Изобретение относится к медицине, а именно к способам моделирования прогредиентного психотического расстройства, и может быть использовано при разработке новых потенциальных антипсихотических средств и уточнения механизмов развития психотических состояний (шизофрения).
Поиск и разработка новых потенциальных антипсихотических средств существенно ограничены отсутствием специфических моделей психотического расстройства. Несмотря на то, что в настоящее время известны экспериментальные методы на животных, позволяющие предполагать потенциальное антипсихотическое действие новых психотропных препаратов, все они не могут рассматриваться как адекватные моделируемой патологии. Прежде всего это связано с тем, что эти модели позволяют оценивать только острые эффекты потенциальных антипсихотиков, поскольку "эквиваленты" психотического расстройства также моделируются в "остром" эксперименте.
Однако, как известно, эндогенная патология, сопровождающаяся появлением психотических симптомов (бред и галлюцинации), редко имеет характер однократного психотического эпизода, а чаще носит хронический прогредиентный тип течения. Связь между активностью центральных дофаминергических процессов и формированием психотических нарушений несомненна [Carlsson A. The current status of the dopamine hypothesis of schizophrenia // Neuropsychopharmacology, 1988, v.1, P.179-203 /1/. Crow T.J. Molecular pathology of schizophrenia: more than one disease process? // British Medical Journal, 1980, v.280, P.66-68 /2/]. Несмотря на трудности моделирования психотических состояний в эксперименте, вышеназванная зависимость подтверждается клиническими данными - антипсихотической активностью средств, снижающих дофаминергическую передачу, именно поэтому поиск новых антипсихотиков проводится в рамках дофаминовой парадигмы развития психотического состояния [Richelson E., Nelson A., Antagonism of neuroleptics of neurotransmitter receptors of normal human brain in vitro // Eur.J.Pharmacol., 1984, v.103, 197-204 /3/].
В соответствии с принятой дофаминовой гипотезой развития психотических нарушений [/3/, Kokkinidis L.,Anisman H. Amphetamine psychosis and schizophrenia: a dual model// Neurosci. Bahav. Res. - 1981. - V.5. - P.449-461 /4/, Snyder S.H. Amphetamine psychosis: A "model" schizophrenia mediated bycatecholamines// Amer. J. Psychiatr. - 1973. - v.130. - P.61-67 /5/] в поведенческих тестах на животных оценивается влияние новых веществ на поведенческие феномены, инициируемые препаратами, действующими на дофаминергическую передачу: галаперидоловая и трифтазиновая каталепсия, фенаминовая стереотипия на крысах и групповая токсичность на мышах, изменение поведения, вызванное апоморфином, резерпином и т.п. Как отмечалось, тесты, связанные с влиянием препаратов на галоперидоловую и трифтазиновую каталепсию, являются обязательными в "батарее тестов" изучения новых нейролептиков [Раевский К.С., Наркевич В.Б. Экспериментальное доклиническое изучение специфической активности новых соединений, предлагаемых для клинических испытаний в качестве нейролептиков (антипсихотических средств) // Методические рекомендации МЗ РФ. - Москва, 1998 /6/].
К настоящему времени достигнуты определенные успехи и в понимании патогенеза психотического расстройства, сопровождающегося появлением патологических симптомов, и в создании новых атипичных антипсихотиков [Blin О. A comparative review of new antipsychotics // Canadian Journal of Psychiatry. - 1999. - Vol.44. - P.235-244. /7/]. Абсолютно установлено, что к развитию патологической психопродукции причастны дофаминергическая, серотонинергическая и глутаматергическая медиаторные системы мозга. Большинство из известных средств антипсихотического спектра действия прямо или косвенно изменяют активность прежде всего дофаминергической передачи. С функциональным изменением ее активности на уровне мезолимбической и мезокортикальной части связывают развитие лечебного, собственно антипсихотического действия, а появление побочных эффектов связывают с нарушением функционирования нигростриатной и тубероинфундибулярной дофаминергической передачи [Stahl, Stephen M. // Essential psychopharmacology: Neurosci basis a clinical application.-Cambridge Univ. Press 1996. - P.379 /8/].
Если рассматривать действие антипсихотиков в соответствии с дофаминовой гипотезой формирования психоза, то можно говорить об их вторичном "нормализующем" влиянии на патологическую нейрохимию "больного мозга", на что указывают следующие факты [Кроу Т.Дж., Джонстоун Е.С. Дофаминергические процессы при шизофрении и механизм антипсихотического действия // Нейротрансмиттерные системы / Под ред. Дж. Легга. - M.: Медицина, 1982. - С.202-214 /9/]:
не выявлено связи между антипсихотической активностью препаратов и изменением уровня пролактина в плазме крови;
нет прямой корреляции между выраженностью экстрапирамидной симптоматики и снижением симптомов психоза;
не обнаружено временной зависимости в развитии антипсихотического эффекта в связи с изменениями фармакокинетических характеристик нейролептических средств.
Любопытно, что эти вопросы остаются до настоящего времени не разрешенными. На наш взгляд во многом это связано с тем, что проведение исследований ограничено отсутствием адекватных поведенческих моделей психотического расстройства на животных. Основное эндогенное психотическое расстройство - шизофрения, как и любое другое хроническое заболевание, течет на протяжении многих лет волнообразно: эпизоды обострения психоза сменяются фазами ремиссии. Современные модели заболевания не учитывают подобного течения болезни. Нам представляется, что подобное состояние может быть воспроизведено в эксперименте на животных путем проведения многократных чередующихся введений миметиков и литиков дофаминергической передачи. Оценка результатов подобной манипуляции может оцениваться в изучении поведения животных, подвергнутых описанной процедуре.
Поскольку любые проявления высших функций, так или иначе, связаны с двигательными реакциями, то по изменению качества и силы последних можно косвенно судить о развитии патологического состояния, а т.к. контроль моторных реакций во многом осуществляется при участии ДА-ергической передачи, то говорить о функциональном состоянии этого нейрохимического звена можно по изменению реакции развития каталепсии.
Следует заметить, что сохранение искусственно приданной животному позы (каталепсия) имеет определенные онто- и филогенетические корни и сам феномен встречается у большинства млекопитающих в периоде раннего онтогенеза. Кроме того, следует заметить, что "застывания" и временное обездвиживание также является достаточно хорошо известным феноменом и отмечается при развитии "Павловской" реакции - "что такое?". В контексте проблемы - использования каталепсии для поиска и оценки эффективности веществ, влияющих на дофаминергические системы мозга, эти факты имеют важное значение, поскольку связаны с глубинными, генетически детерминированными поведенческими реакциями. Кроме того, касаясь другого важного аспекта - хронического течения психотических расстройств, следует заметить, что последующие психотические эпизоды, развивающиеся с определенной частотой и тяжестью, приводят, в конечном счете, к развитию дефицитарных состояний, оцениваемых в рамках шизофрении сочетанием первичных негативных симптомов. Способы моделирования подобных состояний нам не известны, а при их отсутствии нельзя говорить об адекватности этому заболеванию применяемых моделей. В то же время, хорошо известно, что препараты, стимулирующие дофаминовые рецепторы, вызывают развитие нейродегенеративных повреждений [Kelley A.E., Delfs J.M., Chu В. // Neurotoxicity induced by the D-1 agonist SKF 38393 following microinjection into rat brain. - Brain Res. - 1990. - v.532. - N 1-2. - P.342-346 /10/], а подобное использование агонистов дофамина наркоманами сопровождается специфическими расстройствами личности, подобными тем, что описываются в рамках появления негативной симптоматики. Наиболее частыми симптомами являются эмоциональное безразличие, психическая и моторная заторможенность, отсутствие инициативы и т.п. Представляется, что наиболее достоверно подобные симптомы в эксперименте на животных могут быть выявлены в моделях оценки социального поведения, в условиях спровоцированной конфликтной ситуации.
Наиболее удачной в рамках моделируемого состояния может быть применение метода оценки спровоцированной током агрессии в паре мышей.
Наиболее близким по технической сущности и достигаемому результату является способ моделирования психотического расстройства путем инъекции фенамина мышам [Utena Н. // On relapse-liability; schizophrenia. Amphetamine psychosis and animal model. In: Schizophrenia and Schizophrenia-Like Psychosis, Ed. by H. Mitsuda and Т. Fukada, Stuttgard: Georg Thieme Verlag. Tokyo: Igaku Shoin Ltd., 1975, pp.285-287 /11/]. Недостатками известного способа является то, что в условиях острого введения фенамина моделируется состояние интоксикации, а при повторных инъекциях формирование наркоманической зависимости, но не прогредиентного психотического расстройства.
Технический результат, на достижение которого направлено настоящее изобретение, заключается в разработке модели прогредиентно текущего психотического расстройства, которое позволяет моделировать именно хронически прогредиентный тип течения заболевания, приводящий к развитию поведенческих нарушений, связанных с эмоциональной отгороженностью (эмоционально-волевые расстройства).
Указанный технический результат достигается тем, что в способе моделирования психотического расстройства, включающем инъекции фенамина мышам, согласно изобретению, учитывают прогредиентный тип течения психотических состояний, путем проведения поочередных, через день, инъекций фенамина и галоперидола, всего по шесть инъекций каждого препарата, мышам SHR в течение 12 дней, по шесть инъекций каждого препарата:
фенамин - 10 мг/кг (внутрибрюшинно, однократно в сутки утром в 10:00);
галоперидол - 5 мг/кг (внутрибрюшинно, однократно в сутки утром в 10:00) с последующей оценкой продолжительности феномена спонтанной каталепсии через 2 дня после завершения периода инъекций, при этом оценку полученных результатов проводят дважды, при первом измерении устанавливается базовый уровень продолжительности спонтанной каталепсии, отражающий тяжесть вызванных нарушений, а при повторном тестировании, через 2 дня, а также спустя 12 дней после первого измерения этого феномена выявляют стойкость нарушений, компенсаторные возможности работы мозга и определяют динамику изменений состояния в сравнении с базовым уровнем, а через 7 дней после окончания введений фенамина и галоперидола проводят оценку агрессивного поведения методом электрокожного раздражения пары мышей.
Способ выполняют следующим образом:
Всех мышей, необходимых для моделирования патологии, предварительно поделят на 3 группы в соответствии с режимом введения препаратов:
1. Получающие через день физиологический раствор и галоперидол в дозе 5 мг/кг.
2. Получающие через день галоперидол 5 мг/кг и фенамин 10 мг/кг (1-й день галоперидол, 2-й день фенамин, 3-й день галоперидол, 4-й фенамин и т. д.).
3. Получающие в/бр физиологический раствор каждый день.
Необходимые для контроля эксперименты на группе животных, получающих один фенамин, были исключены из работы ввиду того, что при хроническом введении этого препарата наблюдается рост толерантности, развитие зависимости, т.е. формирование симптомов аддитивного поведения [Kramer J.C., Fischmannn V.S., Littlefield D.C. // Amphetamine Abuse, J. Amer. Med. Ass. (JAMA), 1967, v.197, pp.305-309 /12/; Kuhn G.M., Schanberg S.N. // Distribution and metabolism of amphetamine in tolerant animals. - 1977. - In: Cocaine and other stimulants, New-York-London, 1977. - P.161-177 /13/], а при остром однократном введении формирования симптомов интоксикации дофаминомиметиками [Utena Н. // Behavioral aberations in metamphetamine-intoxicated animals and chemical correlates in the brain. In: Progress in Brain Research, Vol.21B, edited by T.Tokizane and J.A.Schade, Amsterdame: Elsevier Publishing Company, 1966, pp.192-207 /14/].
В работе применялись следующие препараты: галоперидол (А.О.Гедеон Рихтер, Венгрия), сернокислый D,L-фенамин (РФ). Первая и третья группы животных были использованы в качестве контрольных. Режим введения галоперидола в первой группе мышей был выбран исходя из данных по фармакокинетике. Известно, что спустя сутки галоперидол уже не определяется в мозге крыс [Cohen B.M., Herschel M., Miller E., Maybery Н. // Radioreceptor assay of haloperidol tissue levels in the rat.- Neuropharmacol.-1980. - v.15. - N 5. - P.663-669 /15/], мы предполагаем, что и мышам свойственна подобная кинетика этого препарата. Физиологический раствор, а также растворы фенамина и галоперидола вводили из расчета 0,1 мл на 10 г веса.
У всех животных перед началом введений препаратов и спустя два дня после окончания инъекций измеряли вес. Спустя 14 дней (срок, в течение которого всем подопытным животным вводили препараты) у мышей оценивали выраженность галоперидоловой и "спонтанной" каталепсии. В одной из групп мышей выраженность каталепсии оценивали при каждом введении галоперидола, всего в семи измерениях.
В остальных группах мышей каталепсию измеряли через 2, 4 и 14 дней после окончания введения препаратов, т.е. оценивали «спонтанную каталепсию», а не каталепсию, вызванную нейролептиком. Для оценки поведения животных использовался тест каталепсии, предложенный К.С.Раевским [6] в следующей модификации: передние лапы животного помещали на горизонтальную проволочную перекладину, расположенную на высоте 4 см, и определяли время удержания данной позы («поза лектора») в течение всего времени наблюдения (5 мин). Затем подсчитывали среднее время сохранения позы для группы животных.
В те же сроки оценивали двигательную активность методом «открытого поля», измеряя количество локомоций и стоек у мышей в течение 1 мин наблюдения. Поведение животных регистрировали зрительно в емкости круглой формы диаметром 50 см, имеющей стандартную диаметрально-круговую разметку.
Оценку агрессивного поведения мышей проводили в паре животных, помещаемых в камеру, имеющую токопроводящий пол. Эксперимент начинали с измерения порогов болевой чувствительности, который фиксировали индивидуально для каждого животного (в вольтах) по первому вздрагиванию (порог чувствительности), а затем драк между животными, развивающимися при увеличении напряжения постоянного электрического тока, подаваемого на лапы через токопроводящий пол. Непосредственно перед помещением животных на токопроводящий пол все лапы его смачивались физиологическим раствором. Болевое раздражение (прямоугольные импульсы постоянного тока 5 Гц, до 50 В) продолжительностью 0,2 с наносили начиная с 1 В, с постепенным увеличением напряжения (1 В в секунду) до регистрации "ложной" драки. В этих условиях у животных возникают вертикальные стойки, атаки с укусами и ударами передними лапами. Регистрация напряжения тока, поданного на лапы мышей и спровоцировавшего драку в паре контрольных или подопытных животных, была объективным критерием, отражающим агрессивность [Blanchard C.D., Blanchard R.J. // Inadequacy of pain-agression hypothesis revealed in naturalistic setting. - Aggres. Behav. 1984. Vol.10, P.33-46 /16/.
Результаты обрабатывали на PC, используя программные продукты «Excel-97» и «Statistica 5,0», подсчитывая M±m, достоверность различий между группами определяли по непараметрическому критерию "U" Вилкоксона-Манна-Уитни.
Результаты исследования
Внутрибрюшинное однократное введение галоперидола в дозе 5 мг/кг вызывало у мышей развитие каталепсии продолжительностью более 7 часов. Причем, спустя один час после введения нейролептика каталепсия практически достигала максимальных значений, оставаясь таковой (250-280 с за 5 мин интервал регистрации) в течение последующего наблюдения за животными.
Повторные введения галоперидола через 48 часов показали, что средняя продолжительность каталепсии за 5 мин наблюдения при первом опыте составляет 247,9 с, а при шестом - 292,9 с (коэффициент корреляции по Пирсону: r=0,85 при Р<0,05).
Интересен тот факт, что введение физиологического раствора также сопровождалось развитием каталепсии (вероятно наличие этого феномена не связано с введением физиологического раствора). Развитие спонтанной каталепсии наблюдали, регистрируя показатели в течение 3,5 часов наблюдения. Выраженность каталептогенной реакции к концу регистрации увеличивалась почти в 20 раз, составляя 1,5 с при первом измерении и 41,7 с через 3,5 часа на восьмом измерении. Как и в предыдущем случае, повторные измерения каталепсии после введения физиологического раствора показали увеличение времени застывания мышей в позе "лектора" (коэффициент корреляции по Пирсону был r=0,97, при Р<0,001).
Итак, поведенческий феномен в виде застывания мышей в позе "лектора" воспроизводится как после введения галоперидола, так и после введения физиологического раствора, однако, продолжительность этого феномена при введении галоперидола через 3 часа составляет около 300 с, а после введения физиологического раствора через тот же временной интервал около 40 с. Является ли этот феномен только рефлексом на обстановку или отражает еще конституциональный "неврологический" статус, было изучено при исследовании корреляционной зависимости исходя из времени измерения каталепсии у одних и тех же животных при введении галоперидола и после введения физиологического раствора. Было выявлено, что такая зависимость существует, значение коэффициента корреляции составило - r=0,29 (n=54; P<0,04), что позволяет рассматривать выраженность феномена "спонтанной каталепсии", как результат, отражающий функциональное состояние дофаминергической передачи в нигростриатной системе.
Результаты этой части работы свидетельствуют также о том, что использование теста каталепсии на мышах в условиях проведения повторных измерений может иметь определенную погрешность, с которой необходимо считаться, если изучаются физиологические механизмы развития самого феномена каталепсии. Естественно это требует более жестких контролирующих мероприятий в случае повторного воспроизведения эксперимента. Если же феномен применяется для отбора активных препаратов, то при наличии соответствующих "слепых контрольных" групп ошибкой, вносимой "спонтанной каталепсией", можно пренебречь ввиду того, что "галоперидоловый радикал" механизма развития каталептогенной реакции намного превышает "физиологическую составляющую" этого феномена.
Поскольку выраженность "спонтанной каталепсии" отражает состояние дофаминергических процессов в нигростриатной системе, но действие как фенамина, так и галоперидола диффузно в отношении всех дофаминергических систем мозга, то можно полагать, что подобные изменения будут определяться и на уровне тубероинфундибулярной, мезолимбической и мезокортикальной систем. Поскольку повышение активности дофаминергических процессов на уровне мезокортикальной и мезолимбической систем связывают с развитием соответственно негативных и позитивных симптомов при психотическом расстройстве, то отражение их функциональной активности на уровне феномена "спонтанной каталепсии" позволяет оценивать "психотическую настроенность" мозга.
Итак, результаты, отражающие выраженность «спонтанной каталепсии» спустя 2, 4 и 14 дней после окончания поочередных введений фенамина 10 мг/кг и галоперидола 5 мг/кг животным в сравнении с контрольными группами мышей (I и III группы), показали, что результаты повторных измерений продолжительности "спонтанной каталепсии" через 2, 4 и через 14 дней воспроизводятся (Таблица 1). Поскольку воспроизводимость выявленных различий определяется спустя 2 недели после окончания введений препаратов - вторая группа мышей значимо отличалась от первой (вводился галоперидол 5 мг/кг через день с физиологическим раствором) и третьей (физиологический раствор), то результаты этого фрагмента исследования показывают то, что «спонтанная каталепсия» в группах мышей, получавших галоперидол и фенамин, выражена существенно слабее, чем в группах мышей, получавших инъекции физиологического раствора или инъекции галоперидола в дозе 5 мг/кг через день с физиологическим раствором.
Таким образом, показано, что способность мышей к застыванию в "позе лектора" наблюдается и при отсутствии введения галоперидола, однако, продолжительность удержания позы существенно короче, чем при введении нейролептика. Наличие положительной корреляционной связи по показателям спонтанной и спровоцированной введением галоперидола каталепсии свидетельствует о наличии общих механизмов, участвующих, как в реализации нейролептической, так и "физиологической" каталептогенной реакции. Вероятно, важное значение в развитии каталепсии имеет кратность помещения животных в опытную ситуацию. Об этом свидетельствуют результаты повторных измерений каталепсии при введении галоперидола. Мы не выявили того, что продолжительность пребывания животных в спонтанной каталепсии как-то связана с двигательной активностью, значения для показателя, отражающего связь каталепсии/локомоции: r=-0,04 (n=30; Р>0,05), а для каталепсии/стойки - r=-0,33 (n=30; Р>0,05). Эти данные свидетельствуют о том, что механизмы формирования двигательного локомоторного поведения не связаны с двигательным каталептогенным поведением. В то же время, развитие каталепсии, как вызванной галоперидолом, так и "спонтанной" имеет общие механизмы развития, которые связаны с активностью дофаминергической передачи (см. выше). Поочередные инъекции фенамина с галоперидолом в течение двух недель приводили к уменьшению продолжительности "спонтанной каталепсии", чего не наблюдалось в группах животных, получавших физиологический раствор и галоперидол.
Представленные данные свидетельствуют о том, что регистрируемые изменения поведения, вызванные поочередными инъекциями галоперидола и фенамина, являются достаточно стойкими и сохраняются, по крайней мере, в течение двух недель. Мы предполагаем, что подобные изменения поведения могут быть вызваны нарушением механизмов нормальной деятельности не только катехоламинергических структур мозга. Поскольку предпринятая процедура не просто стимулирует или тормозит соответствующую медиаторную систему, а вызывает в ее активности периодические полярные колебания, которые могут оказывать дизрегулирующее действие не только на эту систему, но и на вторично связанные с ней нейрохимические процессы. Следует отметить, что мы не выявили существенных нарушений в каталептогенном поведении у мышей 1-й группы, которым вводили только галоперидол. Это может свидетельствовать о том, что функциональное состояние катехоламинергических структур мозга при введении одного нейролептика сильно не пострадало. Известно, что при хроническом введении галоперидола и других нейролептиков развивается повышение адренергической медиации [Schelkunov E.L. // Adrenergic effect of chronic administration ofneuroleptics.-Nature. - 1967. - v.214. - N 5094. - P.1210-1212 /17/, Щелкунов Е.Л. // Повышение тонуса центральных адренергических структур (высших симпатических центров) при хроническом введении нейролептиков.-В сб. "Психофармакология и лечение нервных и психических заболеваний". Л., 1964. - С.23-25 /18/, Минеева М.Ф.// Механизм действия нейролептиков.- Итоги науки и техники, Молекулярные механизмы действия психотропных препаратов, 1987. - т.15. - с.170-228 /19/], что могло отразиться на результатах наших экспериментов. Однако результаты по группе мышей, которым вводили только галоперидол, не подтвердили данного факта. Скорее всего, повторные введения галоперидола через день не успевают за двухнедельный промежуток "раскачать" соответствующую нейрохимическую систему так, чтобы стойкие нарушения определялись в тесте на каталепсию. Однако приходящее уменьшение спонтанной каталепсии было отмечено нами спустя двое суток после последней инъекции препарата, но уже на 4-е сутки достоверных различий не определялось.
Известно, что хроническое введение фенамина вызывает обратный эффект, эффект торможения активности адренергической передачи мозга [Seiden L.S., Fishman M.W., Schuster C.R. // Long-term amphetamine-induced changes in brain catecholamines in tolerant rhesus monkeys. - Drug Alcoh. Dependens, 1976. - V.1. - P.215-219 /20/]. Вероятно разнонаправленные колебания нейрохимических систем в течение 2-х недельного периода выполнения эксперимента вызывают нарушения формирования каталептогенного поведения у мышей.
Любопытно, что животные, получавшие через день только галоперидол в дозе 5 мг/кг на протяжении 14 дней, отличались по весу от контрольных мышей и мышей 2-й и 3-й групп. Прибавка массы тела за 14 дней составила для этих мышей соответственно -2,39±0,67 г (Р<0,01); а для остальных 1,47±0,94 г (Р>0,05); 1,38±0.70 г (Р>0.05) и 1,35±0,88 г (Р>0,05). Это вполне согласуется с эффектами нейролептиков, наблюдаемыми на практике.
Таким образом результаты этих экспериментов свидетельствуют о том, что чередование введений агониста/антагониста дофамина приводит к нарушению механизмов функционирования катехоламинергических процессов. Об этом свидетельствуют изменения каталептогенного поведения животных.
Мы полагаем, что дестабилизирующее влияние полярных лекарственных воздействий на катехоламинергические системы мозга и связанное с ними нарушение каталептогенного поведения можно рассматривать как причину изменений в эмоционально-мотивационной сфере, которая, как известно, страдает у больных с психотическими расстройствами. Последняя наиболее ярко проявляется в заключительной стадии течения болезни, когда на первый план выступают негативные симптомы, отражающие наступление эмоционально-волевого дефекта. Выявленное нами снижение агрессивного поведения мышей при электростимуляции, развившееся три недели спустя после начала введения препаратов и через неделю после окончания инъекций, может указывать на то, что у животных формируется эквивалентное состояние эмоциональных расстройств.
Оценка агрессивности, проведенная через 1 неделю после окончания введений, показала также существенные различия между группами животных (Таблица 2). При оценке агрессивного поведения было не обнаружено изменений порога болевой чувствительности между контрольными и подопытными мышами. Однако выраженные различия между группами были выявлены по уровню напряжения тока, необходимого для провокации "ложной" драки (Таблица 2).
Таким образом, данные таблицы указывают на то, что уровень эмоционального реагирования на болевую провокацию значительно снижен у подопытных мышей, получавших галоперидол поочередно с фенамином.
В связи с полученными результатами, нам бы хотелось обратить внимание на то, что периодическое повышение и последующая блокада дофаминергической активности может иметь некоторые клинические аналогии. К примеру хорошо известно, что один из наиболее тяжелых эндогенных психозов - шизофрения чаще всего течет периодично.
Даже при отсутствии нейролептической терапии, если следовать дофаминовой гипотезе развития психоза, можно предполагать, что вслед за периодом обострения должна наступить фаза функционального истощения этой медиаторной системы, которая скорее всего проявится ремиссией болезни. Затем вновь, после паузы следует период повышения активности и, соответственно, обострение болезни. Как известно, шизофрения это прогредиентное заболевание, каждая фаза обострения оставляет после себя след, который отпечатывается на исходе, т.е. развитии заключительной части болезни - дефицитарной, когда прежде всего на первый план в клинической картине выступают симптомы эмоционально-волевого дефекта. В развитии эмоционального реагирования, так же как и в формировании мотивированных поступков важная роль принадлежит дофаминергической системе. Можно предполагать то, что со временем приходит стойкая недостаточность дофаминовой медиации. Сейчас при описании патогенеза так называемой негативной клинической симптоматики шизофрении говорят о "выгорании нейронов", речь идет о том, что избыточная возбуждающая стимуляция нейронов может привести к их нейродегенерации [8]. Понятно, что в подобной ситуации эффективность и традиционных нейролептических препаратов будет обратно пропорциональна выраженности нейродегенеративных изменений и симптоматики дефицитарного расстройства.
При лечении типичными (преимущественные блокаторы D2 дофаминовых рецепторов) нейролептиками описанная картина может принимать еще более четкие формы. В то же время, появление симптомов эмоционально-волевого дефекта может отодвигаться во времени, поскольку нейролептики сами защищают нейроны от избыточной дофаминергической стимуляции, но этот эффект может маскироваться влянием препаратов, поскольку их прямое действие заключается в торможении дофаминовой передачи. Однако, несмотря на блокаду дофаминовых рецепторов, нейролептики оказывают стимулирующее влияние на нейрохимические звенья адренергической системы, и потому, вслед за блокадой рецепторов следует фаза их повышенной чувствительности. Вновь наблюдается функциональное повышение дофаминергической активности, со всеми описанными выше событиями. Таким образом формируется порочный круг и разрыв его наступает с развитием нейродегенеративных изменений дофаминчувствительных нейронов, но к сожалению эта та фаза заболевания, которая уже не может быть подвергнута никакой фармакологической коррекции.
Сущность способа поясняется примерами.
Пример 1.
Опыты данной серии выполнены в весенний период на 22 мышах самцах SHR (20-26 г), полученных из питомника "Рапполово", которых предварительно поделили на группы:
1. Получающие через день попеременно: галоперидол 5 мг/кг и фенамин 10 мг/кг (1-й день галоперидол, 2-й день фенамин, 3-й день галоперидол, 4-й фенамин и т.д.). Всего по 6 инъекций каждого препарата при повторной инъекции через 48 часов после предыдущей.
2. Получающие в/бр физиологический раствор каждый день.
В работе применялись следующие препараты: галоперидол (А.О.Гедеон Рихтер, Венгрия), сернокислый D,L-фенамин (РФ).
Физиологический раствор, а также растворы фенамина и галоперидола вводили утром в 10 часов из расчета 0,1 мл на 10 г веса.
Ниже представлена схема постановки эксперимента в графическом представлении.
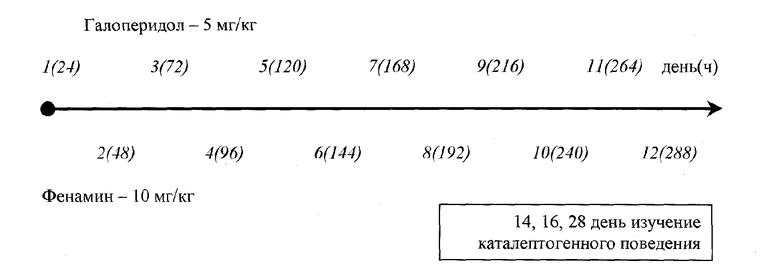
Спустя 14 дней от начала эксперимента у мышей оценивали выраженность "спонтанной" каталепсии, а не каталепсию, вызванную нейролептиком. Для оценки поведения животных использовали тест каталепсии, предложенный К.С.Раевским в следующей модификации: передние лапы животного помещали на горизонтальную проволочную перекладину, расположенную на высоте 4 см, и определяли время удержания данной позы («поза лектора») в течение всего времени наблюдения (5 мин).
Результаты обрабатывали на PC, используя программные продукты «Excel-97» и «Statistica 5,0», подсчитывая M±m, достоверность различий между группами определяли по непараметрическому критерию "U" Вилкоксона-Манна-Уитни.
Результаты проведенного эксперимента представлены в таблицах 3, 4.
Полученные результаты свидетельствуют о том, что между группами, представленными в данном примере, имеются достоверные отличия по критерию "продолжительность каталепсии" (Критерий "U", P<0,02; ANOVA тест LSD, Tukey HSD и др. Р<0,05).
Пример 2.
Данная серии экспериментов выполнена на 20 мышах-самцах линии SHR. Животных, как и в примере 1, поделили на две группы, которым соответственно по той же схеме вводили либо физиологический раствор (контрольная), либо поочередно фенамин и галоперидол в тех же дозах.
Спустя неделю после окончания введений у животных оценивали формирование агрессивного поведения при электрокожном раздражении, через токопроводящий пол. Выраженность агрессии оценивали в паре животных. Эксперимент начинали с измерения порогов болевой чувствительности, которые фиксировали индивидуально для каждого животного (в вольтах) по первому вздрагиванию (порог чувствительности). Порог развития спровоцированной агрессии оценивали при увеличении напряжения постоянного электрического тока до развития драки между мышами. Непосредственно перед помещением животных на токопроводящий пол лапы смачивались физиологическим раствором. Болевое раздражение (прямоугольные импульсы постоянного тока 5 Гц, до 50 В) продолжительностью 0,2 с наносили начиная с 1 В, с постепенным увеличением напряжения (1 В в секунду) до регистрации "ложной" драки (взаимные атаки и удары передними лапами). Регистрация напряжения тока, спровоцировавшего драку, служило критерием оценки агрессивности.
Результаты, отражающие регистрируемые показатели, представлены в таблице 5.
Представленные результаты свидетельствуют о том, что между группами животных наблюдалось незначительное изменение порогов болевой чувствительности и достоверные изменения (Критерий "U", Р<0,02; ANOVA тест LSD, Tukey HSD и др. Р<0,05) по показателю порог агрессивности (напряжение тока, необходимое для развития драки). Мышам, которым вводили галоперидол с фенамином, была необходима большая электростимуляция для развития "ложных" драк.
Выводы:
Повторные, чередующиеся в течение 12 дней инъекции фенамина и галоперидола, вызывают у мышей SHR нарушение формирования феномена "спотанной каталепсии".
Повторные, чередующиеся в течение 12 дней инъекции фенамина и галоперидола снижают спровоцированную электрическим током агрессию у мышей SHR.
Данный способ моделирования психотического расстройства на мышах был апробирован в научно-исследовательском институте им. В.М.Бехтерева, более чем на 600 животных линии SHR. В целом, при точном выполнении методических условий, достигается высокая до 80% воспроизводимость результатов по данному способу. Способ применяется в институте на отделении клинических и экспериментальных исследований новых психотропных средств для уточнения патогенетических звеньев "психотического состояния" и выявления различий в антипсихотическом и побочном механизме действия типичных и атипичных антипсихотиков.
| название | год | авторы | номер документа |
|---|---|---|---|
| Адамантильные производные бензимидазола, обладающие антипаркинсонической активностью | 2023 |
|
RU2820315C1 |
| СРЕДСТВО ДЛЯ ПРЕДУПРЕЖДЕНИЯ И КОРРЕКЦИИ ДВИГАТЕЛЬНЫХ, ЭКСТРАПИРАМИДНЫХ НАРУШЕНИЙ, ВЫЗВАННЫХ НЕЙРОЛЕПТИКАМИ | 2009 |
|
RU2421222C2 |
| СПОСОБ ЛЕЧЕНИЯ СИНДРОМА ПАРКИНСОНИЗМА | 1994 |
|
RU2057529C1 |
| Диметоксибензилсульфопроизводное хиназолин-4(3Н)-она, обладающее анальгезирующей, противопаркинсонической, анксиолитической, психостимулирующей активностью | 2021 |
|
RU2758333C1 |
| АНТИДЕПРЕССАНТНОЕ СРЕДСТВО ДЛЯ КОРРЕКЦИИ НЕЖЕЛАТЕЛЬНЫХ ПОБОЧНЫХ ЭФФЕКТОВ НЕЙРОЛЕПТИКОВ | 2019 |
|
RU2746012C2 |
| СРЕДСТВО, ОБЛАДАЮЩЕЕ СВОЙСТВАМИ НЕЙРОЛЕПТИКА | 1992 |
|
RU2042355C1 |
| НОВЫЕ ТЕРАПЕВТИЧЕСКИЕ КОМБИНАЦИИ МИРТАЗАПИНА И АНТИПСИХОТИЧЕСКИХ СРЕДСТВ ДЛЯ ЛЕЧЕНИЯ ИЛИ ПРОФИЛАКТИКИ ПСИХОТИЧЕСКИХ РАССТРОЙСТВ | 1998 |
|
RU2222330C2 |
| АНТИПСИХОТИЧЕСКОЕ СРЕДСТВО И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2013 |
|
RU2519761C1 |
| СРЕДСТВО, ОБЛАДАЮЩЕЕ АНТИПСИХОТИЧЕСКОЙ АКТИВНОСТЬЮ | 2008 |
|
RU2411248C2 |
| ФАРМАЦЕВТИЧЕСКИЙ СОСТАВ ПРОЛОНГИРОВАННОГО ДЕЙСТВИЯ НА ОСНОВЕ КЛОЗАПИНА ПЕРОРАЛЬНОГО ВВЕДЕНИЯ | 2009 |
|
RU2414903C1 |
Изобретение относится к экспериментальной медицине. Способ включает проведение в течение 12 дней внутрибрюшинных инъекций лекарственных препаратов мышам. Проводят поочередные, через день инъекции фенамина и галоперидола мышам линии SHR, по шесть инъекций каждого препарата. Фенамин вводят по 10 мг/кг однократно в сутки, утром в 10.00. Галоперидол вводят по 5 мг/кг однократно в сутки, утром в 10.00. Способ позволяет моделировать хронически прогредиентный тип течения заболевания. 5 табл.
Способ моделирования прогредиентно текущего психотического расстройства, включающий инъекции фенамина мышам, отличающийся тем, что проводят поочередные через день инъекции фенамина и галоперидола мышам линии SHR в течение 12 дней, по шесть инъекций каждого препарата: фенамин 10 мг/кг внутрибрюшинно, однократно в сутки, утром в 10.00; галоперидол 5 мг/кг внутрибрюшинно, однократно в сутки, утром в 10.00.
| Способ моделирования психоза | 1985 |
|
SU1265831A1 |
| Способ моделирования психомоторных пароксизмов | 1990 |
|
SU1814083A1 |
| Способ моделирования психомоторных пароксизмов | 1988 |
|
SU1562953A1 |

