 или менее нуклеотидов, гомологичную последовательность можно найти в любой области, позволяющей связыванию по Уотсон-Крику формировать такую же вторичную структуру независимо от идентичности последовательностей в указанной области.
Лиганды также можно конструировать так, чтобы они содержали суицидальное положение, обеспечивающее более эффективную регуляцию спаривающимися модуляторами. После связывания лиганда модулятором, суицидальное положение становится одноцепочечным и неустойчивым, таким образом, облегчая расщепление лиганда ферментами, находящимися в крови в природе, такими как эндонуклеазы крови или печени. Это обеспечивает средства для эффективного и по существу немедленного устранения активного лиганда из циркуляции.
Химические модификации
Одной из проблем с которыми сталкиваются при терапевтическом применении нуклеиновых кислот является то, что внутриклеточные и внеклеточные ферменты, такие как эндонуклеазы и экзонуклеазы в жидкостях организма могут быстро разрушать олигонуклеотиды в их фосфодиэфирной форме до реализации желаемого действия. Определенные химические модификации лиганда, представляющего собой нуклеиновую кислоту, могут увеличивать стабильность лиганда, представляющего собой нуклеиновую кислоту, in vivo или увеличивать или опосредовать доставку лиганда, представляющего собой нуклеиновую кислоту. Кроме того, определенные химические модификации могут увеличивать аффинность лиганда, представляющего собой нуклеиновую кислоту, к его мишени, стабилизируя или стимулируя формирование необходимых структурных элементов в лиганде, представляющем собой нуклеиновую кислоту, или обеспечивая дополнительные молекулярные взаимодействия с молекулой-мишенью.
Модификации лигандов в качестве неограничивающих примеров могут включать модификации, обеспечивающие химические группы, обеспечивающие основаниям лиганда, представляющего собой нуклеиновую кислоту, или лиганду как целому дополнительный заряд, поляризуемость, гидрофобность, образование водородных связей, электростатические взаимодействия и функциональность. Такие модификации в качестве неограничивающих примеров включают модификации сахаров в положении 2', модификации пиримидинов в положении 5, модификации пуринов в положении 8, модификации по экзоциклическим аминам, замещение 4-тиоуридином, замещение 5-бром- или 5-йодурацилом, модификации остова, модификации тиофосфатом или алкилфосфатом, метилирование, необычные комбинации при спаривании оснований, такие как изооснования изоцитидин и изогуанидин и т.п. Модификации также могут включать 3'- и 5'-модификации, такие как кэпирование.
Способ SELEX включает идентификацию высокоаффинных лигандов, представляющих собой нуклеиновые кислоты, содержащие модифицированные нуклеотиды, придающие лиганду улучшенные характеристики, такие как улучшенная стабильность in vivo или улучшенные характеристики доставки. Примеры таких модификаций включают химические замены в рибозе и/или фосфате и/или в положениях основания. Идентифицированные посредством SELEX лиганды, представляющие собой нуклеиновые кислоты, содержащие модифицированные нуклеотиды, описаны в патенте США № 5660985, в котором описаны олигонуклеотиды, содержащие производные нуклеотидов, химически модифицированные в 5- и 2'-положениях пиримидинов. В патенте США № 5580737 описаны конкретные лиганды, представляющие собой нуклеиновые кислоты, содержащие один или несколько нуклеотидов, модифицированных 2'-амино (2'-NH2), 2'-фтором (2'-F) и/или 2'-O-метилом (2'-OMe). В патенте США № 5756703 описаны олигонуклеотиды, содержащие различные модифицированные по 2' пиримидины.
Способ SELEX включает комбинирование отобранных олигонуклеотидов с другими отобранными олигонуклеотидами и не являющимися олигонуклеотидами функциональными единицами, как описано в патентах США №№ 5637459 и 5683867. В патенте США № 5637459 описаны высокоспецифичные лиганды, представляющие собой нуклеиновые кислоты, содержащие один или несколько нуклеотидов, модифицированных 2'-амино (2-NH2), 2'-фтором (2'-F) и/или 2'-O-метилом (2'-OMe). Способ SELEX дополнительно включает комбинирование отобранных лигандов, представляющих собой нуклеиновые кислоты, с липофильными или неиммуногенными высокомолекулярными соединениями в диагностический или терапевтический комплекс, как описано в патенте США № 6011020.
Когда лиганды, представляющие собой нуклеиновые кислоты, получают способом SELEX, модификации могут представлять собой модификации до или после SELEX. Посредством модификаций до SELEX можно получать лиганды и со специфичностью к их мишеням, и с улучшенной стабильностью in vivo. Модификации после SELEX, производимые с лигандами, представляющими собой нуклеиновые кислоты, с 2'-гидроксилом (2'-OH), могут приводить к улучшенной стабильности in vivo без неблагоприятного воздействия на связывающую способность лигандов, представляющих собой нуклеиновые кислоты. В одном из вариантов осуществления модификации лигандов включают 3-3' инвертированную фосфодиэфирную связь на 3'-конце молекулы, и модификации 2'-фтором (2'-F), 2'-амино (2-NH2), 2'-дезокси и/или 2'-O-метилом (2'-OMe) некоторых или всех нуклеотидов.
Лиганды, описываемые в настоящем документе, исходно получали посредством SELEX с использованием библиотек транскриптов, в которых остатки C и U являлись 2'-фторзамещенными, а остатки A и G содержали 2'-OH. Хотя такие модификации позволяли получать молекулы лигандов, подходящие для скрининга, высокое содержание 2'-гидроксилов делало их неподходящими для получения кандидатов в лекарственные средства вследствие того факта, что эти положения могут являться очень чувствительными к разрушению нуклеазами in vivo, ограничивая максимальную концентрацию, которую можно достигнуть после парентерального введения, а также их время полужизни в циркуляции. Таким образом, после идентификации функциональных последовательностей, например, способом SELEX, отдельные остатки можно тестировать на устойчивость к заменам, оценивая воздействие этих замен на структуру, функцию и стабильность лигандов.
В определенных вариантах осуществления нуклеиновые кислоты, составляющие лиганд, содержат модифицированные сахара и/или модифицированные основания. В определенных вариантах осуществления модификации включают стабилизирующие модификации, такие как стабилизирующие 2' модификации. В одном из вариантов осуществления стабилизирующие 2' модификации могут включать модификации 2'-фтор, 2'-дезокси или 2'-O-метил в кольце сахара.
В одном из вариантов осуществления конструирование включает снижение содержания 2'-гидроксилов в лиганде или модуляторе, или в обоих. В другом варианте осуществления конструирование включает снижение содержания 2'-фтора в лиганде или модуляторе, или в обоих. В другом варианте осуществления конструирование включает снижение содержания 2'-O-метила в лиганде или модуляторе, или в обоих.
Олигонуклеотид может содержать по меньшей мере одну молекулу модифицированного основания, выбранную из группы, в качестве неограничивающих примеров включающей 5-фторурацил, 5-бромурацил, 5-хлорурацил, 5-йодурацил, гипоксантин, ксантин, 4-ацетилцитозин, 5-(карбоксигидроксиметил)урацил, 5-карбоксиметиламинометилтиоуридин, 5-карбоксиметиламинометилурацил, дигидроурацил, бета-D-галактозилквеозин, инозин, N6-изопентениладенин, 1-метилгуанин, 1-метилинозин, 2,2-диметилгуанин, 2-метиладенин, 2-метилгуанин, 3-метилцитозин, 5-метилцитозин, N6-аденин, 7-метилгуанин, 5-метиламинометилурацил, 5-метоксиаминометил-2α-тиоурацил, β-D-маннозилквеозин, 5'-метоксикарбоксиметилурацил, 5-метоксиурацил, 2-инетилтио-N&изопентениладенин, урацилоксиуксусную кислоту, вайбутоксозин, псевдоурацил, квеозин, 2-тиоцитозин, 5-метилтиоурацил, 2-тиоурацил, 4-тиоурацил, 5-метилурацил, метиловый эфир -урацил-5-оксиуксусной кислоты, урацилоксиуксусную кислоту (v), 5-метилтиоурацил, 3-(3-амино-3-N-карбоксипропил) и 2,6-диаминопурин.
Олигонуклеотиды описанных в настоящем документе лигандов и модуляторов могут содержать модифицированные группы сахаров, например, одну или несколько гидроксильных групп замещают галогеном, алифатическими группами или функционализируют в виде простых эфиров или аминов. В одном из вариантов осуществления положение 2' фуранозного остатка замещают любым из O-метила, O-алкила, O-аллила, S-алкила, S-аллила или галогеновой группы. В другом варианте осуществления лиганд, представляющий собой нуклеиновую кислоту, или модулятор по изобретению могут содержать по меньшей мере одну модифицированную молекулу сахара, выбранную из группы, включающей в качестве неограничивающих примеров, арабинозу, 2-фторарабиноза, ксилулозу, гексозу, 2'-фторрибозу, 2'-O-метилрибозу, 2'-O-метоксиэтилрибозу, 2-O-пропилрибозу, 2'-O-метилтиоэтилрибозу, 2'-O-диэтиламинооксиэтилрибозу, 2'-O-(3-аминопропил)рибозу, 2'-O-(диметиламинопропил)рибозу, 2'-O-(метилацетамидо)рибозу и 2'-O-(диметиламиноктилоксиэтил)рибозу.
Лиганд или модулятор могут содержать по меньшей мере один модифицированный фосфатный остов, выбранный из группы, включающей в качестве неограничивающих примеров тиофосфат, фосфородитиоат, фосфорамидотиоат, фосфорамидат, фосфородиамидат, метилфосфонат, алкилфосфотриэфир и формацеталь или их аналог.
Молекулу лиганда, содержащую стеблевые и петлевые структуры, можно дополнительно стабилизировать для терапевтического применения заменой одой или нескольких петлевых структур нуклеиновой кислоты более стабильной петлевой структурой. Например, для лигандов GPVI, описываемых в настоящем документе, обнаружено, что замена петли 2 гексаэтиленгликолевым спейсером позволяет получать лиганды GPVI со сходной аффинностью и антилиганды GPVI с нуклеотидной петлей. На фиг. 13A представлен исходный фосфорамидит для гексаэтиленгликолевого линкера, используемого при синтезе. На фиг. 13B представлен гексаэтиленгликолевый спейсер встроенный между двумя нуклеотидами лиганда, представляющего собой нуклеиновую кислоту.
В фармацевтических композициях лиганды можно предоставлять в таких формах, как в формах солей, которые улучшают растворимость или биодоступность.
Любой из олигонуклеотидов по изобретению можно синтезировать стандартными известными в данной области способами, например, с использованием автоматического ДНК-синтезатора (такого, как коммерчески доступный, например, в Biosearch, Applied Biosystems).
Лиганды и модификаторы описаны в настоящем документе с использованием сокращений легко понятных специалистам в данной области и означающих следующее: "rA" представляет собой 2'-OH-A или аденозин; "A" представляет собой 2'-дезокси-A или 2'-дезоксиаденозин; "мА" представляет собой 2'-O-метил-A или 2'-метокси-2'-дезоксиаденин; "rG" представляет собой 2-OH-G или гуанозин; "G" представляет собой 2'-дезокси-G или 2'-дезоксигуанозин; "mG" представляет собой 2'-O-метил-G или 2'-метокси-2'-дезоксигуанозин; "fC" представляет собой 2'-фтор-C или 2'-фтор-2'дезоксицитидин; "mC" представляет собой 2'-O-метил-C или метокси-2'-дезоксицитидин; "fU" представляет собой 2'-фтор-U или 2'-фторуридин; "mU" представляет собой 2'-O-метил-U или 2'-метоксиуридин и "iT" представляет собой инвертированный 2'-H-T, (C6L) представляет собой гексиламинолинкер; (6GLY) представляет собой гексаэтиленгликолевый спейсер; (PEG40KGL2-NOF) представляет собой разветвленный PEG массой приблизительно 40 кДа (№ продукта SUNBRIGHT™ GL2-400GS2), (6FAM) представляет собой 6-карбоксифлуоресцеин; (s) представляет собой тиофосфатную связь между двумя нуклеотидами.
Связывание с носителем
Лиганды GPVI также могут содержать модификации, которые улучшают биодоступность или стабильность. Такие модификации могут включать конъюгацию с молекулой-носителем, которая в качестве неограничивающих примеров может содержать гидрофильную или гидрофобную группу. Одним из примеров являются молекулы полиэтиленгликоля, конъюгированные с последовательностью нуклеиновой кислоты. Конъюгация, например, с полимером, как описано ниже, может ограничить распределение плазматическим компартментом и увеличить время полужизни в циркуляции.
Модификации сахаров, как описано выше, могут обеспечивать стабильность, но они не гарантируют лигандам, представляющим собой нуклеиновые кислоты, подходящую фармакокинетику для терапевтической активности. У здоровых индивидуумов лиганды выводятся из плазмы в пределах нескольких минут после в/в инъекции, вероятно, посредством выведения почками. Нахождения интактных лигандов в крови на протяжении от нескольких часов до нескольких суток после инъекции достигают посредством их конъюгации с более крупными макромолекулами, такими как полиэтиленгликоль (PEG). Выведение лигандов из плазмы также снижают посредством введения их в липосомы.
Таким образом, в одном из вариантов осуществления лиганд GPVI, представляющий собой нуклеиновую кислоту, или модулятор лиганда GPVI можно ковалентно связывать или иным образом связывать с неиммуногенным, высокомолекулярным соединением, таким как полиэтиленгликоль (PEG) или другой водорастворимый фармацевтически приемлемый полимер, включая в качестве неограничивающих примеров полиаминоамины (PAMAM); полисахариды, такие как декстран, или полиоксазолины (POZ). Лиганд GPVI, представляющий собой нуклеиновую кислоту, или модулятор лиганда GPVI можно связывать с высокомолекулярным соединением ковалентными связями. Когда используют ковалентное присоединение, высокомолекулярное соединение можно ковалентно связывать с рядом положений на лиганде или модуляторе. В определенных вариантах осуществления лиганд или модулятор для введения нуждающемуся в этом индивидууму можно инкапсулировать в липосоме.
В одном из вариантов осуществления лиганд или модулятор присоединен к полиэтиленгликолю (PEG). Полиэтиленгликоли (PEGs) можно конъюгировать с биологически активными соединениями для того, чтобы они служили в качестве "инертных" носителей для потенциального (1) продления времени полужизни соединения в циркуляции, (2) изменения профиля распределения соединения и/или (3) маскировки соединения, таким образом, уменьшая его иммуногенный потенциал и защищая его от ферментативного расщепления.
Лиганд или модулятор можно присоединять к молекуле PEG посредством ковалентных связей. Например, олигонуклеотидный лиганд или модулятор можно связывать с 5'-тиолом посредством малеинимидной или винилсульфоновой функциональных групп.
Как правило, активированный PEG и другие активированные водорастворимые полимеры активируют подходящей активирующей группой, подходящей для связывания с желаемым участком на терапевтическом средстве. Характерные полимерные реагенты и способы конъюгации этих полимеров с активным средством известны в данной области и дополнительно описаны, например, в Zalipsky, S., et al., "Use of Functionalized Poly(Ethylene Glycols) for Modification of Polypeptides" in Polyethylene Glycol Chemistry: Biotechnical and Biomedical Applications, J. M. Harris, Plenus Press, New York (1992); и в Zalipsky, Advanced Drug Reviews, 1995, 16:157-182. Такие реагенты также являются коммерчески доступными.
Например, в одном из подходов для получения связанного амидом конъюгата, проводят реакцию водорастворимого полимера, несущего активированный сложный эфир, такой как сложный эфир NHS, например, mPEG-сукцинимидил-α-метилбутаноат, с аминогруппой активного средства, таким образом, получая амидную связь между активным средством и водорастворимым полимером. Дополнительные функциональные группы, способные к реакции с реактивными аминогруппами, в числе прочих включают, например, сложные N-гидроксисукцинимидильные эфиры, п-нитрофенилкарбонаты, сукцинимидилкарбонаты, альдегиды, ацетали, N-кетопиперидоны, малеинимиды, карбонилимидазолы, азалактоны, циклические имидотионы, изоцианаты, изотиоцианаты, трезилхлорид и галогенформиаты.
В одном из вариантов осуществления с одной молекулой PEG можно связывать несколько лигандов GPVI или модуляторов лигандов GPVI. Лиганды и модуляторы могут являться одинаковыми или различными последовательностями и модификациями. В еще одном дополнительном варианте осуществления несколько молекул PEG можно связывать друг с другом. В этом варианте осуществления с каждой молекулой PEG можно связывать один или несколько лигандов GPVI или модуляторов лигандов GPVI к одной и той же последовательности-мишени белка GPVI или разным последовательностям-мишеням белка GPVI. В вариантах осуществления, где с PEG связывают несколько лигандов или модуляторов, специфичных к одной и той же мишени, существует возможность приведения одинаковых мишеней в непосредственную близость друг с другом с обеспечением специфичных взаимодействий между одинаковыми мишенями. Когда с PEG связывают несколько лигандов или модуляторов, специфичных к различным мишеням, существует возможность приведения в непосредственную близость друг с другом различных мишеней с обеспечением специфичных взаимодействий между мишенями.
Хотя ряд линкеров и способов конъюгации гидрофильных молекул, таких как молекулы PEG, хорошо известны специалистам в данной области, ниже предоставлены несколько вариантов осуществления. В одном из вариантов осуществления можно использовать аминолинкер, такой как C6-гексиламинолинкер, 6-(трифторацетамидо)гексанол-(2-цианоэтил-N,N-диизопропил)фосфорамидит, представленный на фиг. 14, с добавлением гексиламинолинкера на 5'-конец синтезированного олигонуклеотида. Другие линкерные фосфорамидиты, которые можно использовать для добавления линкеров к синтезированным олигонуклеотидам, описаны ниже:
TFA-амино-C4-CED-фосфорамидит (доступный в ChemGenes, каталожный № CLP-1453) структуры:
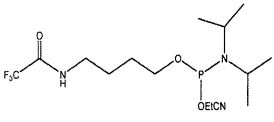 ,
,
5'-аминомодификатор C3-TFA (доступный в Glen Research, каталожный № 10-1923-90) структуры:
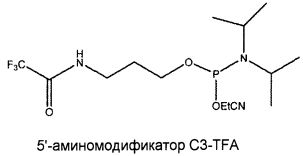 ,
,
5'-аминомодификатор 5 (доступный в Glen Research, каталожный № 10-1905-90) структуры:
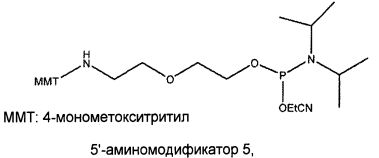
5'-аминомодификатор C12 (доступный в Glen Research, каталожный № 10-1912-90) структуры:
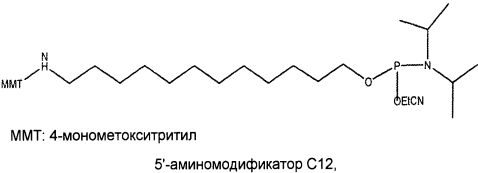
5'-тиолмодификатор C6 (доступный в Glen Research, каталожный № 10-1926-90) структуры:

Модифицированный 5'-тиолом линкер применяют, например, с PEG-малеимидами, PEG-винилсульфоном, PEG-йодацетамидом и PEG-ортопиридилдисульфидом.
Размер PEG может находиться в диапазоне от 5 до 200 кДа, с типичными PEG, используемые в фармацевтических составах в диапазоне 10-60 кДа. Можно получать PEG с линейными цепями массой приблизительно до 30 кДа. Для получения PEG более 30 кДа, можно связывать вместе несколько PEG (PEG с несколькими плечами или "разветвленные" PEG) с получением PEG желаемого размера. Общий синтез соединений с получением разветвленного соединения "mPEG2" (два mPEG связанные посредством аминокислот) описан в Monfardini, et al., Bioconjugate Chem. 1995, 6:62-69. Для получения "разветвленных" PEG, т.е. соединений, содержащей более одного PEG или mPEG, связанных с общей реакционноспособной группой, PEG или mPEG можно связывать вместе посредством аминокислоты, такой как лизин, или их можно связывать, например, посредством глицерина. Для разветвленных PEG, в которых каждый mPEG составляет приблизительно 10, приблизительно 20, или приблизительно 30 кДа, общая масса составляет приблизительно 20, приблизительно 40 или приблизительно 60 кДа и соединение указывают по его общей массе (т.е. mPEG2 массой 40 кДа представляет собой два связанных mPEG массой 20 кДа). PEG с общей молекулярной массой 40 кДа, которые можно использовать в качестве реагентов при получении пегилированного соединения, включают, например, N-гидроксисукцинимид [N 2-(монометокси-полиэтиленгликоль (20 кДа)-карбамоил)-N 6-(монометокси-полиэтиленгликоль (20 кДа)-карбамоил)]лизина структуры:
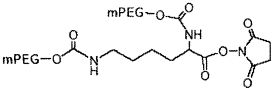 .
.
Дополнительные реагенты PEG, которые можно использовать для получения стабилизированных соединений по изобретению, включают другой разветвленный PEG-N-гидроксисукцинимид (mPEG-NHS) общей формулы:
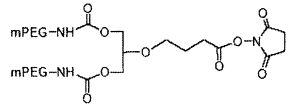

c общей молекулярной массой 40 кДа или 60 кДа (где масса каждого mPEG составляет приблизительно 20 или приблизительно 30 кДа). Как описано выше, разветвленные PEG можно связывать посредством любого подходящего реагента, такого как аминокислота, и в определенных вариантах осуществления они связаны посредством остатков лизина или остатков глицерина.
Они также могут включать неразветвленный mPEG-сукцинимидилпропионат (mPEG-SPA) общей формулы:
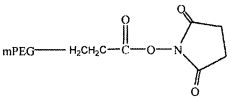 ,
,
в котором масса mPEG составляет приблизительно 20 кДа или приблизительно 30 кДа. В конкретном варианте осуществления реакционноспособный сложный эфир представляет собой -O-CH2CH2-CO2-NHS.
Реагенты также могут включать разветвленный PEG связанный посредством глицерина, такой как серия Sunbright™ из NOF Corporation, Япония. Конкретные, неограничивающие примеры этих реагентов представляют собой:
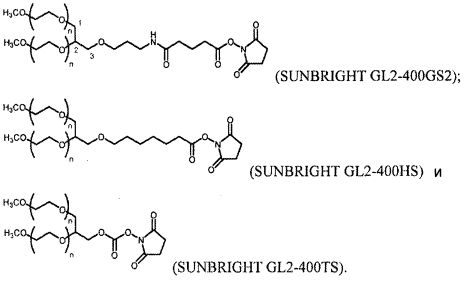
Реагенты также могут включать неразветвленный сукцинимидил-альфа-метилбутаноат (mPEG-SMB) общей формулы:
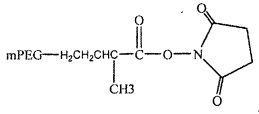 ,
,
в котором масса mPEG составляет от 10 до 30 кДа. В подварианте осуществления, реакционноспособный сложный эфир представляет собой -O-CH2CH2CH(CH3)-CO2-NHS. Соединения этой структуры продаются в Nektar Therapeutics с каталожными номерами № 2M4K0R01.
Реагенты PEG также могут включать PEG, связанные с нитрофенилкарбонатом, например, следующей структуры:
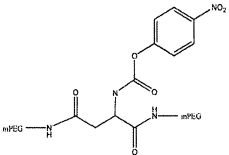 .
.
Соединения этой структуры коммерчески доступны, например, в Sunbio, Inc. Соединения, содержащие нитрофенилкарбонат можно конъюгировать с содержащими первичные амины линкерами. В этой реакции O-нитрофенил служит в качестве уходящей группы, оставляя структуры [mPEG]nNH-CO-NH-линкер-лиганд.
PEG с реагирующими с тиолом группами, которые можно использовать с модифицированным тиолом линкером, как описано выше, включают соединения общей структуры: 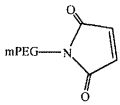 , в которой масса mPEG составляет приблизительно 10, приблизительно 20 или приблизительно 30 кДа. Кроме того, структура может быть разветвленной, такой как
, в которой масса mPEG составляет приблизительно 10, приблизительно 20 или приблизительно 30 кДа. Кроме того, структура может быть разветвленной, такой как 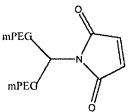 , в которой масса каждого mPEG составляет приблизительно 10, приблизительно 20 или приблизительно 30 кДа, и общая масса составляет приблизительно 20, приблизительно 40 или приблизительно 60 кДа. Разветвленные PEG с реагирующими с тиолами группами, которые можно использовать с модифицированным тиолом линкером, как описано выше, включают соединения, в которых общая масса разветвленного PEG составляет приблизительно 40 или 60 кДа (где масса каждого mPEG составляет 20 или 30 кДа). Реагенты PEG также могут представлять собой следующую структуру:
, в которой масса каждого mPEG составляет приблизительно 10, приблизительно 20 или приблизительно 30 кДа, и общая масса составляет приблизительно 20, приблизительно 40 или приблизительно 60 кДа. Разветвленные PEG с реагирующими с тиолами группами, которые можно использовать с модифицированным тиолом линкером, как описано выше, включают соединения, в которых общая масса разветвленного PEG составляет приблизительно 40 или 60 кДа (где масса каждого mPEG составляет 20 или 30 кДа). Реагенты PEG также могут представлять собой следующую структуру:  . PEG-малеинимид пегилирует тиолы соединения-мишени, где двойная связь малеимидного кольца разрушается для связи с тиолом. Скорость реакции зависит от pH и в одном из вариантов осуществления ее проводят при pH от 6 до 10, или при pH от 7 до 9 или при pH приблизительно 8.
. PEG-малеинимид пегилирует тиолы соединения-мишени, где двойная связь малеимидного кольца разрушается для связи с тиолом. Скорость реакции зависит от pH и в одном из вариантов осуществления ее проводят при pH от 6 до 10, или при pH от 7 до 9 или при pH приблизительно 8.
В одном из вариантов осуществления с одной молекулой PEG можно связывать несколько модуляторов лигандов GPVI. Модуляторы могут быть к одному и тому же или к различным лигандам GPVI, представляющим собой нуклеиновые кислоты. В вариантах осуществления, где присутствуют несколько модуляторов к одному и тому же лиганду, существует увеличение авидности вследствие множественного связывающего взаимодействия с лигандом. В еще одном дополнительном варианте осуществления можно связать друг с другом несколько молекул PEG. В этом варианте осуществления с каждой молекулой PEG можно связать один или несколько модуляторов к одному и тому же лиганду, представляющему собой нуклеиновую кислоту, или к различным лигандам. Это также приводит к увеличению авидности каждого модулятора к его мишени.
В одном из вариантов осуществления лиганд, представляющий собой нуклеиновую кислоту, или его модулятор можно ковалентно связывать с липофильным соединением, таким как холестерин, диалкилглицерин или диацилглицерин. Липофильное соединение или неиммуногенное, высокомолекулярное соединение можно ковалентно связывать или связывать посредством нековалентных взаимодействий с лигандом или модулятор(ами). Связывание лиганда или олигонуклеотидного модулятора с липофильными или неиммуногенными высокомолекулярными соединениями можно проводить непосредственно или с использованием линкеров или спейсеров.
В вариантах осуществления, где используют непосредственное ковалентное связывание, липофильное соединение или неиммуногенное высокомолекулярное соединение можно ковалентно связывать с рядом положений на лиганде или модуляторе, такими как экзоциклическая аминогруппа на основании, положение 5 пиримидинового нуклеотида, положение 8 пуринового нуклеотида, гидроксильная группа фосфата или гидроксильная группа или другая группа на 5'- или 3'-конце.
В вариантах осуществления, где лиганд или модулятор присоединен к липофильному или неиммуногенному высокомолекулярному соединению посредством линкера или спейсера, липофильное соединение или неиммуногенное высокомолекулярное соединение можно связывать с лигандом или модулятором с использованием, например, аминолинкера с шестью атомами углерода.
В другом варианте осуществления между линкером и последовательностью нуклеиновой кислоты можно включать одну или несколько фосфатных групп.
Дополнительные подходящие линкеры и спейсеры для присоединения лиганда или модулятора к липофильному соединению или к неиммуногенному высокомолекулярному соединению описаны в патенте США № 7531524, включенном в настоящий документ в качестве ссылки.
Олигонуклеотиды по изобретению можно модифицировать по остатку основания, остатку сахара или фосфатному остову, например, для улучшения таких свойств, как стабильность молекулы и аффинность к указанной мишени.
G. Способы лечения опосредованных тромбоцитами нарушений
Тромбоциты содержат два физиологически важных рецептора коллагена, GPVI и интегрин α2β1. Из них активацию тромбоцитов в ответ на коллаген опосредует GPVI. Активация тромбоцитов при взаимодействии коллагена с GPVI приводит к высвобождению из тромбоцитов содержимого плотных и альфа-гранул. Содержимое гранул включает ряд агонистов тромбоцитов, а также провоспалительные цитокины, факторы роста, молекулы адгезии и другие белки, включая: АТФ, ОТФ, АДФ, ГДФ, полифосфат, CD63, LAMP2, серотонин, тромбоцитарный фактор 4, β-тромбоглобулин, MIP-1α, RANTES, MCP-3, CCL17, CXCL1, CXCL5, IL-8, BFGF, EGF, HGF, инсулиноподобный фактор роста 1, TGF-β, VEGF-A, VEGF-C, PDGF, P-селектин, vWF, тромбоспондин, фибриноген, интегрины αIIβ3 и αvβ3, фибронектин, альбумин, α1-трипсин, Gas6, богатый гистидином гликопротеин, высокомолекулярный кининоген и белок-предшественник бета-амилоида. Таким образом, активация тромбоцитов посредством взаимодействия GPVI с коллагеном локально создает провоспалительное окружение, способное к стимуляции широкого диапазона находящихся рядом типов клеток.
Взаимодействие GPVI с коллагеном также необходимо для адгезии тромбоцитов к пораженной или поврежденной стенке сосуда. Наконец, активация GPVI коллагеном запускает агрегацию тромбоцитов. Активация GPVI коллагеном играет важную роль в гемостазе, когда повреждение сосуда экспонирует сосудистый коллаген, таким образом, приводя к формированию тромбоцитарной пробки. Хотя роль тромбоцитов при тромбозе хорошо известна и опосредована в основном взаимодействиями GPVI с коллагеном типов I и III, присутствующим в субэндотелиальном матриксе и в больших количествах присутствующим в атеросклеротических бляшках, недавние данные указывают на то, что провоспалительный ответ тромбоцитов при активации в дополнение к тромботическим заболеваниям может лежать в основе большого количества болезненных состояний, включая атеросклероз, диабетическое заболевание сосудов, ревматоидный артрит и склеродермия. Например, взаимодействие GPVI с коллагеном может приводить к болезненным состояниям, где экспрессия GPVI является аномально высокой, или где тромбоциты экспонированы для коллагена на патофизиологических уровнях, или где тромбоциты экспонированы для коллагена с аномальным распределением. Таким образом, в настоящем документе предоставлены способы лечения опосредованных тромбоцитами заболеваний или нарушений с использованием терапевтически эффективных количеств лигандов GPVI. Эти лиганды GPVI могут функционировать посредством связывания с тромбоцитами и ингибирования их активации.
Для понимания заболеваний, в которых могут давать терапевтический эффект лекарственные средства против GPVI, необходимо понимать только специфичность GPVI к различным типам коллагена, находящимся в организме, так как для активации тромбоцитов коллагеном необходим коллаген, способный взаимодействовать с GPVI. Существует 29 различных типов коллагена. Из них 9 идентифицированы, как экспрессирующиеся в сосудистой системе, включая типы I, III, IV, V, VI, VIII, XII, XIII и XIV. Кроме того, из них 7 являются фибриллярными и могут собираться в стабильные тройные спирали и фибриллярные структуры более высокого порядка, и они включают типы I, II, III, IV, V, XI, XXIV и XXVII (Nieswandt et al., Blood, 2003, 102:449-461; Herr et al., J. Biol. Chem., 2009, 284:19781-19785). Показано, что формирующие фибриллы коллагены типов I, II, III и IV, поддерживают активацию, агрегация и адгезия тромбоцитов, тогда как нефибриллярные коллагены типов VI, VII и VIII, индуцируют только слабую адгезию без агрегации тромбоцитов. Таким образом, коллаген типов I-IV может взаимодействовать с GPVI со специфической активацией тромбоцитов. Кроме того, показано, что GPVI специфически связывается с коллагеном типов I-IV (Jung et al., Platelets, 2008; 19:32-42). Таким образом, заболевания при которых происходит высокая экспрессия коллагена типов I-IV или при которых происходит аномальная локализация или презентирование этих типов коллагена тромбоцитам, можно лечить лигандами GPVI.
Структурные требования для связывания GPVI коллагеном и родственными коллагену пептидами хорошо известны (см. пример 6; а также Smethurst et al., J. Biol. Chem., 2007, 282:1296-1304; Smethurst et al., Blood, 2004, 103:903-911; Horii et al., Blood, 2008:936-942). Одним из первичных участков распознавания GPVI в коллагеновых фибриллах является трипептидная последовательность GPO. Кроме того, связывающие коллаген бороздки в GPVI разделены приблизительно 5,5 нм. Таким образом, полагают, что другие типы коллагена, содержащие повторы GPO и формирующие фибриллярные структуры, способные экспонировать повторы GPO с дистанцией ≈5,5 нм, так что они способны к связыванию димеров GPVI, активируют тромбоциты посредством GPVI, приводя к заболеваниям, поддающимся лечению лигандами GPVI.
Как правило, GPVI на тромбоцитах экспрессирован на уровнях от низкого до умеренного, с обычным количеством рецепторов ≈1200/тромбоцит. Однако увеличенные уровни экспрессии GPVI могут делать тромбоциты гиперреактивными в отношении коллагена и, таким образом, предрасполагать индивидуумов к заболеванию, или непосредственно вызывать состояние болезни, опосредованное взаимодействием тромбоцитов-коллагена. Сверхэкспрессия GPVI связана с началом тромботических сосудистых заболеваний, включая транзиторные ишемические атаки (TIA) и острые коронарные синдромы (Bigalke et al., Thromb. Res., 2010:125:e184-189; Bigalke et al., Int. J. Cardiol., 2009, Jan 11 epub; Bigalke et al., Clin. Res. Cardiol., 2010, 99:227-233). Таким образом, лечение цереброваскулярных событий, таких как TIA и инсульт или лечение острых коронарных синдромов лигандами GPVI, может приводить к терапевтическому эффекту.
В основе роли тромбоцитов в гемостазе и воспалительных реакциях лежат общие молекулярные механизмы. Например, при возникновении повреждения сосудистого эндотелия, такого как при диабете и при атеротромботическом заболевании сосудов, существует высокий уровень активации тромбоцитов GPVI. Это приводит к высвобождению цитокинов и активации, локализации и созреванию лейкоцитов. Возникновение и сохранение этого процесса при наличии атеросклеротического заболевания сосудов также приводит к адгезии тромбоцитов к поверхности эндотелия, дестабилизации бляшек и приводит к дальнейшему повреждению сосуда. Эта комбинация событий может приводить к ишемии в сосуде. Таким образом, терапевтические средства, такие как лиганд GPVI, предотвращающие или снижающие нежелательную активацию тромбоцитов, можно использовать для лечения ряда заболеваний, для которых известно, что они ассоциированы, например, с повышением экспрессии GPVI на тромбоцитах или с увеличенной активацией тромбоцитов при увеличенном воздействии коллагена.
Диабет ассоциирован с усиленной опосредованной коллагеном активацией тромбоцитов, и экспрессия GPVI у индивидуумов с диабетом значимо выше по сравнению с индивидуумами без диабета (Cabeza et al., Diabetes, 2004, 53:2117-2121). Высокая экспрессия GPVI у больных диабетом в значительной степени способствует тромбоишемическим осложнениям, ассоциированным с диабетом, и, таким образом, лекарственные средства против GPVI при лечении связанных с диабетом тромбоишемических заболеваний могут обеспечивать терапевтический эффект. Активация GPVI в значительной степени увеличивает поверхностную экспрессию CD40L. CD40L представляет собой мощный цитокин, высвобождаемый из тромбоцитов, вовлеченный в тромбоз и атеросклероз. Кроме того, сверхэкспрессия CD40L в активированных GPVI тромбоцитах усиливает экспрессию на поверхности эндотелия CD62P, αvβ3, молекулы межклеточной адгезии I и секрецию моноцитарного хемотаксического белка 1. Эти результаты указывают на то, что функция рецепторов коллагена GPVI при диабете типа 2 изменена и может играть важную роль в диабетических атеротромботических осложнениях и локализованном заболевании сосудов.
Таким образом, лиганды GPVI можно использовать для лечения ряда опосредованных тромбоцитами нарушений, как правило, ассоциированных с диабетом. В одном из вариантов осуществления предоставлен способ лечения индивидуума, страдающего диабетом, включающий введение лиганда GPVI. Лечение пациентов с высоким риском диабета лигандами GPVI может ослаблять или предотвращать ассоциированные с диабетом нарушения у этих пациентов. Эти нарушения в качестве неограничивающих примеров включают диабетическую ретинопатию, диабетическую васкулопатию, атеросклероз, ишемический инсульт и хроническую почечную недостаточность. Лечение больных диабетом лигандами GPVI также может снижать или ингибировать у этих пациентов формирование микротромбов.
Также предоставлен способ лечения индивидуумов, страдающих опосредованными тромбоцитами воспалительными нарушениями, такими как ревматоидный артрит (RA) или другие воспалительные артритические нарушения. Недавние исследования показали, что у людей, страдающих RA и другими формами воспалительного артрита присутствуют повышенные уровни тромбоцитарных микрочастиц в синовиальных жидкостях их суставов (Boilard et al., Science, 2010, 327:580-583). Тромбоцитарные микрочастицы являются провоспалительными и вызывают воспалительный ответ из клеток окружения (например синовиальные фибробласты). Например, связывание коллагена типа IV с GPVI приводит к высвобождению IL-1 и IL-8.
Исследования механизмов у мышей связывают провоспалительное состояние тромбоцитов с активацией GPVI. Отсутствие GPVI в модели на мышах с нокаутом предотвращало привлечение провоспалительных клеток и, таким образом, привлечение и созревание лейкоцитов в синовиальной оболочке и внеклеточном матриксе.
Воспаление суставов приводит к взаимодействию тромбоцитов с коллагеном во внеклеточном матриксе сустава, что приводит к развитию воспаления посредством опосредованной GPVI активации тромбоцитов близлежащих клеток и манифестации состояний RA/воспалительного артрита, поддающихся лечению лигандами GPVI. Таким образом, лиганды GPVI могут обеспечивать терапевтическое применение для облегчения, ослабления или предотвращения воспалительных заболеваний, таких как RA или другие воспалительные артриты, включая в качестве неограничивающих примеров подагру, псориатический артрит, реактивный артрит, вирусный или поствирусный артрит и спондилоартрит.
Лиганды GPVI также можно использовать для лечения индивидуумов, страдающих склеродермией или системным склерозом. По-видимому склеродермия возникает как аутоиммунный ответ, порождающий опухание (воспаление) мышц и суставов, ассоциированный со сверхпродукцией коллагена. Одним из основных процессов, вовлеченных в системный склероз или склеродермию, является микроповреждение сосудов. Взаимодействие тромбоцитарного рецептора коллагена типов I и III (GPVI) с соответствующим лигандом в доступной в результате постоянного микроповреждения сосудов субэндотелиальной строме у пациентов с системным склерозом приводит к активации и агрегации тромбоцитов с высвобождением провоспалительных медиаторов, которые способствуют повреждению сосудов и воспалению (Chiang et al., Thrombosis, 2006, 117:299-306). При системном склерозе, очаги повреждения сосудов характеризуются артериолярно-капиллярным периваскулитом с инфильтрацией мононуклеарных клеток, который приводит к пролиферации артериальной интимы и облитерации артериол и капилляров с потерей эндотелиальных клеток и базальной мембраны. Характеристикой системного склероза является рецидивирующий характер повреждения эндотелиальных клеток, или базальной мембраны, или обоих. Кроме того, этими событиями управляет сверхпродукция и накопление коллагена в тканях организма, приводящее к обширному уплотнению и рубцеванию тканей во всем организме. Таким образом, использование лигандов GPVI может обеспечивать терапевтическое ослабление такого заболевания, как склеродермия или системный склероз, которое ассоциировано с увеличенными уровнями коллагена и опосредуемым тромбоцитами микроповреждением сосудов. В настоящем документе предоставлен способ лечения индивидуума страдающего склеродермией посредством введения терапевтически эффективного количества лиганда GPVI.
Лиганды GPVI, описываемые в настоящем документе, также можно использовать для лечения индивидуумов, у которых диагностирована злокачественная опухоль. Недавние исследования позволяют предположить, что GPVI опосредует метастазирование опухолей (см., например, Jain et al., J. Thromb. Haemostasis, 2009, 7:1713-1717). Применяя экспериментальный анализ метастазирования in vivo, Jain et al., показали, что мыши с нокаутом GPVI демонстрировали значимое уменьшение метастазирования опухолей по сравнению с контрольными мышами дикого типа. Таким образом, в одном из вариантов осуществления предоставлен способ ингибирования, уменьшения или предотвращения метастазирования у индивидуума, у которого диагностировано наличие первичной злокачественной опухоли, где индивидууму вводят терапевтически эффективное количество лиганда GPVI.
Также предоставлены способы, фармацевтические композиции и применение лигандов GPVI, представляющих собой нуклеиновые кислоты, описываемых в настоящем документе, в качестве регулируемых антитромбоцитарных средств для применения при нарушениях или в схемах лечения, требующих антитромбоцитарной терапии. В определенных вариантах осуществления лечение представляет собой хирургическое вмешательство. Способы могут включать введение лиганда GPVI, представляющего собой нуклеиновую кислоту, нуждающемуся в этом индивидууму, где индивидуум страдает окклюзивным тромботическим заболеванием или нарушением коронарной, черепно-мозговой или периферической сосудистой системы, или подвержен риску их возникновения.
В одном из вариантов осуществления лиганд GPVI ингибирует инициацию активации тромбоцитов. В других вариантах осуществления лиганд GPVI ингибирует активацию тромбоцитов и формирующийся в результате тромбоцитарный провоспалительный ответ. В других вариантах осуществления лиганд GPVI ингибирует адгезию тромбоцитов. В других вариантах осуществления лиганд GPVI ингибирует агрегацию тромбоцитов. В еще одном дополнительном варианте осуществления лиганд GPVI ингибирует образование тромбина.
В одном из вариантов осуществления индивидуум страдает окклюзивным тромботическим заболеванием коронарной, черепно-мозговой и периферической сосудистых систем или подвержен риску их возникновения. В других определенных вариантах осуществления индивидуума готовят подвергнуть или подвергают хирургическому вмешательству, или его подвергли хирургическому вмешательству, подвергающему индивидуума риску окклюзивного тромботического осложнения. В других вариантах осуществления у индивидуума пересадили трансплантат сосуда для обеспечения гемодиализа, который подвержен риску окклюзии вследствие взаимодействия сосуда и тромбоцитов.
В определенных вариантах осуществления предоставлен способ лечения или предотвращения образования сосудистых осложнений, в частности тромботического или тромбоэмболического осложнения, включающий введение лиганда GPVI, представляющего собой нуклеиновую кислоту, по изобретению нуждающемуся в этом индивидууму.
В одном из вариантов осуществления лиганд GPVI, представляющий собой нуклеиновую кислоту, вводят в течение продолжительных периодов времени. В этом случае, модулятор лиганда GPVI можно применять только в экстремальных ситуациях, например, если лечение приводит к кровотечению, включая внутричерепное или желудочно-кишечное кровотечение. В другом варианте осуществления модулятор вводят, когда пациентам, получающим лечение лигандом GPVI, представляющим собой нуклеиновую кислоту, необходимо срочное хирургическое вмешательство. В другом варианте осуществления модулятор вводят для контроля концентрации лиганда GPVI, представляющего собой нуклеиновую кислоту, и, таким образом длительности и интенсивности лечения. В другом варианте осуществления лиганд GPVI, представляющий собой нуклеиновую кислоту, предоставляют в качестве тромбоцитарного анестетика в ходе процедуры искусственного кровообращения. В другом варианте осуществления лиганд GPVI, представляющий собой нуклеиновую кислоту, вводят для обеспечения периода перехода с или на пероральные антитромбоцитарные лекарственные средства, а модулятор применяют для обращения действия лиганда GPVI, представляющего собой нуклеиновую кислоту, после установления терапевтических уровней перорального антитромбоцитарного средства.
H. Фармацевтические композиции
Лиганды GPVI, представляющие собой нуклеиновые кислоты, или модуляторы лигандов GPVI, указанные в настоящем документе, можно формулировать в фармацевтические композиции, которые в качестве неограничивающих примеров могут включать фармацевтически приемлемый носитель, разбавитель или эксципиент. Точные характеристики композиции будут зависеть, по меньшей мере частично, от характера лиганда и/или модулятора, включая любые стабилизирующие модификации, и маршрута введения. Композиции, содержащие модулятор, можно разрабатывать для введения индивидууму, которому вводили лиганд GPVI, представляющий собой нуклеиновую кислоту, для обеспечения модуляции активности лиганда и, таким образом, регуляции антитромбоцитарной активности вводимого лиганда GPVI, представляющего собой нуклеиновую кислоту.
Разработка и получение фармацевтических или фармакологических композиций будет понятна специалистам в данной области с учетом настоящего описания. Как правило, такие композиции можно получать в виде инъецируемых композиций, в виде жидких растворов или суспензий; твердых форм подходящих для растворения или суспендирования в жидкости до инъекции; в виде таблеток или других твердых форм для перорального введения; в виде капсул с замедленным высвобождением, в виде жидких форм для перорального введения; в виде эликсиров, сиропов, суппозиториев, гелей или в любой другой форме, применяемой в данной области, включая глазные капли, кремы, лосьоны, мази, ингаляционные формы и т.п. Также особенно пригодным может являться использование хирургами, терапевтами или патронажными медицинскими работниками стерильных составов, таких как промывания на основе физиологического раствора для лечения конкретной области в оперативном поле. Композиции также можно формулировать для доставки посредством микроустройства, микрочастицы или тампона.
Фармацевтически пригодные композиции, содержащие лиганд GPVI, представляющий собой нуклеиновую кислоту, или модулятор лиганда GPVI по настоящему изобретению, можно формулировать, по меньшей мере частично, посредством смешивания с фармацевтически приемлемым носителем. Примеры таких носителей и способы составления можно найти в Remington: The Science and Practice of Pharmacy, 20th edition (Lippincott Williams & Wilkins, 2000) и Ansel et al., Pharmaceutical Dosage Forms and Drug Delivery Systems, 6th Ed. (Media, Pa.: Williams & Wilkins, 1995).
Подходящие фармацевтические эксципиенты включают стабилизаторы, антиоксиданты, средства регуляции осмоляльности, буферы и средства регуляции pH. Подходящие добавки включают физиологически биологически совместимые буферы (например, гидрохлорид трометамина), добавления хелатирующих средств (таких как, например, ЭДТА, DTPA или DTPA-бисамид) или хелатных комплексов с кальцием (например, как кальций DTPA, CaNaDTPA-бисамид) или, необязательно, добавления кальция или натриевых солей (например, хлорид натрия, хлорид кальция, аскорбат кальция, глюконат кальция или лактат кальция). Фармацевтические композиции по изобретению можно упаковывать для применения в жидкой форме или их можно лиофилизировать.
Для формирования фармацевтически приемлемой композиции, подходящей для эффективного введения, такие композиции должны содержать эффективное количество лиганда, представляющего собой нуклеиновую кислоту, или модулятора. Такие композиции могут содержать смеси более одного соединения. Как правило, композиции содержат приблизительно от 0,1% процента по массе (мас.%) приблизительно до 50 мас.%, приблизительно от 1 мас.% приблизительно до 25 мас.% или приблизительно от 5 мас.% до приблизительно 20 мас.% активного средства (лиганда или модулятора).
По настоящему документу предоставлены фармацевтические композиции для парентерального инъекционного введения, включая подкожные, внутримышечные или внутривенные инъекции и инфузии. Для парентерального введения желательны асептические суспензии и растворы. Когда желательно внутривенное введение, используют изотонические препараты, которые, как правило, содержат подходящие консерванты. Фармацевтические композиции могут являться стерилизованными и/или содержать вспомогательные средства, такие как консерваторы, стабилизаторы, смачивающие средства или эмульгаторы, стимуляторы растворения, соли для регуляции осмотического давления и/или буферы. Жидкие, особенно инъецируемые, композиции можно получать, например, посредством растворения, диспергирования и т.д. Активное соединение растворяют в фармацевтически чистом растворителе, например, таком как вода, буферная вода, физиологический раствор, 0,4% физиологический раствор, 0,3% глицин, гиалуроновая кислота, водная декстроза, глицерин, этанол и т.п., или смешивают с ним, таким образом, получая инъецируемый раствор или суспензию. Кроме того, можно формулировать твердые формы, подходящие для растворения в жидкости перед инъекцией.
Для содействия растворению средства в водной среде можно добавлять поверхностно-активное вещество в виде смачивающего средства. Поверхностно-активные вещества могут включать анионные детергенты, такие как лаурилсульфат натрия, диоктилсульфосукцинат натрия и диоктилсульфонат натрия. Можно использовать катионные детергенты, и они могут включать хлорид бензалкония или хлорид бензетония. Неионные детергенты, которые можно включать в состав в качестве поверхностно-активных веществ в качестве неограничивающих примеров включают лауромакрогол 400, полиоксилстеарат 40, полиоксиэтиленгидрогенизированное касторовое масло 10, 50 и 60, моностеарат глицерина, полисорбат 20, 40, 60, 65 и 80, сложный эфир сахарозы и жирной кислоты, метилцеллюлозу, карбоксиметилцеллюлозу и любой из детергентов-плюроников, таких как плюроник F68 и/или плюроник F127 (например, см. Strappe et al Eur. J. of Pharm. Biopharm., 2005, 61:126-133). Поверхностно-активные вещества могут присутствовать в препарате белка или его производного отдельно или в виде смеси в различных соотношениях.
Для перорального введения в форме таблетки или капсулы компонент активного лекарственного средства можно комбинировать с пероральным, нетоксичным фармацевтически приемлемым инертным носителем, таким как этанол, глицерин, вода и т.п. Кроме того, когда желательно или необходимо, в смесь также можно вводить подходящие связывающие средства, смазочные средства, дезинтегрирующие средства и красители. Подходящие связывающие средства в качестве неограничивающих примеров включают крахмал, желатин, природные сахара, такие как глюкоза или бета-лактоза, кукурузные подсластители, природные и синтетические камеди, такие как гуммиарабик, трагакант или альгинат натрия, карбоксиметилцеллюлоза, полиэтиленгликоль, воски и т.п. Смазочные средства, используемые в этих лекарственных формах, в качестве неограничивающих примеров включают олеат натрия, стеарат натрия, стеарат магния, бензоат натрия, ацетат натрия, хлорид натрия и т.п. Дезинтегрирующие средства в качестве неограничивающих примеров включают, крахмал, метилцеллюлозу, агар, бентонит, ксантановую камедь и т.п.
Для жидких форм, используемых при пероральном введении, компонент активного лекарственного средства можно комбинировать в подходящим образом ароматизированные суспендирующие или диспергирующие средства, такие как синтетические и природные камеди, например, трагакант, гуммиарабик, метилцеллюлоза и т.п. Другие диспергирующие средства, которые можно использовать, включают глицерин и т.п.
Местные препараты, содержащие компонент активного лекарственного средства, можно смешивать с рядом веществ-носителей, хорошо известных в данной области, например, таких как спирты, гель алоэ вера, аллантоин, глицерин, масла с витаминами A и E, минеральное масло, пропионат простого миристилового эфира PPG2 и т.п., с формированием, например, спиртовых растворов, местных очистителей, очищающих кремов, кожных гелей, кожных лосьонов и шампунях в кремовых или гелевых составах.
Соединения по настоящему изобретению также можно вводить в форме липосомных систем доставки, таких как малые однослойные везикулы, большие однослойные везикулы и многослойные везикулы. Липосомы можно составлять из ряда фосфолипидов, таких как холестерин, стериламин или фосфатидилхолины. Активные средства, вводимые непосредственно (например, отдельно) или в липосомных составах описаны, например, в патенте США № 6147204.
Соединения по настоящему изобретению также можно связывать с растворимыми, такими как направляемые носители лекарственных средств. Такие полимеры могут включать поливинил-пирролидон, пирановые сополимеры, полигидроксипропилметакриламидфенол, полигидроксиэтиласпартатамидофенол или полиэтиленоксидполилизин, замещенный пальмитоиловыми остатками. Кроме того, соединения по настоящему изобретению можно связывать (предпочтительно посредством ковалентной связи) с классом биологически разрушаемых полимеров, пригодных для получения лекарственного средства с контролируемым высвобождением, например, с полиэтиленгликолем (PEG), полимолочной кислотой, полиэпсилонкапролактоном, полиоксазолинами, полигидроксимасляной кислотой, сложных полиортоэфиров, полиацеталями, полидигидропиранами, полицианоакрилатами и поперечно-сшитых или амфипатических блок-сополимеров гидрогелей. Для увеличения и продления биодоступности с лигандами, представляющими собой нуклеиновые кислоты, можно связывать холестерин и сходные молекулы.
Липофильные соединения и неиммуногенные высокомолекулярные соединения, с которыми можно формулировать модуляторы по изобретению для применения в настоящем изобретении, и которые можно получать любым из различных способов, известных в данной области или разработанных впоследствии. Как правило, их получают из фосфолипида, например, дистеароилфосфатидилхолина, и они могут содержать другие вещества, такие как нейтральные липиды, например, холестерин, а также поверхностные модификаторы, такие как положительно заряженные (например, стериламиновые или аминоманнозные или аминоманнитные производные холестерина) или отрицательно заряженные (например, диацетилфосфат, фосфатидилглицерин) соединения. Многослойные липосомы можно получать общепринятым способом, т.е. нанося выбранный липид на внутреннюю стенку подходящего контейнера или сосуда посредством растворения липида в подходящем растворителе, а затем выпаривая растворитель, оставляя тонкую пленку внутри сосуда, или посредством сушки распылением. Затем в сосуд добавляют водную фазу циркуляционным движением или встряхиванием, что приводит к образованию MLV. Затем посредством гомогенизации, ультразвуковой обработки и экструзии (через фильтры) MLV можно формировать UV. Кроме того, UV можно формировать способами удаления детергента. В определенных вариантах осуществления настоящего изобретения, комплекс содержит липосому с направленным лигандом(ами), представляющим собой нуклеиновую кислоту, связанным с поверхностью липосомы и инкапсулированным терапевтическим или диагностическим средством. Предварительно сформированные липосомы можно модифицировать, связывая с лигандами, представляющими собой нуклеиновые кислоты. Например, катионную липосому связывают с нуклеиновой кислотой посредством электростатических взаимодействий. Альтернативно, в предварительно сформированные липосомы можно добавлять нуклеиновую кислоту, связанную с липофильным соединением, таким как холестерин, посредством чего холестерин связывается с мембраной липосом. Альтернативно, нуклеиновую кислоту можно связывать с липосомой при формировании липосомы.
В другом варианте осуществления составом, содержащим лиганд GPVI или модулятор лиганда GPVI способами, известными специалистам в данной области можно покрывать стент или медицинское устройство.
Также предусмотрены терапевтические наборы. Наборы содержат реагенты, активные средства и вещества, которые могут быть необходимы для практического осуществления указанных выше способов. Как правило, наборы, в подходящих средствах хранения, содержат фармацевтически приемлемый состав лиганда GPVI и/или модулятора лиганда GPVI. Набор может содержать единственное средство хранения и/или он может содержать несколько различных средств хранения для каждого соединения или каждой реакционной смеси или стадии.
I. Способы введения
Способы введения лигандов GPVI и/или модуляторов лигандов GPVI по настоящему изобретению индивидууму в качестве неограничивающих примеров включают парентеральное (посредством инъекции или постепенной инфузии в течение определенного времени), внутривенное, интрадермальное, внутрисуставное, внутрисиновиальное, интратекальное, внутриартериальное, внутрисердечное, внутримышечное, подкожное, внутриглазное, внутрикапсулярное, интраспинальное, интрастернальное, местное, посредством трансдермального пластыря, посредством ректального, вагинального или уретрального суппозитория, перитонеальное, чрескожное, посредством назального спрея, хирургического имплантата, внутреннего хирургического красителя, инфузионного насоса или посредством катетера. В одном из вариантов осуществления средство и носитель вводят в составе с замедленным высвобождением, таком как имплантат, болюс, микрочастица, микросфера, наночастица или наносфера. В одном из вариантов осуществления лиганд GPVI, представляющий собой нуклеиновую кислоту, доставляют посредством подкожных инъекций или депонирования, включая подкожную инфузию (такую как посредством осмотических насосов).
В одном из вариантов осуществления лиганд GPVI, представляющий собой нуклеиновую кислоту, доставляют посредством подкожного введения, а модулятор доставляют посредством подкожного или внутривенного введения.
Терапевтические композиции, содержащие лиганды и модуляторы по настоящему изобретению, можно вводить внутривенно, например, посредством инъекции стандартной дозы. Термин "стандартная доза", когда его используют по отношению к терапевтической композиции по настоящему изобретению, относится к физически дискретным единицам, подходящим в качестве однократного дозирования индивидууму, где каждая единица содержит предопределенное количество активного вещества, рассчитанного на получение желаемого терапевтического эффекта, в комбинации с необходимым разбавителем; т.е. носителем или транспортером.
Кроме того, в одном из подходов парентерального введения используют имплантацию систем с замедленным высвобождением или с пролонгированным высвобождением, что обеспечивает поддержание постоянного уровня дозы.
Также предусмотрено локальное введение, например, в интерстициальную ткань пораженного сосуда. Локальное введение можно осуществлять посредством инъекции, такой как посредством шприца или другого промышленного изделия, содержащего инъекционное устройство, такое как игла. Скорость введения шприцом можно контролировать посредством регулируемого давления в течение желаемого периода времени распределения содержимого шприца. В другом примере локальное введение можно проводить посредством инфузии, которую можно упрощать, используя насос или другое сходное устройство.
Также предоставлены характерные неограничивающие подходы для местного введения в ткань сосуда, и они включают (1) покрытие или пропитывание ткани кровеносного сосуда гелем, содержащим лиганд, представляющий собой нуклеиновую кислоту, для доставки in vivo, например, посредством имплантации покрытого или пропитанного сосуда вместо поврежденного или больного сегмента ткани сосуда, удаленного или подвергнутого шунтированию; (2) доставку посредством катетера в сосуд, которому желательна доставка; (3) нагнетание композиции в сосуд так, который необходимо имплантировать пациенту. Альтернативно, соединения можно вводить в клетки посредством микроинъекции или посредством инкапсуляции в липосомы.
Также предусмотрено введение индивидууму лигандов GPVI посредством покрытых медицинских устройств, таких как стенты с фармацевтическими композициями, содержащими лиганд. Способы покрытия для обеспечения подходящего высвобождения и введения лиганда известны специалистам в данной области.
Специалист в данной области может легко определить оптимальные режимы дозирования композиций, описываемых в настоящем документе, и они могут варьировать в зависимости от модулятора, пациента и желаемого эффекта. Эффективное количество может варьировать в зависимости от ряда факторов, таких как состояние, масса, пол, возраст индивидуума и количество вводимого лиганда, представляющего собой нуклеиновую кислоту. Другие факторы включают способ введения.
В основном, композиции вводят в дозах, скорректированных на массу тела, например, в дозах в диапазоне приблизительно от 1 мкг/кг массы тела до приблизительно 100 мг/кг массы тела. Более типично, дозы находятся в диапазоне приблизительно от 0,1 мг/кг до приблизительно 20 мг/кг, а более типично - приблизительно от 0,5 мг/кг до приблизительно 10 мг/кг, или приблизительно от 1,0 до приблизительно 5,0 мг/кг, или приблизительно 1,0 мг/кг, приблизительно 2,0 мг/кг, приблизительно 3,0 мг/кг, приблизительно 4,0 мг/кг, приблизительно 5,0 мг/кг, приблизительно 6,0 мг/кг, приблизительно 7,0 мг/кг, приблизительно 8,0 мг/кг, приблизительно 9,0 мг/кг или приблизительно 10,0 мг/кг. Как правило, доза исходно обеспечивает концентрацию лекарственного средства в плазме приблизительно от 0,002 мкг/мл до приблизительно 2000 мкг/мл, более типично - приблизительно от 2,0 мкг/мл до приблизительно 400 мкг/мл, и более типично - приблизительно от 10 мкг/мл до 200 мкг/мл или приблизительно от 20 мкг/мл до приблизительно 100 мкг/мл, приблизительно 20 мкг/мл, приблизительно 40 мкг/мл, приблизительно 60 мкг/мл, приблизительно 80 мкг/мл, приблизительно 100 мкг/мл, приблизительно 120 мкг/мл, приблизительно 140 мкг/мл, приблизительно 160 мкг/мл, приблизительно 180 мкг/мл или приблизительно 200 мкг/мл.
При введении индивидууму модулятора, которому уже вводили лиганд, соотношение модулятора и лиганда можно корректировать на основе желаемого уровня ингибирования лиганда. Дозу модулятора можно рассчитать на основе соответствия с дозой лиганда. В одном из вариантов осуществления массовое отношение доз модулятора к лиганду составляет 1:1. В других вариантах осуществления отношение модулятора и лиганда составляет более 1:1, например, 2:1 или приблизительно 2:1, 3:1 или приблизительно 3:1, 4:1 или приблизительно 4:1, 5:1 или приблизительно 5:1, 6:1 или приблизительно 6:1, 7:1 или приблизительно 7:1, 8:1 или приблизительно 8:1, 9:1 или приблизительно 9:1, 10:1 или приблизительно 10:1 или более. В других вариантах осуществления отношение доз модулятора и лиганда составляет менее чем приблизительно 1:1, например, 0,9:1 или приблизительно 0,9:1, 0,8:1 или приблизительно 0,8:1, 0,7:1 или приблизительно 0,7:1, 0,6:1 или приблизительно 0,6:1, 0,5:1 или приблизительно 0,5:1, 0,45:1 или приблизительно 0,45:1, 0,4:1 или приблизительно 0,4:1, 0,35:1 или приблизительно 0,35:1, 0,3:1 или приблизительно 0,3:1, 0,25:1 или приблизительно 0,25:1, 0,2:1 или приблизительно 0,2:1, 0,15:1 или приблизительно 0,15:1, 0,1:1 или приблизительно 0,1:1 или менее 0,1:1, например, приблизительно 0,005:1 или менее. В определенных вариантах осуществления отношение составляет от 0,5:1 до 0,1:1, или от 0,5:1 до 0,2:1, или от 0,5:1 до 0,3:1. В других вариантах осуществления отношение составляет от 1:1 до 5:1, или от 1:1 до 10:1, или от 1:1 до 20:1.
Лиганды GPVI, представляющие собой нуклеиновые кислоты, по настоящему изобретению можно вводить внутривенно одной суточной дозой, одной дозой через сутки, или общую суточную дозу можно вводить несколькими дробными дозами. Введение лиганда и/или модулятора можно проводить один раз в сутки (q.d.), дважды в сутки (b.i.d.), трижды в сутки (t.i.d.) или более часто по мере необходимости. Затем любыми подходящими способами вводят модулятор для изменения посредством введения модулятора действия лиганда, представляющего собой нуклеиновую кислоту. Лиганды, представляющие собой нуклеиновые кислоты, по настоящему изобретению можно вводить подкожно дважды в неделю, раз в неделю, каждые два недели или раз в месяц. В определенных вариантах осуществления лиганды или модуляторы вводят менее часто, чем раз в сутки. Например, введение лиганда можно проводить через сутки, каждые трое суток, каждые четверо суток, раз в неделю или раз в месяц.
В одном из вариантов осуществления может являться желательным совместное или последовательное введение других средств. Для комбинаторного лечения более чем с одним активным средством, где активные средства находятся в отдельных лекарственная составах, активные средства можно вводить одновременно, или каждое из них можно вводить раздельно в разное время.
Композиции вводят способом, совместимым с лекарственным составом и в терапевтически эффективном количестве. Вводимое количество зависит от подлежащего лечению индивидуума, способности системы индивидуума утилизировать активный ингредиент и степени желаемого терапевтического эффекта. Точные количества активного ингредиента, необходимые для введения, зависят от решения лечащего врача и являются индивидуальными для каждого индивидуума. Однако в настоящем документе описаны подходящие диапазоны доз для системного введения, и они зависят от маршрута введения. Подходящие схемы введения также варьируют, но они характеризуются начальным введением с последующими повторными дозами с интервалами в один или несколько часов посредством последующей инъекции или введения другого вида. Альтернативно предусмотрена непрерывная внутривенная инфузия, достаточная для поддержания в крови концентраций в диапазонах, указанных для терапии in vivo.
Примеры
Пример 1: Идентификация лигандов GPVI, представляющих собой нуклеиновые кислоты
Для получения лигандов, связывающих внеклеточный домен GPVI применяли способ SELEX, как описано и проиллюстрировано на фиг. 1 ниже.
Исходную библиотеку ДНК-кандидатов получали посредством отжига при нагревании и быстром охлаждении 1 нмоль олигонуклеотидов матричной ДНК и 1,5 нмоль олигонуклеотида 5'-праймера ДНК. Последовательность олигонуклеотидов матричной ДНК для получения смеси-кандидата представляют собой: 5'-TCTCGGATCCTCAGCGAGTCGTCTG(N40)CCGCA TCGTCCTCCC TA-3'(SEQ ID NO:4) (N40 представляет собой 40 смежных нуклеотидов, синтезированных при эквимолярных количествах A, T, G и C), олигонуклеотид 5'-праймера и олигонуклеотид 3'-праймера представляют собой, соответственно, 5'-GGGGGAATTCTAATACGACTCACTATAGGG AGGACGATGC GG-3' (SEQ ID NO:5) (последовательность промотора T7 подчеркнута) и 5'-TCTCGGATCC TCAGCGAGTC GTCTG-3' (SEQ ID NO:6). Реакцию проводили с фрагментом Кленова с отсутствием экзонуклеазной активности, останавливали добавлением ЭДТА до конечной концентрации 2 мМ и экстрагировали PCI (фенол:хлороформ:изоамиловый спирт (25:24:1)), а затем хлороформом:изоамиловым спиртом (24:1). Экстракт обессоливали, концентрировали, а невстроившиеся нуклеотиды удаляли на центрифужной колонке Amicon 10. Матричную ДНК использовали в реакции транскрипции для получения исходной библиотеки с остатками 2'-фторпиримидина. Условия транскрипции in vitro составляли 40 мМ Tris-HCl pH 8,0, 4% PEG-800, 12 мМ MgCl2, 1 мМ спермидин, 0,002% Triton, 5 мМ DTT, 1 мМ рГТФ, 1 мМ рАТФ, 3 мМ 2'F-ЦТФ, 3 мМ 2'F-УТФ, 8 мкг/мл неорганической пирофосфатазы, 0,5 мкМ библиотеки ДНК и мутантной полимеразы T7 Y639F. Реакционные смеси с реакцией транскрипции инкубировали в течение ночи при 37°C, обрабатывали ДНКазой, дважды экстрагировали хлороформом:изоамиловым спиртом (24:1), концентрировали на центрифужной колонке Amicon 10, и очищали в геле посредством PAGE в 12% денатурирующем геле. РНК элюировали из геля и буфер заменяли и концентрировали с использованием отмывки в TE (10 мМ Tris pH 7,5, 0,1 мМ ЭДТА) в центрифужной колонке Amicon 10.
Отбор для GPVI начинали с комплексной библиотеки ≈1014 различных последовательностей РНК с 2'-фторпиримидином. Комплексную смесь РНК предварительно очищали с использованием пептида биотин-PEG6-His6, иммобилизованного на магнитных гранулах, покрытых стрептавидином. Предварительно очищенную РНК связывали с очищенным рекомбинантным внеклеточным доменом C-конца меченного His6 белка GPVI (SEQ ID NO:3). Очищенный внеклеточный домен белка GPVI, меченный гистидином, получали в R&D Systems (Minneapolis, MN), каталожный № 3627-GP, и он содержал остатки Gln21-Lys267.
Исходный отбор лигандов GPVI проводили в буфере для связывания "E", и на последующих стадиях отбора жесткость увеличивали до буфера для связывания "F". Буфер для связывания E состоит из 20 мМ HEPES pH 7,4, 50 мМ NaCl, 2 мМ CaCl2 и 0,01% BSA. Буфер для связывания F состоит из 20 мМ HEPES pH 7,4, 150 мМ NaCl, 2 мМ CaCl2 и 0,01% BSA. Комплексы белок-РНК разделяли на 25 мм нитроцеллюлозном диске отмывкой. Связанную РНК удаляли с нитроцеллюлозного диска инкубацией в PCI (25:24:1). Добавляли буфер Tris-ЭДТА и выделяли водную фазу с последующей экстракцией хлороформом:изоамиловым спиртом (24:1). Полученную связавшуюся РНК осаждали этанолом. Одну четверть осажденной РНК отжигали при нагревании с 3'-праймером и подвергали обратной транскрипции с использованием RT AMV. Всю реакционную смесь после RT использовали в ПЦР с 5'- и 3'-праймерами и стандартными условиями ПЦР с получением матричной ДНК для следующей стадии получения РНК. Конкретные условия для каждой стадии отбора представлены на фиг. 2. После стадии 6, отбор разделяли на две ветви, обозначенные "E2" и "EF." При отборе в E2 стадии 1-8 проводили в буфере для связывания E, а стадии 9-10 проводили в буфере для связывания F. При отборе в EF стадии 1-6 проводили в буфере для связывания E, а стадии 7-10 проводили в буфере для связывания F.
Обогащение библиотек лигандов GPVI проверяли в исследованиях прямого связывания с использованием радиоактивно меченого лиганда, представляющего собой РНК, с соответствующих стадий SELEX и растворимого GPVI. Исследования связывания проводили с РНК с концевым мечением маркерным P32, добавляемой к серийным разведениям GPVI в буфере для связывания F. Для получения радиоактивно меченой РНК для исследований связывания сто пикомоль РНК дефосфорилировали бактериальной щелочной фосфатазой при 50°C в течение 1 часа. Реакционную смесь экстрагировали фенолом:хлороформом:изоамиловым спиртом (25:24:1), экстрагировали хлороформом:изоамиловым спиртом (24:1) и осаждали этанолом. Три пмоль дефосфорилированной РНК подвергали концевому мечению полинуклеотидкиназой T4 с поставляемым буфером и 20мкКи γ-P32-АТФ, а затем очищали с использованием центрифужной колонки Biorad MicroBio Spin P-30. Меченую по концам РНК разбавляли до конечной концентрации 2000 имп/мин/мкл и подвергали тепловой денатурации при 65°C в течение 5 минут. Перед использованием разведения РНК и GPVI уравновешивали при 37°C. РНК (5 мкл) добавляли в GPVI (15 мкл) с различными концентрациями при 37°C и совместно инкубировали в течение периода от 5 до 15 минут. Затем смесь с образованными комплексами РНК/белок GPVI в 96-луночной вакуумной распределительной системе с промывкой наносили на нитроцеллюлозную мембрану Protean BA85, наложенную на нейлоновую мембрану Genescreen Plus. Мембраны помещали на экран фосфоимиджера, сканировали и количественно определяли с использованием фосфоимиджера Molecular Dynamics Storm 840. Рассчитывали связавшуюся часть, деля число импульсов на нитроцеллюлозе на общее количество импульсов и корректируя на фоновый уровень. На фиг. 3 представлены результаты связывания обогащенных библиотек лигандов на стадии 10 E2 и стадии 10 EF SELEX по сравнению с не подвергавшейся контакту с GPVI исходной библиотекой лигандов.
Пример 2: Секвенирование и идентификация структурного семейства лигандов GPVI, представляющих собой нуклеиновые кислоты
Конечные продукты ПЦР, представляющие собой обогащенные библиотеки лигандов против GPVI на стадии 10 экспериментов SELEX, описанных в примере 1, расщепляли EcoR1 и BamH1, очищали набором для очистки и направленно клонировали в линеаризованный вектор pUC19. Бактериальные колонии сеяли штрихом с получением единичных клонов и из одиночных колоний инокулировали 5 мл ночных культур. Из единичных колоний получали плазмидную ДНК с использованием наборов Qiagen Plasmid Mini Prep. Сорок плазмид из каждого эксперимента SELEX секвенировали с использованием праймера к вектору. На фиг. 4 приведены последовательности ДНК, полученные из случайной области. Анализ этих последовательностей позволил идентифицировать 6 уникальных последовательностей, которые на фиг. 4 обозначены как "A"-"F". Соответствующие уникальные последовательности ДНК полноразмерных клонов лигандов представлены в таблице 1 ниже, а последовательности, представляющие собой случайную область представлены в таблице 2.



Представление этих уникальных лигандов в виде полноразмерной РНК приведено в таблице 3, и в таблице 4 в виде последовательностей РНК с указанием участков вставки 2'-фторпиримидиновых нуклеотидов, использованных в эксперименте SELEX (f указывает на 2'-фторпиримидиновую модификацию, а r указывает на немодифицированный рибонуклеотид). Полноразмерный относится к последовательностям, полученным способом SELEX, содержащим последовательности, происходящие из случайной части библиотеки лигандов, использованной в способе SELEX, а также последовательности из фиксированных частей последовательностей, фланкирующих случайную область.
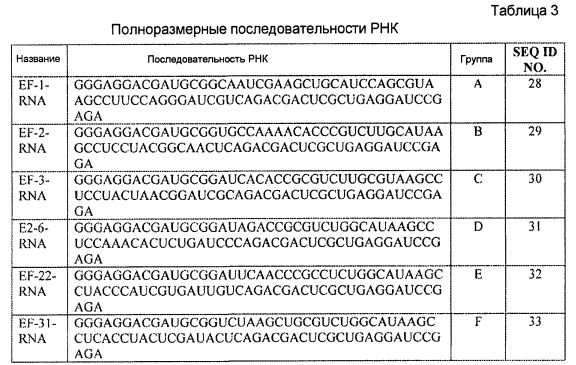
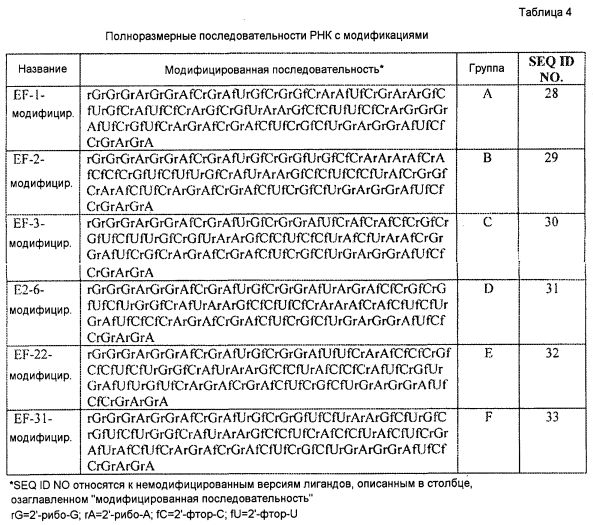
Дополнительное исследование этих шести последовательностей идентифицировало консервативную первичную последовательность, общую для всех этих клонов, состоящую из (A/G)UAA), фланкированных на каждой стороне последовательностью GC (например, GC(A/G)UAAGC), указывая на то, что эти шесть уникальных последовательностей являлись представителями родственного семейства.
Скрининг последовательностей на потенциальные вторичные структуры проводили с использованием сервера mfold (mfold.bioinfo.rpi.edu). Описание этих способов находится на сайте сервера, а также в M. Zuker (2003) "Mfold web server for nucleic acid folding and hybridization prediction." Nucleic Acids Res. 31 (13), 3406-15 и D.H. Mathews, et al., (1999) "Expanded Sequence Dependence of Thermodynamic Parameters Improves Prediction of RNA Secondary Structure" J. Mol. Biol. 288, 911-940. Затем использовали сравнительный анализ последовательностей для уникальных последовательностей с выравниванием последовательностей на основе консервативных консенсусных вторичных структурных элементов для получения предсказанной вторичной структуры лигандов против GPVI (см. фиг. 5 и фиг. 8).
Лиганды против GPVI обладают общей консенсусной вторичной структурой, состоящей из 3 стеблей (S1-S3) и 4 петель (L1-L4), с консервативной последовательностью (A/G)UAA в петле 4 и фланкирующими последовательностями GC, формирующими спаренную область, содержащую стебель 3. Разрешение консенсусной вторичной структуры семейства лигандов против GPVI идентифицировало дополнительные консервативные структурные элементы, как представлено на фиг. 5. Размер и последовательность петли 1 являлись консервативными, где петля 1 состояла из 3 нуклеотидов с последовательностью GAC. Размер и последовательность петли 3 также являлись консервативными, где петля 3 состояла из 2 нуклеотидов, наиболее часто, состоящих из последовательности UU или UG, хотя в одном из представителей семейства присутствовала CA. Стебель 1 состоял из 4-5 пар нуклеотидов, тогда как длина стебля 2 была длиннее, как правило, он состоял из 7-8 пар нуклеотидов. Петля 2 была более вариабельна по размеру, в диапазоне 4-7 нуклеотидов в длину.
Аффинность каждого из лигандов против GPVI к GPVI определяли посредством прямых исследований связывания с использованием радиоактивно меченного маркерного лиганда РНК и растворимого GPVI, с использованием способов связывания, описанных выше в примере 1. Аффинность лигандов против GPVI к GPVI являлась высокой и находилась в диапазоне Kd от ≈11 нМ до ≈50 нМ с ранжированием силы аффинности к GPVI EF-2 ≈ EF-3 ≈ E2-6 ≈ EF-22 ≈ EF-31 >> EF-1. Представляет интерес, что лиганд EF-1, содержащий последовательность CA в петле 3, продемонстрировал наименьшую аффинность к GPVI в соответствии с важностью последовательности UU или UG петли 3 для высокоаффинного связывания GPVI.
Пример 3: Исследование укорочений и мутаций структуры лигандов против GPVI
Консервативная вторичная структура лигандов против GPVI, представленная на фиг. 5, позволяет сделать достоверные предсказания в отношении минимальной последовательности, необходимой для формирования этой структуры и высокоаффинного связывания с GPVI. Начальные эксперименты были направлены на определение требования для 5' и 3' граничных последовательностей для сохранения лиганда с желаемой структурой и функцией.
Консенсусная вторичная структура, представленная на фиг. 5, демонстрирует, что области лиганда, происходящие из 3'-фиксированной области библиотека лигандов с большой долей вероятности не обязательны для связывания GPVI. Важность последовательности, происходящей из 3'-фиксированной области, на взаимодействие с GPVI тестировали посредством отжига комплементарного 3'-праймера ДНК при нагревании с меченой на концах P32 РНК, идентифицированной выше, уравновешивания и проведения исследований связывания (как описано выше в примере 1). Результаты представлены на фиг. 6. Связывание лиганда с отожженным праймером и без него являлось сравнимым, что означает, что эта область является необязательной для связывания с белком GPVI, как предсказано по консенсусной структуре, представленной на фиг. 5.
Получали укороченные соединения для нескольких из лигандов против GPVI, содержащие 5' и 3' последовательности необходимых границ для стебля 1, предсказанные по консенсусной структуре, представленной на фиг. 5 (таблица 5), и определяли их аффинность к GPVI. "RB ID" представляет собой уникальный идентификатор, относящийся к лиганду с последовательностью с конкретными модификациями, указанными в столбце, озаглавленном "модифицированная последовательность". "SEQ ID NO:" относится к соответствующей последовательности нуклеиновой кислоты (ДНК и/или РНК) без модификации.

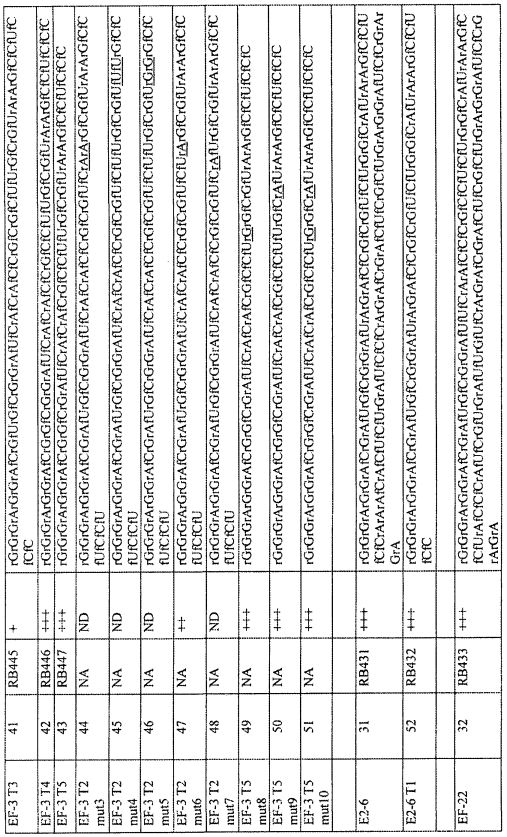
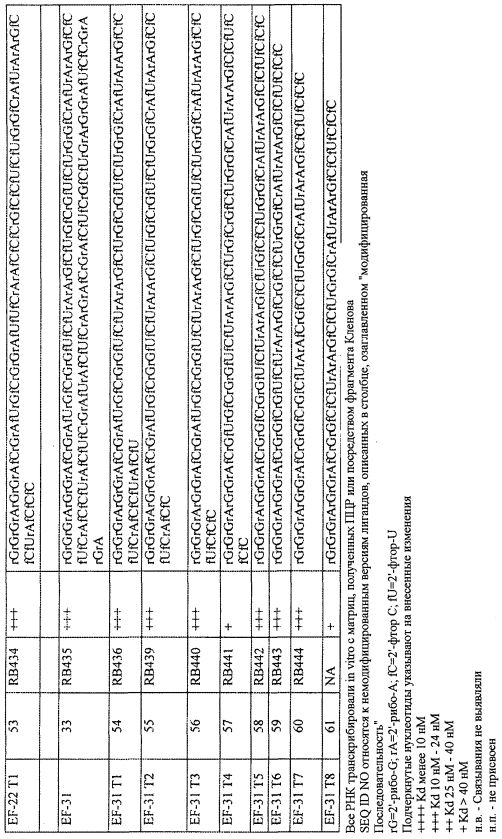
Данные о связывании для нескольких характерных укорочений представлены на фиг. 7. В соответствии с консенсусными 5' и 3' границами, предсказанными по вторичной структуре, определенной на фиг. 5, EF-1 T1, EF2 T2, EF-3 T2, E2-6 T1, EF-22 T1 и EF-31 T2 связывали GPVI с Kd равной соответствующему полноразмерному исходному лиганду, подтверждая, что предсказанные границы 5' и 3'-концов достаточны для формирования стебля 1 и функциональных лигандов против GPVI (см. таблицу 5).
Консенсусная структура, представленная на фиг. 5, позволяет предположить, что стебель 2 может содержать больше пар нуклеотидов, чем необходимо для формирования стабильной структуры стебля. Кроме того, консервативность размера и последовательности петли 1, происходящей из 5'-фиксированной области исходной библиотеки лигандов, в комбинации с размером стебля 2 и отсутствием консервативности длины или последовательности петли 2, позволяет предположить, что стебель 2 и петля 2 могут частично иметь спейсерную функцию для правильного представления петли 1. На основе этой информации можно уменьшать размер стебля 2 при сохранении высокоаффинного связывания с GPVI. Таким образом, получали дополнительные укорочения для определения минимального размера стебля 2, необходимые для высокоаффинного связывания с GPVI (см. фиг. 8A, B, C для обзора представлений вторичной структуры и таблицу 5 для обзора идентификаторов последовательностей). EF-3 T4 и T5, и EF-31 T5, T6 и T7 связывались с GPVI с Kd, равной соответствующему полноразмерному исходному лиганду, подтверждая, что для высокоаффинного связывания с GPVI достаточен стебель 2, настолько короткий как 4 пары нуклеотидов в длину. Эти результаты полностью согласуются с ролью стебля 2 и петли 2 в правильном представлении последовательности GAC петли 1. Кроме того, оба укорочения EF-3 T3 и EF-31 T4, удаляющие пару нуклеотидов A-U в 2 положении основания стебля 2, продемонстрировали сниженное связывание по сравнению с соответствующим полноразмерным исходным лигандом. Изучение состава стебля 2 этих укорочений по сравнению с составом стебля 2 для укорочений, перечисленных выше, которые не демонстрировали сниженной аффинности к GPVI, выявило, что в укорочениях EF-3 T3 и EF-31 T4 в этом положении в стебле 2 уникально расположены U-G, в противоположность паре нуклеотидов G-C или паре нуклеотидов A-U, находящейся в исходных последовательностях. Это указывает на то, что для высокоаффинного связывания GPVI пара пурин-пиримидин в положении 2 пары нуклеотидов в основании стебля 2, характеристика, консервативная в консенсусной вторичной структуре лигандов против GPVI, представленных на фиг. 5., может являться необходимой.
Консенсусная вторичная структура лигандов против GPVI, представленная на фиг. 5, демонстрирует консервативность первичной последовательности в петлях 1, 3 и 4 и отсутствие консервативности по размеру или составу последовательности в петле 2. Для оценки функциональной важности наблюдаемой консервативности или ее отсутствия в последовательностях этих петель конструировали мутации и замены (см. таблицу 5 выше; подчеркнутые участки означают мутантные остатки).
Консервативные UU, выявленные в петле 3 EF-2 T2 или EF-3 T2 подвергали мутации в AA (EF-2 T2mut3 и EF-3 T2mut3, см. фиг. 9 для обзора представлений вторичных структур), что приводило к полной потере измеряемого связывания с GPVI, что согласуется с важностью последовательности UU или UG для петли 3, что предсказано по консенсусной вторичной структуре, представленной на фиг. 5. Консервативные UU, расположенные в петле 3 в EF-3 T2, также подвергали мутации в UA (EF-3 T2mut6) и AU (EF-3 T2mut7). В соответствии с консенсусной структурой лигандов против GPVI мутация первого U петли 3 в A приводила к потере измеряемого связывания с GPVI, тогда как мутация второго U петли 3 в A приводила к небольшому снижению аффинности к GPVI. Таким образом, последовательность UA петли 3, подобно UU или UG, способствует высокоаффинному связыванию лигандов против GPVI с GPVI, и соответствует наблюдаемой консервативности петли 3 в виде 5'-YD-3', где Y представляет собой пиримидин, а в качестве пиримидина в петле 3 высоко предпочтителен U, а D представляет собой U, или G, или A (не C).
Консервативные GUAA EF-3 T2 и AUAA EF-2 T2 в петле 4 подвергали мутации в GUUU (EF-3 T2mut4) и AUUU (EF-2 T2mut4), соответственно, а также в GUGG (EF-3 T2mut5) и AUGG (EF-2 T2mut5) (см. фиг. 9 для обзора представлений вторичных структур). Оба набора замен приводили к полной потере измеряемого связывания с GPVI, что согласуется с важностью последовательности петли 4 AUAA или GUAA для связывания GPVI, что предсказано по консенсусной вторичной структуре, представленной на фиг. 5. Для дальнейшего подтверждения важности консервативных последовательностей в петлях 3 и 4, петлю 3 EF-3 T2 преобразовывали в UG (EF-3 T2mut8), как выявлено в последовательностях EF-22, E2-6 и EF-31, что не приводило к значимому изменению аффинности к GPVI, что соответствовало консервативной структуре лигандов против GPVI, представленной на фиг. 5. Кроме того, петлю 4 EF-3 T2 конвертировали в AUAA (EF-3 T2 mut9), как выявлено в EF-2, EF-22, E2-6 и EF-31, что не приводило к значимому изменению аффинности к GPVI, что соответствовало консервативной структуре лигандов против GPVI, представленной на фиг. 5. Наконец, одновременное преобразованию в EF-3 T2 петли 3 в UG и петли 4 в AUAA (EF-3 T2mutl0), как выявлено в EF-22, E2-6 и EF-31, не приводило к значимому изменению аффинности к GPVI, что соответствовало консервативной структуре лигандов против GPVI, представленной на фиг. 5.
Получали синтетические лиганды против GPVI с одиночными точечными мутациями в петле 1 (RB470-RB472, см. таблицу 6 ниже) с преобразованием последовательности петли 1 в CAC, GUC и GAG, соответственно. Каждая замена приводила к полной потере измеряемого связывания с GPVI, что согласовывается с важностью последовательности петли 1 GAC для связывания GPVI, что предсказано по консенсусной вторичной структуре, представленной на фиг. 5.
Петля 2 не продемонстрировала явной консервативности в длине или составе последовательности, но, вместо этого, она может выполнять в выделенных лигандах роль спейсера, обеспечивая представление в петле 1 последовательности GAC, полученной из фиксированной 5'-последовательности библиотеки лигандов, используемых в SELEX. Это позволяет предсказать, что нуклеотидный состав петли 2 не является необходимым для высокоаффинного связывания с GPVI. В соответствии с этим предсказанием, замена петли 2 EF-31 T7 (R B466) гексаэтиленгликолевым спейсером (см. фигура 13A-B) не приводит к потере аффинности к GPVI по сравнению с исходным лигандом (RB448). Данные связывания RB466 представлены на фигуре 10.
Пример 4: Дальнейшее укорочение и оптимизация лиганда против GPVI 2'-модификацией сахара и фосфодиэфирного скелета анти GPVI-лигандов
Лиганды, выделенные из библиотеки с остатками 2'-фторпиримидина/2'-гидроксипурина демонстрируют значительную устойчивость к нуклеазам при скрининге in vitro. Однако высокое содержание 2'-гидроксила делает их неподходящими для разработке кандидатов в лекарственные вследствие того факта, что эти положения являются очень чувствительными для разрушения нуклеазами in vivo, ограничивая максимальную концентрацию, которой можно добиться после парентерального введения, а также время их полужизни в циркуляции. Таким образом, авторы вели поиск дальнейшей оптимизации лигандов против GPVI посредством дополнительной стабилизации остова посредством замены 2'-гидроксилнуклеотидов 2'-O-метилнуклеотидами или посредством замены 2'-гидроксилнуклеотидов 2'-дезоксинуклеотидами с модификацией остова лиганда посредством замещения фосфортиоатом по мере необходимости с сохранением аффинности к GPVI при увеличении устойчивости к нуклеазам. Также проводили дополнительные замены 2'-фторнуклеотидов 2'-O-метилнуклеотидами для дополнительного увеличения стабильности, снижения стоимости производства и снижения уровня возможных загрязнений, которые могут появляться при нагревании содержащих 2'-фторуридин олигонуклеотидов в процессе получения. Наконец, для дальнейшего увеличения стабильности in vivo проводили "кэпирование" 5'- и 3'-концов, что предотвращает разрушение олигонуклеотидов экзонуклеазами. Консервативность минимальной необходимой последовательности у лигандов против GPVI являлась очень высокой, и, как правило, их аффинности к GPVI являлись сходными. Лиганд EF-31 T7 представлял собой минимальный лиганд против GPVI с высокой аффинностью к GPVI и уменьшенной длиной петли 2, и, таким образом, его выбирали в качестве исходной молекулы для дальнейшей оптимизации лигандов против GPVI в отношении стабильности к нуклеазам и связывания GPVI.
Кэпирование 3'-конца EF-31 T7 проводили посредством синтеза лиганда на подложке CPG, с нанесенным инвертированным дезокситимидином с образованием на 3'-конце лиганда связи 3'-3' (RB448). Эта модификация хорошо переносится, и, таким образом, ее использовали во всех синтетически полученных модификациях этого лиганда. Последовательность RB448 (SEQ ID NO:62) составляла: rGrGrGrArGrGrAfCrGrGfCrGfCfUrArAfCrGfCfCfUrGrGfCrAfUrArArGfCfCfUfCfCfCiT, где "r" представляет собой рибонуклеиновую кислоту, "f представляет собой 2'-фторнуклеотид и "iT" представляет собой инвертированный дезокситимидин.
Исходные замены 2'-гидрокси- и 2'-фторнуклеотидов 2'-O-метилнуклеотидами синтезировали в стеблях 1, 2 и 3 и тестировали в отношении связывания с GPVI в прямых анализах связывания, как описано в примере 1. Результаты анализов связывания представлены в таблице 6 ниже. Замена большинства (RB452) или всех (RB453) 2'-гидроксилов на 2'-O-метилы в стебле 1 переносилась хорошо, при этом наличие всех 2'-O-метилов в составе стебля 1 приводило к увеличению аффинности к GPVI по сравнению с исходным соединением (RB448), что соответствовало консервативной вторичной структуре лигандов против GPVI. Замена 2'-гидроксилов 2'-O-метилами в верхних 2 парах нуклеотидов стебля 2 (RB455) также переносилась хорошо, что соответствовало консервативной вторичной структуре и данным укорочения, что означает, что в лиганде против GPVI в верхней части стебля 2 могут присутствовать несколько комбинаций модификаций пар нуклеотидов и содействовать высокоаффинному связыванию с GPVI. Наконец, замена в стебле 3 пары нуклеотидов G-C с 2'-гидроксилом-2'-фтором парой нуклеотидов G-C с 2'-O-метилом (RB462) приводила к значимому увеличению аффинности к GPVI. То, что эта замена увеличит стабильность стебля 3, ожидалось, и она соответствует консенсусной вторичной структуре лигандов против GPVI, в которой предсказано, что стебель 3 является коротким стеблем из 2 пар нуклеотидов.
Затем синтезировали составную молекулу, состоящую из замен в RB453, RB455, RB462 и RB466 (RB490), и тестировали на связывание с GPVI в прямых анализах связывания (см. фиг. 10). В соответствии с аффинностью композиций, содержащих отдельные группы замен, RB490 связывался с GPVI со значительно большей аффинностью, чем исходное соединение, с Kd для GPVI ≈4-5 нМ по сравнению с ≈14-15 нМ для RB448. Затем, RB490 служило в качестве исходного соединения для дальнейшей оптимизации лиганда против GPVI. Следует отметить, что, хотя замена в первом остатке молекулы на 2'-O-метил (G в положении 1) хорошо переносится, эта модификация значительно снижает эффективность 5'-концевого мечения этих композиций P32, и, таким образом, в этом остатке оставили 2'-гидроксил для оценки большинства оставшихся тестируемых модификаций для облегчения исследования прямого связывания. Представления вторичных структур с указанием модификаций сахаров для описанных ключевых составных композиций, можно найти на фиг. 11A-D.
Так как 5'-половина стебля 2 получена из 5'-фиксированной области библиотеки лигандов, используемой в исследованиях SELEX, состав последовательности стебля может являться менее надежным показателем для вероятности переносимости замен для 2'-гидроксил- или 2'-фторнуклеотидов нижних двух пар нуклеотидов в этом стебле, чем в последовательностях, полученных из случайной области библиотеки лигандов. Таким образом, для двух пар нуклеотидов G-C в основании стебля 2 синтезировали отдельные замены 2'-гидроксила и 2'-фтора на 2'-O-метил (RB497, 498, 499 и 500). Замены 2'-гидроксила на 2'-O-метил хорошо переносились для G в паре нуклеотидов G-C в концевом положении стебля 2 (RB497), но замена 2'-фтора на 2'-O-метил для C этой пары нуклеотидов (RB498) хорошо не переносилась. Замена 2'-гидроксила и 2'-фтора сахара на 2'-O-метил для G и C в паре нуклеотидов G-C во втором положении стебля 2 (RB499 и RB500) переносилась хорошо, хотя для замены 2'-O-метил в G (RB499) отмечено умеренное снижение аффинности.
В нескольких из предсказаний консенсусной вторичной структуры для лигандов против GPVI предполагается, что стебель 1 может являться таким коротким, как 4 пары нуклеотидов. В соответствии с консенсусной вторичной структурой, делеция одной из пар нуклеотидов G-C в стебле 1 переносилась хорошо (RB507), без потери аффинности по сравнению с исходной композицией RB490.
Модификация первой пары нуклеотидов G-C в стебле 3 в пару нуклеотидов, содержащую 2'-O-метилсахара (RB462) приводила к увеличенной аффинности по сравнению с составом с парой нуклеотидов G-C с 2'-гидроксилом-2'-фтором в этом положении. В отличие от этого замена в сахарах на 2'-O-метил в паре нуклеотидов C-G в стебле 3 приводила к потере измеряемой аффинности GPVI (RB463). Таким образом, для дополнительного исследования оптимального профиля модификации сахаров для стебля 3 в сахарах проводили одиночные замены 2'-фтора и 2'-гидроксила на 2'-O-метил, соответственно, в паре нуклеотидов С-G стебля 3 (RB501 и RB502). Замена 2'-O-метилом 2'-гидроксила сахара в G этой пары нуклеотидов C-G переносилась хорошо, без видимой потери аффинности к GPVI (RB502), тогда как замена 2'-O-метилом 2'-фтора в C этой пары нуклеотидов C-G приводила к потере измеряемой аффинности GPVI (RB501), что означает, что данная замена в RB463 была причиной потери связывания GPVI, наблюдаемого в этих композициях. В целом предпочтительный профиль модификации сахаров в стебле 2 соответствует консенсусной структуре лиганда против GPVI и означает, что наличие C в этой паре нуклеотидов C-G стебля 3 является консервативным на уровнях модификации сахара, формирования стебля и первичной последовательности.
Последовательность петли 4 в лигандах против GPVI является высококонсервативной, и, таким образом, в сахарах этих положений в петле 4 проводили отдельные замены на 2'-O-метил (RB503, RB504, RB505 и RB506). Замены на 2'-O-метил хорошо переносились в каждом из этих положений с заменой 2'-гидроксила сахара на 2'-O-метил в каждом A в петле 4, дающей композиции с аффинность к GPVI, эквивалентной исходной композиции RB490 (RB503, RB505 и RB506), тогда как замена 2'-фтора сахара на 2'-O-метил в U в петле 4 (RB504) приводила к умеренной потере аффинности к GPVI. При этом, так как модификация 2'-фтором в сахаре обеспечивает значительную устойчивость к нуклеазам, U в этой петле сохранили в виде 2'-фторнуклеотида.
Анализ мутаций (см. пример 3, RB470, RB471 и RB472) продемонстрировал важность последовательности GAC в петле 1 для высокоаффинного связывания с GPVI. Таким образом, для определения предпочтительного профиля модификация сахаров в нуклеотидах петли 1 в сахарах в каждом из положений в петле 1 проводили отдельные замены на 2'-O-метил и 2'-дезокси (RB491, RB492, RB493, R B494, RB495, и RB496). Замена 2'-O-метилом 2'-гидроксила G в петле 1 переносилась хорошо (RB491) с аффинностью к GPVI, эквивалентной исходной композиции RB490, и, следовательно, замену на 2'-дезокси в этом положении не тестировали (RB494). Замена 2'-гидроксила A в петле 2 на 2'-дезокси (RB495) переносилась лучше, чем замена 2'-O-метилом в этом положении (RB492) с аффинностью к GPVI приблизительно равной исходной композиции RB490. Замена 2'-O-метилом или 2'-дезокси в сахаре для 2'-фторсахара C в петле 2 приводила к умеренной потере аффинности к GPVI, и таким образом в дальнейшем ее не использовали. В соответствии с отдельными заменами в сахарах в петле 1, состав с петлей 1, состоявшей из 2'-O-метил-G, 2'-дезокси-A и 2'-фтор-C (RB519), связывал GPVI с большей аффинностью, чем состав с петлей 1, состоявшей из 2'-O-метил-G, 2'-O-метил-A и 2'-фтор-C (RB518).
Синтезировали комбинированные молекулы, содержащие замены в RB491, RB502, RB503, RB505 и RB506 (RB520) и содержащие замены в RB491, RB502, RB503, RB505, RB506 и RB507 (RB521), для подтверждения того, что введение каждой из этих замен в одну композицию переносится хорошо. Как и ожидалось на основе отдельных замен, RB520 и RB521 (такой же профиль замен в сахарах, как и в RB520, но со стеблем 1 с 4 парами нуклеотидов), связывались с GPVI с Kd, сходной со связыванием RB490 с GPVI (приблизительно 6 нМ в сравнении с 4-5 нМ для RB490). Подобным образом, комбинированная молекула, содержащая эти замены, но с 2'-фтором во 2 паре нуклеотидов G-C стебля 2 (RB524) также связывалась с GPVI с высокой аффинностью.
RB524 в центральном коре молекулы (определяемого верхом стебля 1, петлей 1, основанием стебля 2, петлей 3, стеблем 3 и петлей 4) содержала 4 оставшихся остатка с 2'-гидроксилами. Для определения оптимальных профилей замен сахаров для оставшихся остатков с 2'-гидроксилами относительно сильно модифицированного состава RB524, на основе RB524 проводили одиночные, парные и множественные замены в сахарах, исследованные ранее. Включение последовательности петли 1 с 2'-O-метил-G, 2'-дезокси-A и 2'-фтор-C в сочетании с заменой 2'-дезокси в G в концевой паре нуклеотидов G-C стебля 2 в основу RB524 (RB526) приводило к составу, который сохранял высокоаффинное связывание с GPVI при наличии только двух оставшихся 2'-гидроксилов. Затем, как и ожидалось на основе предыдущего анализа длины стебля 1, делеция пары нуклеотидов G-C в составе RB526 (RB537) приводило к составу с сохраняющимся высокоаффинным связыванием с GPVI. Замена 2'-O-метилом 2'-гидроксисахара в G петли 3 (RB538) в основе 537 также приводила к составу, сохраняющему высокоаффинное связывание с GPVI. Наконец, замена 2'-дезокси одного 2'-гидроксила, оставшегося в G 2 пары нуклеотидов G-C в стебле 2 RB538 (RB540) переносилась хорошо, позволяя получать состав с полным замещением сахаров по положению 2' с Kd для GPVI приблизительно 15 нМ.
Функциональные исследования (см. пример 6) показали, что существует хорошая корреляция между аффинностью к GPVI и ингибированием индуцированной коллагеном или CRP агрегации тромбоцитов. Кроме того, способность лигандов против GPVI блокировать индуцированную CRP агрегацию тромбоцитов позволяет предполагать, что лиганды могут связываться с областью связывающего коллаген домена, перекрывающей участок связывания CRP на GPVI или расположенной близко с ним. Участок связывания коллагена на GPVI имеет суммарный положительный заряд, и большую электроположительную площадь. Таким образом, в областях RB540, содержащих замены 2'-дезокси в петле 1 и стебле 2, проводили замены на тиофосфат, так как замены на тиофосфат увеличивают общую электроотрицательность олигонуклеотидов и, таким образом, могут приводить к увеличению аффинности лигандов к GPVI. Кроме того, замены на тиофосфат в содержащих 2'-дезоксисахара остатках могут обеспечивать дополнительную защиту от разрушения нуклеазами. На основе RB540 проводили отдельные замены на фосфотиоат, фланкирующие A и C петли 1, и G 2 концевых пар нуклеотидов стебля 2 (RB546, RB547, RB548, RB549, RB550). Замена на единичные фосфотиоаты между двумя G в основе стебля 2 (RB549) или с 3'-конца 2 G стебля 2 (RB550) увеличивала аффинность лигандов к GPVI. Включение этих обеих замен на фосфотиоат в комбинированный лиганд против GPVI (RB566) дополнительно увеличивала аффинность лиганда к GPVI, позволяя получать лиганд, демонстрирующий немного увеличенную аффинность к GPVI по сравнению с RB490 с Kd для GPVI приблизительно 2-4 нМ и значительно увеличенной аффинностью по сравнению с RB540 (см. фиг. 12).
Кроме степени стабилизации в отношении нуклеаз, распределение и срок полужизни лигандов после парентерального введения в значительной степени зависит от их молекулярной массы. Конъюгация лигандов с высокомолекулярными носителями, такими как высокомолекулярный полиэтиленгликоль (PEG), ограничивает распределение лиганда в основном плазматическим компартментом, приводя к более высокой Смакс на единицу дозы и значительно ограничивая фильтрацию лиганда почками, и, таким образом, значительно увеличивая эффективность и срок полужизни лиганда in vivo. Принимая во внимание, что для тонкой регуляции эффективности и срока полужизни лигандов против GPVI можно вводить регулирующее средство (см. пример 8), лиганды против GPVI конъюгировали с высокомолекулярным носителем для обеспечения наибольшего возможного срока полужизни с распределением, ограниченным в основном плазматическим компартментом. Пегилирование лигандов можно проводить посредством конъюгации PEG с уникальным участком на лиганде, добавляемым посредством введения в лиганд при синтезе сайт-специфического линкера. Таким образом, оценивали влияние добавления линкера и конъюгации с PEG на RB540, RB549 и RB566. Содержащие 5'-гексиламинолинкер композиции этих соответствующих лигандов (RB542, RB560 и RB567) связывались с GPVI с аффинностями, аффинностями исходных композиций, не содержащих линкер. Следует отметить, что для композиций, содержащих 5'-линкер или 5'-линкер и PEG, сахар G в положении 1 лиганда содержит 2'-O-метил. Кроме того, так как введение линкера в 5'-конец лиганда препятствует мечению 5'-конца P32, аффинность модифицированных линкером и PEG лигандов определяли посредством конкурентного связывания с родственным радиоактивно меченным исходным лигандом преимущественно способами, описанными в примере 1. Конъюгация разветвленного PEG массой 40 кДа с соответствующими лигандами, содержащими линкер (RB569, RB570, RB571) приводила приблизительно к снижению аффинности к GPVI на величину от 1,2 до 1,7 раз по сравнению с не содержащими линкер или содержащими только линкер исходными композициями (например, RB566 для RB571). Таким образом, RB571, состоящий из состава RB566 (с 2'-O-метил-G в положении 1), конъюгированного с PEG массой 40 кДа, демонстрирует Kd для GPVI приблизительно 5 нМ.




Пример 5: Способы оценки антитромбоцитарной активности, специфичности активности и модуляции активности лигандов против GPVI
A. Анализ индуцированной коллагеном агрегации тромбоцитов (CIPA) в PRP и WP
1. Получение обогащенной тромбоцитами плазмы (PRP) и исследования агрегации:
Обогащенную тромбоцитами плазму (PRP) человека получали из свежей цельной крови, собираемой в 60 мл шприцы с использованием в качестве антикоагулянта 0,3 мМ PPACK в физиологическом растворе (9:1 кровь:антикоагулянтная солевая смесь; Biomol, каталожный № PH 117). Кровь центрифугировали при низкоскоростном центрифугировании (250×g) в 50 мл конических пробирках в течение 16 минут. Обогащенную тромбоцитами плазму, отделенную от клеток крови посредством центрифугирования, отбирали 10 мл серологическими пипетками, а бедную тромбоцитами плазму (PPP) получали из оставшейся крови посредством высокоскоростного центрифугирования при 2200×g в течение 10 минут. PPP отбирали и сохраняли в качестве контроля для оптической агрегометрии (LTA).
За агрегацией тромбоцитов в PRP следили с использованием 450 мкл PRP (и 25 мкл физиологического раствора) при 37°C (перемешивая при 1200 об./мин.) в люминесцентном агрегометре Chrono-Log (Havertown, PA) в течение 6 минут. Агрегацию инициировали с использованием в качестве агониста 25 мкл коллагена. В качестве исходного уровня в агрегометре используют 500 мкл бедной тромбоцитами плазмы (PPP). Для скрининга лигандов против GPVI на способность блокировать CIPA 450 мкл PRP в течение 3 минут при 37°C в ячейке агрегометра с постоянным перемешиванием при 1200 об./мин. перед добавлением агониста коллагена инкубируют с 25 мкл раствора, содержащего лиганд против GPVI с концентрацией для получения желаемой конечной концентрации. Агрегацию тромбоцитов инициируют добавлением указанных концентраций коллагена (коллаген типа 1 фибрилл сухожилия лошади, Chronolog, каталожный № 385) с получением процента агрегации 70-90%, и непрерывно записывали светопроницаемость в течение 4-6 мин.
2. Получение отмытых тромбоцитов (WP) и исследования агрегации:
Отмытые тромбоциты человека получали по существу, как описано в Mustard et al. (1972; Br. J. Haematol. 22, 193-204). В кратком изложении кровь человека собирали с одной шестой объема буфера кислота/цитрат/декстроза (ACD) (85 мМ цитрат натрия, 65 мМ лимонная кислота и 110 мМ глюкоза), помещали в водяную баню при 37°C на срок в течение 30 минут, затем центрифугировали при 250×g в течение 16 минут при комнатной температуре. Отбирали обогащенную тромбоцитами плазму и центрифугировали при 2200×g в течение 13 минут при комнатной температуре, затем ресуспендировали в 40 мл забуференного HEPES раствора Тироде (136,5 мМ NaCl, 2,68 мМ KCl, 1 мМ MgCl2, 2 мМ CaCl2, 12 мМ NaHCO3, 0,43 мМ NaH2PO4, 5,5 мМ глюкоза, 5 мМ HEPES pH 7,4, 0,35% бычий сывороточный альбумин), содержащего 10 Ед/мл гепарина и 5 мкМ (конечная концентрация) простагландин I2 (PGI2). Суспензию тромбоцитов инкубировали в водяной бане при 37°C в течение 10 минут, добавляли 5 мкМ (конечная концентрация) PGI2 и смесь центрифугировали при 1900×g в течение 8 минут. Полученный осадок ресуспендируют в 40 мл забуференного HEPES раствора Тироде, содержащего 5 мкМ (конечная концентрация) PGI2, а затем инкубируют в течение 10 минут в водяной бане при 37°C и центрифугируют при 1900×g в течение 8 минут. Осадок ресуспендируют при плотности 3×108 тромбоцитов/мл в забуференном HEPES растворе Тироде, содержащем 0,1 Ед/мл апиразы картофеля, и до использования в агрегометрических исследованиях инкубируют в водяной бане при 37°C в течение 1 часа.
Индуцированную коллагеном агрегацию тромбоцитов WP определяли, измеряя светопропускание через 0,5 мл суспензии перемешиваемых (1200 об./мин.) отмытых тромбоцитов (425 мкл отмытых тромбоцитов, 25 мкл фибриногена, 25 мкл ингибиторов или контролей и 25 мкл коллагена) в люминесцентном агрегометре при 37°C (Chrono-Log Corp. Havertown, PA). Исходный уровень устройства устанавливали с использованием 0,5 мл забуференного HEPES раствора Тироде. Перед измерениями агрегации суспензию тромбоцитов дополняли 1 мг/мл фибриногена. Агрегацию тромбоцитов инициировали добавлением указанной концентрации коллагена (коллаген типа 1 фибрилл сухожилия лошади; Chronolog, каталожный № 385) с получением процента агрегация 70-90%, и непрерывно записывали светопропускание в течение периода по меньшей мере 6 мин. Для скрининга лигандов против GPVI или контролей (различные мутанты лигандов) на способность блокировать CIPA, к суспензии тромбоцитов добавляли лиганды против GPVI при концентрации, позволяющей получить желаемую конечную концентрации, и перед добавлением коллагена инкубировали в течение 3 мин, и ответную реакцию записывали в течение 4-6 мин после добавления коллагена.
Активность коллагена определяли для каждого донора на основе максимальной величины процента агрегации, получаемой на основе кривой доза-ответ с использованием 2× серийного разведения 4 мкг/мл коллагена в физиологическом растворе, и определяли стимулирующую концентрацию. Способность лигандов против GPVI ингибировать CIPA тестировали в препаратах WP и PRP, как описано выше с использованием широкого диапазона концентраций лигандов против GPVI (2 мкМ - 7,8 нМ).
B. Анализ индуцированной CRP агрегации тромбоцитов (CRPIPA) в WP и PRP
Перекрестно сшитый родственный коллагену пептид, CRP-XL [(GPO)10], является селективным мощным агонистом GPVI (Farndale et al., J. Thromb. Haemost. 2004; 2: 561-573; Smethurst PA et al., J. Biol. Chem. 2007; 282: 1296-1304), который можно использовать для специфической инициации агрегации тромбоцитов посредством рецептора GPVI в анализах агрегации в PRP и WP.
Активность CRP-XL определяли для каждого донора на основе максимальной величины процента агрегации, получаемой на основе кривой доза-ответ с использованием 2× серийного разведения 400 нг/мл CRP-XL в физиологическом растворе, и определяли стимулирующую концентрацию. Способность лигандов против GPVI ингибировать CRPIPA тестировали в препаратах WP и PRP, как описано выше, с использованием широкого диапазона концентраций лигандов против GPVI (2 мкМ - 7,8 нМ).
C. Определения процента снижения индуцированной коллагеном или CRP агрегации тромбоцитов и IC 50
Процент снижения индуцированной коллагеном или CRP агрегации тромбоцитов лигандами против GPVI рассчитывали принимая максимальную степень агрегации тромбоцитов при данной активирующей концентрации коллагена или CRP за 100 процентов и строили график с использованием GraphPad Prism.
При тестировании широкого диапазона концентраций лигандов против GPVI (как правило, концентрации от 1 мН до 1 мкМ), также определяли значение IC50. Значения IC50 представляют собой концентрацию лиганда, необходимую для ингибирования агрегации, вызываемой данной концентрацией коллагена или CRP, на 50%.
D. Анализ адгезии тромбоцитов в потоке цельной крови in vitro на активность лигандов против GPVI с использованием Bioflux™ 200 (Fluxion Biosciences, Inc.)
1. Получение тестового планшета, покрытого коллагеном:
Для проточных экспериментов стандартным способом использовали 48-луночные планшеты Bioflux (P/N 0009-0013). Планшеты промывали 0,02 М уксусной кислотой в течение 5 мин при 0,5 Па, а затем из впускной лунки в течение 10 мин при 0,5 Па перфундировали 25 мкг/мл разбавленного фибриллярного коллагена (Chrono-Log P/N 385) в 0,02 М уксусной кислоте. Поток останавливали и планшет инкубировали при комнатной температуре в течение 1 часа. Коллаген отмывали PBS при 0,5 Па в течение 10 мин. Затем покрытый коллагеном планшет блокировали посредством полного заполнения впускной лунки (1 мл) PBS +5% BSA масс./об. и перфундирования раствора в канал при 0,5 Па в течение 15 мин. Поток останавливали и планшет инкубировали в течение дополнительных 10 минут при комнатной температуре. Избыток PBS+BSA удаляли из всех лунок и планшет оставляли при комнатной температуре для использования в течение этих же суток или оставляли при 4°C в PBS+BSA (до двух недель).
2. Получение цельной крови для перфундирования и проточный эксперимент:
Кровь брали у здоровых добровольцев в антикоагулянт PPACK (0,3 мМ)/CTI (60 мкг/мл) в 60 мл шприцы с использованием иглы калибра 193/4''. Кровь сразу же флуоресцентно метили 4 мкМ кальцеина-AM (Invitrogen P/N C3100 MP) в течение 1 часа при 37°C (кальцеин-AM добавляли в кровь очень осторожно переворачивая пробирку несколько раз для смешивания и кровь использовали в течение 3,5 часа после получения). Эксперимент начинали, добавляя 200 мкл меченой крови сверху впускной лунки и сразу же начинали перфузию с использованием настроек тока цельной крови при 37°C 2 Па с использованием программного обеспечения Bioflux™. Данные (флуоресцентные изображения агрегатов тромбоцитов) получали с использованием инвертированного флуоресцентного микроскопа с покадровой съемкой (микроскоп Zeiss 200M Axiovert с присоединенной камерой с полупроводниковой светочувствительной матрицей Axiocam и с программным обеспечением Axiovision) каждые 6 секунд общей длительностью 6 минут. Для тестируемых препаратов (лиганды против GPVI, контрольные лиганды или контрольные антитела) 200 мкл меченой крови перед добавлением во впускную лунку в течение 4 минут при комнатной температуре инкубировали с указанными концентрациями лигандов (или буфером F; в объеме 10 мкл). Для расчета интенсивности флуоресценции с применением программного обеспечения Bioflux Montage™ использовали изображения в формате файла изображений, снабженных метками (tiff), а затем данные экспортировали в Microsoft Excel и наносили на график с использованием Graphpad Prism. Данные нормализовали на флуоресцентный сигнал, получаемый в контрольной камере в момент времени заполнения контрольной камеры флуоресцентными агрегатами тромбоцитов (определяемый как максимум ответной реакции тромбоцитов), что, как правило, происходило через 3-4 минуты, в отсутствии лиганда против GPVI.
E. Анализ индуцированной АДФ агрегации тромбоцитов (AIPA), индуцированная TRAP агрегация тромбоцитов (TIPA), анализ индуцированной арахидоновой кислотой агрегации тромбоцитов (AAIPA) и анализ индуцированной ристоцетином агрегации тромбоцитов (RIPA) для оценки специфичности лиганда против GPVI к GPVI
Специфичность лигандов против GPVI к GPVI определяли, оценивая их действие на индукцию агрегации тромбоцитов агонистами, функция которых опосредована другими хорошо охарактеризованными рецепторами. Для этих исследований при необходимости оценки активности различных агонистов использовали препараты PRP и WP человека. В качестве специфического агониста рецепторов P2Y12 и P2Y1 использовали АДФ, в качестве агониста PAR-1 - TRAP, в качестве агониста рецепторов тромбоксана A2 (TXA2) - арахидоновую кислоту и в качестве агониста взаимодействия vWF-GP1bα - ристоцетин. Эффективность каждого агониста (АДФ, TRAP, арахидоновой кислоты и ристоцетина) определяют на основе максимальной величины процента агрегации, полученной на основе кривой доза-ответ для соответствующего агониста, и определяют активирующую концентрацию для задания EC70-90% для каждого соответствующего агониста.
1. Определение специфичности лигандов против GPVI, представляющих собой нуклеиновые кислоты, посредством AIPA и TIPA в WP:
Для оценки возможного взаимодействия лигандов против GPVI с P2Y12 и P2Y1 для стимуляции агрегации тромбоцитов, как правило, использовали активирующую концентрацию АДФ 5 мкМ, а для оценки возможного взаимодействия с PAR-1 для стимуляции агрегации тромбоцитов, как правило, использовали активирующую концентрацию TFLLRN (TRAP) 2,5 мкМ (конкретную активирующую концентрацию агониста для каждого эксперимента определяют на основе кривой доза-ответ агониста для каждого донора). Индуцированную АДФ и TRAP агрегацию тромбоцитов определяли в препаратах WP, как описано выше. Для демонстрации того, что ингибирование рецептора-мишени в анализах являлось детектируемым, в качестве положительных контролей для каждого рецептора использовали специфичные ингибиторы. В качестве положительного контроля антагонизма PAR-1 использовали SCH79797 (Tocris Biosciences), а в качестве положительного контроля антагонизма P2Y12 использовали INS50589 (Inspire Pharmaceuticals).
2. Определение специфичности лигандов GPVI, представляющих собой нуклеиновые кислоты, посредством RIPA в PRP:
Для оценки возможного взаимодействия лигандов против GPVI с vWF или GP1bα для стимуляции агрегации тромбоцитов использовали активирующую концентрацию ристоцетина (Sigma, каталожный № R7752) 1,0-2,0 мг/мл (конкретную активирующую концентрацию агониста для каждого эксперимента определяют на основе кривой доза-ответ агониста для каждого донора). Индуцированную ристоцетином агрегацию тромбоцитов определяют в препаратах PRP, как описано выше, для демонстрации того, что ингибирование рецептора-мишени GP1bα в анализах являлось детектируемым, в качестве положительного контроля использовали антитело HIP1 к GP1bα (или контрольный изотип IgG, Axxora Bio; конечная концентрация 25 мкг/мл).
3. Определение специфичности лигандов GPVI, представляющих собой нуклеиновые кислоты, посредством AAIPA в PRP:
Для оценки возможного взаимодействия лигандов против GPVI с рецептором TXA2 для стимуляции агрегации тромбоцитов использовали активирующую концентрацию арахидоновой кислоты 0,25-0,5 мг/мл (Helena Biosciences; каталожный № 5364) (конкретную активирующую концентрацию агониста для каждого эксперимента определяют на основе кривой доза-ответ агониста для каждого донора). Индуцированную арахидоновой кислотой агрегацию тромбоцитов в препаратах PRP определяют, как описано выше.
F. Анализы индуцированной коллагеном и CRP агрегации тромбоцитов для тестирования представляющих собой нуклеиновую кислоту модуляторов лигандов против GPVI в WP и PRP:
Индуцированную коллагеном и CRP агрегацию тромбоцитов проводили в препаратах WP и PRP, как описано выше. Для оценки способности представляющих собой нуклеиновую кислоту модуляторов обращать ингибирование агрегации тромбоцитов лигандами против GPVI, тестировали концентрации лигандов против GPVI при их IC95-100 (концентрация лиганда, необходимая для ингибирования 95-100% агрегации, вызываемой данной концентрацией активирующего агониста). Исследования агрегации тромбоцитов проводили, как описано выше, за исключением того, что после исходной инкубации препаратов тромбоцитов с лигандом против GPVI, добавляли различные количества модулятора, обеспечивая молярный избыток модулятора к лиганду в диапазоне от 8:1 до 0,5:1, и перед добавлением агониста совместно инкубировали в течение 10 минут.
G. Исследование длительности обращения действия лигандов против GPVI представляющими собой нуклеиновую кислоту модуляторами в WP:
Для оценки длительности обращения активности против GPVI представляющими собой нуклеиновую кислоту модуляторами в общий объем суспензии WP 4 мл при 37°C добавляли RB490 (конечная концентрация 0,25 мкМ) и RB515 (конечная концентрация 0,75 мкМ) (порядок и сроки добавления, как описано выше для оценки представляющих собой нуклеиновую кислоту модуляторов), в заданные моменты времени (0, 0,16, 0,5, 1, 1,5, 2, 2,5, 3 и 3,5 часа) отбирали аликвоты 450 мкл смеси суспензии WP и проводили индуцированную коллагеном агрегацию тромбоцитов. Для демонстрации активности суспензии WP, активности RB490 и отсутствия препятствий при воздействии RB515 в течение этой инкубации, проводили отдельные инкубации в течение периода времени 3,5 часа, в которых в суспензию WP добавляли только буфер, только RB490 или только RB515 и определяли индуцированную коллагеном агрегацию тромбоцитов.
Пример 6: Ингибирование индуцированной коллагеном и CRP функции тромбоцитов лигандами против GPVI
GPVI, экспрессирующийся только на тромбоцитах, представляет собой основной рецептор коллагена тромбоцитов. GPVI необходим для стабильной адгезии тромбоцитов, и взаимодействие GPVI с коллагеном является одним из наиболее мощных активаторов тромбоцитов, инициируя внутреннюю активацию интегринов тромбоцитов α2β1 и αIIbβ3, а также секрецию содержимого гранул тромбоцитов (что в свою очередь активирует находящиеся в циркуляции рядом тромбоциты). У людей дефицит GPVI вызывает отсутствие активации тромбоцитов в ответ на коллаген, а in vitro - отсутствие агрегации тромбоцитов в ответ на коллаген. Предсказанная структура внеклеточного домена GPVI содержит два Ig-подобных домена, содержащих коллаген-связывающий домен (CBD) с последующим сильно O-гликозилированным стеблем. Являясь типичным рецептором LRC, GPVI ассоциирован с γ-цепью корецептора FcR. Передача сигнала GPVI косвенно опосредована γ-цепью FcR и непосредственно цитоплазматическим доменом GPVI. Для активации GPVI необходима четвертичная структура тканевого коллагена, и опосредованную коллагеном активацию можно воспроизвести посредством поперечно сшитого родственного коллагену пептида (CRP-XL), который состоит из трипептидных повторов последовательности (GPO)n, где G представляет собой глицин, P представляет собой пролин и O представляет собой гидроксипролин.
Определена кристаллическая структура CBD GPVI, состоящая из остатков Q1-T183 (Horii et al., Blood 108; 2006, p936-942). Данные о структуре наряду с данными мутагенеза предоставляют подробную информацию о молекулярном взаимодействии коллагена и GPVI и CRP-XL и GPVI. Такие исследования также облегчают понимание механизмов активации GPVI. Домен CBD состоит из двух Ig-подобных доменов, C2-1 и C2-2 (где C2-1 является N-концевым, а C2-2 - C-концевым), ориентированных под углом 90°. CBD GPVI в кристаллической структуре формирует димер с доменами C2-2 соответствующих CBD, взаимодействующих с образованием димера сдвоенный димер. Каждый из CRP и коллагена частично связаны в поверхностной борозде на C2-1. Эта борозда среди рецепторов LRC уникальна для GPVI, приводя к делеции в GPVI 11 остатков по сравнению с другими рецепторами LRC. Дно этой связывающей борозды сформировано несколькими гидрофобными остатками (L53, F54, P56, L62 и Y66 и алифатической частью K41), с несколькими полярными (S43, S44, Q48, Q50 и S61) и основными (K41, R46, K59 и R166) остатками по периферии. Остатки, непосредственно участвующие в связывании коллагена или CRP, распадаются на два кластера. Первичная область состоит из основных остатков на поверхности C2-1, включая K41, K59, R60 и R166. Второй кластер остатков, участвующих в связывании коллагена или CRP, расположен на дистальном конце C2-1 и содержит L36 (участвующий в связывании коллагена, но не CRP), и V34 и N-гликан, связанный с N72, участвующий в связывании коллагена и CRP. В соответствии с мутационным анализом, компьютерный докинг CRP относительно CBD GPVI демонстрирует, что связывающая CRP борозда расположена в первом кластере основных остатков с непосредственным взаимодействием с K41 и R166, и в пределах образования связи боковых цепей K59 и R60.
Каждый димер GPVI содержит две связывающих коллаген борозды, по одной в пределах каждой субъединицы C2-1 соответствующих CBD. Природные волокна коллагена состоят из псевдошестиугольной структуры параллельных CRP-подобных тройных спиралей, разделенных расстоянием от 1,3 до 1,4 нм, расположение, консервативное в кристаллических структурах растворимых пептидов CRP. Связывающие коллаген борозды в димере GPVI по существу являются параллельными, и они разделены приблизительно 5,5 нм, расстоянием между n и n+4 спиралями в коллагеновом волокне. Геометрическая совместимость связывающих борозд со спиралями коллагена позволяет димеру GPVI одновременно связывать 2 спирали в коллагеновом волокне и подобным образом CRP-XL. Таким образом, геометрия пептидов CRP-XL имитирует геометрию природного коллагена и позволяет использовать CRP-XL в качестве отдельного зонда CBD GPVI.
Для начальной оценки способности лигандов против GPVI блокировать функцию GPVI, оценивали способность лигандов EF-2 T2 и EF-3 T2 (см. фиг. 9) блокировать индуцированную коллагеном активацию тромбоцитов в препаратах WP, как описано в пример 5. В качестве контроля любого возможного неспецифического действия лигандов на индуцированную коллагеном агрегацию тромбоцитов в начальный скрининг в качестве контроля специфичности включали не связывающие GPVI мутантные лиганды, EF-2 T2 M3-M5 и EF-3 T2 M3-M5 (см. фиг. 9). Как показано на фиг. 15, EF-2 T2 и EF-3 T2 эффективно блокировали индуцированную коллагеном агрегацию тромбоцитов, тогда как неактивные мутантные контрольные мутанты не оказывали эффекта.
Затем в индуцированной коллагеном агрегации тромбоцитов в препаратах WP оценивали активность оптимизированных лигандов против GPVI RB490, RB540, RB549 и RB566 (фиг. 16). Каждый из оптимизированных лигандов эффективно блокировал индуцированную коллагеном агрегацию тромбоцитов. Относительная эффективность лигандов GPVI согласовывалась с их соответствующей аффинностью к GPVI, где RB540 (Kd для GPVI ~15 нМ) демонстрировал IC50 ~44 нМ, RB549 (Kd для GPVI ~6-7 нМ) демонстрировал IC50 ~25 нМ, RB490 (Kd для GPVI ~4-5 нМ) демонстрировал IC50 ~23 нМ и RB566 (Kd для GPVI ~2-3 нМ) демонстрировал IC50 of ~20 нМ.
Для дополнительного исследования возможного механизма, посредством которого лиганды против GPVI блокируют функцию GPVI, оценивали способность лигандов RB490, RB549 и RB566 блокировать индуцированную CRP-XL агрегацию тромбоцитов в препаратах WP (фиг. 17), как описано в пример 5. В соответствии с их способностью блокировать индуцированную коллагеном агрегацию тромбоцитов, RB490, RB549 и RB566 блокировали индуцированную CRP агрегацию тромбоцитов зависимым от дозы способом. Как и для коллагена относительная эффективность лигандов согласовывалась с их соответствующей аффинностью к GPVI, с IC50 38 нМ, 79 нМ и 98 нМ для RB566, RB490 и RB549, соответственно. Не является сюрпризом, что IC50 лигандов против GPVI при индуцированной CRP агрегации тромбоцитов является более высокой, так как CRP-XL, используемый в этих анализах является более мощным агонистом тромбоцитов, чем коллаген. Взятые вместе, эти данные соответствуют механизму действия при котором лиганды против GPVI связывают GPVI в связывающей коллаген борозде GPVI или рядом с ней, блокируя, таким образом, связывание коллагена или CRP-XL с GPVI на поверхности тромбоцита.
Также проводили сравнение активности пегилированных лигандов RB570, RB569 и RB571 против GPVI с RB490 в индуцированной коллагеном и CRP агрегации тромбоцитов в препаратах WP (фиг. 18-19). Как и выше, пегилированные лиганды демонстрировали сходное зависимое от дозы ингибирование индуцированной коллагеном и CRP агрегации тромбоцитов. Добавление к лигандам PEG немного влияло на их активность относительно RB490 в индуцированной коллагеном агрегации тромбоцитов, но не в индуцированной CRP агрегации тромбоцитов. Наибольшую эффективность блокирования индуцированной CRP агрегации в препаратах WP демонстрировал RB571 с IC50 ~33 нМ. Подобным образом RB571 демонстрировал эффективное ингибирование индуцированной коллагеном и CRP агрегации тромбоцитов в PRP (см. фиг. 20; для подробностей см. пример 5).
Лиганды против GPVI оценивали на их способность блокировать адгезию, активацию и агрегацию тромбоцитов в ответ на взаимодействие GPVI с покрытыми коллагеном поверхностями в цельной крови при течении с высокой скоростью сдвига, как описано в пример 5. Формат этого анализа отражает много характеристик функции GPVI in vivo, включая ассоциацию GPVI со связанным с поверхностью фибриллярным коллагеном в текущей цельной крови, а также воспроизведение трех ключевых функций, опосредуемых взаимодействием GPVI-коллаген, а именно адгезии, активации и агрегации тромбоцитов. RB566 и RB571 значительно снижали адгезию, активацию и агрегацию тромбоцитов (фиг. 21) по сравнению с неактивным контрольным лигандом RB471 (ответ тромбоцитов в этом анализе сравнивали с RB471 и контрольным буфером).
Таким образом, лиганды против GPVI блокируют функцию GPVI в анализах на выделенных тромбоцитах в статических условиях и в потоке цельной крови, влияя на опосредованную GPVI адгезию, активацию и агрегацию тромбоцитов. Кроме того, в соответствии с механизмом, по которому лиганды против GPVI связываются с участком связывания коллагена на GPVI или рядом с ним, они демонстрируют зависимое от дозы ингибирование индуцированной коллагеном и CRP агрегации тромбоцитов с относительной эффективностью, согласующейся с их с соответствующей аффинностью к GPVI.
Пример 7: Специфичность лигандов против GPVI к GPVI
GPVI представляет собой один из нескольких рецепторов тромбоцитов, посредством которого активация соответствующим ему лигандом стимулирует активацию и агрегацию тромбоцитов. Другие такие рецепторы включают рецепторы тромбоксана A2 (TXA2), рецепторы АДФ P2Y12 и P2Y1, рецептор тромбина PAR-1 и GP1bα, которые активируются связанным коллагеном фактором фон Виллебранда (vWF). Для определения специфичности, с которой лиганды против GPVI связываются с GPVI и блокируют функционирование тромбоцитов, оценивали действие лиганда против GPVI RB571 в анализах индуцированной АДФ, TRAP (агонист PAR-1), арахидоновой кислотой (агонист TXA2) и ристоцетином (агонист взаимодействия vWF-GP1bα) агрегации тромбоцитов, как описано в примере 5. Как представлено на фиг. 22 и 23, RB571 не влиял на индуцированную АДФ или TRAP агрегацию тромбоцитов в препаратах WP, или на индуцированную ристоцетином или арахидоновой кислотой агрегацию тромбоцитов в препаратах PRP. Таким образом, как и ожидалось на основе их высокой аффинности к GPVI и выделения посредством SELEX, лиганды против GPVI являются специфичными к GPVI.
Пример 8: Представляющие собой нуклеиновую кислоту модуляторы лигандов против GPVI
Лиганды кодируют информацию, необходимую для конструирования для них на основе правил спаривания комплементарных оснований Уотсона-Крика модуляторов или регулирующих средств, представляющих собой нуклеиновую кислоту. Эффективность данного регулирующего средства зависит от нескольких факторов, включая доступность области-мишени лиганда для нуклеации с регулирующим средством, а также отсутствие или ограниченность внутренней вторичной структуры в регулирующем средстве, которому перед формированием полного дуплекса с лигандом может быть необходима данатурация. Для определения областей лигандов против GPVI, которые могут являться предпочтительными областями для ассоциации с модуляторами, представляющими собой нуклеиновую кислоту, для EF-2 T2 и EF-3 T2 конструировали ряд регулирующих средств (см. таблицу 7 и фиг. 24).
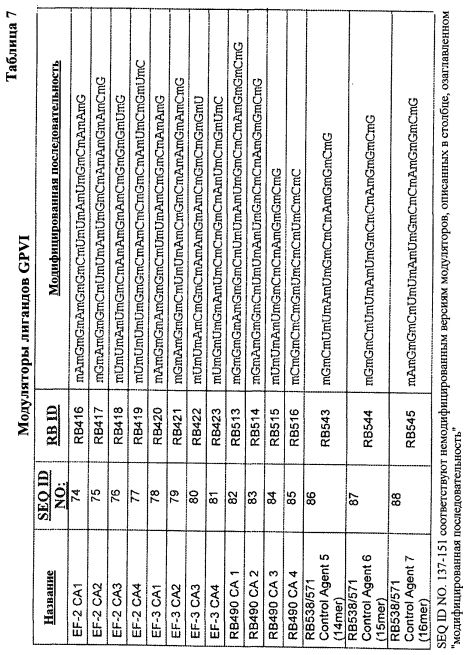
Обращающую активность регулирующих средств RB416-RB419 в отношении EF-2 T2 и RB420-423 в отношении EF-3 T2 оценивали в анализах агрегации тромбоцитов, как описано в примере 5. Как представлено на фиг. 25 и 26, регулирующие средства могли обращать действие лиганда против GPVI в пределах периода времени 10 минут с очень низким молярным избытком относительно лиганда GPVI. У регулирующих средств, сконструированных для EF-2 T2 и EF-3 T2, в отношении комплементарных областей регулирующих средств наблюдали сходную эффективность в отношении лигандов против GPVI. Наибольшую эффективность демонстрировали RB418 и RB422, ассоциирующие со сходными областями лигандов против GPVI, состоящими из 3'-конца стебля 2, петли 3, 5'-конца стебля 3 и петли 4. Ожидалось, что эти регулирующие средства продемонстрируют сходные характеристики эффективности относительно любых описанных лигандов против GPVI, с которыми они могут формировать полностью комплементарные дуплексы.
Сходный набор потенциальных регулирующих средства сконструировали для RB490 (см. таблицу 7). Как представлено на фиг. 27, эффективность, с которой эти модуляторы, представляющие собой нуклеиновую кислоту, обращали активность RB490 против GPVI соответствовала предпочтительным областям спаривания, определенным для регулирующих средств для EF-2T и EF-3T, с наибольшей эффективностью, демонстрируемой RB515. Ключевой особенностью модулятора является то, что после образования комплекса с лигандом, комплекс модулятор:лиганд не диссоциирует, и активность лиганда длительно остается обращенной. Для оценки длительности обращения RB490 посредством RB515 авторы определили длительность обращения активности RB490 против GPVI в индуцированной коллагеном агрегации тромбоцитов, как описано в примере 5. RB515 длительно обращал активность RB490 в течение 3,5 часа (фиг. 28), длительность, в течение которой препараты WP сохраняли полную агрегационную активность в ответ на коллаген. Причем RB490 в отсутствии RB515 продолжал полностью ингибировать индуцированную коллагеном агрегацию тромбоцитов в течение инкубационного периода 3,5 часа. Длительность обращающей активности RB515 в отношении активности RB490 против GPVI является сравнимой с обращающей активностью, определенной в плазме человека in vitro для обращения антикоагулянтной активности лиганда RB006 против FIXa регулирующим средством RB007, пары лиганд:регулирующее средство, которая продемонстрировала длительное обращение in vivo у людей, а также других видов.
RB571 и RB490 обладают одинаковой последовательностью оснований в области, которой комплементарен RB515, и, таким образом, RB515 является потенциальным регулирующим средством для RB571 (см. фиг. 29). Таким образом, обращающую активность RB515 для RB571 оценивали в индуцированной коллагеном и CRP активации тромбоцитов в препаратах WP и PRP. RB515, при низком молярном избытке, эффективно обращал активность RB571 против GPVI в ответ на оба агониста в обоих матриксах (фиг. 30 и 31). Молярный избыток, необходимый для полного обращения активности RB571 посредством RB515 является меньшим, чем молярный избыток, необходимый для полного обращения in vitro RB006 посредством RB007 (молярное отношение 2:1 или меньше по сравнению с 4:1 для обращения RB007 RB006), что указывает на высокую вероятность успеха обращения RB571 посредством RB515 у людей и других видов in vivo.





























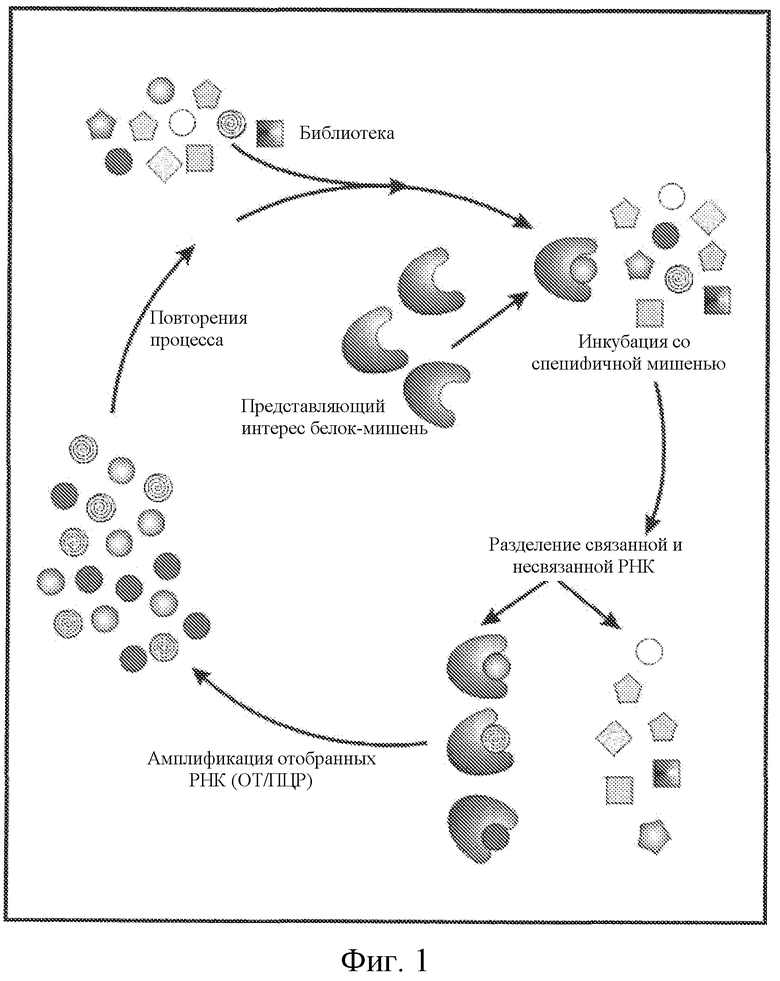

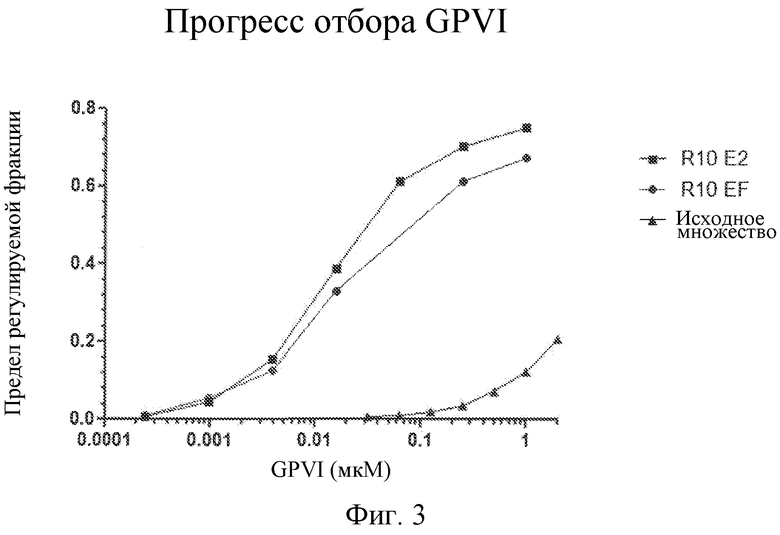
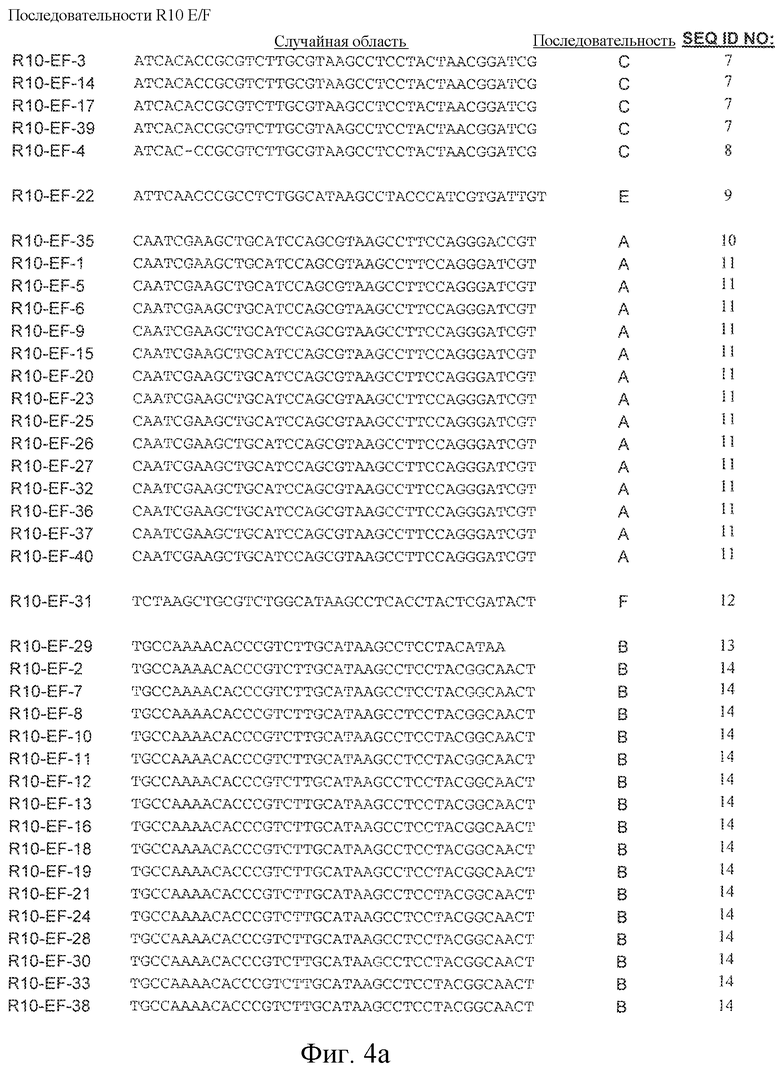

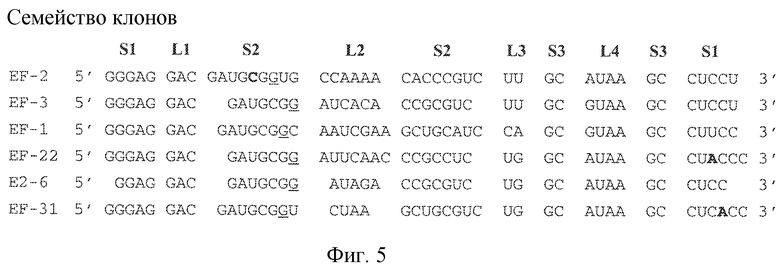
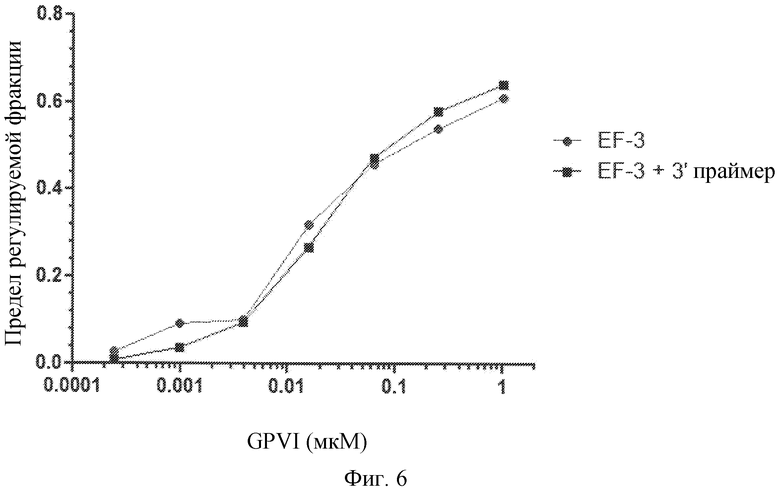
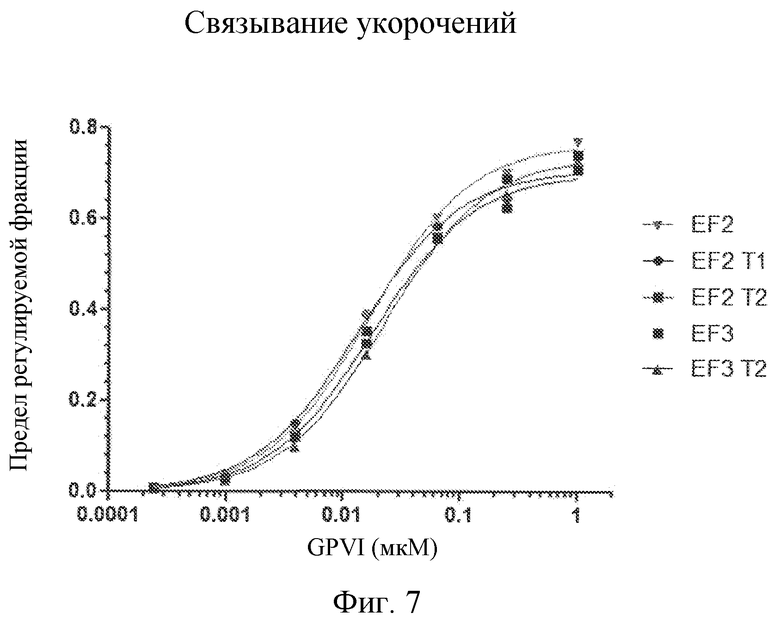
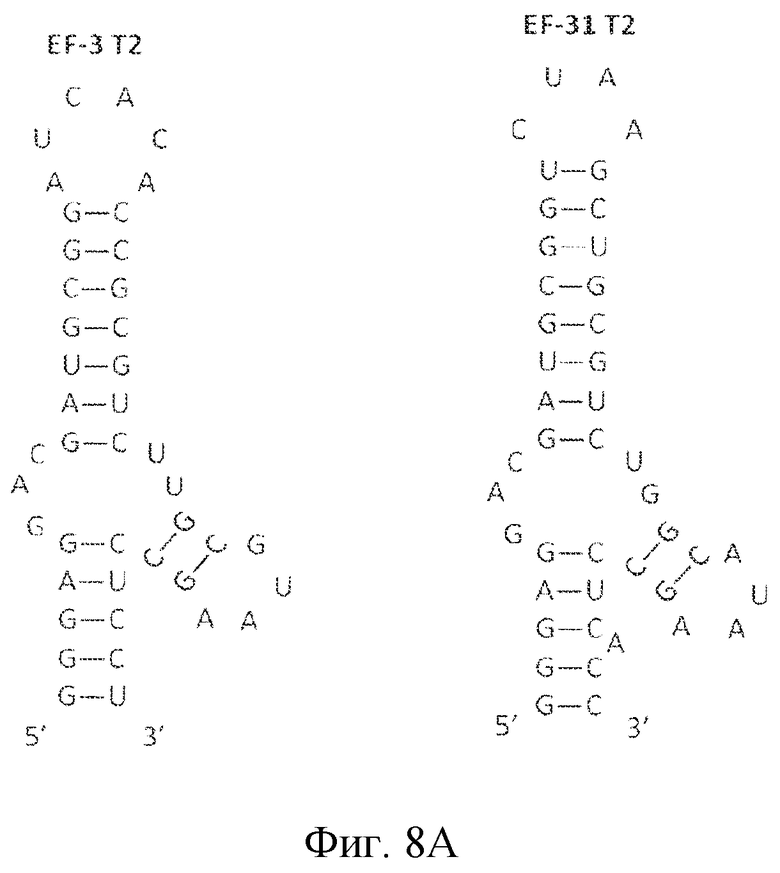
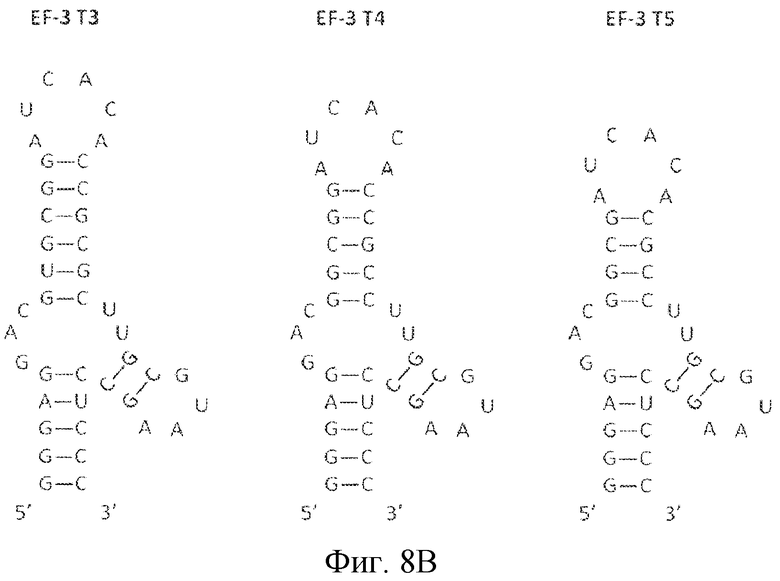
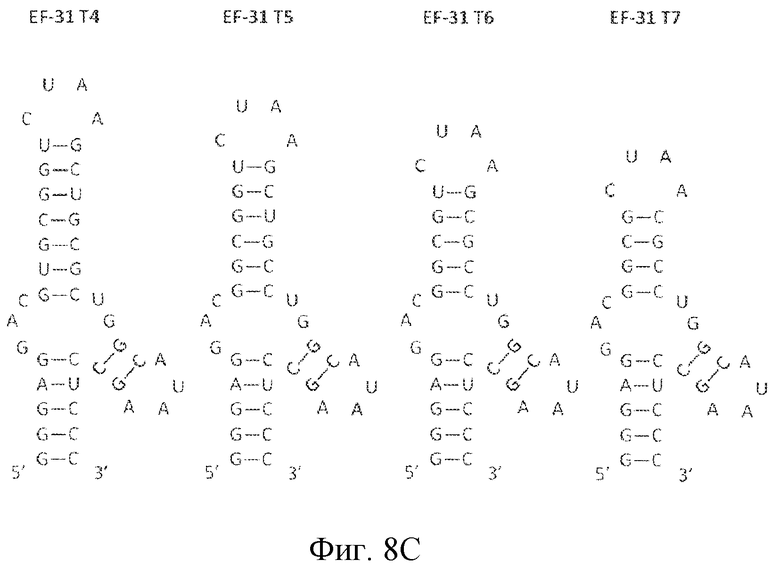
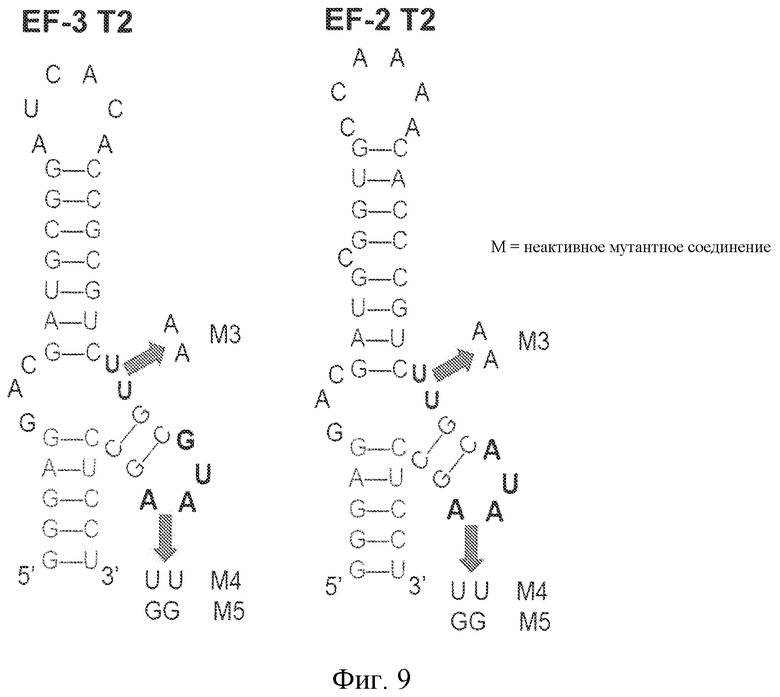
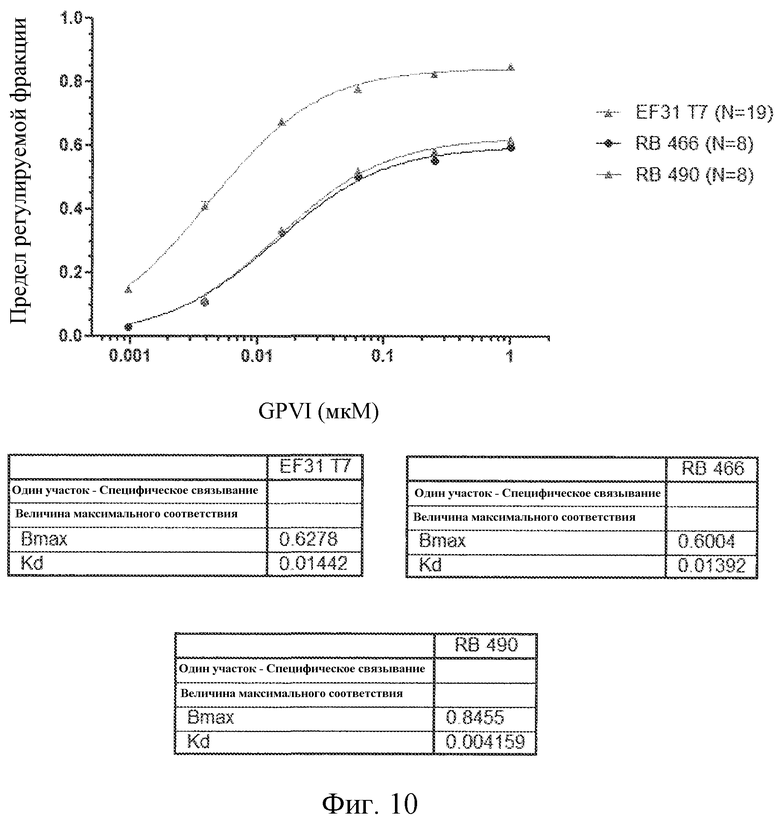
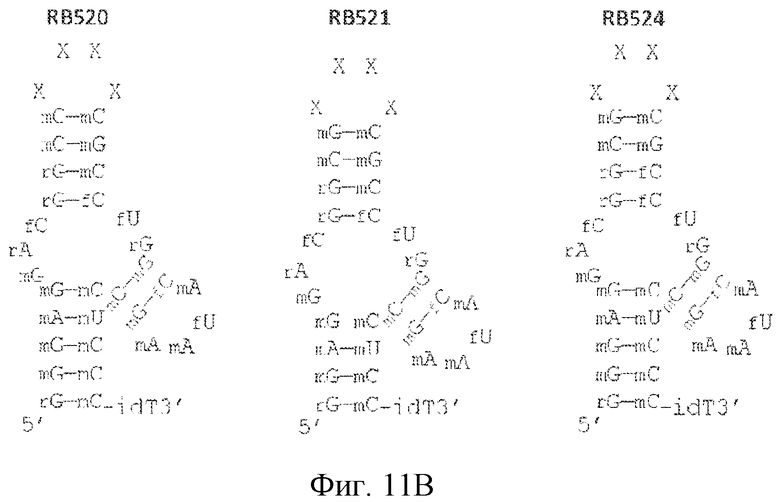
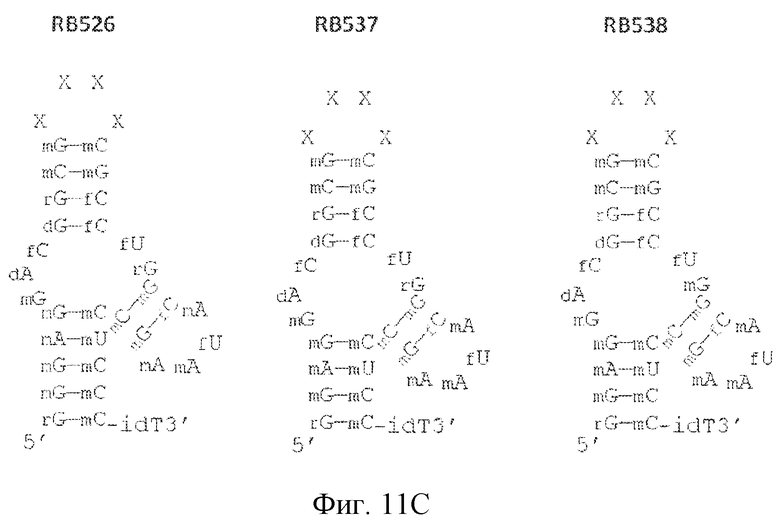
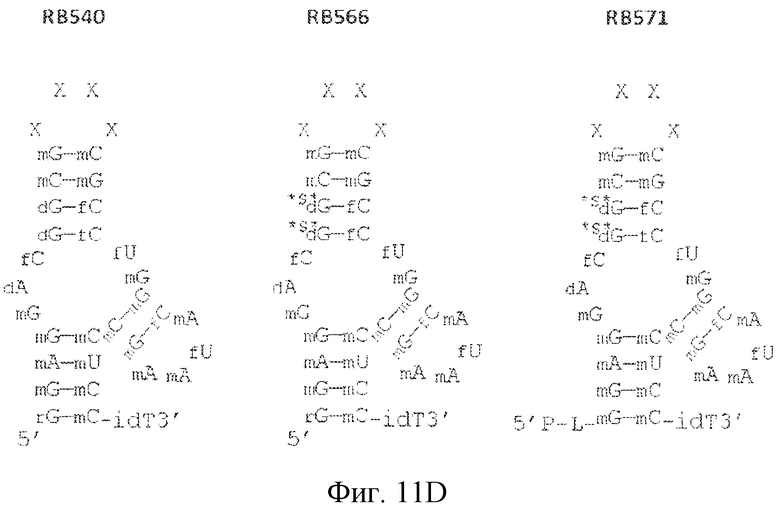
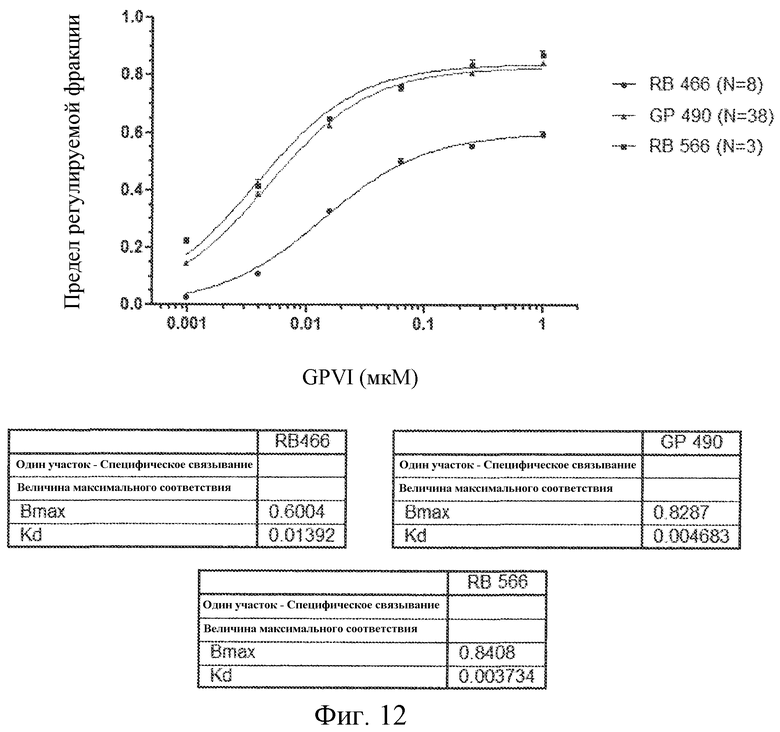
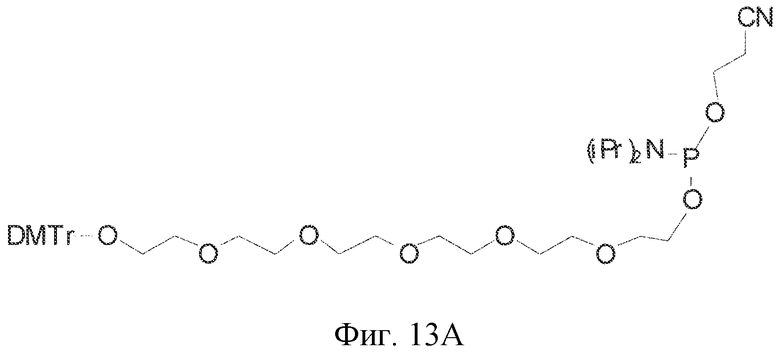
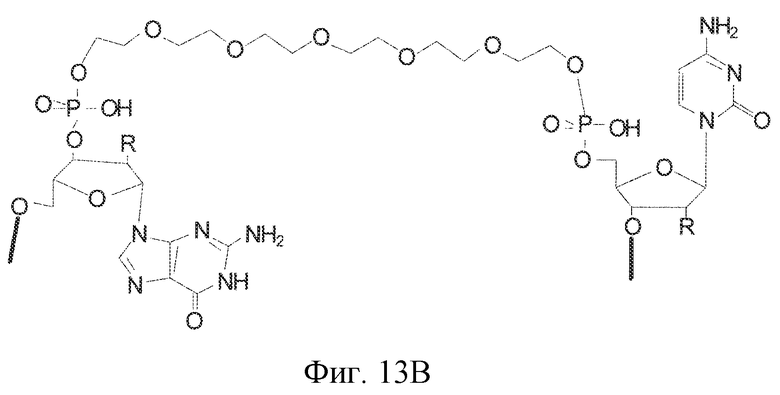
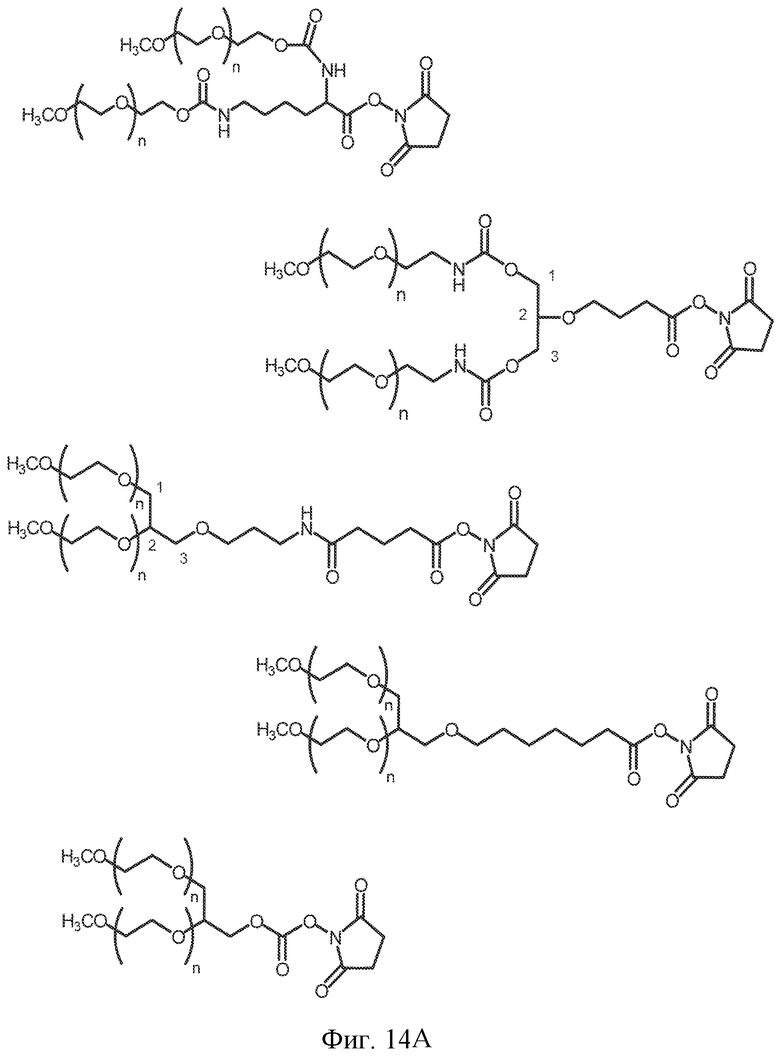
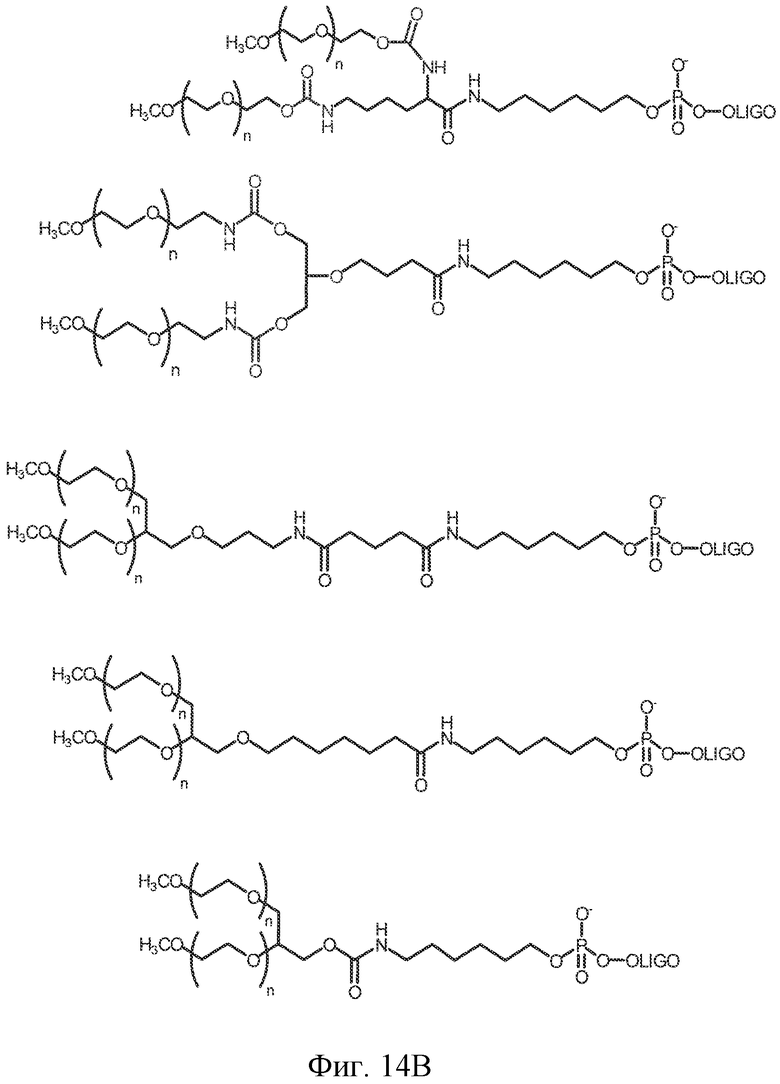
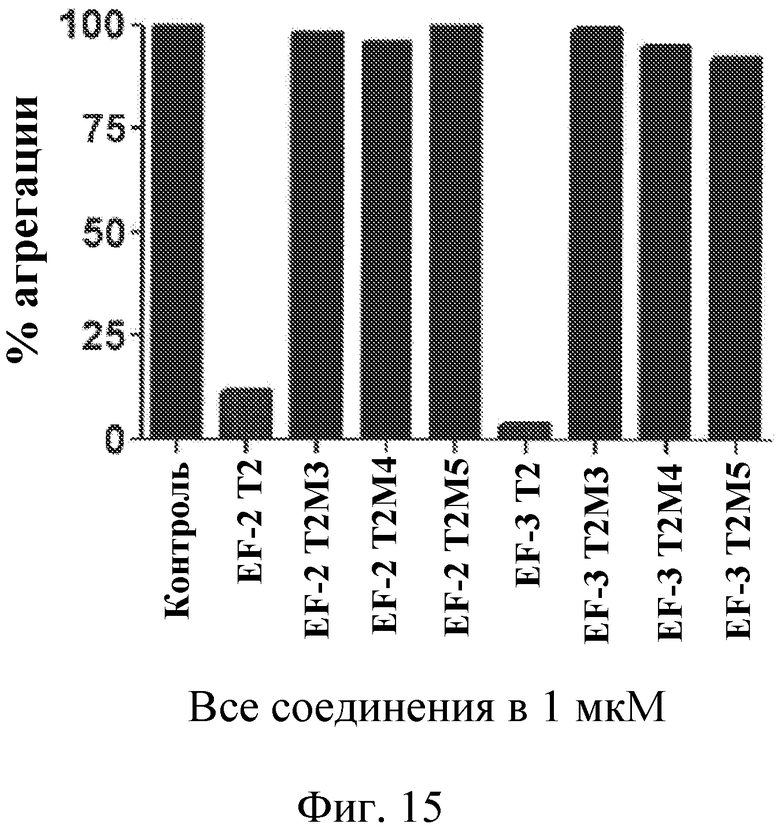
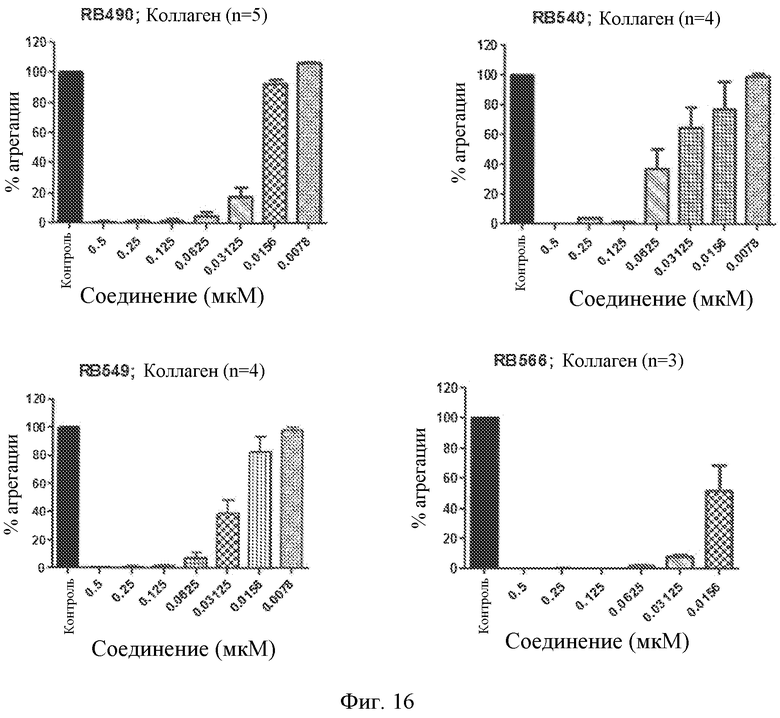
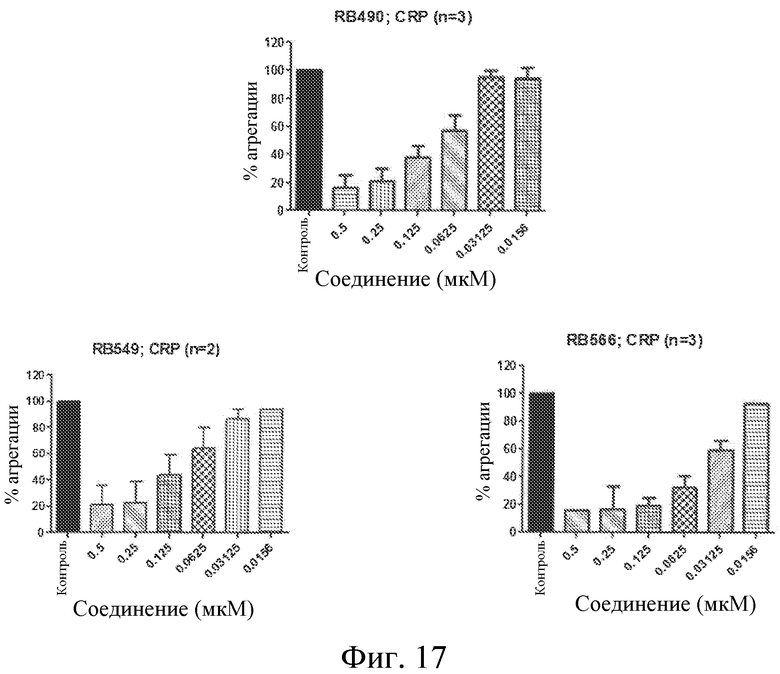
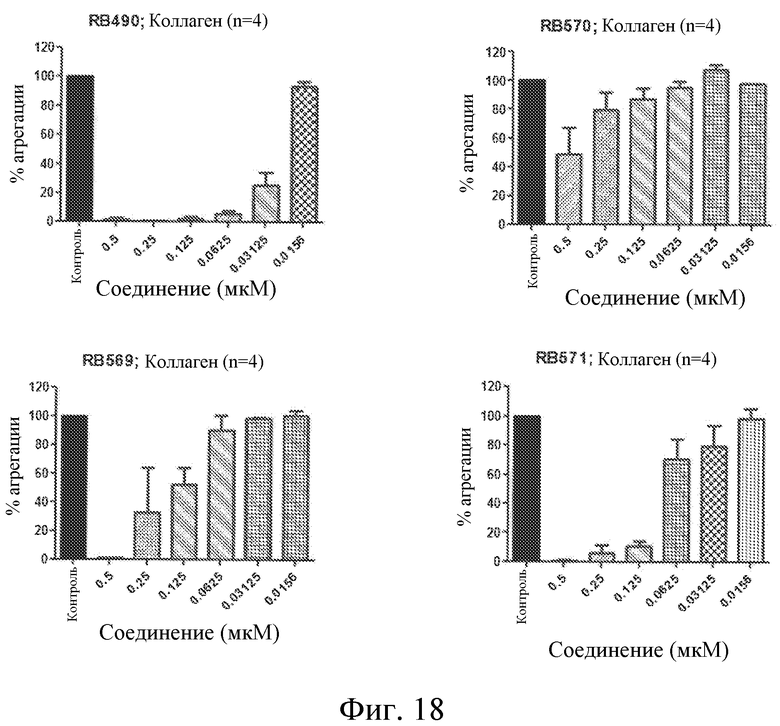
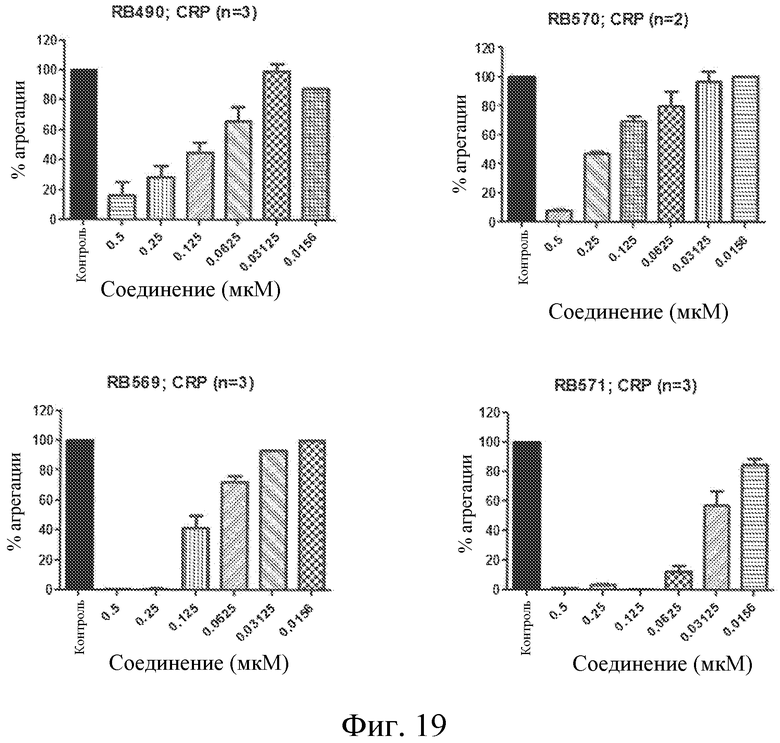
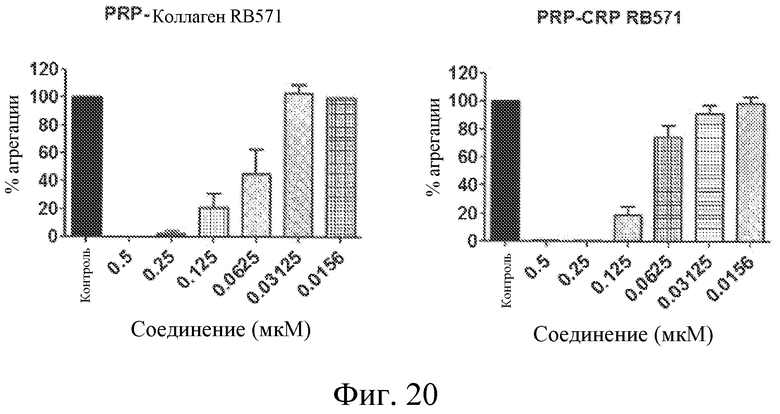
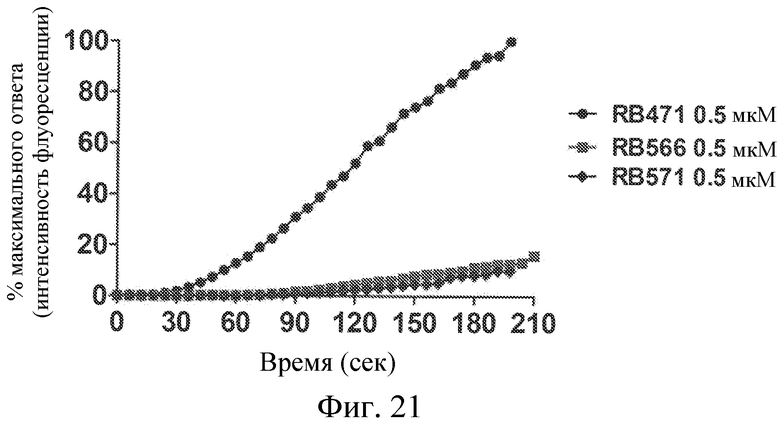

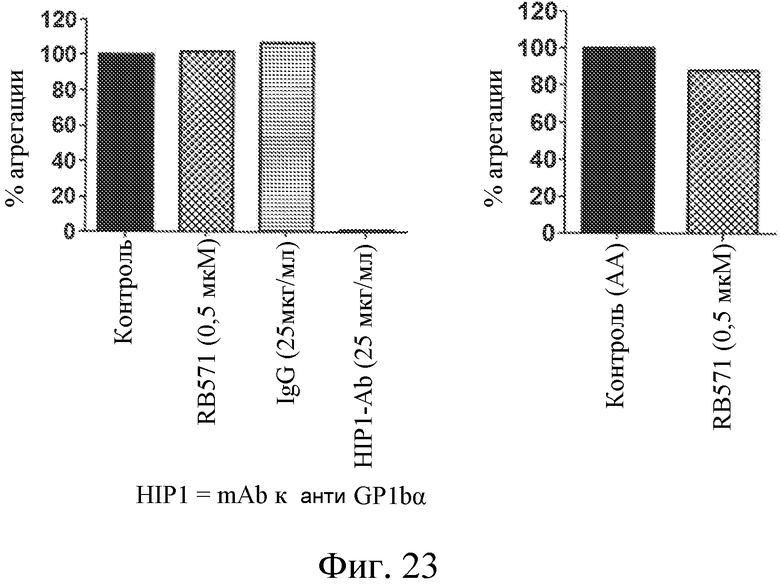
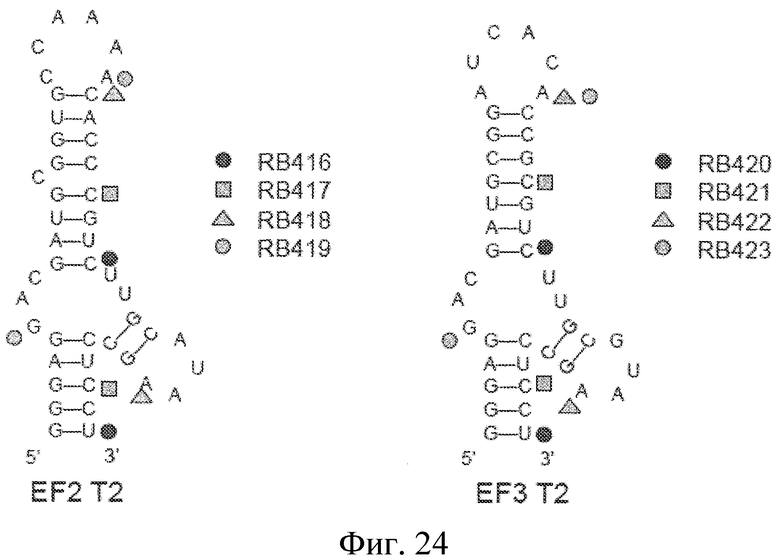


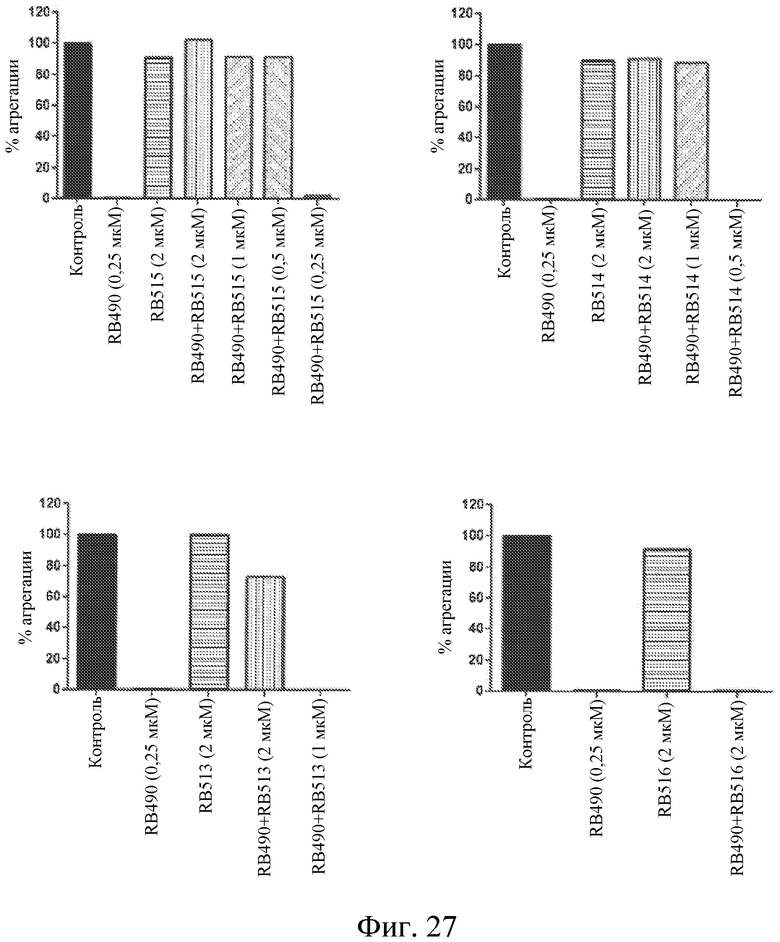
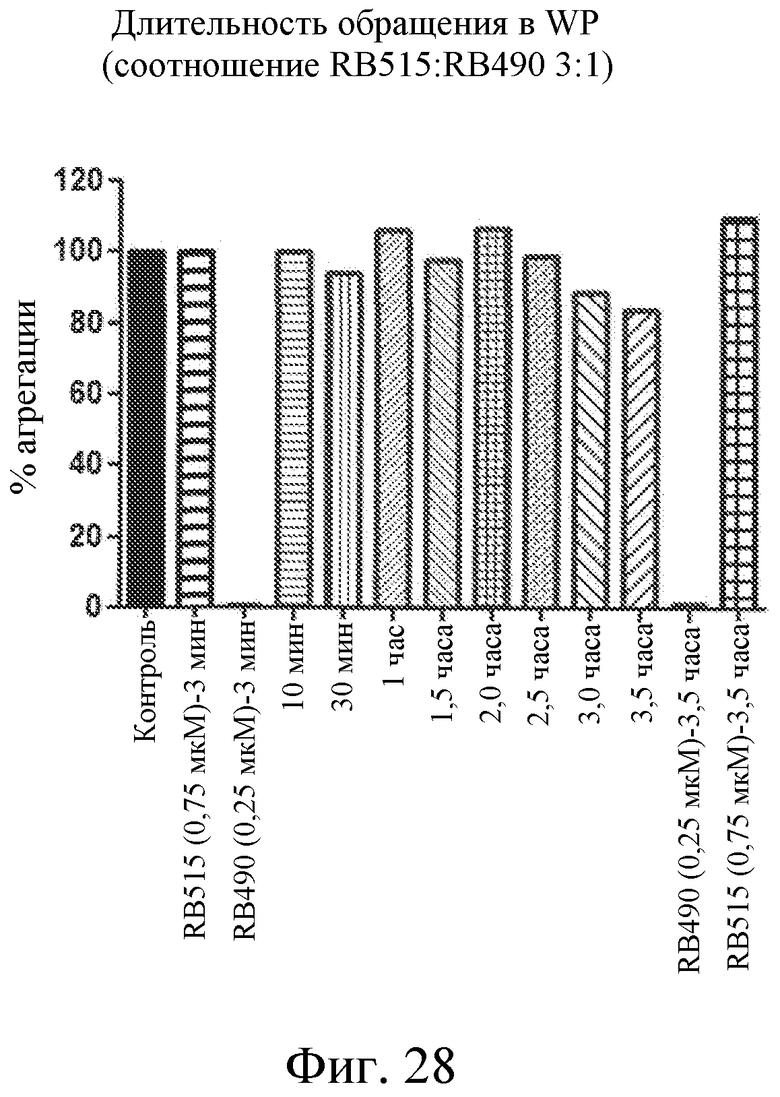
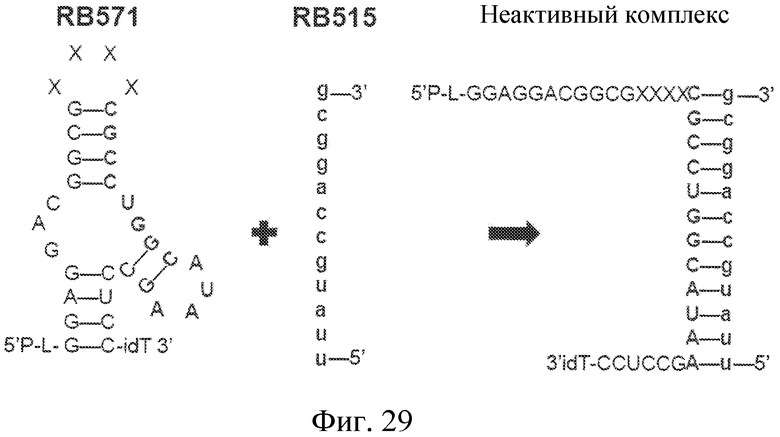
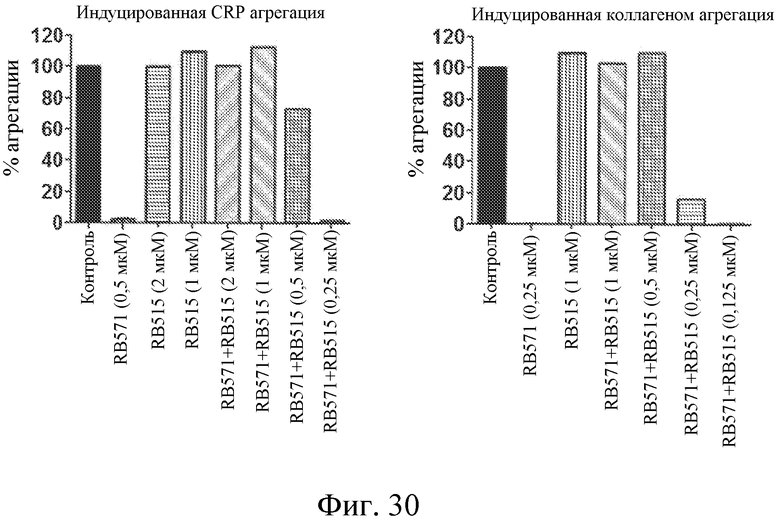
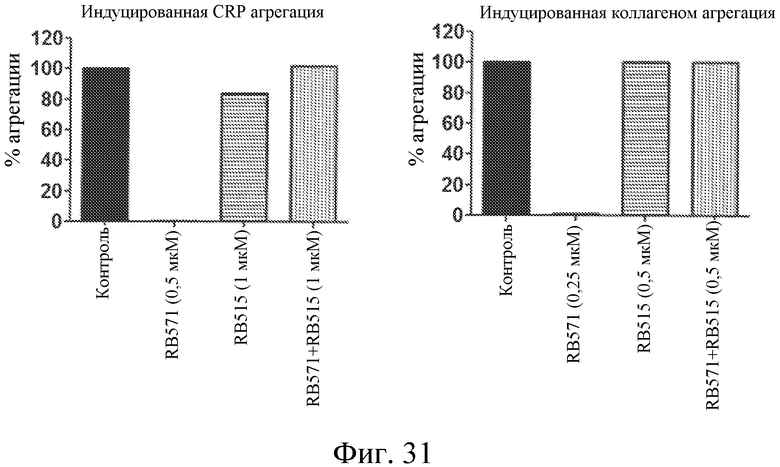
Изобретение относится к области биохимии. Заявлен лиганд гликопротеина VI (GPVI), представляющий собой нуклеиновую кислоту, который взаимодействует с гликопротеином тромбоцитов GPVI и модулирует его активность с регуляцией функции тромбоцитов. Лиганд имеет вторичную структуру, содержащую в направлении от 5′ к 3′ первый стебель, первую петлю, второй стебель, вторую петлю, третью петлю, третий стебель и четвертую петлю. Четвертая петля содержит UAA. Последовательность лиганда модифицирована для увеличения устойчивости к нуклеазам. Лиганд может применяться для ингибирования агрегации тромбоцитов. Также представлены модуляторы, ингибирующие активность лиганда, с восстановлением функции GPVI. Кроме того, изобретение относится к композициям, содержащим лиганд или модулятор, предназначенным для лечения опосредуемых тромбоцитами нарушений. 8 н. и 15 з.п. ф-лы, 39 ил., 7 табл., 8 пр.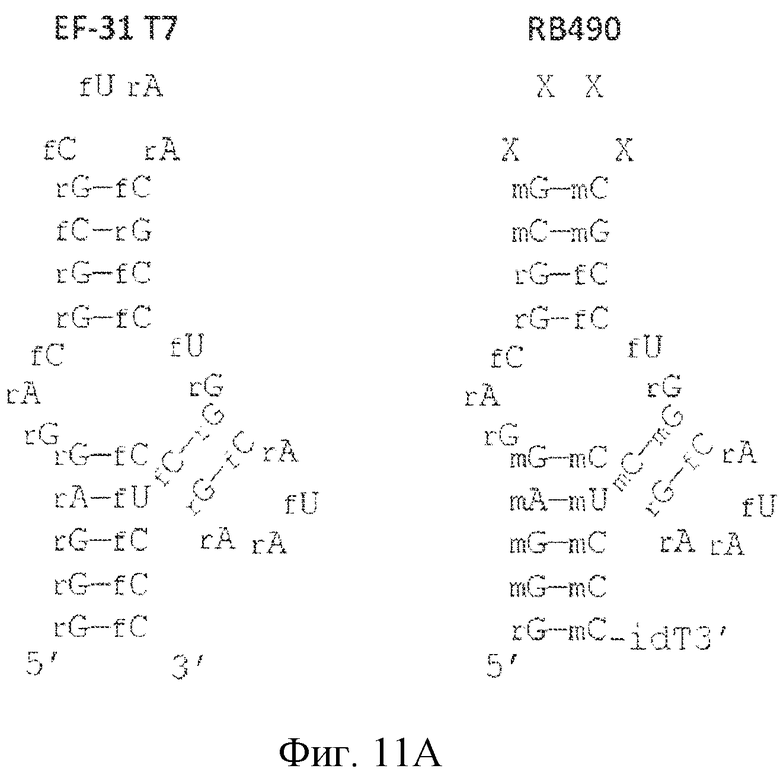
1. Лиганд GPVI, содержащий первую выделенную последовательность нуклеиновой кислоты, где указанный лиганд GPVI связывается с гликопротеином VI (GPVI) и где указанный лиганд GPVI содержит вторичную структуру, содержащую в направлении от 5′ к 3′ первый стебель, первую петлю, второй стебель, вторую петлю, третью петлю, третий стебель и четвертую петлю; и где указанная четвертая петля содержит UAA,
причем указанный лиганд представляет собой последовательность SEQ ID NO: 36, 40, 63 или 69, модифицированную для увеличения устойчивости к нуклеазам,
причем указанный лиганд необязательно содержит молекулу носителя, где молекула носителя необязательно является молекулой полиэтиленгликоля (PEG).
2. Лиганд GPVI по п.1, где вторая петля замещена спейсером.
3. Лиганд GPVI по п.1, содержащий модифицированный фосфатный остов.
4. Лиганд GPVI по п.1, содержащий SEQ ID NO:69, где между 2′-O-метил-G в положении 11 и 2′-O-метил-С в положении 12 SEQ ID NO:69 встроен спейсер.
5. Лиганд GPVI по п.4, содержащий модифицированный фосфатный остов.
6. Лиганд GPVI по п.4 или 5, содержащий молекулу полиэтиленгликоля.
7. Лиганд GPVI по п.1, где указанный лиганд GPVI представляет собой конъюгированную с разветвленным PEG последовательность, выбранную из SEQ ID NO:89, 90 или 91.
8. Фармацевтическая композиция для лечения опосредуемых тромбоцитами нарушений, содержащая терапевтически эффективное количество лиганда GPVI по любому из предшествующих пунктов.
9. Модулятор, специфически связывающийся с лигандом GPVI по п.1,
где указанный модулятор содержит вторую последовательность нуклеиновой кислоты; и
где указанный лиганд GPVI связывается с гликопротеином VI (GPVI); и
где указанный лиганд GPVI содержит первую последовательность нуклеиновой кислоты, содержащую вторичную структуру, содержащую в направлении от 5′ к 3′ первый стебель, первую петлю, второй стебель, вторую петлю, третью петлю, третий стебель и четвертую петлю; и где указанная четвертая петля содержит UAA,
причем указанный модулятор представляет собой последовательность SEQ ID NO: 74, 75, 76, 77, 78, 79, 80, 81, 82, 83 или 84, модифицированную для эффективного связывания с лигандом GPVI по п.1.
10. Фармацевтическая композиция для лечения опосредуемых тромбоцитами нарушений, содержащая терапевтически эффективное количество модулятора по п.9.
11. Модулятор для специфического связывания с лигандом GPVI по п.1, причем указанный модулятор содержит SEQ ID NO: 84, модифицированную для эффективного связывания с лигандом GPVI по п.1.
12. Модулятор по п.11, где указанный модулятор представляет собой последовательность SEQ ID NO: 84.
13. Фармацевтическая композиция для лечения опосредуемых тромбоцитами нарушений, содержащая терапевтически эффективное количество модулятора по п.11 или 12.
14. Применение терапевтически эффективного количества лиганда GPVI по п.1 для лечения опосредуемых тромбоцитами нарушений, включающее введение нуждающемуся в этом индивидууму терапевтически эффективного количества лиганда GPVI.
15. Применение по п.14, где опосредуемое тромбоцитами нарушение выбрано из группы, состоящей из сосудистого нарушения, цереброваскулярного нарушения, опосредуемого тромбоцитами воспалительного нарушения, связанного с диабетом нарушения или злокачественной опухоли.
16. Применение по п.15, где сосудистое нарушение выбрано из группы, состоящей из острых коронарных синдромов, тромбоза, тромбоэмболии, заболевания периферических сосудов и транзиторной ишемической атаки.
17. Применение по п.15, где цереброваскулярное нарушение выбрано из группы, состоящей из транзиторной ишемической атаки, ишемического инсульта и эмболии.
18. Применение по п.15, где опосредуемое тромбоцитами воспалительное нарушение выбрано из группы, состоящей из артрита, ревматоидного артрита, псориатического артрита, реактивного артрита, воспалительного заболевания кишечника, анкилозирующего спондилита и склеродермии.
19. Применение по п.15, где связанное с диабетом нарушение выбрано из группы, состоящей из диабетической ретинопатии, диабетической васкулопатии, атеросклероза, ишемического инсульта, заболевания периферических сосудов, острого повреждения почек и хронической почечной недостаточности.
20. Применение по п.15, где злокачественная опухоль выбрана из группы, состоящей из рака легких, рака молочной железы, рака предстательной железы, рака поджелудочной железы, злокачественной опухоли головного мозга, злокачественной опухоли кости и рака печени.
21. Применение лиганда GPVI по п.1 для ингибирования агрегации тромбоцитов у нуждающегося в этом индивидуума, включающее введение индивидууму эффективного количества лиганда GPVI.
22. Применение по п.21, где индивидууму проводят воздействие на сердечную функцию.
23. Применение по любому из пп.14-22, дополнительно включающее введение индивидууму модулятора по п.9, где указанный модулятор специфически связывается с лигандом GPVI.
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| BIGALKE B | |||
| et al., Platelet collagen receptor glycoprotein VI as a possible novel indicator for the acute coronary syndrome, Am Heart J., 2008, vol.156 no.1, pp.193-200 | |||
| РЕКОМБИНАНТНЫЙ ПОЛИПЕПТИД, СПОСОБ ИНГИБИРОВАНИЯ АГРЕГАЦИИ ТРОМБОЦИТОВ, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК (ВАРИАНТЫ), РЕКОМБИНАНТНЫЙ ШТАММ E.COLI (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ ПОЛИПЕПТИДА | 1991 |
|
RU2109816C1 |

