Предпосылки изобретения
Повреждение кровеносных сосудов запускает ряд событий для репарации этого повреждения и прекращения выделения крови из сосуда. Этот процесс известен как гемостаз. Тромбоциты играют раннюю роль в гемостазе путем образования тромба или пробки для временной репарации повреждения сосуда. В норме тромбоциты не взаимодействуют с эндотелиальной выстилкой стенок сосуда, но повреждение кровеносных сосудов вследствие несчастного случая или во время хирургических процедур может разрушать эндотелиальные клетки. В зависимости от степени повреждения, различные субэндотелиальные элементы, такие как коллагены, эластическая мембрана или клетки гладких мышц с ассоциированными фибриллярными коллагенами, будут экспонироваться протекающей кровью.
При экспонировании субэндотелия после повреждения сосуда тромбоциты, перемещающиеся в локальном кровотоке, взаимодействуют с экспонированным субэндотелиальным матриксом, содержащим коллаген, и замедляются. Последующее взаимодействие между рецепторами на поверхности тромбоцитов и экспонированным слоем коллагена приводит к связыванию и активации тромбоцитов, приводя к задержке локального кровотока. Связанные тромбоциты активируются и образуют агрегаты с тромбоцитами в проходящем кровотоке посредством образования фибриногеновых межтромбоцитных мостиков (Moroi and Jung, Frontiers in Bioscience 3:719-28, 1998; Barnes et al., Atherosclerosis XI, Jacotot et al., eds., Elsevier Science, pp. 299-306, 1998 и Barnes et al., Curr. Opin. Hematol. 5:314-20, 1998).
Гемостатическая реакция состоит из ряда стадий и зависит от степени повреждения кровеносного сосуда, компонентов конкретных экспонированных кровеносных сосудов и условий кровотока в поврежденной зоне (Rand et al., Thrombosis and Haemostasis 78:445-50, 1997). Экспонирование субэндотелиального матрикса (коллагена типа IV и фактора фон Виллебранда), например, во время мягкого васкулярного повреждения, стимулирует низкую степень адгезии и агрегации в зонах с условиями низкого кровотока. Повреждения, которые приводят к большей степени травмы сосуда, и экспонирование дополнительных сосудистых компонентов, таких как внутренняя эластическая мембрана и связанные с эластином микрофибриллы, будет стимулировать образование более прочных агрегатов тромбоцитов. Тяжелая васкулярная травма, обнажающая фибриллярные коллагены, провоцирует тромботическую реакцию тромбоцитов, которая защищает жертву от избыточной потери крови (Rand et al., ibid.).
Ингибиторы гемостаза были бы полезны для увеличения кровотока после васкулярного повреждения и для приведения в порядок коллагеновых поверхностей.
Фактор комплемента C1q состоит из шести копий трех родственных полипептидов (цепей А, В и С), причем каждый из этих полипептидов имеет длину приблизительно 225 аминокислот с расположенным вблизи амино-конца коллагеновым доменом и карбоксиконцевым глобулярным районом. Шесть районов с тройной спиралью образуются коллагеновыми доменами из шести цепей А, шести цепей В и шести цепей С, образуя центральный район и шесть стеблей. Глобулярный участок (головка) образуется связыванием глобулярного карбоксиконцевого домена А-, В- и С-цепи. Таким образом, C1q состоит из шести глобулярных головок, соединенных через шесть коллагеноподобных стеблей с центральным фибриллярным районом. Sellar et al., Biochem. J. 274: 481-90, 1991. Эту конфигурацию часто называют букетом цветов. Асrp30 имеет сходную структуру букета, образуемую из единственного типа полипептидной цепи.
Было обнаружено, что C1q стимулирует защитные механизмы, а также запускает генерирование токсичных разновидностей кислорода, которые могут вызывать повреждение ткани (Tenner, Behring Inst. Mitt. 93:241-53, 1993). Сайты связывания C1q найдены на тромбоцитах. Кроме того, комплемент и C1q играют роль в воспалении. Активация комплемента инициируется связыванием C1q с иммуноглобулинами.
Ингибиторы C1q и пути комплемента были бы полезны для противовоспалительных применений, ингибирования активации комплемента и тромботической активности.
Данное изобретение обеспечивает такие полипептиды для этих и других использований, которые должны быть очевидными специалистам с квалификацией в данной области из приведенных здесь обсуждений.
Сущность изобретения
В одном аспекте данное изобретение обеспечивает способ стимуляции кровотока в сосудистой сети млекопитающего, предусматривающий введение указанному млекопитающему терапевтически эффективного количества белка, родственного комплементу адипоцитов, в фармацевтически приемлемом носителе, посредством чего указанный родственный комплементу адипоцитов белок снижает тромбогенную активность и активность комплемента в указанной сосудистой сети. В одном варианте родственный комплементу адипоцитов белок содержит полипептид, содержащий последовательность аминокислотных остатков, которая является по меньшей мере на 75% идентичной по аминокислотной последовательности остаткам 26-281 SEQ ID NО:2, причем указанная последовательность содержит: повторы Gly-Хаа-Хаа или Gly-Xaa-Pro, образующие коллагеновый домен, где Хаа является любой аминокислотой, и карбоксиконцевую глобулярную часть. В родственном варианте этот полипептид содержит последовательность аминокислотных остатков, которая является по меньшей мере на 90% идентичной по аминокислотной последовательности остаткам 22-281 SEQ ID NО:2. В другом варианте этот полипептид содержит аминокислотную последовательность, которая является по меньшей мере на 90% идентичной по аминокислотной последовательности остаткам 26-281 SEQ ID NО:2. Еще в одном варианте любые различия между указанным полипептидом и SEQ ID NО:2 обусловлены консервативными заменами аминокислот. В другом варианте коллагеновый домен состоит из 13 повторов Gly-Xaa-Xaa и 1 повтора Gly-Xaa-Pro. Еще в одном варианте глобулярный домен состоит из десяти бета-складок. В родственном варианте эти бета-складки ассоциированы с аминокислотными остатками, соответствующими 147-151, 170-172, 178-181, 191-203, 207-214, 219-225, 227-239, 244-250 и 269-274 SEQ ID NO:2. Еще в одном варианте этот полипептид содержит остатки 1-281 SEQ ID NОO2 или остатки 1-281 SEQ ID NО:44.
Данное изобретение обеспечивает также полипептид, находящийся в комплексе с другим полипептидом с образованием олигомера. В одном варианте эти полипептиды образуют комплекс при помощи межмолекулярных дисульфидных связей. В другом варианте этот олигомер является тримером. Еще в одном варианте этот олигомер является гексамером. Еще в одном варианте этот мультимер является 18-мером.
В другом варианте этот полипептид снижает тромбогенную активность и активность комплемента ингибированием пути комплемента и ингибированием опосредованной коллагеном адгезии, активации или агрегации тромбоцитов. В другом варианте полипептид вводят до, во время или после острого сосудистого повреждения в указанном млекопитающем. Еще в одном варианте это повреждение обусловлено хирургической реконструкцией (пластикой) сосудов. В родственном варианте хирургическая пластика сосудов включает в себя пластическую операцию на сосудах (реконструкцию сосудов), трансплантат шунта венечной артерии, эндартерэктомию, микрососудистую репарацию или анастомоз сосудистого трансплантата. В другом родственном варианте повреждение вызвано травмой, ударом или аневризмой.
В другом аспекте данное изобретение обеспечивает способ приведения в порядок (восстановления) поврежденных коллагеновых тканей в млекопитающих, предусматривающий введение указанному млекопитающему терапевтически эффективного количества родственного комплементу адипоцитов белка; посредством чего указанный белок делает поврежденную коллагеновую ткань инертной относительно активации комплемента, тромботической активности или иммунной активации. В одном варианте поврежденные коллагеновые ткани индуцируются повреждением, ассоциированным с ишемией и реперфузией. В другом варианте повреждение включает в себя ишемию, связанную с травматическим повреждением, интестинальную странгуляцию (сдавление) или повреждение, связанное с реакциями перед восстановлением или после восстановления кровотока. Еще в одном варианте этот полипептид вводят млекопитающему, страдающему от ишемии и перерывов в кровотоке, вызываемых искусственным кровообращением, инфаркта миокарда или посттравматического вазоспазама (сужения кровеносных сосудов). В родственном варианте посттравматический вазоспазм включает в себя удар, чрескожную чреспросветную ангиопластику, эндартерэктомию, случайную сосудистую травму или хирургически индуцированную сосудистую травму.
Еще в одном варианте данное изобретение обеспечивает способ приспособления поверхности протезного биоматериала для применения в ассоциации с млекопитающим, предусматривающий введение указанному млекопитающему терапевтически эффективного количества родственного комплементу адипоцитов белка; посредством чего указанный полипептид делает поверхность указанного протезного биоматериала инертной относительно активации комплемента, тромботической активности или иммунной активности. В одном варианте поверхность указанного протезного материала покрывают коллагеном или фрагментами коллагена, желатином, фибрином или фибронектином.
В другом варианте данного изобретения обеспечен способ опосредования заживления репарации ран в млекопитающем, предусматривающий введение указанному млекопитающему терапевтически эффективного количества родственного комплементу адипоцитов белка; посредством чего указанный полипептид ускоряет прогрессирование заживления ран.
Краткое описание графических материалов.
Фиг.1 иллюстрирует множественное сопоставление полипептида zsig37 данного изобретения и HUMUPST2_1 (Maeda et al., Biochem. Biophys. Res. Comm. 221(2): 286-9, 1996); C1QA_HUMAN (Sellar et al., Biochem. J. 274: 481-90, 1991, Reid, Biochem. J. 179: 367-71, 1979; и Reid et al., Biochem. J. 203: 559-69, 1982), HP25_TAMAS (Takamatsu et al., Mol. Cell. Biol. 13: 1516-21, 1993 и Kondo and Kondo, J. Biol. Chem. 267: 473-8, 1992); HP27_TAMAS (Takamatsu et al., и Kondo and Kondo, выше); и CERL_RAT (Wada and Ohtani, Brain Res. Mol. Brain Res. 9: 71-7, 1991).
Фиг.2 представляет собой матрицу, показывающую процентную идентичность аминокислот в сравнении шести белков, показанных во множественном сопоставлении фиг.1.
Фиг.3а показывает связывание zsig37-FITC с коллагеном типа VI.
Фиг.3b показывает конкуренцию немеченого zsig37 с FIТС-меченым zsig37, связанным с коллагеном типа VI.
Фиг.4 показывает связывание комплемента C1q-FITC с zsig37.
Фиг.5 показывает ингибирование активности комплемента человека посредством zsig37.
Фиг.6 показывает процентную агрегацию тромбоцитов коллагеном в присутствии zsig37.
Фиг.7 показывает пролиферацию фибробластов SK5 в присутствии zsig37.
Подробное описание изобретения
Перед подробным изложением этого изобретения для его понимания может быть полезно определить следующие термины.
Термин "аффинная метка" применяется здесь для обозначения пептидного сегмента, который может быть присоединен к полипептиду для обеспечения очистки или обнаружения этого полипептида или обеспечения сайтов для присоединения этого полипептида к субстрату. В принципе, любой пептид или белок, для которого доступны антитело или другой агент специфического связывания, может быть использован в качестве аффинной метки. Аффинные метки включают в себя полигистидиновый участок, протеин A (Nilsson et al., EMBO J. 4:1075 (1985); Nilsson et al., Methods Enzymol. 198:3 (1991), глутатион-S-трансферазу (Smith and Johnson, Gene 67:31 (1988), вещество Р, FLAGTM-пептид (Норр et al., Biotechnology 6:1204-1210 (1988), доступный из Eastman Kodak Co., New Haven, CT), стрептавидинсвязывающий пептид или другой антигенный эпитоп или связывающий домен. См., в общем, Ford et al., Protein Expression and Purification 2:95-107 (1991). ДНК, кодирующие аффинные метки, доступны из коммерческих источников (например, Pharmacia Biotech, Piscataway, NJ).
Термин "комплементы полинуклеотидных молекул" обозначает полинуклеотидную молекулу, имеющую последовательность комплементарных оснований и обратную ориентацию в сравнении со ссылочной последовательностью. Например, последовательность 5' ATGCACGGG 3' является комплементарной последовательности 5' CCCGTGCAT 3'.
Термин "вырожденная последовательность нуклеотидов" обозначает последовательность нуклеотидов, которая включает в себя один или несколько вырожденных кодонов (в сравнении со ссылочной молекулой полинуклеотида, которая кодирует полипептид). Вырожденные кодоны содержат отличающиеся триплеты нуклеотидов, но кодируют один и тот же аминокислотный остаток (т.е. каждый из триплетов GAU и GAC кодирует Asp).
Термин "изолированный" ("выделенный") в применении к полинуклеотиду означает, что этот полинуклеотид был удален из его природной генетической среды и, следовательно, не содержит других посторонних или нежелательных кодирующих последовательностей и находится в форме, пригодной для применения в системах получения генетически сконструированных белков. Такие изолированные молекулы являются молекулами, которые отделены от их природного окружения, и включают в себя клоны к ДНК и геномные клоны. Выделенные молекулы ДНК данного изобретения не содержат других генов, с которыми они обычно связаны, но могут включать в себя природно встречающиеся 5'- и 3'-нетранслируемые районы, такие как промоторы и терминаторы. Идентификация ассоциированных районов будет очевидной для специалиста с обычной квалификацией в данной области. См., например, Dynan and Tijan, Nature 316:774-78 (1985).
"Выделенный" полипептид или белок является полипептидом или белком, который обнаруживается в условиях, иных, чем его нативное окружение, например, отдельно от крови и ткани животного. В предпочтительной форме, выделенный белок является по существу не содержащим других полипептидов, в частности, других полипептидов животного происхождения. Предпочтительно, обеспечивать полипептиды в высокоочищенной форме, а именно, с чистотой более 95%, более предпочтительно с чистотой более 99%. При использовании в этом контексте, термин "выделенный" не исключает присутствия того же самого полипептида в альтернативных физических формах, таких как димеры или альтернативно гликозилированные или дериватизованные формы.
Термин "ортолог" обозначает полипептид или белок, полученный из одного вида, который является функциональной копией полипептида или белка из другого вида. Различия последовательностей среди ортологов являются результатом видообразования.
Термин "полинуклеотид" обозначает одно- или двухцепочечный полимер дезоксирибонуклеотидных или рибонуклеотидных оснований, считываемых от 5'-конца к 3'-концу. Полинуклеотиды включают в себя РНК и ДНК и могут быть изолированы из природных источников, синтезированы in vitro или получены из комбинации природных и синтетических молекул. Размеры полинуклеотидов выражаются как пары нуклеотидов (сокращенно "п.н."), нуклеотиды ("нт") или тысячи пар нуклеотидов ("т.п.н."). Там, где позволяет контекст, два последних термина могут описывать полинуклеотиды, которые являются одноцепочечными или двухцепочечными. При применении этого термина к двухцепочечным молекулам его используют для обозначения общей длины, и должно быть понятно, что он является эквивалентным термину "пары нуклеотидов". Специалистам с квалификацией в данной области будет понятно, что две цепи двухцепочечного полинуклеотида могут слегка отличаться по длине и что их концы могут быть расположены зигзагами в результате ферментативного расщепления; таким образом, не все нуклеотиды в двухцепочечной полинуклеотидной молекуле могут быть спаренными. Такие неспаренные концы обычно не будут превышать длину 20 нт.
"Полипептид" является полимером аминокислотных остатков, соединенных пептидными связями, получен ли он природным путем или синтетическим путем. Полипептиды, имеющие менее приблизительно 10 аминокислотных остатков, обычно называют "пептидами".
"Зондами и/или праймерами", в применении здесь, могут быть РНК или ДНК. ДНК может быть либо кДНК, либо геномной ДНК. Полинуклеотидные зонды и праймеры являются одноцепочечными или двухцепочечными ДНК или РНК, обычно синтетическими олигонуклеотидами, но они могут быть генерированы из клонированных последовательностей кДНК или геномных последовательностей или их комплементов. Аналитические зонды обычно имеют длину по меньшей мере 20 нуклеотидов, хотя могут быть использованы несколько более короткие зонды (14-17 нуклеотидов). ПЦР-праймеры имеют длину по меньшей мере 5 нуклеотидов, предпочтительно 15 или более нуклеотидов, более предпочтительно 20-30 нуклеотидов. Короткие полинуклеотиды могут быть использованы, когда целью анализа является небольшой участок гена. Для валового (gross) анализа генов полинуклеотидный зонд может содержать полный экзон или более. Зонды могут быть мечеными для обеспечения детектируемого сигнала, например, ферментом, биотином, радионуклидом, флуорофором, хемилюминесцирующим соединением, парамагнитной частицей и т.п., которые являются коммерчески доступными из многих источников, таких как Molecular Probes, Inc., Eugene, OR, и Amersham Corp., Arlington Heights, IL, с использованием способов, которые хорошо известны в этой области.
Должно быть понятно, что молекулярные массы и длины полимеров, определяемые неточными аналитическими методами (например, гель-электрофорезом), являются приблизительными величинами. Когда такая величина выражается как "около" X или "приблизительно" X, указанная величина X должна пониматься как величина, определенная с точностью до ±10%.
Данное изобретение основано частично на открытии, что новый гомолог родственного комплементу адипоцитов белка ингибирует опосредованную коллагеном активацию тромбоцитов и путь комплемента, включающий в себя C1q. Этот белок назван zsig37 и полно описан в обычно переданной опубликованной патентной заявке РСТ WO 99/04000.
Нуклеотидная последовательность zsig37 (SEQ ID NО:1) кодирует полипептид (SEQ ID NО:2), имеющий аминоконцевую сигнальную последовательность, смежный N-концевой район не-гомологии, укороченный коллагеновый домен, состоящий из повторов Gly-Xaa-Xaa или Gly-Xaa-Pro, и карбоксиконцевую глобулярную часть. Эта новая полинуклеотидная последовательность содержит также длинный 3'-нетранслируемый район. Эта общая структура полипептида, представленная выше, является общей со структурой Асrр30 и HUMUPST2_1, за исключением того, что коллагеноподобный домен каждого из этих белков является более длинным, чем этот домен полипептидов zsig37. Также последовательность ДНК HUMUPST2_1 характеризуется длинным 3'-нетранслируемым районом. Кроме того, Асrр30 и все последовательности, сопоставленные на фиг.1, за исключением CERL_RAT, имеют общий консервативный остаток цистеина в положении 187 полипептида zsig37, как показано на фиг.1 и в SEQ ID NO:2. Полипептиды zsig37 данного изобретения включают в себя также предполагаемый N-связанный сайт гликозилирования при аминокислоте 93 (Asn) SEQ ID NO:2.
Анализ тканевого распределения мРНК, соответствующей zsig37, показал, что экспрессия была наивысшей в сердце и плаценте, с относительно менее интенсивными сигналами в почке, яичнике, надпочечнике и скелетной мышце и более низкими сигналами в широком разнообразии других тканей, присутствующих на Нозерн-блоте.
Для zsig37 было установлено гомологичное родство с родственным комплементу адипоцитов белком Асrp30 (SEQ ID NO:3) и секретируемым адипоцитами белком арМ1 (HUMUPST2_1 на фиг.1 и 2). Несколько более отдаленная гомология была также идентифицирована с А-цепью компонента C1q, двумя факторами, наблюдаемыми в активном состоянии переносящих зиму в зимней спячке Сибирских сурков (HP25_TAMAS и HP27_TAMAS), и белком мозга крысы (CERL_RAT), как показано на фиг.1 и 2.
Нуклеотидная последовательность zsig37 описана в SEQ ID NO:1, а ее расшифрованная аминокислотная последовательность описана в SEQ ID NO:2. Вырожденная нуклеотидная последовательность, кодирующая полипептид SEQ ID NO:2, обеспечена в SEQ ID NO:23. Как описано в общем виде выше, полипептид zsig37 включает в себя сигнальную последовательность, простирающуюся от аминокислоты 1 (Met) до аминокислотного остатка 21 (Gly). Альтернативная сигнальная последовательность простирается от аминокислоты 1 (Met) до аминокислоты 25 (Ser). Таким образом, зрелый полипептид простирается от аминокислоты 22 (Leu) или 26 (Arg) до аминокислоты 281 (Pro). В зрелом полипептиде обнаружен N-концевой район неизвестной гомологии, расположенный между аминокислотными остатками 22 (Leu) и 98 (Lys). Кроме того, был обнаружен укороченный коллагеновый домен между аминокислотами 99 (Gly) и 140 (Аrg). В укороченном коллагеновом домене наблюдали 1 точный повтор Gly-Xaa-Рrо и 13 неточных повторов Gly-Xaa-Xaa. В противоположность этому, Асrр30 содержит 22 точных или неточных повторов. Полипептид zsig37 включает в себя также карбоксиконцевой глобулярный домен, простирающийся от приблизительно аминокислоты 141 (Cys) до аминокислоты 281 (Pro). Полипептид zsig37, HUMUPST2_1 и Асrр30, по-видимому, являются гомологичными в коллагеновом домене и в глобулярном домене, но не в N-концевой части зрелого полипептида.
Было определено, что глобулярный домен C1q ACRP30 имеет топологию “рулета” из 10 бета-тяжей (Shapiro and Scherer, Curr. Biol. 8:335-8, 1998), которая обнаруживает значительную структурную гомологию с TNF-семейством, а последовательность zsig37, представленная SEQ ID NО:2, содержит все 10 бета-тяжей этой структуры (аминокислотные остатки 147-151, 170-172, 178-181, 185-188, 191-203, 207-214, 219-225, 227-238, 244-250 и 269-274 SEQ ID NO:2). Эти тяжи были названы  соответственно.
соответственно.
Zsig37 имеет две рецепторсвязывающие петли, при аминокислотных остатках 152-180 и 213-226. Аминокислотные остатки 191 (Gly), 193 (Туr), 238 (Leu) и 272 (Gly), по-видимому, являются консервативными во всем этом суперсемействе, в том числе в CD40, TNFα , TNFβ , ACRP30 и zsig37.
Другой аспект данного изобретения включает в себя фрагменты полипептидов zsig37 как ингибиторы гемостаза и иммунных функций. Предпочтительные фрагменты включают в себя коллагеноподобный домен полипептидов zsig37, простирающийся от аминокислоты 99 (Gly) до аминокислоты 140 (Arg) SEQ ID NО:2, часть полипептида zsig37, содержащую коллагеноподобный домен или часть коллагеноподобного домена, способную к димеризации или олигомеризации. Другие предпочтительные фрагменты включают в себя глобулярный домен полипептидов zsig37, простирающийся от аминокислоты 140 (Arg) или 141 (Cys) до 281 (Pro) SEQ ID NО:2, часть полипептида zsig37, содержащую подобный глобулярному домен или активную часть подобного глобулярному домена. Другой фрагмент полипептида zsig37 данного изобретения включает в себя как коллагеноподобный домен, так и подобный глобулярному домен, простирающийся от аминокислотного остатка 99 (Gly) до 281 (Pro) SEQ ID NО:2. Эти фрагменты являются особенно применимыми в ингибировании опосредованной коллагеном активации тромбоцитов и ингибировании комплемента и C1q.
Данное изобретение обеспечивает также применение слитых (гибридных или химерных) белков zsig37. Например, слитые белки данного изобретения включают в себя (1) полипептид, выбранный из группы, включающей в себя: (а) полипептидные молекулы, содержащие последовательность аминокислотных остатков, показанную в SEQ ID NО:2, от аминокислотного остатка 1 (Met), 22 (Leu) или 26 (Arg) до аминокислотных остатков 281 (Pro); (b) полипептидные молекулы, простирающиеся от аминокислоты 99 (Gly) до аминокислоты 140 (Arg) SEQ ID NО:2, причем часть этого полипептида zsig37 содержит коллагеноподобный домен или часть коллагеноподобного домена, способную к димеризации или олигомеризации; (с) полипептидные молекулы, простирающиеся от аминокислоты 140 (Arg) или 141 (Cys) до 281 (Pro) SEQ ID NО:2, причем часть этого полипептида zsig37 содержит подобный глобулярному домен или активную часть подобного глобулярному домена; или (d) полипептидные молекулы, простирающиеся от аминокислоты 99 (Gly) до 281 (Pro), причем часть этого полипептида zsig37 включает в себя коллагеноподобный домен и глобулярный домен; и (2) другой полипептид. Этот другой полипептид может быть альтернативным или дополнительным глобулярным доменом, альтернативным или дополнительным коллагеноподобным доменом, сигнальным пептидом для облегчения секреции гибридного белка или т.п.
В способах данного изобретения применимы также агонисты и антагонисты zsig37. Способы идентификации антагонистов хорошо известны в данной области. Например, антагонисты полипептида zsig37 могут быть идентифицированы обеспечением клеток, чувствительных к полипептиду zsig37, культивированием первой порции этих клеток в присутствии полипептида zsig37, культивированием второй порции этих клеток в присутствии полипептида zsig37 и тест-соединения и детектированием уменьшения в клеточной ответной реакции второй порции клеток в сравнении с первой порцией этих клеток. Кроме этих описанных здесь тестов, пробы могут быть тестированы на ингибирование активности zsig37 в многочисленных анализах, предназначенных для измерения связывания рецептора или стимуляции/ингибирования zsig37-зависимых клеточных ответных реакций. Например, чувствительные к zsig37 клеточные линии могут быть трансфицированы конструкцией с репортерным геном, которая отвечает на стимулируемый zsig37 клеточный путь. Конструкции с репортерным геном этого типа известны в данной области и обычно будут содержать чувствительный к zsig37 ДНК-элемент, функционально связанный с геном, кодирующим детектируемый тестом белок, такой как люцифераза. Чувствительные ДНК-элементы могут включать в себя, но не ограничиваются ими, циклический АМФ-чувствительные элементы (CRE), гормон-чувствительные элементы (HRE), инсулин-чувствительный элемент (IRE) (Nasrin et al., Proc. Natl. Acad. Sci. USA 87:5273-7, 1990) и сыворотка-чувствительные элементы (SRE) (Shaw et al., Cell, 56:563-72, 1989). Циклические АМФ-чувствительные элементы рассматриваются в обзоре Roestler et al., J. Biol. Chem. 263 (19):9063-6; 1988 и Habener, Molec. Endocrinol. 4 (8): 1087-94 (1990). Гормон-чувствительные элементы рассматриваются в Beato, Cell 56:335-44; 1989. Соединения-кандидаты, растворы, смеси или экстракты или кондиционированные среды от различных типов клеток тестируют на их способность ингибировать активность zsig37 на клетках-мишенях, определяемую уменьшением стимуляции zsig37 экспрессии репортерного гена. Тесты этого типа будут детектировать соединения, которые непосредственно блокируют связывание zsig37 с рецепторами поверхности клеток, а также соединения, которые блокируют процессы в клеточном пути, следующие за связыванием рецептор-лиганд. Альтернативно, соединения или другие пробы могут быть тестированы на прямое блокирование связывания zsig37 с рецептором с использованием zsig37, меченого детектируемой меткой (например, 125I, биотином, пероксидазой хрена, FITC и т.п.). В анализах этого типа способность тест-пробы ингибировать связывание меченого zsig37 с рецептором указывает на ингибирующую активность, которая может быть подтверждена при помощи вторичных анализов. Рецепторы, используемые в анализах связывания, могут быть клеточными рецепторами или выделенными иммобилизованными рецепторами.
В способах данного изобретения применимы также антитела, которые специфически связываются с эпитопами, пептидами или полипептидами полипептида zsig37. Способы получения и выделения поликлональных и моноклональных антител хорошо известны в данной области (см., например, Sambrook et al., Molecular Cloning: A Laboratory Manual, Second Edition, Cold Spring Harbor, NY, 1989 и Hurrell, J.G.R., Ed., Monoclonal Hybridoma Antibodies: Techniques and Applications, CRC Press, Inc., Boca Raton, FL, 1982).
Как должно быть очевидно специалисту с обычной квалификацией в данной области, поликлональные антитела могут быть получены инокуляцией различных теплокровных животных, таких как лошади, коровы, козы, овцы, собаки, куры, кролики, мыши, хомячки, морские свинки и крысы, а также трансгенные животные, такие как трансгенные овцы, коровы, козы или свиньи. Антитела могут также экспрессироваться в дрожжах и грибках в модифицированных формах, а также в клетках млекопитающих и насекомых. Полипептид zsig37 или его фрагмент служит в качестве антигена (иммуногена) для инокуляции животного или индукции иммунной реакции. Подходящие антигены могли бы включать в себя полипептид zsig37, кодируемый SEQ ID NО:2, от аминокислотного остатка 22 до аминокислотного остатка 281 SEQ ID NО:2, от аминокислотного остатка 26 до аминокислотного остатка 281 SEQ ID NО:2 или его фрагмент непрерывных аминокислотных остатков 9-281. Иммуногенность полипептида zsig37 может быть увеличена путем применения адъюванта, такого как квасцы (гидроксид алюминия) или полный или неполный адъювант Фрейнда. Полипептиды, применимые для иммунизации, включают в себя также слитые (гибридные) полипептиды, такие как гибриды zsig37 или его части с полипептидом иммуноглобулина или с аффинной меткой. Полипептидный иммуноген может быть полноразмерной молекулой или ее частью. Если эта часть полипептида является "гаптен-подобной", такая часть может быть предпочтительно соединена или связана с макромолекулярным носителем (таким как гемоцианин фиссуреллы (KLH), бычий сывороточный альбумин (БСА) или столбнячный токсоид) для иммунизации.
В применении здесь, термин "антитела" включает в себя поликлональные антитела, аффинно-очищенные поликлональные антитела, моноклональные антитела и антигенсвязывающие фрагменты, такие как протеолитические F(ab')2- и Fab-фрагменты. Также включены генетически сконструированные интактные антитела или фрагменты, такие как химерные антитела, Fv-фрагменты, одноцепочечные антитела и т.п., а также синтетические антигенсвязывающие пептиды и полипептиды. Антитела не из человека могут быть "очеловечены" (гуманизированы) прививкой CDR не человека на каркасные и константные области иммуноглобулина человека или включением полных вариабельных доменов не человека (необязательно "окутыванием" их подобной человеческой поверхностью путем замены экспонированных остатков, результатом чего является "облицованное" антитело). В некоторых случаях "гуманизированные" антитела могут сохранять остатки иммуноглобулина не человека в каркасных доменах вариабельной области человека для усиления характеристик правильного связывания. Посредством "гуманизирования" антител может быть увеличен биологический полупериод существования в организме, и потенциал неблагоприятных иммунных реакций при введении людям уменьшается. Альтернативные способы генерирования или отбора антител, применимые здесь, включают в себя экспонирование in vitro лимфоцитов белку или пептиду zsig37 и отбор библиотек представления антител в фаговых или подобных векторах (например, посредством использования иммобилизованного или меченого белка или пептида zsig37).
Антитела определяются как специфически связывающие, если: 1) они проявляют пороговый уровень связывающей активности и/или 2) они не имеют значимой перекрестной реактивности с родственными полипептидными молекулами. Во-первых, описанные здесь антитела специфически связываются, если они связываются с полипептидом, пептидом или эпитопом zsig37 с аффинностью связывания (Ка) 106 моль-1 или большей, предпочтительно 107 моль-1 или большей, более предпочтительно 108 моль-1 или большей, и наиболее предпочтительно 109 моль-1 или большей. Аффинность связывания антитела может быть легко определена специалистом с обычной квалификацией в этой области, например, при помощи анализа Скетчарда (Scatchard, Ann. NY Acad. Sci. 51: 660-672, 1949).
Во-вторых, антитела специфически связываются, если они не имеют значимой перекрестной реактивности с родственными полипептидами. Антитела не имеют значимой перекрестной реактивности с родственными полипептидными молекулами, например, если они обнаруживают полипептид zsig37, но не обнаруживают известные родственные полипептиды, с использованием стандартного Вестерн-блот-анализа (Ausubel et al., ibid.). Примеры известных родственных полипептидов включают в себя другие члены семейства белков, такие как Асrр30 (SEQ ID NO:1), полипептиды, показанные в сопоставлении фиг.1, и т.п. Они могут также включать, если желательно, ортологи и мутант полипептидов zsig37 человека. Кроме того, антитела могут быть "скринированы против" известных родственных полипептидов для выделения популяции, которая специфически связывается с полипептидами данного изобретения. Например, антитела, индуцированные к полипептидам zsig37 человека, адсорбируют с родственными полипептидами, прикрепленными к нерастворимому матриксу; антитела, специфические для полипептида zsig37 человека, будут протекать через этот матрикс при подходящих буферных условиях. Такой скрининг ("просеивание") позволяет выделить поликлональные и моноклональные антитела, не имеющие перекрестной реактивности с близкородственными полипептидами (Antibodies: A Laboratory Manual, Harlow and Lane (eds.), Cold Spring Harbor Laboratory Press, 1988; Current Protocols in Immunology, Cooligan, et al. (eds.), National Institutes of Health, John Wiley and Sons, Inc., 1995). Скрининг и выделение специфических антител хорошо известны в этой области (см. Fundamental Immunology, Paul (eds.), Raven Press, 1993; Getzoff et al., Adv. in Immunol. 43: 1-98, 1988; Monoclonal Antibodies: Principles and Practice, Coding, J.W. (eds.), Academic Press Ltd., 1996; Benjamin et al., Ann. Rev. Immunol. 2: 67-101, 1984). Характерные примеры таких анализов включают в себя: конкурентный иммуноэлектрофорез, радиоиммуноанализ, радиоиммунопреципитацию, твердофазный иммуноферментный анализ (ELISA), дот-блот или Вестерн-блот анализ, ингибиторный или конкурентный анализ и сэндвич-анализ.
Действие полипептидов, фрагментов, гибридных белков, агонистов или антагонистов zsig37 на гемостаз, в частности, на адгезию и активацию тромбоцитов, приводящие к агрегации тромбоцитов, может быть определено с использованием способов и анализов, обеспеченных здесь, и способов и анализов, известных в данной области. Коллаген является сильным индуктором агрегации тромбоцитов. Это поднимает вопрос о риске для пациентов, восстанавливающихся после сосудистых повреждений. Ингибиторы индуцированной коллагеном агрегации тромбоцитов могли бы быть использованы для таких целей. Было обнаружено, что zsig37 связывается с фибронектином и коллагенами типа I, II, III, V и VI. В частности, zsig37 связывается со специфическими доменами на коллагене VI зависимым от концентрации образом. Было обнаружено, что zsig37 ингибирует опосредованную коллагеном активацию тромбоцитов. Индуцированное zsig37 ингибирование было селективным в отношении активации коллагена, zsig37 не влиял на тромбоциты, активируемые известными активаторами тромбоцитов АДФ или тромбином. Эти результаты описаны более подробно ниже в разделе примеров. Авторы считают, что полипептиды, фрагменты, слитые белки, агонисты или антагонисты zsig37 будут применимы для блокирования связывания тромбоцитов с покрытыми коллагеном поверхностями и уменьшения ассоциированной индуцированной коллагеном агрегации тромбоцитов.
C1q является компонентом пути комплемента, и было обнаружено, что C1q стимулирует защитные механизмы, а также запускает генерирование токсичных разновидностей кислорода, которые могут вызывать повреждение ткани (Tenner, Behring Inst. Mitt. 93:241-53, 1993). Сайты связывания C1q найдены на тромбоцитах. Было обнаружено, что C1q, независимо от партнера иммунного связывания, ингибирует агрегацию тромбоцитов, но не адгезию или изменение формы тромбоцитов. Аминоконцевой район C1q имеет гомологию с коллагеном (Peerschke and Ghebrehiwet, J. Immunol. 145:2984-88, 1990). Zsig37 связывается с C1q комплемента зависимым от концентрации образом. Было обнаружено, что zsig37 является эффективным в ингибировании пути комплемента, включающего C1q, как с сенсибилизированными, так и с несенсибилизированными эритроцитами овцы.
Полипептиды, фрагменты, слитые белки, антитела, агонисты или антагонисты zsig37 данного изобретения могут быть использованы в способах стимуляции кровотока в сосудистой сети млекопитающего путем уменьшения числа тромбоцитов, которые прикрепляются и является активированными, и размера агрегатов тромбоцитов. Такие способы могут включать в себя введение терапевтически эффективного количества полипептидов, фрагментов, слитых белков, антител, агонистов или антагонистов zsig37 млекопитающему, нуждающемуся в таком лечении, посредством чего zsig37 уменьшает тромбогенную активность и активность комплемента в сосудистой сети этого млекопитающего. Как описано ниже, полипептиды zsig37 ингибируют опосредованную коллагеном активацию тромбоцитов и инактивируют фибронектин и коллагены типа I, II, V и VI посредством связывания. Введение zsig37 уменьшает тромбогенную активность в месте сосудистого повреждения уменьшением способов адгезии, активации и агрегации тромбоцитов. Zsig37 ингибирует также путь комплемента и C1q, как описано ниже, уменьшая тем самым активность комплемента в сосудистой сети. Полипептиды, фрагменты, слитые белки, антитела, агонисты или антагонисты zsig37, используемые в таких способах, могут быть введены до, во время или после острого сосудистого повреждения в млекопитающем.
В предпочтительном способе сосудистое повреждение обусловлено хирургической реконструкцией (пластикой) сосудов, в том числе, но не ограничивается ими пластической операцией на сосудах (реконструкцией сосудов), эндартерэктомией, трансплантатом шунта венечной артерии, микрососудистой репарацией или анастомозом сосудистого трансплантата. Также рассматриваются повреждения, вызванные травмой, ударом или аневризмой. В других предпочтительных способах сосудистое повреждение обусловлено разрывом бляшек, деградацией сосудистой сети, осложнениями, связанными с диабетом и атеросклерозом. Разрыв бляшек в венечной артерии индуцирует сердечный приступ, а в мозговой артерии индуцирует удар. Применение полипептидов, фрагментов, гибридных белков, антител, агонистов или антагонистов zsig37 в таких способах было бы также полезным для ослабления всей системы заболеваний сосудистой сети, связанных с иммунной системой, таких как диссеминированная внутрисосудистая коагуляция (DIC) и синдром внезапной младенческой смерти (SIDS). Кроме того, ингибирующая комплемент активность была бы полезной для лечения несосудистых иммунных заболеваний, таких как артериолосклероз.
Была обнаружена корреляция между присутствием C1q в локализованном ишемическом миокарде и накоплением лейкоцитов после венечной окклюзии и реперфузии. Высвобождение клеточных компонентов после повреждения ткани запускает активацию комплемента, которая приводит к токсическим кислородным продуктам, которые могут быть первичной причиной повреждения миокарда (Rossen et al., Circ. Res. 62:572-84, 1998 и Tenner, ibid.). Было обнаружено, что блокирование пути комплемента защищает ишемический миокард от реперфузионного повреждения (Buerke et al., J. Pharm. Exp. Therap. 286:429-38, 1998). Ингибирование комплемента и активности связывания C1q полипептидов zsig37 было бы применимо для таких целей.
Способности связывания коллагена и C1q полипептидом zsig37 было бы полезным для восстановления поврежденных коллагеновых тканей, предотвращая адгезию, активацию или агрегацию тромбоцитов и активацию воспалительных процессов, которые приводят к высвобождению токсических кислородных продуктов. Делая экспонированную (обнаженную) ткань инертной в отношении таких процессов, как активность комплемента, тромботическая активность и иммунная активация, полипептиды, фрагменты, гибридные белки, антитела, агонисты или антагонисты zsig37 были бы полезными в уменьшении повреждающих действий ишемии и реперфузии. В частности, такие повреждения могли бы включать в себя ишемию, вызываемую травматическим повреждением, интестинальную странгуляцию и повреждение, ассоциированное с событиями перед установлением и после установления кровотока. Zsig37 был бы также полезен в лечении ишемии и перерывов в кровотоке, вызываемых искусственным кровообращением, инфаркта миокарда или посттравматического вазоспазама (сужения кровеносных сосудов), например, в случае удара или чрескожной чреспросветной ангиопластики, а также случайной сосудистой травмы или хирургически индуцированной сосудистой травмы.
Полипептиды, фрагменты, слитые белки, антитела, агонисты или антагонисты zsig37 были бы также полезны для приспособления поверхности протезных биоматериалов и хирургического оборудования, делая поверхность таких биоматериалов инертной относительно активации комплемента, тромботической активности или иммунной активности. Такие биоматериалы включают в себя, но не ограничиваются ими, покрытые коллагеном или фрагментами коллагена биоматериалы, покрытые желатином биоматериалы, покрытые фибрином биоматериалы, покрытые фибронектином биоматериалы, покрытые гепарином биоматериалы, покрытые коллагеном и гелем стенты, артериальные трансплантаты, синтетические сердечные клапаны, искусственные органы или любая протезная аппликация, экспонированная кровью, которая будет связывать zsig37 при более чем 1× 108. Покрытие таких материалов может выполняться с использованием способов, известных в данной области (см., например, Rubens, патент США № 5272074).
Комплемент и C1q играют роль в воспалении. Активация комплемента инициируется связыванием C1q с иммуноглобулинами (Johnston, Pediatr. Infect. Dis. J. 12: 933-41, 1993; Ward and Ghetie, Therap. Immunol. 2:77-94. 1995). Ингибиторы C1q и комплемента были бы полезны в качестве противовоспалительных агентов. Такое применение могло бы применяться для предотвращения инфекции. Кроме того, такие ингибиторы могут вводиться индивидууму, страдающему от воспаления, опосредованного активацией комплемента и связыванием иммунных комплексов с C1q. Полипептиды, фрагменты, слитые белки, антитела, агонисты или антагонисты zsig37 были бы также полезны в способах опосредования заживления ран посредством преодоления нарушенного заживления ран. Прогрессирование в заживлении ран будет включать в себя, например, такие элементы, как уменьшение воспаления, рекрутинг фибробластов, затягивание ран и уменьшение инфекции.
Способность опухолевых клеток связываться с коллагеном может способствовать метастазированию опухолей. Ингибиторы связывания коллагена применимы также для медиирования адгезионных взаимодействий и распространения метастазов опухолей (Noeske-Jungbult et al., US Patent №5723312).
Было обнаружено, что zsig37 индуцирует вазодилатацию (расширение сосудов) в сокращенных норэпинеферином кольцах аорты при применении процедур Dainty et al., J. Pharmacol. 100:767, 1990 и Rhee et al., Neurotox. 16:179, 1995, как описано ниже более подробно.
Адгезия, активация и агрегация тромбоцитов может оцениваться с использованием описанных здесь способов или способов, известных в данной области, таких как анализ агрегации тромбоцитов (Chiang et al., Thrombosis Res. 37:605-12, 1985) и анализ адгезии тромбоцитов (Peerschke and Ghebrehiwet, J. Immunol. 144:221-25, 1990). Ингибирование C1q и пути комплемента может быть определено с использованием описанных здесь способов или способов, известных в данной области, таких как описанные в Suba and Csako, J. Immunol. 117:304-9, 1976. Анализы адгезии тромбоцитов с коллагеном и ингибирование индуцированной коллагеном агрегации тромбоцитов могут проводиться с использованием способов, описанных Keller et al., J. Biol. Chem. 268:5450-6, 1993; Waxman and Connolly, J. Biol. Chem. 268:5445-9, 1993; Noeske-Jungblut et al., J. Biol. Chem. 269:5050-3 или Deckmyn et al., Blood 85:712-9, 1995.
Различные модели in vitro и in vivo доступны для оценки эффектов полипептидов, фрагментов, гибридных белков, антител, агонистов или антагонистов zsig37 на ишемию и реперфузионное повреждение. См., например, Shandelya et al., Circulation 88:2812-26, 1993; Weisman et al., Science 249:146-151, 1991; Buerke et al., Circulation 91:393-402, 1995; Horstick et al., Circulation 95:701-8, 1997 и Burke et al., J. Phar. Exp. Therp. 286:429-38, 1998. Анализ агрегации тромбоцитов хомячка ех vivo описан Deckmyn et al., ibid. Время кровотечения в хомячках и павианах может быть измерено после инъекции полипептидов zsig37 с использованием модели, описанной Deckmyn et al., ibid. Образование тромба в ответ на введение белков данного изобретения может быть измерено с использованием модели тромбоза бедренной вены хомячка, обеспеченной Deckmyn et al., ibid. Изменения в адгезии тромбоцитов при проточных условиях после введения zsig37 могут быть измерены с использованием способа, описанного в Harsfalvi et al., Blood 85:705-11, 1995.
Ингибирование комплемента и заживление ран полипептидами, фрагментами, слитыми белками, антителами, агонистами или антагонистами zsig37 могут оцениваться отдельно или в комбинации с другими известными ингибиторами индуцированной коллагеном активации и агрегации тромбоцитов, такими как, например, паллдипин, моубатин или калин.
Полипептиды, фрагменты, слитые белки, антитела, агонисты или антагонисты zsig37 могут оцениваться с использованием описанных здесь способов или способов, известных в данной области, таких как, например, заживление дермальных слоев у свиней (Lynch et al., Proc. Natl. Acad. Sci. USA 84:7696-700, 1987) и ран с полной толщиной кожи в мышах с генетическим диабетом (Greenhalgh et al., Am. J. Pathol. 136:1235-46, 1990). Полипептиды данного изобретения могут анализироваться отдельно или в комбинации с другими известными ингибиторами комплемента, как описано выше.
Кроме того, полипептиды, фрагменты, слитые белки, агонисты или антагонисты zsig37 могут быть терапевтически применимыми для антимикробных приложений. Например, компонент комплемента C1q играет роль в иммунной защите организма против инфекционных агентов, таких как бактерии и вирусы. Известно, что C1q проявляет несколько специализированных функций. Например, C1q запускает каскад комплемента через взаимодействие со связанным антителом или С-реактивным белком (CRP). C1q взаимодействует также непосредственно с некоторыми бактериями, РНК-вирусами, микоплазмой, кристаллами мочевой кислоты, липид А-компонентом бактериального эндотоксина и мембранами определенных внутриклеточных органелл. Считается, что связывание C1q с C1q-рецептором стимулирует фагоцитоз. C1q, по-видимому, усиливает также аспект образования антител иммунной защиты хозяина. См., например, Johnston, Pediatr. Infect. Dis. J. 12(11): 933-41, 1993. Таким образом, растворимые C1q-подобные молекулы могут быть полезны в качестве антимикробных агентов, стимулирующих лизис или фагоцитоз инфекционных агентов.
Было определено, что положительно заряженные, внеклеточные, представляющие собой тройную спираль коллагеновые домены C1q и рецептор акцептора макрофагов играют роль в связывании лигандов, и было показано, что они имеют широкую специфичность связывания для полианионов (Acton et al., J. Biol. Chem. 268:3530-37. 1993). Лизофосфолипидный фактор роста (лизофосфатидиловая кислота, LPA) и другие митогенные анионы локализуются в месте поврежденных тканей и способствуют репарации раны. LPA проявляет многие биологические эффекты, в том числе активацию тромбоцитов и положительную регуляцию сборки матрикса. Считается, что LPA является синергистом относительно других факторов коагуляции крови и опосредует заживление ран.
Известно, что коллагеновые домены белков, таких как C1q и рецептор акцептора макрофагов, связывают кислые фосфолипиды, такие как LPA. 9-мерный район коллагенового домена zsig37, аминокислотные остатки 127-135 SEQ ID NO:2, имеет гомологию последовательности с коллагеновым доменом, обнаруженным на C1q и рецепторе акцептора макрофагов. Взаимодействие полипептидов, фрагментов, слитых белков, агонистов или антагонистов с митогенными анионами, такими как LPA, может быть определено при помощи анализов, известных в данной области, см., например, Acton et al., ibid. Ингибирование воспалительных процессов полипептидами и антителами данного изобретения было бы также полезным в предупреждении инфекции в участке раны.
Для фармацевтического применения белки данного изобретения могут быть приготовлены с фармацевтически приемлемыми носителями для парентерального, перорального, назального, ректального, местного, чрескожного введения или т.п. в соответствии с общепринятыми способами. Предпочтительно введение производят в месте сосудистого повреждения или вблизи от него. Обычно фармацевтические композиции включают в себя белок zsig37 в комбинации с фармацевтически приемлемым носителем, таким как солевой раствор, забуференный солевой раствор, 5% декстроза в воде или т.п. Композиции могут дополнительно включать в себя один или несколько наполнителей, консервантов, солюбилизаторов, буферящих агентов, альбумин для предотвращения потери белка на поверхностях флаконов и т.д. Способы приготовления хорошо известны в данной области и описаны, например, в Remington: The Science and Practice of Pharmacy, Gennaro, ed., Mack Publishing Co., Easton PA, 19th ed., 1995. Терапевтические дозы обычно определяются врачом с учетом характера и тяжести подлежащего лечению состояния, характеристик пациента и т.д. Определение дозы находится на уровне обычной квалификации в данной области.
В применении здесь, “фармацевтически эффективное количество” полипептида, фрагмента, слитого белка, агониста или антагониста zsig37 является количеством, достаточным для индукции желательного биологического результата. Этот результат может быть ослаблением признаков, симптомов или причин заболевания или любым другим изменением биологической системы. Например, эффективное количество полипептида zsig37 является количеством, которое обеспечивает либо субъективное ослабление симптомов, либо объективно идентифицируемое улучшение, отмечаемое врачом или другим квалифицированным наблюдателем. Например, такое эффективное количество полипептида zsig37 обеспечивало бы ингибирование активированной коллагеном активации тромбоцитов и пути комплемента, включающего в себя C1q, увеличение локализованного кровотока в сосудистой сети пациента и/или уменьшение повреждающих эффектов ишемии и реперфузии. Эффективные количества полипептида zsig37 будут в большой степени зависеть от заболевания или симптома, подлежащего лечению.
Количество вводимого полипептида и его концентрация в композициях зависит от выбранного носителя, пути введения, силы конкретного полипептида, клинического состояния пациента, побочных действий и стабильности соединения в композиции. Таким образом, врач должен применять подходящий препарат, содержащий подходящую концентрацию в композиции, а также количество вводимой композиции в зависимости от клинического опыта в отношении конкретного пациента или в отношении сходных пациентов. Такие количества будут отчасти зависеть от конкретного состояния, подлежащего лечению, возраста, веса и общего состояния здоровья пациента и других факторов, очевидных специалистам в данной области. Обычно доза будет находиться в диапазоне 0,1-100 мг/кг веса субъекта. В таких применениях, как баллонные катетеры, типичным диапазоном доз могли бы быть 0,05-5 мг/кг субъекта. Дозы для конкретных соединений могут быть определены из исследований in vitro или ex vivo в сочетании с исследованиями на экспериментальных животных. Концентрации соединений, которые, как было обнаружено, являются эффективными in vitro и in vivo, обеспечивают рекомендацию для исследований на животных, в которых дозы рассчитывают для обеспечения сходных концентраций в месте действия.
Далее данное изобретение иллюстрируется следующими ниже неограничительными примерами.
Пример 1
Удлинение EST-последовательности
Новые полинуклеотиды данного изобретения, кодирующие полипептиды zsig37, первоначально идентифицировали отбором EST из базы данных EST, предсказывая последовательность белка на ее основе, и поиском в базах данных известных последовательностей на секретируемый белок, который наиболее гомологичен предсказанному на основе этой EST белку. EST, которые потенциально кодируют белки, имеющие представляющую интерес гомологию с известными секретируемыми белками, идентифицировали для дальнейшего исследования. Была обнаружена единственная EST-последовательность, которая, как было предсказано, является гомологичной специфическому белку адипоцитов (жировых клеток). См., например, Scherer et al., J. Biol. Chem. 270(45): 26746-9, 1995. Для идентификации соответствующей кДНК для секвенирования использовали клон, который, как считали, содержал, возможно, всю кодирующую последовательность. С использованием набора Invitrogen S.N.A.P.TM Miniprep (Invitrogen, Corp. San Diego, CA) в соответствии с инструкциями изготовителей получали 5 мл ночной культуры в LB+50 мкг/мл ампициллина. Матрицу секвенировали на ДНК-секвенаторе модели 377 ABIPRISMTM (Perkin-Elmer Cetus, Norwalk, Ct.) с использованием набора для реакции секвенирования Dye Terminator Cycle Sequencing Ready Reaction Kit ABIPRISMTM (Perkin-Elmer Corp.) в соответствии с инструкциями изготовителя. Олигонуклеотиды ZC695 (SEQ ID NO:5), ZC694 (SEQ ID NO:6) для промоторов SP6 и Т7 на клонсодержащем векторе использовали в качестве секвенирующих праймеров. Олигонуклеотиды ZC13210 (SEQ ID NO:7), ZC13588 (SEQ ID NO:8), ZC13532 (SEQ ID NO:9), ZC13641 (SEQ ID NO:10), ZC13586 (SEQ ID NO:11), ZC13651 (SEQ ID NO:12), ZC13622 (SEQ ID NO:13), ZC13625 (SEQ ID NO:14), ZC13650 (SEQ ID NO:15), ZC13589 (SEQ ID NO:16), ZC13624 (SEQ ID NO:17), ZC13531 (SEQ ID NO:18), ZC13587 (SEQ ID NO:19) и ZC13623 (SEQ ID NO:20) использовали для комплектования последовательности из этого клона. Реакции секвенирования проводили в термоциклере Hybaid OmniGene Temperature Cycling System (National Labnet Co., Woodbridge, NY). Программное обеспечение для анализа последовательностей SEQUENCHERTM 3.0 (Gene Codes Corporation, Ann Arbor, Ml) использовали для анализа данных. Полученная последовательность 2769 п.н. описана в SEQ ID NO:1. Сравнение исходно полученной EST-последовательности с последовательностью, представленной в SEQ ID NO:1, показало, что была двусмысленность одной пары оснований (неизвестный остаток "N") и не было инсерций пар оснований, что привело к идентификации лейцина в разрешении этой двусмысленности и нулевым смещениям рамки считывания между расшифрованными последовательностями аминокислот.
Пример 2
Тканевое распределение
Нозерн-блот-анализы проводили с использованием блотов множественных тканей человека из Clontech (Palo Alto, CA). Зонд ДНК из 30 нуклеотидов (ZC12447; SEQ ID NO:4) к 5'-концу последовательности нуклеотидов зрелого белка, показанного в SEQ ID NO:1, радиоактивно метили 32P с использованием полинуклеотидкиназы Т4 и буфера для прямой реакции (GIBCO BRL, Gaithersburg, MD) в соответствии с описаниями изготовителя. Зонд очищали с использованием колонки NUCTRAP push column (Stratagene Cloning Systems, La Jolla, CA). Раствор EXPRESSHYB (Clontech, Palo Alto, CA) использовали для предгибридизации и в качестве раствора для гибридизации для Нозерн-блотов. Гибридизация имела место в течение ночи при 50° С и затем блоты промывали в 2Х SSC и 0,1% ДСН при комнатной температуре с последующим промыванием в 1Х SSC и 0,1% ДСН при 68° С (приблизительно на 5° С ниже, чем точка плавления). Размер одного транскрипта наблюдали при приблизительно 2,8 т.п.н. Интенсивность сигнала была наивысшая для сердца и плаценты, с относительно менее интенсивными сигналами в почке, яичнике, надпочечнике и скелетной мышце и более низкими сигналами в большом разнообразии других тканей, присутствующих на Нозерн-блоте.
Дополнительный Нозерн-блот-анализ проводили с Нозерн-блотом ткани кишечника. Блот готовили с использованием мРНК из линии клеток колоректальной аденокарциномы человека SW480 (Clontech, Palo Alto, CA), ткани тонкой кишки человека (Clontech), ткани желудка человека (Clontech), ткани линии клеток гладких мышц кишечника (Hism; ATCC No. CRL-1692: American Type Culture Collection, 12301 Parklawn Drive, Rockville, MD), линии здоровых клеток ободочной кишки человека (FHC; ATCC No. CRL-1831; American Type Culture Collection) и линии здоровых клеток тонкой кишки эмбриона (FHs74 Int.; ATCC No. CCL241; American Type Culture Collection).
Тотальные РНК выделяли из Hism, FHC и FHs74 Int. гуанидий-кислотным способом (Cheomczynski et al., Anal. Biochem, 162: 156-9, 1987). ПoлиA+ РНК отбирали элюцией тотальной РНК через колонку, которая удерживает пoлиA+ РНК (Aviv et al., Proc. Nat. Acad. Sci. 69: 1408-12, 1972). 2 мкг пoлиA+ РНК из каждой пробы отделяли в 1,5% агарозном геле в 2,2 М формальдегиде и фосфатном буфере. Эти РНК переносили на мембрану Nytran (Schleicher and Schuell, Keene, NH) в 20Х SSC в течение ночи. Блот обрабатывали в UV Stratalinker 2400 (Stratagene, La Jolla, CA) при 0,12 Дж. Затем блот отжигали при 80° С в течение одного часа.
Полноразмерную кДНК (показанную в SEQ ID NO:1) амплифицировали при помощи ПЦР и радиоактивно метили 32P-dCTP с использованием набора Rediprime pellet kit (Amersham, Arlington Heights, IL) в соответствии с описаниями изготовителя. Этот блот гибридизовали в EXPRESSHYB (Clontech) при 56° С в течение ночи. Блот промывали при комнатной температуре в 2Х SSC и 0,1% ДСН, затем в 2Х SSC и 0,1% ДСН при 65° С и наконец при 65° С в 0,1Х SSC и 0,1% ДСН. Результаты показали, что zsig37 гибридизовался со всеми тканями, кроме линии клеток гладких мышц кишечника HISM.
Пример 3
Хромосомное картирование гена zsig37
Ген zsig37 картировали в хромосоме 17 человека, районе 17q25.2, при помощи ПЦР с использованием панели номер 2 картирования гибридов соматических клеток человек/грызун NIGMS (National Institute of General Medical Sciences, Coriell Institute of Medical Research). Эта панель состоит из ДНК, выделенной из 24 гибридов соматических клеток человек/грызун, каждый из которых сохранял одну специфическую хромосому человека и родительские ДНК. Для картирования гена zsig37 реакции по 20 мкл устанавливали в пригодном для ПЦР 96-луночном микротитрационном планшете (Stratagene, La Jolla, CA) и использовали термоциклер "RoboCycler Gradient 96" (Stratagene). Каждая из этих 27 ПЦР-реакций состояла из 2 мкл 10Х KlenTaq буфера для ПЦР-реакции (Clontech Laboratories, Inc., Palo Alto, CA), 1,6 мкл смеси dNTP (2,5 мМ каждый, PERKIN-ELMER, Foster City, CA), 1 мкл смыслового праймера (SEQ ID NO:21), 1 мкл антисмыслового праймера (SEQ ID NO:22), 2 мкл "RediLoad" (Research Genetics, Inc.), 0,4 мкл 50Х смеси Advantage KlenTaq Polymerase (Clontech Laboratories, Inc.), 25 нг ДНК из индивидуального гибридного клона или контроля и ddH2O для общего объема 20 мкл. На реакции наслаивали равное количество минерального масла и их герметизировали. Условия ПЦР-циклера были следующими: 1 начальный цикл 5-минутной денатурации при 95° С, 35 циклов 1-минутной денатурации при 95° С, 1-минутный отжиг при 60° С и 1,5-минутное удлинение при 72° С, с последующим конечным 1 циклом удлинения в течение 7 мин при 72° С. Реакции разделяли электрофорезом на 3% агарозном NuSieve GTG геле (FMS Bioproducts, Rockland, ME).
Пример 4
Конструирование экспрессирующих векторов млекопитающих zsig37NEE/pZP9 и zsig37CEE/pZP9
Были изготовлены два экспрессирующих вектора для полипептида zsig37, zsig37NEE/pZP9 и zsig37CEE/pZP9, причем эти конструкции были сконструированы для экспрессии полипептида zsig37, имеющего С- или N-концевую метку Glu-Glu.
zsig37NEE/pZP9
ПЦР-генерируемый ДНК-фрагмент zsig37 800 п.н. получали с использованием ZC15040 (SEQ ID NO:24) и ZC15033 (SEQ ID NO:25) в качестве ПЦР-праймеров и матрицы, описанной в Примере 1 выше. ПЦР-реакцию инкубировали при 94° С в течение 3 мин и затем проводили в течение 5 циклов 94° С в течение 30 с, 30° С в течение 20 с и 72° С в течение 1 мин, с последующими 25 циклами при 94° С в течение 30 с, 64° С в течение 20 с и 72° С в течение 1 мин. Затем следовало 5-минутное удлинение при 72° С. Затем полученный ПЦР-продукт подвергали электрофорезу на 0,9% ТВЕ-агарозном геле с 1× ТВЕ-буфером. Полосу предсказанного размера вырезали и ДНК очищали из геля при помощи смолы Qiaex II® (Qiagen) в соответствии с инструкциями изготовителя. Эту ДНК расщепляли рестриктазами BamHI и XbaI с последующими экстракцией и осаждением.
Вырезанный, расщепленный рестриктазами ДНК-фрагмент zsig37 субклонировали в плазмиду NEE/pZP9, которая была разрезана рестриктазами BamHI и XbaI. Экспрессирующий вектор zsig37NEE/pZP9 включает ТРА-лидер и присоединяет метку Glu-Glu (SEQ ID NO:26) к N-концу последовательности полинуклеотидов, кодирующей полипептид zsig37. Плазмида NEE/pZP9 (депонированная в American Type Culture Collection, 12301 Parklawn Drive, Rockville, MD, ATCC No. 98668) представляет собой экспрессирующий вектор млекопитающих, содержащий экспрессионную кассету, имеющую мышиный промотор металлотионеина-1, лидерный пептид ТРА, за которым следуют последовательность, кодирующая метку Glu-Glu (SEQ ID NO:26), множественные сайты рестрикции для встраивания кодирующих последовательностей и терминатор гормона роста человека. Эта плазмида содержит также точку инициации репликации Е. coli, экспрессионную единицу селектируемого маркера млекопитающих, имеющую промотор SV40, энхансер и точку инициации репликации, ген DHFR и терминатор SV40.
zsig37CEE/pZP9
ПЦР-генерируемый ДНК-фрагмент zsig37 866 п.н. получали в соответствии с процедурой, описанной выше, с использованием ZC15721 (SEQ ID NO:27) и ZC15035 (SEQ ID NO:28) в качестве ПЦР-праймеров. Очищенный ДНК-фрагмент расщепляли рестриктазами EcoRI и BamHI, очищали на геле с использованием смолы Qiaex II, как описано выше.
Вырезанную и расщепленную рестриктазами ДНК zsig37 субклонировали в плазмиду CEE/pZP9, которая была разрезана рестриктазами EcoRI и BamHI. Экспрессирующий вектор zsig37CEE/pZP9 использует сигнальный пептид нативного zsig37 и, Glu-Glu-эпитоп (SEQ ID NO:26) присоединен при С-конце как вспомогательное средство для очистки. Плазмида CEE/pZP29 (депонированная в American Type Culture Collection, 12301 Parklawn Drive, Rockville, MD, ATCC No. 98668) представляет собой экспрессирующий вектор млекопитающих, содержащий экспрессионную кассету, имеющую мышиный промотор металлотионеина-1, множественные сайты рестрикции для встраивания кодирующих последовательностей, последовательность, кодирующую метку Glu-Glu (SEQ ID NO:26), стоп-кодон и терминатор гормона роста человека. Эта плазмида содержит также точку инициации репликации Е. соli, экспрессионную единицу селектируемого маркера млекопитающих, имеющую промотор SV40, энхансер и точку инициации репликации, ген DHFR и терминатор SV40.
Для N- и С-меченых конструкций примерно 30 нг расщепленных рестриктазами вставок и 50 нг соответствующих векторов лигировали при комнатной температуре в течение 4 ч. Один микролитр каждой смеси лигирования независимо вводили электропорацией в компетентные клетки DH10B (GIBCO BRL, Gaithersburg, MD) в соответствии с указаниями изготовителя и высевали на LB-чашки, содержащие 50 мг/мл ампициллина, и инкубировали в течение ночи.
Колонии подвергали скринингу при помощи ПЦР, как описано выше. Для скринингов zsig37NEE/pZP9 и zsig37CEE/pZP9 праймерами были ZC13006 (SEQ ID NO:29) и ZC13007 (SEQ ID NO:20). ПЦР-реакцию инкубировали при 94° С в течение 2,5 мин и затем проводили в течение 25 циклов 94° С в течение 10 с, 58° С в течение 20 с и 72° С в течение 1 мин. Затем следовало 5-минутное удлинение при 72° С. Последовательность вставок позитивных клонов, фрагмент 1013 п.н. для zsig37NEE и фрагмент 950 п.н. для zsig37CEE подтверждали анализом последовательности. Крупномасштабное получение плазмид проводили с использованием набора QIAGEN® Maxi prep kit (Qiagen) в соответствии с инструкциями изготовителя.
Пример 5
Трансфекция и экспрессия полипептидов
zsig37NEE и zsig37CEE
Клетки ВНК 570 (АТСС No. CRL-10314) высевали в чашки 10 см для культуры ткани и давали им расти до приблизительно 50-70% конфлюэнтности в течение ночи при 37° С, 5% СO2, в среде DMEM/ФТС (DMEM, Gibco/BRL High Glucose, (GIBCO BRL, Gaithersburg, MD), 5% фетальная телячья сыворотка (Hyclone, Logan, UT), 2 мкМ L-глутамин (JRH Biosciences, Lenexa, KS), 1 мкМ пируват натрия (Gibco BRL)). Затем эти клетки трансфицировали плазмидой zsig37NEE/pZP9 (М-концевая метка Glu-Glu) или плазмидой zsig37CEE/pZP9 (С-концевая метка Glu-Glu) с использованием ЛипофектаминаTM (Gibco BRL) в бессывороточной композиции среды (SF) (DMEM, Gibco/BRL High Glucose, (GIBCO BRL, Gaithersburg, MD), 2 мМ L-глутамин, 2 мМ пируват натрия, 10 мкг/мл трансферрина, 5 мкг/мл инсулина, 10 мкг/мл фетуина и 2 нг/мл селена). Шестнадцать микрограммов zsig37NEE/pZP9 и 16 мкг zsig37CEE/pZP9 отдельно разводили в пробирки на 15 мл до общего конечного объема 640 мкл среды SF. В отдельных пробирках 35 мкл Липофектамина™ (Gibco BRL) смешивали с 605 мкл среды SF. Смесь с ЛипофектаминомTM добавляли к смеси ДНК и давали инкубироваться приблизительно 30 мин при комнатной температуре. Пять миллилитров среды SF добавляли к смеси ДНК: ЛипофектаминTM. Клетки промывали один раз 5 мл среды SF, отсасывали и добавляли смесь ДНК: ЛипофектаминTM. Клетки инкубировали при 37° С в течение пяти часов, затем в чашку добавляли 6,4 мл DMEM/10% ФТС, 1% среду PSN. Чашку инкубировали при 37° С в течение ночи и смесь ДНК: ЛипофектаминTM заменяли свежей средой ФТС/DMEM на следующий день. В день 2 после трансфекции клетки распределяли в селективную среду (ESTER #1 с 1 мкМ МТХ) в чашки 150 мм при разведениях 1:50, 1:100 и 1:200. В чашки подавали свежую селективную среду в день 5 после трансфекции.
Скрининг колоний
Спустя приблизительно 10-12 дней после трансфекции одну культуральную чашку 150 мм резистентных к метотрексату колоний выбирали из каждой трансфекции, среду отсасывали, чашки промывали 10 мл бессывороточной среды ESTEP 2 (668,7 г/50 л DMEM (Gibco), 5,5 г/50 л пировиноградной кислоты, 96% соль натрия (Mallinckrodt), 185,0 г/50 л NаНСО3 (Mallinkrodt), 5,0 мг/мл, 25 мл/50 л инсулина, 10,0 мг/мл и 25 мл/50 л трансферрина). Промывную среду отсасывали и заменяли 5 мл бессывороточной среды ESTEP 2. Затем стерильную тефлоновую сетку (Spectrum Medical Industries, Los Angeles, CA), предварительно пропитанную в бессывороточной среде ESTEP 2, помещали на клетки. Затем стерильный нитроцеллюлозный фильтр, предварительно пропитанный в бессывороточной среде ESTEP 2, помещали на сетку. Метки для ориентации на нитроцеллюлозе переносили на чашку с культурой. Затем эти чашки инкубировали в течение 5-6 ч в термостате при 37° С и 5% CO2. После инкубирования фильтр удаляли и среду отсасывали и заменяли средой DMEM/5% ФТС, 1Х PSN (Gibco BRL). Затем фильтр помещали в герметизируемый мешок, содержащий 50 мл буфера (25 мМ Трис, 25 мМ глицин, 5 мМ β -меркаптоэтанол), и инкубировали в водяной бане на 65° С в течение 10 мин. Фильтры блокировали в 10% нежирном сухом молоке/буфере Вестерн А (Вестерн А: 50 мМ Трис, рН 7,4, 5 мМ ЭДТА, 0,05% NP-40, 150 мМ NaCl и 0,25% желатин) в течение 15 мин при комнатной температуре на ротационном шейкере. Затем фильтр инкубировали с конъюгатом антитело против Glu-Glu-HRP (пероксидаза хрена) при разведении 1:1000 в 2,5% нежирном сухом молоке/буфере Вестерн А (Вестерн А: 50 мМ Трис, рН 7,4, 5 мМ ЭДТА, 0,05% NP-40, 150 мМ NaCl и 0,25% желатин) в течение ночи при 4° С на ротационном шейкере. Затем фильтр промывали три раза при комнатной температуре в ЗФР плюс 0,1% Твин 20, 5-15 мин на промывку. Фильтр проявляли реагентом ECL (Amersham Corp., Arlington Heights, IL) в соответствии с указаниями изготовителя и экспонировали на пленке (Hyperfilm ECL, Amersham) в течение приблизительно 5 мин.
Пленку сопоставляли с чашкой, содержащей колонии. С использованием этой пленки в качестве направляющего устройства отбирали подходящие колонии. Стерильные диски 3 мм для снятия колоний (PGC Scientific Corp., Frederick, MD) замачивали в трипсине и помещали на эти колонии. Двенадцать колоний для каждой конструкции переносили в 200 мкл селективной среды в 96-луночном планшете. Серию из семи двукратных разведений выполняли для каждой колонии. Эти клетки выращивали в течение одной недели при 37° С и в это время лунки, которые получили самое низкое разведение клеток, которые теперь находятся при оптимальной плотности, отбирали, трипсинизировали и переносили в 12-луночный планшет, содержащий селективную среду. Культуральную чашку 150 мм также трипсинизировали и остальные клетки объединяли и подвергали Вестерн-анализу и тестированию на микоплазму. Объединенный материал замораживали для хранения.
Эти клоны переносили непосредственно из 12-луночного планшета в две колбы Т-75. Одну колбу выдерживали для продолжения роста клеток, клетки во второй колбе выращивали в бессывороточной среде ESTEP 2 и собирали для Вестерн-блот-анализа. Клоны каждой из экспрессионных конструкций, на основе Вестерн-блот-анализа, отбирали, объединяли и переносили в крупномасштабную культуру.
Пример 6
Крупномасштабная экспрессия zsig37CEE клетками млекопитающих
Одну колбу Т-162, содержащую конфлюэнтные клетки, экспрессирующие zsig37CEE, и одну колбу, содержащую zsig37NEE, полученный из процедуры экспрессии, описанной выше, распределяли в четыре колбы Т-162, каждую. Одну из четырех полученных колб использовали для замораживания четырех криофлаконов, а другие, три колбы использовали для генерирования клеточной фабрики Nunc (кассеты биореакторов для крупномасштабного производства клеток).
Клетки из трех колб Т-162 zsig37CEE и zsig37NEE использовали для независимого засева двух клеточных фабрик Nunc (10 слоев, коммерчески доступных из VWR). Вкратце, клетки из колб Т-162, описанных выше, отделяли при помощи трипсина, объединяли и добавляли к 1,5 л среды ESTEP1 (668,7 г/50 л DMEM (Gibco), 5,5 г/50 л пировиноградной кислоты, 96% соль натрия (Mallinkrodt), 185,0 г/50 л NaHCO3 (Mallinkrodt), 5,0 мг/мл и 25 мл/50 л инсулина (JRH Biosciences), 10,0 мг/мл и 25 мл/50 л трансферрина (JRH Biosciences), 2,5 л/50 л фетальной телячьей сыворотки (охарактеризованной) (Hyclone), 1 мкМ МТХ, с рН, доведенным до 7,05 +/-0,05), предварительно нагретой до 37° С. Затем среду, содержащую эти клетки, выливали в клеточные фабрики Nunc через воронку. Клеточные фабрики помещали в термостат с 37° С/5,0% СO2.
При 80-100% конфлюэнтности на содержимом клеточных фабрик Nunc проводили визуальный тест на загрязнение (изменение окраски фенолового красного). Поскольку загрязнения не наблюдали, супернатант из конфлюэнтных фабрик выливали в небольшой резервуар для сбора, брали пробы и выбрасывали. Затем прикрепленные клетки промывали один раз 400 мл ЗФР. Для отделения клеток из фабрик в каждую фабрику добавляли 100 мл трипсина и удаляли, и затем клетки инкубировали в течение 5-10 мин в оставшемся трипсине. Клетки собирали после двух 200 мл-промывок средой ESTEP1. В каждую из десяти бутылей, содержащих среду ESTEP1 (1,5 л каждая, при 37° С), добавляли 40 мл собранных клеток. Затем одну бутыль на 1,5 л использовали для наполнения одной Nunc-фабрики. Каждую клеточную фабрику помещали в термостат при 37° С/5,0% CO2.
При 80-90% конфлюэнтности на содержимом клеточных фабрик Nunc проводили визуальный тест на загрязнение (изменение окраски фенолового красного). Поскольку загрязнения не наблюдали, супернатант из конфлюэнтных фабрик выливали в небольшой резервуар для сбора, брали пробы и выбрасывали. Затем клетки промывали один раз 400 мл ЗФР. 1,5 л среды ESTEP2 (668,7 г/50 л DMEM (Gibco), 5,5 г/50 л пировиноградной кислоты, 96% соль натрия (Mallinckrodt), 185,0 г/50 л NаНСО3 (Mallinkrodt), 5,0 мг/мл, 25 мл/50 л инсулина, 10,0 мг/мл и 25 мл/50 л трансферрина) добавляли в каждую клеточную фабрику Nunc. Эти клеточные фабрики инкубировали при 37° С/5,0% СО2.
При приблизительно 48 ч на содержимом клеточных фабрик Nunc проводили визуальный тест на загрязнение (изменение окраски фенолового красного). Супернатант из каждой фабрики выливали в небольшие резервуары для сбора. Свежую бессывороточную среду (1,5 л) заливали в каждую клеточную фабрику Nunc и фабрики инкубировали при 37° С/5,0% СО2. Один мл собранного супернатанта для каждой конструкции переносили на предметное стекло для микроскопа и подвергали микроскопическому анализу на загрязнение. Содержимое небольших резервуаров для сбора клеток для каждой конструкции объединяли и сразу же фильтровали. Затем проводили второй сбор клеток, по существу, как описано выше при 48 ч, и после этого клеточные фабрики выбрасывали. Асептически собранное устройство с серией фильтров использовали для асептического фильтрования супернатанта собранных клеток (кондиционированной среды). Сборку проводили следующим образом: трубки привязывали проволокой к фильтру Opti-Cap (Millipore Corp., Bedford, MA) и фильтру Gelman Supercap 50 (Gelman Sciences, Ann Arbor, Ml). Фильтр Supercap 50 присоединяли также к стерильному закрытому крышкой резервуару, помещенному в вытяжной шкаф; трубку, расположенную выше относительно фильтра Millipore Opti-cap, вставляли в перистальтический насос и свободный конец этой трубки помещали в большой резервуар для сбора. Перистальтический насос работал между 200 и 300 об/мин до тех пор, пока вся кондиционированная среда не проходила через конечный фильтр 0,22 мкм в стерильный резервуар для сбора. Фильтрат помещали в холодную комнату (4° С) и держали в ней вплоть до очистки. Среду концентрировали в 10 раз с использованием концентратора Millipore с отсечением 5 кДа (Millipore Corp., Bedford, МА) в соответствии с указаниями изготовителя и подвергали Вестерн-блот-анализу с использованием антитела против FLAG-метки (Kodak).
zsig37CEE:
5 колб Т-162=0,12 мг/л, 38 кДа;
1 фабрика, ФТС=0,12 мг/л, 38 кДа;
10 фабрик, ФТС=0,12 мг/л, 38 кДа;
10 фабрик (№1), SF=1,2 мг/л, 38 кДа и
10 фабрик (№2), SF=3,56 мг/л, 38 кДа.
zsig37NEE:
5 колб Т-162=0,137 мг/л, 35 кДа;
1 фабрика, ФТС=0,137 мг/л, 35 кДа;
10 фабрик, ФТС=0,137 мг/л, 35 кДа;
10 фабрик (№1), SF=1,37 мг/л, 35 кДа и
10 фабрик (№2), SF=4,11 мг/л, 35 кДа.
Пример 7
Очистка zsig37 NEE и zsig37 CEE
Если нет других указаний, все операции проводили при 4° С. Следующую процедуру использовали для очистки zsig37, содержащего N-концевую или С-концевую метки Glu-Glu (ЕЕ). В целом 25 л кондиционированной среды от клеток почки детеныша хомяка (ВНК) последовательно стерильно фильтровали через 4-дюймовый, 0,2 мМ-фильтр Millipore (Bedford, MA) OptiCap capsule filter и 0,2 мМ-фильтр Gelman (Ann Arbor, Ml) Supercap 50. Затем этот материал концентрировали до приблизительно 1,3 л при помощи тангенциального проточного концентратора Millipore Proflux A30, снабженного мембраной S10Y3 Amicon (Bedford, MA) с отсечением 3000 кДа. Концентрированный материал опять стерильно фильтровали с фильтром Gelman, описанным выше. Смесь ингибиторов протеаз добавляли к концентрированной кондиционированной среде до конечной концентрации 2,5 мМ этилендиаминтетрауксусной кислоты (ЭДТА, Sigma Chemical Со. St. Louis, МО), 0,001 мМ лейпептина (Boehringer-Mannheim, Indianapolis, IN), 0,001 мМ пепстатина (Boehringer-Mannheim) и 0,4 мМ Pefabloc (Boehringer-Mannheim). Пробу 25,0 мл анти-ЕЕ-Сефарозы, приготовленной, как описано ниже, добавляли к этой пробе для адсорбции партии и смесь осторожно встряхивали на аппарате для роллерной культуры Wheaton (Millville, NJ) в течение 18 ч при 4° С.
Затем эту смесь выливали в колонку 5,0× 20,0 см Econo-Column (Bio-Rad, Laboratories, Hercules, CA) и этот гель промывали 30 колоночными объемами забуференного фосфатом солевого раствора (ЗФР). Неудержанную проходящую через колонку фракцию выбрасывали. Как только поглощение эффлюента при 280 нм становилось менее 0,05, прохождение через колонку уменьшали до нуля и анти-ЕЕ-Сефарозный гель промывали в виде порций 2,0 колоночными объемами ЗФР, содержащего 0,4 мг/мл пептида ЕЕ (AnaSpec, San Jose, CA). Используемый пептид имеет последовательность Glu-Tyr-Met-Pro-Val-Asp, SEQ ID NO:31). После 1,0 ч при 4° С ток возобновляли и элюированный белок собирали. Эту фракцию называли пептидной элюцией. Затем анти-ЕЕ-Сефарозный гель промывали 2,0 колоночными объемами 0,1 М глицина, рН 2,5, и эту глициновую промывку собирали отдельно. рН этой элюированной глицином фракции доводили до 7,0 добавлением небольшого объема 10Х ЗФР и фракцию хранили при 4° С для последующего анализа, если необходимо.
Пептидную элюцию концентрировали до 5,0 мл при помощи мембранного концентратора с отсечением молекулярной массы 15000 (Millipore, Bedford, MA) в соответствии с инструкциями изготовителя. Концентрированную пептидную элюцию отделяли от свободного пептида хроматографией на колонке 1,5× 50 см Сефадекса G-50 (Pharmacia, Piscataway, NJ), уравновешенной в ЗФР, при скорости тока 1,0 мл/мин с использованием BioCad Sprint HPLC (ВЖХ) (PerSeptive BioSystems, Framingham, MA). Фракции по 2 мл собирали и проводили мониторинг поглощения при 280 нм. Собирали первый пик материала, поглощающего при 280 нм и элюирующегося вблизи свободного объема колонки. Эта фракция была чистым zsig37 NEE или zsig37 CEE. Этот чистый материал концентрировали, как описано выше, анализировали при помощи электрофореза в ДСН-ПААГ и Вестерн-блоттинга с антителами против ЕЕ и брали пробы для аминокислотного анализа и N-концевого секвенирования. Остаток этой пробы делили на аликвоты и хранили при -80° С в соответствии с нашими стандартными процедурами.
Электрофорез zsig37 NEE на гелях ДСН-ПААГ в отсутствие восстанавливающих агентов показал одну основную полосу, окрашенную Кумасси синим, со средней молекулярной массой 39000 и несколько минорных компонентов с молекулярными массами между 60000 и 116000. Все эти полосы обнаружили перекрестную реактивность с анти-ЕЕ-антителами на Вестерн-блотах. В присутствии восстанавливающего агента единственной наблюдаемой полосой был белок 39000 кДа и интенсивность его окрашивания Кумасси синим увеличивалась. Эта полоса также обнаружила перекрестную реактивность с анти-ЕЕ-антителом на Вестерн-блотах.
Для zsig37 CEE электрофорез на гелях ДСН-ПААГ в отсутствие восстанавливающих агентов показал одну основную полосу, окрашенную Кумасси синим, со средней молекулярной массой 39000 и несколько минорных компонентов с молекулярными массами между 60000 и 116000. На Вестерн-блотах только полосы со средними молекулярными массами 150000, 116000 и 60000 обнаружили перекрестную реактивность с анти-ЕЕ-антителами. В присутствии восстанавливающих агентов наблюдали только окрашенную Кумасси синим полосу при 39000 кДа, и этот материал обнаружил перекрестную реактивность с анти-ЕЕ-антителом на Вестерн-блотах. В этих условиях небольшое количество перекрестно-реактивного материала наблюдали также при 150000 кДа.
Приготовление анти-ЕЕ-Сефарозы
Объем слоя 100 мл протеин G-Сефарозы (Pharmacia, Piscataway, NJ) промывали 3 раза 100 мл ЗФР, содержащего 0,02% азида натрия, с использованием фильтрующего элемента 0,45 мкм Nalgen на 500 мл. Гель промывали 6,0 объемами 200 мМ триэтаноламина, рН 8,2 (ТЭА, Sigma, St. Louis, МО) и добавляли равный объем раствора антител против ЕЕ, содержащего 900 мг антитела. После инкубирования в течение ночи при 4° С несвязанное антитело удаляли промыванием смолы 5 объемами 200 мМ ТЭА, как описано выше. Смолу ресуспендировали в 2 объемах ТЭА, переносили в подходящий резервуар и добавляли диметилпимилимидат-2НС1 (Pierce, Rockford, IL), растворенный в ТЭА, до конечной концентрации 36 мг/мл геля. Гель качали при комнатной температуре в течение 45 мин и жидкость удаляли при помощи фильтрующего элемента, как описано выше. Затем неспецифические сайты на этом геле блокировали инкубированием в течение 10 мин при комнатной температуре с 5 объемами 20 мМ этаноламина в 200 мМ ТЭА. Гель промывали 5 объемами ЗФР, содержащего 0,02% азида натрия, и хранили в этом растворе при 4° С.
Пример 8
Анализы адгезии и пролиферации
Способность zsig37 стимулировать адгезию (слипание) и распределение клеток TF-1 анализировали следующим образом. Готовили серию разведений из меченого Glu-Glu на С-конце zsig37, от 10 до 0,0625 мкг/мл, либо в ЗФР, либо в ELISA-буфере для сенсибилизации поверхностей (0,1 М NaCO3), и каждое разведение высевали в 96-луночный планшет (Costar, Pleasanton, CA) при 100 мкл/лунку. Планшеты инкубировали при 37° С, 5% СO2 в течение 2 ч. Затем планшеты промывали 3 раза средой RPMI/10% ФТС (RPMI 1640, 2 мМ L-глутамин, 110 мкг/мл пирувата натрия, PSN и 10% инактивированная нагреванием фетальная телячья сыворотка) и проводили блокирование в течение 15 мин.
Клетки TF-1 (полученные из клеток острого миелоидного лейкоза) ресуспендировали в RPMI/10% ФТС и высевали при 10000 клеток/лунку в покрытые zsig37CEE 96-луночные планшеты в конечном объеме 120 мкл/лунку. Планшеты инкубировали при 37° С в атмосфере 5% СО2 в течение 2 ч. Затем планшеты промывали ЗХ ЗФР и добавляли 200 мкл/лунку среды для выращивания (RPMI/10% ФТС, 5 нг/мл GM-CSF). Клетки обследовали микроскопически до и после промывания.
Тест включения красителя использовали также для количественного измерения числа прикрепленных (адгезивных) клеток на основе колориметрического изменения и увеличения флуоресцентного сигнала. В 96-луночные планшеты добавляли Alamar BlueTM (AccuMed, Chicago, IL) и клетки инкубировали при 37° С в атмосфере 5% СO2 в течение ночи. Затем планшеты сканировали с использованием флуорометра с длиной волны возбуждения 544 нм и длиной волны испускания 590 нм. Больше прикрепленных (адгезивных) клеток было на покрытых zsig37СЕЕ-ЗФР планшетах, чем на покрытых zsig37CEE-0,1 M NaCO3 планшетах. Добавление растворимого zsig37 не блокировало адгезию клеток со связанным zsig37.
Второй тест проводили с использованием клеточных линий TF-1, DA-1 (IL-3-зависимой клеточной линии, полученной из лимфатического узла мыши с В-клеточной лимфомой путем выращивания в IL-3-среде (предоставленной доктором Kenneth Kaushanski, University of Washington, Seattle, WA)), пре-В (клеток мышиного мозга р53-/-, IL-7-зависимых, В220+, Thyl-низких, Sca-1+) и A7BaF-3, как описано выше, при 5000 клеток/лунку. Клетки ВНК также высевали при 500 клеток/лунку. Zsig37 усиливал рост клеток A7BaF-3 и слегка ингибировал рост клеток DA-1.
Пример 9
Последовательность мышиного ортолога
Новые полинуклеотиды данного изобретения, кодирующие полипептид zsig37 человека, использовали для скрининга базы данных мышиных EST на гомологичные мышиные последовательности. Была обнаружена и предсказана единственная последовательность EST, гомологичная последовательности zsig37 человека. Для идентификации соответствующей кДНК для секвенирования использовали клон, который, как считали, содержал, возможно, всю кодирующую последовательность. С использованием набора Invitrogen S.N.A.P.TM Miniprep (Invitrogen Corp.) в соответствии с инструкциями изготовителей получали 5 мл ночной культуры в LB + 50 мкг/мл ампициллина. Матрицу секвенировали на ДНК-секвенаторе модели 377 ABIPRISMTM (Perkin-Elmer Cetus, Norwalk, СТ.) с использованием набора для реакции секвенирования Big Dye Terminator Cycle Sequencing Ready Reaction Kit ABIPRISMTM (Perkin-Elmer Corp.) в соответствии с инструкциями изготовителя. Олигонуклеотиды ZC694 (SEQ ID NO:6), ZC6768 (SEQ ID NO:32), ZC18297 (SEQ ID NO:33), ZC18298 (SEQ ID NO:34), ZC18402 (SEQ ID NO:35), ZC18403 (SEQ ID NO:36), ZC18456 (SEQ ID NO:37), ZC18457 (SEQ ID NO:38), ZC18560 (SEQ ID NO:39), ZC18561 (SEQ ID NO:40), ZC18687 (SEQ ID NO:41) и ZC18688 (SEQ ID NO:42) использовали для комплектования этой последовательности из этого клона. Реакции секвенирования проводили в термоциклере Hybaid OmniGene Temperature Cycling System (National Labnet Co., Woodbridge, NY). Программное обеспечение для анализа последовательности SEQUENCHERTM 3.1 (Gene Codes Corporation, Ann Arbor, Ml) использовали для анализа данных. Полученная последовательность 2559 п.н. описана в SEQ ID NO:43 и расшифрованная аминокислотная последовательность в SEQ ID NО:44. Сопоставление с нуклеотидной последовательностью zsig37 человека (SEQ ID NO:1) показывает 77% идентичность на нуклеотидном уровне. Предполагаемая последовательность аминокислот (SEQ ID NO:44) имеет 77% идентичность с последовательностью полипептидов человека (SEQ ID NO:2).
Пример 10
Анализы на основе клеток
Полипептиды zsig37 анализировали в высокопроизводительном анализе in vitro для идентификации веществ, которые избирательно активируют клеточные ответные реакции в иммортализованных линиях остеобластных клеток. В этом анализе использовали линию зрелых остеобластных клеток, полученных из р53-/-(дефектных) мышей, ССС4, которая трансфицирована плазмидой, содержащей индуцируемый элемент сывороточного ответа (SRE), запускающий экспрессию люциферазы. Эти клетки экспрессируют также эндогенные рецепторы РТН, PDGF и bFGF. Стимуляция этого элемента SRE и, следовательно, экспрессии люциферазы в клетках ССС4 указывает на то, что данная химическая частица, по-видимому, стимулирует митогенез в остеобластах.
Линии ССС4 трипсинизировали и доводили до 5× 104 клеток/мл в среде для культивирования (альфа-МЕМ, 1% инактивированная нагреванием фетальная телячья сыворотка, 1 мМ пируват Na и 2 мМ L-глутамат) и высевали (200 мкл/лунку) в непрозрачные белые микротитрационные планшеты Dynatech Microlite (Dynatech, Chantilly, VA) и инкубировали в течение ночи при 37°С, 5,0% СО2. Затем среду для выращивания отсасывали и заменяли 50 мкл/лунку тест-среды (F-12 НАМ, 0,5% бычий сывороточный альбумин, 20 мМ HEPES, 1 мМ пируват Na и 2 мМ L-глутамат). Серийные разведения zsig37 производили в тест-среде (0,29-1000 нг/мл конечная тест-концентрация) и добавляли в лунки. Пробы zsig37 анализировали в трех повторностях. Также использовали сывороточный (негативный) и bFGF (позитивный) контроли. Конечная концентрация bFGF была 3 нг/мл. Контроли анализировали в четырех повторностях. Планшеты инкубировали в течение 4 ч при 37° С, 5,0% СO2. Затем тест-среду отсасывали и планшеты промывали один раз ЗФР. Затем к каждой лунке добавляли 25 мкл лизисного буфера (Luciferase Assay Reagent, E1501, Promega Corp., Madison, WI). Планшеты инкубировали в течение 15 мин при комнатной температуре. Пятьдесят микролитров/лунку субстрата люциферазы (Luciferase Assay Reagent, E1501, Promega Corp.) добавляли и люциферазную активность детектировали при помощи Labsystems LUMINOSKAN® при 2 с/лунку после 1-секундной задержки. Средний базовый (неиндуцированный) сигнал вычитали из всех считываний в виде процента максимальной индукции, производимой 3 нг/мл bFGF.
Zsig37 стимулирует экспрессию люциферазы в этом тесте, что свидетельствует о том, что он стимулирует остеобласты. Zsig37 стимулирует на 73-75% максимально в концентрации 1000 нг/мл.
Проводили дубликатный анализ на имитатор (миметик) фактора роста для определения, действует ли zsig37 как имитатор (миметик) фактора роста, в частности, как лиганды тирозинкиназного рецептора PDGF, bFGF и EGF (инсулин-рецептор-негативные). Клональную клеточную линию Swiss 3T3, полученную из мышей Swiss 3T3, которая трансфицирована плазмидой, содержащей индуцируемый элемент сывороточного ответа (SRE), запускающий экспрессию люциферазы, использовали в этом анализе. Эти клетки экспрессируют также эндогенные рецепторы РМА, EGF и bFGF. Стимуляция этого элемента SRE и, следовательно, экспрессии люциферазы в клетках Swiss 3T3 указывает на то, что данная химическая частица, по-видимому, имитирует активность факторов роста PDGF, bFGF и EGF.
Клетки Swiss 3T3 трипсинизировали и доводили до 5× 104 клеток/мл в среде для культивирования, высевали и инкубировали, как описано выше. Затем среду для выращивания отсасывали и заменяли 50 мкл/лунку тест-среды (F-12 НАМ, 0,5% бычий сывороточный альбумин, 20 мМ HEPES). Серийные разведения zsig37 производили в тест-среде (0,29-1000 нг/мл конечная тест-концентрация) и добавляли в лунки. Пробы zsig37 анализировали в трех повторностях. Также использовали сывороточный (негативный) и bFGF (позитивный) контроли для усиления пролиферации клеток. Конечная концентрация bFGF была 3 нг/мл. Контроли анализировали в четырех повторностях. Планшеты инкубировали в течение 5 ч при 37° С, 5,0% СO2. Затем тест-среду отсасывали и планшеты промывали один раз ЗФР. Затеял к каждой лунке добавляли 25 мкл лизисного буфера (Luciferase Assay Reagent, E1501, Promega Corp., Madison, WI). Планшеты инкубировали в течение 15 мин при комнатной температуре. Сорок микролитров/лунку субстрата люциферазы (Luciferase Assay Reagent, E1501, Promega Corp.) добавляли и люциферазную активность детектировали при помощи Labsystems LUMINOSKAN® при 2 с/лунку после 1-секундной задержки. Средний базовый (неиндуцированный) сигнал вычитали из всех считываний, которые выражали в виде процента максимальной индукции, производимой 3 нг/мл bFGF. Пятичасовая обработка этой клеточной линии bFGF, DDGF, EGF или РМА приводит к 25-50-кратной индукции SRE-экспрессии люциферазы.
Zsig37, по-видимому, не стимулирует экспрессию люциферазы в этом тесте. Zsig37 стимулирует на 0,2-0,1% максимально при 1000 нг/мл.
Пример 11
Введение Zsig37 in vivo посредством адреновирусной доставки
Двадцать четыре самца и 24 самки мышей C57B16/J в возрасте приблизительно 12 недель (Jackson Labs, Bar Harbor, ME) взвешивали, измеряли их температуру тела и наблюдали потребление пищи ежедневно в течение четырех дней перед инъекцией (дни от -4 до -1). В день 0 мышей делили на три группы и они получали 0,1 мл вируса (AdV-пустой 1,8× 1011 вирусных частиц / 0,1 мл или AdV-zsig37-CEE 5× 1011 вирусных частиц/0,1 мл) внутривенной инъекцией в хвостовую вену или вообще не получали инъекции. Инъекция должна была приводить к инфицированию печени хозяина, и экспрессия доставленного вирусом гена должна была начинаться в пределах 24 ч и продолжаться в течение 1-4 недель. Тестировали три группы мышей. Группа 1, необработанные, n=8 для самцов и для самок. Группа 2, AdV-пустой (пустой вирус), n=8 для самцов и для самок. Группа 3, AdV-zsig37 CEE, n=8 для самцов и для самок.
Температуры тела животных, веса и вес принятой внутрь пищи подвергали мониторингу во время трехнедельного исследования. Не было обнаружено различия между группами.
В день 21 самок мышей эвтанизировали и умерщвляли цервикальным смещением и в день 22 умерщвляли самцов. Животных обескровливали и ткани брали для аутопсии.
Стандартную панель сывороточной химии готовили во время умерщвления. Все параметры печени, почки и метаболизма были в пределах нормы. Однако было различие между группой, обработанной zsig37, и группой, обработанной пустым вирусом. Животные с zsig37 имели более высокий (гипер)липемический индекс, чем контроли с пустым вирусом. Это различие не было значимым, однако оно оправдывало дальнейшее исследование. Анализировали общие свободные жирные кислоты на оставшейся сыворотке из каждого животного. Наблюдали статистически значимое различие в уровнях свободных жирных кислот в сыворотке между самцами мышей (р=0,0379), получившими пустой вирус, и самцами мышей, получивших кодирующий zsig37 вирус; мыши с zsig37 имели более высокие уровни. Различие, хотя и статистически незначимое, наблюдали также у самок (р=0,3357). Печень, селезенку, почку, тимус, сердце и мозг взвешивали после их извлечения. Не было обнаружено различие между группами обработок. Гистопатологический анализ этих тканей и костного мозга не выявил различия между группами обработок.
Для подтверждения приведенных выше результатов был проведен второй скрининг, подобный описанному выше, со следующими модификациями. Тестировали три группы: а) не получавшие обработки и голодавшие, b) получавшие AdV-нуль (пустой) и голодавшие, с) получавшие AdV-zsig37-CEE и голодавшие, содержащие 20 мышей C57B16/J, по 10 самцов и 10 самок, каждая. Мышей подвергали голоданию в течение ночи и собирали 100 мкл сыворотки для установления базового уровня (фона) для следующих параметров: глюкоза голодания, ТР, щелочная фосфатаза, холестерин, триглицериды, свободные жирные кислоты и инсулин. Вес тела измеряли три раза в неделю. В день 0 мышей инъецировали в латеральную хвостовую вену 0,1 мл соответствующего раствора вируса. Кровь собирали в день 17 после ночного голодания. После 3 недель мышей умерщвляли и всю кровь извлекали. Порцию крови смешивали с ЭДТА для анализа крови, а остальную часть подвергали повторному анализу и скринингу, как описано выше. Органы собирали, а тушки сохраняли для гистопатологии.
Пример 12
Вазодилатация колец аорты
Действие zsig37 на вазодилатацию колец аорты измеряли в соответствии с процедурами (Dainty et al., J. Pharmacol. 100:767, 1990 и Rhee et al., Neurotox. 16:179, 1995). Вкратце, кольца аорты 4 мм длиной получали из 4-месячных крыс Sprague-Dawley и помещали в модифицированный раствор Кребса (118,5 мМ NaCl, 4,6 мМ KCl, 1,2 мМ MgSO4·7 H2O, 1,2 мМ КН2РO4, 2,5 мМ CaCl2·2H2O, 24,8 мМ NaHCO3 и 10 мМ глюкоза). Затем кольца прикрепляли к изометрическому преобразователю силы (Radnoti Inc., Monrovia, CA), данные регистрировали с использованием платформы для физиологических измерений (Gould Instrument systems, Inc., Valley View, ОН) и кольца помещали в 10 мл оксигенированного (95% О2, 5% СO2) модифицированного раствора Кребса для тканевой бани. Эти ткани доводили до натяжения в покое 1 г и давали им стабилизироваться в течение одного часа перед испытанием. Кольца испытывали добавлениями по 5 мкл 1× 10-7 М норэпинеферина (Sigma, Co., St. Louis, МО) до конечной концентрации приблизительно 1× 10-9 М и карбакола, мускаринового агониста ацетилхолина (Sigma Со.) при конечной концентрации 2× 10-7 М, для испытания целостности колец. После каждого испытания кольца промывали три раза свежим буфером, с 5 минутами между промывками, и давали им стоять в течение одного часа. Для испытания на вазодилатацию кольца сжимали до 2 г и давали стабилизироваться в течение пятнадцати минут. Затем добавляли zsig37 к 1, 2 или 3 баням, без промывки, и регистрировали натяжение на кольцах и сравнивали с контрольными кольцами. Затем эти кольца испытывали на сокращение с норэпинеферином, как описано выше. Кольца испытывали при 323, 162 и 81 нг/мл zsig37, но зависимость от дозы не смогли определить. Для оценки статистической значимости этих данных выполняли испытание на сопряженность признаков (контингенцию) на всех zsig37-кольцах и контрольных кольцах с использованием расширения в качестве детерминанта. 10 из 12 испытанных колец с zsig37 обнаружили вазодилатацию, как и 2 из 7 контролей. Точная величина Фишера Р равна 0,045. Был сделан вывод, что zsig37 индуцировал вазодилатацию в сокращенных норэпинеферином аортальных кольцах.
Пример 13
Связывание zsig37 с белками матрикса
ELISA (твердофазный иммуноферментный анализ) использовали для измерения связывания zsig37 с различными белками матрикса и C1q комплемента. Используемыми белками матрикса были бычий коллаген типа I (Becton Dickinson, Lincoln Park, NJ), ламинин, витронектин, фибронектин, коллаген человека типов II, III, IV, V, VI (Chemicon International, Temecula, CA). БСА V (SigmaCo.) использовали в качестве отрицательного контроля. Непосредственно перед использованием белки разбавляли в 2× ЗФР (забуференном фосфатом солевом растворе, Sigma, Co.) до 100 мкг/мл и доводили до рН 7,2 0,1 н NaOH. Каждую белковую пробу высевали в четырех повторностях (100 мкл на лунку) в 96-луночном планшете. Планшету давали сохнуть в течение ночи в ламинарном боксе и промывали 3 раза 400 мкл 5 мг/мл БСА в 1× ЗФР и промакали насухо. Zsig37 метили FITC в соответствии с инструкцией изготовителя (Pierce, Rockford, IL). В каждую лунку добавляли 100 мкл 1,8 мкг/мл zsig37-FITC в 5% БСА, ЗФР. Планшеты инкубировали в течение 1,5 ч при комнатной температуре, затем промывали 3 раза 5% БСА, ЗФР. Затем в каждую лунку добавляли 100 мкл смеси 1:400 мышиное антитело против ПТС/биотин (Sigma Co.). Планшет инкубировали 1,5 ч при комнатной температуре и промывали 3 раза 5%, БСА, ЗФР. Затем планшет инкубировали со 100 мкл смеси 1:1000 стрептавидин/HRP (пероксидаза хрена) (Amersham, Piscataway, NJ) в течение 1 ч и промывали 3 раза 5% БСА, ЗФР. Затем планшет проявляли с использованием Supersignal® Ultra (Pierce, Rockford, IL) в соответствии с инструкцией изготовителя. После реакции в течение 1 мин избыток жидкости удаляли из планшета переворачиванием планшета и похлопыванием по задней стороне досуха. Планшет экспонировали на рентгеновской пленке (Kodak, Rochester, New York).
Результаты этого скрининга показывают, что только фибронектин и коллагены I, II, III, V и VI связываются значимо с zsig37-FITC. Такое связывание не наблюдали с ламинином, витронектином, коллагеном IV или контролем БСА.
Пример 14
Специфичность связывания zsig37 с коллагеном типа VI
Анализ ELISA для связывания, описанный в примере 13, модифицировали для количественной оценки связывания. Zsig37-FITC, в диапазоне 0,4-4 мкг/мл, связывали с 10 мкг коллагена типа VI (Chemicon International), как описано выше. Люминесценцию из реагента Supersignal® считывали на планшет-ридере Wallac 1420 (Wallac, Gaithersburg MD) и интенсивность использовали в качестве количественной меры zsig37-FITC, связанного с планшетом ELISA.
Связывание zsig37 с коллагеном типа VI совпадает с типичной гиперболической кривой связывания (фиг.3а). Связавшийся zsig37-FITC, помещенный в планшет при 0,4 мкг/мл, мог отдавать конкурентно коллаген при добавлении немеченого zsig37 в диапазоне 0,8-8 мкг/мл (фиг.3b). Эти данные показывают, что связывание является специфическим в отношении доменов коллагена типа VI и зависимым от концентрации.
Пример 15
Связывание zsig37 c C1q комплемента
Было показано, что zsig37-FITC при 0,2 мкг/мл связывается с C1q комплемента (Sigma Co.) при 0,1-10 мкг/мл, по способу, описанному выше в примере 13 (фиг.4). Количество связывания является зависимым от концентрации и насыщаемым.
Пример 16
Ингибирование комплемента с использованием zsiq37
Анализы комплемента проводили в 96-луночных круглодонных планшетах. Желатин-вероналовый буфер, содержащий магний и кальций (141 мМ NaCl, 1,8 мМ натрий-барбитол, 3,1 мМ барбитуровая кислота, 0,1% бычий желатин, 0,5 мМ МgСl2 и 0,15 мМ CaCl2) использовали для всех разведений сыворотки и ингибитора, а также для суспензии эритроцитов. Пятьдесят микролитров стандартизованной комплементсодержащей сыворотки человека (Sigma Co.), разведенной 1/37,5 (для конечного разведения 1/150), добавляли в каждую лунку. Ингибитор добавляли в трех повторностях, 50 мкг на лунку. Сыворотку и ингибитор инкубировали в течение тридцати минут при комнатной температуре. Анализ инициировали добавлением 100 мкл 2× 108/мл несенсибилизированных овечьих эритроцитов (Colorado Serum Co., Denver, CO), сенсибилизированных овечьих эритроцитов, сенсибилизированных с использованием протокола для хемолизина изготовителя (BioWhittaker Inc., Walkersville, MD) и кроличьих эритроцитов, содержащих 16 мМ ЭГТА и 4 мМ Мg++. Ряды разведений от 1/50 до 1/400 сыворотки человека также высевали в качестве контроля активности. Эритроциты, лизированные дистиллированной водой и разведенные до 100, 75, 50, 25 и 12,5% лизиса, использовали для количественного определения процентного лизиса комплемента. Планшет герметизировали и инкубировали при 37° С в течение 1 ч с перемешиванием каждые 15 мин. Реакцию останавливали добавлением 220 мМ ЭДТА, 20 мкл на лунку и планшеты центрифугировали при 1500× g в течение 10 мин. Сто микролитров супернатанта удаляли из каждой лунки и переносили в 96-луночный плоскодонный планшет для анализа. Планшет считывали при 415 нм и рассчитывали процентный лизис.
Zsig37 был эффективным в ингибировании классического пути как с сенсибилизированными, так и с несенсибилизированными эритроцитами овец (фиг.5). Не было видимого ингибирования другого пути, испытанного с кроличьими эритроцитами и ЭГТА. Механизм ингибирования неизвестен, но, поскольку C1q связывает zsig37, наиболее вероятной мишенью является C1q.
Пример 17
Ингибирование посредством zsig37 опосредованной коллагеном активации тромбоцитов
Кровь брали из здоровых волонтеров в пробирки, содержащие цитрат натрия, держали при комнатной температуре и использовали в пределах четырех часов от взятия крови. Цельную кровь анализировали на активацию тромбоцитов с использованием Chrono-Log 560A Whole Blood Lumi-Aggregometer (люминесцентного агрегометра для цельной крови) (Chrono-Log Corp., Haverton, PA) в соответствии с инструкциями изготовителя. Для каждой точки анализа 500 мкл крови добавляли в реакционную пробирку, содержащую стержень мешалки, и 500 мкл изотонического солевого раствора, содержащего zsig37 при концентрации от 0 до 20 мкг/мл. Смесь инкубировали в течение четырех минут с последующей активацией тромбоцитов, инициируемой добавлением 5 мкл 1 мг/мл сшитого коллагена (Chrono-Log Corp.) к смеси кровь/zsig37. Ингибирование активации посредством АДФ (конечная концентрация 10 мкМ) и тромбина (конечная концентрация 1 Е/мл) определяли сходным образом.
Ингибирование опосредованной коллагеном активации тромбоцитов посредством zsig37 показывает зависимую от дозы связь между 5 и 20 мкг/мл (фиг.6а). Ингибирование является селективным в отношении коллагеновой активации и не влияет на активацию, стимулируемую АДФ или тромбином (фиг.6b). Коллагеновая активация не ингибировалась другим родственным C1q комплемента белком zsig39 (находящаяся в процессе совместного рассмотрения заявка на патент 09/140804).
Пример 18
Стимуляция роста фибробластов SK5 посредством zsig37
Фибронектин человека (25 мкг/мл, Gibco BRL, Gaithersburg, MD) помещали в 96-луночные планшеты (Costar, Pleasanton, CA) при 100 мкл на лунку и давали сохнуть в ламинарном боксе в течение ночи. Фибробласты SK5 человека в DMEM (Gibco), содержащей 10% фетальную телячью сыворотку - низкий эндотоксин (Hyclone, Logan, UT), высевали при 5000 клеток/лунку в покрытые фибронектином планшеты и инкубировали при 37° С, 5% CO2 в течение 2-3 дней. Число клеток на планшет корректировали для достижения неконфлюэнтности. Затем клетки промывали дважды бессывороточной DMEM с hi-глюкозой (Gibco) и оставляли голодать в отношении сыворотки в бессывороточной DMEM с hi-глюкозой в течение 24 ч. Zsig37 добавляли в лунки в трех повторностях при концентрациях 312,2 нг/мл - 10000 нг/мл в 100 мкл DMEM. Затем клетки инкубировали в течение 48 ч при 37° С, 5% CO2. Пролиферацию клеток тестировали добавлением 15 мкл раствора МТТ-красителя (набор CellTiter96TM, Promega) в каждую лунку. Планшет инкубировали 4 ч при 37° С, 5% CO2 и реакцию останавливали солюбилизирующим стоп-раствором (набор CellTiter96TM, Promega) в соответствии с инструкциями изготовителя. Планшет инкубировали в течение 1 ч для солюбилизации кристаллов формазана и поглощение измеряли при 570 нм со стандартом при 650 нм с использованием планшет-ридера ELISA.
Результаты (фиг.7) показывают зависимое от дозы увеличение числа фибробластов SK5 на протяжении диапазона испытанных концентраций zsig37. Эти концентрации были в диапазоне величин, обнаруживаемых для митогенных эффектов цепи b фибриногена (Gray, et al., Am. J. Respir. Cell Mol. Biol. 12, 684, 1995 и Gray, et al., J. Biol. Chem. 270, 26602, 1995) и для адгезии фибробластных клеток к помещенному в лунки C1q (Bordin and Ghebrehiwet, J. Immun. 145:2520, 1990), причем считается, что оба взаимодействуют с калретикулином клеточной поверхности.
Пример 19
Получение антисывороток против zsiq37
Кроличьи поликлональные антисыворотки получали иммунизацией двух самок Ново-Зеландских белых кроликов zsig37-CEE, очищенным из клеток ВНК. Белок конъюгировали с белком-носителем гемоцианином фиссуреллы (KLH) глутаровым альдегидом. Каждый кролик получал исходную внутрибрюшинную (ip) инъекцию 200 мкг пептида в полном адъюванте Фрейнда с последующими бустерными ip инъекциями 100 мкг пептида в неполном адъюванте Фрейнда каждые три недели. Спустя семь-десять дней после введения второй бустер-инъекции проводили кровеизвлечение у животных и собирали сыворотку. Затем животным продолжали вводить бустер-инъекции и извлечение крови осуществляли каждые три недели.
Пример 20
Детектирование связывания FIТС-меченого белка zsig37 в тканях
Связывание FIТС-меченого белка zsig37 в тканях детектировали следующим образом:
Залитые в парафин и приготовленные в виде срезов ткани человека или мышиные эмбрионы на предметных стеклах получали или из коммерческих источников (т.е. DAKO Corporation, Carpinteria, CA; BioGenex, San Ramon, CA; Novagen, Madison, Wl и Biomeda, Foster City, CA), или в лаборатории. Срезы тканей человека включали в себя надпочечник, мозг, сердце, тонкую кишку, толстую кишку, почку, печень, легкое, яичник, поджелудочную железу, предстательную железу, селезенку, желудок, яичко, щитовидную железу и матку. Срезы мышиного эмбриона производили на стадии 16 дней.
Срезы тканей освобождали, депарафинировали с использованием стандартных условий 3× 5 мин в ксилоле, 4 мин в 100% этаноле (ЕtOН), 3 мин в 100% ЕtOН и 2 мин в 95% ЕtOН. Затем срезы тканей подвергали 20-минутному процессу возврата антигена при 94° С в соответствии с инструкциями изготовителя (DAKO Corporation), с последующим 20-минутным расщеплением смесью 0,01% пепсин/0,2 н HCl. Срезы тканей промывали дважды в dH2O и один раз в буфере ЗФР/0,05% Твин 20 (Sigma, St. Louis, МО) и затем блокировали в течение 45 мин смесью 1× ЗФР/5% нежирное сухое молоко (Carnation, Los Angeles, CA). После этого следовала стадия блокирования авидин/биотин, выполняемая в соответствии с инструкциями изготовителя (Vector Laboratories, Inc., Burlingame, CA). Срезы тканей промывали 3 раза в буфере 1× ЗФР/0,05% Твин 20 и затем инкубировали с подходящей концентрацией FIТС-меченого белка zsig37 в ЗФР/5% БСА в течение 45-60 мин. После промывания этих срезов тканей 3 раза в 1× ЗФР/0,05% Твин 20 их инкубировали с разведением 1:400 конъюгата МАb анти-FIТС (mo)/биотин (Sigma) в течение 30-60 мин, промывали 3 раза в смеси ЗФР/0,05% Твин 20 и затем инкубировали в течение 30-60 мин с разведением 1:500 стрептавидин-FITC (NEN Life Science Products, Boston, MA), с последующими 2 промывками в буфере 1× ЗФР/0,05% Твин 20 и 1 промывкой в 1× ЗФР без Твина 20. Затем монтировали перпараты срезов ткани со средой против обесцвечивания, содержащей 0,5 мкг/мл иодида пропидия в качестве контрастного вещества.
Значимое связывание наблюдали со стенками сосудов и тканями типа тонких волокнистых соединительных тканей, таких как коллагеновый матрикс, в большинстве исследованных срезов тканей человека и эмбриона.
Пример 21
Zsig37 в кроличьей модели повреждения сонной артерии
Zsig37 вводили в модифицированную кроличью модель повреждения сонной артерии (Folts et al., Circulation 79:116-24, 1989 и Golino et al., Thrombosis and Haemostasis 67:302-5, 1992) для определения степени защиты, предоставляемой в предотвращении окклюзии сосуда после повреждения сосуда раздавливанием.
Тридцать четыре самца Ново-Зеландских белых кроликов в возрасте 3-6 месяцев (R&R Rabbitry, Stanwood, WA) делили на две группы. Пятнадцать кроликов получали дозы zsig37 от 2 до 13,5 мкг/кг, а 19 контрольных кроликов инъецировали ЗФР или эквивалентными количествами ЗФР или zsig39, другим родственным комплементу адипоцитов белком (WO99/10492). Кроликов анестезировали кетамином (50 мг/кг, i.m.) и поддерживали на анестезии ингаляцией галотана на протяжении этого исследования. Шерсть из ушей и с шеи сбривали и ангиокатетер помещали в маргинальную ушную вену для поддержания внутривенного ведения. Делали разрез по средней линии на шее и делали доступ к сонной артерии. Приблизительно 5 см общей сонной артерии проксимально по отношению к раздвоению (бифуркации) на внутреннюю/наружную обнажали посредством тупого отделения от окружающей ткани и любые видимые боковые ответвления каутеризировали (прижигали). Проточный зонд (Transonic Systems, Inc., Ithaca NY) помещали дистально относительно предполагаемого места повреждения и устанавливали фоновый кровоток. Затем участок сосуда 2,5-3,0 см изолировали от кровоснабжения при помощи нетравматических сосудистых зажимов. После удаления крови из этого сегмента сосуда 0,4 мл zsig37 в 0,9% хлориде натрия или 0,04 мл 0,9% хлорида натрия в качестве контроля инъецировали в этот пустой сегмент сосуда при помощи иглы 30 G. Сосуд оставляли непотревоженным в течение 5 мин перед повреждением. Затем наносили повреждение раздавливанием размером 1,0 см в центре этого сегмента сосуда при помощи сдерживаемого кровоостанавливающего зажима и оставляли в покое в течение 10 мин. Затем зажимы для сосудов удаляли и восстанавливали кровоток. Кровоток непрерывно подвергали мониторингу в течение 60 мин, после чего кроликов эвтанизировали и этот сосуд вырезали для гистологического анализа.
Не наблюдали зависимости от дозы при этих концентрациях. Метаанализ всех доз zsig37 показал значимое увеличение времени раскрытого состояния при сравнении с контролями, при непарном t-критерии (Р=0,019).
Среднее процентное время раскрытого состояния сосуда для объединенных групп животных отрицательного контроля, определенное из записей кровотока регистрирующего прибора, было 13,5% со стандартной ошибкой ±1,7%. Среднее процентное время раскрытого состояния сосуда для объединенных обработанных zsig37 групп животных, определенное из записей кровотока регистрирующих приборов, было 37,2% со стандартной ошибкой ±10,3%.
Во второй серии экспериментов флуоресцеинированный zsig37 использовали в модели поврежденной сонной артерии. Самцов Ново-Зеландских белых кроликов анестезировали, как описано выше. Посредством разреза в шее сонную артерию обнажали и приблизительно 5 см этого сосуда выделяли из окружающей последующей ткани. Кровь откачивали из выделенного сегмента и накладывали нетравматические кровоостанавливающие зажимы. Приблизительно 0,05 мл флуоресцеинированного zsig37 (концентрация 100 мкг/мл) инъецировали в этот выделенный сегмент до полного заполнения этого сосуда при помощи иглы 30 G. После периода экспонирования 5 мин сосуд повреждали и экспонирование продолжали еще в течение 110 мин, перед тем как зажимы удаляли и кровоток восстанавливали. Животных эвтанизировали, как описано выше, при 1, 10 и 60 мин повторного установления кровотока и сосуды собирали и фиксировали формалином для гистологической оценки.
Меченый zsig37 преимущественно связывался с рецепторами в среде поврежденных сосудов. Меченый zsig37 не связывался с зонами сосуда, которые не были повреждены. Время кровотока перед сбором сосудов, по-видимому, не влияло на количество zsig37, которое остается связанным с этой тканью, т.е. не было различия в количестве меченого zsig37, связанного с этими тканями за 1 мин, в сравнении с 60-минутной временной точкой сбора. Это может указывать на то, что zsig37 прочно связывается с поврежденным сосудом и не вымывается повторно установленным кровотоком.
Оценивали также действие zsig37 на динамику кровотока после сосудистого повреждения в кроличьей модели повреждения раздавливанием/стеноза подвздошной артерии. Молодых взрослых самцов Ново-Зеландских белых кроликов анестезировали, как описано выше. Посредством брюшного разреза обнажали раздвоение (бифуркацию) на аорту и подвздошную артерию и каждую подвздошную артерию освобождали от окружающих тканей и основные ответвления перевязывали наложением лигатур. Каждую подвздошную артерию снабжали инструментально ультразвуковым проточным зондом для мониторинга кровотока через этот сосуд. На основании данных кровотока, одну подвздошную артерию выбирали для использования для повреждения, а другую катетеризировали для доставки тест-пробы. Кроликов делили на группы доз с 6 кроликами на группу. Дозы тест-проб, содержащие zsig37, увеличивали в приращениях поллогарифма от 3 до 1000 мкг/кг на протяжении выбранного периода инфузии. Инфузию тест-проб начинали после создания критического стеноза, который уменьшал кровоток через этот сосуд приблизительно на 50%. После создания стеноза и периода стабилизации кровотока сосуд повреждали раздавливанием сосуда между зажимами гладкого иглодержателя. Инфузию продолжали после повреждения в течение установленного периода времени, 10-20 мин. Кровоток через поврежденный сосуд подвергали мониторингу в течение 60 мин после повреждения. Животных эвтаназировали в конце периода исследования. Нижнюю часть брюшной аорты и каждую подвздошную артерию собирали и фиксировали формалином для гистологической оценки.
Параметры кровотока, определенные из записей регистрирующего прибора, включали в себя средний кровоток после стеноза, средний кровоток после повреждения и время, когда сосуд оставался раскрытым. Эти результаты предполагают, что существует тенденция, что zsig37 стимулирует увеличенное время раскрытого состояния сосуда увеличенной дозой до 300 мг/кг на протяжении 60-минутного периода.
Пример 22
Релаксация индуцированных серотонином сокращений аортальных колец
Самцов крыс Sprague-Dawley в возрасте приблизительно 3 месяца слегка анестезировали СO2 и затем обезглавливали. Затем грудную аорту быстро извлекали и помещали в модифицированный буфер Кребса-Хенселейта (NaCl, 118,2 мМ; KCl, 4,6 мМ; CaCl2, 2,5 мМ; MgSO4, 1,2 мМ; NaHCO2, 24,8 мМ; КН2РO4, 1,2 мМ и глюкоза, 10,0 мМ). Из каждой крысы нарезали четыре кольцевых отрезка 2-3 мм аорты после выбрасывания неровного конца аорты. В некоторых экспериментах эндотелий денудировали перед нарезением отрезков аорты посредством трения просвета аорты вдоль иглы 21 G. Денудирование эндотелия проверяли добавлением аналога ацетилхолина, карбакола, перед определением зависимых от концентрации ответных реакций на zsig37. В отсутствие эндотелия карбакол не вызывает расширения сокращенных кольцевых отрезков сосуда.
Кольца фиксировали и подсоединяли к преобразователям силового смещения в оксигенированных (95% O2, 5% СO2), имеющих рубашку стеклянных банях для органов, хранящихся при 30° С в модифицированном буфере Кребса-Хенселейта, рН 7,4. Напряжение покоя устанавливали при 1 г и непрерывно повторно корректировали до 1 г на протяжении инкубационного периода 1 ч. Свежий оксигенированный модифицированный буфер Кребса-Хенселейта добавляли в бани каждые пятнадцать минут во время инкубационного периода покоя. В конце 1-часовой инкубации кольцевые отрезки сокращали добавлением 10 мкМ серотонина. После достижения максимального сокращения, приблизительно через 15-20 мин после добавления серотонина, строили кумулятивные концентрационные кривые ответа для zsig37. Zsig37 добавляли в бани на 5 мл в объемах от 5 до 150 мкг/мл, для получения конечных концентраций в диапазоне от 1 нг/мл до 40 мкг/мл. Жизнеспособность кольцевых отрезков проверяли в конце этой концентрационной реакции добавлением форсколина (2,5 мкМ или 25 мкМ) или нитроглицерина (22 мкМ).
Добавление zsig37 индуцировало зависимое от концентрации расширение сокращенных серотонином отрезков крысиной аорты с интактным эндотелием или без интактного эндотелия. Релаксацию в ответ на zsig37 впервые наблюдали при концентрации выше 100 нг/мл. Релаксацию наблюдали приблизительно при 30-60 с после добавлений каждой концентрации zsig37 в баню, и реакции релаксации выходили на плато в пределах 3-5 мин после добавления zsig37. Характер реакции релаксации на zsig37 указывает, что расширение сосудов (вазорелаксация) является событием, опосредованным рецептором-вторичным мессенджером. Кроме того, способность zsig37 вызывать расширение отрезков аорты с денудированным эндотелием указывает на то, что zsig37 действует непосредственно на клетки гладких мышц для индукции реакции вазорелаксации.
Пример 23
Идентификация клеток, экспрессирующих рецептор zsig37, при помощи гибридизации in situ
Выделяли специфические ткани человека и подвергали скринингу на экспрессию zsig37 при помощи гибридизации in situ. Различные ткани человека, приготовленные, нарезанные на срезы и подвергнутые гибридизации in situ, включали в себя аорту, сердце, лимфатический узел, плаценту, предстательную железу, слюнные железы, кожу и яичко. Ткани фиксировали в 10% забуференном формалине (Surgipath, Richmond, IL) и заливали в парапальст X-tra (Oxford Scientific, St. Louis, МО) и делали срезы при 5 мкм при помощи микротома Reichart-Jung 2050 (Leica Instrument GmbH, Nussloch, Germany). Ткани срезали при 4-8 мкм. Ткани готовили с использованием стандартного протокола (“Development of nono-isotopic in situ hydridization”, Laboratory of Experimental Pathology, National Institute of Environmental Health Sciences, Research Park Triangle, NC). Вкратце, срезы тканей депарафинизировали при помощи HistoClear (National Diagnostics, Atlanta, GA) и затем дегидратировали этанолом. Затем эти срезы расщепляли протеиназой К (50 мкг/мл) (Boehringer Mannheim, Indianapolis, IN) при 37° С в течение 2-20 мин. За этой стадией следовали ацетилирование и регидратация тканей.
Три in situ-зонда, генерированные при помощи ПЦР, конструировали против последовательности zsig37 человека. Два набора олигонуклеотидных праймеров конструировали для генерирования зондов для отдельных районов кДНК zsig37: (1) Олигонуклеотид ZC23689 (SEQ ID NO:45) и ZC23694 (SEQ ID NO:46) использовали для генерирования зонда 414 п.н. для zsig37; (2) ZC23703 (SEQ ID NO:47) и ZC23697 (SEQ ID NO:48) использовали для генерирования зонда 896 п.н. для zsig37; (3) ZC24441 (SEQ ID NO:49) и ZC24442 (SEQ ID NO:50) использовали для генерирования зонда 290 п.н. для zsig37. Антисмысловой олигонуклеотид из каждого набора также содержал рабочую последовательность для промотора РНК-полимеразы Т7 для облегчения транскрипции антисмысловых РНК-зондов из этих продуктов. ПЦР-продукты очищали при помощи ротационных колонок Qiagen (Qiagen, Inc., Chatsworth, CA) с последующими экстракцией смесью фенол-хлороформ и осаждением этанолом. Затем зонды метили дигоксигенином (Boehringer) или биотином (Boehringer) с использованием системы транскрипции in vitro (Promega Corp., Madison, WI) в соответствии с инструкциями изготовителя.
Гибридизацию in situ проводили с дигоксигенин- или биотин-меченым зондом zsig37, описанными выше. Этот зонд добавляли к предметным стеклам при концентрации 1-5 пмоль/мл на 12-16 ч при 50-60° С. Предметные стекла затем промывали в 2х SSC и 0,1х SSC при 50-55° С. Сигналы амплифицировали с использованием амплификации с тирамидным сигналом (непрямой набор для амплификации in situ TSA, NEN, Boston, MA) и визуализировали с набором Vector Red Substrat (Vector Laboratories, Burlingame, CA) в соответствии с инструкциями изготовителя. Затем предметные стекла подвергали контрастному окрашиванию гематоксилином (Vector Laboratories).
Положительные сигналы были видны в аорте, сердце, предстательной железе, слюнных железах и яичке человека. Положительно окрашивающиеся клетки были, по-видимому, эндотелиальными клетками сосудов малого диаметра в адвентициальной оболочке, окружающей аорту, мезотелиальными клетками, покрывающими эпикард, гроздевидными клетками слюнных желез и рассеянными мононуклеарными клетками, трофобластами плаценты, эпителиальными клетками предстательной железы и многослойным эпителием семенных канальцев яичка.
Пример 24
SEC-MALLS-анализ zsig37
Zsig37 содержит N-концевой коллаген-подобный домен, а также С-концевой глобулярный район, имеющие гомологию с семейством факторов некроза опухолей, и, подобно другим таким белкам, zsig37, как ожидается, должен мультимеризоваться. Очищенный zsig37, анализируемый с использованием масс-спектрометра с ловушкой ESI-ионов (Finnigan Matt, San Jose, CA), показал присутствие молекулярных частиц, приближающихся к тримерам и 9-мерам, что было неожиданным результатом при сравнении с другими гомологичными белками. Пептидное картирование zsig37 с использованием LC-MS/MS на масс-спектрометре с ловушкой ESI-ионов (Finnigan Matt) выявило, что некоторые остатки цистеина были модифицированы S-цистеинильной группой. Возможно, что модификация ключевых цистеиновых остатков в этом белке zsig37 во время ферментации со свободными цистеинами в среде препятствует правильной олигомерной ассоциации этой молекулы.
Для более глубокого понимания проводили сравнение восстановленного и невосстановленного zsig37 с использованием гель-фильтрационной колонки Biosep S-300 при 1,0 мл/мин (7,8× 3000 мм; Phenomenex, Torrance, CA) на ВЖХ HP 1050 (Hewlett Packard, Heidelberg, Germany). HP 1050 подсоединяли к детекторам светорассеяния и показателя преломления, miniDAWN и Optilab DSP (Waytt Technology, Santa Barbara, CA) для подключенного SEC-MALLS.
Один миллиграмм рекомбинантного zsig37 (1,0 мг/мл) добавляли к ТСЕР при соотношении 10:1 моль/моль ТСЕР к zsig37 и выдерживали при комнатной температуре в течение 70 мин. Шестьдесят микролитров восстановленного zsig37 инъецировали для SEC-MALLS-анализа и остаток восстановленного zsig37 диализовали в кассетах на 0,5-3,0 мл для Slide-A-Lyser, 10K MWCO (Pierce, Rockford, IL) с перемешиванием против ЗФР, рН 7,4 с тремя сменами буфера следующим образом: 1 л ЗФР при комнатной температуре в течение 4 ч, 1 л ЗФР при 4° С в течение ночи и 1 л ЗФР при комнатной температуре в течение 4 ч.
После диализа окислению давали продолжаться при 4° С. Окисление подвергали мониторингу при помощи SEC-MALLS-анализа аликвот, берущихся при трех временных точках. Т=0 ч, Т=24 ч и Т=96 ч. Величины молекулярных масс определяли с использованием LS-RI, двухдетекторного способа.
Анализ восстановленного, диализованного рекомбинантного zsig37, по-видимому, сначала указывает на образование гексамеров и 18-меров, как определено при помощи SEC-MALLS, что более согласуется с олигомерными состояниями, наблюдаемыми в гомологах. Эти формы были также активными в анализах in vitro.
Пример 25
Связывание zsig37 с моноцитами
СD14-положительные моноциты выделяли из продукта аферезиса замороженной периферической крови при помощи способа положительного отбора с гранулами Miltenyi (Miltenyi Biotec Auburn, CA). Очищенные клетки были более чем на 80% СD14-положительными при FACS-окрашивании. Клетки ресуспендировали при 1× 106 клеток/мл в RPMI+10% фетальная телячья сыворотка (ФТС) и высевали в чашки для культуры ткани 100 мм, 5 мл на чашку. Добавляли рекомбинантный человеческий интерферон γ при 100 нг/мл и клетки инкубировали при 5% CO2, 37° С в течение 48 ч.
Клетки удаляли из чашек неферментативными способами с использованием как ЭДТА, так и выскребания, концентрировали центрифугированием, ресуспендировали в буфере для FACS-окрашивания и брали аликвоты при 500000 клеток на пробирку для окрашивания. Неактивированные клетки получали проведением другого отбора на CD14 с тем же самым продуктом аферезиса после 48 ч в интерфероне γ . Клетки инкубировали в варьирующихся концентрациях биотинилированного zsig37 с последующим добавлением стрептавидина РЕ. Все блокирование “холодным” zsig37 выполняли на льду в течение 30 мин. Несвязанный белок удаляли промыванием один раз в FACS-буфере. Связывание определяли количественно при помощи прибора FACS calibur (Becton Dickinson, Lincoln Park, NJ) и выражали в виде сигнала, превышающего контроль (только вторичное антитело). Активацию моноцитов подтверждали приблизительно 1 log-увеличением экспрессии IСАМ-1 в обработанных интерфероном γ клетках.
Связывание zsig37 детектировали как в активированных, так и в неактивированных моноцитах, с увеличением связывания zsig37, наблюдаемым в обработанных интерфероном γ клетках. Связывание детектировали до 1,5 мкг/мл, самой низкой испытанной концентрации. При 15 мкг/мл связывание увеличивалось приблизительно в 4 раза в активированных клетках. Небольшое (приблизительно 10%-ное) уменьшение связывания наблюдали в активированных клетках только в том случае, когда их предобрабатывали 70-кратным избытком “холодного” zsig37. Увеличения связывания zsig37 в активированных моноцитах предполагает, что воспалительные цитокины положительно регулируют связывание моноцитами белков/рецепторов для zsig37. Потенциально это могло бы приводить к участию zsig37 в моноцитарном фагоцитозе, убивании микробов и клеточной цитотоксичности. После двух дней в этой культуре присутствуют макрофаги и zsig37 может быть связан преимущественно с этой субпопуляцией клеток. Нет доступных хороших маркеров макрофагов для определения, происходит ли это. Zsig37 связывался также с мышиной линией моноцитов-макрофагов, RAW 264.7 (АТСС No.CRL-2278), что указывает на специфичность в отношении макрофагов.
Из предыдущего описания должно быть понятно, что, хотя здесь были описаны конкретные варианты данного изобретения для целей иллюстрации, различные модификации могут быть произведены без отклонения от идеи и объема данного изобретения. Таким образом, данное изобретение не ограничивается ничем, кроме прилагаемой формулы изобретения.
Далее приведен список последовательностей.


















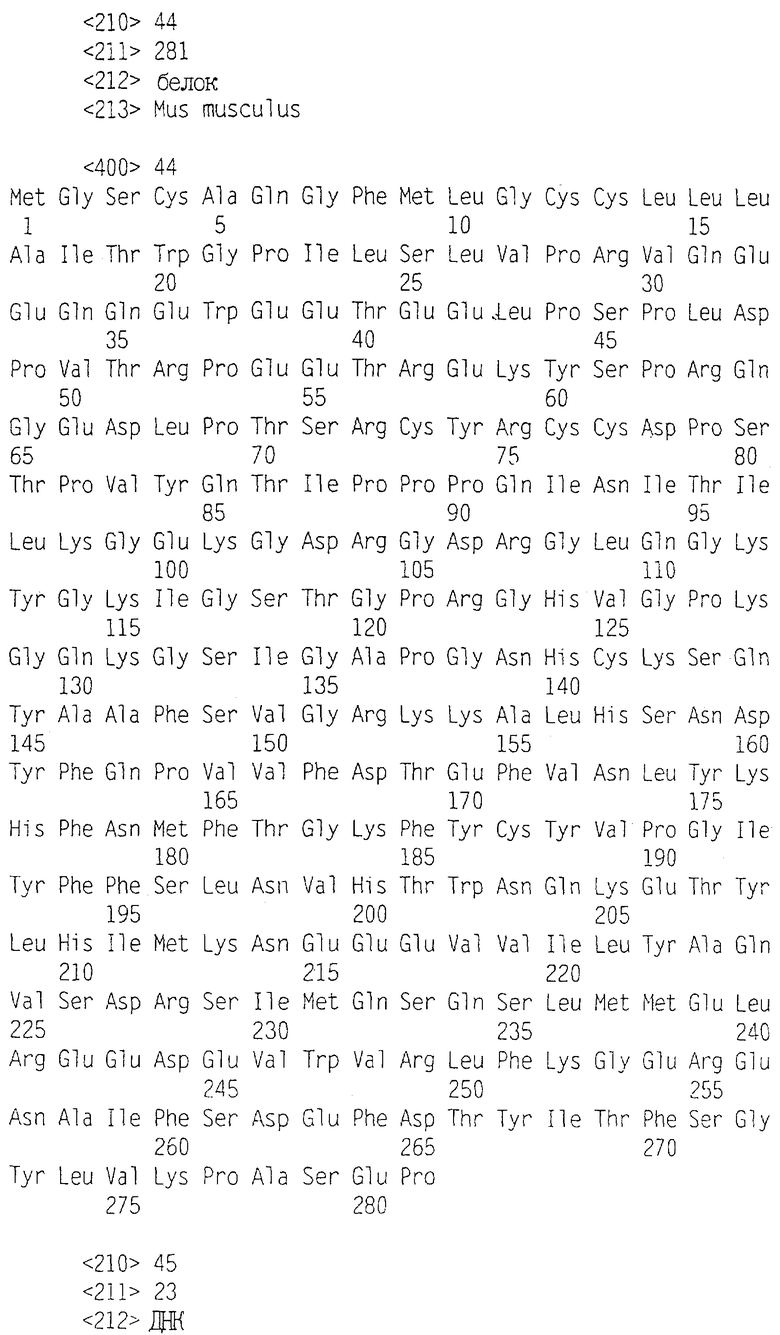


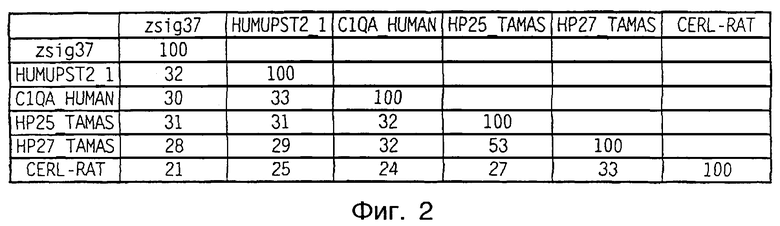
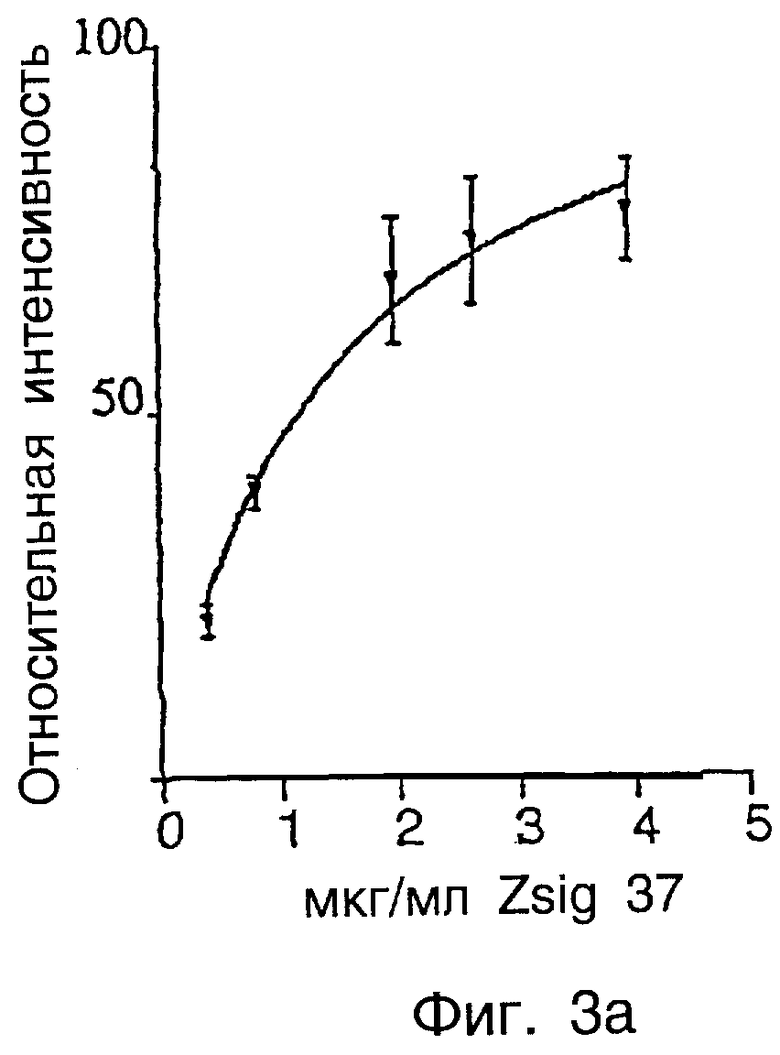

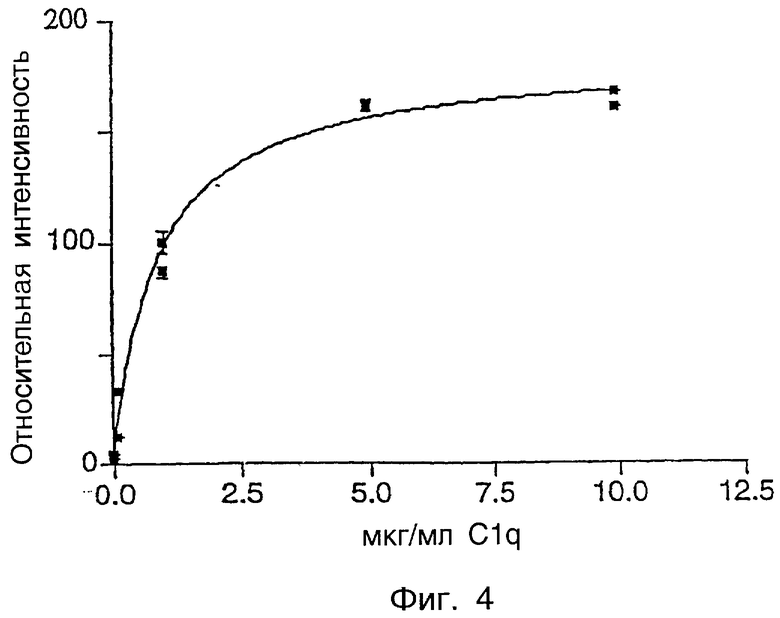
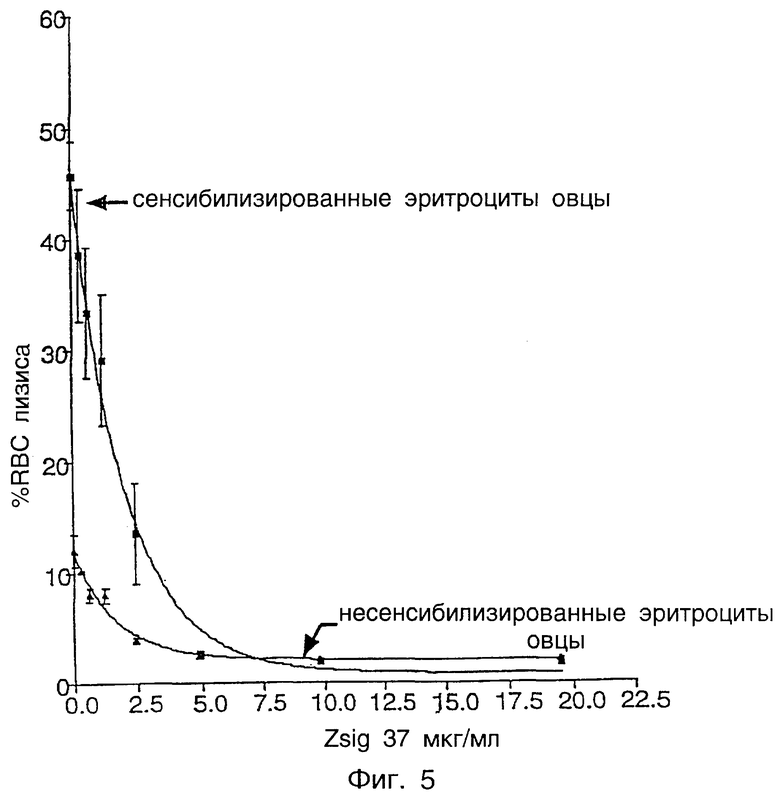
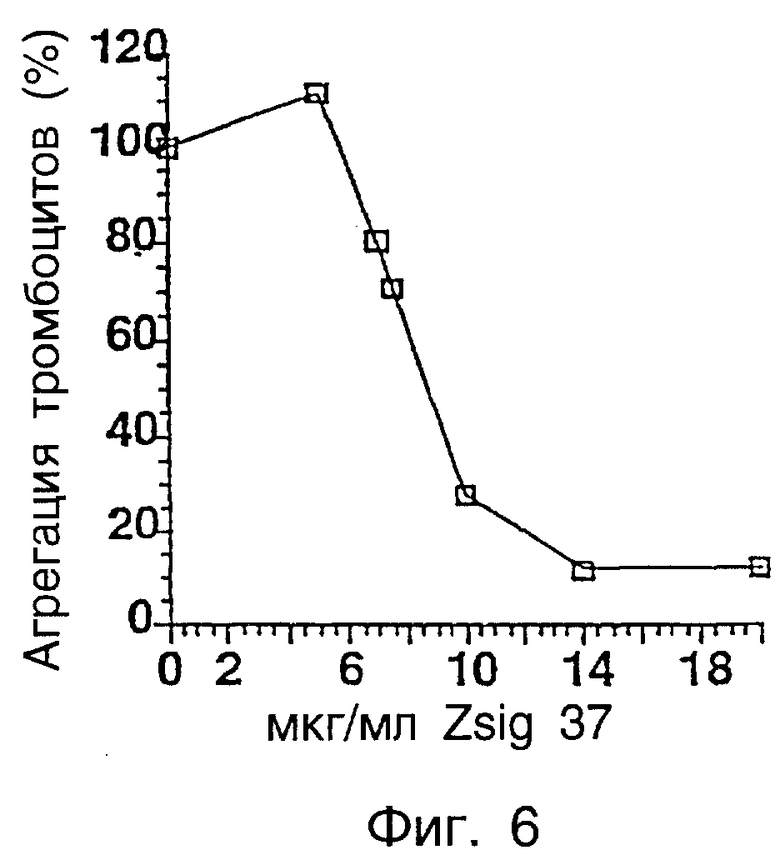
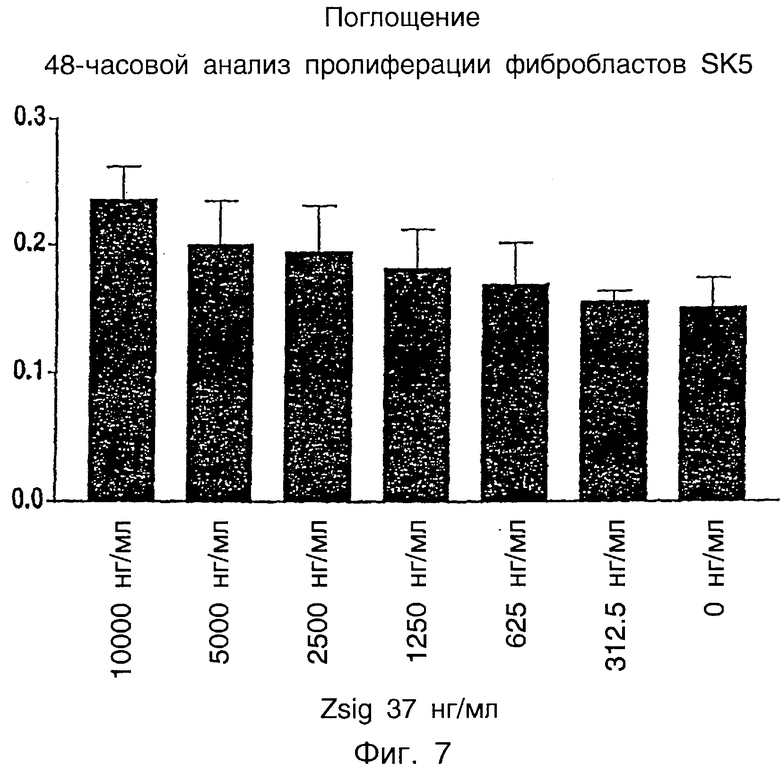
Изобретение относится к медицине и касается ингибиторов для использования при гемостазе и иммунной реакции. Такими ингибиторами являются члены семейства белков, несущих коллагеноподобный и глобулярный домены, а именно различные последовательности zsig 37. Эти ингибиторы пригодны для стимуляции кровотока в сосудистой сети путем уменьшения тромбогенной активности и активности комплемента, а также пригодны для восстановления коллагеновых поверхностей и модуляции заживления ран. Преимущество изобретения заключается в ускорении репарации поврежденных сосудов. 4 н. и 62 з.п. ф-лы, 7 ил.

| US 5869330 А, 09.02.1999.WO 99/04000А, 28.01.1999.MAEDA et al | |||
| Biochem | |||
| Bioplys | |||
| Res | |||
| Commun | |||
| Предохранительное устройство для паровых котлов, работающих на нефти | 1922 |
|
SU1996A1 |

