Предпосылки изобретения
Настоящее изобретение притязает на приоритет предварительной заявки на выдачу патента США с серийным №60/326593, поданной 2 октября 2001 г., полный текст которой включен сюда в качестве ссылки.
1. Область изобретения
Настоящее изобретение относится к усовершенствованным лентивирусным векторам и к их применению в генной доставке желаемых трансгенов в клетки-мишени и их экспрессии в данных клетках на высоком уровне, особенно, в дифференцированных кровяных ростках, происходящих от модифицированных лентивирусным вектором кроветворных стволовых клеток человека (hHSC).
2. Описание связанной области
Генная терапия посредством трансдукции кроветворных стволовых клеток человека (hHSC) представляет собой многообещающий подход для лечения некоторых врожденных и приобретенных лимфогематологических нарушений. Однако стабильные манипуляции генами в hHSC с долгосрочным поддержанием популяции с использованием существующих систем генной доставки не обеспечивали достижения эффективности, совместимой с терапевтическими реалиями. Например, онкоретровирусные векторы, происходящие из вируса мышиного лейкоза Молони (MLV), хотя являются очень привлекательными в связи с тем, что они интегрируют переносимый продукт в хромосомы клетки-мишени, не могут обеспечивать трансдукцию hHSC, которую до этого не обрабатывали индукторами пролиферации (Kohn et al., 1991; Mazurier et al., 1998). Действительно, ядерный транспорт комплекса предварительной интеграции MLV требует разрушения ядерной оболочки, которая происходит при митозе (Roe et al., 1993; Lewis and Emerman, 1994). К несчастью, hHSC, взятые из костного мозга (BM), пуповинной крови (UCB) или мобилизованные в периферической циркуляции, по большей части не делятся и теряют свою плюрипотентность после длительной стимуляции и пролиферации (Bhatia et al., 1997; Dao et al., 1997; Dorrell et al., 2000). Однако в последних сообщениях показано, что существенная доля плюрипотентных клеток, а также клеток, способных к долгосрочному внедрению субъектам без избыточного веса с диабетом/тяжелым комбинированным иммунодефицитом (NOD/SCID), также называемых клетками, поддерживающими популяцию при SCID (SRC), могут поддерживаться, подлежат трансдукции и даже могут размножаться с использованием конкретных условий стимуляции (Dorrell et al., 2000; Dao et al., 1998; Piacibello et al., 1999; Ueda et al., 2000).
Лентивирусы представляют собой подгруппу ретровирусов, которые могут инфицировать неделящиеся клетки благодаря кариофильным свойствам комплекса предварительного интегрирования, которые обеспечивают их активный импорт посредством нуклеопора. Соответственно, лентивирусные векторы, происходящие из вируса иммунодефицита человека типа 1 (HIV-1), могут опосредовать эффективную доставку, интегрирование и долгосрочную экспрессию трансгенов в немитотические клетки in vitro и in vivo (Naldini et al., 1996a; Naldini et al., 1996b; Blomer et al., 1997). В частности, основанные на ВИЧ векторы могут осуществлять эффективную трансдукцию кроветворных CD34+ клеток человека в отсутствие стимуляции цитокинами (Akkina et al., 1996; Sutton et al., 1998; Uchida et al., 1998; Miyoshi et al., 1999; Case et al., 1999). Данные клетки способны к долгосрочному выживанию у мышей NOD/SCID (Miyoshi et al., 1999). Из костного мозга этих первичных реципиентов может образовываться повторная популяция вторичных мышей с трансдуцированными клетками, что подтверждается опосредованной лентивектором генетической модификацией очень примитивных кроветворных предшественников, наиболее вероятно истинных стволовых клеток. Поскольку ни одна из других доступных в настоящее время систем генной доставки не обладает такой способностью, лентивирусные векторы предоставляют ранее незадействованный базис для исследования гемопоэза и генной терапии врожденных и приобретенных нарушений лимфогемопоэза посредством генетической модификации HSC.
Демонстрация данного важного момента, однако, была осуществлена с использованием раннего поколения лентивирусных векторов, не подходящих для терапевтических применений, или поскольку они не могли отвечать требованиям биобезопасности (Akkina et al., 1996; Sutton et al., 1998; Uchida et al., 1998), или поскольку они индуцировали уровни экспрессии трансгена, которые были неприемлемо низкими (Miyoshi et al., 1999; Case et al., 1999; An et al., 2000). Соответственно, имеется значимая необходимость в разработке усовершенствованных лентивирусов для применения в качестве векторов трансдукции, которые способны к эффективной трансдукции кроветворных клеток, в частности, кроветворных клеток-предшественников, и которые способны к экспрессии требуемых трансгенов на высоком уровне.
Оптимальный подход к генной терапии стволовых клеток должен приводить к эффективной трансдукции HSC, и, учитывая пластичность стволовых клеток, к ограниченной экспрессии терапевтических генов в конкретных ростках зрелых кровяных клеток. Лентивирусные векторы третьего поколения в настоящее время представляют собой наиболее оптимизированные инструменты для генной доставки в HSC человека, не находящиеся в клеточном цикле. Более того, предоставляется самоинактивирующаяся конструкция (SIN) для применения тканеспецифических промоторов без помехи для вышележащих LTR.
Сущность изобретения
Настоящее изобретение относится к разработке усовершенствованных лентивирусных векторов, которые отвечают требованиям биобезопасности и которые могут быть стимулированы, если требуется, для индукции высоких уровней экспрессии трансгена в манере, специфичной для ткани или ростка стволовой клетки. Кроме того, настоящее изобретение относится к контролю экспрессии трансгена в подвергнутой трансдукции клетке путем активации или репрессии транскрипции, которая является результатом контактирования промотора или энхансеров с факторами регуляции транскрипции.
Соответственно, настоящее изобретение относится к носителям для переноса генов, которые оказались особенно хорошо приспособленными для трансдукции кроветворных клеток-предшественников (HPC) человека и для экспрессии трансгена в специфично дифференцированных кровяных ростках или под контролем конкретных факторов транскрипции. Данные факторы способствуют дальнейшему применению лентивирусных векторов для генетической манипуляции кроветворными стволовыми клетками, и, в частности, их следует применять в исследовательских и терапевтических применениях. Некоторые примеры рассматриваемых клеточных типов охватывают незрелые кровяные клетки, зрелые кровяные клетки, нейтрофилы, моноциты/макрофаги и гранулоциты.
Однако специалисту в данной области будет понятно, что данное изобретение не ограничено трансдукцией кроветворных клеток и что лентивирусные векторы по изобретению можно использовать для специфичной для клеток экспрессии трансгена также в других клеточных типах. Некоторые примеры других рассматриваемых клеточных типов охватывают окончательно дифференцированные клетки, такие как нейроны, клетки легких, мышечные клетки, клетки печени, клетки поджелудочной железы, клетки эндотелия, кардиоциты, клетки кожи, клетки стромы костного мозга и клетки глаза. Кроме того, также рассматриваются такие стволовые клетки, как клетки протоков поджелудочной железы, предшественники нервных клеток и мезодермальные стволовые клетки.
Таким образом, настоящее изобретение, в общем и целом, относится к усовершенствованным векторам, которые сконструированы так, что они дают возможность трансфекции и трансдукции кроветворных клеток-предшественников человека, или стволовых клеток (hHSC), и обеспечивают в таких клетках экспрессию желаемых трансгенов высокого уровня. Кроме того, настоящее изобретение относится к ограниченной экспрессии данных желаемых трансгенов в том плане, что экспрессия регулируется для достижения экспрессии в конкретных нисходящих ростках HSC или в ответ на активаторы транскрипции. Векторы по настоящему изобретению также могут представлять собой самоинактивирующиеся лентивекторы в том плане, что они могут содержать конкретные характеристики «самоинактивирующейся» конструкции, что делает данные векторы безопасными для применения у человека. Данные характеристики самоинактивирующейся, или SIN-конструкции, могут охватывать модификации LTR вектора, так что предотвращается воссоздание способного к репликации лентивирусного генома. Особенно предпочтительное осуществление такой SIN-конструкции включает в себя делецию нуклеотидов в 3'-области LTR U3.
Лентивекторы по настоящему изобретению впервые предоставляют эффективное средство для достижения контролируемой, специфичной в отношении клеточного типа и высокоуровневой экспрессии желаемых трансгенов в дифференцированном потомстве генетически модифицированных hHSC. HSC человека трудно подвергнуть трансдукции, поскольку, будучи в нестимулированном состоянии, они относительно устойчивы к трансдукции ранее существующими векторными системами. Лентивирусные векторы по настоящему изобретению обладают способностью инфицировать неделящиеся клетки благодаря кариофильным свойствам их комплекса предварительной интеграции, что обеспечивает их активный импорт через нуклеопор. Более того, предпочтительные лентивирусные векторы по настоящему изобретению могут опосредовать эффективную доставку, интегрирование и подходящую или долгосрочную экспрессию трансгенов в немитотические клетки in vitro и in vivo, даже в отсутствие стимуляции цитокинами. Стволовые клетки, трансдуцированные более предпочтительными лентивекторами по настоящему изобретению, способны к долгосрочному приживлению, например, у мышей NOD/SCID. Однако более примечательно то, что более предпочтительные лентивекторы по настоящему изобретению имеют очень необходимые характеристики, которые обеспечивают контролируемую, при этом высокоуровневую экспрессию трансгенов в конкретных ростках человеческих клеток-предшественников и в зрелых, дифференцированных клеточных типах, при этом отвечая требованиям биобезопасности человека.
Поэтому вирусные векторы по настоящему изобретению могут, в общем, быть описаны как рекомбинантные векторы, которые содержат, по меньшей мере, лентивирусные гены gag, pol и rev, или те гены, которые требуются для продукции вирусов, что позволяет осуществлять производство вектора в подходящих количествах с использованием доступных продуктивных клеточных линий. Для соблюдения важной потребности в безопасности для человека, более предпочтительные векторы по настоящему изобретению не содержат каких-либо других активных лентивирусных генов, таких как vpr, vif, vpu, nef, tat. Данные гены могут быть удалены или инактивированы иным образом. Предпочтительным является то, что единственными активными лентивирусными генами, присутствующими на векторе, являются указанные выше гены gag, pol и rev.
Наиболее предпочтительной комбинацией лентивирусных генов и каркаса (т.е. длинных терминальных повторов или LTR), используемой при получении лентивекторов по настоящему изобретению, является та, которая происходит из вируса иммунодефицита человека (ВИЧ), и, более конкретно, из ВИЧ-1. Таким образом, гены gag, pol и rev предпочтительно являются генами ВИЧ и, более предпочтительно, генами ВИЧ-1. Однако для некоторых применений по настоящему изобретению могут использоваться гены gag, pol и rev и LTR-области из других лентивирусов, включая гены и LTR ВИЧ-2, вируса иммунодефицита обезьян (ВИО), вируса иммунодефицита кошек, вируса иммунодефицита крупного рогатого скота, вируса инфекционной анемии лошадей, вируса артрита-энцефалита коз и тому подобное. Такие конструкции могут быть использованы, например, когда требуется модификация некоторых клеток, происходящих не от человека. Однако основанные на ВИЧ векторные каркасы (т.е. LTR ВИЧ и гены gag, pol и rev ВИЧ) в общем являются предпочтительными в большинстве аспектов настоящего изобретения, так как основанные на ВИЧ конструкции являются наиболее эффективными при трансдукции кроветворных клеток-предшественников человека.
Вирусные векторы по настоящему изобретению также содержат экспрессирующую кассету, включающую в себя трансген, находящийся под контролем промотора, который активен в плане обеспечения выявляемой транскрипции трансгена в клетке человека. В предпочтительном осуществлении промотор активен в плане обеспечения транскрипции трансгена в кроветворной клетке-предшественнике человека. Более предпочтительные осуществления включают промоторы, которые активны в плане обеспечения транскрипции в конкретных клеточных типах или нисходящих ростках клеток-предшественников. Дальнейшие предпочтительные осуществления относятся к промоторам, которые являются субъектами контроля за счет активации или супрессии факторами контроля транскрипции, или активаторами и репрессорами.
Примеры промоторов, которые могут быть предпочтительно задействованы в связи с настоящим изобретением, охватывают gp91-phox, gp47-phox, CD11b, EF1-α, PGK, промотор бета-глобина, промоторы MHC класса II, фактора свертывания IX, инсулина, промотор PDX1, промоторы CD11, CD4 и CD2. Из них особенно предпочтительным является промотор gp91-phox. Промотор gp91-phox является примером промотора, который обеспечивает контролируемую экспрессию, ограниченную конкретными требуемыми клеточными типами, так как он способствует экспрессии трансгена в основном в моноцитах и гранулоцитах и так как его активность может модулироваться путем контакта промотора с активаторами, в частности, с интерфероном-гамма (ИФН-гамма). Так или иначе, воплощение настоящего изобретения, тем не менее, не ограничено указанными выше промоторами, если промотор активен в предшественнике, кроветворной или другой клетке, которую требуется применить в качестве мишени, или он реагирует на контроль транскрипции.
Для определения того, может ли использоваться конкретный промотор, выбранный промотор тестируют в конструкции in vitro в выбранной клетке-предшественнике, и, если промотор может обеспечивать экспрессию трансгена с выявляемым отношением сигнал/шум, он, в общем, может быть использован по настоящему изобретению. Требуемое отношение сигнал/шум составляет примерно от 10 до 200, более предпочтительное отношение сигнал/шум составляет примерно от 40 до 200, и еще более предпочтительное отношение сигнал/шум составляет примерно от 150 до 200. Одним из способов тестирования такого промотора, описанным здесь ниже более подробно, является применение трансгена, генерирующего сигнал, такого как зеленый флуоресцентный белок (GFP).
Настоящее изобретение, кроме того, относится к увеличенной эффективности трансдукции путем включения в вектор центрального полипуриновго тракта (cPPT). Эффективность трансдукции может составлять примерно 20%, 30%, 40%, 50%, 60%, 70%, или включительно до 80% трансдукции. В предпочтительном осуществлении cPPT располагают выше промотора последовательности. Примером cPPT является последовательность нуклеотидов, описанная SEQ ID NO:1.
Дальнейшие предпочтительные аспекты изобретения относятся к множественным уникальным участкам клонирования. Уникальные участки клонирования представляют собой участки последовательностей, распознаваемых ферментами рестрикции, которые являются уникальными в пределах векторной последовательности. Некоторые из таких участков, кластеризованные вместе, предоставляют множественные уникальные участки клонирования. Данные участки предпочтительно располагают между cPPT и промотором, или выше cPPT, хотя они могут находиться там, где это будет удобно для перемещения полинуклеотидов в вектор или из него. Например, данные множественные уникальные участки клонирования легко обеспечивают введение в вектор элементов последовательности, которые используются дополнительно и благоприятны для воплощения изобретения.
Указанные выше промоторы могут охватывать дополнительные элементы, требуемые для транскрипции, и, таким образом, являться частью транскрипционной кассеты. Транскрипционная кассета определяется как содержащая один или несколько промоторных элементов, соединенных с энхансерами и/или областями контроля локуса для того, чтобы гарантировать мощную или ограниченную тканью экспрессию трансгена. Один или несколько энхансеров могут быть расположены в векторе там, где они являются наиболее активными в отношении модулирования экспрессии трансгена. Для достижения высокого уровня экспрессии трансгена в целевых дифференцированных клеточных ростках энхансеры также могут быть специфичными в отношении целевых дифференцированных ростков. Специфичные в отношении ростков энхансеры включают в себя HS-участки. HS-участки известны для бета-глобина, CD2 и gp91, но могут быть идентифицированы дополнительные HS-участки или участки HS-типа. Например, это энхансер GATA-1 для эритробластов. Доступность последовательности генома человека должна сильно облегчить идентификацию таких элементов, которые рассматриваются как часть настоящего изобретения.
Особенно предпочтительной группой энхансерных и инсуляторных элементов являются те, что локализованы в области контроля локуса (LCR) и могут быть идентифицированы как гиперчувствительные к ДНКазе участки. Координированная энхансерная активность данных HS-участков, как полагают, ответственна за активность по открытию хроматинового домена, что, таким образом, облегчает доступность в хроматине фактора(ов) транскрипции, стимулирует белок-белковые взаимодействия между факторами связывания энхансера и промотора и необходима для определения границ домена. HS-участки, присутствующие в цис-положении относительно кассет промотор-ген, приводят к высокоуровневой, не зависящей от участка интегрирования экспрессии. Данные элементы могут располагаться в количестве одного или нескольких выше или ниже кассеты трансгена. В наиболее предпочтительном осуществлении изобретения HS-элементы располагают так, чтобы они прилегали к cPPT-элементу выше и ниже его и полностью выше промотора. Поэтому данные HS-участки могут быть введены в позиции множественных уникальных участков клонирования, описанных выше. Под прилеганием подразумевается, что указанный элемент, например, cPPT-элемент, является первым функционально значимым элементом, встречающимся при сканировании векторной последовательности от границ базового элемента, например промоторного элемента.
Для некоторых применений, например, в случае промоторов, которые лишь умеренно активны в клетках, подлежащих для трансдукции, потребуется задействовать посттранскрипционную регуляторную последовательность, расположенную так, чтобы способствовать экспрессии трансгена. Одним из типов посттранскрипционной регуляторной последовательности внутри экспрессирующей кассеты является интрон, который может служить для стимуляции генной экспрессии. Однако интроны, расположенные таким образом, могут экспонировать лентивирусный РНК-транскрипт для нормальных клеточных механизмов сплайсинга и процессинга. Так, в конкретных осуществлениях может потребоваться локализовать интрон-содержащие трансгены в ориентации, противоположной таковой векторного геномного транскрипта.
Более предпочтительным способом усиления экспрессии трансгена является применение посттранскрипционного регуляторного элемента, который не относится к событиям сплайсинга, такого как элемент посттранскрипционного процессинга вируса простого герпеса, посттранскрипционный регуляторный элемент вируса гепатита B (HPRE) или таковой вируса гепатита североамериканского лесного сурка (WPRE), который содержит дополнительный, действующий в цис-положении элемент, не обнаруженный в HPRE. Регуляторный элемент расположен внутри вектора, так что он входит в состав РНК-транскрипта трансгена, но за стоп-кодоном трансляционной единицы трансгена. Обнаружено, что применение таких регуляторных элементов особенно предпочтительно в контексте умеренных промоторов, но может быть противопоказанным в случае очень высокоэффективных промоторов.
Особенно предпочтительно применять в лентивекторах по настоящему изобретению LTR-область, которая обладает сниженной промоторной активностью по отношению к LTR дикого типа, поскольку такие конструкции обеспечивают «самоинактивирующуюся» (SIN) характеристику биобезопасности. Самоинактивирующиеся векторы являются векторами, в которых продукция полноразмерной векторной РНК в трансдуцированных клетках сильно снижена или вовсе прекращена. Данная характеристика сильно снижает риск того, что возникнут компетентные в плане репликации рекомбинанты (RCR). Более того, она снижает риск того, что клеточные кодирующие последовательности, локализованные в непосредственной близости к участку интегрирования вектора, будут неправильно экспрессироваться. Более того, SIN-конструкция снижает возможность взаимной помехи между LTR и промотором, который направляет экспрессию трансгена. Поэтому особенно подходящим является выявление полного потенциала внутреннего промотора.
Самоинактивация предпочтительно достигается путем введения делеции в U3-область 3'-LTR векторной ДНК, т.е. ДНК, используемой для продукции векторной РНК. Таким образом, во время обратной транскрипции данная делеция переносится на 5'-LTR провирусной ДНК. Требуется элиминировать количество последовательности U3, достаточное для сильного снижения или полной отмены транскрипционной активности LTR, с сильным снижением или отменой продукции полноразмерной векторной РНК в трансдуцированных клетках. Однако, в общем, требуется сохранение тех элементов LTR, которые участвуют в полиаденилировании вирусной РНК, функции, распределенной между U3, R и U5. Соответственно, требуется элиминировать столько транскрипционно значимых мотивов из LTR, сколько возможно при сохранении детерминант полиаденилирования. В случае лентивекторов, основанных на ВИЧ, обнаружено, что такие векторы переносят значительные делеции U3, включая удаление TATA-бокса LTR (например, делеции от -418 до -18), без значительного снижения титров вектора. Данные делеции приводят LTR-область, по существу, в транскрипционно неактивное состояние, так как транскрипционная способность LTR снижается примерно на 90% или ниже. В предпочтительных осуществлениях транскрипция LTR снижается примерно на 95%-99%. Таким образом, LTR может становиться от примерно на 90%, 91%, 92%, 93%, 94%, 95% 96% 97%, 98% транскрипционно неактивным до транскриционно неактивного примерно на 99%.
Полагают, что лентивекторы по настоящему изобретению могут использоваться для доставки любого желаемого трансгена, в зависимости от применения. В случае доставки кроветворных клеток-предшественников, обычно следует выбирать трансген, который будет придавать таким клеткам требуемую функцию, включая, например, гены глобинов, кроветворных факторов роста, которые охватывают эритропоэтин (EPO), интерлейкинов (таких как интерлейкин-1 (IL-1), интерлейкин-2 (IL-2), интерлейкин-3 (IL-3), интерлейкин-6 (IL-6), интерлейкин-12 (IL-12), и т.д.) и колониестимулирующих факторов (таких как гранулоцитарный колониестимулирующий фактор, гранулоцитарный/макрофагальный колониестимулирующий фактор или колониестимулирующий фактор стволовых клеток), специфический тромбоцитарный интегрин αIIbβ), гены множественной лекарственной резистентности, гены gp91-phox или gp47, дефектные у больных хроническим гранулематозом (CGD), антивирусные гены, придающие клеткам резистентность к инфекциям таких патогенов, как вирус иммунодефицита человека, гены, кодирующие факторы свертывания крови VIII или IX, мутантные у гемофиликов, лиганды, задействованные в опосредованные T-клетками иммунные ответы, такие как рецепторы T-клеточных антигенов, рецепторы B-клеточных антигенов (иммуноглобулины), а также комбинации рецепторов T- и B-клеточных антигенов отдельно или в комбинации с одноцепочечными антителами, такими как ScFv, фактор некроза опухоли (TNF), IL-2, IL-12, гамма-интерферон, CTLA4, B7 и тому подобное, гены, экспрессируемые в опухолевых клетках, такие как Melana, гены MAGE (такие как MAGE-1, MAGE-3), P198, P1A, gp100 и т.д.
В предпочтительном осуществлении трансген, подлежащий введению для лечения, представляет собой ген gp91-phox (Dinauer, et al. 1987). В дополнительном предпочтительном осуществлении трансген представляет собой ген gp91-phox, функционально связанный с промотором gp91-phox, введенным для лечения CGD. В наиболее предпочтительном осуществлении промотор gp91-phox обеспечивает экспрессию гена gp91-phox в моноцитах и гранулоцитах и, кроме того, обеспечивает модулирование экспрессии gp91-phox под действием активатора ИФН-гамма. В дополнительном предпочтительном осуществлении посттранскрипционный регуляторный элемент WPRE располагают в векторе для усиления экспрессии гена gp91-phox. В аналогично предпочтительном осуществлении трансген, подлежащий введению для лечения, представляет собой ген gp47-phox.
Основным применением настоящих трансгенов является доставка желаемых трансгенов в кроветворные клетки по некоторым возможным причинам. Они могут охватывать, конечно, без ограничения, лечение миелосупрессии и нейтропений, которые могут быть результатом химиотерапии или иммуносупрессивной терапии, или инфекций, таких как СПИД, генетических нарушений, злокачественных опухолей и тому подобное.
Рассматриваемые типичные генетические нарушения кроветворных клеток охватывают серповидно-клеточную анемию, талассемии (включая бета-талассемию), гемоглобинопатии, тромбастению Гланцманна, лизосомальные болезни накопления (такие как болезнь Фабри, болезнь Гоше, болезнь Ниманна-Пика и синдром Вискотта-Олдрича), синдромы тяжелого комбинированного иммунодефицита (SCID), дефицит адгезии лейкоцитов (LAD), а также заболевания, являющиеся результатом недостатка системной продукции секретируемого белка, например, фактора коагуляции VIII и/или IX. В таких случаях может потребоваться введение трансгенов, таких как гены глобинов (включая бета-глобины), альфа-галактозидазы A, глюкоцереброзидазы, сфингомиелинфосфодиэстеразы-1, цитокинового рецептора, CD18-интегриновой субъединицы, кроветворных факторов роста, которые включают в себя эритропоэтин (EPO), интерлейкинов (особенно интерлейкин-1, интерлейкин-2, интерлейкин-3, интерлейкин-6, интерлейкин-12 и т.д.) и колониестимулирующих факторов (таких как гранулоцитарный колониестимулирующий фактор, гранулоцитарный/макрофагальный колониестимулирующий фактор или колониестимулирующий фактор стволовых клеток), специфического тромбоцитарного интегрина αIIbβ), гены множественной лекарственной резистентности, гены gp91-phox или gp47-phox, антивирусные гены, придающие клеткам резистентность к инфекциям таких патогенов, как вирус иммунодефицита человека, гены, кодирующие факторы свертывания крови VIII или IX, мутантные у гемофиликов, лиганды, задействованные в опосредованные T-клетками иммунные ответы, такие как рецепторы T-клеточных антигенов, рецепторы B-клеточных антигенов (иммуноглобулины), а также комбинации рецепторов T- и B-клеточных антигенов отдельно и/или в комбинации с одноцепочечными антителами (ScFv), IL-2, IL-12, TNF, гамма-интерферон, CTLA4, B7 и тому подобное, гены, экспрессируемые в опухолевых клетках, такие как Melana, гены MAGE (такие как MAGE-1, MAGE-3), P198, P1A, gp100 и т.д.
Типичные злокачественные опухоли представляют собой те, что имеют кроветворное происхождение, например, возникают из миелоидного, лимфоидного или эритроидного ростков, или их клеток-предшественников. Типичные миелоидные нарушения охватывают, без ограничения, острый промиелогенный лейкоз (APML), острый миелогенный лейкоз (AML) и хронический миелогенный лейкоз (CML). Лимфоидные злокачественные опухоли, которые можно лечить с использованием лентивекторов по настоящему изобретению, охватывают, без ограничения, острый лимфобластный лейкоз (ALL), который включает в себя B-клеточный ALL и T-клеточный ALL, хронический лимфоцитарный лейкоз (CLL), пролимфоцитарный лейкоз (PLL), волосатоклеточный лейкоз (HLL) и макроглобулинемию Вальденстрема (WM). Дополнительные формы злокачественных лимфом, рассматриваемые в качестве кандидатов на лечение с использованием лентивирусных векторов по настоящему изобретению, охватывают, без ограничения, неходжкинскую лимфому и ее варианты, периферические Т-клеточные лимфомы, Т-клеточный лейкоз/лимфому взрослых (ATL), кожную Т-клеточную лимфому (CTCL), крупноклеточный гранулярный лимфоцитарный лейкоз (LGF) и болезнь Ходжкина.
В других осуществлениях настоящее изобретение относится к клеткам-хозяевам, которые были трансдуцированы одним из указанных выше лентивекторов. Полагают, что лентивекторы по настоящему изобретению могут быть использованы для трансдукции, по существу, любой клетки. Типичные клетки охватывают, без ограничения, CD4+T-клетку, лимфоцитарную клетку периферической крови, мононуклеарную клетку периферической крови, кроветворную стволовую клетку, клетку пуповинной крови, фибробласт, клетку головного мозга, клетку легких, клетку печени, клетку мышцы, клетку поджелудочной железы, эндотелиальную клетку, сердечную клетку, клетку кожи, клетку стромы костного мозга и клетку глаза, клетку протока поджелудочной железы, нервную клетку-предшественник, мезодермальную стволовую клетку и тому подобное. Трансдуцированные клетки могут, кроме того, по происхождению относиться к клеткам примата, мыши, свиньи или человека, или быть получены от другого вида животных.
Для продукции вирусных частиц можно использовать любую клетку, совместимую с экспрессией лентивирусных генов gag и pol, или любую клетку, которая может быть сконструирована для поддержки такой экспрессии. Например, могут быть использованы такие клетки-продуценты, как клетки 293T и клетки HT1080.
Как отмечалось выше, лентивекторы по изобретению, несомненно, могут использоваться, в частности, при трансдукции кроветворных клеток-предшественников человека или кроветворных стволовых клеток, полученных из костного мозга, периферической крови или пуповинной крови, а также при трансдукции CD4+T-клеток, B- или T-лимфоцитов периферической крови, мононуклеарных клеток периферической крови, дендритных клеток и моноцитарных клеток. Особенно предпочтительными мишенями являются CD34+-клетки, включая те, что выделены из мобилизованной периферической крови.
Еще в одном осуществлении настоящее изобретение относится к способу трансдукции кроветворных стволовых клеток человека, включающему контактирование популяции человеческих клеток человека, которая содержит кроветворные стволовые клетки, с одним из приведенных ниже лентивекторов в условиях, подходящих для проведения трансдукции кроветворных клеток-предшественников человека в указанной популяции данным вектором. Стволовые клетки могут быть трансдуцированы in vivo или in vitro, что зависит от конечного применения. Даже в контексте генной терапии человека, такой как генная терапия стволовых клеток человека, можно трансдуцировать стволовые клетки in vivo или, альтернативно, трансдуцировать их in vitro с последующей инфузией трансдуцированной стволовой клетки субъекту-человеку. В одном из аспектов данного изобретения стволовая клетка человека может быть выделена из организма человека, например, пациента-человека, с использованием способов, хорошо известных специалистам в данной области, и может быть трансдуцирована, как указано выше. Трансдуцированные стволовые клетки затем вводят обратно тому же или другому человеку.
При лечении субъекта-человека непосредственно путем введения вектора субъекту лечение обычно проводят путем внутривенного введения вектора. Когда клетки, например, CD34+-клетки, дендритные клетки, клетки периферической крови или опухолевые клетки трансдуцируют ex vivo, частицы вектора инкубируют с клетками, используя дозы, в общем, порядка от 1 до 50 множественности заражения (MOI), что также соответствует 1·105-50·105 единицам трансдукции вирусного вектора на 105 клеток. Этот интервал, без сомнения, охватывает количество вектора, соответствующее 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 35, 40, 45 и 50 MOI. Обычно количество вектора может быть выражено в терминах единиц трансдукции (TU) HeLa. Другие пути введения вектора включают в себя внутриартериальный, эндоскопический, в область повреждения, чрескожный, подкожный, внутримышечный, интратекальный, интраорбитальный, внутрикожный, внутрибрюшинный, транстрахеальный, субкутикулярный, путем внутристеблевой инъекции, путем ингаляции или интраназального распыления, эндотрахеальный путь и тому подобное. В осуществлениях, относящихся к способам лечения опухоли/рака векторами по изобретению, экспрессирующий вектор может быть доставлен путем прямой инъекции в опухоль или в сосудистое русло опухоли.
Предпочтительным примером генной терапии ex vivo является пациент, страдающий от заболевания хронического гранулематоза (CGD), CD34+-клетки которого могут быть выделены из костного мозга или периферической крови и трансдуцированы ex vivo лентивектором, экспрессирующим ген gp91-phox под контролем промотора gp91-phox, перед обратной имплантацией. Аналогичный подход может применяться при лечении пациентов, страдающих от талассемий, например, бета-талассемии, где клетки могут быть трансдуцированы лентивектором, экспрессирующим бета-глобин под контролем бета-глобинового или другого подходящего промотора. Аналогично, рассматриваются лентивекторы по настоящему изобретению, экспрессирующие подходящую комбинацию гена и промотора для лечения дефицита адгезии лейкоцитов (LAD).
В случае пациентов, страдающих от тяжелого комбинированного иммунодефицита (SCID), авторы изобретения рассматривают аналогичный подход с использованием лентивекторов по изобретению, экспрессирующих ген, дефектный у пациента, например, ген, кодирующий общую гамма-цепь интерлейкинового рецептора, функционально связанную с подходящим промотором, обеспечивающим специфичность в отношении подходящей ткани или клетки и функциональный контроль. Для генетического лечения инфекции ВИЧ авторы настоящего изобретения предполагают внутриклеточную иммунизацию, где клеткам придают устойчивость к вирусу ВИЧ путем введения противовирусных генов. В осуществлениях внутриклеточной иммунизации против ВИЧ мишени лентивекторов по изобретению охватывают кроветворные предшественники, CD4+Т-клетки периферической крови и моноциты. Как понятно специалисту в данной области, подобные способы внутриклеточной иммунизации могут быть использованы также для других вирусных инфекций. Для иммунотерапии злокачественных опухолей опухолевые клетки или антигенпрезентирующие клетки, такие как дендритные клетки, конструируют генетически с использованием лентивекторов по изобретению. Для способов лечения злокачественных опухолей некоторые трансгены, которые могут использоваться в лентивекторных конструкциях по изобретению, представляют собой те, которые могут ингибировать, и/или уничтожать, и/или предотвращать пролиферацию, и/или опосредовать апоптоз раковых/опухолевых клеток и/или генов, таких как TNF.
Описанные здесь лентивекторы также могут использоваться in vivo путем прямой инъекции в кровь или в конкретный орган. Например, в одном из осуществлений для лечения болезни Паркинсона может применяться инъекция в головной мозг лентивекторов, экспрессирующих происходящий из глиальной клетки фактор роста нервов (GDNF). В другом примере предполагается введение в воротную вену лентивектора, экспрессирующего фактор свертывания VIII, для коррекции гемофилии A. Еще в одном примере предполагается внутривенная или внутримышечная инъекция лентивектора по настоящему изобретению, экспрессирующего ген дистрофина, для лечения мышечной дистрофии Дюшенна. В дальнейшем, предпочтительном примере, лентивектор, экспрессирующий gp91-phox, вводят для лечения заболевания хронического гранулематоза (CGD). В особенно предпочтительном осуществлении лентивектор, экспрессирующий gp91-phox под контролем промотора gp91-phox, можно инъецировать для лечения CGD. Так, обычному специалисту в данной области очевидно интенсивное применение лентивекторных конструкций по настоящему изобретению в понятиях способов генной терапии.
Как используется здесь в спецификации или формуле изобретения, при применении слова «содержащий», слова в единственном числе означают один предмет или несколько. Используемое здесь выражение «еще один» означает, по меньшей мере, второй предмет или более.
Другие объекты, характеристики и преимущества настоящего изобретения будут понятны из следующего подробного описания. Однако следует понимать, что подробное описание и конкретные примеры, хотя и указывают на предпочтительные осуществления изобретения, приведены только для иллюстрации, следовательно, специалистам в данной области из данного подробного описания станут понятны различные изменения и модификации в пределах сущности и объема изобретения.
Краткое описание чертежей
Следующие чертежи составляют часть настоящей спецификации и включены для дальнейшей демонстрации некоторых аспектов настоящего изобретения. Изобретение может быть лучше понято при ссылке на один или несколько чертежей в комбинации с подробным описанием представленных здесь осуществлений.
Фиг.1A. Лентивекторы, содержащие промотор gp91-phox. Схематичные карты лентивирусных векторов, содержащих промотор gp91-phox (1540 н.п.) (pHPP91-GFP) и последовательности WPRE (pWPP91-GFP).
Фиг.1B. Модель регуляции промотора gp91-phox. Транскрипционный репрессор CDP конкурирует за связывание активирующих факторов транскрипции в четырех элементах. ДНК-связывающая активность CDP подлежит отрицательной регуляции во время конечного развития фагоцитов, что обеспечивает взаимодействие активаторов транскрипции с промотором gp91-phox (Luo W, Skalnik D G. JBC, 271: 18203, 1996).
Фиг.2A. Индуцируемая интерфероном-(экспрессия GFP в происходящих из UCB CD34+ моноцитах, управляемая промотором gp91-phox. CD34+клетки из UCB трансдуцировали лентивекторами pWPT-GFP и pWPP91-GFP (MOI 10). Клетки дифференцировали in vitro в присутствии GM-CSF (гранулоцитарно-макрофагальный колониестимулирующий фактор) в моноциты (CD14+клетки) в течение 3 недель. Дифференцированные клетки стимулировали ИФН-((1000 ед/мл) в течение 6 суток и метили конъюгированными с PE моноклональными антителами. Экспрессию GFP в PE-положительной популяции анализировали с использованием FACS. Значения означают процентную долю клеток в квадрантах.
Фиг.2B. Индуцируемая интерфероном-(экспрессия GFP в происходящих из UCB CD34+гранулоцитах, управляемая промотором gp91-phox. CD34+клетки из UCB трансдуцировали лентивекторами pWPT-GFP и pWPP91-GFP (MOI 10). Клетки дифференцировали in vitro в присутствии G-CSF (гранулоцитарный колониестимулирующий фактор) в моноциты (CD15+клетки) в течение 3 недель. Дифференцированные клетки стимулировали ИФН-((1000 ед/мл) в течение 6 суток и метили конъюгированными с PE моноклональными антителами. Экспрессию GFP в PE-положительной популяции анализировали с использованием FACS. Значения означают процентную долю клеток в квадрантах.
Фиг.3A. Экспрессия GFP в костном мозге мышей NOD/SCID, которым трансплантировали трансдуцированные лентивектором CD34+клетки UCB. Не трансдуцированные CD34+клетки UCB вводили внутривенно сублетально облученным мышам NOD/SCID (375 кГи) (в возрасте 8-10 недель). Через 8 недель клетки костного мозга, полученные из бедер трансплантированных мышей, метили конъюгированным с PerCP антителом против человеческого CD45 для мечения прижившихся клеток, далее идентифицировали конкретные ростки человеческих клеток с использованием конъюгированных с PE антител против CD34 (кроветворные клетки-предшественники), CD19 (B-лимфоциты), CD33 (нейтрофилы), CD14 (моноциты), CD15 (гранулоциты), CD42b (мегакариоциты) и гликофорина A (эритробласты). Экспрессию GFP анализировали в выявленных как CD45+ и PE-положительных клетках. Значения означают процентную долю клеток в квадрантах.
Фиг.3B. Экспрессия GFP в костном мозге мышей NOD/SCID, которым трансплантировали трансдуцированные лентивектором CD34+клетки UCB. Трансдуцированные лентивекторами pHPT-GFP (MOI 10) CD34+клетки UCB вводили внутривенно сублетально облученным мышам NOD/SCID (375 кГи) (в возрасте 8-10 недель). Через 8 недель клетки костного мозга, полученные из бедер трансплантированных мышей, метили конъюгированным с PerCP антителом против человеческого CD45 для мечения прижившихся клеток, далее идентифицировали конкретные ростки человеческих клеток с использованием конъюгированных с PE антител против CD34 (кроветворные клетки-предшественники), CD19 (B-лимфоциты), CD33 (нейтрофилы), CD14 (моноциты), CD15 (гранулоциты), CD42b (мегакариоциты) и гликофорина A (эритробласты). Экспрессию GFP анализировали в выявленных как CD45+ и PE-положительных клетках. Значения означают процентную долю клеток в квадрантах.
Фиг.3C. Экспрессия GFP в костном мозге мышей NOD/SCID, которым трансплантировали трансдуцированные лентивектором CD34+клетки UCB. Трансдуцированные лентивекторами pHPP91-GFP (MOI 10) CD34+клетки UCB вводили внутривенно сублетально облученным мышам NOD/SCID (375 кГи) (в возрасте 8-10 недель). Через 8 недель клетки костного мозга, полученные из бедер трансплантированных мышей, метили конъюгированным с PerCP антителом против человеческого CD45 для мечения прижившихся клеток, далее идентифицировали конкретные ростки человеческих клеток с использованием конъюгированных с PE антител против CD34 (кроветворные клетки-предшественники), CD19 (B-лимфоциты), CD33 (нейтрофилы), CD14 (моноциты), CD15 (гранулоциты), CD42b (мегакариоциты) и гликофорина A (эритробласты). Экспрессию GFP анализировали в выявленных как CD45+ и PE-положительных клетках. Значения означают процентную долю клеток в квадрантах.
Фиг.3D. Экспрессия GFP в костном мозге мышей NOD/SCID, которым трансплантировали трансдуцированные лентивектором CD34+клетки UCB. Трансдуцированные лентивекторами pWPT-GFP (MOI 10) CD34+клетки UCB вводили внутривенно сублетально облученным мышам NOD/SCID (375 кГи) (в возрасте 8-10 недель). Через 8 недель клетки костного мозга, полученные из бедер трансплантированных мышей, метили конъюгированным с PerCP антителом против человеческого CD45 для мечения прижившихся клеток, далее идентифицировали конкретные ростки человеческих клеток с использованием конъюгированных с PE антител против CD34 (кроветворные клетки-предшественники), CD19 (B-лимфоциты), CD33 (нейтрофилы), CD14 (моноциты), CD15 (гранулоциты), CD42b (мегакариоциты) и гликофорина A (эритробласты). Экспрессию GFP анализировали в выявленных как CD45+ и PE-положительных клетках. Значения означают процентную долю клеток в квадрантах.
Фиг.3E. Экспрессия GFP в костном мозге мышей NOD/SCID, которым трансплантировали трансдуцированные лентивектором CD34+клетки UCB. Трансдуцированные лентивекторами pWPP91-GFP (MOI 10) CD34+клетки UCB вводили внутривенно сублетально облученным мышам NOD/SCID (375 кГи) (в возрасте 8-10 недель). Через 8 недель клетки костного мозга, полученные из бедер трансплантированных мышей, метили конъюгированным с PerCP антителом против человеческого CD45 для мечения прижившихся клеток, далее идентифицировали конкретные ростки человеческих клеток с использованием конъюгированных с PE антител против CD34 (кроветворные клетки-предшественники), CD19 (B-лимфоциты), CD33 (нейтрофилы), CD14 (моноциты), CD15 (гранулоциты), CD42b (мегакариоциты) и гликофорина A (эритробласты). Экспрессию GFP анализировали в выявленных как CD45+ и PE-положительных клетках. Значения означают процентную долю клеток в квадрантах.
Фиг.4. WPRE сохраняет экспрессию GFP, направляемую промотором gp91-phox, в миелоидных клетках in vivo. Данные из фиг.3B, 3C и 3D; вычтена фоновая флуоресценция контрольных клеток. Neu - нейтрофилы, Mo - моноциты, Gr - гранулоциты.
Фиг.5A, 5B, 5C и 5D. Лентивирусные векторы, несущие маркер GFP под контролем конститутивного промотора EF-1-альфа (5A, 5B) или под контролем миелоидспецифичного промотора gp91-phox (5C, 5D). Векторы 5B и 5D содержат последовательности WPRE.
Фиг.6A, 6B и 6C. Конструкция лентивирусного вектора, несущая фрагмент промотора gp91-phox (500 н.п.) и участки множественного клонирования (MCS) и вставку gp91-phox-специфических энхансеров, начиная с pHPT-GFP. На первой стадии фрагмент размером 500 н.п.промотора человеческого gp91-phox генерировали путем PCR с использованием фрагмента размером 1540 н.п. в качестве матрицы (последовательность MCS-I включали в состав 5'-концевого прямого праймера), и встраивали в XhoI-BamHI pHPT-GFP вместо короткого промотора EF-1 (с получением промежуточного вектора pHP500-GFP). Затем PCR-фрагмент, содержащий MCS-II, cPPT, MCS-I и фрагмент gp91-phox размером 500 н.п., созданный с использованием вектора pHP500-GFP в качестве матрицы (последовательность MCS-II добавляли к 5'-прямому праймеру), встраивали в участки ClaI-BamHI вектора pHOX-GFP с образованием вектора pHPX-GFP (фиг.6B). gp91-phox-Специфические энхансеры встраивали последовательно в оба MCS выше или ниже последовательности cPPT с образованием вектора pHPHS-GFP (фиг.6C). Наконец, вектор pWPHS-GFP создавали путем вставки последовательности WPRE в вектор pHPHS-GFP. Позиции амплифицированных путем PCR элементов HS (+1 старт транскрипции gp91-phox) в последовательности ДНК генома человека: HS-12 (-11503, -13244), HS-14 (-13244, -14715), HS-26 (-25345, -26529), HS-27 (-26529, -27656), HS-28 (-27657, -28893) (фиг.6C). Описанные фрагменты слегка крупнее, чем рассчитанные вследствие введения участков рестрикции на концах для облегчения клонирования (HS-12 и HS-14; gta для получения 5' SnaBI после клонирования в данный участок в pHPX-GFP; HS-26, HS-27, HS-28 SalI; gcgtcgac и XhoI; ctcgagcggc).
Фиг.7. Вставка cPPT-элемента в лентивирусные векторы. Центральный полипуриновый тракт (cPPT) получали из вектора pRRLsinb.hPGK.EGFP (см. Follenzi, A, Ailles, L. E., Bakovic, S., Geuna, M., Naldini, L. (2000) Gene Transfer by Lentiviral Vecotrs is Limited by Nuclear Translocation and Rescued by HIV-1 pol Sequences. Nat. Genet. 25: 217-22.). Фрагмент NotI-EcorRV, содержащий cPPT-элемент, клонировали в участки NotI-ClaI вектора pHOX-GFP с образованием pHPT-GFP. Участки NotI и ClaI помещали в оба фрагмента лигирования, так что NotI не сохранялся в pHPT-GFP. Участок ClaI сохранялся, но становился метилированным за счет dam. Следовательно, данный плазмидный вектор для использования данного участка ClaI должен выращиваться в dam(-) бактериях.
Описание иллюстративных осуществлений
Хотя лентивирусные векторы представляют большой потенциал для генной терапии и, особенно, для трансдукции кроветворных стволовых клеток человека (hHSC), разработанные до сих пор векторы не удовлетворяли стандартам биобезопасности и пока неэффективны в плане экспрессии трансгенов. Например, хотя содержащие CMV-промотор, происходящие из ВИЧ векторы могут индуцировать высокие уровни экспрессии трансгена в центральной нервной системе (Naldini et al., 1996a; Naldini et al., 1996b; Blomer et al., 1997) и обеспечивают начальную демонстрацию того, что плюрипотентные кроветворные предшественники могут быть эффективно трансдуцированы данным инструментом доставки генов, они, по большей части, бесполезны в смысле переноса терапевтических генов в большинство клеток лимфогемопоэза, поскольку в данных мишенях их транскрипционная активность слишком мала (Miyoshi et al., 1999; Case et al., 1999; An et al., 2000). Современные лентивирусные векторы содержат множественно ослабленные гены вирулентности ВИЧ, что снимает потенциальную возможность их реконструкции вирусом дикого типа путем рекомбинации (Zufferey et al., 1997; Dull et al., 1998). Самоинактивирующаяся конструкция делает данные векторы еще более биологически безопасными путем удаления транскрипционных элементов ВИЧ (Zufferey et al., 1998). Однако это может отрицательно воздействовать на экспрессию трансгена, очевидно, путем снижения эффективности полиаденилирования (DeZazzo et al., 1991; Valsamakis et al., 1991; Brown et al., 1991; Cherrington and Ganem, 1992; Valsamakis et al., 1992; Gilmartin et al., 1992).
Настоящее изобретение преодолевает такие и другие недостатки, имеющиеся в данной области, и относится к разработке усовершенствованных векторов, происходящих из ВИЧ, которые оптимизированы в плане биобезопасности и повышенной генной экспрессии. Таким образом, в воплощении настоящего изобретения клетки человека могут трансдуцироваться происходящими из ВИЧ лентивекторами, которые содержат элементы, препятствующие образованию компетентных в плане репликации рекомбинантов (RCR), и далее содержат внутренний промоторный элемент, который индуцирует высокие уровни экспрессии трансгена в кроветворных предшественниках и дифференцированных in vitro ростках кровяных клеток, а также первичных Т-клеток. Например, с использованием векторов по данному изобретению можно трансдуцировать CD34+-клетки человека, а также других кроветворных ростков человека.
Промоторные элементы описанных векторов содержат промотор gp91-phox, промотор бета-талассемии, промоторы gp47-phox и CD4, промотор EF1 или промотор CD11b, хотя, как может понять специалист в данной области, может быть использован почти любой промоторный элемент.
Промотор gp91-phox был активен в плане обеспечения экспрессии в конкретных клеточных типах, а именно, в дифференцированных гранулоцитах и моноцитах. Более того, промотор gp91-phox может активироваться путем контакта промотора с активаторами, такими как ИФН-гамма. Также рассматриваются анализы приживления и повторной популяции у мышей NOD/SCID с данными векторами, для подтверждения стабильности экспрессии с данных промоторов in vivo.
Элемент, который предотвращает RCR в лентивекторах по настоящему изобретению, представляет собой самоинактивирующуюся (SIN) конструкцию. Она достигается путем делеции основной части U3 в 3'-концевом LTR векторной плазмиды, что приводит к образованию самоинактивирующейся (SIN) конфигурации (Zufferey et al., 1998). Данная делеция также предотвращает потенциальную помеху между LTR и внутренними промоторными элементами. Однако SIN может индуцировать снижение экспрессии трансгена, особенно в не очень сильных промоторах, таких как промотор PGK. Изобретение далее относится к способам сохранения уровней трансгена в лентивекторных конструкциях, которые не имеют сильных промоторов, путем вставки в вектор других регуляторных элементов, таких как посттранскрипционный регуляторный элемент вируса гепатита североамериканского лесного сурка (WPRE) или регуляторный элемент вируса гепатита B (HPRE), непосредственно выше делетированного 3' LTR. Вставка элемента WPRE не влияет на специфичность экспрессии промоторных элементов.
Дополнительные преимущества могут быть достигнуты путем функционального включения в векторы HS-элементов. Например, включение HS-серии энхансерных элементов в pHPX-GFP может приводить к более высокой генной экспрессии и меньшего разброса в уровне экспрессии вследствие сайленсерной активности HS. См., например, May, et al. (2000) «Therapeutic haemaglobin synthesis in beta-thalassaemic mice expressing lentivirus-encoded human beta-globin», Nature 406: 82-86, включенную сюда в качестве ссылки. May, et al. (2000) описывают HS-элементы, включенные в состав лентивектора выше промотора бета-глобина, для управления более высокого, и к тому же менее вариабельного уровня экспрессии кДНК бета-глобина.
Лентивекторы по данному изобретению могут эффективно трансдуцировать клетки некоторых ростков крови человека, включая CD34+-клетки, с использованием условий, при которых неэффективны основанные на MLV векторы. Более того, также было продемонстрировано, что человеческие CD34+-клетки могут эффективно трансдуцироваться с относительно низкой MOI, хотя эффективность генного переноса достигает примерно 60-70% трансдуцированных клеток. Например, MOI, равную 10, использовали для достижения оптимальной трансдукции, что существенно ниже, чем описано в предыдущих исследованиях, где она изменялась в пределах 60-300 и 1000-3000 (Miyoshi et al., 1999; Case et al., 1999). Частично, это может быть следствием повышенной вероятности встречи вектор-мишень, поскольку способы по настоящему изобретению задействуют воздействие на CD34+-клетки векторных частиц в малом объеме (105 клеток в 200 мкл) и в течение примерно 6-24 ч.
Таким образом, настоящее изобретение относится к происходящим из ВИЧ векторам, которые являются безопасными, высоко эффективными и очень действенными в плане экспрессии трансгенов в кроветворных клетках-предшественниках человека, а также в других производных кровяных клеток, даже в самоинактивирующейся конфигурации. Более того, данные векторы обеспечивают специфичную в отношении клеточных типов экспрессию и контролируемую экспрессию в дифференцированных клетках. Поэтому данные векторы предоставляют подходящие инструменты для генетического лечения таких заболеваний, как наследственные и приобретенные лимфогематологические нарушения, для генной терапии злокачественных опухолей, особенно гематологических злокачественных опухолей, для лечения и профилактики ВИЧ-инфекции, а также для исследования гемопоэза через опосредованную лентивекторами модификацию человеческих HSC человека.
1. Лентивирусные векторы и генная терапия
Лентивирусы представляют собой сложные ретровирусы, которые, в дополнение к обычным ретровирусным генам gag, pol и env, содержат другие гены с регуляторной или структурной функцией. Более высокая сложность позволяет вирусу модулировать свой жизненный цикл, как в случае латентной инфекции. Некоторые примеры лентивируса охватывают вирусы иммунодефицита человека ВИЧ-1, ВИЧ-2 и вирус иммунодефицита обезьян: ВИО. Лентивирусные векторы были получены множественным ослаблением генов вирулентности ВИЧ, например, гены env, vif, vpr, vpu и nef делетировали, делая вектор биологически безопасным.
Лентивирусные векторы предоставляют большие преимущества для генной терапии. Они стабильно интегрируются в хромосомы клеток-мишеней, что требуется для долгосрочной экспрессии. Далее, они не переносят вирусные гены, таким образом, избегая проблемы получения трансдуцированных клеток, которые могут быть разрушены цитотоксическими Т-клетками. Более того, они характеризуются относительно большой способностью к клонированию, достаточной для большинства предполагаемых клинических применений. Кроме того, лентивирусы, в отличие от других ретровирусов, способны трансдуцировать неделящиеся клетки. Это очень важно в контексте генной терапии тканей, таких как система кроветворения, головной мозг, печень, легкие и мышцы. Например, векторы, происходящие от ВИЧ-1, обеспечивают эффективную доставку in vivo и ex vivo, интегрирование и стабильную экспрессию трансгенов в клетки, такие как нейроны, гепатоциты и миоциты (Blomer et al., 1997; Kafri et al., 1997; Naldini et al., 1996; Naldini et al., 1998).
Лентивирусный геном и провирусная ДНК содержат три гена, обнаруженные в ретровирусах: gag, pol и env, которые фланкированы двумя последовательностями длинных концевых повторов (LTR). Ген gag кодирует внутренние структурные белки (матрикс, капсид и нуклеокапсид); ген pol кодирует РНК-направленную ДНК-полимеразу (обратную транскриптазу), протеазу и интегразу; и ген env кодирует вирусные оболочечные гликопротеины. 5'- и 3'-LTR служат для обеспечения транскрипции и полиаденилирования РНК вириона. LTR содержат все другие последовательности, действующие в цис-направлении, необходимые для репликации вируса. Лентивирусы содержат дополнительные гены, включая vif, vpr, tat, rev, vpu, nef и vpx.
К 5'-LTR прилегают последовательности, необходимые для обратной транскрипции генома (участок связывания тРНК-праймера) и для эффективной инкапсидации вирусной РНК в частицы (участок Psi). Если последовательности, необходимые для инкапсидации (или упаковки ретровирусной РНК в инфекционные вирионы), теряются из вирусного генома, цис-дефект предотвращает инкапсидацию геномной РНК. Однако полученные в результате мутанты способны направлять синтез всех белков вириона.
В данной области известны лентивирусные векторы, см. Naldini et al., (1996 and 1998); Zufferey et al., (1997); Dull et al., (1998), Ramezani et al., (2000), все включенные сюда в качестве ссылки. Также см. патенты США №5994136; 6013516; 6165782; 6207455; 6218181; 6218186; и 6277633; все они включены сюда в качестве ссылки. В общем, данные векторы основаны на плазмидах или вирусах, и они конфигурированы так, что несут последовательности, необходимые для включения чужеродной нуклеиновой кислоты, для отбора и переноса нуклеиновой кислоты в клетку-хозяина.
Два компонента участвуют в образовании основанной на вирусе системе генной доставки: во-первых, элементы упаковки, охватывающие структурные белки, а также ферменты, необходимые для образования инфекционной частицы, и во-вторых, вектор сам по себе, т.е. генетический материал, подлежащий переносу. В конструкцию обоих этих компонентов могут быть введены элементы биобезопасности. Таким образом, единица упаковки основанных на ВИЧ векторов первого поколения включала в себя все белки ВИЧ-1, кроме оболочечных белков (Naldini et al., 1998). Впоследствии было показано, что делеция четырех дополнительных вирусных генов, которые ответственны за вирулентность, включая vpr, vif, vpu и nef, не изменяла применимость векторной системы (Zufferey et al., 1997). Также было показано, что Tat, основной трансактиватор ВИЧ, также является несущественным для образования полностью эффективного вектора (Dull et al., 1998). Таким образом, системы упаковки третьего поколения основанных на ВИЧ лентивирусных векторов содержат только три гена родительского вируса: gag, pol и rev, что отменяет возможность реконструкции вируса дикого типа путем рекомбинации.
Данная система была усовершенствована далее путем удаления из вектора транскрипционных единиц ВИЧ (Zufferey et al., 1998). В данной работе было продемонстрировано, что введение делеции в U3-область 3'-LTR ДНК, использованное для продукции векторной РНК, генерировало самоинактивирующиеся (SIN) векторы. В течение обратной транскрипции данная делеция переносится 5'-LTR провирусной ДНК. Достаточная часть последовательности была удалена, включая удаление TATA-бокса, что прекращало транскрипционную активность LTR, предотвращая продукцию полноразмерной векторной РНК в трансдуцированных клетках. Однако это не влияет на титры вируса или на свойства вируса in vitro или in vivo.
Настоящее изобретение относится к некоторым усовершенствованиям существующих лентивекторов, как описано выше и в других частях данной спецификации. Введение лентивектора, предоставляющее клетке упаковки гетерологичный ген, такой как гены для лечения нарушений кроветворения и лимфогемопоэза, по данному изобретению, дает продуцирующую клетку, которая высвобождает инфекционные вирусные частицы, несущие интересующий чужеродный ген.
Ген env может происходить из любого вируса, включая ретровирусы. Предпочтительно, env представляет собой амфотропный оболочечный белок, который обеспечивает трансдукцию клеток человека и других видов. Примеры происходящих из ретровируса генов env охватывают, без ограничения: вирус мышиного лейкоза Молони (MoMuLV или MMLV), вирус мышиной саркомы Харви (HaMuSV или HSV), вирус опухоли молочной железы мыши (MuMTV или MMTV), вирус лейкоза обезьяны гиббона (GaLV или GALV), вирус иммунодефицита человека (ВИЧ) и вирус саркомы Рауша (RSV). Также могут использоваться другие гены env, такие как белок G (VSV G) вируса везикулярного стоматита (VSV), таковой из вирусов гепатита и гриппа.
Хотя белок VSV G является предпочтительным геном env, поскольку VSV G наделяет рекомбинантный вирус широким интервалом хозяев, VSV G может приносить вред клетке-хозяину. Так, когда используется такой ген, как VSV G, предпочтительным является использование индуцируемой промоторной системы, так что экспрессия VSV G может регулироваться с минимизацией токсичности в отношении хозяина, когда экспрессия VSV G не требуется. Например, регулируемая тетрациклином генная экспрессирующая система по Gossen & Bujard, (1992), может использоваться для обеспечения индуцируемой экспрессии VSV G, когда из трансформированных клеток удаляется тетрациклин. Таким образом, трансактиватор tet/VP16 присутствует на первом векторе, и кодирующую последовательность VSV G клонируют ниже промотора, контролируемого операторными последовательностями tet на другом векторе.
Вектор, предоставляющий вирусную нуклеиновую последовательность env, функционально ассоциирован с регуляторными последовательностями, например, промотором или энхансером. Регуляторная последовательность может представлять собой любой эукариотический промотор или энхансер, включая, например, EF1α, PGK, промоторный-энхансерный элемент вируса мышиного лейкоза Молони, энхансер цитомегаловируса человека, промотор коровьей оспы P7.5 или подобные (также см. примеры, перечисленные в таблицах 1 и 2 ниже). В некоторых случаях, таких как промоторный-энхансерный элемент вируса мышиного лейкоза Молони, промоторные-энхансерные элементы располагаются внутри последовательностей LTR или прилегают к ним. Предпочтительно, регуляторная последовательность не является эндогенной для лентивируса, из которого сконструирован вектор. Таким образом, если вектор создан из ВИО, регуляторная последовательность ВИО, обнаруженная в LTR ВИО, будет заменена регуляторным элементом, который не происходит из ВИО.
Можно далее нацелить рекомбинантный вирус путем связывания оболочечного белка с антителом или конкретным лигандом для нацеливания на рецептор конкретного клеточного типа. При вставке интересующей последовательности (включая регуляторную область) в вирусный вектор вместе с другим геном, который кодирует лиганд для рецептора на конкретной клетке-мишени, например, вектор становится специфичным в отношении мишени. Ретровирусные векторы могут быть сделаны специфичными в отношении мишени путем вставки, например, гликолипида или белка. Нацеливание часто осуществляется путем использования антигенсвязывающей части антитела или рекомбинантной молекулы типа антитела, такой как одноцепочечное антитело, для нацеливания ретровирусного вектора. Специалистам в данной области известны конкретные способы достижения доставки ретровирусного вектора к конкретной мишени, или они могут легко выявить их без излишнего экспериментирования.
Гетерологичная или чужеродная последовательность нуклеиновой кислоты, такая как полинуклеотидная последовательность, кодирующая такой ген, как ген для лечения врожденных или приобретенных кроветворных нарушений, указанных здесь, функционально связана с регуляторной последовательностью нуклеиновой кислоты. Предпочтительно, гетерологичная последовательность связана с промотором, с получением в результате химерного гена.
Маркерные гены могут использоваться в анализе наличия вектора и, таким образом, для подтверждения инфекции и интегрирования. Наличие маркерного гена гарантирует отбор и рост только тех клеток-хозяев, которые экспрессируют вставки. Обычные гены отбора кодируют белки, которые обеспечивают устойчивость к антибиотикам и другим токсичным веществам, например, гистидинол, пуромицин, гигромицин, неомицин, метотрексат и маркеры клеточной поверхности.
Рекомбинантный вирус по изобретению способен к переносу последовательности нуклеиновой кислоты в клетку млекопитающего. Термин «последовательность нуклеиновой кислоты» относится к любой молекуле нуклеиновой кислоты, предпочтительно, ДНК, как подробно обсуждается здесь. Молекула нуклеиновой кислоты может быть выделена из разнообразных источников, включая ДНК, кДНК, синтетическую ДНК, РНК или их комбинации. Такие последовательности нуклеиновой кислоты могут содержать геномную ДНК, которая может включать в себя встречающиеся в природе интроны и может не содержать их. Более того, такая геномная ДНК может быть получена в ассоциации с промоторными областями, поли-A-последовательностями или с другими ассоциированными последовательностями. Геномная ДНК может быть экстрагирована и очищена из подходящих клеток средствами, хорошо известными в данной области. Альтернативно, РНК-мессенджер (мРНК) может быть выделена из клеток и использована для продукции кДНК посредством обратной транскрипции или другими способами.
Векторы вводят путем трансфекции или инфекции в упаковывающую клеточную линию. Упаковывающая клеточная линия продуцирует вирусные частицы, которые содержат векторный геном. Способы трансфекции или инфекции хорошо известны специалисту в данной области. После совместной трансфекции упаковывающих векторов и вектора переноса в упаковывающую клеточную линию, рекомбинантный вирус выделяют из культуральной среды и титруют стандартными способами, используемыми специалистами в данной области. Таким образом, упаковывающие конструкции могут быть введены в линии клеток человека путем трансфекции фосфатом кальция, липофекции или электропорации, в основном вместе с доминантным селектируемым маркером, таким как неомицин, DHFR, глутаминсинтетаза или ADA, с последующей селекцией в присутствии подходящего лекарственного средства и выделением клонов. Селектируемый маркерный ген может быть физически связан в конструкцию с упаковывающими генами.
Известны стабильные клеточные линии, в которых сконфигурированы упаковывающие функции для экспрессии подходящими упаковывающими клетками. Например, см. патент США №5686279; и Ory et al., (1996), где описаны упаковывающие клетки. Упаковывающие клетки с введенным в них лентивирусным вектором образуют клетки-продуценты. Клетки-продуценты, таким образом, представляют собой клетки или клеточные линии, которые могут продуцировать или высвобождать упакованные инфекционные вирусные частицы, несущие интересующий терапевтический ген. Данные клетки, кроме того, могут быть зависимыми от прикрепления, и это означает, что данные клетки растут, выживают или оптимально поддерживают функцию, будучи прикрепленными к поверхности, такой как стекло или пластик. Клетки-продуценты могут также представлять собой неопластически трансформированные клетки. Некоторыми примерами зависимых от прикрепления клеточных линий, используемых в качестве клеток упаковки лентивирусного вектора, при том, что вектор компетентен в плане репликации, являются клетки HeLa или 293 и клетки PERC.6.
В некоторых применениях, особенно, в тех, где вирус подлежит использованию для приложений генной терапии, предпочтительно, чтобы вектор характеризовался дефицитом репликации (или дефектом репликации) во избежание неконтролируемой пролиферации вируса у подверженного лечению субъекта. В таких случаях выбирают клеточные линии млекопитающих, которые были сконструированы путем модификации генома клетки-продуцента, чтобы он кодировал присущие вирусу функции, или путем совместной инфекции клетки-продуцента хелперным вирусом, для экспрессии белков, замещающих эффект последовательностей, удаленных из вирусного генома. Например, для векторов, происходящих из ВИЧ-1, может использоваться клеточная линия упаковки ВИЧ-1, PSI422, как описано в Corbeau, et al. (1996). Аналогично, где вирусный вектор должен продуцироваться ретровирусом, может быть использована происходящая от 293 упаковывающая ретровирусы человеческая клеточная линия (293GPG), способная продуцировать высокие титры ретровирусных частиц, как описано в Ory, et al. (1996). При продукции минимальных векторных систем конструируют клетку-продуцент (путем модификации вирусного генома или путем использования вспомогательного вируса или космиды) для дополнения функций родительского вируса, что обеспечивает репликацию и упаковку в вирионы в линии клеток-продуцентов.
Лентивирусные векторы переноса по Naldini et al., (1996), использовали для инфицирования клеток человека с остановкой роста in vitro и для трансдукции нейронов после прямой инъекции в мозг взрослых крыс. Вектор был эффективным при переносе маркерных генов in vivo в нейроны, и была достигнута долгосрочная экспрессия в отсутствие детектируемой патологии. Животных анализировали через десять месяцев после единичной инъекции вектора, и было показано отсутствие снижения среднего уровня экспрессии трансгена и отсутствие признаков патологии ткани или иммунной реакции (Blomer et al., 1997).
2. SIN-конструкция
SIN-конструкция повышает биобезопасность лентивирусных векторов. Большая часть LTR ВИЧ состоит из последовательностей U3. Область U3 содержит энхансерный и промоторный элементы, которые модулируют базовую и индуцированную экспрессию генома ВИЧ в инфицированных клетках и в ответ на активацию клеток. Некоторые из данных промоторных элементов являются существенными для репликации вируса. Некоторые из энхансерных элементов высоко консервативны среди вирусных изолятов и задействованы в качестве критических факторов вирулентности в вирусном патогенезе. Энхансерные элементы могут действовать, влияя на скорость репликации в различных клеточных мишенях вируса (Marthas et al., 1993).
Поскольку вирусная транскрипция начинается на 3'-конце U3-области 5'-LTR, те последовательности, которые не являются частью вирусной мРНК и ее копии, из 3'-LTR действуют в качестве матрицы для получения обоих LTR в интегрированном провирусе. Если 3'-копия U3-области в ретровирусной векторной конструкции изменена, векторная РНК все еще продуцируется с интактной 5'-LTR в клетках-продуцентах, но не может регенерироваться в клетках-мишенях. Трансдукция такого вектора приводит к инактивации обоих LTR в вирусах потомства. Таким образом, ретровирус является самоинактивирующимся (SIN), и такие векторы известны как SIN-векторы переноса.
SIN-конструкция более подробно описана в Zufferey et al., 1998, и патенте США №5994136, причем оба источника включены сюда в качестве ссылки. Однако, как описано здесь, имеются пределы делеции при 3'-LTR. Вначале, 5'-конец U3-области выполняет иную существенную функцию при переносе вектора, что требуется для интегрирования (концевой динуклеотид + последовательность att). Таким образом, концевой динуклеотид и последовательность att может представлять собой 5'-концевую границу последовательностей U3, которая может быть удалена. Кроме того, некоторые неточно определенные области могут влиять на активность нижележащего участка полиаденилирования в R-области. Избыточная делеция последовательности U3 из 3'-LTR может снижать полиаденилирование векторных транскриптов с неблагоприятными последствиями как для титра вектора в клетках-продуцентах, так и для экспрессии трансгена в клетках-мишенях. С другой стороны, ограниченные делеции могут не нарушать транскрипционную активность LTR в трансдуцированных клетках.
Описанные здесь лентивирусные векторы могут нести делеции U3-области 3'-LTR, простирающиеся от нуклеотида -418 до -18. Это наиболее обширная делеция, и она продлевается до TATA-бокса, нарушая любую транскрипционную активность LTR в трансдуцированных клетках. Титр вектора в клетках-продуцентах, а также экспрессия трансгена в клетках-мишенях в таких векторах не подвергались воздействию. Таким образом, данная конструкция обеспечивает исключительное увеличение безопасности вектора.
Векторы SIN-типа с такими обширными делециями U3-области могут генерироваться для ретровирусных векторов, основанных на вирусе мышиного лейкоза (MLV) или вирусе некроза селезенки (SNV), без ослабления эффективности трансдукции.
Удаление нуклеотидной последовательности от -418 до -18 прекращает транскрипционную активность LTR, отменяя, таким образом, продукцию полноразмерной векторной РНК в трансдуцированных клетках. Однако в происходящих из ВИЧ лентивекторах ни свойства in vitro, ни свойства in vivo не нарушались за счет SIN-конструкции.
3. Посттранскрипционные регуляторные элементы (PRE)
Увеличение экспрессии трансгена может потребоваться для некоторых осуществлений, особенно тех, где используются лентивирусные конструкции по настоящему изобретению с умеренными промоторами.
Одним из типов PRE является интрон, расположенный внутри экспрессирующей кассеты, который может стимулировать генную экспрессию. Однако интроны могут быть вырезаны в течение событий жизненного цикла лентивируса. Следовательно, если интроны используют в качестве PRE, их следует поместить в противоположной ориентации по отношению к геномному транскрипту вектора.
Посттранскрипционные регуляторные элементы, которые не имеют отношения к событиям сплайсинга, предоставляют то преимущество, что они не удаляются во время жизненного цикла вируса. Некоторыми примерами являются элементы посттранскрипционного процессинга вируса простого герпеса, посттранскрипционный регуляторный элемент вируса гепатита B (HPRE) и вируса гепатита североамериканского лесного сурка (WPRE). Из них наиболее предпочтительным является WPRE, поскольку он содержит дополнительный, действующий в цис-направлении элемент, не обнаруженный в HPRE (Donello et al., 1998). Данный регуляторный элемент расположен внутри вектора, так что он включен в РНК-транскрипт трансгена, но вне стоп-кодона трансляционной единицы трансгена. Как продемонстрировано в настоящем изобретении и в Zufferey et al., 1999, WPRE-элемент является подходящим инструментом для стимуляции и усиления генной экспрессии желаемых трансгенов в контексте лентивирусных векторов.
WPRE охарактеризован и описан в патенте США №6136597, включенном сюда в качестве ссылки. Как описано здесь, WPRE представляет собой элемент экспорта РНК, который опосредует эффективный транспорт РНК из ядра в цитоплазму. Он усиливает экспрессию трансгенов путем вставки действующей в цис-направлении последовательности нуклеиновой кислоты, так что данный элемент и трансген содержатся в одном транскрипте. Присутствие WPRE в смысловой ориентации, как было показано, усиливает экспрессию трансгена в 7-10 раз. Ретровирусные векторы переносят последовательности в форме кДНК вместо полных, содержащих интрон генов, поскольку интроны в основном вырезаются в течение последовательности событий, ведущей к образованию ретровирусной частицы. Интроны опосредуют взаимодействие первичных транскриптов с аппаратом сплайсинга. Поскольку процессинг РНК аппаратом сплайсинга способствует их экспорту в цитоплазму вследствие соединения между аппаратами сплайсинга и транспорта, кДНК часто экспрессируются неэффективно. Так, включение в вектор WPRE приводит к усиленной экспрессии трансгенов.
4. Промоторы и энхансеры
«Промотор» представляет собой контрольную последовательность, которая является областью последовательности нуклеиновой кислоты, в которой контролируется инициация и скорость транскрипции. Он может содержать генные элементы, в которых могут связываться регуляторные белки и молекулы, такие как РНК-полимераза и другие факторы транскрипции, для инициации специфической транскрипции последовательности нуклеиновой кислоты. Выражения «функционально расположенный», «функционально связанный», «под контролем» и «под транскрипционным контролем» означают, что промотор находится в корректном функциональном расположении и/или ориентации по отношению к последовательности нуклеиновой кислоты для контроля инициации транскрипции и/или экспрессии такой последовательности.
Промотор, в общем, содержит последовательность, которая функционирует с расположением участка старта для синтеза РНК. Ее лучшим известным примером является TATA-бокс, но в некоторых промоторах, лишенных TATA-бокса, таких как, например, промотор гена терминальной дезоксинуклеотидилтрансферазы млекопитающего и промотор поздних генов SV40, другой элемент, прилегающий к участку старта, способствует точному определению места инициации. Дополнительные промоторные элементы регулируют частоту инициации транскрипции. Обычно они локализованы в области на 30-110 н.п. выше участка старта, хотя некоторые промоторы, как было показано, содержат также функциональные элементы ниже участка старта. Для помещения кодирующей последовательности «под контроль промотора» располагают 5'-конец участка инициации транскрипции в транскрипционной рамке считывания «ниже» (т.е. в 3'-направлении) выбранного промотора. «Вышележащий» промотор стимулирует транскрипцию ДНК и способствует экспрессии кодируемой РНК.
Разрядка между элементами промотора часто является изменчивой, так что функция промотора сохраняется, когда его элементы обращены или сдвинуты относительно друг друга. В промоторе tk разрядка между элементами промотора может увеличиваться на 50 н.п., до тех пор, пока не начнет снижаться его активность. В зависимости от промотора, оказывается, что отдельные элементы могут функционировать по активации транскрипции кооперативно или независимо. Промотор может использоваться в связи с «энхансером», что означает действующую в цис-направлении регуляторную последовательность, участвующую в активации транскрипции последовательности нуклеиновой кислоты, или энхансера может не быть.
Промотор может быть ассоциирован в природе с последовательностью нуклеиновой кислоты, которая может быть получена путем выделения 5'-некодирующих последовательностей, локализованных выше кодирующего сегмента и/или экзона. Такой промотор может обозначаться как «эндогенный». Также и энхансер может быть в природе ассоциирован с последовательностью нуклеиновой кислоты, расположенной ниже или выше такой последовательности. Альтернативно, некоторые преимущества можно получить путем расположения кодирующего сегмента нуклеиновой кислоты под контролем рекомбинантного или гетерологичного промотора, который относится к промотору, обычно не ассоциированному с последовательностью нуклеиновой кислоты в ее природном окружении. Рекомбинантный или гетерологичный энхансер также относится к энхансеру, обычно не ассоциированному с последовательностью нуклеиновой кислоты в ее природном окружении. Такие промоторы или энхансеры могут включать в себя промоторы или энхансеры других генов и промоторы или энхансеры, выделенные из какого-либо другого вируса, или прокариотической, или эукариотической клетки, и промоторы или энхансеры, не «встречающиеся в природе», т.е. содержащие различные элементы различных областей регуляции транскрипции, и/или мутации, которые изменяют экспрессию. Например, промоторы, которые наиболее часто используют в конструкции рекомбинантной ДНК, включают в себя промоторные системы β-лактамазы (пенициллиназы), оперонов лактозы и триптофана (trp). В дополнение к синтетической продукции последовательностей нуклеиновой кислоты промоторов и энхансеров, последовательности можно продуцировать, используя рекомбинантное клонирование и/или технологию амплификации нуклеиновой кислоты, включая PCR™, в сочетании с описанными здесь композициями (см. патенты США №4683202 и 5928906, каждый из которых включен сюда в качестве ссылки). Более того, подразумевается, что могут также использоваться контрольные последовательности, которые направляют транскрипцию и/или экспрессию последовательностей в неядерных органеллах, таких как митохондрии, хлоропласты и тому подобное. Контрольные последовательности, охватывающие промоторы, энхансеры и другие элементы контроля/модулирования локуса или транскрипции, также обозначаются как «транскрипционные кассеты».
Естественно, важно использовать промотор и/или энхансер, который эффективно направляет экспрессию сегмента ДНК в органелле, клеточном типе, ткани, органе или организме, выбранном для экспрессии. Специалистам в области молекулярной биологии, в основном, известно применение промоторов, энхансеров и комбинаций клеточных типов для экспрессии белка (см., например, Sambrook et al., 1989, включенный сюда в качестве ссылки). Используемые промоторы могут быть конститутивными, зависимыми от ткани, индуцируемыми и/или могут использоваться в условиях, подходящих для направления высокоуровневой экспрессии введенного сегмента ДНК, такими как те, что предпочтительны для генной терапии или таких применений, как крупномасштабная продукция рекомбинантных белков и/или пептидов. Промотор может быть гетерологичным или эндогенным.
Применение цитоплазматической системы экспрессии T3, T7 или SP6 является другим возможным осуществлением. Эукариотические клетки могут поддерживать транскрипцию в цитоплазме с других бактериальных промоторов, если предоставляется подходящая бактериальная полимераза, как часть комплекса доставки или как дополнительная конструкция генной экспрессии.
В таблице 1 перечислены неограничивающие примеры элементов/промоторов, которые могут использоваться в контексте настоящего изобретения для регуляции экспрессии РНК. В таблице 2 предоставлены неограничивающие примеры индуцируемых элементов, которые представляют собой области последовательности нуклеиновой кислоты, которые могут активироваться в ответ на конкретный стимул.
Промотор и/или энхансер
Индуцируемые элементы
Poly(rc)
Отличительные черты тканеспецифических промоторов или элементов, а также анализы для характеристики их активности хорошо известны специалистам в данной области. Неограничивающие примеры таких областей охватывают человеческий ген LIMK2 (Nomoto et al., 1999), ген соматостатинового рецептора 2 (Kraus et al., 1998), ген связывающего ретиноевую кислоту белка придатков яичка мыши (Lareyre et al., 1999), человеческий CD4 (Zhao-Emonet et al., 1998), мышиный альфа-2-(XI)-коллаген (Tsumaki, et al., 1998), ген дофаминового рецептора D1A (Lee, et al., 1997), инсулиноподобный фактор роста II (Wu et al., 1997) и молекулу-1 адгезии тромбоцитов-эндотелиальных клеток человека (Almendro et al., 1996).
Лентивирусные векторы по настоящему изобретению конструируют вначале для трансформации клеток терапевтическим геном под контролем регулируемых эукариотических промоторов. Хотя предпочтительным является промотор gp91-phox, могут использоваться также другие промоторные и регуляторные сигнальные элементы, описанные выше в таблицах 1 и 2. Кроме того, также может использоваться любая комбинация промотор/энхансер (согласно базе данных эукариотических промоторов EPDB) для направления экспрессии структурных генов, кодирующих интересующий терапевтический ген, который используется в контексте с лентивирусными векторами по настоящему изобретению. Альтернативно, тканеспецифический промотор для генной терапии злокачественных опухолей или для прицельного воздействия на опухоль может использоваться с лентивирусными векторами по настоящему изобретению для лечения злокачественных опухолей, особенно, гематологических злокачественных заболеваний.
Обычно промоторы и энхансеры, которые контролируют транскрипцию белка, кодирующего гены в эукариотических клетках, составлены из множественных генетических элементов. Клеточный аппарат способен объединить и интегрировать регуляторную информацию, которую несет каждый элемент, что позволяет различным генам образовывать различные, зачастую комплексные профили транскрипционной регуляции. Активация или репрессия промоторных и энхансерных элементов может осуществляться путем контакта данных элементов с подходящими транскрипционными активаторами или репрессорами, такими как описанные на фиг.1B для промотора gp91-phox и раскрытые в Luo and Skalnik (1996) J. Biol. Chem. 271: 18203-210, and Luo and Skalnik (1996) J. Biol. Chem. 271: 23445-23451, включенных сюда в качестве ссылки. В отношении промотора gp91-phox, активность интерферона-гамма в модулировании транскрипции и экспрессии экспрессионной кассеты является примером того, как такие промоторные или энхансерные элементы и факторы, которые с ними взаимодействуют, могут быть использованы в воплощении настоящего изобретения.
Энхансеры вначале были выявлены как генетические элементы, которые повышали транскрипцию промотора, локализованные в отдаленном положении на той же молекуле ДНК. Данная способность действовать на большом расстоянии редко имела место при классических исследованиях регуляции транскрипции у прокариот. Последующая работа показала, что области ДНК с энхансерной активностью организованы во многом подобно промоторам. То есть они состоят из многих отдельных элементов, каждый из которых связывается с одним или несколькими транскрипционными белками. См., например, модель регуляции промотором gp91-phox, представленную на фиг.1B. Типичными энхансерами, рассматриваемыми в настоящем изобретении, являются гиперчувствительные к ДНКазе элементы и их гомологи, описанные в публикации Lien LL, Lee Y, Orkin SH, (1997) «Regulation of the myeloid-cell-expressed human gp91-phox gene as studied by transfer of yeast artificial chromosome clones into embryonic stem cells: suppression of a variegated cellular pattern of expression requires a full complement of distant cis elements», Mol Cell Biol. 17(4): 2279-90, специально включенной сюда полностью в качестве ссылки. Под влиянием данных энхансерных элементов генная экспрессия может увеличиваться (вследствие энхансерной активности HS) и быть менее вариабельной (вследствие сайленсерной активности HS).
Аналоги HS-элементов gp91-phox активны в других промоторных-энхансерных системах. См., например, публикацию May C, Rivella S, Callegari J, Heller G, Gaensler K M, Luzzatto L, Sadelain M, (2000) Therapeutic haemoglobin synthesis in beta-thalassaemic mice expressing lentivirus-encoded human beta-globin. Nature 406(6791): 82-6, специально включенную сюда полностью в качестве ссылки, где аналогичные HS-элементы бета-глобина встроены в лентивектор выше промотора бета-глобина для направления экспрессии кДНК бета-глобина.
Промоторы и энхансеры характеризуются одинаковой общей функцией активации транскрипции в клетке. Они часто перекрываются и смежны по отношению друг к другу, и часто представляется, что они имеют сходную модульную организацию. Данные объединенные соображения указывают на то, что энхансеры и промоторы представляют собой гомологичные объекты и что белки-активаторы транскрипции, связывающиеся с данными последовательностями, могут взаимодействовать с клеточным аппаратом транскрипции фундаментально одним и тем же образом. Основное отличие энхансеров и промоторов является практическим. Энхансерная область как целое должна быть способна к стимуляции транскрипции на расстоянии; это может не быть обязательным для промоторной области или составляющих ее элементов. С другой стороны, промотор должен иметь один или несколько элементов, которые направляют инициацию синтеза РНК в конкретном участке и в конкретной ориентации, в то время как энхансеры не обладают такими конкретными чертами. Без этого практического отличия энхансеры и промоторы представляют собой очень сходные объекты. Конструкции элементов, которые контролируют транскрипцию и экспрессию, могут поэтому состоять из различных элементов, систематизированных так, что предоставляются способы контроля увеличенной полезности и эффективности.
Сигнал, который может оказаться применимым, представляет собой сигнал полиаденилирования (hGH, BGH, SV40). Использование элементов внутренних участков связывания рибосомы (IRES) применяли для создания мультигенных или полицистронных мРНК. IRES-элементы способны обходить модель сканирования рибосомой 5'-метилированной кэп-зависимой трансляции на внутренних участках (Pelletier and Sonenberg, 1988). Описаны IRES-элементы двух представителей семейства пикорнавирусов (полиомиелит и энцефаломиокардит) (Pelletier and Sonenberg, 1988), а также IRES из мРНК млекопитающих (Macejak and Sarnow, 1991). IRES-элементы могут связываться с гетерологичными открытыми рамками считывания. Множественные открытые рамки считывания могут транскрибироваться совместно, каждая отделена IRES, с образованием полицистронных мРНК. При наличии IRES-элемента каждая открытая рамка считывания доступна рибосомам для эффективной трансляции. Множественные гены могут эффективно экспрессироваться с использованием единственного промотора/энхансера для транскрипции одной мРНК.
В любом случае, понятно, что промоторы представляют собой элементы ДНК, которые при расположении выше гена приводят к экспрессии данного гена. Большая часть трансгенов, которыми трансформируют с использованием лентивирусных векторов по настоящему изобретению, функционально расположены ниже промоторного элемента.
Конкретный сигнал инициации также может потребоваться для эффективной трансляции кодирующих последовательностей. Данные сигналы охватывают инициирующий кодон ATG или прилегающие последовательности. Экзогенные сигналы контроля трансляции, включая инициирующий кодон ATG, могут оказаться необходимыми. Обычный специалист в данной области может легко определить их и предоставить необходимые сигналы. Хорошо известно, что инициирующий кодон должен находиться «в рамке» с открытой рамкой считывания желаемой кодирующей последовательности для обеспечения трансляции целой вставки. Сигналы экзогенного контроля трансляции и инициирующие кодоны могут быть природными или синтетическими. Эффективность экспрессии может быть усилена включением подходящих энхансерных элементов транскрипции.
5. Краткое описание списков последовательностей
SEQ ID NO:1 предоставляет нуклеотидную последовательность элемента центрального полипуринового тракта ВИЧ, взятого из ВИЧ, о котором сообщали Follenzi, A, Ailles, L.E., Bakovic, S., Geuna, M., Naldini, L. (2000) "Gene Transfer by Lentiviral Vecotrs is Limited by Nuclear Translocation and Rescued by HIV-1 pol Sequences," Nat. Genet. 25:217-22. SEQ ID NOS:2-6 предоставляют нуклеотидные последовательности, охватывающие участки, гиперчувствительные к ДНКазе, и используются в качестве энхансеров в некоторых осуществлениях изобретения. См. Lien LL, Lee Y, and Orkin SH, "Regulation of the myeloid-cell-expressed human gp91-phox gene as studied by transfer of yeast artificial chromosome clones into embryonic stem cells: suppression of a variegated cellular pattern of expression requires a full complement of distant cis elements," Mol. Cell. Biol. 17(4): 2279-90 (1997).
SEQ ID NOS:7-16 предоставляют нуклеотидные последовательности PCR-праймеров, используемых для получения HS-элементов SEQ ID NOS:2-6. Последовательности PCR-праймеров и HS-элементов основаны на последовательности генома человека, опубликованной в рамках проекта «Геном человека» (контиг NT-011844; http://www.ncbi.nlm.nih.gov/genome/guide/human/). Положения амплифицированных путем PCR HS-элементов (+1 - старт транскрипции gp91-phox) в последовательности ДНК генома человека представляют собой: HS-12 (-11503, -13244), HS-14 (-13244, -14715), HS-26 (-25345, -26529), HS-27 (-26529, -27656), HS-28 (-27657, -28893). Используемые фрагменты немного превышают рассчитанные вследствие включения на концах участков рестрикции для облегчения клонирования (HS-12 и HS-14; gta для восстановления 5'-концевого SnaBI после клонирования в данный участок pHPX-GFP; HS-26, HS27, HS-28; SalI; gcgtcgac и XhoI; ctcgagcggc).
Другие последовательности, включенные в конкретные осуществления настоящего изобретения, охватывают те, которые кодируют gp91-phox и их гомологи, включая нуклеотидные последовательности, раскрытые под инвентарным номером Генбанка NM000397 для gp91-phox (Dinauer, et al., 1987), SEQ ID NO:18 и полипептиды последовательности SEQ ID NO:19. Также включены в конкретные осуществления промоторы gp91-phox, кодируемые нуклеотидной последовательностью под инвентарным номером Генбанка M66390, SEQ ID NO:17 и промоторы CD11b, кодируемые нуклеотидной последовательностью с инвентарным номером Генбанка M82856, SEQ ID NO:20.
6. Нуклеиновые кислоты
Одним из осуществлений настоящего изобретения является перенос нуклеиновых кислот, кодирующих терапевтический ген, особенно ген, который обеспечивает лечение нарушений кроветворения и лимфогемопоэза, таких как приобретенные или врожденные нарушения, описанные выше. В одном из осуществлений нуклеиновые кислоты кодируют полноразмерную, по существу полноразмерную, или функционально эквивалентную форму такого гена.
Таким образом, в некоторых осуществлениях настоящего изобретения лечение нарушений кроветворения и лимфогемопоэза охватывает введение лентивирусного вектора по изобретению, содержащего экспрессирующую конструкцию терапевтической нуклеиновой кислоты, в клетку кроветворного происхождения. Предполагается, что кроветворные клетки захватывают данную конструкцию и экспрессируют терапевтический полипептид, кодируемый нуклеиновой кислотой, восстанавливая, таким образом, нормальный фенотип клеток.
Нуклеиновая кислота может быть получена любым способом, известным обычному специалисту в данной области. Не ограничивающие примеры синтетической нуклеиновой кислоты, в частности, синтетического олигонуклеотида, охватывают нуклеиновую кислоту, полученную путем химического синтеза in vitro с использованием фосфотриэфирной, фосфитной или фосфорамидитной химии и твердофазных способов, таких как описанные в EP 266032, включенном сюда в качестве ссылки, или посредством дезоксинуклеозид-H-фосфонатовых интермедиатов, как описано Froehler et al., 1986, и в патенте США №5705629, каждый из которых включен сюда в качестве ссылки. Неограничивающий пример продуцированной ферментно нуклеиновой кислоты охватывает такую кислоту, продуцированную ферментами в реакциях амплификации, таких как PCR (см., например, патенты США №4683202 и 4682195, каждый из которых включен сюда в качестве ссылки), или синтез олигонуклеотидов, описанный в патенте США №5645897, включенном сюда в качестве ссылки. Не ограничивающий пример биологически продуцированной нуклеиновой кислоты охватывает продукцию рекомбинантной нуклеиновой кислоты в живых клетках (см., например, Sambrook et al. 1989, включенный сюда в качестве ссылки).
Нуклеиновую кислоту можно очищать на полиакриламидных гелях, на градиентах центрифугирования в хлориде цезия, или любым другим способом, известным обычному специалисту в данной области (см., например, Sambrook et al. 1989, включенный сюда в качестве ссылки).
Термин «нуклеиновая кислота» в общем относится, по крайней мере, к одной молекуле или цепи ДНК, РНК или их производного, или их миметика, включающей, по крайней мере, одно нуклеиновое основание, такое как, например, встречающееся в природе пуриновое или пиримидиновое основание, обнаруженное в ДНК (например, аденин «A», гуанин «G», тимин «T» и цитозин «C») или в РНК (например A, G, урацил «U» и C). Термин «нуклеиновая кислота» охватывает термины «олигонуклеотид» и «полинуклеотид». Термин «олигонуклеотид» относится, по крайней мере, к одной молекуле длиной примерно от 3 до 100 нуклеиновых оснований. Термин «полинуклеотид» относится, по крайней мере, к одной молекуле длиной более чем примерно 100 нуклеиновых оснований. Данные определения в основном относятся, по крайней мере, к одной одноцепочечной молекуле, но в конкретных осуществлениях также охватывают, по крайней мере, одну дополнительную цепь, которая частично, по существу или полностью комплементарна, по крайней мере, одной одноцепочечной молекуле. Таким образом, нуклеиновая кислота может охватывать, по крайней мере, одну двухцепочечную молекулу или, по крайней мере, одну трехцепочечную молекулу, которая содержит одну или несколько комплементарных цепей или «комплементов» конкретной последовательности, содержащей цепь данной молекулы.
В конкретных осуществлениях «ген» относится к нуклеиновой кислоте, которая транскрибируется. Как используется здесь, «генный сегмент» означает сегмент нуклеиновой кислоты гена. В некоторых аспектах ген охватывает регуляторные последовательности, участвующие в транскрипции или продукции или композиции мРНК. В конкретных осуществлениях ген содержит транскрибируемые последовательности, которые кодируют белок, полипептид или пептид. В других конкретных аспектах ген содержит нуклеиновую кислоту и/или кодирует кодирующие полипептид или пептид последовательности гена, который является дефектным или мутантным при нарушении кроветворения или лимфогемопоэза. В рамках описанной здесь терминологии «выделенный ген» может содержать транскрибированную(ые) нуклеиновую(ые) кислоту(ы), регуляторные последовательности, кодирующие последовательности или тому подобное, выделенные по существу отдельно от других таких последовательностей, таких как встречающиеся в природе гены, регуляторные последовательности, последовательности, кодирующие полипептид или пептид и т.д. В этом отношении термин «ген» применяется попросту для ссылки на нуклеиновую кислоту, содержащую нуклеотидную последовательность, которая транскрибируется, и комплементарную ей последовательность. В конкретных аспектах транскрибируемая нуклеотидная последовательность содержит, по крайней мере, одну функциональную единицу, кодирующую белок, полипептид и/или пептид. Как понятно специалистам в данной области, данный функциональный термин «ген» охватывает обе геномные последовательности, последовательности РНК или кДНК, или меньшие сконструированные сегменты нуклеиновой кислоты, включая сегменты нуклеиновой кислоты не транскрибируемой части гена, включая, без ограничения, не транскрибируемые промоторные или энхансерные области гена. Меньшие сконструированные генные сегменты нуклеиновой кислоты могут экспрессировать или могут быть адаптированы для экспрессии белков, полипептидов, доменов, пептидов, белков слияния, мутантов и/или тому подобное, с использованием технологии манипуляции нуклеиновой кислотой. Таким образом, «укороченный ген» относится к последовательности нуклеиновой кислоты, в которой не хватает участка следующих друг за другом остатков нуклеиновой кислоты.
Различные сегменты нуклеиновой кислоты могут быть сконструированы на основе конкретной последовательности нуклеиновой кислоты и могут быть любой длины. Путем присвоения последовательности численных значений, где, например, первый остаток представляет собой 1, второй остаток представляет собой 2, и т.д., может быть создан алгоритм, определяющий все сегменты нуклеиновой кислоты:
от n до n+y,
где n представляет собой целое число от 1 до последнего числа последовательности, и y представляет собой длину сегмента нуклеиновой кислоты минус один, где n+y не превышает последнего числа последовательности. Таким образом, для 10-членника сегменты нуклеиновой кислоты соответствуют основаниям от 1 до 10, от 2 до 11, от 3 до 12... и/или так далее. Для 15-членника сегменты нуклеиновой кислоты соответствуют основаниям от 1 до 15, от 2 до 16, от 3 до 17... и/или так далее. Для 20-членника сегменты нуклеиновой кислоты соответствуют основаниям от 1 до 20, от 2 до 21, от 3 до 22... и/или так далее.
Нуклеиновая(ые) кислота(ы) по настоящему изобретению, независимо от длины последовательности как таковой, может(гут) комбинироваться с другими последовательностями нуклеиновой кислоты, включая, без ограничения, промоторы, энхансеры, сигналы полиаденилирования, участки ферментов рестрикции, множественные участки клонирования, кодирующие сегменты и тому подобное, для создания одной или нескольких конструкций нуклеиновой кислоты. Общая длина конструкций нуклеиновой кислоты может значительно варьировать. Таким образом, могут использоваться сегменты нуклеиновой кислоты почти любой длины с общей длиной, которая предпочтительно ограничена простотой получения или применением в соответствии с нужной методикой рекомбинантных нуклеиновых кислот.
Термин «вектор» применяется для ссылки на несущую молекулу нуклеиновой кислоты, в которую может быть встроена последовательность нуклеиновой кислоты для введения в клетку, где она может реплицироваться. Векторы по настоящему изобретению основаны на лентивирусах, как описано выше и в других частях спецификации. Молекулы нуклеиновой кислоты, переносимые векторами по изобретению, кодируют терапевтические гены и будут использованы для проведения способов генной терапии. Специалист в данной области снабжен всем необходимым для конструирования такого терапевтического вектора с использованием стандартных рекомбинантных способов (см., например, Maniatis et al., 1988 и Ausubel et al., 1994, причем оба источника включены в качестве ссылки).
Термин «экспрессирующий вектор» относится к любому типу генетической конструкции, содержащей нуклеиновую кислоту, кодирующую РНК, способную транскрибироваться. В некоторых случаях молекулы РНК затем транслируются в белок, полипептид или пептид. В других случаях данные последовательности не транслируются, например, при продукции антисмысловых молекул или рибозимов. Экспрессирующие векторы могут содержать различные «контрольные последовательности», которые относятся к последовательностям нуклеиновой кислоты, необходимым для транскрипции и, возможно, трансляции функционально связанной кодирующей последовательности в конкретной клетке-хозяине. В дополнение к контрольным последовательностям, которые управляют транскрипцией и трансляцией, векторы и экспрессирующие векторы могут содержать последовательности нуклеиновой кислоты, которые выполняют также другие функции, и описаны ниже.
A. Множественные участки клонирования
Векторы по настоящему изобретению могут включать в себя множественный участок клонирования (MCS), представляющий собой область нуклеиновой кислоты, которая содержит множественные участки рестрикционных ферментов, любой из которых можно использовать в сочетании со стандартной рекомбинантной технологией для расщепления вектора (см., например, Carbonelli et al., 1999, Levenson et al., 1998, и Cocea, 1997, включенные сюда в качестве ссылок). «Расщепление рестрикционным ферментом» означает каталитическое расщепление молекулы нуклеиновой кислоты ферментом, который функционирует только в определенном положении в молекуле нуклеиновой кислоты. Большинство из этих рестрикционных ферментов являются коммерчески доступными. Применение таких ферментов в значительной степени понятно специалистам в данной области. Зачастую, вектор линеаризуется или фрагментируется с использованием рестрикционного фермента, который разрезает молекулу в MCS, чтобы предоставить возможность экзогенным последовательностям лигироваться с вектором. «Лигирование» означает процесс образования фосфодиэфирных связей между двумя фрагментами нуклеиновых кислот, которые могут быть соседними друг с другом или нет. Способы, задействующие рестрикционные ферменты и реакции лигирования, хорошо известны специалистам в области рекомбинантной технологии.
В. Участки сплайсинга
Большинство транскрибируемых эукариотических молекул РНК подвергаются сплайсингу РНК для удаления интронов из первичных транскриптов. Для векторов, содержащих геномные эукариотические последовательности, могут требоваться донорные и/или акцепторные участки сплайсинга для обеспечения надлежащего процессинга транскрипта для экспрессии белка (см., например, Chandler et al., 1997, включенную сюда в качестве ссылки).
С. Сигналы терминации
Векторы или конструкции по настоящему изобретению, как правило, будут включать в себя, по крайней мере, один терминальный сигнал. «Терминальный сигнал» или «терминатор» находится в последовательностях ДНК, вовлеченных в специфическую терминацию транскрипта РНК РНК-полимеразой. Таким образом, в некоторых осуществлениях рассматривается сигнал терминации, который завершает образование транскрипта РНК. Терминатор может быть необходим in vivo для достижения требуемых уровней мРНК.
В эукариотических системах область терминации может также включать в себя определенные последовательности ДНК, которые допускают сайт-специфическое расщепление нового транскрипта с тем, чтобы сделать доступным участок полиаденилирования. Это дает сигнал определенной эндогенной полимеразе добавить участок приблизительно в 200 остатков А (поли A) к 3'-концу транскрипта. Молекулы РНК, модифицированные при помощи такого конца поли A, по-видимому, являются более стабильными и транслируются более эффективно. Таким образом, в других осуществлениях, касающихся эукариот, предпочтительно, чтобы терминатор включал в себя сигнал для расщепления РНК, и наиболее предпочтительно, чтобы терминаторный сигнал способствовал полиаденилированию мРНК. Терминатор и/или элементы участка полиаденилирования могут служить для увеличения уровней мРНК и для минимизации сквозного считывания с кассеты в других последовательностях.
Терминаторы, рассматриваемые для применения в изобретении, включают в себя любой терминатор транскрипции, описанный здесь или известный обычному специалисту в данной области, включая, без ограничения, например, последовательности терминации генов, таких как, например, терминатор бычьего гормона роста или вирусные последовательности терминации, такие как, например, терминатор SV40. В некоторых осуществлениях сигналом терминации может быть отсутствие последовательности, подходящей для транскрипции или трансляции, например, в результате укорочения последовательности.
D. Сигналы полиаденилирования
При экспрессии эукариотического гена он обычно включает в себя сигнал полиаденилирования для осуществления надлежащего полиаденилирования транскрипта. Предполагается, что природа сигнала полиаденилирования не является решающей для успешной практики изобретения, и может применяться любая такая последовательность. Некоторые примеры включают в себя сигнал полиаденилирования SV40 или сигнал полиаденилирования бычьего гормона роста, подходящий и, как известно, хорошо функционирующий в различных клетках-мишенях. Полиаденилирование может увеличивать стабильность транскрипта или может облегчать цитоплазматический транспорт.
E. Точки инициации репликации
Для того чтобы внедрить вектор по изобретению в клетку-хозяина, он может содержать один или более участков инициации репликации (часто называемых «ori»), представляющих собой определенную последовательность нуклеиновой кислоты, в которой инициируется репликация. Альтернативно, можно применять автономно реплицирующуюся последовательность (ARS), если клеткой-хозяином являются дрожжи.
F. Маркеры, подходящие для селекции и скрининга
В некоторых осуществлениях изобретения клетки, трансдуцированные лентивекторами по настоящему изобретению, можно идентифицировать in vitro или in vivo путем включения маркера в экспрессирующий вектор. Такие маркеры придают опознаваемые изменения трансдуцированным клеткам, позволяя легко определить содержащие экспрессирующий вектор клетки. В основном, подходящим для селекции маркером является маркер, придающий свойство, которое позволяет проводить селекцию. Положительным подходящим для селекции маркером является тот, в котором наличие маркера позволяет проводить его селекцию, в то время как подходящим для селекции отрицательным маркером является тот, в котором его наличие предотвращает его селекцию. Примером положительного подходящего для селекции маркера является маркер резистентности к лекарственному средству.
Обычно включение маркера селекции по лекарственному средству способствует клонированию и определению трансформантов, например, генетические конструкции, которые придают резистентность к неомицину, пиромицину, гигромицину, DHFR, GPT, зеоцину и гистидинолу, являются применимыми, подходящими для селекции маркерами. В дополнение к маркерам, придающим фенотип, который позволяет проводить дифференциацию трансформантов, основанную на реализации условий, рассматриваются также другие типы маркеров, включающие в себя такие подходящие для скрининга маркеры, как GFP, чьей основой является колориметрический анализ. Альтернативно, можно использовать такие подходящие для скрининга ферменты, как тимидинкиназа вируса простого герпеса (tk) или хлорамфениколацетилтрансфераза (CAT). Специалистам в данной области также известно, как применять иммунологические маркеры, возможно в комбинации с анализом FACS. Полагают, что используемый маркер не важен, при условии, что он способен экспрессироваться одновременно с нуклеиновой кислотой, кодирующей генный продукт. Дальнейшие примеры маркеров, пригодных для селекции и скрининга, хорошо известны специалистам в данной области.
7. Клетки-хозяева
Используемые здесь термины «клетка», «клеточная линия» и «клеточная культура» могут применяться взаимозаменяемо. Все из этих терминов также включают в себя их потомство, которое охватывает любые или все последующие поколения. Понятно, что все потомство не может быть идентичным из-за предумышленных или непреднамеренных изменений. В контексте экспрессии гетерологичной последовательности нуклеиновой кислоты, «клетка-хозяин» означает прокариотическую или эукариотическую клетку, и она включает в себя какие-либо трансформируемые организмы, которые способны реплицировать вектор и/или экспрессировать гетерологичную нуклеиновую кислоту, кодируемую вектором по данному изобретению. Клетка-хозяин может быть использована и используется в качестве реципиента для векторов. Клетка-хозяин может быть «трансфицированной» или «трансформированной», что означает процесс, посредством которого экзогенная нуклеиновая кислота переносится или вводится в клетку-хозяина. Трансформированные клетки включают в себя первичную клетку субъекта и ее потомство. Используемые здесь термины «конструированные» и «рекомбинантные» клетки или клетки-хозяева предназначены для обозначения клетки, в которую была введена экзогенная последовательность нуклеиновой кислоты, такая как, например, лентивектор по изобретению, несущий терапевтическую генную конструкцию. Поэтому рекомбинантные клетки отличаются от встречающихся в природе клеток, которые не содержат рекомбинантно введенной нуклеиновой кислоты.
В некоторых осуществлениях рассмотрено, что последовательности РНК или белка могут совместно экспрессироваться с другими отобранными последовательностями РНК или белка в той же самой клетке-хозяине. Совместной экспрессии можно достичь путем совместной трансфекции клетки-хозяина двумя или более различными рекомбинантными векторами. Альтернативно, один рекомбинантный вектор можно конструировать так, чтобы он содержал множественные различные кодирующие области для РНК, которые могли бы затем экспрессироваться в клетке-хозяине, трансфицированной одним вектором.
Клетки-хозяева могут происходить из прокариот или эукариот, в зависимости от того, является ли желаемым результатом репликация вектора или экспрессия части или всех кодируемых вектором последовательностей нуклеиновых кислот. Многочисленные клеточные линии и культуры доступны для использования в качестве клетки-хозяина, и их можно получить через Американскую коллекцию типовых культур (ATCC), являющуюся организацией, которая служит в качестве архива для живых культур и генетического материала (www.atcc.org). Некоторые примеры клеток-хозяев, используемых в данном изобретении, включают в себя, без ограничения, клетки, упаковывающие вирусы, клетки-продуценты вируса, клетки 293T, кроветворные клетки-предшественники человека, кроветворные стволовые клетки человека, клетки CD34+, клетки CD4+и тому подобное.
A. Ткани и клетки
Ткань может включать в себя клетку-хозяина или клетки, которые будут трансформированы или которые будут контактировать с композицией, доставляющей нуклеиновую кислоту, и/или дополнительным агентом. Ткань может быть частью организма или может быть выделена из организма. В некоторых осуществлениях ткань и ее составляющие клетки могут включать в себя, без ограничения, кровь (например, кроветворные клетки (такие как кроветворные клетки-предшественники человека, кроветворные стволовые клетки человека, клетки CD34+, клетки CD4+), лимфоциты и другие клетки ростков крови), костный мозг, мозг, стволовые клетки, кровеносные сосуды, печень, легкие, кость, молочную железу, хрящ, шею, толстую кишку, роговицу, эмбриональную ткань, эндометрий, эндотелий, эпителий, пищевод, фасции, фибробласты, ткань фолликулов, клетки ганглиев, глиальные клетки, бокаловидные клетки, почки, лимфатический узел, мышцы, нейрон, яичники, поджелудочную железу, периферическую кровь, простату, кожу, тонкую кишку, селезенку, желудок, яички.
B. Организмы
В некоторых осуществлениях клетка-хозяин или ткань может включать в себя, по крайней мере, один организм. В некоторых осуществлениях организм может быть организмом человека, примата или мыши. В других осуществлениях организмом может быть любой эукариот или даже прокариот (например, эубактерия, архея), как должно быть понятно специалисту в данной области (см., например, web-страницу http://phylogeny.arizona.edu/tree/phylogeny.html). Некоторые лентивекторы по изобретению могут использовать контрольные последовательности, которые позволяют им реплицироваться и/или экспрессироваться как в прокариотических, так и в эукариотических клетках. Специалистам в данной области следует далее понимать условия, при которых инкубируют все из перечисленных выше клеток-хозяев для их содержания и для возможности репликации вектора. Также понятными и известными являются способы и условия, которые позволяют осуществлять крупномасштабную продукцию лентивекторов по изобретению, так же как и продукцию нуклеиновых кислот, кодируемых лентивекторами, и родственных им полипептидов, белков или пептидов, часть из которых является терапевтическими генами или белками, которые будут использоваться для генной терапии.
C. Подходящие для инъекций композиции и фармацевтические составы
Для проведения генной терапии с использованием композиций лентивирусных векторов по настоящему изобретению, в общем, следует обеспечить контакт нуждающейся в этом клетки с лентивирусным вектором, содержащим терапевтический ген. Клетка далее будет введена в организм, такой как человек, нуждающийся в генной терапии. Способы введения, естественно, будут меняться в зависимости от локализации и природы заболевания и будут включать в себя, например, внутривенный, внутриартериальный, внутрикожный, трансдермальный, внутримышечный, внутриносовой, подкожный, чрескожный, внутритрахейный, внутрибрюшинный, внутриопухолевый способы введения, перфузию и лаваж. Иногда клетки будут выделяться из организмов, подвергаться взаимодействию с лентивектором ex vivo и затем заново имплантироваться.
Введение конструкций лентивирусных нуклеиновых кислот по изобретению можно проводить при помощи шприца или любого другого способа, используемого для введения раствора, поскольку экспрессирующая конструкция может проходить через иглу определенного размера, необходимого для инъекции. Недавно была описана новая безыгольная система инъекции (патент США №5846233), имеющая насадку, определяющую границы ампульной камеры для хранения раствора, и энергетическое устройство для выдавливания раствора через насадку к месту доставки. Также для использования в генной терапии была описана шприцевая система, которая позволяет осуществлять многократное введение предварительно определенных количеств раствора точно на любую глубину (патент США №5846225).
Растворы нуклеиновых кислот в виде свободных оснований или фармакологически приемлемых солей можно приготовить в воде, соответствующим образом смешанной с сурфактантом, таким как гидроксипропилцеллюлоза. Дисперсии также можно приготовить в глицерине, жидком полиэтиленгликоле и в их смеси и в маслах. При обычных условиях хранения и использования эти препараты содержат консерванты для предотвращения роста микроорганизмов. Фармацевтические формы, подходящие для применения в виде инъекций, включают в себя стерильные водные растворы или дисперсии и стерильные порошки для приготовления предназначенных для немедленного приема стерильных растворов или дисперсий, подходящих для инъекций (патент США №5466468, конкретно включенный сюда полностью в качестве ссылки). Во всех случаях форма должна быть стерильной и должна быть жидкой до такой степени, чтобы легко вводиться шприцем. Она должна быть устойчивой в условиях производства и хранения и должна охраняться от загрязняющего действия таких микроорганизмов, как бактерии и грибы. Носителем может быть растворитель или дисперсионная среда, содержащая, например, воду, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и тому подобное), их подходящие смеси и/или растительные масла. Надлежащая текучесть может поддерживаться, например, посредством использования покрытия, такого как лецитин, сохраняя необходимый размер частиц в случае дисперсии, и посредством использования сурфактантов. Предотвращение действия микроорганизмов можно осуществить, используя различные антибактериальные и противогрибковые агенты, например, парабены, хлорбутанол, фенол, сорбиновую кислоту, тимерозал и тому подобное. В большинстве случаев, предпочтительным будет включение изотонических агентов, например, сахаров или хлорида натрия. Продолжительная абсорбция композиций для инъекций может осуществляться посредством использования агентов, замедляющих абсорбцию, например, моностеарата алюминия и желатина.
Для парентерального введения в водном растворе, например, раствор, при необходимости, следует соответствующим образом буферизовать и сначала сделать жидкий разбавитель изотоническим с достаточным количеством соли или глюкозы. Эти конкретные водные растворы в основном подходят для внутривенного, внутриартериального, внутримышечного, подкожного, внутриопухолевого и внутрибрюшинного введения. В этой связи, стерильные водные среды, которые могут использоваться, хорошо известны специалистам в данной области в свете настоящего описания. Например, одну дозу можно растворить в 1 мл изотонического раствора NaCl и либо добавить к 1000 мл жидкости для гиподермоклизиса, либо ввести в предложенное место инфузии, (см., например, «Remington's Pharmaceutical Sciences» 15th Edition, страницы 1035-1038 и 1570-1580). Некоторые изменения дозировок непременно будут иметь место в зависимости от условий лечения пациента. В любом случае, лицо, ответственное за введение, будет определять соответствующую дозу для индивидуального пациента. Кроме того, для введения человеку, препараты должны соответствовать стандартам стерильности, пирогенности, общей безопасности и чистоты по требованию FDA Office of Biologics standards.
Стерильные подходящие для инъекций растворы приготавливают путем объединения активных соединений в требуемом количестве в соответствующем растворителе с различными другими перечисленными выше ингредиентами, с последующей, по требованию, стерилизацией фильтрованием. Дисперсии, как правило, приготавливают путем объединения различных стерилизованных активных ингредиентов в стерильном носителе, который содержит основную дисперсионную среду и необходимые другие из перечисленных выше ингредиенты. В случае стерильных порошков для приготовления стерильных растворов, подходящих для инъекций, предпочтительными способами приготовления являются способы вакуумной сушки и сушки замораживанием, которые приводят к образованию порошка активного ингредиента совместно с любым дополнительным необходимым ингредиентом из его раствора, предварительно стерилизованного фильтрованием.
Описанные здесь композиции можно приготовить в нейтральной или солевой форме. Фармацевтически приемлемые соли включают в себя дополнительные кислые соли и соли, образованные с неорганическими кислотами, такими как, например, хлористоводородная или фосфорная кислоты, или с такими органическими кислотами, как уксусная, щавелевая, винная, миндальная кислота и тому подобное. Соли, образованные со свободными карбоксильными группами, можно также получать из неорганических оснований, таких как, например, гидроксиды натрия, калия, аммония, кальция или железа, и таких органических оснований, как изопропиламин, триметиламин, гистидин, прокаин и тому подобное. При изготовлении состава растворы будут вводиться способом, совместимым с дозированным составом и в таком количестве, которое является терапевтически эффективным. Составы легко вводятся в разнообразных дозированных формах, таких как подходящие для инъекций растворы, капсулы, высвобождающие лекарственное средство, и тому подобное.
Используемый здесь термин «носитель» охватывает любой или все растворители, дисперсионные среды, носители, покрытия, разбавители, антибактериальные и противогрибковые агенты, изотонические и замедляющие абсорбцию агенты, буферы, растворы носителей, суспензии, коллоиды и тому подобное. Применение таких сред и агентов для фармацевтически активных веществ хорошо известно в данной области. Кроме тех случаев, когда общепринятая среда или агент несовместимы с активным ингредиентом, рассмотрено их использование в терапевтических составах. Дополнительные активные ингредиенты могут также быть добавлены в композиции.
Фраза «фармацевтически приемлемый» или «фармакологически приемлемый» означает молекулярные вещества и композиции, которые не вызывают аллергических или подобных тяжелых реакций при введении их человеку. Приготовление водной композиции, которая содержит белок в качестве активного ингредиента, хорошо понятна в данной области. Обычно, такие композиции приготавливают как подходящие для инъекций, либо в виде жидких растворов, либо суспензий; также можно приготовить твердые формы, подходящие для растворения или получения суспензии в жидкой фазе перед введением.
Термины «контактирующий» и «подвергающийся воздействию», применяемые в отношении клетки, используются здесь для того, чтобы описать процесс, посредством которого терапевтический лентивирусный вектор доставляется в клетку-мишень.
Для генной терапии изолированных твердых доступных опухолей конкретно рассмотрена внутриопухолевая инъекция или инъекция в сосудистую систему опухоли. Местное, региональное или системное введение также может быть подходящим. Для опухолей >4 см, объем, который должен быть введен, будет составлять приблизительно 4-10 мл (предпочтительно 10 мл), тогда как для опухолей <4 см, будет использован объем приблизительно 1-3 мл (предпочтительно 3 мл). Многократные инъекции, доставляемые в качестве единичной дозы, содержат объемы приблизительно от 0,1 до 0,5 мл. Вирусные частицы преимущественно могут контактировать посредством введения многократных инъекций в опухоль, разнесенных интервалом приблизительно в 1 см. Системное введение предпочтительно для таких условий, как гематологическая злокачественность.
Продолжительное введение также может применяться там, где оно подходит. Предпочтительной является доставка через шприц или катетер. Такая продолжительная перфузия может иметь место в течение периода времени от приблизительно 1-2 часов до приблизительно 2-6 часов, до приблизительно 6-12 часов, до приблизительно 12-24 часов, до приблизительно 1-2 дней, до приблизительно 1-2 недель или дольше после начала лечения. Как правило, доза терапевтической композиции через продолжительную перфузию будет эквивалентна дозе, которая дается посредством единичной или многократных инъекций, введенных за период времени, в течение которого осуществлялась перфузия.
Режим лечения может тоже изменяться, и он часто зависит от типа заболевания и расположения больной ткани, и таких факторов, как общее состояние организма и возраст пациента. Клиницисты наилучшим образом подходят к принятию такого решения, основанного на известной эффективности и токсичности (если она имеет место) терапевтических составов на основе лентивирусных векторов по настоящему изобретению.
Лечение может включать в себя различные «единичные дозы». Единичная доза определяется как доза, содержащая предварительно определенное количество терапевтической композиции, содержащей лентивирусный вектор по настоящему изобретению. Количество, которое необходимо ввести, и конкретный путь введения и состав известны специалистам в клинической области. Нет необходимости вводить единичную дозу как одну инъекцию, но она может включать в себя продолжительную перфузию за установленный период времени. Единичная доза по настоящему изобретению легко может быть описана на основе единиц трансдукции (T.U.) лентивекторов, определенных титрованием вектора в клеточной линии, такой как HeLa или 293. Единичные дозы находятся в пределах от 103, 104, 105, 106, 107, 108, 109, 1010, 1011, 1012, 1013 T.U. и выше.
8. ПРИМЕРЫ
Следующие примеры включены для демонстрации предпочтительных осуществлений изобретения. Специалистам в данной области следует понимать, что способы, описанные в последующих примерах, представляют собой предложенные авторами изобретения процедуры, которые подходящим образом функционируют при воплощении изобретения, и, таким образом, их следует рассматривать как составляющие предпочтительные пути его воплощения. Однако специалистам в данной области следует, в свете данного описания, понимать, что можно произвести множество изменений в определенных осуществлениях, которые описаны и при этом дают подобный или схожий результат без отклонения от сущности и объема изобретения.
A. Материалы и методология, используемые в примерах 1-3
1. Получение вектора
Получение происходящих из ВИЧ векторов, ложно классифицированных с оболочечным белком G вируса везикулярного стоматита (VSV), проводили посредством транзитной совместной трансфекции трех плазмид в эпителиальную клеточную линию 293T, как описано у Naldini et al 1996a. Используемой происходящей из ВИЧ упаковочной конструкцией являлась pCMVΔR8.91, которая кодирует предшественники Gag и Pol HIV-1, а также регуляторные белки Tat и Rev (Zufferey et al., 1997). G VSV экспрессировали из pMD.G.
Плазмиды вектора ВИЧ получали из исходной основной цепи pHR' (Naldini et al., 1996a) с последующими модификациями. Самоинактивирующиеся векторы получали из ранее описанного вектора SIN-18, который содержит делецию в области U3 3'LTR от нуклеотида -418 до нуклеотида -18, удаляющую всю транскрипционно активную последовательность (Zufferey et al., 1998). В кратком изложении, плазмиды pHR'SIN получали, как изложено ниже; фрагмент KpnI-XbaI, содержащий полипуриновый участок и 3'-LTR, удаляли из плазмиды pHR' и субклонировали в соответствующие участки pUC18. Данную плазмиду полностью расщепляли EcoRV и частично PvuII и лигировали саму на себя. Плазмиду, несущую делецию в 400 нуклеотидов из U3, восстанавливали. В участок EcoRI делеции плазмиды вставляли линкер XhoI, и фрагмент Xhol-XbaI клонировали обратно в плазмиду CMVlacZ pHR', расщепленную соответствующими ферментами. Все другие плазмиды SIN-18 получали посредством замены генов-репортеров (кодирующих люциферазу, GFP и Neo) для lacZ. Используемые плазмиды вектора pHR' отличались от первоначально описанных (Naldini et al., 1996) делецией XhoI-KpnI, удаляющей 118 нуклеотидов из последовательности, кодирующей Nef выше полипуринового участка, и делецией 1456 нуклеотидов человеческой последовательности ниже 3'LTR. Данная человеческая последовательность оставалась от первоначального клонирования провирусного генома HXB2. Две делеции не оказывали влияния на титры векторов или трансгенную экспрессию в делящихся клетках 293T.
Для получения вектора pHR-EF1-GFP-W-SIN была выполнена вставка EF1-альфа посредством вставки кассеты ClaI-BamHI, содержащей вставку EF1-GFP, в участок ClaI-BamHI pHR'-GFP-W-SIN (Zufferey et al. 1999). Производные этого вектора обладали некоторыми функциональными отличиями. Функциональными отличиями являются промотор EF1-alfa-short, участок loxP в 3'LTR и cPPT. Участок loxP в 3'LTR дуплицируется до 5'LTR в трансдуцированных клетках и, в присутствии дополнительно экспрессированной рекомбиназы Cre, позволяет осуществить удаление объединенного провируса. Тем не менее, участок loxP и опосредованное Cre возбуждение не являются необходимыми для активности протомора gp91-phox и не будут необходимы и не будут использоваться в любом потенциальном применении векторов в клинических испытаниях, основанных на кроветворных стволовых клетках. Отличием между EF1-альфа, представленным в pHR-EF1-GFP-(+/-)W-SIN, и промотором EF1-альфа-укороченным в pHPT-GFP и pWPT-GFP является то, что более короткая версия не содержит интрон, который делает вектор меньше.
Карты показаны на фиг.5А-5D, изображающих плазмидные конструкции pHPT-GFP, pWPT-GFP, pHPP91-GFP и pWPP91-GFP.
После транзитной трансфекции плазмид фосфатом кальция в клетки 293T, супернатант собирали, концентрировали ультрацентрифугированием, используя градиент сахарозы, ресуспендировали в бессывороточной среде Cellgro(SCGM (Cellgenix, Германия) и фильтровали через фильтр SpinX 0,45 мкм. Вирусный маточный раствор хранили при температуре -70°C и титры определяли посредством трансдукции и анализа путем проточной цитометрии экспрессии GFP в клетках HeLa, как описано ранее (Zufferey et al., 1997). Титры составляли между 5×107 и 108 единиц трансдукции (TU) в HeLa на мл.
2. Очистка и трансдукция клеток CD34+
Образцы пуповинной крови (CB) получали согласно установленному руководству, и клетки CD34+очищали, как описано (Arrighi et al., 1999). Вкратце, мононуклеарные клетки CB, полученные после центрифурирования в градиенте Ficoll-Paque (Pharmacia, Uppsala, Sweden), инкубировали на льду с Dynabeads M450 против CD34 (Dynal, Norway), как описано производителем. После нескольких отмывок для удаления несвязавшихся клеток, клетки CD34+снимали с шариков посредством инкубации в течение 15 минут при температуре 37°C с препаратом «Detach-a-bead», включенным в набор. Клетки сразу же отмывали и анализировали при помощи проточной цитометрии. Процентное соотношение очищенных клеток CD34+составляло 89±7,0. Для трансдукции 105 клеток выращивали в 96-луночных планшетах в 100 мкл среды Cellgro (SCGM, дополненной антибиотиками (Gibco BRL, Life Technologies LTD, Paisley, Scotland, U.K.), 10-4М дитиотреитолом (Fluka Biochemika, Buchs, Switzerland) и TPO (10 нг/мл), SCF (50 нг/мл), Flt3L (50 нг/мл). После инкубации в течение ночи 106 (обычно) или 105-5×106 (для анализа ответа на дозу) единиц трансдукции (TU) вектора в HeLa добавляли в ячейку и доводили объем до 200 мкл средой Cellgro (SCGM, содержащей TPO. Через 24 часа клетки отмывали, разводили в 400 мкл среды Cellgro (SCGM, дополненной антибиотиками, 10-4M дитиотреитолом и TPO (10 нг/мл), SCF (50 нг/мл), Flt3L (50 нг/мл) в течение 3 дней. Клетки либо непосредственно анализировали на экспрессию GFP и CD34, либо культивировали далее с 3 факторами роста.
3. Цитокины
Все цитокины представляли собой рекомбинантный человеческий материал и были получены от Peprotech (London, U.K.).
4. Антитела и иммунореагенты
Все антитела были получены от Becton Dickinson Pharmingen (USA). mIgG против CD34, обработанные препаратом Dynabead M450, были получены от Dynal A/S (Oslo, Norway).
5. Дифференцировка in vitro и стимуляция INF-гамма
Дифференцировку проводили in vitro в присутствии GM-CSF и SCF для дифференцировки моноцитов и G-CSF или G-CSF в течение 3 недель и SCF для дифференцировки гранулоцитов. Дифференцированные клетки стимулировали INF-((1000 ед/мл) в течение 6 дней, метили конъюгированными с PE моноклональными антителами и при помощи FACS анализировали экспрессию GFP в положительной для РЕ популяции. Числа указывают процентное соотношение клеток в квадрантах.
6. Анализ путем проточной цитометрии
Клетки анализировали, как описано (Arrighi et al, 1999), на приборе FACScalibur (Becton-Dickinson) с небольшими модификациями. FL-1 применяли для GFP, FL-2 для PE-меченных Mabs, FL-3 для Mabs, меченных PercP. Перед анализом суспензии клеток доводили до 0,5% параформальдегида. Данные анализировали при помощи программы WINMDI, написанной J. Trotter из Scripps Institute (La Jolla, CA), и программы CellQuest (Becton-Dickinson).
B. Примеры 1-4
Пример 1: Трансдукция кроветворных клеток-предшественников человека с основанным на ВИЧ лентивектором, содержащим экспрессирующую кассету под контролем промотора gp91-phox, приводящего к ограниченной экспрессии в моноцитах и гранулоцитах.
Лентивирусные векторы pWPP91-GFP (с WPRE) и pHPP91-GFP (без WPRE) создавали путем замены промотора EF-1 (фрагментом 1540 н.п. субъединицы gp91-phox промотора(1) NADPH-оксидазы фагоцита в лентивирусных векторах pWPT-GFP и pHPT-GFP, соответственно. Как pWPT-GFP, так и pHPT-GFP, являются SIN (самоинактивирующимися) лентивирусными векторами и для увеличения эффективности трансдукции клеток-мишеней содержат центральные последовательности полипуриновых участков (cPPT) (фиг.1 и 5). Рекомбинантные лентивекторы получали и концентрировали 100 (ранее описанными стандартными способами и использовали их для трансдукции кроветворных CD34+клеток-предшественников/стволовых клеток человека (HSC). См. Salmon P, Kindler V, Ducrey O, Chapuis B, Zubler RH, Trono D., «High-level transgene expression in human hematopoietic progenitors and differentiated blood lineages after transduction with improved lentiviral vectors», Blood 96(10): 3392-8 (2000), включенную сюда в качестве ссылки.
Высокое процентное соотношение трансдукции UCB HSC, вплоть до 80%, достигалось при помощи лентивектора, в котором центральный полипуриновый участок был помещен выше экспрессирующей кассеты EF1-альфа-GFP. Большое количество GFP-положительных клеток сохранялось после дифференцировки трансдуцированных HSC in vitro (цитокиновый коктейль) или in vivo (трансплантация мышам NOD/SCID).
Трансдукция HSC лентивектором, несущим кассету gp91-phox-GFP, которая сопровождается их дифференцировкой in vitro (в цитокиновом коктейле) или in vivo (трансплантация сублетальным облученным мышам NOD/SCID), приводила к экспрессии GFP, ограниченной исключительно зрелыми моноцитами и гранулоцитами. Экспрессию GFP под промотором gp91-phox в зрелых нейтрофилах увеличивали посредством вставки WPRE без потери специфичности.
Промотор gp91-phox, переданный в HP/SC посредством лентивектора, с WPRE или без него, проявлял свою физиологическую реакционноспособность по отношению к INF- (в зрелых нейтрофилах in vitro (фиг.2).
Пример 2: Элементы энхансера HS вызывают более высокую и менее разнородную экспрессию.
Важным результатом генетического лечения различных лимфогематологических нарушений является уровень экспрессии предположительно терапевтического трансгена в соответствующем ряде дифференцированных клеток. Для того чтобы получить более высокую и однородную экспрессию гена, в лентивирусный вектор клонировали последовательности специфичного для gp91 энхансера, расположенного на 30 н.п. выше гена gp91-phox внутри четырех участков гиперчувствительности (HS) к ДНКазе.
Сначала, для того, чтобы уменьшить размер вирусного генома, фрагмент промотора gp91 величиной в 1540 н.п. заменяли функционально равнозначным фрагментом величиной 0,5 тыс.н.п. (Skalnik, et al. 1991), образуя вектор pHP500-GFP (фиг.6A). Для облегчения клонирования вводили множественные уникальные участки рестрикции, позволяющие осуществлять введение энхансера на оба участка cPPT (которые должны сохранять свое центральное положение и функциональность после клонирования энхансера), образуя вектор pHPX-GFP (фиг.6B).
Фрагменты, перекрывающие 4 индивидуальных участка HS, получали посредством ПЦР с использованием геномной ДНК в качестве матрицы. Последовательность праймеров для ПЦР была основана на последовательности из генома человека, опубликованного в проекте «Геном человека» (контиг NT_011844; http://www.ncbi.nlm.nih.gov/genome/guide/human/). См. SEQ ID NOS:2-16. Расположение участков gp91-phoxHS было основано на опубликованных данных. См. Lien LL, Lee Y, and Orkin SH (1997). Последовательности приблизительно в 1-1,5 тыс.н.п., фланкирующие определенный участок, амплифицировали и клонировали в лентивектор, получая в конечном счете pHPHS-GFP. См. SEQ ID NOS: 2-16. Также был получен вариант с последовательностями WPRE (pWPHS-GFP, фиг.6C). При трансдукции HSC и дифференцировке включение элементов HS в эту и другие конфигурации приведет к более высокой полной экспрессии и/или менее разнородной экспрессии трансгена.
Пример 3: Разработка лечения хронического гранулематоза (CGD)
Хронический гранулематоз сильно коррелирует с отсутствием гена gp91-phox, который кодирует субъединицу NADPH (Dinauer et al., 1987). Ген gp91-phox, сцепленный с X-хромосомой, дефектен почти у 60% пациентов с CGD. Для обзора см. Malech HL, «Progress in gene therapy for chronic granulomatous disease», J. Infect. Dis. 179 (Suppl.2):S 318-25, 1999. Один подход к лечению будет включать в себя трансдукцию соответствующих клеток с тем, чтобы, по крайней мере, одна функциональная копия gp91-phox была введена пациенту. Как обсуждалось выше, данный тип лечения может включать в себя выделение клеток у пациента, трансдукцию ex vivo, затем перемещение этих клеток обратно пациенту.
Клетки CD34 выделяют у пациентов с CGD и трансдуцируют вектором по настоящему изобретению, который несет функциональную копию гена gp91-phox, например, ген gp91-phox, кодируемый последовательностью нуклеотидов, состоящей из нуклеотидов SEQ ID NO:18, или любой нуклеотидной последовательности, которая кодирует функциональные полипептиды аминокислотных последовательностей SEQ ID NO:19. Соответствующую экспрессию полипептида gp91-phox получают под контролем промотора gp91-phox, введенного в вектор. Повышенную экспрессию можно получить при помощи включения элемента WPRE и энхансеров HS, как описано выше.
Клетки трансплантируются в тестовые системы, такие как мыши SCID-NOD, или пациентам по показаниям. Дальнейшая оценка эффективности данного подхода будет получена при трансплантации трансдуцированных клеток в соответствующие линии нокаут-мышей.
Пример 4: Терапевтические подходы при недостаточности адгезии лейкоцитов (LAD)
Один подход к лечению будет включать в себя трансдукцию соответствующих клеток с тем, чтобы, по крайней мере, одна функциональная копия миеломоноцитарного лейкоцитарного интегрина была введена пациенту. Как обсуждалось выше, данный тип лечения может включать в себя выделение клеток у пациента, трансдукцию ex vivo, затем перемещение этих клеток обратно пациенту.
Клетки CD34 выделяют у пациентов с LAD и трансдуцируют векторами по настоящему изобретению, которые несут функциональную копию гена интегрина. Соответствующую экспрессию полипептида интегрина получают под контролем промотора CD11b, введенного в вектор. Промотор CD11b выбирают из промоторов, кодируемых полинуклеотидной последовательностью SEQ ID NO:20. См. Hickstein, et al. 1992. Повышенную экспрессию можно получить при помощи включения элемента WPRE и энхансеров HS, как описано выше.
Все описанные и заявленные здесь композиции и/или способы могут быть получены и выполнены без излишнего экспериментирования в свете настоящего описания. В то время как композиции и способ по данному изобретению описаны в плане предпочтительных осуществлений, специалисту в данной области ясно, что к композициям и/или способам и стадиям или последовательностям стадий описанного здесь способа могут применяться вариации без отклонения от концепции, сущности и объема изобретения. Более конкретно, понятно, что некоторые средства, родственные химически и физиологически, могут замещать описанные здесь средства с достижением тех же или подобных результатов. Предполагается, что все такие подобные замены и модификации, понятные специалистам в данной области, соответствуют сущности, объему и концепции изобретения, как определено прилагаемой формулой изобретения.
Ссылки
Следующие ссылки конкретно включены сюда в качестве ссылки в той степени, в которой они предоставляют методические или иные подробности, дополняющие приведенные здесь.
Патент США №4682195
Патент США №4683202
Патент США №5466468
Патент США №5645897
Патент США №5686279
Патент США №5705629
Патент США №5846225
Патент США №5846233
Патент США №5925565
Патент США №5928906
Патент США №5935819
Патент США №5994136
Патент США №6013516
Патент США №6136597
Патент США №6165782
Патент США №6207455
Патент США №6218181
Патент США №6218186
EP 266032
Akkina, Walton, Chen, Li, Planelles, Chen, «High-efficiency gene transfer into CD34+cells with a human immunodeficiency virus type 1-based retroviral vector pseudotyped with vesicular stomatitis virus envelope glycoprotein G», J. Virol; 70: 2581-2585, 1996.
Almendro et al, «Cloning of the human platelet endothelial cell adhesion molecule-1 promoter and its tissue-specific expression. Structural and functional characterization», J. Immunol., 157(12): 5411-5421, 1996.
An, Wersto, Agricola, Metzger, Lu, Amado, Chen, Donahue, «Marking and gene expression by a lentivirus vector in transplanted human and nonhuman primate CD34(+) cells», J. Virol., 74: 1286-1295, 2000.
Angel, Bauman, Stein, Dellus, Rahmsdorf, and Herrlich, «12-0-tetradecanoyl-phorbol-13-acetate Induction of the Human Collagenase Gene is Mediated by an Inducible Enhancer Element Located in the 5' Flanking Region», Mol. Cell. Biol., 7: 2256, 1987a.
Angel, Imagawa, Chiu, Stein, Imbra, Rahmsdorf, Jonat, Herrlich, and Karin, «Phorbol Ester-Inducible Genes Contain a Common cis Element Recognized by a TPA-Modulated Trans-acting Factor», Cell, 49: 729, 1987b
Arrighi, Hauser, Chapuis, Zubler, Kindler, «Long-term culture of human CD34(+) progenitors with FLT3-ligand, thrombopoietin, and stem cell factor induces extensive amplification of a CD34(-)CD14(-) and CD34(-)CD14(+) dendritic cell precursor», Blood, 93: 2244-2252, 1999.
Atchison and Perry, «Tandem Kappa Immunoglobulin Promoters are Equally Active in the Presence of the Kappa Enhancer: Implications for Model of Enhancer Function», Cell, 46: 253, 1986.
Atchison and Perry, «The Role of the Kappa Enhancer and its Binding Factor NF-kappa B in the Developmental Regulation of Kappa Gene Transcription», Cell, 48: 121, 1987.
Banerji et al., «Expression of a Beta-Globin Gene is Enhanced by Remote SV40 DNA Sequences», Cell, 27: 299, 1981.
Banerji, Olson, and Schaffner, «A lymphocyte-specific cellular enhancer is located downstream of the joining region in immunoglobulin heavy-chain genes», Cell, 35: 729, 1983.
Berkhout, Silverman, and Jeang, «Tat Trans-activates the Human Immunodeficiency Virus Through a Nascent RNA Target», Cell, 59: 273, 1989.
Bhatia, Bonnet, Kapp, Wang, Murdoch, Dick, «Quantitative analysis reveals expansion of human hematopoietic repopulating cells after short-term ex vivo culture», J. Exp.Med, 186: 619-624, 1997.
Blanar, Baldwin, Flavell, and Sharp, «A gamma-interferon-induced factor that binds the interferon response sequence of the MHC class I gene, H-2Kb», EMBO J., 8: 1139, 1989.
Blomer, Naldini, Kafri, Trono, Verma, Gage, «Highly efficient and sustained gene transfer in adult neurons with a lentivirus vector», J. Virol., 71: 6641-6649, 1997.
Bodine and Ley, «An enhancer element lies 3' to the human a gamma globin gene», EMBO J., 6: 2997,1987.
Boshart, Weber, Jahn, Dorsch-Hasler, Fleckenstein, and Schaffner, «A very strong enhancer is located upstream of an immediate early gene of human cytomegalovirus», Cell, 41: 521, 1985.
Bosze, Thiesen, and Charnay, «A transcriptional enhancer with specificity for erythroid cells is located in the long terminal repeat of the friend murine leukemia virus», EMBO J., 5: 1615,1986.
Braddock, Chambers, Wilson, Esnouf, Adams, Kingsman, and Kingsman, «HIV-I Tat activates presynthesized RNA in the nucleus», Cell, 58: 269, 1989.
Brown, Tiley, Cullen, «Efficient polyadenylation within the human immunodeficiency virus type 1 long terminal repeat requires flanking U3-specific sequences», J. Virol., 65: 3340-3343, 1991.
Bulla and Siddiqui, «The hepatitis B virus enhancer modulates transcription of the hepatitis B virus surface-antigen gene from an internal location», J. Virol., 62: 1437, 1986.
Campbell and Vllarreal, «Functional analysis of the individual enhancer core sequences of polyoma virus: cell-specific uncoupling of DNA replication from transcription», Mol. Cell. Biol., 8: 1993, 1988.
Campere and Tilghman, «Postnatal repression of the alpha-fetoprotein gene is enhancer independent», Genes and Dev., 3:37, 1989.
Campo, Spandidos, Lang, Wilkie, «Transcriptional control signals in the genome of bovine papilloma virus type I,» Nature, 303: 77, 1983.
Carbonelli et al. «A plasmid vector for isolation of strong promoters in E.coli» FEMS Microbiol Lett. 177(1): 75-82, 1999.
Case, Price, Jordan, Yu, Wang, Bauer, Haas, Xu, Stripecke, Naldini, Kohn, Crooks, «Stable transduction of quiescent CD34(+)CD38(-) human hematopoietic cells by HIV-1 based lentiviral vectors,» Proc. Natl. Acad. Sci. USA, 96: 2988-2993, 1999.
Celander and Haseltine, «Glucocorticoid Regulation of Murine Leukemia Virus Transcription Elements is Specified by Determinants Within the Viral Enhancer Region,» J. Virology, 61: 269, 1987.
Celander, Hsu, and Haseltine, «Regulatory Elements Within the Murine Leukemia Virus Enhancer Regions Mediate Glucocorticoid Responsiveness,». J. Virology, 62: 1314, 1988.
Chandler, Maler, and Yamamoto, «DNA Sequences Bound Specifically by Glucocorticoid Receptor in vitro Render a Heterlogous Promoter Hormone Responsive in vivo,» Cell, 33: 489, 1983.
Chandler et al., «RNA splicing specificity determined by the coordinated action of RNA recognition motifs in SR proteins,» Proc Natl Acad Sci USA. 94(8): 3596-3601, 1997.
Chang, Erwin, and Lee, "Glucose-regulated Protein (GRP94 and GRP78) Genes Share Common Regulatory Domains and are Coordinately Regulated by Common Trans-acting Factors," Mol. Cell. Biol., 9:2153, 1989.
Chatterjee, Lee, Rentoumis, and Jameson, «Negative Regulation of the Thyroid-Stimulating Hormone Alpha Gene by Thyroid Hormone: Receptor Interaction Adjacent to the TATA Box,» Proc Natl. Acad Sci. U.S.A., 86: 9114, 1989.
Chen and Okayama, «High-efficiency transformation of mammalian cells by plasmid DNA,» Mol. Cell. Biol. 7: 2745-2752, 1987.
Cherrington and Ganem, «Regulation of polyadenylation in human immunodeficiency virus (HIV): contributions of promoter proximity and upstream sequences,» Embo. J., 11: 1513-1524,1992.
Choi, Chen, Kriegler, and Roninson, «An altered pattern of cross-resistance in multi-drug-resistant human cells results from spontaneous mutations in the mdr-1 (p-glycoprotein) gene,» Cell, 53: 519, 1988.
Cocea, «Duplication of a region in the multiple cloning site of a plasmid vector to enhance cloning-mediated addition of restriction sites to a DNA fragment,» Biotechniques, 23: 814-816, 1997.
Cohen, Walter, and Levinson, «A Repetitive Sequence Element 3' of the Human c-Ha-ras1 Gene Has Enhancer Activity,» J. Cell. Physiol., 5: 75, 1987.
Corbeau, et al., PNAS (U.S.A.,) 93(24): 14070-14075, 1996.
Costa, Lai, Grayson, and Darnell, «The Cell-Specific Enhancer of the Mouse Transthyretin (Prealbumin) Gene Binds a Common Factor at One Site and a Liver-Specific Factor(s) at Two Other Sites,» Mol. Cell. Biol., 8: 81, 1988.
Cripe, Haugen, Turk, Tabatabai, Schmid, Durst, Gissmann, Roman, and Turek, «Transcriptional Regulation of the Human Papilloma Virus-16 E6-E7 Promoter by a Keratinocyte-Dependent Enhancer, and by Viral E2 Trans-Activator and Represser Gene Products: Implications for Cervical Carcinogenesis,» EMBO J., 6: 3745, 1987.
Culotta and Hamer, «Fine Mapping of a Mouse Metallothionein Gene Metal-Response Element,» Mol. Cell. Biol., 9: 1376, 1989.
Dandolo, Blangy, and Kamen, «Regulation of Polyoma Virus Transcription in Murine Embryonal Carcinoma Cells,» J. Virology, 47: 55, 1983.
Dao, Hannum, Kohn, Nolta, «FLT3 ligand preserves the ability of human CD34+progenitors to sustain long-term hematopoiesis in immune-deficient mice after ex vivo retroviral-mediated transduction,» Blood, 89: 446-456, 1997.
Dao, Hashino, Kato, Nolta, «Adhesion to fibronectin maintains regenerative capacity during ex vivo, culture and transduction of human hematopoietic stem and progenitor cells,» Blood, 92: 4612-4621, 1998.
Deschamps, Meijlink, and Verma, «Identification of a Transcriptional Enhancer Element Upstream From the Proto-Oncogene Fos,» Science, 230: 1174, 1985.
De Villiers, Schaffner, Tyndall, Lupton, and Kamen, «Polyoma Virus DNA Replication Requires an Enhancer,» Nature, 312: 242, 1984.
DeZazzo, Kilpatrick, Imperiale, «Involvement of long terminal repeat U3 sequences overlapping the transcription control region in human immunodeficiency virus type 1 mRNA 3' end formation,» Mol. Cell. Biol., 11: 1624-1630, 1991.
Dinauer MC, Orkin SH, Brown R, Jesaitis AJ, Parkos CA, «The glycoprotein encoded by the X-linked chronic granulomatous disease locus is a component of the neutrophil cytochrome b complex,» Nature 327(6124): 717-20 (1987).
Donello, Loeb, Hope, «Woodchuck hepatitis virus contains a tripartite posttranscriptional regulatory element,». J. Virol, 72: 5085-5092, 1998.
Dorrell, Gan, Pereira, Hawley, Dick, «Expansion of human cord blood CD34(+) CD38(-) cells in ex vivo culture during retroviral transduction without a corresponding increase in SCID repopulating cell (SRC) frequency: dissociation of SRC phenotype and function,» Blood, 95: 102-110, 2000.
Edbrooke, Burt, Cheshire, and Woo, «Identification of cis-acting sequences responsible for phorbol ester induction of human serum amyloid a gene expression via a nuclear-factor-kappa β-like transcription factor,» Mol. Cell. Biol., 9: 1908, 1989.
Edlund, Walker, Barr, and Rutter, «Cell-specific expression of the rat insulin gene: evidence for role of two distinct 5' flanking elements,» Science, 230: 912, 1985.
Fechheimer, Boylan, Parker, Sisken, Patel and Zimmer, «Transfection of mammalian cells with plasmid DNA by scrape loading and sonication loading,» Proc Nat'l. Acad. Sci. USA 84: 8463-8467, 1987.
Feng and Holland, «HIV-I Tat Trans-Activation Requires the Loop Sequence Within Tar,» Nature, 334: 6178, 1988.
Firak and Subramanian, «Minimal Transcription Enhancer of Simian Virus 40 is a 74-Base-Pair Sequence that Has Interacting Domains,» Mol. Cell. Biol., 6: 3667, 1986.
Foecking and Hofstetter, «Powerful and Versatile Enhancer-Promoter Unit for Mammalian Expression Vectors,» Gene, 45(1): 101-105, 1986.
Follenzi, A, Ailles, L.E., Bakovic, S., Geuna, M., Naldini, L. (2000) «Gene Transfer by Lentiviral Vecotrs is Limited by Nuclear Translocation and Rescued by HIV-1 pol Sequences,» Nat. Genet., 25: 217-22.
Fujita, Shibuya, Hotta, Yamanishi, and Taniguchi, «Interferon-Beta Gene Regulation: Tandemly Repeated Sequences of a Synthetic 6-bp Oligomer Function as a Virus-Inducible Enhancer,» Cell, 49: 357, 1987.
Gilles, Morris, Oi, and Tonegawa, «A tissue-specific transcription enhancer element is located in the major intron of a rearranged immunoglobulin heavy-chain gene,» Cell, 33: 717, 1983.
Gilmartin, Fleming, Oetjen, «Activation of HIV-1 pre-mRNA 3' processing in vitro requires both an upstream element and TAR,» Embo. J., 11: 4419-4428, 1992.
Gloss, Bernard, Seedorf, and Klock, «The Upstream Regulatory Region of the Human Papilloma Virus-16 Contains an E2 Protein-Independent Enhancer Which is Specific for Cervical Carcinoma Cells and Regulated by Glucocorticoid Hormones,» EMBO J., 6: 3735, 1987.
Godbout, Ingram, and Tilghman, «Fine-Structure Mapping of the Three Mouse Alpha-Fetoprotein Gene Enhancers,» Mol. Cell. Biol., 8: 1169, 1988.
Goodbourn, Burstein, and Maniatis, «The Human Beta-Interferon Gene Enhancer is Under Negative Control,» Cell, 45: 601, 1986.
Goodbourn and Maniatis, «Overlapping Positive and Negative Regulatory Domains of the Human β-Interferon Gene,» Proc. Natl. Acad. Sci. USA, 85: 1447, 1988.
Gopal, «Gene transfer method for transient gene expression, stable transformation, and cotransformation of suspension cell cultures,» Mol. Cell. Biol. 5: 1188-1190, 1985.
Gossen and Bujard, Proc. Natl. Acad. Sci., 89: 5547-5551, 1992.
Graham and Van Der Eb, «A new technique for the assay of infectivity of human adenovirus 5 DNA,» Virology 52: 456-467, 1973.
Greene, Bohnlein, and Ballard, «HIV-1, and Normal T-Cell Growth: Transcriptional Strategies and Surprises,» Immunology Today, 10: 272, 1989.
Grosschedl and Baltimore, «Cell-Type Specificity of Immunoglobulin Gene Expression is Regulated by at Least Three DNA Sequence Elements,» Cell, 41: 885, 1985.
Haslinger and Karin, «Upstream Promoter Element of the Human Metallothionein-II Gene Can Act Like an Enhancer Element» Proc Natl. Acad. Sci. U.S.A., 82: 8572, 1985.
Hauber and Cullen, «Mutational Analysis of the Trans-Activiation-Responsive Region of the Human Immunodeficiency Virus Type I Long Terminal Repeat,» J. Virology, 62: 673, 1988.
Hen, Borrelli, Fromental, Sassone-Corsi, and Chambon, «A Mutated Polyoma Virus Enhancer Which is Active in Undifferentiated Embryonal Carcinoma Cells is not Repressed by Adenovirus-2 E1A Products,» Nature, 321: 249, 1986.
Hensel, Meichle, Pfizenmaier, and Kronke, «PMA-Responsive 5' Flanking Sequences of the Human TNF Gene,» Lymphokine Res., 8: 347, 1989.
Herr and Clarke, «The SV40 Enhancer is Composed of Multiple Functional Elements That Can Compensate for One Another,» Cell, 45: 461, 1986.
Hickstein, D.D., Baker, D.M., Gollahon, K.A. and Back, A.L., «Identification of the promoter of the myelomonocytic leukocyte integrin CDllb,» Proc. Natl. Acad. Sci. U.S.A. 89 (6): 2105-2109, 1992.
Hirochika, Browker, and Chow, «Enhancers and Trans-Acting E2 Transcriptional Factors of Papilloma Viruses,» J. Virol., 61: 2599, 1987.
Hirsch, Gaugler, Deagostini-Bauzin, Bally-Cuif, and Gordis, «Identification of Positive and Negative Regulatory Elements Governing Cell-Type-Specific Expression of the Neural-Cell-Adhesion-Molecule Gene,» Mol. Cell. Biol., 10: 1959, 1990.
Holbrook, Gulino, and Ruscetti, «cis-Acting Transcriptional Regulatory Sequences in the Gibbon Ape Leukemia Virus (GALV) Long Terminal Repeat,» Virology, 157: 211, 1987.
Horlick and Benfield, «The upstream muscle-specific enhancer of the rat muscle creatine kinase gene is composed of multiple elements,» Mol. Cell. Biol., 9: 2396, 1989.
Huang, Ostrowski, Berard, and Hagar, «Glucocorticoid regulation of the ha-musv p21 gene conferred by sequences from mouse mammary tumor virus,» Cell, 27: 245, 1981.
Hug, Costas, Staeheli, Aebi, and Weissmann, «Organization of the Murine Mx Gene and Characterization of its Interferon- and Virus-Inducible Promoter,» Mol. Cell. Biol., 8: 3065, 1988.
Hwang, Lim, and Chae, «Characterization of the S-Phase-Specific Transcription Regulatory Elements in a DNA-Replication-Independent Testis-Specific H2B (TH2B) Histone Gene,» Mol. Cell. Biol., 10: 585, 1990.
Imagawa, Chiu, and Karin, «Transcription Factor AP-2 Mediates Induction by Two Different Signal-Transduction Pathways: Protein Kinase C and cAMP,» Cell, 51: 251, 1987.
Imbra and Karin, «Phorbol Ester Induces the Transcriptional Stimulatory Activity of the SV40 Enhancer,» Nature, 323: 555, 1986.
Imler, Lemaire, Wasvlyk, and Waslyk, «Negative Regulation Contributes to Tissue Specificity of the Immunoglobulin Heavy-Chain Enhancer,» Mol. Cell. Biol, 7: 2558, 1987.
Imperiale and Nevins, «Adenovirus 5 E2 Transcription Unit: an ElA-Inducible Promoter with an Essential Element that Functions Independently of Position or Orientation,» Mol. Cell. Biol., 4: 875, 1984.
Jakobovits, Smith, Jakobovits, and Capon, «A Discrete Element 3' of Human Immunodeficiency Virus 1 (HIV-1) and HIV-2 mRNA Initiation Sites Mediates Transcriptional Activation by an HIV Trans-Activator,» Mol. Cell. Biol., 8: 2555, 1988.
Jameel and Siddiqui, «The Human Hepatitis B Virus Enhancer Requires Transacting Cellular Factor(s) for Activity,» Mol. Cell. Biol., 6: 710, 1986.
Jaynes, Johnson, Buskin, Gartside, and Hauschka, «The Muscle Creatine Kinase Gene is Regulated by Multiple Upstream Elements, Including a Muscle-Specific Enhancer,» Mol. Cell. Biol., 8: 62, 1988.
Johnson, Wold, and Hauschka, «Muscle creatine kinase sequence elements regulating skeletal and cardiac muscle expression in transgenic mice,» Mol. Cell. Biol., 9: 3393, 1989.
Kadesch and Berg, «Effects of the Position of the Simian Virus 40 Enhancer on Expression of Multiple Transcription Units in a Single Plasmid,» Mol. Cell. Biol., 6: 2593, 1986.
Karin, Haslinger, Heguy, Dietlin, and Cooke, «Metal-Responsive Elements Act as Positive Modulators of Human Metallothionein-IIA Enhancer Activity,» Mol. Cell. Biol., 7: 606, 1987.
Katinka, Yaniv, Vasseur, and Blangy, «Expression of Polyoma Early Functions in Mouse Embryonal Carcinoma Cells Depends on Sequence Rearrangements in the Beginning of the Late Region,» Cell, 20: 393, 1980.
Kawamoto, Makino, Niw, Sugiyama, Kimura, Anemura, Nakata, and Kakunaga, «Identification of the Human Beta-Actin Enhancer and its Binding Factor,» Mol. Cell. Biol., 8: 267,1988.
Kiledjian, Su, Kadesch, «Identification and characterization of two functional domains within the murine heavy-chain enhancer», Mol. Cell. Biol., 8: 145, 1988.
Klamut, Gangopadyhay, Worton, and Ray, «Molecular and Functional Analysis of the Muscle-Specific Promoter Region of the Duchenne Muscular Dystrophy Gene,» Mol. Cell. Biol., 10: 193, 1990.
Klein et al., «High-velocity microprojectiles for delivering nucleic acids into living cells,» Nature, 327: 70-73, 1987.
Koch, Benoist, and Mathis, «Anatomy of a new β-cell-specific enhancer,» Mol. Cell. Biol., 9: 303, 1989.
Kraus et al., «Alternative promoter usage and tissue specific expression of the mouse somatostatin receptor 2 gene,» FEBS Lett., 428(3): 165-170, 1998.
Kohn, Nolta, Weinthal, Bahner, Yu, Lilley, Crooks, «Toward gene therapy for Gaucher disease,» Hum. Gene Ther., 2: 101-105, 1991.
Kriegler and Botchan, «A retrovirus LTR contains a new type of eukaryotic regulatory element,» In: Eukaryotic Viral Vectors, Gluzman (ed.). Cold Spring Harbor, Cold Spring Harbor Laboratory, NY, 1982.
Kriegler et al., «Promoter substitution and enhancer augmentation increases the penetrance of the sv40 a gene to levels comparable to that of the harvey murine sarcoma virus ras gene in morphologic transformation,» In: Gene Expression, Alan Liss (Ed.), Hamer and Rosenberg, New York, 1983.
Kriegler et al., «Viral Integration and Early Gene Expression Both Affect the Efficiency of SV40 Transformation of Murine Cells: Biochemical and Biological Characterization of an SV40 Retrovirus,» In: Cancer Cells 2/Oncogenes and Viral Genes, Van de Woude et al. (eds). Cold Spring Harbor, Cold Spring Harbor Laboratory, 1984.
Kriegler, Perez, Defay, Albert and Liu, «A Novel Form of TNF/Cachectin Is a Cell-Surface Cytotoxix Transmembrane Protein: Ramifications for the Complex Physiology of TNF,» Cell, 53: 45, 1988.
Kuhl, De La Fuenta, Chaturvedi, Parinool, Ryals, Meyer, and Weissman, «Reversible Silencing of Enhancers by Sequences Derived From the Human IFN-alpha Promoter,» Cell, 50: 1057, 1987.
Kunz, Zimmerman, Heisig, and Heinrich, «Identification of the Promoter Sequences Involved in the Interleukin-6-Dependent Expression of the Rat Alpha-2-Macroglobulin Gene,» Nucl. Acids Res., 17: 1121, 1989.
Lareyre et al., «A 5-kilobase pair promoter fragment of the murine epididymal retinoic acid-binding protein gene drives the tissue-specific, cell-specific, and androgen-regulated expression of a foreign gene in the epididymis of transgenic mice,» J Biol Chem., 274(12): 8282-8290, 1999.
Larsen, Harney, and Moore, «Repression medaites cell-type-specific expression of the rat growth hormone gene," Proc Natl. Acad. Sci. USA., 83:8283, 1986.
Laspia, Rice, and Mathews, "HIV-1 Tat protein increases transcriptional initiation and stabilizes elongation,» Cell, 59: 283, 1989.
Latimer, Berger, and Baumann, «Highly conserved upstream regions of the alpha..sub.1-antitrypsin gene in two mouse species govern liver-specific expression by different mechanisms,» Mol. Cell. Biol, 10: 760, 1990.
Lee, Mulligan, Berg, and Ringold, «Glucocorticoids Regulate Expression of Dihydrofolate Reductase cDNA in Mouse Mammary Tumor Virus Chimaeric Plasmids,» Nature, 294: 228,1981.
Lee et al., «Activation of beta3-adrenoceptors by exogenous dopamine to lower glucose uptake into rat adipocytes,» J Auton Nerv Syst. 74(2-3): 86-90, 1997.
Levenson et al., «Internal ribosomal entry site-containing retroviral vectors with green fluorescent protein and drug resistance markers,» Human Gene Therapy, 9: 1233-1236, 1998.
Levinson, Khoury, VanDeWoude, and Gruss, «Activation of SV40 Genome by 72-Base-Pair Tandem Repeats of Moloney Sarcoma Virus,» Nature, 295: 79, 1982. Lewis and Emerman, «Passage through mitosis is required for oncoretroviruses but not for the human immunodeficiency virus,» J. Virol., 68: 510-516, 1994.
Lien LL, Lee Y, Orkin SH, (1997) «Regulation of the myeloid-cell-expressed human gp91-phox gene as studied by transfer of yeast artificial chromosome clones into embryonic stem cells: suppression of a variegated cellular pattern of expression requires a full complement of distant cis elements,» Mol. Cell Biol. 17(4): 2279-90.
Lin, Cross, Halden, Dragos, Toledano, and Leonard, «Delineation of an enhancerlike positive regulatory element in the interleukin-2 receptor alpha-chain gene,» Mol. Cell. Biol., 10: 850,1990.
Luo W, and Skalnik DG, «CCAAT Displacement Protein Competes with Multiple Transcriptional Activators for Binding to Four Sites in the Proximal gp91-phox Promoter», J. Biol. Chem. 271: 18203-210, 1996.
Luo W, and Skalnik DG, «Interferon Regulatory Factor-2 Directs Transcription from the gp91-phox Promoter,» J. Biol. Chem. 271: 23445-23451, 1996.
Luria, Gross, Horowitz, and Givol, «Promoter Enhancer Elements in the Rearranged Alpha-Chain Gene of the Human T-Cell Receptor,» EMBO J., 6: 3307, 1987.
Lusky, Berg, Weiher, and Botchan, «Bovine Papilloma Virus Contains an Activator of Gene Expression at the Distal End of the Early Transcription Unit,» Mol. Cell. Biol. 3: 1108, 1983.
Lusky and Botchan, «Transient Replication of Bovine Papilloma Virus Type 1 Plasmids: cis and trans Requirements,» Proc Natl. Acad. Sci. U.S.A., 83: 3609, 1986.
Majors and Varmus, «A Small Region of the Mouse Mammary Tumor Virus Long Terminal Repeat Confers Glucocorticoid Hormone Regulation on a Linked Heterologous Gene,» Proc. Natl. Acad. Sci. U.S.A., 80: 5866, 1983.
Malech HL, «Progress in gene therapy for chronic granulomatous disease,» J. Infect. Dis. 179 (Suppl.2): S 318-25, 1999.
Marthas et al. J. Virol.. 67: 6047-6055, 1993.
Mazurier, Moreau-Gaudry, Maguer-Satta, Salesse, Pigeonnier-Lagarde, Ged, Belloc, Lacombe, Mahon, Reiffers, de Verneuil, «Rapid analysis and efficient selection of human transduced primitive hematopoietic cells using the humanized S65T green fluorescent protein,» Gene Ther., 5: 556-562, 1998.
McNeall, Sanchez, Gray, Chesterman, and Sleigh, «Hyperinducible Gene Expression From a Metallotionein Promoter Containing Additional Metal-Responsive Elements,» Gene, 76: 81, 1989.
Miksicek, Heber, Schmid, Danesch, Posseckert, Beato, and Schutz, «Glucocorticoid Responsiveness of the Transcriptional Enhancer of Moloney Murine Sarcoma Virus,» Cell, 46: 203, 1986.
Miyoshi, Smith, Mosier, Verma, Torbett, «Transduction of human CD34+cells that mediate long-term engraftment of NOD/SCID mice by HIV vectors,» Science, 283: 682-686, 1999.
Mizushima and Nagata, «pEF-BOS, a powerful mammalian expression vector,» Nucleic Acids Res., 18: 5322, 1990.
Mordacq and Linzer, «Co-localization of Elements Required for Phorbol Ester Stimulation and Glucocorticoid Repression of Proliferin Gene Expression,» Genes and Dev., 3: 760, 1989.
Moreau, Hen, Wasylyk, Everett, Gaub, and Chambon, «The SV40 base-repair repeat has a striking effect on gene expression both in sv40 and other chimeric recombinants,» Nucl. Acids Res., 9: 6047, 1981.
Muesing et al., Cell, 48: 691, 1987.
Naldini, Blomer, gallay, Ory, Mulligan, Gage, Verma, Trono, «In vivo gene delivery and stable transduction of nondividing cells by a lentiviral vector,» Science, 272: 263-267, 1996a.
Naldini, Blomer, Gage, Trono, Verma, «Efficient transfer, integration, and sustained long-term expression of the transgene in adult rat brains injected with a lentiviral vector,» Proc. Natl. Acad. Sci. USA, 93: 11382-11388, 1996b.
Ng, Gunning, Liu, Leavitt, and Kedes, «Regulation of the Human Beta-Actin Promoter by Upstream and Intron Domains,» Nuc. Acids Res., 17: 601, 1989.
Nomoto et al, «Cloning and characterization of the alternative promoter regions of the human LIMK2 gene responsible for alternative transcripts with tissue-specific expression,» Gene, 236(2): 259-271, 1999.
Omitz, Hammer, Davison, Brinster, and Palmiter, «Promoter and enhancer elements from the rat elastase i gene function independently of each other and of heterologous enhancers,» Mol. Cell. Biol. 7: 3466. 1987.
Ondek, Sheppard, and Herr, «Discrete Elements Within the SV40 Enhancer Region Display Different Cell-Specific Enhancer Activities,» EMBO J., 6: 1017, 1987.
Ory et al., Proc. Natl. Acad. Sci., 93: 11400-11406, 1996.
Palmiter, Chen, and Brinster, «Differential regulation of metallothionein-thymidine kinase fusion genes in transgenic mice and their offspring,» Cell, 29: 701, 1982.
Pech, Rao, Robbins, and Aaronson, «Functional identification of regulatory elements within the promoter region of platelet-derived growth factor 2,» Mol. Cell. Biol., 9: 396,1989.
Perez-Stable and Constantini, «Roles of fetal γ-globin promoter elements and the adult β-globin 3' enhancer in the stage-specific expression of globin genes,» Mol. Cell. Biol., 10: 1116, 1990.
Piacibello, Sanavio, Severino, Dane, Gammaitoni, Fagioli, Perissinotto, Cavalloni, Kollet Lapidot, Aglietta, «Engraftment in nonobese diabetic severe combined immunodeficient mice of human CD34(+) cord blood cells after ex vivo expansion: evidence for the amplification and self-renewal of repopulating stem cells,» Blood, 93: 3736-3749, 1999.
Picard and Schaffner, «A Lymphocyte-Specific Enhancer in the Mouse Immunoglobulin Kappa Gene,» Nature, 307: 83, 1984.
Pinkert, Omitz, Brinster, and Palmiter, «An albumin enhancer located 10 kb upstream functions along with its promoter to direct efficient, liver-specific expression in transgenic mice,» Genes and Dev., 1: 268, 1987.
Ponta, Kennedy, Skroch, Hynes, and Groner, «Hormonal Response Region in the Mouse Mammary Tumor Virus Long Terminal Repeat Can Be Dissociated From the Proviral Promoter and Has Enhancer Properties,» Proc. Natl. Acad. Sci. U.S.A., 82: 1020, 1985.
Porton, Zaller, Lieberson, and Eckhardt, «Immunoglobulin heavy-chain enhancer is required to maintain transfected.gamma.2a gene expression in a pre-b-cell line,» Mol. Cell. Biol., 10: 1076, 1990.
Potter et al., «Enhancer-dependent expression of human (immunoglobulin genes introduced into mouse pre-B lymphocytes by electroporation,» Proc Nat'l Acad. Sci. USA, 81: 7161-7165, 1984.
Queen and Baltimore, «Immunoglobulin Gene Transcription is Activated by Downstream Sequence Elements,» Cell, 35: 741, 1983.
Quinn, Farina, Gardner, Krutzsch, and Levens, «Multiple components are required for sequence recognition of the ap1 site in the gibbon ape leukemia virus enhancer,» Mol. Cell. Biol., 9: 4713, 1989.
Ramezani A, Hawley TS, Hawley RG, «Lentiviral Vectors for Enhanced Gene Expression in Human Hematopoietic Cells,» Mol. Ther. 2(5): 458-69, 2000.
Redondo, Hata, Brocklehurst, and Krangel, «A T-Cell-Specific Transcriptional Enhancer Within the Human T-Cell Receptor.delta. Locus,» Science, 247: 1225, 1990.
Resendez Jr., Wooden, and Lee, «Identification of highly conserved regulatory domains and protein-binding sites in the promoters of the rat and human genes encoding the stress-inducible 78-kilodalton glucose-regulated protein,» Mol. Cell. Biol., 8: 4579, 1988.
Reisman and Rotter, «Induced Expression From the Moloney Murine Leukemia Virus Long Terminal Repeat During Differentiation of Human Myeloid Cells is Mediated Through its Transcriptional Enhancer.» Mol. Cell. Biol., 9: 3571, 1989.
Remington's Pharmaceutical Sciences, 15th Ed., pages 1035-1038 and 1570-1580.
Ripe, Lorenzen, Brenner, and Breindl, «Regulatory elements in the 5' flanking region and the first intron contribute to transcriptional control of the mouse alpha-1-type collagen gene,» Mol. Cell. Biol., 9: 2224, 1989.
Rippe, Brenner and Leffert, «DNA-mediated gene transfer into adult rat hepatocytes in primary culture,» Mol. Cell Biol., 10: 689-695, 1990.
Rittling, Coutinho, Amarm, and Kolbe, «AP-1/jun-binding Sites Mediate Serum Inducibility of the Human Vimentin Promoter,» Nuc. Acids Res., 17: 1619, 1989.
Roe, Reynolds, Yu, Brown, «Integration of murine leukemia virus DNA depends on mitosis,» Embo. J., 12: 2099-2108, 1993.
Rosen, Sodroski, and Haseltine, «The location of cis-acting regulatory sequences in the human t-cell lymphotropic virus type III (HTLV-III/LAV) long terminal repeat,» Cell, 41: 813, 1988.
Sakai, Helms, Carlstedt-Duke, Gustafsson, Rottman, and Yamamoto, «Hormone-Mediated Repression: A Negative Glucocorticoid-Response Element From the Bovine Prolactin Gene,» Genes and Dev., 2: 1144, 1988.
Salmon P, Kindler V, Ducrey O, Chapuis B, Zubler RH, Trono D., «High-level transgene expression in human hematopoietic progenitors and differentiated blood lineages after transduction with improved lentiviral vectors,» Blood 96(10): 3392-8, 2000.
Sambrook, Fritsch, Maniatis, In: Molecular Cloning: A Laboratory Manual 2 rev. ed., Cold Spring Harbor, Cold Spring Harbor Laboratory Press, 1(77): 19-17.29, 1989.
Satake, Furukawa, and Ito, «Biological activities of oligonucleotides spanning the f9 point mutation within the enhancer region of polyoma virus DNA,» J. Virology, 62: 970, 1988.
Scharfmann, Axelrod, Verma, «Long-term in vivo expression of retrovirus-mediated gene transfer in mouse fibroblast implants,» Proc. Natl. Acad. Sci. USA, 88: 4626-4630, 1991.
Schaffner, Schirm, Muller-Baden, Wever, and Schaffner, «Redundancy of Information in Enhancers as a Principle of Mammalian Transcription Control,» J. Mol. Biol., 201: 81, 1988.
Schmid, Uittenbogaart, Keld, Giorgi, «A rapid method for measuring apoptosis and dual-color immunofluorescence by single laser flow cytometry,» J. Immunol. Methods, 170: 145-157,1994.
Searle, Stuart, and Palmiter, «Building a metal-responsive promoter with synthetic regulatory elements,» Mol. Cell. Biol., 5:1480, 1985.
Sharp and Marciniak, «HIV Tar: an RNA Enhancer?» Cell, 59: 229, 1989.
Shaul and Ben-Levy, «Multiple Nuclear Proteins in Liver Cells are Bound to Hepatitis B Virus Enhancer Element and its Upstream Sequences,» EMBO J., 6: 1913, 1987.
Sherman, Basta, Moore, Brown, and Ting, «Class II Box Consensus Sequences in the HLA-DR.alpha. Gene: Transcriptional Function and Interaction with Nuclear Proteins,» Mol. Cell. Biol., 9: 50, 1989.
Skalnik DG, Dorfman DM, Perkins AS, Jenkins NA, Copeland NG, Orkin SH, (1991) «Targeting of transgene expression to monocyte / macrophages by the gp91-phox promoter and consequent histiocytic malignancies». Proc. Natl. Acad. Sci. USA 88: 8505-09.
Sleigh and Lockett, «SV40 Enhancer Activation During Retinoic-Acid-Induced Differentiation of F9 Embryonal Carcinoma Cells», J. EMBO, 4: 3831, 1985.
Spalholz, Yang, and Howley, «Transactivation of a Bovine Papilloma Virus Transcriptional Regulatory Element by the E2 Gene Product,» Cell, 42: 183, 1985.
Spandau and Lee, «Trans-Activation of Viral Enhancers by the Hepatitis B Virus X Protein», J. Virology, 62: 427, 1988.
Spandidos and Wilkie, «Host-Specificities of Papilloma Virus, Moloney Murine Sarcoma Virus and Simian Virus 40 Enhancer Sequences,» EMBO J., 2: 1193, 1983.
Stephens and Hentschel, «The Bovine Papilloma Virus Genome and its Uses as a Eukaryotic Vector,» Biochem. J., 248: 1, 1987.
Stuart, Searle, and Palmiter, «Identification of Multiple Metal Regulatory Elements in Mouse Metallothionein-I Promoter by Assaying Synthetic Sequences,» Nature, 317: 828, 1985.
Sullivan and Peterlin, «Transcriptional Enhancers in the HLA-DQ Subregion,» Mol. Cell. Biol., 7: 3315, 1987.
Sutton, Reitsma, Uchida, Brown, «Transduction of human progenitor hematopoietic stem cells by human immunodeficiency virus type 1-based vectors is cell cycle dependent,» J. Virol., 73: 3649-3660, 1999.
Sutton, Wu, Rigg, Bohnlein, Brown, «Human immunodeficiency virus type 1 vectors efficiently transduce human hematopoietic stem cells,» J. Virol., 72: 5781-5788, 1998.
Swartzendruber and Lehman, «Neoplastic Differentiation: Interaction of Simian Virus 40 and Polyoma Virus with Murine Teratocarcinoma Cells,» J. Cell. Physiology, 85: 179, 1975.
Takebe, Seiki, Fujisawa, Hoy, Yokota, Arai, Yoshida, and Arai, «SR. alpha. Promoter: An Efficient and Versatile Mammalian cDNA Expression System Composed of the Simian Virus 40 Early Promoter and the R-U5 Segment of Human T-Cell Leukemia Virus Type 1 Long Terminal Repeat,» Mol. Cell. Biol., 8: 466, 1988.
Taylor and Kingston, «E1A Trans-Activation of Human HSP70 Gene Promoter Substitution Mutants is Independent of the Composition of Upstream and TATA Elements,» Mol. Cell. Biol., 10: 176, 1990.
Taylor and Kingston, «Factor Substitution in a Human HSP70 Gene Promoter: TATA-Dependent and TATA-Independent Interactions,» Mol. Cell. Biol., 10: 165, 1990.
Taylor, Solomon, Weiner, Paucha, Bradley, and Kingston, «Stimulation of the Human Heat-Shock Protein 70 Promoter in vitro by Simian Virus 40 Large T Antigen,» J. Biol. Chem., 264: 15160, 1989.
Tavernier, Gheysen, Duerinck, Can Der Heyden, and Fiers, «Deletion Mapping of the Inducible Promoter of Human IFN-beta Gene,» Nature, 301: 634, 1983.
Thiesen, Bosze, Henry, and Charnay, «A DNA Element Responsible for the Different Tissue Specificities of Friend and Moloney Retroviral Enhancers,» J. Virology, 62: 614, 1988.
Tronche, Rollier, Bach, Weiss, and Yaniv, «The Rat Albumin Promoter: Cooperation with Upstream Elements is Required When Binding of APF/HNF 1 to the Proximal Element is Partially Impaired by Mutation or Bacterial Methylation,» Mol. Cell. Biol., 9: 4759, 1989.
Tronche, Rollier, Herbomel, Bach, Cereghini, Weiss, and Yaniv, «Anatomy of the Rat Albumin Promoter,» Mol. Biol. Med., 7: 173, 1990.
Trudel and Constantini, «A 3' Enhancer Contributes to the Stage-Specific Expression of the human Beta-Globin Gene,» Genes and Dev., 6: 954, 1987.
Tsumaki et al, «Modular arrangement of cartilage- and neural tissue-specific cis-elements in the mouse alpha2(XI) collagen promoter,» J Biol Chem. 273(36): 22861-22864, 1998.
Tur-Kaspa, Teicher, Levine, Skoultchi and Shafritz, «Use of electroporation to introduce biologically active foreign genes into primary rat hepatocytes,» Mol. Cell Biol., 6: 716-718, 1986.
Tyndall, La Mantia, Thacker, Favaloro, and Kamen, «A Region of the Polyoma Virus Genome Between the Replication Origin and Late Protein-Coding Sequences is Required in cis for Both Early Gene Expression and Viral DNA Replication,» Nuc. Acids. Res., 9: 6231, 1981.
Uchida, Sutton, Friera, He, Reitsma, Chang, Veres, Scollay, Weissman, «HIV, but not murine leukemia virus, vectors mediate high efficiency gene transfer into freshly isolated G0/G1 human hematopoietic stem cells,» Proc. Natl. Acad Sci. USA, 95: 11939-11944, 1998.
Ueda, Tsuji, Yoshino, Ebihara, Yagasaki, Hisakawa, Mitsui, Manabe, Tanaka, Kobayashi, Ito, Yasukawa, Nakahata, «Expansion of human NOD/SCID-repopulating cells by stem cell factor, Flk2/Flt3 ligand, thrombopoietin, IL-6, and soluble IL-6 receptor,» J. Clin. Invest., 105: 1013-1021, 2000.
Unutmaz, Kewal, Ramani, Marmon, Littman, «Cytokine signals are sufficient for HIV-1 infection of resting human T lymphocytes,» J. Exp.Med., 189: 1735-1746, 1999.
Valsamakis, Schek, Alwine, «Elements upstream of the AAUAAA within the human immunodeficiency virus polyadenylation signal are required for efficient polyadenylation in vitro,» Mol. Cell Biol., 12: 3699-3705, 1992.
Valsamakis, Zeichner, Carswell, Alwine, «The human immunodeficiency virus type 1 polyadenylylation signal: a 3' long terminal repeat element upstream of the AAUAAA necessary for efficient polyadenylylation,» Proc. Natl. Acad Sci. USA, 88: 2108-2112, 1991.
Vannice and Levinson, «Properties of the Human Hepatitis B Virus Enhancer: Position Effects and Cell-Type Nonspecificity,» J. Virology, 62: 1305, 1988.
Vasseur, Kress, Montreau, and Blangy, «Isolation and Characterization of Polyoma Virus Mutants Able to Develop in Multipotential Murine Embryonal Carcinoma Cells,» Proc Natl. Acad. Sci. U.S.A., 77: 1068, 1980.
Wang and Calame, «SV40 enhancer-binding factors are required at the establishment but not the maintenance step of enhancer-dependent transcriptional activation,» Cell, 47: 241, 1986.
Watanabe et al., «Gene transfection of mouse primordial germ cells in vitro and analysis of their survival and growth control, Experimental Cell Research», 230: 76-83, 1997.
Weber, De Villiers, and Schaffner, «An SV40 'Enhancer Trap' Incorporates Exogenous Enhancers or Generates Enhancers From its Own Sequences,» Cell, 36: 983, 1984.
Weinberger, Jat, and Sharp, «Localization of a Repressive Sequence Contributing to B-cell Specificity in the Immunoglobulin Heavy-Chain Enhancer,» Mol. Cell. Biol., 8: 988, 1984.
Winoto and Baltimore, «αβ-lineage-specific Expression of the (T-Cell Receptor Gene by Nearby Silencers,» Cell, 59: 649, 1989.
Wu et al., «Promoter-dependent tissue-specific expressive nature of imprinting gene, insulin-like growth factor II, in human tissues,» Biochem Biophys Res Commun. 233(1): 221-226, 1997.
Yang, Burkholder, Roberts, Martinell and McCabe, «In vivo and in vitro gene transfer to mammalian somatic cells by particle bombardment,» Proc Nat'l Acad Sci. USA, 87: 9568-9572, 1990.
Yutzey, Kline, and Konieczny, «An Internal Regulatory Element Controls Troponin I Gene Expression,» Mol. Cell. Biol., 9: 1397, 1989.
Zhao-Emonet et al., «The equine herpes virus 4 thymidine kinase is a better suicide gene than the human herpes virus 1 thymidine kinase,» Gene Ther. 6(9): 1638-1642, 1999.
Zufferey, Nagy, Mandel, Naldini, Trono, «Multiply attenuated lentiviral vector achieves efficient gene delivery in vivo,» Nat. Biotechnol., 15: 871-875, 1997.
Zufferey, Dull, Mandel, Bukovsky, Quiroz, Naldini, Trono, «Self-inactivating lentivirus vector for safe and efficient in vivo gene delivery,» J. Virol., 72: 9873-9880, 1998.
Zufferey, Donello, Trono, Hope, «Woodchuck hepatitis virus posttranscriptional regulatory element enhances expression of transgenes delivered by retroviral vectors,» J. Virol., 73: 2886-2892, 1999.








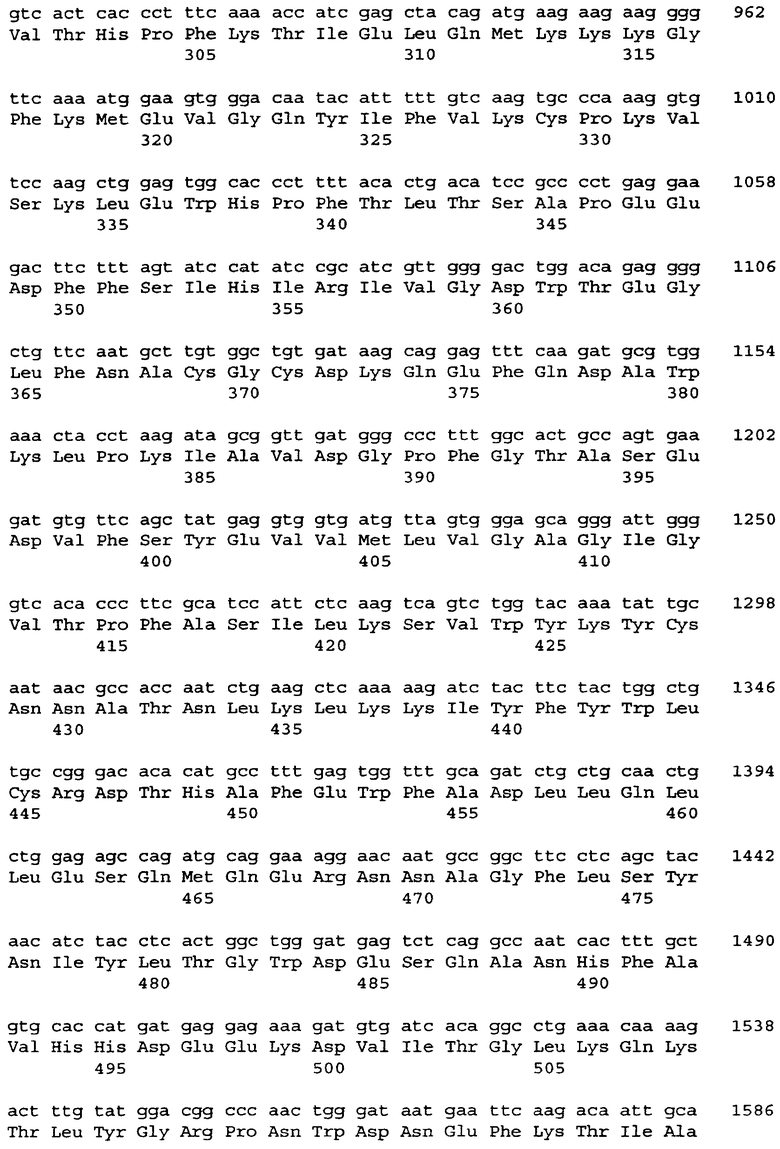




| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛУСТАБИЛЬНОЕ ПОЛУЧЕНИЕ ЛЕНТИВИРУСНЫХ ВЕКТОРОВ | 2011 |
|
RU2577982C2 |
| БИОПРОИЗВОДСТВО ЛЕНТИВИРУСНЫХ ВЕКТОРОВ | 2016 |
|
RU2729385C2 |
| СТАБИЛЬНОЕ ПОЛУЧЕНИЕ ЛЕНТИВИРУСНЫХ ВЕКТОРОВ | 2011 |
|
RU2577979C2 |
| СПОСОБЫ СТАБИЛЬНОЙ ТРАНСДУКЦИИ КЛЕТОК ВИРУСНЫМИ ВЕКТОРАМИ | 2001 |
|
RU2280074C2 |
| ЛЕНТИВИРУСНЫЕ ВЕКТОРЫ, ПСЕВДОТИПИРОВАННЫЕ МУТАНТНЫМИ BaEV ГЛИКОПРОТЕИНАМИ | 2012 |
|
RU2618864C2 |
| ВСПОМОГАТЕЛЬНЫЙ ПЛАЗМИДНЫЙ ЛЕНТИВИРУСНЫЙ ЭКСПРЕССИОННЫЙ ВЕКТОР ДЛЯ ПОЛУЧЕНИЯ ВЫСОКИХ ТИТРОВ VPX-СОДЕРЖАЩИХ ЛЕНТИВИРУСНЫХ ЧАСТИЦ, ОБЕСПЕЧИВАЮЩИЙ ЭФФЕКТИВНОЕ ЗАРАЖЕНИЕ МОНОЦИТОВ И ДЕНДРИТНЫХ КЛЕТОК ЧЕЛОВЕКА | 2017 |
|
RU2697781C2 |
| ХОРОШО ПЕРЕНОСИМАЯ И ВЫСОКОСПЕЦИФИЧЕСКАЯ АДАПТИРОВАННАЯ РЕКОМБИНАЗА ДЛЯ РЕКОМБИНИРОВАНИЯ АСИММЕТРИЧНЫХ УЧАСТКОВ-МИШЕНЕЙ ВО МНОЖЕСТВЕ ШТАММОВ РЕТРОВИРУСОВ | 2015 |
|
RU2706194C2 |
| ВЕКТОРЫ AAV | 2018 |
|
RU2793112C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ЛЕЧЕНИЯ ГЕМОГЛОБИНОПАТИЙ | 2015 |
|
RU2707540C2 |
| ЛЕНТИВИРУСНЫЕ ВЕКТОРЫ ДЛЯ ДОСТАВКИ PKLR ДЛЯ ЛЕЧЕНИЯ ДЕФИЦИТА ПИРУВАТКИНАЗЫ | 2018 |
|
RU2777934C2 |
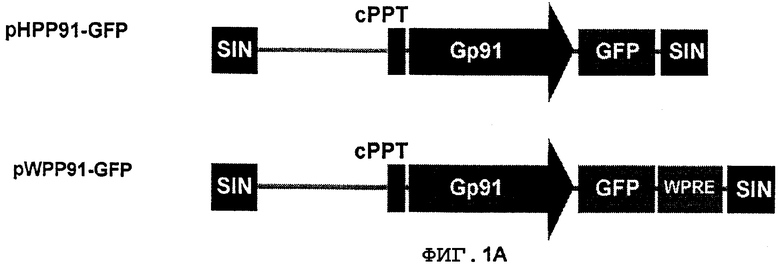
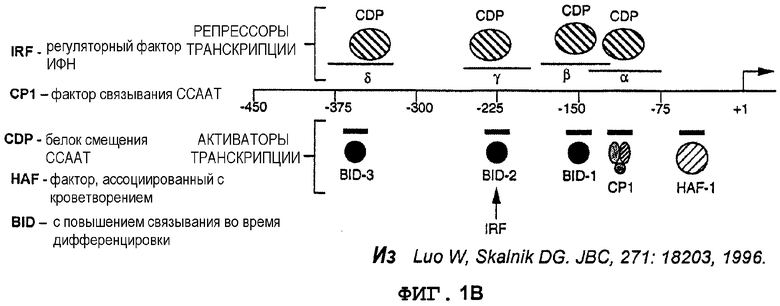
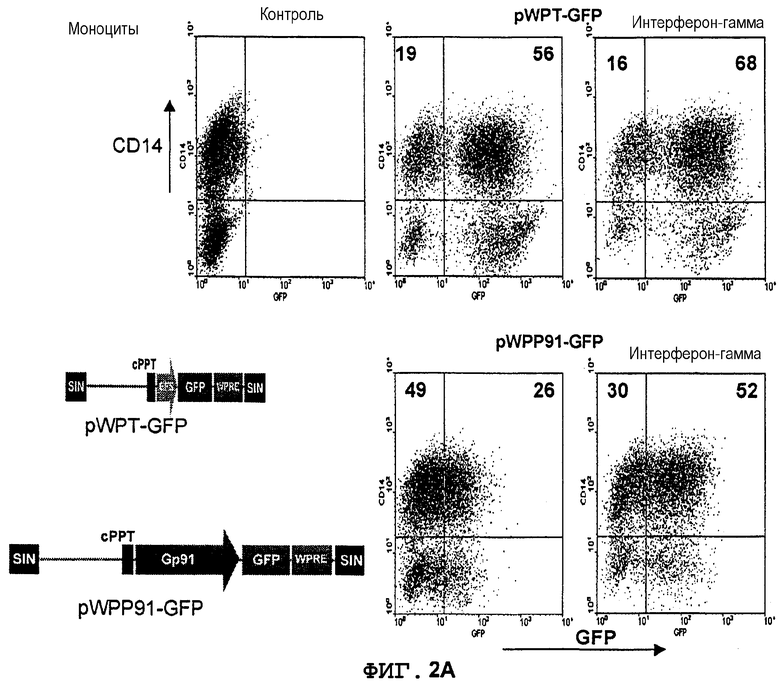
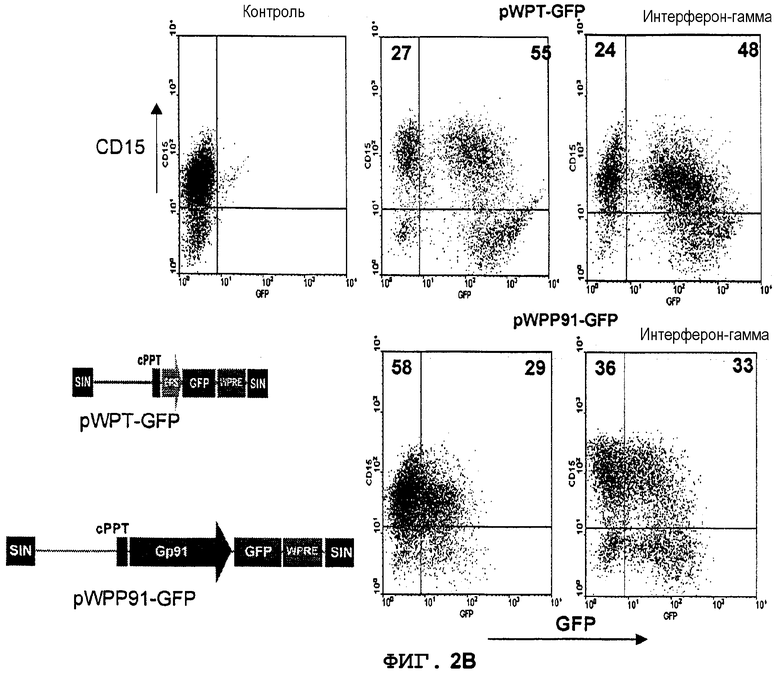
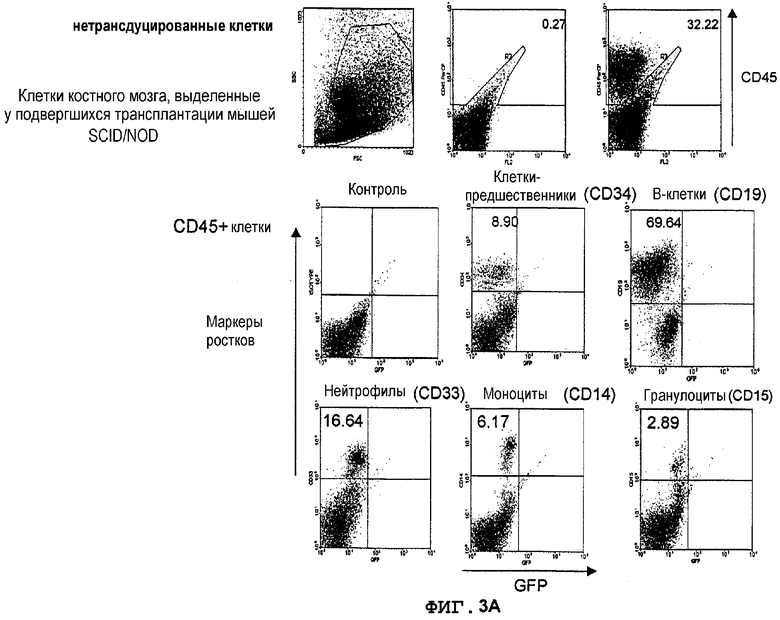
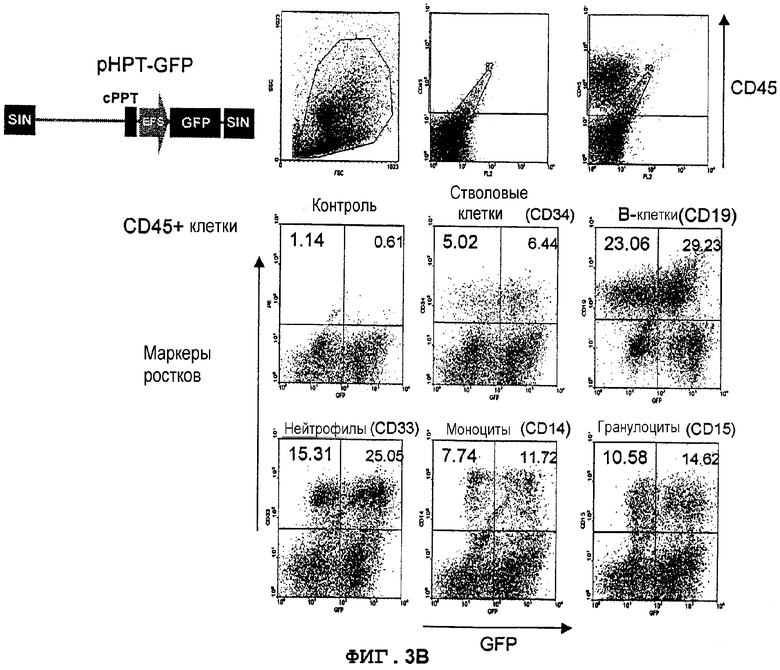
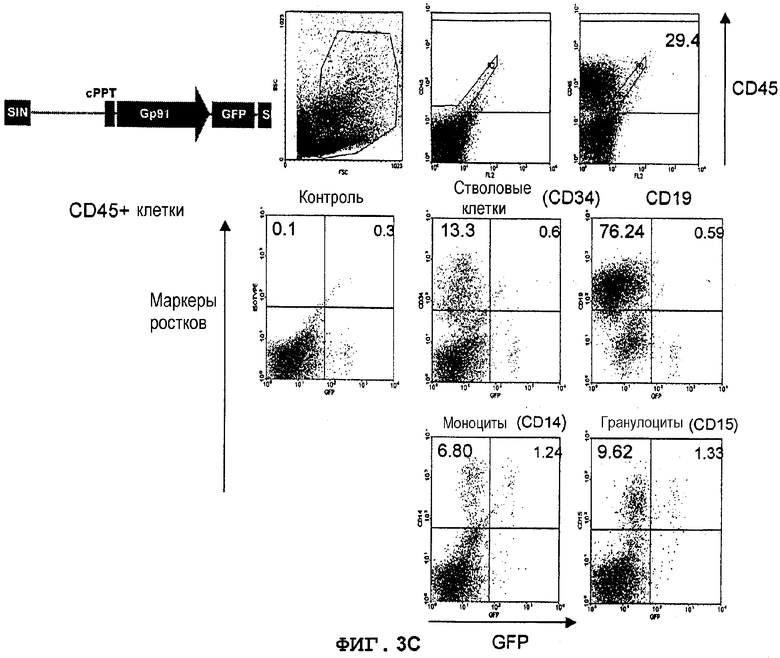
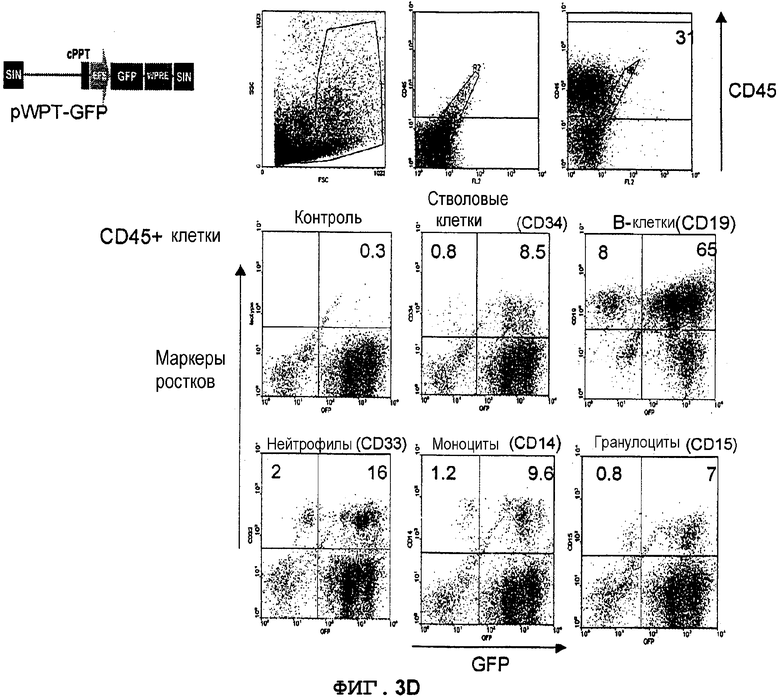
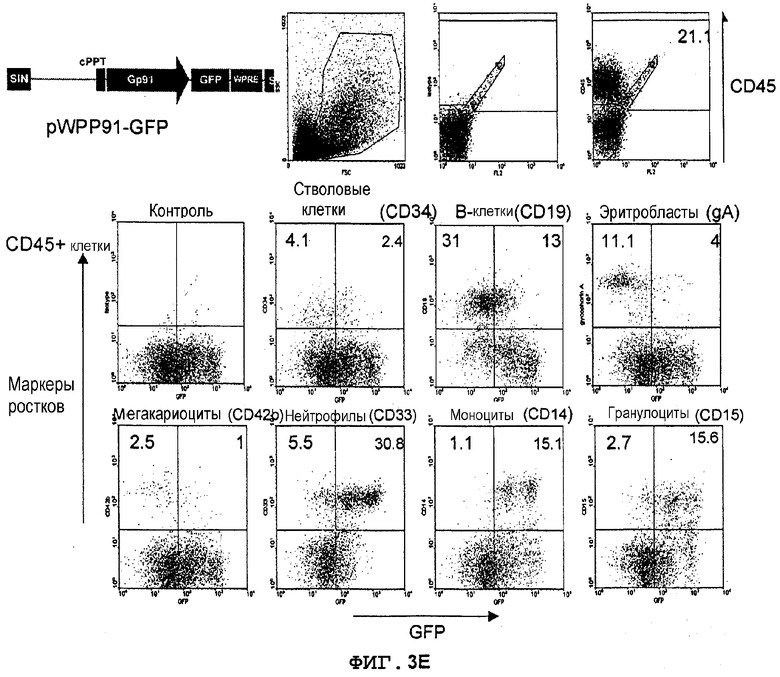
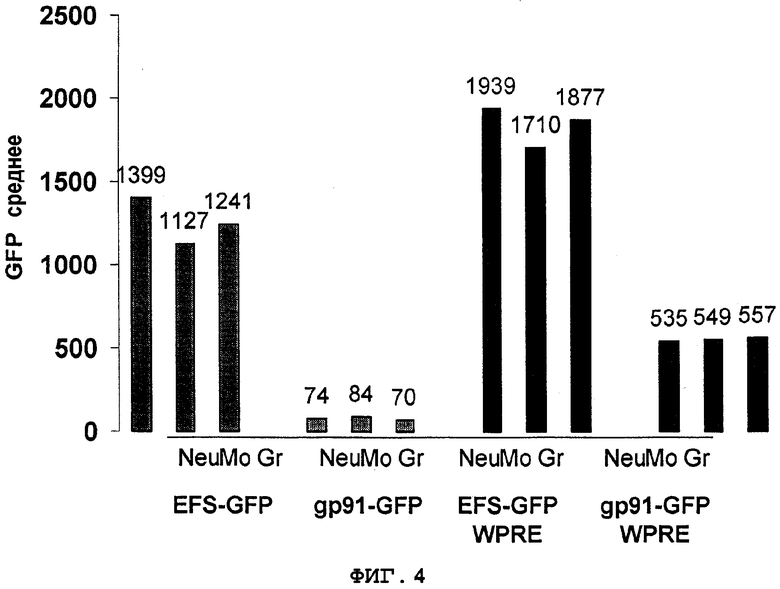
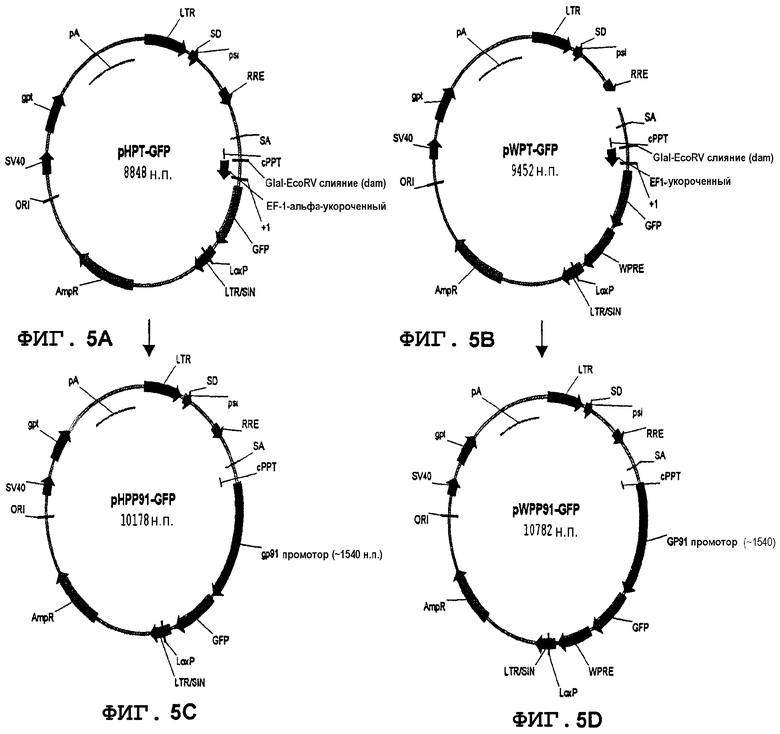
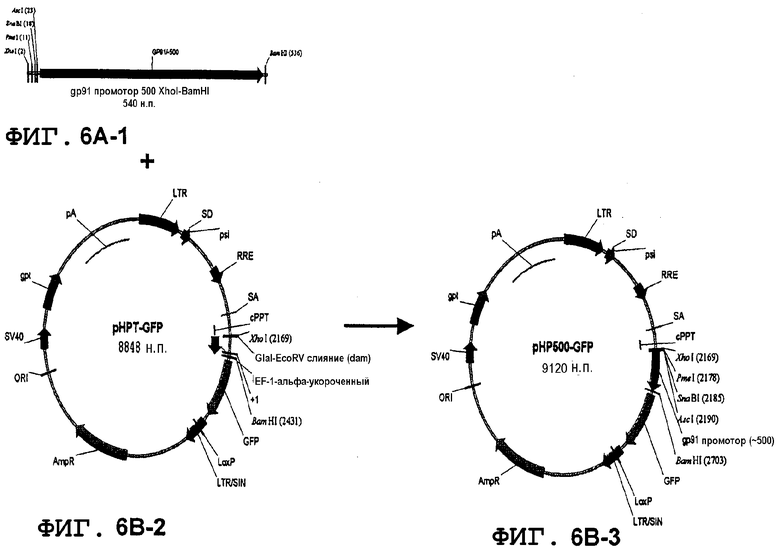
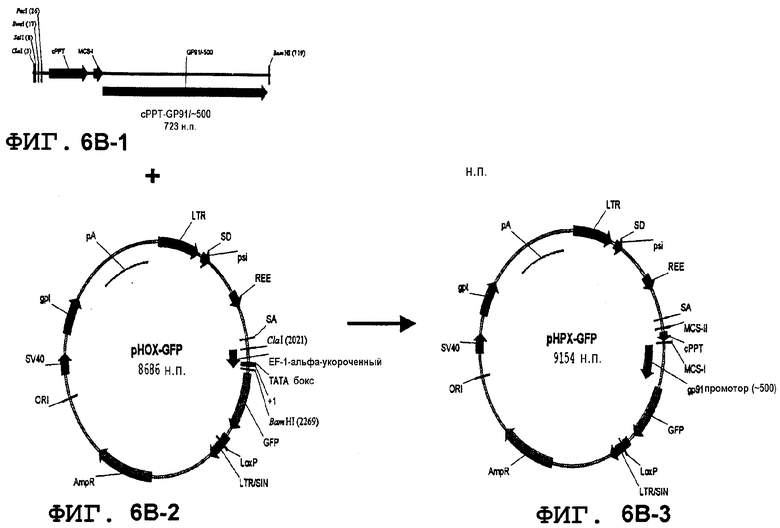

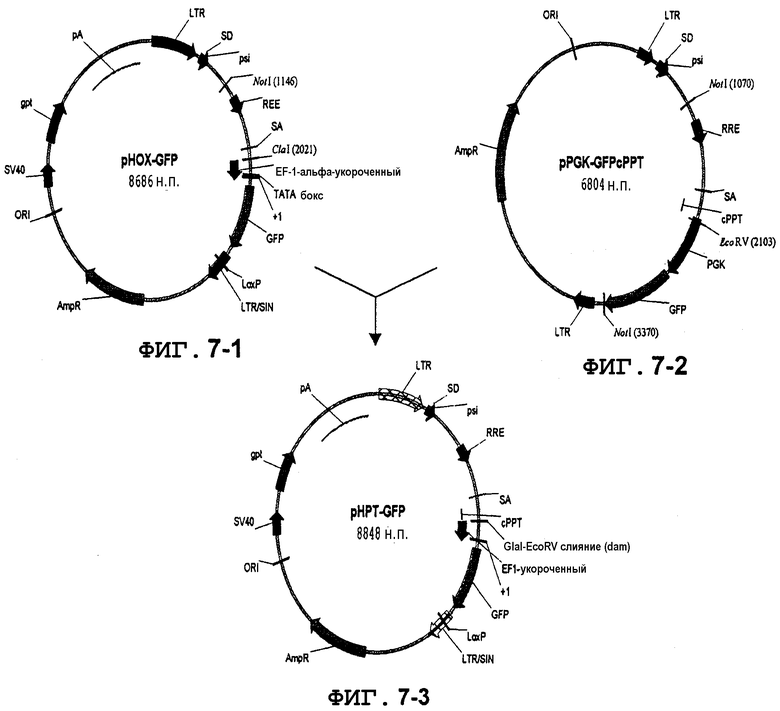
Изобретение относится к области генной инженерии. Вектор содержит экспрессионную кассету, содержащую трансген, расположенный под контролем промотора, и центральный полипуриновый тракт (сРРТ), расположенный выше кассеты. Вектор обеспечивает доставку желаемых трансгенов в клетки-мишени и их экспрессию в данных клетках на высоком уровне. Также предложены клетка-хозяин, трансдуцированная лентивирусным вектором, способ ее трансдукции и применение такой клетки. Изобретение может быть использовано в генной терапии. 24 н. и 5 з.п. ф-лы, 7 ил., 2 табл.
(a) экспрессионную кассету, содержащую трансген, расположенный под контролем промотора, который обладает активностью в отношении стимуляции выявляемой транскрипции трансгена в человеческих клетках человека; и
(b) центральный полипуриновый тракт (сРРТ), расположенный выше экспрессирующей кассеты.
| US 6165782, 26.12.2000 | |||
| US 6207455, 27.03.2001 | |||
| CHANG LJ | |||
| et al., «Efficacy and safety analyses of a recombinant human immunodeficiency virus type 1 derived vector system» | |||
| Gene Ther | |||
| Металлический водоудерживающий щит висячей системы | 1922 |
|
SU1999A1 |

