Изобретение относится к медицине, а именно к способам моделирования деперсонализационного расстройства, и может быть использовано при разработке новых потенциальных психотропных средств.
Деперсонализация традиционно рассматривается как чисто «человеческое» расстройство, которому присущи симптомы, описать которые способен только человек. Ее определяют как расстройство осознания собственной личности, при котором происходит отчуждение всех психических процессов (Cattel J.P. Depersonalization phenomena // America Handbook of psychiatry, v.30 /Ed. by S.Ariety/, 1966, Basic Book, Inc, Publishers. NY, pp.88-102 [1]; DSM-IY [2]). Подчеркивается, что деперсонализация носит сугубо субъективный характер (Tucker G.J., Harrow M., Quinlan D. Depersonalization, dysphoria and though disorder // Am.J.Psychiatry, 1973, v.130, pp.702-706 [3]). Поэтому неудивительно, что нам неизвестны специальные исследования механизмов патогенеза деперсонализации в эксперименте на животных. Попыток моделирования этого состояния не предпринималось, что и определяет актуальность настоящей разработки.
Деперсонализационное расстройство не специфично и встречается при многих психических заболеваниях, височной эпилепсии, пограничных состояниях, а также в легкой и непродолжительной форме у психически здоровых людей при больших эмоциональных нагрузках, усталости, после родов, при отравлениях лекарственными и наркотическими веществами. Очень часто развитию деперсонализации предшествует выраженная острая психотравма, а в преморбиде большинству больных, перенесших деперсонализационное расстройство, свойственны тревожность, впечатлительность, ранимость, т.е. гиперэмоциональность. При деперсонализации же практически всегда отмечается снижение или утрата эмоционального компонента психических процессов (Hollander E., Carrasco J.L; Mullen L.S. et al. // Biol.Psychiatry, 1992, v.31, N 11, pp.1157-1162 [4]). Некоторые авторы считают это сутью данного феномена (Нуллер Ю.Л., Михаленко И.Н. Аффективные психозы. Л., 1988 [5]), направленного, по-видимому, в качестве защитной реакции против отрицательного эмоционального состояния, такого, например, как тревога и депрессия (Нуллер Ю.Л. Парадигмы в психиатрии. Киев. 1993 [6]), [3].
Существенно, что деперсонализация, являясь, по выражению Mayer-Gross (1935), «преформированным функциональным ответом мозга», который может быть вызван многочисленными факторами, описана не только у психически больных, но и у здоровых людей. При этом имеются свидетельства об идентичности психопатологического и кратковременного, возникшего вследствие острой стрессовой ситуации, например, при автомобильной аварии, состояний (Roberts W.W. Normal and abnormal depersonalization // J.Ment Sci., I960. v.106, pp.478-493 [7]). Это указывает на ключевую роль стресса в развитии деперсонализации.
Следующий важный момент заключается в том, что у многих больных с деперсонализацией отмечается притупление болевой чувствительности вплоть до ее полного исчезновения [1], [5] (Мороз Б.Т., Нуллер Ю.Л., Устимова И.Н., Андреев Б.В. Исследование болевой чувствительности по показателям электроодонтометрии у больных с деперсонализацией и депрессией. //Журнал невропатологии и психиатрии им. С.С.Корсакова. 1990, т.90, N10, с.81-82 [8]) и снижение реакции на введение морфина [5], т.е. имеют место симптомы, отражающие повышение активности эндогенной антиноцицептивной системы. Описан и положительный опыт использования налоксона - блокатора опиатных рецепторов, при лечении симптомов деперсонализации (Нуллер Ю.Л., Морозова М.Г., Дубинина Е.Е., Кушнир О.Н., Гампер Н.Л. Использование налоксона для лечения деперсонализации и исследования ее патогенеза. // Обозрение психиатрии и медицинской психологии им. В.М.Бехтерева. 1999. N2, с.56-59 [9]).
Все это позволяет предположить, что в основе данной патологии лежат фундаментальные механизмы деятельности мозга, свойственные не только человеку, но и животным, что, в свою очередь, открывает возможности для моделирования этого «чисто человеческого страдания».
В настоящее время стрессу отводят пусковую роль в возникновении и развитии ряда психических заболеваний. Стресс - неспецифическая реакция организма, она всегда комплексная и включает изменения со стороны различных функциональных систем и поведения в целом. Одной из составляющих адаптационной реакции на стрессорное воздействие является и подавление болевого восприятия. Этот феномен, получивший название «аналгезия, вызванная стрессом» (Amit Z. Galina Z.H. Stress Induced analgesia play an adaptive role in the organization of behavioral responding. // Brain Research Bulletin. 1988, v.21, N6, pp.955-958 [10]), описан как у животных, так и у человека. В эксперименте к настоящему времени он широко и разносторонне изучен. Показано влияние на развитие аналгезии, вызванной стрессом, разного рода факторов, в первую очередь, интенсивности и продолжительности стрессорного воздействия при ключевом условии его неизбегаемости. Показано, что выраженность феномена у животных может быть использована как селекционной признак. Это указывает на генетическую детерминированность данной реакции. Существенно, что продолжительность и степень повышения болевых порогов зависят как от физических параметров стрессора, так и его эмоциональной значимости. Более того, выраженность негативного аффекта влияет на сам характер изменения болевого восприятия. Так, показано, что развитие гипоалгезии связано с сильным негативным аффектом (состояние страха), при средней же степени его выраженности (тревоге) отмечается понижение болевых порогов (обострение болевой чувствительности - гипералгезия) (Rhudy J.L., Meagher M.W. Negative affect: effects on an evaluative measure of human pain // Pain. 2003, v.104, N3, pp.617-626 [11]).
Поскольку боль, по определению, является неприятным сенсорным и эмоциональным переживанием, выделяют сенсорный и аффективный компоненты боли. Хорошо известно, что отнюдь не всегда эти компоненты однозначно взаимосвязаны. Состояние деперсонализации является примером именно такой диссоциации, поскольку показано, что у больных с деперсонализацией по сравнению с контрольными группами значительно повышены именно пороги переносимости боли, отражающие главным образом эмоциональный ответ (патент на изобретение №2211711, кл. А61К 1/32, 2002 г. [12]). Очевидно, моделирование деперсонализационного расстройства должно учитывать как эмоциональную, так и собственно ноцицептивную составляющую болевого восприятия.
Экспериментальное моделирование психопатологических состояний на животных связано в первую очередь с применением разнообразных моделей стресса.
Наиболее близким по технической сущности и достигаемому результату является самая распространенная при использовании лабораторных грызунов (мышей и крыс) методика принудительного (неизбегаемого) плавания. Существует огромное разнообразие методических приемов, базирующихся на этой модели, но все они, по сути, являются модификациями известного теста Порсолта (Porsolt R.D., Lenegre A., McArthur R.A. Pharmacological models of depression. // In: Animal Models in Pharmacology. Advances in Pharmacological Sciences. Birkhanser Verlag Basel. 1991, pp.137-159 [13]). Основной показатель этого теста - длительность иммобилизации во время плавания - широко вошел в исследовательскую практику и используется не только при скрининге новых антидепрессантов (изначальное предназначение теста), но и для исследования нейрохимических процессов, оценки поведения животных при стрессе. Методика принудительного плавания применяется и для анализа механизмов аналгезии, вызванной стрессом. Однако при этом используется только сам методический прием - плавание как неизбегаемое стрессорное воздействие.
При экспериментальном изучении реакций на стресс выделяют два основных типа: активный (борьба/бегство) и пассивный (иммобилизация). Иммобилизация трактуется многими авторами вслед за создателями теста, как «поведение отчаяния» и эквивалент депрессии человека. Вместе с тем, несмотря на широкое распространение самого методического приема, интерпретация форм двигательной активности животных в момент тестирования остается предметом дискуссии (Korte S.M., De Kloet E.R., Buwalda В., et al. Antisense to the glucocorticoid receptor in hippocampal dentate gyrus reduces immobility in forced swim test. // Europ J Pharmacol.1996, v.301, N1-3, pp.19-25 [14]).
Между тем в клинике часто описывается коморбидность депрессии и деперсонализации, в том числе с выделением депрессивно-деперсонализационного расстройства [6] (Simeon D., Gross S., Guralnik O., Stein D.J., Schmeidler J., Hollander E. Feeling Unreal: 30 cases of DSM-Ш-R depersonalization disorder // Am J Psychiatry. 1997, v.158, N 8, pp.1107-1113 [15]), вытекает предположение о том, что эти два состояния представляют континуум адаптационных реакций организма человека при нарастании тревоги (стресса). Исходя из этого представляется, что сочетанная регистрация двигательного поведения, в частности продолжительности состояния иммобилизации в условиях стрессорного воздействия, и выраженности формирующейся аналгезии позволяет более полно представить и адекватно оценить реакцию животных на стресс, включая ее эмоциональный компонент.
Технический результат, на достижение которого направлено настоящее изобретение, заключается в разработке модели деперсонализационного расстройства, которая позволит изучать механизмы формирования и разрабатывать методы терапии состояния, проявляющегося притуплением эмоциональной сферы личности, эмоциональной отгороженностью («скорбное бесчувствие»).
Указанный технический результат достигается тем, что в способе моделирования деперсонализационного расстройства, согласно изобретению, у мышей или крыс определяют болевые пороги (чувствительности и переносимости), затем животных подвергают неизбегаемому стрессорному воздействию в течение 4-6 минут, при этом регистрируют суммарную продолжительность иммобилизации, и через 2-10 минут после окончания стрессорного воздействия повторно определяют болевые пороги, при этом по разнице болевых порогов судят о выраженности аналгезии, вызванной стрессом, а чувствительность животных к стрессу и выраженность "деперсонализации" определяют по характеру сочетанных изменений болевых порогов и продолжительности иммобилизации.
Кроме того, грызунам за 20-30 минут перед стрессорным воздействием могут проводить инъекции нейротропными средствами в расчете 0,1 мл раствора препарата на 10 г массы, при этом через 15-25 минут непосредственно перед стрессорным воздействием повторно замеряют болевые пороги.
В качестве нейротропного средства могут использовать коразол (пентилентетразол, 20 мг/кг) или диазепам (0,25-0,5 мг/кг).
Способ выполняют следующим образом.
Все эксперименты начинали с определения болевых порогов, которые фиксировали индивидуально для каждого животного по первому вздрагиванию (порог чувствительности) и по реакции вокализации (порог переносимости), появляющихся при увеличении напряжения постоянного электрического тока, подаваемого на предварительно смоченные физиологическим раствором лапы животного через токопроводящий пол. Болевое раздражение (прямоугольные импульсы постоянного тока 1,5 Гц до 50 В) продолжительностью 0,2 с наносили, начиная с 1 В с постепенным увеличением напряжения (1 В в 5 секунд). За величину порога принимали среднее из значений, полученных в трех последовательных измерениях, проводимых с 60-секундными интервалами.
Сразу после определения болевых порогов животных подвергали стрессорному воздействию в виде принудительного плавания в холодной воде.
При использовании теста принудительного плавания в холодной воде мышь помещали в сосуд высотой 30 см и диаметром 9 см, наполовину заполненный водой, температура которой составляла 11°С, и в течение 4 минут наблюдали за ее поведением, фиксируя суммарную продолжительность пассивного плавания (иммобилизацию). По окончании теста животное извлекали из воды и обтирали теплым полотенцем, а спустя 10 минут вновь измеряли пороги болевой чувствительности, по их изменению судили о выраженности аналгезии, вызванной стрессом.
При использование в качестве объекта наблюдения крыс тестирование проводили в сосуде размером 50×40 см. Наблюдение вели в течение 6 минут аналогично описанному выше.
При исследовании влияния нейротропных средств на развитие поведенческих и аналгетической реакций на стресс препараты вводили животным после определения у них исходных значений болевых порогов, а непосредственно перед стрессорным воздействием болевые пороги измеряли повторно. Таким образом, в течение эксперимента у каждого животного уровень болевых порогов определяли трижды. По их изменению между первым и вторым замерами судили о влиянии препарата, а между вторым и третьим - о выраженности аналгезии, вызванной стрессом.
В работе применялись следующие препараты: коразол (пентилентетразол, RBI), диазепам (реланиум, Polfa). Физиологический раствор, а также растворы коразола и диазепама вводили из расчета 0,1 мл раствора препарата на 10 г массы.
Статистическую обработку результатов осуществляли на PC с помощью программных продуктов Excel 2002, «Statistica 5.0 for Windows» и "Origin 6.1 for Windows», при анализе полученных данных использованы методы ANOVA и линейной регрессии.
Результаты исследования
Стресс, связанный с неизбегаемым плаванием в холодной воде, приводит к развитию у лабораторных грызунов (мышей и крыс) постстрессорной аналгезии, проявляющейся увеличением порога переносимости боли. Спустя 10 минут по окончании плавания значение показателя возрастало в среднем в группе крыс на 5,6±1,6 В (n=23, Р<0.01), в группах мышей SHR на 4,4±1,0 В (n=32, Р<0.001), NMRI на 11±1,9 В (n=29, Р << 0.001). Интересно, что при этом не отмечено увеличения порогов чувствительности боли. У крыс и мышей NMRI они практически не изменялись по сравнению с исходными значениями (-0,1±0,3 В), а у мышей SHR даже снижались (-1,3±0,4В, Р<0,01).
Полученные данные полностью соответствуют данным литературы о том, что острая неизбегаемая стрессорная ситуация приводит к развитию аналгезии (см. выше). В то же время снижение болевой чувствительности оказывается связанным только с увеличением порога переносимости боли, порог же чувствительности практически не изменяется, что указывает на подавление именно эмоциональной компоненты болевой реакции в условиях стресса.
Регистрация двигательного поведения животных в условиях неизбегаемого стресса выявила существенные различия между мышами разных линий. Если для беспородных животных, как и для крыс, была характерна преимущественно пассивная стратегия поведения, то мыши линии NMRI реагировали на него существенно более активно, но при этом заметно тяжелее переносили тестирование. В среднем в состоянии иммобилизации они проводили лишь треть времени экспозиции (32±3%), в то время как беспородные мыши 61,6±2,1%, а крысы 54,2±2,7%.
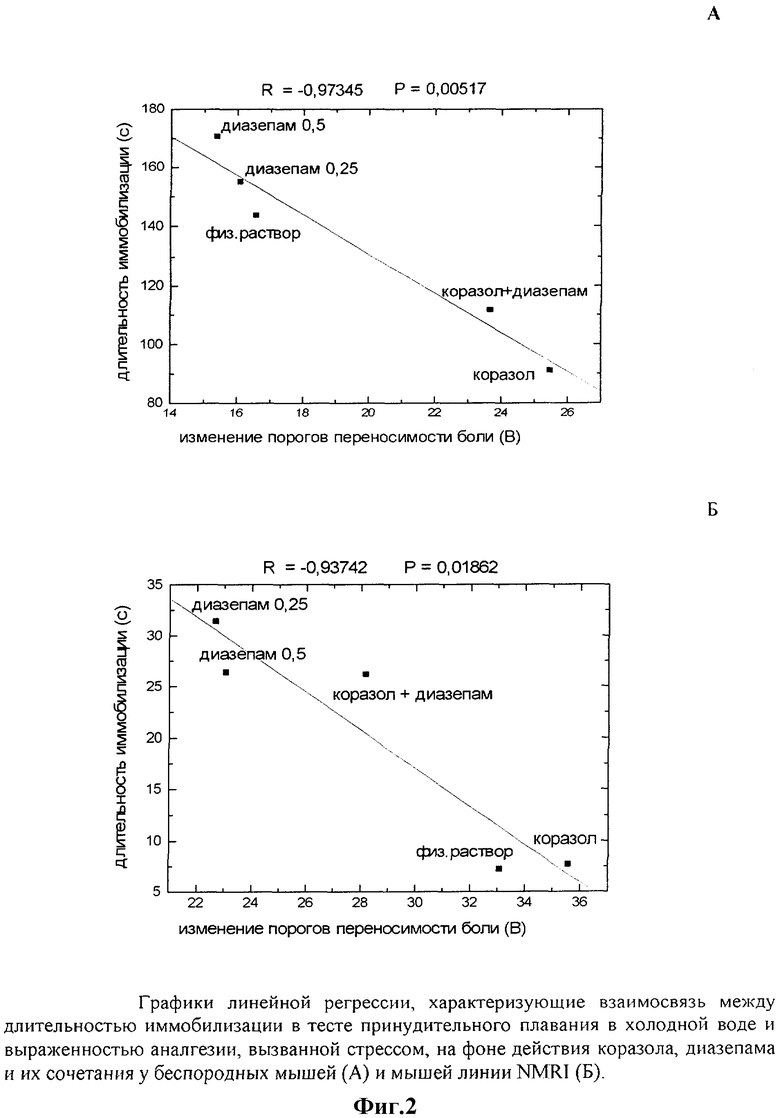
Индивидуальные значения показателей продолжительности иммобилизации во время принудительного плавания и изменения порогов переносимости боли в результате неизбегаемого стресса варьировали в довольно широком диапазоне во всех опытных группах. Имеет ли место зависимость между поведенческой реакцией на стресс и развитием вызванной им аналгезии, или эти феномены самостоятельны? Для ответа на этот вопрос была проанализирована связь между регистрируемыми показателями и выявлена отрицательная корреляционная зависимость между ними (фиг.1). Чем больше у животного преобладала пассивная стратегия поведения во время принудительного плавания, тем меньше была выражена аналгезия, вызванная этим стрессом, оцениваемая по изменению порога переносимости боли. Значение коэффициента корреляции составило для группы крыс r=-0,497 (n=23; Р<0,02), мышей SHR r=-0,378 (n=32; Р=0,04), мышей NMRI r=-0,526 (n=29; Р<0,01).
Таким образом, обнаружена тесная связь между двигательной активностью животных при стрессе и выраженностью постстрессорной аналгезии, которая, по-видимому, носит универсальный характер, так как выявлена у животных разных видов (мышей и крыс) и линий (мыши SHR и NMRI), и позволяет рассматривать эти феномены как единое целое в рамках общей реакции на стресс.
К настоящему времени однозначно установлено, что выраженность стрессорной реакции зависит как от физических параметров внешнего воздействия, так и от состояния животного. С одной стороны, более тяжелый стресс приводит к развитию более сильной аналгезии (Mogil J.C., Sternberg W.F., Balian H., Liebeskind J.C., Sadowski B. Opioid and nonopioid swim-induced analgesia: a parametric analysis in mice. // Physiol Behav. 1996, v.59, N1, pp.123-132 [16]).
С другой, при одном и том же физическом воздействии реакция на стресс зависит от субъективной оценки его тяжести, что показано не только в отношении человека [11], но и животных (Janssen S.A. Negative affect and sensitization to pain // Scand J Physiol, 2002, v.43, pp.131-137 [17]).
Очевидно, что выявленные в работе как внутригрупповая, так и межлинейная вариативности регистрируемых показателей, проявляющиеся в одних и тех же методических условиях, отражают в первую очередь психологическую составляющую стрессорной реакции. Таким образом, можно предполагать, что более сильная аналгезия, развивающаяся у мышей линии NMRI в ответ на неизбегаемое плавание в холодной воде, указывает на более субъективно тяжелый для них стресс, а, следовательно, и на их большую чувствительность к нему или, иначе говоря, на их большую тревожность.
Подчеркнем, что повышенная реактивность, тревожность или гиперэмоциональность больных отмечается практически во всех клинических исследованиях состояния деперсонализации /15/ (Sierra M., Berrios G.E. Depersonalization: neurobioligical perspectives // Biol Psychiatry. 1998, v.44, N9, pp.898-908 [18]).
Исследование эмоционального статуса в развитии изучаемых феноменов в условиях эксперимента традиционно проводят с использованием фармакологических агентов, влияющих на уровень тревоги. Результаты серии экспериментов в нашем случае показали, что предварительное введение стандартного анксиогена пентилентетразола (коразола, 20 мг/кг, подкожно, за 20 минут до тестирования) потенцирует стрессорное воздействие. Это проявилось увеличением выраженности аналгезии, вызванной стрессом, и сокращением продолжительности иммобилизации у мышей в тесте принудительного плавания в холодной воде (Табл.1).
Влияние коразола на продолжительность состояния иммобилизации и изменение болевых порогов под влиянием стресса
При этом переносимость стресса заметно утяжелялась: на фоне предварительно введенного анксиогена во время плавания у животных часто отмечали нарушение координации движений, погружение головы под воду выше линии ушей, а по окончании тестирования - экстензии конечностей и более длительное боковое положение.
Предварительное (за 30 минут до принудительного плавания) введение эталонного анксиолитика диазепама (0,25 мг/кг, внутрибрюшинно) в целом приводило к противоположным в сравнении с коразолом эффектам: выраженность аналгезии, вызванной стрессом, уменьшалась по сравнению с данными контрольных групп, а продолжительность иммобилизации во время стресса возрастала (Табл.2). Наиболее ярко эффект анксиолитика проявился у мышей NMRI. Под влиянием диазепама заметно облегчалась и переносимость животными тяжелых условий стресса: проведя часть времени тестирования в состоянии «поплавка», по окончании теста они сохраняли компактную позу, способность передвигаться, быстрее высыхали.
Влияние диазепама на продолжительность состояния иммобилизации и изменение болевых порогов под влиянием стресса у мышей линии NMR.I
Вновь обращает на себя внимание тот факт, что выраженность постстрессорной аналгезии обусловлена изменением только порогов переносимости боли при полном отсутствии изменений со стороны порогов чувствительности, что с определенностью указывает на изменение эмоциональной составляющей восприятия боли под влиянием стресса.
Результаты, полученные на этом этапе исследования, свидетельствуют о том, что изменение «тревожного статуса» животных с помощью анксиотропных препаратов влияет как на поведение животных во время стресса, так и на развитие вызванной этим стрессом аналгезии. При этом, с одной стороны, введение коразола и диазепама приводило к полярным изменениям регистрируемых показателей, с другой, в отношении друг друга показатели изменялись реципрокно, что снова указывает на наличие между ними взаимосвязи, подобной выявленной нами ранее на уровне отдельных групп (фиг.2).
Таким образом, поскольку стратегия поведения при стрессе оказывается тесно связанной с развитием аналгезии, вызванной этим стрессом, вполне правомерно говорить о единстве механизмов, лежащих в основе этих двух проявлений реакции на стресс. При этом очевидно, что тревога играет роль модулирующего фактора, от выраженности которого в значительной степени зависит формирование ответного паттерна на стресс.
Нейрохимические процессы, лежащие в основе реакций, запускаемых стрессом, сложны и многоплановы. В реализации адаптационного ответа на вызовы окружающей среды, который включает в том числе как изменения двигательного поведения, так и развитие аналгезии, задействованы гипоталамо-гипофизарно-надпочечниковая, моноамин-, опиат-, ВАКергические и другие системы мозга, что затрудняет интерпретацию полученных данных с точки зрения выявления «ответственной за феномен» системы (Bremner J.D., Vermetten E. Stress and development: Behavioral and biological consequences // Development and Psychopatology. 2001, v.13, pp.473-489 [19]; Wiedenmayer C.P. Adaptation or pathologies? Long-term changes in brain and behavior after a single exposure to severe threat // Neurosci Biobehav Rev. 2004, v.28, pp.1-12 [20]). Тем не менее, можно предполагать, что одна из ключевых ролей принадлежит ГАМКергической системе, которую рассматривают в качестве основной при осуществлении тормозных процессов в ЦНС. Именно с активацией ГАМКергической передачи связывают эффекты стандартных противотревожных препаратов, в то время как анксиогенный эффект коразола объясняют противоположным действием в отношении ГАМКергических нейронов. Исследование влияния сочетанного введения коразола и диазепама на изменение регистрируемых показателей, проведенное в нашей работе, показало, что во всех случаях оно сопровождалось взаимным снижением количественных эффектов исследуемых препаратов (фиг.2), что вполне согласуется с общеизвестным конкурентным действием веществ через ГАМКергические системы.
Роль тревоги хорошо известна в отношении развития психопатологических состояний у человека, в частности, депрессии и деперсонализации [6] (Young L.Т., Cooke R.G., Robb J.C. et al. Anxious and non-anxious bipolar disorder. // J Affect Disord. 1993, v.29, N1, pp.49-52 [21]), возникновению которых тоже, как правило, предшествует перенесение стресса [3]. Наши результаты вполне соотносятся с тем, что наблюдается в клинической практике при развитии деперсонализационного расстройства. Чем сильнее стресс или острее его восприятие, тем выше вероятность развития деперсонализации. Лечение этого расстройства бензодиазепиновыми анксиолитиками возможно на ранних стадиях [5] и снижение последствий стресса подобными препаратами может предотвратить ее развитие.
К сожалению, терапия сформированного деперсонализационного синдрома остается практически неразработанной, и эмпирически применяемые препараты оказываются чаще малоактивными. Предлагаемая модель заболевания может способствовать пониманию механизмов его развития и оптимизации лечения.
Сущность способа поясняется примерами.
Пример 1.
Опыт выполнен в осенний период на 15 мышах самцах SHR (20-26 г), полученных из питомника "Рапполово".
Эксперимент начинали с определения болевых порогов, которые фиксировали индивидуально для каждого животного по первому вздрагиванию (порог чувствительности) и по реакции вокализации (порог переносимости), появляющихся при увеличении напряжения постоянного электрического тока, подаваемого на предварительно смоченные физиологическим раствором лапы животного через токопроводящий пол. Болевое раздражение (прямоугольные импульсы постоянного тока 1,5 Гц до 50 В) продолжительностью 0,2 с наносили, начиная с 1 В с постепенным увеличением напряжения (1 В в 5 секунд). За величину порога принимали среднее из значений, полученных в трех последовательных измерениях, проводимых с 60-секундными интервалами.
Сразу после определения болевых порогов мышь помещали в сосуд высотой 30 см и диаметром 9 см, наполовину заполненный водой, температура которой составляет 11°С, и в течение 4 минут наблюдали за ее поведением, фиксируя суммарную продолжительность пассивного плавания (иммобилизацию). По окончании теста животное извлекали из воды и обтирали теплым полотенцем, а спустя 10 минут вновь измеряли пороги болевой чувствительности.
Статистическую обработку результатов осуществляли на PC с помощью программных продуктов Excel 2002 и "Origin 6.1 for Windows», при анализе полученных данных использованы методы ANOVA и линейной регрессии.
Результаты проведенного эксперимента представлены в таблицах.
Пороги болевой чувствительности, измеренные у каждого подопытного животного непосредственно перед тестом принудительного плавания в холодной воде
Пороги болевой чувствительности, измеренные у каждого подопытного животного спустя 10 минут по окончании теста принудительного плавания в холодной воде
Длительность иммобилизации во время неизбегаемого плавания в холодной воде и изменения порогов болевой чувствительности вследствие этого стресса
Полученные результаты свидетельствуют о том, что в результате неизбегаемого стресса у группы животных, представленной в данном примере, произошло достоверное повышение порогов переносимости боли на фоне практически неизменившихся порогов чувствительности. Статистический анализ связи между величиной изменения порога переносимости боли и продолжительностью состояния иммобилизации во время принудительного плавания показал наличие отрицательной корреляции между показателями (R=-0,601; Р<0,02).
Пример 2.
Данная серия экспериментов выполнена на 23 мышах-самцах линии NMRI (вес 22-26 г) в зимний период. Животных поделили на две группы. После определения исходных значений болевых порогов за 20 минут до плавания мышам первой группы вводили физиологический раствор, а второй коразол в дозе 20 мг/кг (подкожно). Через 15 минут после введения и непосредственно перед стрессорным воздействием у животных повторно измеряли болевые пороги, после чего их помещали в условия неизбегаемого плавания, а спустя 10 минут после описанной ранее процедуры тестирования вновь приступали к измерению порогов болевой чувствительности. Таким образом, в течение данного эксперимента у каждого животного уровень болевых порогов определяли трижды по схеме, аналогичной представленной в предыдущем примере. По изменению порогов между первым и вторым замерами судили о влиянии коразола на болевое восприятие, а между вторым и третьим - о выраженности аналгезии, вызванной стрессом.
Результаты, отражающие влияние стандартного анксиогена на регистрируемые показатели, представлены в таблицах (где в качестве болевых порогов приведены индивидуальные средние значения, полученные на основании трех измерений).
Длительность иммобилизации в тесте неизбегаемого плавания в холодной воде и изменения порогов болевой чувствительности у мышей после предварительного введения физиологического раствора
Длительность иммобилизации в тесте неизбегаемого плавания в холодной воде и изменения порогов болевой чувствительности у мышей после предварительного введения коразола
Представленные результаты свидетельствуют о том, что между группами животных, не отличавшихся исходно по порогам болевой чувствительности, наблюдалось существенное различие по реагированию на стрессорное воздействие. Предварительное введение коразола, не повлиявшее на болевое восприятие, заметно потенцировало у мышей второй группы повышение порогов переносимости боли (аналгезию, вызванную стрессом) и привело к значимому (Р<0.001) сокращению длительности иммобилизации в тесте принудительного плавания в холодной воде по сравнению с контрольной группой.
Пример 3.
Данная серия экспериментов выполнена в зимний период на 20 мышах-самцах линии NMR I (вес 22-26 г) по схеме, аналогичной представленной в предыдущем примере. Животных поделили на две группы. После определения исходных значений болевых порогов за 30 минут до плавания мышам первой группы вводили физиологический раствор, а второй диазепам в дозе 0,25 мг/кг (внутрибрюшинно). Через 25 минут после введения и непосредственно перед стрессорным воздействием у животных повторно измеряли болевые пороги, после чего их помещали в условия неизбегаемого плавания, а спустя 10 минут после описанной ранее процедуры тестирования вновь приступали к измерению порогов болевой чувствительности. Таким образом, и в течение данного эксперимента у каждого животного уровень болевых порогов определяли трижды. По изменению порогов между первым и вторым замерами судили о влиянии диазепама на болевое восприятие, а между вторым и третьим - о выраженности аналгезии, вызванной стрессом.
Результаты, отражающие влияние стандартного анксиолитика на регистрируемые показатели, представлены в таблице, где приведены средние для групп значения болевых порогов.
Влияние предварительного введения диазепама (0,25 мг/кг) на длительность иммобилизации в тесте неизбегаемого плавания в холодной воде и изменения порогов болевой чувствительности у мышей линии NMRI
Выводы
Аналгезия, вызванная стрессом, связана с повышением порогов переносимости боли, при этом пороги чувствительности не изменяются.
Стратегия поведения животных в условиях неизбегаемого стресса тесно взаимосвязана с анальгезией, развивающейся вследствие этого стресса.
Анксиоген коразол увеличивает выраженность аналгезии, вызванной стрессом, и сокращает продолжительность иммобилизации у мышей в тесте принудительного плавания в холодной воде. Введение анксиолитика диазепама приводит к противоположным эффектам и противодействию коразолу. При введении анксиотропных препаратов изменения регистрируемых показателей (длительности иммобилизации в условиях неизбегаемого стресса и аналгезии, вызванной этим стрессом) носят реципрокный характер.
Данный способ моделирования деперсонализационного расстройства на лабораторных грызунах был апробирован в Научно-исследовательском институте им. В.М.Бехтерева на 50 крысах и более чем на 600 мышах разных линий (SHR, NMRI, С57Bl, BALB). В целом, при точном выполнении методических условий, достигается высокая до 85% воспроизводимость результатов по данному способу. Способ применяется в Институте на отделении клинических и экспериментальных исследований новых психотропных средств для исследования патогенетических механизмов развития аффективных состояний и разработки новых подходов к терапии резистентных состояний.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ДИФФЕРЕНЦИАЛЬНОЙ ДИАГНОСТИКИ ДЕПЕРСОНАЛИЗАЦИИ И ДЕПРЕССИИ | 2002 |
|
RU2211711C1 |
| СПОСОБ ВЫЯВЛЕНИЯ ПСИХОТРОПНОЙ АКТИВНОСТИ ЛЕКАРСТВЕННЫХ И НЕЛЕКАРСТВЕННЫХ ВЕЩЕСТВ | 2012 |
|
RU2506649C1 |
| СПОСОБ МОДЕЛИРОВАНИЯ ПСИХОТИЧЕСКОГО РАССТРОЙСТВА | 2004 |
|
RU2366464C2 |
| 9-[2-(4-ИЗОПРОПИЛФЕНОКСИ)ЭТИЛ]АДЕНИН, ОБЛАДАЮЩИЙ АНТИДЕПРЕССАНТНЫМ И ПРОТИВОСТРЕССОРНЫМ ДЕЙСТВИЕМ | 2013 |
|
RU2529817C1 |
| ФЕНИЛГИДРАЗИД (4-ФЕНИЛ-2-ПИРРОЛИДОН-1-ИЛ)-УКСУСНОЙ КИСЛОТЫ, ОБЛАДАЮЩИЙ НООТРОПНОЙ, АНТИДЕПРЕССИВНОЙ И АНКСИОЛИТИЧЕСКОЙ АКТИВНОСТЬЮ | 2010 |
|
RU2440981C1 |
| СПОСОБ МОДЕЛИРОВАНИЯ ПАРОКСИЗМАЛЬНОГО РАССТРОЙСТВА | 2009 |
|
RU2434306C2 |
| СРЕДСТВО, ОБЛАДАЮЩЕЕ АНТИДЕПРЕССАНТНЫМ, АНКСИОЛИТИЧЕСКИМ, НЕЙРОПРОТЕКТОРНЫМ И ИММУНОСТИМУЛИРУЮЩИМ ДЕЙСТВИЕМ | 2010 |
|
RU2429834C1 |
| СРЕДСТВО, ОБЛАДАЮЩЕЕ АНТИДЕПРЕССИВНЫМ, АНКСИОЛИТИЧЕСКИМ И НООТРОПНЫМ ДЕЙСТВИЕМ | 2010 |
|
RU2437659C1 |
| СПОСОБ ДИАГНОСТИКИ НЕВРОЛОГИЧЕСКИХ БОЛЬНЫХ С ЦЕРЕБРОВАСКУЛЯРНОЙ ПАТОЛОГИЕЙ | 2005 |
|
RU2341190C2 |
| (2,2,3,3,4,4,4,-Гептафторбутил)-N-(диметилметоксисилил)-азиридин, обладающий противосудорожным и транквилизирующим действием | 1991 |
|
SU1781221A1 |
Изобретение относится к медицине, а именно к способам моделирования деперсонализационного расстройства, и может быть использовано при проверке новых психотропных средств. Способ заключается в том, что у грызунов определяют болевые пороги (БП) чувствительности и переносимости (БПЧ и БПП). Затем за 20-30 минут перед стрессорным воздействием проводят инъекцию нейротропного средства. Через 15-25 минут непосредственно перед стрессорным воздействием повторно замеряют БПЧ и БПП и подвергают грызунов неизбегаемому стрессорному воздействию в течение 4-6 минут. При этом регистрируют суммарную продолжительность иммобилизации. Через 2-10 минут после окончания стрессорного воздействия повторно определяют БПЧ и БПП. По характеру сочетания продолжительности иммобилизации и изменений БПЧ и БПП судят о чувствительности грызунов к стрессу и выраженности деперсонализации. По изменению БП между первым и вторым их измерениями судят о влиянии препарата на болевое восприятие, а между вторым и третьим измерениями БП - о выраженности аналгезии, вызванной стрессом. Изобретение позволяет изучать механизмы формирования деперсонализационного расстройства и разрабатывать методы его терапии. 8 табл., 2 ил.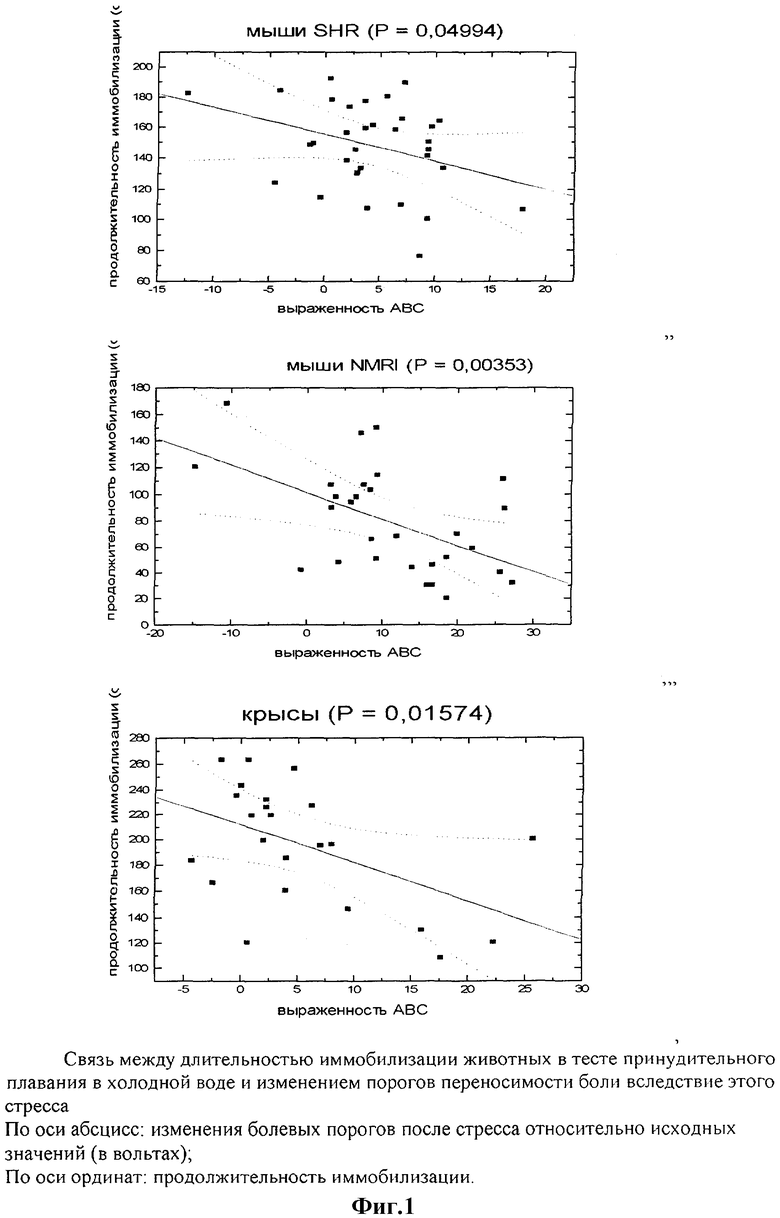
Способ исследования деперсонализационного расстройства на его модели у грызунов, отличающийся тем, что у грызунов определяют болевые пороги (БП) чувствительности и переносимости (БПЧ и БПП), затем за 20-30 мин перед стрессорным воздействием проводят инъекцию нейротропного средства, через 15-25 мин непосредственно перед стрессорным воздействием повторно замеряют БПЧ и БПП, затем подвергают грызунов неизбегаемому стрессорному воздействию в течение 4-6 мин, при этом регистрируют суммарную продолжительность иммобилизации, а через 2-10 мин после окончания стрессорного воздействия повторно определяют БПЧ и БПП, при этом по разнице их измерений судят о выраженности аналгезии, вызванной стрессом, а по характеру сочетания продолжительности иммобилизации и изменений БПЧ и БПП судят о чувствительности грызунов к стрессу и выраженности деперсонализации, причем по изменению БП между первым и вторым их измерениями судят о влиянии препарата на болевое восприятие, а между вторым и третьим измерениями БП - о выраженности аналгезии, вызванной стрессом.
| КЕНУНЕН О.Г | |||
| и др | |||
| Выраженность аналгезии, вызванной стрессом, связана с активностью животных в тесте неизбегаемого плавания | |||
| Журнал высшей нервной деятельности им | |||
| И.П.Павлова | |||
| Перекатываемый затвор для водоемов | 1922 |
|
SU2001A1 |
| СПОСОБ ДИФФЕРЕНЦИАЛЬНОЙ ДИАГНОСТИКИ ДЕПЕРСОНАЛИЗАЦИИ И ДЕПРЕССИИ | 2002 |
|
RU2211711C1 |
| СПОСОБ ЛЕЧЕНИЯ АФФЕКТИВНО-ПСИХОТИЧЕСКИХ РАССТРОЙСТВ | 2000 |
|
RU2166943C1 |
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| МОРОЗ Б.Т | |||
| и др | |||
| Исследование болевой | |||

