Настоящее изобретение относится к олигонуклеотидам-ловушкам и антисмысловым олигонуклеотидам с последовательностью нуклеиновой кислоты в соответствии с SEQ ID NO:1-43, а также их применению в качестве лекарственного средства.
Одна из важных целей расшифровки человеческого генома заключается в том, чтобы идентифицировать патологические гены (на основании механизма действия их продуктов) и патологические изменения в структуре этих генов и отнести их к определенной клинической картине болезни. Поэтому этиотропная терапия большинства заболеваний, по-видимому, является достижимой, если принимать, что их причиной является определенное количество генных продуктов, которые экспрессируются слишком сильно, слишком слабо или с ошибками. В действительности уже известен для целого ряда наследственных заболеваний (например, муковисцидоза), как правило, единичный генетический дефект (моногенетические заболевания), тогда как ситуация для других заболеваний (например, гипертония) представляется намного более сложной. Последние, очевидно, являются результатом не единичных, а множественных генетических дефектов (полигенетические заболевания), которые предопределяют развитие болезни у пораженных людей при воздействии определенных факторов окружающей среды. Несмотря на это ограничение, направленное вмешательство в экспрессию одного или более чем одного гена дает возможность для этиотропной, а не только симптоматической терапии.
Транскрипционные факторы представляют собой ДНК-связывающие белки, которые присоединяются к промоторному участку одного или более чем одного гена в ядре клетки и таким образом регулируют экспрессию, то есть синтез белка, который этот ген кодирует. Наряду с физиологически важным контролем процессов развития и дифференцировки в человеческом организме, транскрипционные факторы имеют высокий болезнетворный потенциал, прежде всего, когда экспрессия гена активируется не в тот момент. Дополнительно (при тех же обстоятельствах) транскрипционные факторы могут блокировать гены с защитными функциями и действовать как предопределяющие развитие какой-либо болезни. В этом отношении описанный ниже принцип терапии против транскрипционных факторов направлен на ингибирование патологических генов и, напротив, на активирование защитных генов.
Воспаление представляет собой защитную реакцию организма и его тканей от повреждающих раздражителей с целью устранения повреждения или по меньшей мере локального ограничения, а также устранения причины повреждения (например, проникнувшие бактерии или инородные тела). Инициирующим фактором воспаления могут быть микроорганизмы (бактерии, вирусы, грибы или паразиты), инородные тела (пыльца, кристаллы асбеста или силикатные кристаллы), разрушение ткани посредством механического повреждения, химические повреждающие факторы и физические влияния, а также факторы собственного организма (распадающиеся опухолевые клетки, внесосудистая кровь, аутоиммунные реакции) или кристаллы веществ, выпавших в осадок в организме (мочевая кислота, оксалат и фосфат кальция, холестерин).
Быстрая активация тучных клеток (в ткани) или базофильных гранулоцитов в крови является примером инициирования очень сильной острой воспалительной реакции и характерной для иммунологической реакции гиперчувствительности немедленного типа (гуморальная аллергия 1 типа). Если организм уже вступил в контакт с антигеном (или аллергеном в случае гиперчувствительности), то в качестве ответной реакции на него будут сенсибилизироваться В-лимфоциты, которые преобразуются в плазмоциты при кооперации с предварительно также сенсибилизированными CD4-положительными Т-хелперами 2 типа (Тп2-клетки) и продуцируют антитела типа IgE против данного антигена. При данном процессе дифференцировки костимуляция В-лимфоцитов через СD40-рецептор Тh2-клетками, экспрессирующими соответствующие лиганды (CD 154), имеет решающее значение. При связывании lgE-антител, нагруженных антигеном, с соответствующими рецепторами (тип 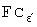 ) на тучных клетках высвобождаются различные медиаторы воспаления, в частности гистамин, интерлейкин-8, лейкотриен и фактор некроза опухоли-α (ФНОα). Вследствие этого происходит привлечение профессиональных воспалительных клеток, в особенности эозинофильных и нейтрофильных гранулоцитов, и моноцитов, а также Т-лимфоцитов в место события (хемотаксис). Одновременно происходит, прежде всего, гистамин-зависимое расширение кровеносных сосудов и увеличение проницаемости клеток эндотелия, выстилающих стенки сосудов. Вследствие расширения сосудов скорость течения падает, что облегчает установление физического контакта привлеченных лейкоцитов с клетками эндотелия. Эти клетки эндотелия, уже активированные посредством экспозиции цитокинов (например ФНОα), экспрессируют на своей люминальной поверхности увеличенное количество селектинов (например Е-селектина), которые вызывают роллинг лейкоцитов вдоль клеток эндотелия и вследствие этого активацию дальнейших молекул адгезии (интегринов; например, молекулы межклеточной адгезии 1 [1САМ-1] или васкулярно-клеточной молекулы адгезии 1 [VCAM-1]). Лейкоциты могут теперь прилипать к стенке сосуда (маргинация), а увеличение проницаемости, обусловленное гистамином (ослабление связи клеток эндотелия), способствует их последующему выходу во внесосудистое пространство (диапедез). Одновременно увеличенное количество жидкости, богатой белками (воспалительный экссудат), достигает интерстициального пространства, образуя отек. Посредством увеличивающегося давления ткани и посредством дальнейших медиаторов, образованных воспалительными клетками, будут раздражаться окружающие нервные окончания, которые вызывают боли, тем самым приводя к повреждению ткани.
) на тучных клетках высвобождаются различные медиаторы воспаления, в частности гистамин, интерлейкин-8, лейкотриен и фактор некроза опухоли-α (ФНОα). Вследствие этого происходит привлечение профессиональных воспалительных клеток, в особенности эозинофильных и нейтрофильных гранулоцитов, и моноцитов, а также Т-лимфоцитов в место события (хемотаксис). Одновременно происходит, прежде всего, гистамин-зависимое расширение кровеносных сосудов и увеличение проницаемости клеток эндотелия, выстилающих стенки сосудов. Вследствие расширения сосудов скорость течения падает, что облегчает установление физического контакта привлеченных лейкоцитов с клетками эндотелия. Эти клетки эндотелия, уже активированные посредством экспозиции цитокинов (например ФНОα), экспрессируют на своей люминальной поверхности увеличенное количество селектинов (например Е-селектина), которые вызывают роллинг лейкоцитов вдоль клеток эндотелия и вследствие этого активацию дальнейших молекул адгезии (интегринов; например, молекулы межклеточной адгезии 1 [1САМ-1] или васкулярно-клеточной молекулы адгезии 1 [VCAM-1]). Лейкоциты могут теперь прилипать к стенке сосуда (маргинация), а увеличение проницаемости, обусловленное гистамином (ослабление связи клеток эндотелия), способствует их последующему выходу во внесосудистое пространство (диапедез). Одновременно увеличенное количество жидкости, богатой белками (воспалительный экссудат), достигает интерстициального пространства, образуя отек. Посредством увеличивающегося давления ткани и посредством дальнейших медиаторов, образованных воспалительными клетками, будут раздражаться окружающие нервные окончания, которые вызывают боли, тем самым приводя к повреждению ткани.
Гранулоциты, которые мигрируют к месту воспаления, и моноциты, передифференцированные в макрофаги, пытаются элиминировать агенты, вызвавшие воспаление, посредством фагоцитоза или лизиса, при этом высвобождаются среди прочего протеолитические ферменты и кислородные радикалы, которые могут повредить также окружающие здоровые ткани. В особенности этому может способствовать активация макрофагов разнообразными способами (например, высвобождением цитокинов, таких как интерлейкин-1β или интерлейкин-6), так что целый организм включается в изначально местную воспалительную реакцию в форме ответа острой фазы. Типичными признаками ответа острой фазы являются усталость, подавленность и лихорадка, повышенное высвобождение лейкоцитов из костного мозга (лейкоцитоз), обнаружение белков острой фазы в крови (например, С-реактивного белка), стимуляция иммунной системы, а также потеря веса ввиду измененного обмена веществ.
Если причина воспаления может быть устранена, то начинается процесс заживления, при котором разрушенная ткань восстанавливается. В наилучшем случае она будет полностью восстановлена (восстановление до целого), тогда как при больших повреждениях или чрезмерном образовании соединительной ткани (в особенности коллагена), образуется рубец, который может быть ассоциирован со значительными функциональными нарушениями из-за пораженной ткани. Если причина не может быть немедленно устранена (инородные тела или раневая инфекция), то заживление раны замедляется из-за одновременного усиления иммиграции и активности фагоцитов с последующей гибелью ткани (некроз) вплоть до формирования полостей (абсцесс). Результатом практически всегда является рубцовая перестройка ткани с соответствующей потерей функции. Если не происходит локального ограничения воспаления, обусловленного возбудителем, оно распространяется через лимфатическую систему на весь организм, результатом чего является сепсис с возможным смертельным исходом (септический шок).
Нарушение заживления раны также имеет место тогда, когда процессы воспаления и заживления находятся в равновесии. Результатом является хроническое воспаление, которое может быть фиброзным (чрезмерный синтез коллагена) или гранулематозным (организация воспалительных клеток в виде грануляционной ткани) и которое, как правило, приводит к непрерывному разрушению или возрастающему ограничению функций пораженных тканей.
Помимо описанных общих воспалительных реакций, которые могут переходить в хронические, существуют воспалительные заболевания, которые обнаруживают общие особенности, но также и различия в отношении патогенеза, лежащего в их основе. Двумя такими воспалительными заболеваниями являются, например, осложнения после кардиохирургического вмешательства и иммунологические реакции несовместимости, которым посвящается больше места в данных материалах заявки ввиду их огромного клинического значения и разнообразия.
Механическое расширение с помощью баллонного катетера (чрескожная ангиопластика), а также расширение атеросклеротических суженных артерий с помощью венозных шунтов по-прежнему представляет собой терапию выбора у пациентов с коронарными и периферическими нарушениями кровообращения для предотвращения угрожающего инфаркта или органной недостаточности. Однако частота повторной закупорки артерий (рестеноза), которые были механически расширены и (в большинстве случаев) снабжены металлическими эндопротезами сосудов (стентами), неприемлемо повышается на 20-50% за 6 месяцев. Также частота закупорки аортокоронарных и периферических венозных шунтов, составляющая 50-70% за 5 месяцев, является более чем неудовлетворительной, в особенности на фоне риска во время процедуры и послеоперационного риска у оперируемых пациентов. Вероятно, из-за механического повреждения стенки сосудов (в данном случае пораженными являются как клетки эндотелия, так и мышечные клетки сосудов) после ангиопластики рестеноз проявляет, в особенности в ранней стадии, выраженный воспалительный компонент, который характеризуется среди прочего инфильтрацией профессиональных воспалительных клеток (прежде всего, моноциты и Т-лимфоциты) в стенке сосуда. Формирование фибропролиферативного стеноза (гиперплазия внутренней оболочки сосуда) аортокоронарных, а также периферических венозных шунтов также основано на воспалительной реакции, причиной которой являются, в частности, механические и физические патогенные факторы. Также давно известно, что к разрушению ткани, обусловленному воспалением, приводят так называемые ишемические/реперфузионные повреждения при хирургических вмешательствах или трансплантации органов, при которых решающую роль играет, в особенности, взаимодействие клеток эндотелия и профессиональных воспалительных клеток (прежде всего, гранулоцитов, но также моноцитов и Т-клеток), а также высвобождение веществ, повреждающих ткань (кислородных радикалов, цитокинов).
В отношении упомянутых сердечно-сосудистых осложнений важным является то, что существуют защитные механизмы, прежде всего в клетках эндотелия и клетках гладкой мускулатуры стенки сосуда, которые помогают ограничить степень воспалительной реакции и последующее адаптивное восстановление ткани. К таким механизмам относится, например, синтез оксида азота (NO) посредством NO-синтазы в клетках эндотелия. NO, возможно в качестве эндогенного антагониста супероксид кислородных радикалов, ингибирует среди прочего экспрессию провоспалительных хемокинов (например, моноцитарного хемоаттрактантного белка-1, МСР-1) и молекул адгезии (например, 1САМ-1) в клетках эндотелия, экспрессию рецепторов для факторов роста в клетках гладкой мускулатуры (например, рецептора эндотелина В), а также высвобождение факторов роста из лейкоцитов. Поэтому легко понять, что механическое, так же как и функциональное повреждение эндотелия (например, посредством цитокин-индуцированного снижения экспрессии NO-синтазы в этих клетках), противодействует процессам воспаления и последующей фибропролиферативной перестройки стенки сосуда, лежащим в основе упомянутых сердечно-сосудистых осложнений.
Все предыдущие попытки медикаментозно локализовать рестеноз после ангиопластики не достигли желаемого эффекта у большинства пациентов. В настоящее время преимущественно используют два локальных принципа терапии: уже разрешенная сосудистая брахитерапия, способ сдерживания клеточного роста посредством кратковременного радиоактивного облучения расширенного отрезка сосуда, а также еще находящиеся на клиническом испытании стенты, элюирующие лекарства. При этом способе вводят покрытые полимером стенты, которые «импрегнированы» лекарствами, ингибирующими рост (цитостатиками, иммуносупрессорами), и эти лекарства медленно высвобождаются в течение нескольких недель. Новейшие клинические исследования подтверждают, что оба терапевтических подхода, несмотря на первоначально обнадеживающие результаты, не исключают некоторых серьезных проблем (например, тромбозы в стентах с опасностью инфаркта).
Помимо уже описанных иммунологических реакций несовместимости типа I существуют в принципе четыре другие формы аллергии, а также нарушений иммунорегуляции. Реакцию типа I саму по себе можно разделить, в принципе, на две фазы после произошедшей сенсибилизации: быстрое высвобождение и синтез активных в отношении сосудов медиаторов воспаления из обогащенных IgE тучных клеток, а также реакция замедленного типа, опосредованная привлеченными эозинофильными и нейтрофильными гранулоцитами. Реакция типа I в общей сложности может протекать местно или генерализованно, в зависимости от экспозиции аллергена. Аллергены во вдыхаемом воздухе запускают реакции в дыхательном тракте, обычно сопровождающиеся отеками слизистой оболочки и гиперсекрецией (аллергическая ринопатия, вазомоторный ринит), а также бронхоспазмом (астма), а пищевые аллергены, напротив, в первую очередь вызывают такие желудочно-кишечные симптомы, как тошнота, рвота и диарея. Кожа реагирует на аллергены зудом, припухлостью и крапивницей (уртикария), а также атопическим дерматитом (нейродермит). Если аллерген, напротив, попадает непосредственно в кровяное русло (например, инфузия продуктов крови, лекарственных препаратов), или экспозиция аллергена особенно сильная, то в результате развивается системная реакция немедленного типа, которая в некоторых условиях вызывает опасное для жизни снижение кровяного давления (анафилактический шок).
При реакции II типа в центре внимания находятся антигенно активные клетки (например, чужеродные клетки крови) или внеклеточные белки (например, индуцированное лекарствами изменение поверхности собственных клеток). После аллергизации при вторичном контакте образуются аллерген-специфичные антитела типа IgG и IgM, которые в большом количестве связываются с поверхностью аллергической клетки (опсонизация). Вследствие этого активируются система комплемента (образование мембранатакующего комплекса) и специальная субпопуляция лимфоцитов, натуральные киллеры (НК-клетки). Результатом является разрушение аллергенной клетки посредством цитолиза. Похожая реакция развивается, когда аутоантитела присоединяются к структурам собственного организма, таким как, например, базальная мембрана капилляров почечных клубочков, и вследствие этого развивается быстро прогрессирующий гломерулонефрит с угрозой почечной недостаточности. Помимо Т-хелперов I типа (Тh1-клетки, см. выше) активированные НК-клетки являются основными источниками интерферона-γ, цитокина, который значительно усиливает воспалительную реакцию, особенно посредством активирования макрофагов.
Реакция III типа характеризуется образованием и осаждением иммунокомплексов (комплекс антиген-антитело) с последующим активированием системы комплемента и фагоцитов (гранулоцитов, макрофагов). Они циркулируют в крови и постепенно накапливаются, прежде всего, в капиллярах почечных клубочков, но также в суставах или в коже. Воспалительная реакция, развившаяся в результате этого, может привести среди прочего к (иммунокомплексному) гломерулонефриту, болях в суставах, а также к крапивнице. Также системную реакцию III типа могут вызвать инфекции, если иммунной системе не удалось элиминировать возбудителя (например, стрептококков) полностью. Типичными местными реакциями III типа являются так называемые реакции Артюса в коже после прививки или экзогенный аллергический альвеолит при осаждении комплексов антиген-антитело в легких (например, болезнь любителей голубей). Также реакцией III типа является системная красная волчанка, правда, в смысле аутоиммунного заболевания посредством образования аутоантител.
Реакция IV типа в противоположность упомянутым выше реакциям гиперчувствительности является не гуморальной, а клеточной и достигает своего максимума, как правило, впервые через несколько дней (замедленный тип реакции, или гиперчувствительность замедленного типа). Запускающими факторами являются прежде всего белки проникнувших чужеродных организмов (бактерий, вирусов, грибов и паразитов), другие чужеродные белки (например, глиадин из пшеницы в случае целиакии), а также гаптены (лекарства, металлы (например, никель в случае контактного дерматита), косметика и части растений). Также реакцией IV типа является первичное отторжение трансплантированного органа. Антиген подвергается фагоцитозу (тканевыми) макрофагами, процессингу и презентации наивными Т-хелперами (СDd4-положительными); сенсибилизация Т-хелперов продолжается несколько дней. При вторичном контакте сенсибилизированные таким образом Т-хелперы превращаются в Тh1-клетки; при этом важную роль играет CD154-опосредованная костимуляция антиген-презентирующих клеток (они экспрессируют СD40-рецептор), так как через этот сигнальный путь происходит высвобождение интерлейкина-12 из макрофагов. Интерлейкин-12 запускает дифференцировку и пролиферацию Т-хелперов. Th1-клетки в свою очередь посредством определенных факторов роста (например, GM-CSF) вызывают образование моноцитов в костном мозге, привлекают их с помощью определенных хемокинов (например, MIF) и активируют их посредством высвобождения интерферона-γ (ИФН-γ). Развивающаяся в результате этого очень сильная воспалительная реакция может в большом объеме разрушать ткань собственного организма (например, туберкулез) или трансплантированную ткань. Более того, в отторжении трансплантата участвуют CDS-положительные цитотоксические Т-клетки (цитолиз), которые, как и СD4-положительные ТП1-клетки, способны распознавать свою мишень (поверхность чужеродной клетки) только посредством предшествующей презентации антигена и, соответственно, «вооружаться».
Нарушение иммунорегуляции, подобное реакции IV типа, лежит в основе, например, ревматоидного артрита или рассеянного склероза (аутореактивные Тh1-клетки), а также сахарного диабета (аутореактивные цитотоксические Т-клетки). Помимо соответствующей генетической предрасположенности (МНС-белки, дисбаланс Тh1/Th2) и бактериальных суперантигенов (например, возбудитель Tbc) при этих аутоиммунных заболеваниях могут играть роль Т-клетки, направленные против определенных антигенов возбудителей (например, стрептококков), которые перекрестно реагируют с (гомологичными) аутоантигенами (молекулярная мимикрия). Реакции V типа, напротив, вызываются среди прочего посредством активирующих или блокирующих аутоантител рецепторов гормонов (например, тиреотропина при базедовой болезни) или нейротрансмиттеров (например, ацетилхолина при тяжелой псевдопаралитической миастении).
Сравнимой с отторжением трансплантата, правда, в обратном смысле является реакция «трансплантат против хозяина» (GHVD), которая встречается у примерно 40% реципиентов в ходе трансплантаций аллогенного костного мозга (от генетически не идентичных индивидуумов). В течение острой фазы, продолжающейся до трех месяцев, Т-клетки донора, пересаженные вместе со стволовыми клетками, атакуют организм хозяина, возникающая в результате при этих обстоятельствах тяжелая воспалительная реакция проявляется прежде всего в коже, в желудочно-кишечном тракте и в печени.
Для лечения острых воспалительных заболеваний в зависимости от предполагаемых причин используют, как правило, нестероидные противовоспалительные средства (НПВС, среди прочего, ингибирование синтеза простагландина) и/или противоинфекционную (для уничтожения бактерий, грибов или паразитов) или противовирусную химиотерапию, также возможно местное применение глюкокортикоидов (общее ингибирование экспрессии генов). При серьезных или хронических рецидивирующих воспалительных заболеваниях системно применяют глюкокортикоиды, или иммуносупрессоры (ингибирование активации Т-клеток), или цитостатики, такие как метотрексат. Это используют также при трансплантации органов или костного мозга. Несмотря на их неоспоримое терапевтическое значение, системное применение упомянутых выше лекарственных средств, в особенности при длительном применении, может вызвать тяжелые побочные действия. Например, у 25% пациентов, принимающих метотрексат в течение двух или более лет, развивается тяжелый цирроз печени. Новые активные вещества, в особенности назначаемые при хронических рецидивирующих воспалительных заболеваниях, блокируют провоспалительное действие ФНОα-антитела против самого цитокина или его рецептора, низкомолекулярные антагонисты рецептора, такие как растворимые рецепторные белки, полученные рекомбинантным способом, которые захватывают цитокин. Однако возрастает количество ссылок на повышенное число инфекционных заболеваний при лечении рецепторными белками (среди них туберкулез), а примерно 40%, по-видимому, совсем не реагируют на лечение (не восприимчивы). Кроме того, для разрешенных гуманизированных ФНОα-антител имеется соответствующее предупреждение относительно появления инфекций вплоть до сепсиса через 2-4 года после начала лечения. Более того, противопоказано совместное применение обоих лекарственных препаратов при обострении воспаления. В дополнение, низкомолекулярные антагонисты рецепторов, разрешенные для лейкотриенов, которые прежде всего находят применение при лечении астмы, а также ингибиторы циклооксигеназы-2 - новый класс нестероидных противовоспалительных средств (НПВП) со значительно сниженными желудочно-кишечными побочными явлениями по сравнению с классическими НПВП. Более того, имеется ряд дополнительных подходов, как правило, на основе гуманизированных антител или олигонуклеотидов против молекул адгезии лейкоцитов или клеток эндотелия, цитокиновых рецепторов Т-хелперов или lgE-антител, которые находятся на различных фазах клинического испытания. Чтобы отказаться от глюкокортикоидов и противоинфекционных средств как группы, упомянутые лекарства в целом должны быть направлены специфично против терапевтически релевантной молекулы-мишени.
Таким образом, задача настоящего изобретения заключается в том, чтобы предложить средство для предотвращения или лечения выраженных воспалительных реакций и ассоциированных с ними последствий заболеваемости и смертности пораженных пациентов.
Данная задача решена посредством объекта, определенного в формуле изобретения.
Данное изобретение поясняется подробнее следующими фигурами.
На фиг.1 показано ингибирование цитокин-стимулированной экспрессии CD40 (а, с, d и е), Е-селектина и МСР-1 (а) и индукция лигандом CD40 экспрессии интерлейкина-12р40 (b) в культивируемых клетках эндотелия человека посредством нейтрализации транскрипционного фактора STAT-1 с помощью соответствующей цис-элемент ловушки (SEQ ID NO: 33). (а) Репрезентативный анализ ПЦР с обратной транскриптазой экспрессии м-РНК Е-селектина, МСР-1 и CD40 (включая денситометрический анализ («интенсивность»), данный в процентах от стимулированного контроля и отнесенный к внутреннему стандарту EF-1) в клетках эндотелия, которые предварительно инкубировали в течение 4 часов со STAT-1 (SEQ ID NO: 33) или NF-κВ цис-элемент ловушкой (10 мкмоль) и затем инкубировали в течение 9 часов с 100 Е/мл фактора некроза опухоли-α и 1000 Е/мл интерферона-γ. (b) Репрезентативный анализ экспрессии мРНК интерлейкина-12р40 путем полимеразной цепной реакции (ПЦР) с обратной транскриптазой (включая денситометрический анализ («интенсивность»), данный в процентах от стимулированного контроля и отнесенный к внутреннему стандарту гр132) в клетках эндотелия, которые предварительно инкубировали в течение 4 часов со STAT-1 цис-элемент ловушкой (10 мкмоль; SEQ ID NO: 33) с последующей инкубацией в течение 12 часов с примерно 670000 Р3×ТВ.А7-клетками/мл (эти клетки миеломы мышей стабильно экспрессируют лиганды к CD40 человека - CD154) и 1000 Е/мл интерферона-γ. (с) Репрезентативный анализ экспрессии мРНК CD40 путем ПЦР с обратной транскриптазой (включая денситометрический анализ («интенсивность»), данный в процентах от стимулированного контроля и отнесенный к внутреннему стандарту EF-1) в клетках эндотелия, которые в течение 4 часов предварительно инкубировали со STAT-1 цис-элемент ловушкой (SEQ ID NO: 33) или с соответствующим контрольным олигонуклеотидом (STAT-1-25mut) (концентрация 10 мкмоль) с последующей инкубацией в течение 9 часов с 100 Е/мл фактора некроза опухоли-α и 1000 Е/мл интерферона-γ. (а) Статистическое обобщение 5 независимых экспериментов по действию STAT-1 цис-элемент ловушки (SEQ ID NO: 33) на цитокин-стимулированную экспрессию мРНК CD40 в культивированных клетках эндотелия (*Р<0,05 против стимулированных контрольных клеток), (е) Репрезентативный Вестерн-блот анализ, включая денситометрический анализ («интенсивность», данная в процентах от стимулированного контроля и отнесенная к внутреннему стандарту β-актину) действия STAT-1 цис-элемент ловушки (SEQ ID NO: 33) на цитокин-стимулированную экспрессию белка CD40 в культивированных клетках эндотелия через 24 часа. Сравнимые результаты были получены в дальнейших экспериментах.
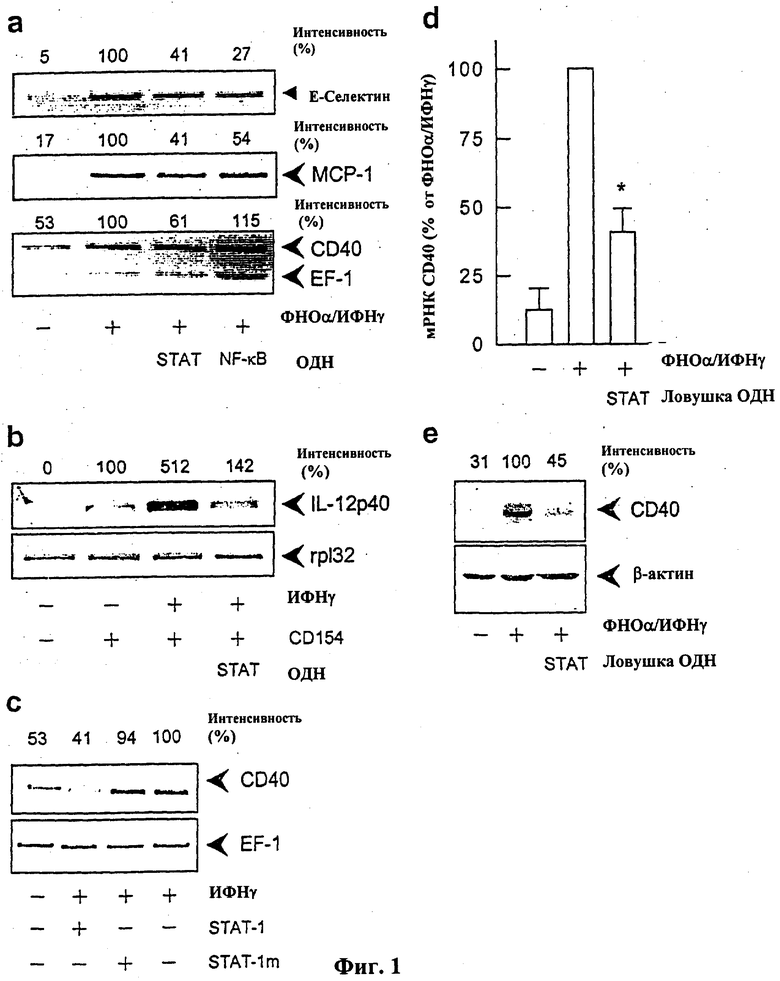
На фиг.2 показано ингибирование цитокин-индуцированной экспрессии гена CD40 в культивированных клетках эндотелия человека посредством основанной на антисмысловом олигонуклеотиде регуляции экспрессии транскрипционного фактора STAT-1 по механизму отрицательной обратной связи, (а) Экспрессия белков CD40 и STAT-1 в состоянии покоя и после 14-часовой инкубации клеток с 100 Е/мл фактора некроза опухоли-α и 1000 Е/мл интерферона-γ. На левой половине чертежа показано статистическое обобщение 2-4 экспериментов с различными сериями клеток, на правой половине чертежа показан в каждом случае репрезентативный Вестерн-блот анализ, включая денситометрический анализ («интенсивность»), данный в процентах от стимулированного контроля и отнесенный к внутреннему стандарту β-актину (*Р<0,05 против нестимулированных клеток). (b) Сравнительное ингибирование экспрессии белков CD40 и STAT-1 в стимулированных клетках эндотелия посредством 24-часовой предварительной обработки STAT-1 антисмысловым олигонуклеотидом (1 мкмоль; SEQ ID NO: 33). Обобщение 2 экспериментов (левая половина чертежа; *Р<0,05 против стимулированных контрольных клеток) и репрезентативный Вестерн-блот анализ (правая половина чертежа).
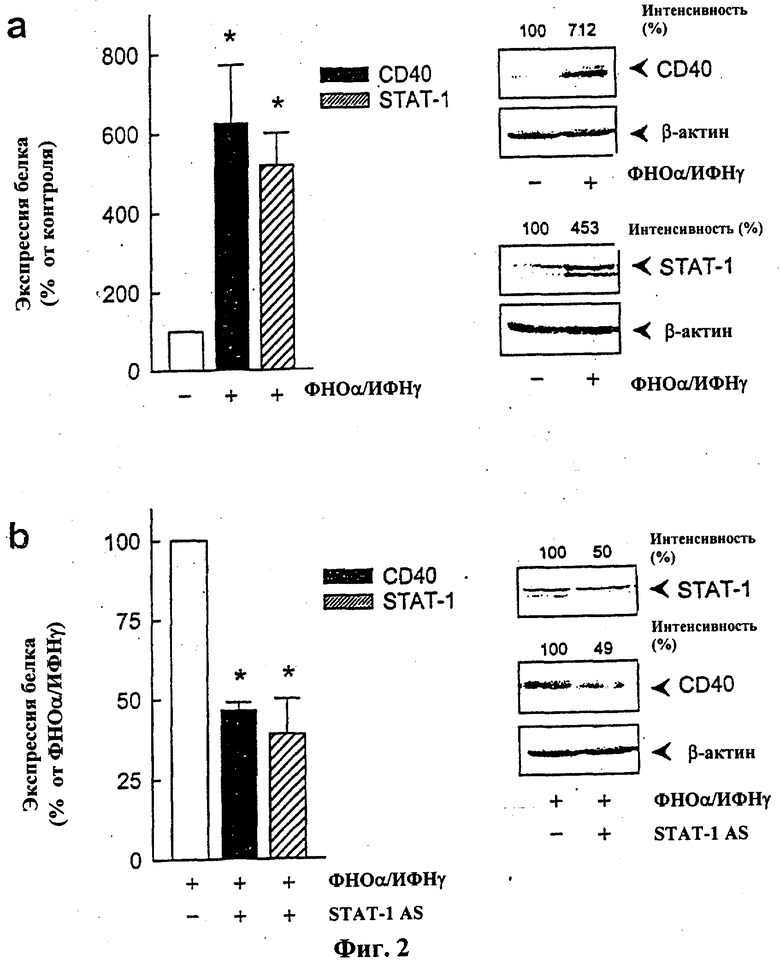
На фиг.3 показано ингибирование экспрессии транскрипционного фактора IRF-1 в клеточной линии моноцитов ТНР-1 (а), а также индуцибельной изоформы NO-синтазы в культивированных клетках гладких мышц человека (b) посредством нейтрализации транскрипционного фактора STAT-1 с помощью соответствующей цис-элемент ловушки (SEQ ID NO: 33). (а) Репрезентативный Вестерн-блот анализ, включая денситометрический анализ («интенсивность»), данный в процентах от стимулированного контроля и отнесенный к внутреннему стандарту β-актину. Культивированные ТНР-1 клетки предварительно инкубировали в течение 4 часов с цис-элемент ловушкой (10 мкмоль) и затем инкубировали в течение 3 часов с 100 Е/мл фактора некроза опухоли-α и 1000 Е/мл интерферона-γ. (b) Левая половина чертежа: статистическое обобщение 3 экспериментов с различными линиями культивированных клеток гладкой мускулатуры человека, которые предварительно инкубировали в течение 4 часов со STAT-1 (SEQ ID NO: 33), NF-κВ или GATA-1 цис-элемент ловушкой (10 мкмоль) и затем инкубировали в течение 9 часов с 1000 Е/мл интерферона-γ, 60 Е/мл интерлейкина-1р, 100 Е/мл фактора некроза опухоли-а и 1 мкг/мл бактериального липополисахарида. Анализ экспрессии мРНК для индуцибельной изоформы NO-синтазы путем ПЦР с обратной транскриптазой (*Р<0,05 против стимулированных клеток =100%). Правая половина чертежа: статистическое обобщение 3 экспериментов с различными клеточными линиями и репрезентативный Вестерн-блот анализ ингибирования цитокин-стимулированной экспрессии белков NO-синтазы (через 20 часов экспозиции) посредством предварительной инкубации со STAT-1 (SEQ ID NO: 33) и NF-κВ цис-элемент ловушкой (*Р<0,05 против стимулированных клеток = 100%).
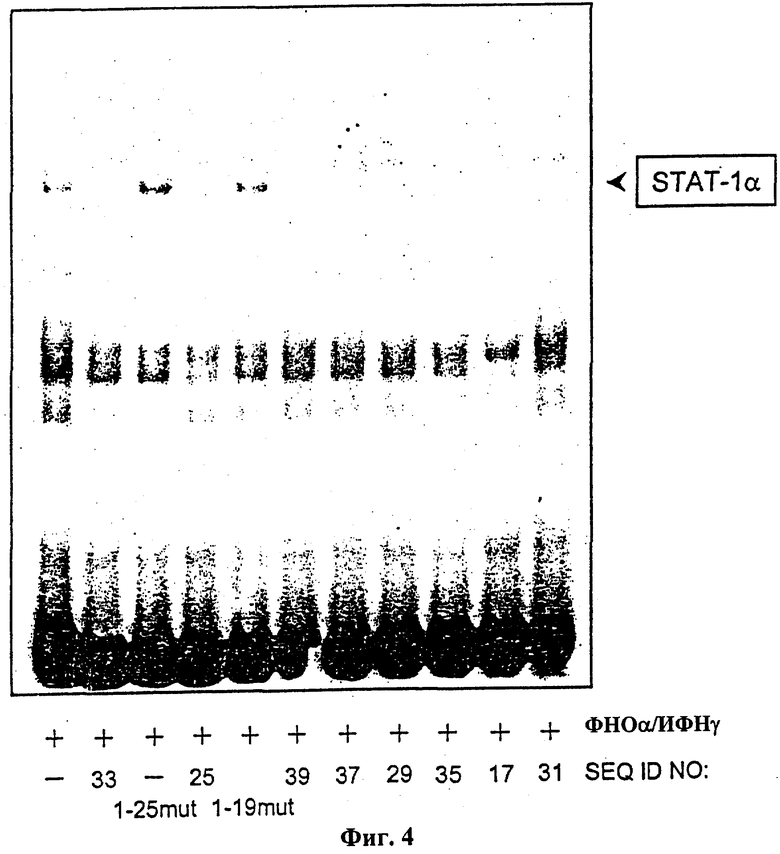
На фиг.4 показана нейтрализация эндогенного STAT-1 в клеточном ядерном экстракте клеточной линии моноцитов ТНР1 посредством различных цис-элемент ловушек (SEQ ID NO: 17, 25, 29, 31, 33, 35, 37, 39 и мутированные контрольные олигонуклеотиды STAT-1-19 mut и STAT-1-25 mut). Репрезентативный анализ изменения электрофоретической подвижности (EMSA). Культивированные ТНР-1 клетки инкубировали 3 часа с 100 Е/мл фактора некроза опухоли-α и 1000 Е/мл интерферона-γ и далее использовали для приготовления ядерного экстракта. Ядерный экстракт клеток инкубировали вместе с двунитевым SIE-олигонуклеотидом, меченным [^Р] (Santa Cruz Biotechnologie, Heidelberg, Германия), и соответствующей цис-элемент ловушкой или контрольным олигонуклеотидом в течение 20 минут при комнатной температуре и в дальнейшем подвергали анализу изменения электрофоретической подвижности.
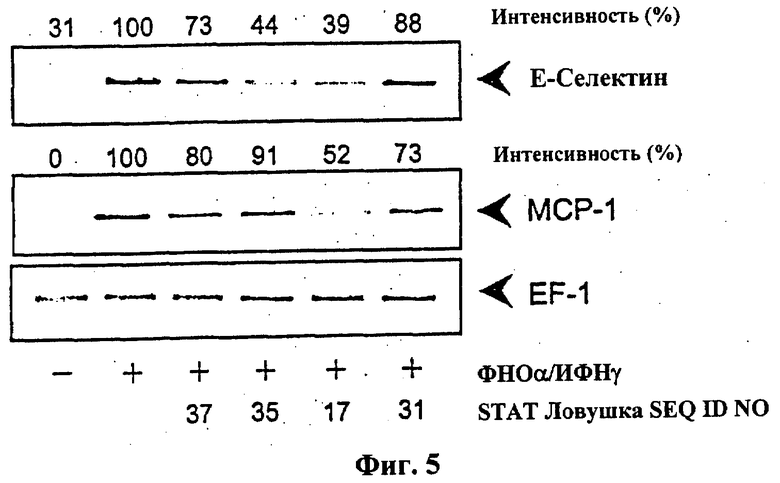
На фиг.5 показан эффект выбранных STAT-1 цис-элемент ловушек (SEQ ID NO: 17, 31, 35, 37) на экспрессию мРНК Е-селектина и МСР-1 в клетках гладкой мускулатуры человека из вены тимуса. Культивированные клетки (Passage 2) предварительно инкубировали 4 часа с соответствующей цис-элемент ловушкой и затем инкубировали 9 часов с 100 Е/мл фактора некроза опухоли-α и 1000 Е/мл интерферона-γ. Репрезентативный анализ ПЦР с обратной транскриптазой, сравнимые результаты были получены в дальнейших экспериментах.
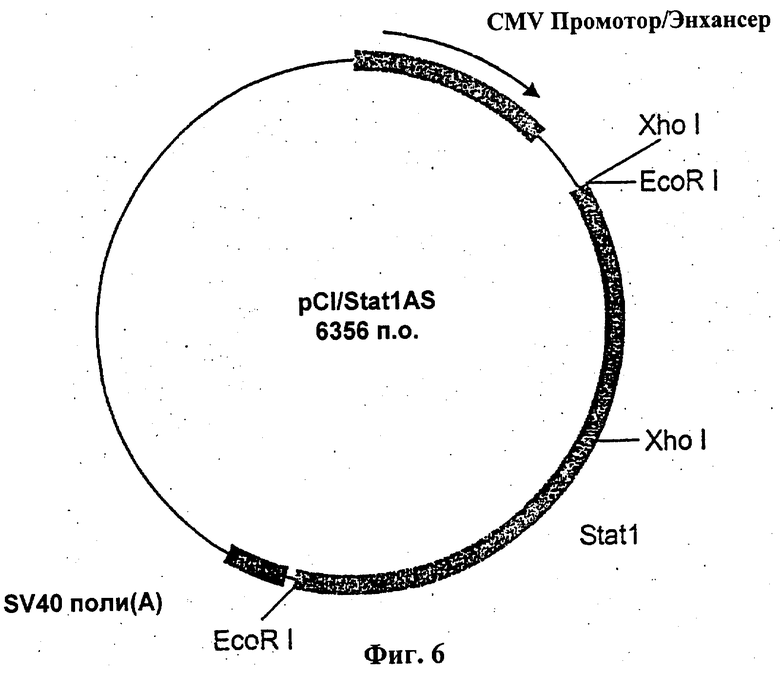
На фиг.6 схематически показана структура антисмыслового вектора экспрессии STAT-1 pCI/ STAT-1 AS в виде генетической карты.
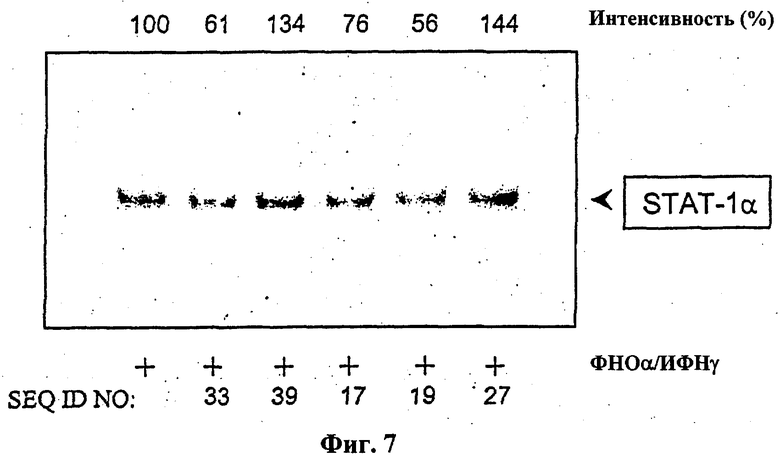
На фиг.7 показан результат нейтрализации STAT-1 в культивированных клетках эндотелия человека посредством различных цис-элемент ловушек (SEQ ID NO: 17, 19, 27, 33 и 39). Репрезентативный анализ EMSA, включая денситометрический анализ («интенсивность»). Культивированные клетки эндотелия инкубировали 4 часа с олигонуклеотидами-ловушками (10 мкмоль/л) и затем стимулировали в течение 30 минут 100 Е/мл фактора некроза опухоли-α и 1000 Е/мл интерферона-γ. Для анализа EMSA использовали ядерный экстракт стимулированных клеток и двунитевые SIE-олигонуклеотиды, меченные [32Р] (Santa Cruz Biotechnologie, Heidelberg, Германия).
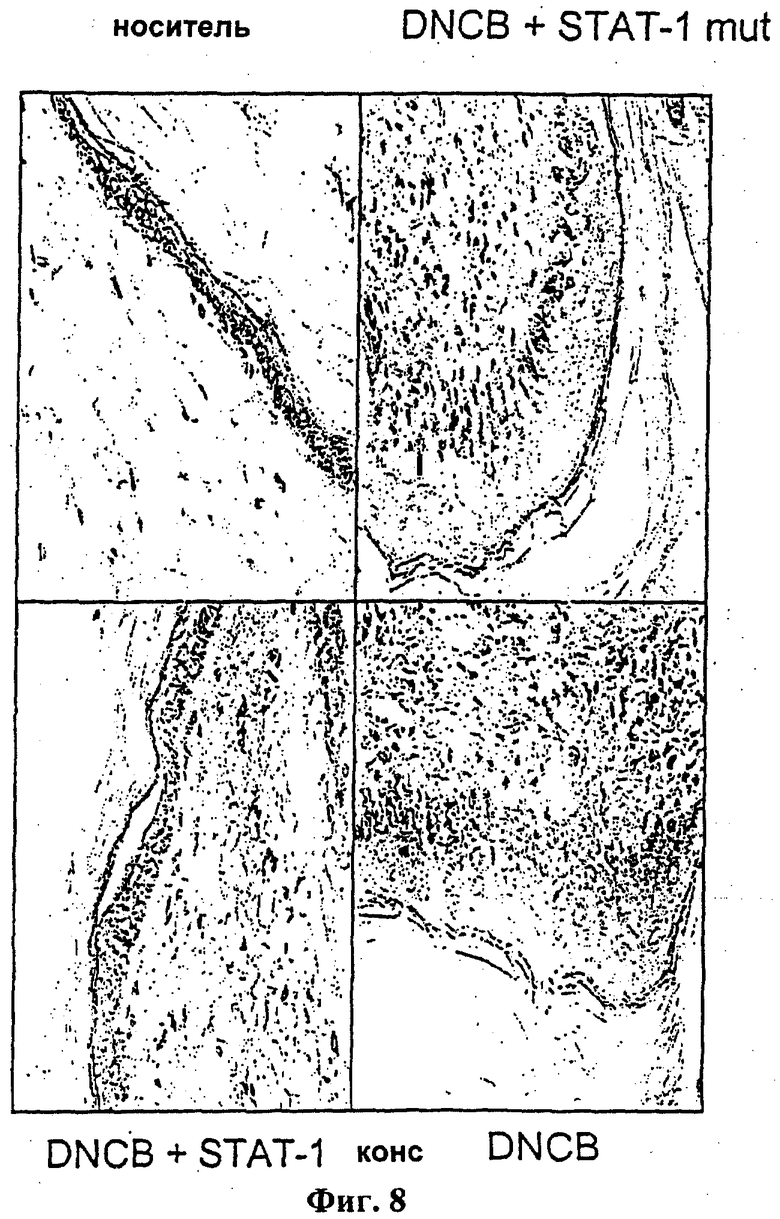
На фиг.8 показан гистологический анализ эффекта STAT-1 олигонуклеотида-ловушки (STAT-1 конс, 10 нмоль, SEQ ID NO: 19), но не мутированные контрольные олигонуклеотиды (STAT-1 mut, 10 нмоль, SEQ ID NO: 61), на DNCB-индуцированный контактный дерматит у самцов морской свинки (оригинал ×400, типичный результат для, в общей сложности, 17 подопытных морских свинок).
Авторы изобретения охарактеризовали транскрипционные факторы, принимающие участие в опосредованном цитокинами усилении экспрессии провоспалительных генных продуктов (CD40, Е-селектина, индуцибельной изоформы NO-синтазы, интерлейкина-12[р40], МСР-1) в клетках эндотелия, клетках гладкой мускулатуры человека, а также в моноцитах. При этом показано, что при стимуляции культивированных клеток эндотелия ФНО-а или CD154 в комбинации с ИФН-γ имеет место синергизм между транскрипционными факторами: ядерным фактором кВ (NF-κВ) и переносчиком сигнала и активатором транскрипции-1 (STAT-1). То же самое справедливо и для культивированных клеток гладкой мускулатуры и моноцитов.
ИФН-γ сам по себе оказался способным усиливать экспрессию CD40 в клетках эндотелия человека, но не экспрессию Е-селектина или интерлейкина-12. Для экспрессии обоих этих генных продуктов, которые почти не экспрессируются в клетках эндотелия или экспрессируются не конститутивно, существенной является одновременная стимуляция клеток ФНО-α (для Е-селектина) и CD154 (для интерлейкина-12). К тому же, для опосредованного ИФН-γ усиления экспрессии CD40, но не Е-селектина, в клетках эндотелия и моноцитах необходима de novo экспрессия еще одного транскрипционного фактора - интерферон-регулирующего фактора-1 (IRF-1). В рамках этих исследований показано, что экспрессия белка IRF-1 при моностимуляции клеток ИФНγ, и в особенности ФНОα, существенно слабее, чем в присутствии обоих цитокинов. Также при транскрипции гена IRF-1 транскрипционные факторы NF-κВ и STAT-1, следовательно, вместе действуют синергически (Ohmori et al., J. Biol. Chem., (1997), 272, 14899).
STAT-1 (GenBank Accession Number NM007315 и ХМ010893, а также http://transfac.gbf.de/cgi-bin/Qt/getEntrv.pl7t0149) относится к группе транскрипционных факторов, содержащей по меньшей мере 6 членов. Продукт гена STAT-1 экспрессируется в большинстве клеток конститутивно, но, как правило, находится в цитоплазме в виде неактивного мономерного белка (размером 91 кДа). Тирозин-фосфорилирование этой р91-субъединицы и последующее объединение (димеризация) двух таких р91-субъединиц (называемых STAT-1α) делает возможным транспорт теперь активного транскрипционного фактора в ядро клетки. Гетеродимеризация с р84-субъединицей STAT-1 p (дифференциально сплайсированным продуктом того же гена) также является возможной. Фосфорилирование конститутивно присутствующих субъединиц осуществляется посредством цитоплазматических Janus-киназ в зависимости от стимула. Обе Janus-киназы (Jak1 и Jak2) стимулирует ИФНα (лучше привлекаемый на рецептор интерферона); (пато)физиологический важнейший стимул для активации STAT-1 ИФНγ, напротив, стимулирует только Jak2. Также.STAT-1 активируют различные факторы роста и пептидные гормоны (например, ангиотензин II); кроме внутренних (фактор роста) рецептор-тирозинкиназ здесь также играет роль митоген-активируемая протеинкиназа (МАР-киназа). В противоположность STAT-1α STAT-1β не обладает трансактивирующей, то есть стимулирующей экспрессию генов активностью.
STAT-1 участвует в экспрессии ряда потенциальных провоспалительных генных продуктов в лейкоцитах, клетках эндотелия и клетках гладкой мускулатуры, где, как правило, активация транскрипционных факторов происходит зависимым от ИФНγ образом. Исключением является, в особенности, зависимая от STAT-1 экспрессия интерлейкина-6 в клетках гладкой мускулатуры, стимулированных ангиотензином II (Schieffer et al., Circ Res, (2000), 87, 1195). Активность белков, к которым также относится STAT-1, может быть ингибирована различными способами. Для этого могут быть использованы, например, анти-STAT-l-антитела, а также природные или синтетические вещества, которые ослабляют взаимодействие STAT-1 с ДНК, то есть ослабляют трансактивирующую активность. Кроме того, можно ингибировать сигнальный путь, который ведет к активации STAT-1 (Jak1, Jak2, рецепторы тирозинкиназ, МАР-киназы). Предпочтительными способами специфического ингибирования активности STAT-1 являются:
1) нейтрализация (подавление) активированного транскрипционного фактора посредством олигонуклеотида-ловушки,
2) ингибирование экспрессии белка STAT-1 с помощью антисмыслового олигонуклеотида и
3) ингибирование экспрессии белка STAT-1 с помощью антисмыслового вектора экспрессии.
Используемое здесь выражение "олигонуклеотид-ловушка" или "цис-элемент ловушка" означает двунитевую молекулу ДНК или двунитевой ДНК-олигонуклеотид. Обе нити ДНК имеют комплементарную последовательность. В настоящем изобретении цис-элемент ловушка имеет последовательность, которая соответствует природной кор-связывающей последовательности STAT-1 в геноме или сходную с ней, и связывается в клетке с транскрипционным фактором STAT-1. Эта цис-элемент ловушка действует, таким образом, как молекула для конкурентного ингибирования (лучше нейтрализации) STAT-1.
Аспект настоящего изобретения состоит в том, чтобы предложить двунитевой ДНК-олигонуклеотид, который способен связываться с транскрипционным фактором STAT-1 специфично к последовательности и имеет одну из следующих последовательностей, причем в каждом случае здесь показана одна нить двунитевого ДНК-олигонуклеотида, а комплементарная нить также содержится:
5'-NTTNCBGDAAN-3' (SEQ ID NO:1),
5'-ATTACCGGAAG-3' (SEQ ID NO:3),
5'-ATTCCGGTAAG-3' (SEQ ID NO:5),
5'-ATTCCTGGAAG-3' (SEQ ID NO:7),
5'-ATTCCTGTAAG-3' (SEQ ID NO:9),
5'-GTTCCAGGAAC-3' (SEQ ID NO:11),
5'-GTTCCCGGAAG-3' (SEQ ID NO: 13),
5'-GTTCCGGGAAC-3' (SEQ ID NO:15),
5'-TGTGAATTACCGGAAGTGAGA-3'(SEQ ID NO: 17),
5'-TGTGAATTACCGGAAGTG-3'(SEQ ID NO:19),
5'-AGTCAGTTCCAGGAACTGACT-3'(SEQ ID NO:21),
5'-ATGTGAGTTCCCGGAAGTGAACT-3' (SEQ ID NO:23),
5'-ACAGTTCCGGGAACTGTC-3'(SEQ ID NO:25),
5'-GACAGTTCCGGGAACTGTC-3(SEQ ID NO:27),
5'-GTGTATTCCGGTAAGTGA-3'(SEQ ID NO:29),
5'-TTATGTGAATTCCTGGAAGTG-3'(SEQ ID NO:31),
5'-CATGTTATGCATATTCCTGTAAGTG-3'(SEQ ID NO:33),
5'-TGTGAATTCCTGTAAGTGAGA-3'(SEQ ID NO:35),
5'-TGCATATTCCTGTAAGTG-3' (SEQ ID NO:37),
5'-ATATTCCTGTAAGTG-3'(SEQ ID NO:39).
Когда используют олигонуклеотид-ловушку против STAT-1 по настоящему изобретению, но не соответствующий контрольный олигонуклеотид, в клетках эндотелия человека, цитокин-индуцированная экспрессия CD40 (и при моностимуляции ИФКγ, и при комбинации ИФНγ и ФНОα) ингибируется значительно более чем на 50%. Это справедливо также для экспрессии Е-селектина и МСР-1, а также интерлейкина-12 (р40) при стимуляции клеток ИФНγ и ФНОα, а также CD154. Выключение активности STAT-1 в соответствии с этим приводит к высокозначимому ингибированию экспрессии группы провоспалительных генных продуктов в клетках эндотелия человека. Поэтому при таком терапевтическом подходе нужно учитывать значительное ослабление взаимодействия эндотелий-лейкоциты (Е-селектин, МСР-1), а также взаимодействия антиген-презентирующих клеток (например, макрофагов и В-лимфоцитов) с Т-лимфоцитами (CD40, интерлейкин-12) при воспалительных заболеваниях. По смыслу это также справедливо и для показанного ослабления цитокин-индуцированной экспрессии IRF-1 в ТНР-1 моноцитах (и, следовательно, последовательной экспрессии IRF-1-зависимых генов), а также цитокин-индуцированной экспрессии упомянутых генных продуктов, включая индуцибельную МО-синтазу в клетках гладкой мускулатуры человека.
Предпочтительным способом специфического ингибирования активности STAT-1 является, таким образом, использование двунитевых ДНК-олигонуклеотидов по настоящему изобретению, также называемых цис-элемент-ловушкой или олигонуклеотид-ловушкой, которые содержат сайт связывания для STAT-1. Экзогенное введение в клетку большого количества сайтов связывания транскрипционного фактора, в особенности гораздо большего количества, чем в геноме, приводит к ситуации, в которой большое количество определенного транскрипционного фактора специфично связывается с соответствующей цис-элемент ловушкой, а не с эндогенными сайтами связывания - мишенями. Этот подход к ингибированию связывания транскрипционных факторов с их эндогенными сайтами связывания также называется подавлением (squelching). Подавление (или также нейтрализация) транскрипционных факторов при использовании цис-элемент ловушек было успешно использовано для ингибирования роста клеток. Для этого использовали фрагменты ДНК, содержащие специфичные сайты связывания транскрипционных факторов для транскрипционного фактора E2F (Morishita et al., PNAS, (1995) 92, 5855).
Последовательность нуклеиновой кислоты, которую применяют для предотвращения связывания транскрипционного фактора STAT-1, например, представляет собой последовательность, с которой STAT-1 связывается в клетке естественным образом. STAT-1 специфично связывается с мотивом с последовательностью 5'-NNNSANTTCCGGGAANTGNSN-3', где N=А, Т, С или G, и S=С или G. Для эффективного связывания STAT-1 важно точное соответствие с подчеркнутыми основаниями и расстоянием между этими основаниями. Поэтому цис-элемент ловушка по настоящему изобретению может иметь следующую 11-членную консенсусную кор-связывающую последовательность: 5'-NTTNCBGDAAN-3' (SEQ ID NO:1), где В=С, G или Т, D=А, G или Т, и N = А, Т, С или G. Цис-элемент ловушка может, кроме того, быть длиннее, чем 11-членная кор-связывающая последовательность, и может быть удлинена на 5'-конце и/или на 3'-конце. Соответствующие мутации в пределах кор-связывающей последовательности приводят к потере связывания STAT-1 с олигонуклеотидом-ловушкой.
Поскольку цис-элемент ловушка представляет собой двунитевую нуклеиновую кислоту, ДНК-олигонуклеотид по настоящему изобретению содержит в каждом случае не только смысловую или прямую последовательность, но и комплементарную антисмысловую или обратную последовательность. Предпочтительный ДНК-олигонуклеотид по настоящему изобретению содержит 11-членную кор-связывающую последовательность для STAT-1, такую как последовательность, содержащаяся в SEQ ID NO:3 по SEQ IDNO:16.
Цис-элемент ловушка также может иметь последовательность, отличающуюся от предыдущей последовательности и быть длиннее, чем 11-членная. Особенно предпочтительными являются последовательности, которые содержатся в SEQ ID NO:17 no SEQ ID NO:40. Такие цис-элемент ловушки содержат в каждом случае по два сайта связывания для STAT-1.
Выражение «2 сайта связывания» относится к смысловой и антисмысловой нити. Этим перечень предпочтительных последовательностей не ограничивается. Специалисту очевидно, что в качестве ингибиторов STAT-1 может быть использовано множество последовательностей, пока они проявляют указанные выше условия 11-членной консенсусной кор-связывающей последовательности и аффинность к STAT-1.
Аффинность связывания последовательности нуклеиновой кислоты со STAT-1 может быть определена путем анализа изменения электрофоретической подвижности (EMSA) (Sambrook et al. (1989) Molecular Cloning. Cold Spring Harbor Laboratory Press; Krzesz et al. (1999) FEBS Lett. 453, 191). Эта тест-система подходит для контроля качества нуклеиновых кислот, которые предназначены для использования в способе по настоящему изобретению, или для определения оптимальной длины сайта связывания. Она также подходит для идентификации других последовательностей, которые связываются со STAT-1. Для EMSA, предназначенного для выделения новых сайтов связывания, больше всего подходят очищенные или рекомбинантные экспрессируемые варианты STAT-1, которые используют в нескольких чередующихся циклах ПЦР-амплификации и отбора посредством EMSA (Thiesen und Bach (1990) Nucleic Acids Res. 18, 3202).
Гены, для которых известно, что в их промоторных или энхансерных участках содержатся сайт связывания STAT-1, или для которых уже имеется функциональное доказательство значения STAT-1 для их экспрессии и которые поэтому являются предполагаемыми мишенями для специфичного подавления согласно способу по настоящему изобретению, представляют собой, помимо генов CD40, Е-селектина, индуцибельной NO-синтазы, интерлейкина-12 (р40) и МСР-1, и другие провоспалительные гены, например, ИФНγ самого по себе, цитокина интерлейкин-6, молекул адгезии 1САМ-1, РЕСАМ-1 (эндотелиальная молекула клеточной адгезии тромбоцитов - 1), RANTES (регулируемые активацией, экспрессируемые нормальными Т-клетками; предположительно секретируемые; секретируются Т-лимфоцитами в растворенном виде) и VCAM-1, хемокина интерлейкин-8, IP-10 (интерферон-индуцибельный белок-10) и Mig (монокин, индуцируемый гамма-интерфероном), а также МНС-белков I и II. При этом не играет никакой роли, регулируется ли экспрессия этого гена прямо или опосредованно через STAT-1 (например, через зависимую от STAT-1 экспрессию IRF-1).
Способ по настоящему изобретению модулирует транскрипцию гена или генов таким образом, что этот ген или эти гены, например, Е-селектина не экспрессируются вовсе или экспрессируются слабее. Сниженная или подавленная экспрессия в объеме настоящего изобретения означает, что уровень транскрипции снижен по сравнению с клетками, которые не обработаны двунитевым ДНК-олигонуклеотидом по настоящему изобретению. Такое снижение может быть установлено, например, посредством Нозерн-блоттинга (Sambrook et al., 1989) или ПЦР с обратной транскриптазой (Sambrook et al., 1989). Такое снижение, как правило, является по меньшей мере 2-кратным, предпочтительно по меньшей мере 5-кратным, еще более предпочтительно по меньшей мере 10-кратным. Потеря активации может быть достигнута, например, когда STAT-1 в качестве транскрипционного фактора воздействует на определенный ген, и, таким образом, это подавление активатора приводит к потере экспрессии гена-мишени.
Более того, способ по настоящему изобретению делает возможным отмену ингибирования экспрессии гена, поскольку эта экспрессия блокируется постоянно активным или (после соответствующей стимуляции клетки) активированным транскрипционным фактором. Примером этого является отмена ингибирования экспрессии гена препро-эндотелина-1 в нативных клетках эндотелия кролика V. jugularis посредством цис-элемент ловушки против белка, связывающего транскрипционный фактор ССААТ/энхансер (Lauth et al., J. Mol. Med., (2000), 78, 441). Таким же способом может быть осуществлена отмена ингибирования экспрессии генов, продукты которых оказывают защитное действие, например, против воспалительных заболеваний. Таким образом, например, осуществляется регуляция по типу отрицательной обратной связи посредством ИФНγ эндотелиальной изоформы МО-синтазы, продукт NO которой играет решающую роль при подавлении экспрессии провоспалительных молекул адгезии и хемокинов в клетках эндотелия (Rosenkranz-Weiss et al., (1994) J. Clin. Invest. 93, 1875). Цис-элемент ловушка против STAT-1 может отменить этот нежелательный эффект путем предотвращения связывания STAT-1 с соответствующим сайтом связывания в промоторе гена эндотелиальной NO-синтазы.
Олигонуклеотиды, как правило, быстро разрушаются эндо- и экзонуклеазами, в частности ДНКазами и РНКазами, в клетке. Вследствие этого ДНК-олигонуклеотиды могут быть модифицированы для стабилизации от разрушения, так чтобы в течение длительного времени в клетке сохранялась высокая концентрация олигонуклеотида. Обычно такая стабилизация может быть получена путем введения одной или более чем одной модифицированной межнуклеотидной связи.
Успешным образом стабилизированный ДНК-олигонуклеотид содержит модификацию не обязательно в каждой межнуклеотидной связи. Предпочтительно, чтобы межнуклеотидные связи были модифицированы на соответствующих концах обоих олигонуклеотидов цис-элемент ловушки. При этом могут быть модифицированы последние шесть, пять, четыре, три, две или последняя либо одна или более чем одна межнуклеотидная связь в пределах последних шести межнуклеотидных связей. Кроме того, могут быть введены различные модификации межнукпеотидных связей в нуклеиновой кислоте, и полученные таким образом двунитевые ДНК-олигонуклеотиды тестируют на специфичное к последовательности связывание со STAT-1, используя рутинные тест-системы EMSA. Эта тест-система позволяет определить константу связывания цис-элемент ловушки и таким образом определить, изменилась ли аффинность вследствие модификации. Модифицированные цис-элемент ловушки, которые все еще обладают достаточным связыванием, могут быть отобраны, при этом достаточное связывание означает по меньшей мере приблизительно 50% или по меньшей мере приблизительно 75% и особенно предпочтительно приблизительно 100% от связывания немодифицированной нуклеиновой кислоты.
Цис-элемент ловушки с модифицированными межнуклеотидными связями, которые все еще проявляют достаточное связывание, могут быть проверены в отношении того, являются ли они более стабильными в клетке по сравнению с немодифицированными цис-элемент ловушками. Клетки, «трансфицированные» цис-элемент ловушками по настоящему изобретению, проверяют в различные моменты времени в отношении количества все еще присутствующих цис-элемент ловушек. При этом предпочтительно используют цис-элемент ловушку, меченную флуоресцентным красителем (например, Техасский красный), или цис-элемент ловушку с радиоактивной меткой (например, 32Р) с последующей цифровой флуоресцентной микроскопией и авторадиографией или сцинтиграфией. Успешно модифицированная цис-элемент ловушка имеет период полужизни в клетке больше, чем у немодифицированных цис-элемент ловушек, предпочтительно по меньшей мере приблизительно 48 часов, более предпочтительно приблизительно 4 суток и наиболее предпочтительно по меньшей мере приблизительно 7 суток.
Подходящие модифицированные межнуклеотидные связи обобщены в Uhlmann und Peyman ((1990) Chem. Rev. 90, 544). Модифицированные межнуклеотид-фосфатные остатки и/или не фосфорные мостики в нуклеиновой кислоте, которую можно использовать в способе по настоящему изобретению, содержат, например, метилфосфат, фосфоротиоат, фосфородитиоат, фосфорамидат, фосфатный сложный эфир, в то время как межнуклеотидные аналоги, не содержащие фосфора, содержат, например, силоксановые мостики, карбонатные мостики, карбоксиметилэфирные мостики, ацетамидатные мостики и/или тиоэфирные мостики. При использовании фосфоротиоат-модифицированных межнуклеотидных связей они предпочтительно не должны находиться между основаниями цитозином и гуанином, так как это может привести к активированию клеток-мишеней цис-элемент ловушки.
Еще одним воплощением данного изобретения является стабилизация нуклеиновых кислот путем введения в нуклеиновые кислоты структурных особенностей, которые увеличивают период полужизни нуклеиновой кислоты. Такие структуры, содержащие ДНК-шпильки и колоколообразные ДНК, описаны в US 5683985. Одновременно с вышеуказанными структурами могут быть введены модифицированные межнуклеотид-фосфатные остатки и/или не фосфатные мостики. Полученные таким образом нуклеиновые кислоты могут быть проверены в отношении связывания и стабильности с помощью тест-системы, описанной выше.
Кор-связывающая последовательность может находиться не только в цис-элемент ловушке, но также в векторе. Предпочтительной формой воплощения вектора является плазмидный вектор, и в особенности плазмидный вектор, способный реплицироваться аутосомно, посредством этого увеличивая стабильность введенной двунитевой нуклеиновой кислоты.
Цис-элемент ловушка по настоящему изобретению быстро проникает в клетку. Достаточное проникновение характеризуется модулированием экспрессии одного или более чем одного гена, которые находятся под контролем STAT-1. Цис-элемент ловушка по настоящему изобретению предпочтительно модулирует транскрипцию одного гена или генов приблизительно через 4 часа после контакта с клеткой, более предпочтительно приблизительно через 2 часа, приблизительно через 1 час, приблизительно через 30 минут и наиболее предпочтительно приблизительно через 10 минут. Типичная смесь, которую используют в таком эксперименте, содержит 10 мкмоль/л цис-элемент ловушки.
Кроме того, настоящее изобретение относится к применению цис-элемент ловушки по настоящему изобретению для производства лекарства. Настоящее изобретение также относится к применению цис-элемент ловушки по настоящему изобретению для производства лекарства для предупреждения или терапии сердечно-сосудистых осложнений (например, рестеноза после чрескожной ангиопластики, стеноза венозных шунтов), отторжения трансплантата, болезни "трансплантат против хозяина" (GVHD), ишемических/реперфузионных повреждений при хирургических операциях, иммунологических реакций гиперчувствительности (тип I-IV), аутоиммунных заболеваний (например, сахарного диабета, рассеянного склероза, ревматоидного артрита), а также всех других форм острых, подострых и хронических воспалительных заболеваний, в особенности суставов (например, артрит), органов дыхания (например, бронхиальная астма, хронический бронхит), кожи (например, псориаз, нейродермит) и желудочно-кишечного тракта (например, неспецифический язвенный колит, болезнь Крона), включая септический шок.
Настоящее изобретение относится также к способу модулирования транскрипции по меньшей мере одного гена в клетках, в особенности в клетках эндотелия, клетках эпителия, лейкоцитах, клетках гладкой мускулатуры, кератиноцитах или фибробластах, включающему в себя стадию, на которой указанные клетки приводят в контакт со смесью, содержащей одну или более чем одну двунитевую нуклеиновую кислоту по настоящему изобретению, обладающую способностью связываться специфично к последовательности с транскрипционным фактором STAT-1. Предпочтительным способом является, например, обработка ex vivo донорского костного мозга, содержащего Т-лимфоциты, перед пересадкой в организм реципиента.
Цис-элемент ловушки по настоящему изобретению может быть, кроме того, назначен пациентам в составе композиции или использован в способе по настоящему изобретению. Композицию (в дальнейшем называемая смесью), содержащую цис-элемент ловушки по настоящему изобретению, приводят в контакт с клетками-мишенями (например, клетками эндотелия, клетками эпителия, лейкоцитами, клетками гладкой мускулатуры, кератиноцитами или фибробластами). Целью этого приведения в контакт является перенос цис-элемент ловушек, которые связывают STAT-1, в клетку-мишень (то есть, в клетку, которая экспрессирует STAT-1-зависимые провоспалительные генные продукты). Поэтому в объеме настоящего изобретения могут быть использованы модификация нуклеиновой кислоты и/или добавка или вспомогательное вещество, для которых известно, что они увеличивают проникновение через мембрану (Uhlmann und Peyman (1990) Chem. Rev. 90, 544).
Смесь по настоящему изобретению содержит в предпочтительном воплощении только нуклеиновую кислоту и буфер. Подходящая концентрация цис-элемент ловушек находится в диапазоне по меньшей мере от 0,1 до 100 мкмоль, предпочтительно до 10 мкмоль, причем добавлен один или более чем один подходящий буфер. Примером такого буфера является раствор Рингера, содержащий 145 ммоль/л Na+ 5 ммоль/л К+, 156 ммоль/л Cl+, 2 ммоль/л Са+, 1 ммоль/л Mg+, 10 ммоль/л Hepes, 10 ммоль/л D-глюкозы, рН 7,4.
В еще одном воплощении данного изобретения смесь дополнительно содержит по меньшей мере одну добавку и/или вспомогательное вещество. Добавки и/или вспомогательные вещества, такие как липиды, катионные липиды, полимеры, липосомы, наночастицы, аптамеры нуклеиновых кислот, пептиды и белки, которые связываются с ДНК, или синтетические молекулы пептид-ДНК, предназначены для того, чтобы, например, усиливать включение нуклеиновых кислот в клетку, чтобы смесь достигла только одной подгруппы клеток, чтобы препятствовать разрушению нуклеиновой кислоты в клетке, чтобы облегчить хранение смеси нуклеиновых кислот перед использованием. Примерами пептидов и белков или синтетических молекул пептид-ДНК являются, например, антитела, фрагменты антител, лиганды, молекулы адгезии, которые все могут быть модифицированными или немодифицированными.
Добавки, которые стабилизируют цис-элемент ловушки в клетке, представляют собой, например, вещества, конденсирующие нуклеиновую кислоту, такие как катионные полимеры, поли-L-лизин или полиэтиленимин.
Смесь, которую используют в способе по настоящему изобретению, предпочтительно применяют местно путем инъекции, при помощи катетера, суппозитория («свечи»), аэрозоля (ингаляция спреями в нос или ротовую полость), троакара, бомбардирующих частиц (projectiles), гелей из плуроника, полимеров с длительным высвобождением лекарства или любым другим средством, делающим возможным местный доступ. Также ex vivo применение смеси, используемое в способе по настоящему изобретению, обеспечивает местный доступ.
Ингибированию активности STAT-1, однако, можно препятствовать не только на уровне белков в описанном выше способе, а также на него можно воздействовать во время трансляции белка транскрипционного фактора. Ингибирование может быть осуществлено посредством так называемого антисмыслового олигонуклеотида. Антисмысловые олигонуклеотиды могут ингибировать синтез гена-мишени на трех различных уровнях: при транскрипции (предотвращение синтеза гетерогенной ядерной РНК (гяРНК)), при процессинге (сплайсинге) гяРНК в мРНК и при трансляции белка с мРНК на рибосомах. Способ ингибирования экспрессии генов посредством антисмысловых олигонуклеотидов описан в уровне техники и хорошо известен специалистам в данной области. Еще одним аспектом по настоящему изобретению является антисмысловой олигонуклеотид, который специфично ингибирует экспрессию STAT-1 и предпочтительно имеет следующие последовательности:
5'-TACCACTGAGACATCCTGCCAC-3'(SEQ ID NO:41),
5'-AACATCATTGGCACGCAG-3'(SEQ ID NO:42),
5'-GTGAACCTGCTCCAG-3'(SEQ ID NO:43).
Антисмысловой олигонуклеотид может представлять собой однонитевую молекулу ДНК, молекулу РНК или гибридную однонитевую молекулу ДНК/РНК. Антисмысловой олигонуклеотид может также иметь одну или более чем одну модифицированную межнуклеотидную связь, например, такие как описаны выше для цис-элемент ловушки. Особенно следует обратить внимание, что в случае антисмыслового олигонуклеотида, стабилизированного фосфоротиоат-модифицированными межнуклеотидными связями, между основаниями цитозином и гуанином не вводят никакие фосфоротиоат-модифицированные межнуклеотидные связи, так как это приведет к ИФНγ-подобной активации, в особенности, иммунокомпетентных клеток (например, клеток эндотелия) и таким образом расстроит желаемый лечебный эффект, по меньшей мере частично.
Антисмысловой олигонуклеотид может быть введен не только в виде однонитевой молекулы нуклеиновой кислоты, но также может быть представлен в виде вектора. Предпочтительным воплощением является плазмидный вектор, и в особенности плазмидный вектор, способный аутосомно реплицироваться, тем самым увеличивая стабильность введенной однонитевой нуклеиновой кислоты.
Таким образом, еще одним аспектом настоящего изобретения является антисмысловой вектор экспрессии, который после трансфекции в клетки-мишени экспрессируется ими и специфически ингибирует экспрессию STAT-1. При этом может быть применен любой доступный эукариотический вектор экспрессии в соответствии с уровнем техники. Предпочтительно это касается pCI-плазмиды фирмы "Promega" (Каталог No. E1731, GenBank Accession Number U47119), у которой, например, участок гена STAT-1 размером 2350 п.о. (от -121 до +2229, GenBank Accession Number (XM010893) клонирован в обратном направлении (3'→5'). Этот участок STAT-1-гена фланкирован двумя сайтами рестрикции EcoR1 и содержит один сайт рестрикции Xho1. Его экспрессия находится под контролем CMV-промоторов. Вся плазмида целиком (обозначена pCI/Stat1 AS) содержит 6365 п.о.
Ослабление экспрессии белка STAT-1, основанное на антисмысловом олигонуклеотиде, также ингибирует цитокин-индуцированную экспрессию CD40 в клетках эндотелия человека в той же степени, что и олигонуклеотид-ловушка.
Настоящее изобретение также относится к применению антисмысловых олигонуклеотидов по настоящему изобретению для изготовления лекарств. Кроме того, настоящее изобретение относится к применению антисмысловых олигонуклеотидов по настоящему изобретению для изготовления лекарства для предупреждения или терапии сердечно-сосудистых осложнений (например, рестеноза после чрескожной ангиопластики, стеноза венозных шунтов), отторжения трансплантата, болезни «трансплантат против хозяина» (GHVD), ишемических/реперфузионных повреждений при хирургических операциях, иммунологических реакций гиперчувствительности (тип I-IV), аутоиммунных заболеваний (например, сахарного диабета, рассеянного склероза, ревматоидного артрита), а также всех других форм острых, подострых и хронических воспалительных заболеваний, в особенности суставов (например, артрит), органов дыхания (например, бронхиальная астма, хронический бронхит), кожи (например, псориаз, нейродермит) и желудочно-кишечного тракта (например, неспецифический язвенный колит, болезнь Крона), включая септический шок.
Также антисмысловые олигонуклеотиды по настоящему изобретению могут быть использованы в составе композиции и введены пациентам. Композиция может также содержать стабилизирующие добавки или вспомогательные вещества, которые облегчают, например, введение антисмыслового нуклеотида в клетку, нацеливая, например, смесь только на одну подгруппу клеток, предотвращая, например, разрушение антисмыслового олигонуклеотида в клетке или облегчая, например, хранение антисмыслового олигонуклеотида перед использованием.
Следующие фигуры и примеры служат только для пояснения и никаким образом не ограничивают объем изобретения.
1. Клеточная культура
Клетки эндотелия человека выделяли из пупочных вен путем обработки 1,6 Е/мл диспазы в Hepes-модифицированном растворе Тироде в течение 30 мин при 37°С и культивировали на покрытых желатином 6-луночных культуральных планшетах (2 мг/мл желатина в 0,1 М HCI в течение 30 мин при температуре окружающей среды) в 1,5 мл среды М199 (Gibco Life Technologies, Карлсруе, Германия), содержащей 20% фетальную телячью сыворотку, 50 Е/мл пенициллина, 50 мкг/мл стрептомицина, 10 Е/мл нистатина, 5 мМ HEPES и 5 мМ TES, 1 мкг/мл гепарина и 40 мкг/мл эндотелиального фактора роста. Их идентифицировали по их типичной морфологии, напоминающей брусчатку, положительному иммунному окрашиванию для фактора фон Виллебрандта (vWF) и флуориметрической детекции (FACS) PECAM-1 (CD31), а также по отрицательному иммунному окрашиванию для α-актина гладких мышц (Krzesz et al., (1999), FEBS Lett. 453191). Клеточную линию моноцитов человека ТНР-1 (АТСС TIB 202) культивировали в среде RPMI 1640 (Life Technologies), содержащей 10% фетальную телячью сыворотку, 50 Е/мл пенициллина, 50 мкг/мл стрептомицина и 10 Е/мл нистатина. Клетки гладких мышц человека выделяли из препарированной вены тимуса посредством технологии эксплантации (Krzesz et al., (1999), FEBS Lett. 453191) и культивировали на покрытых желатином 6-луночных культуральных планшетах (см. выше) в 1,5 мл модифицированной по Дульбекко среды Игла, содержащей 15% фетальной телячьей сыворотки, 50 Е/мл пенициллина, 50 мкг/мл стрептомицина и 10 Е/мл нистатина. Их идентифицировали по положительному иммунному окрашиванию для α-актина гладких мышц.
2. Анализ путем ПЦР с обратной транскриптазой
Эндотелиальную общую РНК выделяли при помощи набора Qiagen RNeasy Kit (Qiagen, Hilden, Deutschland) с последующим синтезом кДНК с максимальным количеством 3 мкг РНК и 200 Е обратной транскриптазы Superscript™ II (Life Technologies) в общем объеме 20 мкл в соответствии с инструкциями производителя. Для выравнивания нагрузки по кДНК использовали 5 мкл (приблизительно 75 нг кДНК) получающегося в результате раствора кДНК и пары праймеров (Gibco) для ПЦР фактора элонгации 1 (EF-1) с 1 Е Taq ДНК-полимеразы (Gibco) в общем объеме 50 мкл. EF-1 использовали в качестве внутреннего стандарта для ПЦР. Продукты ПЦР разделяли в 1,5% агарозных гелях, содержащих 0,1% этидиум бромид, и интенсивности полос определяли денситометрически при помощи системы камер CCD и One-Dscan Gelanalyse-Software от Scanalytics (Billerica, MA, USA) для приведения в соответствие с объемом кДНК в последующих анализах ПЦР.
Все реакции ПЦР проводили отдельно для каждой пары праймеров в термоциклере Hybaid OmnE (AWG; Heidelberg, Германия). Индивидуальные условия ПЦР для кДНК из клеток эндотелия пупочных вен человека были следующими: CD40 (размер продукта 381 п.о., 25 циклов, температура отжига 60°С, (прямой праймер) 5'-CAGAGTTCACTGAAACGGAATGCC-3' (SEQ ID NO:44), (обратный праймер) 5'-TGCCTGCCTGTTGCACAACC-3' (SEQ ID NO:45)); Е-селектин (размер продукта 304 п.о., 33 цикла, температура отжига 60°С, (прямой праймер) 5'-AGCAAGGCATGATGTTAACC-3' (SEQ ID NO:46), (обратный праймер) 5'-GCATTCCTCTCTTCCAGAGC-3' (SEQ ID NO:47)); EF-1 (размер продукта 220 п.о., 22 цикла, температура отжига 55°С, (прямой праймер) 5'-TCTTAATCAGTGGTGGAAG-3' (SEQ ID NO:48), (обратный праймер) 5'-TTTGGTCAAGTTGTTTCC-3' (SEQ ID NO:49)); ИЛ-12р40 (размер продукта 281 п.о., 30 циклов, температура отжига 62°С, (прямой праймер) 5'-GTACTCCACATTCCTACTTCTC-3' (SEQ ID NO:50), (обратный праймер) 5'-TTTGGGTCTATTCCGTTGTGTC-3' (SEQ ID NO:51)); rpl32 (размер продукта 368 п.о., 20 циклов, температура отжига 60°С, (прямой праймер) 5'-GTTCATCCGGCACCAGTCAG-3' (SEQ ID NO:52), (обратный праймер) 5'-ACGTGCACATGAGCTGCCTAC-3' (SEQ ID NO:53)); MCP-1 (размер продукта 330 п.о., 22 цикла, температура отжига 63°С, (прямой праймер) 5'-GCGGATCCCCTCCAGCATGAAAGTCTCT-3' (SEQ ID NO:54), (обратный праймер) 5'-ACGAATTCTTCTTGGGTTGTGGAGTGAG-3' (SEQ ID NO:55)).
3. Анализ изменения электрофоретической подвижности (EMSA)
Ядерные экстракты и [32-Р]-меченные двунитевые консенсусные олигонуклеотиды (Santa Cruz Biotechnologie, Heidelberg, Германия) подвергали не денатурирующему электрофорезу в полиакриламидном геле, авторадиографии и анализу Supershift, как описано Krzesz et al., (1999), FEBS Lett. 453, 191. При этом использовали двунитевой ДНК-олигонуклеотид со следующей однонитевой последовательностью (кор-связывающая последовательность подчеркнута): SIE, 5'-GTGCATTTCCCGTAAATCTTGTC-3' (SEQ ID NO:56). Для анализа вытеснения эндогенного STAT-1 в ядерных экстрактах цитокин-стимулированных ТНР-1 клеток (клеточная линия премоноцитов человека) посредством различных цис-элемент ловушек в условиях EMSA-связывания было выбрано соотношение 30:1 (STAT-1 цис-элемент ловушка: [32-Р]-меченный SIE олигонуклеотид (11 frnol)).
4. Технология олигонуклеотидов-ловушек
Двунитевые олигонуклеотиды-ловушки получали из комплементарных однонитевых фосфоротиоат-связанных олигонуклеотидов (Eurogentec, Koln, Deutschland) в соответствии с тем, как описано в Krzesz et al (1999) FEBS Lett. 453, 191. Культивированные клетки эндотелия человека предварительно инкубировали в течение 4 часов с соответствующим олигонуклеотидом-ловушкой в концентрации 10 мкМ. Эти условия были такими же, как условия, оптимизированные ранее на основе анализов EMSA и ПЦР с обратной транскриптазой. Среду, содержащую олигонуклеотид-ловушку, после этого обычно замещали свежей средой. Однонитевые последовательности олигонуклеотида были следующими (подчеркнутые буквы означают фосфоротиоат-связанные основания, все последовательности приведены в направлении 5'-3'):
GATA-2, CACTTGATAACAGAAAGTGATAACTCT (SEQ ID NO:57);
NF-κВ, AGTTGAGGGGACTTTCCCAGGC (SEQ ID NO:58);
STAT-1, CATGTTATGCATATTCCTGTAAGTG (SEQ ID NO:33);
STAT-1-19mut, GACAGTGCAGTGAACTGTC (SEQ ID NO:59);
STAT-1 -25mut, CATGTTATGCAGACCGTAGTAAGTG (SEQ ID NO:60).
5. Технология антисмысловых олигонуклеотидов (ODN)
Для эксперимента с антисмысловыми последовательностями к 100 мл культуральной среды OPTI-MEM®I добавляли 15 мкл липофектина (Gibco Life Technologie, Karlsruhe, Германия) и инкубировали при комнатной температуре (КГ) в течение 45 минут (раствор А). После этого добавляли антисмысловой ODN (Eurogentec, Koln, Германия) в конечной концентрации 0,5 мкМ в 100 мкл культуральной среды OPTI-MEM®I (раствор В). После объединения растворов А и В дальнейшее инкубирование осуществляли в течение 15 минут (КГ). В начале эксперимента 0,8 мл общепринятой клеточной культуральной среды для культуры эндотелиальных клеток (без гепарина и эндотелиального фактора роста) добавляли в пробирку типа Эппендорф, содержащую комплекс липофектин-антисмысловой ODN, и клеточную культуральную среду для культуры эндотелиальных клеток заменяли средой Antisense-Lipofectin-Medium. Среду Antisense-Lipofectin-Medium удаляли через 4 часа и замещали свежей культуральной средой (с гепарином и эндотелиальным фактором роста). Последовательность STAT-1-антисмыслового ODN представляла собой 5'-T*A*CCA*C*T*G*A*G*A*C*A*T*CC*T*GCC*A*C-3' (*фосфоротиоат-модифицированное основание; SEQ ID NO:41).
6. Вестерн-блот анализ
Клетки эндотелия пуповины человека и гладкомышечные клетки из вены тимуса разрушали посредством пяти последовательных циклов замораживания в жидком азоте и оттаивания при 37°С (термоблок, Kleinfelden, Германия). Белковые экстракты получали, как описано Hecker et al., (1994), Biochem J. 299, 247. 20-30 мкг белка разделяли путем электрофореза в 10% полиакриламидном геле в денатурирующих условиях в присутствии додецилсульфата натрия (SDS), следуя стандартному протоколу, и переносили на BioTrace™ Polyvinylidene Fluoride Transfermembran (Pall Corporation, Ro[3dorf, Deutschland). Для иммунологической детекции белка использовали следующие первичные антитела: белок CD40 (поликлональное, разведение 1:2000, Research Diagnostics Inc., Flanders NJ, США), белок STAT-1 (моноклональное, разведение 1:5000, BD Transduction Laboratories, Heidelberg, Германия), белок IRF-1 (поликлональное, разведение 1:2000, Santa Cruz Biotechnology, Heidelberg, Германия), белок iNOS (поликлональное, разведение 1:3000, BD Transduction Laboratories, Heidelberg, Германия). Полосы белка обнаруживали после добавления антикроличьих IgG, связанных с пероксидазой, и в случае использования моноклонального первичного антитела посредством соответствующего анти-мышиного IgG (1:3000, Sigma, Deisenhofen, Германия) с использованием хемилюминесцентного метода (SuperSignal хемилюминесцентный субстрат; Pierce Chemical, Rockford, IL, США) и последующей авторадиографией (Hyperfilm™MP, Amersham Phamnacia, Biotech, Buckinghamshire, Англия). Нанесение и перенос одинаковых количеств белка были показаны после «разборки» мембраны для переноса (5 минут 0,2 н. NaOH, с последующей отмывкой Н20, 3х10 минут) обнаружением одинаковых белка β-актина с моноклональным первичным антителом и связанным с пероксидазой анти-мышиным IgG (оба от Sigma-Aldrich, разведение 1:3000).
7. Статистический анализ
Если не указано иного, все данные на фигурах и в тексте представлены как среднее ± среднеквадратическое отклонение (С.О.) из n экспериментов. Статистический анализ осуществляли с использованием t-критерия Стьюдента для непарных данных с р-значением 0,05, который считали статистически значимым.
8. Подтверждение эффекта олигонуклеотидов-ловушек экспериментами на животных
8.1 Мышь
Для доказательства эффективности терапевтического подхода, основанного на олигонуклеотидах-ловушках, разработанных в настоящей заявке, было проведено экспериментальное "proof-of-concept" исследование на животных с использованием мышей, 8-10 животных на группу, для выявления антиген-индуцированного артрита (модель см. у Henzgen et al., Exp. Toxicol. Pathol. (1996), 48, 255). Однократное введение 0,25 нмоль олигонуклеотида-ловушки STAT-1 (SEQ ID NO:33) непосредственно в сустав (внутрисуставная инъекция) значительно снижало индуцированное антигеном опухание сустава (на 35%), интенсивность воспалительной реакции (на 70%), разрушение сустава (на 80%), общее количество артритных очагов (на 70%) и концентрацию провоспалительных цитокинов в сыворотке (например, интерлейкина-6, на 80%) за период 3-14 суток. В противоположность этому соответствующий контрольный олигонуклеотид не обладал никаким терапевтическим эффектом.
Более того, заслуживающим внимания в этом исследовании является то, что контактный дерматит (реакция IV типа), развившийся на коже через 14 дней после индуцирования артрита, тем что антиген введен еще одной инъекцией животным подкожно, также значительно снизился (на 35%) у мышей, подвергнутых лечению олигонуклеотидом-ловушкой.
8.2 Морская свинка
После двукратной аллергизации морских свинок (Hartley, самец, масса тела 350 г) в течение периода 7 суток (в 1-е сутки - в одно ухо, во 2-е сутки - в другое ухо по 50 мкл 10% DNCB-раствора в 50% ацетоне/50% оливковом масле; на 7-е сутки - повторная иммунизация в кожу шеи 15 мкл 2% раствора DNCB-раствора в 95% ацетоне/5% оливковом масле) контактный дерматит вызывали повторным применением 2,4-динитрохлорбензола (DNCB; 10 мкп 0,5% раствора DNCB в 95% ацетоне/5% оливковом масле) на 13-е сутки на одном или более чем одном участке размером примерно 1 см2 бритых спин животных и через 24 часа проводили макроскопическую и гистологическую оценку. Вызванный таким образом контактный дерматит гистологически (окрашивание красителем Гимза) характеризуется образованием ярко выраженного отека и межклеточного отека в области эпидермиса, увеличением апоптотических клеток, а также обширной инфильтрацией лейкоцитов (фиг.8). Внутрикожное введение олигонуклеотида-ловушки STAT-1 (SEQ ID NO:19), но не мутантного контрольного олигонуклеотида (5'-TGTGGACCGTAGGAAGTG-3', SEQ ID NO:61) за час до окончательного выдерживания в DNCB приводило к выраженному сокращению упомянутых гистологических параметров, то есть в общем к значительному ослаблению воспалительной реакции.













Изобретение относится к области молекулярной биологии и может быть использовано в медицине. Предложены двунитевые ДНК-олигонуклеотиды (олигонуклеотиды-ловушки), специфически связывающие транскрипционный фактор STAT-1 и нейтрализующие его действие в клетке. Олигонуклеотиды-ловушки по изобретению имеют размер от 15 до 21 п.н. и характеризуются последовательностью, которая включает 2 сайта связывания белка STAT-1. Показаны возможности применения новых ДНК-олигонуклеотидов в качестве лекарственного средства для предупреждения и/или терапии иммунологических реакций гиперчувствительности. 2 н. и 2 з.п. ф-лы, 8 ил.
| HUAHG J.ET AL., Biochem, J., 342, pt 1, 231-238, 1999 | |||
| Прибор для очистки паром от сажи дымогарных трубок в паровозных котлах | 1913 |
|
SU95A1 |

