Область техники
Настоящее изобретение относится к выделенным полипептидам, обладающим протеазной активностью, и выделенным последовательностям нуклеиновых кислот, кодирующим указанные полипептиды. Данное изобретение относится также к конструкциям на основе нуклеиновых кислот, векторам и клеткам-хозяевам, содержащим указанные последовательности нуклеиновых кислот, а также к способам получения и применения указанных полипептидов.
Предпосылки изобретения
Клонирование гена пенициллолизина (plnC) из штамма Penicillium citrinum описано в статье Matsumoto et al., in Biochim. Biophys. Acta 1218: 469 (1994). Последовательность была включена в базу данных EMBL под номером D25535.
В заявке WO 97/46689 описаны нижеследующие (кодирующие) последовательности протеазы, выделенные из специфических штаммов Aspergillus:
SEQ ID No.1 в заявке WO 97/46689 представляет неполный ген рерН Aspergillus niger;
SEQ ID No.2 в заявке WO 97/46689 представляет неполную кДНК рерН Aspergillus niger;
SEQ ID No.3 в заявке WO 97/46689 представляет неполную аминокислотную последовательность РЕРН Aspergillus niger;
SEQ ID No.4 в заявке WO 97/46689 представляет ген pepI Aspergillus nidulans;
SEQ ID No.5 в заявке WO 97/46689 представляет кДНК pepI Aspergillus nidulans; и
SEQ ID No.6 в заявке WO 97/46689 представляет аминокислотную последовательность PEPI Aspergillus nidulans.
В статье Ramesh et al., Gene 165(1):121-125 (1995), описано клонирование и исследование генов, кодирующих протеазы из штаммов Aspergillus flavus и Aspergillus fumigatus. Указанные последовательности включены в базу данных EMBL соответственно под номерами L7524 и U24146.
Последовательность вариантной нейтральной протеазы, созданной на основе нейтральной протеазы II из штамма Aspergillus oryzae, описана в патенте Японии 05-168479. Клонирование родительской нейтральной протеазы II Aspergillus oryzae описано в статье Hiroki Tatsumi et al., Mol. Gen. Genet. (1991), 228, p. 97-103.
Зрелые пептидные части вышеуказанных протеаз идентичны зрелой пептидной части протеазы Thermoascus по настоящему изобретению на 75,7% или меньше.
Нуклеотидные последовательности, соответствующие зрелым пептидным частям вышеуказанных протеаз, идентичны нуклеотидной последовательности, соответствующей зрелой пептидной части протеазы Thermoascus по настоящему изобретению, на 68,4% или меньше.
Целью настоящего изобретения является получение альтернативных протеаз, предназначенных для использования в корме для животных.
Краткое изложение существа изобретения
Настоящее изобретение относится к выделенным полипептидам, обладающим протеазной активностью, которые выбраны из группы, состоящей из:
(а) полипептида, содержащего аминокислотную последовательность, которая по крайней мере на 80% идентична аминокислотам -178-177, -159-177 или +1-177 SEQ ID No.2;
(b) полипептида, кодируемого последовательностью нуклеиновой кислоты, которая гибридизирует в условиях низкой (степени) строгости (жесткости) с:
(i) зрелой протеазой, кодирующей часть плазмиды, находящейся в штамме DSM 14652 Escherichia coli;
(ii) нуклеотидами 25-1089, 1-1089, 1-1344, 25-1344, 559-1344 или 559-1089 SEQ ID No.1;
(iii) субпоследовательностью (i) или (ii), содержащей по крайней мере 100 нуклеотидов, или
(iv) комплементарной цепью последовательности (i), (ii) или (iii);
(с) варианта полипептида, содержащего аминокислотную последовательность, включающую аминокислоты -178-177, -159-177 или +1-177 SEQ ID No.2, включающего замены, делеции и/или инсерции одной или более аминокислот;
(d) аллельного варианта полипептида (а) или (b);
(е) фрагмента полипептида (а), (b) или (d), обладающего протеазной активностью.
SEQ ID No.1 представляет последовательность кДНК протеазы из штамма CGMCC № 0670 Thermoascus aurantiacus (SEQ ID No.1), и SEQ ID No.2 представляет выведенную из нее аминокислотную последовательность.
Настоящее изобретение относится также к выделенным последовательностям нуклеиновых кислот, кодирующим полипептиды, и к конструкциям на основе нуклеиновых кислот, векторам и клеткам-хозяевам, содержащим последовательности нуклеиновых кислот, а также к способам получения и применения полипептидов, в частности, в корме для животных.
Краткое описание чертежей
На фигуре 1 показаны результаты испытания на ингибирование с использованием протеазы штамма CGMCC №0670 Thermoascus aurantiacus.
На фигуре 2 показан температурный профиль вышеуказанного испытания.
На фигуре 3 показан профиль показателей рН вышеуказанного испытания.
На фигуре 4 показана устойчивость к показателям рН в вышеуказанном испытании.
На фигуре 5 показаны результаты испытания на субстратную специфичность в вышеуказанном испытании.
Подробное описание изобретения
Авторы настоящего изобретения неожиданно обнаружили новый класс протеаз, которые могут быть пригодны для использования в корме для животных. Большинство известных кормовых протеаз являются сериновыми протеазами, причем многие из указанных протеаз не обладают достаточной кислотостойкостью, чтобы избежать разрушения в кислотной среде желудка. Протеазы по данному изобретению представляют собой кислотостойкую металлопротеазу, выделенную из Thermoascus aurantiacus. Зрелая полипептидная часть указанной протеазы содержит аминокислоты 1-177 SEQ ID No.2. При выполнении исследований in vitro, имитирующих пищеварение у животных с однокамерным желудком и рыб, было установлено, что протеаза Thermoascus aurantiacus может увеличивать количество растворимого и расщепляемого белка и повышать степень гидролиза белка. Гомологичный новый полипептид обнаружен в Aspergillus oryzae (аминокислоты 177-353 SEQ ID No.11).
Полипептиды, обладающие протеазной активностью
Протеазы иногда именуются также пептидазами, протеиназами, пептидными гидролазами или протеолитическими ферментами. Протеазы могут относиться к экзо-типу, в соответствии с которым гидролиз пептидов происходит, начиная с любого конца, или к эндо-типу, в соответствии с которым гидролиз происходит внутри полипептидных цепей (эндопептидазы). Эндопептидазы активно воздействуют на пептидные субстраты, блокированные у N- и C-конца, что характерно для специфичности рассматриваемой протеазы.
Термин "протеаза" в используемом здесь значении означает фермент, который гидролизует пептидные связи. Протеаза означает любой фермент, относящийся к группе ферментов ЕС 3.4 (включая ферменты тринадцати подклассов). Номер ЕС относится к номенклатуре ферментов, представленной в Enzyme Nomenclature 1992 NC-IUBMB, Academic Press, San Diego, California, включая дополнения 1-5, опубликованные соответственно в журналах Eur. J. Biochem. 1994, 223, 1-5; Eur. J. Biochem. 1995, 232, 1-6; Eur. J. Biochem. 1996, 237, 1-5; Eur. J. Biochem. 1997, 250, 1-6; и Eur. J. Biochem. 1999, 264, 610-650. Данная номенклатура регулярно дополняется и обновляется; см., например, в Интернете (WWW) на сайте http://www.chem.gmw.ac.uk/iubmb/enzyme/index.html).
Протеазы классифицированы с учетом их каталитического механизма в следующие группы: сериновые протеазы (S), цистеиновые протеазы (С), аспарагиновые протеазы (А), металлопротеазы (М) и неизвестные или еще не классифицированные протеазы (U); см. справочник Handbook of Proteolytic Enzymes, A.J. Barrett, N.D. Rawlings, J.F. Woessner (eds.), Academic Press (1998), в частности, общее введение.
В конкретных вариантах осуществления изобретения протеазы по данному изобретению выбраны из группы, состоящей из:
(а) протеаз, относящихся к металлоэндопептидазам ЕС 3.4.24;
(b) металлопротеаз, относящихся к группе М, приведенных в вышеуказанном справочнике;
(с) металлопротеаз, еще не отнесенных к какому-либо классу (обозначение: Clan MX) или относящихся к одному из классов МА, МВ, МС, MD, ME, MF, MG, MH (рассмотренные на стр. 989-991 вышеуказанного справочника);
(d) других семейств металлопротеаз (рассмотренных на стр. 1448-1452 вышеуказанного справочника);
(е) металлопротеаз с фрагментом HEXXH;
(f) металлопротеаз с фрагментом HEFTH;
(g) металлопротеаз, относящихся к одному из семейств М3, М26, М27, М32, М34, М35, М36, М41, М43 или М47 (рассмотренные на стр. 1448-1452 вышеуказанного справочника); и
(h) металлопротеаз, относящихся к семейству М35 (рассмотренные на стр. 1492-1495 вышеуказанного справочника).
В других конкретных вариантах осуществления изобретения металлопротеазы являются гидролазами, в которых нуклеофильное воздействие на пептидную связь опосредовано молекулой воды, активируемой катионом двухвалентного металла. Примерами катионов двухвалентных металлов являются цинк, кобальт или марганец. Ион металла может удерживаться в требуемом положении лигандами аминокислот. Число лигандов может быть равно пяти, четырем, трем, двум, одному или нулю. В конкретном варианте осуществления изобретения число лигандов равно двум или трем, предпочтительно трем.
Чтобы определить, является ли данная протеаза металлопротеазой или нет, следует обратиться к вышеуказанному справочнику и к изложенным в нем принципам. Такое определение можно произвести для всех типов протеаз независимо от того, относится такая протеаза к природному или дикому типу, является генетически сконструированной или синтетической.
Активность протеазы можно измерить при помощи любого анализа, в котором используется субстрат, имеющий пептидные связи, соответствующие специфичности исследуемой протеазы. Показатель рН и температуру анализа необходимо также привести в соответствие с исследуемой протеазой. Примерами значений рН для анализа являются рН 6, 7, 8, 9, 10 или 11. Примерами температур для анализа являются 30, 35, 37, 40, 45, 50, 55, 60, 65, 70 или 80°С.
Примером субстрата для протеазы является казеин, в частности казеин, сшитый с азурином (AZCL-казеин). Два анализа протеазы описаны в примере 1, из которых так называемый анализ с использованием AZCL-казеина является более предпочтительным для достижения целей настоящего изобретения.
Не существует ограничений, относящихся к происхождению протеазы по данному изобретению. Таким образом, термин "протеаза" включает не только протеазы природного или дикого типа, полученные из микроорганизмов любого рода, но также любые мутанты, варианты, фрагменты и т.д., обладающие протеазной активностью, а также синтетические протеазы, такие как инвертированные протеазы и консенсусные протеазы. Такие генетически сконструированные протеазы можно получить методами, известными в данной области, например, при помощи сайт-направленного мутагенеза, полимеразной цепной реакции (PCR) (используя фрагмент PCR, содержащий требуемую мутацию в качестве одной из затравок при выполнении реакций PCR) или неспецифического мутагенеза. Получение консенсусных белков описано, например, в европейском патенте ЕР 897985. Термин "полученный из" применительно к данному источнику означает, что полипептид, кодируемый последовательностью нуклеиновой кислоты, продуцирован источником или клеткой, в которых присутствует данная последовательность нуклеиновой кислоты. В предпочтительном варианте осуществления изобретения полипептид секретируется вне клетки.
В конкретном варианте осуществления изобретения протеаза является слабоаллергенным вариантом, вызывающим пониженную иммунологическую реакцию при воздействии на животных, включая человека. Термин "иммунологическая реакция" означает любую реакцию иммунной системы животного под действием данной протеазы. Одним типом иммунологической реакции является аллергическая реакция, вызывающая повышение уровней IgE у подвергнутого такому воздействию животного. Слабоаллергенные варианты можно получить методами, известными в данной области. Например, протеаза может быть конъюгирована с полимерами, защищающими части или эпитопы протеазы, вызывающие иммунологическую реакцию. Конъюгация с полимерами может включать химическое связывание полимера с протеазой in vitro, как это описано, например, в заявках WO 96/17929, WO 98/30682, WO 98/35026 и/или WO 99/00489. Конъюгация может дополнительно или альтернативно включать связывание полимеров с протеазой in vivo. Такая конъюгация может быть достигнута путем создания методами генетической инженерии нуклеотидной последовательности, кодирующей протеазу, введения в протеазу консенсусных последовательностей, кодирующих дополнительные сайты гликозилирования, и экспрессии протеазы в хозяине, способном гликозилировать протеазу, как это описано, например, в заявке WO 00/26354. Другим способом получения слабоаллергенных вариантов является создание методами генетической инженерии нуклеотидной последовательности, кодирующей протеазу, которая вызывает аутоолигомеризацию протеазы, благодаря которой одни мономеры протеазы могут защищать эпитопы других мономеров протеазы и уменьшать, таким образом, антигенность олигомеров. Такие продукты и их получение описаны, например, в заявке WO 96/16177. Эпитопы, вызывающие иммунологическую реакцию, могут быть идентифицированы разными методами, такими как метод выявления фагов, описанный в заявках WO 00/26230 и WO 01/83559, или неспецифический подход, описанный в ЕР №561907. После идентификации эпитопа его аминокислотную последовательность можно изменить с получением измененных иммунологических свойств протеазы, используя известные методы манипуляции генами, такие как сайтнаправленный мутагенез (см., например, заявки WO 00/26230, WO 00/26354 и/или WO 00/22103), и/или конъюгируя полимер в достаточной близости к эпитопу, чтобы защитить эпитоп.
Настоящее изобретение относится к выделенным полипептидам, содержащим аминокислотную последовательность, которая идентична аминокислотам -178-177, -159-177 или предпочтительно аминокислотам 1-177 (зрелый полипептид) SEQ ID No.2 по крайней мере примерно на 64% или по крайней мере примерно на 65%, по крайней мере примерно на 70% или по крайней мере примерно на 75%, по крайней мере примерно на 76% или по крайней мере примерно на 77%, по крайней мере примерно на 78% или по крайней мере примерно на 79%, по крайней мере примерно на 80% или по крайней мере примерно на 82%, по крайней мере примерно на 85%, или по крайней мере примерно на 90%, или по крайней мере примерно на 95%, или по крайней мере примерно на 97%, и обладает протеазной активностью (далее именуемые "гомологичные полипептиды"). В конкретных вариантах осуществления изобретения полипептиды по данному изобретению i) имеют или ii) состоят из аминокислотной последовательности с вышеуказанной степенью идентичности.
Протеаза Thermoascus aurantiacus, зрелый полипептид которой содержит аминокислоты 1-177 SEQ ID No.2, несомненно, является одним примером полипептида по данному изобретению.
Другой полипептид по данному изобретению, полученный из Aspergillus oryzae, содержит SEQ ID No.11 или аминокислоты -23-353, -23-374, -23-397, 1-353, 1-374, 1-397, 177-353, 177-374 или 177-397 и кодируется SEQ ID No.10 или соответственно ее нуклеотидами 2-1129, 2-1195, 2-1267, 71-1129, 71-1195, 71-1267, 599-1129, 599-1195 или 599-1267.
В соответствии с целями настоящего изобретения степень идентичности двух аминокислотных последовательностей, а также степень идентичности двух нуклеотидных последовательностей можно определить при помощи программы "Align", которая представляет собой сопоставление по Нидлману-Вуншу (то есть глобальное сопоставление). Указанная программа используется для сопоставления полипептидных и нуклеотидных последовательностей. Для сопоставления полипептидов используется матрица подсчета по умолчанию BLOSUM50 и для сопоставления нуклеотидов используется матрица идентичности по умолчанию. За первый остаток разрыва цепи начисляется -12 очков для полипетидов и -16 очков для нуклеотидов. За последующие остатки разрыва цепи начисляется -2 очка для полипептидов и -4 очка для нуклеотидов.
Программа "Align" является частью пакета программ FASTA версии v20u6 (см. W.R. Pearson and D.J. Lipman (1988), "Improved Tools for Biological Sequence Analysis", PNAS 85:2444-2448, and W.R. Pearson (1990) "Rapid and Sensitive Sequence Comparison with FASTP and FASTA", Methods in Enzymology 183:63-98). В программе сопоставления белков FASTA использован алгоритм Смита-Уотермана без ограничения величины разрыва цепи (см. "Smith-Waterman algorithm", T.F. Smith and M.S. Waterman (1981) J. Mol. Biol. 147:195-197).
В конкретном варианте осуществления изобретения гомологичные полипептиды имеют аминокислотную последовательность, которая отличается сорока, тридцатью пятью, тридцатью, двадцатью пятью, двадцатью или пятнадцатью аминокислотами. В другом варианте осуществления изобретения гомологичные полипептиды имеют аминокислотную последовательность, которая отличается десятью, девятью, восьмью, семью, шестью или пятью аминокислотами. В другом конкретном варианте осуществления изобретения гомологичные полипептиды отличаются четырьмя, тремя или двумя аминокислотами либо одной аминокислотой из аминокислот -178-177, -159-177 или +1-177 SEQ ID No.2.
В конкретных вариантах осуществления изобретения полипептиды по настоящему изобретению а) имеют или b) состоят из
i) аминокислотной последовательности, состоящей из аминокислот -178-177, -159-177 или +1-177 SEQ ID No.2;
ii) аминокислотной последовательности, состоящей из аминокислот -23-353, -23-374, -23-397, 1-353, 1-374, 1-397, 177-353, 177-374 или 177-397 SEQ ID No.11; или
аллельных вариантов или фрагментов последовательностей i) и ii), обладающих протеазной активностью.
Фрагмент аминокислот -178-177, -159-177 или +1-177 SEQ ID No.2 или аминокислот -23-353, -23-374, -23-397, 1-353, 1-374, 1-397, 177-353, 177-374 или 177-397 SEQ ID No.11 представляет полипептид, в котором одна или более аминокислот удалены из амино- и/или карбоксильного конца указанных аминокислотных последовательностей. В одном варианте осуществления изобретения фрагмент содержит по крайней мере 75 аминокислотных остатков, или по крайней мере 100 аминокислотных остатков, или по крайней мере 125 аминокислотных остатков, или по крайней мере 150 аминокислотных остатков, по крайней мере 160 аминокислотных остатков, или по крайней мере 165 аминокислотных остатков, или по крайней мере 170 аминокислотных остатков, или по крайней мере 175 аминокислотных остатков.
Аллельный вариант представляет собой одну из двух или более альтернативных форм гена, занимающего тот же хромосомный локус. Аллельный вариант образуется в результате мутации и может привести к полиморфизму в популяциях. Генные мутации могут быть "молчащими" (отсутствие изменений в кодированном полипептиде) или могут кодировать полипептиды, содержащие измененные аминокислотные последовательности. Аллельным вариантом полипептида является полипептид, кодированный аллельным вариантом гена.
Настоящее изобретение относится также к выделенным полипептидам, обладающим протеазной активностью, которые кодированы последовательностями нуклеиновых кислот, гибридизирующими в условиях очень низкой, низкой, средней, средне-высокой, высокой или очень высокой строгости с зондом нуклеиновой кислоты, который гибридизирует в тех же условиях (а) с нуклеотидами 25-1089, 1-1089, 1-1344, 25-1344, 559-1344 или предпочтительно с нуклеотидами 559-1089 SEQ ID No.1, (b) с последовательностью кДНК, находящейся в нуклеотидах 559-1089 SEQ ID No.1, (с) с субпоследовательностью последовательности (а) или (b), (d) с комплементарной цепью последовательности (а), (b) или (с) (J. Sambrook, E.F. Fritsch, and T. Maniatis, 1989, Molecular Cloning, A Laboratory Manual, 2nd edition, Cold Spring Harbor, New York). В одном конкретном варианте осуществления изобретения зонд нуклеиновой кислоты выбирают из вышеуказанных последовательностей нуклеиновых кислот (а), (b), (c) или (d).
Последовательности нуклеотидов 559-1089, 25-1089, 1-1089, 1-1344, 25-1344 или 559-1344 SEQ ID No.1 могут содержать по крайней мере 100 нуклеотидов или в другом варианте осуществления изобретения по крайней мере 200 нуклеотидов. Кроме того, указанная последовательность может кодировать фрагмент полипептида, обладающий протеазной активностью.
Последовательности нуклеиновых кислот, содержащие нуклеотиды 559-1089, 25-1089, 1-1089, 1-1344, 25-1344 или 559-1344 SEQ ID No.1 или их субпоследовательность, а также аминокислотные последовательности, содержащие аминокислоты -178-177, -159-177 или +1-177 SEQ ID No.2 или их фрагмент, можно использовать для создания зонда нуклеиновой кислоты для идентификации и клонирования ДНК, кодирующей полипептиды, обладающие протеазной активностью, из штаммов других родов или видов методами, хорошо известными в данной области. В частности, такие зонды можно использовать для гибридизации с геномной или кДНК представляющего интерес рода или вида стандартными методами саузерн-блоттинга, чтобы идентифицировать и выделить соответствующий ген. Такие зонды могут быть значительно короче всей последовательности, но должны содержать по крайней мере 15, предпочтительно по крайней мере 25 и более предпочтительно по крайней мере 35 нуклеотидов. Можно также использовать более длинные зонды. Можно использовать ДНК- и РНК-зонды. Зонды обычно метят для обнаружения соответствующего гена (например, 32Р, 3Н, 35S, биотином или авидином). Такие зонды входят в объем настоящего изобретения.
Таким образом, можно произвести скрининг библиотеки геномной ДНК или кДНК, полученной из других подобных организмов, для скрининга ДНК, гибридизирующейся с вышеописанными зондами и кодирующей полипептид, обладающий протеазной активностью. Геномную или другую ДНК из других подобных организмов можно выделить электрофорезом в агарозном или полиакриламидном геле или другими методами выделения. ДНК из библиотек или выделенную ДНК можно перенести или иммобилизовать на нитроцеллюлозе или другом приемлемом носителе. Чтобы идентифицировать клон или ДНК, гомологичную SEQ ID No.1 или ее субпоследовательности, носитель используют при выполнении анализа методом саузерн-блоттинга. В соответствии с целями настоящего изобретения гибридизация показывает, что последовательность нуклеиновой кислоты гибридизирует с меченым зондом нуклеиновой кислоты, соответствующим последовательности нуклеиновой кислоты, представленной в SEQ ID No.1, ее комплементарной цепи или субпоследовательности, в условиях от очень низкой до очень высокой строгости. Молекулы, с которыми гибридизирует зонд нуклеиновой кислоты в указанных условиях, можно обнаружить при помощи рентгеновской пленки.
В конкретном варианте осуществления изобретения зонд нуклеиновой кислоты представляет собой последовательность нуклеиновой кислоты, кодирующую аминокислоты -178-177, -159-177 или +1-177 SEQ ID No.2, или ее субпоследовательности. В другом варианте осуществления изобретения зонд нуклеиновой кислоты содержит нуклеотиды 25-1089, 1-1089, 1-1344, 25-1344, 559-1344 или предпочтительно нуклеотиды 559-1089 SEQ ID No.1 (кодирующая зрелый полипептид область SEQ ID No.1). В другом предпочтительном варианте осуществления изобретения зонд нуклеиновой кислоты представляет собой последовательность нуклеиновой кислоты или предпочтительно ее область, кодирующую зрелый полипептид, которая находится в плазмиде, находящейся в Escherichia coli DSM 14652, при этом последовательность нуклеиновой кислоты кодирует полипептид, обладающий протеазной активностью.
Для получения длинных зондов, содержащих по крайней мере 100 нуклеиотидов, условия от очень низкой до очень высокой строгости определяются как прегибридизация и гибридизация при 42°С в 5-кратном объеме SSPE, 0,3% SDS, 200 мкг/мл фрагментированной и денатурированной ДНК спермы лосося и 25% формамида для условий очень низкой и низкой строгости, 35% формамида для условий средней и средне-высокой строгости или 50% формамида для условий высокой и очень высокой строгости в соответствии со стандартными методами саузерн-блоттинга.
Для получения длинных зондов, содержащих по крайней мере 100 нуклеотидов, носитель трижды промывают по 15 минут, используя 2-кратный объем SSC, 0,2% SDS, предпочтительно по крайней мере при 45°С (условия очень низкой строгости), более предпочтительно по крайней мере при 50°С (условия низкой строгости), более предпочтительно по крайней мере при 55°С (условия средней строгости), более предпочтительно по крайней мере при 60°С (условия средне-высокой строгости), еще предпочтительнее по крайней мере при 65°С (условия высокой строгости) и наиболее предпочтительно по крайней мере при 70°С (условия очень высокой строгости).
Для получения коротких зондов длиной от около 15 нуклеотидов до около 70 нуклеотидов строгие условия определяются как прегибридизация, гибридизация и промывка после гибридизации при температуре на 5°С-10°С ниже вычисленной температуры Тm при выполнении вычислений Болтона и МакКартни (1962, Proceedings of the National Academy of Sciences USA 48:1390) в 0,9 М NaCl, 0,09 М трис-HCl c рН 7,6, 6 мМ EDTA, 0,5% NP-40, 1 объем раствора Денгардта, 1 мМ пирофосфата натрия, 1 мМ одноосновного фосфата натрия, 0,1 мМ АТФ и 0,2 мг РНК дрожжей на мл в соответствии со стандартными методами саузерн-блоттинга.
Для получения коротких зондов длиной от около 15 нуклеотидов до около 70 нуклеотидов носитель один раз промывают в 6-кратном объеме SCC с 0,1% SDS в течение 15 минут и дважды промывают 6-кратным объемом SSC в течение 15 минут при температуре на 5°С-10°С ниже вычисленной температуры Тm.
Настоящее изобретение относится также к вариантам полипептида, содержащего аминокислотную последовательность, состоящую из аминокислот 1-177, -159-177 или -178-177 SEQ ID No.2, включающим замену, делецию и/или инсерцию одной или более аминокислот.
Аминокислотные последовательности вариантных полипептидов могут отличаются от аминокислотной последовательности, состоящей из аминокислот 1-177, -159-177 или -178-177 SEQ ID No.2, вследствие инсерции или делеции одного или более аминокислотных остатков и/или замены одного или более аминокислотных остатков другими аминокислотными остатками. Замены аминокислот предпочтительно должны быть незначительными, например консервативные аминокислотные замены, которые не оказывают существенного влияния на укладку цепи и/или активность белка, небольшие делеции обычно от одной до около 30 аминокислот, небольшие удлиняющие фрагменты у амино- или карбоксильного конца, такие как аминоконцевой остаток метионина, небольшой линкерный пептид, содержащий примерно 20-25 остатков; или небольшой удлиняющий фрагмент, облегчающий очистку благодаря изменению суммарного заряда или другой функции, такой как полигистидиновый хвост, антигенный эпитоп или домен связывания.
Примеры консервативных замен относятся к группе основных аминокислот (аргинин, лизин и гистидин), кислых аминокислот (глутаминовая кислота или аспарагиновая кислота), полярных аминокислот (глутамин и аспарагин), гидрофобных аминокислот (лейцин, изолейцин и валин), ароматических аминокислот (фенилаланин, триптофан и тирозин) и аминокислот с короткой цепью (глицин, аланин, серин, треонин и метионин). Замены аминокислот, которые фактически не изменяют удельную активность, известны в данной области и описаны, например, в публикации H. Neurath and R.L. Hill, 1979, In The Proteins, Academic Press, New York. Наиболее часто встречающимися заменами являются Ala/Ser, Val/Ile, Asp/Glu, Thr/Ser, Ala/Gly, Ala/Thr, Ser/Asn, Ala/Val, Ser/Gly, Tyr/Phe, Ala/Pro, Lys/Arg, Asp/Asn, Leu/Ile, Leu/Val, Ala/Glu и Asp/Gly, а также указанные замены в обратном порядке.
Настоящее изобретение относится также к выделенным полипептидам, обладающим протеазной активностью, которые выбраны из группы, состоящей из:
(f) полипептидов, имеющих
(i) вычисленную молекулярную массу от около 17 до около 22 кДа, предпочтительно от около 18 до около 21 кДа или от около 19 до около 20 кДа; или экспериментально установленную молекулярную массу (методом SDS-PAGE) от около 19 до около 27 кДа, предпочтительно от около 20 до около 26 кДа, от около 21 до около 25 кДа или от около 22 до около 24 кДа;
(ii) pI от около рН 7 до около рН 10, предпочтительно от около рН 7,5 до около рН 9,5 или от около рН 8,0 до около рН 9,0;
(iii) оптимальный показатель рН от около рН 5,5 до около рН 8,0, предпочтительно от около рН 5,5 до около рН 7,0 или от около рН 5,5 до около рН 6,5, определяемый при помощи анализа с использованием AZCL-казеина при 45°С и буферной системы с янтарной кислотой; и/или
(iv) оптимальную температуру от около 45°С до около 90°С, предпочтительно от около 50°С до около 85°С или от около 60°С до около 80°С, определяемую при помощи анализа с использованием AZCL-казеина при рН9;
(g) полипептидов, не ингибируемых
(v) SSI (ингибитором субтилизина Streptomyces) и/или
(vi) EDTA
при определении при помощи анализа с использованием AZCL-казеина при рН 9 и 45°С;
(h) полипептидов, устойчивых в интервале от около рН 3,5 до около рН 10,5, предпочтительно в интервале от около рН 3,5 до около рН 9,0 или в интервале от около рН 4,0 до рН 5,0 после инкубации в течение 2 часов при 37°С в буферной системе с янтарной кислотой, при этом остаточную активность измеряют при помощи анализа с использованием AZCL-казеина при рН9 и 45°С;
(k) полипептидов, синтезированных с использованием N-концевого пропептида; и
(l) полипептидов, имеющих мотив НЕХХН.
Стабильность полипептида по (h) означает, что остаточная активность равна по крайней мере 40%, 50%, 60%, 70% или 80% по сравнению с контрольным образцом, который не подвергали предварительной инкубации в течение 2 часов. Полипептиды, которые являются устойчивыми в соответствии с данным определением, могут быть определены как кислотостойкие полипептиды.
Полипептид по настоящему изобретению может быть бактериальным полипептидом. Например, такой полипептид может быть полипептидом грамоположительных бактерий, таким как полипептид Bacillus или полипептид Streptomyces; или полипептидом грамотрицательных бактерий, например полипептидом E. coli или Pseudomonas sp.
Полипептид по настоящему изобретению может быть грибковым полипептидом и более предпочтительно дрожжевым полипептидом, таким как полипептид Candida, Kluyveromyces, Pichia, Saccharomyces, Schizosaccharomyces или Yarrowia; или более предпочтительно полипептидом нитевидного грибка, таким как полипептид Acremonium, Aspergillus, Aureobasidium, Cryptococcus, Filibasidium, Fusarium, Humicola, Magnaporthe, Mucor, Myceliophthora, Neocallimastix, Neurospora, Paecilomyces, Penicillium, Piromyces, Schizophyllum, Talaromyces, Thermoascus, Thielavia, Tolypocladium или Trichoderma.
В другом варианте осуществления изобретения полипептид является полипептидом Aspergillus aculeatus, Aspergillus awamori, Aspergillus foetidus, Aspergillus japonicus, Aspergillus nidulans, Aspergillus niger, Aspergillus oryzae, Fusarium bactridioides, Fusarium cerealis, Fusarium crookwellense, Fusarium culmorum, Fusarium graminearum, Fusarium graminum, Fusarium heterosporum, Fusarium negundi, Fusarium oxysporum, Fusarium reticulatum, Fusarium roseum, Fusarium sambucinum, Fusarium sarcochroum, Fusarium sporotrichioides, Fusarium sulphureum, Fusarium torulosum, Fusarium trichothecioides, Fusarium venenatum, Humicola insolens, Humicola lanuginosa, Mucor miehei, Myceliophthora thermophila, Neurospora crassa, Penicillium purpurogenum, Thermoascus aurantiacus, Trichoderma harzianum, Trichoderma koningii, Trichoderma longibrachiatum, Trichoderma reesei или Trichoderma viride.
В другом варианте осуществления изобретения полипептид является полипептидом, выделенным из нитевидного грибка типа Ascomycota, предпочтительно класса Pezizomycotina, более предпочтительно отряда Eurotiomycetes, еще предпочтительнее подсемейства Eurotiales и наиболее предпочтительно семейства Trichocomaceae.
В еще одном варианте осуществления изобретения полипептид выделяют из грибка рода Thermoascus, например вида Thermoascus aurantiacus, такого как штамм CGMCC №0670 Thermoascus aurantiacus, например, полипептид с аминокислотной последовательностью, содержащей аминокислоты -178-177, -159-177 или +1-177 SEQ ID No.2.
Очевидно, что в объем данного изобретения входят вышеуказанные виды в совершенном и дефектном состояниях и другие таксономические эквиваленты, например анаморфы, независимо от названия вида, под которым они известны. Специалисты в данной области могут легко установить идентичность соответствующих эквивалентов.
Штаммы указанных видов могут быть получены в ряде коллекций культур, таких как Американская коллекция типовых культур (АТСС), Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (DSM), Centraalbureau Voor Schimmelcultures (CBS) и Agricultural Research Service Patent Culture Collection, Norhern Regional Research Center (NRRL).
Кроме того, такие полипептиды могут быть идентифицированы и получены из других источников, включающих микроорганизмы, выделенных из природных источников (например, почвы, компоста, воды и т.д.) с использованием вышеуказанных зондов. Методы выделения микроорганизмов из естественных мест обитания хорошо известны в данной области. Последовательность нуклеиновой кислоты может быть выделена в результате аналогичного скрининга библиотеки геномной ДНК или кДНК другого микроорганизма. После обнаружения при помощи зонда (зондов) последовательности нуклеиновой кислоты, кодирующей полипептид, указанную последовательность можно выделить или клонировать методами, известными специалистам в данной области (см., например, Sambrook et al., 1989, приведена выше).
Как указано в данном описании изобретения, "выделенный" полипептид является полипептидом, который по существу не содержит других непротеазных полипептидов, например, указанный полипептид является чистым по крайней мере примерно на 20%, предпочтительно по крайней мере примерно на 40%, более предпочтительно примерно на 60%, еще предпочтительнее примерно на 80%, наиболее предпочтительно примерно на 90% и особенно предпочтительно примерно на 95% при определении методом SDS-PAGE.
Полипептиды, кодированные последовательностями нуклеиновых кислот по настоящему изобретению, включают также слитые полипептиды или расщепляемые слитые полипептиды, в которых другой полипептид слит с N-концом или C-концом данного полипептида или его фрагмента. Слитый полипептид получают слиянием последовательности нуклеиновой кислоты (или ее части), кодирующей другой полипептид, с последовательностью нуклеиновой кислоты (или ее частью) по настоящему изобретению. Методы получения слитых полипептидов известны в данной области и включают лигирование последовательностей, кодирующих полипептиды, при котором они находятся в рамке считывания и экспрессия слитого полипептида контролируется тем же промотором (промоторами) и терминатором.
Последовательности нуклеиновых кислот
Настоящее изобретение относится также к выделенным последовательностям нуклеиновых кислот, которые кодируют полипептид по настоящему изобретению. Типичные последовательности нуклеиновых кислот по данному изобретению содержат нуклеотиды 25-1089, 1-1089, 1-1344, 25-1344, 559-1344 и, в частности, нуклеотиды 559-1089 SEQ ID No.1, причем последние нуклеотиды соответствуют области, кодирующей зрелый полипептид. Другой типичной последовательностью нуклеиновых кислот по данному изобретению является последовательность, предпочтительно ее область, кодирующая зрелый полипептид, которая встроена в плазмиду, введенную в депонированный микроорганизм Escherichia coli DSM 14652. В объем настоящего изобретения входят также последовательности нуклеиновых кислот, которые кодируют полипептид, содержащий аминокислотную последовательность, включающую аминокислоты 1-177, -159-177 или -178-177 SEQ ID No.2, которые отличаются от соответствующих частей SEQ ID No.1 вследствие вырожденности генетического кода. Настоящее изобретение относится также к субпоследовательностям SEQ ID No.1, которые кодируют фрагменты SEQ ID No.2, обладающие протеазной активностью.
Настоящее изобретение относится также к выделенным последовательностям нуклеиновых кислот, которые кодируют полипептид по настоящему изобретению. Типичными последовательностями нуклеиновых кислот по данному изобретению являются SEQ ID No.10 или нуклеотиды 2-1129, 2-1195, 2-1267, 71-1129, 71-1195, 71-1267, 599-1129, 599-1195 или 599-1267, нуклеотиды 71-1129 SEQ ID No.10, соответствующие области, кодирующей зрелый полипептид.
Субпоследовательность SEQ ID No.1 является последовательностью нуклеиновой кислоты, входящей в SEQ ID No.1, за исключением делеции одного или более нуклеотидов у 5'- и/или 3'-конца. Субпоследовательность предпочтительно содержит по крайней мере 225 нуклеотидов, более предпочтительно по крайней мере 300 нуклеотидов, еще предпочтительнее по крайней мере 375, 450, 500, 531, 600, 700, 800, 900, 1000, 1100, 1200 или 1300 нуклеотидов.
Настоящее изобретение относится также к нуклеотидным последовательностям с степенью идентичности к нуклеотидам 25-1089, 1-1089, 1-1344, 25-1344, 559-1344 или предпочтительно нуклеотидам 559-1089 SEQ ID No.1, равной по крайней мере 69%, 70%, 72%, 74%, 76% или 80%. В конкретном варианте осуществления изобретения степень идентичности равна по крайней мере 85%, по крайней мере 90%, по крайней мере 95% или по крайней мере 97%. Для определения степени идентичности нуклеотидов можно использовать вышеуказанную программу "align".
Конкретный вариант осуществления изобретения относится к нуклеотидным последовательностям со степенью идентичности к нуклеотидам 25-1089, 1-1089, 1-1344 или 25-1344 SEQ ID No.1, равной по крайней мере 55%, 60%, 62%, 64%, 66%, 68%, 70%, 72%, 74%, 76% или 80%. В конкретном варианте осуществления изобретения степень идентичности равна по крайней мере 85%, по крайней мере 90%, по крайней мере 95% или по крайней мере 97%. Для определения степени идентичности нуклеотидов можно использовать вышеуказанную программу "align".
Настоящее изобретение относится также к мутантным последовательностям нуклеиновых кислот, содержащим по крайней мере одну мутацию в нуклеотидах 25-1089, 1-1089, 1-1344, 25-1344, 559-1344 или 559-1089 SEQ ID No.1, где мутантная последовательность нуклеиновой кислоты кодирует полипептид, который (i) состоит из аминокислот -178-177, -159-177 или +1-177 SEQ ID No.2, (ii) является вариантом любой последовательности (i), который включает замену, делецию и/или инсерцию одной или более аминокислот, (iii) является аллельным вариантом любой последовательности (i) или (iv) является фрагментом любой последовательности (i).
Методы, используемые для выделения или клонирования последовательности нуклеиновой кислоты, кодирующей полипептид, известны в данной области и включают выделение из геномной ДНК, получение из кДНК или комбинацию указанных методов. Последовательности нуклеиновых кислот по настоящему изобретению можно клонировать из такой геномной ДНК, например, при помощи хорошо известной полимеразной цепной реакции (PCR) или серологического скрининга библиотек экспрессируемых последовательностей для обнаружения фрагментов клонированной ДНК с родственными структурными признаками. См., например, Innis et al., 1990, PCR: A Guide to Methods and Application, Academic Press, New York. Можно использовать другие методы амплификации нуклеиновых кислот, такие как лигазная цепная реакция (LCR), лигированная активированная транскрипция (LAT) и амплификация на основе последовательности нуклеиновой кислоты (NASBA). Последовательность нуклеиновой кислоты может быть клонирована из штамма Thermoascus, другого или родственного организма и, таким образом, например, может быть аллельным или видовым вариантом кодирующей полипептид области последовательности нуклеиновой кислоты.
Термин "выделенная последовательность нуклеиновой кислоты" в используемом здесь значении означает последовательность нуклеиновой кислоты, которая по существу не содержит других последовательностей нуклеиновых кислот, например является чистой по крайней мере примерно на 20%, предпочтительно по крайней мере примерно на 40%, более предпочтительно по крайней мере примерно на 60%, еще предпочтительнее по крайней мере примерно на 80% и наиболее предпочтительно по крайней мере примерно на 90% при определении электрофорезом в агарозном геле. Например, выделенную последовательность нуклеиновой кислоты можно получить стандартными методами клонирования, используемыми в генетической инженерии для перемещения последовательности нуклеиновой кислоты из естественного положения на другой сайт, где она будет репродуцироваться. Методы клонирования могут включать вырезание и выделение требуемого фрагмента нуклеиновой кислоты, включающего последовательность нуклеиновой кислоты, кодирующую полипептид, вставку фрагмента в молекулу вектора и введение рекомбинантного вектора в клетку-хозяина, где будут реплицированы многочисленные копии или клоны данной последовательности нуклеиновой кислоты. По происхождению последовательность нуклеиновой кислоты может быть геномной, кДНК, РНК, полусинтетической, синтетической или может представлять комбинацию указанных последовательностей.
Настоящее изобретение относится также к последовательностям нуклеиновых кислот, кодирующим активный полипептид, которые обладают степенью гомологии с кодирующей зрелый полипептид последовательностью SEQ ID No.1 (то есть нуклеотиды 559-1089), равной по крайней мере примерно 70%, предпочтительно примерно 75%, предпочтительно примерно 80%, более предпочтительно примерно 90%, еще предпочтительнее примерно 95% и наиболее предпочтительно примерно 97%. В соответствии с целями настоящего изобретения степень гомологии между двумя последовательностями нуклеиновых кислот может быть определена при помощи вышеуказанной программы "align".
Модификация последовательности нуклеиновой кислоты, кодирующей полипептид по настоящему изобретению, может быть необходима для синтеза полипептидов, по существу подобных данному полипептиду. Термин "по существу подобный" применительно к полипептиду означает не встречающиеся в природе формы полипептида. При конструировании такие полипептиды могут отличаться от полипептида, выделенного из естественного источника, например, они могут представлять собой варианты, отличающиеся удельной активностью, термостойкостью, оптимальным показателем рН или подобными параметрами. Вариантная последовательность может быть создана на основе последовательности нуклеиновой кислоты, которая является кодирующей указанный полипептид частью SEQ ID No.1, например ее субпоследовательностью, и/или путем введения нуклеотидных замен, которые не образуют другую аминокислотную последовательность полипептида, кодированного последовательностью нуклеиновой кислоты, но соответствуют используемому кодону организма хозяина, используемого для продуцирования протеазы, или путем введения нуклеотидных замен, которые могут образовывать другую аминокислотную последовательность. Для общего ознакомления с заменой нуклеотидов, см., например, Ford et al., 1991, Protein Expression and Purification 2: 95-107.
Специалистам в данной области должно быть очевидно, что такие замены могут быть произведены за пределами областей, имеющих существенное значение для функционирования молекулы, позволяя при этом получить активный полипептид. Аминокислотные остатки, имеющие важное значение для активности полипептида, кодированного выделенной последовательностью нуклеиновой кислоты по данному изобретению и поэтому предпочтительно не подлежащие замене, могут быть идентифицированы методами, известными в данной области, такими как сайтнаправленный мутагенез или аланин-сканирующий мутагенез (см., например, Cunningham and Wells, 1989, Science 244: 1081-1085). При использовании последнего метода мутации вводят в каждый положительно заряженный остаток в молекуле и полученные мутантные молекулы испытывают на протеазную активность для идентификации аминокислотных остатков, которые существенны для активности молекулы. Сайты взаимодействия субстрат-протеаза можно также определить при помощи анализа трехмерной структуры такими методами, как ядерный магнитный резонанс, кристаллография или фотоаффинное мечение (см., например, de Vos et al., 1992, Science 255: 306-312; Smith et al., 1992, Journal of Molecular Biology 224: 899-904; Wlodaver et al., 1992, FEBS Letters 309: 59-64).
Настоящее изобретение относится также к выделенным последовательностям нуклеиновых кислот, кодирующим полипептид по настоящему изобретению, который гибридизирует в условиях очень низкой строгости, предпочтительно в условиях низкой строгости, более предпочтительно в условиях средней строгости, более предпочтительно в условиях средне-высокой строгости, еще предпочтительнее в условиях высокой строгости и наиболее предпочтительно в условиях очень высокой строгости с зондом нуклеиновой кислоты, который гибридизирует в тех же условиях с последовательностью нуклеиновой кислоты SEQ ID No.1 или ее комплементарной цепью, или аллельными вариантами и субпоследовательностями (Sambrook et al., 1989, см. выше), как определено здесь.
Настоящее изобретение относится также к выделенным последовательностям нуклеиновых кислот, полученным (а) путем гибридизации ДНК в условиях очень низкой, низкой, средней, средне-высокой, высокой или очень высокой строгости с (i) нуклеотидами 559-1089, 25-1089, 1-1089, 1-1344, 25-1344 или 559-1344 SEQ ID No.1, (ii) последовательностью кДНК, содержащейся в нуклеотидах 559-1089, 25-1089, 1-1089, 1-1344, 25-1344 или 559-1344 SEQ ID No.1, (iii) субпоследовательностью последовательности (i) или (ii) либо (iv) комплементарной цепью последовательности (i), (ii) или (iii); и (b) путем выделения последовательности нуклеиновой кислоты. Субпоследовательность предпочтительно является последовательностью, содержащей по крайней мере 100 нуклеотидов, такой как последовательность, кодирующая полипептидный фрагмент, обладающий протеазной активностью.
Способы получения мутантных последовательностей нуклеиновых кислот
Настоящее изобретение далее относится к способам получения мутантной последовательности нуклеиновой кислоты, которые включают введение по крайней мере одной мутации в кодирующую зрелый полипептид последовательность SEQ ID No.1 или ее субпоследовательность, причем мутантная последовательность нуклеиновой кислоты кодирует полипептид, состоящий из аминокислот 1-177, -159-177 или -178-177 SEQ ID No.2, или его фрагмента, обладающего протеазной активностью.
Введение мутации в последовательность нуклеиновой кислоты для замены одного нуклеотида другим нуклеотидом может быть произведено путем сайтнаправленного мутагенеза при помощи любых методов, известных в данной области. Особенно пригодным является метод, в котором используется вектор на основе суперспиральной, двухцепочечной ДНК с представляющей интерес вставкой и двумя синтетическими затравками, содержащими требуемую мутацию. Олигонуклеотидные затравки, комплементарные противоположным цепям вектора, удлиняются во время циклического изменения температуры при помощи ДНК-полимеразы Pfu. При встраивании затравок образуется мутированная плазмида, содержащая неравномерно расположенные одноцепочечные разрывы. Вслед за циклическим изменением температуры продукт обрабатывают Dpnl, который специфически воздействует на метилированную и полуметилированную ДНК, расщепляя родительскую матричную ДНК и отбирая синтезированную ДНК, содержащую мутацию. Можно также использовать другие методы, известные в данной области.
Конструкции на основе нуклеиновых кислот
Настоящее изобретение относится также к конструкциям на основе нуклеиновых кислот, которые содержат последовательность нуклеиновой кислоты по настоящему изобретению, функционально связанную с одной или более регуляторными последовательностями, направляющими экспрессию кодирующей последовательности в приемлемой клетке-хозяине в условиях, сопоставимых с регуляторными последовательностями. Понятно, что экспрессия включает любые стадии, осуществляемые при продуцировании полипептида, которые включают, не ограничиваясь ими, транскрипцию, посттранскрипционную модификацию, трансляцию, посттрансляционную модификацию и секрецию.
Термин "конструкция на основе нуклеиновой кислоты" в используемом здесь значении означает молекулу одноцепочечной или двухцепочечной нуклеиновой кислоты, которая выделена из природного гена или модифицирована с включением сегментов нуклеиновой кислоты, объединенных и расположенных в таком порядке, в каком они никогда не встречаются в природе. Термин "конструкция на основе нуклеиновой кислоты" синонимичен термину "экспрессирующий кластер", когда конструкция на основе нуклеиновой кислоты содержит все регуляторные последовательности, необходимые для экспрессии кодирующей последовательности по настоящему изобретению. Термин "кодирующая последовательность" в используемом здесь значении означает последовательность нуклеиновой кислоты, которая непосредственно определяет аминокислотную последовательность ее белкового продукта. Границы кодирующей последовательности обычно определяются сайтом связывания рибосомы (прокариоты) или инициирующим кодоном ATG (эукариоты), расположенным выше открытой рамки считывания у 5'-конца мРНК, и последовательностью терминации транскрипции, расположенной ниже открытой рамки считывания у 3'-конца мРНК. Кодирующая последовательность может включать, не ограничиваясь ими, ДНК, кДНК и рекомбинантные последовательности нуклеиновых кислот.
Выделенную последовательность нуклеиновой кислоты, кодирующую полипептид по настоящему изобретению, можно изменять разными способами для достижения экспрессии полипептида. Изменение последовательности нуклеиновой кислоты до ее введения в вектор может быть желательно или необходимо в зависимости от экспрессирующего вектора. Методы модификации последовательностей нуклеиновых кислот при помощи методов рекомбинантных ДНК хорошо известны в данной области.
Термин "регуляторные последовательности" в используемом здесь значении означает все компоненты, которые необходимы или благоприятны для экспрессии полипептида по настоящему изобретению. Каждая регуляторная последовательность может быть нативной или чужеродной для последовательности нуклеиновой кислоты, кодирующей полипептид. Такие регуляторные последовательности включают, не ограничиваясь ими, лидерную последовательность, последовательность полиаденилирования, пропептидную последовательность, промотор, последовательность сигнального пептида и терминатор транскрипции. Регуляторные последовательности включают как минимум промотор и сигналы терминации транскрипции и трансляции. Регуляторные последовательности могут иметь линкеры для введения специфических сайтов рестрикции, облегчающих лигирование регуляторных последовательностей с кодирующей областью последовательности нуклеиновой кислоты, кодирующей полипептид. Термин "функционально связанный" в используемом здесь значении относится к конфигурации, в которой регуляторная последовательность находится в требуемом положении относительно кодирующей последовательности ДНК, позволяющем регуляторной последовательности направлять экспрессию полипептида.
Регуляторная последовательность может быть приемлемой промоторной последовательностью, представляющей последовательность нуклеиновой кислоты, которая распознается клеткой-хозяином для экспрессии последовательности нуклеиновой кислоты. Промоторная последовательность содержит последовательности, регулирующие транскрипцию, которые опосредуют экспрессию полипептида. Промотор может быть любой последовательностью нуклеиновой кислоты, которая проявляет транскрипционную активность в выбранной клетке-хозяине, включая мутантные, усеченные и гибридные промоторы, и может быть получена из генов, кодирующих внеклеточные или внутриклеточные полипептиды, гомологичные или гетерологичные клетке-хозяину.
Примерами приемлемых промоторов, используемых для направления транскрипции конструкций на основе нуклеиновых кислот по настоящему изобретению в клетке-хозяине, представляющей нитевидный грибок, являются промоторы, полученные из генов ТАКА-амилазы Aspergillus oryzae, аспарагиновой протеиназы Rhizomucor miehei, нейтральной альфа-амилазы Aspergillus niger, кислотостойкой альфа-амилазы Aspergillus niger, глюкоамилазы Aspergillus niger (glaA) или Aspergillus awamori, липазы Rhizomucor miehei, щелочной протеазы Aspergillus oryzae, триозофосфатизомеразы Aspergillus oryzae, ацетамидазы Aspergillus nidulans и трипсиноподобной протеазы Fusarium oxysporum (заявка WO 96/00787), а также промотор NA2-tpi (гибрид промоторов из генов нейтральной альфа-амилазы Aspergillus niger и триозофосфатизомеразы Aspergillus oryzae) и их мутантные, усеченные и гибридные промоторы.
Регуляторная последовательность может также быть приемлемой последовательностью терминации транскрипции, последовательностью, распознаваемой клеткой-хозяином для терминации транскрипции. Терминаторная последовательность функционально связана с 3'-концом последовательности нуклеиновой кислоты, кодирующей полипептид. В настоящем изобретении можно использовать любой терминатор, который является функциональным в выбранной клетке-хозяине.
Предпочтительные терминаторы для клеток-хозяев, представляющих нитевидные грибки, получают из генов ТАКА-амилазы Aspergillus oryzae, глюкоамилазы Aspergillus niger, антранилат-синтазы Aspergillus nidulans, альфа-глюкозидазы Aspergillus niger и трипсиноподобной протеазы Fusarium oxysporum.
Регуляторная последовательность может быть также приемлемой лидерной последовательностью, нетранслируемой областью мРНК, которая имеет важное значение для трансляции клеткой-хозяином. Лидерная последовательность функционально связана с 5'-концом последовательности нуклеиновой кислоты, кодирующей полипептид. В настоящем изобретении можно использовать любую лидерную последовательность, которая является функциональной в выбранной клетке-хозяине.
Предпочтительные лидерные последовательности для клеток-хозяев нитевидных грибков получают из генов ТАКА-амилазы Aspergillus oryzae и триозофосфатизомеразы Aspergillus nidulans.
Регуляторная последовательность может быть также последовательностью полиаденилирования, функционально связанной с 3'-концом последовательности нуклеиновой кислоты, которая в процессе трансляции распознается клеткой-хозяином в качестве сигнала о добавлении остатков полиаденозина к транскрибированной мРНК. В настоящем изобретении можно использовать любую последовательность полиаденилирования, которая является функциональной в выбранной клетке-хозяине.
Предпочтительные последовательности полиаденилирования для клеток-хозяев нитевидных грибков получают из генов ТАКА-амилазы Aspergillus oryzae, глюкоамилазы Aspergillus niger, антранилат-синтазы Aspergillus nidulans, трипсиноподобной протеазы Fusarium oxysporum и альфа-глюкозидазы Aspergillus niger.
Регуляторная последовательность может быть также кодирующей сигнальный пептид областью, которая кодирует аминокислотную последовательность, связанную с аминоконцом полипептида, и направляет кодированный полипептид в секреторный путь клетки. 5'-конец кодирующей последовательности, являющейся частью последовательности нуклеиновой кислоты, может содержать кодирующую сигнальный пептид область, естественно связанную в рамке считывания трансляции с сегментом кодирующей области, который кодирует секретируемый полипептид. Альтернативно, 5'-конец кодирующей последовательности может содержать кодирующую сигнальный пептид область, которая чужеродна кодирующей последовательности. Чужеродная область, кодирующая сигнальный пептид, может быть необходима в том случае, когда кодирующая последовательность не содержит в естественном состоянии область, кодирующую сигнальный пептид. Альтернативно, чужеродная область, кодирующая сигнальный пептид, может просто заменять естественную область, кодирующую сигнальный пептид, для усиления секреции полипептида. Однако в настоящем изобретении можно использовать любую область, кодирующую сигнальный пептид, которая направляет экспрессируемый полипептид в секреторный путь выбранной клетки-хозяина.
Эффективными областями, кодирующими сигнальный пептид, для клеток-хозяев нитевидных грибков являются кодирующие сигнальный пептид области, полученные из генов ТАКА-амилазы Aspergillus oryzae, нейтральной амилазы Aspergillus niger, глюкоамилазы Aspergillus niger, аспарагиновой протеиназы Rhizomucor miehei, целлюлазы Humicola insolens и липазы Humicola lanuginosa.
В предпочтительном варианте осуществления изобретения область, кодирующая сигнальный пептид, включает нуклеотиды 25-81 SEQ ID No.1, которые кодируют аминокислоты 1-19 SEQ ID No.2.
Регуляторной последовательностью может быть также кодирующая пропептид область, которая кодирует аминокислотную последовательность, расположенную у аминоконца полипептида. Полученный полипептид известен как пропротеаза или прополипептид (или зимоген в некоторых случаях). Прополипептид обычно является неактивным и может быть превращен в зрелый активный полипептид в результате каталитического или аутокаталитического отщепления пропептида от прополипептида. Область, кодирующую пропептид, можно получить из генов щелочной протеазы (aprE) Bacillus subtilis, нейтральной протеазы (nprT) Bacillus subtilis, альфа-фактора Saccharomyces cerevisiae, аспарагиновой протеиназы Rhizomucor miehei и лакказы Myceliophthora thermophila (заявка WO 95/33836).
В предпочтительном варианте осуществления изобретения область, кодирующая пропептид, включает нуклеотиды 82-558 SEQ ID No.1, которые кодируют аминокислоты 20-178 SEQ ID No.2.
Когда области сигнального пептида и пропептида находятся у аминоконца полипептида, область пропептида располагается рядом с аминоконцом полипептида и область сигнального пептида располагается рядом с аминоконцом области пропептида.
Кроме того, может быть желательно добавить регуляторные последовательности, которые регулируют экспрессию полипептида относительно роста клетки-хозяина. Примерами регуляторных систем являются такие системы, которые включают или выключают экспрессию гена под действием химического или физического стимула, включая наличие регуляторного соединения. Регуляторные системы в прокариотических системах включают lac-, tac- и trp-операторные системы. В дрожжах можно использовать систему ADH2 или систему GAL1. В нитевидных грибках в качестве регуляторных последовательностей можно использовать ТАКА-альфа-амилазный промотор, глюкоамилазный промотор Aspergillus niger и глюкоамилазный промотор Aspergillus oryzae. Другие примеры регуляторных последовательностей включают такие последовательности, которые обеспечивают амплификацию гена. В эукариотических системах указанные последовательности включают ген дигидрофолат-редуктазы, который амплифицирован в присутствии метотрексата, и гены металлотионеина, которые амплифицированы тяжелыми металлами. В таких случаях последовательность нуклеиновой кислоты, кодирующая полипептид, должна быть функционально связана с регуляторной последовательностью.
Экспрессирующие векторы
Настоящее изобретение относится также к рекомбинантным экспрессирующим векторам, содержащим последовательность нуклеиновой кислоты по настоящему изобретению, промотор и сигналы терминации транскрипции и трансляции. Разные описанные выше последовательности нуклеиновых кислот и регуляторные последовательности могут быть связаны друг с другом с образованием рекомбинантного экспрессирующего вектора, который может включать один или более обычных сайтов рестрикции, обеспечивающих вставку или замену последовательности нуклеиновой кислоты, кодирующей полипептид на таких сайтах. Альтернативно, последовательность нуклеиновой кислоты по настоящему изобретению может быть экспрессирована путем введения последовательности нуклеиновой кислоты или конструкции на основе нуклеиновой кислоты, включающей последовательность, в соответствующий вектор для экспрессии. При создании экспрессирующего вектора кодирующую последовательность располагают в векторе так, чтобы кодирующая последовательность была функционально связана с соответствующими регуляторными последовательностями для обеспечения экспрессии.
Рекомбинантный экспрессирующий вектор может быть любым вектором (например, плазмидой или вирусом), стандартно изменяемым методами рекомбинантных ДНК и вызывающим экспрессию последовательности нуклеиновой кислоты. Выбор вектора обычно зависит от совместимости вектора с клеткой-хозяином, в которую необходимо ввести вектор. Векторы могут быть линейными или замкнутыми кольцевыми плазмидами.
Вектор может быть автономно реплицирующим вектором, то есть вектором, который существует в виде внехромосомного элемента, репликация которого не зависит от репликации хромосом, такого как, например, плазмида, внехромосомный элемент, минихромосома или искусственная хромосома. Вектор может содержать любые элементы, обеспечивающие ауторепликацию. Альтернативно, вектор может быть таким вектором, который при введении в клетку-хозяина встраивается в геном и реплицирует вместе с хромосомой, в которую он был введен. Кроме того, можно использовать один вектор или плазмиду, два или более векторов или плазмид, которые вместе содержат полную ДНК, предназначенную для введения в геном клетки-хозяина, или транспозон.
Векторы по настоящему изобретению предпочтительно содержат один или более селектируемых маркеров, которые позволяют производить быструю селекцию трансформированных клеток. Селектируемый маркер является геном, продукт которого обладает биоцидной или вирусной устойчивостью, устойчивостью к тяжелым металлам, прототрофией к ауксотрофам и подобными свойствами. Примерами бактериальных селектируемых маркеров являются dal-гены из Bacillus subtilis или Bacillus licheniformis или маркеры, сообщающие устойчивость к антибиотикам, таким как ампициллин, канамицин, хлорамфеникол или тетрациклин. Приемлемыми маркерами для дрожжевых клеток-хозяев являются ADE2, HIS3, LEU2, LYS2, MET3, TRP1 и URA3. Селектируемые маркеры, пригодные для использования в клетке-хозяине нитевидного грибка, включают, не ограничиваясь ими, amdS (ацетамидаза), argB (орнитинкарбамоилтрансфераза), bar (фосфинотрицин-ацетилтрансфераза), hygB (гигромицин-фосфотрансфераза), niaD (нитрат-редуктаза), pyrG (оротидин-5'-фосфат-декарбоксилаза), sC (сульфат-аденилтрансфераза), trpC (антранилат-синтаза), а также их эквиваленты. В клетке Aspergillus желательно использовать гены amdS и pyrG Aspergillus nidulans или Aspergillus oryzae и ген bar Streptomyces hygroscopicus.
Векторы по настоящему изобретению предпочтительно содержат элемент(ы), который обеспечивает устойчивую интеграцию вектора в геном клетки-хозяина или автономную репликацию вектора в клетке независимо от генома.
Для интеграции в геном клетки-хозяина вектор может использовать последовательность нуклеиновой кислоты, кодирующую полипептид, или любой другой элемент вектора, обеспечивающий устойчивую интеграцию вектора в геном путем гомологичной или негомологичной рекомбинации. Альтернативно, вектор может содержать дополнительные последовательности нуклеиновых кислот, направляющие интеграцию в геном клетки-хозяина путем гомологичной рекомбинации. Дополнительные последовательности нуклеиновых кислот позволяют вектору встраиваться в геном клетки-хозяина в точное место хромосомы. Чтобы увеличить вероятность интеграции в точно определенное место, интегрирующие элементы предпочтительно должны содержать достаточное число нуклеиновых кислот, например 100-1500 пар оснований, предпочтительно 400-1500 пар оснований и наиболее предпочтительно 800-1500 пар оснований, которые высоко гомологичны соответствующей последовательности-мишени, для усиления вероятности гомологичной рекомбинации. Встроенные элементы могут быть любой последовательностью, которая гомологична последовательности-мишени в геноме клетки-хозяина. Кроме того, интегрирующие элементы могут быть некодирующими или кодирующими последовательностями нуклеиновых кислот. С другой стороны, вектор может быть встроен в геном клетки-хозяина путем негомологичной рекомбинации.
Для автономной репликации вектор может далее включать точку начала репликации, позволяющую вектору реплицировать автономно в рассматриваемой клетке-хозяине. Примерами бактериальных точек начала репликации являются точки начала репликации плазмид pBR322, pUC19, pACYC177 и pAСYC184, обеспечивающие репликацию в E. coli, и pUB110, рЕ194, pTA1060 и рАМβ1, обеспечивающие репликацию в Bacillus. Примерами точек начала репликации, предназначенных для использования в дрожжевой клетке-хозяине, являются двухмикронная точка начала репликации, ARS1, ARS4, комбинация ARS1 и CEN3 и комбинация ARS4 и CEN6. Точка начала репликации может иметь мутацию, которая заставляет ее функционировать в зависимости от температуры в клетке-хозяине (см., например, Ehrlich, 1978, Proceedings of the National Academy of Sciences USA 75:1433).
В клетку-хозяин можно ввести более одной копий последовательности нуклеиновой кислоты по настоящему изобретению для увеличения продуцирования генного продукта. Число копий последовательности нуклеиновой кислоты можно увеличить путем встраивания по крайней мере одной дополнительной копии последовательности в геном клетки-хозяина или введения амплифицируемого селектируемого гена-маркера в последовательность нуклеиновой кислоты, после чего клетки, содержащие амплифицированные копии селектируемого гена-маркера, а следовательно, и дополнительные копии последовательности нуклеиновой кислоты, могут быть отобраны в результате культивирования клеток в присутствии соответствующего селектируемого агента.
Методы, используемые для лигирования вышеописанных элементов с созданием рекомбинантных экспрессирующих векторов по настоящему изобретению, хорошо известны специалистам в данной области (см., например, Sambrook et al., 1989, приведена выше).
Протеаза по данному изобретению может быть также совместно экспрессирована по крайней мере с одним другим ферментом, используемым в корме для животных, таким как фитаза, ксиланаза, галактаназа и/или бета-глюканаза. Ферменты могут быть совместно экспрессированы из разных векторов, из одного вектора или при использовании комбинации обоих методов. При использовании разных векторов такие векторы могут иметь разные селектируемые маркеры и разные точки начала репликации. При использовании только одного вектора гены могут быть экспрессированы из одного или более промоторов. При клонировании с регуляцией одним промотором (ди- или мультицистрон) на порядок, в котором происходит клонирование генов, могут влиять уровни экспрессии белков. Протеаза может быть также экспрессирована в виде слитого белка, то есть ген, кодирующий протеазу, слит в рамке считывания с геном, кодирующим другой белок. Указанный белок может быть другим ферментом или функциональным доменом другого фермента.
Клетки-хозяева
Настоящее изобретение относится также к рекомбинантным клеткам-хозяевам, содержащим последовательность нуклеиновой кислоты по данному изобретению, которые успешно используются в рекомбинантном продуцировании полипептидов. Вектор, содержащий последовательность нуклеиновой кислоты по настоящему изобретению, вводят в клетку-хозяин так, чтобы вектор был интегрирован в хромосому или представлял собой аутореплицирующийся внехромосомный вектор, описанный выше. Термин "клетка-хозяин" означает любое потомство родительской клетки, которое неидентично родительской клетке вследствие мутаций, возникающих в процессе репликации. Выбор клетки-хозяина в значительной степени зависит от гена, кодирующего полипептид, и источника его получения.
Клетка-хозяин может быть одноклеточным микроорганизмом, например прокариотом, или многоклеточным микроорганизмом, например эукариотом.
Пригодными клетками одноклеточных микроорганизмов являются бактериальные клетки, такие как грамположительные бактерии, которые включают, не ограничиваясь ими, клетку Bacillus, клетку Streptomyces, клетки молочно-кислых бактерий; или грамотрицательные бактерии, такие как E. coli и Pseudomonas sp. Кисло-молочные бактерии включают, не ограничиваясь ими, виды родов Lactococcus, Lactobacillus, Leuconostoc, Streptococcus, Pediococcus и Enterococcus.
Вектор можно ввести в бактериальную клетку-хозяин путем трансформации протопласта (см., например, Chang and Cohen, 1979, Molecular General Genetics 168:111-115) при помощи компетентных клеток (см., например, Young and Spizizin, 1961, Journal of Bacteriology 81: 823-829, or Dubnau and Davidoff-Abelson, 1971, Journal of Molecular Biology 56: 209-221), электропорации (см., например, Shigekawa and Dower, 1988, Biotechniques 6: 742-751) или конъюгации (см., например, Koehier and Thorne, 1987, Journal of Bacteriology 169: 5771-5278).
Клетка-хозяин может быть эукариотической, такой как животная клетка, отличная от человеческой, клетка насекомого, растительная клетка или грибковая клетка.
В одном конкретном варианте осуществления изобретения клетка-хозяин является грибковой клеткой. Термин "грибки" в используемом здесь значении означает такие виды, как Ascomycota, Basidiomycota, Chytridiomycota и Zygomycota (описанные в справочнике Hawksworth et al., In Ainsworth and Bisby's Dictionary of The Fungi, 8th edition, 1995, CAB International, University Press, Cambridge, UK), а также Oomycota (описанный в справочнике Hawksworth et al., 1995, supra, page 171) и все митоспоровые грибки (Hawksworth et al., 1995, см. выше).
В другом конкретном варианте осуществления изобретения гибковая клетка-хозяин является дрожжевой клеткой. Термин "дрожжи" в используемом здесь значении означает аскоспорогенные дрожжи (Endomycetales), базидиоспорогенные дрожжи и дрожжи, относящиеся к виду Fungi Imperfecti (Blastomycetes). Так как в будущем классификация дрожжей может измениться, в целях данного изобретения дрожжи должны быть классифицированы в соответствии с определением, приведенным в Biology and Activities of Yeast (Skinner, F.A., Passmore, S.M. and Davenport, R.R. eds, Soc. App. Bacteriol. Symposium Series № 9, 1980).
Дрожжевая клетка-хозяин может быть клеткой Candida, Hansenula, Kluyveromyces, Pichia, Saccharomyces, Schizosaccharomyces или Yarrowia.
Грибковая клетка-хозяин может быть клеткой нитевидного грибка. В определение термина "нитевидные грибки" входят все нитевидные формы подтипов Eumycota и Oomycota (описанные в приведенном выше справочнике Hawksworth et al., 1995). Нитевидные грибки отличаются наличием стенки мицелия, состоящей из хитина, целлюлозы, глюкана, хитозана, маннана и других сложных полисахаридов. Вегетативный рост таких грибков происходит путем удлинения гифов, при этом диссимиляция углерода должна быть обязательно аэробной. В отличие от этого вегетативный рост дрожжей, таких как Saccharomyces cerevisiae, происходит путем почкования одноклеточного таллома, при этом диссимиляция углерода может быть ферментативной.
Примерами клеток-хозяев нитевидных грибков являются клетки нижеследующих видов, которые включают, не ограничиваясь ими, Acremonium, Aspergillus, Fusarium, Humicola, Mucor, Myceliophthora, Neurospora, Penicillium, Thielavia, Tolypocladium или Trichoderma.
Грибковые клетки могут быть трансформированы способом, включающим образование протопласта, трансформацию протопластов и регенерацию клеточной оболочки известными методами. Приемлемые методы трансформации клеток-хозяев Aspergillus описаны в ЕР №238023 и в Yelton et al., 1984, Proceedings of the National Academy of Sciences USA 81:1470-1474. Приемлемые методы трансформации видов Fusarium описаны в статье Malardier et al., 1989, Gene 78:147-156 и в заявке WO 96/00787. Дрожжи могут быть трансформированы методами, описанными в следующих документах Becker and Guarente, In Abelson, J.N. and Simon, M.I., editors, Guide to Yeast Genetics and Molecular Biology, Methods in Enzymology, Volume 194, pp. 182-187, Academic Press, Inc., New York; Ito et al., 1983, Journal of Bacteriology 153:163; and Hinnen et al., 1978, Proceedings on the National Academy of Sciences USA 75:1920.
Способы получения
Настоящее изобретение относится также к способам получения полипептида по данному изобретению, которые включают (а) культивирование штамма, который способен продуцировать полипептид в диком виде, с получением супернатанта, содержащего полипептид; и (b) выделение полипептида. В одном варианте осуществления изобретения указанный штамм относится к роду Thermoascus, например к виду Thermoascus aurantiacus, такому как штамм Thermoascus aurantiacus CGMCC № 0670.
Настоящее изобретение относится также к способам получения полипептида по данному изобретению, которые включают (а) культивирование клетки-хозяина в условиях, пригодных для продуцирования полипептида; и (b) выделение полипептида.
Настоящее изобретение относится также к способам получения полипептида по данному изобретению, которые включают (а) культивирование клетки-хозяина в условиях, пригодных для продуцирования полипептида, при этом клетка-хозяин включает мутантную последовательность нуклеиновой кислоты, содержащую по крайней мере одну мутацию в нуклеотидах 25-1089, 1-1089, 1-1344, 25-1344, 559-1344 или 559-1089 SEQ ID No.1 и кодирующую полипептид, который (i) состоит из аминокислот -178-177, -159-177 или +1-177 SEQ ID No.2, (ii) является вариантом любой последовательности (i), при этом вариант включает замену, делецию и/или инсерцию одной или более аминокислот, (iii) является аллельным вариантом любой последовательности (i) или (iv) является фрагментом любой последовательности (i).
Настоящее изобретение относится также к способам получения полипептида по данному изобретению, которые включают (а) культивирование клетки-хозяина в условиях, пригодных для продуцирования полипептида, при этом клетка-хозяин содержит мутантную последовательность нуклеиновой кислоты, имеющую по крайней мере одну мутацию в нуклеотидах SEQ ID No.10 или нуклеотидах 2-1129, 2-1195, 2-1267, 71-1129, 71-1195, 71-1267, 599-1129, 599-1195 или 599-1267 указанной последовательности или кодирующую полипептид, который (i) состоит из SEQ ID No.11 или аминокислот -23-353, -23-374, -23-397, 1-353, 1-374, 1-397, 177-353, 177-374 или 177-397 указанной последовательности.
При осуществлении способов получения по настоящему изобретению клетки культивируют в питательной среде, пригодной для получения полипептида методами, известными в данной области. Например, клетки можно культивировать во встряхиваемой колбе или путем мелкомасштабной или крупномасштабной ферментации (включая непрерывную, периодическую, порционную или твердофазную ферментацию) в лабораторных или промышленных ферментерах в приемлемой среде и в условиях, обеспечивающих экспрессию и/или выделение полипептида. Культивирование осуществляют в приемлемой питательной среде, содержащей источники углерода и азота и неорганические соли, методами, известными в данной области. Приемлемые среды можно приобрести у коммерческих поставщиков или получить, используя опубликованную рецептуру (например, в каталогах Американской коллекции типовых культур). Полипептид, секретируемый в питательную среду, можно выделить непосредственно из среды. Несекретируемый полипептид можно выделить из клеточных лизатов.
Полипептиды можно обнаружить методами, известными в данной области, которые являются специфическими для полипептидов. Такие методы обнаружения могут включать использование специфических антител, образование протеазного продукта или удаление протеазного субстрата. Например, анализ протеазы можно использовать для определения активности описанного полипептида.
Полученный полипептид можно выделить методами, известными в данной области. Например, полипептид можно выделить из питательной среды обычными методами, которые включают, не ограничиваясь ими, центрифугирование, фильтрацию, экстракцию, сушку распылением, упаривание или осаждение.
Полипептиды по настоящему изобретению можно очистить разными методами, известными в данной области, которые включают, не ограничиваясь ими, хроматографию (например, ионообменную, аффинную, гидрофобную, хроматофокусирующую и вытеснительную), электрофорез (например, препаративное изоэлектрическое фокусирование), дифференциальную солюбилизацию (например, осаждение сульфатом аммония), SDS-PAGE или экстракцию (см., например, Protein Purification, J.-C. Janson and Lars Ryden, editors, VCH Publishers, New York, 1989).
Растения
Настоящее изобретение относится также к трансгенному растению, части растения или растительной клетке, которые были трансформированы последовательностью нуклеиновой кислоты, кодирующей полипептид, обладающий протеазной активностью, по настоящему изобретению для экспрессии и продуцирования полипептида в выделяемых количествах. Полипептид может быть выделен из растения или части растения. Альтернативно, растение или часть растения, содержащие рекомбинантный полипептид, можно использовать в том виде, как есть, для улучшения качества пищи или корма, например, для повышения пищевой ценности, улучшения вкусовых качеств и реологических свойств или для разрушения несъедобного фактора.
В конкретном варианте осуществления изобретения полипептид направленно воздействует на вакуоли семян, содержащие эндосперм. Такой результат достигается в результате синтеза данного полипептида в качестве предшественника с приемлемым сигнальным пептидом, как это описано в статье Horvath et al., in PNAS, Feb. 15, 2000, vol. 97, no. 4, p. 1914-1919.
Трансгенное растение может быть двудольным или однодольным. Примерами однодольных растений являются травы, такие как мятлик луговой (голубая трава, Роа), кормовая трава, такая как festuca, lolium, полевица, такая как Agrostis, и злаки, например, пшеница, овес, рожь, ячмень, рис, сорго и маис (кукуруза). Примерами двудольных растений являются табак, растения из семейства бобовых, такие как люпин, картофель, сахарная свекла, горох, фасоль и соя, и крестоцветные растения (семейство Brassicaceae), такие как капуста цветная, рапс и близко родственное растение Arabidopsis thaliana. Растения с низким содержанием фитата, описанные в патентах США №5689054 и 6111168, являются примерами генетически созданных растений.
Примерами частей растений являются стебель, каллюс, листья, корень, плоды, семена и клубни. Частями растения считаются также специфические растительные ткани, такие как хлоропласт, апопласт, митохондрия, вакуоль, пероксисомы и цитоплазма. Кроме того, любая растительная клетка независимо от происхождения ткани считается частью растения.
В объем настоящего изобретения входит также потомство таких растений, частей растений и растительных клеток.
Трансгенное растение или растительная клетка, экспрессирующая полипептид по настоящему изобретению, может быть создана методами, известными в данной области. Вкратце, растение или растительную клетку получают путем введения в геном растения-хозяина одной или более экспрессирующих конструкций, кодирующих полипептид по настоящему изобретению, и размножения полученного модифицированного растения или растительной клетки с получением трансгенного растения или растительной клетки.
Экспрессирующей конструкцией обычно является конструкция на основе нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты, кодирующую полипептид по настоящему изобретению, которая функционально связана с соответствующими регуляторными последовательностями, необходимыми для экспрессии последовательности нуклеиновой кислоты в выбранном растении или части растения. Кроме того, экспрессирующая конструкция может включать селектируемый маркер, пригодный для идентификации клеток-хозяев, в которые встроена данная экспрессирующая конструкция, и последовательности ДНК, необходимые для введения конструкции в данное растение (последнее условие зависит от используемого метода введения ДНК).
Выбор регуляторных последовательностей, таких как промоторные и терминаторные последовательности, и необязательно сигнальных или транзитных последовательностей определяется с учетом того, когда, где и как данный полипептид должен быть экспрессирован. Например, экспрессия гена, кодирующего полипептид по настоящему изобретению, может быть конститутивной, индуцируемой или специфической для стадии развития или ткани, причем генный продукт может направленно воздействовать на определенную ткань или часть растения, такую как семена или листья. Регуляторные последовательности описаны, например, в Tague et al., 1988, Plant Physiology 86:506.
Для конститутивной экспрессии можно использовать промотор 35S-CaMV (Franck et al., 1980, Cell 21:285-294). Органспецифическим промотором может быть, например, промотор из запасающих тканей, таких как семена, клубни картофеля и плоды (Edwards & Coruzzi, 1990, Ann. Rev. Genet. 24: 275-303), или метаболических тканей, таких как меристемы (Ito et al., 1994, Plant Mol. Biol. 24:863-878), семяспецифический промотор, такой как промотор риса, глутелина, проламина, глобулина или альбумина (Wu et al., 1998, Plant and Cell Physiology 39:885-889), промотор Vicia faba из легумина В4 и неизвестный ген семенного белка из Vicia faba (Conrad et al., 1998, Journal of Plant Physiology 152:708-711), промотор из белка масляного тельца семени (Chen et al., 1998, Plant and Cell Physiology 39:935-941), промотор napA запасающего белка из Brassica napus или любой другой семяспецифический промотор, известный в данной области, например, описанный в заявке WO 91/14772. Кроме того, промотор может быть листспецифическим промотором, таким как промотор rbcs риса или томата (Kyozuka et al., 1993, Plant Physiology 102:991-1000), промотор гена аденинметилтрансферазы вируса хлореллы (Mitra and Higgins, 1994, Plant Molecular Biology 26:85-93), промотор гена aldP риса (Kagaya et al., 1995, Molecular and General Genetics 248:668-674) или индуцируемый повреждением промотор, такой как промотор pin2 картофеля (Xu et al., 1993, Plant Molecular Biology 22:573-588).
Можно также использовать энхансерный элемент промотора для достижения более высокой экспрессии протеазы в растении. Например, энхансерным элементом промотора может быть интрон, который вводят между промотором и нуклеотидной последовательностью, кодирующей полипептид по настоящему изобретению. В приведенной выше статье Xu et al., 1993, описано использование первого интрона гена актина 1 риса для усиления экспрессии.
Кроме того, для улучшения экспрессии можно оптимизировать использование кодона для рассматриваемых видов растений (см. приведенную выше статью Horvath et al.).
Селектируемый ген-маркер и любые другие части экспрессирующей конструкции могут быть выбраны из имеющихся в данной области.
Конструкцию на основе нуклеиновой кислоты встраивают в геном растения обычными методами, известными в данной области, включая Agrobacterium-опосредуемую трансформацию, вирусопосредуемую трансформацию, микроинъекцию, бомбардировку частицами, биолистическую трансформацию и электропорацию (Gasser et al., 1990, Science 244:1293; Potrykus, 1990, Bio/Technology 8:535; Shimamoto et al., 1989, Nature 338:274).
В настоящее время наиболее часто используемым методом создания трансгенных двудольных растений является перенос гена, опосредуемый Agrobacterium tumefaciens (для ознакомления с данным методом см. Hooykas and Schilperoort, 1992, Plant Molecular Biology 19:15-38). Однако указанный метод можно также использовать для трансформации однодольных растений, хотя для таких растений обычно используют другие методы трансформации. В настоящее время наиболее часто используемым методом создания трансгенных однодольных растений является бомбардировка частицами (микроскопическими частицами золота или вольфрама, покрытыми трансформирующей ДНК) эмбриональных каллюсов или развивающихся зародышей (Christou, 1992, Plant Journal 2:275-281; Shimamoto, 1994, Current Opinion Biotechnology 5:158-162; Vasil et al., 1992, Bio/Technology 10:667-674). Альтернативным методом трансформации однодольных растений является трансформация протопласта, описанная в статье Omirulleh et al., 1993, Plant Molecular Biology 21:415-428.
Полученные в результате трансформации трансформанты, содержащие экспрессирующую конструкцию, отбирают и регенерируют из них цельные растения методами, хорошо известными в данной области.
Настоящее изобретение относится также к способам получения полипептида по данному изобретению, которые включают (а) культивирование трансгенного растения или растительной клетки, содержащей последовательность нуклеиновой кислоты, кодирующую полипептид, обладающий протеазной активностью, по настоящему изобретению в условиях, пригодных для продуцирования полипептида; и (b) выделение полипептида.
Животные
Настоящее изобретение относится также к трансгенным животным кроме человека и их продуктам или элементам, примерами которых являются жидкости организма, такие как молоко и кровь, органы, плоть и животные клетки. Методы экспрессии белков, например, в клетках млекопитающих известны в данной области; см., например, справочник Protein Expression: A Practical Approach, Higgins and Hames (eds.), Oxford University Press (1999) и три других справочника в данной серии, относящиеся к транскрипции гена, процессингу РНК и посттрансляционному процессингу. В целом, чтобы получить трансгенное животное, отобранные клетки выбранного животного трансформируют последовательностью нуклеиновой кислоты, кодирующей полипептид, обладающий протеазной активностью, по настоящему изобретению, так, чтобы экспрессировать и продуцировать полипептид. Полипептид может быть получен у животного, например выделен из молока самки животного, или полипептид может быть экспрессирован с пользой для самого животного, например для улучшения пищеварения животного. Примеры животных приведены ниже в разделе, озаглавленном "Корм для животных".
Чтобы получить трансгенное животное с целью выделения протеазы из молока животного, ген, кодирующий протеазу, можно ввести в оплодотворенные яйцеклетки исследуемого животного, например, при помощи вектора, экспрессирующего трансген, который содержит приемлемый промотор белка молока и ген, кодирующий протеазу. Вектор, экспрессирующий трансген, микроинъекцируют в оплодотворенные яйцеклетки и предпочтительно стабильно встраивают в хромосому. Как только яйцеклетка начинает расти и делиться, потенциальный эмбрион имплантируют суррогатной матери и идентифицируют животных, несущих трансген. Полученное животное затем можно размножить обычными методами разведения. Полипептид может быть очищен от молока животного методами, описанными, например, в публикации Meade, H.M. et al. (1999): Expression of recombinant proteins in the milk of transgenic animals, Gene expression systems: Using nature for the art of expression, J. M. Fernandez and J. P. Hoeffler (eds.). Academic Press.
В качестве альтернативы получения трансгенного животного кроме человека, несущего в геноме соматических и/или зародышевых клеток последовательность нуклеиновой кислоты, включающую гетерологичную трансгенную конструкцию, содержащую трансген, кодирующий протеазу, трансген можно функционально связать с первой регуляторной последовательностью для специфической экспрессии протеазы в слюнной железе, как это описано в заявке WO 2000064247.
Композиции
Настоящее изобретение относится также к композициям, содержащим полипептид по настоящему изобретению.
Полипептидные композиции можно получить методами, известными в данной области, в виде жидкой или сухой композиции. Например, полипептидная композиция может иметь форму гранул или микрогранул. Полипептид, вводимый в такую композицию, может быть стабилизирован методами, известными в данной области.
В конкретном варианте осуществления изобретения полипептидные композиции по данному изобретению могут быть защищены от разложения под действием кислоты, например, для лучшего сохранения полипептида при прохождении через желудок животного. Защита может представлять кислотостойкое покрытие, примеры которого можно найти в фармацевтических справочниках.
Ниже приведены примеры предпочтительных применений полипептидов или полипептидных композиций по данному изобретению.
Корм для животных
Настоящее изобретение относится также к способам применения полипептидов, обладающих протеазной активностью, в корме для животных, а также к кормовым композициям и кормовым добавкам, содержащим полипептиды по данному изобретению.
Термин "животное" означает всех животных, включая человека. Примерами животных являются нежвачные животные и жвачные животные, такие как коровы, овцы и лошади. В конкретном варианте осуществления изобретения такое животное является нежвачным животным. Нежвачные животные включают животных с однокамерным желудком, например поросят или свиней (включая, но не ограничиваясь ими, новорожденных поросят, растущих поросят и свиноматок), домашнюю птицу, такую как индейки и куры (включая, но не ограничиваясь ими, бройлерных цыплят, несушек); телят и рыбу (включая лосося, но не ограничиваясь им).
Термин "корм" или "кормовая композиция" означает любое соединение, препарат, смесь или композицию, пригодную или предназначенную для скармливания животному.
В соответствии с целями данного изобретения протеазу можно скармливать животному до, после или одновременно с кормом. Последнее является предпочтительным.
В конкретном варианте осуществления изобретения протеаза в той форме, в какой ее добавляют в корм или кормовую добавку, должна иметь определенный состав. Термин "определенный состав" означает, что протеазный препарат должен быть чистым по крайней мере на 50% по результатам вытеснительной хроматографии (см. пример 12 заявки WO 01/58275). В других конкретных вариантах осуществления изобретения протеазный препарат должен быть чистым по крайней мере на 60%, 70%, 80%, 85%, 88%, 90%, 92%, 94% или 95% при определении вышеуказанным методом.
Протеазный препарат определенного состава является предпочтительным. Например, гораздо легче точно дозировать протеазу, вводимую в корм, которая по существу не загрязнена другими протеазами. Термин "точное дозирование" означает, в частности, получение сопоставимых и постоянных результатов и возможность оптимизировать дозу с учетом достижения требуемого эффекта.
Однако для использования в корме для животных данная протеаза необязательно должна быть совершенно чистой; она может включать другие ферменты, представляя в данном случае протеазный препарат.
Протеазный препарат можно (а) добавлять непосредственно в корм (или использовать непосредственно в процессе обработки растительных белков) или (b) использовать для получения одной или более промежуточных композиций, таких как кормовые добавки или премиксы, которые затем добавляют в корм (или используют в процессе обработки). Вышеуказанная степень чистоты относится к чистоте исходного протеазного препарата, используемого в соответствии с пунктами (а) или (b).
Протеазные препараты с чистотой указанного порядка величины получают, в частности, методами рекомбинантных ДНК, так как такие препараты труднее получить и они характеризуются более высокими расхождениями между партиями при получении традиционными методами ферментации.
Такой протеазный препарат может быть смешан с другими ферментами.
В другом конкретном варианте осуществления изобретения протеаза, используемая в соответствии с данным изобретением, может:
увеличивать количество расщепленного белка, степень гидролиза (DH) и/или количество солюбилизованного белка в полной модели пищеварения с однокамерным желудком in vitro, описанной в нижеследующем примере 6;
увеличивать количество расщепленного белка, DH и/или количество солюбилизованного белка в кишечном отделе модели пищеварения с однокамерным желудком in vitro, описанной в нижеследующем примере 6; и/или
увеличивать растворимость белка, расщепляемость белка и/или DH в модели пищеварения аквакультуры in vitro, описанной в нижеследующем примере 7.
Термин "растительные белки" в используемом здесь значении означает любое соединение, композицию, препарат или смесь, которые включают по крайней мере один белок, выделенный или происходящий из растения, включая модифицированные белки и производные белков. В конкретных вариантах осуществления изобретения содержание растительных белков составляет по крайней мере 10%, 20%, 30%, 40%, 50% или 60% (мас./мас.).
Растительные белки могут быть получены из таких источников растительного белка, как бобовые и злаки, например растения семейств Fabaceae (Leguminosae), Cruciferaceae, Chenopodiaceae и Poaceae, в частности соевая, люпиновая и рапсовая мука.
В конкретном варианте осуществления изобретения источником растительного белка являются растения семейства Fabaceae, например соя, люпин, горох или фасоль.
В другом конкретном варианте осуществления изобретения источником растительного белка являются одно или более растений семейства Chenopodiaceae, например свекла, сахарная свекла, шпинат или квиноя.
Другими примерами источников растительного белка являются рапсовое семя и капуста.
Соя является предпочтительным источником растительного белка.
Другими примерами источников растительного белка являются злаки, такие как ячмень, пшеница, рожь, овес, маис (кукуруза), рис и сорго.
Обработка растительных белков по крайней мере одной протеазой по данному изобретению повышает солюбилизацию растительных белков.
Ниже приведены примеры количества солюбилизованного белка в процентах, полученного при использовании протеаз по данному изобретению: по крайней мере 100,0%, 100,1%, 100,2%, 100,4%, 100,5%, 100,6%, 100,7%, 100,8%, 100,9%, 101,0%, 101,1%, 101,2%, 101,3% или 101,4% по сравнению с контрольным и ссылочным образцом при использовании полной модели пищеварения с однокамерным желудком in vitro по примеру 6.
Ниже приведены примеры количества расщепляемого белка в процентах, полученного при использовании протеаз по данному изобретению: по крайней мере 100% или 101%, 102%, 103% или 104% по сравнению с контрольным и ссылочным образцом при использовании полной модели пищеварения с однокамерным желудком in vitro по примеру 6.
Ниже приведены примеры i) % солюбилизованного и ii) % расщепленного белка, полученного с использованием протеаз по данному изобретению: по крайней мере 100%, 101%, 102% или 103% по сравнению с контрольным и ссылочным образцом при использовании кишечного отдела модели пищеварения с однокамерным желудком in vitro по примеру 6.
Ниже приведены примеры степеней гидролиза (DH) в процентах, полученных при использовании протеаз по данному изобретению: по крайней мере 100%, 101%, 102%, 103%, 104%, 105%, 106%, 107%, 108%, 109%, 110%, 111% или 112% по сравнению с контрольным и ссылочным образцом при использовании модели пищеварения аквакультуры in vitro по примеру 7.
Ниже приведены примеры i) % солюбилизованного и ii) % расщепленного белка, полученного с использованием протеаз по данному изобретению: по крайней мере 100%, 101%, 102%, 103%, 104%, 105%, 106%, 107%, 108%, 109%, 110%, 111%, 112% или 113% по сравнению с контрольным и ссылочным образцом при использовании модели пищеварения аквакультуры in vitro по примеру 7.
Термин "солюбилизация белков" в основном означает перевод белков в раствор. Такая солюбилизация может происходить вследствие опосредуемого протеазой высвобождения белка из других компонентов комплексных природных композиций, таких как корм. Солюбилизацию можно измерить по увеличению количества солюбилизованных белков по сравнению с контрольными образцами, обработанными только пепсином и панкреатином (см. примеры 6 и 7).
Термин "расщепление белков" в основном означает протеолитический гидролиз солюбилизованных белков под действием протеаз, в результате которого образуются низкомолекулярные пептиды и аминокислоты. Расщепление солюбилизованных белков можно измерить по увеличению количества солюбилизованных пептидов и аминокислот с молекулярной массой, равной 1500 Дальтон или меньше, по сравнению с контрольными образцами, обработанными только пепсином и панкреатином (см. примеры 6 и 7).
В конкретном варианте осуществления способа обработки рассматриваемая протеаза(ы) воздействует (или оказывает солюбилизирующее действие) на растительные белки или источники белка. Для достижения такого действия растительный белок или источник белка обычно суспендируют в растворителе, например водном растворителе, таком как вода, и значения рН и температуры регулируют с учетом характеристик исследуемого фермента. Например, обработка может быть произведена при таком значении рН, при котором активность фактической протеазы равна по крайней мере 50%, 60%, 70%, 80% или 90%. Аналогичным образом, например, указанную обработку можно производить при такой температуре, при которой активность фактической протеазы равна по крайней мере 50%, 60%, 70%, 80% или 90%. Приведенные выше процентные показатели активности относятся к максимальным активностям. Ферментативная реакция продолжается до достижения требуемого результата, после чего она может быть прекращена или не прекращена инактивацией фермента, например, в результате выполнения стадии тепловой обработки.
В другом конкретном варианте осуществления способа обработки по данному изобретению действие протеазы может быть отсрочено, вследствие чего, например, протеазу добавляют к растительным белкам или источникам белка, но она оказывает солюбилизирующее действие, или, иными словами, запускается, позже при создании приемлемых условий солюбилизации, или при инактивации ингибиторов фермента, или при использовании любого другого средства, замедляющего действие фермента.
В одном варианте осуществления изобретения обработка является предварительной обработкой корма для животных или растительных белков, предназначенных для использования в корме для животных, то есть белки солюбилизируют до их поглощения.
Термин "повышение пищевой ценности корма для животных" означает лучшую доступность белков, в результате чего увеличивается экстракция белка, достигается более высокий выход белка и/или улучшается утилизация белков. Таким образом повышается пищевая ценность корма и улучшаются показатели скорости роста, привеса и/или превращения корма (то есть отношение массы потребленного корма к привесу) у животного.
Протеазу можно добавлять к корму в любой форме, например в виде относительно чистой протеазы или в смеси с другими компонентами, добавляемыми в корм для животных, то есть в форме добавок в корм для животных, таких как так называемые премиксы корма для животных.
Другим объектом настоящего изобретения являются композиции, предназначенные для использования в корме для животных и добавках к корму для животных, например премиксы.
Помимо протеазы по данному изобретению добавки к корму для животных по данному изобретению содержат по крайней мере один жирорастворимый витамин, по крайней мере один водорастворимый витамин, и/или по крайней мере один микроминерал, и/или по крайней мере один макроминерал.
Кроме того, необязательными ингредиентами, добавляемыми в корм, являются красители, ароматизаторы, стабилизаторы, антимикробные пептиды и/или по крайней мере один другой фермент, выбираемый из таких ферментов, как фитазы ЕС 3.1.3.8 или 3.1.3.26; ксиланазы ЕС 3.2.1.8; галактаназы ЕС 3.2.1.89 и/или бета-глюканазы ЕС 3.2.1.4.
В конкретном варианте осуществления изобретения указанные другие ферменты являются хорошо известными ферментами (описанными выше для протеазных препаратов).
Примерами антимикробных пептидов (АМР) являются САР18, лейкоцин А, тритптицин, протегрин-1, танатин, дефензин и овиспирин, такой как новиспирин (Robert Lehrer, 2000), плектазины и статины, включая соединения и полипептиды, описанные в РСТ/DK02/00781 и PCT/DK02/00812, а также варианты или фрагменты вышеуказанных веществ, обладающие антимикробной активностью.
Примерами противогрибковых полипептидов (AFP) являются пептиды Aspergillus giganteus и Aspergillus niger, а также их варианты и фрагменты, обладающие противогрибковой активностью, описанные в заявках WO 94/01459 и WO 02/090384.
Жиро- и водорастворимые витамины и микроминералы обычно составляют часть так называемого премикса, предназначенного для добавления к корму, в то время как макроминералы добавляют в корм отдельно. Любая из указанных композиций, обогащенных протеазой по данному изобретению, является добавкой к корму для животных по данному изобретению.
В конкретном варианте осуществления изобретения добавку к корму для животных по данному изобретению вводят (или предписывают для введения) в рацион или корм для животных в количестве от 0,01 до 10,0%, более конкретно от 0,05 до 5,0% или от 0,2 до 1,0% (% означает количество добавки в граммах на 100 г корма). Указанные количества относятся также к премиксам.
Ниже приведены неограничивающие примеры указанных компонентов.
Примерами жирорастворимых витаминов являются витамин А, витамин D3, витамин Е и витамин К, например витамин К3.
Примерами водорастворимых витаминов являются витамин В12, биотин и холин, витамин В1, витамин В2, витамин В6, ниацин, фолиевая кислота и пантотенат, например Са-D-пантотенат.
Примерами микроминералов являются марганец, цинк, железо, медь, иод, селен и кобальт.
Примерами макроминералов являются кальций, фосфор и натрий.
Пищевые требования, предъявляемые к указанным компонентам (на примере домашней птицы и поросят/свиней), приведены в таблице А заявки WO 01/58275. Термин "пищевые требования" означает, что указанные компоненты должны присутствовать в рационе в указанных концентрациях.
В качестве альтернативы следует отметить, что добавка к корму для животных по данному изобретению включает по крайней мере один из компонентов, указанных в таблице А заявки WO 01/58275. Термин "по крайней мере один" означает один или более, например один, два, три, четыре и так далее до тринадцати или пятнадцати, отдельных компонентов. Более конкретно, по крайней мере один отдельный компонент вводят в добавку по данному изобретению в таком количестве, которое необходимо для достижения концентрации в корме в пределах диапазона, указанного в четвертом, пятом или шестом столбце таблицы А.
Настоящее изобретение относится также к композициям корма для животных. Композиции или рационы корма для животных характеризуются относительно высоким содержанием белка. Рационы для домашней птицы и поросят могут быть аналогичны приведенным в таблице В заявки WO 01/58275, столбцы 2-3. Рационы для рыбы могут быть аналогичны приведенным в 4-м столбце таблицы В. Кроме того, такие рационы для рыбы обычно включают неочищенный жир в количестве 200-310 г/кг.
Композиция корма для животных по данному изобретению содержит сырой белок в количестве 50-800 г/кг, а также по крайней мере одну протеазу по данному изобретению.
Кроме того или в качестве альтернативы (вышеуказанному содержанию сырого белка), композиция корма для животных по данному изобретению характеризуется обменной энергией, равной 10-30 МДж/кг, содержит кальций в количестве 0,1-200 г/кг, доступный фосфор в количестве 0,1-200 г/кг, и/или метионин в количестве 0,1-100 г/кг, и/или метионин с цистеином в количестве 0,1-150 г/кг, и/или лизин в количестве 0,5-50 г/кг.
В конкретных вариантах осуществления изобретения величина обменной энергии и содержание сырого белка, кальция, фосфора, метионина, метионина с цистеином и/или лизина соответствуют одному из диапазонов 2, 3, 4 или 5, приведенных в таблице B заявки WO 01/58275 (R. 2-5).
Сырой белок высчитывают, умножая количество азота (N) на коэффициент 6,25, то есть сырой белок (г/кг) = N (г/кг) х 6,25. Содержание азота определяют методом Kjeldahl (A.O.A.C., 1984, Official Methods of Analysis, 14th ed. Association of Official Analytical Chemists, Washington DC).
Величину обменной энергии можно вычислить на основании данных, приведенных в публикациях NRC "Nutrient Requirements in Swine", ninth revised edition 1988, subcommittee on swine nutrition, committee on animal nutrition, board of agriculture, national research council. National Academy Press, Washington, D.C., pp. 2-6, и the European Table of Energy Values for Poultry Feed-stuffs, Spelderholt centre for poultry research and extension, 7361 DA Beekbergen, The Netherlands, Grafisch bedrijf Ponsen & Iooijen bv, Wageningen. ISBN 90-71463-12-5.
Содержание кальция, доступного фосфора и аминокислот в полных рационах для животных вычисляют на основании кормовых таблиц, таких как Veevoedertabel 1997, gegevens over chemische samenstelling, verteerbaarheid en voederwaarde van voedermiddelen, Central Veevoederbureau, Runderweg 6, 8219 pk Lelystad. ISBN 90-72839-13-7.
В конкретном варианте осуществления изобретения композиция корма для животных по данному изобретению содержит по крайней мере один вышеуказанный растительный белок или источник белка.
В других конкретных вариантах осуществления изобретения композиция корма для животных по данному изобретению содержит 0-80% кукурузы, 0-80% сорго, 0-70% пшеницы, 0-70% ячменя, 0-30% овса, 0-40% соевой муки: 0-10% рыбной муки и/или 0-20% сыворотки.
Корм для животных можно получить в виде дробленого корма (негранулированного) или гранулированного корма. Измельченные корма обычно смешивают и добавляют достаточные количества незаменимых витаминов и минералов в соответствии с требованиями, предъявляемыми данным видом животных. Ферменты можно добавлять в виде твердых или жидких ферментных препаратов. Например, твердый ферментный препарат обычно добавляют до или во время стадии смешивания; и жидкий ферментный препарат обычно добавляют после стадии гранулирования. Фермент можно также вводить в кормовой добавке или премиксе.
Конечная концентрация фермента в рационе находится в пределах 0,01-200 мг белка фермента на кг рациона, например в диапазоне 5-30 мг белка фермента на кг корма для животных. В конкретных вариантах осуществления изобретения концентрация фермента в корме составляет 0,1-180, 0,1-160, 0,1-150, 0,2-125, 0,2-100, 1-180, 2-160, 5-150, 10-100, 10-90, 10-80, 10-70 или 10-50 мг белка фермента на кг корма для животных.
В конкретных вариантах применения способа по данному изобретению для обработки растительных белков выполняют дополнительную стадию добавления фитазы. В других конкретных вариантах осуществления изобретения помимо совместного добавления фитазы и протеазы в композицию могут быть также введены другие ферменты, причем указанные ферменты выбирают из группы, включающей другие протеазы, фитазы, липолитические ферменты и ферменты глюкозидазы/карбогидразы. Примеры таких ферментов приведены в заявке WO 95/28850.
Протеазу, конечно, необходимо вводить в эффективном количестве, то есть в количестве, необходимом для улучшения солюбилизации и/или повышения пищевой ценности корма. В настоящее время считается, что указанный фермент нужно вводить в одном или более нижеследующих количеств (диапазоны доз), равных 0,01-200, 0,01-100, 0,05-100, 0,05-50 или 0,10-10, выраженных в мг белка протеазы на кг корма (частях на миллион).
Для определения содержания белка протеазы в мг на кг корма протеазу очищают от композиции корма и определяют удельную активность очищенной протеазы при помощи соответствующего анализа (см. в разделе "Протеазная активность, субстраты и анализы"). Протеазную активность композиции корма также определяют при помощи вышеуказанного анализа и на основании полученных двух определений вычисляют дозу белка протеазы в мг на кг корма.
Аналогичные принципы применяют к определению содержания белка протеазы в мг в кормовых добавках. При наличии образца протеазы, используемой для получения кормовой добавки или корма, удельную активность определяют на основании данного образца (не нужно очищать протеазу от композиции корма или добавки).
Настоящее изобретение далее будет описано со ссылкой на нижеследующие примеры, которые не следует рассматривать как ограничивающие объем данного изобретения.
Примеры
Реагенты, среды и оборудование
Реагенты
За исключением особо оговоренных случаев используемые химикаты являются коммерчески производимыми продуктами по крайней мере с чистотой реактивов.
AZCL-казеин (Megazyme, № по каталогу I-AZCAS). Для анализа эндопротеазы используют сшитый азурином казеин, полученный путем окрашивания и сшивания высоко очищенного казеина. Указанный казеин поставляется в виде тонко измельченного порошка, который не растворим в буферном растворе, но быстро гидратирует с образованием частиц геля, которые легко и быстро гидролизуются эндопротеазами, высвобождая растворимый меченный красителем фрагмент.
pNA-субстраты:
(AAPF) Ala-Ala-Pro-Phe-pNa (Sigma S-7388)
(GGF) Gly-Gly-Phe-pNa (Sigma S-1899)
(FGL) Phe-Gly-Leu-pNA (Sigma S-6768)
Ba-pNA (Sigma B-4875)
(AAPE) Ala-Ala-Pro-Glu-pNa (BACHEM C-1710)
SSI (ингибитор субтилизина Streptomyces): SSI был очищен от ферментирующего супернатанта Streptomyces albogriseolus FERM P-1205 (S-3253) при помощи хроматографии. Препарат SSI имел чистоту более 95% при определении методом SDS-PAGE. Альтернативно SSI можно приобрести в фирме Wako в Японии, № по каталогу 303-05201, производства компании Daiwa Kasei K.K. (см., например:
http://search.wako-chem.co.jp/lifedb_e/lifedocs_e/44834.asp).
EDTA (Gibco BRL, № по каталогу 15576-028)
IPTG (Promega, № по каталогу V3951)
Х-gal (Promega, № по каталогу V3941)
Натриевая соль ампициллина (GIBCOL., № по каталогу 11593-019)
LMP-агароза (Promega, № по каталогу V2111)
Название "трипсин" служит для обозначения трипсиноподобной протеазы, полученной из штамма Fusarium oxysporum. Получение указанного вещества описано в ЕР №471265, примеры 1 и 2 ("протеаза II").
SAVINASE™ является субтилизин-протеазой, полученной из Bacillus clausii (ранее именуемой Bacillus lentus NCIB 10309), которая коммерчески производится фирмой Novozymes A/S, Krogshoejvej, DK-2880 Bagsvaerd, Denmark. Получение указанного вещества описано в патенте США №3723250.
Пепсин (Sigma P-7000; 539 ед/мг, твердое вещество)
Панкреатин (Sigma P-7545; 8xU.S.P (фармакопея США))
Цитохром С (12500 Дальтон, Boehringer Mannheim 103870)
Апротинин (6500 Дальтон, Sigma A-1153)
Гастрин I (2126 Дальтон, Sigma G-1276)
Вещество Р (1348 Дальтон, Sigma S-6883)
Среды
Буферная система (рН 3-рН 11): 100 мМ янтарной кислоты, 100 мМ HEPES, 100 мМ CHES, 100 мМ CABS, 1 мМ CaCl2, 150 мМ KCl, 0,01% Тритон® Х-100 со значениями рН, доведенными до 2,0, 2,5, 3,0, 3,5, 4,0, 4,5, 5,0, 6,0, 7,0, 8,0, 9,0, 10,0 или 11,0 при помощи HCl или NaOH (далее кратко именуется "буферная система с янтарной кислотой").
Бура/NaH2PO4, рН 9: получена в результате смешивания 8,25 мл 0,05 М буры с 1,75 мл 0,1 М NaH2PO4.
1 х буфер ТАЕ: 0,04 М трис-ацетат; 0,001М EDTA.
0,5 х буфер ТВЕ: 0,045 М трис-борат; 0,001 М EDTA.
1 мМ pNa-субстратов: взвешивают 5 мг pNa-субстрата и растворяют в 100 мкл ДМСО, затем разбавляют 10 мл смеси бура/NaH2PO4, буфер с pH 9,0.
Среда СВН1:
80 мл помещают в 500 мл колбы Эрленмейера и обрабатывают в автоклаве в течение 20 минут при 121°С.
Жидкая среда LB: к 950 мл деионизированной Н2О добавляют 10 г бактотриптона, 5 г бакто-дрожжевого экстракта, 10 г NaCl. Встряхивают до растворения растворяющихся веществ. Доводят до рН 7,0, добавляя 5 н. раствор NaOH (˜0,2 мл). Объем раствора доводят до 1 литра, добавляя де-ионизированную Н2О. Смесь стерилизируют в автоклаве в течение 20 минут под давлением 1 атм (15 фунтов/кв.дюйм) в жидком цикле.
Планшеты для LB с ампициллином/IPTG/X-Gal: 15 г агара добавляют к одному литру среды LB. Вводят ампициллин в конечной концентрации 100 мкг/мл, затем добавляют 0,5 мМ IPTG и 80 мкг/мл X-gal и выливают на планшеты.
1% LMP-агарозный гель: 1 г LMP-агарозы добавляют к 100 мл однократного объема буфера ТАЕ.
Исходный раствор IPTG (0,1 М):
Дистиллированную воду добавляют к 1,2 г IPTG до достижения конечного объема, равного 50 мл, стерилизуют через фильтр и хранят при 4°С.
Рацион на основе кукурузы/SBM (SBM = соевая мука): соотношение кукурузы и SBM равно 6:4 (мас./мас.), содержание белка равно 43% (мас./мас.) в SBM и 8,2% (мас./мас.) в кукурузной муке. Общее количество белка в 10 г рациона на основе кукурузы/SBM равно 2,21 г.
Вытесняемая SBM: содержание белка равно 45%. Общее количество белка в 10 г вытесняемой SBM равно 4,5 г.
Оборудование, включающее разные наборы
Мембрана 5К (Millipore BIOMAX-5, 13442AM)
Фильтр 0,45 мкм (Nalgene 190-2545)
Колонка Q Sepharose FF (Amersham Pharmacia 17-0510-01)
Колонка Superdex75 (Amersham Pharmacia 17-1047-01)
IEF-гель (Amersham Pharmacia 80-1124-80)
Термосмеситель Comfort (Eppendorf)
Мининабор RNeasy (QIAGEN, № по каталогу 74904)
Набор 3' RACE (GIBCO, № по каталогу 18373-019), включающий затравку Adapter и смесь dNTP AUAP (100 мМ, Promega, № по каталогу U1330)
Система TaqДНК-полимеразы (Promega, № по каталогу М1661), включающая буфер для PCR (200 мМ трис-HCl (рН 8,4), 500 мМ KCl)
Система очистки ДНК PCR Preps (Promega, № по каталогу А7170)
Система векторов pGEM-T (Promega, № по каталогу А3600), включающая двукратный объем буфера с ДНК-лигазой Т4
Высокоэффективные компетентные клетки JM109 (Promega, № по каталогу L1001)
Система очистки ДНК Minipreps (Promega, № по каталогу А7100)
Набор для секвенирования BigDye Terminator Cycle Sequencing Ready Reaction (PE Applied Biosystems, № по каталогу 4303149)
Секвенатор ДНК ABI Prism 377 (PE)
Система 5' RACE (GIBCO, № по каталогу 18374-058), включающая якорную затравку с мостиковой связью
Система ДНК-полимеразы pfu (Promega, № по каталогу М7741)
Поликарбонатные фильтры 0,45 мкм (Sartorius)
Колонка для гель-фильтрации Superdex Peptide PE (7,5 x 300 мм) (Global)
Спектрофотометр DU7500 (Beckman)
Система для PCR 9700 GeneAmp (PE)
Лабораторный вакуумный коллектор Vac-Man Jr. (Promega, № по каталогу А7660)
Пример 1. Анализы протеазы
Были выполнены нижеследующие анализы активности протеазы.
Анализ с использованием AZCL-казеина
Раствор 0,2% синего субстрата AZCL-казеина суспендируют при перемешивании в буфере со смесью бура/NaH2PO4 с рН 9. (Для достижения профиля показателей рН по примеру 4 использовали буферную систему с рН 3-рН 11). Перемешиваемый раствор распределяют по лункам титрационного микропланшета (100 мкл в каждую лунку), добавляют 30 мкл образца фермента и планшеты инкубируют в термосмесителе фирмы Eppendorf в течение 30 минут при 45°С со скоростью вращения 600 оборотов/мин. Денатурированный образец фермента (кипящий при 100°С в течение 20 минут) используют в качестве контрольного образца. После инкубации реакцию прекращают, перенося титрационный микропланшет на лед, и окрашенный раствор отделяют от твердого вещества центрифугированием со скоростью 3000 оборотов/мин в течение 5 минут при 4°С. 60 мкл супернатанта переносят на титрационный микропланшет и измеряют оптическую плотность при 595 нм в спектрофотометре для считывания микропланшетов BioRad.
Анализ с использованием pNA-субстрата
50 мкл содержащего протеазу образца помещают на титрационный микропланшет и выполняют анализ, добавляя 100 мкл 1 мМ pNA-субстрата (5 мг растворяют в 100 мкл ДМСО и разводят до 10 мл буфером со смесью бура/NaH2PO4, рН 9,0). Активность протеазы определяют, измеряя увеличение оптической плотности OD405 при комнатной температуре.
Пример 2. Культивирование штамма CGMCC №0670 Thermoascus aurantiacus
Штамм Thermoascus aurantiacus CGMCC № 0670 выращивали при 45°С в течение 60 часов во встряхиваемых колбах со средой СВН1. Культуральный бульон собирали центрифугированием (7000 оборотов/мин в течение 20 минут при 4°С). Было получено в общей сложности 1500 мл культурального бульона.
Пример 3. Очистка протеазы, штамма Thermoascus aurantiacus CGMCC №0670
1500 мл супернатанта, полученного в примере 2, осаждали сульфатом аммония (80% насыщение) и повторно растворяли в 40 мл 25 мМ буфера с трис-HCl, рН 7,4. Полученный раствор подвергали ультрафильтрации через мембрану 5К для удаления солей и заменяли буфер 25 мМ трис-HCl с рН 7,4, после чего раствор фильтровали через фильтр 0,45 мкм. Конечный объем был равен 30 мл. Раствор вводили в 20 мл колонку Q Sepharose FF, уравновешенную 25 мМ трис-HCl, рН 7,4, и белки элюировали линейным градиентом NaCl (0-0,4 М). Фракции, элюированные из колонки, анализировали в отношении активности протеазы с использованием AZCL-казеина, рН 9,0, с SSI или без него. Собирали фракции, обладающие протеазной активностью и не ингибируемые SSI. Затем собранный раствор вводили в колонку Superdex75, уравновешенную 25 мМ трис-HCl, рН 7,4, и элюировали белки тем же буфером. Содержащие протеазу фракции анализировали методом SDS-PAGE и собирали чистые фракции.
Степень чистоты очищенной протеазы проверяли методом SDS-PAGE и с использованием IEF-геля. Образец содержал только одну протеазу, которой было присвоено название АР025. Молекулярная масса равна примерно 23 кДа, и pI соответствует рН 8,5.
Пример 4. Исследование протеазы, штамма Thermoascus aurantiacus CGMCC №0670
Испытание на ингибирование
Очищенную протеазу АР025 испытывали на ингибирование разными ингибиторами, включая SSI и EDTA. Протеазу инкубировали с 1,12 мг/мл SSI или 0,5 М EDTA в течение 5 минут при комнатной температуре и определяли остаточную активность при помощи анализа с использованием AZCL-казеина по примеру 1 (рН 9, 45°С). В качестве контрольных образцов использовали трипсин и SAVINASE™. Как показывают результаты, приведенные на фиг.1, ни SSI, ни EDTA, по-видимому, не ингибируют активность АР025.
Температурный профиль
Взаимосвязь между температурой и активностью протеазы определяли при помощи анализа с использованием AZCL-казеина (20 минут при встряхивании в термосмесителе). Полученный температурный профиль показан на фиг.2.
Протеаза АР025 является активной в широком диапазоне температур от 20°С до 90°С, причем оптимальная температура равна примерно 70°С. Но даже при 90°С указанная активность составляет по крайней мере 60% по сравнению с активностью при 55°С.
Профиль показателей рН
Взаимосвязь между показателем рН и активностью протеазы определяли при помощи анализа с использованием AZCL-казеина по примеру 1 (буферная система с рН 3-11). Полученный профиль показателей рН приведен на фиг.3.
Протеаза АР025 является активной в широком диапазоне показателей рН от 5 до 10. Оптимальный показатель рН равен примерно 6.
Устойчивость к показателям рН
Для измерения устойчивости к показателям рН смесь 15 мл образца фермента и 200 мл буфера с рН 3, 4, 5, 6, 7, 8, 9, 10, 11 инкубировали при 37°С в течение 2 часов и затем определяли остаточную активность фермента при помощи анализа с использованием AZCL-казеина по примеру 1 (добавляют 900 мкл AZCL-казеина в буфер со смесью бура/NaH2PO4, рН 9,0, инкубируют смесь при 45°С со скоростью вращения 1200 оборотов/мин в течение 10 минут и измеряют оптическую плотность при 595 нм). Результаты показаны на фиг.4. Протеаза является устойчивой в очень широком диапазоне показателей рН от 4 до 10, но оптимальная устойчивость соответствует примерно рН 4.
Специфичность pNA-субстрата
Специфичность субстрата протеазы АР025 определяли, используя более pNA-субстратов, включающих AAPF, AAPE, GGF, BA и FGL, а также SAVINASE™ и трипсин в качестве контрольных образцов.
Результаты, приведенные на фиг.5, показывают, что протеаза АР025 не обладает существенной активностью ни на одном из указанных субстратов.
Секвенирование N-концевой последовательности
N-концевая аминокислотная последовательность протеазы АР025 представляла собой: QRISSCSGSRQSALTTALRN (SEQ ID No.3).
Пример 5. Клонирование гена, кодирующего протеазу, штамма Thermoascus aurantiacus CGMCC 0670
Грибковый штамм и его выращивание
Штамм Thermoascus aurantiacus CGMCC №0670 выращивали в среде СВН1 при 45°С со скоростью вращения 165 оборотов/мин в течение 3 дней. Мицелий собирали центрифугированием со скоростью 7000 оборотов/мин в течение 30 минут. Собранный мицелий хранили при -80°С до использования для экстракции РНК.
Экстракция полной РНК
Полную РНК экстрагировали из 100 мг мицелия с использованием мининабора RNeasy.
Выращенные затравки
Нижеследующие выращенные затравки были созданы на основе части N-концевой аминокислотной последовательности QSALTTA:
T025-3 5' CA(A/G) TC(T/C/A/G) GC(T/C/A/G) CT(T/C/A/G) AC(T/C) AC(A/G) GC 3' (SEQ ID No.4)
T025-12 5' CA(A/G) AG(T/C) GC(T/C/A/G) CT(T/C/A/G) AC(A/G) AC(A/G) GC 3' (SEQ ID No.5)
Клонирование 3'-конца протеазы АР025
Для синтеза кДНК протеазы АР025 использовали набор 3' RACE. Примерно 5 мг полной РНК использовали в качестве матрицы и затравку Adapter использовали для синтеза первой цепи кДНК. Затем кДНК протеазы АР025 амплифицировали методом PCR, используя вышеуказанные выращенные затравки.
Были использованы реакционная система для выполнения PCR и условия реакции, приведенные ниже:
Условия
В результате анализа продукта PCR в геле при использовании затравок АР025-3 и АР025-12 была обнаружена специфическая полоса, соответствующая фрагменту длиной около 800 п.о. Полученные продукты выделяли из 1% LMP-агарозного геля, очищали инкубацией на бане с 70°С с последующим использованием системы для очистки ДНК PCR Preps. Концентрации очищенных продуктов определяли, измеряя оптическую плотность А260 и А280 в спектрофотометре. Затем очищенные фрагменты лигировали с вектором pGEM-T, используя соответствующий набор Promega:
Реакционные смеси инкубировали в течение ночи при 4°С.
2-4 мкл продуктов лигирования вводили в 50 мкл высокоэффективных компетентных клеток JM109 методом "теплового шока" (J. Sambrook, E.F., Fritsch, T. Maniatis (1989) Molecular Cloning 1.74, 1.84).
Трансформированные культуры помещали на планшеты LB со смесью ампициллин/IPTG/X-Gal и инкубировали в течение ночи при 37°С. Рекомбинантные клоны идентифицировали путем цветового скрининга на индикаторных планшетах и скрининга колоний методом PCR следующим образом.
Система скрининга колоний методом PCR
В белую колонию погружают кончик пипетки и пипетируют колонию в смесь для PCR, используемой в качестве матрицы.
Условия выполнения PCR
Положительные клоны инокулировали в 3 мл жидкой среды LB и инкубировали при встряхивании (примерно 250 оборотов/мин) в течение ночи при 37°С. Клеточный осадок отделяли центрифугированием с ускорением 10000 х g в течение 5 минут и из полученного клеточного осадка получали образцы плазмид, используя систему очистки ДНК Minipreps. И наконец, плазмиды секвенировали при помощи набора для секвенирования BigDye Terminator Cycle Sequencing Ready Reaction и секвенатора АВI377. Реакцию секвенирования выполняли следующим образом.
Результаты секвенирования показали, что полоса PCR, полученная при использовании затравки АР025-3 и затравки АР025-12, является 3'-концевой последовательностью АР025.
Клонирование 5'-конца АР025
На основании 3'-концевой последовательности АР025 были созданы три специфические затравки, которые использовали для клонирования 5'-конца последовательности.
Систему 5' RACE использовали для синтеза 5'-концевого фрагмента АР025. Для синтеза первой цепи добавляли 5 мкг полной РНК и затравки 5'-1. Затем кДНК очищали при помощи системы для очистки ДНК (входящей в комплект системы 5' RACE) и к 3'-концу кДНК добавляли полиС-концевой фрагмент. Для синтеза второй цепи использовали затравку 5'-2.
Были использованы система и условия PCR для получения кДНК с dC-концевым фрагментом, приведенные ниже.
Условия PCR
Для получения специфического продукта авторы настоящего изобретения амплифицировали 5'-конец при помощи вложенной PCR, используя затравку АР025-5'-3 (Frohman, M.A. (1990) PCR Protocols: A Guide to Methods and Applications (Innis, M.A., Gelfand, D.H., Sninsky, J.J., and White, T.J., eds.) p. 28, Academic Press, San Diego.
Условия PCR
При использовании затравки АР025-5'-3 в системе 5' RACE была получена специфическая полоса, относящаяся к фрагменту длиной 1000 п.о. Продукты PCR очищали, лигировали в вектор pGEM-T, трансформировали в компетентные клетки JM109 и секвенировали. Результаты секвенирования подтвердили, что был клонирован 5'-концевой фрагмент АР025.
Клонирование полного гена АР025
Затравка для клонирования полноразмерной последовательности была создана на основе вышеуказанных 3'- и 5'-концевых последовательностей.
Затравку АР025-CDS-2 и AUAP использовали для амплификации полноразмерного гена из кДНК АР025. Были использованы реакционная система и условия выполнения PCR, приведенные ниже.
Условия
В результате указанной амплификации была получена специфическая полоса длиной около 1,4 т.п.о. Для добавления полиА-концевого фрагмента использовали TaqДНК-полимеразу и инкубировали реакционную смесь при 72°С в течение 30 минут. dA-концевой фрагмент выделяли из геля при помощи системы очистки ДНК PCR Preps. Затем очищенный фрагмент лигировали в вектор pGEM-T и трансформировали в компетентные клетки (JM109). Скрининг некоторых положительных клонов был произведен при помощи PCR для колоний.
Система выполнения PCR для колоний
В белую колонию погружают кончик пипетки и пипетируют колонию в смесь для PCR, используемой в качестве матрицы.
Условия
Затем плазмиду экстрагировали из указанных клонов при помощи системы очистки ДНК Minipreps. Плазмиду секвенировали, используя набор для секвенирования BigDye Terminator Cycle Sequencing Ready Reaction, в результате чего была получена полноразмерная последовательность.
Анализ последовательности клона кДНК показал, что последовательность содержит кодирующую область из 1065 нуклеотидов. Продуктом трансляции указанной кодирующей области является пептид, содержащий 355 аминокислот. Выведенная аминокислотная последовательность данного гена с сигнальным пептидом (аа 1-19), пропептидом (20-178) и зрелым пептидом (аа 179-335) показана на фиг.6 и 7.
Пример 6. Действие протеазы, штамма Thermoascus aurantiacus CGMCC 0670, в модели пищеварения с однокамерным желудком in vitro
Протеаза АР025 была экспрессирована в и выделена из Aspergillus oryzae. Очищенная протеаза характеризовалась показателем А280, равным 1,15, и А260, равным 0,52. Действие очищенной протеазы испытывали в модели in vitro, имитирующей пищеварение у животных с однокамерным желудком. В частности, испытывали способность протеазы улучшать солюбилизацию и расщепление белков кукурузы/SBM (кукурузы/соевой муки).
Система in vitro состояла из 10 колб, в которых первоначально инкубировали субстрат кукурузы/SBM с HCl/пепсином, имитируя пищеварение в желудке, и затем с панкреатином, имитируя пищеварение в кишечнике. В начале фазы пищеварения в желудке протеазу АР025 вводили в пять колб, при этом остальные колбы служили в качестве контрольных колб. В конце фазы инкубации, соответствующей пищеварению в кишечнике, образцы переваренного корма in vitro удаляли и анализировали в отношении содержания солюбилизованного и расщепленного белка.
Схема пищеварения in vitro
41 мл HCl (0,105 М)
(3000 ед/г субстрата), протеаза АР025 (100 мг белка фермента протеазы/кг рациона)
Условия
Растворы
0,39 М NaOH
0,105 М HCl
0,105 М HCl, содержащий 6000 ед. пепсина на 5 мл
1 М NaHCO3, содержащий 16 мг панкреатина на мл
125 мМ буфера с NaAc, рН 6,0
Определение белка фермента
Количество белка фермента протеазы вычисляют с учетом значений А280 и аминокислотных последовательностей (аминокислотных композиций) на основании принципов, приведенных в статье S.C.Gill & P.H. von Hippel, Analytical Biochemistry 182, 319-326 (1989).
Экспериментальная методика использования модели in vitro
Экспериментальная методика была аналогична описанной выше. Показатель рН измеряли в периоды времени, соответствующие 1, 2,5 и 5,5 часам. Инкубацию заканчивали по истечении 6 часов, после чего 30 мл образцы удаляли и помещали на лед до центрифугирования (10000 х g, 10 минут, 4°С). Супернатанты удаляли и хранили при -20°С.
Анализ
Все образцы анализировали в отношении степени гидролиза (DH) в процентах методом на основе ОРА, а также в отношении содержания солюбилизованного и расщепленного белка методом гель-фильтрации.
Определение степени гидролиза методом на основе ОРА
Степень гидролиза (DH) белка в разных образцах определяли колориметрическим методом с использованием полуавтоматического титрационного микропланшета (Nielsen, P.M.; Petersen, D.; Dambmann, C. Improved method for determining food protein degree of hydrolysis. J. Food Sci. 2001, 66, 642-646). Реагент ОРА получали следующим образом: 7,620 г декагидрата динатрийтетрабората и 200 мг додецилсульфата натрия (SDS) растворяли в 150 мл деионизированной воды. Реагенты полностью растворяли. Затем 160 мг 97% о-фталдиальдегида (ОРА) растворяли в 4 мл этанола. Раствор ОРА переносили в количественном отношении в вышеуказанный раствор, осуществляя промывку деионизированной водой. К раствору добавляли 176 мг 99% дитиотреитола (DTT), который доводили до 200 мл при помощи деионизированной воды. Сериновый эталон (0,9516 мэкв/л) получали, растворяя 50 мг серина (Merck, Germany) в 500 мл деионизированной воды.
Раствор образца получали, разводя каждый образец до достижения оптической плотности (280 нм), равной примерно 0,5. В целом супернатанты разводили (в 100 раз) при помощи автоматической установки для разведения Tecan (Mannedorf, Switzerland). Все другие показания спектрофотометра считывали при 340 нм, используя деионизированную воду в качестве контрольного образца. 25 мкл образца, эталон и контрольный образец диспергировали в титрационном микропланшете. Титрационный микропланшет помещали в спектрофотометр для считывания планшетов (Labsystems, Finland) и автоматически диспергировали 200 мкл реагента ОРА. Планшеты встряхивали (2 минуты; 700 оборотов/мин) до измерения оптической плотности. Затем вычисляли степень гидролиза. Определение степени гидролиза всех образцов выполняли по восемь раз.
Определение солюбилизованного и расщепленного белка
Содержание в супернатантах солюбилизованного белка из расщепленных образцов in vitro определяли путем количественного измерения сырого белка (СР) при помощи ВЭЖХ с гель-фильтрацией. Супернатанты оттаивали, фильтровали через поликарбонатные фильтры 0,45 мкм и разводили (1:50, об/об) Н2О. Разведенные образцы хроматографировали при помощи ВЭЖХ с использованием колонки Superdex Peptide PE (7,5 x 300 мм) для гель-фильтрации (Global). В качестве элюента для изократического элюирования использовали 50 мМ буфера с фосфатом натрия (рН 7,0), содержащего 150 мМ NaCl. Общий объем элюента, используемый в одном цикле, был равен 26 мл при скорости потока 0,4 мл/мин. Профили элюирования регистрировали при 214 нм и общую площадь под профилями определяли интегрированием. Для определения содержания белка на интегрированных участках строили калибровочную кривую (R2=0,9993) на основании ряда разведений in vitro расщепленного контрольного образца смеси кукуруза/SBM с известным общим содержанием белка. Определение белка в указанном контрольном образце выполняли методом Kjeldahl (определение в % азота; А.О.А.С. (1984) Official Methods of Analysis 14th ed., Washington DC).
Содержание расщепленного белка определяли интегрированием площади хроматограммы, соответствующей пептидам и аминокислотам с молекулярной массой 1500 Дальтон или меньше (Savoie, L.; Gauthier, S.F. Dialysis Cell For The In-vitro Measurement of Protein Digestibility. J. Food Sci. 1986, 51, 494-498; Babinszky, L.; Van, D.M.J.M.; Boer, H.; Den, H.L.A. An In-vitro Method for Prediction of The Digestible Crude Protein Content in Pig Feeds. J. Sci. Food Agr. 1990, 50, 173-178; Boisen, S.; Eggum, B.O. Critical Evaluation of In-vitro Methods for Estimating Digestibility in Simple-Stomach Animals. Nutrition Research Reviews 1991, 4, 141-162). Для определения линии деления, соответствующей 1500 Да, колонку для гель-фильтрации калибровали, используя цитохром С, апротинин, гастрин I и вещество Р в качестве эталонов молекулярной массы.
Результаты
Результаты, приведенные в нижеследующих таблицах 1 и 2, показывают, что протеаза АР025 значительно увеличивала количество расщепляемого белка, при этом степень гидролиза и количество растворимого белка возрастали в численном отношении по сравнению с контрольным образцом.
При испытании только в части указанной модели, имитирующей пищеварение в кишечнике, протеаза АР025 (в аналогичной дозе, равной 100 мг белка фермента на кг субстрата) значительно повышала уровень солюбилизованного и расщепленного белка (соответственно 103,0 и 103,1% по сравнению с контрольным образцом), при этом степень гидролиза не увеличивалась (96,5% по сравнению с контрольным образцом).
Разные буквенные обозначения в одном и том же столбце означают существенные различия (однонаправленный метод ANOVA, критерий Tukey-Kramer, P<0,05). SD = стандартное отклонение. %CV = коэффициент вариации = (стандартное отклонение/среднее значение) х 100%.
троль
Разные буквенные обозначения в одном и том же столбце означают существенные различия (однонаправленный метод ANOVA, критерий Tukey-Kramer, P<0,05). SD = стандартное отклонение. %CV = коэффициент вариации = (стандартное отклонение/среднее значение) х 100%.
Пример 7. Действие протеазы, штамма Thermoascus aurantiacus CGMCC 0670, в модели пищеварения аквакультуры in vitro
Препарат протеазы АР025, аналогичный описанному в примере 6, испытывали в модели аквакультуры in vitro, имитирующей пищеварение у рыбы, обитающей в холодной воде. По сравнению с моделью пищеварения в однокамерном желудке испытание системы аквакультуры производили при несколько других показателях рН (рН 3,3 в фазе пищеварения в желудке и рН 7,2-7,8 в фазе пищеварения в кишечнике). Температура инкубации была ниже (15°С вместо 40°С) и периоды инкубации продолжительнее (указаны ниже). Десять колб, содержащих вытесненную SBM в качестве субстрата, сначала инкубировали с HCl/пепсином, имитируя пищеварение в желудке, и затем с панкреатином, имитируя пищеварение в кишечнике. В пять колб протеазу АР025 вводили в начале фазы пищеварения в желудке, остальные колбы служили в качестве контрольных образцов. В конце фазы инкубации, соответствующей пищеварению в кишечнике, образцы переваренного корма in vitro удаляли и анализировали в отношении солюбилизованного и расщепленного белка.
Схема исследования аквакультуры in vitro, имитирующая пищеварение у рыбы, обитающей в холодной воде
Условия
Растворы
1,10 М NaOH
0,18 М HCl, содержащей 652 ед. пепсина на мл
1 М NaHCO3, содержащий 16 мг панкреатина на мл
125 мМ буфера с NaAc, рН 6,0
Экспериментальная методика использования модели пищеварения аквакультруры in vitro
Экспериментальная методика была аналогична описанной выше. Показатель рН измеряли через 1, 5, 8 и 23 часа. Инкубацию заканчивали по истечении 24 часов, после чего отбирали 30 мл образцы и помещали их на лед до центрифугирования (10000 х g, 10 минут, 4°С). Супернатанты удаляли и хранили при -20°С.
Анализ
Все супернатанты анализировали методом на основе ОРА (степень гидролиза в процентах), солюбилизованный и расщепленный белок исследовали при помощи ВЭЖХ Akta (см. пример 6).
Результаты
Результаты, приведенные в нижеследующих таблицах 3 и 4, показывают, что протеаза АР025 значительно увеличивает растворимость белка, расщепляемость белка и степень гидролиза по сравнению с контрольным образцом.
Разные надстрочные индексы в одном и том же столбце означают существенные различия (однонаправленный метод ANOVA, критерий Tukey-Kramer, P<0,05).
Разные надстрочные индексы означают существенные различия (метод ANOVA, критерий Tukey-Kramer, P<0,05).
Депонированный биологический материал
Нижеследующий биологический материал был депонирован в соответствии с положениями Будапештского договора в коллекции DSMZ (DSMZ-Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH, Mascheroder Weg 1b, D-38124 Braunschweig, Germany) и CGMCC (Китайский центр коллекций микробиологических культур, Институт микробиологии, Китайская академия наук, Haidian, Beijing 100080, China), где указанному материалу были присвоены нижеследующие номера доступа:
Штамм Escherichia coli включает плазмиду, содержащую последовательность нуклеиновой кислоты протеазы Thermoascus aurantiacus (то есть SEQ ID No.1, кодирующую SEQ ID No.2).
Указанные штаммы были депонированы на условиях, гарантирующих доступ к культурам в течение всего срока рассмотрения данной заявки на патент лицу, определяемому уполномоченным по патентам и товарным знакам в соответствии со статьей 37 C.F.R (Свод федеральных постановлений США), §1.14 и статьей 35 U.S.C. (Кодекс законов США), §122. Депозиты представляют по существу чистую культуру депонированных штаммов. Депозиты предоставляются в соответствии с требованиями иностранного патентного законодательства в странах, где находится на рассмотрении данная заявка или последующие заявки. Однако совершенно очевидно, что доступ к депозиту не предполагает предоставление лицензии на использование данного изобретения в нарушение патентных прав, предусмотренных государственным правом.
Штамм №CGMCC 0670 Thermoascus aurantiacus был выделен из образца почвы, полученного 21 июля 1998 г в провинции Юньнань, г.Сишуан-банна, Китай.
Изобретение, представленное в данном описании изобретения и формуле изобретения, не ограничивается приведенными конкретными вариантами его осуществления, которые служат только для иллюстрации некоторых объектов данного изобретения. Все подобные варианты осуществления изобретения входят в объем данного изобретения. Из приведенного описания изобретения специалистам в данной области должны быть очевидны другие модификации изобретения помимо рассмотренных выше. Такие модификации также входят в объем прилагаемой формы изобретения. В случае возникновения противоречий следует обратиться к описанию изобретения, содержащему определения терминов.
Все приведенные материалы полностью входят в описание изобретения в качестве ссылки.










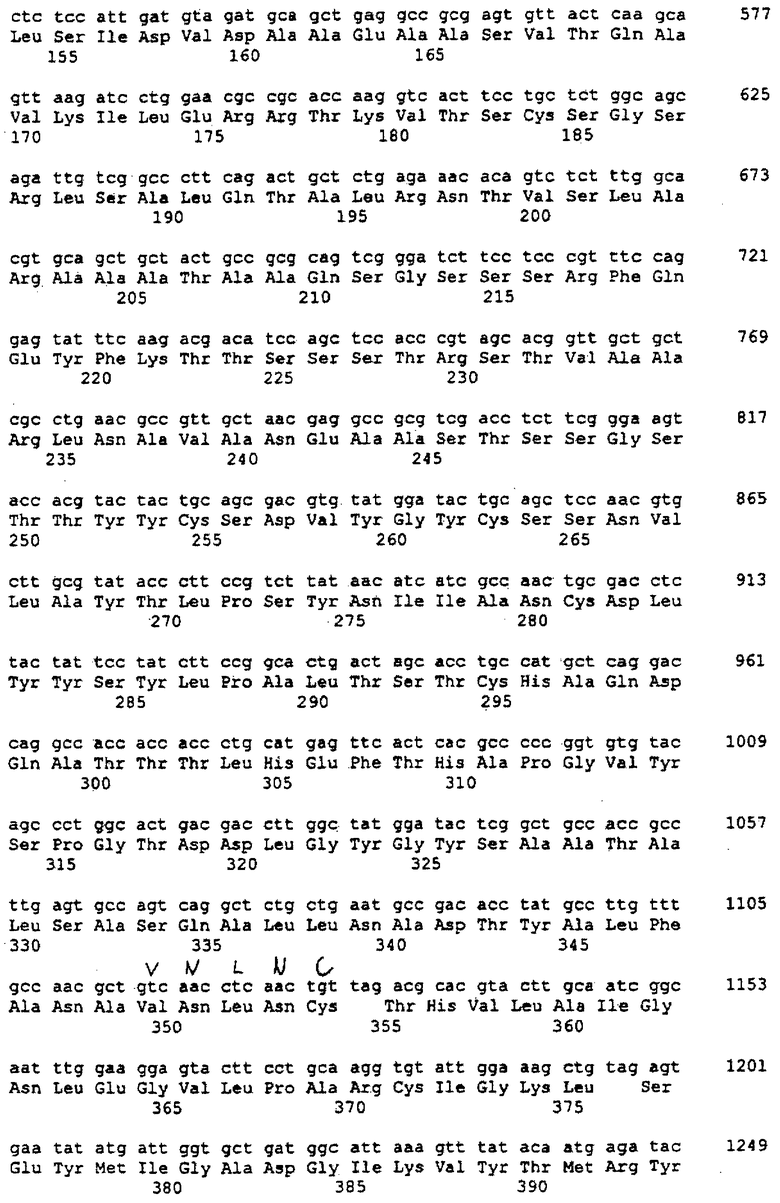


| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЕ ГЕНЫ, КОДИРУЮЩИЕ НОВЫЕ ПРОТЕОЛИТИЧЕСКИЕ ФЕРМЕНТЫ | 2002 |
|
RU2423525C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ ПРОТИВОМИКРОБНЫМ ДЕЙСТВИЕМ, И КОДИРУЮЩИЕ ИХ ПОЛИНУКЛЕОТИДЫ | 2006 |
|
RU2403260C2 |
| НОВЫЕ ГЕНЫ, КОДИРУЮЩИЕ НОВЫЕ ПРОТЕОЛИТИЧЕСКИЕ ФЕРМЕНТЫ | 2002 |
|
RU2296160C2 |
| ПОЛИПЕПТИДЫ С ПРОТИВОМИКРОБНОЙ АКТИВНОСТЬЮ И КОДИРУЮЩИЕ ИХ ПОЛИНУКЛЕОТИДЫ | 2005 |
|
RU2393224C2 |
| ПОЛИПЕПТИДЫ С ЛИЗОЦИМНОЙ АКТИВНОСТЬЮ И ПОЛИНУКЛЕОТИДЫ, КОДИРУЮЩИЕ ИХ | 2012 |
|
RU2619051C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ ЭНДОПЕПТИДАЗНОЙ АКТИВНОСТЬЮ, И КОДИРУЮЩИЕ ИХ ПОЛИНУКЛЕОТИДЫ | 2011 |
|
RU2583293C2 |
| ПРОТИВОМИКРОБНЫЕ ПОЛИПЕПТИДЫ ИЗ PSEUDOPLECTANIA NIGRELLA | 2002 |
|
RU2336278C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ АНТИМИКРОБНОЙ АКТИВНОВНОСТЬЮ, И ПОЛИНУКЛЕОТИДЫ, КОДИРУЮЩИЕ ИХ | 2006 |
|
RU2512525C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ АНТИМИКРОБНОЙ АКТИВНОСТЬЮ, И ПОЛИНУКЛЕОТИДЫ, КОДИРУЮЩИЕ ИХ | 2006 |
|
RU2415150C2 |
| СПОСОБЫ ПОВЫШЕНИЯ УСИЛИВАЮЩЕЙ ЦЕЛЛЮЛОЛИТИЧЕСКОЙ АКТИВНОСТИ ПОЛИПЕПТИДА | 2008 |
|
RU2510417C2 |
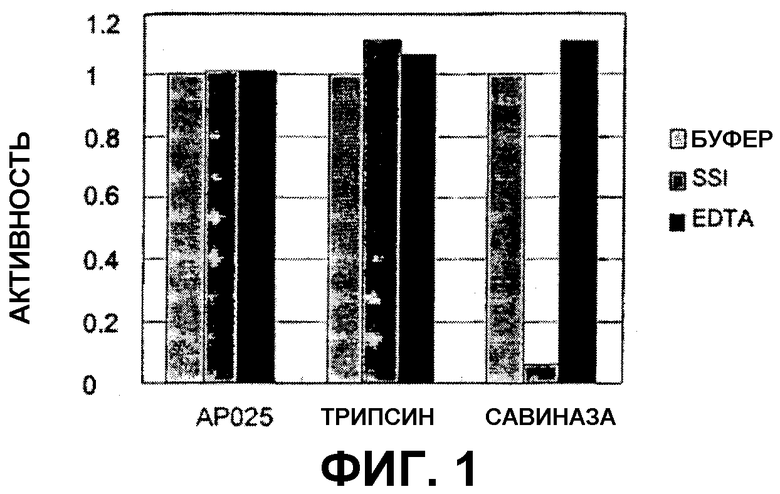
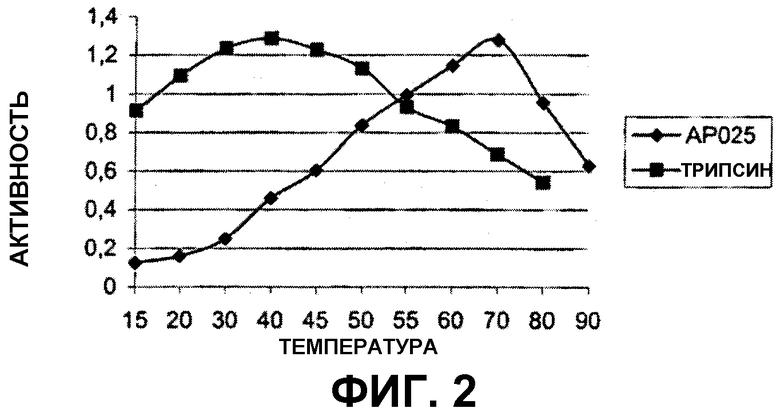
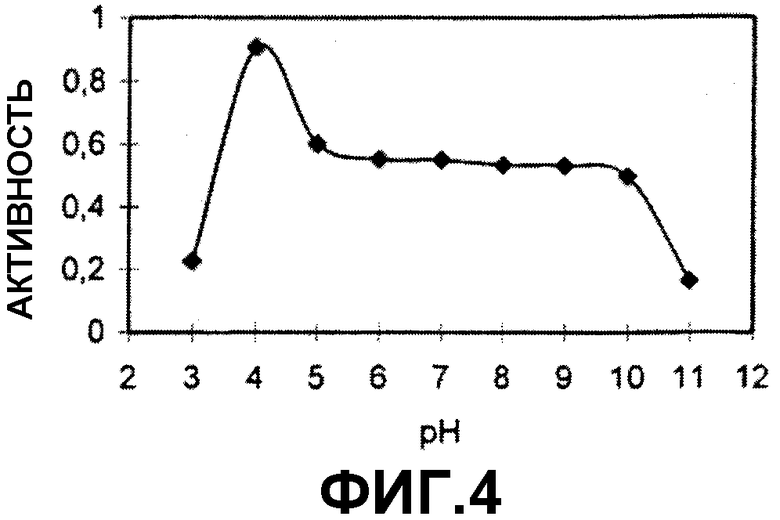
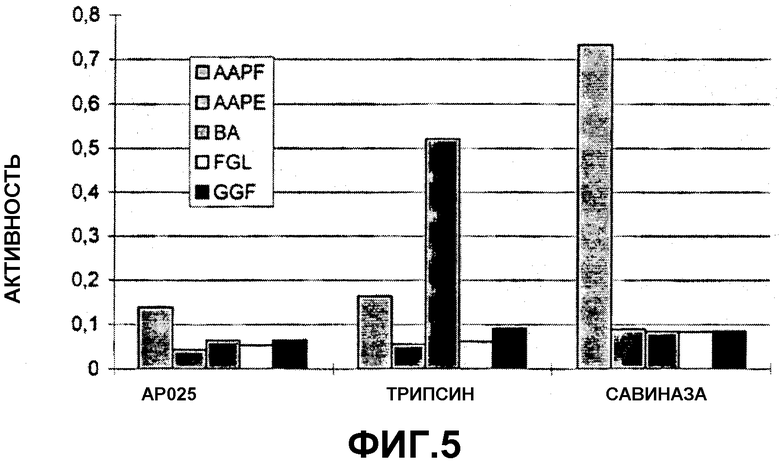
Изобретение относится к биотехнологии. Предложен выделенный полипептид, являющийся кислотостойкой металлопротеазой, выделенной из Thermoascus aurantiacus. Описаны: штамм Thermoascus aurantiacus CGMCC №0670 для получения полипептида и способ получения полипептида с использованием указанного штамма. Раскрыт способ обработки растительных белков с целью повышения их усвояемости с использованием предложенного полипептида. Настоящее изобретение позволяет получить протеазу, которая обладает достаточной кислотостойкостью, чтобы избежать разрушения в желудке и может найти применение в производстве кормов. 4 н. и 2 з.п. ф-лы, 5 ил, 4 табл.
(a) полипептида с аминокислотной последовательностью, степень идентичности которой с аминокислотами от -178 до 177, от -159 до 177 или от +1 до 177 последовательности SEQ ID No.2 составляет по меньшей мере 80%; и
(b) полипептида, кодируемого последовательностью нуклеиновой кислоты, которая гибридизуется в условиях средней степени жесткости
(i) с частью плазмиды, находящейся в штамме Escherichia coli DSM 14652, которая кодирует зрелую протеазу;
(ii) с нуклеотидами 25-1089, 1-1089, 1-1344, 25-1344, 559-1344 или 559-1089 последовательности SEQ ID No.l;
(iii) с комплементарной цепью указанного в (i) или (ii);
(c) аллельного варианта полипептида (а) или (b) и
(d) фрагмента полипептида (а), (b) или (с), обладающего протеазной активностью.
(i) является металлоэндопептидазой ЕС 3.4.24;
(ii) является металлопротеазой семейства М и/или
(iii) содержит мотив HEFTH.
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| Амплитудно-фазовый измеритель для геоэлектроразведки | 1984 |
|
SU1236405A1 |
| RU 94028712 A1, 10.03.1997 | |||
| RU 2070575 C1, 20.12.1996 | |||
| TATSUMI H et al: "Cloning and expression in yeast of a complementary dna clone encoding aspergillus-oryzae neutral protease ii a unique metalloprotease" molecular & general genetics, vol | |||
| Приспособление для нагрузки тендеров дровами | 1920 |
|
SU228A1 |

