Настоящее изобретение относится к способу приготовления вектора экспрессии, кодирующего хорошо переносимую и высокоспецифическую адаптированную рекомбиназу, где адаптированная рекомбиназа способна к рекомбинированию асимметричных последовательностей-мишеней в пределах длинного концевого повтора (LTR) провирусной ДНК множества штаммов ретровирусов, которые могут быть встроены в геном клетки-хозяина, а также к полученному вектору экспрессии, клеткам, трансфицированным им, экспрессированной рекомбиназе и фармацевтическим композициям, содержащим клетки и/или рекомбиназу. Фармацевтические композиции пригодны, напр., в лечении и/или предотвращении ретровирусной инфекции, в частности, инфекции ВИЧ. В частности, изобретение относится к хорошо переносимым и высокоспецифическим адаптированным рекомбиназам, способным к комбинированию асимметричных последовательностей-мишеней в более чем 90% штаммов ВИЧ-1, посредством этого вырезая последовательности ВИЧ-1, и экспрессионным векторам, кодирующим их.
Ретровирусные инфекции, такие как, например, инфекции вирусом иммунодефицита человека (ВИЧ), все еще являются одним из наиболее серьезных и наиболее широко распространенных человеческих заболеваний.
Один из подходов к лечению ретровируса, напр., ВИЧ, заключается в нацеливании на провирус, встроенный в геном клетки-хозяина. Вырезание провирусной ДНК из генома хозяина, например, может предотвратить дальнейшую репликацию ВИЧ и отличается от существующих в настоящее время методологий, в которых он имеет потенциал уничтожения даже латентного вируса, представленного в геноме хозяина.
Одним из классов белков, который был рассмотрен для применения в этом альтернативном подходе, являются специфическые к участку рекомбиназы (FLOWERS et al, 1997). Специфическые к участку рекомбиназы опосредуют в природе множество функций от перестановки гена до сегрегации генома, такие как, например, вырезание, инверсия или интеграция определенных блоков ДНК (обзор в STARK et al, 1992).
Одной из наиболее простых и наиболее изученных рекомбиназ является рекомбиназа Cre от бактериофага PI, которая расщепляет геномные димеры в мономеры посредством рекомбинации между двумя идентичными, т.е., симметричными, участками двухцепочечных ДНК конкретной последовательности (HOESS & ABREMSKI, 1985). Рекомбиназа Cre нашла широкое применение в генетике мышей (NAGY, 2000). Cre представляет собой белок из 38 кДа, который назван в соответствии с его функцией, так как он вызывает рекомбинацию (STERNBERG & HAMILTON, 1981). Предпосылкой для этой рекомбинации является выравнивание двух рекомбинацинных участков, распознаваемых Cre в антипараллельной ориентации, которые затем бывают связаны четырьмя идентичными субъединицами Cre, которые связываются с образованием кольца, в котором каждая субъединица контактирует с двумя смежными субъединицами и одной половиной участка одного рекомбинационного участка (HOESS & ABREMS I, 1985). Рекомбинационный участок, распознаваемый Cre, представляет последовательность двухцепочечной ДНК из 34 п.о., известную как loxP (from locus of crossing over (x), P1; STERNBERG & HAMILTON, 1981), которая является палиндромной, за исключением ее восьми центральных пар оснований (называемых спейсер), что обеспечивает направленность на участок.
Некоторые специфическые к участку системы рекомбинации, включая систему Cre/loxP функционируют без акцессорных белков или кофакторов и функционируют в широком разнообразии клеточных условий. Однако, так как специфическые к участку рекомбиназы функционируют посредством специфических взаимодействий субъединицы фермента рекомбиназы с их когнатными последовательностями ДНК мишеней, применение этих ферментов ограничено тем условием, что области ДНК, на которые они направлены, должны содержать подходящим образом расположенные участки-мишени (LEWANDOSKI, 2001). К настоящему времени не было идентифицировано рекомбиназы дикого типа, которая распознает природные ретровирусные последовательности, в качестве их ДНК последовательностей-мишеней.
В последние годы были осуществлены экстенсивные мутационные и структурные анализы специфическых к участку рекомбиназ для изменения их свойств и для достижения наилучшего понимания многосложных механизмов этих ферментов (обзор см. в VAN DUYNE, 2001; и COATES et al, 2005). Множество исследований сфокусировано на рекомбиназе Cre для исследования возможности ее развития. Несколько исследований продемонстрировало, что специфичность Cre к мишени может быть изменена, когда несколько нуклеотидов в ее участке распознавания loxP были изменены (BUCHHOLZ & STEWART, 2001; SANTORO & SCHULTZ, 2002; RUFER & SAUER, 2002). Дальнейшее исследование направлено на конструирование мутированных участков-мишеней loxP, содержащих последовательности из LTR ВИЧ-1 для получения возможных участков-мишеней для применения Cre в качестве антивирусной стратегии (LEE & PARK, 1998; LEE et al., 2000).
Способ направленной эволюции является эффективным способом выбора ферментов с измененными специфичностями (обзор в Yuan et al., 2005; и JOHANNES & ZHAO, 2006). Сначала этот способ был использован для выделения улучшенных ферментов на основании РНК посредством выбора молекул РНК с измененными участками субстрата. Применение основанных на ПЦР способов позволяет скринирование очень больших библиотек и выделение удачных кодирующих областей из совокупности кандидатов. В направленной эволюции белков, в противоположность этому, для скринирования и выделения улучшенных мутантов, которые идентифицированы посредством изменений свойств белка, требуется способ извлечения последовательности нуклеиновой кислоты, кодирующей белок. Связь между белком и его кодирующей последовательностью часто бывает сохранена посредством компартментализации. Следовательно, скринирование библиотеки в направленной белковой эволюции было ограничено подходами "один за другим", которые сохраняют компартменты, и преимущества, связанные со скринированиями совокупности кандидатов, являлись недоступными.
Это ограничение было преодолено посредством разработки способов, которые позволяют перекрестное связывание белков с их соответствующими матричными РНК (мРНК) с применением слияний мРНК-белок и рибосомного дисплея. Таким образом, функциональные скринирования для улучшенных свойств белка были связаны с прямым извлечением соответствующих кодирующих молекул, и большие совокупности подвергали скринированию in vitro (см., например, BUCHHOLZ et al, 1998). Дальнейшее усовершенствование направленной эволюции белков было достигнуто посредством так называемой эволюции связанных с субстратом белков (SLiPE; BUCHHOLZ & STEWART, 2001), при которой субстрат рекомбиназы помещали в ту же молекулу ДНК, что и кодирующую белок область, таким образом, когда рекомбиназа была экспрессирована внутри компартмента, ее действие изменяло субстрат ДНК, следующий за ее собственной кодирующей областью. Следовательно, библиотека могла быть скринирована в виде совокупности посредством ПЦР для амплификации только кодирующих областей-кандидатов, которые были расположены после измененного субстрата. Это делает удобным скринирование больших библиотек для быстрого получения удачных кодирующих областей. Этот способ применяли для изменения ДНК-специфичности рекомбиназы Cre и адаптации ее к новому участку распознавания мишени (BUCHHOLZ & STEWART, 2001).
Принимая во внимание возможности сайт-специфической рекомбиназы и потребности в обнаружении лечения против СПИДа, удаляющего провирус ВИЧ-1 из генома клетки-хозяина, в WO 2008/083931 раскрыто генерирование адаптированной рекомбиназы (TRE), которая способна к рекомбинированию асимметричных участков-мишеней внутри LTR провирусной ДНК ретровируса, встроенного в геном клетки-хозяина, таким образом, вырезая провирус из генома клетки-хозяина. Сконструированная рекомбиназа Tre, раскрытая в примерах, распознает специфический асимметричный участок, представленный в конкретном штамме ВИЧ-1. Асимметричный участок-мишень имеет некоторую гомологию с симметричным участком loxP, распознаваемым Cre. В WO 2008/083931 принято во внимание то, что из-за высокой вариабельности последовательностей ретровирусов, в частности, ВИЧ, для лечения пациента с другим штаммом ВИЧ, другая адаптированная рекомбиназа должна быть адаптирована, или приготовлена коллекция рекомбиназ, содержащих адаптированные рекомбиназы, специфические для множества последовательностей-мишеней.
В отличие от этого WO 2011/147590 A2 относится к адаптированной рекомбиназе, способной к вырезанию множества ретровирусов, напр., штаммов ВИЧ. Таким образом, генерированная рекомбиназа может быть применена от множества инфекций ВИЧ без генерирования новой рекомбиназы для каждого штамма. Изобретатели обнаружили, что, несмотря на высокую вариабельность последовательности ретровирусов, с применением инновационного подхода было возможно идентифицировать асимметричные последовательности-мишени, представленные в большом количестве вирусов конкретного субтипа. Неожиданно, было возможно идентифицировать последовательность-мишень, представленную в 96% штаммах ВИЧ-1 субтипа B, т.е., преобладающих штаммов в Европе и Америке (SEQ ID NO:1). Также была идентифицированы дополнительная последовательность-мишень, представленная в более низком проценте штаммов ВИЧ-1 (SEQ ID NO:2). С применением Cre (SEQ ID NO: 6 в качестве основы для направленной молекулярной эволюции, авторы также идентифицировали несколько адаптированных рекомбиназ, способных к рекомбинированию указанных асимметричных последовательностей-мишеней, и предоставили консенсусные последовательности этих адаптированных рекомбиназ, напр., SEQ ID NO:7 или Tre 3.0 (SEQ ID NO:8, способная к рекомбинированию SEQ ID NO: 1).
В свете этого, изобретатели исследовали проблему получения улучшенной адаптированной рекомбиназы, способной к рекомбинированию асимметричных последовательностей-мишеней, представленных во множестве штаммов ВИЧ-1. Изобретатели неожиданно обнаружили, что адаптированные рекомбиназы, имеющие последовательность, отличающуюся от консенсусных последовательностей SEQ ID NO: 8, как раскрыто в WO 2011/147590 A2, также являются высокоактивными в отношении асимметричной последовательности-мишени SEQ ID NO: 1, представленной в 96% всех штаммов ВИЧ-1 субтипа B, т.е., в преобладающих штаммах в Европе и Америке, и имеют улучшенные характеристики. Адаптированные рекомбиназы в соответствии с изобретением предпочтительно содержат консенсусную аминокислотную последовательность SEQ ID NO: 9, более предпочтительно, более специфическую консенсусную последовательность SEQ ID NO: 10 или любую из SEQ ID NO: 11-13. Адаптированные рекомбиназы по настоящему изобретению имеют улучшенную специфичность и, следовательно, являются лучше переносимыми людьми, в частности, в человеческих T-клетках, чем адаптированные рекомбиназы в соответствии с современным уровнем техники. Рекомбиназы предпочтительно являются высокоспецифическими, так как они не обладают какой-либо поддающейся обнаружению остаточной активностью в отношении известных последовательностей-мишеней рекомбиназы, из которой они были сконструированы, напр., в отношении loxP (SEQ ID NO:4), loxH (SEQ ID NO:5) или также в отношении loxLTR Tre 1.0 (SEQ ID NO:3).
Настоящее изобретение впервые относится к способу генерирования вектора экспрессии, кодирующего хорошо переносимую и высокоспецифическую адаптированную рекомбиназу, способную к рекомбинированию асимметричных последовательностей-мишеней внутри LTR провирусной ДНК множества штаммов ретровирусов одних видов, встроенных в геном клетки-хозяина. Рекомбиназы были адаптированы для распознавания асимметричных участков-мишеней, отличных от их природных симметричных участков-мишеней, которые могут быть представлены во множестве штаммов ретровирусов, посредством разделения субстрата на ряд новых подгрупп с меньшими отличиями от первоначальных мишеней и посредством поэтапной адаптации рекомбиназ для распознавания этих подгрупп (WO 2008/083931 и WO 2011/147590). Комбинаторный подход позволяет отбор функциональных молекул, распознающих асимметричный участок-мишень в пределах определенной последовательности. Таким образом, посредством прохождения через промежуточные субстраты во время направленной молекулярной эволюции, было возможно продуцировать ферменты с сильно отличающимися новыми специфичностями к асимметричным мишеням. Этот подход также применен в представленном изобретении. Настоящее изобретение является добавлением к способам, раскрытым в WO 2008/083931 и WO 2011/147590, так как оно вводит стадии отбора адаптированных рекомбиназ, хорошо переносимых человеческими клетками, в частности, человеческими T-клетками.
Конкретно, изобретение относится к способу получения вектора экспрессии, кодирующего хорошо переносимую адаптированную рекомбиназу, в котором адаптированная рекомбиназа способна к рекомбинированию асимметричных последовательностей-мишеней внутри LTR провирусной ДНК множества штаммов ретровирусов, которая может быть встроена в геном клетки-хозяина, содержащему стадии идентифицирования в последовательности LTR провирусной ДНК множества штаммов ретровирусов последовательности с гомологией по меньшей мере 30% с последовательностью левой половины участка и последовательностью правой половины участка по меньшей мере одного известного участка-мишени рекомбиназы, при этом гомологичные последовательности разделены спейсером из 5-12 нуклеотидов, и при этом асимметричная последовательность-мишень обнаружена во множестве штаммов ретровирусов; и генерирование, посредством следующих повторяющихся стадий:
i) направленная молекулярная эволюция для по меньшей мере одной рекомбиназы, распознающей известный гомологичный участок-мишень с применением в качестве субстрата модифицированных последовательностей-мишеней, основанных на последовательности асимметричной последовательности-мишени, но модифицированной для содержания только ограниченного количества вариаций из известных последовательностей-мишеней; при этом в каждом цикле последовательность-мишень может отличаться от последовательности-мишени, на которую, как известно, рекомбиназа действует в одном в одном, двух или трех нуклеотидах; и
ii) перестановка в библиотеках рекомбиназ для получения библиотек рекомбиназ, способных рекомбинировать последовательности-мишени, более гомологичные асимметричной последовательности-мишени;
до тех пор пока не получают по меньшей мере одну рекомбиназу, которая является активной в отношении асимметричной последовательности-мишени внутри LTR ретровирусной ДНК;
повторяющиеся стадии негативного отбора против рекомбинации известного участка-мишени посредством направленной молекулярной эволюции и перестановки библиотек;
отбор указанной адаптированной рекомбиназы или рекомбиназ посредством экспрессии библиотеки в человеческих клетках, в частности, человеческих T-клетках, и культивирование указанных человеческих клеток, экспрессирующих адаптированную рекомбиназу(ы) в течение по меньшей мере 1 недели, предпочтительно, по меньшей мере 2 недель, и выделение нуклеиновой кислоты(кислот) рекомбиназы(рекомбиназ) из культивированных клеток, экспрессирующих селектируемый маркер;
и, необязательно, клонирование нуклеиновой кислоты, кодирующей рекомбиназу(ы) в подходящем векторе экспрессии.
Изобретение в частности относится к способу приготовления нуклеиновой кислоты или вектора экспрессии, кодирующего хорошо переносимую и высокоспецифическую адаптированную рекомбиназу, где адаптированная рекомбиназа способна к рекомбинированию асимметричных последовательностей-мишеней внутри LTR провирусной ДНК множества штаммов ретровирусов, содержащему следующие стадии:
(a) идентифицирование последовательности с гомологией по меньшей мере 30% с последовательностью левой половины участка и последовательностью правой половины участка по меньшей мере одного известного участка-мишени рекомбиназы в последовательности LTR провирусной ДНК множества штаммов ретровирусов, при этом гомологичные последовательности разделены спейсером из 5-12 нуклеотидов, и при этом асимметричная последовательность-мишень обнаружена во множестве штаммов ретровирусов;
(b) идентифицирование двух последовательностей, при этом первая последовательность соответствует последовательности асимметричной последовательности-мишени стадии (a), гомологичной левой половине участка указанного известного участка-мишени и называется "последовательность 1 половины участка", и при этом вторая последовательность соответствует последовательности асимметричной последовательности-мишени стадии (a), гомологичной правой половине участка, и называется "последовательность 2 половины участка";
(c) определение нуклеотидов внутри последовательностей стадии (b), отличающихся от соответствующих гомологичных последовательностей левой половины участка и правой половины участка по меньшей мере одного известного гомологичного участка-мишени стадии (a);
(d) генерирование первой подгруппы двух мишеневых нуклеиновых кислот, содержащих последовательности-мишени, при этом первая последовательность-мишень обозначена субучасток 1 и содержит смежно друг другу и в порядке от 5' к 3' последовательность 1 половины участка стадии (b), спейсерную последовательность асимметричной последовательности-мишени и инвертированный повтор последовательности 1 половины участка, и при этом вторая последовательность-мишень обозначена субучасток 2 и содержит смежно друг другу и в порядке от 5ʹ к 3ʹ инвертированный повтор последовательности 2 половины участка, спейсерную последовательность асимметричной последовательности-мишени и последовательность 2 половины участка стадии (b);
(e) генерирование второй подгруппы мишеневых нуклеиновых кислот, содержащих модифицированные последовательности-мишени на основании последовательностей-мишеней в первой подгруппе стадии (d),
при этом в последовательностях, основанных на субучастке 1 в последовательности левой половины участка, участок нуклеотидов, отличающихся от последовательности соответствующей гомологичной половины участка по меньшей мере одного известного участка-мишени стадии (a), замещен природными нуклеотидами, обнаруженными в указанном известном участке-мишени, при условии что последовательность указанной половины участка содержит один, два или три нуклеотида, отличающихся от указанного известного участка-мишени, при этом правая половина участка указанной модифицированной последовательности-мишени образована инвертированным повтором указанной модифицированной последовательности левой половины участка, которая отделена от указанной модифицированной последовательности левой половины участка спейсерной последовательностью асимметричной последовательности-мишени, и
при этом в последовательностях, основанных на субучастке 2, в последовательности правой половины участка, участок нуклеотидов, отличающихся от последовательности соответствующей гомологичной половины участка по меньшей мере одного известного участка-мишени стадии (a), замещен природными нуклеотидами, обнаруженными в указанном известном участке-мишени, при условии что последовательность указанной половины участка содержит один, два или три нуклеотида, отличающихся от указанного известного участка-мишени, при этом левая половина участка указанной модифицированной последовательности-мишени образована инвертированным повтором указанной последовательности модифицированной правой половины участка, которая отделена от указанной последовательности модифицированной правой половины участка спейсерной последовательностью асимметричной последовательности-мишени,
так, что во всех последовательностях модифицированных половин участка, происходящих от одной последовательности-мишени первой подгруппы стадии (d), в совокупности могут быть обнаружены все отличающиеся нуклеотиды, тогда как ни одна из указанных модифицированных последовательностей половины участка не содержит отдельно все отличающиеся нуклеотиды,
(f) использование направленной молекулярной эволюции отдельно для по меньшей мере одной рекомбиназы, распознающей известный гомологичный участок-мишень в соответствии со стадией (a), с применением каждой нуклеиновой кислоты второй подгруппы, полученной на стадии (e) в качестве субстрата;
(g) перестановка в библиотеках рекомбиназ, полученных на стадии (f), при этом все библиотеки рекомбиназ полученные для последовательностей, основанных на субучастке 1, комбинируют и подвергают перестановкам, и при этом все библиотеки рекомбиназ полученные для последовательностей, основанных на субучастке 2, комбинируют и подвергают перестановкам;
(h) использование направленной молекулярной эволюции, предпочтительно, связанной с субстратом эволюции белка, для библиотек с перестановками, полученных на стадии (g), с применением каждой нуклеиновой кислоты подгруппы в соответствии со стадией (d) в качестве субстрата;
(i) перестановка в библиотеках рекомбиназ, полученных на стадии (h);
(j) использование направленной молекулярной эволюции, предпочтительно, связанной с субстратом эволюции белка, для библиотеки с перестановками, полученной на стадии (g), с применением нуклеиновой кислоты, содержащей асимметричную последовательность-мишень стадии (a) в качестве субстрата, при условии, что получена по меньшей мере одна рекомбиназа, которая активна в отношении асимметричной последовательности-мишени внутри LTR ретровирусной ДНК стадии (a);
(k) выделение нуклеиновой кислоты, кодирующей по меньшей мере одну рекомбиназу, полученную на стадии (j), из библиотеки и клонирование ее в эволюционном векторе, который позволяет негативный отбор адаптированных рекомбиназ рекомбинированием известного участка-мишени в соответствии со стадией (a), посредством этого получая библиотеку;
(l) использование направленной молекулярной эволюции, предпочтительно, связанной с субстратом эволюции белка, для библиотеки, полученной на стадии (k);
(m) перестановка в библиотеке, полученной на стадии (l);
(n) выделение нуклеиновой кислоты, кодирующей по меньшей мере одну адаптированную рекомбиназу, полученную на стадии (m), и клонирование ее в вектор для экспрессии кодируемой рекомбиназы и селектируемого маркера в человеческой клетке, посредством этого получая библиотеку векторов,
(o) трансформирование человеческих клеток, предпочтительно, человеческих T-клеток, указанной библиотекой векторов, полученной на стадии (n);
(p) культивирование клеток, экспрессирующих указанный селектируемый маркер, в течение по меньшей мере 1 недели и отбор для высокой экспрессии селектируемого маркера;
(q) выделение нуклеиновой кислоты(кислот), кодирующей рекомбиназу из клеток, экспрессирующих указанный селектируемый маркер, полученных на стадии (p);
(r) отбор на нуклеиновую кислоту, кодирующую рекомбиназу, способную к рекомбинированию асимметричной последовательности-мишени стадии (a);
(s) выделение нуклеиновой кислоты, кодирующей по меньшей мере одну рекомбиназу, полученную на стадии (f), из библиотеки; и,
(t) необязательно, клонирование нуклеиновой кислоты, полученной на стадии(ях), в подходящем векторе экспрессии.
На стадии (a) способа настоящего изобретения последовательность LTR провирусной ДНК может быть определена, например, посредством секвенирования ДНК с применением ингибиторов терминации цепи (SANGER et al, 1977). Однако, если последовательность LTR ретровирусной ДНК, встроенной в геном хозяина, уже была определена, то последовательность может быть определена посредством ссылки на базу данных. На основании информации о последовательности выполнен основанный на применении компьютера анализ информации о последовательности для идентифицирования в ней последовательностей с гомологией по меньшей мере 30% с последовательностью левой половины участка и правой половины участка известных участков-мишеней, соответственно, известных рекомбиназ, которые разделены подходящим спейсером из 5-12 нуклеотидов, при этом асимметричная последовательность-мишень обнаружена во множестве штаммов ретровирусов. Предпочтительно, гомология последовательности левой половины участка и правой половины участка известных участков-мишеней составляет по меньшей мере 40% или по меньшей мере 50%. Предпочтительно, эти штаммы ретровирусов принадлежат одному виду или его одному субтипу. Предпочтительно, множество штаммов содержит более чем 10 штаммов, более предпочтительно, более чем 100 штаммов, более чем 130 штаммов, более чем 200 штаммов или более чем 300 штаммов, напр., штаммов ВИЧ. Штаммы могут принадлежать одному субтипу вируса, напр., ВИЧ-1, субтип A, B и C ВИЧ-1, предпочтительно, субтипу B ВИЧ-1. Таким образом, полученная рекомбиназа или вектор экспрессии, кодирующий ее, может быть применена для лечения инфекции с множеством штаммов, напр., более чем 50%, более чем 70%, более чем 80%, более чем 90% или всех известных штаммов ретровируса или его субтипов.
Термин "рекомбиназа", как применено в данном документе, относится к белку, задействованному в рекомбинации. По существу рекомбиназы распознают и связывают две специфические последовательности ДНК, обозначенные "участки рекомбинации" или "участки-мишени", и опосредуют рекомбинацию между этими двумя участками-мишенями. Соответственно, термин "рекомбиназа" подразумевается для обозначения любого белкового компонента любой рекомбинантной системы, которая опосредует перегруппировки ДНК в специфическом локусе ДНК. Встречающиеся в природе рекомбиназы распознают симметричные участки-мишени, состоящие из двух идентичных последовательностей, обозначенных "половины участков", из приблизительно 9-20 п.о., образующих инвертированный повтор, при этом последовательности половины участка разделены спейсерной последовательностью из 5-12 п.о. Рекомбиназы из семейства тирозиновых интеграз характеризуются наличием тирозина в качестве нуклеофила активного участка, что бывает применено для расщепления ДНК, тогда как рекомбиназы из семейства сериновых интеграз применяют серин вместо тирозина.
В одном варианте осуществления настоящего изобретения по меньшей мере одна известная рекомбиназа, последовательность-мишень которой применена на стадии (a) и по отношению к которой применена направленная молекулярная эволюция в этапах (h) и (j), принадлежит к семейству сериновых интеграз. Предпочтительные рекомбиназы, принадлежащие к семейству сериновых интеграз, выбраны из группы, состоящей из интегразы phiC31 (COMBES et al., 2002), любого компонента систем рекомбинации Gin или Hin, Tn3 резольвазы (KRASNOW & COZZARELLI, 1983) или любого другого члена больших сериновых рекомбиназ, Rag1, Rag2 или любого другого компонента рекомбинационной системы VDJ или их вариантов.
В другом варианте осуществления указанная рекомбиназа принадлежит к семейству тирозиновых интеграз. Предпочтительные рекомбиназы, принадлежащие к семейству тирозиновых интеграз, выбраны из группы, состоящей из Cre из Phage P1 (ABREMSKI et al, 1983, 1984), рекомбиназы FLP из дрожжей (VOLERT & BROACH, 1986), Dre из фага D6 (SAUER & MCDERMOTT, 2004), рекомбиназы R из Zygosaccharomyces rouxii плазмиды pSR1, рекомбиназы A из Kluveromyces drosophilarium плазмиды pKDl, рекомбиназы A из Kluveromyces waltii плазмиды pKW1, TnpI из транспозона Tn4430 Bacillus, любого компонента из рекомбинационной системы λ Int или их вариантов. Предпочтительно указанная рекомбиназа является рекомбиназой Cre или ее вариантом.
Термин вариант в этом контексте относится к белкам, которые получены из вышеприведенных белков делецией, заменой и/или добавлением аминокислот и которые сохраняют некоторые или все из функций, свойственных белку, из которого они получены.
В предпочтительном варианте осуществления известная рекомбиназа является химерной рекомбиназой, полученной, например, "перестановками семейств", как описано CRAMERI et al. (1998). Предпосылкой для задействования перестановки семейств является значительная гомология между рекомбиназами, применяемыми для генерирования химерных рекомбиназ. Примером химерной рекомбиназы, которая может быть применена в настоящем изобретении, является химерная рекомбиназа, состоящая из последовательности рекомбиназы Cre и рекомбиназы Dre, соответственно.
В более предпочтительном варианте осуществления рекомбиназа является рекомбиназой Cre, распознающей симметричный участок-мишень из 34 п.о., известный как loxP. Участок loxP (а также другие участки рекомбинации рекомбиназ дикого типа) является палиндромным с двумя повторами из 13 п.о., разделенными восемью центральными парами оснований, которые представляют собой так называемый спейсер, который придает участку направленность. Рекомбинация происходит посредством расщепления внутри последовательности спейсера. В зависимости от относительного расположения и ориентации двух принимающих участие участков loxP Cre катализирует интеграцию, вырезание или перестановку ДНК (HOESS & ABREMSKI, 1985).
Одной из пригодных рекомбиназ является Zre, выделенная из Salmonella enterica, или ее варианты, фрагменты и гомологи, напр., имеющие гомологию по меньшей мере приблизительно 70%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 90% или по меньшей мере приблизительно 95% с последовательностью дикого типа и имеющие рекомбиназную функцию. Рекомбиназы Zre рекомбинируют ДНК в участках zox. Они могут быть применены в качестве начального способа по изобретению или отдельно, или в контексте библиотеки.
В наиболее предпочтительном варианте осуществления библиотека рекомбиназ применена в качестве исходной точки для молекулярной эволюции, напр., библиотека рекомбиназ, содержащей другие рекомбиназы дикого типа и/или адаптированные/рекомбиназы с перестановками, напр., как описано, напр., в примере 2 WO 2011/147590 A2. Такая библиотека предпочтительно бывает применена в качестве исходной точки, используемой для генерирования адаптированных рекомбиназ, способных к распознаванию SEQ ID NO:1 или альтернативно SEQ ID NO:2.
Адаптированная рекомбиназа, полученная посредством способа по изобретению, способна к рекомбинированию асимметричных последовательностей-мишеней внутри LTR провирусной ДНК множества штаммов ретровирусов. Провирусная ДНК, к которой специфична рекомбиназа, может быть встроена в геном клетки-хозяина. Альтернативно, адаптированная рекомбиназа изобретения может рекомбинировать асимметричные последовательности-мишени внутри LTR провирусной ДНК множества штаммов ретровирусов, которые еще не являются интегрированными в геном клетки-хозяина, т.е. которые представлены в виде неинтегрированного преинтеграционного комплекса (PIC). Таким образом, ВИЧ, которая еще не интегрировалась в геном клетки-хозяина, так же как и ВИЧ, которая уже интегрировалась, может быть инактивирована посредством адаптированной рекомбиназы по изобретению.
Необходимо отметить, что в настоящем изобретении, а также в данной области, термины "последовательность-мишень", "участок-мишень" и "рекомбинационный участок" применяются взаимозаменяемо.
В противоположность встречающимся в природе рекомбиназам, распознающим симметричный участок-мишень, способ по настоящему изобретению относится к адаптированным рекомбиназам, распознающим участки-мишени, которые не состоят из палиндромных последовательностей, разделенных спейсером. Вместо этого, в асимметричных участках-мишенях последовательности не образуют симметричный инвертированный повтор. Соответственно, адаптированная рекомбиназа, способная к распознаванию асимметричного участка-мишени, должна распознавать и рекомбинировать участки-мишени, состоящие из половин участков варьирующейся последовательности.
Внутри асимметричного участка-мишени последовательности, называемые "левая половина участка" и "правая половина участка", соответственно, определены по их гомологии с левой и правой половиной участка известного участка-мишени. Последовательность, расположенная между последовательностями, гомологичными левой и правой половине участка известного участка-мишени, называется спейсер.
Однако если последовательности обнаружены в LTR, которая имеет гомологию только с последовательностью или левой, или правой половины участка известного участка-мишени, эти последовательности, тем не менее, могут быть применены при воплощении на практике настоящего изобретения. Размер участка-мишени, принадлежащего рекомбиназе, природная последовательность-мишень которого демонстрирует гомологию с последовательностями внутри LTR, известен специалисту в данной области. Например, если гомология обнаружена внутри последовательности LTR с последовательностью-мишенью, распознаваемой рекомбиназой Cre, то асимметричный участок-мишень, который должен быть распознан рекомбиназой Cre, должен состоять из 34 нуклеотидов с двумя последовательностями половины участка из 13 нуклеотидов каждый, разделенными спейсером из 8 нуклеотидов. Соответственно, гомологичная последовательность внутри LTR определена в виде или левой, или правой половины участка или спейсера асимметричного участка-мишени в зависимости от гомологии с последовательностью известного участка-мишени. Таким образом, последовательности с гомологией с левой половиной участка известной последовательности-мишени определены в виде левой половины участка, последовательности с гомологией с правой половиной участка известной последовательности-мишени определены в виде правой половины участка. Исходя из этого определения другие части асимметричных участков-мишеней определены при рассмотрении структуры известного участка-мишени. Таким образом, определив, например, последовательность правой половины участка внутри LTR по гомологии c участком loxP (распознаваемым рекомбиназой Cre), могут быть легко определены другие последовательности, соответствующие последовательности спейсера и левой половине участка асимметричной последовательности-мишени. Спейсерная последовательность, например, определена посредством подсчета 8 нуклеотидов в обратном направлении от 5' конца последовательности, определенной как последовательность правой половины участка, тогда как последовательность левой половины участка определена подобным образом посредством подсчета 13 нуклеотидов в обратном направлении от 5' конца ранее определенной последовательности спейсера.
Гомология в этом контексте, а также во всей заявке, означает идентичность последовательности. Предпочтительным сравнением в целях выявления гомологии является сравнение по меньшей мере двух последовательностей с использованием стандартных технологий, известных в данной области, включая, но без ограничения ими, алгоритм локальной гомологии SMITH & WATERMAN (1981), алгоритм выравнивания по гомологии NEEDLEMAN & WUNSCH (1970) или способ поиска сходства PEARSON & LIPMAN (1988). В целях настоящей заявки, если не указано иное, гомология последовательности предпочтительно определена с применением компьютерной программы ClustalW, доступной от European Bioinformatics Institute (EBI).
С точки зрения требования, что два идентичных участка-мишени должны быть представлены в геноме провируса для позволения вырезания рекомбиназой последовательности между этими двумя участками-мишенями, последовательности провирусной ДНК, сканированные на стадии (a) способа настоящего изобретения, должны быть представлены в геноме по меньшей мере два раза. Такие последовательности, например, представляют собой последовательности LTR провирусной ДНК. Соответственно, последовательность LTR предпочтительно бывает сканирована, так как 5'-LTR и 3'-LTR провирусной ДНК являются идентичными. Асимметричный участок-мишень, представленный в 5'-LTR, также представлен в 3'-LTR и, таким образом, позволяет вырезание провирусной ДНК, расположенной между LTR.
Из последовательностей, идентифицированных внутри последовательности LTR, имеющей достаточную гомологию с известными участками-мишенями, предпочтительно выбраны последовательности, которые имеют наибольшую гомологию с последовательностью участка-мишени известных рекомбиназ. Однако также возможно выбирать последовательности, отличающиеся от последовательностей, имеющих другую наибольшую гомологию, напр., последовательности, которые представлены в наибольшем количестве штаммов ретровирусов, или в штаммах ретровирусов, представляющих интерес, напр., если пациент инфицирован с конкретным штаммом.
Необходимо отметить, что возможности способа по настоящему изобретению позволяют адаптировать даже рекомбиназы, которые распознают асимметричные участки-мишени с менее чем 30% гомологии с известными участками-мишенями, напр., по меньшей мере 11% или по меньшей мере 20% гомологии. Однако, для обеспечения наличия остаточной рекомбинационной активности для соответствующего асимметричного участка-мишени или его субучастков, предпочтительно сканировать на последовательности, имеющие гомологию по меньшей мере 30% с последовательностями левой половины участка и правой половины участка известных участков-мишеней известных рекомбиназ. В дополнительных предпочтительных вариантах осуществления выбраны асимметричные последовательности-мишени, имеющие гомологию по меньшей мере 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80%, более предпочтительно 85%, особенно предпочтительно 90% и наиболее предпочтительно 95% с последовательностями левой половины участка и правой половины участка известных участков-мишеней известных рекомбиназ.
В одном варианте осуществления настоящего изобретения последовательность, выбранная внутри LTR, имеет гомологию с симметричными участками-мишенями loxP, распознаваемыми специфической к участку рекомбиназой Cre.
В предпочтительном варианте осуществления библиотека рекомбиназ применена в качестве исходной точки для молекулярной эволюции, напр., библиотека рекомбиназ, содержащая другие рекомбиназы дикого типа и/или адаптированные/рекомбиназы с перестановками, такие как библиотека, описанная в примере 2 WO 2011/147590 A2. Иллюстративная библиотека содержит Cre и рекомбиназы, полученные из нее. Она также может содержать рекомбиназы Tre, Dre, рекомбиназы из Salmonella и Shewanella и/или рекомбиназы, полученные из них. Библиотека может содержать, напр., Cre, Dre, Dre "Cre-ed", рекомбиназу Shewanella (Shew), Shew "Cre-ed", и/или Zre, как раскрыто в WO 2011/147590 A2. Tre представляет собой адаптированную рекомбиназу, как раскрыто WO 2008/083931, которая также в дальнейшем называется Tre 1.0.
В одном варианте осуществления все рекомбиназы в библиотеке распознают последовательность-мишень с одинаковой длиной спейсера. Общая длина последовательностей половины участка 1 и 2, включая спейсер, предпочтительно составляет 34 нуклеотида.
Если по меньшей мере одна рекомбиназа является библиотекой рекомбиназ, то гомология является гомологией с совокупностью известных участков-мишеней рекомбиназ (т.е., гомология в определенном положении с по меньшей мере одной из последовательностей-мишеней определена как гомология). Следовательно, на стадии (c) только те нуклеотиды, которые не соответствуют нуклеотиду в по меньшей мере одной из известных последовательностей-мишеней, определены как отличающиеся нуклеотиды. В случае библиотеки рекомбиназ "природный нуклеотид" на стадии (e) может являться нуклеотидом, представленным в этом положении в любой из известных последовательностей-мишеней, предпочтительно, он является нуклеотидом, представленным в этом положении в нескольких или большинстве из известных последовательностей-мишеней.
Для идентифицирования последовательностей-мишеней, представленных во множестве штаммов ретровирусов, могут быть применены известные участки распознавания рекомбиназ, которые были описаны в литературе, в качестве поискового запроса для поиска на консервативные асимметричные последовательности-мишени против геномного удлинения. Однако учитывая повторяемую природу областей, применение стандартного средства поиска подобия последовательностей исключено. Sarkar et al., 2007, применяли BLAST (ALTSCHUL et al., 1997) для обнаружения подобного lox связывающего участка в штаммах ВИЧ. Поиск BLAST на участок, подобный lox при выполнении по последовательностям LTR ВИЧ-1 привел в результате к обнаружению только одного участка, представленного в единственном штамме. BLAST не бывает выполнен хорошо с такими короткими избыточными последовательностями, также было доказано, что альтернативные программы, такие как HMMER (EDDY et al, 1998), RepeatMasker или палиндромная программа из пакета Emboss suite, также не являются неподходящими. Со специфической программой с применением алгоритма, основанного на позиционной весовой матрице на фланкирующие области, основанной на известном участке распознавания рекомбиназы, и с применением бинарных операций на последовательности после того, как они преобразованы в битовые строки, были идентифицированы асимметричные последовательности-мишени, обнаруженные во множестве штаммов ретровирусов, WO 2011/147590 A2.
Для ВИЧ-1 были определены подходящие асимметричные последовательности-мишени, имеющие последовательность, представленную далее как SEQ ID NO:1 или SEQ ID NO:2. Это делает возможным генерирование рекомбиназ, которые пригодны практически в качестве терапевтических агентов против ретровирусных геномов для значительного количества пациентов, так как эти рекомбиназы нацелены на участки распознавания, представленные в настолько большом количестве штаммов ретровируса, насколько это возможно.
Последовательности левой половины участка и правой половины участка SEQ ID NO: 1 и 2 подчеркнуты, и спейсер напечатан жирным шрифтом:
SEQ ID NO: l AACCCACTGCTTAAGCCTCAATAAAGCTTGCCTT SEQ ID NO: 2 CTGGGCGGGACTGGGGAGTGGCGAGCCCTCAGAT
SEQ ID NO: l представлена в 96% штаммов ВИЧ-1 субтипа B, подвергнутых поиску (1024/1067), в 92% штаммов ВИЧ-1 субтипа C, подвергнутых поиску (624/679), и в 82% штаммов ВИЧ-1 субтипа A, подвергнутых поиску (71/87). SEQ ID NO: 2 идентична в более низком проценте штаммов субтипа B и C.
SEQ ID NO: l имеет 54% гомологию с совокупностью известных участков-мишеней рекомбиназы, и SEQ ID NO:2 имеет 42% гомологии с совокупностью этих последовательностей (по отношению к левой и правой половине участков, соответственно). Гомология с индивидуальными известными участками-мишенями является более низкой, напр., по меньшей мере 30% для SEQ ID NO: l и по меньшей мере 11% для SEQ ID NO:2. В частности в случае низкой индивидуальной гомологии с известными участками-мишенями, может иметь преимущество использование библиотеки рекомбиназ в качестве исходного материала, напр., для генерирования адаптированной рекомбиназы, способной к рекомбинированию SEQ ID NO: l или SEQ ID NO:2, при этом библиотека содержит Cre, Fre, Dre, Zre и Tre.
На стадии (b) способа по изобретению последовательность асимметричного участка-мишени внутри LTR провирусов, которая является гомологичной левой половине участка известного участка-мишени определена как "последовательность 1 половины участка". Последовательность асимметричного участка-мишени внутри LTR провирусов, которая гомологична правой половине участка известного участка-мишени, определена как последовательность 2 половины участка. Последовательность между последовательностями, представляющими левую и правую половину участка, называется спейсер.
На стадии (c) нуклеотиды внутри "последовательности 1 половины участка" и "последовательности 2 половины участка", соответственно последовательностей стадии (b), отличающихся от последовательностей соответствующей гомологичной левой половины участка и правой половины участка известных мишеней, определены посредством выравнивания последовательностей и сравнения последовательностей. В этом контексте последовательность "последовательности 1 половины участка" сравнивают с соответствующей природной половиной участка, которая предпочтительно является последовательностью левой половины участка, тогда как последовательность "последовательности 2 половины участка" сравнивают с другой половиной участка, образующей палиндромный природный участок-мишень, которая предпочтительно является последовательностью правой половины участка.
Фиг. 1 WO 2011/147590 A2 демонстрирует результат этого сравнения для SEQ ID NO: l и 2, по сравнению с библиотекой рекомбиназ. Отличающиеся нуклеотиды продемонстрированы перед темным фоном.
Это сравнение не должно быть обязательно выполнено после стадии (b) и перед стадией (d) способа по изобретению, а может также быть выполнено в другой фазе способа после стадии (a) и перед стадией (e).
На стадии (d) генерирована первая подгруппа из двух мишеневых нуклеиновых кислот, содержащая последовательности-мишени, при этом первая последовательность-мишень обозначена субучасток 1 и содержит смежно друг другу и в порядке от 5ʹ к 3ʹ последовательность 1 половины участка стадии (b), спейсерную последовательность асимметричной последовательности-мишени и инвертированный повтор последовательности 1 половины участка, и при этом вторая последовательность-мишень обозначена субучасток 2 и содержит смежно друг другу и в порядке от 5ʹ к 3ʹ инвертированный повтор последовательности 2 половины участка, спейсерную последовательность асимметричной последовательности-мишени и последовательность 2 половины участка стадии (b). Последовательности-мишени первой подгруппы являются палиндромными олигонуклеотидным последовательностям, имеющими структуру симметричного участка-мишени. Эти искусственные симметричные участки-мишени синтезированы на основании последовательностей половины участка стадии (b) посредством дополнения отсутствующей последовательности половины участка в каждой олигонуклеотидной последовательности, в виде инвертированного повтора, в котором последовательность "последовательности 1 половины участка" и "последовательности 2 половины участка", соответственно, применена для дополнения последовательности второй половины участка на противоположном конце последовательности спейсера. Соответственно, первая последовательность-мишень в первой подгруппе (называемой "субучасток 1") содержит инвертированный повтор, состоящий из "последовательности 1 половины участка" и повторяющейся в обратном порядке "последовательности половины участка", разделенные спейсерной последовательностью, тогда как вторая последовательность-мишень в первой подгруппе (называемая "субучасток 2") содержит инвертированный повтор, состоящий из повторяющейся в обратном порядке "последовательности 2 половины участка"' и "последовательности 2 половины участка", разделенные спейсерной последовательностью. В "субучастке 1" последовательности расположены следующим образом: 5'-"последовательность 1 половины участка"-спейсер -"инвертированный повтор последовательности 1 половины участка"-3', в "субучастке 2" последовательности расположены следующим образом: 5'-"инвертированный повтор последовательности 2 половины участкаʺ-спейсер -"последовательность 2 половины участка"- 3'.
Спейсерные последовательности внутри каждых двух синтетических последовательностей-мишеней первой подгруппы предпочтительно являются идентичными и соответствуют последовательности LTR, представляющей или определенной как спейсерная последовательность асимметричного участка-мишени. Однако в дополнительном варианте осуществления спейсерные последовательности могут содержать одну или две девиации последовательности, возникшие в результате нуклеотидных замен.
Обычно, эта стадия представляет собой первое разделение последовательностей асимметричного участка-мишени, выбранных для адаптации специфической рекомбиназы (см. фигуру 1 WO 2008/083931, которая полностью включена в настоящее описание посредством ссылки, и Фиг. 2 WO 2011/147590 A2, которая также полностью включена в настоящее описание посредством ссылки). На этой стадии генерированы последовательности, содержащие симметричные участки-мишени, полученные из половин участков асимметричного участка-мишени, выбранного для адаптации специфической рекомбиназы. Вследствие этого, каждая мутация (т.е. отличие от участка-мишени, распознаваемого первоначальной (дикого типа) рекомбиназой(ами)), представленными в одной половине участка указанного асимметричного участка-мишени, теперь была распределена между симметричными последовательностями -мишенями в первой подгруппе.
На стадии (e) способа по изобретению вторая подгруппа мишеневых нуклеиновых кислот, содержащая модифицированные последовательности-мишени, генерирована на основании последовательностей-мишеней в первой подгруппе стадии (d). В последовательностях, основанных на субучастке 1 в последовательности левой половины участка, участок нуклеотидов, отличающихся от последовательности соответствующей гомологичной половины участка по меньшей мере одного известного участка-мишени стадии (a), замещен природными нуклеотидами, обнаруженными в указанном известном участке-мишени, при условии, что последовательность указанной половины участка содержит один, два или три (предпочтительно, два) нуклеотида, отличающихся от указанного известного участка-мишени, при этом правая половина участка указанной модифицированной последовательности-мишени образована инвертированным повтором указанной модифицированной последовательности левой половины участка, которая отделена от указанной модифицированной последовательности левой половины участка спейсерной последовательностью асимметричной последовательности-мишени.
В последовательностях, основанных на субучастке 2 в последовательности правой половины участка, участок нуклеотидов, отличающийся от последовательности соответствующей гомологичной половины участка по меньшей мере одним известным участком-мишенью стадии (a), замещен природными нуклеотидами, обнаруженными в указанном известном участке-мишени, при условии, что последовательность указанной половины участка содержит один, два или три (предпочтительно, два) нуклеотида, отличающихся от указанного известного участка-мишени, при этом левая половина участка указанной модифицированной последовательности-мишени образована инвертированным повтором указанной последовательности модифицированной правой половины участка, которая отделена от указанной последовательности модифицированной правой половины участка спейсерной последовательностью асимметричной последовательности-мишени.
Например, если один субучасток содержит шесть отличающихся нуклеотидов, такие как оба субучастка, основанные на SEQ ID NO:1, или субучасток 2 SEQ ID NO:2 по отношению к библиотеке рекомбиназ, показанной на фиг. 1 WO 2011/147590 A2, то три модифицированные последовательности-мишени могут быть генерированы на основании субучастка, каждая из которых содержит два (различных) отличающихся нуклеотида в левой половине участка (если она основана на субучастке 1) или правой половине участка (если она основана на субучастке 2). Следовательно, в каждой модифицированной последовательности-мишени, последовательность соответствующего субучастка модифицирована для соответствия последовательности известной последовательности-мишени (или по меньшей мере одной известной последовательности-мишени) в четырех нуклеотидах (фиг. 2 WO 2011/147590 A2). Несомненно, также возможно генерировать шесть модифицированных последовательностей-мишеней, каждая из которых содержит один из отличающихся нуклеотидов, или две последовательности-мишени, каждая из которых содержит три из отличающихся нуклеотидов.
В другом примере если один субучасток содержит девять отличающихся нуклеотидов, таких как субучасток 1 SEQ ID NO:2 по отношению к библиотеке рекомбиназ, показанной на фиг. 1 WO 2011/147590 A2, могут быть генерированы три модифицированные последовательности-мишени на основании субучастка, каждая из которых содержит три (различных) отличающихся нуклеотида в половине участка.
Вследствие этого во всех модифицированных последовательностях половины участка, происходящих от одной последовательности-мишени первой подгруппы стадии (d), в совокупности могут быть обнаружены все отличающиеся нуклеотиды могут, тогда как ни одна из указанных модифицированных последовательностей половины участка не содержит отдельно все отличающиеся нуклеотиды.
Инвертированный повтор снова генерирован на основании модифицированной последовательности половины участка так, что спейсерная последовательность разделяет обе последовательности, образующие инвертированный повтор (см. фиг. 2 WO 2011/147590 A2). Спейсерные последовательности внутри каждой из модифицированных последовательностей-мишеней новой подгруппы, являющиеся полученными из последовательности-мишени подгруппы более высокого порядка, предпочтительно являются идентичными и соответствуют последовательности LTR, представляющей собой или определенной как спейсерная последовательность асимметричного участка-мишени. Однако в дополнительном варианте осуществления спейсерные последовательности могут содержать одну или две девиации последовательности, происходящие от нуклеотидных замен. С применением этого подхода количество мутаций (т.е. отличия с участком-мишенью, распознаваемым рекомбиназой дикого типа) в мишеневых последовательностях, представляющих каждую подгруппу, бывает меньше чем в исходной асимметричной последовательности-мишени, но все мутации все еще представлены в одной из последовательностей-мишеней (см. фигуру 1 WO 2008/083931, фиг. 2 WO 2011/147590 A2).
Термин "отличающийся нуклеотид", как применено в данном документе, относится к нуклеотиду внутри асимметричной последовательности-мишени, идентифицированному или определенному внутри LTR или внутри последовательности-мишени подгруппы, генерированной в соответствии с настоящим изобретением, который отличается (т.е. отличен) от нуклеотида, представленного в таком же положении в соответствующей гомологичной последовательности известной гомологичной симметричной последовательности-мишени известной рекомбиназы, выбранной на стадии (a) способа настоящего изобретения. В этом контексте термины "отличающиеся нуклеотиды" и "мутации" применяются взаимозаменяемо.
WO 2008/083931 описывает, что рекомбиназы могут быть адаптированы с применением направленной молекулярной эволюции с применением последовательностей-мишеней в качестве субстрата, если последовательность-мишень, применяемая в качестве субстрата, отличается по более чем на 3 нуклеотида от природной последовательности-мишени. Таким образом, генерирование подгрупп других порядков, описанное выше, служит для снижения количества отличающихся нуклеотидов в расчете на последовательность-мишень до 3 или менее (см. фигуру 1 WO 2008/083931). Поэтапное снижение количества отличающихся нуклеотидов, в конечном счете, приводит к количеству подгрупп последовательностей-мишеней других порядков с уменьшением количеств отличающихся нуклеотидов при условии, что создана конечная подгруппа, которая может быть применена в качестве субстрата для направленной молекулярной эволюции. При создании других подгрупп, посредством этого снижая количество отличающихся нуклеотидов, отличия c участком-мишенью, распознаваемым рекомбиназой дикого типа, бывают распределены между несколькими последовательностями-мишенями, каждая из которых не содержит более чем 3 этих отличающихся нуклеотида, тогда как в последовательности-мишени конечного порядка в совокупности все еще представлены все отличающиеся нуклеотиды.
Необязательно, в способе по изобретению могут быть генерированы дополнительные подгруппы последовательностей-мишеней исходя из последовательностей-мишеней второй подгруппы посредством поэтапного повторения процесса стадии (e), т.е. разделением последовательностей-мишеней на соответствующие последовательности половины участка и генерированием новых палиндромных структур на основании этих последовательностей половины участка после изменения последовательности половины участка, полученного из последовательности-мишени второй подгруппы, каждый раз генерируя новую подгруппу последовательностей-мишеней, при этом последовательности половины участка, применяемые для генерирования инвертированных повторов содержат меньше нуклеотидов, отличаются от последовательности соответствующей гомологичной половины участка по меньшей мере одним известным участком-мишенью. Эти дополнительные последовательности-мишени могут быть применены для дополнительных стадий направленной молекулярной эволюции и перестановки библиотек рекомбиназ. Несомненно, такая дополнительная стадия может также быть выполнена только для некоторых последовательностей, напр., для последовательностей, в которых получены рекомбиназы с низкой эффективностью рекомбинации. Если генерированы дополнительные подгруппы и на них получены рекомбиназы, то полученную библиотеку рекомбиназ примененяют на стадии (f) способа по изобретению.
Исходя из второй подгруппы последовательностей-мишеней, полученных на стадии (e), третья подгруппа может быть генерирована, с последующими четвертой, пятой, шестой и т.д. подгруппами, если необходимо. Однако генерирование третьей подгруппы обычно бывает необходимо только, если последовательности-мишени второй подгруппы все еще содержат более чем три отличающихся нуклеотида. То же самое относится к генерированию следующих подгрупп, которые бывают необходимы только, если последовательности-мишени предшествующей подгруппы все еще содержат более чем три отличающихся нуклеотида. Необходимо отметить, что в одном варианте осуществления подгруппы последовательностей-мишеней будут генерированы при условии, что последовательности-мишени конечной подгруппы содержат только один отличающийся нуклеотид. Соответственно, в зависимости от количества отличающихся нуклеотидов в каждой последовательности половины участка, может различаться количество подгрупп, генерированное для каждой последовательности половины участка асимметричного участка-мишени,. Может, например, быть необходимо генерирование только двух подгрупп для последовательности левой половины участка, тогда как должны быть генерированы три или четыре подгруппы для правой половины участка для того, чтобы распределить отличающиеся нуклеотиды между несколькими последовательностями-мишенями так, чтобы ни одна последовательность-мишень не содержала более чем 3 этих отличающихся нуклеотида.
Принцип генерирования дополнительных подгрупп последовательностей-мишеней для снижения количества отличающихся нуклеотидов до количеств ниже трех проиллюстрирован на фигуре 1 WO 2008/083931, и фиг. 2 WO 2011/147590 A2 относится к конкретному примеру модифицированных последовательностей-мишеней.
На стадии (f) способ направленной молекулярной эволюции применен для по меньшей мере одной рекомбиназы, распознающей известный гомологичный участок-мишень стадии (a), с применением последовательности-мишени конечной или второй подгруппы, полученной на стадии (e), содержащей один, два или три нуклеотида, отличающихся от последовательности соответствующей гомологичной половины участка указанного известного гомологичного участка-мишени, в качестве субстрата.
Термин "конечная подгруппа", как применено в данном документе, относится к последней подгруппе, генерированной на стадии (e), т.е., если не бывает генерировано дополнительных подгрупп на вторую подгруппу. В зависимости от количества отличающихся нуклеотидов в асимметричном участке-мишени и количества подгрупп, которые должны были быть генерированы, для снижения количества отличающихся нуклеотидов в расчете на последовательность-мишень после ниже 3, "конечная подгруппа" может соответствовать любой подгруппе, например, второй, третьей, четвертой или последней подгруппе, и может отличаться по последовательностями половины участка асимметричной последовательности-мишени внутри LTR. Если ранее были получены рекомбиназы на дополнительные подгруппы модифицированных последовательностей-мишеней, имеющих меньше нуклеотидов, отличающихся от последовательности соответствующей гомологичной половины участка указанного известного гомологичного участка-мишени, то применяют рекомбиназу, полученную на этой стадии.
Несомненно, возможно начать процесс изобретения со специфической рекомбиназой в отношении специфической модифицированной последовательности-мишени, и с другой рекомбиназой (или библиотекой) в отношении другой специфической модифицированной последовательности-мишени. В данной области известны способы направленной молекулярной эволюции, также называемые лабораторная эволюция или эволюция in vitro, (для обзора см. в YUAN et al, 2005 и ссылки в ней; JOHANNES & ZHAO, 2006).
В первой стадии направленной молекулярной эволюции бывают генерированы библиотеки случайным образом мутированных рекомбиназных последовательностей посредством способов, известных в данной области, напр., посредством применения ПЦР пониженной точности и перестановки ДНК (обзор в, напр., YUAN et al., 2005), или способов, раскрытых в международной заявке на патент WO 2002/44409. Плазмиды каждой библиотеки, содержащие мутированную рекомбиназу, также содержат одну из последовательностей-мишеней конечной подгруппы, полученной на стадии (f). После трансфекции генерированной плазмидной библиотеки в соответствующие клетки, бывает позволена экспрессия рекомбиназы, и молекулярная направленная эволюция бывает осуществлена так, как известно специалисту в данной области.
В предпочтительном варианте осуществления молекулярная направленная эволюция, используемая на стадии (f) способа по настоящему изобретению, представляет собой эволюцию связанных с субстратом белков (SLiPE; Buchholz & Stewart, 2001; международная заявка на патент WO 02/44409). Эволюция связанных с субстратом белков может быть осуществлена, как описано подробно в примерах WO 2008/083931 или WO 2011/147590 A2. В кратком изложении, последовательности-мишени, полученные на стадии (e), клонируют в плазмиду (так называемый эволюционный вектор) вместе со случайным образом мутированной кодирующей последовательностью для рекомбиназы. Случайная мутация бывает внесена посредством ПЦР пониженной точности (см. BUCHHOLZ & STEWART, 2001). Затем генерированная плазмидная библиотека бывает трансфицирована в клетки E. coli для позволения экспрессии рекомбиназы. Посредством применения индуцируемого промотора для направления экспрессии рекомбиназы, возможно регулировать уровни экспрессии. После инкубации в течение ночи плазмидную ДНК выделяют из клеток и расщепляют с NdeI для разрезания плазмид, которые не были рекомбинированы и затем только рекомбинированные плазмиды бывают амплифицированы с праймерами. Продукт ПЦР рекомбинированной формы плазмиды дает полосу 1,7 т.п.о. Продукт ПЦР бывает расщеплен с BsrGI и XbaI и субклонирован обратно в схожим образом расщепленный эволюционный вектор для следующего цикла эволюции.
На стадии (g) библиотеки рекомбиназ, полученные на стадии (f), комбинируют и подвергают перестановкам. Технология перестановки ДНК известна в данной области (для обзора см. MINSHULL & STEMMER, 1999; STEMMER, 1994). Библиотеки рекомбиназ, полученные на модифицированные последовательности-мишени, основанные на субучастке 1, комбинируют и подвергают перестановкам, и, отдельно, комбинируют и подвергают перестановкам библиотеки рекомбиназ, полученные на модифицированные последовательности-мишени, основанные на субучастке 2. Затем комбинированные библиотеки и библиотеки с перестановками клонируют в новое поколение векторов, содержащих последовательности-мишени следующей подгруппы более высокого порядка, т.е., если генерированы две подгруппы, то подгруппа генерирована на стадии (d). Например, вектор для библиотеки, полученный на последовательности, основанный на субучастке 1, содержит последовательность субучастка 1 в качестве последовательности-мишени, и вектор для библиотеки, полученный на последовательности, основанный на субучастке 2, содержит последовательность субучастка 2 в качестве последовательности-мишени.
На стадии (h) способ направленной молекулярной эволюции применяют в отношении библиотек с перестановками, полученных на стадии (g), с применением последовательности-мишени следующей подгруппы более высокого порядка, что, как было обсуждено, может являться подгруппами в соответствии со стадией (d). На этой стадии, такой же способ направленной молекулярной эволюции, как способ применяемый ранее на стадии (f), может быть применен, но также возможно применять другой способ направленной молекулярной эволюции на этой стадии способа настоящего изобретения. Примеры других способов направленной молекулярной эволюции описаны, например, в YUAN et al. (2005). Предпочтительно, способ эволюции связанных с субстратом белков также применяют в отношении комбинированных и библиотек с перестановками.
Эта стадия приводит к рекомбиназам, распознающим и рекомбинирующим последовательности-мишени, несущие комбинацию (и, таким образом, возрастающие количества) мутаций из других последовательностей-мишеней подгруппы более низкого порядка. Комбинация мутаций из других библиотек более низкой подгруппы последовательностей-мишеней приводит в результате к синергическим эффектам и приводит к генерированию рекомбиназ, которые теперь рекомбинируют последовательности-мишени подгруппы более высокого порядка, демонстрируя, что эволюциионная стратегия, проходящая через промежуточные субстраты, может быть применена для достижения требуемой активности.
На стадии (i), повторяют стадии (g), т.е. комбинирование и перестановка библиотек рекомбиназ, и (j), т.е. использование направленной молекулярной эволюции на комбинированные и перетасованные библиотеки, при условии что получают по меньшей мере одну рекомбиназу, которая является активной в отношении асимметричной последовательности-мишени, представленной в LTR провирусной ДНК.
В способе, в котором было необходимо генерирование двух подгрупп последовательностей-мишеней для генерирования последовательности-мишени только с одной, двумя или тремя нуклеотидными девиациями, полученные библиотеки рекомбиназ, например, измененные для второй подгруппы последовательностей-мишеней, комбинируют и осуществляют перестановки и применяют молекулярную направленную эволюцию в отношении этой библиотеки с перестановками с применением последовательностей-мишеней первой подгруппы. На следующей стадии асимметричную последовательность-мишень стадии (a) внутри LTR провирусной ДНК применяют для изменения библиотеки рекомбиназ, содержащей рекомбиназы, распознающие последовательности-мишени первой подгруппы, посредством направленной молекулярной эволюции для получения по меньшей мере одной рекомбиназы, которая является активной в отношении асимметричной последовательности-мишени внутри LTR ретровирусной ДНК. На этой стадии способ направленной молекулярной эволюции предпочтительно представляет собой способ связанной с субстратом эволюции белка.
ʺПо меньшей мере одна рекомбиназаʺ относится к тому факту, что способ изобретения должен привести к одной или более (отдельных) рекомбиназ, каждая из которых сама по себе является активной в рекомбинировании асимметричной последовательности-мишени. Не подразумевается, что это охватывает некоторые другие рекомбиназы, которые только вместе способны к рекомбинированию асимметричной последовательности-мишени. На самом деле, способ изобретения не должен привести к отбору рекомбиназ, которые должны быть комбинированы с другими отличающимися рекомбиназами для рекомбинирования асимметричной последовательности-мишени, потому что бывает экспрессирована только одна рекомбиназа в расчете на индивидуальную клетку.
Стадии (a)-(j) способа в сущности известны в данной области, в частности из WO 2011/147590.
После стадии (j) библиотеку адаптированных рекомбиназ отбирают негативно для рекомбинации известного симметричного участка-мишени в соответствии со стадией (a), напр., для рекомбинации loxP и/или loxH.
Этот отбор может быть осуществлен посредством по меньшей мере одного цикла одной или более стадий, содержащих специфическую направленную молекулярную эволюцию и перестановку в библиотеке векторов.
С этой целью нуклеиновая кислота, кодирующая по меньшей мере одну адаптированную рекомбиназу, полученная в предшествующих этапах, может быть выделена из вектора, применяемого там, и клонирована в подходящий эволюционный вектор. Указанный вектор позволяет негативный отбор адаптированных рекомбиназ, рекомбинирующих известный участок-мишень в соответствии со стадией (a), напр., для рекомбинации loxP (SEQ ID NO:4) и/или loxH (SEQ ID NO:5). Посредством этого получают библиотеку векторов. Затем выполняют молекулярную направленную эволюцию, предпочтительно, связанную с субстратом эволюцию белков (SLiPE), как известно специалисту в данной области и в соответствии с принципами, описанными выше.
Например, эволюционный вектор может быть сконструирован так, что он содержит как конечную асимметричную последовательность-мишень (напр., SEQ ID NO:1), так и известный участок-мишень (напр., loxP и/или loxH), по два раза каждый, для позволения рекомбинации. Рекомбинация в известном участке-мишени и последующее рестрикционное расщепление приводит к линейному продукту, который не содержит два специфических участка праймера в порядке, позволяющем амплификацию ПЦР продукта. Если совсем не происходит рекомбинации, вектор является линеризированным рестрикционным расщеплением, и не происходит амплификации посредством ПЦР. В противоположность этому, рекомбинация в конечном асимметричном участке-мишени (напр., SEQ ID NO:1) вырезает рестрикционный участок, т.е., вектор не бывает линеризирован рестрикционным расщеплением, и адаптированная рекомбиназа может быть амплифицирована посредством ПЦР. Для генерирования вариабельности применяют ПЦР, которая неотъемлемо вносит ошибки. Эволюция может быть осуществлена в E. coli.
Библиотека, полученная после одного или более, предпочтительно, после приблизительно десяти циклов направленной молекулярной эволюции, может быть подвержена перестановкам. Может быть осуществлен один или более дополнительных циклов направленной молекулярной эволюции и/или перестановки.
Необязательно, негативный отбор для рекомбинации нескольких известных участков-мишеней, напр., для рекомбинации loxP и loxH может быть осуществлен попеременно, напр., один цикл эволюции с негативным отбором на loxP можно чередовать с одним циклом эволюции с негативным отбором на loxH. Например, могут быть осуществлены от приблизительно 10 до приблизительно 30 или приблизительно 15-20 циклов негативного отбора в комбинации с приблизительно двумя циклами перестановки ДНК. Между этими эволюционными циклами, количество активатора транскрипции L-арабинозы может быть варьировано, напр., от 100 мкг/мл до 1 мкг/мл. Предпочтительные векторы и способы показаны на фиг. 2 и примеры.
Специфичность адаптированных рекомбиназ в отношении конечной асимметричной последовательности-мишени и потенциальная остаточная активность в отношении известных последовательностей-мишеней может быть проверена для одного или более клонов после того, как было осуществлено некоторое количество эволюционных циклов.
Если специфичность все еще не является удовлетворительной, то должны быть осуществлены дополнительные циклы эволюции.
Как показано на фиг. 3 негативный отбор удаляет остаточную активность адаптированных рекомбиназ, как описано в WO 2011/147590 A2 (TRE3) в отношении известных последовательностей-мишеней loxP и loxH. Не бывает обнаружено остаточной активности генерированных рекомбиназ по изобретению на известные участки-мишени, напр., loxP и/или loxH, даже при наличии высоких количеств (50 или 100 мкг/мл) активатора транскрипции L-арабинозы, т.е., в присутствии высоких количеств рекомбиназы. Это демонстрирует, что полученные адаптированные рекомбиназы являются высокоспецифическими по отношению к последовательности-мишени, в этом случае по отношению к SEQ ID NO:1, так как специфическая асимметричная последовательность-мишень бывает рекомбинирована, а известная симметричная последовательность-мишень нет.
Это имеет преимущество, заключающееся в том, что когда адаптированная рекомбиназа по изобретению применяется для лечения людей, риск перекрестной реакции и рекомбинации человеческой последовательности в клетке-хозяина является минимальным. Это является одним из факторов, вносящих вклад в переносимость адаптированной рекомбиназы в человеческих клетках. Однако предметом обсуждения являются не только кратковременные последствия экспрессии рекомбиназы, но также вопросы безопасности, такие как возможные онкогенные эффекты неспецифической рекомбинации даже с низкой эффективностью. Таким образом, исключение даже остаточной активности адаптированной рекомбиназы, вносит вклад в безопасность и надежность полученной в результате адаптированной рекомбиназы в терапевтических планах.
За отбором против рекомбинации известных симметричных последовательностей-мишеней следует отбор на адаптированную рекомбиназу, хорошо переносимую в человеческих клетках.
На стадии (n) способа по изобретению нуклеиновую кислоту, кодирующую по меньшей мере одну адаптированную рекомбиназу, полученную на стадии (m), выделяют и клонируют в векторе для экспрессии кодируемой рекомбиназы и селектируемого маркера в эукариотических, предпочтительно, человеческих клетках, посредством этого получая векторную библиотеку. Нуклеиновая кислота может быть выделена из соответствующей плазмиды внутри библиотеки с применением соответствующих рестрикционных ферментов. Способы расщепления рестрикционной эндонуклеазой известны специалисту. Затем нуклеиновая кислота, кодирующая рекомбиназу, может быть извлечена известными способами, такими как, например, гель-электрофорез. Она может быть клонирована в соответствующем векторе экспрессии для экспрессии в эукариотических, напр., человеческих, клетках, как известно в современном уровне техники или описано далее. Например, может быть использован ретровирусный, напр., лентивирусный вектор, напр., как показано на фиг. 4A. Экспрессия кодируемой адаптированной рекомбиназы и селектируемого маркера предпочтительно является конститутивной, или она может быть индуцирована подходящими агентами.
Селектируемый маркер может придавать устойчивость к антибиотику, или он может представлять собой флуоресцентный белок, такой как зеленый флуоресцентный белок (GFP) или его производное (напр., EBFB, ECFP, YFP). Флуоресцентные белки, такие как GFP, позволяют легкую сортировку клеток в зависимости от силы экспрессии.
На стадии (l) эукариотические, предпочтительно, человеческие клетки, предпочтительно, человеческие T-клетки, преобразуют с указанной библиотекой векторов, полученной на стадии (k). Могут быть применены способы, известные в современном уровне техники. Трансформированные клетки обычно являются человеческими клетками, однако, если предполагается лечение не являющегося человеком пациента, рекомендуется тестировать переносимость в клетках, этих видов пациентов. Человеческие клетки предпочтительно являются гемопоэтическими клетками, напр., предпочтительно, T-клетками, в частности T-клетками CD4+, но также могут быть применены стволовые клетки, такие как стволовые клетки CD34+. Первичные клетки, напр., первичные T-клетки, предпочтительно, первичные T-клетки CD4+, могут быть использованы, но также может быть применена a клеточная линия, такая как T-клетки Jurkat.
На стадии (p) клетки, экспрессирующие указанный селектируемый маркер, культивируют в течение промежутка времени, достаточного для отбора рекомбиназ TRE, хорошо переносимых человеческой клеткой. Отбор основан на предположении, что экспрессия маркера и экспрессия адаптированной рекомбиназы являются коррелирующими. Для экспрессии маркера выбраны клетки, напр., выбраны GFP-позитивные клетки, предпочтительно, клетки с сильной экспрессией GFP. Так как экспрессия селектируемого маркера и экспрессия адаптированной рекомбиназы коррелируют, эти клетки также будут экспрессировать адаптированную рекомбиназу. Следовательно, клетки, экспрессирующие адаптированную рекомбиназу, которая является пагубной для их выживаемости или их способности к размножению, бывают элиминированы или их количество снижено. Предпочтительно, клетки, экспрессирующие маркер, культивируют в течение по меньшей мере 1 недели, по меньшей мере двух недель, по меньшей мере 3 недель или по меньшей мере 4 недель. Соответственно, адаптированные рекомбиназы, экспрессированные в T-клетках, будут хорошо переносимыми человеческими клетками, напр., человеческими T-клетками, т.е., они не будут токсичны для указанных клетках, или, предпочтительно, также не будут иным образом пагубными для выживаемости и размножения указанных клеток. Предпочтительно во время этого периода культивирования клетки отобраны по меньшей мере один раз, предпочтительно 2, 3, или 4 раза, для высокой экспрессии селектируемого маркера. Например, для флуоресцентного белка может быть выполнен отбор посредством сортировки клеток с активированной флуоресценцией. С геном устойчивости к антибиотикам к культуральной среде могут быть добавлены возрастающие количества антибиотиков.
Хотя экспрессия рекомбиназы Cre дикого типа в человеческих клетках была разработана довольно давно, и было продемонстрировано, что экспрессия с приемлемыми уровнями не представляет сложностей, сверхэкспрессия Cre может являться токсичной (LOONSTRA et al., 2001). Изобретатели обнаружили, что значительное количество мутированных адаптированных рекомбиназ в соответствии с настоящим изобретение являлось пагубным для выживаемости и/или размножения человеческих T-клеток при сильной сверхэкспрессии. Хотя это можно было ожидать, интересно, что относительная низкая специфичность адаптированных рекомбиназ и остаточная активность в отношении участков-мишеней, таких как loxP и loxH (последовательность, представленная в человеческом геноме) приводит к низкой переносимости в человеческих клетках, отбор на переносимость в человеческих T-клетках отдельно без предшествующего отбора на высокую специфичность не являлся достаточным для элиминирования остаточной перекрестной реактивности в отношении loxP или loxH. Таким образом, только комбинация обоих стадий отбора с ранее известным способом изобретения приводит к адаптированной рекомбиназе, которая является как хорошо переносимой, так и высокоспецифической.
На стадии (q) за стадиями культивирования и отбора следует выделение нуклеиновой кислоты, кодирующей по меньшей мере одну рекомбиназу, из клеток, экспрессирующих указанный селектируемый маркер, полученных на стадии (p).
Стадия (r) является необязательной и добавляет другой отбор в отношении нуклеиновой кислоты, кодирующей рекомбиназу, способную к рекомбинированию асимметричной последовательности-мишени стадии (a), предпочтительно для рекомбинации с высокой активностью. Рекомбинационную активность предпочтительно тестируют в человеческих клетках, в частности, человеческих клетках CD4+ T, но она также может быть тестирована в E. coli.
На стадии (s) из библиотеки выделена нуклеиновая кислота рекомбиназы, имеющая активность на асимметричную последовательность-мишень стадии (a) внутри LTR ретровирусной ДНК. Нуклеиновая кислота может быть выделена из соответствующей плазмиды внутри библиотеки с применением соответствующих рестрикционных ферментов. Способы расщепления рестрикционной эндонуклеазой известны специалисту. Нуклеиновая кислота, кодирующая рекомбиназу, может затем быть регенерирована известными способами, такими как, например, гель-электрофорез.
Нуклеиновая кислота может храниться (предпочтительно при температурах ниже -80°C) или может необязательно быть клонирована на стадии (t) в векторе экспрессии для применения в дополнительном анализе, в способах экспрессии белка или для введения индивиду для лечения и/или предотвращения ретровирусной инфекции, в частности, инфекции ВИЧ и/или СПИДа. Подходящие векторы экспрессии известны в современном уровне техники или раскрыты далее.
Разработка адаптированных рекомбиназ, которые специфически нацелены на асимметричные последовательности-мишени, такие как SEQ ID NO:1, внутри множества LTR ВИЧ-1 позволяет вырезание соответствующего провируса из его хромосомной интеграции для большинства индивидов, инфицированных с ВИЧ-1. Вектор экспрессии, кодирующий такую рекомбиназу, клетки, трансфицированные с ним, и/или рекомбиназный белок, полученный из них, имеют медицинские применения, напр., в лечении и/или предотвращении инфекции ВИЧ-l. Предпочтительные способы приготовления такой адаптированной рекомбиназы или кодирующего ее вектора экспрессии раскрыты в WO 2011/147590. Авторы настоящего открытия добавили к способу, описанному в WO 2011/147590, стадию активного отбора на высокую специфичность, т.е., отсутствие поддающейся обнаружению перекрестной реактивности на известную последовательность-мишень стадии (a) (или на, напр., loxP и loxH), и на переносимость адаптированной рекомбиназы в человеческих клетках, таких как человеческие T-клетки. Как описано, это значительно улучшает медицинское использование адаптированной рекомбиназы для вырезания геномов провируса ВИЧ из человеческих T-клеток.
Провирусная ДНК, которая может быть встроена в геном клетки-хозяина или которая может еще не являться встроенной, предпочтительно представляет собой ДНК ретровируса. Ретровирусы содержат большое и разнообразное семейство РНК вирусов в оболочке. Отличительным свойством семейства является его репликационная стратегия, которая содержит в качестве необходимых стадий обратную транскрипцию вирусной РНК в линейную двухцепочечную ДНК и последующую интеграцию этой ДНК (провирусная ДНК) в геном клетки-хозяина. Ретровирусы делятся семь групп, определенных по эволюционному родству. Пять из этих групп (альфа-, бета-, дельта-, эпсилон- и гамма-ретровирус) представляют ретровирусы с онкогенным потенциалом, и другие две группы представляют собой лентивирусы и спумавирусы. Человеческие патогенные вирусы лейкоза человеческих T-клеток клетка типа I и типа II (HTLV-I и HTLV-II) принадлежат к группе дельта-ретровирусов, тогда как вирусы СПИДа вируса иммунодефицита человека типа 1 и типа 2 (HIV-1 и ВИЧ-2) принадлежат к группе лентивирусов (обзор см. в стандартном руководство "Retroviruses" COFFIN JM, HUGHES SH, VAR US HE (Eds.) 1997, Cold Spring Harbor Laboratory Press, New York).
В одном варианте осуществления провирусная ДНК, которая может быть встроена в геном клетки-хозяина, представляет собой ДНК ретровируса, выбранную из группы, состоящей из вируса мышиной опухоли молочной железы (MMTV), обезьяньего вируса Mason pfizer (MPMV), человеческого вируса Т-клеточного лейкоза типа I (HTLV-I), человеческого вируса Т-клеточного лейкоза типа II (HTLV-II), обезьяньего вируса Т-клеточного лейкоза типа I (STLV-I), обезьяньего вируса Т-клеточного лейкоза типа II (STLV-II), вируса коровьего лейкоза (BLV), вируса лейкоза кошек (FeLV) и вируса мышиного лейкоза Молони (MoMLV).
В другом варианте осуществления ретровирус представляет собой лентивирус, выбранный из группы, состоящей из вируса иммунодефицита человека типа 1 (HIV-1), вируса иммунодефицита человека типа 2 (HIV-2), обезьяньего вируса иммунодефицита (SIV), кошачьего вируса иммунодефицита (FIV), бычьего вируса иммунодефицита (BIV), вируса Maedi-visna (MVV), вируса инфекционной анемии лошадей (EIAV) и вируса артроэнцефалита козлов (CAEV). Как указано выше, ВИЧ, в частности ВИЧ-1, является предпочтительным.
Асимметричная последовательность-мишень, идентифицированная на стадии (a), способа по настоящему изобретению расположена как в 5'-LTR, так и в 3'-LTR провируса ВИЧ. Предпочтительно, указанная асимметричная последовательность-мишень, расположенная как в 5'-LTR, так и в 3'-LTR провируса ВИЧ, имеет последовательность, представленную далее как SEQ ID NO:1 или SEQ ID NO:2.
В предпочтительном варианте осуществления способ направленной молекулярной эволюции, примененный в способе по настоящему изобретению, представляет собой способ эволюции связанных с субстратом белков (SLiPE; BUCHHOLZ & STEWART, 2001; см. также WO 02/44409).
Посредством осуществления способа изобретения, как описано в настоящем описании, изобретатели генерировали несколько нуклеиновых кислот, кодирующих хорошо переносимую адаптированную рекомбиназу, и сами адаптированные рекомбиназы. Изобретение, таким образом, относится к хорошо переносимой адаптированной рекомбиназе, содержащей последовательность в соответствии с любой из SEQ ID NO:9-13 или составляющей их, или нуклеиновым кислотам, кодирующим их.
Неожиданно было обнаружено, что эти адаптированные рекомбиназы отличаются от консенсусных последовательностей SEQ ID NO:7 и 8 адаптированных рекомбиназ, способных к рекомбинированию асимметричных последовательностей-мишеней, как описано в WO 2011/147590, и от всех других ранее известных рекомбиназ.
В частности, новые анализируемые хорошо переносимые адаптированные рекомбиназы, способные к рекомбинированию асимметричной последовательности-мишени SEQ ID NO: 1 с высокой специфичностью, неожиданно содержат мутацию Q89L.
В одном варианте осуществления адаптированные рекомбиназы по настоящему изобретению содержат последовательность в соответствии с SEQ ID NO:9 (Tre 3.1 консенсусная последовательность 85%). SEQ ID NO:9 представляет консенсусную последовательность, с каждой мутацией (по сравнению с аминокислотной последовательностью Cre), представленной с вероятностью 85%. Для этого определения 100 индивидуальных клонов, генерированных способом настоящего изобретения, анализировали секвенированием по Сэнгеру, а также всю генерированную библиотеку Tre 3.1 секвенированием нового поколения (33000 считывания уникальной последовательности из 200 п.о.). Вариабельные аминокислоты представлены посредством X, что может означать любую встречающуюся в природе аминокислоту (см. фиг. 1). Приблизительно треть аминокислот SEQ ID NO:9 является высоковариабельной, т.е., нет потребности сохранять эти положения для того, чтобы рекомбиназа была способна как к рекомбинированию асимметричной последовательности-мишени SEQ ID NO: 1, так и была хорошо переносима людьми. С другой стороны приблизительно две трети аминокислотных положений, по-видимому, являлись важными для рекомбинирования асимметричной последовательности-мишени SEQ ID NO: 1 с высокой специфичностью и/или для хорошей переносимости людьми.
В предпочтительном варианте осуществления адаптированная рекомбиназа по настоящему изобретению содержит последовательность в соответствии с любой из SEQ ID NO:11-13, наиболее предпочтительно SEQ ID NO:11. Эти адаптированные рекомбиназы были выбраны по их специфичности, т.е., хотя другие рекомбиназы, генерированные в соответствии с изобретением, все еще имели низкую, но поддающуюся обнаружению рекомбинационную активность в отношении loxP, loxH или lox LTR 1.0, такая рекомбинационная активность не была обнаруживаема с рекомбиназами SEQ ID NO: 11-13, как показано в примерах. Изобретение, таким образом, относится к хорошо переносимой и высокоспецифической адаптированной рекомбиназе, способной к рекомбинированию асимметричных последовательностей-мишеней внутри LTR провирусной ДНК множества штаммов ретровирусов, которые могут быть встроены в геном клетки-хозяина (т.е., функциональная адаптированная рекомбиназа), которая предпочтительно содержит SEQ ID NO:11-13, предпочтительно, SEQ ID NO:11. Такую адаптированную рекомбиназу получают, напр., в соответствии со способом по изобретению.
В одном варианте осуществления адаптированная рекомбиназа может содержать последовательность в соответствии с SEQ ID NO:10. Эта последовательность, Tre 3.1 консенсусная последовательность 100% (3 клона), является консенсусной последовательностью трех предпочтительных рекомбиназ SEQ ID NO:11-13.
Адаптированная рекомбиназа может также иметь по меньшей мере 95% аминокислотной идентичности, предпочтительно, по меньшей мере 99% аминокислотной идентичности или 100% аминокислотной идентичности с SEQ ID NO: 10, или она может отличаться от SEQ ID NO: 10 только по одной или двум аминокислотам, и она содержит последующие определенные аминокислотные замены по сравнению с последовательностью Cre (SEQ ID NO: 6): V7L, P12S, P15L, M30V, H40R, M44V, S51T, Y77H, K86N, Q89L, G93A, S108G, C155G, A175S, A249V, R259D, E262R, T268A, D278G, P307A, N317T, I320S. Она также может содержать замены: N160T, R241Q, K244I, N319E. Предпочтительно, она содержит последующие аминокислотные замены по сравнению с Cre: N3I, V7L, N10S, P12S, P15L, V23A, M30V, F31L, H40R, M44V, S51T, Y77H, K86N, Q89L, G93A, S102F, S108G, N111D, K122R, A131T, S147A, D153E, C155G, N160T, F163L, I166V, I174V, A175S, V182I, G198S, D232S, R241Q, K244I, A249V, Q255R, R259D, A260V, E262R, G263K, T268A, D278G, P307A, N317T, N319E, I320S.
Эти специфические замены делают фермент особенно подходящим для рекомбинации в последовательность-мишень SEQ ID NO: 1 или последовательность-мишень, имеющую высокую идентичность последовательности с SEQ ID NO: 1 (напр., по меньшей мере 80%, по меньшей мере 90% или по меньшей мере 95% идентичности последовательности с SEQ ID NO: 1). Неожиданно изобретателям удалось продемонстрировать, что одна аминокислотная вариация, а именно, Q89L, обеспечивает то, что адаптированная рекомбиназа бывает как хорошо переносима в человеческих клетках, напр., человеческие гемопоэтические клетки или человеческие T-клетки, так и имеет высокую специфичность, так как она не имеет поддающуюся обнаружению активность в рекомбинировании первоначальной последовательности-мишени, loxP или loxH, и, предпочтительно, также не имеет поддающуюся обнаружению активность на loxLTR Tre 1. Активность может быть детектирована посредством гель-электрофореза образца, содержащего loxP (SEQ ID NO:4), loxH (SEQ ID NO:5), или для сравнения loxLTR Tre 3, содержащего SEQ ID NO:1, каждый из которых был приведен в контакт с адаптированной рекомбиназой, напр., посредством индукции ее экспрессии из подходящего вектора, как показано в примерах и на Фиг. 3. Как можно увидеть на указанной фигуре, рекомбиназа Tre 3.0 (полученная в соответствии с WO 2011/147590), несмотря на то, что она является довольно специфической по сравнению с другими рекомбиназами, имеет остаточную активность в отношении loxP и loxH при показанных условиях, тогда как с uTre, рекомбиназа настоящего изобретения, рекомбинированный продукт может быть виден только для loxLTR, содержащей SEQ ID NO: 1. Эта высокая специфичность минимизирует риск нежелательной рекомбинации в человеческий геном.
Последовательность адаптированной рекомбиназы изобретения не раскрыта в WO 2008/083931 или WO 2011/147590. В частности, предшествующий уровень техники не описывает или предполагает, что адаптированная рекомбиназа, способная к рекомбинированию асимметричной последовательности-мишени SEQ ID NO: 1, имеет аминокислотную замену Q89L. В противоположность этому WO 2011/147590 подробно описывает, что это положение должно быть сохранено как Q (см. все специфические последовательности или консенсусные последовательности указанной публикации). Последовательность адаптированной рекомбиназы изобретения также отличается от встречающихся в природе рекомбиназ, таких как Cre, Dre, Fre или Zre, что является очевидным исходя из того свойства, что она способна к рекомбинированию асимметричных последовательностей-мишеней, предпочтительно SEQ ID NO: 1, внутри LTR провирусной ДНК множества штаммов ретровирусов, которые могут быть встроены в геном клетки-хозяина.
Если адаптированная рекомбиназа, способная к рекомбинированию асимметричных последовательностей-мишеней внутри LTR провирусной ДНК множества штаммов ретровирусов, встроенной в геном клетки-хозяина, должна рекомбинировать последовательность-мишень SEQ ID NO: 1, то она предпочтительно содержит консенсусные последовательности Tre 3.1 85%, SEQ ID NO: 9, или Tre 3.1 консенсусную последовательность 100%, SEQ ID NO: 10, или одну из специфических последовательностей SEQ ID NO: 11-13.
Функциональные адаптированные рекомбиназы, способные к рекомбинированию асимметричных последовательностей-мишеней внутри LTR провирусной ДНК множества штаммов ретровирусов, которые могут быть встроены в геном клетки-хозяина, которые могут, напр., быть получены посредством способа по изобретению, могут отличаться от указанных последовательностей, но последовательности представляют ценные указания для специалиста в данной области для получения адаптированной рекомбиназы, способной к рекомбинированию асимметричных мишеневых сторон, таких как SEQ ID NO: 1, даже без осуществления способа по изобретению.
Предпочтительно, аминокислотные замены в референсной последовательности представляют собой консервативные замены, которые хорошо известны специалисту в данной области (напр., Creighton (1984), Proteins. W. H. Freeman and Company (Ed.)). Например, консервативные замены замещают одну аминокислоту из группы негативно заряженных аминокислот другой аминокислотой. Наиболее предпочтительно замены приводят к одной аминокислоте, представленной в любой из SEQ ID NO:11-13 в соответствующем положении.
Адаптированные рекомбиназы, способные к рекомбинированию асимметричных последовательностей-мишеней внутри LTR провирусной ДНК множества штаммов ретровирусов, которые могут быть встроены в геном клетки-хозяина, могут также содержать комбинацию 2 или более последовательностей, выбранных из группы, состоящей из SEQ ID NO:11-13, напр., C-концевая часть любой этих последовательностей, напр., SEQ ID NO:11, и N-концевая часть любой другой из этих последовательностей, напр., SEQ ID NO:12. C-концевая часть может иметь длину 1-342 аминокислот. В комбинации из двух последовательностей N-концевая часть может иметь длину из 1-342 аминокислот. Адаптированная рекомбиназа может представлять собой комбинацию из трех или более частей, полученных из этих последовательностей. Комбинация содержит консенсусный мотив TRE 3.1, напр., SEQ ID NO:10, или предпочтительно, SEQ ID NO:9.
Изобретение также относится к нуклеиновой кислоте, кодирующей адаптированную рекомбиназу, способную к рекомбинированию асимметричных последовательностей-мишеней, таких как SEQ ID NO: 1 внутри LTR провирусной ДНК множества штаммов ретровирусов, которые могут быть встроены в геном клетки-хозяина, при этом адаптированная рекомбиназа содержит аминокислотную последовательность, как определено выше.
В контексте изобретения нуклеиновая кислота или белок, содержащий последовательность, могут состоять из указанной последовательности.
Она может альтернативно содержать дополнительные последовательности, напр., сигнальную последовательность, обеспечивающую экспрессию/локализацию в специфическом клеточном компартменте, такую как сигнал ядерной локализациии, как в SEQ Id NO:14 (сигнал ядерной локализации представлен в положениях 2-9 SEQ ID NO:14) a. Если белок должен быть использован в фармацевтической композиции, предпочтительно чтобы он был экспрессирован в виде слитого белка с доменом белковой трансдукции, таким как домен белковой трансдукции tat, который позволяет белковую трансдукцию мишеневых клеток. Предпочтительно адаптированная рекомбиназа изобретения, которая должна быть применена в фармацевтической композиции, получена в виде белка слияния с последовательностью ядерной локализацией и с доменом белковой трансдукции, напр. из tat, и нуклеиновая кислота, кодирующая адаптированную рекомбиназу изобретения, может кодировать такой слитый белок. Например, последующие домены белковой трансдукции могут быть применены в слитом белке с адаптированной рекомбиназой по изобретению, которая предпочтительно дополнительный содержит сигнал ядерной локализации:
- Основной домен трансактиватора Tat ВИЧ-1 (Fawell S, Seery J, Daikh Y, Moore C, Chen LL, Pepinsky B, Barsoum J., Tat-mediated delivery of heterologous proteins into cells., Proc Natl Acad Sci U S A. 1994 Jan 18;91(2):664-8.)
- Гомеодомен Drosohila antennapedia (Antp) Derossi D, Joliot AH, Chassaing G, Pro-chiantz A., The third helix of the Antennapedia homeodomain translocates through biological membranes., J Biol Chem. 1994 Apr 8;269(14): 10444-50.)
- Фактор транскрипции HSV VP22 (Elliott G, O'Hare P., Intercellular trafficking and protein delivery by a herpesvirus structural protein., Cell. 1997 Jan 24;88(2):223-33.) - Транслокационный проникающий в клетку мотив (TLM) поверхностного антигена PreS2 вируса гепатита B (HBV) (Oess S, Hildt E., Novel cell permeable motif derived from the PreS2-domain of hepatitis-B virus surface antigens., Gene Ther. 2000 May;7(9):750-8.).
В случае если белок должен быть очищен, к белку также может быть добавлена облегчающая очистку метка, такая как гистидиновая метка.
Использование кодонов нуклеиновой кислоты изобретения, кодирующей рекомбиназу Tre, как определено выше, может быть выбрано специалистом. Например, может быть выбрано использование кодонов, подходящее для экспрессии в человеческой клетке, в частности, если предусмотрена экспрессия в человеческой клетке, напр., в терапевтических целях. Использование кодонов может также быть основано на использовании кодонов, напр., рекомбиназы Cre.
Адаптированная рекомбиназа или нуклеиновая кислота, кодирующая указанную адаптированную рекомбиназу, может быть получена посредством способа по изобретению, как описано в настоящем описании, или она может быть получена этим способом. Она также может быть получена, на основании последовательностей, раскрытых в настоящем описании, необязательно, посредством комбинирования и/или дополнительного варьирования этих последовательностей, необязательно тестируя на активность в рекомбинации асимметричных участков-мишеней, таких как SEQ ID NO:1.
Изобретение дополнительно относится к композиции, напр., библиотеке, содержащей две или более нуклеиновых кислот, кодирующих адаптированную рекомбиназу, как определено выше, напр., кодирующих две или более адаптированных рекомбиназ, содержащих другие последовательности в соответствии с SEQ ID NO:9, предпочтительно, в соответствии с SEQ ID NO:10 или любую из SEQ ID NO:11-13. В одном варианте осуществления композиция содержит нуклеиновые кислоты, кодирующие адаптированные рекомбиназы, содержащие две или более, три или более, четыре или более, пять или более, десять или более, 20 или более или 25 или более рекомбиназ, содержащих последовательности в соответствии с любой из SEQ ID NO:9-13 или комбинации этих последовательностей. Такие композиции, в частности композиции, в которых нуклеиновая кислота представляет собой вектор экспрессии, могут являться особенно подходящими в качестве фармацевтических композиций, как описано далее.
В способе настоящего изобретения нуклеиновая кислота, кодирующая адаптированную рекомбиназу, которая является активной в отношении асимметричной последовательности-мишени внутри LTR ретровирусной ДНК, предпочтительно бывает клонирована в векторе экспрессии. Векторы экспрессии представляют собой генетические конструкции для экспрессии белков, кодируемых нуклеиновыми кислотами внутри вектора. Такие векторы экспрессии могут являться или самореплицирующимися экстрахромосомными векторами или векторами, которые интегрируются в геном хозяина. Обычно, эти векторы экспрессии содержат транскрипционную и трансляционную регулятивную нуклеиновую кислоту, функционально связанную с нуклеиновой кислотой, кодирующей адаптированную рекомбиназу настоящего изобретения.
Термин "контрольные последовательности" относится к последовательностям ДНК, необходимым для экспрессии функционально связанной кодирующей последовательности в конкретном организме хозяина. Контрольные последовательности, которые являются подходящими для прокариотов, например, содержат промотор, необязательно операторную последовательность и связывающий рибосому участок. Известно, что эукариотические клетки используют промоторы, сигналы полиаденилирования и энхансеры.
Нуклеиновая кислота является "функционально связанной", когда она приведена в функциональную взаимосвязь с другой последовательностью нуклеиновой кислоты. Например, промотор или энхансер является функционально связанным с кодирующей последовательностью, если он воздействует на транскрипцию последовательностей; или связывающий рибосому участок функционально связан с кодирующей последовательностью, если он расположен так, что это облегчает трансляцию. Связывание выполнено лигированием в подходящие рестрикционные участки. Если такие участки не представлены, то применены синтетические олигонуклеотидные адапторы или линкеры в соответствии с общепринятой практикой. Транскрипционная и трансляционная регулятивная нуклеиновая кислота будет обычно соответствовать клетке-хозяина, применяемой для экспрессии адаптированной рекомбиназы. Многочисленные типы соответствующих экспрессионных векторов и подходящие регулятивные последовательности известны в данной области для множества клеток-хозяев.
Вектор экспрессии, применяемый в настоящем изобретении, может представлять собой ретровирусный вектор, лентивирусный вектор, спумавирусный вектор, аденовирусный вектор, или адено-ассоциированный вирусный вектор. Однако в предпочтительном варианте осуществления вектор экспрессии представляет собой лентивирусный вектор, выбранный из группы, состоящей из полученных из лентивирусных векторов ВИЧ-1, SIV, FIV или EIAV. Лентивирусные векторы описаны, например, в SCHAMBACH et al. (2006) или в европейской патентной заявке No. 1 1000751.5.
В предпочтительных вариантах осуществления по настоящему изобретению вектор экспрессии содержит клеточный, бактериальный, вирусный или гибридный промотор.
В целом, в целях настоящего изобретения промотор может являться конститутивным или индуцируемым промотором. Дополнительно, промоторы могут или представлять собой встречающийся в природе промотор, такой как бактериальный, клеточный или вирусный промотор, или гибридный промотор. Гибридные промоторы, которые комбинируют элементы более чем одного промотора, известны в данной области, и являются пригодными в настоящем изобретении. Дополнительно, промотор, применяемый в настоящем изобретении, может также представлять собой производное встречающегося в природе промотора. "Производное" встречающегося в природе промотора, как применено в данном документе, может представлять собой комбинацию цис-активных элементов, полученных из промоторов или последовательностей другого происхождения или, альтернативно, может быть получено посредством делеции или мутации цис-активных элементов внутри специфического встречающегося в природе промотора (EDELMAN et al, 2000; ALPER et al, 2006; HARTENBACH & FUSSENEGGER, 2006).
В одном варианте осуществления настоящего изобретения конститутивный промотор или его производное выбран или получен из группы, состоящей из промоторов цитомегаловируса, вируса саркомы Рауса, связанных с вирусом лейкемии мышей ретровирусов, гена фосфоглицерокиназы, мышиного вируса некроза селезенки или человеческого элонгационного фактора 1 альфа.
В дополнительном варианте осуществления настоящего изобретения индуцируемый промотор или его производное выбран или получен из группы, состоящей из LTR или ее производных, полученных из лентивирусв, спумавирусов и дельтаретровирусов.
В этом контексте термин "LTR" относится к как 5', так и к 3' длинным концевым повторам провируса, имеющим промоторную функцию (обзор см. в стандартном руководстве "Retroviruses" (COFFIN JM, HUGHES SH, VARMUS HE (Eds.) 1997, Cold Spring Harbor Laboratory Press, New York)).
Предпочтительно индуцируемый промотор или его производное выбран или получен из LTR или его производное получено из ВИЧ-1, ВИЧ-2, MVV, EIAV, CAEV, SIV, FIV, BIV, HTLV- I и HTLV-II.
Настоящее изобретение дополнительно относится к способу плучения адаптированной рекомбиназы, при этом указанный способ содержит вышеупомянутый способ получения вектора экспрессии, кодирующего адаптированную рекомбиназу, и дополнительной стадии экспрессии указанной адаптированной рекомбиназы (или слитого полипептида, содержащего аминокислотную последовательность указанной адаптированной рекомбиназы) из указанного вектора экспрессии в подходящей клетке-хозяина.
Предпочтительно полученные в конечном счете рекомбиназы тестируют в клетках млекопитающих для обеспечения того, что они функционируют в среде клетки млекопитающих. Дополнительно для получения хорошей экспрессии в клетках млекопитающих рекомбиназы могут быть оптимизированы для экспрессии в этих клетках (напр. оптимизация использования кодонов с применением хорошо известных в данной области способов, см., например, SHIMSHE et al, 2002), или могут быть добавлены сигнальные последовательности, необходимые для направление белка в ядро клетки млекопитающих, такие как последовательность NLS (MACARA, 2001), к нуклеиновой кислоте адаптированной рекомбиназы. Экспрессия нуклеиновой кислоты, кодирующей адаптированную рекомбиназу, клонированную в векторе экспрессии, напр., в соответствии со стадией (1) способа приготовления вектора экспрессии, кодирующего адаптированную рекомбиназу, может быть осуществлена с применением, например, системы экспрессии бактерий, насекомых или млекопитающих. Однако также могут быть применены другие известные в данной области системы экспрессии. Способы введения экзогенной нуклеиновой кислоты млекопитающим, насекомым или бактериальным хозяевам, а также другим хозяевам, также известны в данной области и будут варьироваться с используемой клеткой-хозяина. Технологии содержат опосредованную декстраном трансфекцию, осаждение фосфатом кальция, опосредованную полибреном трансфекцию, слияние протопластов, электропорацию, вирусную инфекцию, инкапсуляцию полинуклеотида(ов) в липосомы и прямую микроинъекцию ДНК в ядра.
Слитые белки приготовлены посредством хорошо известных в данной области способов. Например, вектор экспрессии, в который уже клонирована нуклеиновая кислота, кодирующая адаптированную рекомбиназу, содержит нуклеиновую последовательность, кодирующую второй полипептид или белок. Посредством клонирования нуклеиновой кислоты, кодирующей адаптированную рекомбиназу в рамке с последовательностью второго полипептида или белка, обе последовательности будут экспрессированы в виде слитого белка.
Клетки-хозяева, применяемые для экспрессии адаптированной рекомбиназы из вектора экспрессии, содержат прокариотические клетки, такие как, например, бактериальные клетки или дрожжевые клетки, или, предпочтительно, эукариотические клетки, такие как, например, клетки насекомых или млекопитающих, наиболее предпочтительно, человеческие клетки. Клетка-хозяина может являться гемопоэтической клеткой, напр., зрелая гемопоэтическая стволовая клетка или T-клетка, напр., клетка CD4+. Клетка может быть получена у индивида, инфицированного с ретровирусом, и клетка может быть введена обратно индивиду после трансформации, и, необязательно, культивирования и/или размножения.
Настоящее изобретение дополнительно относится к способу приготовления трансформированной зрелой стволовой клетки, при этом указанный способ содержит вышеупомянутый способ приготовления вектора экспрессии, кодирующего адаптированную рекомбиназу и дополнительную стадию введения вектора экспрессии, полученного в вышеупомянутом способе приготовления вектора экспрессии, кодирующего адаптированную рекомбиназу in vitro в подходящую зрелую стволовую клетку.
В дополнительном аспекте настоящее изобретение относится к нуклеиновой кислоте, как раскрыто в настоящем описании, и/или полученной вышеупомянутым способом настоящего изобретения. Настоящее описание также относится к нуклеиновым кислотам, кодирующим адаптированную рекомбиназу, определенную последовательностью.
"Нуклеиновая кислота", как применено в данном документе, является полимерным соединением, составленным из ковалентно связанных субъединиц, называемых нуклеотиды. Нуклеиновая кислота содержит полирибонуклеиновую кислоту (РНК), напр., мРНК, и полидезоксирибонуклеиновую кислоту (ДНК), обе из которых могут являться одноцепочечными или двухцепочечными. ДНК содержит кДНК, геномную ДНК, синтетическую ДНК и полусинтетическую ДНК.
В дополнительном аспекте настоящее изобретение также относится к вектору экспрессии, полученному вышеупомянутым способом настоящего изобретения, и к вектору экспрессии, содержащему нуклеиновую кислоту, кодирующую адаптированную рекомбиназу, как определено в настоящем описании.
Термин "белок", как применено в данном документе, содержит белки, полипептиды и пептиды. Специалисту в данной области следует иметь в виду, что последовательности нуклеиновых кислот по изобретению могут быть применены для генерирования белковых последовательностей. Дополнительным объектом изобретения является белок адаптированная рекомбиназа, получаемый, напр., вышеупомянутым способом настоящего изобретения, где рекомбиназа может необязательно являться слитым белком, содержащим функциональную рекомбиназу. В одном варианте осуществления белок адаптированная рекомбиназа может быть приготовлен в виде слитого полипептида с применением хорошо известных в данной области технологий. В предпочтительном варианте осуществления белок адаптированная рекомбиназа связан со вторым полипептидом. Предпочтительно слитый полипептид получен вышеупомянутым способом настоящего изобретения, при этом адаптированная рекомбиназа связана со вторым полипептидом.
В одном варианте осуществления белок адаптированная рекомбиназа приготовлен в виде слитого полипептида для увеличения экспрессии. В дополнительном варианте осуществления, белок адаптированная рекомбиназа получен в виде слитого полипептида для позволения введения полипептида в живые клетки. Как правило, очищенные белки не могут проникнуть в клетки, так как они не способны проходить через клеточную мембрану из-за их размера. Однако слияние специфических последовательностей пептидов с белками может привести в результате к поглощению этих слитых белков в клетки. Затем белок в клетке выполняет свою функцию. Участок специфический рекомбиназы, включая рекомбиназу Cre, был успешно доставлен в клетки с этим подходом (PEITZ et al., 2002). Проникающие в клетки рекомбиназы дополнительно были описаны NOLDEN et al. (2006) и LIN et al. (2004). Следовательно, эта стратегия может быть применена для доставки адаптированных рекомбиназ в клетки для удаления провируса из инфицированных клеток. Таким образом, второй полипептид в слитом полипептиде может содержать сигнальный пептид. Сигнальный пептид может представлять собой домен белковой трансдукции, такой как пептид TAT или пептид из третьей спирали гомеодомена Antennapedia (DEROSSI et al, 1994, 1996; VIVES et al, 1997; VIVES, 2003; RICHARD et al, 2005) или NLS (последовательность ядерной локализации) для доставки слитого полипептида в ядро эукариотической клетки (MACARA, 2001).
Дополнительный аспект настоящего изобретения относится к зрелой стволовой клетке, получаемой вышеупомянутым способом приготовления трансформированной зрелой стволовой клетки настоящего изобретения. Стволовые клетки предпочтительно бывают инфицированы или трансфицированы вектором экспрессии в соответствии с изобретением. В предпочтительном варианте осуществления зрелая стволовая клетка является стволовой клеткой из гемопоэтической линии, экспрессирующей адаптированную рекомбиназу, вышеупомянутый слитый полипептид или содержащей вышеупомянутый вектор экспрессии. Гемопоэтические стволовые клетки (HSC) являются полученными из костного мозга клетками CD34+, которые, напр., могут быть очищены из G-CSF-мобилизованной периферической крови доноров (напр., ВИЧ-инфицированных пациентов) посредством рутинного лейкафереза (SCHERR & EDER, 2002). Затем генетически модифицированные in vitro клетки могут быть составлены для реинфузии пациентам.
В современном уровне техники термин "стволовые клетки" обозначает клетки, которые (a) имеют способность к самообновлению и (b) способность образовывать по меньшей мере один и часто несколько специализированных клеточных типов из-за их способности к асимметричному делению (DONOVAN & GEARHART, 2001). Зрелые стволовые клетки могут быть выделены из различных тканей взрослого, т.е. из дифференцированных индивидуумов. Такие стволовые клетки в современном уровне техники называются "мультипотентные зрелые стволовые клетки". Основное различие между эмбриональными плюрипотентными стволовыми клетками и зрелыми мультипотентными стволовыми клетками заключается в количестве дифференцированных тканей, которое может быть получено из соответствующих клеток.
В дополнительном варианте осуществления вектор экспрессии по настоящему изобретению применен для трансформирования T-клеток, напр., первичных клеток CD4+ (клетки крови) ретровируса (напр., ВИЧ)-инфицированных пациентов.
Альтернативно, адаптированная рекомбиназа изобретения может быть составлена для доставки вирусоподобными частицами (VLP). VLP могут быть применены для упаковки мРНК Tre, белка Tre, напр., слитого белка, или ДНК, напр., экспрессирующих ДНК Tre плазмид или конструкций, содержащей промотор-кДНК Tre-участок polyA. Соответственно, нуклеиновая кислота изобретения может дополнительно содержать сигнал упаковки.
В дополнительной стадии способа настоящего изобретения нуклеиновая кислота по изобретению, вектор экспрессии, содержащий последовательность нуклеиновой кислоты, кодирующую адаптированную рекомбиназу по изобретению, белок рекомбиназа, слитый белок или зрелая стволовая клетка, полученные способами по настоящему изобретению, составлены в фармацевтическую композицию для применения в предотвращении и/или лечении ретровирусной инфекции и/или для снижения вирусной нагрузки у индивида, инфицированного ретровирусом, напр., ВИЧ, в частности, ВИЧ-1. Дополнительной задачей изобретения является фармацевтическая композиция, полученная вышеупомянутым способом. Фармацевтическая композиция предпочтительно представлена в форме раствора, подходящего для внутривенного применения (вливания).
Фармацевтический состав может дополнительно содержать один или более фармацевтически приемлемых носителей, наполнителей и/или адъювантов. В данной области известны носители, наполнители и адъюванты, подходящие для применения в фармацевтической композиции.
Фармацевтическая композиция по настоящему изобретению предпочтительно снижает вирусную нагрузку у индивида, инфицированного ретровирусом до ниже 5000 геномных эквивалентов/мл плазмы, предпочтительно до ниже 500 геномных эквивалентов/мл плазмы и более предпочтительно до ниже 50 геномных эквивалентов/мл плазмы при введении индивиду. Таким образом, фармацевтическая композиция по настоящему изобретению, содержащая вектор экспрессии, кодирующий адаптированную рекомбиназу (или адаптированную рекомбиназу в виде белка или слитого полипептида, или стволовую клетку, содержащую вектор экспрессии), способна к снижению вирусной нагрузки у индивида, инфицированного ретровирусом, посредством удаления генетического резервуара ретровирусов внутри клеток-хозяев, посредством этого предотвращая дополнительные циклы жизни вируса.
Термин "вирусная нагрузка", как применено в данном документе, относится, напр., к эквивалентам РНК ВИЧ (т.е. геномам), которые связаны с 1 мл плазмы пациента (DYBUL et al, 2002). Таким образом, вирусная нагрузка определена измерением содержания вирусной ДНК в образце, взятом у пациента. В настоящее время существуют три основных типа анализа вирусной нагрузки:
1) полимеразная цепная реакция с обратной транскрипцией РНК ВИЧ (ОТ-ПЦР): Amplicor (TM) HIV-1 Monitor Test; Roche Diagnostics
2) ответвление цепи ДНК (bDNA): Versant(TM) HIV RNA Assay; Bayer Diagnostics; и
3) основанная на последовательности нуклеиновой кислоты амплификация (NASBA): NucliSens (TM) Assay; bioMerieux.
В предпочтительном варианте осуществления фармацевтическая композиция по настоящему изобретению способна к снижению вирусной нагрузки у индивида, инфицированного ретровирусом до ниже 5000 геномных эквивалентов/мл плазмы, предпочтительно ниже 500 геномных эквивалентов/мл плазмы и более предпочтительно ниже 50 геномных эквивалентов/мл плазмы. Пациент с вирусной нагрузкой ниже 5000 геномных эквивалентов/мл плазмы считается являющися относительно хорошо адаптированным к медицинскому лечению. Однако цель существующей в настоящее время терапии против СПИДа заключается в снижении вирусной нагрузки ниже предела обнаружения анализов вирусной нагрузки, который настоящее время составляет ниже чем приблизительно 50 геномных эквивалентов/мл плазмы. Фармацевтическая композиция предпочтительно снижает вирусную нагрузку ретровирусов выбранных из группы, состоящей из вируса Мышиной опухоли молочной железы (MMTV), обезьяньего вируса Мэйзена-Пфейзера (MPMV), человеческого вируса Т-клеточного лейкоза типа I (HTLV-I), человеческого вируса Т-клеточного лейкоза типа II (HTLV-II), обезьяньего вируса Т-клеточного лейкоза типа I (STLV-I), обезьяньего вируса Т-клеточного лейкоза типа II (STLV-II), вируса коровьего лейкоза (BLV), вируса лейкоза кошек (FeLV) и вируса мышиного лейкоза Молони (MoMLV). В еще одном дополнительном предпочтительном варианте осуществления ретровирусом для лечения фармацевтическим средством по настоящему изобретению является лентивирус. Указанный лентивирус предпочтительно выбран из группы, состоящей из вируса иммунодефицита человека типа 1 (HIV-1), вируса иммунодефицита человека типа 2 (HIV-2), обезьяньего вируса иммунодефицита (SIV), кошачьего вируса иммунодефицита (FIV), бычьего вируса иммунодефицита (BIV), вируса Maedi-visna (MVV), вируса инфекционной анемии лошадей (EIAV) и вируса артроэнцефалита козлов (CAEV). Наиболее предпочтительно, ретровирус представляет собой ВИЧ, в частности ВИЧ-1. Однако, специалисту данной области очевидно, что настоящее изобретение также применимо к ретровирусным инфекциям ретровирусами, отличными от указанных выше.
Индивид, инфицированный ретровирусом, которому должна быть введена фармацевтическая композиция, выбран из группы, состоящей из людей, приматов, обезьян, крупного рогатого скота, лошадей, коз, овец и домашних кошек. Однако предпочтительно индивид является человеком.
В целом, индивиду должно быть введено эффективное количество вектора экспрессии, адаптированной рекомбиназы или трансформированных клеток. Введение может являться, напр., внутривенным или внутримышечным введением.
В одном варианте осуществления фармацевтическая композиция составлена для сопутствующего введения с другими активными агентами высокоактивной противоретровирусной лечения (ВААРТ). Высокоактивная противоретровирусная терапия ВААРТ является комбинированной терапией, воздействующей на вирусную обратную транскриптазу, протеазу и слияние (GULIC et al, 1997; LALEZARI et al, 2003).
В другом варианте осуществления фармацевтическая композиция составлена для введения параллельно с или после терапии, направленной на общую активацию иммунной системы или специфическую активацию экспрессии генов провируса. Предпосылкой направленной на общую активацию иммунной системы лечения является предположение, что преднамеренная активация латентно инфицированных ВИЧ клеток может ускорить уничтожение персистирующих вирусных резервуаров. Уничтожение происходит посредством иммунной элиминации посредством программируемой смерти клеток, активно экспрессирующих продукты ВИЧ-1 (проапоптотические) (KULKOSKY & BRAY, 2006). Общая активация иммунной системы (активация иммунных клеток, включая покоящиеся клетки) обычно достигается посредством, например, введения иммунотоксинов, цитокинов (напр., IL-2) или активирующих Т-клетки антител (напр., OKT3).
Ввиду того факта, что активация иммунной системы, проводимая для преднамеренного активирования устойчивых к ВААРТ латентных резервуаров, к сожалению не смогла полностью элиминировать ВИЧ-1, и возобновления связанных с вирусом симптомов (для обзора см. KULOSKY & BRAY 2006; MARCELLO, 2006; SHEHU-XHILAGA et al, 2005) из-за того факта, что общая активация T-клеток предположительно также индуцирует вирусную репликацию и увеличивает количество потенциальных клеток-мишеней ВИЧ-1 до уровня выше, чем может быть сдерживать ВААРТ (FRASER et al, 2000), для лечения ВИЧ необходимы дополнительные способы специфического лечения. Один из подходов заключается в активации транскрипции неактивных в иных случаях вирусных геномов. Специфическая активация экспрессии гена латентного провируса может быть достигнута посредством введения форболового эфира простратина или человеческого цитокина IL-7, оба из которых, по-видимому, реактивируют латентный ВИЧ-1 в отсутствии клеточной пролиферации (MARCELLO, 2006). Кроме того, селективная транскрипционная активность ВИЧ-1 также может быть достигнута посредством ингибиторов гистоновой деацетилазы (HDACl) таких как, например, вальпроевая кислота, что в конечном счете индуцирует почкование ВИЧ-1 из покоящихся клеток в отсутствии клеточной активации (MARCELLO, 2006; LEHRMAN et al, 2005).
Однако лечение с общей активацией иммунной системы или специфической активацией экспрессии генов провирусов или подобные стратегии лечения значительно улучшаются от одновременного удаления провирусной ДНК, посредством этого снижая у пациента пул инфицированных клеток.
Настоящее изобретение также относится к способу лечения и/или предотвращения ретровирусной инфекции, в частности инфекции ВИЧ, у индивида. В одном варианте осуществления анализируют последовательность ретровируса, инфицирующего индивид, в образце, полученном у индивида, и по меньшей мере один вектор экспрессии, кодирующий адаптированную рекомбиназу, по меньшей мере одну адаптированную рекомбиназу, или по меньшей мере одна клетка, трансформированная с указанным вектором экспрессии, напр., одна зрелая стволовая клетка, должна быть введена индивиду, если провирусная ДНК индивида содержит асимметричную последовательность-мишень, идентифицированную на стадии (a), на которую была отобрана рекомбиназа. Образец, полученный у индивида, может являться образцом крови, напр., содержащим инфицированные клетки CD4+.
Краткое описание фигур
Фиг. 1: Фиг. 1 относится к выравниванию белковых последовательностей
(a) рекомбиназы Cre SEQ ID NO:6;
(b) общей консенсусной последовательности Tre, SEQ ID NO:7, консенсусной последовательности рекомбиназы Tre, специфической в отношении асимметричных участков-мишеней внутри LTR ВИЧ-1 в соответствии с WO 2011/147590;
(c) консенсусной последовательности рекомбиназы Tre 3.0, SEQ ID NO:8, рекомбиназы Tre являются специфичными в отношении SEQ ID NO:1 в соответствии с WO 2011/147590;
(d) консенсусная последовательность 85% рекомбиназы Tre 3.1, SEQ ID NO:9, индивидуальные мутации в консенсусной последовательности по сравнению с последовательностью Cre представлены в 85% всех клонов, генерированных посредством способа по настоящему изобретению и анализированных секвенированием с высокой пропускной способностью (33000 считывания уникальной последовательности из 200 п.о.);
(e) консенсусной последовательности рекомбиназы Tre 3.1 100%, SEQ ID NO:10, консенсусной последовательности трех Tre рекомбиназ 3.1, отобранных на высокую специфичность в отношении SEQ ID NO:1 (отсутствие активности на loxP, loxH или loxLTR Tre 1.0 (SEQ ID NOs: 3-5) и переносимость людьми; и
(f) ииллюстративной рекомбиназы Tre 3.1 uTre, SEQ ID NO:11, высокоспецифической в отношении SEQ ID NO:1 и хорошо переносимой людьми. Адаптированная рекомбиназа в соответствии с этой последовательностью обозначена uTre, соответственно ʺуниверсальная Treʺ.
Выделенные полужирным шрифтом буквы указывают консервативные аминокислоты, вариабельные и неустановленные положения обозначены X. Положения Cre, мутированные в SEQ ID NO: 9. 10 и/или 11, подчеркнуты в последовательности Cre, и положение мутации представлено над последовательностями. Замена Q89L, обнаруженная только в Tre 3.1, выделена курсивом.
Фиг. 2 демонстрирует иллюстративный эволюционный вектор для эволюционного отбора против рекомбинации на loxP. Соответствующие векторы для отбора против рекомбинации на loxH легко могут быть сконструированы. loxLTR содержит SEQ ID NO: 1. Циклы эволюции 51-69 Tre3 выполняли с pEVOloxLTR-loxP и pEVOloxLTR-loxH, попеременно, 100-1 мкг/мл активатора транскрипции L-ara, включая два цикла перестановки ДНК).
Фиг. 3 демонстрирует высокую специфичность uTre в сравнении с Tre3. Tre3: Клон выделяли из библиотеки Tre3.0, цикл 43. uTre: Клон выделяли из Tre3.1 библиотеки, цикл 71. Рекомбинированный продукт обозначен одним треугольником, нерекомбинированный двумя треугольниками. При условиях, когда адаптированная рекомбиназа экспрессирована (индукция L-арабинозозой), uTre рекомбинирует loxLTR, содержащую SEQ ID NO: 1 в E. coli. В противоположность этому, Tre3 рекомбинирует loxLTR и loxP и loxH, т.е., она имеет относительно ослабленную специфичность.
Фиг. 4 демонстрирует иллюстративный лентивирусный вектор экспрессии для конститутивной экспрессии селектируемого маркера, EGFP, и библиотеки tre в человеческих клетках. Фиг. 4B демонстрирует блок-схему для клеточного скринирования на хорошо переносимую высокоспецифическую Tre. Конечное скринирование на высокоактивную Tre подтверждает активность в человеческих клетках. Одиночные клоны рекомбиназ были выбраны и подвержены дополнительным анализам.
Фиг. 5 демонстрирует антивирусную активность uTre в тканевой культуре в двух иллюстративных культурах. T-клетки PM1 трансдуцировали векторами, кодирующими отдельно GFP (контроль, незаштрихованные кружки) или кодирующими uTre и GFP (uTre, заштрихованные квадраты). Впоследствии культуры инфицировали с ВИЧ-1 и наблюдали за вирусной нагрузкой в течение некоторого времени с применением ELISA с антигеном p24. Эксперимент демонстрирует, что адаптированная рекомбиназа является эффективной в снижении вирусной нагрузки. После нескольких недель (8 или 9 недель) вирусную нагрузку больше невозможно обнаружить посредством ELISA с антигеном p24.
Фиг. 6 Показана выраженная антивирусная активность uTre в первичных человеческих клетках CD4+, полученных от инфицированного ВИЧ пациента. Клетки трансдуцировали вектором, экспрессирующим отдельно GFP (контрольный эксперимент; левая панель), или вектором, экспрессирующим uTre и GFP (uTre; правая панель). За репликацией вируса наблюдали посредством высвобождения антигена p24 ВИЧ-1 (незаштрихованные кружки), и за процентом трансдуцированных (GFP+) человеческих клеток CD4+ (заштрихованные квадраты) наблюдали посредством FACS. Экспрессия uTre привела в результате к выраженному антивирусному эффекту и защите клеток CD4+. Стоит отметить, что снижение вирусной нагрузки между днем 15 и днем 20 в контрольном эксперименте отражает клеточную смерть из-за неингибируемой репликации вируса.
Фиг. 7 демонстрирует антивирусную активность uTre в ВИЧ-инфицированных гуманизированных мышах. Иммунодефицитным мышам трансплантировали человеческие гемопоэтические стволовые клетки CD34+/HSC (контроль) или экспрессирующие uTre CD34+ HSC (Animal#1 и #2). Впоследствии животных инфицировали ВИЧ-1 и наблюдали за вирусной нагрузкой (детектированной как РНК ВИЧ-1 копии/мл; незаштрихованные кружки) и процентом человеческих клеток CD45+CD4+ из всех лимфоцитов (заштрихованные квадраты) в течение некоторого времени.
Примеры
Пример 1
Использовали материалы и способы, как описано WO 2008/083931, WO 2011/147590 и BUCHHOLZ & STEWART, 2001, если особо не указано иное. Адаптированные рекомбиназы, способные к рекомбинированию асимметричных последовательностей-мишеней во множестве других штаммов ВИЧ-1, получали, как описано в WO 2011/147590. Полученные в результате библиотеки tre применяли в дальнейших экспериментах.
Пример 2
Для усиления специфичности uTre выполняли дополнительные циклы эволюции отбора против рекомбинационной активности на loxP и loxH. С этой целью созданную библиотеку Tre, полученную из цикла эволюции 50, клонировали в эволюционном векторе, содержащем два участка loxLTR (SEQ ID NO: 1), расположенных переплетено с двумя участками loxP или двумя участками loxH, соответственно. Иллюстративный вектор показан на фиг. 2. При индукции экспрессии рекомбиназы рекомбинация на loxLTR привела в результате к удалению только представленного участка NdeI, тогда как рекомбинация на loxP или loxH не привела. Плазмидную ДНК, выделенную после каждого цикла эволюции, расщепляли с NdeI, и кодирующие рекомбиназу последовательности, которые содержали успешно рекомбинированную loxLTR, а не loxP или loxH, амплифицировали посредством ПЦР и субклонировали обратно в эволюционном векторе для следующего цикла эволюции. В общем, выполняли 19 дополнительных циклов эволюции Tre3, включая два цикла перестановки, отбирая попеременно против рекомбинации в отношении loxP и loxH.
Пример 3
Для скринирования на рекомбиназы uTre со значительно уменьшенной клеточной токсичностью (т.е. цитопатические свойства) библиотеки tre лигировали в лентивирусный вектор, который конститутивно экспрессирует EGFP из внутреннего промотора SFFV LTR и библиотеку tre из конститутивного промотора EF1 альфа (Фиг. 4A). Трансдукция клеток Jurkat T позволяла последовательную сортировку (посредством FACS) высоко экспрессирующих GFP клеток и последующее выделение нетоксичных клонов uTre (Фиг. 4B). Для этого клеточные сортировки в отношении трансдуцированных T-клеточных культур выполняли в день 3, день 10 и день 24 после трансдукции с увеличенной строгостью в отношении экспрессии EGFP. После еще одной недели культивирования оставшуюся библиотеку tre выделяли и выбранные клоны анализировали с точки зрения усиленной активности Tre.
Пример 4
Для анализа активности uTre в клеточных линиях культуры T-клеток PM-1 трансдуцировали с полученными из ASLV ретровирусными векторами, экспрессирующими или uTre и GFP, или отдельно GFP (негативный контрольный вектор). Стоит отметить, что экспрессия GFP позволяла отслеживание трансдуцированных клеток. На 10 день после трансдукции клетки инфицировали с HIV-1Bal. За эффектом экспрессии uTre на репликацию ВИЧ-1 наблюдали посредством еженедельных измерений посредством ELISA количества вирусного антигена p24 в культуральных супернатантах. Как показано (фигура 5), высвобождение p24 снижалось значительно в трансдуцированных uTre культурах, тогда как оно оставалось стабильным или даже возрастало в контрольных культурах (экспрессирование GFP отдельно).
Пример 5
Анализ активности uTre в первичных клетках CD4+, полученных от ВИЧ-1-инфицированного пациента. Клетки CD4+ стимулировали с магнитными шариками CD3/CD28 в течение 48 ч. Впоследствии клетки трансдуцировали лентивирусными векторами, экспрессирующими или GFP отдельно (в качестве негативного контроля), или экспрессирующими uTre вместе с GFP. Клетки культивировали в присутствии 100 МЕ IL2 в течение 20 дней. Наблюдали за вирусными нагрузками (измеренные посредством ELISA с антигеном p24) и за количеством человеческих трансдуцированных клеток CD4+ (анализировали посредством FACS) в указанные дни после трансдукции. Как показано на фигуре 6, экспрессия uTre приводила в результате к выраженному антивирусному эффекту (показано посредством незаштрихованных кружков) и защите клеток CD4+ (показано посредством заштрихованных квадратов). В противоположность этому снижение вирусной нагрузки между днем 15 и днем 20 в контрольном эксперименте отражало смерть клеток из-за неингибируемой вирусной репликации.
Пример 6
Анализ активности uTre in vivo. Иммунодефицитным мышам NOG (NOD.Cg-PrkdcscidIL2rgtm1Wjl/SzJ) трансплантировали человеческие гемопоэтические стволовые клетки CD34+/HSC (контроль), или экспрессирующие uTre CD34+ HSC. Впоследствии животных инфицировали с HIV-1Bal и наблюдали за вирусной нагрузкой (детектированной посредством сверхчувствительного основанного на ПЦР анализа) и процентом человеческих клеток CD45+ CD4+ (анализированным посредством FACS) в течение некоторого времени. Как показано (фигура 7) экспрессия uTre привела в результате к значительным антивирусным активностям in vivo.
Список ссылок
Abremski K, Hoess RH, Sternberg N (1983) "Studies on the properties of PI site-specific recombination: evidence for topologically unlinked products following recombination." Cell 32, 1301-1311.
Abremski K, Hoess R (1983) "Bacteriophage PI site-specific recombination. Purification and properties of the Cre recombinase protein." J Biol. Chem. 259, 1509-1514.
Adachi A, Gendelman HE, Koenig S, Folks T, Willey R, Rabson A, Martin MA (1986) "Production of acquired immunodeficiency syndrome-associated retrovirus in human and nonhu- man cells transfected with an infectious molecular clone." J Virol. 59, 284-291.
Alper H, Fischer C, Nevoigt E, Stephanopoulos G (2006) "Tuning genetic control through promoter engineering" Proc. Natl. Acad. Sci. USA 102, 12678-12683.
Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang, Z., Miller, W. and Lipman, D.J. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic acids research, 25, 3389-3402.
Beyer WR, Westphal M, Ostertag W, von Laer D (2002)"Oncoretrovirus and lentivirus vectors pseudotyped with lymphocytic choriomeningitis virus glycoprotein: generation, concentration and broad host range." J Virol. 76, 1488-1495.
Blackard JT, Renjifo BR, Mwakagile D, Montano MA, Fawzi WW, Essex M (1999) "Transmission of human immunodeficiency type 1 viruses with intersubtype recombinant long terminal repeat sequences." Virology 254, 220-225.
Bloom JD, Meyer MM, Meinhold P, Otey CR, MacMillan D, Arnold FH (2005) "Evolving strategies for enzyme engineering." Curr. Opin. Struct. Biol. 15, 447-452.
Buchholz F, Ringrose L, Angrand PO, Rossi F, Stewart AF (1996) "Different thermostabilities of FLP and Cre recombinases: implications for applied site-specific recombination." Nucl. Acids Res. 24, 4256-4262.
Buchholz F, Angrand PO, Stewart AF (1998) "Improved properties of FLP recombinase evolved by cycling mutagenesis." Nat. Biotechnol. 16, 657-662.
Buchholz F, Stewart AF (2001) "Alteration of Cre recombinase site specificity by substrate- linked protein evolution." Nat. Biotechnol. 19, 1047-1052.
Chiu YL, Soros VB, Kreisberg JF, Stopak K, Yonemoto W, Greene WC (2005) "Cellular APOBEC3G restricts HIV-1 infection in resting CD4+ T cells." Nature 435, 108-114
Chun T-W, Engel D, Berrey MM, Shea T, Corey L, Fauci AS (1998) "Early establishment of a pool of latently infected, resting CD4+ T cells during primary HIV-1 infection." Proc. Natl. Acad. Sci. USA 95, 8869-8873.
Coates CJ, Kaminski JM, Summers JB, Segal DJ, Miller AD, Kolb AF (2005) "Site-directed genome modification: derivatives of DNA-modifying enzymes as targeting tools." Trends Biotechnol. 23, 407-419.
Collins CH, Yokobayashi Y, Umeno D, Arnold FH, (2003) "Engineering proteins that bind, move, make and break DNA." Curr. Opin. Biotechnol. 14, 665.
Combes P, Till R, Bee S, Smith MC (2002) "The streptomyces genome contains multiple pseudo-attB sites for the (phi)C31 -encoded site-specific recombination system." J Bacteriol. 184, 5746-5752.
Crameri A, Raillard SA, Bermudez E, Stemmer WP (1998) "DNA shuffling of a family of genes from diverse species accelerates directed evolution." Nature 391, 288-291. Derossi D, Joliot AH, Chassaing G, Prochiantz A (1994) "The thrid helix of the Antennapedia homeodomain translocates through biological membranes." J Biol Chem. 269, 10444-10450.
Derossi D, Calvet S, Trembleau A, Chassaing G, Prochiantz A (1996) "Cell internalization of the third helix of the Antennapedia homeodomain is receptor-independent." J Biol Chem 271, 18188-18193.
Donovan, P.J., Gearhart, J. (2001) "The end of the beginning for pluripotent stem cells." Nature 414, 92-97.
Donzella GA, Schols D, Lin SW, Este JA, Nagashima KA, Maddon PJ, Allaway GP, Sakmar TP, Henson G, De Clercq E, Moore JP (1998) "AMD3100, a small molecule inhibitor of HIV-1 entry via the CXCR4 co-receptor." Nature Medicine 4, 72-77.
Dybul M, Fauci AS, Bartlett JG, Kaplan JE, Pau AK (2002) "Guidelines for using antiretrovi- ral agents among HIV-infected adults and adolescents." Annals of Internal Medicine 137, 381-433.
Eddy, S.R. (1998) Profile hidden Markov models. Bioinformatics (Oxford, England), 14, 755- 763.
Edelman GM, Meech R, Owens GC, Jones FS (2000) "Synthetic promoter elements obtained by nucleotide sequence variation and selection for activity." Proc. Natl. Acad. Sci. USA 97, 3038-3043.
Elliott G, O'Hare P., Intercellular trafficking and protein delivery by a herpesvirus structural protein., Cell. 1997 Jan 24;88(2):223-33
Emerman M, Malim MH (1998) "HIV-l regulatory/accessory genes: keys to unraveling viral and host cell biology." Science 280, 1880-1884.
Fawell S, Seery J, Daikh Y, Moore C, Chen LL, Pepinsky B, Barsoum J., Tat-mediated delivery of heterologous proteins into cells., Proc Natl Acad Sci U S A. 1994 Jan 18;91(2):664-8.
Finzi D, Hemankova M, Pierson T, Carruth LM, Buck C, Chaisson RE, Quinn TC, Chadwick K, Margolick J, Brookmeyer R, Gallant J, Markowitz M, Ho DD, Richman DD, Siliciano RF (1997) "Identification of a reservoir for HIV-1 in patients on highly active antiretro viral therapy." Science 278, 1295-1300.
Flowers CC, Woffendin C, Petryniak J, Yang S, Nabel GJ (1997) "Inhibition of recombinant human immunodeficiency virus type 1 replication by a site-specific recombinase." J. Virol. 71, 2685-2692.
Gulick RM, Mellors JW, Havlir D, Eron JJ, Gonzalez C, McMahon D, Richman DD, Valentine FT, Jonas L, Meibohm A, Emini EA, Chodakewitz JA (1997) "Treatment with indinavir, zidovudine, and lamivudine in adults with human immunodeficiency virus infection and prior antiretroviral therapy." N Engl. J. Med. 337, 734-739.
Guzman LM, Belin D, Carson MJ, Beckwith J (1995) "Tight regulation, modulation, and high-level expression by vectors containing the arabinose PBAD promoter." J. Bacteriol. Ill, 4121-4130.
Hartenbach S, Fussenegger M (2006) "A novel synthetic mammalian promoter derived from an internal ribosome entry site." Biotechnology and Bioengineering 95, 547-559.
Hauber I, Bevec D, Heukeshoven J, Kratzer F, Horn F, Choidas A, Harrer T, Hauber J (2005) "Identification of cellular deoxyhypusine synthase as a novel target for antiretroviral therapy." J. Clin. Invest. 115, 76-85.
Hazuda DJ, Young SD, Guare JP, Anthony NJ, Gomez RP, Wai JS, Vacca JP, Handt L, Motzel SL, Klein HJ, Dornadula G, Danovich RM, Witmer MV, Wilson KA, Tussey L, Schleif WA, Gabryelski LS, Jin L, Miller MD, Casimiro DR, Emini EA, Shiver JW (2004) "Integrase inhibitors and cellular immunity suppress retroviral replication in rhesus macaques." Science 305, 528-532.
Hoess RH, Abremski K (1985) " Mechanism of strand cleavage and exchange in the Cre-lox site-specific recombination system." J. Mol. Biol. 181, 351-362. Johannes TW, Zhao H (2006) "Directed evolution of enzymes and biosynthetic pathways." Curr. Opin. Microbiol. 9, 261-267.
Krasnow MA, Cozzarelli NR (1983) "Site-specific relaxation and recombination by the Tn3 resolvase: Recognition of the DNA path between oriented res sites." Cell 32, 1313-1324.
Kulkosky J, Bray S (2006) "HAART-persistent HIV-1 latent reservoirs: their origin, mechanisms of stability and potential strategies for eradication." Curr. HIV Res. 4, 199-208.
Lalezari JP, Henry K, O'Hearn M, Montaner JS, Piliero PJ, Trottier B, Walmsley S, Cohen C, Kuritzkes DR, Eron Jr. JJ, Chung J, DeMasi R, Donatacci L, Drobnes C, Delehanty J, Salgo M (2003) "Enfuvirtide, an HIV-1 fusion inhibitor, for drug-resistant HIV infection in North and South America." N. Engl. J. Med. 348, 2175-2185.
Lee YS, Park JS (1998) "A novel mutant lox? containing part of long terminal repeat of HIV - 1 in spacer region: presentation of possible target site for antiviral strategy using site-specific recombinase." Biochem. Biophys. Res. Comm. 253, 588-593.
Lee YS, Kim ST, Kim GW, Lee M, Park JS (2000) "An engineered lox sequence containing part of a long terminal repeat of HIV-1 permits Cre recombinase-mediated DNA excision." Biochem. Cell Biol. 78, 653-658.
Lehrman G, Hogue IB, Palmer S, Jennings C, Spina CA, Wiegand A, Landay AL, Coombs RW, Richman DD, Mellors JW, Coffin JM, Bosch RJ, Margolis DM (2005) "Depletion of latent HIV-1 infection in vivo: a proof-of-concept study" Lancet 366, 549-555.
Lewandoski, M. (2001) "Conditional control of gene expression in the mouse." Nat. Rev. Genet. 2, 743-755.
Lin Q, Jo D, Gebre-Amlak KD, Ruley HE (2004) "Enhanced cell-permeant Cre protein for site-specific recombination in cultured cells." BMC Biotechnol. 4, 25.
Little SJ, Holte S, Routy JP, Daar ES, Markowitz M, Collier AC, Koup RA, Mellors JW, Connick E, Conway B, Kilby M, Wang L, Whitcomb JM, Hellmann NS, Richman DD (2002) "Antiretroviral-drug resistance among patients recently infected with HIV." N. Engl. J. Med. 347, 385-394.
Loonstra A, Vooijs M, Beverloo HB, Allak BA, van Drunen E, Kanaar R, Berns A, Jonkers J (2001) ʺGrowth inhibition and DNA damage induced by Cre recombinase in mammalian cells.ʺ PNAS 98, 9209-9214.
Macara IG (2001) "Transport into and out of the nucleus." Microbiology and molecular biology reviews 65, 570-594.
Malim MH, Hauber J, Fenrick R, Cullen BR (1988) "Immunodeficiency virus rev trans- activator modulates the expression of the viral regulatory genes." Nature 335, 181-183.
Marcello A (2006) "Latency: the hidden HIV-1 challenge." Retrovirology 3, 7.
Matsumura I, Ellington AD (2001) "In vitro evolution of beta-glucuronidase into a beta- galactosidase proceeds through non-specific intermediates." J. Mol. Biol. 305, 331-339.
Minshull J, Stemmer WP. (1999) "Protein evolution by molecular breeding." Curr. Opin. Chem. Biol. 3, 284-290.
Nagy A (2000) "Cre recombinase: the universal reagent for genome tailoring." Genesis 26, 99-109.
Needleman SB, Wunsch CD (1970) "A general method applicable to the search for similarities in the amino acid sequence of two proteins." J. Mol. Biol. 48, 443-453.
Nolden L, Edenhofer F, Haupt S, Koch P, Wunderlich FT, Siemen H, Brustle O. (2006) "Site- specific recombination in human embryonic stem cells induced by cell-permeant Cre recombinase." Nat. Methods 3, 461-467.
Oess S, Hildt E., Novel cell permeable motif derived from the PreS2-domain of hepatitis-B virus surface antigens., Gene Ther. 2000 May;7(9):750-8
O'Doherty U, Swiggard WJ, Malim MH (2000) "Human immunodeficiency virus type 1 spinoculation enhances infection through virus binding." J Virol. 74, 10074-10080. Pearson WR, Lipman DJ (1988) "Improved tools for biological sequence comparison." Proc Natl Acad Sci USA 85, 2444-2448.
Peitz M, Pfannkuche K, Rajewsky K, Edenhofer F. (2002) "Ability of the hydrophobic FGF and basic TAT peptides to promote cellular uptake of recombinant Cre recombinase: A tool for efficient genetic engineering of mammalian genomes." Proc. Natl. Acad. Sci. USA 99, 4489-4494.
Ratner L, Starcich B, Josephs SF, Hahn BH, Reddy EP, Livak KJ, Petteway SR, Jr., Pearson ML, Haseltine WA, Arya SK, (1985) "Polymorphism of the 3' open reading frame of the virus associated with the acquired immune deficiency syndrome, human T-lymphotropic virus type III." Nucl. Acids Res. 13, 8219-8229.
Richard JP, Melikov K, Brooks H, Prevot P, Lebleu B, Chemomordik LV (2005) "Cellular uptake of the unconjugated TAT peptide involves clathrin-dependent endocytosis and heparin sulfate receptors." J. Biol. Chem. 280, 15300-15306.
Rufer AW, Sauer B (2002) "Non-contact positions impose site selectivity on Cre recombinase." Nucl. Acids Res. 30, 2764-2771.
Ruhl M, Himmelspach M, Bahr GM, Hammerschmid F, Jaksche H, Wolff B, Aschauer H, Farrington GK, Probst H, Bevec D, Hauber J (1993) "Eukaryotic initiation factor 5 A is a cellular target of the human immunodeficiency virus type 1 Rev activation domain mediating trans-activation" J Cell Biol. 123, 1309-1320.
Sanger F, Nickler S, Coulson AR (1977) "DNA sequencing with chain-terminating inhibitors." Proc. Natl. Acad. Sci. USA 74, 5463-5467.
Santoro SW, Schultz PG (2002) "Directed evolution of the site specificity of Cre recombinase." Proc. Natl. Acad. Sci. USA 99, 4185-4190.
Saraf-Levy T, Santoro SW, Volpin H, Kushnirsky T, Eyal Y, Schultz PG, Gidoni D, Carmi N (2006) "Site-specific recombination of asymmetric lox sites mediated by a heterotetrameric Cre recombinase complex." Bioorg. Med. Chem. 14, 3081-3089.
Sauer B, McDermott J (2004) "DNA recombination with a heterospecific Cre homolog identified from comparison of the pac-cl regions of PI -related phages." Nucl. Acids. Res. 32, 6086- 6095.
Schambach A, Bohne J, Chandra S, Will E, Margison GP, Williams DA, Baum C (2006) "Equal potency of gammaretro viral and lenti viral SIN vectors for expression of O6- methylguanine-DNA methyltransferase in hematoietic cells." Molecular Therapy 13, 391- 400.
Scherr M, Eder M (2002) "Gene Transfer into Hematopoietic Stem Cells Using Lentiviral Vectors." Current Gene Therapy 2, 45-55.
Shehu-Xhilaga M, Tachedjian G, Crowe SM, Kedzierska K. (2005) "Antiretro viral compounds: mechanisms underlying failure of HAART to eradicate HIV-1." Curr. Med. Chem. 12, 1705-1719.
Shimshek DR, Kim J, Hubner MR, Spergel DJ, Buchholz F, Casanova E, Stewart AF, See- burg PH, Sprengel R (2002) "Codon-improved Cre recombinase (iCre) expression in the mouse." Genesis 32(1), 19-26.
Smith Tf, Waterman MS (1981) "Overlapping genes and information theory." J Theor. Biol. 91, 379-380.
Stark WM, Boocock MR, Sherratt DJ (1992) "Catalysis by site-specific recombinases." Trends Genet. 8, 432-439. Stemmer WPC (1994) "Rapid evolution of a protein in vitro by DNA shuffling." Nature 370, 389-391.
Sternberg N, Hamilton D (1981) "Bacteriophage PI site-specific recombination. I. Recombination between loxP sites." J. Mol. Biol. 150, 467-486.
Van Duyne GD (2001) "A structural view of cre-loxp site-specific recombination." Annu. Rev. Biophys. Biomol. Struct. 30, 87-104.
Vives E, Brodin P, Lebleu B (1997) "A truncated HIV-1 Tat protein basic domain rapidly translocates through the plasma membrane and accumulates in the cell nucleus." J. Biol. Chem. 272, 16010-16017.
Vives E (2003) "Cellular uptake of the TAT peptide: an endocytosis mechanism following<"> ionic interactions." J. Mol. Recognit. 16, 265-271.
Volkert FC, Broach JR (1986) "Site-specific recombination promotes plasmid amplification in yeast." Cell 46, 541-550.
Voziyanov Y, Konieczka JH, Stewart AF, Jayaram M (2003) "Stepwise manipulation of DNA specificity in Flp recombinase: progressively adapting Flp to individual and combinatorial mutations in its target site." J. Mol. Biol. 326, 65-76.
Yuan L, Kurek I, English J, Keenan R (2005) "Laboratory-directed protein evolution" Microbiol. Mol. Biol. Rev. 69, 373-92.
WO 2002/44409
WO 2008/083931
WO 2011/147590.
| название | год | авторы | номер документа |
|---|---|---|---|
| АДАПТИРОВАННАЯ РЕКОМБИНАЗА ДЛЯ РЕКОМБИНАЦИИ АСИММЕТРИЧНЫХ УЧАСТКОВ-МИШЕНЕЙ ВО МНОЖЕСТВЕ ШТАММОВ РЕТРОВИРУСОВ | 2011 |
|
RU2617968C2 |
| ПРИМЕНЕНИЕ АДАПТИРОВАННЫХ РЕКОМБИНАЗ ДЛЯ ЛЕЧЕНИЯ РЕТРОВИРУСНЫХ ИНФЕКЦИЙ | 2008 |
|
RU2501860C2 |
| ВЕКТОРНАЯ СИСТЕМА НА ОСНОВЕ ASLV | 2010 |
|
RU2566563C2 |
| СПОСОБЫ И СОСТАВЫ ДЛЯ ПРОИЗВОДСТВА ЛИМФОЦИТОВ И ИХ РЕГУЛИРУЕМОГО УВЕЛИЧЕНИЯ | 2017 |
|
RU2755059C2 |
| УСОВЕРШЕНСТВОВАНИЕ ГЕНЕТИЧЕСКИХ КОНСТРУКЦИЙ ДЛЯ ПОВЫШЕНИЯ ЭФФЕКТИВНОСТИ АНТИВИЧ ТЕРАПИИ | 2013 |
|
RU2533817C1 |
| ИММОРТАЛИЗОВАННАЯ КЛЕТОЧНАЯ ЛИНИЯ ПТИЦ | 2008 |
|
RU2475536C2 |
| ГЕНЕТИЧЕСКИЕ КОНСТРУКЦИИ ДЛЯ АНТИВИЧ-ТЕРАПИИ | 2010 |
|
RU2426788C1 |
| СПОСОБ БЫСТРОГО ОТБОРА ВАРИАНТОВ GP-120 ВИЧ | 2012 |
|
RU2603732C2 |
| ТЕЛОМЕРАЗНАЯ ОБРАТНАЯ ТРАНСКРИПТАЗА ПТИЦ | 2007 |
|
RU2415867C2 |
| БИОПРОИЗВОДСТВО ЛЕНТИВИРУСНЫХ ВЕКТОРОВ | 2016 |
|
RU2729385C2 |

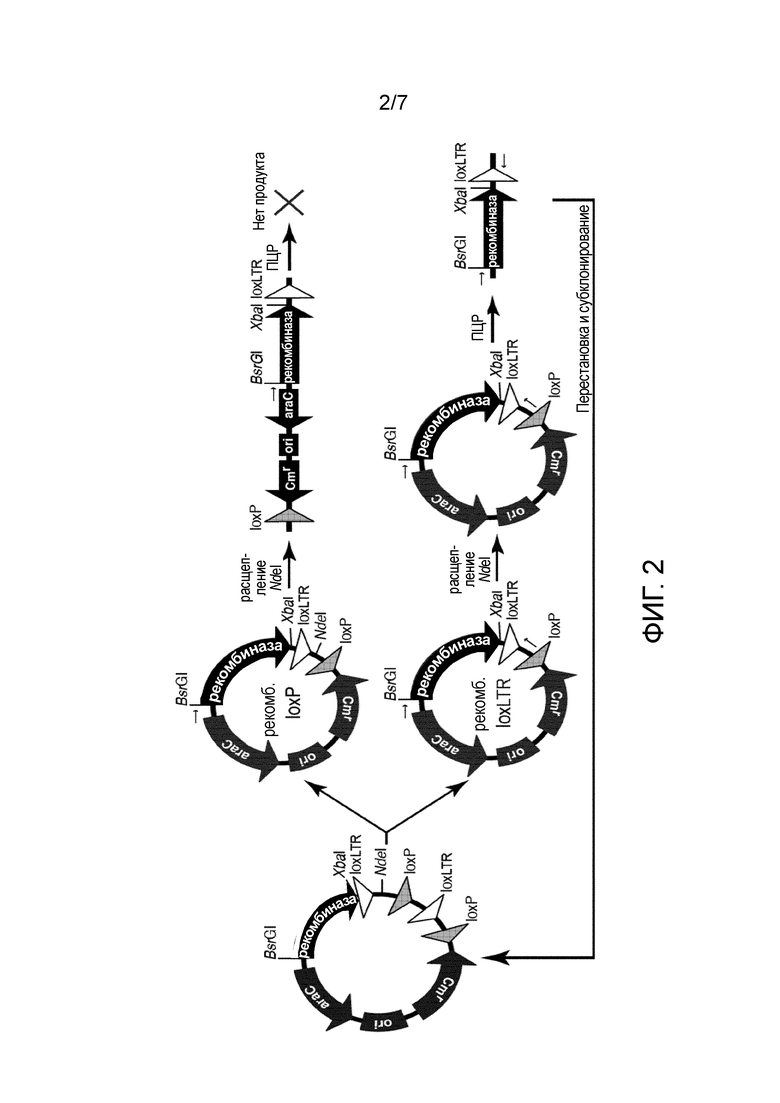
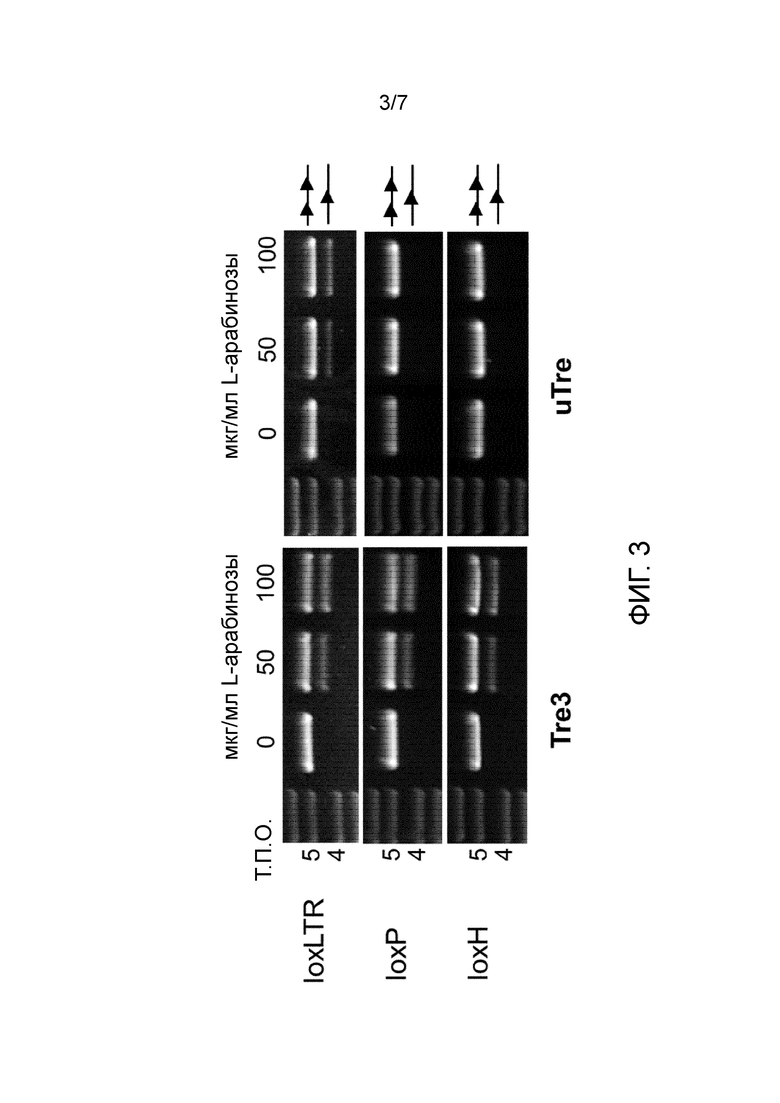
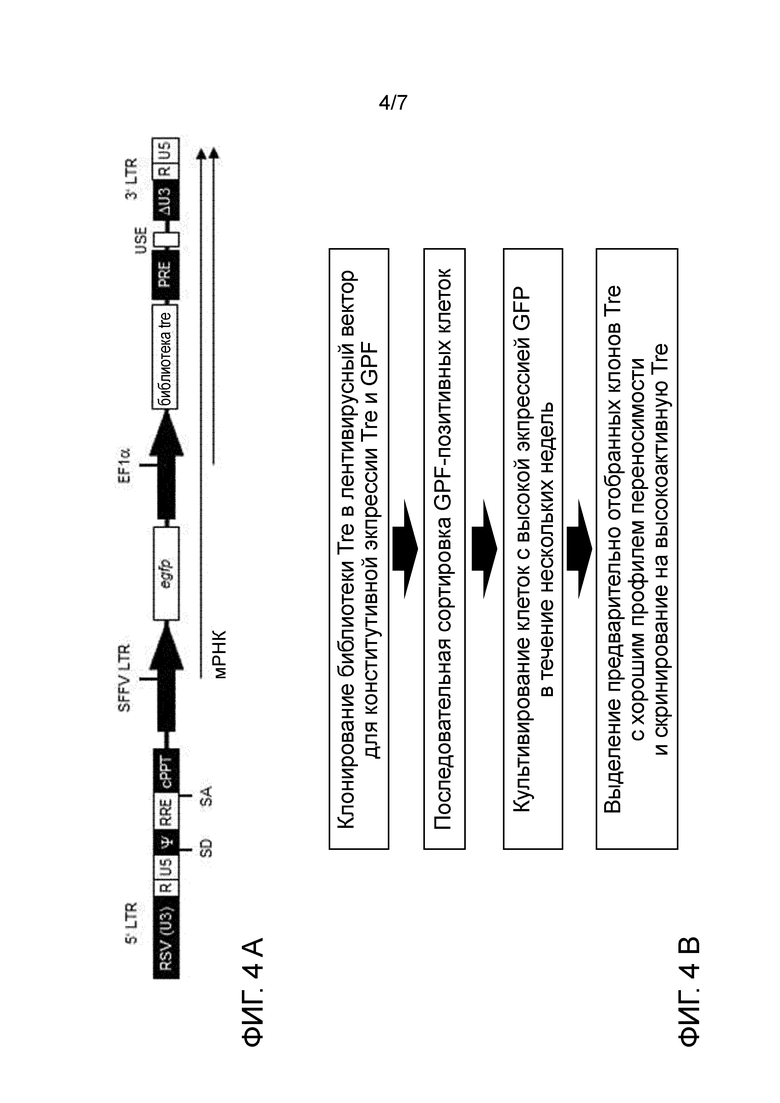
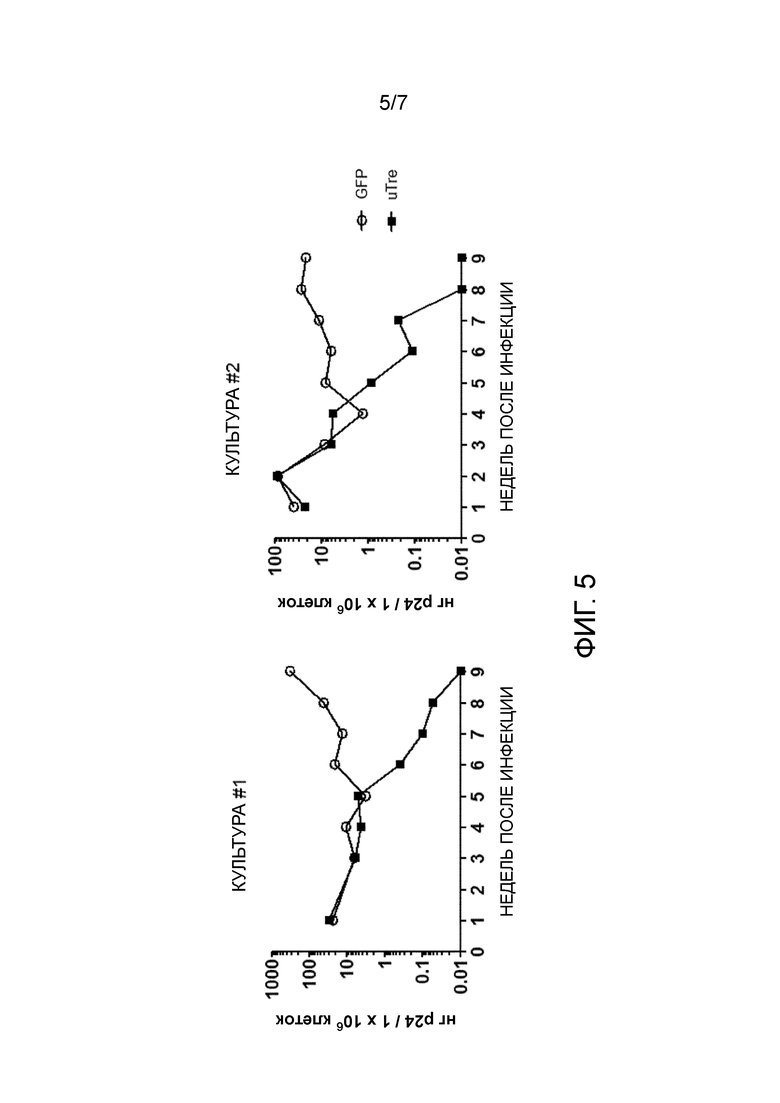
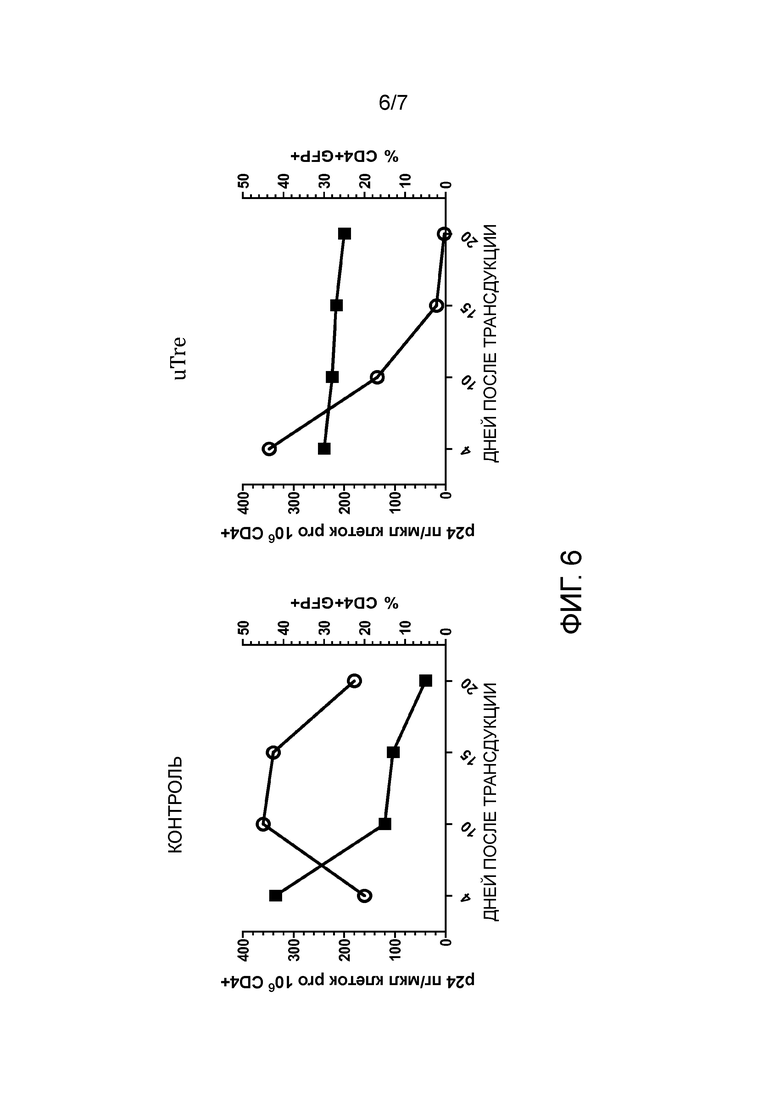
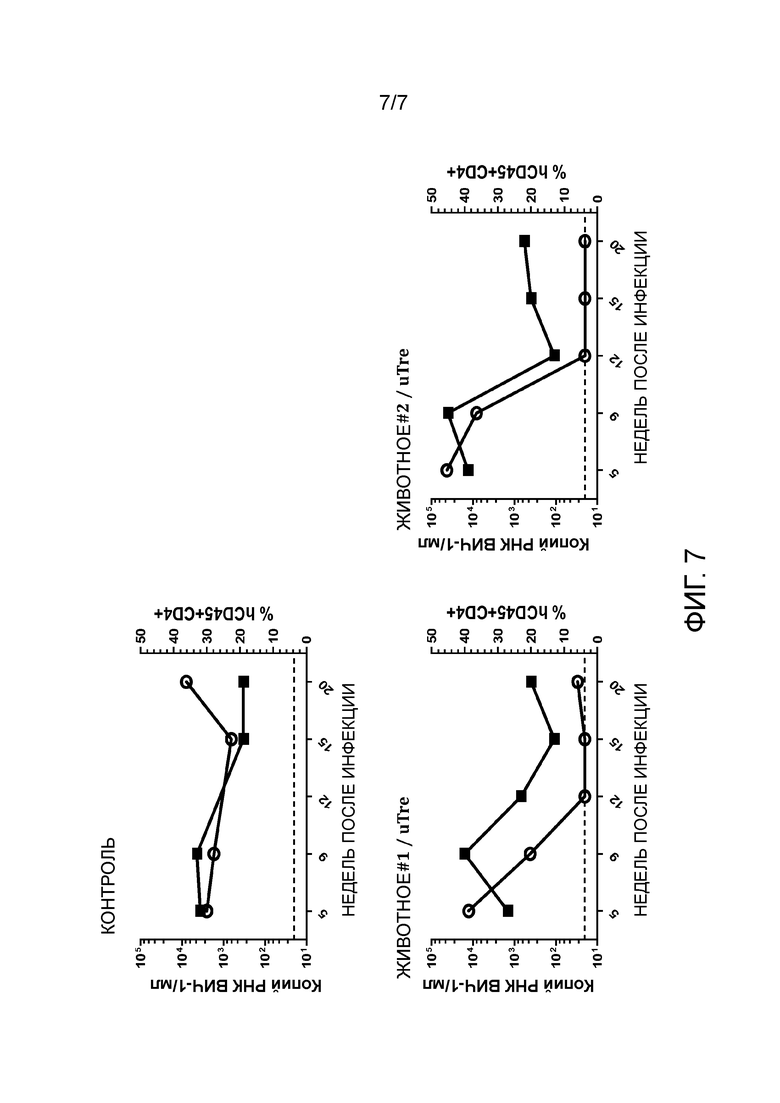
Изобретение относится к биотехнологии. Описана нуклеиновая кислота, кодирующая адаптированную рекомбиназу, где адаптированная рекомбиназа способна к рекомбинированию асимметричных последовательностей-мишеней SEQ ID NO:1 внутри длинного концевого повтора провирусной ДНК множества штаммов ВИЧ-1, причем аминокислотная последовательность адаптированной рекомбиназы имеет по меньшей мере 95% идентичности последовательности с последовательностью в соответствии с SEQ ID NO:10, причем указанная адаптированная рекомбиназа содержит определенные аминокислотные замены по сравнению с SEQ ID NO:6: V7L, P12S, P15L, M30V, H40R, M44V, S51T, Y77H, K86N, Q89L, G93A, S108G, C155G, A175S, A249V, R259D, E262R, T268A, D278G, P307A, N317T, I320S. Описана адаптированная рекомбиназа, кодируемая описанной нуклеиновой кислотой, которая необязательно экспрессирована в виде слитого белка. Представлена трансформированная клетка для лечения или предотвращения инфекции ВИЧ-1, содержащая описанную нуклеиновую кислоту. Описана фармацевтическая композиция для лечения или предотвращения инфекции ВИЧ-1, содержащая описанную нуклеиновую кислоту, описанную адаптированную рекомбиназу и/или представленную трансформированную клетку и фармацевтически приемлемый(е) носитель(и), наполнитель(и) и/или адъювант(ы). Описан способ получения описанной нуклеиновой кислоты. Описан способ получения описанной адаптированной рекомбиназы. Изобретение позволяет получать хорошо переносимую и высокоспецифическую адаптированную рекомбиназу. 7 н. и 14 з.п. ф-лы, 7 ил., 6 пр.
1. Нуклеиновая кислота, кодирующая адаптированную рекомбиназу, где адаптированная рекомбиназа способна к рекомбинированию асимметричных последовательностей-мишеней SEQ ID NO:1 внутри длинного концевого повтора провирусной ДНК множества штаммов ВИЧ-1, причем аминокислотная последовательность адаптированной рекомбиназы имеет по меньшей мере 95% идентичности последовательности с последовательностью в соответствии с SEQ ID NO:10, причем указанная адаптированная рекомбиназа содержит определенные аминокислотные замены по сравнению с SEQ ID NO:6: V7L, P12S, P15L, M30V, H40R, M44V, S51T, Y77H, K86N, Q89L, G93A, S108G, C155G, A175S, A249V, R259D, E262R, T268A, D278G, P307A, N317T, I320S.
2. Нуклеиновая кислота по п.1, отличающаяся тем, что аминокислотная последовательность адаптированной рекомбиназы имеет по меньшей мере 99% идентичности последовательности с последовательностью в соответствии с SEQ ID NO:10.
3. Нуклеиновая кислота по п.1, отличающаяся тем, что адаптированная рекомбиназа содержит аминокислотную последовательность в соответствии с SEQ ID NO:10.
4. Нуклеиновая кислота по любому из пп.1-3, отличающаяся тем, что адаптированная рекомбиназа содержит аминокислотную последовательность в соответствии с SEQ ID NO: 11-13, или комбинацию любых из указанных последовательностей.
5. Нуклеиновая кислота по любому из пп.1-4, отличающаяся тем, что адаптированная рекомбиназа является высокоспецифической в отношении рекомбинации SEQ ID NO: 1, так как указанная адаптированная рекомбиназа не рекомбинирует последовательности loxP (SEQ ID NO:4) или loxH (SEQ ID NO:5) с поддающейся обнаружению активностью.
6. Адаптированная рекомбиназа, кодируемая нуклеиновой кислотой по любому из пп.1-5, которая необязательно экспрессирована в виде слитого белка.
7. Трансформированная клетка для лечения или предотвращения инфекции ВИЧ-1, содержащая нуклеиновую кислоту по любому из пп.1-5.
8. Трансформированная клетка по п.7, отличающаяся тем, что указанная клетка является стволовой клеткой из гемопоэтической линии.
9. Фармацевтическая композиция для лечения или предотвращения инфекции ВИЧ-1, содержащая нуклеиновую кислоту по любому из пп.1-5, адаптированную рекомбиназу по п.6 и/или трансформированную клетку по п.7 или 8 и фармацевтически приемлемый(е) носитель(и), наполнитель(и) и/или адъювант(ы).
10. Фармацевтическая композиция по п.9, отличающаяся тем, что фармацевтическая композиция составлена для введения индивиду, если провирусная ДНК, обнаруженная в образце, полученном от индивида, содержит асимметричную последовательность-мишень, идентифицированную на стадии (a), на которую была отобрана рекомбиназа.
11. Способ получения нуклеиновой кислоты по любому из пп. 1-5, включающий следующие стадии:
(a) идентификация последовательности с гомологией по меньшей мере 30% с последовательностью левой половины участка и последовательностью правой половины участка по меньшей мере одного известного участка-мишени рекомбиназы в последовательности LTR провирусной ДНК множества штаммов ретровирусов, в которой гомологичные последовательности разделены спейсером из 5-12 нуклеотидов, и в которой асимметричная последовательность-мишень обнаружена во множестве штаммов ретровирусов;
(b) идентификация двух последовательностей, причем первая последовательность соответствует последовательности асимметричной последовательности-мишени стадии (a), гомологичной левой половине участка указанного известного участка-мишени, и называется "последовательность 1 половины участка", и в которой вторая последовательность соответствует последовательности асимметричной последовательности-мишени стадии (a), гомологичной правой половине участка, и называется "последовательность 2 половины участка";
(c) определение нуклеотидов внутри последовательностей стадии (b), отличающихся от соответствующих гомологичных последовательностей левой половины участка и правой половины участка по меньшей мере одного известного гомологичного участка-мишени стадии (a);
(d) генерация первой подгруппы двух мишеневых нуклеиновых кислот, содержащей последовательности-мишени, в которой первая последовательность-мишень обозначена субучасток 1 и содержит смежно друг другу и в порядке от 5' к 3' последовательность 1 половины участка стадии (b), спейсерную последовательность асимметричной последовательности-мишени и инвертированный повтор последовательности 1 половины участка, и в которой вторая последовательность-мишень обозначена субучасток 2 и содержит смежно друг другу и в порядке от 5' к 3' инвертированный повтор последовательности 2 половины участка, спейсерную последовательность асимметричной последовательности-мишени и последовательность 2 половины участка стадии (b);
(e) генерация второй подгруппы мишеневых нуклеиновых кислот, содержащей модифицированные последовательности-мишени на основании последовательностей-мишеней в первой подгруппе стадии (d),
причем в последовательностях, основанных на субучастке 1 в последовательности левой половины участка участок нуклеотидов, отличающихся от последовательности соответствующей гомологичной половины участка по меньшей мере одним известным участком-мишенью стадии (a), замещен природными нуклеотидами, обнаруженными в указанном известном участке-мишени, при условии, что последовательность указанной половины участка содержит один, два или три нуклеотида, отличающихся от указанного известного участка-мишени, причем правая половина участка указанной модифицированной последовательности-мишени образована инвертированным повтором указанной модифицированной последовательности левой половины участка, которая отделена от указанной модифицированной последовательности левой половины участка спейсерной последовательностью асимметричной последовательности-мишени, и
причем в последовательностях, основанных на субучастке 2 в последовательности правой половины участка, участок нуклеотидов, отличающихся от последовательности соответствующей гомологичной половины участка по меньшей мере одного известного участка-мишени стадии (a), замещен природными нуклеотидами, обнаруженными в указанном известном участке-мишени, при условии, что последовательность указанной половины участка содержит один, два или три нуклеотида, отличающихся от указанного известного участка-мишени, причем левая половина участка указанной модифицированной последовательности-мишени образована инвертированным повтором указанной последовательности модифицированной правой половины участка, которая отделена от указанной последовательности модифицированной правой половины участка спейсерной последовательностью асимметричной последовательности-мишени,
таким образом, во всех модифицированных последовательностях половины участка, происходящих от одной последовательности-мишени первой подгруппы стадии (d), в совокупности могут быть обнаружены все отличающиеся нуклеотиды, тогда как ни одна из указанных модифицированных последовательностей половины участка не содержит отдельно все отличающиеся нуклеотиды,
(f) применение направленной молекулярной эволюции отдельно для по меньшей мере одной рекомбиназы, распознающей известный гомологичный участок-мишень в соответствии со стадией (a), с применением каждой нуклеиновой кислоты второй подгруппы, полученной на стадии (e), в качестве субстрата;
(g) перестановка в библиотеках рекомбиназ, полученных на стадии (f), причем все библиотеки рекомбиназ, полученные на последовательности, основанные на субучастке 1, комбинировали и подвергали перестановкам, и причем все библиотеки рекомбиназ, полученные на последовательности, основанные на субучастке 2, комбинировали и подвергали перестановкам;
(h) применение направленной молекулярной эволюции для библиотек с перестановками, полученных на стадии (g), с применением каждой нуклеиновой кислоты подгруппы в соответствии со стадией (d) в качестве субстрата;
(i) перестановка в библиотеках рекомбиназ, полученных на стадии (h);
(j) применение направленной молекулярной эволюции для библиотеки с перестановками, полученной на стадии (g), с применением нуклеиновой кислоты, содержащей асимметричную последовательность-мишень стадии (a) в качестве субстрата, при условии, что получена по меньшей мере одна рекомбиназа, которая является активной в отношении асимметричной последовательности-мишени внутри LTR ретровирусной ДНК стадии (a);
(k) выделение нуклеиновой кислоты, кодирующей по меньшей мере одну рекомбиназу, полученную на стадии (j), из библиотеки и клонирование ее в эволюционный вектор, который позволяет негативный отбор адаптированных рекомбиназ, рекомбинирующих известный участок-мишень в соответствии со стадией (a), посредством этого получая библиотеку;
(l) применение направленной молекулярной эволюции для библиотеки, полученной на стадии (k);
(m) перестановка в библиотеках, полученных на стадии (l);
(n) выделение нуклеиновой кислоты, кодирующей по меньшей мере одну адаптированную рекомбиназу, полученную на стадии (m), и клонирование в векторе для экспрессии кодируемой рекомбиназы и селектируемого маркера в человеческой клетке, посредством этого получая векторную библиотеку,
(o) трансформирование человеческих клеток указанной библиотекой векторов, полученной на стадии (n);
(p) культивирование клеток, экспрессирующих указанный селектируемый маркер, в течение по меньшей мере 1 недели, и отбор для высокой экспрессии селектируемого маркера;
(q) выделение нуклеиновой кислоты(кислот), кодирующей рекомбиназу из клеток, экспрессирующих указанный селектируемый маркер, полученных на стадии (p);
(r) отбор на нуклеиновую кислоту, кодирующую рекомбиназу, способную к рекомбинированию асимметричной последовательности-мишени стадии (a);
(s) выделение нуклеиновой кислоты, кодирующей по меньшей мере одну рекомбиназу, полученную на стадии (f), из библиотеки; и,
(t) необязательно, клонирование нуклеиновой кислоты, полученной на стадии (s), в подходящем векторе экспрессии.
12. Способ по п.11, в котором направленная молекулярная эволюция на стадиях (h), (j) и (l) является связанной с субстратом эволюцией белка.
13. Способ по п.11, в котором на стадии (o) трансформируют человеческие T-клетки.
14. Способ по п.11, в котором ретровирус представляет собой ВИЧ и в котором асимметричная последовательность-мишень имеет последовательность, представленную SEQ ID NO:1 или SEQ ID NO:2.
15. Способ по п.14, в котором ВИЧ представляет собой ВИЧ-1.
16. Способ по п.14, в котором асимметричная последовательность-мишень имеет последовательность, представленную SEQ ID NO:1.
17. Способ по любому из пп.11-16, в котором по меньшей мере одна известная рекомбиназа, последовательность-мишень которой применена на стадии (a) и на основе которой применена молекулярная направленная эволюция на стадии (f), является частью библиотеки рекомбиназ.
18. Способ по любому из пп.11-17, в котором вектор экспрессии на стадии (t) выбран из группы, состоящей из ретровирусных векторов, лентивирусных векторов, спумавирусных векторов, аденовирусных векторов и адено-ассоциированных вирусных векторов.
19. Способ получения адаптированной рекомбиназы, причем указанный способ включает способ по любому из пп.11-18 и дополнительную стадию экспрессии адаптированной рекомбиназы из нуклеиновой кислоты, кодирующей рекомбиназу, встроенную в вектор экспрессии, в подходящей клетке-хозяина, причем рекомбиназа необязательно экспрессирована в виде слитого полипептида, содержащего аминокислотную последовательность адаптированной рекомбиназы.
20. Способ получения трансформированной клетки, причем указанный способ включает способ по любому из пп.11-19 и дополнительную стадию введения вектора экспрессии в клетку in vitro, причем клетка предпочтительно является зрелой стволовой клеткой.
21. Способ по любому из пп.11-20, дополнительно включающий стадию получения вектора экспрессии, рекомбиназы или клетки в виде фармацевтической композиции.
| WO2011147590 A2, 01.12.2011 | |||
| WO2008083931 A1, 17.07.2008 | |||
| FRANK BUCHHOLZ et al., In vitro evolution and analysis of HIV-1 LTR-specific recombinases, Methods Vol | |||
| Веникодробильный станок | 1921 |
|
SU53A1 |
| Транспортер для перевозки товарных вагонов по трамвайным путям | 1919 |
|
SU102A1 |
| INDRANI SARKAR et al., HIV-1 Proviral DNA Excision Using an Evolved Recombinase, Science Vol | |||
| Способ амидирования жидких сульфохлоридов ароматического ряда | 1921 |
|
SU316A1 |
| Аппарат для электролиза воды | 1924 |
|
SU1912A1 |
| RU 2012157303 | |||

