ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способу модуляции альтернативного сплайсинга пре-мРНК, кодирующей рецептор конечных продуктов глубокого гликирования (RAGE) или его часть, с использованием переключающих сплайсинг антисмысловых олигонуклеотидов (AON) для модификации экспрессии и/или активности изоформ RAGE и к способам лечения связанных RAGE расстройств с использованием указанных модуляторов.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Рецептор конечных продуктов глубокого гликирования (RAGE) представляет собой многовалентный трансмембранный гликопротеин типа I, принадлежащий к сверхсемейству иммуноглобулинов (Ig). Ген человеческого RAGE (Ager) находится в пределах области главного комплекса гистосовместимости класса III на хромосоме 6. Он содержит 11 экзонов и 10 интронов, и 5' фланкирующую область, которая регулирует его транскрипцию. Транскрибированная мРНК RAGE имеет размер порядка 1,4 т.п.н с короткой 3'UTR (3'-нетранслируемая область).
50-55 кДа гликозилированный белок RAGE конститутивно экспрессируется в ограниченном диапазоне клеток (например, сосудистый эндотелий, пневмоциты типа I, лейкоциты), хотя экспрессия RAGE может индуцироваться в большинстве типов клеток и тканей после ранения, стресса, гипоксии или воспаления, обеспечивая канал провоспалительной и пропролиферативной сигнализации. Экспрессия RAGE, вследствие этого, подвергается повышающей регуляции при воспалительных и метаболических расстройствах, включающих нейродегенеративное заболевание, рак, сердечно-сосудистое заболевание, диабет, аутоиммунное и ишемическое поражение, но не ограничивающихся ими, при которых RAGE также вовлечен в развитие и прогрессирование.
RAGE связывали с целым рядом мозговых расстройств, включающих болезнь Альцгеймера; боковой амиотрофический склероз; болезнь Хантингтона; болезнь Крейтцфельда-Якоба; нейродегенеративные состояния, такие как диабетическая нейропатия, семейная амилоидная полинейропатия, нейроартропатия Шарко и васкулитная нейропатия; невропатическая боль; развитие и прогрессирование глиомы; ишемическое поражение/инсульт мозга и рассеянный склероз.
RAGE связывали со многими аспектами биологии опухолей, включая рост, миграцию и инвазию опухолевых клеток. При многих раковых заболеваниях экспрессируются высокие уровни RAGE (иллюстративными примерами являются рак молочной железы, толстой кишки, почки и желудка). Исключением является рак легкого, при котором экспрессия RAGE снижается, так как RAGE теряется по мере того, как клетки легкого дифференцируются и становятся более злокачественными.
В клетках глиомы С6 объем опухоли заметно уменьшается в опухолях, состоящих из клеток, в которых RAGE был заблокирован. В отличие от этого, опухоли, сверхэкспрессирущие RAGE дикого типа, быстро росли и очень эффективно вторгались в окружающую ткань. Было бы желательным иметь терапевтические средства для блокирования сигнализации RAGE в качестве лечения рака в отношении многих обычных раковых заболеваний, таких как глиома/полиморфная медуллобластома; рак поджелудочной железы; меланома; рак предстательной железы; рак молочной железы; рак печени/гепатома и рак толстой кишки.
В здоровом состоянии уровень легочной экспрессии RAGE является наивысшим из всех тканей. Однако экспрессия RAGE в легком обычно наблюдается только в пневмоцитах типа I. Повышающую регуляцию сигнализации RAGE в легком в других клетках и в других местах связывали с целым рядом легочных расстройств, включающих следующие: хроническое обструктивное заболевание легких (COPD)/эмфизема; астма; поражение, обусловленное курением сигарет/загрязнением; острое поражение легких/острый респираторный дистресс-синдром и легочный фиброз.
RAGE также критически участвует в целом ряде воспалительных состояний, таких как воспалительный артрит; остеоартрит; заболевание сетчатки; атеросклероз; кальциноз сосудов; ишемическая болезнь сердца/ремоделирование сердца/фиброз; сердечная недостаточность; диабетическое и недиабетическое заболевание почек; воспалительное заболевание кишечника; преэклампсия; поликистоз яичников; жировая дегенерация печени, фиброз, ишемическое и неишемическое поражение печени; мышечная дистрофия; повреждение спинного мозга; воспаление и старение кожи; и кератит.
Человеческий RAGE состоит из иммуноглобулиноподобного эктодомена, одного трансмембранного домена и короткого (42 аминокислоты) цитозольного хвоста. Данный эктодомен (также известный как внеклеточный домен) RAGE включает три иммуноглобулиноподобные области: N-концевой домен V-типа с последующими двумя доменами С-типа (именуемыми С и С или, альтернативно С1 иС2).
Связывание лигандов в виде конечных продуктов глубокого гликирования (AGE) и лигандов, не являющихся AGE, с эктодоменом RAGE активирует каскады внутриклеточной трансдукции сигналов, вовлеченные в воспаление, ранение, пролиферацию и дифференциацию клеток. Активация RAGE также запускает петлю положительной обратной связи, при которой взаимодействие лиганд-рецептор RAGE увеличивает экспрессию RAGE посредством активации NFκB (ядерный фактор каппа би), посредством этого увеличивая последующую индуцированную RAGE клеточную активацию. На самом деле, единственным известным для авторов данного изобретения способом сильной понижающей регуляции экспрессии RAGE является уменьшение активации RAGE. Данная ситуация расходится с другими рецепторами, для которых повышенные уровни лиганда снижают экспрессию рецептора.
У человека цитозольный хвост RAGE имеет длину 43 аминокислоты (от остатка 362 до остатка 404). Данный цитозольный хвост содержит мотивы, которые являются критически важными для RAGE-зависимой клеточной активации, но не для связывания лиганда. Цитозольный хвост RAGE может быть транс-активирован после активации колокализованных рецепторов, связанных с G-белком, посредством их когнатных лигандов без связывания лигандов в виде AGE или не являющихся AGE с эктодоменом RAGE (также известная как лиганд-независимая активация RAGE), приводя к активации тех же самых путей (Pickering et al. J Clin Invest 2019; 129: 406-421). Лиганднезависимая активация RAGE цитозольного хвоста RAGE посредством определенных колокализованных активированных рецепторов, связанных с G-белком, по-видимому, является важным путем, посредством которого RAGE активируется in vivo.
Как при лигандзависимой активации RAGE, так и при лиганднезависимой активации RAGE внутриклеточная сигнализация опосредуется цитозольным доменом RAGE, который взаимодействует с целым рядом партнеров сигнализации.
Альтернативный сплайсинг RAGE также является важным для регуляции активности RAGE посредством генерации изоформ RAGE, которые имеют измененную способность к активации лигандзависимым и лиганднезависимым путями сигнализации. Альтернативный сплайсинг RAGE изменяется при болезненных состояниях, включающих злокачественные образования, диабет и болезнь Апьцгеймера. Но, в то время как альтернативный сплайсинг, по-видимому, является важным для регуляции/неправильной регуляции RAGE, не было способа специфичного направленного действия на данный механизм.
Именно относительно данного предшествующего уровня техники описывается настоящий способ применения переключающих сплайсинг AON (антисмысловой олигонуклеотид) для модуляции сплайсинга RAGE в направлении предпочтительной генерации цитопротективных изоформ RAGE по всей длине RAGE.
Приведенное выше обсуждение предшествующего уровня техники предназначено только для того, чтобы облегчать понимание настоящего изобретения. Данное обсуждение не является признанием или допущением того, что любой материал, на который дается ссылка, представляет собой или был частью обычного общего знания по состоянию на дату приоритета данной заявки.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В широком смысле, согласно одной форме данного изобретения предложен выделенный или очищенный AON, который используется для модуляции альтернативного сплайсинга транскрипта пре-мРНК гена, кодирующего рецептор конечных продуктов глубокого гликирования (RAGE) или его часть.
В одном аспекте данного изобретения предложен AON из 10-50 нуклеотидов, содержащий нацеливающую последовательность, комплементарную области около или в пределах интрона пре-мРНК RAGE.
В одном аспекте данного изобретения предложен AON из 10-50 нуклеотидов, содержащий нацеливающую последовательность, комплементарную сайту сплайсинга или участку рядом с сайтом сплайсинга пре-мРНК RAGE.
Из-за таких факторов, как вторичная структура РНК, конкуренция между AON и белками SR, гетерогенными ядерными рибонуклеопротеинами (hnRNP) и/или другими элементами, которые составляют сплайсосому и могут влиять на действие AON, AON, направленные на ключевые сайты донора и акцептора сплайсинга, не всегда будут изменять сплайсинг. Следовательно, в одном аспекте данного изобретения предложен AON из 10-50 нуклеотидов, содержащий нацеливающую последовательность, комплементарную цис-действующим элементами РНК или смежным с ними участкам в пре-мРНК RAGE, которые действуют как энхансеры или сайленсеры, который, при связывании элементами сплайсосомы (например, белками-факторами сплайсинга, uRNA, IncRNA), модулирует сплайсинг близлежащего экзона.
В одном аспекте данного изобретения предложен AON из 10-50 нуклеотидов, содержащий нацеливающую последовательность, комплементарную пре-мРНК RAGE, который модулирует вторичную структуру указанной мРНК с влиянием на выбор сайта сплайсинга.
В одной форме данного изобретения предложен выделенный или очищенный AON для индукции исключения (также известного как пропуск) одной или более экзонных последовательностей в транскрипте гена RAGE или его части.
В одной форме данного изобретения предложен выделенный или очищенный AON для индукции сохранения интронных последовательностей в транскрипте гена RAGE или его части.
В одной форме данного изобретения AON является химически модифицированным для предупреждения деградации комплекса пре-мРНК-AON, включающим фосфородиамидатные морфолиноолигомеры (РМО), 2'-O-метилфосфоротиоатные олигонуклеотиды (2OMe) и 2'-O-метоксиэтил фосфоротиоатные олигонуклеотиды (2OMe), AON, модифицированные запертыми нуклеиновымие кислотами (LNA), термостабильную скрученную интеркалирующую нуклеиновую кислоту (TINA) и пептидные нуклеиновые кислоты (PNA), но не ограничивающимся ими.
В одной форме данного изобретения AON являются конъюгированными с группировками для усиления их доставки, включающими проникающие в клетку пептиды (СРР), vivo-морфолины (VMO) или пептидные фосфородиамидатные морфолиноолигомеры (РРМО), но не ограничивающимися ими.
Предпочтительно AON выбран из группы, содержащей последовательности, изложенные в любой из Таблиц 3a-3d. Предпочтительно AON выбран из списка, содержащего SEQ ID NO: 1-31. Более предпочтительно AON представляет собой SEQ ID NO: 11, 18, 19 или 20.
AON по изобретению может быть выбран в качестве AON, способного к связыванию с выбранным сайтом-мишенью, где данный сайт-мишень представляет собой предположительный сайт сплайсинга мРНК, выбранный из сайта донора сплайсинга, сайтов акцепторов сплайсинга, последовательностей энхансера сплайсинга, последовательностей сайленсера сплайсинга или сайтов, которые модулируют вторичную структуру пре-мРНК. Данный сайт-мишень также может включать некоторые фланкирующие интронные последовательности при нацеливании на сайты донора или акцептора сплайсинга.
Более конкретно AON может быть выбран из группы, состоящей из любой одной или более чем одной SEQ ID NO: 1-31 и/или последовательностей, изложенных в любой из Таблиц 3a-3d, и их комбинаций или смесей. Более предпочтительно данный AON представляет собой SEQ ID NO: 11, 18, 19 или 20. Комбинация AON предпочтительно представляет собой комбинацию SEQ ID NO: 11 и 10 или SEQ ID NO: 11 и 13. Это включает последовательности, которые могут гибридизоваться с такими последовательностями при жестких условиях гибридизации, комплементарные им последовательности, последовательности, содержащие модифицированные основания, модифицированные остовы и их функциональные усечения или удлинения, которые обладают активностью процессинга пре-мРНК или модулируют активность процессинга пре-мРНК в транскрипте гена RAGE.
В некоторых воплощениях AON могут быть на 100% комплементарными последовательности-мишени или могут включать несоответствия, например, для приспособления к вариантам, при условии, что гетеродуплекс, образованный между олигонуклеотидом и последовательностью-мишенью, является достаточно стабильным для противостояния действию клеточных нуклеаз и других способов деградации, которые могут наблюдаться in vivo. Следовательно, определенные олигонуклеотиды могут иметь примерно или по меньшей мере примерно 70%-ную комплементарность последовательности, например, 70%-ную, 71%-ную, 72%-ную, 73%-ную, 74%-ную, 75%-ную, 76%-ную, 77%-ную, 78%-ную, 79%-ную, 80%-ную, 81%-ную, 82%-ную, 83%-ную, 84%-ную, 85%-ную, 86%-ную, 87%-ную, 88%-ную, 89%-ную, 90%-ную, 91%-ную, 92%-ную, 93%-ную, 94%-ную, 95%-ную, 96%-ную, 97%-ную, 98%-ную, 99%-ную или 100%-ную комплементарность последовательности между олигонуклеотидом и последовательностью-мишенью.
Данное изобретение также распространяется на комбинацию двух или более чем двух AON, способных к связыванию с выбранной мишенью для модуляции альтернативного сплайсинга пре-мРНК RAGE, включая конструкцию, содержащую два или более чем два таких AON. Данные конструкции могут использоваться совместно для комбинированной терапии на основе AON. Комбинация AON предпочтительно представляет собой комбинацию SEQ ID NO: 11 и 10 или SEQ ID NO: 11 и 13.
Данное изобретение распространяется согласно его еще одному другому аспекту на кДНК или клонированные копии последовательностей AON по изобретению, а также на векторы, содержащие последовательности AON по изобретению. Данное изобретение также дополнительно распространяется на клетки, содержащие такие последовательности и/или векторы.
Также предложен способ манипулирования сплайсингом транскрипта гена RAGE, включающий стадию:
а) предоставления одного или более чем одного AON, как описано в данном документе, и обеспечения связывания данного(ных) олигомера(ров) с сайтом нуклеиновой кислоты-мишени.
Также предложена фармацевтическая, профилактическая или терапевтическая композиция для лечения, предупреждения или снижения интенсивности эффектов заболевания, связанных с экспрессией RAGE у пациента, содержащая:
a) один или более чем один AON, как описано в данном документе; и
b) один или более чем один фармацевтически приемлемый носитель и/или разбавитель.
Данная композиция может содержать примерно от 1 нМ до 1000 нМ каждого из желательных AON по изобретению. Предпочтительно данная композиция может содержать примерно от 10 нМ до 500 нМ, наиболее предпочтительно от 1 нМ до 10 нМ каждого из AON по изобретению.
Также предложен способ лечения, предупреждения или купирования эффектов заболевания, ассоциированных с экспрессией RAGE, включающий стадию:
а) введения пациенту эффективного количества одного или более чем одного AON или фармацевтической композиции, содержащей один или более чем один AON, как описано в данном документе.
Также предложено применение очищенных и выделенных AON, как описано в данном документе, для изготовления лекарственного средства для лечения, предупреждения или купирования эффектов заболевания, ассоциированного с экспрессией и/или активностью RAGE.
Также предложен набор для лечения, предупреждения или купирования эффектов заболевания, ассоциированного с экспрессией RAGE у пациента, который содержит по меньшей мере один AON, как описано в данном документе, и его комбинации или смеси, упакованные в подходящий контейнер, совместно с инструкциями для его применения.
Предпочтительно заболевание, ассоциированное с экспрессией RAGE у пациента, представляет собой нейродегенеративное заболевание, рак, легочное расстройство или воспалительное заболевание.
Субъектом с заболеванием, ассоциированным с экспрессией RAGE, может быть млекопитающее, включая человека.
Другие аспекты данного изобретения теперь будут описаны со ссылкой на сопровождающие неограничивающие примеры и графические материалы.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Другие характеристики настоящего изобретения более полно описаны в следующем описании его нескольких неограничивающих воплощений. Данное описание включается единственно в целях иллюстрации настоящего изобретения примерами. Его не следует понимать как ограничение широкого краткого изложения, раскрытия или описания изобретения, как изложено выше. Данное описание будет сделано со ссылкой на сопровождающие графические материалы, в которых:
На Фиг. 1а показан альтернативный сплайсинг RAGE, который генерирует RAGE и RAGE_v1. Экзоны показаны в виде прямоугольников. Кодирующие последовательности являются черными, а некодирующая последовательность является белой. Линии с углами обозначают события сплайсинга. S - кодон преждевременной терминации.
На Фиг. 1b показана кратность изменения экспрессии любой сплайсоформы мРНК RAGE (всего человеческого RAGE) и сплайсоформы мРНК RAGE, содержащие последовательность человеческого RAGE 9b, в клетках А579, при выявлении ПЦР-ОТ (полимеразная цепная реакция, сопряженная с обратной транскрипцией) в реальном времени, с последующей обработкой отобранными AON, нацеленными на экзон 10, или контрольных клеток (обработанные скремблированной РНК).
На Фиг. 1с показаны кратность изменения экспрессии мРНК любой сплайсоформы мРНК RAGE (всего человеческого RAGE) и сплайсоформы человеческой мРНК RAGE, содержащие экзон 10, при выявлении ПЦР-ОТ в реальном времени с последующей обработкой клеток А579 отобранными AON, нацеленными на экзон 10, или контрольных клеток (обработанные скремблированной РНК).
На Фиг. 1d показаны кратность изменения экспрессии мРНК любой сплайсоформы RAGE (всего человеческого RAGE) и сплайсоформы человеческой мРНК RAGE, содержащие экзон 10, при выявлении ПЦР-ОТ в реальном времени, с последующей обработкой клеток А579 отобранными AON, также нацеленными на экзон 10, смежный с предположительным сайтом связывания hnRNPF, или контрольные клетки (обработанные скремблированной РНК).
На Фиг. 1е показана процентная доля клонов RAGE, кодирующих конструкции, содержащие фрагменты экзонов 8-10 разных размеров, обозначающая относительную экспрессию разных сплайсоформ мРНК RAGE после обработки клеток А579 специфичными AON, нацеленными на экзон 10, или контрольных клеток (обработанные скремблированной РНК). Например, полоса 300 обозначает сплайсоформу, содержащую экзон 8, 9, 10 и 11 (т.е. сплайсоформы, способные к сигнализации), тогда как полоса 250 обозначает сплайсоформу, содержащую RAGE 9b, а отсутствие полосы обозначает сплайсоформу мРНК RAGE, ассоциированную с потерей экзона 8.
Фиг. 1f представляет собой репрезентативное изображение геля ПЦР-продуктов ДНК, охватывающих экзоны 8-11, с полосами разных размеров или отсутствием полос (стрелки), обозначающим присутствие других сплайсоформ.
На Фиг. 1g показано число копий мРНК RAGE, которая содержит экзон 10, при измерении посредством цифровой ПЦР после обработки клеток А579 с использованием AON 3779 по сравнению с контрольными клетками (обработанные скремблированной РНК).
На Фиг. 1h показана концентрация эндогенного растворимого белка RAGE, выявленного в среде клеток после обработки клеток А579 отобранными AON, нацеленными на экзон 10, по сравнению с контрольными клетками (обработанные скремблированной РНК).
На Фиг. 1i показана экспрессия белка RAGE в среде клеток (слева) и клеточном лизате (справа) после трансфекции клеток СНО (яичники китайского хомяка) с использованием ДНК, кодирующей геномную последовательность человеческого RAGE (gRAGE), с или без AON3779 или AON87, или контрольной (CTL) РНК посредством вестерн-блоттинга с детекцией с использованием антитела, специфичного в отношении RAGE. Полоса М представляет собой маркер, полоса 2 представляет собой контрольный вектор (не экспрессируется эндогенный RAGE), полоса 3 представляет собой gRAGE плюс контроль (скремблированная РНК), полоса 4 представляет собой gRAGE плюс AON3779, и полоса 5 представляет собой gRAGE плюс AON87. Стрелки обозначают размер меньшего растворимого RAGE в среде клеток (красная) и большего полноразмерного RAGE в клеточном лизате (белая).
На Фиг. 1j показана экспрессия растворимого белка RAGE в среде клеток после трансфекции клеток СНО с использованием ДНК, кодирующей геномную последовательность человеческого RAGE (gRAGE), с или без трансфекции AON, нацеленными на экзон 10, при измерении ELISA (твердофазный иммуноферментный анализ).
На Фиг. 1k показана экспрессия растворимого белка RAGE в среде после трансфекции клеток СНО с использованием ДНК, кодирующей геномную последовательность человеческого RAGE (gRAGE), с или без отобранных AON, нацеленных на экзон 10 рядом или с перекрытием с сайтом, на который также нацелен AON 3779, при измерении ELISA.
На Фиг. 1l показана дозозависимая индукция экспрессии растворимого RAGE в среде после трансфекции клеток СНО с использованием ДНК, кодирующей геномную последовательность человеческого RAGE, с или без AON 3779 при измерении ELISA. Пунктирная линия представляет экспрессию растворимого RAGE в клетках СНО, трансфицированных скремблированной РНК (10 нМ).
На Фиг. 1m показана индукция экспрессии гена ICAM-1 после обработки клеток А549 лигандом RAGE - S100A8/9 (0,6 мкг/мл) и ее модуляция посредством предварительной трансфекции отобранными AON, нацеленными на экзон 10, по сравнению с контрольными клетками (обработанными скремблированной РНК).
На Фиг. 1n показана индукция экспрессии гена ICAM-1 после обработки первичных человеческих эндотелиальных клеток аорты (НАЕС) лигандом RAGE -S100A8/9 (0,6 мкг/мл) и ее модуляция посредством предварительной трансфекции отобранными AON, нацеленными на экзон 10, по сравнению с контрольными клетками (обработанными скремблированной РНК).
На Фиг. 1о показана индукция экспрессии гена ICAM-1 после обработки клеток А549 с использованием Ang II, который способен индуцировать трансактивацию RAGE. Примечательно то, что данная индукция экспрессии гена ICAM-1 также модулируется посредством предварительной трансфекции клеток А579 отобранными AON, нацеленными на экзон 10, по сравнению с контрольными клетками (обработанными скремблированной РНК).
На Фиг. 1р показана индукция экспрессии гена ICAM-1 после обработки первичных человеческих эндотелиальных клеток аорты (НАЕС) ангиотезином II (1 мкМ) и ее модуляция посредством предварительной трансфекции отобранными AON, нацеленными на экзон 10, по сравнению с контрольными клетками (обработанными скремблированной РНК).
На Фиг. 1q показаны кратность изменения экспрессии любой сплайсоформы мРНК RAGE (всего человеческого RAGE) и сплайсоформы мРНК человеческого RAGE, содержащие экзон 10, при выявлении ПЦР-ОТ в реальном времени после обработки НМЕС AON, специфично нацеленными на экзон 10, смежный с предположительным сайтом связывания hnRNP-F/H1, или контрольных клеток (обработанные скремблированной РНК).
На Фиг. 1r показана кратность изменения экспрессии мРНК сплайсоформ мРНК RAGE, содержащих последовательность человеческого RAGE 9b после обработки НМЕС AON, нацеленными на экзон 10, смежный с предположительным сайтом связывания hnRNPF, или контрольных клеток (обработанные скремблированной РНК).
На Фиг. 1s показано влияние обработки первичных человеческих эндотелиальных клеток аорты (НАЕС) виво-морфолино препаратом AON 3779 (0,1-1 мкМ) на экспрессию сплайсоформ мРНК RAGE, содержащих экзон 9b, по сравнению с ненацеленным морфолино AON контролем.
На Фиг. 1t показано влияние обработки первичных человеческих эндотелиальных клеток аорты (НАЕС) виво-морфолино препаратом AON 3779 (0,1-1 мкМ) на экспрессию растворимого белка RAGE в культуральной среде по сравнению с ненацеленным морфолино AON контролем.
На Фиг. 1u показан эффект обработки клеток СНО, экспрессирующих геномный человеческий RAGE, виво-морфолино препаратом AON 3779 (1 мкМ) на экспрессию растворимого белка RAGE в культуральной среде по сравнению с ненацеленным морфолино AON контролем.
На Фиг. 1v показана последовательность ДНК человеческого RAGE, включающая и ограничивающая экзон 10, с обозначением разных AON, используемых в вышеописанных экспериментах, и их комплементарных мишеней. Пурпурный прямоугольник обозначает предположительную мишень полиG связывания hnRNPF.
На Фиг. 1w показано отсутствие влияния после мутации предположительной мишени полиС hnRNP на альтернативный сплайсинг экспресируемого человеческого RAGE (мутант gRAGE М2), на наблюдаемое увеличение растворимого RAGE в ответ на странсфекцию AON 3779.
На Фиг. 2а показана генетическая последовательность человеческого AGER, включающего и ограничивающего экзон 9, с обозначением разных AON, используемых ниже, и их комплементарных мишеней на пре-мРНК RAGE.
На Фиг. 2b показана экспрессия любых сплайсоформ мРНК RAGE (всего человеческого RAGE) и человеческих сплайсоформ мРНК RAGE, содержащих экзон 10, при выявлении ПЦР-ОТ в реальном времени после обработки клеток А579 отобранными AON, нацеленными на экзон 9, или контрольных клеток (обработанные скремблированной РНК).
Фиг. 2с представляет собой репрезентативное изображение геля ПЦР-продуктов ДНК, охватывающих экзон 8-11, с полосами разных размеров или без полос, обозначая присутствие разных сплайсоформ.
На Фиг. 2d представлена процентная доля клонов мРНК RAGE, кодирующих конструкции, содержащие фрагменты экзона 8-11 разных размеров, с обозначением относительной экспрессии разных сплайсоформ мРНК после обработки клеток А579 отобранными AON, нацеленными на экзон 10, или контрольной скремблированной РНК (CTL). Например, полоса 300 обозначает сплайсоформу мРНК RAGE, содержащую экзон 8, 9, 10 и 11 (т.е. сплайсоформы, способные к сигнализации), тогда как полоса 250 обозначает сплайсоформу, содержащую RAGE 9b, а отсутствие полосы обозначает сплайсоформу мРНК RAGE, ассоциированную с потерей экзона 8.
На Фиг. 2е показана экспрессия растворимого белка RAGE в среде после трансфекции клеток СНО ДНК, кодирующей геномную последовательность человеческого RAGE, с или без отобранных AON, нацеленных на экзон 9, при измерении посредством ELISA, контрольные клетки (обработанные скремблированной РНК). ND - не выявляется.
На Фиг. 2f показана индукция экспрессии гена TLR-4 после обработки лигандом RAGE - S100A8/9 (0,6 мкг/мл) и ее модуляция после трансфекции отобранными AON, нацеленными на экзон 9, по сравнению с контрольными клетками (обработанными скремблированной РНК).
На Фиг. 2g показана индукция экспрессии гена ICAM-1 после обработки Ang II (1 мкМ), который способен индуцировать трансактивацию RAGE. Примечательно то, что данная индукция экспрессии гена ICAM-1 модулируется после трансфекции клеток А579 отобранными AON, нацеленными на экзон 9, по сравнению с контрольными клетками (обработанными скремблированной РНК).
На Фиг. 2h показана кратность изменения экспрессии любых сплайсоформ мРНК RAGE (всего человеческого RAGE) и человеческих сплайсоформ мРНК RAGE, содержащих экзон 10, при выявлении ПЦР-ОТ в реальном времени после обработки клеток НМЕС1 отобранными AON, нацеленными на экзон 9, или контрольных клеток (обработанные скремблированной РНК).
На Фиг. 3а показана кратность изменения экспрессии любых сплайсоформ мРНК RAGE (всего RAGE) и человеческих сплайсоформ мРНК RAGE, содержащих экзон 10, при выявлении ПЦР-ОТ в реальном времени после обработки клеток А579 отобранными AON, нацеленными на интрон 9, AON 3779 (в качестве позитивного контроля) или контрольных клеток (обработанные скремблированной РНК).
На Фиг. 3b показана кратность изменения экспрессии сплайсоформ мРНК RAGE, содержащих последовательность экзона 9b, при выявлении ПЦР-ОТ в реальном времени после обработки клеток А579 отобранными AON, нацеленными на экзон 9, AON 3779 (в качестве позитивного контроля) или контрольных клеток (обработанные скремблированной РНК).
На Фиг. 3с показана экспрессия любых сплайсоформ мРНК RAGE (всего человеческого RAGE) и человеческих сплайсоформ мРНК RAGE, содержащих экзон 10, при выявлении ПЦР-ОТ в реальном времени после обработки клеток НМЕС1 отобранными AON, нацеленными на экзон 9, AON 3779 (в качестве позитивного контроля) или контрольных клеток (обработанные скремблированной РНК).
На Фиг. 3d показана кратность изменения экспрессии сплайсоформ мРНК RAGE, содержащих экзон 9b, при выявлении ПЦР-ОТ в реальном времени после обработки клеток НМЕС1 отобранными AON, нацеленными на экзон 9, AON 3779 (в качестве позитивного контроля) или контрольных клеток (обработанные скремблированной РНК).
На Фиг. 3е показана концентрация растворимого RAGE при выявлении ELISA в среде после обработки клеток А579, экспрессирующих геномный человеческий RAGE, специфичными AON, нацеленными на интрон 9, AON 3779 (в качестве позитивного контроля) или контрольных клеток (обработанные скремблированной РНК).
На Фиг. 3f показана последовательность ДНК человеческого RAGE, включающая и ограничивающая интрон 9, с обозначением AON и их комплементарных мишеней.
На Фиг. 4а показана концентрация растворимого RAGE при выявлении ELISA в среде после обработки клеток СНО, экспрессирующих геномный мышиный RAGE, AON, нацеленными на пре-мРНК RAGE, или контрольных клеток (обработанные скремблированной РНК).
На Фиг. 4b показана кратность изменения экспрессии любых сплайсоформ мРНК RAGE (всего мышиного RAGE) и сплайсоформ мРНК RAGE, содержащих экзон 10 и 11, на ПЦР-ОТ в реальном времени после обработки РМАЕС отобранными AON (50 нМ), нацеленными на экзон 9 мышиного RAGE, или контрольных клеток (обработанные скремблированной РНК).
На Фиг. 4с показана кратность изменения экспрессии любых сплайсоформ мРНК RAGE (всего мышиного RAGE) и сплайсоформ мРНК RAGE, содержащих экзон 10 и 11, после обработки РМАЕС отобранными AON (10 нМ), нацеленными на экзон 9 мышиного RAGE, или контрольных клеток (обработанные скремблированной РНК).
На Фиг. 4d показана концентрация растворимого белка RAGE в среде после обработки клеток РМАЕС специфичными AON (10 нМ), нацеленными на экзон 9 мышиного RAGE, или контрольных клеток (обработанные скремблированной РНК).
На Фиг. 4е показана кратность изменения экспрессии любых сплайсоформ мРНК RAGE (всего мышиного RAGE) и сплайсоформ мРНК RAGE, содержащих экзон 10 и 11, на ПЦР-ОТ в реальном времени после обработки РМАЕС AON m3779 (10 нМ) или контрольных клеток (обработанные скремблированной РНК).
На Фиг. 4f показана концентрация растворимого белка RAGE в среде после обработки клеток СНО, экспрессирующих геномный мышиный RAGE, AON 3779 (10 нМ), AON m3779 или контрольных клеток (обработанные скремблированной РНК).
На Фиг. 4g показана концентрация растворимого белка RAGE в среде после обработки клеток СНО, экспрессирующих геномный человеческий RAGE, AON 3779 (10 нМ), AON m3779 или контрольных клеток (обработанные скремблированной РНК).
На Фиг. 4h показана экспрессия любых сплайсоформ мРНК RAGE (всего RAGE) и сплайсоформ мРНК RAGE, содержащих экзон 10 и 11, на ПЦР-ОТ в реальном времени после обработки точно нарезанных кусочков легкого от мышей AON m3779 или ненацеленным морфолино AON в течение 48 часов.
На Фиг. 4i показана зависимая от времени экспрессия любых сплайсоформ мРНК RAGE (всего RAGE) и сплайсоформ мРНК RAGE, содержащих экзон 9b, на ПЦР-ОТ в реальном времени после обработки точно нарезанных кусочков легкого от мышей AON m3779 или носителем (физиологический раствор).
На Фиг. 4j показана кратность изменения легочной экспрессии любых сплайсоформ мРНК RAGE (всего мышиного RAGE) и мышиных сплайсоформ мРНК RAGE, содержащих экзон 9b или экзон 10, на ПЦР-ОТ в реальном времени через одну неделю после подкожной инъекции AON m3779 мышам C57BI6 по сравнению с контролем в виде ненацеленного AON.
На Фиг. 4k показана кратность изменения легочной экспрессии любых сплайсоформ мРНК RAGE (всего мышиного RAGE) и мышиных сплайсоформ мРНК RAGE, содержащих экзон 10 или 11, на ПЦР-ОТ в реальном времени через 48 часов после внутритрахеального закапывания морфолинового препарата AON m3779 мышам C57BI6 по сравнению с контролем в виде ненацеленного AON.
На Фиг. 4l показана кратность изменения легочной экспрессии любых сплайсоформ мРНК RAGE (всего мышиного RAGE) и мышиных сплайсоформ мРНК RAGE, содержащих экзон 10 или 11, на ПЦР-ОТ в реальном времени через 48 часов после внутритрахеального закапывания 2-О'Ме препарата AON m3779 мышам C57BI6 по сравнению с контролем в виде носителя. * обозначает р равно 0,01 относительно исходного уровня.
На Фиг. 4m показана кратность изменения (%) растворимого RAGE в системе кровообращения по сравнению с отбором крови в исходный момент времени через двое суток после внутритрахеальной инъекции AON m3779, AON 4105 или контроля в виде стерильной воды мышам C57BI6.
На Фиг. 5а показана экспрессия сплайсоформ мРНК RAGE, содержащих экзон 9b, при измерении ПЦР-ОТ в реальном времени после трансфекции клеток СНО ДНК, кодирующей геномную последовательность человеческого RAGE (gRAGE), комбинациями или без комбинаций выбранных AON, нацеленных на разные области экзона 10, или контрольных клеток (обработанные скремблированной РНК).
На Фиг. 5b показана экспрессия растворимого белка RAGE в среде после трансфекции клеток СНО ДНК, кодирующей геномную последовательность человеческого RAGE (gRAGE), комбинациями или без комбинаций выбранных AON, нацеленных на сайт 5' сплайсинга экзона 10, или контроля (обработанный скремблированной РНК) при измерении ELISA.
На Фиг. 5с показана экспрессия растворимого белка RAGE в среде после трансфекции клеток СНО с ДНК, кодирующей геномную последовательность человеческого RAGE (gRAGE), комбинациями или без комбинаций выбранных AON, нацеленных на сайт 3' и 5' сплайсинга экзона 10, или контрольной РНК при измерении ELISA. Прямоугольники указывают обработки, включающие AON 3779.
На Фиг. 5d показана экспрессия сплайсоформ мРНК RAGE, содержащих экзон 9b, после трансфекции клеток СНО ДНК, кодирующей геномную последовательность человеческого RAGE, комбинациями и без комбинаций отобранных AON, нацеленных на смежные области экзона 10, или контрольных клеток (обработанные скремблированной РНК) при измерении ПЦР-ОТ в реальном времени.
На Фиг. 5е показана экспрессия растворимого белка RAGE в среде после трансфекции клеток СНО ДНК, кодирующей мышиную геномную последовательность RAGE, комбинациями и без комбинаций отобранных AON, или контроля (скремблированная РНК) при измерении ELISA.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Подробное описание изобретения
Антисмысловые олигонуклеотиды (AON) представляют собой короткие, синтетические, антисмысловые, модифицированные нити ДНК или РНК, которые могут селективно гибридизоваться с пре-РНК/мРНК посредством образования пар оснований по Уотсону-Крику и селективно модулировать функцию РНК-мишени.
При применении AON для модуляции альтернативного сплайсинга мРНК их часто называют олигонуклеотидами, переключающими сплайсинг (SSO). В настоящем изобретении термины AON и SSO могут использоваться взаимозаменяемо. SSO образуют пары оснований с пре-мРНК и нарушают нормальный репертуар сплайсинга транскрипта посредством блокирования образования пар оснований РНК-РНК или взаимодействий связывания белок-РНК, которые происходят между компонентами аппарата сплайсинга и пре-мРНК. SSO могут индуцировать «пропуск» выбранных экзонов и/или сохранение интронных последовательностей для подуляции продукта трансляции. Это может достигаться посредством непосредственного нацеливания на сайты сплайсинга или посредством нацеливания на цис-действующие последовательности, участвующие в усилении или сайленсинге сплайсинга посредством модуляции связывания специфичных белков или изменения вторичной структуры пре-мРНК.
Терапевтические SSO можно использовать для лечения генетических расстройств, для пропуска испорченных или неправильно выровненных отрезков, обеспечивая получение внутренне делетированного, но теперь функционального белка в качестве терапии.
Альтернативный сплайсинг считается важным уровнем посттранскрипционной регуляции генов в отношении рецептора конечных продуктов глубокого гликирования (RAGE). Хотя большая часть RAGE и экспрессируется в его полноразмерную изоформу, посредством альтернативного сплайсинга генерируется целый ряд разных кодирующих изоформ (также известных как сплайсоформы), включая сплайсоформы с N-концевыми усечениями, С-концевыми усечениями и сплайсоформы, сохраняющие интронные последовательности. Эти разные сплайсоформы могут действовать в качестве возможных регуляторов полноразмерного рецептора RAGE либо посредством конкурентного связывания лиганда, либо посредством вытеснения полноразмерного белка от партнеров связывания. Идентифицировали свыше двадцати сплайсоформ в разных тканях, таких как легкое, печень, почка, гладкие мышцы, эндотелиальные клетки и мозг.
Разные сплайсированные варианты гена RAGE были названы RAGE, от RAGE_v1 до RAGE_v19 согласно Комитету по номенклатуре человеческих генов. Например, (запуск) сохранения интрона 9 (экзона 9b) приводит к преждевременной остановке (трансляции) и полной потере трансмембранного и цитоплазматического доменов, генерируя С-усеченную растворимую сплайсоформу (RAGE_v1 - эндогенный секреторный RAGE или esRAGE), которая составляет примерно 5% RAGE в системе кровообращения у человека. При отсутствие любых элементов сигнализации или трансмембранного домена esRAGE способен действовать в качестве рецептора-ловушки, конкурирующего с полноразмерным RAGE за лиганды или увеличивая клиренс лигандов. Более высокие уровни esRAGE в системе кровообращения ассоциированы с улучшенными результатами для здоровья и долголетием, тогда как меньшие уровни esRAGE ассоциированы со многими болезненными состояниями, включающими атеросклероз, диабет, метаболический синдром, смертность от сердечно-сосудистых заболеваний, анемию, аутизм и разные онкогенные состояния, но не ограничиваясь ими. Лечение диабетических мышей рекомбинантным esRAGE уменьшает атеросклероз, воспаление сосудов, почечное и ретинальное повреждение.
У RAGEΔ (также известного как DN RAGE или RAGEv20) отсутствуют 16 аминокислот во внутриклеточном домене, но сохраняется связывание лигандов и трансмембранные домены таким образом, что он действует как доминантно-негативный ингибитор RAGE клеточной поверхности. События сплайсинга, приводящие к изменениям внеклеточного домена, также могут влиять на домен связывания лиганда посредством вставки, делеции или удаления некоторой части или всего домена Ig-V RAGE. Например, N-RAGE начинается в альтернативном сайте инициации трансляции в экзоне 3 таким образом, что он не имеет сигнального пептида или V-домена, требующегося для связывания лиганда.
Нарушенный сплайсинг RAGE (и, следовательно, нарушенная сигнализация RAGE) был описан при диабете, некоторых видах рака и болезни Альцгеймера.
Пропуск экзона 10 при сплайсинге по типу esRAGE приписывают ограничению длины интрона у высших эукариот. Порядка 45 нуклеотидов должны отделять сайт 5' сплайсинга и точку ветвления, и минимальное расстояние между точкой ветвления и сайтом 3' сплайсинга, по-видимому, составляет приблизительно 18 нуклеотидов соответственно. Следовательно, интроны короче, чем 70 нуклеотидов являются крайне редкими у млекопитающих и не могут подвергаться эффективному сплайсингу. При выборе сайта 5' сплайсинга esRAGE расстояние между данным сайтом и сайтом 3' сплайсинга, который ограничивает экзон 10, составляет 46 нуклеотидов, что значительно короче, чем нижний предел длины интрона. Следовательно, использование расположенного ниже сайта 5' сплайсинга esRAGE интрона 9 и включение экзона 10 были бы взаимоисключающими. Среди известных проанализированных сплайсированных вариантов все варианты, в которых использовался расположенный ниже сайт 5' сплайсинга esRAGE в интроне 9, пропускали экзон 10; в отличие от этого, все варианты, в которых использовался расположенный выше сайт 5' сплайсинга RAGE в интроне 9, включали экзон 10. Таким образом, доступное доказательство указывает на то, что выбор любого одного из двух альтернативных сайтов 5' сплайсинга в интроне 9 связан с включением или исключением экзона 10. Способы регуляции данного сплайсинга или внешние способы модуляции раньше не были известными.
В настоящем изобретении используются SSO для селективной манипуляции картиной альтернативного сплайсинга пре-мРНК RAGE, приводя к генерации одной из двух природных сплайсоформ мРНК RAGE, которые либо являются нефункциональными, либо которые действуют как рецептор-ловушка с антагонистическим эффектом на лигандзависимую активацию и лиганднезависимую трансктивацию полноразмерного RAGE.
Примечательно то, что в данных сайтах сплайсинга нет обычных полиморфизмов RAGE. Последовательность RAGE является высококонсервативной. Следовательно, не требуется персонализация или индивидуализированная модификация последовательности, в отличие от управления генетическими расстройствами с использованием технологий пропуска экзонов.
Согласно настоящему изобретению предложен альтернативный способ лечения, предупреждения или купирования эффектов заболеваний, в которых RAGE вовлечен в развитие или прогрессирование, включая нейродегенеративные заболевания, рак, легочные расстройства или воспалительные заболевания, но не ограничиваясь ими, посредством разработки AON, которые модулируют альтернативный сплайсинг пре-мРНК RAGE или его части.
В широком смысле согласно одному аспекту данного изобретения предложен выделенный или очищенный AON для осуществления модуляции сплайсинга пре-мРНК в транскрипте гена рецептора конечных продуктов глубокого гликирования (RAGE) или его части. Предпочтительно предложен выделенный или очищенный AON для индукции исключения экзона и/или сохранения интрона в пре-мРНК RAGE или ее части.
Согласно данному изобретению предложен AON, способный к связыванию с выбранной мишенью на транскрипте гена рецептора конечных продуктов глубокого гликирования (RAGE), для модуляции сплайсинга пре-мРНК в транскрипте гена RAGE или его части.
Например, в одном аспекте данного изобретения предложен AON из 10-50 нуклеотидов, содержащий нацеливающую последовательность, комплементарную области пре-мРНК RAGE или ее части, ассоциированной со связыванием белка, участвующего в регуляции альтернативного сплайсинга мРНК.




В отличие от других терапий на основе AON согласно настоящему изобретению не индуцируется повышенная деградация РНК посредством рекрутинга РНКазы Н, где РНКаза Н предпочтительно связывается и деградирует РНК, связанную в дуплексе с ДНК гена RAGE. Оно не основывается ни на гибридизации AON с геномной ДНК RAGE, ни на связывании AON с мРНК для модуляции количества продуцированного белка RAGE посредством вмешательства в нормальные функции, такие как репликация, транскрипция, транслокация и трансляция. Скорее данные AON используются для модуляции сплайсинга пре-мРНК в транскрипте гена RAGE или его части и индуцируют «пропускание» экзона или (запуск) сохранения интронных последовательностей. Данная стратегия предпочтительно уменьшает экспрессию сплайсоформ мРНК RAGE, способных опосредовать RAGE-зависимую сигнализацию, и/или увеличивает генерацию сплайсоформ мРНК RAGE, у которых отсутствуют функциональные домены, способные опосредовать RAGE-зависимую сигнализацию.
Согласно первому аспекту данного изобретения предложены AON, способные к связыванию с выбранной мишенью на транскрипте гена RAGE для модуляции сплайсинга пре-мРНК в транскрипте гена RAGE или его части. В широком смысле, предложен выделенный или очищенный AON для индуцирования целевого исключения экзона/сохранения интрона в транскрипте гена RAGE или его части.
Под «выделенным» подразумевают вещество, которое в значительной степени или по существу на содержит компонентов, которые обычно сопровождают его в его природном состоянии. Например, термины «выделенный полинуклеотид» или «выделенный олигонуклеотид» в том виде, в котором они используются в данном документе, могут относиться к полинуклеотиду, который был очищен или удален от последовательностей, которые фланкируют его в состоянии, встречающемся в природе, например, фрагмент ДНК, который удаляется от последовательностей, которые являются смежными с данным фрагментом в геноме. Термин «осуществление выделения» в том виде, в котором он относится к клеткам, относится к очистке клеток (например, фибробластов, лимфобластов) из субъекта-источника (например, субъекта с заболеванием, связанным с полинуклеотидом с повторами). В контексте ДНК, мРНК или белка термин «осуществление выделения» относится к извлечению ДНК, мРНК или белка из источника, например, клеток.
Можно сказать, что AON «направлен на» или «нацелен против» последовательности-мишени, с которой он гибридизуется. В некоторых воплощениях последовательность-мишень включает область, включающую сайт 3' или 5' сплайсинга мРНК до процессинга, точку ветвления или другие последовательности, участвующие в регуляции сплайсинга, включая энхансеры сплайсинга и сайленсеры сплайсинга, и сайты, определяющие вторичную структуру РНК, которые влияют на сплайсинг. Данная последовательность-мишень может находиться в пределах экзона или в пределах интрона, или охватывать соединение интрон/экзон.
В некоторых воплощениях AON имеет достаточную комплементарность последовательности с РНК-мишенью (т.е. РНК, в отношении которой модулируется выбор сайта сплайсинга) для эффективного блокирования области РНК-мишени (например, пре-мРНК). В типичных воплощениях такое блокирование пре-мРНК RAGE служит для модулирования сплайсинга либо посредством маскировки сайта связывания сплайсосомного белка, который, в противном случае, модулировал бы сплайсинг, и/либо посредством изменения структуры РНК-мишени. В некоторых воплощениях РНК-мишень представляет собой пре-мРНК-мишень (например, пре-мРНК гена RAGE).
Фраза «AON, имеющий достаточную комплементарность последовательности с последовательностью РНК-мишени для модуляции сплайсинга РНК-мишени» означает то, что данный AON имеет достаточную последовательность для запуска маскировки сайта связывания природного белка, который, в противном случае, модулировал бы сплайсинг и/или изменял бы трехмерную структуру РНК-мишени.
Отобранные AON могут быть сделаны более короткими, например, примерно 12 оснований или длиннее, например, примерно 50 оснований, и включают небольшое число несоответствий, при условии, что данная последовательность является достаточно комплементарной для осуществления модуляции сплайсинга при гибридизации с последовательностью-мишенью и возможно образует с РНК гетеродуплекс, имеющий Tm (температура плавления) 45°С или больше.
Предпочтительно AON выбран из группы, содержащей SEQ ID NO: 1-31 и/или последовательности, изложенные в любой из Таблиц 3а-3d. Более предпочтительно данный AON представляет собой SEQ ID NO: 11, 18, 19 или 20.
В некоторых воплощениях степень комплементарности между последовательностью-мишенью и AON является достаточной для образования стабильного дуплекса. Область комплементарности AON с последовательностью РНК-мишенью может быть такой короткой как 8-11 оснований, но может составлять 12-15 оснований или больше, например, 10-50 оснований, 10-40 оснований, 12-30 оснований, 12-25 оснований, 15-25 оснований, 12-20 оснований или 15-20 оснований, включая все целые числа между данными интервалами. AON из примерно 16-17 оснований обычно является достаточно длинным для наличия уникальной комплементарной последовательности. В некоторых воплощениях может потребоваться минимальная длина комплементарных оснований для достижения необходимой Tm связывания, как обсуждается в данном документе.
В некоторых воплощениях подходящими могут быть олигонуклеотиды длиной 50 оснований, где по меньшей мере минимальное число оснований, например, 10-12 оснований, является комплементарным последовательности-мишени. В общем, однако, облегченное или активное поглощение в клетки оптимизировано при длине олигонуклеотидов меньше, чем примерно 30 оснований. Для фосфороамидатморфолино олигомера (РМО) AON, описанного в данном документе далее, оптимальный баланс стабильности связывания и поглощения обычно наблюдается при длине 18-25 оснований. Включенными являются AON (например, РМО, РМО-Х, PNA, LNA, TINA, 2'-ОМе), которые состоят из примерно 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49 или 50 оснований.
В некоторых воплощениях AON могут быть на 100% комплементарными последовательности-мишени или могут включать несоответствия, например, для приспособления вариантов, при условии, что гетеродуплекс, образованный между олигонуклеотидом и последовательностью-мишенью является достаточно стабильным для того, чтобы противостоять действию клеточных нуклеаз и других способов деградации, которые могут встречаться in vivo. Следовательно, определенные олигонуклеотиды могут иметь примерно или по меньшей мере примерно 70%-ную комплементарность последовательности, например, 70%-ную, 71%-ную, 72%-ную, 73%-ную, 74%-ную, 75%-ную, 76%-ную, 77%-ную, 78%-ную, 79%-ную, 80%-ную, 81%-ную, 82%-ную, 83%-ную, 84%-ную, 85%-ную, 86%-ную, 87%-ную, 88%-ную, 89%-ную, 90%-ную, 91%-ную, 92%-ную, 93%-ную, 94%-ную, 95%-ную, 96%-ную, 97%-ную, 98%-ную, 99%-ную или 100%-ную комплементарность последовательности между олигонуклеотидом и последовательностью-мишенью.
Несоответствия, при их присутствии, типично являются менее дестабилизирующими в направлении к концевым областям гибридного дуплекса, чем в середие. Чисто допустимых несоответствий будет зависеть от длины олигонуклеотида, процентной доли пар оснований G:C в дуплексе и положения несоответствия(вий) в дуплексе согласно хорошо понятным принципам стабильности дуплексов. Хотя такой AON и не обязательно является на 100% комплементарным последовательности-мишени, он является эффективным для того, чтобы стабильно и специфично связываться с последовательностью-мишенью, таким образом, что модулируется сплайсинг пре-РНК-мишени.
Стабильность дуплекса, образующегося между AON и последовательностью-мишенью, является функцией Tm связывания и чувствительности дуплекса к клеточному ферментативному расщеплению. Tm олигонуклеотида по отношению к РНК с комплементарной последовательностью может быть измерена традиционными способами, такими как способы, описанные Hames et al., Nucleic Acid Hybridization, IRL Press, 1985, pp. 107-108, или как описано в Miyada С.G. and Wallace R.В., 1987, Oligonucleotide Hybridization Techniques, Methods Enzymol. Vol. 154 pp. 94-107. В некоторых воплощениях AON могут иметь Tm связывания по отношению к РНК с комплементарной последовательностью больше, чем температура тела и предпочтительно больше, чем примерно 45°С или 50°С. Также включается Tm в интервале 60-80°С или больше.
Дополнительные примеры вариантов включают AON, имеющие примерно или по меньшей мере примерно 70%-ную идентичность или гомологию последовательности, например, 70%-ную, 71%-ную, 72%-ную, 73%-ную, 74%-ную, 75%-ную, 76%-ную, 77%-ную, 78%-ную, 79%-ную, 80%-ную, 81%-ную, 82%-ную, 83%-ную, 84%-ную, 85%-ную, 86%-ную, 87%-ную, 88%-ную, 89%-ную, 90%-ную, 91%-ную, 92%-ную, 93%-ную, 94%-ную, 95%-ную, 96%-ную, 97%-ную, 98%-ную, 99%-ную или 100%-ную идентичность или гомологию последовательности по всей длине любой из SEQ ID NO: 1-31 и/или последовательностей, изложенных в любой из Таблиц 3a-3d. Более предпочтительно AON представляет собой SEQ ID NO: 11, 18, 19 или 20.
Более конкретно, предложен AON, способный к связыванию с выбранным сайтом-мишенью, для модификации сплайсинга пре-мРНК в транскрипте гена RAGE или его части. AON предпочтительно выбран из SEQ ID NO: 1-31 и/или последовательностей, изложенных в любой из Таблиц 3а-3d. Более предпочтительно AON представляет собой SEQ ID NO: 11, 18, 19 или 20.
Модификация сплайсинга пре-мРНК предпочтительно включает «пропускание» или удаление одного или более чем одного экзона или сохранение интронов мРНК. Образующийся белок предпочтительно имеет более короткую длину по сравнению с родительским полноразмерным белком RAGE из-за либо внутреннего усечения, либо преждевременной термнации. Данные усеченные белки RAGE могут называться сплайсоформами полноразмерного белка RAGE.
Остальные экзоны генерированной мРНК могут находиться в рамке считывания и могут продуцировать более короткий белок с последовательностью, которая является аналогичной последовательности, родительского полноразмерного белка за исключением того, что она имеет внутреннее усечение в области между исходными 3'- и 5'-концами. При другой возможности пропуск экзона может индуцировать сдвиг рамки считывания, который приводит к белку, в котором первая часть данного белка является по существу идентичной родительскому полноразмерному белку, но в котором вторая часть данного белка имеет отличную последовательность (например, бессмысленная последовательность) из-за сдвига рамки считывания. В качестве альтернативы, пропуск экзона может индуцировать продукцию преждевременно терминированного белка из-за нарушения рамки считывания и присутствия преждевременной терминации трансляции. Преждевременно терминированный белок может быть результатом мРНК, которая преждевременно терминируется (например, пропуск экзонов 10 и/или 11), или может быть результатом продолжения в интрон (например, RAGE 9b) или бессмысленный пропуск, который дает мРНК, которая содержит экзон 10 и/или 11 мРНК, но которая не обеспечивает экспрессию белка, кодируемого данными экзонами.
Пропуск индивидуальных экзонов - экзонов 1-9 - будет предпочтительно нарушать рамку считывания транскрипта RAGE. Это будет приводить к усиленной деградации РНК посредством нонсенс-опосредованного распада.
Пропуск индивидуальных экзонов - экзонов 1-11 - будет предпочтительно сохранять интактную рамку считывания. Это будет предпочтительно приводить к трансляции в белок с внутренним усечением. Данный усеченный белок или сплайсоформа мРНК RAGE может иметь полностью устраненную функцию, может иметь пониженную функцию или может действовать в качестве рецептора-ловушки.
Предпочтительно данные усеченные, нонсенсные или преждевременно терминированные белки не имеют одного или более чем одного функционального домена, участвующего в индукции путей внутриклеточной сигнализации посредством лигандов RAGE или независимой от лигандов трансактивации RAGE посредством колокализованных GPCR (рецепторы, связанные с G-белками). Например, экзон 10 кодирует трансмембранный домен, и удаление данного экзона может генерировать растворимый белок RAGE, который мог бы потенциально действовать в качестве растворимой ловушки или конкурентного антагониста лиганд-индуцированной сигнализации через RAGE. Усеченные, нонсенсные или преждевременно терминированные белки, кроме того, могут не иметь присоединения или сайта связывания других факторов, удаление которых может приводить к уменьшению взаимодействия белка RAGE с релевантными путями сигнализации.
В качестве альтернативы, удаление одного или более чем одного экзона может приводить к неправильному фолдингу белка RAGE и уменьшению способности данного белка к успешной транспортировке через мембрану.
Присутствие внутренне усеченных белков (т.е. белков, не имеющих аминокислот, кодируемых одним или более чем одним экзоном) является предпочтительным. При ингибировании белка RAGE могут быть проблемы с увеличением транскрипции RAGE, т.к. организм пытается компенсировать уменьшение общего количества белка RAGE. В отличие от этого, присутствие внутренне усеченного белка (предпочтительно не имеющего один или более чем один элемент полного белка RAGE) должно быть достаточным для предупреждения повышенной транскрипции, но все еще обеспечивает терапевтическое преимущество из-за уменьшения общего количества функционального белка RAGE.
Индуцированный AON пропуск экзона по настоящему изобретению не обязательно должен полностью или даже существенно устранять функцию белка RAGE. Предпочтительно модуляция альтернативного сплайсинга посредством способа пропуска экзона приводит к сниженной или нарушенной функциональности белка RAGE.
Разные изоформы RAGE, продуцированные с использованием разных стратегий пропуска, могли бы приводить к белкам с устраненной или пониженной активностью сигнализации, которые предпочтительно могли бы использоваться для лечения или предупреждения разных заболеваний, ассоциированных с антивностью RAGE, таких как нейродегенеративные заболевания, рак, легочные расстройства или воспалительные заболевания. Стратегии альтернативного сплайсинга могут образовать усеченные белки или белки с ослабленными функциями, которые предпочтительно могут использоваться в качестве лечений конкретных аспектов, форм или прогрессирования заболеваний, ассоциированных с экспрессией и активностью RAGE.
Способ пропуска по настоящему изобретению с использованием AON может исключать (пропускать) индивидуальный экзон или может приводить сразу к пропуску двух или более чем двух экзонов.
Способ пропуска на настоящему изобретению с использованием AON может включать сохранение интронных последовательностей с или без непосредственного пропуска одного или более чем одного экзона.
AON по данному изобретению могут представлять собой комбинацию двух или более чем двух AON, способных к связыванию с выбранной мишенью для индукции исключения экзона в транскрипте гена RAGE. Данная комбинация может представлять собой смесь двух или более чем двух AON и/или конструкцию, содержащую два или более чем два AON, связанных друг с другом.






Согласно данному изобретению дополнительно предложен способ модулирования альтернативного сплайсинга в транскрипте гена RAGE, включающий стадию:
предоставления одного или более чем одного AON, как описано в данном документе, и обеспечения связывания данного(ных) олигомера(ров) с сайтом нуклеиновой кислоты-мишени.
Согласно еще одному другому аспекту данного изобретения предложена последовательность нуклеиновой кислоты-мишени для модулирования альтернативного сплайсинга пре-мРНК RAGE, содержащая ДНК-эквиваленты последовательностей нуклеиновой кислоты, выбранные из группы, состоящей из SEQ ID NO: 1-31 и/или последовательностей, изложенных в любой из Таблиц 3а-3d, и последовательностей, комплементарных им. Более предпочтительно данный AON представляет собой SEQ ID NO: 11, 18, 19 или 20. Данный AON может представлять собой комбинацию AON, предпочтительно комбинацию SEQ ID NO: 11 и 10 или SEQ ID NO: 11 и 13.
Конструирование AON для полной маскировки консенсусных сайтов сплайсинга не обязательно может генерировать изменение сплайсинга экзона-мишени. Кроме того, авторы данного изобретения открыли то, что сам размер или длина AON не всегда является первичным фактором при конструировании AON. С некоторыми мишенями такие короткие AON, как из 20 оснований, могли индуцировать некоторое включение экзона, в некоторых случаях более эффективно, чем другие более длинные (например, 25 оснований) олигомеры, направленные на тот же самый экзон.
Авторы данного изобретения также открыли, что, по-видимому, нет какого-либо стандартного мотива, который может быть блокирован или маскирован AON для перенаправления сплайсинга. Обнаружили, что для каждого гена-мишени должны быть сконструированы AON и должна эмпирически оцениваться их индивидуальная эффективность.
Более конкретно AON могут быть выбраны из AON, изложенных в любой из Таблиц 3a-3d. Данные последовательности предпочтительно выбраны из группы, состоящей из любой одной или более чем одной SEQ ID NO: 1-31 и их комбинаций или смесей. Более предпочтительно данный AON представляет собой SEQ ID NO: 11, 18, 19 или 20. Комбинация AON предпочтительно представляет собой комбинацию SEQ ID NO: 11 и 10 или SEQ ID NO: 11 и 13. Это включает последовательности, которые могут гибридизоваться с такими последовательностями при жестких условиях гибридизации, комплементарные им последовательности, последовательности, содержащие модифицированные основания, модифицированные остовы и их функциональные усечения или удлинения, которые обладают или модулируют активность процессинга пре-мРНК в транскрипте гена RAGE.
Олигомер и ДНК, кДНК или РНК являются комплементарными друг другу, когда достаточное число соответствующих положений в каждой молекуле заняты нуклеотидами, которые могут связываться друг с другом водородными связями. Таким образом, «специфично гибридизуемый» и «комплементарный» представляют собой термины, которые используются для указания достаточной степени комплементарности или образования пар, таким образом, что происходит стабильное и специфичное связывание между олигомером и ДНК, кДНК или РНК-мишенью. В данной области понятно то, что последовательность AON не должна быть на 100% комплементарной ее последовательности-мишени для того, чтобы быть специфично гибридизуемой. AON является специфично гибридизуемым, когда связывание соединения с молекулой ДНК- или РНК-мишени препятствует нормальной функции данного целевого ДНК- или РНК-продукта, и имеется достаточная степень комплементарности для того, чтобы избегать неспецифичного связывания данного AON с нецелевыми последовательностями при условиях, при которых специфичное связывание является желательным, т.е. при физиологических условиях в случае анализов in vivo или терапевтического лечения, и в случае анализов in vitro при условиях, в которых проводятся данные анализы.
Селективная гибридизация может осуществляться при условиях низкой, умеренной или высокой жесткости, но предпочтительно при условиях высокой жесткости. Специалистам в данной области будет понятно то, что на жесткость гибридизации будут влиять такие условия, как концентрация соли, температура или органические растворители, помимо состава оснований, длины комплементарных нитей и числа несоответствий нуклеотидных оснований между гибридизуемыми нуклеиновыми кислотами. Жесткие температурные условия обычно будут включать температуры, превышающие 30°С, типично превышающие 37°С и предпочтительно превышающие 45°С, предпочтительно по меньшей мере 50°С и типично 60°С-80°С или выше. Жесткие солевые условия обычно будут представлять собой меньше, чем 1000 мМ, типично меньше, чем 500 мМ и предпочтительно меньше, чем 200 мМ. Однако значительно более важной является комбинация параметров, чем мера любого одиночного параметра. Примером жестких условий гибридизации являются 65°С и 0,1 SSC (1 SSC - 0,15 М NaCl, 0,015 М цитрат натрия, рН 7,0). Таким образом, AON по настоящему изобретению могут включать олигомеры, которые селективно гибридизуются с последовательностями, приведенными в любой из Таблиц 3a-3d или в SEQ ID NO: 1-31. Более предпочтительно данный AON представляет собой SEQ ID NO: 11, 18, 19 или 20.
Будет понятно то, что организации кодонов на конце экзонов в структурных белках могут не всегда прерываться на конце кодона, следовательно, может быть необходимо удалять более чем один экзон из пре-мРНК для обеспечения считывания мРНК в рамке считывания. При таких обстоятельствах может быть нужно выбирать целый ряд AON способом по изобретению, где каждый направлен на другую область, ответственную за индукцию включения желательного экзона и/или интрона. При данной ионной силе и рН Tm представляет собой температуру, при которой 50% последовательности-мишени гибридизуется с комплементарным полинуклеотидом. Такая гибридизация может происходить с «близкой» или «существенной» комплементарностью AON с последовательностью-мишенью, а также сточной комплементарностью.
Типично селективная гибридизация будет происходить, когда имеется по меньшей мере примерно 55%-ная идентичность по отрезку из по меньшей мере примерно 14 нуклеотидов, предпочтительно по меньшей мере примерно 65%-ная, более предпочтительно по меньшей мере примерно 75%-ная и наиболее предпочтительно по меньшей мере примерно 90%-ная, 95%-ная, 98%-ная или 99%-ная идентичность с нуклеотидами AON. Длина сравнения гомологии, как описано, может осуществляться на более длинных отрезках, и в некоторых воплощениях часто будет осуществляться на отрезке из по меньшей мере примерно девяти нуклеотидов, обычно по меньшей мере примерно 12 нуклеотидов, более обычно примерно 20, часто по меньшей мере примерно 21, 22, 23 или 24 нуклеотидов, по меньшей мере примерно 25, 26, 27 или 28 нуклеотидов, по меньшей мере примерно 29, 30, 31 или 32 нуклеотидов, по меньшей мере примерно 36 или больше нуклеотидов.
Таким образом, последовательности AON по изобретению предпочтительно имеют по меньшей мере 75%-ную, более предпочтительно по меньшей мере 85%-ную, более предпочтительно по меньшей мере 86, 87, 88, 89 или 90%-ную гомологию с последовательностями, показанными в перечнях последовательностей в данном документе. Более предпочтительно имеется по меньшей мере 91, 92, 93, 94 или 95%-ная, наиболее предпочтительно по меньшей мере 96, 97, 98%-ная или 99%-ная гомология. Обычно чем короче длина AON, тем большая гомология требуется для получения селективной гибридизации. Следовательно, где AON по изобретению состоит из меньше, чем примерно 30 нуклеотидов, предпочтительным является то, что процентная доля идентичности больше, чем 75%, предпочтительно больше, чем 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95%, 96, 97, 98% или 99% по сравнению с AON, изложенными в данном документе в перечнях последовательностей. Сравнения гомологии нуклеотидов могут проводиться программами сравнения последовательностей, такими как программа GCG Wisconsin Bestfit или GAP (Deveraux et al., 1984, Nucleic Acids Research 12, 387-395). Данным способом могли бы сравниваться последовательности аналогичной или существенно разной длины с последовательностями, процитированными в данном документе, посредством вставок пробелов в выравнивание, причем такие пробелы определяются, например, посредством алгоритма сравнения, используемого GAP.
AON по настоящему изобретению могут иметь области пониженной гомологии и области точной гомологии с последовательностью-мишенью. Для олигомера не нужно иметь точную гомологию по всей его длине. Например, данный олигомер может иметь непрерывные отрезки из по меньшей мере 4 или 5 оснований, которые являются идентичными последовательности-мишени, предпочтительно непрерывные отрезки из по меньшей мере 6 или 7 оснований, которые являются идентичными последовательности-мишени, более предпочтительно непрерывные отрезки из по меньшей мере 8 или 9 оснований, которые являются идентичными последовательности-мишени. Данный олигомер может иметь отрезки из по меньшей мере 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или 26 оснований, которые являются идентичными последовательности-мишени. Остальные отрезки последовательности олигомера могут быть перемежающе идентичными последовательности-мишени; например, остальная последовательность может иметь идентичное основание с последующим неидентичным основанием, с последующим идентичным основанием. В качестве альтернативы (или также), последовательность олигомера может иметь несколько отрезков идентичной последовательности (например, 3, 4, 5 или 6 оснований), перемежающихся с отрезками с менее, чем совершенной гомологией. Такие несоответствия последовательности предпочтительно не будут иметь или будут иметь очень малую потерю активности переключения сплайсинга.
Термин «модулировать» или «модулирует» включает «увеличивать» или «уменьшать» один или более чем один количественно оцениваемый параметр, возможно на определенное и/или статистически значимое количество. Термины «увеличивать» или «увеличивающий», «усиливать» или «усиливающий», или «стимулировать», или «стимулирущий» в общем относятся к способности одного или более чем одного AON или композиций продуцировать или вызывать больший физиологический ответ (например, эффекты ниже) в клетке или у субъекта относительно ответа, вызванного либо без AON, либо контрольным соединением. Термины «уменьшающий» или «уменьшать» в общем относятся к способности одного или более чем одного AON или композиций продуцировать или вызывать меньший физиологический ответ (например, эффекты ниже) в клетке или у субъекта относительно ответа, вызванного либо без AON, либо контрольным соединением.
Релевантные физиологические или клеточные ответы (in vivo или in vitro) будут очевидными специалистам в данной области и могут включать увеличения исключения конкретных экзонов в пре-мРНК, кодирующей RAGE, уменьшения количества пре-мРНК, кодирующей RAGE, или снижения экспрессии функционального белка RAGE в клетке, ткани или у субъекта, нуждающегося в этом. «Увеличенное» или «повышенное» количество типично представляет собой статистически значимое количество и может включать увеличение, которое составляет в 1,1, 1,2, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 30, 40, 50 или более раз (например, 500, 1000 раз) (включая все целые числа и десятые доли между ними и больше 1, например, 1,5; 1,6; 1,7; 1,8) относительно количества, продуцируемого при отсутствии AON (отсутствие агента) или в присутствии контрольного соединения. Термин «уменьшать» или «ингибировать» может, в общем, относиться к способности одного или более чем одного AON или композиций «снижать» релевантный физиологический или клеточный ответ, такой как симптом заболевания или состояния, описанного в данном документе, при измерении согласно традиционным методикам в области диагностики. Релевантные физиологические или клеточные ответы (in vivo или in vitro) будут очевидными специалистам в данной области и могут включать уменьшения симптомов или патологии заболевания, такого как рак, нейродегенеративные заболевания, легочные расстройства и другие воспалительные заболевания. «Уменьшение» ответа может быть статистически значимым по сравнению с ответом, продуцируемым без AON или контрольной композицией, и может включать уменьшение на 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 11%, 12%, 13%, 14%, 15%, 16%, 17%, 18%, 19%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100%, включая все целые числа между ними.
Длина AON может варьировать, при условии, что он способен к селективному связыванию с намеченным местом в пределах молекулы пре-мРНК. Длина таких последовательностей может определяться согласно методикам выбора, описанным в данном документе. В общем, AON будет от примерно 10 нуклеотидов в длину вплоть до примерно 50 нуклеотидов в длину. Однако будет понятно то, что в данном способе можно использовать любую длину нуклеотидов в пределах данного интервала. Предпочтительно длина AON составляет от 10 до 40, от 10 до 35, от 15 до 30 нуклеотидов в длину или от 20 до 30 нуклеотидов в длину, наиболее предпочтительно примерно от 25 до 30 нуклеотидов в длину. Например, данный олигомер может иметь 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 или 30 нуклеотидов в длину.
Термин «AON» в том виде, в котором он используется в данном документе, относится к линейной последовательности нуклеотидов или аналогов нуклеотидов, которая обеспечивает гибридизацию нуклеиновых оснований с последовательностью-мишенью в РНК посредством образования пар оснований по Уотсону-Крику, с образованием гетеродуплекса олигонуклеотид : РНК в пределах последовательности-мишени. Термины «AON», «олигомер» и «антисмысловое соединение» могут использоваться взаимозаменяемо для названия олигонуклеотида. Циклические субъединицы могут быть основаны на рибозе или другом пентозном сахаре, или, в некоторых воплощениях, на морфолино группе (см. описание морфолино олигонуклеотидов ниже). Также рассматриваются пептидные нуклеиновые кислоты (PNA), запертые нуклеиновые кислоты (LNA) и 2'-О-метилолигонуклеотиды, среди других антисмысловых агентов, известных в данной области.
Включенными являются не встречающиеся в природе AON или «аналоги олигонуклеотидов», включая AON или олигонуклеотиды, имеющие (i) модифицированную структуру остова, например, остов, отличный от стандартной фосфодиэфирной связи, находящейся во встречающихся в природе олиго- и полинуклеотидах, и/или (ii) модифицированные сахарные группировки, например, морфолино группировки, а не рибозные или дезоксирибозные группировки. Аналоги олигонуклеотидов поддерживают основания, способные к связыванию водородными связями посредством образования пар по Уотсону-Крику с основаниями стандартных полинуклеотидов, где остов аналога представляет основания таким образом, чтобы обеспечивать такое связывание водородными связями способом, специфичным в отношении последовательности, между молекулой аналога олигонуклеотида и основаниями в стандартном полинуклеотиде (например, одноцепочечная РНК или одноцепочечная ДНК). Предпочтительными аналогами являются аналоги, имеющие по существу незаряженный фосфоросодержащий остов.
Одним способом получения AON является метилирование положения 2'-гидроксирибозы, а включение фосфоротиоатного остова дает молекулы, которые имеют поверхностное сходство с РНК, но которые являются значительно более устойчивыми к нуклеазной деградации, хотя специалистам в области данного изобретения будут известны другие формы подходящих остовов, которые могут быть применимыми для целей данного изобретения.
Для того чтобы избежать деградации пре-мРНК во время образования дуплекса с AON, AON, используемые в данном способе, могут быть адаптированы для минимизации или предупреждения расщепления эндогенной РНКазой Н. Данное свойство является весьма предпочтительным, так как обработка РНК неметилированными олигомерами, либо внутриклеточными, либо в грубых экстрактах, которые содержат РНКазу Н, приводит к деградации дуплексов пре-мРНК:AON. В настоящем способе можно использовать любую форму модифицированных AON, которая способна обходить или не индуцировать такую деградацию. Устойчивость к нуклеазам может достигаться посредством модифицирования AON по изобретению таким образом, что они содержат частично ненасыщенную алифатическую углеводородную цепь и одну или более чем одну полярную или заряженную группу, включающую карбоксильные группы, сложноэфирные группы и спиртовые группы.
Примером AON, которые при нахождении с дуплексе с РНК не расщепляются клеточной РНКазой Н, являются 2'-О-метильные производные. Такие 2'-О-метил-олигорибонуклеотиды являются стабильными в клеточной среде и в животных тканях, и их дуплексы с РНК имеют более высокие значения Tm, чем у их рибо- или дезоксирибо-аналогов. В качестве альтернативы, нуклеазорезистентные AON по изобретению могут иметь фторированный по меньшей мере один из последних 3'-концевых нуклеотидов. Кроме того, альтернативно, нуклеазорезистентные AON по изобретению имеют фосфоротиоатные связи, осуществляющие связь между по меньшей мере двумя последними нуклеотидными основаниями 3'-конца, предпочтительно имеющие фосфоротиоатные связи, осуществляющие связь между последними четырьмя 3'-концевыми нуклеотидными основаниями.
Усиленное переключение сплайсинга также может достигаться с использованием альтернативных химических соединений олигонуклеотидов. Например, AON может быть выбран из списка, содержащего: фосфорамидатный или фосфородиамидатный морфолино олигомер (РМО); РМО-Х; РРМО; пептидную нуклеиновую кислоту (PNA); запертую нуклеиновую кислоту (LNA) и производные, включающие альфа-L-LNA, 2'-амино-LNA, 4'-метил-LMA и 4'-O-метил-LNA; нуклеиновые кислоты с этиленовым мостиком (ENA) и их производные; фосфоротиоатный олигомер; трицикло-ДНК олигомер (тцДНК); трициклофосфоротиоатный олигомер; 4'-О-метил-модифицированный олигомер (2'-ОМе); 2-О-метоксиэтил (2'-МОЕ); 2'-фтор-, 2'-фторарабино (FANA); незапертую нуклеиновую кислоту (UNA); термостабильную скрученную интеркалирующую нуклеиновую кислоту (TINA); гекситолнуклеиновую кислоту (HNA); циклогексенилнуклеиновую кислоту (CeNA); 2'-амино (2'-NH2); 2'-О-этиленаминовую или любую комбинацию вышеописанных в виде миксмеров или в виде гэпмеров. Для дальнейшего улучшения эффективности доставки вышеупомянутые модифицированные нуклеотиды часто конъюгируют с жирными кислотами/липидом/холестерином/аминокислотами/углеводами/полисахаридами/наночастицами и т.д. с сахарной группировкой или группировкой нуклеинового основания. Данные конъюгированные производные нуклеотидов также можно использовать для конструирования AON для пропуска экзонов. В индуцированной антисмысловым олигонуклеотидом модификации сплайсинга транскриптов человеческого гена RAGE обычно использовали один из олигорибонуклеотидов, PNA, 2ОМе- или МОЕ-модифицированные основания на фосфоротиоатном остове. Хотя 2ОМеАО и используют для конструирования олигонуклеотидов, из-за их эффективного поглощения in vitro при доставке в виде катионных липоплексов, данные соединения являются чувствительными к деградации нуклеазой и не считаются идеальными для in vivo или клинических применений. При использовании альтернативных химических соединений для получения AON по настоящему изобретению урацил (U) последовательностей, предложенных в данном документе, можно заменять тимином (Т).
Включенными в пределы AON по настоящему изобретению являются не встречающиеся в природе олигомеры или «аналоги олигонуклеотидов», включающие олигомеры, имеющие (i) модифицированную структуру остова, например, остов, отличный от стандартной фосфодиэфирной связи, находящейся во встречающихся в природе олиго- и полинуклеотидах, и/или (ii) модифицированные сахарные группировки, например, морфолино группировки, а не рибозные или дезоксирибозные группировки. Аналоги олигомеров поддерживают основания, способные к образованию водородных связей посредством образования пар по Уотсону-Крику со стандартными основаниями полинуклеотидов, где остов аналога представляет основания таким образом, чтобы обеспечивать такое образование водородных связей способом, специфичным в отношении последовательности, между молекулой аналога олигомера и основаниями в стандартном полинуклеотиде (например, одноцепочечной РНК или одноцепочечной ДНК). Предпочтительными аналогами являются аналоги, имеющие по существу неизменный фосфоросодержащий остов.
Антисмысловые олигонуклеотиды, которые не активируют РНКазу Н, можно получать согласно известным методикам (см., например, патент США 5149797). Такие AON, которые могут представлять собой дезоксирибонуклеотидные или рибонуклеотидные последовательности, просто содержат любую структурную модификацию, которая стерически мешает или предотвращает связывание РНКазы Н с молекулой дуплекса, содержащего олигомер в качестве его одного члена, структурная модификация которого по существу не мешает или нарушает образование дуплекса. Поскольку части олигомера, участвующие в образовании дуплекса, существенно отличаются от тех частей, которые участвуют в связывании с ними РНКазы Н, доступны многочисленные AON, которые не активируют РНКазу Н. Например, такие AON могут представлять собой олигомеры, в которых по меньшей мере один или все из образующих межнуклеотидные мостики остатков фосфата представляют собой модифицированные фосфаты, такие как метилфосфонаты, метилфосфоротиоаты, фосфороморфолидаты, фосфоропиперазидаты, боранофосфаты, амидные связи и фосфорамидаты. Например, каждый второй из образующих межнуклеотидные мостики остатков фосфата может быть модифицирован, как описано выше. В другом неограничивающем примере такие AON представляют собой молекулы, в которых по меньшей мере один или все из нуклеотидов содержат 2' низшую алкильную группировку (такую как, например, С1-С4 линейный или разветвленный, насыщенный или ненасыщенный алкил, такой как метил, этил, этенил, пропил, 1-пропенил, 2-пропенил и изопропил). Например, каждый второй из нуклеотидов может быть модифицирован, как описано.
В то время как описанные выше AON представляют собой предпочтительную форму AON по настоящему изобретению, настоящее изобретение включает другие олигомерные антисмысловые молекулы, включающие олигомерные миметики, такие как описанные ниже, но не ограничивающиеся ими.
Конкретные примеры предпочтительных полезных AON в данном изобретении включают олигомеры, содержащие модифицированные остовы или неприродные межнуклеозидные связи. Как определено в данном описании изобретения, олигомеры, имеющие модифицированные остовы, включают олигомеры, которые сохраняют атом фосфора в остове, и олигомеры, которые не имеют атома фосфора в остове. Для целей данного описания изобретения, и как иногда приводится в данной области, модифицированные олигомеры, которые не имеют атома фосфора в их межнуклеозидном остове, также могут рассматриваться как AON.
В других предпочтительных олигомерных миметиках и сахар, и межнуклеозидная связь, т.е. остов нуклеотидных звеньев, заменяются новыми группами. Звенья оснований сохраняются для гибридизации с подходящим соединением нуклеиновой кислоты-мишени. Одно такое олигомерное соединение - олигомерный миметик, который, как было показано, имеет превосходные свойства гибридизации, называется пептидной нуклеиновой кислотой (PNA). В соединениях PNA сахарный остов олигомера заменяется амидсодержащим остовом, в частности, аминоэтилглициновым остовом. Нуклеиновые основания сохраняются и связываются прямо или опосредованно с аза атомами азота амидной части остова.
Другой предпочтительной химией являются олигомерные соединения на основе фосфородиамидатморфолино олигомера (РМО), которые не деградируются любой известной нуклеазой или протеазой. Данные соединения являются незаряженными, не активируют активность РНКазы Н при связывании с нитью РНК и, как было показано, оказывают длительную модуляцию сплайсинга после введения in vivo (Summerton and Weller, Antisense Nucleic Acid Drug Development, 7, 187-197).
Модифицированные олигомеры также могут содержать одну или более чем одну замещенную сахарную группировку. Олигомеры также могут включать нуклеиновое основание (часто именуемое в данной области просто как «основание»), модификации или замены. Определенные нуклеиновые основания являются особенно полезными для увеличения аффинности связывания олигомерных соединений по изобретению. Они включают 5-замещенные пиримидины, 6-азапиримидины, N-2, N-6 и О-6 замещенные пурины, включая 2-аминопропиладенин, 5-пропинилурацил и 5-пропинилцитозин. Было показано то, что замены 5-метилцитозином увеличивают стабильность дуплекса нуклеиновой кислоты на 0,6-1,2°С, даже более конкретно при объединении с 2'-О-метоксиэтильными модификациями сахара.
Другая модификация олиомеров по изобретению включает химическое связывание с олигомером одной или более чем одной группировки или конъюгата, которые увеличивают активность, клеточное распределение или клеточное поглощение данного олигомера. Такие группировки включают липидные группировки, такие как холестериновая группировка, холиновая кислота, тиоэфир, например, гексил-S-тритилтиол, тиохолестерин, алифатическую цепь, например, додекандиольные или ундецильные остатки, фосфолипид, например, ди-гексадецил-рац-глицерин или триэтиламмония 1,2-ди-О-гексадецил-рац-глицеро-3-Н-фосфонат, полиамин или цепь полиэтиленгликоля, или адамантануксусная кислота, пальмитильная группировка, миристильная или октадециламиновая или гексиламино-карбонил-оксихолестериновая группировка, но не ограничиваются ими.
Проникающие в клетку пептиды добавляли к фосфородиамидатморфолино олигомерам для усиления поглощения в клетку и ядерной локализации. Было показано то, что разные пептидные метки влияют на эффективность поглощения и специфичность в отношении ткани-мишени, как продемонстрировано в Jearawiriyapaisarn et al. (2008), Mol. Ther. 16 9, 1624-1629.
В данном соединении нет необходимости однородно модифицировать все положения, и, на самом деле, в одно соединение или даже в один нуклеозид в пределах олигомера можно включать более чем одну из вышеупомянутых модификаций. Настоящее изобретение также включает AON, которые представляют собой химерные соединения. «Химерные» AON или «химеры» в контексте данного изобретения представляют собой AON, в частности, олигомеры, которые содержат две или более чем две химически отличные области, причем каждая составлена из по меньшей мере одного мономерного звена, т.е. нуклеотида в случае олигомерного соединения. Данные олигомеры типично содержат по меньшей мере одну область, где олигомер модифицируется таким образом, чтобы придавать олигомеру или AON повышенную устойчивость к нуклеазной деградации, повышенное поглощение в клетку, и дополнительную область для повышенной аффинности связывания в отношении нуклеиновой кислоты-мишени.
Активность AON и их вариантов можно оценивать согласно традиционным в данной области методикам. Например, сплайсоформы и уровни экспрессии рассматриваемых РНК и белков можно анализировать любым из широкого спектра хорошо известных способов для выявления сплайсоформ и/или экспрессии транскрибированной нуклеиновой кислоты или белка. Неограничивающие примеры таких способов включают ПЦР-ОТ (полимеразная цепная реакция, сопряженная с обратной транскрипцией) сплайсоформ РНК с последующим разделением ПЦР-продуктов по размеру, способы гибридизации нуклеиновых кислот, например, норзерн-блоттинг и/или применение чипов нуклеиновых кислот; способы амплификации нуклеиновых кислот; иммунологические способы выявления белков; способы очистки белков; и анализы функции или активности белков.
Уровни экспрессии РНК можно оценивать посредством получения мРНК/кДНК (т.е. транскрибированного полинуклеотида) из клетки, ткани или организма и посредством гибридизации данной мРНК/кДНК с эталонным полинуклеотидом, который является комплементарным анализируемой нуклеиновой кислоте или ее фрагменту. кДНК может, возможно, быть амплифицирована с использованием любого из целого ряда способов полимеразной цепной реакции или транскрипции in vitro перед гибридизацией с комплементарным полинуклеотидом; предпочтительно она не амплифицируется. Экспрессия одного или более чем одного транскрипта также может быть выявлена с использованием количественной ПЦР для оценки уровня экспрессии транскрипта(тов).
Согласно настоящему изобретению предложен AON, индуцирующий переключение сплайсинга транскрипта гена RAGE, клинически релевантные химические соединения олигомера и системы доставки для управления манипуляциями сплайсингом RAGE до терапевтических уровней. Существенные уменьшения количества полноразмерной мРНК RAGE и, следовательно, белка RAGE от транскрипции гена RAGE достигаются посредством:
а) уточнения олигомера in vitro с использованием линий клеток фибробластов посредством экспериментальной оценки (i) интронных энхансерных целевых мотивов, (ii) длины AON и разработки смесей олигомеров, (iii) выбора химического соединения и (iv) добавления добавления проникающих в клетку пептидов (СРР) для усиления доставки олигомера; и
b) подробной оценки нового подхода для генерации транскриптов RAGE с одним или более чем одним отсутствующим экзоном.
В таком качестве в данном документе демонстрируется то, что альтернативный сплайсинг пре-мРНК RAGE может модулироваться специфичными AON. Данным способом могут быть получены функционально значимые снижения количества полноразмерного (способного к сигнализации) белка RAGE и/или могут достигаться увеличения несигнализирующей сплайсоформы мРНК рецептора-ловушки RAGE (RAGE_v1) или других рецепторов-ловушек, снижая, посредством этого, тяжесть патологии, ассоциированной с такими заболеваниями, как нейродегенеративные заболевания, рак, легочные расстройства и другие воспалительные заболевания.
AON, используемые согласно данному изобретению, могут быть с удобством получены посредством хорошо известной методики твердофазного синтеза. Оборудование для такого синтеза продается несколькими поставщиками, включая, например, Applied Biosystems (Foster City, Calif.). Один способ осуществления синтеза олигомеров на модифицированной твердой подложке описывается в патенте США №4458066.
Дополнительно или альтернативно могут использоваться любые другие способы такого синтеза. Применение аналогичных методик хорошо известно для получения олигомеров, таких как фосфоротиоаты и алкилированные производные. В одном таком автоматизированном воплощении в качестве исходных веществ используют диэтил-фосфорамидиты, и они могут синтезироваться, как описано Beaucage, et al., (1981) Tetrahedron Letters, 22: 1859-1862.
AON по изобретению синтезируются in vitro и не включают антисмысловые композиции биологического происхождения или конструкции генетических векторов, разработанные для управления синтезом AON in vivo. Молекулы по данному изобретению также можно смешивать, инкапсулировать, конъюгировать или иным способом ассоциировать с другими молекулами, молекулярными структурами или смесями соединений, такими как, например, липосомы, молекулы, нацеленные на рецептор, пероральные, ректальные, местные или иные препараты, для помощи в поглощении, распределении и/или абсорбции.
AON по настоящему изобретению также можно использовать в качестве профилактического или терапевтического средства, которое может использоваться с целью лечения заболевания. Соответственно, в одном воплощении согласно настоящему изобретению предложены AON, которые связываются с выбранной мишенью в пре-мРНК RAGE для индукции эффективного и прогнозируемого пропуска экзона, как описано в данной документе, в терапевтически эффективном количестве, смешанные с фармацевтически приемлемым носителем, разбавителем или эксципиентом.
Согласно данному изобретению, следовательно, предложена фармацевтическая, профилактическая или терапевтическая композиция для лечения, предупреждения или купирования эффектов заболевания, ассоциированных с экспрессией RAGE у пациента, включающая:
a) один или более чем один AON, как описано в данном документе, и
b) один или более чем один фармацевтически приемлемый носитель и/или разбавитель.
Предпочтительно заболевание, ассоциированное с экспрессией RAGE, выбрано из списка, содержащего: неврологические расстройства, раковые заболевания; сердечно-сосудистые расстройства; расстройства пищеварения; дыхательные расстройства, скелетно-мышечные расстройства, расстройства соединительной ткани, почечные расстройства, половые расстройства, кожные расстройства, глазные расстройства и эндокринные расстройства.
В одной форме данного изобретения связанное с RAGE расстройство представляет собой сердечно-сосудистое расстройство, выбранное из следующей группы: атеросклероз, ишемическое заболевание сердца, миокардит, эндокардит, кардиомиопатия, острая ревматическая лихорадка, хроническая ревматическая болезнь сердца, заболевание сосудов мозга/инсульт, сердечная недостаточность, кальциноз сосудов, заболевание периферических сосудов и лимфангит.
В одной форме данного изобретения связанное с RAGE расстройство представляет собой расстройство пищеварительной системы, выбранное из следующей группы: периодонтит, эзофагит, гастрит, язва желудка и двенадцатиперстной кишки, болезнь Крона, язвенный колит, ишемический колит, энтерит и энтероколит, перитонит, алкогольная болезнь печени, гепатит, токсическое отравление печени, билиарный цирроз печени, фиброз/цирроз печени, неалкогольная болезнь печени/неалкогольный стеатогепатит (NAFLD/NASH), травма печени и восстановление после повреждения, травмы или хирургии печени.
В одной форме данного изобретения связанное с RAGE расстройство представляет собой раковое заболевание, выбранное из следующей группы: злокачественные новообразования губы, полости рта и глотки, злокачественные новообразования органов пищеварения, злокачественные новообразования дыхательных и внутригрудных органов, злокачественные новообразования кости и суставного хряща, меланома и другие злокачественные новообразования кожи, злокачественные новообразования мезотелиальной и мягкой ткани, злокачественное новообразование молочной железы, злокачественные новообразования женских половых органов, злокачественные новообразования мужских половых органов, злокачественные новообразования мочевых путей, злокачественные новообразования глаза, мозга и других частей центральной нервной системы, злокачественные новообразования щитовидной железы и других эндокринных желез, злокачественные новообразования лимфоидной, гематопоэтической и родственной ткани, злокачественные новообразования в прохо определенных, вторичных и/или неспецифичных местах.
В одной форме данного изобретения связанное с RAGE расстройство представляет собой неврологическое расстройство, и оно выбрано из следующей группы: воспалительные заболевания центральной нервной системы, системные атрофии, воздействующие, главным образом, на центральную нервную систему, экстрапирамидальные расстройства и расстройства движения, болезнь Паркинсона, демиелинизирующие заболевания центральной нервной системы, болезнь Альцгеймера, синдром Пика, деменция с тельцами Леви, эпилепсия, мигрень, невропатическая боль, диабетическая невропатия, полинейропатии, развитие и прогрессирование глиомы, травма спинного мозга, ишемическое поражение мозга/инсульт, травма мозга и восстановление от повреждения, травмы или хирургии мозга.
В одной форме данного изобретения связанное с RAGE расстройство представляет собой скелетно-мышечное расстройство, и оно выбрано из следующей группы: мышечная дистрофия, врожденная и приобретенная миопатия, полимиозит, тяжелая миастения, дерматомиозит, миозит с тельцами включения, мышечная атрофия и мышечное повреждение.
В одной форме данного изобретения связанное с RAGE расстройство представляет собой умственное расстройство, и оно выбрано из следующей группы: деменция, болезнь Альцгеймера, сосудистая деменция, аддикция, шизофрения, большое аффективное расстройство, депрессия, мания, биполярное расстройство и тревожное расстройство.
В одной форме данного изобретения связанное с RAGE расстройство представляет собой дыхательное (легочное) расстройство, и оно выбрано из следующей группы: острые инфекции верхних дыхательных путей, ринит, назофарингит, синусит, ларингит, грипп и пневмония, острый бронхит, острый бронхиолит, астма, хроническая обструктивная болезнь легких (ХОБЛ), бронхоэктаз, эмфизема, хронические заболевания легкого из-за внешних агентов, острый респираторный дистресс-синдром (ARDS), легочная эозинофилия, плевральная, легочная травма и восстановление от повреждения, травмы или хирургии легкого.
В одной форме данного изобретения связанное с RAGE расстройство представляет собой расстройство соединительной ткани, и оно выбрано из следующей группы: остеоартрит, инфекционный артрит, ревматоидный артрит, псориатические и энтеропатические артропатии, хронический полиартрит у детей, подагра и другие кристаллические артропатии, диабетическая артропатия, узелковый полиартериит, синдром Чарга-Стросса, слизисто-кожный лимфоузелковый синдром [Кавасаки], гиперчувствительный ангиит, синдром Гудпасчера, тромботическая микроангиопатия, гранулематоз Вегенера, синдром дуги аорты [Такаясу], гигантоклеточный артериит, ревматическая полимиалгия, микроскопический полиангиит, васкулит с гипокомплементемией, системная красная волчанка, дерматополимиозит, полимиозит, системый склероз, синдром CR(E)ST, синдром Сикка [Шегрена], смешанное заболевание соединительной ткани, болезнь Бехчета, травматическое повреждение мышцы, растяжение связки, растяжение сухожилия и перелом.
В одной форме данного изобретения связанное с RAGE расстройство представляет собой почечное расстройство, и оно выбрано из следующей группы: гломерулонефрит, нефрит, диабетическое почечное заболевание, интерстициальный нефрит, обструктивная и рефлюкс нефропатия, острая почечная недостаточность и хроническое почечное заболевание.
В одной форме данного изобретения связанное с RAGE расстройство представляет собой половое расстройство, и оно выбрано из следующей группы: простатит, гипертрофия простаты, дисплазия простаты, сальпингит, оофорит, воспаление органов таза (PID), поликистоз яичников, цервицит, дисплазия шейки матки, вагинит, вульвит.
В одной форме данного изобретения связанное с RAGE расстройство представляет собой кожное расстройство, выбранное из следующей группы: дерматит, экзема, обыкновенная пузырчатка/пемфигоид, псориаз, розовый лишай, лишай Вильсона, уртикария, мультиформная эритема, узелковая эритема, солнечный ожог, кератоз, образование язв на коже в связи с фотостарением, поверхностное повреждение кожи и открытая рана.
В одной форме данного изобретения связанное с RAGE расстройство представляет собой глазное расстройство, выбранное из следующей группы: кератит, конъюнктивит, ретинит, глаукома, склерит, эписклерит, хориоретинальное воспаление, диабетическая ретинопатия, макулярный отек, ретинопатия недоношенных, неврит зрительного нерва, травма глаза и восстановление от повреждения, травмы или хирургии глаза.
В одной форме данного изобретения связанное с RAGE расстройство представляет собой эндокринное расстройство, выбранное из следующей группы: сахарный диабет, инсулинорезистентность, нарушение переносимости глюкозы и тиреоидит.
Данная композиция может содержать примерно от 1 нМ до 1000 нМ каждого из желательных AON по изобретению. Предпочтительно данная композиция может содержать примерно от 1 нМ до 500 нМ, от 10 нМ до 500 нМ, от 50 нМ до 750 нМ, от 10 нМ до 500 нМ, от 1 нМ до 100 нМ, от 1 нМ до 50 нМ, от 1 нМ до 40 нМ, от 1 нМ до 30 нМ, от 1 нМ до 20 нМ, наиболее предпочтительно от 1 нМ до 10 нМ каждого из AON по изобретению.
Данная композиция может содержать примерно 1 нМ, 2 нМ, 3 нМ, 4 нМ, 5 нМ, 6 нМ, 7 нМ, 8 нМ, 9 нМ, 10 нМ, 20 нМ, 50 нМ, 75 нМ, 100 нМ, 150 нМ, 200 нМ, 250 нМ, 300 нМ, 350 нМ, 400 нМ, 450 нМ, 500 нМ, 550 нМ, 600 нМ, 650 нМ, 700 нМ, 750 нМ, 800 нМ, 850 нМ, 900 нМ, 950 нМ или 1000 нМ каждого из желательных AON по изобретению.
Согласно настоящему изобретению дополнительно предложен один или более чем один AON, адаптированный для помощи в профилактическом или терапевтическом лечении, предупреждении или уменьшении интенсивности симптомов заболевания, такого как заболевание или патология, связанная с экспрессией RAGE, в подходящей форме для доставки пациенту.
Фраза «фармацевтически приемлемый» относится к химическим соединениям и композициям, которые являются физиологически переносимыми и типично не продуцируют аллергическую или аналогичную нежелательную реакцию, такую как расстройство желудка и тому подобное, при введении пациенту. Термин «носитель» относится к разбавителю, адъюванту, эксципиенту или наполнителю, с которым вводится данное соединение. Такие фармацевтические носители могут представлять собой стерильные жидкости, такие как вода и масла, включающие жидкости нефтяного, животного, растительного или синтетического происхождения, такие как арахисовое масло, соевое масло, минеральное масло, кунжутное масло и тому подобные. В качестве носителей предпочтительно используются вода или физиологические растворы, водная декстроза и растворы глицерина, в частности, для инъецируемых растворов. Подходящие фармацевтические носители описываются в Martin, Remington's Pharmaceutical Sciences, 18th Ed., Mack Publishing Co., Easton, PA, (1990).
В более конкретной форме данного изобретения предложены фармацевтические композиции, содержащие терапевтически эффективные количества одного или более чем одного AON по изобретению совместно с фармацевтически приемлемыми разбавителями, консервантами, солюбилизаторами, эмульгаторами, адъювантами и/или носителями. Такие композиции включают разбавители с разным содержанием буфера (например, Tris-HCl, ацетатный, фосфатный), рН и ионной силой, и добавки, такие как детергенты и солюбилизаторы (например, Tween 80, полисорбат 80), антиоксиданты (например, аскорбиновая кислота, метабисульфит натрия), консерванты (например, тиомерсал, бензиловый спирт) и объемообразующие вещества (например, лактоза, маннит). Данное вещество может быть включено в препараты в виде частиц полимерных соединений, таких как полимолочная кислота, полигликолевая кислота и т.д. или в липосомы. Также может использоваться гиалуроновая кислота. Такие композиции могут влиять на физическое состояние, стабильность, скорость высвобождения in vivo и скорость клиренса in vivo настоящих белков и производных. См., например, Martin, Remington's Pharmaceutical Sciences, 18th Ed. (1990, Mack Publishing Co., Easton, PA 18042) страницы 1435-1712, которая включена в данный документ посредством ссылки. Данные композиции могут быть получены в жидкой форме или могут находиться в форме сухого порошка, как, например, в лиофилизированной форме.
Будет понятно, что фармацевтические композиции, предложенные согласно настоящему изобретению, можно вводить любыми способами, известными в данной области. Предпочтительно фармацевтические композиции для введения вводятся посредством инъекции, перорально, местно или посредством легочного или назального пути. Подходящий путь может быть определен обычным специалистом в данной области сообразно состоянию субъекта, которого лечат.
В некоторых воплощениях AON по данному раскрытию могут доставляться легочным или назальным путями (например, посредством распыляемого физиологического раствора, включающего AON). Наивысшая эндогенная экспрессия мРНК RAGE в тканях здорового человека обнаруживается в легком, и она доступна через дыхательные пути. Ингалируемые олигонуклеотиды представляют собой перспективное терапевтическое воздействие против респираторных заболеваний. Дыхательные пути уникальным образом выстланы легочными поверхностно-активными веществами, которые состоят, главным образом, из цвиттер-ионных липидов. Данные поверхностно-активные липиды обладают катионными свойствами при рН респираторного тракта. При ингаляции анионных олигонуклеотидов они имеют тенденцию к поглощению поверхностно-активными веществами, приводя к частицам в новом препарате, которые, как предполагалось, эффективно поглощаются бронхиальными и альвеолярными эпителиальными в клетки легкого. Примечательно, что было показано то, что AON способны выдерживать процесс распыления.
В некоторых воплощениях AON более предпочтительно доставляются посредством внутривенного, внутриартериального, внутрибрюшинного, внутримышечного или подкожного путей введения. Сосудистое или внесосудистое кровообращение, кровеносная или лимфатическая система и спинномозговая жидкость являются некоторыми неограничивающими сайтами, где могут вводиться AON.
В некоторых воплощениях может использоваться прямая доставка в ЦНС, например, в качестве путей введения можно использовать внутримозговое, вентрикулярное или подоболочечное введение.
Препараты для местного введения включают препараты, в которых олигомеры по данному раскрытию находятся в смеси со средством местной доставки, таким как липиды, липосомы, жирные кислоты, сложные эфиры жирных кислот, стероиды, хелаторы и поверхностно-активные вещества. Липиды и липосомы включают нейтральные (например, диолеоилфосфатидилэтаноламин (DOPE), димиристоилфосфатидилхолин (DMPC), дистеароилфосфатидилхолин), отрицательно заряженные (например, димиристоилфосфатидилглицерин (DMPG)) и катионные (например, диолеоилтетраметиламинопропил (DOTAP) и диолеоилфосфатидилэтаноламин (DOTMA)). Для местного или другого введения олигомеры по данному раскрытию могут быть инкапсулированы в липосомы или могут образовать комплексы с ними, в частности, с катионными липосомами. В качестве альтернативы, олигомеры могут быть комплексированы с липидами, в частности, с катионными липидами. Жирные кислоты и сложные эфиры, их фармацевтически приемлемые соли и их применения дополнительно описываются в патенте США №6287860 и/или в заявке на патент США с сер. №09/315298, поданной 20 мая 1999 г.
В некоторых воплощениях AON по данному раскрытию могут доставляться чрескожными способами (например, посредством включения AON, например, в эмульсии, возможно с такими AON, упакованными в липосомы). Такие чрескожные и опосредованные эмульсией/липосомами способы доставки описыватся в данной области для доставки AON, например, в патенте США №6965025.
AON, описанные в данном документе, также могут доставляться посредством имплантируемого устройства. Конструирование такого устройства является известным в данной области способом, например, конструирование синтетического импланта, описанное, например, в патенте США №6969400.
Композиции и препараты для перорального введения включают порошки или гранулы, микрочастицы, наночастицы, суспензии или растворы в воде или неводных средах, капсулы, гелевые капсулы, пакетики, таблетки или минитаблетки. Могут быть желательными загустители, корригенты, разбавители, эмульгаторы, диспергирующие вспомогательные средства или связующие вещества. Пероральными препаратами являются препараты, в которых олигомеры по данному раскрытию вводятся в сочетании с одним или более чем одним усилителем проникновения, поверхностно-активным веществом и хелатором. Поверхностно-активные вещества включают жирные кислоты и/или их сложные эфиры или соли, желчные кислоты и/или их соли. Желчные кислоты/соли и жирные кислоты и их применения дополнительно описываются в патенте США №6287860. В некоторых воплощениях согласно настоящему раскрытию предложены комбинации усилителей проникновения, например, жирных кислот/солей в комбинации с желчными кислотами/солями. Типичной комбинацией является натриевая соль лауриновой кислоты, каприновой кислоты и UDCA (урсодезоксихолиевая кислота). Другие усилители проникновения включают полиоксиэтилен-9-простой лауриловый эфир, полиоксиэтилен-20-простой цетиловый эфир. Олигомеры по данному раскрытию могут доставляться перорально, в гранулярной форме, включая частицы, высушенные распылением, или в комплексе с образованием микро- или наночастиц. Агенты комплексирования олигомеров и их применения дополнительно описываются в патенте США №6287860. Пероральные препараты для олигомеров и их получение подробно описываются в US 6887906, 09/315298, поданных 20 мая 1999, и/или в US 20030027780.
Композиции и препараты для парентерального, подоболочечного или интравентрикулярного введения могут включать стерильные водные растворы, которые также могут содержать буферы, разбавители и другие подходящие добавки, такие как усилители проникновения, соединения-носители и другие фармацевтически приемлемые носители или эксципиенты, но не ограничиваются ими.
Доставка терапевтически полезного количества AON может достигаться ранее опубликованными способами. Например, внутриклеточная доставка AON может осуществляться посредством композиции, содержащей смесь AON и эффективного количества блоксополимера. Пример данного способа описывается в заявке на патент США US 20040248833. Другие способы доставки AON в ядро описываются в Mann CJ et al. (2001) Proc, Natl. Acad. Science, 98 (1) 42-47 и в Gebski et al. (2003) Human Molecular Genetics, 12 (15): 1801-1811. Способ введения молекулы нуклеиновой кислоты в клетку посредством экспрессионного вектора либо в виде голой ДНК, либо в комплексе с липидными носителями описывается в US 6806084.
Может быть желательной доставка AON в коллоидной дисперсионной системе. Коллоидные дисперсионные системы включают макромолекулярные комплексы, нанокапсулы, микросферы, шарики и системы на основе липидов, включающие эмульсии типа «масло в воде», мицеллы, смешанные мицеллы и липосомы или липосомные препараты. Данные коллоидные дисперсионные системы можно использовать в изготовлении терапевтических фармацевтических композиций.
Липосомы представляют собой искусственные мембранные везикулы, которые являются полезными в качестве носителей доставки in vitro и in vivo. Данные препараты могут иметь нетто катионные, анионные или нейтральные характеристики заряда и имеют полезные характеристики для способов доставки in vitro, in vivo и ex vivo. Было показано то, долю что большие однослойные везикулы могут инкапсулировать значительную процентную водного буфера, содержащего большие макромолекулы. РНК и ДНК могут быть инкапсулированы в пределах водного внутреннего содержимого и могут доставляться в клетки в биологически активной форме (Fraley, et al., Trends Biochem. Sci. 6: 77, 1981).
Для того чтобы липосома была эффективным носителем для переноса генов, должны присутствовать следующие характеристики: (1) инкапсуляция интересующих AON с высокой эффективностью при отсутствие нарушения их биологической активности; (2) предпочтительное и существенное связывание с клеткой-мишенью по сравнению с клетками, не являющимися мишенями; (3) доставка водного содержимого везикулы в цитоплазму клетки-мишени с высокой эффективностью; и (4) точная и эффективная экспрессия генетической информации (Mannino, et al., Biotechniques, 6: 682, 1988). Композиция липосомы обычно представляет собой комбинацию фосфолипидов, в частности, фосфолипидов с высокой температурой фазового перехода, обычно в комбинации со стероидами, особенно холестерином. Также можно использовать другие фосфолипиды или другие липиды. Физические характеристики липосом зависят от рН, ионной силы и присутствия дивалентных катионов. Катионные липосомы представляют собой положительно заряженные липосомы, которые, как считается, взаимодействуют с отрицательно заряженными молекулами ДНК с образованием стабильного комплекса. Считается, что липосомы, которые являются рН-чувствительными или отрицательно заряженными, захватывают ДНК, а не образуют с ней комплекс. И катионные, и некатионные липосомы использовали для доставки ДНК в клетки.
Липосомы также включают «стерически стабилизированные» липосомы - термин, который в том виде, в котором он используется в данном документе, относится к липосомам, содержащим один или более чем один специализированный липид, который, при включении в липосомы, приводит к повышенному времени жизни в системе кровообращения относительно липосом, лишенных таких специализированных липидов. Примерами стерически стабилизированных липосом являются липосомы, в которых часть образующей везикулу липидной части липосомы содержит один или более чем один гликолипид или представляет собой производное с одним или более чем одним гидрофильным полимером, таким как группировка полиэтиленгликоля (PEG). Липосомы и их применения дополнительно описываются в US 6287860.
AON, описанные в данном документе, также могут доставляться посредством имплантируемого устройства. Конструирование такого устройства является известным в данной области способом, с использованием, например, конструкции синтетического импланта, описанной, например, в патенте США №6969400, содержание которого включается в данный документ во всей его полноте.
Антисмысловые олигонуклеотиды могут вводиться в клетки с использованием известных в данной области методик (например, трансфекция, электропорация, слияние, липосомы, коллоидные полимерные частицы и вирусные, и невирусные векторы, а также другие способы, известные в данной области). Выбранный способ доставки будет зависеть по меньшей мере от клеток, подлежащих обработке, и от расположения данных клеток, и будет очевиден специалисту. Например, локализация может достигаться посредством липосом со специфичными маркерами на поверхности для направления липосомы, прямой инъекции в ткань, содержащую клетки-мишени, поглощения, опосредованного специфичным рецептором, или тому подобного.
Как известно в данной области, AON могут доставляться с использованием, например, способов с участием липосомоопосредованного поглощения, липидных конъюгатов, опосредованного полилизином поглощения, опосредованного наночастицами поглощения и опосредованного рецептором эндоцитоза, а также дополнительными неэндоцитозными способами доставки, такими как микроинъекция, пермеабилизация (например, пермеабилизация стрептолизином-О, пермеабилизация анионным пептидом), электропорация и разные неинвазивные неэндоцитозные способы доставки, которые известны в данной области (см. Dokka and Rojanasakul, Advanced Drug Delivery Reviews 44, 35-49, включенную посредством ссылки во всей ее полноте).
AON также можно объединять с другими фармацевтически приемлемыми носителями или разбавителями с получением фармацевтической композиции. Подходящие носители и разбавители включают изотоничные физиологические растворы, например, фосфатно-солевой буферный раствор. Данная композиция может быть приготовлена для парентерального, внутримышечного, внутривенного, подкожного, внутриглазного, перорального или чрескожного введения.
Описанные пути введения предназначены только в качестве руководства, так как квалифицированный практик легко сможет определить оптимальный путь введения и любую дозировку для любого конкретного животного и состояния.
Пытались осуществить многие подходы для введения нового функционального генетического материала в клетки как in vitro, так и in vivo (Friedmann (1989) Science, 244: 1275-1280). Данные подходы включают интеграцию гена, подлежащего экспрессии, в модифицированные ретровирусы (Friedmann (1989) выше; Rosenberg (1991) Cancer Research 51 (18), suppl.: 5074S-5079S); интеграцию в неретровирусные векторы (Rosenfeld, et al. (1992) Cell, 68: 143-155; Rosenfeld, et al. (1991) Science, 252: 431-434); или доставку трансгена, связанного с гетерологичным промоторным-энхансерным элементом, посредством липосом (Friedmann (1989), выше; Brigham, et al. (1989) Am. J. Med. Sci., 298: 278-311; Nabel, et al. (1990) Science, 249: 1285-1288; Hazinski, et al. (1991) Am. J. Resp. Cell Molec. Biol., 4: 206-209 и Wang and Huang (1987) Proc. Natl. Acad. Sci. (USA), 84: 7851-7855); связывание с лигандспецифичными транспортными системами на основе катионов (Wu and Wu (1988) J. Biol. Chem., 263: 14621-14624) или применение голой ДНК, экспрессионных векторов (Nabel et al. (1990), выше); Wolff et al. (1990) Science, 247: 1465-1468). Прямая инъекция трансгенов в ткань продуцирует только локализованную экспрессию (Rosenfeld (1992) выше); Rosenfeld et al. (1991) выше; Brigham et al. (1989) выше; Nabel (1990) выше; и Hazinski et al. (1991) выше). Группа Brigham et al. (Am. J. Med. Sci. (1989) 298: 278-311 and Clinical Research (1991) 39 (реферат)) описали in vivo трансфекцию только легких мышей после либо внутривенного, либо внутритрахеального введения комплекса ДНК-липосома. Примером обзорной статьи по методикам генотерапии человека является: Anderson, Science (1992) 256: 808-813; Barteau et al. (2008), Curr Gene Ther; 8 (5): 313-23; Mueller et al. (2008). Clin Rev Allergy Immunol; 35 (3): 164-78; Li et al. (2006) Gene Ther., 13 (18): 1313-9; Simoes et al. (2005) Expert Opin Drug Deliv; 2 (2): 237-54.
AON по изобретению охватывают любые фармацевтически приемлемые соли, сложные эфиры или соли таких эфиров, или любое другое соединение, которое, при введении животному, включая человека, способно предоставлять (прямо или опосредованно) биологически активный метаболит или его остаток. Соответственно, в качестве примера, данное раскрытие также направлено на пролекарства и фармацевтически приемлемые соли соединений по изобретению, фармацевтически приемлемые соли таких пролекарств и другие биоэквиваленты.
Термин «фармацевтически приемлемые соли» относится к физиологически и фармацевтически приемлемым солям соединений по данному изобретению: т.е. солям, которые сохраняют желательную биологическую активность родительского соединения и не придают ему нежелательных токсикологических эффектов. Для олигомеров предпочтительные примеры фармацевтически приемлемых солей включают (а) соли, образованные с катионами, такими как натрий, калий, аммоний, магний, кальций, полиамины, такие как спермин и спермидин и т.д.; (b) соли присоединения кислоты, образованные с неорганическими кислотами, например, соляной кислотой, бромистоводородной кислотой, серной кислотой, фосфорной кислотой, азотной кислотой и тому подобными; (с) соли, образованные с органическими кислотами, такими как, например, уксусная кислота, щавелевая кислота, винная кислота, янтарная кислота, малеиновая кислота, фумаровая кислота, глюконовая кислота, лимонная кислота, яблочная кислота, аскорбиновая кислота, бензойная кислота, дубильная кислота, пальмитиновая кислота, альгиновая кислота, полиглутаминовая кислота, нафталинсульфоновая кислота, метансульфоновая кислота, п-толуолсульфоновая кислота, нафталиндисульфоновая кислота, полигалактуроновая кислота и тому подобные; и (d) соли, образованные из элементарных анионов, таких как хлор, бром и йод, но не ограничиваются ими. Фармацевтические композиции по настоящему изобретению могут вводиться целым рядом способов, в зависимости от того, является ли желательным местное или системное лечение, и от области, подлежащей лечению. Введение может быть местным (включая глазное и на слизистую, а также ректальную доставку), легочным, например, посредством ингаляции или вдувания порошков или аэрозолей (включая посредством небулайзера, внутритрахеальное, интраназальное, эпидермальное и чрескожное), пероральным или парентеральным. Парентеральное введение включает внутривенную, внутриартериальную, подкожную, внутрибрюшинную или внутримышечную инъекцию или инфузию; или внутричерепное, например, подоболочечное или внутрижелудочковое введение. Считается, что особенно полезными для перорального введения являются олигомеры с по меньшей мере одной 2'-O-метоксиэтильной модификацией. Предпочтительно AON доставляется посредством подкожного или внутривенного пути.
Фармацевтические препараты по настоящему изобретению, которые могут быть с удобством представлены в стандартной лекарственной форме, могут быть получены согласно традиционным методикам, хорошо известным в фармацевтической промышленности. Такие методики включают стадию приведения в ассоциацию активных ингредиентов с фармацевтическим(кими) носителем(лями) или эксципиентом(тами). В общем, данные препараты получают посредством приведения в однородную и тесную ассоциацию активных ингредиентов с жидкими носителями или мелко измельченными твердыми носителями, или и теми, и другими, и затем, если это необходимо, формовки продукта.
В одном воплощении AON вводится в эффективном количестве и эффективным способом для того, чтобы приводить к пиковой концентрации в крови по меньшей мере 200-400 нМ AON. Типично вводят одну или более чем одну дозу AON, обычно с равными промежутками времени, в течение периода примерно от одной до двух недель. Предпочтительные дозы для перорального введения составляют от примерно 1 мг до 1000 мг олигомера на 70 кг. В некоторых случаях могут быть необходимы дозы больше, чем 1000 мг олигомера/пациента. Для в.в. введения предпочтительные дозы составляют от примерно 0,5 мг до 1000 мг олигомера на 70 кг. Для внутривенного или подкожного введения AON может вводиться в дозировке примерно 120 мг/кг ежесуточно или еженедельно.
AON может вводиться с равными промежутками времени в течение короткого периода времени, например, ежесуточно в течение двух недель или меньше. Однако в некоторых случаях данный олигомер вводится периодически в течение более длительного периода времени. После введения может следовать или происходить одновременно с ним введение антибиотика или другое терапевтическое лечение. Схема лечения может корректироваться (доза, частота, путь и т.д.), как указано, на основе результатов иммуноанализов, других биохимичеких анализов и физиологического обследования субъекта, которого лечат.
Дозирование зависит от тяжести и восприимчивости болезненного состояния, подлежащего лечению, причем курс лечения длится от нескольких суток до нескольких месяцев, или пока не осуществляется излечение, или не достигается ослабление болезненного состояния. Оптимальные схемы дозирования могут быть рассчитаны из измерений накопления лекарственного средства в организме пациента. Обычные специалисты могут легко определять оптимальные дозировки, методологии дозирования и частоту повторения. Оптимальные дозировки могут варьировать, в зависимости от относительной эффективности индивидуальных олигомеров, и обычно могут оцениваться на основе ЕС50 (полумаксимальная эффективная концентрация), обнаруженных как эффективные в животных моделях in vitro и in vivo. В общем, дозировка составляет от 0,01 мкг до 100 г на кг массы тела, и ее можно давать один или более чем один раз ежесуточно, еженедельно, ежемесячно или ежегодно, даже один раз каждые 2-20 лет. Обычные специалисты в данной области могут легко определять частоты повторения для дозирования на основе измеренного времени нахождения и концентраций лекарственного средства в жидкостях организма или тканях. После успешного лечения может быть желательным то, что пациент подвергается поддерживающей терапии для предупреждения рецидива болезненного состояния, где олигомер вводится в поддерживающих дозах, варьирующих от 0,01 мкг до 100 г на кг массы тела, от одного или более чем одного раза в сутки до одного раза каждые 20 лет.
Эффективная схема лечения in vivo с использованием AON по данному изобретению может варьировать согласно продолжительности, дозе, частоте и пути введения, а также состоянию субъекта, подвергающегося лечению (т.е. профилактическое введение относительно введения в ответ на локализованную или системную инфекцию). Соответственно, такая терапия in vivo часто будет требовать мониторинга посредством подходящих анализов для конкретного типа расстройства, подвергающегося лечению, и соответствующих корректировок в дозе или схеме лечения для того, чтобы добиться оптимального терапевтического результата.
Лечение можно отслеживать, например, посредством общих показателей заболевания, известных в данной области. Эффективность AON по изобретению, вводимых in vivo, можно определять из биологических образцов (ткань, кровь, моча и т.д.), отобранных у субъекта до, во время и после введения AON. Анализы таких образцов включают (1) отслеживание присутствия или отсутствия образования гетеродуплекса с целевыми и нецелевыми последовательностями с использованием методик, известных специалистам в данной области, например, анализа электрофоретической подвижности в геле; (2) отслеживание количества мутантной мРНК по отношению к контрольной нормальной мРНК или белку при определении стандартными методиками, такими как ПЦР-ОТ, норзерн-блоттинг, ELISA (твердофазный иммуноферментный анализ) или вестерн-блоттинг.
Внутриядерная доставка олигомера является главным вызовом для AON. Разные проникающие в клетку пептиды (СРР) в разной степени локализуют РМО в разных условиях и линиях клеток, и новые СРР оценивались авторами данного изобретения на их способность доставлять РМО в клетки-мишени. Термины СРР или «пептидная группировка, которая усиливает клеточное поглощение» используются взаимозаменяемо и относятся катионным проникающим в клетку пептидам, также именуемым «транспортными пептидами», «пептидами-носителями» или «пептидными доменами трансдукции». Данные пептиды, как показано в данном документе, имеют способность индуцировать проникновение в клетку в пределах примерно или по меньшей мере примерно 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% клеток данной популяции культуры клеток и обеспечивают транслокацию макромолекул в пределах многих тканей in vivo при системном введении. СРР хорошо известны в данной области и раскрываются, например, в заявке США №2010/0016215, которая включается посредством ссылки во всей ее полноте.
Согласно настоящему изобретению, следовательно, предложены AON по настоящему изобретению в комбинации с проникающими в клетку пептидами для изготовления терапевтических фармацевтических композиций.
Согласно еще одному другому аспекту данного изобретения предложен один или более чем один AON, как описано в данном документе, для применения в терапии на основе AON. Предпочтительно данная терапия осуществляется в отношении состояния, связанного с экспрессией RAGE. Более предпочтительно терапия в отношении состояния, связанного с экспрессией RAGE, представляет собой терапию против заболевания, выбранного из рака, нейродегенеративных заболеваний, легочных расстройств и воспалительных заболеваний.
Более конкретно, данный AON может быть выбран из группы, состоящей из любого одного или более чем одного AON, перечисленного в любой из Таблиц 3а-3d и/или SEQ ID NO: 1-31, и их комбинаций или смесей. Более предпочтительно данный AON представляет собой SEQ ID NO: 11, 18, 19 или 20. Это включает последовательности, которые могут гибридизоваться с такими последовательностями при жестких условиях гибридизации, комплементарные им последовательности, последовательности, содержащие модифицированные основания, модифицированные остовы и их функциональные усечения или удлинения, которые обладают или модулируют активность процессинга пре-мРНК в транскрипте гена RAGE.
Данное изобретение также распространяется на комбинацию двух или более чем двух AON, способных к связыванию с выбранной мишенью для индукции исключения экзона в транскрипте гена RAGE. Данная комбинация может представлять собой смесь двух или более чем двух AON, конструкцию, содержащую два или более чем два AON, соединенных друг с другом для применения в терапии на основе AON. Данная комбинация AON предпочтительно представляет собой комбинацию SEQ ID NO: 11 и 10 или SEQ ID NO: 11 и 13.
Согласно данному изобретению предложен способ лечения, предупреждения или купирования эффектов заболевания, ассоциированного с экспрессией RAGE, включающий стадию:
а) введения пациенту эффективного количества одного или более чем одного AON, или фармацевтической композиции, содержащей один или более чем один AON, как описано в данном документе.
Кроме того, согласно данному изобретению предложен способ лечения, предупреждения или купирования эффектов рака, нейродегенеративных заболеваний, легочных заболеваний и воспалительных заболеваний, включающий стадию:
а) введения пациенту эффективного количества одного или более чем одного AON, или фармацевтической композиции, содержащей один или более чем один AON, как описано в данном документе.
Предпочтительно данную терапию используют для уменьшения уровней функционального белка RAGE посредством стратегии пропуска экзона. Уменьшение уровней RAGE предпочтительно достигается посредством уменьшения уровня транскриптов через модификацию сплайсинга пре-мРНК в транскрипте гена RAGE или его части.
Уменьшение RAGE будет предпочтительно приводить к уменьшению количества, продолжительности или тяжести симптомов связанного с RAGE состояния или патологии, таких как нейродегенеративные заболевания, рак, легочные расстройства и воспалительные заболевания.
Термин «лечение» субъекта (например, млекопитающего, такого как человек) или клетки в том виде, в котором он используется в данном документе, представляет собой любой тип вмешательства, используемый в попытке изменить природное течение у индивида или клетки. Лечение включает введение фармацевтической композиции, но не ограничивается им, и может осуществляться либо профилактически, либо после инициации патогенного события или контакта с возбудителем. Также включаются «профилактические» лечения, которые могут быть направлены на уменьшения скорости прогрессирования заболевания или состояния, которое лечат, задержку начала данного заболевания или состояния, или уменьшение тяжести его начала. «Лечение» или «профилактика» не обязательно указывает полное устранение, излечение или предупреждение заболевания или состояния, или симптомов, ассоциированных с ним.
Согласно другому аспекту данного изобретения предложено применение одного или более чем одного AON, как описано в данном документе, в изготовлении лекарственного средства для модуляции или контроля заболевания, ассоциированного с экспрессией RAGE.
Согласно данному изобретению также предложено применение очищенных или выделенных AON, как описано в данном документе, для изготовления лекарственного средства для лечения заболевания, ассоциированного с экспрессией RAGE.
Предложено применение очищенных или выделенных AON, как описано в данном документе, для изготовления лекарственного средства для лечения, предупреждения или купирования эффектов заболевания, ассоциированного с экспрессией RAGE.
Предпочтительно связанная с RAGE патология или заболевание представляет собой нейродегенеративные заболевания, рак, легочные расстройства или воспалительные заболевания.
Данное изобретение распространяется согласно его еще одному другому аспекту на кДНК или клонированные копии последовательностей AON по изобретению, а также на векторы, содержащие последовательности AON по изобретению. Данное изобретение дополнительно также распространяется на клетки, содержащие такие последовательности и/или векторы.
AON по настоящему изобретению может вводиться совместно с другой терапевтической молекулой. Например, данный AON может вводиться со вторым терапевтическим средством, которое представляет собой соединение, такое как пептид, который способен модулировать цитозольный хвост RAGE. Такие соединения включают следующие: IQGAP-1, прозрачный-1, протеинкиназа С дзета (PKCζ), Dock7, MyD88, TIRAP, ERK1/2, обонятельный рецептор 2Т2, АДФ/АТФ транслоказа 2, протеинфосфатаза 1G, IRAK4, белок DJ-1 (PARK7), кальпонин-3, дребрин, филамин В, родственный Ras белок Rab-13, радиксин/эзрин/моезин, протеолипидный белок 2, коронин, S100 А11, субъединица альфа сукцинил-КоА лигазы [ГДФ-образующая], белок, взаимодействующий с Hsc70, ингибитор апоптоза 5, нейропилин, фактор стимуляции расщепления, белок 2, связанный с рецептором фактора роста, субъединица бета sec61 или Nck1.
Согласно данному изобретению также предложены наборы для лечения, предупреждения или купирования заболевания или состояния, ассоциированного с экспрессией RAGE у пациента, которые содержат по меньшей мере выделенный или очищенный AON для модификации сплайсинга пре-мРНК в транскрипте гена RAGE или его части, упакованный в подходящий контейнер, наряду с инструкциями для его применению.
В предпочтительном воплощении данные наборы будут содержать по меньшей мере один AON, как описано в данном документе, любую одну или более чем одну SEQ ID NO: 1-31 и/или последовательности, изложенные в любой из Таблиц 3a-3d, или смесь AON, как описано в данном документе. Данные наборы также могут содержать второстепенные реактивы, такие как буферы, стабилизаторы и т.д. Более предпочтительно AON представляет собой SEQ ID NO: 11, 18, 19 или 20.
Следовательно, предложен набор для лечения, предупреждения или купирования заболевания или состояния, ассоциированного с экспрессией RAGE у пациента, который содержит по меньшей мере AON, описанный в данном документе, любую одну или более чем одну SEQ ID NO: 1-31 и/или последовательности, изложенные в любой из Таблиц 3a-3d, и их комбинации или смеси, упакованный в подходящий контейнер, наряду с инструкциями для его применения. Более предпочтительно AON представляет собой SEQ ID NO: 11, 18, 19 или 20. Данная комбинация AON предпочтительно представляет собой комбинацию SEQ ID NO: 11 и 10 или SEQ ID NO: 11 и 13.
Предпочтительно данное заболевание или состояние выбрано из списка, содержащего: рак, нейродегенеративные заболевания, легочные расстройства или воспалительные заболевания.
Содержимое данного набора может быть лиофилизированным, и данный набор может дополнительно содержать подходящий растворитель для растворения лиофилизированных компонентов. Индивидуальные компоненты данного набора были бы упакованы в отдельные контейнеры, и с такими контейнерами может быть ассоциировано объявление в виде, предписанном правительственным агенством, регулирующим изготовление, применение или продажу фармацевтических средств или биологических продуктов, причем данное объявление отражает одобрение данным агентством изготовления, применения или продажи для введения человеку.
При предоставлении компонентов набора в одном или более чем одном жидком растворе данный жидкий раствор может представлять собой водный раствор, например, стерильный водный раствор. Для применения in vivo экспрессионная конструкция может быть приготовлена в фармацевтически приемлемой отбираемой шприцом композиции. В данном случае само контейнерное средство может представлять собой ингалятор, шприц, пипетку, глазную пипетку или другой такой подобный прибор, из которого препарат может быть нанесен на пораженную область животного, как, например, в легкие, инъцировано животному или даже нанесено или смешано с другими компонентами набора.
Компоненты данного набора также могут быть предоставлены в высушенной или лиофилизированной формах. При предоставлении реактивов или компонентов в высушенной форме растворение обычно осуществляется посредством добавления подходящего растворителя. Рассматривается то, что растворитель также может быть предоставен в другом контейнерном устройстве. Независимо от числа или типа контейнеров наборы по изобретению также могут содержать или быть упакованы с прибором для помощи в инъекции/введении или помещении конечной сложной композиции в пределы организма животного. Такой прибор может представлять собой ингалятор, шприц, пипетку, пинцет, мерную ложку, глазную пипетку или любое такое одобренное для медицинского применения средство доставки.
Обычные специалисты в данной области должны понимать то, что применения приведенного выше способа имеют широкое приложение для идентификации подходящих AON для применения в лечении многих других заболеваний.
AON по настоящему изобретению также могут использоваться в сочетании с альтернативными терапиями, такими как лекарственные терапии.
Согласно настоящему изобретению, следовательно, предложен способ лечения, предупреждения или снижения интенсивности эффектов заболевания или состояния, ассоциированного с экспрессией RAGE, где AON по настоящему изобретению вводится последовательно или сопутствующие с другой альтернативной терапией, ассоциированной с лечением, предупреждением или уменьшением интенсивности эффектов заболевания или состояния, ассоциированного с экспрессией RAGE. Предпочтительно данное заболевание или состояние выбрано из списка, содержащего: нейродегенеративные заболевания, рак, легочные расстройства или воспалительные заболевания.
Общее
Специалистам в данной области будет понятно то, что изобретение, описанное в данном документе, подвержено вариациям и модификациям, отличным от конкретно описанных вариаций и модификаций. Следует понимать то, что данное изобретение включает все такие вариации и модификации. Данное изобретение также включает все стадии, характеристики, композиции и соединения, на которые дается ссылка, или которые указаны в описании изобретения, индивидуально или коллективно, и любые и все комбинации или любые две или более чем две стадии или характеристики.
Настоящее изобретение не ограничивается объемом конкретных воплощений, описанных в данном документе, которые предназначены только для цели иллюстрации примером. Функционально эквивалентные продукты, композиции и способы явно находятся в пределах объема изобретения, как описано в данном документе.
Полные раскрытия всех публикаций (включая патенты, патентные заявки, журнальные статьи, лабораторные руководства, книги или другие документы), процитированных в данном документе, являются тем самым включенными посредством ссылки. Не делается допущения о том, что любые из данных ссылок составляют предшествующий уровень техники или представляют собой часть обычного общего знания работающих в области, к которой относится данное изобретение.
Каждый документ, ссылка, патентная заявка или патент, процитированные в данном тексте, прямо включаются в данный документ во всей их полноте посредством ссылки, что означает то, что читателю следует их читать и рассматривать как часть данного текста. То, что документ, ссылка, патентная заявка или патент, процитированные в данном тексте, не повторяются в данном тексте делается просто по причине краткости.
Любые инструкции, описания, спецификации продукта изготовителя и описания продукта для любых продуктов, упомянутых в данном документе или в любом документе, включенном в данный документ посредством ссылки, являются тем самым включенными в данный документ посредством ссылки и могут использоваться в воплощении данного изобретения на практике.
Термин «происходящий» и «полученный из» в том виде, в котором он используется в данном документе, следует принимать как указывающий на то, что конкретное целое может быть получено из конкретного источника, хотя и не обязательно непосредственно изданного источника.
Формы единственного числа в том виде, в котором они используются в данном документе, включают ссылки во множественном числе, если контекст явно не диктует иное.
Во всем данном описании изобретения, если контекст на требует иного, будет понятно, что слово «содержать» или вариации, такие как «содержит» или «содержащий», подразумевает включение заявленного целого или группы целых, но не исключение какого-либо другого целого или группы целых.
В отличие от рабочего примера или где указано иначе, все числа, выражающие количества ингредиентов, условий реакции и так далее, используемые в описании и формуле изобретения, следует понимать как во всех случаях модифицированные термином «примерно». Соответственно, если не указано противоположное, числовые параметры, изложенные в описании и формуле изобретения, представляют собой приближения, которые могут варьировать, в зависимости от желательных свойств, которые стараются получить посредством настоящего изобретения. Следовательно «примерно 80%» означает «примерно 80%» и также «80%». Самое меньшее каждый числовой параметр следует истолковывать в свете числа значимых чисел и обычных подходов округления.
Несмотря на то, что числовые интервалы и параметры, излагающие широкой объем данного изобретения, представляют собой приближения, числовые значения, изложенные в конкретных примерах, приводятся настолько точно, насколько возможно. Любое числовое значение; однако, по своему характеру содержит определенные ошибки с необходимостью возникающие из-за стандартного отклонения, обнаруженного в их соответствующих опытных измерениях.
Другие определения для выбранных терминов, использованных в данном документе, можно найти в пределах подробного описания данного изобретения и они применяются везде. Если не определено иначе, все другие научные и технические термины, использованные в данном документе, имеют такое же значение, которое обычно понятно обычному специалисту в области, к которой принадлежит данное изобретение.
Номера идентификации последовательности («SEQ ID NO:»), содержащие информацию по нуклеотидной и аминокислотной последовательности, включенной в данное описание изобретения, собраны в конце данного описания и были получены с использованием программы Patentln Version 3.0. Каждая нуклеотидная или аминокислотная последовательность идентифицируется в перечне последовательностей посредством числового индикатора <210> с последующим идентификатором последовательности (например, <210> 1, <210> 2 и т.д.). Длина, тип последовательности и организм-источник для каждой нуклеотидной или аминокислотной последовательности указываются посредством информации, приведенной в полях числового индикатора <211>, <212> и <213> соответственно.
Была предложена и опубликована система номенклатуры антисмыслового олигомера для различения разных антисмысловых олигомеров (см. Mann et al., (2002) J Gen Med 4, 644-654). Данная номенклатура становится особенно релевантной при анализе нескольких слегка отличных антисмысловых олигомеров, причем все направлены на ту же самую область-мишень, как показано ниже:
Н # A/D (х:у)
причем первая буква обозначает вид (например, Н: человеческое, М: мышиное)
«#» обозначает номер экзона-мишени
«А/D» указывает акцепторный или донорный сайт сплайсинга в начале/конце экзона соответственно
(х у) представляет координаты отжига, где «-» или «+» указывают интронные или экзонные последовательности соответственно. В качестве примера, А(-6+18) указывало бы последние 6 оснований интрона, предшествующего целевому экзону, и первые 18 оснований целевого экзона. Ближайший сайт сплайсинга был бы акцептором таким образом, что данным координатам предшествовало бы «А». Описание координат отжига в донорном сайте сплайсинга могло бы быть D(+2-18), где последние 2 экзонных основания и первые 18 интронных оснований соответствуют сайту отжига антисмыслового олигомера. Полностью экзонные координаты отжига, которые были бы представлены А(+65+85), то есть сайт между 65-ым и 85-ым нуклеотидом включительно с начала данного экзона.
Следующие примеры более полно описывают способ применения вышеописанного изобретения, а также излагают лучшие способы осуществления разных аспектов данного изобретения. Следует понимать то, что данные способы не ограничивают объем данного изобретения, но скорее представлены для иллюстративных целей.
ПРИМЕРЫ
В каждом из следующих примеров применяются следующие общие материалы и методы, если контекст не требует иного.
Человеческие микрососудистые эндотелиальные клетки (НМЕС1) культивировали в среде MCDB 131 (10% FCS (фетальная телячья сыворотка) с 10 мМ глутамином, EGF (эпидермальный фактор роста) и гидрокортизоном). Эптелиальные клетки легкого человека (А549) (карцинома) культивировали в среде F-12K (10% FBS с 2 мМ глутамином). Первичные эндотелиальные клетки аорты (РМАЕС) выделяли из аорты мышей C57bI6 (дикого типа) и нокаутированных по AGER мышей, и культивировали в среде Игла, модифицированной по Дульбекко (ОМЕМ)/среде F12, дополненной добавкой для роста эндотелиальных клеток (ECGS).
Для трансфекции AON эпителиальные или эндотелиальные клетки высевали в 48-луночные планшеты на 24 или 48 часов и затем трансфицировали с использованием реактива липофектамин 3000 (0,15 мкл/лунку липофектамина 3000; 0,4 мкл/лунку Р3000/лунку) с одним из AON, нацеленным на экзон 9 человеческого AGER (для НМЕС1 и А549) и мышиный AGER для мышиных клеток РМАЕС или соответствующих контролей (Таблицы 3a-3d). Клетки инкубировали с AON/катионными липоплексами при 37° в течение 24 ч, после чего клетки лизировали, экстрагировали РНК и получали кДНК с использованием либо методики TRIZOL, либо способа клетки в СТ.
Для измерения влияния на альтернативный сплайсинг пре-мРНК человеческого RAGE определяли экспрессию с использованием праймеров ПЦР-ОТ в реальном времени к одному из 11-го экзона, экзона 9b или охватывающих экзоны 8-10 RAGE, отмечая экспрессию (мРНК), сохраняющуюся во всех вариантах сплайсинга мРНК RAGE, сохранение экзона 9b или экспрессию изоформы RAGEv1, кодирующей трансмембранный-цитозольный хвост соответственно. Уменьшение сигнала ПЦР экзона 8-10 цитозольного хвоста относительно экзона 11 указывает на то, что меньше сплайсоформ мРНК RAGE содержат экзон 10, и клеткой продуцируется меньше изоформ полноразмерного белка RAGE (способных к сигнализации). В мышиных клетках и тканях праймеры, охватывающие экзон 10-11 мРНК мышиного RAGE, использовали для того, чтобы отмечать экспрессию сплайсоформ мРНК RAGE, содержащих экзон 10.
Также отбирали среды, и определяли уровни растворимого RAGE посредством ELISA. Среды (6 мл) концентрировали с использованием фильтра с порогом отсечения молекулярной массы перед проведением анализа посредством ELISA.
Клетки яичника китайского хомяка (СНО) экспрессируют рекомбинантные белки на высоком уровне, делая их идеальной системой для исследования регуляции сплайсинга пре-мРНК с измененной секрецией белка в качестве ожидаемого результата. В качестве модельной системы для лучшего исследования влияния AON на сплайсинг RAGE, приводящего к экспрессии и секреции внеклеточного растворимого RAGE, клетки СНО, выращенные в среде F12 (дополненной 10% FBS), трансфицировали плазмидой, кодирующей последовательность человеческого геномного AGER (gRAGE) (исключая природные 5' и 3' нетранслируемые области), наряду с AON, используя липофектамин 2000. Через 2 суток среды отбирали, концентрировали с использованием фильтра с порогом отсечения молекулярной массы и проверяли вестерн-блоттингом с использованием антитела против hRAGE (R&D systems).
Для определения точной последовательности вариантов RAGE, выявленных после введения AON, РНК выделяли с использованием способа с Trizol, а кДНК получали с использованием олиго dT праймеров перед ПЦР с использованием RAGE-специфичных праймеров к 5' и 3' UTR последовательностям RAGE и полугнездовой ПЦР для амплификации смеси последовательностей RAGE. Полосы RAGE, очищенные на геле, клонировали в векторы клонирования ТА перед трансформацией в клетки Тор10 Е. coli. Для характеристики отбирали больше, чем 50 колоний с использованием анализатора последовательностей MultiNA, используя праймеры, охватывающие экзоны 8-11 для определения размера вставки RAGE. Пять клонов каждого размера высылали для секвениования по Сэнджеру для подтверждения последовательности сплайсоформ RAGE, соответствующих размеру полос.
Для измерения функционального влияния вмешательства лигандзависимой и лиганднезависимой активации RAGE клетки подвергали воздействию когнатного лиганда AT1R, Ang II (1 мкМ) или лиганда RAGE - S100A8/A9 (0,6 мкг/мл клеток) в течение 4 часов. Клетки затем хранили замороженными до экстракции мРНК, и синтезировали кДНК с использованием способа от клеток до Ct. Изменения экспрессии гена субъединицы NFκB, р65 (ReIA) или NFκB-активированных генов-мишеней (например, ICAM-1) оценивали посредством количественной ПЦР-ОТ в реальном времени, проведенной с использованием системы TaqMan на основе выявления в реальном времени накопленной флуоресценции (ABI Prism 7700, Perkin-Elmer Inc, РЕ Biosystems, Foster City, CA, США). Экспрессию генов нормировали к 18S мРНК и приводили как кратность изменения по сравнению с уровнем экспрессии необработанных контрольных мышей/клеток, которой придается произвольное значение 1.
ПРИМЕР 1. МОДУЛЯЦИЯ АЛЬТЕРНАТИВНОГО СПЛАЙСИНГА RAGE В ЧЕЛОВЕЧЕСКИХ КЛЕТКАХ С ИСПОЛЬЗОВАНИЕМ АНТИСМЫСЛОВЫХ ОЛИГОНУКЛЕОТИДОВ, НАЦЕЛЕННЫХ НА ЦИС-ДЕЙСТВУЮЩИЕ ЭЛЕМЕНТЫ РНК В ЭКЗОНЕ 10
В данном примере продемонстрировано то, что сплайсинг пре-мРНК RAGE может модулироваться с использованием AON, нацеленных на цис-действующие элементы РНК в экзоне 10 пре-мРНК RAGE.
пре-мРНК RAGE в естественных условиях подвергается альтернативному сплайсингу, генерируя целый ряд сплайсоформ мРНК. Большинство данных сплайсоформ содержит и экзон 10, и 11, кодирующие трансмембранный и цитозольный домены RAGE, соответственно, и изоформы белка, которые продуцируются, следовательно, способны к активации лигандами RAGE и к трансактивации солокализующимся GPCR с индукцией провоспалительной и пропролиферативной сигнализации. Выбор альтернативного сайта 5' сплайсинга в экзоне 9 также может приводить к 83-нуклеотидному удлинению экзона 9 (экзон 9В; Фиг. 1а). Поскольку расстояние между данным сайтом альтернативного сплайсинга и сайтом 3' сплайсинга (акцепторным) в начале экзона 10 составляет только 46 нуклеотидов, что значительно короче, чем нижняя граница длины интронов у эукариот, экзон 10, следовательно, пропускается, и вместо этого выбирается 3' сайт (акцепторный) в начале экзона 11. Данный альтернативный сплайсинг также вводит кодон преждевременной терминации, но не подвергается нонсенс-опосредованному распаду из-за его тесной (29 п.н.) близости с последним контактом экзон-экзон. Зрелая мРНК, образующаяся в результате этого альтернативного сплайсинга (RAGE_v1, Фиг. 1а), следовательно, кодирует изоформу белка, у которой отсутствуют элементы, требующиеся для удерживания в мембране и цитоплазматической сигнализации, кодируемые экзоном 10 и 11, соответственно, таким образом, она секретируется и естественно обнаруживается в кровотоке, бронхо-альвеолярной и спинномозговой жидкостях. Данный esRAGE может действовать в качестве рецептора-ловушки, конкурируя с полноразмерным RAGE клеточной поверхности за лиганды или увеличивая клиренс лиганда. Новый 17-аминокислотный С-конец, кодируемый экзоном 9b, также может иметь независимое действие на сигнализацию или мультимеризацию RAGE.
Трансфекция человеческих эпителиальных клеток легкого (А549) 10 нМ AON, нацеленными на экзон 10, приводила к изменениям в экспрессии сплайсоформ мРНК RAGE, содержащих экзон 9b (Фиг. 1b). Некоторые AON, в частности AON 3779, увеличивали экспрессию сплайсоформ мРНК RAGE, содержащих экзон 9b, по сравнению с контрольными (обработанные скремблированной РНК) клетками при измерении посредством ПЦР-ОТ в реальном времени. Другие AON, также нацеленные на экзон 10, в частности, AON 3777 и 3781, не имели влияния.
Трансфекция человеческих эпителиальных клеток легкого (А549) 10 нМ концентрациями AON, нацеленных на экзон 10, также приводила к изменениям в экспрессии сплайсоформ мРНК RAGE таким образом, что некоторые AON, в частности AON 3779, 3780, 3781, 82, 83 и 87 снижали экспрессию сплайсоформ мРНК RAGE, содержащих экзон 10 (т.е. кодирующий элементы сигнализации цитозольного хвоста), по сравнению с контрольными (обработанные скремблированной РНК) клетками при измерении посредством ПЦР-ОТ в реальном времени (Фиг. 1с и 1d). Другие AON, также нацеленные на экзон 10, в частности, AON 3777, 84 и 85, не имели влияния.
Все сплайсоформы мРНК RAGE, экспрессируемые в человеческих эпителиальных клеток легкого (А549) в присутствии и в отсутствие отобранных AON, нацеленных на экзон 10, затем выделяли и клонировали в клетки Тор10 Е. coli. С использованием праймеров, охватывающих участок между экзоном 8 и экзоном 11, определяли размер каждой вставки RAGE. Обработка отобранными AON, нацеленными на экзон 10, в частности, AON 3779 и 3778, приводила к повышенной экспрессии сплайсоформ мРНК RAGE, содержащих фрагменты размером 250 т.п.н. (Фиг. 1е), отмечая сохранение экзона 9b.
Все AON, нацеленные на экзон 10, также снижали процентную долю экспрессии клонов мРНК, экспрессирующих полосу 300 т.п.н., обозначающую полную экспрессию элементов, способных к сигнализации, содержащихся в экзонах 8-10. В частности, AON 3779 приводил к 49%-ному снижению уровня сплайсоформ мРНК RAGE, содержащих фрагменты размера 300 т.п.н. (Фиг. 1f).
Все AON, нацеленные на экзон 10, также индуцировали экспрессию de novo сплайсоформ мРНК RAGE с полным отсутствием экзона 8 (подтвержденных секвенированием по Сэнджеру и отмеченных пустой полосой на анализаторе MultiNA, указанной стрелками на геле; Фиг. 1f) и фрагментом «без полосы» (Фиг. 1е).
Трансфекция человеческих эпителиальных клеток легкого (А549) AON, нацеленными на экзон 10, в частности, AON 3777 и 3778, также приводила к экспрессии de novo сплайсоформ мРНК RAGE, содержащих фрагменты размером 276 и 430 т.п.н. (Фиг. 1е), обозначая нетипичное удаление экзона 9 и сохранение экзона 9, 9b и экзона 10 соответственно.
Трансфекция человеческих эпителиальных клеток легкого (А549) 10 нМ AON, нацеленных на экзон 10, в частности, AON 3779, также приводила к изменениям числа копий сплайсоформ мРНК RAGE, содержащих экзон 10, в отличие от контрольных клеток (обработанных скремблированной РНК), при измерении на цифровой ПЦР (Фиг. 1g).
Трансфекция человеческих эпителиальных клеток легкого (А549) 10 нМ AON, нацеленных на экзон 10, в частности, AON 3779, также увеличивала высвобождение эндогенного растворимого RAGE в среду (Фиг. 1h), согласуясь с повышенной экспрессией сплайсоформы мРНК RAGE, содержащей экзон 9b.
Трансфекция клеток СНО плазмидой, кодирующей геномную последовательность человеческого RAGE (гДНК) приводила к альтернативному сплайсингу продукта гена, включая небольшое количество эндогенного RAGE, секретируемого в среды клеток (красная стрелка), при выявлении посредством вестерн-блоттинга, согласуясь с нормальной картиной сплайсинга RAGE (Фиг. 1i). Количество эндогенного растворимого RAGE (esRAGE), секретируемого в среду, увеличивалось, при сотрансфекции данных клеток СНО, экспрессирующих человеческий геномный RAGE, с использованием AON 3779 или AON 87 (Фиг. 1i). В то же самое время полноразмерный RAGE, экспрессированный и сохраняющийся в клетках (белая стрелка), снижался при сотрансфекции клеток СНО, экспрессирующих человеческий геномный RAGE, с использованием AON 3779 или AON 87. Клетки также содержали внутри клеточно повышенное количество С-усеченного RAGE.
Количественное значение растворимого RAGE при измерении на ELISA также увеличивалось в клетках СНО, экспрессирующих человеческий геномный RAGE, после трансфекции отобранными AON, нацеленными на экзон 10, включающими AON 3779, AON 82, 83, 84 и 85 (Фиг. 1j). Из них AON 3779 имел наибольший эффект.
Количественное значение растворимого RAGE при измерении на ELISA также увеличивалось некоторыми другими AON, нацеленными на экзон 10 в области, аналогичной AON 3779, включая AON 90 и 93 (Фиг. 1k), на аналогичное AON 3779 (10 нМ) значение. Количественное значение растворимого RAGE, высвобожденного из клеток СНО, экспрессирующих человеческий геномный RAGE, дозозависимо возрастало после трансфекции AON 3779 (Фиг. 11).
Трансфекция человеческих эпителиальных клеток легкого (А549) 10 нМ AON, нацеленными на экзон 10, в частности, AON 3779, также модулировала индукцию провоспалительной сигнализации после активации RAGE лигандом RAGE S100A8/A9 (0,6 мкг/мл), приводя к индукции экспрессии ICAM-1 (Фиг. 1m).
Трансфекция человеческих эндотелиальных клеток аорты (НАЕС) 10 нМ AON, нацеленными на экзон 10, в частности, AON 3779, 3780, 81, 82 и 83, также предотвращала индукцию экспрессии ICAM-1 после воздействия лиганда RAGE S100A8/A9 (0,6 мкг/мл; Фиг. 1m).
Трансфекция человеческих эпителиальных клеток легкого (А549) 10 нМ AON, нацеленными на экзон 10, в частности, AON 3779, также модулировала индукцию провоспалительной сигнализации после трансактивации RAGE после активации рецептора АТ1 ангиотензином II (1 мкМ), приводя к индукции экспрессии ICAM-1 (Фиг. 1о).
Трансфекция человеческих эндотелиальных клеток аорты (НАЕС) 10 нМ AON, нацеленными на экзон 10, в частности, AON 3777, 3779, 3780, 3780, 3782 и 3783, также модулировала индукцию провоспалительной сигнализации, включая индукцию экспрессии ICAM-1 после независимой от лиганда трансактивации RAGE после активации рецептора АТ1 ангиотензином II (1 мкМ; Фиг. 1р). Другие AON, нацеленные на экзон 10, в частности, AON 3778 и 84, не имели влияния.
Трансфекция человеческих эндотелиальных клеток микрососудов (НМЕС1) 10 нМ AON, нацеленными на экзон 10, в частности, AON 3779, 3780, 82, 83 и 87, приводила к изменениям экспрессии сплайсоформ мРНК RAGE, включая уменьшение экспрессии сплайсоформ мРНК RAGE, содержащих экзон 10 (Фиг. 1q). Трансфекция человеческих эндотелиальных клеток микрососудов (НМЕС1) 10 нМ AON, нацеленными на экзон 10, в частности, AON 3779 и 87, также индуцировала увеличение экспрессии сплайсоформ мРНК RAGE, содержащих экзон 9b (Фиг. 1r).
Морфолиноолигонуклеотиды с ковалентно связанным октагуанидиновым дендримером (известные как виво-морфолино) могут использоваться in vitro и in vivo без необходимости в трансфекционных реактивах. Обработка человеческих эндотелиальных клеток аорты (НАЕС) виво-морфолино препаратом AON 3779 (1 мкМ) приводила к изменениям экспрессии сплайсоформ мРНК RAGE, включая увеличение уровня сплайсоформ мРНК RAGE, содержащих экзон 9b, при измерении ПЦР-ОТ (Фиг. 1s). Кроме того, экспрессия esRAGE в среде клеток также возрастала по сравнению с клетками, обработанными контрольным морфолино AON (Фиг. 1t). Обработка AON 79 без трансфекционных реактивов не имела влияния. Трансфекция AON 3779 с трансфекционными реактивами показана в качестве позитивного контроля.
Обработка клеток СНО, экспрессирующих геномный человеческий RAGE, vivo-морфолино препаратом AON 3779 (1 мкМ) также увеличивала экспрессию растворимого RAGE при измерении на ELISA (Фиг. 1u).
Известно то, что отрезки поли(G), особенно мотивы GGGG, могут действовать в качестве сайленсеров экспрессии, и они часто связываются hnRNP Н и F (Sohail et al., ВМС Genomics. 2014). Предполагается то, что сплайсинг RAGE частично регулируется G-богатыми цис-элементами и гетерогенным ядерным рибонуклеопротеином Н в пределах интрона 9/экзона 9b, которые требуются для предпочтительного использования расположенного выше 5' сайта сплайсинга RAGE. Отрезки поли(G) благоприятствуют пропуску экзона, так как связывание hnRNP Н требует непрерывного отрезка G. AON обычно имеют плохую аффинность и пониженную эффекивность в G-богатых областях. Экзон 10 содержит три мотива GGGG, один из которых распознется как мишень для связывания hnRNPF и фланкируется AON 3779, 3787 и AON 3782 (Фиг. 1v). Однако мутирование данной области (мутант RAGE М3) не имело влияния на сплайсинг экзона или ответ на AON 3779 (Фиг. 1w), свидетельствуя о том, что авторы данного изобретения модулируют новый сайленсер сплайсинга RAGE посредством нацеливания на данные области AON с модуляцией сплайсинга RAGE, что не могло быть спрогнозировано из предшествующего уровня техники.
ПРИМЕР 2. МОДУЛИРОВАНИЕ АЛЬТЕРНАТИВНОГО СПЛАЙСИНГА RAGE В ЧЕЛОВЕЧЕСКИХ КЛЕТКАХ С ИСПОЛЬЗОВАНИЕМ АНТИСМЫСЛОВЫХ ОЛИГОНУКЛЕОТИДОВ, НАЦЕЛЕННЫХ НА ЦИС-ДЕЙСТВУЮЩИЕ ЭЛЕМЕНТЫ РНК В ЭКЗОНЕ 9
Данный пример демонстрирует то, что альтернативный сплайсинг пре-мРНК RAGE может модулироваться с использованием AON, нацеленных на цис-действующие элементы РНК в экзоне 9 пре-мРНК RAGE (Фиг. 2а).
Трансфекция человеческих эпителиальных клеток легкого (А549) 10 нМ концентрациями некоторых AON, нацеленных на экзон 9, в частности, AON 3668, 3669, 3670, 4103, 4104, 4105, уменьшала экспрессию сплайсоформ мРНК RAGE, содержащих экзон 10, по сравнению с контрольными клетками (обработанными скремблированной РНК) при измерении посредством ПЦР-ОТ в реальном времени (Фиг. 2b). Другие AON, нацеленные на экзон 9, в частности AON 4106, не имели влияния. Однако общая экспрессия мРНК RAGE слегка повышалась после обработки всеми AON, нацеленными на экзон 9.
Все сплайсоформы мРНК RAGE, экспрессируемые в человеческих эпителиальных клетках легкого (А549) в присутствии и в отсутствие отобранных AON, нацеленных на экзон 9, выделяли и клонировали в клетки Тор10 Е. coli. С использованием праймеров, охватывающих экзоны 8-11, определяли размер каждой вставки RAGE (Фиг. 2с). Все AON, нацеленные на экзон 9, снижали процентную долю экспрессии клонов мРНК, экспрессирующих полосу 300 т.п.н. (Фиг. 2d), обозначая полную экспрессию элементов, спосбных к сигнализации, содержащихся в экзоне 10 и 11. В частности, AON 3670 приводил к снижению уровня сплайсоформ мРНК RAGE, содержащих фрагменты размером 300 т.п.н. (Фиг. 2d).
Трансфекция человеческих эпителиальных клеток легкого (А549) AON, нацеленными на экзон 9, в частности, AON 3669, 3670, 4103, также индуцировала экспрессию de novo сплайсоформ мРНК RAGE, в которых экзон 8 был пропущен (обозначено пустыми полосами на анализаторе multiNA; Фиг. 2с).
Трансфекция человеческих эпителиальных клеток легкого (А549) AON, нацеленными на экзон 9, в частности, AON 3669, 3670, 4103 и 4105, также приводила к экспрессии de novo сплайсоформ мРНК RAGE, содержащих фрагменты 276 т.п.н., обозначая удаление экзоне 9 из сплайсоформ (Фиг. 2d).
В клетках СНО, трансфицированных плазмидой, содержащей геномную последовательность человеческого RAGE (gRAGE), количество растворимого RAGE, секретируемого в среду при измерении на ELISA, умеренно повышалось некоторыми AON, нацеленными на сайт сплайсинга RAGE в экзоне 9, в частности, AON 3669 и 3671 (Фиг. 2е), по сравнению с контрольными клетками (обработанными скремблированной РНК).
Трансфекция человеческих эпителиальных клеток легкого (А549) разными концентрациями AON, нацеленных на экзон 9, в частности, AON 3669, 3670 и 4103, также ингибировала индукцию провоспалительной сигнализации после активации RAGE лигандом RAGE S100A8/A9 (0,6 мкг/мл), включая индукцию экспрессии гена TLR-4 (Фиг. 2f). Другие AON, нацеленные на экзон 9, в частности, AON 3668 и 4104, не имели влияния.
Трансфекция человеческих эпителиальных клеток легкого (А549) 10 нМ AON, нацеленными на экзон 9, в частности, AON 3669, 3670 и 4104, также модулировала индукцию провоспалительной сигнализации посредством независимой от лиганда трансактивации RAGE после активации рецептора АТ1 ангиотензином II (1 мкМ; Фиг. 2g). Другие AON, нацеленные на экзон 9, в частности, AON 3668, 3671 и 4103, не имели влияния.
Трансфекция человеческих эндотелиальных клеток микрососудов (НМЕС) 10 нМ AON, нацеленными на экзон 9, в частности, AON 3669 и 3670, также снижала экспрессию эндогенных сплайсоформ мРНК RAGE, содержащих экзон 10 (Фиг. 2h).
ПРИМЕР 3. МОДУЛЯЦИЯ АЛЬТЕРНАТИВНОГО СПЛАЙСИНГА RAGE В ЧЕЛОВЕЧЕСКИХ КЛЕТКАХ С ИСПОЛЬЗОВАНИЕМ АНТИСМЫСЛОВЫХ ОЛИГОНУКЛЕОТИДОВ. НАЦЕЛЕННЫХ НА ЦИС-ДЕЙСТВУЮЩИЕ ЭЛЕМЕНТЫ РНК В ЭКЗОНЕ 9В
Данный пример демонстрирует то, что альтернативный сплайсинг RAGE может модулироваться с использованием AON, нацеленных на элементы РНК в пределах экзона 9b в пре-мРНК RAGE.
Экзон 9b удаляется у большинства изоформ RAGE альтернативным сплайсингом (Фиг. 1е). Для данного удаления потенциально требуется взаимодействие сплайсосомы с мотивами в экзоне 9b. Авторы данного изобретения предположили то, что модулирование данных элементов связывания с использованием AON также могло бы модулировать альтернативный сплайсинг пре-мРНК RAGE.
Трансфекция человеческих эпителиальных клеток легкого (А549) 10 нМ концентрациями AON, нацеленных на экзон 9b, приводила к изменениям в экспрессии мРНК сплайсоформ мРНК RAGE. Общая экспрессия RAGE слегка повышалась после обработки обоими AON, нацеленными на экзон 9b (Фиг. 3а). Кроме того, экспрессия сплайсоформ мРНК RAGE, содержащих экзон 10, слегка снижалась после трансфекции AON 88 по сравнению с контрольными клетками, обработанными скремблированной РНК, при измерении ПЦР-ОТ в реальном времени (Фиг. 3а). Экспрессия сплайсоформ мРНК RAGE, содержащих экзон 9b, также умеренно повышалась AON 88 и 89 (Фиг. 3b). Трансфекция AON 3779, нацеленным на экзон 10, показана в качестве позитивного контроля.
Трансфекция человеческих эндотелиальных клеток микрососудов (НМЕС) 10 нМ концентрациями AON, нацеленных на экзон 9b, также приводила к пониженной экспрессии сплайсоформ мРНК RAGE, содержащих экзон 10 (Фиг. 3с), и повышенной экспрессии сплайсоформ мРНК RAGE, содержащих экзон 9b (Фиг. 3d). Опять-таки, общая экспрессия мРНК RAGE слегка повышалась после обработки обоими AON, нацеленными на экзон 9b (Фиг. 3с). Трансфекция AON 3779, нацеленным на экзон 10, показана в качестве позитивного контроля.
В клетках СНО, трансфицированных плазмидой, содержащей геномную последовательность человеческого RAGE (gRAGE), количество растворимого RAGE, секретируемого в среду, при измерении на ELISA, умеренно увеличивалось некоторыми AON, нацеленными на экзон 9b, в частности, AON 89 (Фиг. 3е), по сравнению с контрольной РНК. Трансфекция AON 3779, нацеленным на экзон 10, показана в качестве позитивного контроля.
Последовательность интрона 9 человеческого RAGE и комплементарное нацеливание для AON 88 и 89 показано на Фиг. 3f.
ПРИМЕР 4. МОДУЛЯЦИЯ АЛЬТЕРНАТИВНОГО СПЛАЙСИНГА МЫШИНОГО RAGE С ИСПОЛЬЗОВАНИЕМ АНТИСМЫСЛОВЫХ ОЛИГОНУКЛЕОТИДОВ. НАЦЕЛЕННЫХ НА ЦИС-ДЕЙСТВУЮЩИЕ ЭЛЕМЕНТЫ РНК В ПРЕ-мРНК RAGE
Данный пример демонстрирует то, что альтернативный сплайсинг пре-мРНК RAGE также может модулироваться с использованием определенных AON, нацеленных на цис-действующие элементы РНК в экзоне 9, 9В и 10 в пре-мРНК мышиного RAGE, и что данная модуляция может быть продемонстрирована in vitro, ex vivo и in vivo.
Для анализа влияния AON на альтернативный сплайсинг мышиного RAGE клетки СНО трансфицировали плазмидой, содержащей геномную последовательность мышиного RAGE. Это приводило к секреции небольшого количества esRAGE. Трансфекция AON, нацеленными на мышиный RAGE, в частности, AON m3779 (AON, сконструированный для нацеливания на экзон 10 последовательности мышиного RAGE в сравнимой последовательности с той, на которую нацелен AON 3779 в последовательности человеческого RAGE, подробно описанной в Примере 1) и AON m101, увеличивала секрецию esRAGE в среду клеток (Фиг. 4а).
Первичные мышиные клетки эндотелия аорты (РМАЕС) трансфицировали AON (50 мМ), нацеленными на экзон 9 RAGE (Таблица 3d). Данная обработка приводила к умеренным изменениям в экспрессии сплайсоформ мРНК RAGE по сравнению с соответствующим им негативным контролем. В частности, AON 3674 и 3675 могли уменьшать экспрессию сплайсоформ мРНК RAGE, содержащих экзон 10, при измерении ПЦР-ОТ в реальном времени (Фиг. 4b).
Однако трансфекция РМАЕС данными AON, использованными в концентрации 10 нМ, не демонстрировала значимого влияния на экспрессию сплайсоформ мРНК RAGE (Фиг. 4с). Уровни растворимого RAGE, секретированного в среду, умеренно возрастали через 24 часа после трансфекции культивируемых РМАЕС AON 3674 и 3675 в концентрации 50 нМ (Фиг. 4d).
Трансфекция клеток СНО, экспрессирующих мышиный геномный RAGE, m3779 (10 нМ) уменьшала экспрессию сплайсоформ мРНК RAGE, содержащих экзон 10, при оценке ПЦР-ОТ в реальном времени; Фиг. 4е) и повышала секрецию esRAGE по сравнению с контрольными клетками (обработанными скремблированной РНК) (Фиг. 4f). AON 3779, нацеленный на последовательности человеческого RAGE, также повышал секрецию esRAGE, но не так сильно, как AON m3779.
При трансфекции AON, сконструированных для нацеливания на экзон 10 мышиной последовательности RAGE, в клетки СНО, экспрессирующие геномную последовательность человеческого RAGE из плазмиды, AON m3779 (сконструированный для нацеливания на экзон 10 мышиного RAGE) также индуцировал увеличение уровня растворимого человеческого RAGE в среде клеток (Фиг. 4g). Однако AON 3779 (специфично нацеленный на экзон 10 человеческого RAGE) был более эффективным в модулировании человеческого геномного RAGE, чем AON m3779. Эти данные демонстрируют то, что AON, нацеленные на специфическую последовательность в экзоне 10, могут быть эффективными в модулировании сплайсинга RAGE, несмотря на области с отсутствием идентичности.
Для анализа эффекта AON, нацеленного на RAGE в мышиной ткани, получали точно нарезанные кусочки легкого (PCLS) от мышей и затем культивировали ex vivo. Вкратце, мышей гуманно умерщвляли, экспланты их легких инфундировали агарозой, отбирали цилиндрические стержни и затем нарезали ножом для получения срезов ткани, получая срезы легкого однородного диаметра и толщины. PCLS затем погружали в культуральную среду в многолуночных планшетах при условиях культуры тканей. Через 24 часа PCLS трансфицировали виво-морфолино AON 3779 или ненацеленным контролем в течение 48 часов. Обработка AON 3779 приводила к значимому увеличению экспрессии мРНК RAGE, содержащей экзон 9b, и уменьшала экспрессию сплайсоформ мРНК RAGE, содержащих экзон 10 (Фиг. 4h), с наибольшим эффектом на экзон 9b, наблюдаемым через 72 часа после дозирования (Фиг. 4i).
Для подтверждения релевантности in vivo AON, нацеленных на сплайсинг RAGE, самцам мышей C57bl6 подкожно инъецировали AON m3779 (1 мг/кг/сутки) в течение 7 суток. Экспрессия сплайсоформ мРНК RAGE, содержащих экзон 9b, увеличивалась в легких мышей, а уровень сплайсоформ, содержащих экзон 10, уменьшался после подкожной обработки AON m3779 в течение одной недели (Фиг. 4j).
Для подтверждения их потенциала в качестве ингалируемого терапевтического средства, живых мышей анестезировали перед доставкой 30 мкл 11 мкМ виво-морфолино-AON m3779 или ненацеленного виво-морфолино контроля (в качестве негативного контроля) в воде, не содержащей РНКаз, в легкое через трахею с использованием канюли или одной воды. Анестетик обращали, и мышей умерщвляли через 48 ч. Внутритрахеальная обработка m3779 приводила к значимому снижению легочной экспрессии сплайсоформ мРНК RAGE, содержащих экзон 10, по сравнению с одной водой, которая не имела значимого эффекта. Экспрессия изоформ мРНК RAGE, содержащих экзон 9b, также возрастала (Фиг. 4k). Аналогичное уменьшение экспрессии сплайсоформ мРНК RAGE, содержащих экзон 10, наблюдали после внутритрахеальной доставки 30 мкл 11 мкМ 2-O'-Ме-фосфоротиоатного AON m3779 по сравнению с одним носителем (стерильная вода) (Фиг. 41). Кроме того, также наблюдали увеличение уровней растворимого RAGE в системе кровообращения после обработки мышей AON m3779 по сравнению со стерильной водой или неэффективным AON 4105, нацеленным на человеческий RAGE (Фиг. 4m).
ПРИМЕР 5. МОДУЛЯЦИЯ АЛЬТЕРНАТИВНОГО СПЛАЙСИНГА RAGE С ИСПОЛЬЗОВАНИЕМ КОМБИНАЦИИ АНТИСМЫСЛОВЫХ ОЛИГОНУКЛЕОТИДОВ, НАЦЕЛЕННЫХ НА ПРЕ-мРНК RAGE
Данный пример демонстрирует то, что альтернативный сплайсинг RAGE может модулироваться с использованием комбинаций AON, нацеленных на разные цис-действующие элементы РНК в пре-мРНК RAGE.
Полагают то, что определение экзона регулируется сборкой многокомпонентного «кросс-экзонного» комплекса распознавания, который облегчает сохранене или исключение мишени. После связывания AON 3779 с экзоном 10 в пре-мРНК RAGE для увеличения образования esRAGE авторы данного изобретения предполагают то, что другие регуляторные мишени в экзоне 10 могут стать более критически важными, включая прогнозируемые сайты сплайсинга, но не ограничиваясь ими. Кроме того, в присутствии AON 3779 посредством селективного нацеливания на данные цис-действующие элементы РНК в экзоне 10 в пре-мРНК RAGE с использованием других AON авторы данного изобретения предупредили бы какое-либо уклонение от исключения экзона и дополнительно модулировали бы сплайсинг RAGE в направлении предпочтительного образования esRAGE.
Прогнозируется то, что вторичная струкура экзона 10 RAGE содержит две петли шпилек, связанных центральной шарнирной областью. Предполагается то, что нацеливание на последовательности экзонного энхансера в 3' шпильке с использованием AON 3779 увеличивало бы важность мишеней в альтернативной 5' шпильке, включая расположенный выше 5' сайт сплайсинга в экзоне 10.
Для подтверждения данной гипотезы, клетки СНО, содержащие плазмиду, кодирующую человеческий геномный RAGE, трансфицировали AON 3779 (10 нМ) в комбинации с другими AON, нацеленными на прогнозируемый 5' (акцепторный) сайт сплайсинга в экзоне 10 (также 10 нМ). Данная комбинированная обработка приводила к изменениям экспрессии сплайсоформ мРНК RAGE таким образом, что некоторые комбинации, в частности, AON 3779 плюс 3777, 3779 плюс 3778, значимо увеличивали экспрессию изоформ мРНК RAGE, содержащих экзон 9b, по сравнению с одним AON 3779 или контрольными клетками, обработанными скремблированной РНК, при измерении ПЦР-ОТ в реальном времени (Фиг. 5а). Данная комбинация также увеличивала продукцию белка esRAGE, измеренную в среде посредством ELISA (Фиг. 5b). Комбинация AON 3997 и AON 3778 также была более эффективной при низкой дозе (5 нМ плюс 5 нМ). Примечательно то, AON 3777 и AON 3778 имели слабый эффект или не имели эффекта сами по себе в концентрации 10 нМ.
В другом эксперименте клетки СНО, содержащие плазмиду, кодирующую человеческий геномный RAGE, трансфицировали AON 3779 в комбинации с другими AON, нацеленными либо на 3', либо на 5' сайты сплайсинга в экзоне 10. Некоторые комбинации также приводили к изменениям экспрессии сплайсоформ мРНК RAGE таким образом, что некоторые комбинации, в частности AON 3779 плюс 3778, 3779 плюс 3781 и 3779 плюс 3778 плюс 3781 значимо увеличивали экспрессию растворимого RAGE по сравнению с одним AON 3779 или контрольными клетками, обработанными скремблированной РНК, при измерении ELISA (Фиг. 5с). Данное увеличение было аналогичным при использовании AON 93 или AON 87 вместо 3779, хотя в целом комбинации с AON 3779 (обозначенные в прямоугольниках) были самыми сильными и оказывающими самый согласованный эффект.
Трансфекция человеческих эпителиальных клеток легкого (А549) другими комбинациями AON, также нацеленными на смежные области в экзоне 10, в частности AON 3779 плюс 3782, 3779 плюс 3784, 3779 плюс 3785, не имели значимого влияния на экспрессию изоформ RAGE, содержащих экзон 9b, помимо одного AON 3779 (Фиг. 5d). Примечательно то, что данные AON также нацелены на 5' шпильку (аналогично AON 3779) и/или центральную шарнирную область, в отличие от AON 3777 или AON 3778, которые были эффективными в комбинации, которые нацелены на экзонные энхансеры на другой (3') шпильке.
Клетки СНО, содержащие плазмиду, кодирующую геномную последовательность мышиного RAGE, трансфицировали AON m3779 в комбинации с AON, нацеленным либо на 3', либо на 5' сайты сплайсинга в экзоне 10. Данная комбинированная обработка приводила к изменениям в экспрессии сплайсоформ мРНК RAGE таким образом, что некоторые комбинации, в частности, AON m3779 плюс m102, значимо увеличивали секрецию растворимого RAGE в среду клеток при измерении ELISA по сравнению с AON m3779 индивидуально (Фиг. 5е).
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Monash University
Murdoch University
<120> МОДУЛЯТОРЫ И МОДУЛЯЦИЯ РНК РЕЦЕПТОРА КОНЕЧНЫХ ПРОДУКТОВ ГЛУБОКОГО
ГЛИКИРОВАНИЯ
<130> N423236EP
<140> 20806464.2
<141> 2020-05-07
<150> AU2019901641
<151> 2019-05-14
<150> AU2019902095
<151> 2019-06-17
<150> AU2019902772
<151> 2019-08-02
<150> AU2019903900
<151> 2019-10-16
<160> 77
<170> PatentIn version 3.5
<210> 1
<211> 25
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 3668
<400> 1
ccuccucgcc ugguucugga agaca 25
<210> 2
<211> 20
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 3669
<400> 2
uuggccccuc cucgccuggu 20
<210> 3
<211> 20
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 3670
<400> 3
cugcaguugg ccccuccucg 20
<210> 4
<211> 20
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 3671
<400> 4
ucaaaccccu caccugcagu 20
<210> 5
<211> 25
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 4103
<400> 5
ccugcaguug gccccuccuc gccug 25
<210> 6
<211> 25
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 4104
<400> 6
ccccucaccu gcaguuggcc ccucc 25
<210> 7
<211> 17
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 4105
<400> 7
gcaguuggcc ccuccuc 17
<210> 8
<211> 25
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 4106
<400> 8
caccugcagu uggccccucc ucgcc 25
<210> 9
<211> 25
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 3777
<400> 9
auccucccac agagccugua cggag 25
<210> 10
<211> 25
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 3778
<400> 10
cccugauccu cccacagagc cugua 25
<210> 11
<211> 25
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 3779
<400> 11
ggcguugccg ccuuugccac aagau 25
<210> 12
<211> 25
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 3780
<400> 12
cucaccucuc cucuccucgg cguug 25
<210> 13
<211> 25
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 3781
<400> 13
cuccacucac cucuccucuc cucgg 25
<210> 14
<211> 20
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 82
<400> 14
agcagggcgg cuguccccag 20
<210> 15
<211> 25
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 83
<400> 15
gccacaagau gaccccaaug agcag 25
<210> 16
<211> 25
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 84
<400> 16
ccuuugccac aagaugaccc caaug 25
<210> 17
<211> 25
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 85
<400> 17
ugccgccuuu gccacaagau gaccc 25
<210> 18
<211> 25
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 87
<400> 18
uccucggcgu ugccgccuuu gccac 25
<210> 19
<211> 13
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 90
<400> 19
uuugccacaa gau 13
<210> 20
<211> 20
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 93
<400> 20
ccgccuuugc cacaagauga 20
<210> 21
<211> 25
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 88
<400> 21
uuucuuguug accauccccc caguc 25
<210> 22
<211> 25
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 89
<400> 22
ccaucccccc agucacaugu guugg 25
<210> 23
<211> 25
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 3672
<400> 23
ccuccucgcc ugguucugga agaca 25
<210> 24
<211> 20
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 3673
<400> 24
cuggccccuc aucgccgguu 20
<210> 25
<211> 20
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 3674
<400> 25
cuucagcugg ccccucaucg 20
<210> 26
<211> 20
<212> RNA
<213> Artificial Sequence
<220>
<223> AON 3675
<400> 26
uccagucccu caccuucagc 20
<210> 27
<211> 25
<212> RNA
<213> Artificial Sequence
<220>
<223> AON m3779
<400> 27
uggguugucg uuuucgccac aggau 25
<210> 28
<211> 23
<212> RNA
<213> Artificial Sequence
<220>
<223> AON m97
<400> 28
acuacuccca ggccucccag gau 23
<210> 29
<211> 22
<212> RNA
<213> Artificial Sequence
<220>
<223> AON m98
<400> 29
gcagggcuac uacucccagg ca 22
<210> 30
<211> 25
<212> RNA
<213> Artificial Sequence
<220>
<223> AON m101
<400> 30
ucucacucac cucuccucac gccug 25
<210> 31
<211> 25
<212> RNA
<213> Artificial Sequence
<220>
<223> AON m102
<400> 31
ucucacucac cucuccucac gccug 25
<210> 32
<211> 320
<212> DNA
<213> Homo sapiens
<400> 32
gataaagtca gggaagcaga agatagcccc caacacatgt gactgggggg atggtcaaca 60
agaaaggaat ggtgagtggt ggtggctgtg ctctcaattt tccctgtctc cgtacaggct 120
ctgtgggagg atcagggctg ggaactctag ccctggccct ggggatcctg ggaggcctgg 180
ggacagccgc cctgctcatt ggggtcatct tgtggcaaag gcggcaacgc cgaggagagg 240
agaggtgagt ggagaaagcc agacccctca gacctagggc ttccaggcag caagcgaaga 300
ggggtcgggg ggtggaacga 320
<210> 33
<211> 320
<212> DNA
<213> Homo sapiens
<400> 33
tcgttccacc ccccgacccc tcttcgcttg ctgcctggaa gccctaggtc tgaggggtct 60
ggctttctcc actcacctct cctctcctcg gcgttgccgc ctttgccaca agatgacccc 120
aatgagcagg gcggctgtcc ccaggcctcc caggatcccc agggccaggg ctagagttcc 180
cagccctgat cctcccacag agcctgtacg gagacaggga aaattgagag cacagccacc 240
accactcacc attcctttct tgttgaccat ccccccagtc acatgtgttg ggggctatct 300
tctgcttccc tgactttatc 320
<210> 34
<211> 53
<212> PRT
<213> Homo sapiens
<400> 34
Ile Lys Ser Gly Lys Gln Lys Ile Ala Pro Asn Thr Cys Asp Trp Gly
1 5 10 15
Asp Gly Gln Gln Glu Arg Asn Gly Glu Trp Trp Trp Leu Cys Ser Gln
20 25 30
Phe Ser Leu Ser Pro Tyr Arg Leu Cys Gly Arg Ile Arg Ala Gly Asn
35 40 45
Ser Ser Pro Gly Pro
50
<210> 35
<211> 6
<212> PRT
<213> Homo sapiens
<400> 35
Ser Gln Gly Ser Arg Arg
1 5
<210> 36
<211> 45
<212> PRT
<213> Homo sapiens
<400> 36
Pro Pro Thr His Val Thr Gly Gly Met Val Asn Lys Lys Gly Met Val
1 5 10 15
Ser Gly Gly Gly Cys Ala Leu Asn Phe Pro Cys Leu Arg Thr Gly Ser
20 25 30
Val Gly Gly Ser Gly Leu Gly Thr Leu Ala Leu Ala Leu
35 40 45
<210> 37
<211> 13
<212> PRT
<213> Homo sapiens
<400> 37
Asp Lys Val Arg Glu Ala Glu Asp Ser Pro Gln His Met
1 5 10
<210> 38
<211> 10
<212> PRT
<213> Homo sapiens
<400> 38
Leu Gly Gly Trp Ser Thr Arg Lys Glu Trp
1 5 10
<210> 39
<211> 24
<212> PRT
<213> Homo sapiens
<400> 39
Val Val Val Ala Val Leu Ser Ile Phe Pro Val Ser Val Gln Ala Leu
1 5 10 15
Trp Glu Asp Gln Gly Trp Glu Leu
20
<210> 40
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Синтетическая ДНК
<400> 40
gctatcttct gcttccctga c 21
<210> 41
<211> 160
<212> DNA
<213> Homo sapiens
<400> 41
caaaagtgaa actccatctc aaaaaaaaaa agaaagggaa agactccact ggggctccca 60
ctaaataacc ctctctcaac ccgaagtctt cctttctgac tggatccaac tttgtcttcc 120
agaaccaggc gaggaggggc caactgcagg tgaggggttt 160
<210> 42
<211> 160
<212> DNA
<213> Homo sapiens
<400> 42
aaacccctca cctgcagttg gcccctcctc gcctggttct ggaagacaaa gttggatcca 60
gtcagaaagg aagacttcgg gttgagagag ggttatttag tgggagcccc agtggagtct 120
ttccctttct tttttttttt gagatggagt ttcacttttg 160
<210> 43
<211> 143
<212> DNA
<213> Homo sapiens
<400> 43
agaagatagc ccccaacaca tgtgactggg gggatggtca acaagaaagg aatggtgagt 60
ggtggtggct gtgctctcaa ttttccctgt ctccgtacag gctctgtggg aggatcaggg 120
ctgggaactc tagccctggc cct 143
<210> 44
<211> 143
<212> DNA
<213> Homo sapiens
<400> 44
agggccaggg ctagagttcc cagccctgat cctcccacag agcctgtacg gagacaggga 60
aaattgagag cacagccacc accactcacc attcctttct tgttgaccat ccccccagtc 120
acatgtgttg ggggctatct tct 143
<210> 45
<211> 50
<212> DNA
<213> Homo sapiens
<400> 45
aggactcttg tcccaaaggc atgaattcct agcattccct gtgacaagac 50
<210> 46
<211> 154
<212> DNA
<213> Homo sapiens
<400> 46
gactgaaaga tgggggctgg agagagggtg caggccccac ctagggcgga ggccacagca 60
gggagagggg cagacagagc caggaccctg gaaggaagca ggatggcagc cggaacagca 120
gttggagcct gggtgctggt cctcagtctg tggg 154
<210> 47
<211> 25
<212> DNA
<213> Homo sapiens
<400> 47
gtgagccact ccctcaaccc cactg 25
<210> 48
<211> 25
<212> DNA
<213> Homo sapiens
<400> 48
cctctaccat ggtgctatct cccag 25
<210> 49
<211> 107
<212> DNA
<213> Homo sapiens
<400> 49
gggcagtagt aggtgctcaa aacatcacag cccggattgg cgagccactg gtgctgaagt 60
gtaagggggc ccccaagaaa ccaccccagc ggctggaatg gaaactg 107
<210> 50
<211> 25
<212> DNA
<213> Homo sapiens
<400> 50
gtaagcgggg ctcctgttgc agcct 25
<210> 51
<211> 25
<212> DNA
<213> Homo sapiens
<400> 51
ttaggccctg cttctctgct tctag 25
<210> 52
<211> 196
<212> DNA
<213> Homo sapiens
<400> 52
aacacaggcc ggacagaagc ttggaaggtc ctgtctcccc agggaggagg cccctgggac 60
agtgtggctc gtgtccttcc caacggctcc ctcttccttc cggctgtcgg gatccaggat 120
gaggggattt tccggtgcca ggcaatgaac aggaatggaa aggagaccaa gtccaactac 180
cgagtccgtg tctacc 196
<210> 53
<211> 25
<212> DNA
<213> Homo sapiens
<400> 53
gtaagaattc cagggtcttc tccaa 25
<210> 54
<211> 25
<212> DNA
<213> Homo sapiens
<400> 54
tctgactgga tttttcctcc ttcag 25
<210> 55
<211> 65
<212> DNA
<213> Homo sapiens
<400> 55
agattcctgg gaagccagaa attgtagatt ctgcctctga actcacggct ggtgttccca 60
ataag 65
<210> 56
<211> 25
<212> DNA
<213> Homo sapiens
<400> 56
gtagtggaag aaagcaggag aagta 25
<210> 57
<211> 25
<212> DNA
<213> Homo sapiens
<400> 57
tctgaggtca ccactctttc cccag 25
<210> 58
<211> 88
<212> DNA
<213> Homo sapiens
<400> 58
gtggggacat gtgtgtcaga gggaagctac cctgcaggga ctcttagctg gcacttggat 60
gggaagcccc tggtgcctaa tgagaagg 88
<210> 59
<211> 25
<212> DNA
<213> Homo sapiens
<400> 59
gtgagtccta aggtgccccc caagc 25
<210> 60
<211> 25
<212> DNA
<213> Homo sapiens
<400> 60
aatttgtctt atcctcccat catag 25
<210> 61
<211> 183
<212> DNA
<213> Homo sapiens
<400> 61
gagtatctgt gaaggaacag accaggagac accctgagac agggctcttc acactgcagt 60
cggagctaat ggtgacccca gcccggggag gagatccccg tcccaccttc tcctgtagct 120
tcagcccagg ccttccccga caccgggcct tgcgcacagc ccccatccag ccccgtgtct 180
ggg 183
<210> 62
<211> 25
<212> DNA
<213> Homo sapiens
<400> 62
gtgagcatag gtggggaggg cccca 25
<210> 63
<211> 25
<212> DNA
<213> Homo sapiens
<400> 63
acctcaaaac ccttccaact cccag 25
<210> 64
<211> 131
<212> DNA
<213> Homo sapiens
<400> 64
agcctgtgcc tctggaggag gtccaattgg tggtggagcc agaaggtgga gcagtagctc 60
ctggtggaac cgtaaccctg acctgtgaag tccctgccca gccctctcct caaatccact 120
ggatgaagga t 131
<210> 65
<211> 25
<212> DNA
<213> Homo sapiens
<400> 65
gtgagtgacc tggagagagg ggctg 25
<210> 66
<211> 25
<212> DNA
<213> Homo sapiens
<400> 66
gtctcctctc cccttccccc accag 25
<210> 67
<211> 142
<212> DNA
<213> Homo sapiens
<400> 67
ggtgtgccct tgccccttcc ccccagccct gtgctgatcc tccctgagat agggcctcag 60
gaccagggaa cctacagctg tgtggccacc cattccagcc acgggcccca ggaaagccgt 120
gctgtcagca tcagcatcat cg 142
<210> 68
<211> 25
<212> DNA
<213> Homo sapiens
<400> 68
gtgagacctc tccccaagcc ctaca 25
<210> 69
<211> 25
<212> DNA
<213> Homo sapiens
<400> 69
gactggatcc aactttgtct tccag 25
<210> 70
<211> 27
<212> DNA
<213> Homo sapiens
<400> 70
aaccaggcga ggaggggcca actgcag 27
<210> 71
<211> 25
<212> DNA
<213> Homo sapiens
<400> 71
gtgaggggtt tgataaagtc aggga 25
<210> 72
<211> 25
<212> DNA
<213> Homo sapiens
<400> 72
ctcaattttc cctgtctccg tacag 25
<210> 73
<211> 127
<212> DNA
<213> Homo sapiens
<400> 73
gctctgtggg aggatcaggg ctgggaactc tagccctggc cctggggatc ctgggaggcc 60
tggggacagc cgccctgctc attggggtca tcttgtggca aaggcggcaa cgccgaggag 120
aggagag 127
<210> 74
<211> 25
<212> DNA
<213> Homo sapiens
<400> 74
gtgagtggag aaagccagac ccctc 25
<210> 75
<211> 25
<212> DNA
<213> Homo sapiens
<400> 75
cattcccccc aatctttctc ctcag 25
<210> 76
<211> 272
<212> DNA
<213> Homo sapiens
<400> 76
gaaggcccca gaaaaccagg aggaagagga ggagcgtgca gaactgaatc agtcggagga 60
acctgaggca ggcgagagta gtactggagg gccttgaggg gcccacagac agatcccatc 120
catcagctcc cttttctttt tcccttgaac tgttctggcc tcagaccaac tctctcctgt 180
ataatctctc tcctgtataa ccccaccttg ccaagctttc ttctacaacc agagcccccc 240
acaatgatga ttaaacacct gacacatctt gc 272
<210> 77
<211> 50
<212> DNA
<213> Homo sapiens
<400> 77
tcttgtgtgt ctgtgtgtgt gtatgagaca caacctcacc cctataccct 50
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ ДЛЯ МОДУЛИРОВАНИЯ ЭКСПРЕССИИ C9ORF72 | 2016 |
|
RU2736574C2 |
| СОЕДИНЕНИЕ ДЛЯ МОДУЛЯЦИИ ЭКСПРЕССИИ ГЕНА C9ORF72 И ЕГО ПРИМЕНЕНИЕ | 2013 |
|
RU2730677C2 |
| НОВЫЙ СПОСОБ ЛЕЧЕНИЯ ПИГМЕНТНОГО РЕТИНИТА | 2020 |
|
RU2817702C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ СНИЖЕНИЯ ЭКСПРЕССИИ TAU | 2016 |
|
RU2747734C2 |
| ЛЕЧЕНИЕ СВЯЗАННЫХ С ГЕНОМ-СУПРЕССОРОМ ОПУХОЛЕЙ ЗАБОЛЕВАНИЙ ПОСРЕДСТВОМ ИНГИБИРОВАНИЯ ПРИРОДНОГО ТРАНСКРИПТА В АНТИСМЫСЛОВОЙ ОРИЕНТАЦИИ ОТНОСИТЕЛЬНО ЭТОГО ГЕНА | 2009 |
|
RU2746478C2 |
| МОДУЛЯЦИЯ ЭКСПРЕССИИ ФАКТОРА 11 | 2014 |
|
RU2739594C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ СНИЖЕНИЯ ЭКСПРЕССИИ TAU | 2016 |
|
RU2777570C2 |
| Система направленного изменения сплайсинга в гене MARK2 | 2023 |
|
RU2810907C1 |
| ИНГИБИТОР СИГНАЛА МИОСТАТИНА | 2019 |
|
RU2820270C2 |
| КОМПОЗИЦИИ И СПОСОБЫ МОДУЛЯЦИИ SMN2 СПЛАЙСИНГА У СУБЪЕКТА | 2019 |
|
RU2793459C2 |












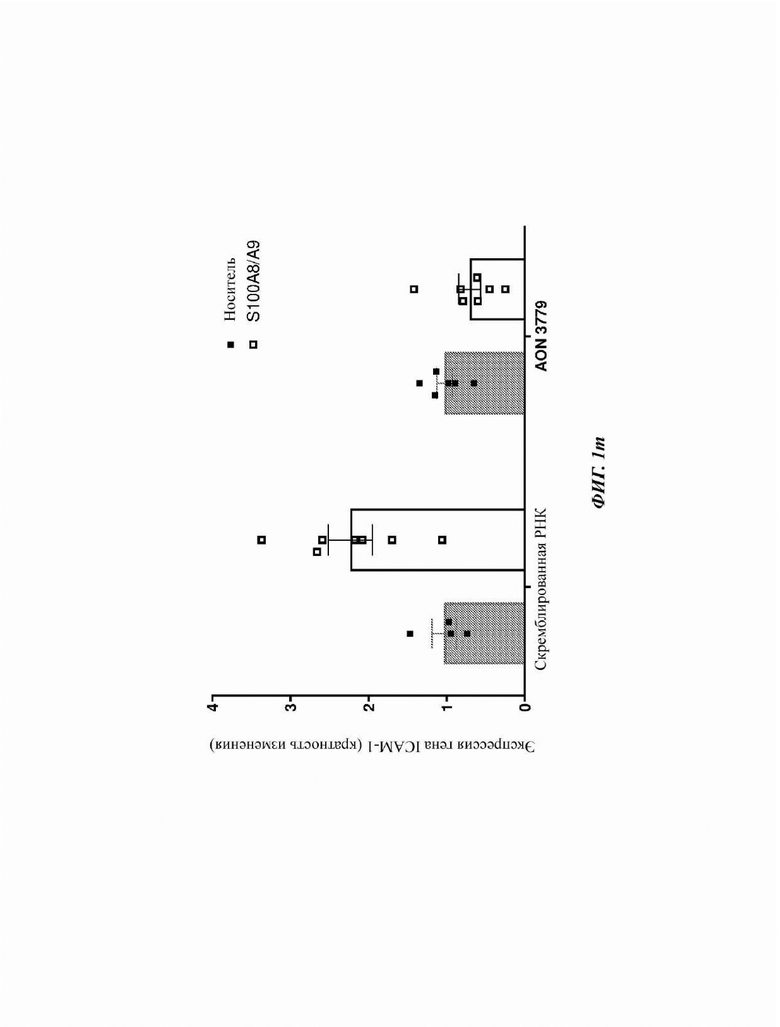

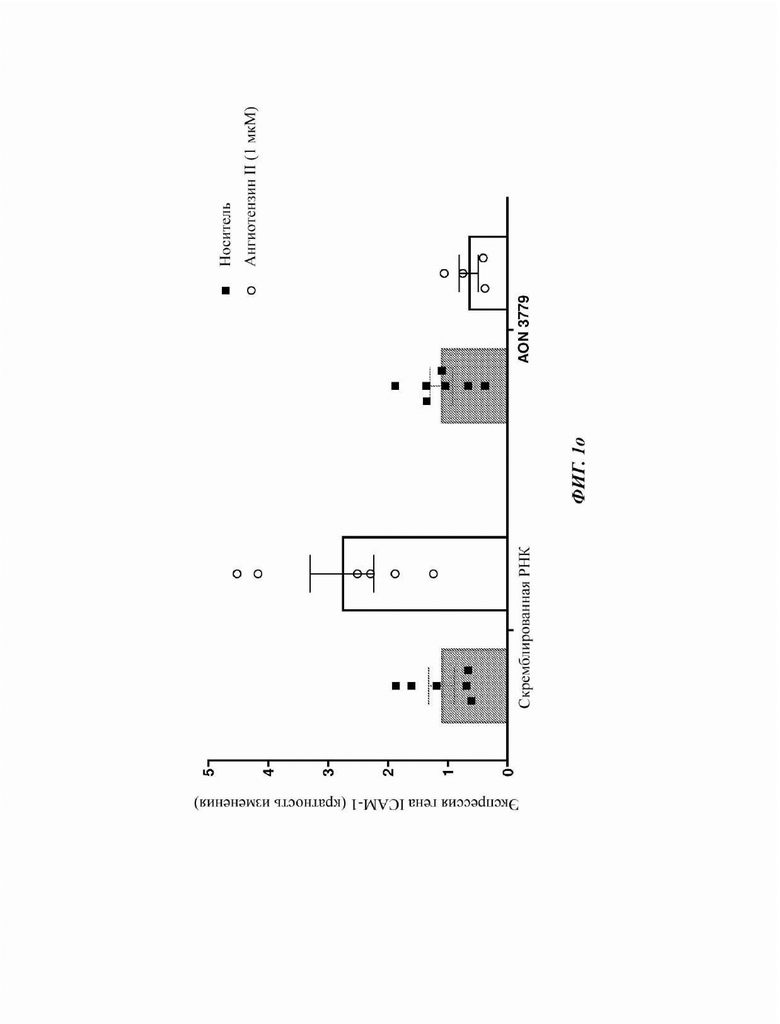







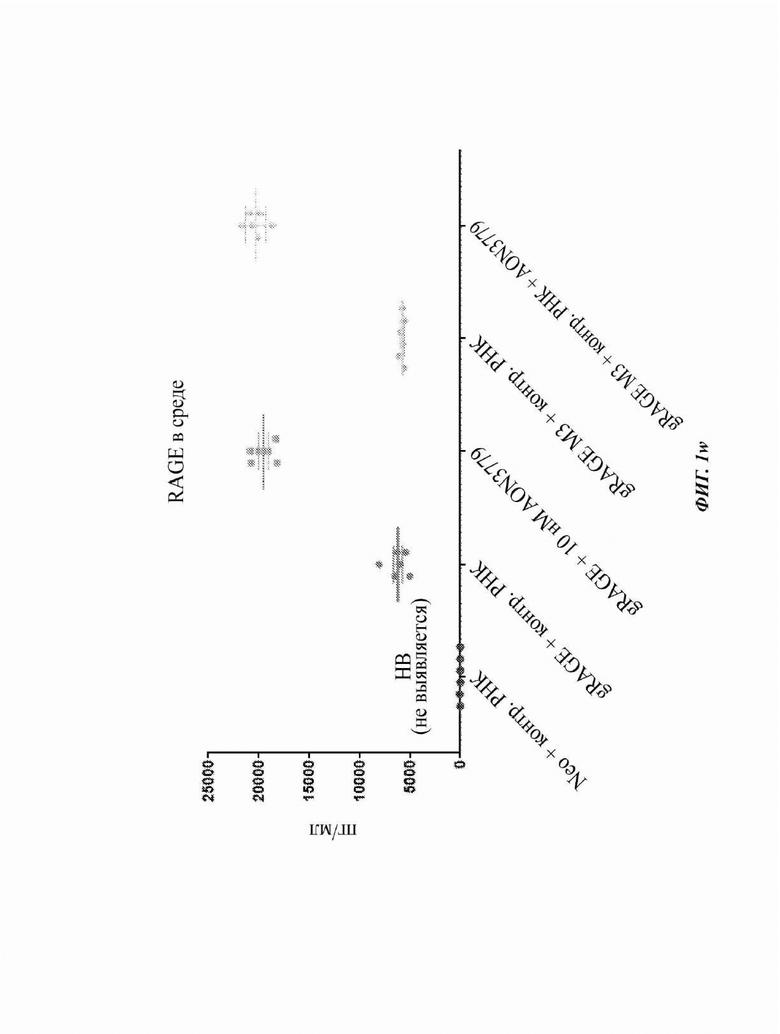


















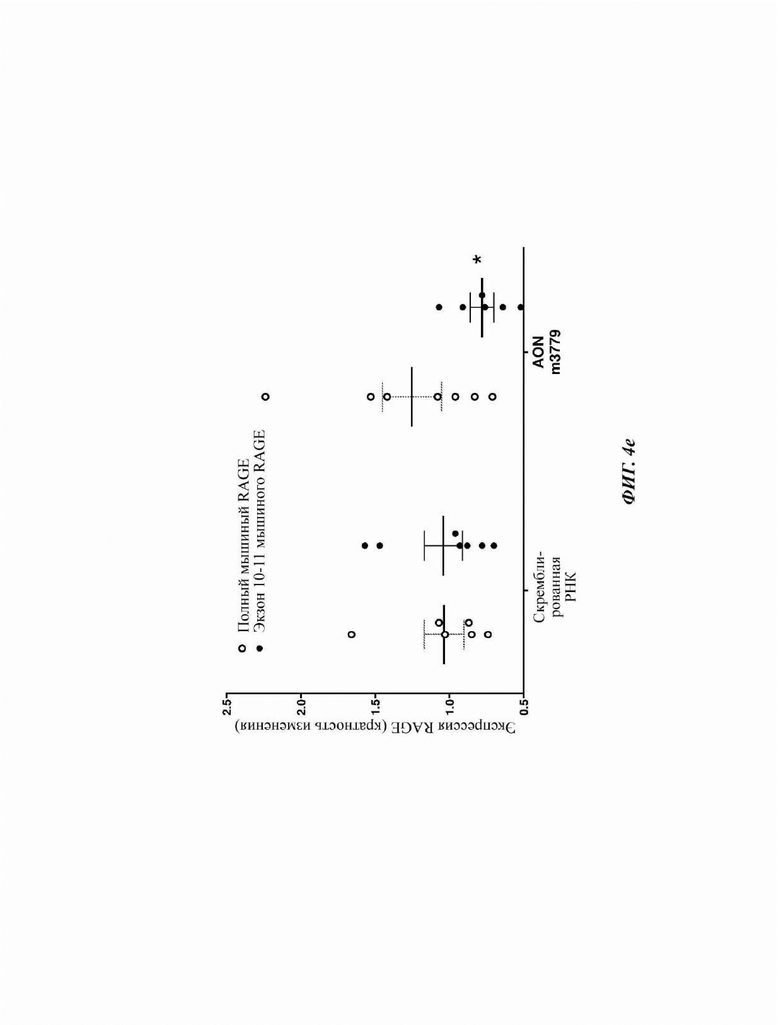





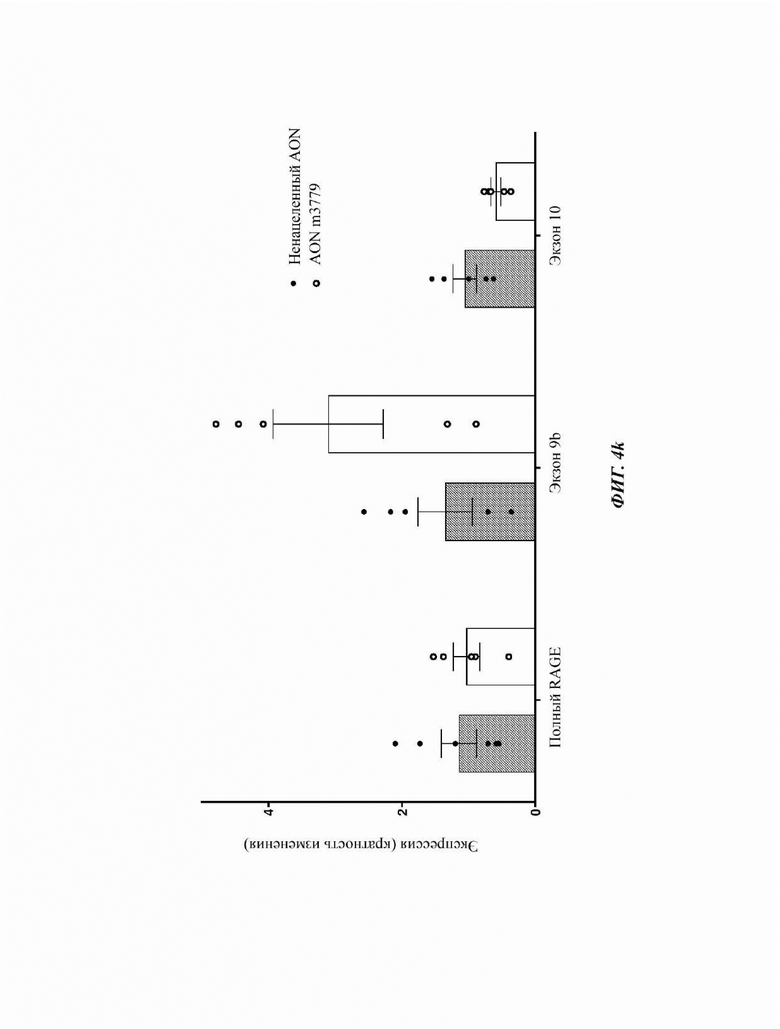




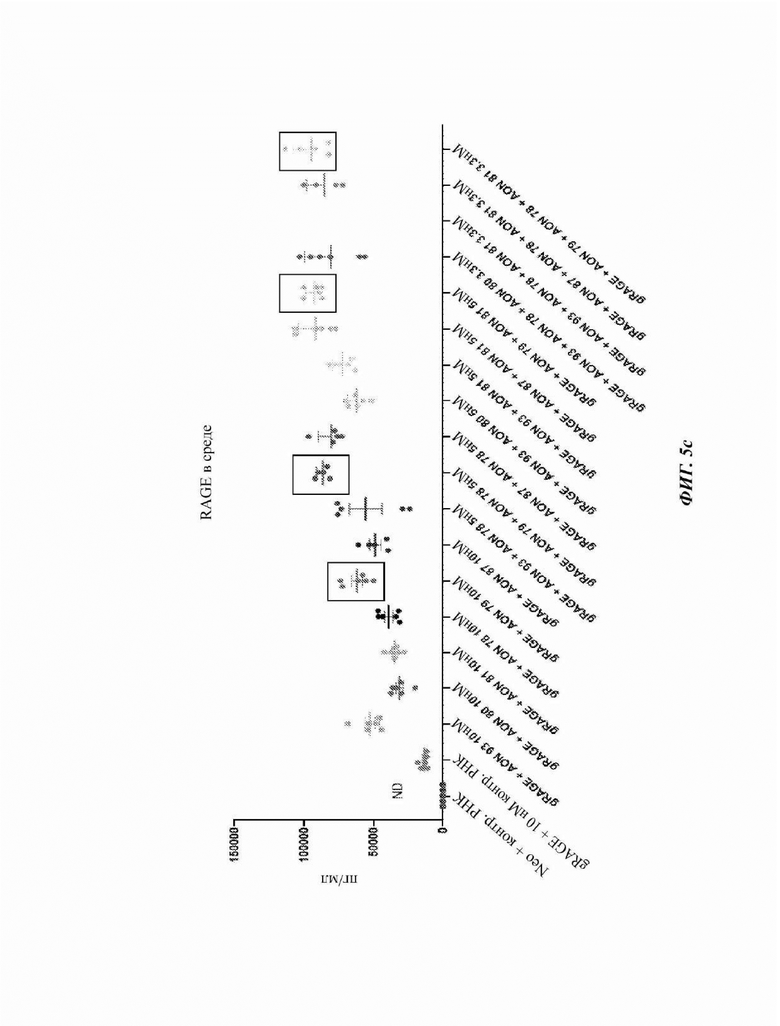


Настоящее изобретение относится к области биотехнологии, в частности к антисмысловым олигонуклеотидам и может быть использовано в медицине. Изобретение раскрывает новый антисмысловой олигонуклеотид для модифицирования сплайсинга пре-мРНК у рецептора конечных продуктов глубокого гликирования (RACE). Антисмысловые олигонуклеотиды согласно настоящему изобретению способны селективно снижать экспрессию провоспалительного трансмембранного RAGE (mRAGE; RAGE, содержащий экзон 10), одновременно увеличивая экспрессию внеклеточногорастворимого RAGE (esRAGE; пропуск экзона 10 приводит к сохранению интрона 9b). Изобретение может быть применимо в медицине для лечения, предупреждения или купирования эффектов заболевания, ассоциированных с экспрессией RAGE. 4 н. и 32 з.п. ф-лы, 5 ил., 3 табл., 5 пр.
1. Антисмысловой олигонуклеотид для манипулирования сплайсингом пре-мРНК рецептора конечных продуктов глубокого гликозилирования (RAGE), приводящему к пропуску экзона 10, содержащий:
нуклеотидную последовательность, по меньшей мере на 80% комплементарную области-мишени экзона 10 пре-мРНК RAGE по всей длине антисмыслового олигонуклеотида,
где крайний 5'-нуклеотид области-мишени является нуклеотидом в положении 88 или 114 или находится между положениями нуклеотидов 88 и 114 экзона 10 пре-мРНК RAGE,
где нумерация положений нуклеотидов экзона 10 пре-мРНК RAGE соответствует SEQ ID NO: 73,
при этом указанный антисмысловой олигонуклеотид имеет от 8 до 40 нуклеотидов в длину,
и при этом указанный антисмысловой олигонуклеотид содержит по меньшей мере один модифицированный нуклеотид.
2. Антисмысловой олигонуклеотид по п. 1, где крайний 5'-нуклеотид области-мишени находится:
а) в положении 88 или 113, или между положениями нуклеотидов от 88 до 113 экзона 10 пре-мРНК RAGE;
б) в положении 90 или 113, или между положениями нуклеотидов от 90 до 113 экзона 10 пре-мРНК RAGE;
в) в положении 88 или 108, или между положениями нуклеотидов от 88 до 108 экзона 10 пре-мРНК RAGE;
г) в положении 90 или 108, или между положениями нуклеотидов от 90 до 108 экзона 10 пре-мРНК RAGE; или
д) в положении 88 или 95, или между положениями нуклеотидов от 88 до 95 экзона 10 пре-мРНК RAGE.
3. Антисмысловой олигонуклеотид по п. 1 или 2, где область-мишень находится:
а) между положениями 88 и 113 экзона 10 пре-мРНК RAGE;
б) между положениями нуклеотидов от 90 до 113 экзона 10 пре-мРНК RAGE;
в) между положениями нуклеотидов от 88 до 108 экзона 10 пре-мРНК RAGE;
г) между положениями нуклеотидов от 90 до 108 экзона 10 пре-мРНК RAGE;
или
д) между положениями нуклеотидов от 88 до 95 экзона 10 пре-мРНК RAGE.
4. Антисмысловой олигонуклеотид по любому из пп. 1-3, содержащий нуклеотидную последовательность, по меньшей мере на 90% комплементарную области-мишени экзона 10 пре-мРНК RAGE.
5. Антисмысловой олигонуклеотид по любому из пп. 1-4, содержащий нуклеотидную последовательность, по меньшей мере на 95% комплементарную области-мишени экзона 10 пре-мРНК RAGE.
6. Антисмысловой олигонуклеотид по любому из пп. 1-5, содержащий нуклеотидную последовательность, на 100% комплементарную области-мишени экзона 10 пре-мРНК RAGE.
7. Антисмысловой олигонуклеотид по любому из пп. 1-6, длина которого составляет от 15 до 25 нуклеотидов.
8. Антисмысловой олигонуклеотид по любому из пп. 1-7, длина которого составляет 18 нуклеотидов.
9. Антисмысловой олигонуклеотид по любому из пп. 1-7, нуклеотидная последовательность которого является SEQ ID NO: 11 или нуклеотидной последовательностью, идентичной ей по меньшей мере на 85%, 90% или 95%.
10. Антисмысловой олигонуклеотид по любому из пп. 1-7, нуклеотидная последовательность которого является SEQ ID NO: 19 или нуклеотидной последовательностью, идентичной ей по меньшей мере на 85%, 90% или 95%.
11. Антисмысловой олигонуклеотид по любому из пп. 1-7, нуклеотидная последовательность которого является SEQ ID NO: 20 или нуклеотидной последовательностью, идентичной ей по меньшей мере на 85%, 90% или 95%.
12. Антисмысловой олигонуклеотид по любому из пп. 1-11, где модифицированный нуклеотид содержит одно или более из следующего: морфолиногруппа, модифицированная связь и модифицированная сахарная группировка.
13. Антисмысловой олигонуклеотид по п. 12, где модифицированная сахарная группировка включает метилирование в 2'-положении гидроксирибозы.
14. Антисмысловой олигонуклеотид по п.12, где модифицированная связь представляет собой фосфортиоатную связь.
15. Антисмысловой олигонуклеотид по любому из пп. 1-14, где если в антисмысловом олигонуклеотиде присутствует урацил, то урацил (U) антисмыслового олигонуклеотида заменен на тимин (Т).
16. Способ манипулирования сплайсингом в пре-мРНК рецептора конечных продуктов глубокого гликозилирования (RAGE), приводящий к пропуску экзона 10, включающий:
приведение клетки в контакт с одним или несколькими антисмысловыми олигонуклеотидами, содержащими нуклеотидную последовательность, по меньшей мере на 80% комплементарную области-мишени экзона 10 пре-мРНК RAGE по всей длине антисмыслового олигонуклеотида,
где крайний 5'-нуклеотид области-мишени является нуклеотидом в положении 88 или 114 или находится между положениями нуклеотидов 88 и 114 экзона 10 пре-мРНК RAGE,
где нумерация положений нуклеотидов экзона 10 пре-мРНК RAGE соответствует SEQ ID NO: 73,
при этом указанный антисмысловой олигонуклеотид имеет от 8 до 40 нуклеотидов в длину, и
при этом указанный антисмысловой олигонуклеотид содержит по меньшей мере один модифицированный нуклеотид; и
манипулирование сплайсингом в пре-мРНК RAGE, приводящее к пропуску экзона 10, происходит посредством связывания одного или нескольких антисмысловых олигонуклеотидов с областью-мишенью экзона 10 в пре-мРНК RAGE.
17. Способ лечения заболевания, связанного с экспрессией RAGE, включающий:
введение пациенту эффективного количества антисмыслового олигонуклеотида, содержащего нуклеотидную последовательность, по меньшей мере на 80% комплементарную области-мишени экзона 10 пре-мРНК RAGE по всей длине антисмыслового олигонуклеотида,
где крайний 5'-нуклеотид области-мишени является нуклеотидом в положении 88 или 114 или находится между положениями нуклеотидов 88 и 114 экзона 10 пре-мРНК RAGE,
где нумерация положений нуклеотидов экзона 10 пре-мРНК RAGE соответствует SEQ ID NO: 73,
при этом указанный антисмысловой олигонуклеотид имеет от 8 до 40 нуклеотидов в длину, и
при этом указанный антисмысловой олигонуклеотид содержит по меньшей мере один модифицированный нуклеотид, что представляет собой лечение пациента.
18. Способ по п. 16 или 17, где крайний 5'-нуклеотид области-мишени находится:
а) в положении 88 или 113, или между положениями нуклеотидов от 88 до 113 экзона 10 пре-мРНК RAGE;
б) в положении 90 или 113, или между положениями нуклеотидов от 90 до 113 экзона 10 пре-мРНК RAGE;
в) в положении 88 или 108, или между положениями нуклеотидов от 88 до 108 экзона 10 пре-мРНК RAGE;
г) в положении 90 или 108, или между положениями нуклеотидов от 90 до 108 экзона 10 пре-мРНК RAGE; или
д) в положении 88 или 95, или между положениями нуклеотидов от 88 до 95 экзона 10 пре-мРНК RAGE.
19. Способ по любому из пп. 16-18, где антисмысловой нуклеотид имеет нуклеотидную последовательность, по меньшей мере на 90%, по меньшей мере на 95% или на 100% комплементарную области-мишени экзона 10 пре-мРНК RAGE.
20. Способ по любому из пп. 16-19, где длина антисмыслового олигонуклеотида составляет от 15 до 25 нуклеотидов или 18 нуклеотидов.
21. Способ по любому из пп.16-20, где
антисмысловой олигонуклеотид имеет нуклеотидную последовательность SEQ ID NO: 11 или нуклеотидную последовательность, идентичную ей по меньшей мере на 85%, 90% или 95%; или
антисмысловой олигонуклеотид имеет нуклеотидную последовательность SEQ ID NO: 19 или нуклеотидную последовательность, идентичную ей по меньшей мере на 85%, 90% или 95%; или
антисмысловой олигонуклеотид имеет нуклеотидную последовательность SEQ ID NO: 20 или нуклеотидную последовательность, идентичную ей по меньшей мере на 85%, 90% или 95%.
22. Способ по любому из пп.16-21, где модифицированный нуклеотид содержит одно или более из следующего: морфолиногруппа, модифицированная связь и модифицированная сахарная группировка.
23. Способ по п. 22, где модифицированная сахарная группировка содержит метилирование в 2'- положении гидроксирибозы.
24. Способ по п. 22, где модифицированная связь представляет собой фосфортиоатную связь.
25. Способ по любому из пп. 16-24, где если в антисмысловом олигонуклеотиде присутствует урацил, то урацил (U) антисмыслового олигонуклеотида заменен на тимин (Т).
26. Способ по любому из пп. 16-25, где заболевание, связанное с экспрессией RAGE, выбрано из группы, состоящей из нейродегенеративных заболеваний, рака, легочных расстройств, воспалительных заболеваний, сердечно-сосудистых заболеваний; расстройств пищеварения; респираторных заболеваний, скелетно-мышечных заболеваний, заболеваний соединительной ткани, заболеваний почек, заболеваний половых органов, кожных заболеваний, заболеваний глаз и эндокринных заболеваний.
27. Применение эффективного количества антисмыслового олигонуклеотида при производстве лекарственного средства для лечения заболевания, связанного с экспрессией RAGE, где антисмысловой олигонуклеотид содержит:
нуклеотидную последовательность, по меньшей мере на 80% комплементарную области-мишени экзона 10 пре-мРНК RAGE по всей длине антисмыслового олигонуклеотида,
где крайний 5'-нуклеотид области-мишени является нуклеотидом в положении 88 или 114 или находится между положениями нуклеотидов 88 и 114 экзона 10 пре-мРНК RAGE,
где нумерация положений нуклеотидов экзона 10 пре-мРНК RAGE соответствует SEQ ID NO: 73,
при этом указанный антисмысловой олигонуклеотид имеет от 8 до 40 нуклеотидов в длину, и
при этом указанный антисмысловой олигонуклеотид содержит по меньшей мере один модифицированный нуклеотид.
28. Применение по п. 27, где крайний 5'-нуклеотид области-мишени находится:
а) в положении 88 или 113, или между положениями нуклеотидов от 88 до 113 экзона 10 пре-мРНК RAGE;
б) в положении 90 или 113, или между положениями нуклеотидов от 90 до 113 экзона 10 пре-мРНК RAGE;
в) в положении 88 или 108, или между положениями нуклеотидов от 88 до 108 экзона 10 пре-мРНК RAGE;
г) в положении 90 или 108, или между положениями нуклеотидов от 90 до 108 экзона 10 пре-мРНК RAGE; или
д) в положении 88 или 95, или между положениями нуклеотидов от 88 до 95 экзона 10 пре-мРНК RAGE.
29. Применение по п. 27 или 28, где нуклеотидная последовательность указанного антисмыслового олигонуклеотида по меньшей мере на 90%, по меньшей мере на 95% или на 100% комплементарна области-мишени экзона 10 пре-мРНК RAGE.
30. Применение по любому из пп. 27-29, где длина антисмыслового олигонуклеотида составляет от 15 до 25 нуклеотидов или 18 нуклеотидов.
31. Применение по любому из пп.26-30, где
антисмысловой олигонуклеотид имеет нуклеотидную последовательность SEQ ID NO: 11 или нуклеотидную последовательность, идентичную ей по меньшей мере на 85%, 90% или 95%; или
антисмысловой олигонуклеотид имеет нуклеотидную последовательность SEQ ID NO: 19 или нуклеотидную последовательность, идентичную ей по меньшей мере, на 85%, 90% или 95%; или
антисмысловой олигонуклеотид имеет нуклеотидную последовательность SEQ ID NO: 20 или нуклеотидную последовательность, идентичную ей по меньшей мере, на 85%, 90% или 95%.
32. Применение по любому из пп. 27-31, где указанный модифицированный нуклеотид содержит одно или более из следующего: морфолиногруппа, модифицированная связь и модифицированная сахарная группировка.
33. Применение по п. 32, где модифицированная сахарная группировка содержит метилирование в 2'-положении гидроксирибозы.
34. Применение по п. 32, где модифицированная связь представляет собой фосфортиоатную связь.
35. Применение по любому из пп. 27-34, где если в антисмысловом олигонуклеотиде присутствует урацил, то урацил (U) антисмыслового олигонуклеотида заменен на тимин (Т).
36. Применение по любому из пп. 27-35, где заболевание, связанное с экспрессией RAGE, выбрано из группы, состоящей из нейродегенеративных заболеваний, рака, легочных расстройств, воспалительных заболеваний, сердечно-сосудистых заболеваний; расстройств пищеварения; респираторных заболеваний, скелетно-мышечных заболеваний, заболеваний соединительной ткани, заболеваний почек, заболеваний половых органов, кожных заболеваний, заболеваний глаз и эндокринных заболеваний.
| KALEA A | |||
| et al.: "Alternative splicing of RAGE: roles in biology and disease", Front Biosci | |||
| (Landmark Ed), 2011, v | |||
| Устройство для электрической сигнализации | 1918 |
|
SU16A1 |
| AARTASMA-RUS A | |||
| et al.: "Antisense-mediated exon skipping: a versatile tool with therapeutic and research applications", RNA, 2007, v | |||
| Насос | 1917 |
|
SU13A1 |
| ANDO R | |||
| et al.: "Involvement of advanced glycation end | |||
