ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение относится к новым полипептидам, проявляющим активность гранулоцитарного колониестимулирующего фактора (G-CSF), к конъюгатам между полипептидом, проявляющим активность G-CSF, и неполипептидной частью молекулы, к способам получения таких полипептидов или конъюгатов и применению таких полипептидов или конъюгатов в терапии, в частности, для лечения лейкопении.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Процесс, посредством которого лейкоциты растут, делятся и дифференцируются в костном мозгу, называют гемопоэзом (Dexter and Spooncer, Ann. Rev. Cell. Biol., 3:423, 1987). Каждый тип клеток крови возникает из плюрипотентных стволовых клеток. Обычно имеются три класса клеток крови, продуцируемых in vivo: красные кровяные клетки (эритроциты), тромбоциты и белые кровяные клетки (лейкоциты), причем большая часть последних участвует в иммунной защите хозяина. Пролиферация и дифференцировка гемопоэтических клеток-предшественников регулируется семейством цитокинов, в том числе колониестимулирующими факторами (CSF), такими как G-CSF и интерлейкины (Arai et al., Ann. Rev. Biochem., 59:783-836, 1990). Основной биологический эффект G-CSF in vivo состоит в стимуляции роста и развития определенных лейкоцитов, известных как нейтрофильные гранулоциты или нейтрофилы (Welte et al., PNAS-USA 82:1526-1530, 1985, Souza et al., Science, 232:61-65, 1986). При высвобождении в кровоток функцией нейтрофильных гранулоцитов является борьба с бактериальной инфекцией.
Аминокислотная последовательность G-CSF человека (hG-CSF) сообщалась Nagata et al., Nature 319:415-418, 1986). hG-CSF является мономерным белком, который димеризует G-CSF-рецептор образованием комплекса 2:2 из 2 молекул G-CSF и 2 рецепторов (Horan et al. (1996), Biochemistry 35(15): 4886-96). Aritomi et al., Nature 401:713-717, 1999 описали рентгеновскую (кристаллическую) структуру комплекса между hG-CSF и доменами BN-BC G-CSF-рецептора. Они идентифицируют следующие остатки hG-CSF как часть пограничных слоев связывания рецептора: G4, Р5, А6, S7, S8, L9, Р10, Q11, S12, L15, К16, Е19, Q20, L108, D109, D112, Т115, Т116, Q119, Е122, Е123 и L124. Сообщалась экспрессия rhG-CSF в Escherichia coli, Saccharomyces cerevisiae и клетках млекопитающих (Souza et al., Science 232:61-65, 1986, Nagata et al., Nature 319: 415-418, 1986, Robinson and Wittrup, Biotechnol. Prog. 11:171-177, 1985).
Рекомбинантный G-CSF человека (rhG-CSF) обычно используют для лечения различных форм лейкопении. Так, коммерческие препараты rhG-CSF доступны под названиями филграстим (Gran® и Neupogen®, ленограстим (Neutrogin® и Granocyte®) и нартограстим (Neu-up®). Gran® и Neupogen® являются негликозилированными и продуцируются в рекомбинантных клетках Е. coli. Neutrogin® и Granocyte® являются гликозилированными и продуцируются в рекомбинантных клетках СНО, а Neu-up® является негликозилированным, причем пять аминокислот заменены при N-концевом районе интактного rhG-CSF, продуцируемого в рекомбинантных клетках Е. coli.
Сообщались несколько вариантов hG-CSF со сконструированным белком (Патенты США US 5581476, US 5214132, US 5362853, US 4904584 и Riedhaar-Olson et al. Biochemistry 35: 9034-9041, 1996). Была предложена модификация hG-CSF и других полипептидов таким образом, чтобы ввести, по меньшей мере, одну дополнительную углеводную цепь в сравнении с нативным полипептидом (US 5218092). Утверждается, что аминокислотная последовательность этого полипептида может быть модифицирована заменой аминокислоты, делецией аминокислоты или инсерцией аминокислоты таким образом, чтобы осуществить добавление дополнительной углеводной цепи. Кроме того, сообщались полимерные модификации нативного hG-CSF, включающие в себя присоединение ПЭГ-групп (Satake-Ishikawa et al., Cell Structure and Function 17:157-160, 1992, US 5824778, US 5824784, WO 96/11953, WO 95/21629, WO 94/20069).
Bowen et al., Experimental Hematology 27 (1999), 425-432 описывают исследование взаимосвязи между массой молекулы и продолжительностью активности ПЭГ-конъюгированного мутеина G-CSF. Явная обратная корреляция предполагалась между молекулярной массой ПЭГ-частей молекулы, конъюгированных с этим белком, и активностью in vitro, тогда как активности in vivo увеличивались с увеличением молекулярной массы. Предполагается, что более низкая аффинность этих конъюгатов вызывает увеличение периода полувыведения, поскольку опосредованный рецептором эндоцитоз является важным механизмом, регулирующим уровни гемопоэтических факторов роста.
Коммерчески доступный rhG-CSF имеет краткосрочное фармакологическое действие и часто должен вводиться более одного раза в день во время протекания лейкопенического состояния. Молекула с более продолжительным полупериодом существования в кровотоке могла бы уменьшить число введений, необходимое для ослабления лейкопении, и предотвращать последующие инфекции. Другой проблемой доступных в настоящее время продуктов rG-CSF является проявление зависимых от дозы болей в костях. Поскольку боль в костях воспринимается пациентами как существенное побочное действие лечения с использованием rG-CSF, было бы желательным обеспечение продукта rG-CSF, который не вызывает боли в костях, посредством продукта, который либо не имеет этого действия, либо является эффективным в достаточно малой дозе, которая не вызывает боли в костях. Таким образом, существует явная необходимость в улучшенных рекомбинантных G-CSF-подобных молекулах.
Что касается полупериода существования, одним из путей увеличения полупериода существования белка в кровотоке является обеспечение снижения клиренса этого белка, в частности, посредством почечного клиренса и опосредованного рецептором клиренса. Это может быть достигнуто конъюгацией данного белка с химической частью молекулы, которая способна увеличивать кажущийся размер, уменьшая тем самым почечный клиренс и увеличивая полупериод существования in vivo. Кроме того, присоединение химической части к белку может эффективно блокировать протеолитические ферменты от физического контакта с белком, предотвращая таким образом деградацию посредством неспецифического протеолиза. Полиэтиленгликоль (ПЭГ) является одной такой химической частью молекулы, которую использовали в получении терапевтических белковых продуктов. Молекула G-CSF, модифицированная единственной N-терминально связанной группой ПЭГ, названная SD/01, подвергается в настоящее время клиническому испытанию. Хотя было показано, что эта ПЭГилированная молекула G-CSF имеет увеличенный полупериод существования в сравнении с не-ПЭГилированным G-CSF, она имеет низкую активность и, следовательно, должна вводиться в довольно высоких дозах. В результате проблема боли в костях, обсуждаемая выше, не была решена с использованием этой молекулы. Кроме того, все еще имеется существенное поле действия для улучшения известных молекул G-CSF в отношении биологического полупериода существования.
Таким образом, все еще существует потребность в обеспечении новых молекул, проявляющих активность G-CSF, которые применимы в лечении лейкопении и которые являются улучшенными в отношении увеличенного полупериода существования и уменьшения побочных действий. Данное изобретение относится к таким молекулам.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Более конкретно, данное изобретение относится к специфическим конъюгатам, содержащим полипептид, проявляющий активность G-CSF, и неполипептидную часть молекулы, способам их получения и их применению в лекарственном лечении и в приготовлении фармацевтических средств. Таким образом, в первом аспекте данное изобретение относится к различным специфическим конъюгатам, содержащим полипептид, проявляющий активность G-CSF и имеющий аминокислотную последовательность, которая отличается от известной аминокислотной последовательности G-CSF человека, показанной в SEQ ID NO:1, по меньшей мере, в одном указанном аминокислотном остатке, содержащем группу присоединения для неполипептидной части молекулы, и имеющим, по меньшей мере, одну неполипептидную часть молекулы, присоединенную к группе присоединения полипептида. Конъюгат данного изобретения имеет одно или несколько улучшенных свойств в сравнении с коммерчески доступным rhG-CSF, в том числе увеличенный функциональный полупериод существования in vivo, увеличенный полупериод существования в сыворотке, уменьшенные побочные действия, уменьшенную иммуногенность и/или увеличенную биодоступность. Следовательно, медицинское лечение конъюгатом данного изобретения предоставляет ряд преимуществ над доступными в настоящее время соединениями G-CSP.
В следующем аспекте данное изобретение относится к полипептидам, проявляющим активность G-CSF и образующим часть конъюгата данного изобретения. Предполагается, что полипептиды данного изобретения применимы как таковые для терапевтических, диагностических или других целей, но особый интерес представляют в качестве промежуточных продуктов для получения конъюгата данного изобретения.
В следующем аспекте данное изобретение относится к конъюгату полипептида, содержащему полипептид, проявляющий активность G-CSF, который имеет аминокислотную последовательность, отличающуюся от аминокислотной последовательности hG-CSF (аминокислотная последовательность показана в SEQ ID NO:1) в, по меньшей мере, одном аминокислотном остатке, выбранном из введенного или удаленного аминокислотного остатка, содержащем группу присоединения для неполипептидной части молекулы, и достаточные число или тип неполипептидной части молекулы для обеспечения конъюгата с увеличенным полупериодом существования в сравнении с известными рекомбинантными продуктами G-CSF.
В следующих аспектах данное изобретение относится к способам получения конъюгата данного изобретения, в том числе нуклеотидным последовательностям, кодирующим полипептид данного изобретения, экспрессирующим векторам, содержащим такую нуклеотидную последовательность, и клеткам-хозяевам, содержащим также нуклеотидную последовательность или экспрессирующий вектор.
В конечных аспектах данное изобретение относится к композиции, содержащей конъюгат или полипептид данного изобретения, способу приготовления фармацевтической композиции, применению конъюгата или композиции данного изобретения в качестве фармацевтического средства и способу лечения млекопитающего такой композицией. В частности, полипептид, конъюгат или композиция данного изобретения могут быть использованы для предотвращения инфекции у раковых пациентов, подвергающихся некоторым типам лучевой терапии, химиотерапии и трансплантациям костного мозга, для мобилизации клеток-предшественников для сбора в трансплантациях клеток-предшественников периферической крови, для лечения тяжелой хронической или относительной лейкопении, независимо от причины, и для поддерживающей терапии пациентов с острым миелоидным лейкозом. Кроме того, полипептид, конъюгат или композиция данного изобретения могут быть использованы для лечения СПИДа или других иммунодефицитных заболеваний, а также бактериальных инфекций.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В контексте данной заявки и изобретения применяются следующие определения:
Термин "конъюгат" предназначен для обозначения гетерогенной молекулы, образованной ковалентным присоединением одного или нескольких полипептидов, обычно единственного полипептида, к одной или нескольким неполипептидным частям молекулы, таким как полимерные молекулы, липофильные соединения, углеводные части молекулы или органические дериватизующие агенты. Термин ковалентное присоединение обозначает, что полипептид и неполипептидная часть молекулы непосредственно ковалентно соединены друг с другом или альтернативно опосредованно ковалентно присоединены друг к другу через промежуточные часть или части, такие как мостик, спейсер, или связывающие часть или части молекулы. Предпочтительно, конъюгат является растворимым при уместных концентрациях и условиях, т.е. растворимым в физиологических жидкостях, таких как кровь.
Термин "неконъюгированный полипептид" может быть использован в отношении полипептидной части конъюгата.
Термин "полипептид" может быть использован взаимозаменяемо с термином "белок".
"Полимерная молекула" является молекулой, образованной ковалентной связью двух или нескольких мономеров, где ни один из мономеров не является аминокислотным остатком, за исключением случая, когда полимер является человеческим альбумином или другим обильным белком плазмы. Термин "полимер" может быть использован взаимозаменяемо с термином "полимерная молекула". Термин предназначен для обозначения углеводных молекул, хотя, обычно, этот термин не предназначен для обозначения типа углеводной молекулы, которая присоединена к полипептиду N- или O-гликозилированием (как дополнительно описано ниже), так как такую молекулу называют здесь "олигосахаридной частью молекулы". За исключением случаев, когда число полимерных молекул особо указывается, каждая ссылка на "полимер", "полимерную молекулу", "данный полимер" или "данную полимерную молекулу", содержащиеся в полипептиде данного изобретения, или ссылка, используемая иначе в данном изобретении, должна быть ссылкой на одну или несколько полимерных молекул.
Термин "группа присоединения" обозначает группу аминокислотного остатка полипептида, способную связываться с соответствующей неполипептидной частью молекулы. Например, для конъюгации полимера, в частности, с ПЭГ часто используемой группой присоединения является ε-аминогруппа лизина или N-концевая аминогруппа. Другие группы присоединения полимера включают в себя свободную карбоксильную группу (например, группу С-концевого аминокислотного остатка или остатка аспарагиновой кислоты или глутаминовой кислоты), подходящим образом активированные карбонильные группы, окисленные углеводные части молекулы и меркаптогруппы. Применимые группы присоединения и совместимые с ними непептидные части можно видеть из таблицы ниже.
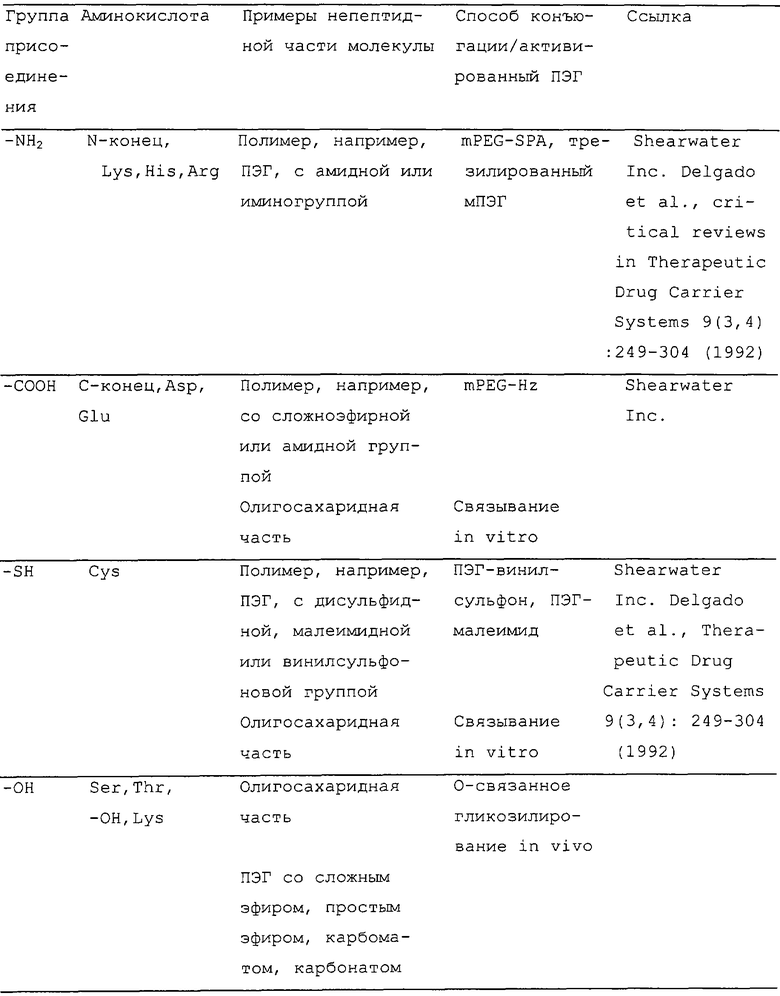

Для N-гликозилирования in vivo термин "группа присоединения" используют необычным путем для указания аминокислотных остатков, составляющих сайт N-гликозилирования (с последовательностью N-X'-S/T/C-X'', где X' обозначает любой аминокислотный остаток, кроме пролина, X'' обозначает любой аминокислотный остаток, который может быть или может не быть идентичным X' и который предпочтительно является отличающимся от пролина, N обозначает аспарагин и S/T/C обозначает либо серин, треонин, либо цистеин, предпочтительно серин или треонин и наиболее предпочтительно треонин). Хотя остаток аспарагина сайта N-гликозилирования находится там, где олигосахаридная часть присоединяется во время гликозилирования, такое присоединение не может быть достигнуто, если не присутствуют другие аминокислотные остатки сайта N-гликозилирования. Таким образом, когда непептидная часть молекулы является олигосахаридной частью и конъюгация должна достигаться N-гликозилированием, термин "аминокислотный остаток, включающий в себя группу присоединения для непептидной части молекулы", в применении в связи с изменениями аминокислотной последовательности представляющего интерес полипептида, должен пониматься в значении, что один или несколько аминокислотных остатков, составляющих сайт N-гликозилирования, должны быть изменены таким образом, что функциональный сайт N-гликозилирования вводят в эту аминокислотную последовательность или его удаляют из указанной последовательности.
В данной заявке названия аминокислот и названия атомов (например, СА, СВ, NZ, N, О, С и т.д.) используются, как определено Банком данных белков Protein DataBank (PDB) (www.pdb.org), который основывается на номенклатуре IUPAC (IUPAC Nomenclature and Symbolism for Amino Acids and Peptides (названия остатков, названия атомов и т.д.), Eur. J. Biochem., 138, 9-37 (1984) вместе с их поправками в Eur. J. Biochem., 152, 1 (1985). Термин "аминокислотный остаток" прежде всего предназначен для указания аминокислотного остатка, содержащегося в группе, состоящей из 20 природно встречающихся аминокислот, т.е. остатков аланина (Ala или А), цистеина (Cys или С), аспарагиновой кислоты (Asp или D), глутаминовой кислоты (Glu или Е), фенилаланина (Phe или F), глицина (Gly или G), гистидина (His или Н), изолейцина (Ile или I), лизина (Lys или К), лейцина (Leu или L), метионина (Met или М), аспарагина (Asn или N), пролина (Pro или Р), глутамина (Gln или Q), аргинина (Arg или R), серина (Ser или S), треонина (Thr или Т), валина (Val или V), триптофана (Trp или W) и тирозина (Tyr или Y).
Терминология, используемая для идентификации аминокислотных положений/замен, иллюстрируется следующим образом; F13 указывает номер положения 13, занимаемого остатком фенилаланина в ссылочной аминокислотной последовательности. F13K указывает на то, что остаток фенилаланина положения 13 был заменен остатком лизина. Если нет других указаний, нумерация аминокислотных остатков, используемая здесь, указывается относительно аминокислотной последовательности hG-CSF, показанной в SEQ ID NO:1. Альтернативные замены указываются «/», например Q67D/E означает аминокислотную последовательность, в которой глутамин в положении 67 заменен либо аспарагиновой кислотой, либо глутаминовой кислотой. Множественные замены указываются «+», например S53N+G55S/T означает аминокислотную последовательность, которая содержит замену остатка серин в положении 53 остатком аспарагина и замену остатка глицина в положении 55 остатком серина или треонина.
Термин "нуклеотидная последовательность" обозначает последовательный отрезок двух или нескольких нуклеотидных молекул. Нуклеотидная молекула может быть геномной, кДНК, РНК, полусинтетической или синтетической или любой их комбинацией.
Термин "полимеразная цепная реакция" или "ПЦР" обычно обозначает способ амплификации желаемой нуклеотидной последовательности in vitro, как описано, например, в патенте США US 4683195. Обычно способ ПЦР включает в себя повторяемые циклы синтеза с удлинением праймеров, с использованием олигонуклеотидных праймеров, способных гибридизоваться преимущественно с матричной нуклеиновой кислотой.
"Клетка", "клетка-хозяин", "клеточная линия" и "культура клеток" используются здесь взаимозаменяемо, и все подобные термины, как должно быть понятно, включают в себя потомство, происходящее из роста или культивирования клетки. Термины "трансформация" и "трансфекция" используют взаимозаменяемо для обозначения введения ДНК в клетку.
"Функционально (оперативно) связанный" обозначает ковалентное соединение двух или нескольких нуклеотидных последовательностей посредством ферментативного лигирования или иным образом, в такой конфигурации относительно друг друга, что может выполняться нормальная функция этих последовательностей. Например, нуклеотидная последовательность, кодирующая пре-последовательность или секреторный лидер, функционально связана с нуклеотидной последовательностью для полипептида, если он экспрессируется в виде пре-белка, который участвует в секреции полипептида; промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию этой последовательности; сайт связывания рибосомы функционально связан с кодирующей последовательностью, если он расположен таким образом, чтобы облегчать трансляцию. В общем "функционально связанный" обозначает, что нуклеотидные связываемые последовательности являются смежными и в случае секреторного лидера - смежными в рамке считывания. Связывание выполняют лигированием в удобных сайтах рестрикции. Если такие сайты не существуют, то используют синтетические олигонуклеотидные адаптеры или линкеры вместе со стандартными способами рекомбинантных ДНК.
Термин "вводить" относится к введению аминокислотного остатка, содержащего группу присоединения для неполипептидной части молекулы, в частности, посредством замены существующего аминокислотного остатка или альтернативно инсертированием дополнительного аминокислотного остатка. Термин "удалять" относится к удалению аминокислотного остатка, содержащего группу присоединения для неполипептидной части молекулы, в частности, заменой удаляемого аминокислотного остатка другим аминокислотным остатком или альтернативно делецией (без замены) удаляемого аминокислотного остатка.
Когда замены выполняют в отношении исходного полипептида, они предпочтительно являются "консервативными заменами", другими словами, замены выполняют в группах аминокислот со сходными характеристиками, например небольших аминокислот, кислых аминокислот, полярных аминокислот, основных аминокислот, гидрофобных аминокислот и ароматических аминокислот.
Предпочтительные замены в данном изобретении могут быть, в частности, выбраны из групп консервативных замен, приведенных в таблице ниже.
Термин "иммуногенность", в применении в связи с конкретным веществом, обозначает способность этого вещества индуцировать ответную реакцию иммунной системы. Иммунная реакция может быть клеточно-опосредованной или опосредованной антителами реакцией (см., например, Roitt: Essential Immunology (8th) Edition, Blackwell в отношении дополнительного определения иммуногенности). Обычно, пониженная реактивность антител будет указанием на пониженную иммуногенность. Пониженная иммуногенность может быть определена с использованием любого подходящего способа, известного в данной области, например in vivo или in vitro.
Термин "функциональный полупериод существования in vivo" используется в его обычном значении, т.е. как время, при котором 50% биологической активности полипептида или конъюгата все еще присутствуют в теле/органе-мишени, или как время, при котором активность полипептида или конъюгата составляет 50% первоначальной величины. В качестве альтернативы определению функционального полупериода существования in vivo может определяться "сывороточный полупериод существования", т.е. время, в течение которого 50% молекул полипептида или конъюгата циркулируют в плазме или кровотоке, перед клиренсом. Альтернативные термины для сывороточного полупериода существования включают в себя "полупериод существования в плазме", "полупериод существования в кровотоке", "сывороточный клиренс", "плазменный клиренс" и "полупериод клиренса". Полипептид или конъюгат удаляются посредством действия одной или нескольких ретикулоэндотелиальных систем (RES), почки, селезенки или печени, опосредованной рецепторами деградации или специфическим или неспецифическим протеолизом, в частности действием опосредованного рецепторами клиренса и почечного клиренса. Обычно клиренс зависит от размера (относительно отсечения для гломерулярной фильтрации), заряда, присоединенных углеводных цепей и присутствия клеточных рецепторов для конкретного белка. Функциональность, которая должна сохраняться, обычно выбирают из пролиферативной или рецепторсвязывающей активности. Функциональный полупериод существования in vivo и полупериод существования в сыворотке могут быть определены любым подходящим способом, известным в данной области, как дополнительно обсуждается в разделе Материалы и Способы ниже.
Термин "увеличенный" в применении относительно функционального полупериода существования in vivo или полупериода существования в сыворотке используется для указания на то, что соответствующий полупериод существования конъюгата или полипептида является статистически значимо увеличенным относительно полупериода существования ссылочной молекулы, такой как неконъюгированный hG-CSF (например, Neupogen®), при определении в сравнимых условиях. Например, соответствующий полупериод существования может увеличиваться, по меньшей мере, на приблизительно 25%, например, по меньшей мере, на приблизительно 50%, например, по меньшей мере, на приблизительно 100%, 200%, 500% или 1000%.
Термин "почечный клиренс" используется в его обычном значении для указания любого клиренса, выполняемого почками, например гломерулярной фильтрации, канальцевой экскреции или канальцевой элиминации. Почечный клиренс зависит от физических характеристик конъюгата, в том числе размера (диаметра), симметрии, формы/жесткости и заряда. Пониженный почечный клиренс может быть установлен любым подходящим анализом, например, установленным анализом in vivo. Обычно почечный клиренс определяют введением меченого (например, радиоактивно или флуоресцентно меченного) конъюгата полипептида пациенту и измерением активности метки в моче, взятой у этого пациента. Пониженный почечный клиренс определяют относительно соответствующего ссылочного полипептида, например соответствующего неконъюгированного полипептида, неконъюгированного соответствующего полипептида дикого типа или другого конъюгированного полипептида (такого как конъюгированный полипептид, не относящийся к данному изобретению) в сравнимых условиях. Предпочтительно, скорость почечного клиренса конъюгата уменьшается на приблизительно 50%, предпочтительно, по меньшей мере, на 75%, наиболее предпочтительно на, по меньшей мере, 90% в сравнении с соответствующим ссылочным полипептидом.
Обычно активация рецептора сопряжена с опосредованным рецептором клиренсом (RMC), так что связывание полипептида с его рецептором без активации не ведет к RMC, тогда как активация рецептора приводит к RMC. Этот клиренс обусловлен интернализацией связанного рецептором полипептида с последующей деградацией лизосомами. Пониженный RMC может достигаться конструированием конъюгата таким образом, чтобы можно было связывать и активировать достаточное число рецепторов для получения оптимальной биологической реакции in vivo и избежать активации большего числа рецепторов, чем это необходимо для получения такой реакции. Это может отражаться в снижении биоактивности in vitro и/или увеличенной скорости отсоединения полипептидного конъюгата от его рецептора.
Обычно пониженная биоактивность in vitro отражает пониженную действенность/эффективность и/или пониженную силу и может быть определена любым подходящим способом определения любого из этих свойств. Например, биоактивность in vitro может быть определена в анализе на основе люциферазы (см. раздел Материалы и Способы). Биоактивность in vitro конъюгата может быть понижена, по меньшей мере, на 30%, например, по меньшей мере, на 50%, 60% или 75%, например, по меньшей мере, на 90%, в сравнении с биоактивностью in vitro соответствующего ссылочного полипептида, определенной при сравнимых условиях. Другими словами, конъюгат может иметь биоактивность in vitro, которая является такой низкой, как 1%, обычно, по меньшей мере, приблизительно 2%, например, по меньшей мере, приблизительно 5% биоактивности соответствующего неконъюгированного полипептида или соответствующего полипептида дикого типа. Например, биоактивность in vitro может быть в диапазоне приблизительно 1-50% биоактивности ссылочного полипептида, например, около 2-40%, например, около 5-25%, при определении в сравнимых условиях. В случаях, когда уменьшенная биоактивность in vitro является желательной для уменьшения опосредованного рецептором клиренса, будет ясно, что достаточная биоактивность для получения желаемой активации рецептора должна быть тем не менее сохранена.
Предпочтительно, увеличенная скорость отсоединения полипептидного конъюгата от его рецептора является величиной, приводящей к высвобождению полипептидного конъюгата из его рецептора, до того как какая-либо существенная интернализация комплекса рецептор-лиганд имела место. Аффинность связывания рецептор-полипептид (в том числе скорость отсоединения) может быть определена, как описано здесь в разделе Материалы и Способы. Опосредованный рецептором клиренс (RMC) in vitro может быть определен мечением (например, радиоактивным или флуоресцентным мечением) полипептидного конъюгата, стимуляцией клеток, содержащих рецептор для этого полипептида, промыванием клеток и измерением активности метки. Альтернативно, конъюгат может быть подвергнут действию клеток, экспрессирующих соответствующий рецептор. После подходящего времени инкубации супернатант удаляют и переносят в лунку, содержащую подобные клетки. Биологический ответ этих клеток на супернатант определяют относительно неконъюгированного полипептида или другого ссылочного полипептида, и это является мерой степени уменьшенного RMC.
Обычно уменьшенную биоактивность конъюгата in vitro получают как следствие его модификации неполипептидной частью молекулы. Однако для дополнительного уменьшения биоактивности in vitro или для других причин может представлять интерес модифицировать дополнительно полипептидную часть конъюгата. Например, в одном варианте, по меньшей мере, один аминокислотный остаток, расположенный при сайте связывания рецептора полипептида или около него, может быть заменен другим аминокислотным остатком в сравнении с соответствующим полипептидом дикого типа, чтобы получить уменьшенную биоактивность in vitro. Этот аминокислотный остаток, который должен быть введен заменой, может быть любым аминокислотным остатком, способным уменьшать биоактивность конъюгата in vitro. Удобно, чтобы вводимый аминокислотный остаток содержал группу присоединения для неполипептидной части молекулы, определенной здесь. В частности, когда неполипептидная часть молекулы является полимерной молекулой, такой как ПЭГ, вводимый аминокислотный остаток может быть остатком лизина.
Термин "проявляющий активность G-CSF" указывает на то, что полипептид или конъюгат имеет одну или несколько функций нативного G-CSF, в частности hG-CSF с аминокислотной последовательностью, показанной в SEQ ID NO:1, в том числе способность связываться с рецептором G-CSF (Fukunaga et al., J. Biol. Chem, 265:14008, 1990). Активность G-CSF удобно анализировать с использованием первичного анализа, описанного далее в разделе Материалы и Способы. Считают, что полипептид, "проявляющий" активность G-CSF, имеет такую активность, когда он проявляет измеримую функцию, например измеряемую пролиферативную активность или рецепторсвязывающую активность (например, определяемую первичным анализом, описанным в разделе Материалы и Способы). Полипептид, проявляющий активность G-CSF, может также называться здесь "молекулой G-CSF".
Термин "исходный G-CSF" или "исходный полипептид" обозначает молекулу, подлежащую модификации в соответствии с данным изобретением. Исходный G-CSF обычно является hG-CSF или его вариантом. "Вариант" представляет собой полипептид, который отличается в одном или нескольких аминокислотных остатках от исходного полипептида, обычно в 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 аминокислотных остатках. Примеры rhG-CSF включают в себя филграстим (Gran® и Neupogen®), ленограстим (Neutrogin® и Granocyte®) и нартограстим (Neu-up®).
Конъюгат изобретения
Как указывалось выше, в первом аспекте данное изобретение относится к конъюгату, содержащему полипептид, проявляющий активность G-CSF, который содержит аминокислотную последовательность, которая отличается от аминокислотной последовательности SEQ ID NO:1 в, по меньшей мере, одном аминокислотном остатке, выбранном из указанных введенных или удаленных аминокислотных остатков, содержащем группу присоединения для неполипептидной части молекулы, и, по меньшей мере, одну неполипептидную часть молекулы, присоединенную к группе присоединения этого полипептида. Вводимые и/или удаляемые аминокислотные остатки описаны более подробно в следующих разделах. Должно быть понятно, что сам конъюгат также проявляет активность G-CSF.
Удалением и/или введением аминокислотного остатка, содержащего группу присоединения для неполипептидной части молекулы, можно специфически приспособить этот полипептид таким образом, чтобы сделать эту молекулу более восприимчивой к конъюгации с выбранной неполипептидной частью молекулы, для оптимизации характера конъюгации (например, обеспечения оптимального распределения неполипептидных частей молекулы на поверхности молекулы G-CSF и для обеспечения присутствия в этой молекуле только групп присоединения, предназначенных для конъюгации, и получения тем самым новой молекулы конъюгата, которая имеет активность G-CSF и, кроме того, одно или несколько улучшенных свойств, в сравнении с доступными в настоящее время молекулами G-CSF).
Хотя этот полипептид может быть любого происхождения, в частности, происходить из млекопитающего, в настоящее время он является предпочтительно полипептидом, происходящим из человека.
В предпочтительном варианте данного изобретения более чем один аминокислотный остаток полипептида с активностью G-CSF является измененным, например изменение включает в себя удаление, а также введение аминокислотных остатков, содержащих группу присоединения для выбранной неполипептидной части молекулы.
Должно быть понятно, что, кроме описанных здесь изменений аминокислот, нацеленных на удаление и/или введение сайтов присоединения для неполипептидной части молекулы, аминокислотная последовательность полипептида данного изобретения может, если желательно, содержать другие изменения, которые не должны быть обязательно связанными с введением или удалением сайтов присоединения, т.е. другие замены, инсерции или делеции. Они могут, например, включать в себя укорочение N- и/или С-конца на один или несколько аминокислотных остатков или добавление одного или нескольких лишних остатков на N- и/или С-конце, например добавление остатка метионина на N-конце.
Конъюгат данного изобретения имеет одно или несколько следующих улучшенных свойств в сравнении с hG-CSF, в частности в сравнении с rhG-CSF (например, филграстимом, ленограстимом или нартограстимом) или известными вариантами hG-CSF: увеличенный функциональный полупериод существования in vivo, увеличенный полупериод существования в плазме, пониженный почечный клиренс, пониженный опосредованный рецептором клиренс, уменьшенные побочные эффекты, такие как боль в костях, и уменьшенную иммуногенность.
Должно быть понятно, что аминокислотный остаток, содержащий группу присоединения для неполипептидной части молекулы, должен ли он удаляться или вводиться, будет выбираться на основе характера выбранной неполипептидной части молекулы и, в большинстве случаев, на основе способа, посредством которого должна достигаться конъюгация между полипептидом и неполипептидной частью молекулы. Например, когда неполипептидная часть молекулы является полимерной молекулой, такой как произведенная из полиэтиленгликоля или полиалкиленоксида молекула, аминокислотные остатки, содержащие группу присоединения, могут быть выбраны из группы, состоящей из лизина, цистеина, аспарагиновой кислоты, глутаминовой кислоты, гистидина и аргинина. Когда должна быть получена конъюгация с остатком лизина, подходящей активированной молекулой является, например, mPEG-SPA из Shearwater Polymers, Inc., оксикарбонилокси-N-дикарбоксиимид-ПЭГ (US 5122614) или ПЭГ, доступный из PolyMASC Pharmaceuticals plc. Первая из них будет дополнительно иллюстрироваться ниже.
Во избежание слишком большого разрушения структуры и функции исходной молекулы hG-CSF, общее число изменяемых аминокислотных остатков в соответствии с данным изобретением, например, как описано в последующих разделах здесь (в сравнении с аминокислотной последовательностью, показанной в SEQ ID NO:1), обычно не будет превышать 15. Точное число и тип вводимых или удаляемых аминокислотных остатков зависит, в частности, от желаемого характера и степени конъюгации (например, идентичности неполипептидной части молекулы, от того, сколько неполипептидных частей молекулы желательно или возможно конъюгировать с полипептидом, где конъюгация является желательной или где ее надо избежать, и т.д.). Предпочтительно полипептидная часть конъюгата данного изобретения или полипептид данного изобретения содержит аминокислотную последовательность, которая отличается на 1-15 аминокислотных остатков от аминокислотной последовательности, показанной в SEQ ID NO:1, обычно на 2-10 аминокислотных остатков, например, на 3-8 аминокислотных остатков, например, на 4-6 аминокислотных остатков, от аминокислотной последовательности, показанной в SEQ ID NO:1. Таким образом, обычно полипептидная часть конъюгата или полипептид данного изобретения содержит аминокислотную последовательность, которая отличается от аминокислотной последовательности, показанной в SEQ ID NO:1 на 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 аминокислотных остатков.
Полипептидная часть конъюгата обычно будет иметь аминокислотную последовательность с, по меньшей мере, приблизительно 80%-ной идентичностью относительно SEQ ID NO:1, предпочтительно, по меньшей мере, приблизительно 90%-ной, например, по меньшей мере, приблизительно 95%-ной идентичностью. Гомологию/идентичность аминокислотной последовательности удобным образом определяют из сопоставленных последовательностей, с использованием, например, программы ClustalW, версии 1.8, июнь 1999 года, с использованием параметров по умолчанию (Thompson et al., 1994, ClustalW: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice, Nucleic Acids Research, 22: 4673-4680) или версии 4.0 (http://pfam.wustl.edu/) PFAM families Database (Nucleic Acids Res. 1999 Jan 1; 27(1):260-2) с использованием версии 2,5 GENEDOC (Nicholas, К.В., Nicholas H.B. Jr., and Deerfield, D.W. II 1997 GeneDoc: Analysis and Visualization of Genetic Variation, EMBNEW.NEWS 4:14; Nicholas, K.B. and Nicholas H.B. Jr. 1997 GeneDoc: Analysis and Visualization of Genetic Variation).
В предпочтительном варианте одним различием между аминокислотной последовательностью полипептида и аминокислотной последовательностью, показанной в SEQ ID NO:1, является то, что, по меньшей мере, один и часто несколько, например 1-15, аминокислотных остатков, содержащих группу присоединения для неполипептидной части молекулы, были введены, предпочтительно, посредством замены в эту аминокислотную последовательность. Таким образом, полипептидную часть изменяют в содержании специфических аминокислотных остатков, с которыми связывается выбранная неполипептидная часть, в результате чего достигается более эффективная, специфическая и/или интенсивная конъюгация. Например, когда общее число аминокислотных остатков, содержащих группу присоединения для выбранного неполипептида, изменено до оптимизированного уровня, клиренс конъюгата обычно значимо уменьшается вследствие измененной формы, размера и/или заряда молекулы, достигаемых конъюгацией. Кроме того, когда общее число аминокислотных остатков, содержащих группу присоединения для выбранного неполипептида, увеличивается, большая доля молекулы полипептида защищается выбранными неполипептидными частями молекулы, что приводит к более низкой иммунной реакции.
Термин "одно различие", в применении в данной заявке, подразумевает возможность присутствия дополнительных различий. Таким образом, кроме указанного различия аминокислоты, другие аминокислотные остатки, чем указанные остатки, могут быть мутированными.
В дополнительном предпочтительном варианте одним различием между аминокислотной последовательностью полипептида и аминокислотной последовательностью, показанной в SEQ ID NO:1, является то, что, по меньшей мере, один и часто несколько, например 1-15, аминокислотных остатков, содержащих группу присоединения для неполипептидной части молекулы, были удалены, предпочтительно, посредством замены из этой аминокислотной последовательности. Посредством удаления одного или нескольких аминокислотных остатков, содержащих группу присоединения для неполипептидной части молекулы, можно избежать конъюгации с неполипептидной частью молекулы в частях полипептида, в которых такая конъюгация является невыгодной, например в аминокислотных остатках, расположенных в функциональном сайте полипептида или вблизи него (так как конъюгация в таком сайте приводит к инактивации или уменьшенной активности G-CSF полученного конъюгата вследствие нарушенного узнавания рецептора). В данном контексте, термин "функциональный сайт обозначает один или несколько аминокислотных остатков, которые являются существенными для функции или производительности hG-CSF или иным образом участвуют в них. Такие аминокислотные остатки являются частью функционального сайта. Функциональный сайт может быть определен способами, известными в данной области, и его предпочтительно идентифицируют анализом структуры полипептида, находящегося в комплексе с соответствующим рецептором, таким как hG-CSF-рецептор (см. Aritomi et al. Nature 401:713-717, 1999).
В следующем предпочтительном варианте, аминокислотная последовательность полипептида отличается от аминокислотной последовательности, показанной в SEQ ID NO:1, тем, что а) по меньшей мере, один указанный аминокислотный остаток, содержащий группу присоединения для неполипептидной части молекулы и присутствующий в аминокислотной последовательности, показанной в SEQ ID NO:1, был удален, предпочтительно посредством замены, и b) по меньшей мере, один указанный аминокислотный остаток, содержащий группу присоединения для неполипептидной части молекулы, был введен в эту аминокислотную последовательность, предпочтительно, посредством замены, причем указанные аминокислотные остатки являются любыми из аминокислотных остатков, описанных в последующих разделах здесь. Считается, что этот вариант представляет особенный интерес, заключающийся в том, что можно специально сконструировать полипептид таким образом, чтобы получить оптимальную конъюгацию с неполипептидной частью молекулы. Например, посредством введения и удаления выбранных аминокислотных остатков, как описано в следующих разделах, можно обеспечить оптимальное распределение групп присоединения для выбранной неполипептидной части молекулы, что приводит к получению конъюгата, в котором неполипептидные части молекулы размещены таким образом, чтобы а) эффективно защитить эпитопы и другие части поверхности полипептида и b) обеспечить оптимальный радиус Стокса конъюгата, без вызывания слишком большого структурного разрушения и нарушения тем самым функции полипептида.
Конъюгат данного изобретения обычно содержит достаточные число и тип неполипептидных частей молекулы для обеспечения конъюгата с увеличенными функциональным полупериодом существования in vivo и/или полупериодом существования в сыворотке в сравнении с hG-CSF, например филграстимом, ленограстимом или нартограстимом, и предпочтительно в сравнении с rhG-CSF, содержащим единственную присоединенную на N-конце ПЭГ-часть 20 кДа. Увеличенный полупериод существования in vivo подходящим образом определяют, как описано здесь в разделе Материалы и Способы.
Конъюгат данного изобретения может содержать, по меньшей мере, одну неконъюгированную, "способную к образованию конъюгата группу присоединения" для неполипептидной части молекулы. В данном контексте термин "способная к образованию конъюгата группа присоединения" обозначает группу присоединения, которая расположена в положении полипептида, где она доступна для конъюгации, и которая, за исключением особых предостережений, конъюгируется с соответствующей неполипептидной частью молекулы при подвергании конъюгации. Например, такая группа присоединения может быть частью аминокислотного остатка, участвующей в проявлении активности полипептида или иным образом необходимая для проявления активности полипептида. Подходящим способом исключения конъюгации, в противном случае способной к конъюгации группы присоединения, является защита этой группы присоединения посредством хелперной молекулы, например, как описано в разделе, озаглавленном "Блокирование функционального сайта". Должно быть понятно, что число неконъюгированных, способных к конъюгации групп присоединения зависит от конкретного полипептида G-CSF и местоположения способных к конъюгации групп присоединения. Например, конъюгат полипептида содержит одну или две неконъюгированные, способные к конъюгации группы присоединения и, по меньшей мере, одну, предпочтительно, две или более конъюгированных групп присоединения.
Конъюгат данного изобретения, в котором неполипептидная часть молекулы присоединена к лизину или N-концевому аминокислотному остатку
В одном аспекте данное изобретение относится к конъюгату полипептида, содержащему i) полипептид, проявляющий активность G-CSF, содержащий аминокислотную последовательность, которая отличается от аминокислотной последовательности, показанной в SEQ ID NO:1, по меньшей мере, одной заменой, выбранной из группы, состоящей из Т1К, Р2К, L3K, G4K, Р5К, А6К, S7K, S8K, L9K, Р10К, Q11K, S12K, F13K, L14K, L15K, Е19К, Q20K, V21K, Q21K, G26K, D27K, А29К, А30К, Е33К, А37К, Т38К, Y39K, L41K, Н43К, Р44К, Е45К, Е46К, V48K, L49K, L50K, Н52К, S53K, L54K, I56K, Р57К, Р60К, L61K, S62K, S63K, Р65К, S66K, Q67K, А68К, L69K, Q70K, L71K, А72К, G73K, S76K, Q77K, L78K, S80K, F83K, Q86K, G87K, Q90K, Е93К, G94K, S96K, Р97К, Е98К, L99K, G100K, P101K, T102K, D104K, T105K, Q107K, L108K, D109K, A111K, D112K, F113K, T115K, T116K, W118K, Q119K, Q120K, M121K, E122K, E123K, L124K, M126K, A127K, P128K, A129K, L130K, Q131K, P132K, T133K, Q134K, G135K, A136K, M137K, P138K, A139K, A141K, S142K, A143K, F144K, Q145K, S155K, H156K, Q158K, S159K, L161K, E162K, V164K, S164K, Y165K, V167K, L168K, H170K, L171K, A172K, Q173K и Р174К, и
ii) по меньшей мере, одну неполипептидную часть молекулы, присоединенную к остатку лизина этого полипептида.
hG-CSF содержит четыре остатка лизина, из которых К16 расположен в рецепторсвязывающем домене, а другие расположены в положениях 23, 34 и 40, соответственно, все относительно близко к рецепторсвязывающему домену. Во избежание конъюгации с одним или несколькими остатками лизина (так как это может инактивировать или серьезно снизить активность полученного конъюгата) может быть желательным удаление, по меньшей мере, одного остатка лизина, например двух, трех или всех четырех этих остатков. Таким образом, в другом, более предпочтительном аспекте, данное изобретение относится к конъюгату полипептида, описанному выше, где, по меньшей мере, один из аминокислотных остатков, выбранных из группы, состоящей из К16, К23, К34 и К40, был делегирован или заменен другим аминокислотным остатком. Предпочтительно, по меньшей мере, К16 заменен другим аминокислотным остатком.
Примеры предпочтительных аминокислотных замен включают в себя одну или несколько из замен Q70K, Q90K, Т105К, Q120K и Т133К, например, две, три или четыре из этих замен, например: Q70K+Q90K, Q70K+T105K, Q70K+120K, Q70K+T133K, Q90K+T105K, Q90K+Q120K, Q90K+T133K, T105K+Q120K, Т105К+Т133К, Q120K+T133K, Q70K+Q90K+T105K, Q70K+Q90K+Q120K, Q70K+Q90K+T133K, Q70K+T105K+T120K, Q70K+T105K+T133K, Q70K+Q120K+T133K, Q90K+T105K+Q120K, Q90K+T105K+T133K, Q90K+Q120K+T133K, Т105К+Т120К+Т133К, Q90K+T105K+Q120K+T133K, Q70K+T105K+Q120K+T133K, Q70K+Q90K+Q120K+T133K, Q70K+Q90K+T105K+T133K или Q70K+Q90K+T105K+Q120K.
Полипептид конъюгата в соответствии со вторым из аспектов, описанных в данном разделе (т.е. по меньшей мере, с одним введенным и одним удаленным лизином), предпочтительно содержит, по меньшей мере, один, например один, два, три или четыре, из замен, выбранных из группы, состоящей из K16R, K16Q, K23R, K23Q, K34R, K34Q, K40R и K40Q, более предпочтительно по меньшей мере одну из замен K16R и K23R, посредством этого конъюгация этих остатков может избегаться. Предпочтительно, этот полипептид содержит, по меньшей мере, одну замену, выбранную из группы, состоящей из K16R+K23R, K16R+K34R, K16R+K40R, K23R+K34R, K23R+K40R, K34R+K40R, K16R+K23R+K34R, K16R+K23R+K40R, K23R+K34R+K40R, K16R+K34R+K40R и K16R+K23R+K34R+K40R. Эти замены способны приводить к появлению, по меньшей мере, структурного различия в сравнении с нативным или исходным полипептидом.
Кроме замен, перечисленных выше, полипептидный конъюгат данного изобретения может также включать в себя одну или несколько замен, выбранных из R22K, R146K, R147K, R166K и R169K.
Хотя неполипептидная часть молекулы конъюгата в соответствии с этим аспектом данного изобретения может быть любой молекулой, которая при применении конкретного способа конъюгации имеет лизин в качестве группы присоединения, такой как углеводная часть молекулы, предпочтительно, чтобы неполипептидная часть молекулы была молекулой полимера. Молекула полимера может быть любой из молекул, упомянутых в разделе "Конъюгация с молекулой полимера", но, предпочтительно, выбрана из группы, состоящей из линейного или разветвленного полиэтиленгликоля или другого полиалкиленоксида. Предпочтительными молекулами полимеров являются, например, mPEG-SPA из Shearwater Polymers, Inc. или оксикарбонилокси-N-дикарбоксиимид-ПЭГ (US 5122614).
Должно быть понятно, что любая из аминокислотных замен, в частности замены, описанные в данном разделе, может комбинироваться с любой из аминокислотных замен, предпочтительно замен, указанных в других разделах здесь, описывающих конкретные аминокислотные замены, в том числе с введением и/или удалением сайтов гликозилирования.
Конъюгат данного изобретения, в котором неполипептидная часть молекулы является молекулой, которая имеет цистеин в качестве группы присоединения
В другом аспекте данное изобретение относится к конъюгату, содержащему
i) полипептид, проявляющий активность G-CSF, который содержит аминокислотную последовательность, которая отличается от аминокислотной последовательности, показанной в SEQ ID NO:1, по меньшей мере, одной заменой, выбранной из группы, состоящей из T1C, P2C, L3C, G4C, Р5С, А6С, S7C, S8C, L9C, Р10С, Q11C, S12C, F13C, L14C, L15C, Е19С, Q20C, V21C, R22C, Q25C, G25C, D27C, А29С, А30С, Е33С, А37С, Т38С, Y39C, L41C, Н43С, Р44С, Е45С, Е46С, V48C, L49C, L50C, Н52С, S53C, L54C, I56C, Р57С, Р60С, L61C, S62C, S63C, Р65С, S66C, Q67C, А68С, L69C, Q70C, L71K, А72С, G73C, S76C, Q77C, L78C, S80C, F83C, Q86C, G87C, Q90C, Е93С, G94C, S96C, Р97С, Е98С, L99C, G100C, Р101С, Т102С, D104C, Т105С, Q107C, L108C, D109C, А111С, D112C, F113C, Т115С, Т116С, W118C, Q119C, Q120C, М121С, Е122С, Е123С, L124C, М126С, А127С, Р128С, А129С, L130C, Q131C, Р132С, Т133С, Q134C, G135C, А136С, М137С, Р138С, А139С, А141С, S142C, А143С, F144C, Q145C, R146C, R147C, S155C, Н156С, Q158C, S159C, L161C, Е162С, V163C, S164C, Y165C, R166C, V167C, L168C, R169C, Н170С, L171C, А172С, Q173C и Р174С, и
ii) по меньшей мере, одну неполипептидную часть молекулы, присоединенную к остатку цистеина этого полипептида.
Рецепторсвязывающий домен hG-CSF содержит остаток цистеина в положении 17, который не участвует в цистине и который может быть выгодным образом удален, чтобы избежать конъюгации неполипептидной части молекулы с указанным цистеином. Таким образом, в другом, более предпочтительном аспекте данное изобретение относится к конъюгату, содержащему
i) полипептид, проявляющий активность G-CSF, который содержит аминокислотную последовательность, которая отличается от аминокислотной последовательности, показанной в SEQ ID NO:1, по меньшей мере, одной заменой, выбранной из группы, состоящей из T1C, Р2С, L3C, G4C, Р5С, А6С, S7C, S8C, L9C, Р10С, Q11C, S12C, F13C, L14C, L15C, Е19С, Q20C, V21C, R22C, Q25C, G26C, D27C, А29С, А30С, Е33С, А37С, Т38С, Y39C, L41C, Н43С, Р44С, Е45С, Е46С, V48C, L49C, L50C, Н52С, S53C, L54C, I56C, Р57С, Р60С, L61C, S62C, S63C, Р65С, S66C, Q67C, А68С, L69C, Q70C, L71K, А72С, G73C, S76C, Q77C, L78C, S80C, F83C, Q86C, G87C, Q90C, Е93С, G94C, S96C, Р97С, Е98С, L99C, G100C, Р101С, Т102С, D104C, Т105С, Q107C, L108C, D109C, А111С, D112C, F113C, Т115С, Т116С, W118C, Q119C, Q120C, М121С, Е122С, Е123С, L124C, М126С, А127С, Р128С, А129С, L130C, Q131C, Р132С, Т133С, Q134C, G135C, А136С, М137С, Р138С, А139С, А141С, S142C, А143С, F144C, Q145C, R146C, R147C, S155C, Н156С, Q158C, S159C, L161C, Е162С, V163C, S164C, Y165C, R166C, V167C, L168C, R169C, Н170С, L171C, А172С, Q173C и Р174С, в сочетании с удалением С17, предпочтительно заменой С17 любым другим аминокислотным остатком, например остатком серина, и
ii) по меньшей мере, одну неполипептидную часть молекулы, которая имеет остаток цистеина в качестве группы присоединения.
Предпочтительными заменами согласно этому аспекту данного изобретения являются замены аргинина цистеином, например одна или несколько из R146C, R147C, R166C и R169C.
Должно быть понятно, что любая из модификаций аминокислот, в частности замены, указанные в этом разделе, могут комбинироваться с любым из изменений аминокислот, в частности заменами, указанными в других разделах здесь, описывающих конкретные аминокислотные модификации, в том числе введение и/или удаление сайтов гликозилирования.
Конъюгат данного изобретения, в котором неполипептидная часть молекулы связывается с кислотной группой или С-концевым аминокислотным остатком
Еще в одном дополнительном аспекте данное изобретение относится к конъюгату, содержащему
i) полипептид, проявляющий активность G-CSF, который содержит аминокислотную последовательность, которая отличается от аминокислотной последовательности, показанной в SEQ ID NO:1, по меньшей мере, одной заменой, выбранной из группы, состоящей из T1D, P2D, L3D, G4D, P5D, A6D, S7D, S8D, L9D, P10D, Q11D, S12D, F13D, L14D, L15D, K16D, Q20D, V21D, R22D, K23D, Q25D, G26D, A29D, A30D, K34D, A37D, T38D, Y39D, K40D, L41D, H43D, P44D, V48D, L49D, L50D, H52D, S53D, L54D, I56D, P57D, P60D, L61D, S62D, S63D, P65D, S66D, Q67D, A68D, L69D, Q70D, L71D, A72D, G73D, S76D, Q77D, L78D, S80D, F83D, Q86D, G87D, Q90D, G94D, S96D, P97D, L99D, G100D, P101D, T102D, T105D, Q107D, L108D, A111D, F113D, T115D, T116D, W118D, Q119D, Q120D, M121D, L124D, M126D, A127D, P128D, A129D, L130D, Q131D, P132D, T133D, Q134D, G135D, A136D, M137D, P138D, A139D, A141D, S142D, A143D, F144D, Q145D, R146D, R147D, S155D, H156D, Q158D, S159D, L161D, V163D, S164D, Y165D, R166D, V167D, L168D, R169D, H170D, L171D, A172D, Q173D и P174D, или, по меньшей мере, одной заменой, выбранной из группы, состоящей из T1E, P2E, L3E, G4E, Р5Е, А6Е, S7E, S8E, L9E, Р10Е, Q11E, S12E, F13E, L14E, L15E, К16Е, Q20E, V21E, R22E, К23Е, Q25E, G26E, А29Е, А30Е, К34Е, А37Е, Т38Е, Y39E, К40Е, L41E, Н43Е, Р44Е, V48E, L49E, L50E, Н52Е, S53E, L54E, I56E, Р57Е, Р60Е, L61E, S62E, S63E, Р65Е, S66E, Q67E, А68Е, L69E, Q70E, L71E, А72Е, G73E, S76E, Q77E, L78E, S80E, F83E, Q86E, G87E, Q90E, G94E, S96E, Р97Е, L99E, G100E, Р101Е, Т102Е, Т105Е, Q107E, L108E, А111Е, F113E, Т115Е, Т116Е, W118E, Q119E, Q120E, М121Е, L124E, М126Е, А127Е, Р128Е, А129Е, L130E, Q131E, Р132Е, Т133Е, Q134E, G135E, А136Е, М137Е, Р138Е, А139Е, А141Е, S142E, А143Е, F144E, Q145E, R146E, R147E, S155E, Н156Е, Q158E, S159E, L161E, V163E, S164E, Y165E, R166E, V167E, L168E, R169E, Н170Е, L171E, А172Е, Q173E и Р174Е; и
ii) по меньшей мере, одну неполипептидную часть молекулы, имеющую остаток аспарагиновой кислоты или глутаминовой кислоты в качестве группы присоединения.
Примеры предпочтительных замен в соответствии с данным аспектом изобретения включают в себя Q67D/E, Q70D/E, Q77D/E, Q86D/E, Q90D/E, Q120D/E, Q131D/E, Q134D/E, Q145D/E и Q173D/E.
Кроме перечисленных выше замен, полипептид конъюгата в соответствии с любым из указанных выше аспектов может содержать удаление, предпочтительно посредством замены, по меньшей мере, одного из аминокислотных остатков, выбранных из группы, состоящей из D27, D104, D109, D112, Е19, Е33, Е45, Е46, Е93, Е98, Е122, Е123 и Е163. Эта замена может быть для любого другого аминокислотного остатка, в частности для остатка аспарагина или глутамина, посредством чего может избегаться конъюгация этих остатков. В частности, полипептид может содержать, по меньшей мере, одну из следующих замен: D27N, D104N, D109N, D112N, E19Q, E33Q, E45Q, E46Q, E93Q, E98Q, E122Q, E123Q и E163Q. Предпочтительно, аминокислотная замена в одном или нескольких из указанных положений может, кроме того, комбинироваться с, по меньшей мере, одной из следующих замен: D109N, D112N, E19Q, E122Q и E123Q. Замена любого из этих аминокислотных остатков способна привести к появлению, по меньшей мере, структурного различия.
Хотя неполипептидная часть молекулы конъюгата в соответствии с этим аспектом изобретения, которая имеет кислотную группу в качестве группы присоединения, может быть любой неполипептидной частью с таким свойством, в настоящее время предпочтительно, чтобы неполипептидная часть молекулы была молекулой полимера или органическим дериватизующим агентом, в частности молекулой полимера, а конъюгат готовят, например, как описано Sakane and Pardridge, Pharmaceutical Research, Vol.14, No.8, 1997, pp.1085-1091.
Должно быть понятно, что любая из аминокислотных замен, в частности замены, описанные в этом разделе, могут комбинироваться с любыми из изменений аминокислот, в частности заменами, описанными здесь в других разделах, описывающих аминокислотные изменения, в том числе введение и/или удаление сайтов гликозилирования.
Другие конъюгаты изобретения
Кроме неполипептидных частей молекулы, описанных выше, например, в разделах, озаглавленных "Конъюгат данного изобретения...", конъюгат данного изобретения может содержать дополнительные углеводные части молекулы как следствие того, что полипептид экспрессируется в гликозилирующей клетке-хозяине, и гликозилирование осуществляется в этих сайтах гликозилирования или во введенных сайтах гликозилирования.
Конъюгат данного изобретения, где неполипептидной частью молекулы является углеводная часть молекулы
В следующем аспекте данное изобретение относится к конъюгату, содержащему гликозилированный полипептид, проявляющий активность G-CSF, который содержит аминокислотную последовательность, которая отличается от аминокислотной последовательности, показанной в SEQ ID NO:1, тем, что, по меньшей мере, один не встречающийся в природе сайт гликозилирования был введен в эту аминокислотную последовательность посредством, по меньшей мере, одной замены, выбранной из группы, состоящей из L3N+P5S/T, P5N, A6N, S8N+P10S/T, P10N, Q11N+F13S/T, S12N+L14S/T, F13N+L15S/T, L14N+K16S/T, K16N+L18S/T, E19N+V21S/T, Q20N+R22S/T, V21N+K23S/T, R22N+I24S/T, K23N+Q25S/T, Q25N+D27S/T, G26N+G28S/T, D27N+A29S/T, A29N+L31S/T, A30N+Q32S/T, E33N+L35S/T, A37N+Y39S/T, T38N+K40S/T, Y39N+L41S/T, P44N+E46S/T, E45N+L47S/T, E46N+V48S/T, V48N+L50S/T, L49N+G51S/T, L50N+H52S/T, H52N+L54S/T, S53N+G55S/T, P60N, L61N, S63N+P65S/T, P65N+Q67S/T, S66N+A68S/T, Q67N+L69S/T, A68N+Q70S/T, L69N+L71S/T, Q70N+A72S/T, L71N+G73S/T, G73N+L75S/T, S76N+L78S/T, Q77N+H79S/T, L78N, S80N+L82S/T, F83N+Y85S/T, Q86N+L88S/T, G87N+L89S/T, Q90N+L92S/T, E93N+I95S/T, P97N+L99S/T, L99N+P101S/T, P101N+L103S/T, T102N+D104S/T, D104N+L106S/T, T105N+Q107S/T, Q107N+D109S/T, L108N+V110S/T, D109N+A111S/T, A111N+F113S/T, D112N+A114S/T, F113N, T115N+I117S/T, T116N+W118S/T, W118N+Q120S/T, Q119N+M121S/T, Q120N+E122S/T, M121N+E123S/T, E122N+L124S/T, E123N+G125S/T, L124N+M126S/T, M126N+P128S/T, P128N+L130S/T, L130N+P132S/T, P132N+Q134S/T, T133N+G135S/T, Q134N+A136S/T, A136N+P138S/T, P138N+F140S/T, A139N+A141S/T, A141N+A143S/T, S142N+F144S/T, A143N+Q145S/T, F144N+R146S/T, Q145N+R147S/T, R146N+A148S/T, R147N+G149S/T, S155N+L157S/T, H156N+Q158S/T, S159N+L161S/T, L161N+V163S/T, E162N, V163N+Y165S/T, S164N+R166S/T, Y165N+V167S/T, R166N+L168S/T, V167N+R169S/T, L168N+H170S/T, R169N+L171S/T и H170N+A172S/T, где S/T обозначает остаток S или Т, предпочтительно остаток Т.
Должно быть понятно, что для получения конъюгата согласно этому аспекту данного изобретения полипептид должен быть экспрессирован в гликозилирующей клетке-хозяине, способным присоединять олигосахаридные части молекулы в сайте (сайтах) гликозилирования или альтернативно подвергаться гликозилированию in vitro. Примеры гликозилирующих клеток-хозяев приведены в разделе ниже, озаглавленном "Связывание с олигосахаридной частью молекулы".
Альтернативно, конъюгат в соответствии с этим аспектом данного изобретения содержит полипептид, проявляющий активность G-CSF, который содержит аминокислотную последовательность, которая отличается от аминокислотной последовательности, показанной в SEQ ID NO:1, по меньшей мере, одной заменой, выбранной из группы, состоящей из P5N, A6N, P10N, P60N, L61N, L78N, F113N и E162N, в частности из группы, состоящей из P5N, A6N, P10N, P60N, L61N, F113N и E162N, например из группы, состоящей из P60N, L61N, F113N и E162N.
Альтернативно, конъюгат в соответствии с этим аспектом данного изобретения содержит полипептид, проявляющий активность G-CSF, который содержит аминокислотную последовательность, которая отличается от аминокислотной последовательности, показанной в SEQ ID NO:1, по меньшей мере, одной заменой, выбранной из группы, состоящей из D27N+A29S, D27N+A29T, D104N+L106S, D104N+L106T, D109N+A111S, D109N+A111T, D112N+A114S и D112N+A114T, более предпочтительно из группы, состоящей из D27N+A29S, D27N+A29T, D104N+L106S, D104N+L106T, D112N+A114S и D112N+A114T, например из группы, состоящей из D27N+A29S, D27N+A29T, D104N+L106S и D104N+L106T.
Кроме углеводной молекулы, конъюгат в соответствии с этим аспектом изобретения, описанным в данном разделе, может содержать дополнительные неполипептидные части молекулы, в частности молекулу полимера, как описано в данной заявке, конъюгированную с одной или несколькими группами присоединения, присутствующими в полипептидной части этого конъюгата.
Должно быть понятно, что любая из аминокислотных замен, в частности замены, описанные в этом разделе, могут комбинироваться с любыми из изменений аминокислот, в частности заменами, описанными в других разделах здесь, раскрывающих конкретные изменения аминокислот.
Кольцеобразно пермутированные варианты (Варианты с кольцевой перестановкой)
В следующем варианте полипептидная часть конъюгата полипептида данного изобретения может быть в форме кольцеобразно пермутированного варианта полипептидной последовательности, описанной в других отношениях здесь. В таком кольцеобразно пермутированном полипептиде первоначальные N-конец и С-конец соединены вместе либо непосредственно пептидной связью, либо опосредованно через пептидный линкер, тогда как новые N- и С-концы образуются между соседними аминокислотными остатками, которые первоначально были соединены пептидной связью. Поскольку первоначальные N- и С-концы будут обычно локализованы на некотором расстоянии друг от друга, они будут обычно связываться посредством пептидного линкера, имеющего подходящие длину и состав, так что структура и активность данного конъюгата не подвергаются повреждающему воздействию. Должно быть ясно, что новые N-конец и С-конец не должны образовываться между парой аминокислотных остатков, где это могло бы мешать активности данного полипептида. Кольцеобразно пермутированные агонисты G-CSF-рецептора описаны в патенте США US 6100070, на который делается ссылка в отношении дополнительной информации по выбору пептидных линкеров и местоположения новых N-конца и С-конца, а также способов получения таких вариантов.
Действие конъюгатов данного изобретения на образование лейкоцитов и нейтрофилов
В следующем варианте полипептидный конъюгат данного изобретения может быть охарактеризован как конъюгат, проявляющий активность G-CSF и содержащий полипептид с аминокислотной последовательностью, которая отличается, по меньшей мере, на один аминокислотный остаток от аминокислотной последовательности, показанной в SEQ ID NO:1, и имеет, по меньшей мере, одну неполипептидную часть молекулы, присоединенную к группе присоединения полипептида, причем этот полипептидный конъюгат дополнительно удовлетворяет, по меньшей мере, одному из следующих критериев (A)-(D):
(А) после одного подкожного введения 100 микрограммов на кг веса тела крысам (в расчете на вес полипептидной части конъюгата) он:
i) увеличивает образование лейкоцитов с, по меньшей мере, той же самой скоростью и до, по меньшей мере, приблизительно того же самого уровня (измеренного в виде числа клеток на литр крови), что и введение 100 микрограммов неконъюгированного hG-CSF на кг веса тела, в течение 12 часов после введения, и
ii) увеличивает уровень лейкоцитов (измеренный в виде числа лейкоцитов на литр крови) выше уровня лейкоцитов перед введением в течение, по меньшей мере, приблизительно 96 часов, предпочтительно в течение, по меньшей мере, приблизительно 120 часов;
(B) после одного подкожного введения 25 микрограммов на кг веса тела крысам (в расчете на вес полипептидной части конъюгата) он:
i) увеличивает образование лейкоцитов с, по меньшей мере, той же самой скоростью и до, по меньшей мере, приблизительно того же самого уровня (измеренного в виде числа клеток на литр крови), что и введение 100 микрограммов неконъюгированного hG-CSF на кг веса тела, в течение 12 часов после введения, и
ii) увеличивает уровень лейкоцитов (измеренный в виде числа лейкоцитов на литр крови) выше уровня лейкоцитов перед введением в течение, по меньшей мере, приблизительно 72 часов, предпочтительно, по меньшей мере, приблизительно 96 часов, более предпочтительно в течение, по меньшей мере, приблизительно 120 часов;
(C) после одного подкожного введения 100 микрограммов на кг веса тела крысам (в расчете на вес полипептидной части конъюгата) он:
i) увеличивает образование нейтрофилов с, по меньшей мере, той же самой скоростью и до, по меньшей мере, приблизительно того же самого уровня (измеренного в виде числа клеток на литр крови), что и введение 100 микрограммов неконъюгированного hG-CSF на кг веса тела, в течение 12 часов после введения, и
ii) увеличивает уровень нейтрофилов (измеренный в виде числа клеток на литр крови) выше уровня нейтрофилов перед введением в течение, по меньшей мере, приблизительно 96 часов, предпочтительно, в течение, по меньшей мере, приблизительно 120 часов;
(D) после одного подкожного введения 25 микрограммов на кг веса тела крысам (в расчете на вес полипептидной части конъюгата) он:
i) увеличивает образование нейтрофилов с, по меньшей мере, той же самой скоростью и до, по меньшей мере, приблизительно того же самого уровня (измеренного в виде числа клеток на литр крови), что и введение 100 микрограммов неконъюгированного hG-CSF на кг веса тела, в течение 12 часов после введения, и
ii) увеличивает уровень нейтрофилов (измеренный в виде числа клеток на литр крови) выше уровня нейтрофилов перед введением в течение, по меньшей мере, приблизительно 72 часов, предпочтительно, по меньшей мере, приблизительно 96 часов, более предпочтительно в течение, по меньшей мере, приблизительно 120 часов.
Неполипептидная часть конъюгата данного изобретения
Как указывалось дополнительно выше, неполипептидная часть конъюгата данного изобретения предпочтительно выбрана из группы, состоящей из молекулы полимера, липофильного соединения, углеводной части (например, присоединенной посредством гликозилирования in vivo) и органического дериватизующего агента. Все эти агенты могут придавать желаемые свойства полипептидной части конъюгата, в частности увеличенный функциональный полупериод существования in vivo и/или увеличенный полупериод существования в сыворотке. Полипептидная часть конъюгата обычно конъюгирована только с одним типом неполипептидной части, но может быть также конъюгирована с двумя или большим числом различных типов неполипептидных частей, например с молекулой полимера и олигосахаридной частью молекулы, с липофильной группой и олигосахаридной частью молекулы, с органическим дериватизующим агентом и олигосахаридной частью, с липофильной группой и молекулой полимера, и т.д. Конъюгация двух или большего числа различных неполипептидных частей может выполняться одновременно или последовательно.
Способы получения конъюгата данного изобретения
В следующих разделах "Конъюгация с липофильным соединением", "Конъюгация с молекулой полимера", "Конъюгация с олигосахаридной частью" и "Конъюгация с органическим дериватизующим агентом" описана конъюгация с конкретными типами неполипептидных частей молекулы. Обычно полипептидный конъюгат в соответствии с данным изобретением может быть получен культивированием подходящей клетки-хозяина в условиях, пригодных для экспрессии полипептида, и извлечением полипептида, причем а) полипептид содержит, по меньшей мере, один сайт N- или О-гликозилирования, а клетка-хозяин является эукариотической клеткой-хозяином, способной к гликозилированию in vivo, и/или b) полипептид подвергают конъюгации с неполипептидной частью молекулы in vitro.
Конъюгация с липофильным соединением
Полипептид и липофильное соединение могут быть конъюгированы друг с другом либо непосредственно, либо с использованием линкера. Липофильным соединением может быть природное соединение, такое как насыщенная или ненасыщенная жирная кислота, дикетон жирной кислоты, терпен, простагландин, витамин, каротиноид или стероид, или синтетическое соединение, такое как угольная кислота, спирт, амин и сульфоновая кислота с одним или несколькими алкильными, арильными, алкенильными группами, или другие полиненасыщенные соединения. Конъюгация между полипептидом и липофильным соединением, необязательно через линкер, может быть выполнена в соответствии со способами, известными в данной области, например, как описано Bodanszky в Peptide Synthesis, John Wiley, New York, 1976 и в WO 96/12505.
Конъюгация с молекулой полимера
Молекулой полимера для связывания с полипептидом может быть любая подходящая полимерная молекула, например природный или синтетический гомополимер или гетерополимер, обычно с молекулярной массой в диапазоне приблизительно 300-100000 Да, например, приблизительно 500-20000 Да, более предпочтительно в диапазоне приблизительно 1000-15000 Да, даже более предпочтительно в диапазоне приблизительно 2000-12000 Да, например, приблизительно 3000-10000. При применении для молекул полимеров здесь слово "приблизительно" указывает приблизительную среднюю молекулярную массу и отражает тот факт, что обычно в конкретном препарате полимера будет иметься определенный разброс молекулярных масс.
Примеры гомополимеров включают в себя полиол (т.е. поли-ОН), полиамин (т.е. поли-NH2) и поликарбоновую кислоту (т.е. поли-СООН). Гетерополимер представляет собой полимер, который содержит различные связывающие группы, такие как гидроксильная группа и аминогруппа.
Примеры подходящих молекул полимеров включают в себя молекулы полимеров, выбранные из группы, состоящей из полиалкиленоксида (РАО), в том числе полиалкиленгликоля (ПАГ), такого как линейный или разветвленный полиэтиленгликоль (ПЭГ) и полипропиленгликоль (ППГ), поливинилового спирта (ПВС), поликарбоксилата, поливинилпирролидона, сополимера полиэтилена и ангидрида малеиновой кислоты, сополимера полистирола и ангидрида малеиновой кислоты, декстрана, в том числе карбоксиметилдекстрана, или любого другого биополимера, пригодного для уменьшения иммуногенности и/или увеличения функционального полупериода существования in vivo и/или полупериода существования в сыворотке. Другим примером молекулы полимера является человеческий альбумин или другой изобильный белок плазмы. Обычно, произведенные из полиалкиленгликоля полимеры являются биосовместимыми, нетоксичными, неантигенными, неиммуногенными, имеют разные свойства водорастворимости и легко экскретируются из живых организмов.
ПЭГ является предпочтительной полимерной молекулой, так как он имеет только несколько реакционноспособных групп, способных к сшиванию, в сравнении с полисахаридами, такими как декстран. В частности, монофункциональный ПЭГ, например метоксиполиэтиленгликоль (мПЭГ), представляет интерес, так как способ его химического связывания является относительно простым (только одна реактивная группа является доступной для конъюгации с группами присоединения на полипептиде). Поэтому риск сшивания исключается, полученные конъюгаты полипептида являются более гомогенными, и реакцию этих молекул полимера с полипептидом легче контролировать.
Для выполнения ковалентного присоединения полимерных молекул к полипептиду обеспечивают гидроксильные концевые группы молекулы полимера в активированной форме, т.е. с реакционноспособными функциональными группами. Подходящие активированные полимерные молекулы являются коммерчески доступными, например, из Shearwater Polymers, Inc., Huntsville, AL, USA или из PolyVASC Pharmaceutical plc, UK. Альтернативно, молекула полимера может быть активирована общепринятыми способами, известными в данной области, например, как описано в WO 90/13540. Конкретные примеры активированных линейных или разветвленных полимерных молекул для применения в данном изобретении, описаны в Shearwater Polymers, Inc. 1997 и 2000 Catalogs (Functionalized Biocompatible Polymers for Research and pharmaceuticals, Polyethylen Glycol and Derivatives, включенных здесь в качестве ссылки). Конкретные примеры активированных ПЭГ-полимеров включают в себя следующие линейные ПЭГ: NHS-PEG (например, SPA-PEG, SSPA-PEG, SBA-PEG, SS-PEG, SSA-PEG, SC-PEG, SG-PEG и SCM-PEG) и NOR-PEG, BTC-PEG, EPOX-PEG, NCO-PEG, NPC-PEG, CDI-PEG, ALD-PEG, TRES-PEG, VS-PEG, IODO-PEG и MAL-PEG, и разветвленные ПЭГ, такие как PEG2-NHS и описанные в US 5932462 и US 5643575, оба включены здесь в качестве ссылки. Кроме того, следующие публикации, включенные здесь в качестве ссылки, описывают применимые полимерные молекулы и/или химические способы ПЭГилирования: US 5824778, US 5476653, WO 97/32607, ЕР 229108, ЕР 402378, US 4902502, US 5281698, US 5122614, US 5219564, WO 92/16555, WO 94/04193, WO 94/14758, WO 94/17039, WO 94/18247, WO 94/28024, WO 95/00162, WO 95/11924, WO 95/13090, WO 95/33490, WO 96/00080, WO 97/18832, WO 98/41562, WO 98/48837, WO 99/32134, WO 99/32139, WO 99/32140, WO 99/32140, WO 96/40791, WO 98/32466, WO 95/06058, ЕР 439508, WO 97/03106, WO 96/21469, WO 95/13312, ЕР 921131, US 5736625, WO 98/05363, ЕР 809996, US 5629384, WO 96/41813, WO 96/07670, US 5473034, US 5516673, ЕР 605963, US 5382657, ЕР 510356, ЕР 400472, ЕР 183503 и ЕР 154316.
Конъюгацию полипептида и активированных молекул полимеров проводят с использованием любого общепринятого способа, например, как описано в следующих ссылках (которые описывают также подходящие способы для активации полимерных молекул): R.F.Taylor (1991), "Protein immobilization. Fundamental and applications". Marcel Dekker, N.Y.; S.S.Wong (1992), "Chemistry of Protein Conjugation and Crosslinking", CRC Press, Boca Raton; G.T.Hermanson et al. (1993), "Immobilization Affinity Ligand Techniques", Academic Press, N.Y.). Специалисту с квалификацией в данной области будет ясно, что способ активации и/или способ конъюгации, которые должны использоваться, зависят от группы (групп) присоединения полипептида (примеры которых даны дополнительно выше), а также функциональных групп полимера (например, являющихся аминогруппой, гидроксилом, карбоксилом, альдегидом, сульфгидрилом, сукцинимидилом, малеимидом, винилсульфоном или галогенацетатом). ПЭГилирование может быть направлено на конъюгацию для всех доступных групп присоединения на полипептиде (т.е. таких групп присоединения, которые экспонированы на поверхности полипептида) или может быть направлено на одну или несколько конкретных групп присоединения, например на N-концевую аминогруппу (US 5985265). Кроме того, конъюгация может достигаться в одной стадии или постадийным способом (например, как описано в WO 99/55377).
Должно быть понятно, что ПЭГилирование планируется таким образом, чтобы получить оптимальную молекулу с учетом числа присоединенных молекул ПЭГ, размера и формы таких молекул (например, являются ли они линейными или разветвленными), и с учетом того, где в полипептиде присоединяются такие молекулы. Молекулярную массу используемого полимера выбирают с учетом желаемого эффекта, который должен быть получен. Например, если первичной целью конъюгации является получение конъюгата, имеющего высокую молекулярную массу и более крупный размер (например, для уменьшения почечного клиренса), можно выбрать для конъюгации одну или несколько высокомолекулярных полимерных молекул или ряд полимерных молекул с более низкой молекулярной массой для получения желаемого действия. Однако предпочтительно использовать несколько полимерных молекул с меньшей молекулярной массой. Когда желательна высокая степень защиты эпитопа, это может быть достигнуто применением достаточно высокого числа молекул низкомолекулярных полимеров (например, с молекулярной массой около 5000 Да) для эффективной защиты всех или большинства эпитопов полипептида. Например, могут быть использованы 2-8 таких полимеров. Как показывают приведенные ниже примеры, может быть выгодным иметь большее число полимерных молекул более низкой молекулярной массы (например, 4-6 с MW 5000), в сравнении с меньшим числом полимерных молекул с более высокой молекулярной массой (например, 1-3 с MW 12000-20000) для улучшения функционального полупериода существования in vivo конъюгата полипептида, даже когда общая молекулярная масса присоединенных полимерных молекул в этих двух случаях является одинаковой. Считается, что присутствие большего числа меньших полимерных молекул обеспечивает полипептид с большим диаметром или средним размером, чем, например, единственная более крупная молекула полимера, по меньшей мере, в том случае, когда молекулы полимера относительно однородно распределены на поверхности полипептида.
Хотя конъюгация только единственной полимерной молекулы с единственной группой присоединения на белке не является предпочтительной, в том случае, когда присоединена только одна молекула полимера, обычно будет выгодным, чтобы эта полимерная молекула, которая может быть линейной или разветвленной, имела относительно высокую молекулярную массу, например около 20 кДа.
Обычно конъюгацию полимера выполняют в условиях, нацеленных на реакцию по возможности многих доступных групп присоединения полимера с полимерными молекулами. Это достигается посредством подходящего молярного избытка полимера относительно полипептида. Обычные молярные отношения молекул активированного полимера к полипептиду составляют до около 1000-1, например, до около 200-1 или до около 100-1. В некоторых случаях это отношение может быть несколько более низким, однако до приблизительно 50-1, 10-1 или 5-1.
Данное изобретение обсуждает также связывание молекул полимеров через линкер. Подходящие линкеры хорошо известны специалисту в данной области. Предпочтительным примером является цианурхлорид (Abuchowski et al., (1977). J. Biol. Chem., 252, 3578-3581; US 4179337; Shafer et al., (1986), J. Polym. Sci. Polym. Chem. Ed., 24, 375-378.
После конъюгации остаточные активированные молекулы полимера блокируют в соответствии со способами, известными в данной области, например, добавлением первичного амина к реакционной смеси, и полученные инактивированные молекулы полимера удаляют подходящим способом (см. Материалы и Способы).
В предпочтительном варианте конъюгат полипептида данного изобретения содержит молекулу ПЭГ, присоединенную к некоторым, большинству или, предпочтительно, по существу ко всем остаткам лизина в полипептиде, доступным для ПЭГилирования, в частности линейную или разветвленную молекулу ПЭГ, например, с молекулярной массой около 1-15 кДа, обычно около 2-12 кДа, например, около 3-10 кДа, например, около 5 или 6 кДа.
Должно быть понятно, что в зависимости от обстоятельств, например аминокислотной последовательности полипептида, природы используемого активированного ПЭГ-соединения и конкретных условий ПЭГилирования, в том числе молярного отношения ПЭГ к полипептиду, могут быть получены варьирующиеся степени ПЭГилирования, причем более высокую степень ПЭГилирования обычно получают с более высоким отношением ПЭГ к полипептиду. ПЭГилированные полипептиды, получаемые из любого конкретного процесса ПЭГилирования, будут, однако, обычно иметь случайное распределение конъюгатов полипептида, имеющих слегка различные степени ПЭГилирования.
Еще в одном варианте конъюгат полипептида данного изобретения может содержать молекулу ПЭГ, присоединенную к остаткам лизина в полипептиде, доступным для ПЭГилирования, и, кроме того, к N-концевому аминокислотному остатку полипептида.
Связывание с олигосахаридной частью
Конъюгация с олигосахаридной частью может происходить in vivo или in vitro. Для получения гликозилирования in vivo молекулы G-CSF, содержащей один или несколько сайтов гликозилирования, нуклеотидная последовательность, кодирующая этот полипептид, должна быть встроена в гликозилирующего, эукариотического экспрессирующего хозяина. Экспрессирующая клетка-хозяин может быть выбрана из грибных клеток (нитчатые грибы или дрожжи), клеток насекомых или клеток животных или из трансгенных клеток растений. В одном варианте клетка-хозяин является клеткой млекопитающего, такой как клетка СНО, клетка ВНК или НЕК, например НЕК 293, или клеткой насекомого, такой как клетка SF9, или дрожжевой клеткой, например S. cerevisiae или Pichia pastoris, или любой из клеток-хозяев, приведенных далее. Может быть также использовано ковалентное связывание in vitro (например, декстрана) с аминокислотными остатками полипептида, например, как описано в WO 87/05330 и в Aplin et al., CRC Crit Rev. Biochem., pp.259-306, 1981.
Связывание in vitro олигосахаридных частей молекулы или ПЭГ со связанными в белке или пептиде остатками Gln может проводиться трансглутаминазами (ТГазами). Трансглутаминазы катализируют перенос донорных аминогрупп к белок- и пептид-связанным остаткам Gln в так называемой реакции сшивания. Донорными аминогруппами могут быть белок- или пептид-связанные аминогруппы, например ε-аминогруппа в остатках Lys, или они могут быть частью небольшой или большой органической молекулы. Примером небольшой органической молекулы, функционирующей в качестве аминодонора в катализируемом трансглутаминазой сшивании, является путресцин (1,4-диаминобутан). Примером более крупной органической молекулы, функционирующей в качестве аминодонора в катализируемом трансглутаминазой сшивании, является аминосодержащий ПЭГ (Sato et al., Biochemistry 35, 13072-13080).
Трансглутаминазы (ТГазы) являются обычно высокоспецифическими ферментами, и не каждый остаток Gln, открытый на поверхности белка, является доступным катализируемому ТГазой сшиванию с аминосодержащими веществами. Напротив, только небольшое количество остатков Gln функционируют в природе в качестве субстратов ТГазы, но точные характеристики, определяющие, какие остатки Gln являются хорошими субстратами для ТГазы, остаются неизвестными. Таким образом, для того чтобы сделать белок доступным для катализируемых ТГазой реакций сшивания, обязательным условием часто является добавление в подходящих положениях отрезков аминокислотной последовательности, о которых известно, что они очень хорошо функционируют в качестве субстратов ТГазы. Известны несколько аминокислотных последовательностей, которые являются превосходными природными субстратами ТГазы или содержат превосходные природные субстраты ТГазы, например вещество Р, элафин, фибриноген, фибронектин, ингибитор α2-плазмина, α-казеины и β-казеины.
Связывание с органическим дериватизующим агентом
Ковалентная модификация полипептида, проявляющего активность G-CSF, может быть выполнена реакцией одной или нескольких групп присоединения полипептида с органическим дериватизующим агентом. Подходящие дериватизующие агенты хорошо известны в данной области. Например, цистеиновые остатки наиболее часто реагируют с α-галогенацетатами (и соответствующими аминами), такими как хлоруксусная кислота или хлорацетамид, с образованием карбоксиметил- или карбоксиамидометилпроизводных. Цистеиновые остатки также дериватизуются реакцией с бромтрифторацетоном, α-бром-β-(4-имидазоил)пропионовой кислотой, хлорацетилфосфатом, N-алкилмалеимидом, 3-нитро-2-пиридилдисульфидом, метил-2-пиридилдисульфидом, п-хлормеркурбензоатом, 2-хлормеркур-4-нитрофенолом или хлор-7-нитробензо-2-окса-1,3-диазолом. Гистидиновые остатки дериватизуются реакцией с диэтилпирокарбонатом при рН 5,5-7,0, так как этот агент является относительно специфическим в отношении гистидильной боковой цепи. Применим также пара-бромфенацилбромид. Реакцию проводят предпочтительно в 0,1 М какодилате натрия при рН 6,0. Лизиновые и аминоконцевые остатки реагируют с ангидридами янтарной или другой карбоновой кислоты. Дериватизация этими агентами приводит к обращению заряда лизиновых остатков. Другие подходящие реагенты для дериватизации α-аминосодержащих остатков включают в себя имидоэфиры, такие как метилпиколинимидат, пиридоксальфосфат, пиридоксаль, хлороборгидрид, тринитробензолсульфоновую кислоту, O-метилизомочевину, 2,4-пентандион и катализируемую трансглутаминазой реакцию с глиоксилатом. Аргининовые остатки модифицируют реакцией с одним или несколькими общепринятыми реагентами, среди них находятся фенилглиоксаль, 2,3-бутандион, 1,2-циклогександион и нингидрин. Дериватизация остатков аргинина требует, чтобы реакция выполнялась в щелочных растворах вследствие высокого рКа функциональной группы гуанидина.
Кроме того, эти реагенты могут взаимодействовать с группами лизина, а также гуанидиногруппой аргинина. Карбоксильные боковые группы (аспартил или глутамил) избирательно модифицируются реакцией с карбодиимидами (R-N=C=N-R'), где R и R' являются различными алкильными группами, такими как 1-циклогексил-3-(3-морфолинил-4-этил)карбодиимид или 1-этил-3-(4-азониа-4,4-диметилпентил)карбодиимид. Кроме того, аспартиловый и глутамиловый остатки превращают в аспарагиниловый и глутаминиловый остатки реакцией с ионами аммония.
Блокирование функционального сайта
Сообщалось, что избыточная конъюгация с полимером может приводить к потере активности полипептида, с которым конъюгирован этот полимер. Эта проблема может быть устранена, например, удалением групп присоединения, локализованных на функциональном сайте или блокированием функционального сайта перед конъюгацией таким образом, что данный функциональный сайт является блокированным во время конъюгации. Последняя стратегия составляет следующий вариант данного изобретения (первая стратегия приведена дополнительно в качестве примера выше, например, удаление остатков лизина, которые могут быть расположены близко к функциональному сайту). Более конкретно, согласно второй стратегии конъюгацию между полипептидом и неполипептидной частью проводят в условиях, в которых функциональный сайт блокирован хелперной молекулой (молекулой-помощником), способной связываться с функциональным сайтом полипептида.
Предпочтительно хелперной молекулой является молекула, которая специфически узнает функциональный сайт полипептида, такая как рецептор, в частности, G-CSF-рецептор или часть G-CSF-рецептора.
Альтернативно, хелперная молекула может быть антителом, в частности моноклональным антителом, узнающим полипептид, проявляющий активность G-CSF. В частности, хелперной молекулой может быть нейтрализующее моноклональное антитело.
Полипептиду дают взаимодействовать с хелперной молекулой перед выполнением конъюгации. Это гарантирует, что функциональный сайт полипептида является закрытым или защищенным и, следовательно, недоступным для дериватизации неполипептидной частью молекулы, такой как полимер. После его элюции от хелперной молекулы конъюгат между неполипептидной частью и полипептидом может быть извлечен с, по меньшей мере, частично защищенным функциональным сайтом.
Последующую конъюгацию полипептида, имеющего блокированный функциональный сайт, с полимером, липофильным соединением, олигосахаридной частью, органическим дериватизующим агентом или любым другим соединением проводят обычным путем, например, как описано в разделах выше, озаглавленных "Конъюгация с...".
Независимо от природы хелперной молекулы, используемой для защиты функционального сайта полипептида от конъюгации, желательно, чтобы хелперная молекула была свободна от групп присоединения или содержала немного групп присоединения для выбранной неполипептидной части в той части (частях) этой молекулы, где конъюгация с такими группами могла бы препятствовать десорбции конъюгированного полипептида из хелперной молекулы. Таким образом, может быть получена селективная конъюгация с группами присоединения, присутствующими в незащищенных частях полипептида, и можно повторно использовать хелперную молекулу для повторяемых циклов конъюгации. Например, если неполипептидная часть является молекулой полимера, такой как ПЭГ, которая имеет эпсилон-аминогруппу лизина или N-концевой аминокислотный остаток в качестве группы присоединения, желательно, чтобы эта хелперная молекула не содержала конъюгируемых эпсилон-аминогрупп, предпочтительно, не содержала никаких эпсилон-аминогрупп. Таким образом, в предпочтительном варианте хелперной молекулой является белок или пептид, способный связываться с функциональным сайтом полипептида, причем этот белок или пептид не содержит никаких групп присоединения для выбранной неполипептидной части.
Особый интерес в этой связи представляет вариант данного изобретения, в котором конъюгаты полипептида получают из разнообразной популяции нуклеотидных последовательностей, кодирующих представляющий интерес полипептид, блокирование функциональной группы выполняют в микропланшетах для титрования перед конъюгацией, например, высевом варианта экспрессируемого полипептида в микропланшет для титрования, содержащий иммобилизованную блокирующую группу, такую как рецептор, антитело или т.п.
В следующем варианте хелперную молекулу сначала ковалентно связывают с твердой фазой, такой как упаковывающие материалы для колонок, например, Сефадекс или гранулы агарозы, или поверхность, например реакционный сосуд. Затем, полипептид загружают на материал колонки, несущий хелперную молекулу, и конъюгацию проводят в соответствии со способами, известными в данной области, например, как описано в разделах, озаглавленных "Конъюгация с ...". Эта процедура позволяет конъюгату полипептида отделиться от хелперной молекулы при элюции. Конъюгат полипептида элюируют общепринятыми способами при физико-химических условиях, которые не приводят к существенной деградации конъюгата полипептида. Жидкую фазу, содержащую конъюгат полипептида, отделяют от твердой фазы, с которой остается ковалентно связанной хелперная молекула. Это отделение может быть достигнуто разными путями: например хелперная молекула может быть дериватизована второй молекулой (например, биотином), которая может узнаваться специфическим связывающим веществом (например, стрептавидином). Специфическое связывающее вещество может быть связано с твердой фазой, что позволяет отделение конъюгата полипептида от комплекса хелперная молекула - вторая молекула пропусканием через твердофазную колонку со вторым хелпером, которая будет удерживать, при последующей элюции, комплекс хелперная молекула - вторая молекула, но не конъюгат полипептида. Конъюгат полипептида может высвобождаться из хелперной молекулы любым подходящим способом. Удаление защиты может быть достигнуто обеспечением условий, в которых хелперная молекула диссоциирует от функционального сайта G-CSF, с которым она связана. Например, комплекс между антителом, с которым конъюгирован полимер, и антиидиотипическое антитело могут быть диссоциированы коррекцией рН до кислотного или щелочного рН.
Конъюгация меченого полипептида
В альтернативном варианте полипептид экспрессируется в виде слитого с меткой белка, т.е. с аминокислотной последовательностью или пептидным отрезком, изготовленным обычно из до 1-30, например 1-20 аминокислотных остатков. Кроме возможности быстрой и легкой очистки, эта метка является удобным инструментом для достижения конъюгации между меченым полипептидом и неполипептидной частью конъюгата. В частности, эта метка может быть использована для получения конъюгации в микропланшетах для титрования или в других носителях, таких как парамагнитные гранулы, на которых меченый полипептид может быть иммобилизован посредством этой метки. Конъюгация с меченым полипептидом, например, в микропланшетах для титрования имеет то преимущество, что меченый полипептид может быть иммобилизован в микропланшетах для титрования непосредственно из культурального бульона (в принципе без всякой очистки) и подвергнут конъюгации. Кроме того, метка может функционировать в качестве спейсерной молекулы, гарантируя улучшенную доступность к иммобилизованному полипептиду, который должен быть конъюгирован. Конъюгация с использованием меченого полипептида может осуществляться с любой из неполипептидных частей, описанных здесь, например с молекулой полимера, такой как ПЭГ.
Идентичность специфической метки, которая должна быть использована, не является решающей до тех пор, пока эта метка способна экспрессироваться с полипептидом и способна иммобилизоваться на поверхности носителя или материала носителя. Ряд подходящих меток являются коммерчески доступными, например, из Unizyme Laboratories, Denmark. Например, метка может состоять из любой из следующих последовательностей:
His-His-His-His-His-His
Met-Lys-His-His-His-His-His-His
Met-Lys-His-His-Ala-His-His-Gln-His-His
Met-Lys-His-Gln-His-Gln-His-Gln-His-Gln-His-Gln-His-GIn
Met-Lys-His-Gln-His-Gln-His-Gln-His-Gln-His-Gln-His-Gln-GIn
или любой из следующих последовательностей:
EQKLI SEEDL (С-концевая метка, описанная в Mol. Cell. Biol. 5:3610-16, 1985)
DYKDDDDK (С- или N-концевая метка)
YPYDVPDYA
Антитела против приведенных выше меток являются коммерчески доступными, например, из ADI, Aves Lab and Research Diagnostics.
Удобный способ применения меченого полипептида для ПЭГилирования дается в разделе Материалы и Способы ниже. Последующее отщепление метки от полипептида может выполняться с использованием коммерчески доступных ферментов.
Способы получения полипептида данного изобретения или полипептидной части конъюгата данного изобретения
Полипептид данного изобретения или полипептидная часть конъюгата данного изобретения, необязательно в гликозилированной форме, могут быть получены любым подходящим способом, известным в данной области. Такие способы включают в себя конструирование нуклеотидной последовательности, кодирующей этот полипептид, и экспрессию этой последовательности в подходящем трансформированном или трансфицированном хозяине. Однако полипептиды данного изобретения могут быть получены, хотя менее эффективно, химическим синтезом или комбинацией химического синтеза и технологии рекомбинантных ДНК.
Нуклеотидная последовательность, кодирующая полипептид или полипептидную часть данного изобретения, может быть сконструирована выделением или синтезом нуклеотидной последовательности, кодирующей исходный G-CSF, например, hG-CSF с аминокислотной последовательностью, показанной в SEQ ID NO:1, и затем изменением этой нуклеотидной последовательности таким образом, чтобы осуществить введение (т.е. инсерцию или замену) или делецию (т.е. удаление или замену) соответствующего аминокислотного остатка (остатков).
Нуклеотидную последовательность просто модифицируют сайт-направленным мутагенезом согласно общепринятым способам. Альтернативно, нуклеотидную последовательность получают химическим синтезом, например, с использованием олигонуклеотидного синтезатора, где олигонуклеотиды конструируют на основе аминокислотной последовательности желаемого полипептида, с предпочтительным отбором тех кодонов, которые являются предпочтительными в клетке-хозяине, в которой будет продуцироваться рекомбинантный полипептид. Например, несколько небольших олигонуклеотидов, кодирующих части желаемого полипептида, могут быть синтезированы и собраны при помощи ПЦР, лигирования или лигазной цепной реакции (LCR) (Barany, PNAS 88:189-193, 1991). Индивидуальные олигонуклеотиды обычно содержат 5'- или 3'-выступающие концы для комплементарной сборки.
Альтернативные способы модификации нуклеотидной последовательности являются доступными для получения полипептидных вариантов для высокопроизводительного скрининга, например способы, которые включают в себя перетасовку генов, т.е. рекомбинацию между двумя или несколькими гомологичными нуклеотидными последовательностями, приводящую к новым нуклеотидным последовательностям, имеющим ряд нуклеотидных изменений, в сравнении с исходными нуклеотидными последовательностями. Перестановка (перетасовка) генов (также известная как перетасовка ДНК) включает в себя один или несколько циклов случайной фрагментации и повторной сборки нуклеотидных последовательностей с последующим скринингом для отбора нуклеотидных последовательностей, кодирующих полипептиды с желаемыми свойствами. Для того чтобы имела место основанная на гомологии перетасовка нуклеиновых кислот, релевантные части нуклеотидных последовательностей являются предпочтительно, по меньшей мере, на 50% идентичными, например, по меньшей мере, на 60% идентичными, более предпочтительно, по меньшей мере, на 70% идентичными, например, по меньшей мере, на 80% идентичными. Рекомбинация может выполняться in vitro или in vivo.
Примеры подходящих способов перетасовки генов in vitro описаны Stemmer et al. (1994) Proc. Natl. Acad. Sci. USA; vol.91, pp.10747-10751; Stemmer (1994), Nature, vol.370, pp.389-391; Smith (1994), Nature vol.370, pp.324-325; Zhao et al., Nat. Biotechnol. 1998, Mar; 16(3): 258-61; Zhao H. and Arnold, FB, Nucleic Acids Research, 1997, Vol.25, No.6 pp.1307-1308; Shao et al., Nucleic Acids Research 1998, Jan 15; 26(2): pp.681-83; и WO 95/17413.
Пример подходящего способа перетасовки in vivo описан в WO 97/07205. Другие способы мутагенеза последовательностей нуклеиновых кислот рекомбинацией in vitro или in vivo описаны, например, в WO 97/20078 и патенте США US 5837458. Примеры конкретных способов перетасовки включают в себя "перетасовку в семействе генов", "синтетическую перетасовку" и "рекомбинацию in silico". Перетасовка в семействе генов включает в себя подвергание семейства гомологичных генов из разных видов одному или нескольким циклам перетасовки и последующий скрининг или отбор. Способы "семейной" перетасовки описаны, например, Crameri et al. (1998), Nature, vol.391, pp.288-291; Christians et al. (1999), Nature Biotechnology, vol.17, pp.259-264; Chang et al. (1999), Nature Biotechnology, vol.17, pp.793-797; и Ness et al. (1999), Nature Biotechnology, vol.17, 893-896. Синтетическая перетасовка включает в себя обеспечение библиотек перекрывающихся синтетических олигонуклеотидов, основанных, например, на сопоставлении последовательностей представляющих интерес гомологичных генов. Синтетически генерируемые олигонуклеотиды рекомбинируют, и полученные рекомбинантные последовательности нуклеиновых кислот подвергают скринингу и, если желательно, используют для дополнительных циклов перетасовки. Способы синтетической перетасовки описаны в WO 00/42561. Перетасовка in silico относится к процедуре перетасовки ДНК, которая выполняется или моделируется с использованием компьютерной системы, посредством чего частично или почти полностью избегается необходимость физического манипулирования нуклеиновыми кислотами. Способы для перетасовки in silico описаны в WO 00/42560.
После сборки (с использованием синтеза, сайт-направленного мутагенеза или другого способа) нуклеотидную последовательность, кодирующую полипептид, вставляют в рекомбинантный вектор и функционально связывают с регуляторными последовательностями, необходимыми для экспрессии G-CSF в желаемой трансформированной клетке-хозяине.
Конечно, должно быть понятно, что не все векторы и регуляторные последовательности экспрессии функционируют одинаково хорошо для экспрессии нуклеотидной последовательности, кодирующей описанный здесь полипептид. Также не все хозяева функционируют одинаково хорошо с одной и той же системой экспрессии. Однако специалист в данной области сможет сделать выбор среди этих векторов, регуляторных последовательностей экспрессии и хозяев без чрезмерного экспериментирования. Например, в выборе вектора должен учитываться хозяин, поскольку вектор должен в нем реплицироваться или должен быть способен интегрироваться в хромосому. Копийность вектора, способность регулировать эту копийность и экспрессия любых других белков, кодируемых этим вектором, таких как маркеры для устойчивости к антибиотикам, должны также учитываться. В выборе регуляторной последовательности экспрессии, должны также учитываться множество факторов. Они включают в себя, например, относительную силу этой последовательности, ее регулируемость и ее совместимость с нуклеотидной последовательностью, кодирующей полипептид, в частности, в отношении потенциальной вторичной структуры. Хозяева должны выбираться с учетом их совместимости с выбранным вектором, токсичности продукта, кодируемого этой нуклеотидной последовательностью, их характеристик секреции, их способности правильно укладывать полипептид, их требований к ферментации или культивированию и легкости очистки продуктов, кодируемых данной нуклеотидной последовательностью.
Рекомбинантный вектор может быть автономно реплицирующимся вектором, т.е. вектором, который существует в виде внехромосомного элемента, репликация которого не зависит от репликации хромосом, например, плазмидой. Альтернативно, применим вектор, который при введении в клетку-хозяина интегрируется в геном клетки-хозяина и реплицируется вместе с хромосомой (хромосомами), в которые он был интегрирован.
Вектор предпочтительно является экспрессирующим вектором, в котором нуклеотидная последовательность, кодирующая полипептид данного изобретения, функционально связана с дополнительными сегментами, требующимися для транскрипции этой нуклеотидной последовательности. Этот вектор обычно произведен из плазмиды или вирусной ДНК. Ряд подходящих экспрессирующих векторов для экспрессии в упомянутых здесь клетках-хозяевах являются коммерчески доступными и описаны в литературе. Применимые экспрессирующие векторы для эукариотических хозяев включают в себя, например, векторы, содержащие регуляторные последовательности экспрессии из SV40, вируса бычьей папилломы, аденовируса и цитомегаловируса. Конкретными векторами являются, например, pCDNA3.1(+)/Hyg (Invitrogen, Carlsbad, CA, USA) и pCl-neo (Stratagene, La Jolla, CA, USA). Применимые экспрессирующие векторы для дрожжей включают в себя плазмиду 2μ и ее производные, вектор РОТ1 (US 4931373), вектор pJSO37, описанный в Okkels, Ann. New York Acad. Sci. 782, 202-207, 1996, и pPICZ а, В или С (Invitrogen). Полезными векторами для инсерции клеток являются pVL941, pBG311 (Cate et al., "Isolation of the Bovine and Human Genes for Mullerian Inhibiting Substance And Expression of the Human Gene In Animal Cells", Cell, 45, pp.685-98 (1986), pBluebac 4.5 и pMelbac (обе доступны из Invitrogen). Применимые экспрессирующие векторы для бактериальных хозяев включают в себя известные бактериальные плазмиды, такие как плазмиды из Е. coli, в том числе pBR322, рЕТЗа и рЕТ12а (обе доступны из Novagen Inc., WI, USA), плазмиды более широкого диапазона, такие как RP4, ДНК фагов, например многочисленные производные фага лямбда, например NM989, и другие ДНК-фаги, такие как М13 и нитчатые одноцепочечные ДНК-фаги.
Другие векторы для применения в данном изобретении включают в себя векторы, которые позволяют нуклеотидной последовательности, кодирующей полипептид, амплифицироваться в числе копий. Такие амплифицируемые векторы хорошо известны в данной области. Они включают в себя, например, векторы, способные амплифицироваться DHFR-амплификацией (см., например, Kaufman, US Pat. No. 4470461, Kaufman and Sharp, "Constuction Of A Modular Dihydrafolate Reductase cDNA Gene: Analysis Of Signals Utilized For Efficient Expression", Mol. Cell. Biol., 2, pp.1304-19 (1982)) и глутаматсинтетазной ("GS") амплификацией (см., например, US 5122464 и ЕР 338841).
Рекомбинантный вектор может дополнительно содержать последовательность ДНК, позволяющую вектору реплицироваться в рассматриваемой клетке-хозяине. Примером такой последовательности (когда клеткой-хозяина является клетка млекопитающего) является начало репликации (ориджин) SV40. Когда клеткой-хозяином является дрожжевая клетка, пригодными последовательностями, позволяющими вектору реплицироваться, являются гены репликации REP 1-3 дрожжевой плазмиды 2μ и начало репликации.
Вектор может также содержать селектируемый маркер, например ген, продукт которого восполняет дефект в клетке-хозяине, такой как ген, кодирующий дигидрофолатредуктазу (DHFR), или ген TRI Schizosaccharomyces pombe (описанный P.R.Russel, Gene 40, 1985, pp.125-130), или ген, который придает устойчивость к лекарственному средству, например ампициллину, канамицину, тетрациклину, хлорамфениколу, неомицину, гигромицину или метотрексату. Для Saccharomyces cerevisiae селектируемые маркеры включают в себя ura3 и leu2. Для нитчатых грибов селектируемые маркеры включают в себя amdS, pyrG, arcB, niaD и sC.
Термин "регуляторные последовательности" обозначает здесь все компоненты, которые являются необходимыми или выгодными для экспрессии полипептида данного изобретения. Каждая регуляторная последовательность может быть природной или чужеродной для последовательности нуклеиновой кислоты, кодирующей полипептид. Такие регуляторные последовательности включают в себя, но не ограничиваются ими, лидерную последовательность, последовательность полиаденилирования, последовательность пропептида, промотор, энхансер или расположенную слева активирующую последовательность, последовательность сигнального пептида и терминатор транскрипции. Как минимум, регуляторные последовательности включают в себя промотор.
В данном изобретении может быть использовано большое разнообразие регуляторных последовательностей экспрессии. Такие применимые регуляторные последовательности экспрессии включают в себя регуляторные последовательности экспрессии, связанные со структурными генами предыдущих экспрессирующих векторов, а также любую последовательность, о которой известно, что она регулирует экспрессию генов прокариотических или эукариотических клеток или их вирусов и их различные комбинации.
Примеры подходящих регуляторных последовательностей для управления транскрипцией в клетках млекопитающих включают в себя ранний и поздний промоторы SV40 и аденовируса, например, основной поздний промотор аденовируса 2, промотор МТ-1 (гена металлотионеина), промотор немедленно раннего гена цитомегаловируса человека (CMV), промотор фактора элонгации 1α (EF-1α) человека, минимальный промотор белка 70 теплового шока Drosophila, промотор вируса саркомы Рауса (RSV), промотор убиквитина С (UbC) человека, терминатор гормона роста человека, сигналы полиаденилирования района Elb SV40 или аденовируса и консенсусная последовательность Козака (Kozak, M.J. Mol. Biol. 1987 Aug 20; 196(4):947-50).
Для улучшения экспрессии в клетках млекопитающих синтетический интрон может быть инсертирован в 5'-нетранслируемый район нуклеотидной последовательности, кодирующей полипептид. Примером синтетического интрона является синтетический интрон из плазмиды pCI-Neo (доступный из Promega Corporation, WI, USA).
Примеры подходящих регуляторных последовательностей для управления транскрипцией в клетках насекомых включают в себя промотор полиэдрина, промотор Р10, промотор основного белка вируса полиэдроза Autographa californica, промотор немедленно раннего гена бакуловируса, промотор задержанно раннего гена бакуловируса 39К и последовательность полиаденилирования SV40. Примеры подходящих регуляторных последовательностей для применения в дрожжевых клетках-хозяевах включают в себя промоторы дрожжевой системы α-скрещивания, промотор триозофосфатизомеразы (TPI) дрожжей, промоторы из гликолитических генов или генов алкогольдегидрогеназы дрожжей, промотор ADH2-4c и индуцируемый промотор GAL. Примеры подходящих регуляторных последовательностей для применения в нитчатых грибных клетках-хозяевах включают в себя промотор и терминатор ADH3, промотор, происходящий из генов, кодирующих ТАКА-амилазу, триозофосфатизомеразу или щелочную протеазу Aspergillus oryzae, α-амилазу A. niger, глюкоамилазу А. niger или A. nidulans, ацетамидазу A. nidulans, аспартатпротеиназу или липазу Rhizomucor miehei, терминатор ТРI1 и терминатор ADH3. Примеры подходящих регуляторных последовательностей для применения в бактериальных клетках-хозяевах включают в себя промоторы lac-системы, trp-системы, ТАС- или TRC-системы и основные промоторные районы фага лямбда.
Присутствие или отсутствие сигнального пептида будет, например, зависеть от экспрессирующей клетки-хозяина, используемой для получения полипептида, который должен быть экспрессирован (является ли он внутриклеточным или внеклеточным полипептидом и желательно ли, чтобы он был секретируемым). Для применения в нитчатых грибах сигнальный пептид может быть удобным образом получен из гена, кодирующего амилазу или глюкоамилазу Aspergillus sp., гена, кодирующего липазу или протеазу Rhizomucor miehei, или липазу Humicola lanuginosa. Сигнальный пептид предпочтительно получают из гена, кодирующего ТАКА-амилазу A. oryzae, нейтральную α-амилазу A. niger, кислотоустойчивую амилазу А. niger или глюкоамилазу А. niger. Для применения в клетках насекомых сигнальный пептид может быть удобным образом произведен из гена насекомого (см. WO 90/05783), такого как гены предшественника адипокинетического гормона Lepidopteran manduca sexta (см. US 5023328), мелитина медоносной пчелы (Invitrogen), экдистероид-УДФ-гликозилтрансферазы (egt) (Murphy et al., Protein Expression and Purification 4, 349-357 (1993) или панкреатической липазы (hpl) человека (Methods in Enzymology 284, pp.262-272, 1997). Предпочтительным сигнальным пептидом для применения в клетках млекопитающих является сигнальный пептид hG-CSF или сигнальный пептид легкой цепи каппа Ig (Coloma, M (1992) J. Imm. Methods 152:89-104). Для применения в дрожжевых клетках подходящими сигнальными пептидами, как было обнаружено, являются сигнальный пептид α-фактора из S. cerevisiae (см. US 4870008), модифицированный сигнальный пептид карбоксипептидазы (см. L.A.Valls et al., Cell 48, 1987, pp.887-897), сигнальный пептид BAR1 дрожжей (см. WO 87/02670), сигнальный пептид дрожжевой аспартатпротеазы 3 (YAP3) (см. М. Egel-Mitani et al., Yeast 6, 1990, pp.127-137) и синтетическая лидерная последовательность ТА57 (WO 98/32867). Для применения в клетках Е. coli подходящим сигнальным пептидом, как было обнаружено, является сигнальный пептид ompA.
Нуклеотидная последовательность данного изобретения, кодирующая полипептид, проявляющий активность G-CSF, полученная сайт-направленным мутагенезом, синтезом, ПЦР или другими способами, может необязательно включать в себя также нуклеотидную последовательность, которая кодирует сигнальный пептид. Сигнальный пептид присутствует, когда полипептид должен секретироваться из клеток, в которых он экспрессируется. Такой сигнальный пептид, если он присутствует, должен быть пептидом, узнаваемым клеткой, выбранной для экспрессии этого полипептида. Сигнальный пептид может быть гомологичным (например, пептидом, обычно ассоциированным с hG-CSF) или гетерологичным (т.е. происходящим из другого источника, чем hG-CSF) относительно полипептида или может быть гомологичным или гетерологичным относительно клетки-хозяина, т.е. быть сигнальным пептидом, обычно экспрессируемым из клетки-хозяина, или быть сигнальным пептидом, который обычно не экспрессируется из этой клетки-хозяина. Таким образом, сигнальный пептид может быть прокариотическим, например, полученным из бактерии, например, Е. coli, или эукариотическим, например, полученным из клетки млекопитающего или клетки насекомого или дрожжей.
Любой подходящий хозяин может быть использован для получения полипептида или полипептидной части конъюгата данного изобретения, в том числе бактерии, грибы (в том числе дрожжи), клетки растений, насекомых, млекопитающих или другие подходящие клетки или клеточные линии животных, а также трансгенные животные или растения. Примеры бактериальных клеток-хозяев включают в себя грамположительные бактерии, такие как штаммы Bacillus, например В. brevis или В. subtilis, Pseudomonas или Streptomyces, или грамотрицательные бактерии, такие как штаммы Е. coli. Введение вектора в бактериальную клетку-хозяина может, например, выполняться трансформацией протопластов (см., например, Chang and Cohen, 1979, Molecular General Genetics 168: 111-115), с использованием компетентных клеток (см., например, Young and Spizizin, 1961, Journal of Bacteriology 81: 823-829 или Dubnau and Davidoff-Abelson, 1971, Journal of Molecular Biology 56: 209-221), электропорацией (см., например, Shigekawa and Dower, 1988, Biotechniques 6: 742-751), или конъюгацией (см., например, Koehler and Thorne, 1987, Journal of Bacteriology 169: 5771-5278). Примеры подходящих нитчатых грибных клеток-хозяев включают в себя штаммы Aspergillus, например A. oryzae, A. niger или A. nidulans, Fusarium или Trichoderma. Грибные клетки могут быть трансформированы способом, включающим в себя образование протопластов, трансформацию протопластов и регенерацию клеточной стенки известным per se способом. Подходящие процедуры для трансформации клеток-хозяев Aspergillus описаны в ЕР 238023 и US 5679543. Подходящие способы для трансформации видов Fusarium описаны Malardier et al., 1989, Gene 78:147-156 и WO 96/00787. Примеры подходящих дрожжевых клеток-хозяев включают в себя штаммы Saccharomyces, например S. cerevisiae, Schizosaccharomyces, Klyveromyces, Pichia, например P. pastoris или Р. methanolica, Hansenula, например Н. polymorpha или Yarrowia. Дрожжи могут быть трансформированы с использованием процедур, описанных Becker and Guarente, In Abelson, J.N. and Simon, M.I., editors. Guide to Yeast Genetics and Molecular Biology, Methods in Enzymology, Volume 194, pp.182-187, Academic Press, Inc., New York; Ito et al., 1983, Journal of Bacteriology 153: 163; Hinnen et al., 1978, Proceedings of National Academy of Sciences USA 75: 1920; и, как описано Clontech Laboratories, Inc, Palo Alto, CA, USA (в протоколе для набора системы трансформации дрожжей Yeastmaker™). Примеры подходящих клеток-хозяев насекомых включают в себя клеточную линию Lepidoptera, например, Spodoptera frugiperda (Sf9 или Sf21) или клетки Trichoplusioa ni (High Five) (US 5007214). Трансформация клеток насекомых и получение гетерологичных полипептидов могут выполняться, как описано Invitrogen. Примеры подходящих клеток-хозяев млекопитающих включают в себя клеточные линии яичника Китайского хомячка (СНО), (например, СНО-К1; АТСС CCL-61), клеточные линии зеленой мартышки (COS) (например, COS-1 (АТСС CRL-1650), COS 7 (АТСС CRL-1651)); клетки мышей (например, NS/O), клеточные линии почки детеныша хомячка (ВНК) (например, АТСС CRL-1632 или АТСС CCL-10) и клетки человека (например, НЕК 293 (АТСС CRL-1573)), а также клетки растений в культуре ткани. Дополнительные подходящие клеточные линии известны в данной области и доступны из публичных депозитариев, таких как Американская Коллекция типовых культур, Rockville, Maryland. Способы введения экзогенной ДНК в клетки-хозяина млекопитающих включают в себя опосредованную кальций-фосфатом трансфекцию, электропорацию, опосредованную ДЭАЭ-декстраном трансфекцию, опосредованную липосомами трансфекцию, вирусные векторы и способ трансфекции, описанный в Life Technologies Ltd, Paisley, UK с использованием липофектамина 2000. Эти способы хорошо известны в данной области и описаны, например, в Ausubel, F.M. et al., 1996, "Current Protocols in Molecular Biology", John Wiley and Sons, New York, USA. Культивирование клеток млекопитающих проводят в соответствии с установленными способами, например, как описано в (Animal Cell Biotechnology, Methods and Protocols, Edited by Nigel Jenkins, 1999, Human Press Inc, Totowa, New Jersey, USA и Harrison MA and Rae IF, General Techniques of Cell Culture, Cambridge University Press 1997).
В способах получения данного изобретения клетки культивируют в питательной среде, пригодной для получения полипептида, с использованием способов, известных в данной области. Например, клетка может культивироваться во встряхиваемых колбах, ферментацией малого масштаба или крупномасштабной ферментацией (в том числе непрерывной, периодической, периодической с подачей среды или ферментацией на твердой основе) в лаборатории или в промышленных ферментерах, в подходящей среде и при условиях, позволяющих полипептиду экспрессироваться и/или позволяющих выделить полипептид. Культивирование происходит в подходящей питательной среде, содержащей источники углерода и азота и неорганические соли, с использованием процедур, известных в данной области. Подходящие среды доступны из коммерческих источников или могут быть приготовлены в соответствии с опубликованными составами (например, в каталогах Американской Коллекции Типовых Культур). Если полипептид секретируется в питательную среду, полипептид может быть извлечен непосредственно из этой среды. Если полипептид не секретируется, он может быть извлечен из клеточных лизатов.
Полученный полипептид может быть извлечен способами, известными в данной области. Например, полипептид может быть извлечен из питательной среды общепринятыми процедурами, в том числе, но не только, центрифугированием, фильтрованием, экстракцией, распылительной сушкой, упариванием или осаждением.
Полипептиды могут быть очищены различными процедурами, известными в данной области, в том числе, но не только, хроматографией (например, ионообменной, аффинной, гидрофобной хроматографией, хроматографическим фокусированием и гель-фильтрацией), электрофоретическими процедурами (например, препаративным изоэлектрическим фокусированием), с использованием дифференциальной растворимости (например, осаждением сульфатом аммония), электрофорезом в ПААГ-ДСН или экстракцией (см., например, Protein Purification, J.-C. Janson and Lars Ryden, editors, VCH Publishers, New York, 1989). Конкретные способы для очистки полипептидов, проявляющих активность G-CSF, описаны D.Metcalf and N.A.Nicola in The hemopoietic colony-stimulating factors, p.50-51, Cambridge University Press (1995), C.S. Bae et al., Appl. Microbiol. Biotechnol, 52:338-344 (1999) и патенте США US 4810643.
Фармацевтическая композиция данного изобретения и ее применение
В следующем аспекте данное изобретение относится к композиции, содержащей полипептид или конъюгат, описанные здесь, и, по меньшей мере, один фармацевтически приемлемый носитель или наполнитель.
Полипептид, конъюгат или фармацевтическая композиция данного изобретения могут быть использованы для приготовления лекарственного средства для лечения заболеваний, в частности предупреждения инфекции у раковых пациентов, подвергающихся некоторым типам химиотерапии, лучевой терапии и трансплантациям костного мозга, мобилизации клеток-предшественников для сбора в трансплантациях клеток-предшественников периферической крови, лечения тяжелой хронической или относительной лейкопении, лечения пациентов с острым миелоидным лейкозом, лечения СПИДа или других связанных с иммунодефицитом заболеваний и для противогрибковой терапии, в частности для лечения системного или инвазивного кандидамикоза (кандидоза).
В другом аспекте полипептид, конъюгат или фармацевтическую композицию, в соответствии с данным изобретением, используют в способе лечения млекопитающего, имеющего общее гемопоэтическое нарушение, в том числе нарушение, возникающее от лучевой терапии или от химиотерапии, в частности лейкопении, СПИДа или других связанных с иммунодефицитом заболеваний, предусматривающем введение млекопитающему, нуждающемуся в этом, таких полипептида, конъюгата или фармацевтической композиции.
Полипептиды и конъюгаты данного изобретения будут вводиться пациентам в "терапевтически эффективной дозе", т.е. дозе, которая является достаточной для получения желаемых эффектов в отношении состояния, для которого ее вводят. Точная доза будет зависеть от подлежащего лечению нарушения и будет определяться специалистом с квалификацией в данной области с использованием известных способов. Полипептиды или конъюгаты данного изобретения могут, например, вводиться в дозе, сходной с дозой, применяемой в терапии rhG-CSF, например, Neupogen®. Предполагается, что подходящая доза конъюгата данного изобретения находится в диапазоне приблизительно 5-300 мкг/кг веса тела (в расчете на вес белковой части конъюгата), например 10-200 мкг/кг, например 25-100 мкг/кг. Специалистам в данной области должно быть очевидно, что эффективное количество полипептида, конъюгата или композиции данного изобретения зависит, inter alia, от заболевания, схемы введения дозы, от того, вводится ли полипептид, конъюгат или композиция отдельно или в сочетании с другими терапевтическими агентами, полупериода существования в сыворотке этих композиций, общего состояния здоровья пациента и частоты введения. Предпочтительно, полипептид, конъюгат, препарат или композиция данного изобретения вводится в эффективной дозе, в частности, в дозе, которая является достаточной для нормализации числа лейкоцитов, в частности нейтрофилов, в обсуждаемом пациенте. Нормализация числа лейкоцитов может быть определена простым счетом числа лейкоцитов при регулярных интервалах в соответствии с установленной практикой.
Полипептид или конъюгат данного изобретения предпочтительно вводят в виде композиции, включающей в себя один или несколько фармацевтически приемлемых носителей или наполнителей. Полипептид или конъюгат могут быть приготовлены в виде фармацевтических композиций известным per se в данной области способом для получения полипептидного фармацевтического средства, которое является достаточно стабильным при хранении и пригодно для введения людям или животным. Фармацевтическая композиция может быть приготовлена в виде разнообразных форм, в том числе в виде жидкости или геля, или лиофилизирована, или в любой другой подходящей форме. Предпочтительная форма будет зависеть от конкретного, подлежащего лечению состояния и будет очевидной для специалиста в данной области.
Таким образом, данное изобретение обеспечивает композиции и способы для лечения различных форм лейкопении. В частности, полипептид, конъюгат или композиция данного изобретения могут быть использованы для предупреждения инфекции у раковых пациентов, подвергающихся некоторым типам лучевой терапии, химиотерапии и трансплантациям костного мозга, для мобилизации клеток-предшественников для сбора в трансплантациях клеток-предшественников периферической крови, лечения тяжелой хронической или относительной лейкопении и для поддерживающей терапии пациентов с острым миелоидным лейкозом. Кроме того, полипептид, конъюгат или композиция данного изобретения могут быть использованы для лечения СПИДа или других связанных с иммунодефицитом заболеваний и для противогрибковой терапии, в частности для лечения системного или инвазивного кандидамикоза (кандидоза), и для лечения бактериальных инфекций.
Форма лекарственного средства
Полипептид или конъюгат данного изобретения может быть использован "как есть" и/или в форме его соли. Подходящие соли включают в себя, но не ограничиваются ими, соли со щелочными металлами или щелочно-земельными металлами, такие как соли натрия, калия, кальция и магния, а также, например, соли цинка. Эти соли или комплексы могут присутствовать в виде кристаллической и/или аморфной структуры.
Наполнители
"Фармацевтически приемлемые" означает носитель или наполнитель, которые в применяемых дозах и концентрациях не вызывают никаких неблагоприятных эффектов у пациентов, которым их вводят. Такие фармацевтически приемлемые носители и наполнители хорошо известны в данной области (см. Remington's Pharmaceutical Sciences, 18th edition, A.R.Gennaro, Ed., Mack Publishing Company, [1990]; Pharmaceutical Formulation Development of Peptides and Proteins, S.Frokjaer and L.Hovgaard, Eds., Taylor and Fransis [2000]; и Handbook of Pharmaceutical Excipients, 3rd edition, A.Kibbe, Ed., Pharmaceutical Press [2000]).
Смеси лекарственных средств
Фармацевтическая композиция данного изобретения может вводиться отдельно или в сочетании с другими терапевтическими агентами. Эти агенты могут быть включены в виде части одной и той же фармацевтической композиции или могут вводиться отдельно от полипептида или конъюгата данного изобретения, либо одновременно, либо по другой схеме введения. Кроме того, полипептид, конъюгат или фармацевтическая композиция данного изобретения могут использоваться в качестве адъюванта для других терапий.
Пациенты
"Пациент" для цели данного изобретения включает в себя как людей, так и других млекопитающих. Таким образом, данные способы применимы как для терапии человека, так и для ветеринарных применений.
Путь введения
Введение композиций данного изобретения может выполняться различными путями, в том числе, но не только, перорально, подкожно, внутривенно, интрацеребрально, интраназально, трансдермально, внутрибрюшинно, внутримышечно, внутрилегочно, вагинально, ректально, внутриглазно или любым другим приемлемым образом. Композиции могут вводиться непрерывно инфузией, хотя является приемлемой и болюсная инъекция, с использованием способов, хорошо известных в данной области.
Парентеральные формы
Примером фармацевтической композиции является раствор, предназначенный для парентерального введения. Хотя во многих случаях фармацевтические композиции в виде растворов обеспечивают в жидкой форме, пригодной для немедленного использования, такие парентеральные композиции могут быть также обеспечены в замороженной или лиофилизированной форме. В первом случае композиция должна оттаиваться перед использованием. Последнюю форму часто используют для повышения стабильности активного соединения, содержащегося в этой композиции при широком разнообразии условий хранения, так как специалисты с квалификацией в данной области считают, что лиофилизированные препараты обычно являются более стабильными, чем их жидкие копии. Такие лиофилизированные препараты восстанавливают перед использованием добавлением одного или нескольких подходящих фармацевтически приемлемых разбавителей, таких как стерильная вода для инъекций или стерильный физиологический солевой раствор.
В случае парентеральных препаратов их готовят для хранения в виде лиофилизированных композиций или водных растворов смешиванием соответствующим образом полипептида, имеющего желаемую степень чистоты, с одним или несколькими приемлемыми носителями, наполнителями или стабилизаторами, обычно используемыми в данной области (все они называются "наполнителями" (эксципиентами)), например буферящими агентами, стабилизирующими агентами, консервантами, изотонизирующими агентами, неионогенными детергентами, антиоксидантами и/или другими разносторонними добавками.
Буферящие агенты помогают поддерживать рН в диапазоне, который приближается к физиологическим условиям. Они обычно присутствуют в концентрации в диапазоне от приблизительно 2 мМ до приблизительно 50 мМ. Подходящие буферящие агенты для применения с данным изобретением включают в себя как органические, так и неорганические кислоты и их соли, такие как нитратные буферы (например, смесь мононатрийцитрат-динатрийцитрат, смесь лимонная кислота-тринатрийцитрат, смесь лимонная кислота-мононатрийцитрат и т.д.), сукцинатные буферы (например, смесь янтарная кислота-мононатрийсукцинат, смесь янтарная кислота-гидроксид натрия, смесь янтарная кислота-динатрийсукцинат и т.д.), тартратные буферы (например, смесь винная кислота-тартрат натрия, смесь винная кислота-тартрат калия, смесь винная кислота-гидроксид натрия и т.д.), фумаратные буферы (например, смесь фумаровая кислота-мононатрийфумарат, смесь фумаровая кислота-динатрийфумарат, смесь мононатрийфумарат-динатрийфумарат и т.д.), глюконатные буферы (например, смесь глюконовая кислота-глюконат натрия, смесь глюконовая кислота-гидроксид натрия, смесь глюконовая кислота-глюконат калия и т.д.), оксалатные буферы (например, смесь щавелевая кислота-оксалат натрия, смесь щавелевая кислота-гидроксид натрия, смесь щавелевая кислота-оксалат калия и т.д.), лактатные буферы (например, смесь молочная кислота-лактат натрия, молочная кислота-гидроксид натрия, молочная кислота-лактат калия и т.д.) и ацетатные буферы (например, смесь уксусная кислота-ацетат натрия, смесь уксусная кислота-гидроксид натрия и т.д.). Дополнительными возможностями являются фосфатные буферы, гистидиновые буферы и соли триметиламина, например Трис.
Консерванты добавляют для замедления микробного роста, и их обычно добавляют в количествах около 0,2%-1% (мас./об.). Подходящие консерванты для применения с данным изобретением включают в себя фенол, бензиловый спирт, метакрезол, метилпарабен, пропилпарабен, октадецилдиметилбензиламмонийхлорид, бензалконийгалогениды (например, бензалконийхлорид, бромид или иодид), гексаметонийхлорид, алкилпарабены, например, метил- и пропилпарабен, катехин, резорцин, циклогексанол и 3-пентанол.
Изотонизирующие агенты добавляют для гарантии изотоничности жидких композиций, и они включают в себя сахароспирты, предпочтительно трехатомные или более высокоатомные сахароспирты, такие как глицерин, эритрит, арабит, ксилит, сорбит и маннит. Многоатомные спирты могут присутствовать в количестве от 0,1% до 25% по весу, обычно 1%-5%, с учетом относительных количеств других ингредиентов.
Стабилизаторами называют широкую категорию эксципиентов, которая может по ее функции находиться в диапазоне от наполнителя до добавки, которая солюбилизирует терапевтический агент или способствует предотвращению денатурации или прилипания к стенке контейнера. Типичными стабилизаторами могут быть многоатомные сахароспирты (перечисленные выше); аминокислоты, такие как аргинин, лизин, глицин, глутамин, аспарагин, гистидин, аланин, орнитин, L-лейцин, 2-фенилаланин, глутаминовая кислота, треонин и т.д., органические сахара или сахароспирты, такие как лактоза, трегалоза, стахиоза, маннит, сорбит, ксилит, рибит, миоинозит, галактит, глицерин и т.п., в том числе циклиты, такие как инозит; полиэтиленгликоль; полимеры аминокислот; серусодержащие восстанавливающие агенты, такие как мочевина, глутатион, тиоктовая кислота, тиогликолат натрия, тиоглицерин, α-монотиоглицерин и тиосульфат натрия; низкомолекулярные полипептиды (т.е. <10 остатков); белки, такие как сывороточный альбумин человека, бычий сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры; дисахариды, такие как лактоза, мальтоза и сахароза; трисахариды, такие как рафиноза, и полисахариды, такие как декстран. Стабилизаторы обычно присутствуют в диапазоне от 0,1 до 10000 частей по весу в расчете на вес активного белка.
Неионогенные поверхностно-активные вещества или детергенты (также известные как "увлажняющие агенты") могут присутствовать для улучшения солюбилизации терапевтического агента, а также для защиты терапевтического полипептида против вызываемой перемешиванием агрегации, что позволяет также композиции подвергаться сдвигающему поверхностному натяжению без денатурации полипептида. Подходящие неионогенные поверхностно-активные вещества включают в себя полисорбаты (20, 80 и т.д.), полиоксамеры (184, 188 и т.д.), полиолы Pluronic®, простые моноэфиры полиоксиэтиленсорбитана (Твин®-20, Твин®-80 и т.д.).
Дополнительные разнообразные эксципиенты включают в себя образующие объем агенты или наполнители (например, крахмал), хелатообразователи (например, ЭДТА), антиоксиданты (например, аскорбиновую кислоту, метионин, витамин Е) и сорастворители.
Активный ингредиент может быть также заключен в микрокапсулы, приготовленные, например, способами коацервации или межфазной полимеризации, например, микрокапсулы из гидроксиметилцеллюлозы, желатина или поли(метилметацилата), в коллоидальные системы доставки лекарственных средств (например, липосомы, альбуминовые микросферы, микроэмульсии, наночастицы и нанокапсулы) или в макроэмульсии. Такие способы описаны в Remington's Pharmaceutical Sciences, supra.
Парентеральные композиции для использования при введении in vivo должны быть стерильными. Это легко выполняется, например, фильтрованием через стерильные фильтрующие мембраны.
Препараты с пролонгированным действием
Подходящие примеры препаратов с пролонгированным действием включают в себя полупроницаемые матриксы твердых гидрофобных полимеров, содержащие полипептид или конъюгат, причем эти матриксы имеют подходящую форму, например находятся в виде пленки или микрокапсулы. Примеры матриксов пролонгированного высвобождения включают в себя полиэфиры, гидрогели (например, поли(2-гидроксиэтилметакрилат) или поли(виниловый спирт)), полилактиды, сополимеры L-глутаминовой кислоты и этил-L-глутамата, недеградируемый сополимер этилена и винилового спирта, деградируемые сополимеры молочной кислоты и гликолевой кислоты, например, технология ProLease® или Lupron-Depot® (инъецируемые микросферы, состоящие из сополимера молочной кислоты и гликолевой кислоты и лейпролидацетата), и поли-D-(-)-3-гидроксимасляную кислоту. Хотя полимеры, такие как этилен-винилацетат и молочная кислота-гликолевая кислота делают возможным высвобождение молекул в течение продолжительных периодов времени, таких как до или более 100 дней, некоторые гидрогели высвобождают белки в течение более коротких периодов. Когда инкапсулированные полипептиды остаются в теле в течение продолжительного времени, они могут денатурироваться или агрегировать в результате подвергания действию влаги при 37°С, что приводит к потере биологической активности и возможным изменениям иммуногенности. Разумные стратегии могут быть придуманы для стабилизации в зависимости от участвующего в этих процессах механизма. Например, если обнаруживается, что механизмом агрегации является образование межмолекулярных связей S-S посредством тио-дисульфидного взаимообмена, стабилизация может быть достигнута модификаций сульфгидрильных остатков, лиофилизацией из кислотных растворов, регуляцией содержания влаги, использованием подходящих добавок и разработкой специфических композиций полимерного матрикса.
Легочная доставка
Композиции, пригодные для применения с использованием распылителя, либо инжекторного, либо ультразвукового, обычно будут содержать полипептид или конъюгат, растворенные в воде в концентрации, например, около 0,01 до 25 мг конъюгата на мл раствора, предпочтительно около 0,1-10 мг/мл. Композиция может также включать в себя буфер и простой сахар (например, для стабилизации белка и регуляции осмотического давления), и/или сывороточный альбумин в диапазоне концентраций от 0,1 до 10 мг/мл. Примерами буферов, которые могут быть использованы, являются ацетат, цитрат натрия и глицин. Предпочтительно, буфер будет иметь состав и молярность, подходящие для доведения раствора до рН в диапазоне 3-9. Обычно молярности буфера от 1 мМ до 50 мМ являются подходящими для этой цели. Примерами сахаров, которые могут быть использованы, являются лактоза, мальтоза, маннит, сорбит, трегалоза и ксилоза, обычно в количествах в пределах от 1% до 10% по весу композиции.
Композиция для распылителя может также содержать поверхностно-активное вещество для уменьшения или предотвращения поверхностно индуцируемой агрегации белка, вызываемой атомизацией (пульверизацией) раствора в образовании аэрозоля. Могут использоваться различные общепринятые поверхностно-активные вещества, такие как эфиры и спирты полиоксиэтилена и жирных кислот и эфиры полиоксиэтиленсорбитана и жирных кислот. Количества обычно будут в диапазоне от 0,001% до 4% по весу композиции. Особенно предпочтительным поверхностно-активным веществом для целей данного изобретения является моноолеат полиоксиэтиленсорбитана.
Конкретные композиции и способы генерирования подходящих дисперсий частиц жидкости данного изобретения описаны в WO 94/20069, US 5915378, US 5960792, US 5957124, US 5934272, US 5915378, US 5855564, US 5826570 и US 5552385, которые включены тем самым в качестве ссылки.
Композиции для применения с ингаляторным устройством с отмеренными дозами обычно будет содержать мелкоизмельченный порошок. Этот порошок может быть получен лиофилизацией и последующим размалыванием композиции жидкого конъюгата и может также содержать стабилизатор, например, человеческий сывороточный альбумин (ЧСА). Обычно добавляют более 0,5% (масса/масса) ЧСА. Кроме того, один или несколько сахаров или сахароспиртов могут быть добавлены к препарату, если это необходимо. Примеры включают в себя лактозу, мальтозу, маннит, сорбит, сорбитозу, трегалозу, ксилит и ксилозу. Количество, добавляемое к композиции, может варьироваться в диапазоне от приблизительно 0,01 до 200% (масса/масса), предпочтительно от приблизительно 1 до 50% присутствующего конъюгата. Затем такие композиции лиофилизируют и перемалывают до желаемого размера частиц.
Затем частицы правильного размера суспендируют в пропелленте с использованием поверхностно-активного вещества. Пропеллентом может быть любой общепринятый материал, используемый для этой цели, такой как хлорфторуглерод, гидрохлорфторуглерод, гидрофторуглерод или углеводород, в том числе трихлорфторметан, дихлордифторметан, дихлортетрафторэтанол и 1,1,1,2-тетрафторэтан или их комбинации. Подходящие поверхностно-активные вещества включают в себя сорбитантриолеат и лецитин сои. Олеиновая кислота может быть также применима в качестве поверхностно-активного вещества. Затем эту смесь загружают в устройство доставки. Примером коммерчески доступного ингалятора с отмеренными дозами, пригодного для применения в данном изобретении, является ингалятор с отмеренными дозами Ventolin, изготовляемый Glaxo Inc., Research Triangle Park, N.C.
Композиции для порошковых ингаляторов будут содержать тонкоизмельченный сухой порошок, содержащий конъюгат, и может также включать в себя образующий объем агент, такой как лактоза, сорбит, сахароза или маннит в количествах, которые облегчают рассеивание порошка из устройства, например 50%-90% в расчете на вес композиции. Частицы порошка будут иметь аэродинамические свойства в легком, соответствующие частицам с плотностью около 1 г/см2, имеющим медиану диаметра менее 10 микрометров, предпочтительно между 0,5 и 5 микрометрами, наиболее предпочтительно, между 1,5 и 3,5 микрометрами. Примером порошкового ингалятора, пригодного для использования в соответствии с данным изобретением, является порошковый ингалятор Spinhaler, изготовляемый Fisons Corp., Bedford, Mass.
Порошки для этих устройств могут быть получены и/или доставлены способами, описанными в патентах США US 5997848, US 5993783, US 5985248, US 5976574, US 5922354, US 5785049 и US 5654007.
Механические устройства, сконструированные для легочной доставки терапевтических продуктов, включают в себя, но не ограничиваются ими, распылители, ингаляторы с отмеренными дозами и порошковые ингаляторы, причем все они знакомы специалистам в данной области. Конкретными примерами коммерчески доступных устройств, пригодных для практики данного изобретения, являются распылитель Ultravent, изготовляемый Malinckrodt, Inc., St. Louis, Missouri; распылитель Acorn II, изготовляемый Marquest Medical Products, Englewood, Colorado; распылитель с отмеренными дозами Ventolin, изготовляемый Glaxo Inc., Research Triangle Park, North Carolina; порошковый ингалятор Spinhaler, изготовляемый Fisons Corp., Bedford, Massachusetts; устройство "постоянного облака" Inhale Therapeunic Systems, San Carlos, California; ингалятор AIR, изготовляемый Alkermes, Cambridge, Massachusetts; и система легочной доставки лекарственного средства AIR, изготовляемая Aradigm Corporation, Hayward, California.
В изобретении раскрывается также применение нуклеотидной последовательности, кодирующей полипептид данного изобретения, в генотерапии. В частности, представляет интерес применение нуклеотидной последовательности, кодирующей полипептид, как описано в разделе выше, озаглавленном "Конъюгат изобретения, в котором неполипептидная часть является углеводной частью". Таким образом, гликозилирование полипептидов достигается во время курса генотерапии, т.е. после экспрессии нуклеотидной последовательности в теле человека.
Рассматриваемые применения в генотерапии включают в себя лечение заболеваний, когда ожидается, что этот полипептид будет обеспечивать эффективную терапию. Они включают в себя лечение различных форм лейкопении, в частности предупреждение инфекции у раковых пациентов, подвергающихся определенным типам химиотерапии и трансплантации костного мозга, мобилизации клеток-предшественников для сбора в трансплантациях клеток-предшественников периферической крови, для лечения тяжелой хронической или относительной лейкопении, независимо от причины, для терапии пациентов с острым миелоидным лейкозом, для лечения СПИДа или других иммунодефицитных заболеваний.
Локальная доставка G-CSF с использованием генотерапии может обеспечивать терапевтический агент в зоне-мишени с избеганием проблем потенциальной токсичности, связанной с неспецифическим введением.
Далее раскрываются методологии генотерапии как in vitro, так и in vivo.
Известны несколько способов переноса потенциально терапевтических генов для определенных клеточных популяций. В качестве дополнительной ссылки см., например, Mulligan "The Basic Science Of Gene Therapy", Science, 260, pp.926-31 (1993). Эти способы включают в себя:
Прямой перенос генов, например, как описано Wolff et al., "Direct Gene transfer Into Mouse Muscle In vivo", Science 247, pp.1465-68 (1990);
Опосредованный липосомами перенос ДНК, например, как описано Caplen et al., "Liposome-mediated CFTR Gene Transfer to the Nasal Epithelium Of Patients With Cystic Fibrosis" Nature Med., 3, pp.39-46 (1955); Crystal, "The Gene As A Drug", Nature Med., 1, pp.15-17 (1995); Gao and Huang, "A Novel Cationic Liposome Reagent For Efficient Transfection of Mammalian Cells", Biochem. Biophys. Res. Commun., 179, pp.280-85 (1991);
Опосредованный ретровирусом перенос ДНК, например, как описано Кау et al., "In vivo Gene Therapy of Hemophilia В: Sustained Partial Correction In Factor IX-Deficient Dogs", Science, 262, pp.117-19 (1993); Anderson, "Human Gene Therapy", Science, 256, pp.808-13 (1992);
Опосредованный ДНК-вирусом перенос ДНК. Такие ДНК-вирусы включают в себя аденовирусы (предпочтительно векторы на основе Ad-2 или AD-5), вирусы герпеса (предпочтительно векторы на основе вируса простого герпеса) и парвовирусы (предпочтительно "дефектные" и неавтономные векторы на основе парвовируса, более предпочтительно, векторы на основе аденоассоциированного вируса, наиболее предпочтительно векторы на основе AAV-2). См., например, Ali et al., "The Use Of DNA Viruses as Vectors for Gene Therapy", Gene Therapy, 1, pp.367-84 (1994); патент США US 4797-368 и US 5139941.
Далее данное изобретение описывается в неограничительных примерах ниже.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1: Полупериод существования in vivo rhG-CSF (Neupogen®) и SPA-PEG 5000-конъюгированного hG-CSF K16R K34R K40R Q70K Q90K Q120K.
Фигура 2: Биологические активности in vivo rhG-CSF (Neupogen®), SPA-PEG 5000-конъюгированного hG-CSF K16R K34R K40R Q70K Q120K и SPA-PEG 5000-конъюгированного hG-CSF K16R K34R K40R Q70K Q90K Q120K.
Фигура 3: Биологические активности in vivo rhG-CSF (Neupogen®), SPA-PEG 12000-конъюгированного hG-CSF K16R K34R K40R и различных доз SPA-PEG 5000-конъюгированного hG-CSF K16R K34R K40R Q70K Q90K Q120K.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Прилагаемый список последовательностей содержит следующие последовательности:
SEQ ID NO:1: Аминокислотная последовательность G-CSF человека.
SEQ ID NO:2: Синтетическая последовательность ДНК, кодирующая G-CSF человека, с использованием кодонов, оптимизированных для экспрессии в Е. coli.
SEQ ID NO:3: Аминокислотная последовательность сигнальной последовательности OmpA.
SEQ ID NO:4: Синтетическая последовательность ДНК, кодирующая сигнальную последовательность OmpA.
SEQ ID NO:5: Синтетическая гистидиновая метка.
SEQ ID NO:6: Синтетическая последовательность ДНК, кодирующая гистидиновую метку SEQ ID NO:5.
SEQ ID NO:7: Аминокислотная последовательность сигнального пептида G-CSF человека.
SEQ ID NO:8: Синтетическая последовательность ДНК, кодирующая G-CSF человека, в том числе сигнальный пептид SEQ ID NO:7, с использованием кодонов, оптимизированных для экспрессии в клетках СНО.
МАТЕРИАЛЫ И СПОСОБЫ
Способы, используемые для определения аминокислот, которые должны быть амплифицированы
Доступная площадь поверхности (ASA)
3D (трехмерный) ансамбль из 10 структур, определенных ЯМР-спектроскопией (Zink et al. (1994) Biochemistry 33: 8453-8463), является доступным из банка данных белков Protein Data Bank (PDB) (www.rcsb/org/pdb/). Эта информация может быть введена в компьютерную программу Access (В.Lee and F.M.Richards, J.Mol. Biol. 55: 379-400 (1971)) version 2 (© 1983 Yale University) и использована для вычисления доступной площади поверхности (ASA) индивидуальных атомов в этой структуре. Этот способ обычно использует размер зонда 1,4 Å и определяет доступную площадь поверхности (ASA) как площадь, образуемую центром этого зонда. Перед этим расчетом все молекулы воды и все атомы водорода должны быть удалены из системы координат, как и другие атомы, не связанные непосредственно с этим белком.
Парциальная ASA боковой цепи
Парциальную ASA атомов боковой цепи вычисляют делением суммы ASA атомов в боковой цепи на величину, представляющую ASA атомов боковой цепи этого типа остатка в растянутом трипептиде ALA-x-ALA. См. Hubbard, Campbell and Thornton (1991) J. Mol. Biol. 220, 507-530. Для этого примера атом СА рассматривается как часть боковой цепи остатков глицина, но не в отношении остальных остатков. Величины в следующей таблице используются в качестве стандарта 100% ASA для боковой цепи:
Остатки, не детектируемые в этой структуре, определяют как имеющие 100% экспонирование, так как считают, что они находятся в гибких районах.
Определение расстояний между атомами:
Расстояние между атомами наиболее легко определить с использованием программного обеспечения молекулярной графики, например, Insight® v. 98.0, MSI INC.
Общие соображения относительно аминокислотных остатков, которые должны быть модифицированы
Как объяснялось выше, аминокислотные остатки, которые подлежат модификации, в соответствии с данным изобретением, предпочтительно являются аминокислотными остатками, боковые цепи которых экспонированы (открыты) на поверхности, в частности остатки, имеющие приблизительно более 25% боковой цепи, экспонированной на поверхности молекулы. Другим соображением является то, что остатки, локализованные в пограничных поверхностях рецепторов, предпочтительно исключаются таким образом, чтобы избежать или, по меньшей мере, минимизировать взаимодействие со связыванием или активацией рецепторов. Следующим соображением является то, что остатки, которые находятся меньше чем на 10 Å, от ближайшего Lys (Glu, Asp) CB-CB (CA для Gly), должны быть также исключены. Наконец, предпочтительными положениями для модификации являются, в частности, положения, которые имеют гидрофильный и/или заряженный остаток, т.е. Asp, Asn, Glu, Gln, Arg, His, Tyr, Ser и Thr, причем положения, которые имеют остаток аргинина, являются особенно предпочтительными.
Идентификация аминокислотных остатков G-CSF для модификации
Приведенная ниже информация иллюстрирует факторы, которые обычно должны учитываться при идентификации аминокислотных остатков, которые должны быть модифицированы, в соответствии с данным изобретением.
Трехмерные структуры сообщались для G-CSF человека при использовании рентгеновской кристаллографии (Hill et al. (1993) Proc. Natl. Acad. Sci. USA 90: 5167-5171) и ЯМР-спектроскопии (Zink et al. (1994) Biochemistry 33: 8453-8463). Как упоминалось выше, Aritomi et al. (Nature 401:713-717, 1999), идентифицировали следующие остатки hG-CSF как являющиеся частью рецепторсвязывающих граничных поверхностей: G4, Р5, А6, S7, S8, L9, Р10, Q11, S12, L15, К16, Е19, Q20, L108, D109, D112, Т115, Т116, Q119, Е122, Е123 и L124. Таким образом, хотя можно модифицировать эти остатки, предпочтительно, чтобы эти остатки были исключены из модификации.
С использованием 10 ЯМР-структур G-CSF, идентифицированных Zink et al. (1994) в качестве входных структур, после вычисления средней ASA боковой цепи, следующие остатки были идентифицированы как имеющие более чем 25% ASA: М0, Т1, Р2, L3, G4, Р5, А6, S7, 38, L9, Р10, Q11, S12, F13, L14, L15, К16, С17, Е19, Q20, V21, R22, К23, Q25, G26, D27, А29, А30, Е33, К34, С36, А37, Т38, Y39, К40, L41, Н43, Р44, Е45, Е46, V48, L49, L50, Н52, S53, L54, I56, Р57, Р60, L61, S62, S63, Р65, S66, Q67, А68, L69, Q70, L71, А72, G73, С74, 376, Q77, L78, S80, F83, Q86, G87, Q90, Е93, G94, S96, Р97, Е98, L99, G100, P101, T102, D104, T105, Q107, L108, D109, A111, D112, F113, T115, T116, W118, Q119, Q120, M121, E122, E123, L124, M126, A127, P128, A129, L130, Q131, P132, T133, Q134, G135, A136, M137, P138, A139, A141, S142, A143, F144, Q145, R146, R147, S155, H156, Q158, S159, L161, E162, V163, S164, Y165, R166, V167, L168, R169, H170, L171, A172, Q173, P174.
Подобным образом, следующие остатки имеют более чем 50% ASA: М0, Т1, Р2, L3, G4, Р5, А6, S7, S8, L9, Р10, Q11, S12, F13, L14, L15, К16, С17, Е19, Q20, R22, К23, G26, D27, А30, Е33, К34, Т38, К40, L41, Н43, Р44, Е45, Е46, L49, L50, S53, Р57, Р60, L61, S62, S63, Р65, S66, Q67, А68, L69, Q70, L71, А72, G73, S80, F83, Q90, G94, Р97, Е98, P101, D104, T105, L108, D112, F113, T115, T116, Q119, Q120, E122, E123, L124, M126, P128, A129, L130, Q131, P132, T133, Q134, G135, A136, A139, A141, S142, A143, F144, R147, S155, S159, E162, R166, V167, R169, H170, L171, A172, Q173, P174.
Программу молекулярной графики Insight® v. 98.0 использовали для определения остатков, имеющих их атом СВ (СА в случае глицина) на расстоянии более чем 15 Å от ближайшей аминогруппы, определяемой как NZ-атомы лизина и N-атом N-концевого остатка Т1. Следующий перечень включает в себя остатки, которые удовлетворяют этому критерию в, по меньшей мере, 10 ЯМР-структурах. G4, Р5, А6, S7, S8, L9, Р10, Q11, L14, L15, L18, V21, R22, Q25, G26, D27, G28, А29, Q32, L35, С36, Т38, Y39, С42, Н43, Р44, Е45, Е46, L47, V48, L49, L50, G51, Н52, S53, L54, G55, I56, Р57, W58, А59, Р60, L61, S62, S63, С64, Р65, S66, Q67, А68, L69, Q70, L71, А72, G73, С74, L75, S76, Q77, L78, Н79, S80, G81, L82, F83, L84, Y85, Q86, G87, L88, L89, Q90, А91, L92, Е93, G94, I95, S96, Р97, Е98, L99, G100, P101, T102, L103, D104, T105, L106, Q107, L108, D109, V110, A111, D112, F113, A114, T115, T116, I117, W118, Q119, Q120, M121, E122, E123, L124, G125, M126, A127, P128, A129, L130, Q131, P132, T133, Q134, G135, A136, M137, P138, A139, F140, A141, S142, A143, F144, Q145, R146, R147, A148, G149, G150, V151, L152, V153, A154, S155, H156, L157, Q158, S159, F160, L161, E162, V163, S164, Y165, R166, V167, L168, R169, H170, L171, A172, Q173, P174.
Программу Insight® v. 98.0 использовали подобным образом для определения остатков, имеющих их СВ атом (СА атом в случае глицина) на расстоянии более чем 10 Å от ближайшей кислотной группы, определяемой как атомы CG аспарагиновой кислоты, атомы CD глутаминовой кислоты и атом С С-концевого остатка Р174. Следующий перечень включает в себя остатки, которые удовлетворяют этому критерию в, по меньшей мере, одной из 10 ЯМР-структур. М0, T1, P2, L3, G4, Р5, А6, S7, S8, L9, Р10, Q11, S12, F13, L14, Т38, Y39, К40, L41, С42, L50, G51, Н52, S53, L54, G55, I56, Р57, W58, А59, Р60, L61, S62, S63, С64, Р65, S66, Q67, А68, L69, Q70, L71, А72, G73, С74, L75, S76, Q77, L78, Н79, S80, G81, L82, F83, L84, Y85, Q86, G87, L88, I117, М126, А127, Р128, А129, L130, Q131, Р132, Т133, Q134, G135, А136, М137, Р138, А139, F140, А141, S142, А143, F144, Q145, R146, R147, А148, G149, G150, V151, L152, V153, А154, S155, Н156, L157, V167, L168, R169, Н170, L171.
Комбинированием и сравнением приведенных выше перечней можно отобрать отдельные аминокислотные остатки для модификации, чтобы получить перечень, содержащий ограниченное число аминокислотных остатков, модификация которых в конкретном полипептиде G-CSF способна привести к желаемым свойствам.
Способы ПЭГилирования hG-CSF и его вариантов
ПЭГилирование hG-CSF и его вариантов в растворе
G-CSF человека и его варианты ПЭГилируют при концентрации 250 мкг/мл в 50 мМ фосфате натрия, 100 мМ NaCl, pH 8,5. Молярный избыток ПЭГ является 100-кратным относительно сайтов ПЭГилирования на белке. Реакционную смесь помещают в термосмеситель на 30 минут при 37°С и при 1200 об/мин. Спустя 30 минут реакцию гасят добавлением молярного избытка глицина.
Катионообменную хроматографию применяют для удаления избытка ПЭГ и других побочных продуктов из реакционной смеси. Реакционную смесь ПЭГилирования разбавляют 20 мМ цитратом натрия pH 2,5, пока ионная сила меньше 7 мС/см. pH доводят до 2,5 при помощи 5 н. HCl. Смесь наносят на SP-сефарозную FF-колонку, уравновешенную 20 мМ цитратом натрия pH 2,5. Несвязанный материал вымывают из колонки 4 колоночными объемами буфера для уравновешивания. ПЭГилированный белок элюируют в трех колоночных объемах добавлением 20 мМ цитрата натрия, 750 мМ хлорида натрия. Чистый ПЭГилированный G-CSF концентрируют и смену буфера выполняют с использованием концентрационных устройств VivaSpin, отсечение молекулярной массы (mwco): 10 кДа.
ПЭГилирование в микропланшетах для титрования меченого полипептида с G-CSF-активностью
Полипептид, проявляющий G-CSF-активность, экспрессируют с подходящей меткой, например, любой из меток, приведенных в качестве примеров в общем описании выше, и культуральный бульон переносят в одну или несколько лунок микропланшета для титрования, способного иммобилизировать меченый полипептид. Когда меткой является Met-Lys-His-Gln-His-Gln-His-Gln-His-Gln-His-Gln-His-Gln-Gln, может быть использован микропланшет никель-нитрилотриуксусная кислота (Ni-NTA) HisSorb, коммерчески доступный из QIAGEN.
После иммобилизации меченого полипептида на этом микропланшете для титрования лунки промывают буфером, пригодным для связывания и последующего ПЭГилирования с последующей инкубацией лунок с выбранным активированным ПЭГ. В качестве примера использовали M-SPA-5000 из Shearwater Polymers. Молярное отношение активированного ПЭГ к полипептиду должно быть оптимизированным, но обычно будет больше чем 10:1, например, до приблизительно 100:1 или выше. После подходящего времени реакции при температуре окружающей среды, обычно около 1 часа, реакцию останавливают удалением раствора активированного ПЭГ. Конъюгированный белок элюируют из планшета инкубацией с подходящим буфером. Подходящие буферы для элюции могут содержать имидазол, избыток NTA или другого хелатообразователя. Конъюгированный белок анализируют на биологическую активность и иммуногенность соответствующим образом. Метка может быть необязательно отщеплена с использованием способа, известного в данной области, например с использованием диаминопептидазы. Gln в положении - 1 может быть превращен в пироглутамил с GCT (глутамилциклотрансферазой) и в конце концов отщеплен при помощи PGAP (пироглутамиламинопептидазы) с получением немеченого белка. Способ включает в себя несколько стадий аффинной хроматографии с хелатом металла. Альтернативно, меченый полипептид может быть конъюгирован.
ПЭГилирование полипептида, проявляющего hG-CSF-активность и имеющего блокированный сайт связывания рецептора
Для оптимизации ПЭГилирования hG-CSF по способу исключения ПЭГилирования остатков лизина, участвующих в узнавании рецептора, был разработан следующий способ:
Очищенный hG-CSF получают, как описано в примере 3. Гомодимерный комплекс, состоящий из полипептида hG-CSF и растворимого домена G-CSF-рецептора в стехиометрии 2:2, образуется в забуференном фосфатом солевом растворе (ЗФР-буфере) при рН 7. Концентрация полипептида hG-CSF равна приблизительно 20 мкг/мл или 1 мкМ, и рецептор присутствует в эквимолярной концентрации.
M-SPA-5000 из Shearwater Polymers, Inc. добавляют при 3 различных уровнях концентрации, соответствующих 5-, 20- и 100-молярному избытку полипептида hG-CSF. Время реакции равно 30 мин при комнатной температуре. После периода реакции 30 мин рН реакционной смеси доводят до рН 2,0 и реакционную смесь наносят на колонку Vydac C18 и элюируют градиентом ацетонитрила, по существу, как описано (Utsumi et al., J. Biochem., Vol.101, 1199-1208 (1987). Альтернативно, может быть использован градиент изопропанола.
Фракции анализируют с использованием первичного анализа-скрининга, описанного здесь, и активный ПЭГилированный полипептид hG-CSF, полученный этим способом, хранят при -80°С в ЗФР, рН 7, содержащем 1 мг/мл человеческого сывороточного альбумина (ЧСА).
Способы, используемые для характеристики конъюгированного и неконъюгированного hG-CSF и его вариантов
Определение молекулярного размера hG-CSF и его вариантов
Молекулярную массу конъюгированного или неконъюгированного hG-CSF или его вариантов определяют при помощи электрофореза в ДСН-ПААГ, гель-фильтрации, лазерной десорбционной масс-спектрометрии с использованием матрикса или равновесным центрифугированием.
Определение концентрации полипептида
Концентрация полипептида может быть измерена при помощи измерений оптической плотности при 280 нм, твердофазным иммуноферментным анализом (ELISA), радиоиммуноанализом (РИА) или другими подобными способами иммунодетектирования, хорошо известными в данной области. Кроме того, концентрация полипептида в пробе может быть измерена прибором Biacore® с использованием чипа Biacore®, покрытого антителом, специфическим для данного полипептида.
Такое антитело может быть связано ковалентно с чипом Biacore® различными химическими способами. Альтернативно, антитело может быть связано нековалентно, например, с использованием антитела, специфического для Fc-белка антитела против полипептида. Fc-специфическое антитело сначала связывают ковалентно с чипом. Затем антитело против полипептида протекает через чип и связывается с первым антителом непосредственно. Кроме того, биотинилированные антитела могут быть иммобилизованы при помощи покрытой стрептавидином поверхности (например, сенсорный чип Biacore Sensor Chip SA®) (Real-Time Analysis of Biomolecular Interactions, Nagata and Handa (Eds.), 2000, Springer Verlag, Tokyo; Biacore 2000 Instrument Handbook, 1999, Biacore AB).
При протекании пробы через чип полипептид будет связываться с антителом покрытия и увеличение в массе может быть измерено. С использованием препарата полипептида в известной концентрации можно построить стандартную кривую и затем можно определять концентрацию полипептида в пробе. После каждой инъекции пробы сенсорный чип регенерируют при помощи подходящего элюента (например, буфера с низким рН), который удаляет связанный аналит.
Обычно применяемые антитела должны быть моноклональными антителами, индуцированными против полипептида дикого типа. Введение мутаций или другие манипуляции с полипептидом дикого типа (экстра-гликозилирования или конъюгации с полимером) могут изменять узнавание такими антителами. Кроме того, такие манипуляции, которые приводят к появлению увеличенной молекулярной массы полипептида, будут приводить к увеличенному сигналу плазменного резонанса. Поэтому необходимо построение стандартной кривой для каждой тестируемой молекулы.
Способы, применяемые для определения активности in vitro и in vivo конъюгированного и неконъюгированного hG-CSF и его вариантов
Первичный анализ 1 - анализ активности G-CSF in vitro
Пролиферация мышиной клеточной линии NFS-60 (полученной от доктора J. Ihle, St. Jude Children's Research Hospital, Tennesee, USA) зависит от присутствия активного G-CSF в среде для выращивания. Таким образом, биологическая активность in vitro hG-CSF и его вариантов может быть определена измерением числа делящихся клеток NFS-60 после добавления пробы G-CSF к среде для выращивания с последующим инкубированием на протяжении фиксированного периода времени.
Клетки NFS-60 поддерживают в среде DME Исков, содержащей 10% масса/масса ФТС (фетальную телячью сыворотку), 1% масса/масса Пен/Стреп, 10 мкг/л hG-CSF и 2 мМ Глутамакс. Перед взятием проб клетки промывают дважды в среде для выращивания без hG-CSF и разбавляют до концентраци 2,2×105 клеток на мл. 100 мкл клеточной суспензии добавляют в каждую лунку 96-луночного микропланшета (Corning).
Пробы, содержащие конъюгированный или неконъюгированный G-CSF или его варианты, разбавляют до концентраций между 1,1×10-6 М и 1,1×10-13 М в среде для выращивания. 10 мкл каждой пробы добавляют в 3 лунки, содержащие клетки NFS-60. Контроль, состоящий из 10 мкл среды для выращивания клеток млекопитающих, добавляют в 8 лунок на каждом микропланшете. Клетки инкубируют в течение 48 часов (37°С, 5% СО2), и число делящихся клеток в каждой лунке количественно определяют с использованием агента пролиферации клеток WST-1 (Roche Diagnostics GmbH, Mannheim, Germany). 0,01 мл WST-1 добавляют в лунки с последующим инкубированием в течение 150 мин при 37°С в атмосфере воздуха с 5% СО2. Расщепление тетразолиевой соли WST-1 митохондриальными дегидрогеназами в жизнеспособных клетках приводит к образованию формазана, который определяют количественно измерением поглощения при 450 нм. Таким образом количественно определяют число жизнеспособных клеток в каждой лунке.
На основе этих измерений рассчитывают кривые доза-ответ для каждой конъюгированной и неконъюгированной молекулы G-CSF или ее вариантов, после чего может быть определена величина ЕС50 для каждой молекулы. Эта величина равна количеству активного белка G-CSF, которое необходимо для получения 50% максимальной активности пролиферации неконъюгированного G-CSF человека. Таким образом, величина ЕС50 является прямым измерением активности in vitro конкретного белка.
Первичный анализ 2 - анализ активности G-CSF in vitro
Линию BaF3 мышиных гемопоэтических клеток трансфицируют плазмидой, несущей G-CSF-рецептор человека и промотор регулятора транскрипции, fos, впереди репортерного гена люциферазы. При стимуляции такой клеточной линии пробой G-CSF ряд внутриклеточных реакций приводит к стимуляции экспрессии fos и, следовательно, к экспрессии люциферазы. Эту стимуляцию подвергают мониторингу с использованием системы анализа люциферазы Steady-Glo™ (Promega, Cat. No. E2510), посредством чего может быть количественно определена активность in vitro пробы G-CSF.
Клетки BaF3/hGCSF-R/pfos-lux поддерживают при 37°С в увлажненной атмосфере 5% СО2 в полной культуральной среде (RPMI-1640/HEPES (Gibco/BRL, Cat. No. 22400), 10% ФТС (Hyclone, characterized), 1x пенициллин/стрептомицин (Gibco/BRL, Cat. No. 15140-122), 1x L-глутамин (Gibco/BRL, Cat. No. 25030-081), 10% кондиционированная среда WEHI-3 (источник muIL-3)) и выращивают до плотности 5×105 клеток/мл (конфлюентных). Клетки пересевают при приблизительно 2×104 клеток/мл каждые 2-3 дня.
За один день перед анализом клетки логарифмической фазы роста ресуспендируют при 2×105 клеток/мл в минимальной среде (DMEM/F-12 (Gibco BRL, Cat. No. 11039), 1% БСА (Sigma, Cat. No. 25030-081), 1× пенициллин/стрептомицин (Gibco/BRL, Cat. No. 15140-122), 1× L-глутамин (Gibco/BRL, Cat. No. 25030-081), 0,1% кондиционированная среда WEHI-3)) и дают голодать в течение 20 часов. Эти клетки промывают дважды ЗФР и испытывают на жизнеспособность с использованием окрашивания на жизнеспособность Трипановым синим. Клетки ресуспендируют в среде для анализа (RPMI-1640 (без фенолового красного, Gibco/BRL, Cat. No. 11835), 25 мМ HEPES, 1% БСА (Sigma, Cat. No. A3675), 1x пенициллин/стрептомицин (Gibco/BRL, Cat. No. 15140-122), 1x L-глутамин (Gibco/BRL, Cat. No. 25030-081) при 4×106 клеток/мл) и аликвоты 50 мкл добавляют в каждую лунку 96-луночного микропланшета (Corning). Пробы, содержащие конъюгированный или неконъюгированный G-CSF или его варианты, разбавляют до концентраций между 1,1×10-7 M и 1,1×10-12 М в среде для анализа. 50 мкл каждой пробы добавляют в 3 лунки, содержащие клетки BaF3/hGCSF-R/pfos-lux. Негативный контроль, состоящий из 50 мкл среды, добавляют в 8 лунок на каждом микропланшете. Планшеты медленно перемешивают и инкубируют в течение 2 часов при 37°С. Люциферазную активность измеряют согласно следующему протоколу Promega Steady-Glo™ (Система анализа люциферазы Promega Steady-Glo™, Cat. No. E2510). 100 мкл субстрата добавляют на лунку с последующим медленным перемешиванием. Люминесценцию измеряют на люминометре TopCount (Packard) в режиме SPC (счета отдельных фотонов).
На основе этих измерений рассчитывают кривые доза-ответ для каждой конъюгированной и неконъюгированной молекулы G-CSF или его вариантов, после чего может быть определена величина ЕС50 для каждой молекулы.
Вторичный анализ - аффинность связывания G-CSF или его вариантов с hG-CSF-рецептором
Связывание rhG-CSF или его вариантов с hG-CSF-рецептором исследуют с использованием стандартных анализов связывания. Рецепторы могут быть очищенными внеклеточными доменами рецепторов, рецепторами, связанными с очищенными мембранами клеточной цитоплазмы или целыми клетками, - источниками клеток являются либо клеточные линии, которым присуща экспрессия G-CSF-рецепторов (например, NFS-60), либо клетки, трансфицированные кДНК, кодирующей рецепторы. Способность rhG-CSF или его вариантов конкурировать за сайты связывания с природным G-CSF анализируют инкубированием с меченым G-CSF-аналогом, например биотинилированным hG-CSF или радиоиодинированным hG-CSF. Пример такого анализа описан Yamasaki et al. (Drugs. Exptl. Clin. Res. 24:191-196 (1998)).
Внеклеточные домены G-CSF-рецептора могут быть необязательно связаны с Fc и иммобилизованы в 96-луночных планшетах. Затем добавляют rhG-CSF или его варианты и их связывание детектируют с использованием либо специфических анти-hG-CSF-антител, либо биотинилированного или радиоиодинированного hG-CSF.
Измерение полупериода существования in vivo конъюгироваиного и неконъюгированного rhG-CSF и его вариантов
Важным аспектом данного изобретения является пролонгированный биологический полупериод существования, который получают конструированием hG-CSF с конъюгацией или без конъюгации полипептида с полимерной частью молекулы. Быстрое уменьшение сывороточных концентраций hG-CSF сделало важным оценку биологических реакций на обработку конъюгированным или неконъюгированным hG-CSF и его вариантами. Предпочтительно, конъюгированный и неконъюгированный hG-CSF и его варианты данного изобретения имеют пролонгированные сывороточные полупериоды существования также после введения i.v., что позволяет измерять их, например, по способу ELISA или при помощи анализа первичного скрининга. Измерение биологического полупериода существования in vivo проводили, как описано ниже.
Использовали самцов крыс Sprague-Dawley (в возрасте 7 недель). В день введения измеряли вес животных (280-310 г/животное). 100 мкг на кг веса тела проб неконъюгированного и конъюгированного hG-CSF инъецировали по отдельности внутривенно в хвостовую вену трех крыс. На 1 минуте, 30 минутах, 1, 2, 4, 6 и 24 часе после инъекции брали 500 мкл крови из глаз каждой крысы при анестезии CO2. Пробы крови хранили при комнатной температуре в течение 1,5 ч с последующим выделением сыворотки центрифугированием (4°С, 18000хg в течение 5 мин). Пробы сыворотки хранили при -80°С до дня анализа. Количество активного G-CSF в пробах сыворотки определяли при помощи анализов активности G-CSF in vitro (см. первичные анализы 1 и 2) после оттаивания проб на льду.
Другой пример анализа для измерения полупериода существования in vivo G-CSF или его вариантов описан в патенте США US 5824778, содержание которого тем самым включено в качестве ссылки.
Измерение биологической активности in vivo конъюгированного и неконъюгированного hG-CSF и его вариантов
Измерение биологических эффектов hG-CSF в крысах SPF Sprague-Dawley (купленных в М & В A/S, Denmark) используют для оценки биологической эффективности конъюгированного и неконъюгированного G-CSF и его вариантов.
В день прибытия крыс случайным образом распределяют на группы из 6 животных. Животных акклиматизируют в течение 7 дней, причем индивидов в плохом состоянии или с крайними весами исключают из исследования. Диапазон веса крыс в начале периода акклиматизации был 250-270 г.
В день введения крыс подвергают голоданию в течение 16 часов с последующей подкожной инъекцией 100 мкг на кг веса тела hG-CSF или его варианта. Каждую пробу hG-CSF инъецируют группе из 6 отобранных крыс. Пробы крови 300 мкл ЭДТА-стабилизированной крови берут из хвостовой вены крыс перед введением и на 6, 12, 24, 36, 48, 72, 96, 120 и 144 ч после введения доз. Пробы крови анализируют на следующие гомологические параметры: гемоглобин, число эритроцитов, гематокрит, средний объем клеток, среднюю концентрацию клеточного гемоглобина, средний клеточный гемоглобин, число лейкоцитов, лейкоцитарную формулу крови (нейтрофилы, лимфоциты, эозинофилы, базофилы, моноциты). На основе этих измерений оценивают биологическую эффективность конъюгированного и неконъюгированного hG-CSF и его вариантов.
Дополнительные примеры анализа для измерения биологической активности in vivo hG-CSF или его вариантов описаны в патентах США US 5681720, US 5795968, US 5824778, US 5985265 и Bowen et al., Experimental Hematology 27: 425-432 (1999).
Определение рецепторсвязывающей аффинности полипептида (в связанном и диссоциированном состоянии комплекса лиганд-рецептор (on-rate и off-rate)
Сила связывания между рецептором и лигандом может быть измерена при помощи твердофазного иммуноферментного анализа (ELISA), радиоиммуноанализа (РИА) или других способов иммунодетектирования, хорошо известных в данной области. Связывающее взаимодействие лиганд-рецептор может быть также измерено на приборе Biacore®, который использует детектирование плазменного резонанса (Zhou et al., Biochemistry, 1993, 32, 8193-98; Faegerstram and О'Shannessy, 1993, In Handbook of Affinity Chromatography, 229-52, Marcel Dekker, Inc., NY).
Способ Biacore® позволяет связывать рецептор с поверхностью золота и давать лиганду протекать через него. Детектирование плазменного резонанса дает прямое количественное определение количества массы, связанной с поверхностью в режиме реального времени. Этот способ дает константы как в связанном, так и в диссоциированном состоянии (on- и off-константы), и, следовательно, может быть непосредственно определена константа диссоциации и константа аффинности комплекса лиганд-рецептор.
Тест иммуногенности in vitro hG-CSF-конъюгатов
Уменьшенная иммуногенность конъюгата данного изобретения может быть определена при помощи способа ELISA, измеряющего иммунореактивность конъюгата относительно контрольной молекулы или контрольного препарата. Контрольные молекула или препарат являются обычно препаратом рекомбинантного G-CSF человека, таким как Neupogen®, или препаратом другого рекомбинантного G-CSF человека, например N-терминально ПЭГилированной молекулы rhG-CSF, как описано в патенте США US 5824784. Способ ELISA основан на антителах пациентов, получавших один из этих рекомбинантных препаратов G-CSF. Считается, что иммуногенность является уменьшенной, когда конъюгат данного изобретения имеет статистически значимую более низкую реакцию в этом анализе, чем контрольные молекула или препарат.
Нейтрализация активности в биоанализе G-CSF
Нейтрализацию конъюгатов hG-CSF сыворотками против G-CSF анализируют с использованием биоанализа G-CSF, описанного выше.
Используют сыворотки пациентов, обработанных контрольной молекулой G-CSF, или иммунизированных животных. Сыворотки добавляют либо в фиксированной концентрации (разбавление 1:20-1:500 (сыворотки пациентов) или 20-1000 нг/мл (сыворотки животных)), либо в пятикратных серийных разведениях, начиная от 1:20 (сыворотки пациентов), или 1000 нг/мл (сыворотки животных). Конъюгат hG-CSF добавляют либо в виде семикратных разведений, начиная от 10 нМ, либо в фиксированной концентрации (1-100 пМ) в общем объеме 80 мкл среды DMEM + 10% ФТС. Сыворотки инкубируют в течение 1 часа при 37°С с конъюгатом hG-CSF.
Пробы (0,01 мл) переносят затем в 96-луночные культуральные планшеты, содержащие клетки NFS-60 в 0,1 мл среды DMEM. Культуры инкубируют в течение 48 часов при 37°С в атмосфере воздуха с 5% СО2. К этим культурам добавляют 0,01 мл WST-1 (агент пролиферации клеток WST-1, Roche Diagnostics GmbH, Mannheim, Germany) и инкубируют в течение 150 мин при 37°С в атмосфере воздуха с 5% СО2. Расщепление тетразолиевой соли WST-1 митохондриальными дегидрогеназами в жизнеспособных клетках приводит к образованию формазана, который определяют количественно измерением поглощения при 450 нм.
При титровании проб hG-CSF-конъюгата в присутствии фиксированного количества сыворотки нейтрализующее действие определяют как ингибирование в какое-либо число раз (FI), определяемое количественно в виде ЕС50 (с сывороткой)/ЕС50 (без сыворотки). Уменьшение нейтрализации антителами вариантных белков G-CSF определяют как

ПРИМЕР 1
Конструирование и клонирование синтетических генов, кодирующих hG-CSF
Следующие ДНК-фрагменты синтезировали при помощи общей процедуры, описанной Stemmer et al. (1995), Gene 164, pp.49-52:
Фрагмент 1, состоящий из сайта расщепления BamHI, последовательности, кодирующей сигнальный пептид YAP3 (WO 98/32867), последовательности, кодирующей лидерную последовательность ТА57 (WO 98/32867), последовательности, кодирующей сайт узнавания протеазы КЕХ2 (AAAAGA), последовательности, кодирующей hG-CSF, с ее использованием кодонов, оптимизированных для экспрессии в Е. coli (SEQ ID NO:2), и сайта расщепления XbaI.
Фрагмент 2, состоящий из сайта расщепления BamHI, последовательности, кодирующей сигнальный пептид YAP3 (WO 98/32867), последовательности, кодирующей лидерную последовательность ТА57 (WO 98/32867), последовательности, кодирующей гистидиновую метку (SEQ ID NO:5), последовательности, кодирующей сайт расщепления протеазы КЕХ2 (AAAAGA), последовательности, кодирующей hG-CSF, с ее использованием кодонов, оптимизированных для экспрессии в Е. coli, (SEQ ID NO:2), и сайта расщепления XbaI.
Фрагмент 3, состоящий из сайта расщепления NdeI, последовательности, кодирующей сигнальный пептид OmpA (SEQ ID NO:3), последовательности, кодирующей hG-CSF с ее использованием кодонов, оптимизированных для экспрессии в Е. coli (SEQ ID NO:2), и сайта расщепления BamHI.
Фрагмент 2, состоящий из сайта расщепления BamHI, консенсусной последовательности Козака (Kozak, M. J. Mol. Biol. 1987 Aug 20; 196(4): 947-50), последовательности, кодирующей сигнальный пептид hG-CSF (SEQ ID NO:7) и hG-CSF, с использованием ее кодона, оптимизированного для экспрессии в клетках СНО (SEQ ID NO:8), и сайта расщепления XbaI.
Фрагменты ДНК 1 и 2 встраивали в сайты расщепления BamHI и Xbal в плазмиде pJSO37 (Okkels, Ann. New York Acad. Sci. 782: 202-207, 1996) с использованием стандартной ДНК-техники. Это приводило к плазмидам pG-CSFcerevisiae и pHISG-CSFcerevisiae.
ДНК-фрагмент 3 встраивали в сайты расщепления Ndel и BamHI в плазмиде рЕТ12а (Invitrogen) при помощи стандартной ДНК-техники. Это приводило к плазмиде pG-CSFcoli.
ДНК-фрагмент 4 встраивали в сайты расщепления BamHI и Xbal плазмиды pcDNA3.1(+) (Invitrogen) при помощи стандартной ДНК-техники. Это приводило к плазмиде pG-CSFCHO.
ПРИМЕР 2
Экспрессия hG-CSF в S. cerevisiae и E.coli
Трансформацию Saccharomyces cerevisiae YNG318 (доступную из Американской Коллекции Типовых культур, VA, USA ATCC 208973) либо плазмидой pG-CSFcerevisiae, либо плазмидой pHISG-CSFcerevisiae, выделение трансформантов, содержащих любую из этих двух плазмид, и последующую внеклеточную экспрессию hG-CSF без HIS-метки и с HIS-меткой, соответственно, выполняли при помощи стандартных способов, описанных в литературе. Трансформацмю E.coli BL21 (DE3) (Novagen, Cat. No. 69387-3) плазмидой pG-CSFcoli, выделение трансформантов, содержащих эту плазмиду, и последующую экспрессию hG-CSF в супернатанте и в периплазме клетки выполняли, как описано в рЕТ System Manual (8th edition) из Novagen.
Экспрессию hG-CSF S. cerevisiae и E.coli подтверждали Вестерн-блот-анализом с использованием набора ImmunePure Ultra-Sensitive ABC Rabbit IgG Staining kit (Pierce) и поликлонального антитела против hG-CSF (Pepro Tech EC Ltd.). Наблюдали, что этот белок имел правильный размер.
Уровни экспрессии hG-CSF с N-концевой гистидиновой меткой или без метки в S. cerevisiae и E.coli определяли количественно с использованием коммерчески доступного G-CSF-специфического ELISA-набора (Quantikine Human G-CSFIinmunoassay, R&D Systems Cat. No. DCS50). Измеренные величины приведены ниже.
ПРИМЕР 3
Очистка hG-CSF и его вариантов из культуральных супернатантов S. cerevisiae
Очистку hG-CSF выполняли следующим образом:
Клетки удаляют центрифугированием. Затем супернатант с удаленными клетками стерилизуют посредством фильтрации через фильтр 0,22 мкм. Простерилизованный посредством фильтрации супернатант разбавляют в 5 раз в 10 мМ ацетате натрия рН 4,5. рН доводят добавлением 10 мл концентрированной уксусной кислоты на 5 литров разбавленного супернатанта. Ионная сила должна быть менее 8 мС/см перед нанесением на катионообменную колонку.
Разбавленный супернатант наносят при линейной скорости тока 90 см/ч на SP-сефарозную FF (Pharmacia) колонку, уравновешенную 50 мМ ацетатом натрия, рН 4,5, пока эффлюент из колонки не достигает стабильных фоновых УФ и проводимости. Для удаления любого несвязанного материала колонку промывают при помощи буфера для уравновешивания, пока эффлюент из колонки не достигает стабильного уровня в отношении поглощения в УФ и проводимости. Связанный белок G-CSF элюируют из колонки при помощи линейного градиента; 30 колоночных объемов; 0-80% буфер В (50 мМ NaAc, pH 4,5, 750 мМ NaCl) при скорости тока 45 см/ч. На основе электрофореза в ДСН-ПААГ, фракции, содержащие G-CSF, объединяют. Добавляют хлорид натрия, пока ионная сила раствора не будет более чем 80 мС/см.
Белковый раствор наносят на колонку Phenyl Toyo Pearl 650S, уравновешенную 50 мМ NaAc, pH 4,5, 750 мМ NaCl. Любой несвязанный материал вымывают из колонки буфером для уравновешивания. Элюцию G-CSF выполняют нанесением ступенчатого градиента воды MilliQ. Фракции, содержащие G-CSF, объединяют. С использованием этой стратегии 2-стадийной понижающей обработки можно получить G-CSF с чистотой более 90%. Затем очищенный белок определяют количественно с использованием спектрофотометрических измерений при 280 нм и/или аминокислотным анализом.
Фракции, содержащие G-CSF, объединяют. Замену буфера и концентрирование выполняют с использованием концентраторов VivaSpin (mwco (отсечение mw): 5 кДа).
ПРИМЕР 4
Идентификация и количественное определение неконъюгированного и конъюгированного hG-CSF и его вариантов
Электрофорез в ДСН-полиакриламидном геле
Очищенный концентрированный G-CSF анализировали при помощи электрофореза в ДСН-ПААГ. Доминирующей была единственная полоса, имеющая среднюю молекулярную массу приблизительно 17 кДа.
Поглощение
Приближенную оценку концентрации G-CSF получают спектрофотометрическими способами. Измерением поглощения при 280 нм и с использованием теоретического коэффициента экстинкции 0,83 рассчитывают концентрацию белка.
Аминокислотный анализ
Более точное определение белка получают аминокислотным анализом. Аминокислотный анализ, выполненный на очищенном G-CSF, показал, что экспериментально определенный аминокислотный состав находится в согласии с ожидаемым на основе ДНК-последовательности аминокислотным составом.
ПРИМЕР 5
MALDI-TOF-масс-спектрометрия ПЭГилированного G-CSF дикого типа и вариантов G-CSF
MALDI-TOF-масс-спектрометрию использовали для оценки числа ПЭГ-групп, присоединенных к ПЭГилированному G-CSF дикого типа и к выбранным ПЭГилированным вариантам G-CSF.
G-CSF дикого типа содержит 5 первичных аминокислот, которые являются ожидаемыми сайтами присоединения для SPA-PEG (N-концевая группа и ε-аминогруппа на К16, К23, К34 и К40). После ПЭГилирования G-CSF дикого типа SPA-PEG 5000, MALDI-TOF-масс-спектрометрия показала присутствие разновидностей G-CSF дикого типа в основном с присоединенными 4, 5 и 6 ПЭГ-группами. Кроме того, G-CSF дикого типа с присоединенными 7 ПЭГ-группами был ясно виден, хотя и в минорных количествах.
Вариант G-CSF, имеющий замены K16R, K34R, K40R, Q70K, Q90K и Q120K, также содержит 5 первичных аминогрупп (N-концевую аминогруппу и ε-аминогруппу на К23, К70, К90 и К120). После ПЭГилирования этого варианта G-CSF SPA-PEG5000 MALDI-TOF-масс-спектрометрия показала присутствие разновидностей G-CSF-вариантов в основном с присоединенными 4, 5 и 6 ПЭГ-группами. Кроме того, G-CSF дикого типа с присоединенными 7 ПЭГ-группами был ясно виден, хотя и в минорных количествах.
Вариант G-CSF, имеющий замены K16R, K34R и K40R, содержит 2 первичные аминогруппы (N-концевую аминогруппу и ε-аминогруппу на К23). После ПЭГилирования этого варианта G-CSF SPA-PEG5000 MALDI-TOF-масс-спектрометрия показала присутствие разновидностей G-CSF-варианта в основном с присоединенными 2 и 3 ПЭГ-группами. Кроме того, G-CSF-вариант с присоединенными 4 ПЭГ-группами был ясно виден, хотя и в минорных количествах.
Эти наблюдения ясно показывают, что, кроме аминокислотных остатков, содержащих аминогруппы, другие аминокислотные остатки являются иногда ПЭГилированными в использованных условиях ПЭГилирования. Это также показывает, что для ПЭГилирования важно, где введены аминогруппы. Это наблюдали также с использованием анализа электрофорезом в ДСН-ПААГ G-CSF дикого типа и G-CSF-вариантов.
Как описано в примере 11, было показано, что гистидин 170 является полностью ПЭГилированным при применении химического способа SPA-PEG. Кроме того, К23 и S159 являются частично ПЭГилированными. Это объясняет присутствие 1-2 лишних сайтов ПЭГилирования, наряду с первичными аминогруппами в hG-CSF и вариантах, которые были приготовлены.
ПРИМЕР 6
Пептидное картирование ПЭГилированных и не-ПЭГилированных вариантов G-CSF
Для картирования дополнительных сайтов присоединения для SPA-PEG на G-CSF и вариантах G-CSF использовали следующую стратегию.
G-CSF-вариант с низким числом аминогрупп был выбран для уменьшения числа ожидаемых сайтов ПЭГилирования до минимума. Выбранный G-CSF-вариант имеет замены K16R, K34R, K40R и H170Q. Кроме ε-аминогруппы на К23, которая, как показали предыдущие результаты, не ПЭГилируется до большой степени, этот вариант содержит только одну первичную аминогруппу на N-конце. Таким образом, фоновое ПЭГилирование на аминогруппах значительно уменьшено в этом варианте G-CSF. Этот вариант G-CSF ПЭГилировали при помощи SPA-PEG 5000. После ПЭГилирования G-CSF-вариант денатурировали, дисульфидные связи восстанавливали, полученные тиоловые группы алкилировали и алкилированный и ПЭГилированный белок деградировали специфической для глутаминовой кислоты протеазой. Наконец, полученные пептиды разделяли обращенно-фазовой ВЖХ.
Параллельно с этим, не-ПЭГилированную версию G-CSF-варианта с заменами K16R, K34R и K40R обрабатывали идентично для создания контрольной ВЖХ-хроматограммы.
Сравнение ВЖХ-хроматограмм деградации ПЭГилированного G-CSF-варианта и не-ПЭГилированного G-CSF-варианта должно затем выявить, какие пептиды исчезают при ПЭГилировании. Затем идентификация этих пептидов секвенированием N-концевых аминокислот пептида из не-ПЭГилированного G-CSF-варианта непосредственно указывает на положения, которые являются ПЭГилированными.
В принципе, было бы предпочтительно использовать не-ПЭГилированную версию G-CSF-варианта, имеющую все замены K16R, K34R, K40R и H170Q, но для всех практических целей это не имеет значения.
Более конкретно, приблизительно 1 мг ПЭГилированного G-CSF-варианта, имеющего замены K16R, K34R, K40R и H170Q, и приблизительно 500 мкг не-ПЭГилированного G-CSF-варианта, имеющего замены K16R, K34R и K40R, сушили в концентраторе SpeedVac. Эти две пробы растворяли, каждую, в 400 мкл 6 М гуанидинина, 0,3 М Трис-HCl, рН 8,3 и денатурировали в течение ночи при 37°С. После денатурации дисульфидные связи в белках восстанавливали добавлением 50 мкл белка 0,1 М DTT в 6 М гуанидина, 0,3 М Трис-HCl, рН 8,3. После 2 ч инкубирования при температуре окружающей среды присутствующие тиоловые группы алкилировали добавлением 50 мкл 0,6 М иодоацетамида в 6 М гуанидинине, 0,3 М Трис-HCl, рН 8,3. Алкилирование происходило в течение 30 мин при температуре окружающей среды, после чего у восстановленных и алкилированных белков заменяли буфер на 50 мМ NH4НСО3 с использованием колонок NAP5. Объемы проб уменьшали до приблизительно 200 мкл в концентраторе SpeedVac перед добавлением 20 мкг и 10 мкг специфической для глутаминовой кислоты протеазы, соответственно. Деградации специфической для глутаминовой кислоты протеазой проводили в течение 16 ч при 37°С. Полученные пептиды разделяли обращенно-фазовой ВЖХ с использованием колонки C18 Phenomenex Jupiter (0,2*5 см) с элюцией линейным градиентом ацетонитрила в 0,1% водной ТФУ. Собранные фракции анализировали MALDI-TOF-масс-спектрометрией, и затем отобранные пептиды подвергали секвенированию N-концевых аминокислот.
Сравнение ВЖХ-хроматограмм деградаций ПЭГилированного G-CSF-варианта и не-ПЭГилированного G-CSF-варианта показало, что только две фракции исчезают при ПЭГилировании. Секвенирующий анализ N-концевых аминокислот этих двух фракций из не-ПЭГилированного G-CSF-варианта показал, что оба эти пептида происходили из N-конца G-CSF. Один пептид состоял из аминокислот 1-11, генерированных неожидаемым расщеплением после Gln11. Другой пептид состоял из аминокислотных остатков 1-19, генерированных ожидаемым расщеплением после Glu19.
Ожидалось, что N-концевой пептид G-CSF мог бы быть идентифицирован с использованием этого подхода, так как N-концевая аминогруппа является легко ПЭГилируемой. Однако ни один из дополнительных сайтов присоединения для SPA-PEG не был идентифицирован с использованием этого подхода.
Альтернативой этой непрямой идентификации сайтов присоединения ПЭГ 5000 является прямая идентификация сайтов присоединения в ПЭГилированных пептидах. Однако фракции, содержащие ПЭГилированные пептиды в ВЖХ-разделении деградированного ПЭГилированного G-CSF-варианта, являются плохо отделяемыми друг от друга и от нескольких фракций, содержащих не-ПЭГилированные пептиды. Таким образом, анализ N-концевой аминокислотной последовательности этих фракций не приводил к каким-либо применимым данным для указания на то, что К23 мог бы быть частично ПЭГилированным.
Для преодоления этой проблемы два пула ПЭГилированных пептидов получали из фракций первого ВЖХ-разделения. Эти два пула сушили в концентраторе SpeedVac, растворяли в 200 мкл свежеприготовленного 50 мМ МН4НСО3 и дополнительно деградировали 1 мкг химотрипсина. Полученные пептиды разделяли обращенно-фазовой ВЖХ с применением колонки C18 Phenomenex Jupiter (0,2*5 см) с элюцией линейным градиентом ацетонитрила в 0,1% водной ТФУ. Собранные фракции анализировали MALDI-TOF-масс-спектрометрией, и затем отобранные пептиды подвергали секвенированию М-концевых аминокислот.
Из определений N-концевой аминокислотной последовательности можно было определить, что К23 так же, как S159, является частично ПЭГилированным. Можно было определить точную степень ПЭГилирования в этих двух положениях, но ПЭГилирование является только частичным, так как пептиды, в которых К23 и S159 являются немодифицированными, были идентифицированы и секвенированы из первоначального ВЖХ-разделения.
ПРИМЕР 7
Гликозилирование G-CSF дикого типа и G-CSF-вариантов
Постоянно обнаруживается при анализе очищенных G-CSF дикого типа и G-CSF-вариантов при помощи MALDI-TOF-масс-спектрометрии присутствие дополнительного компонента с массой приблизительно на 324 Да большей, чем масса анализируемой молекулы G-CSF. Поскольку компонент с самой низкой массой постоянно имеет массу молекулы G-CSF и поскольку молекулы G-CSF имеют правильную N-концевую аминокислотную последовательность, был сделан вывод, что дополнительный компонент является модифицированной молекулой G-CSF, несущей два гексозных остатка. Во многих случаях немодифицированная молекула G-CSF вызывает наиболее интенсивный сигнал, но в некоторых случаях интенсивность сигнала для модифицированной молекулы G-CSF является наивысшей.
Во время анализа пептидов, генерированных с целью идентификации дополнительных сайтов ПЭГилирования, в каждой из деградации были идентифицированы два представляющих интерес пептида для идентификации сайта гликозилирования.
В обоих ВЖХ-разделениях эти два пептида элюируются близко друг к другу и MALDI-TOF-масс-спектрометрия показывает различие по массе между этими двумя пептидами приблизительно 324 Да. Данные масс-спектрометрии показывают, что пептид охватывает аминокислотные остатки 124-162. Анализ N-концевой аминокислотной последовательности всех четырех пептидов показал, что это соотнесение является правильным и что Thr133 является единственным сайтом модификации. В пептидах с массой немодифицированного пептида, Thr133 ясно виден в этой последовательности, тогда как невозможно обнаружить аминокислотный остаток в положении 133 в пептидах с дополнительной массой 324 Да. Поскольку все другие аминокислотные остатки можно было соотнести в этой последовательности, был сделан вывод, что Thr133 является единственным сайтом модификации. Этот сайт гликозилирования, как сообщалось ранее, используется в рекомбинантном G-CSF, экспрессируемом в клетках СНО, хотя этот гликан отличается от присоединяемого дрожжами.
Негликозилированный wt G-CSF отделяли от гликозилированного wt G-CSF с использованием обращенно-фазовой ВЖХ с применением колонки C18 Vydac (0,21*5 см) с изократической элюцией 51% ацетонитрилом в 0,1% водной ТФУ в виде фракции, которая, как показано MALDI-TOF-масс-спектрометрией, содержит только негликозилированную форму wt G-CSF.
ПРИМЕР 8
Разделение молекул G-CSF с различными числами ковалентно присоединенных молекул ПЭГ
Разделение молекул G-CSF, ковалентно присоединенных к 4, 5 или 6 ПЭГ-группам, получали следующим образом. ПЭГилированный белок в 20 мМ цитрате натрия, рН 2,5 наносили на SP-сефарозную FF колонку, уравновешенную 20 мМ цитратом натрия рН 2,5. Любой несвязанный материал вымывали из колонки. Элюцию проводили с использованием градиента рН. ПЭГилированный G-CSF начинал элюироваться из колонки при приблизительно рН 3,8 и продолжал элюироваться во фракциях, охватывающих диапазон рН от 3,8 до 4,5.
Эти фракции подвергали электрофорезу в ДСН-ПААГ и масс-спектрометрическому анализу. Эти анализы указывают на то, что G-CSF, имеющий наивысшую степень ПЭГилирования, локализуется во "фракциях низкого рН". ПЭГилированный G-CSF, имеющий более низкую степень ПЭГилирования, элюируется во "фракциях высокого рН".
Аминокислотный анализ, выполненный на ПЭГилированном G-CSF, показал хорошее соответствие между теоретически и экспериментально определенным коэффициентом экстинкции.
ПРИМЕР 9
Конструирование hG-CSF-вариантов
Специфические замены существующих аминокислот в hG-CSF другими аминокислотными остатками, например, специфические замены, обсуждаемые выше в общем описании, вводили с использованием стандартных ДНК-способов, известных в данной области. Новые G-CSF-варианты получали с использованием плазмиды pG-CSFcerevisiae, содержащей ген, кодирующий hG-CSF без HIS-метки, в качестве ДНК-матрицы в реакции ПЦР. Варианты экспрессировали в S. cerevisiae и очищали, как описано в примере 3. Некоторые из сконструированных G-CSF-вариантов перечислены ниже (см. примеры 10 и 11).
ПРИМЕР 10
Ковалентное присоединение SPA-PEG к hG-CSF или его вариантам
G-CSF человека и его варианты ковалентно связывали с SPA-PEG 5000, SPA-PEG 12000 и SPA-PEG 20000 (Shearwater), как описано выше ("ПЭГилированиие hG-CSF и его вариантов в растворе"). Активности in vitro этих конъюгатов показаны в виде списка в примере 12.
ПРИМЕР 11
Идентификация сайтов присоединения SPA-PEG в G-CSF сайт-направленным мутагенезом с последующим ПЭГилированием очищенных вариантов
SPA-PEG может быть присоединен к другим остаткам, чем лизин в G-CSF. Для определения, был ли SPA-PEG присоединен к гистидинам, серинам, треонинам и аргининам, готовили варианты, в которых эти аминокислоты заменяли на лизин, аланин или глутамин. Эти варианты экспрессировали в S. cerevisiae, очищали и ПЭГилировали с последующим анализом числа присоединенных молекул SPA-PEG при электрофорезе в ДСН-ПААГ. Уменьшение числа присоединенных молекул SPA-PEG после замены конкретной аминокислоты глутамином или аланином является явным указанием на то, что эта аминокислота является ПЭГилированной SPA-PEG, и это наблюдение дополнительно подтверждается неизмененной степенью ПЭГилирования после замены этой аминокислоты на лизин. Анализируемые варианты перечислены ниже.
Эти данные показывают, что, наряду с N-концом, К16, К34 и К40, SPA-PEG также ковалентно связан с Н170. Кроме того, эти данные показывают, что только 10% доступных аминокислотных остатков К23 являются ПЭГилированными и что приблизительно 10% S159 являются ПЭГилированными.
ПРИМЕР 12
Биологическая активность in vitro неконъюгированного и конъюгированного hG-CSF и его вариантов
Биологические активности конъюгированного и неконъюгированного hG-CSF и его вариантов измеряли, как описано выше в разделе "Первичный анализ 2 - анализ активности hG-CSF in vitro". Измеренные величины ЕС50 для каждого варианта с конъюгацией и без конъюгации SPA-PEG 5000 с доступными сайтами ПЭГилирования перечислены ниже. Величины нормализовали в отношении величины ЕС50 неконъюгированного hG-CSF (Neupogen®). Эту величину измеряли одновременно с вариантами каждый раз в идентичных условиях анализа. Величина ЕС50 hG-CSF в описанном анализе равна 30 пМ.
Эти данные показывают, что замена К23 на аргинин не увеличивает активность конъюгированного белка. Это обусловлено тем фактом, что только 10% К23 являются ПЭГилированными, в результате чего конъюгированный K23R-вариант имеет по существу то же самое число ПЭГ-групп, присоединенных к нему, и имеет то же самое расположение сайтов ПЭГилирования, что и hG-CSF. Удаление остальных лизинов в положении К16, К34 и К40 приводило к G-CSF-варианту со значительной активностью после ПЭГилирования. Конъюгация SPA-PEG 5000 с этим вариантом не уменьшала активность значимо в сравнении с неконъюгированным вариантом. Таким образом, ПЭГилирование М-конца и Н170 SPA-PEG 5000 не уменьшало активность hG-CSF. Авторы решили использовать hG-CSF K16R K34R K40R в качестве скелета для инсертирования новых сайтов ПЭГилирования. В этот скелет были введены 21 новый сайт ПЭГилирования между остатками Е45 и Н159. Эти остатки распределены на частях hG-CSF, которые не взаимодействуют с G-CSF-рецептором. Введение новых сайтов ПЭГилирования в положениях Q70, Q90, Т105, Q120, Т133 и S142 приводило к hG-CSF-вариантам, которые сохраняли значительное количество активности после ПЭГилирования SPA-PEG 5000. Таким образом, эти новые сайты ПЭГилирования комбинировали в hG-CSF-вариантах, которые имели 2 или 3 новых сайта ПЭГилирования. Наиболее активными из этих вариантов после ПЭГилирования SPA-PEG 500 были hG-CSF K16R K34R K40R Q70K Q120K и hG-CSF K16R K34R K40R Q70K Q90K Q120K.
Кроме того, SPA-PEG 5000, SPA-PEG 12000 и SPA-PEG 20000 присоединяли к hG-CSF K16R K34R K40R. Величины ЕС50 in vitro были 35%, 10% и 1%, соответственно, от неконъюгированного hG-CSF (Neupogen®).
ПРИМЕР 13
Полупериод существования in vivo неконъюгированного и конъюгированного hG-CSF и его вариантов
Полупериоды существования in vivo неконъюгированного hG-CSF (Neupogen®) и SPA-PEG 5000-конъюгированного hG-CSF K16R K34R K40R Q70K Q90K Q120K измеряли, как описано выше ("Измерения полупериода существования конъюгированного и неконъюгированного rhG-CSF и его вариантов"). Эти результаты показаны на фигуре 1. Полупериод существования in vivo Neupogen® и SPA-PEG 5000-конъюгированного hG-CSF K16R K34R K40R Q70K Q90K Q120K были определены как 2,01 ч и 12,15 ч соответственно. Таким образом, введение новых сайтов ПЭГилирования в hG-CSF и конъюгирование SPA-PEG с ним приводило к значимому увеличению полупериода существования in vivo. В более раннем эксперименте полупериод существования in vivo Neupogen® сравнивали с hG-CSF с более крупной N-терминально конъюгированной молекулой ПЭГ (10 кДа). В этом случае полупериоды существования in vivo были 1,75 ч и 7,01 ч соответственно. Таким образом, SPA-PEG 5000-конъюгированный hG-CSF K16R K34R K40R Q70K Q90K Q120K имеет значительно более длинный полупериод существования, чем Neupogen® и hG-CSF с N-терминально конъюгированной молекулой ПЭГ 10 кДа.
ПРИМЕР 14
Биологическая активность in vivo неконъюгированного и конъюгированного hG-CSF и его вариантов
Биологические активности in vivo неконъюгированного hG-CSF (Neupogen®), SPA-PEG 5000-конъюгированного hG-CSF K16R K34R K40R Q70K Q120K и SPA-PEG 5000-конъюгированного hG-CSF K16R K34R K40R Q70K Q90K Q120K измеряли, как описано выше ("Измерения полупериода существования конъюгированного и неконъюгированного rhG-CSF и его вариантов"). Эти результаты показаны на фигуре 2. Не смогли обнаружить активность Neupogen® на 48 часе после инъекции 100 мкг на кг веса тела в t=0 часов. Активность SPA-PEG 5000-конъюгированного hG-CSF K16R K34R K40R Q70K Q120K могла быть обнаружена до 72 часов после первоначальной инъекции, тогда как SPA-PEG 5000 конъюгированный hG-CSF K16R K34R K40R Q70K Q90K Q120K оставался активным in vivo до 96 часов после первоначальной инъекции. Таким образом, было показано, что оба конъюгированных варианта имели более продолжительную биологическую активность in vivo, чем Neupogen®, и что SPA-PEG 5000-конъюгированный hG-CSF K16R K34R K40R Q70K Q90K Q120K оставался в два раза дольше активным in vivo, чем Neupogen®.
Кроме того, измеряли биологические активности in vivo Neupogen®, SPA-PEG 12000-конъюгированного hG-CSF K16R K34R K40R и различных доз SPA-PEG 5000-конъюгированного hG-CSF K16R K34R K40R Q70K Q90K Q120K. Эти результаты показаны на фигуре 3. Как наблюдали ранее, не смогли обнаружить активность Neupogen® на 48 часе после первоначальной инъекции 100 мкг на кг веса тела. Введение 5 мкг на кг веса тела SPA-PEG 5000-конъюгированного hG-CSF K16R K34R K40R Q70K Q90K Q120K приводило к немного более продолжительной биологической активности in vivo, чем Neupogen®, тогда как введение 25 мкг на кг веса тела и 100 мкг на кг веса тела этого соединения приводило к активности hG-CSF до 72 и 96 часов соответственно после первоначальной инъекции. SPA-PEG-12000-конъюгированный hG-CSF K16R K34R K40R оставался активным in vivo до 72 часов после введения 100 мкг на кг веса тела. Как описано в примере 5, SPA-PEG 12000-конъюгированный hG-CSF K16R K34R K40R имеет 2 или 3 присоединенные группы SPA-PEG 12000, тогда как SPA-PEG 5000-конъюгированный hG-CSF K16R K34R K40R Q70K Q90K Q120K имеет 5 или 6 присоединенных групп SPA-PEG 5000. Таким образом, молекулярные массы этих двух соединений равны 42-54 кДа и 43-48 кДа, соответственно. Более продолжительная биологическая активность in vivo SPA-PEG 5000-конъюгированного hG-CSF K16R K34R K40R Q70K Q90K Q120K в сравнении с SPA-PEG 12000-конъюгированным hG-CSF K16R K34R K40R по существу с той же самой молекулярной массой показывает, что распределение присоединенных молекул ПЭГ является важным для продолжительности биологической активности in vivo.



| название | год | авторы | номер документа |
|---|---|---|---|
| МОЛЕКУЛЫ, ПОДОБНЫЕ ФАКТОРУ VII ИЛИ VIIA | 2001 |
|
RU2278123C2 |
| КОНЪЮГАТЫ γ-ИНТЕРФЕРОНА | 2000 |
|
RU2268749C2 |
| НОВЫЙ ВАРИАНТ ЭКСЕНДИНА И ЕГО КОНЪЮГАТ | 2010 |
|
RU2528734C2 |
| КОНЪЮГАТЫ G-CSF | 2002 |
|
RU2307126C2 |
| ВАРИАНТЫ ПОЛИПЕПТИДА ГАММА-ИНТЕРФЕРОНА | 2002 |
|
RU2296130C2 |
| ГЛИКОПЭГИЛИРОВАННЫЙ ГРАНУЛОЦИТАРНЫЙ КОЛОНИЕСТИМУЛИРУЮЩИЙ ФАКТОР | 2004 |
|
RU2400490C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПОЛИПЕПТИДА, КОНЪЮГИРОВАННОГО С ПОЛИ(ЭТИЛЕНГЛИКОЛЕВЫМ) ФРАГМЕНТОМ, НУКЛЕИНОВАЯ КИСЛОТА И СЛИТЫЙ ПОЛИПЕПТИД, ПРЕДНАЗНАЧЕННЫЕ ДЛЯ ПРИМЕНЕНИЯ В СПОСОБЕ | 2011 |
|
RU2584572C2 |
| БЕЛКОВЫЙ КОМПЛЕКС, ПОЛУЧЕННЫЙ С ИСПОЛЬЗОВАНИЕМ ФРАГМЕНТА ИММУНОГЛОБУЛИНА, И СПОСОБ ПОЛУЧЕНИЯ ТАКОГО КОМПЛЕКСА | 2004 |
|
RU2356909C2 |
| КОНЪЮГАТ ПОЛИПЕПТИДА ФАКТОРА VII, СПОСОБ ЕГО ПОЛУЧЕНИЯ, ЕГО ПРИМЕНЕНИЕ И СОДЕРЖАЩАЯ ЕГО ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 2003 |
|
RU2362807C2 |
| КОНЪЮГИРОВАННЫЕ МОЛЕКУЛЫ ФАКТОРА VIII | 2009 |
|
RU2573587C2 |
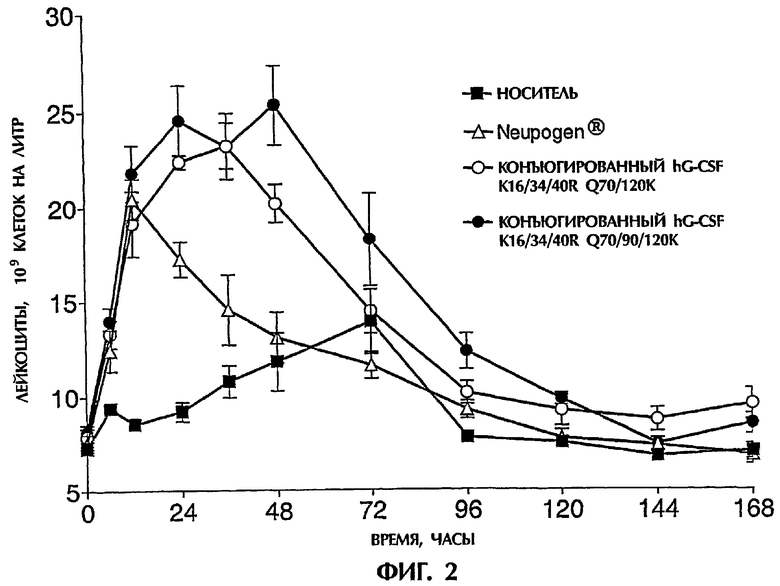
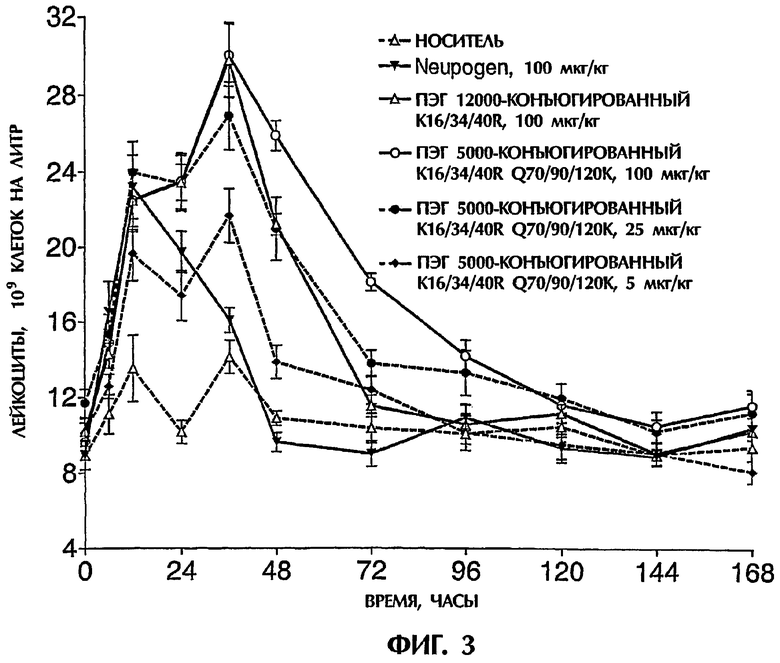
Изобретение относится к области биотехнологии, в частности к получению конъюгатов полипептидов с полиэтиленгликолем, и может быть использовано для получения конъюгатов мутантной формы G-CSF с ПЭГ. Мутантный пептид, обладающий активностью G-CSF, получают путем замен не более чем 15 аминокислотных остатков в определенных положениях аминокислотной последовательности G-CSF дикого типа, преимущественно на остаток лизина. Затем полученный мутантный пептид G-CSF конъюгируют с молекулой ПЭГ. Изобретение позволяет увеличить полупериод существования и повысить биологическую активность G-CSF in vivo, по сравнению с неконъюгированным G-CSF дикого типа. 14 з.п. ф-лы, 3 ил.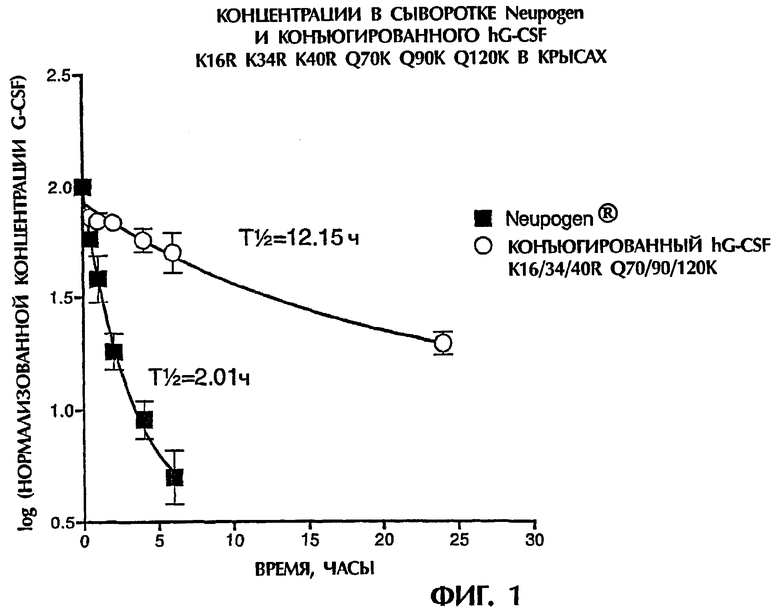
i) полипептид, проявляющий активность G-CSF, причем этот полипептид по существу соответствует аминокислотной последовательности, которая отличается от аминокислотной последовательности, приведенной в SEQ ID NO:1, не более чем по 15 аминокислотным остаткам, и включает по меньшей мере одну замену, выбранную из группы, состоящей из Т1К, Р2К, L3K, G4K, Р5К, А6К, S7K, S8K, L9K, Р10К, Q11K, S12K, F13K, L14K, L15K, Е19К, Q20K, V21K, Q25K, G26K, D27K, А29К, А30К, Е33К, А37К, Т38К, Y39K, L41K, Н43К, Р44К, Е45К, Е46К, V48K, L49K, L50K, Н52К, S53K, L54K, I56K, Р57К, Р60К, L61K, S62K, S63K, Р65К, S66K, Q67K, А68К, L69K, Q70K, L71K, А72К, G73K, S76K, Q77K, L78K, S80K, F83K, Q86K, G87K, Q90K, Е93К, G94K, S96K, Р97К, Е98К, L99K, G100K, Р101К, Т102К, D104K, Т105К, Q107K, L108K, D109K, А111К, D112K, F113K, Т115К, Т116К, W118K, Q119K, Q120K, М121К, Е122К, Е123К, L124K, М126К, А127К, Р128К, А129К, L130K, Q131K, Р132К, Т133К, Q134K, G135K, А136К, М137К, Р138К, А139К, А141К, S142K, А143К, F144K, Q145K, S155K, Н156К, Q158K, S159K, L161K, Е162К, V163K, S164K, Y165K, V167K, L168K, Н170К, L171K, А172К, Q173K и Р174К, и дополнительно при необходимости включает делецию или замену другим аминокислотным остатком по меньшей мере одного из аминокислотных остатков К16, К23, К34 и К40 SEQ ID NO:1, и
ii) по меньшей мере, одну молекулу полиэтиленгликоля, присоединенную к остатку лизина этого полипептида.
(A) после одного подкожного введения 100 мкг на 1 кг веса тела крысам (в расчете на вес полипептидной части конъюгата) он:
i) увеличивает образование лейкоцитов с, по меньшей мере, той же самой скоростью и до по меньшей мере приблизительно того же самого уровня (измеренного в виде числа клеток на литр крови), что и введение 100 мкг неконъюгированного hG-CSF на 1 кг веса тела, в течение 12 ч после введения, и
ii) увеличивает уровень лейкоцитов (измеренный в виде числа лейкоцитов на литр крови) выше уровня лейкоцитов перед введением в течение, по меньшей мере, приблизительно 96 ч;
(B) после одного подкожного введения 25 мкг на 1 кг веса тела крысам (в расчете на вес полипептидной части конъюгата) он:
i) увеличивает образование лейкоцитов с, по меньшей мере, приблизительно той же самой скоростью и до, по меньшей мере, приблизительно того же самого уровня (измеренного в виде числа клеток на литр крови), что и введение 100 мкг неконъюгированного hG-CSF на 1 кг веса тела, в течение 12 ч после введения, и
ii) увеличивает уровень лейкоцитов (измеренный в виде числа лейкоцитов на литр крови) выше уровня лейкоцитов перед введением в течение, по меньшей мере, приблизительно 72 ч;
(C) после одного подкожного введения 100 мкг на 1 кг веса тела крысам (в расчете на вес полипептидной части конъюгата) он:
i) увеличивает образование нейтрофилов с, по меньшей мере, той же самой скоростью и до, по меньшей мере, приблизительно того же самого уровня (измеренного в виде числа клеток на литр крови), что и введение 100 мкг неконъюгированного hG-CSF на 1 кг веса тела, в течение 12 ч после введения, и
ii) увеличивает уровень нейтрофилов (измеренный в виде числа клеток на литр крови) выше уровня нейтрофилов перед введением в течение, по меньшей мере, приблизительно 96 ч;
(D) после одного подкожного введения 25 мкг на 1 кг веса тела крысам (в расчете на вес полипептидной части конъюгата) он:
i) увеличивает образование нейтрофилов с, по меньшей мере, той же самой скоростью и до, по меньшей мере, приблизительно того же самого уровня (измеренного в виде числа клеток на литр крови), что и введение 100 мкг неконъюгированного hG-CSF на 1 кг веса тела, в течение 12 ч после введения, и
ii) увеличивает уровень нейтрофилов (измеренный в виде числа клеток на литр крови) выше уровня нейтрофилов перед введением в течение, по меньшей мере, приблизительно 72 ч.
| US 4904584, 27.02.1990 | |||
| HERSHFIELD M.S | |||
| et al., Use of site-directed mutagenesis to enhance the epitope-shielding effect of covalent modification of proteins with polyethylene glycol, Proc | |||
| Natl | |||
| Acad | |||
| Sci., USA, 1991, v.88, n.16, p.7185-7189 | |||
| ПОЛИЭТИЛЕНГЛИКОЛЕВОЕ ПРОИЗВОДНОЕ ПОЛИПЕПТИДОВ НЕЙРОТРОФИЧЕСКОГО ФАКТОРА И СПОСОБ ИХ ПОЛУЧЕНИЯ | 1995 |
|
RU2136694C1 |

