Область, к которой относится изобретение
Настоящее изобретение относится к новым идентифицированным полинуклеотидным последовательностям, включающим гены, кодирующие новые протеазы, выделенные из Aspergillus niger. Настоящее изобретение относится к полноразмерной нуклеотидной последовательности указанных новых генов, к кДНК-последовательностям, включающим эти полноразмерные кодирующие последовательности новых протеаз, а также к аминокислотным последовательностям полноразмерных функциональных белков, а также к их фрагментам и вариантам. Настоящее изобретение также относится к способам использования этих ферментов в промышленности и к способам диагностики грибковых инфекций. Настоящее изобретение также относится к клеткам, трансформированным полинуклеотидом настоящего изобретения, и к клеткам, в которых протеаза настоящего изобретения является генетически модифицированной в целях усиления или снижения ее активности и/или уровня экспрессии.
Предшествующий уровень техники
Протеолитические ферменты
Белки могут рассматриваться как гетерополимеры, которые состоят из аминокислотных структурных элементов, соединенных пептидной связью. Повторяющимся элементом в белках является центральный атом альфа-углерода с аминогруппой и карбоксильной группой. Так называемая боковая цепь аминокислоты, за исключением глицина, замещает один из двух оставшихся атомов водорода у альфа-углерода. Боковая цепь аминокислоты делает центральный атом альфа-углерода асимметричным. В составе белков, в основном, обнаруживаются L-энантиомеры аминокислоты. Различные типы полимеризованных аминокислот определяют нижеследующими терминами. “Пептиды” представляют собой короткие цепи аминокислотных остатков с определенной последовательностью. Хотя, фактически, нельзя указать точно максимальное число остатков, однако этот термин обычно обозначает цепь, которая обладает свойствами, определяемыми, главным образом, ее аминокислотным составом, и которая не имеет фиксированной трехмерной конформации. Термин “полипептид” обычно используется для обозначения более длинных цепей, имеющих обычно определенную последовательность и длину, которая, в принципе, является достаточной для образования трехмерной структуры. Термин “белок” обычно означает полипептиды, встречающиеся в природе и имеющие определенную трехмерную структуру. В случае если главной функцией белков является катализ химической реакции, то обычно такие белки называют ферментами. Протеазами называют ферменты, которые катализируют гидролиз пептидной связи в (поли)пептидах и в белках.
В физиологических условиях протеазы катализируют гидролиз пептидной связи. Международный союз по биохимии и молекулярной биологии (1984) рекомендует использовать термин “пептидаза” для подгруппы гидролазпептидной связи (подкласс Е.С.3.4). Термины “протеаза” и “пептидогидролаза” являются синонимами термина “пептидаза” и также могут быть использованы в настоящем описании. Протеазы включают два класса ферментов: эндопептидазы и экзопептидазы, которые расщепляют пептидные связи в определенных точках в белке и последовательно удаляют аминокислоты либо из N-, либо из С-конца соответственно. Термин “протеиназа” используется как синоним термина “эндопептидаза”. Пептидная связь может присутствовать в контексте ди-, три-, тетрапептидов, пептидов, полипептидов или белков. В основном, аминокислотные составы природных пептидов и полипептидов включают 20 различных аминокислот, которые имеют L-конфигурацию (за исключением глицина, у которого отсутствует хиральный центр). Однако протеолитическая активность протеаз не ограничивается пептидами, которые содержат лишь 20 природных аминокислот. Могут быть также расщеплены пептидные связи между так называемыми неприродными аминокислотами и пептидные связи между модифицированными аминокислотами или аминокислотными аналогами. Некоторые протеазы распознают D-энантиомеры аминокислот в некоторых положениях. В основном, значительная стереоселективность протеаз является очень важным фактором, который позволяет использовать их в способах химического разделения. Многие протеазы обладают другими представляющими интерес активностями, такими как эстеразная активность, тиолэстеразная активность и (де)амидазная активность. Эти вспомогательные активности обычно не ограничиваются лишь аминокислотами и могут быть с успехом использованы в реакциях биологических превращений в области тонкого химического синтеза.
Особый интерес к протеазам нитчатых грибов, эукариотических микроорганизмов, вызван рядом причин. Очевидно, что основной процесс гидролитического расщепления пептидных связей в белках является дорогостоящим, и если он соответствующим образом не регулируется, то может оказывать негативное влияние на микроорганизм. Желаемое ограничение протеолитического действия может быть достигнуто посредством специфичности протеиназ; компартментализации протеаз и субстратов в клетке; модификации субстратов, распознаваемых соответствующими протеазами, регуляции посредством активации зимогенов и благодаря присутствию или отсутствию специфических ингибиторов, а также посредством регуляции экспрессии гена протеазы. В грибах протеазы также участвуют в других фундаментальных клеточных процессах, включая метаболизм внутриклеточного белка, процессинг, транслокацию, споруляцию, развитие и дифференцировку. Действительно, Aspergillus nidulans и Neurospora crassa были использованы как микроорганизмы-модели для анализа молекулярной основы ряда физиологических процессов и процессов развития. Их генетика открывает прямой доступ к биохимическим и генетическим исследованиям в определенных условиях питания и культивирования. Кроме того, была выделена большая группа грибов, патогенных для человека, крупного рогатого скота и сельскохозяйственных культур, и было высказано предположение, что в их патогенности (проникновение в организм хозяина, противодействие механизмам защиты хозяина и/или инфицирование в процессе питания) определенную роль играет протеолиз. Протеазы также часто используются в лабораторных, клинических и промышленных способах; при этом как микробные, так и немикробные протеазы широко используются в пищевой промышленности (в хлебопечении, пивоварении, в производстве сыра и в тендеризации мяса), в кожевенной промышленности и в промышленном производстве биологических детергентов (Aunstrup, 1980). В последнее время, в связи с промышленным применением некоторых нитчатых грибов, а в частности Aspergillus, в качестве хозяев для продуцирования гомологичных и гетерологичных белков, снова возрос интерес к грибковым протеазам (van Brunt, 1986ab). Протеазы часто затрудняют гетерологичную экспрессию и гомологичную сверхэкспрессию белков в грибах. В частности, на гетерологичную экспрессию негативно влияет протеолитическое расщепление продуктов экспрессии гомологичными протеазами. Указанный коммерческий интерес явился стимулом для детального исследования протеолитического спектра протеаз и конструирования дефицитных по протеазе штаммов, что привело к получению более глубоких знаний об экспрессии протеаз и их регуляции в указанных микроорганизмах. Поэтому в настоящее время назрела необходимость в идентификации и элиминации новых протеаз в нитчатых грибах.
Микроорганизмы, такие как, например, грибы, являются особенно подходящими для крупномасштабного продуцирования белков, а особенно, если такие белки секретируются в среду. В этих процессах продуцирования определенную роль играют протеолитические ферменты. С одной стороны, конкретные протеолитические ферменты, в основном, необходимы для “правильного” процессинга белка-мишени и благоприятного осуществления метаболических функций хозяина-продуцента. С другой стороны, протеолитическое расщепление может значительно снизить выход секретированных белков. Неправильная укладка в пути секреции может приводить к расщеплению внутриклеточными протеазами. В связи с этим при продуцировании гетерологичных белков могут возникнуть определенные трудности. Точные механизмы протеолитических процессов, которые ответственны за расщепление белков, которые отклоняются от секреторного пути в грибках, пока еще не совсем ясны. В эукариотах деградация клеточных белков достигается с использованием протеасомы и обычно осуществляется мечением разлагаемых белков убихитином. В грибках протеосомные и вакуолярные протеазы, вероятно, также являются кандидатами для протеолитического расщепления плохо уложенных секреторных белков. Протеолитическая деградация, вероятно, является цитоплазматической, но нельзя исключать и существование протеаз в эндоплазматическом ретикулуме. С точки зрения усовершенствования хозяйского штамма-продуцента, протеолитическая система может служить интересной целью для усовершенствования генетического конструирования и продуцирования штаммов. Дополнительные копии генов протеаз, сверхэкспрессия некоторых протеаз, модификация транскрипционной регуляции, а также “нокаут”-процедуры для делеции генов протеаз позволят лучше понять функции данной протеазы. Делеция генов, кодирующих протеазу, может служить ценной стратегией для модификации штамма-хозяина, осуществляется в целях увеличения выхода продуцирования гомологичных, а также гетерологичных белков.
Эукариотические микробные протеазы были описаны North (1982). Совсем недавно Suarez Rendueles и Wolf (1988) описали протеазы S. cerevisiae и их функции.
Помимо гидролитического расщепления связей, протеазы могут также участвовать в образовании связей. В этом аспекте понятие “связи” включает не только пептидные и амидные связи, но также и сложноэфирные связи. Способность протеаз катализировать расщепление или образование конкретной связи, в первую очередь, зависит от термодинамики реакции. Такой фермент, как протеаза, не влияет на равновесие реакции. Равновесие реакции зависит от конкретных условий, при которых осуществляется данная реакция. В физиологических условиях термодинамика данных реакций может оказаться благоприятной для гидролиза пептида благодаря в высокой степени термодинамически стабильной структуре цвиттерионного продукта. Опираясь на физико-химические законы, влияющие на равновесие реакции, либо изменяя концентрации или природу реагентов и продуктов, либо подбирая кинетические параметры ферментативной реакции, можно использовать протеазы в целях синтеза пептидных связей. Добавление смешиваемых с водой органических растворителей приводит к снижению степени ионизации карбоксильного компонента и, тем самым, к повышению концентрации субстрата, доступного для реакции. Для увеличения выхода часто используют двухфазные системы, водные миметики, обращенные мицеллы, безводные среды или модифицированные амино- и карбоксильные группы, стимулирующие преципитацию продуктов. Если бы имелись в распоряжении протеазы с подходящими свойствами, то использование таких протеаз для синтеза дало бы значительные преимущества. Поскольку протеазы являются стереоселективными, а также региоселективными, то чувствительные группы на реагентах обычно не нуждаются в защите и эти реагенты необязательно должны быть оптически чистыми. Если условия ферментативного синтеза являются мягкими, то можно избежать рацемизации и разложения лабильных реагентов или продуктов. Кроме того, помимо связей между аминокислотами, соответствующим образом выбранные протеазы могут также связывать другие соединения, имеющие первичную аминогруппу, тиоловую группу или карбоксильную группу. Кроме того, некоторыми протеазами могут быть синтезированы сложные эфиры, тиоловые эфиры и амиды. Было показано, что протеазы обладают региоселективностью при ацилировании моно-, ди- и трисахаридов, нуклеозидов и рибофлавина. Проблемы, связанные со стабильностью, которые иногда возникают при жестких реакционных условиях, могут быть решены путем правильного подбора реагентов. Инкапсуляция и иммобилизация не только стабилизируют ферменты, но также позволяют легко отделить и выделить их из реакционной среды. Густота сшивок, обработка альдегидами или покрытие поверхности некоторыми полимерами, такими как декстраны, полиэтиленгликоль и полиимины, может значительно увеличить срок службы биокатализатора.
Природная роль протеаз
Традиционно протеазы считались протеолитическими ферментами, которые способны расщеплять белки на небольшие пептиды и/или аминокислоты и роль которых заключается в гидролизе пищевых белков или в участии в метаболизме клеточных белков. Кроме того, было показано, что протеазы также играют ключевые роли в клеточных процессах широкого ряда в соответствии с механизмами селективной модификации путем ограниченного протеолиза, а поэтому они обладают важными регуляторными функциями (Holzer & Tschensche 1979; Holzer & Heinrich, 1980). Предполагается, что специфичность протеиназы тесно связана с ее физиологической функцией и с типом ее экспрессии. Что касается функции конкретной протеазы, то ее локализация в большинстве случаев является очень важной; так, например, серия вакуолярных и периплазматических протеаз участвует в разложении белка, тогда как многие мембрано-ассоциированные протеазы играют важную роль в процессинге белка (Suarez Rendueles & Wolf, 1988). Различные роли протеаз во многих клеточных процессах могут быть подразделены на четыре основных функции протеаз: 1) деградация белка, 2) посттрансляционный процессинг и (ин)активация специфических белков, 3) морфогенез и 4) патогенез.
Очевидная роль протеаз в организмах, которые используют белок в качестве питательного источника, заключается в гидролизе питательных веществ. В грибах это должно приводить к их деградации за пределами клетки под действием внеклеточных протеаз с широкой специфичностью. Деградация белка также имеет важное значение для быстрого метаболизма клеточных белков и позволяет удалять из клетки ненужные белки и адаптировать их комплемент белка к изменению физиологических условий. Обычно протеазы с достаточно широкой специфичностью должны очень хорошо регулироваться для защиты клеточных белков, не являющихся белками-мишенями, от случайной деградации.
В противоположность гидролизу синтез полипептидов происходит in vivo посредством АТФ-управляемого процесса на рибосоме. В конечном счете, последовательность, в которой аминокислоты являются связанными, определяется информацией, обеспечиваемой геномом. Этот процесс известен как транскрипция. Первичные продукты трансляции обычно являются более длинными, чем конечные функциональные продукты, и после транскрипции обычно необходим процессинг таких белков-предшественников под действием протеаз. Протеазы играют ключевую роль в созревании таких белков-предшественников и в образовании конечного функционального белка. В противоположность в высокой степени регулируемому “урезанию” и “реконструированию” белков протеазы могут оказывать в высокой степени деструктивное действие и могут приводить к полному разложению полипептидов на пептиды и аминокислоты. Для предотвращения такой свободной протеолитической активности, до того как это потребуется, протеазы должны подвергаться интенсивной регуляции. Многие протеазы синтезируются как более крупные предшественники, известные как зимогены, которые, при необходимости, становятся активированными. Следует отметить, что такая активация всегда происходит посредством протеолиза. Помимо прямого участия в процессинге, селективная активация и инактивация отдельных белков являются хорошо известными процессами, катализируемыми специфическими протеазами.
Очевидно, что селективность ограниченного протеолиза почти всегда наблюдается при взаимодействии протеиназа-субстрат. Эта специфичность может обеспечиваться протеолитическим ферментом, который распознает только специфические аминокислотные последовательности-мишени. С другой стороны, она также может быть обусловлена селективным воздействием на “сайт процессинга” при определенных условиях, таких как рН, ионная сила или вторичные модификации, что позволяет неспецифическим протеазам, тем или иными способом, катализировать в высокой степени специфическое событие. Примером события такого типа является активация вакуолярных зимогенов при ограниченном протеолизе.
Морфогенез или дифференциация могут быть определены как регулируемая серия событий, приводящих к переходу из одного состояния в организме в другое состояние. Хотя, во многих случаях, прямая взаимосвязь между протеазами и морфологическими эффектами не была установлена, однако имеются данные, которые позволяют предположить о значительной роли протеаз в морфогенезе грибов, то есть, помимо наблюдаемого интенсивного метаболизма белка в процессе дифференцировки, споруляции и прорастании спор, протеазы, очевидно, непосредственно участвуют в нормальных процессах, таких как ветвление кончиков гифов и образование перегородок (Deshpande, 1992).
Считается, что виды Aspergillus, а в частности A.fumigatus и A.flavus, являются этиологическими факторами ряда заболеваний у человека и животных, называемых аспергиллезом (Bodey & Vartivarian, 1989). И снова было сделано предположение, что протеазы играют определенную роль в вирулентности A.fumigatus и A.flavus, подобно тому, как это наблюдалось во многих исследованиях по взаимосвязи секретированных протеаз и вирулентности бактерий. Действительно, большинство инфекций человека, вызываемых видами Aspergillus, характеризуются интенсивной деградацией паренхимы легкого, которая состоит, главным образом, из коллагена и эластина (Campbell et al., 1994). Исследования были направлены на изучение предполагаемой роли секретированных протеаз в вирулентности A.fumigatus и A.flavus, которые являются главными патогенами человека и, как известно, обладают эластинолитической и коллагеновой активностями (Kolattukudy et al., 1993). Было показано, что эти эластинолитические активности коррелируют in vitro с инфекционностью мышей (Kothary et al., 1984). Известно, что две секретированных протеазы, щелочная сериновая протеаза (ALP) и нейтральная металлопротеаза (МЕР), продуцируются A.fumigatus и A.flavus. Оба гена, кодирующие эти протеазы из A.fumigatus, были выделены, охарактеризованы и подвергнуты дизрупции (Reicherd et al., 1990; Tang et al., 1992, 1993; Jaton-Ogay et al., 1994). Однако было показано, что двойные alp-mep-мутанты по своей патогенности не отличаются от штаммов дикого типа. Поэтому был сделан вывод, что секретированные протеазы A.fumigatus, идентифицированные in vitro, не являются главными факторами для инвазии в ткань (Jaton Ogay et al., 1994). Хотя A.fumigatus ответственен за образование лишь небольшой части спор воздушной плесени, этот грибок чаще всего выделяется из легких и из мокроты (Schmitt et al., 1991). Другим объяснением вирулентности грибков может служить то, что условия в бронхах (температура и питательная среда) являются более благоприятными для паразитарного роста A.fumigatus. Следовательно, если защита хозяина от патогенов ослаблена в результате введения иммунодепрессантов или заболеваниями, такими как СПИД, то инвазивный аспергиллез может быть вызван факторами окружающей среды.
Известны четыре главных класса протеаз, которые определяются главными функциональными группами в их активном сайте, а именно “сериновые”, “тиоловые” или “цистеиновые”, “аспарагиновые” или “карбоксильные” протеазы и “металлопротеазы”. Подробное описание этих больших и малых классов протеаз и неклассифицированных протеаз можно найти в “Methods in Enzymology”, part 244 and 248 (A.J.Barrett ed., 1994 and 1995).
Специфичность протеаз
Помимо каталитического механизма действия протеаз, другим важным аспектом протеолитических ферментов является специфичность протеаз. Специфичность протеазы указывает на конкретные субстраты протеазы, которые, по всей вероятности, гидролизует данная протеаза. Двадцать природных аминокислот дают широкие возможности для конструирования пептидов. Например, с использованием двадцати аминокислот можно сконструировать примерно 400 дипептидов и 800 различных трипептидов и т.п. С использованием более длинных пептидов число возможных конструкций становится почти неограниченным. Некоторые протеазы гидролизуют только конкретные последовательности в высокой степени специфическом положении. Взаимодействие протеазы с пептидным субстратом может охватывать от одного до десяти аминокислотных остатков пептидного субстрата. В случае крупных белковых субстратов взаимодействовать с протеазами может даже большее число остатков субстрата. Однако это, вероятно, приводит к менее специфическим взаимодействиям с остатками протеазы, находящимися за пределами связывающего кармана активного центра. В основном, специфическое распознавание ограничивается линейным пептидом, который связывается в активном сайте протеазы.
Номенклатура для описания взаимодействия субстрата с протеазой была разработана в 1967 году Schechter & Berger (Biochem. Biophys. Res. Com. 1967, 27, 157-162) и в настоящее время широко используется в литературе. Считается, что в этой системе аминокислотные остатки полипептидного субстрата связываются с так называемыми субсайтами в активном сайте. Для удобства эти субсайты на протеазе были обозначены S (для субсайтов), а соответствующие аминокислотные остатки были обозначены Р (для пептида). Аминокислотные остатки с N-концевой стороны от расщепляющейся связи были обозначены Р3, Р2, Р1, а остатки с С-концевой стороны были обозначены P1', P2', P3'. Остатки Р1 или Р1' представляют собой аминокислотные остатки, расположенные возле расщепляющейся связи. Остатки субстрата, расположенные около сайта расщепления, могут быть затем пронумерованы до Р8. Соответствующие субсайты на этой протеазе, которые комплементируют остатки, связывающиеся с субстратом, были пронумерованы S3, S2, S1, S1', S2', S3' и т.п. Предпочтительность субсайтов в сайте связывания с пептидом определяется предпочтением протеазы в расщеплении определенных специфических аминокислотных последовательностей в конкретном месте. Аминокислотная последовательность субстрата должна соответствовать предпочтительности, проявляемой субсайтами. Специфичность по отношению к определенному субстрату явно зависит от аффинности связывания с субстратом и от скорости, при которой эта расщепляемая связь гидролизуется. Поэтому специфичность протеазы к определенному субстрату обычно определяется отношением kcat/Km, которое более известно под термином “константа специфичности”. В этой константе специфичности, kcat означает скорость метаболизма, а Кm означает константу диссоциации.
Помимо аминокислотных остатков, участвующих в катализе и связывании, протеазы содержат множество других основных аминокислотных остатков. Некоторые остатки имеют важное значение для укладки, некоторые остатки поддерживают всю трехмерную структуру протеазы, некоторые остатки могут участвовать в регуляции протеолитической активности, а некоторые остатки могут нацеливать протеазу на конкретный участок. Многие протеазы содержат, за пределами активного сайта, один или более сайтов связывания с ионами металлов. Эти ионы металлов, в большинстве случаев, играют определенную роль в стабилизации структуры. Кроме того, секретированные эукариотические микробные протеазы могут быть в высокой степени гликозилированы. При этом может происходить как N-, так и О-связанное гликозилирование. Гликозилирование может способствовать укладке белка, может повышать растворимость, а также предотвращать агрегацию и, таким образом, стабилизировать зрелый белок. Кроме того, степень гликозилирования может влиять на секрецию, а также на связывание с водой под действием белка.
Регуляция протеолитической активности
Во избежание нежелательного протеолитического расщепления большое число протеаз подвергается интенсивной регуляции протеолитической активности. До определенной степени эта регуляция происходит на транскрипционном уровне. Так, например, в грибах транскрипция генов секретированной протеазы очевидно является чувствительной к внешним углеродным и азотным источникам, тогда как гены, кодирующие внутриклеточные протеазы, не являются чувствительными к таким источникам. Грибы чувствительны к внеклеточному рН, и некоторые гены регулируются рН. В этом процессе транскрипционные регуляторные белки играют решающую роль. Протеолитический процессинг таких регуляторных белков часто служит “переключателем”, который либо “включает”, либо “отключает” регуляторные белки.
Протеазы подвергаются внутримолекулярной, а также межмолекулярной регуляции. Это означает, что некоторые аминокислоты в молекуле протеолитического фермента играют главную роль в такой регуляции. Протеазы обычно синтезируются в виде более крупных предшественников, известных как зимогены, которые являются каталитически неактивными. Обычно удлинение пептидной цепи, которое делает протеазу-предшественника неактивной, локализуется у аминоконца данной протеазы. Этот предшественник больше известен как про-белок. Поскольку многие протеазы, процессированные таким способом, секретируются из клеток, то они, кроме того, содержат сигнальную последовательность (пре-последовательность) для того, чтобы полноразмерный предшественник синтезировался как пре-про-белок. Помимо того, что такая протеаза является неактивной, этот про-пептид часто играет важную роль в опосредовании укладки продукта. Примерами протеаз являются сериновые протеазы (альфалитическая протеаза, субтилизин, аквализин, прогормон-конвертаза), тиоловые протеазы (катепсин L и крузиан), аспарагиновая протеаза (протеиназа А и катепсин D) и металлопротеазы. Кроме того, про-пептид может играть определенную роль в клеточном транспорте либо отдельно, либо в сочетании с сигнальными пептидами. Он может облегчать взаимодействие с клеточными шаперонами, либо он может облегчать транспорт через мембрану. Размер удлиняющего участка в пре-про-белке-предшественнике может значительно варьироваться, от короткого пептидного фрагмента до полипептида, и этот участок может существовать в виде автономной единицы укладки. В частности, как наблюдается в большинстве случаев, эти более крупные удлиняющие участки являются сильными ингибиторами протеазы даже после отщепления от протеазы. Наблюдалось, что даже после отщепления такие про-пептиды могут способствовать соответствующей укладке протеазы. Можно предположить, что такие про-пептиды действуют как молекулярные шапероны и отдельная или совокупная ко-экспрессия таких про-пептидов может оказаться предпочтительной для продуцирования протеазы.
В уровнях регуляции протеаз, которые секретируются в среду, и протеаз, которые остаются внутри клетки, имеется значительное различие. Обычно, после активации, протеазы, секретируемые в среду, больше не подвергаются регуляции, а поэтому они имеют относительно простую молекулярную структуру, состоящую из одного глобулярного модуля. Внутриклеточные протеазы должны подвергаться непрерывной регуляции во избежание повреждения клеток. В отличие от зимогенов секретируемых протеаз, в более сложных регуляторных протеазах, между сигнальной последовательностью и доменом активации зимогена протеолитического модуля могут быть введены очень крупные полипептидные сегменты. Исследование структуры и функции указывает на то, что такие непротеазные части могут участвовать во взаимодействиях с макроскопическими структурами, мембранами, кофакторами, субстратами, эффекторами, ингибиторами и ионами, которые регулируют активность и активацию протеолитического(их) модуля(ей) или его (их) зимогена(ов). Такие непротеолитические модули значительно варьируются по своим размерам и структурам. Многие из этих модулей могут существовать независимо от протеолитического модуля. Поэтому можно считать, что такие модули не зависят от структурных и функциональных единиц, которые являются автономными в отношении укладки. Ценность такой модульной организации заключается в том, что приобретение новых модулей может наделять протеазу-реципиента новой специфичностью связывания и может приводить к резкому изменению ее активности, регуляции и нацеливания. Принцип модульной организации протеолитических ферментов может быть также использован в методах молекулярной биологии для обеспечения новых взаимодействий, регуляции, специфичности и/или нацеливания путем “перетасовки” модулей. Хотя, в основном, такие дополнительные модули присутствуют как N- или С-концевые удлинения, однако наблюдались также крупные вставки во внешних петлях каталитического домена. Очевидно, что в этом случае также главная укладка протеазы представляет основную топологию для образования функциональной протеолитической молекулы и что эта вставка может рассматриваться как субструктура, уложенная на поверхности протеолитического модуля.
Молекулярная структура
В принципе, главной темой обсуждения является модульная организация более крупных природных белков. В частности, в более крупных мультимодульных каркасах типичные протеолитические модули имеют размеры в среднем от 100 до 400 аминокислот. Это соответствует среднему размеру большинства глобулярных протеолитических ферментов, которые секретируются в среду. Как обсуждалось выше, полипептидными модулями являются полипептидные фрагменты, которые могут быть уложены и могут функционировать как независимые молекулы. Другим названием таких модулей является “домены”. Однако термин “домен” используется в более широком контексте, чем модуль. Используемый здесь термин “домен” обычно означает часть полипептидной цепи, которая имеет типичную топологию укладки в трехмерной структуре. В белке домены взаимодействуют на различных уровнях, но менее интенсивно, чем структурные элементы внутри доменов. В литературе также используются и другие термины, такие как субдомен и единица укладки. Фактически наблюдалось, что многие белки, которые имеют одни и те же конкретные функциональные группы, могут иметь одинаковые домены. Такие домены могут распознаваться по их первичной структуре, которая может иметь последовательности определенного типа, являющиеся характерными для данного домена. Типичными примерами являются мононуклеотид-связывающая складка; домены, связывающиеся с целлюлозой; мотив связывания ДНК “спираль-виток-спираль”; домены “цинковые пальцы”; ЕF-плечи; мембранные якори. Термин “модули” означает домены, которые, как предполагается, способны к автономной укладке и функции. Специалисту известны способы идентификации конкретных доменов в первичной структуре с использованием общедоступных компьютерных программ для указанной структуры и гомологичных последовательностей, происходящих от других организмов или видов.
Хотя мультимодульные или мультидоменные белки могут представлять собой цепочку типа “нитки бус”, однако наблюдалась и значительно более сложная архитектура. В случае если на одной и той же полипептидной цепи присутствуют различные “бусинки”, то эти “бусинки” обычно называются модулями или доменами. Если “бусинки” не присутствуют на одной и той же полипептидной цепи, но образуют упорядоченную структуру посредством нековалентных взаимодействий, то в этом случае используют термин “субъединица”. Субъединицы могут транскрибироваться одним и тем же геном или различными генами. После транскрипции мультимодульный белок может подвергаться протеолитическому процессингу, приводящему к образованию множества субъединиц. Отдельные субъединицы могут состоять из множества доменов. В основном, более мелкие глобулярные белки, содержащие 100-300 аминокислот, обычно состоят только из одного домена.
Молекулярная классификация протеолитических ферментов
В основном, протеазы классифицируют по их молекулярным свойствам или по их функциональным свойствам. Молекулярная классификация основана на первичной структуре протеазы. Первичная структура белка представляет его аминокислотную последовательность, которая может происходить от нуклеотидной последовательности соответствующего гена. Интенсивные исследования, касающиеся сходства первичных структур, могут привести к выявлению сходства каталитического механизма и других свойств, которые могут даже относиться к функциональным свойствам. Используемый здесь термин “семейство” означает группу протеаз, которые обнаруживают эволюционное родство, устанавливаемое исходя из сходства их первичных структур. Очевидно, что члены такого семейства произошли от одного и того же предшественника, но затем подверглись дивергенции в процессе эволюции. Кроме того, внутри семейства эти протеазы подразделяются на подгруппы по своим первичным структурам, которые были установлены путем более детального уточнения их последовательности, исходя из результатов сравнения подсемейств. Классификация по трехмерной укладке протеаз может включать вторичную структуру, третичную структуру и четвертичную структуру. В общих чертах, классификация по вторичной структуре ограничивается содержанием и приблизительной ориентацией элементов вторичной структуры. Сходство третичных структур позволяет идентифицировать суперсемейства или кланы. Суперсемейство или клан представляет собой группу семейств, которые, очевидно, имеют общего “предка”, поскольку они обнаруживают аналогичную 3-мерную укладку. В основном, трехмерная структура является более консервативной, чем первичная структура. Следовательно, сходство первичной структуры не всегда отражает аналогичные функциональные свойства. Действительно, функциональные свойства могут претерпевать значительные изменения, что может приводить к появлению новых представляющих интерес свойств. В настоящее время четвертичная структура не используется для классификации различных протеаз. Это может быть обусловлено некоторым смещением структурных баз данных в сторону простых глобулярных протеаз. Многие протеолитические системы, которые подвергаются активации, регуляции или сложным каскадам реакций, вероятно, состоят из множества доменов или субъединиц. Общие тенденции структурной организации таких протеазных систем могут приводить к новым типам классификации.
Классификация в соответствии со специфичностью
В отсутствие информации о последовательности протеаз они могут быть классифицированы по различным типам их функций. Классификация и названия ферментов в соответствии с реакциями, которые они катализируют, являются главным принципом в номенклатуре ферментов. Такой подход также основан на принципе ЕС-нумерации ферментов (Enzyme Nomenclature 1992 Academic Press, Orlando). Два типа протеаз (ЕС 3.4) могут быть определены в соответствии с номенклатурой ферментов Enzyme Nomenclature 1992, т.е. экзопептидазы (ЕС 3.4.11-19) и эндопептидазы (ЕС 3.4.21-24, 3.4.99). Эндопептидазы расщепляют пептидные связи на внутренних участках пептидной цепи, которые находятся далеко от ее конца. Экзопептидазы отщепляют остатки лишь от концов пептидной цепи. Экзопептидазы, действующие у свободного N-конца, могут высвобождать один аминокислотный остаток, дипептид или трипептид и называются соответственно аминопептидазами (ЕС 3.4.11), дипептидилпептидазами (ЕС 3.4.14) и трипептидилпептидазами (ЕС 3.3.14). Протеазы, запускающие процессинг пептида от карбоксильного конца, приводящий к высвобождению одной аминокислоты, называются карбоксипептидазами (ЕС 3.4.16-18). Пептидил-дипептидазы (ЕС 3.4.15) отщепляют дипептид от карбоксильного конца. Экзо- и эндопептидазы, в целом, представляют собой дипептидазы (ЕС 3.4.13), которые специфически расщепляют только дипептиды в их двух аминокислотных половинах. Омега-пептидазы (ЕС 3.4.19) удаляют концевые остатки, которые являются замещенными, циклическими или присоединены изопептидными связями.
Помимо положения, в котором данная протеаза расщепляет пептидную цепь, для каждого типа протеаз возможно и дополнительное разделение в соответствии с природой предпочтительных аминокислотных остатков в субстрате. В основном, протеазы могут быть классифицированы по широкой, средней и узкой специфичности. Некоторые протеазы просто называют по специфическим белкам или полипептидам, которые они гидролизуют, например, кератиназа, коллагеназа, эластаза. Узкая специфичность может ограничиваться одной конкретной аминокислотой или одной конкретной последовательностью, которая может быть удалена или расщеплена, соответственно. Если протеаза обнаруживает особое предпочтение одной аминокислоте в Р1- или Р1'-положении, то название этой аминокислоты может быть ее определителем. Так, например, пролиламинопептидаза удаляет пролин из аминоконца пептида (пролин представляет собой остаток Р1). Х-Pro или пролин используются в том случае, если расщепляется связь на боковой иминоцепи этого пролина (пролин представляет собой остаток Р1'), например, пролинкарбоксипептидаза удаляет пролин у карбоксиконца. Пролилэндопептидаза (или Pro-Х) расщепляет в положении позади пролина, а пролинэндопептидаза (Х-Pro) расщепляет в положении впереди пролина. Аминокислотный остаток, находящийся перед расщепляемой пептидной связью, означает аминокислотный остаток, который вводит карбоксильную группу в пептидную связь. Аминокислотный остаток, находящийся позади расщепляемой пептидной связи, означает аминокислотный остаток, который вводит аминогруппу в пептидную связь. Согласно общепринятому соглашению, аминокислотная цепь простирается от аминоконца (начало) до карбоксиконца (конец), и в соответствии с этим осуществляют нумерацию аминокислот. Эндопротеазы могут также оказывать явное предпочтение конкретной аминокислоте в положении Р1 или Р1', например, глицилэндопептидаза, пептидиллизинэндопептидаза, глутамилэндопептидаза. Кроме того, протеазы могут оказывать предпочтение определенной группе аминокислот, которые обладают некоторым сходством. Такая группа предпочитаемых аминокислот может включать гидрофобные аминокислоты, только объемные гидрофобные аминокислоты, небольшие гидрофобные аминокислоты или только небольшие аминокислоты, крупные положительно заряженные аминокислоты и т.п. Помимо предпочтения к остаткам Р1 и Р1', может также наблюдаться особая или исключительная предпочтительность к остаткам, предпочитаемым другими субсайтами на данной протеазе. Такая множественная предпочтительность может приводить к тому, что протеазы будут в высокой степени специфичными только к последовательностям, которые одновременно удовлетворяют множеству требований к связыванию. Вообще говоря, следует отметить, что протеазы являются скорее неспецифическими ферментами. Даже очень специфическая протеаза может расщеплять пептиды, которые не являются объектом обычно наблюдаемого предпочтения протеазы. Кроме того, следует отметить, что на предпочтения протеаз могут влиять окружающие условия, такие как рН, температура, ионная сила, активность воды, присутствие растворителей, присутствие конкурирующих субстратов или ингибиторов. Окружающие условия могут влиять не только на протеазу, но и также на путь представления белкового субстрата протеазе.
Классификация по каталитическому механизму
Протеазы могут быть классифицированы на основе их каталитических механизмов. Следует отметить, что для каждого каталитического механизма вышеуказанная классификация протеаз, основанная на их специфичности, приводит к последующему подразделению протеаз по механизму их действия. Известны четыре основных класса протеаз, которые были названы по их главным функциональным группам в активном сайте, а именно сериновые протеазы (ЕС 3.4.21 эндопептидаза, ЕС 3.4.16 карбоксипептидаза), тиоловые или цистеиновые протеазы (ЕС 3.4.22 эндопептидаза, ЕС 3.4.18 карбоксипептидаза), карбоксильная или аспарагиновая протеазы (ЕС 3.4.23 эндопептидаза) и металлопротеаза (ЕС 3.4.24 эндопептидаза, ЕС 3.4.18 карбоксипептидаза). Для членов каждого каталитического типа протеазы имеются характерные ингибиторы. Эти небольшие ингибиторы обратимо модифицируют аминокислотный остаток активного сайта протеазы. Так, например, сериновая протеаза инактивируется фенилметансульфонилфторидом (PMSF) и диизопропилфторфосфатом (DFP), которая реагирует с активным серином, тогда как хлорметилкетоновые производные реагируют с гистидином каталитической триады. Фосфорамидон и 1,10-фенантролин обычно ингибируют металлопротеазы. Ингибирование пепстатином обычно указывает на аспарагиновую протеазу. Е64 специфически ингибирует тиоловую протеазу. Амастатин и бестатин ингибируют различные аминопептидазы. При этом наблюдаются значительные различия в восприимчивости протеаз к ингибиторам, даже внутри одного каталитического класса. До некоторой степени это может быть связано со специфичностью протеазы. В случае, когда архитектура сайта связывания препятствует механизму, в соответствии с которым ингибитор воздействует на каталитический сайт, то такая протеаза “ускользает” от ингибирования, и идентификация по типу механизма ингибирования не может быть осуществлена. Так, например, химостатин является сильным ингибитором сериновой протеазы со специфичностью, подобной специфичности химотрипсина. Эластатинал ингибирует эластазоподобные сериновые протеазы и не реагирует с трипсином или химотрипсином; 4-амидо-PMSF (АРМSF) ингибирует только сериновые протеазы со специфичностью, подобной трипсину. Широкое обсуждение использования ингибиторов в классификации протеаз приводится в работах Barret and Salvesen, Proteinase Inhibitors, Elsevier Amstardam, 1986; Bond and Beynon (eds), Proteolytic Enzymes, A Practical Approach, IRL Press, Oxford, 1989; Methods in Enzymology, eds E.J.Barret, volume 244, 1994 and volume 248, 1995; E.Shaw, Cysteinyl proteinases and their selective inactivation, Adv Enzymol. 63:271-347 (1990).
Классификация по оптимальным условиям функционирования
Каталитический механизм действия протеаз и требования к их конформационной целостности определяют, главным образом, условия, при которых могут быть использованы данные протеазы. Установление оптимальных условий применения протеазы является главной задачей. Часто условия, при которых протеазы должны функционировать, не являются оптимальными, и необходимо найти компромисс между идеальными условиями для данного конкретного применения и условиями, которые должны быть наиболее подходящими для данной протеазы. Помимо конкретных свойств протеазы, следует также учесть, что презентация белковых субстратов зависит от условий, и, по существу, она также определяет условия, которые являются наиболее эффективными для протеолиза. Параметрами фермента, подходящего для его использования, являются, например, зависимость от рН, зависимость от температуры, чувствительность к ионам металла или зависимость от ионов металла, ионная сила, концентрация соли и совместимость с растворителем. Другим наиболее важным фактором является специфическая активность протеазы. Чем большую специфическую активность имеет данный фермент, тем меньше он нуждается в специфической конверсии. Чем ниже требования к ферменту, тем ниже его стоимость и ниже уровни белковых примесей.
рН является главным параметром, который определяет функционирование протеазы, используемой в определенных целях. Поэтому зависимость от рН является важным параметром для группы протеаз. Основными группами, на которые обычно делят протеазы, являются кислотные протеазы, нейтральные протеазы, щелочные протеазы и в высокой степени щелочные протеазы. Оптимальный рН соответствует протеолитическому механизму лишь до определенной степени, например, в большинстве случаев аспарагиновая протеаза оптимально действует лишь при кислотном рН; металлопротеазы и тиоловые протеазы оптимально действуют примерно от нейтрального до слабощелочного рН; сериновые протеазы являются активными, главным образом, в щелочной и высокощелочной среде. Для каждого класса протеаз известны исключения. Помимо этого, определенную роль играет активность воды в данной системе. Оптимальный рН протеазы определяется как диапазон рН, при котором данная протеза имеет оптимальную скорость гидролиза для большинства из ее субстратов в конкретной среде и в конкретных условиях. Этот диапазон может быть низким, например в одну рН-единицу, а также достаточно широким, в 3-4 единицы. В основном, оптимальный рН также зависит от природы белкового субстрата. Скорость функционального цикла протеазы, а также ее специфичность могут варьироваться в зависимости от рН. Для некоторых целей может оказаться желательным использовать протеазу при рН, значения которых далеки от оптимальных, например, в том случае, когда необходимо избежать продуцирования менее желательных пептидов. Такими менее желательными пептидами могут быть, например, очень короткие пептиды или пептиды, дающие горький привкус. Кроме того, более узкая специфичность может служить основанием для выбора условий, которые не соответствуют оптимальным условиям в отношении скорости функционального цикла. В зависимости от рН специфичность протеазы может быть узкой, например, она может расщеплять пептидную цепь лишь в одном конкретном положении впереди или позади конкретной аминокислоты, либо она может быть более широкой, например, когда протеаза расщепляет цепь во множестве положений, либо впереди или позади аминокислот множества типов. Действительно, зависимость от рН может служить важным инструментом для регуляции протеолитической активности в определенных целях. В случае, когда рН изменяется в процессе протеолиза, такой протеолиз может прекращаться спонтанно и не требует дополнительной обработки для инактивации протеазы. В некоторых случаях сам протеолиз может быть регулятором изменения рН.
При использовании протеаз очень важным фактором является их технологическая и функциональная стабильность. Поскольку стабильность протезы в значительной степени зависит от рабочей температуры, то этот параметр часто также называют термостабильностью. Вообще говоря, стабильность протеазы указывает на срок сохранения протеолитической активности протеазы в данных конкретных условиях. Такие конкретные условия могут включать условия ферментации, условия во время выделения и последующей обработки фермента, условия хранения, условия приготовления и функционирования фермента или условия его применения. В случае если конкретные условия включают стабильность при повышенных температурах, то такая стабильность, в основном, называется термостабильностью. Помимо общеизвестных причин инактивации фермента, таких как химическая модификация, развертывание молекулы фермента, ее агрегация и т.п., главная проблема заключается в том, что эти ферменты легко подвергаются аутодеградации. Поэтому при использовании протеаз оптимальная температура является важным параметром для группы протеаз. Хотя существуют различные определения, однако, с экономической точки зрения, наиболее часто используемым определением оптимальной температуры является температура, при которой протеаза является наиболее продуктивной для данной цели применения. Продуктивность протеазы зависит как от стабильности, так и от скорости ее функционального цикла. В то время как повышенная температура, в основном, увеличивает скорость цикла, быстрая инактивация приводит к снижению скорости цикла и, в конечном счете, к низкой продуктивности. Конформационная стабильность протеазы в данных условиях процесса будет определять ее максимальную рабочую температуру. Температура, при которой данная протеаза теряет свою активную конформацию, часто рассматривается как точка развертывания или плавления и может быть определена различными методами, например, с помощью ЯМР, спектроскопии кругового дихроизма, дифференциальной сканирующей калориметрии и т.п. Развертывание молекулы протеазы обычно сопровождается резким увеличением скорости ее аутодеградации.
В случаях если требуются низкие температуры, то при выборе протеазы следует ориентироваться на ее высокую внутреннюю активность при низких или умеренных температурах. Поскольку при таких условиях инактивация протеазы происходит относительно медленно, то активность при этих условиях может в значительной степени определять ее продуктивность. В процессах, где активность протеазы требуется только в очень короткий период времени, стабильность протеазы может быть использована для “включения” или “отключения” протеазы. В этом случае вместо в высокой степени термостабильной протеазы может оказаться предпочтительней использовать более лабильную протеазу.
Другими параметрами окружающей среды, которые могут играть определенную роль в выборе подходящей протеазы, может быть ее чувствительность к солям. Совместимость с ионами металлов, которые часто присутствуют в низких концентрациях в различных природных веществах, может оказаться решающим фактором для некоторых применений. Что касается металлопротеаз, то некоторые ионы могут заменять каталитический ион металла и снижать или даже полностью подавлять активность протеазы. При некоторых применениях, для предотвращения утечки ионов металла, координированных с протеазой, необходимо добавить эти ионы металлов. Хорошо известно, что для поддержания стабильности фермента и сохранения срока его действия необходимо добавлять ионы кальция в целях предупреждения диссоциации кальция, связанного с белком.
Большинство микроорганизмов обнаруживают определенную толерантность и адаптируются к изменениям условий окружающей среды. Следовательно, как было показано исходя из протеолитического спектра такого микроорганизма, он способен вырабатывать, по крайней мере, аналогичную толерантность. Такой протеолитический спектр может покрываться многими протеазами, которые в совокупности закрывают “окно” спектра, либо он только может покрываться несколькими протеазами широкого спектра. Если принять во внимание весь протеолитический спектр микроорганизма, то важное значение может иметь локализация.
Клеточная локализация и характеризация протеолитического процессинга и деградации
С точки зрения промышленного применения, протеазы, которые экскретируются из клетки, имеют конкретные преимущества в отношении крупномасштабной продуктивности и толерантности к стрессам, поскольку они являются жизнеспособными в отсутствие клеточной защиты. Большая группа клеточных протеаз может быть, кроме того, подразделена на растворимые и мембрано-ассоциированные протеазы. Группа мембрано-ассоциированных протеаз может включать протеазы, ассоциированные с мембраной как с внутренней, так и с внешней стороны. Внутриклеточные растворимые протеазы могут быть, кроме того, подразделены в соответствии со специфическими компартментами клетки, в которых они присутствуют. Поскольку клетка, до некоторой степени, защищает протеазы от воздействия внешней среды и поскольку клетка регулирует имеющиеся в ней условия, то внутриклеточная протеаза может оказаться более чувствительной к значительным изменениям условий окружающей среды, и, в этом случае, оптимальные параметры могут лучше коррелировать со специфическими условиями внутри клетки. Знание условий клеточного компартмента, в котором присутствует данная протеаза, может указывать на ее предпочтительность. Если внеклеточная протеаза, в основном, не требует какой-либо регуляции сразу после ее экскрекции из клетки, то внутриклеточные протеазы часто подвергаются более тонкому контролю и регуляции.
Что касается функции конкретной протеазы, то ее локализация часто имеет очень важное значение, так, например, большое число вакуолярных и периплазматических протеаз участвует в разложении белка, тогда как мембрано-ассоциированные протеазы играют важную роль в процессинге белка (Suarez Rendueles & Wolf, 1988).
Полный обзор биологических свойств и эволюции протеаз был опубликован в работе van den Homberg: Thesis Landbouwuniversitet Wageningen: An analysis of proteolytic system in Aspergillus in order to improve protein production ISBN 90-5485-545-2, которая вводится в настоящее описание посредством ссылки.
Проблемы, связанные с использованием протеаз
Важной причиной интереса, проявляемого к микробным протеазам, являются проблемы, связанные с экспрессией протеаз в некоторых экспрессирующих хозяевах, используемых в отрасли промышленности, ориентированной на биологические процессы. В последние годы, в связи с возрастанием применения гетерологичных хозяев в целях продуцирования белков методами рекомбинантных ДНК, возникли определенные проблемы, поскольку, как оказалось, гетерологичные белки более подвержены протеолизу (Archer et al., 1992; van den Homberg et al., 1996b).
Что касается применения S. cerevisiae, то еще в восьмидесятые годы возникли проблемы и трудности, связанные с использованием некоторых протеаз, а поэтому для решения этой проблемы был разработан способ, предусматривающий дизрупцию целевого гена-мишени. Во время секреции белок подвергается нескольким протеолитическим активностям, действующим на данном секреторном пути. Кроме того, у прототрофного микроорганизма, такого как Aspergillus, секретированный белок может подвергаться нескольким внеклеточным протеолитическим активностям.
Проблема деградации гетерологически экспрессированных белков в Aspergillus хорошо освещена в литературе (van den Hombergh: Thesis Landbouwuniversitet Wageningen: An analysis of the proteolitic system in Aspergillus in order to improve protein production ISBN 90-5485-545-2), и сообщалось, что такая деградация наблюдалась при экспрессии коровьего прохимозина, человеческого интерферона α-2-tHA, GМCSF, IL6, лактоферрина, лизоцима куриного яичного белка, свиного рIА2, пектин-лиазы В A. niger, энтеротоксина В и β-глюкоронидазы E.coli и пектат-лиазы 3 Erwinia carobotovora.
Проблема протеолиза может быть связана с несколькими стадиями продуцирования белка. Специалисты по биологическим процессам, которые сталкиваются с проблемой протеолиза, могут проводить обработку при низких температурах, осуществлять раннее выделение продукта и протеазы (протеаз) или использовать ингибиторы протеазы. Все это может приводить к успешному решению данной проблемы. Однако все эти трудности не могут быть полностью устранены, поскольку, в большинстве случаев, деградация происходит in vivo во время продуцирования белка.
Для осуществления регуляции протеолиза в клетке важно понять механизм запуска протеолиза. До определенной степени (гетерологичные) белки являются протеолитически чувствительными и распознаваемыми как аберрантные белки из-за их ошибочной укладки или, в случае правильной укладки, как “чужеродные” белки из-за того, что они не обладают важными для их стабильности признаками, которые являются специфическими для хозяина. Различные типы стрессов могут вызывать общее значительное увеличение протеолиза в клетке. Известными факторами увеличения скорости протеолиза являются клеточное “голодание” и различные типы стрессов (то есть повышение температуры, осмотический стресс, токсические вещества и экспрессия некоторых гетерологичных белков). Для успешного решения проблем, связанных с протеолизом in vivo, было предложено несколько способов, которые будут обсуждаться ниже. Однако авторы настоящего изобретения обращают внимание на то, что не может существовать абсолютно “непротеолитических клеток”, поскольку протеолиз под действием внутриклеточных протеаз является звеном многих важных метаболических и конститутивных реакций (реакций “домашнего хозяйства”). Поэтому снижение степени протеолиза всегда представляет собой процесс, в котором происходит изменение генетического фона, приводящее к снижению протеолитической активности, и этот процесс необходимо анализировать на возможные побочные эффекты, которые могут приводить к снижению продуцирования белка (например, на снижение скорости роста или споруляцию).
Дизрупция протеаз, экспрессирующихся в нитчатых грибах-хозяевах
Berka и сотрудниками (1990) было описано клонирование и дизрупция гена рерА А.awamori. Совсем недавно были описаны три дизруптированные аспартилпротеазы в A.niger. Были описаны дизруптанты как для основных внеклеточных аспартилпротеаз, так и для основных вакуолярных аспартилпротез. Двойные и тройные дизруптанты были генерированы посредством рекомбинации и протестированы на протеазный спектр, а также на экспрессию и секрецию пектинлиазы белка РЕLВ A.niger, который является очень чувствительным к протеолитической деградации (van den Hombergh et al., 1995). Дизрупция генов рерА и рерВ приводила к снижению активностей этих внеклеточных протеаз на 80% и на 6% соответственно. В дизруптанте ΔрерЕ, активности других (вакуолярных) протеаз были также значительно снижены, что обусловлено инактивацией протеолитического каскада реакций для других вакуолярных протеаз. Пониженные внеклеточные активности коррелировали со снижением уровня in vitro деградации РЕLВ и с увеличением уровня in vivo экспрессии реlВ (van den Hombergh et al., 1996f).
Дефицитные по протеазе (prt) мутанты нитчатых грибов
Несколько дефицитных по протеазе мутантов Aspergillus были исследованы на увеличение уровня продуцирования белка. Archer и сотрудниками был описан пониженный уровень протеолиза лизоцима куриного яичного белка в супернатантах prt-двойного мутанта A.niger, генерированного Mattern и сотрудниками (1992), и был сделан вывод, что хотя деградация имела место, однако она была значительно снижена. Van den Hombergh et al. (1995) показали, что in vitro-деградация РЕLВ A.niger была снижена у всех выделенных ими семи групп с prt-комплементацией. Фактически у prtВ-, prtF- и prtG-мутантов не наблюдалось какой-либо деградации. Недавно было показано, что экспрессия гена реlВ усиливалась у шести протестированных комплементированных групп (prtА-F), и самый высокий уровень экспрессии наблюдался у prtВ-, prtF- и prtG-мутантов. Помимо мутантов, дефицитных по одному гену, которые содержали остаточные внеклеточные протеолитические активности, варьирующиеся от 2% до 80% по сравнению с активностью дикого типа, были генерированы двойные мутанты посредством рекомбинации и дополнительных раундов мутагенеза. Такими способами были отобраны и охарактеризованы несколько двойных prt-мутантов, и было показано, что они обнаруживали еще более низкую степень деградации РЕLВ по сравнению с родительскими штаммами.
Кроме подавления протеазной активности посредством дизрупции или мутагенеза, снижение протеолиза может быть достигнуто посредством негативной регуляции ненужной протеолитической активности. Это может быть осуществлено путем генетической модификации промотора или других регуляторных последовательностей гена. Как было показано Fraissinet-Tachet и сотрудниками (1996), все внеклеточные протеазы в A.niger регулировались ингибированием углеродного катаболита и ингибированием азотного метаболита. Клеточное голодание также приводило к общему значительному увеличению уровня протеолиза в клетке, что имело важное значение для клеток, не содержащих питательные вещества, но содержащих белки, которые не являются необходимыми в условиях голодания или являются необходимыми только в небольших количествах. Сообщалось, что при использовании стратегий экспрессии, которые дают высокие уровни экспрессии на средах, содержащих высокие концентрации глюкозы и аммония, протеолиз снижался. Некоторые конститутивные гликолитические промоторы (gpd и pkiA) обеспечивают высокую степень экспрессии в этих условиях и могут быть также использованы для регуляции экспрессии (гетерологичного) гена при непрерывной ферментации. Тип принудительного клеточного голодания может в различной степени влиять на различные протеазы, что указывает на важность условий питания клетки в данном процессе в зависимости от типа протеолиза. Поэтому специфический протеолиз может быть индуцирован условиями субстратного ограничения, которые часто используются в процессах крупномасштабной промышленной ферментации.
В настоящее время для решения проблемы, связанной с протеазой, обращаются к одной или более из вышеперечисленных стратегий. Однако остаточная протеолитическая активность еще неидентифицированных протеолитических ферментов пока составляет главную проблему для специалистов. Для дополнительного снижения уровня нежелательного протеолиза крайне необходимо идентифицировать новые протеазы, ответственные за деградацию гомологично и гетерологично экспрессированных белков. Настоящее изобретение относится к таким новым последовательностям генов протеазы, кодирующих новые протеазы. Как только станет известна первичная последовательность гена новой протеазы, то одна или более из вышеуказанных стратегий рекомбинантной ДНК могут быть использованы для продуцирования (нокаут)-мутантов со сниженной протеолитической активностью.
Несмотря на широкое применение протеаз в большинстве промышленных процессов, известные ферменты также имеют множество недостатков, касающихся, по крайней мере, одного из нижеследующих свойств.
При добавлении в пищу животного обычно используемые протеазы не обладают достаточной резистентностью к расщепляющим ферментам, присутствующим в желудочно-кишечном (ЖК) тракте, например, свиньи или домашней птицы.
Что касается других аспектов, то имеющиеся ферменты не обладают достаточной устойчивостью к высоким температурам и высокому давлению, которые обычно используются во время операций вытеснения или осаждения.
Кроме того, обычно используемые ферменты недостаточно активны при рН в пределах 3-7, то есть в условиях приготовления многих пищевых продуктов и напитков, а также в ЖК-тракте большинства животных.
В соответствии с другим аспектом специфичность обычно используемых протеаз очень ограничена, что приводит к неспособности присутствующих ферментов разлагать или растворять некоторые “протеазо-резистентные” белки и, тем самым, к низкому выходу пептидов или аминокислот. Кроме того, протеазы с низкой специфичностью могут способствовать синтезу новых пептидов.
Еще одним недостатком обычно используемых ферментов является их низкая специфическая активность.
Исходя из этого, очевидно, что в большинстве случаев требуется использование протеаз, которые являются более резистентными к пищеварительным ферментам, к высоким температурам и/или высокому давлению и которые обладают новыми специфичностями по отношению к сайтам гидролиза. Настоящее изобретение относится к таким ферментам.
Цель изобретения
Целью настоящего изобретения является получение новых полинуклеотидов, кодирующих новые протеазы. Другой целью настоящего изобретения является получение природных и рекомбинантно продуцируемых протеаз, а также рекомбинантных штаммов, продуцирующих эти протеазы. Такие штаммы могут быть также использованы для продуцирования продуктов в процессе классической ферментации с более высокой скоростью или с более высоким выходом. Еще одной целью настоящего изобретения является получение штамма нитчатых грибов, дефицитных по продуцированию протеаз настоящего изобретения. Такие штаммы могут быть использованы для более эффективного продуцирования гетерологичных или гомологичных белков. Настоящее изобретение также включает антитела и гибридные белки, а также способы получения и использование полинуклеотидов и полипептидов настоящего изобретения.
Краткое описание изобретения
Настоящее изобретение относится к новым полинуклеотидам, кодирующим новые протеазы.
Более конкретно, настоящее изобретение относится к полинуклеотидам, имеющим нуклеотидную последовательность, которая гибридизуется (предпочтительно в условиях высокой жесткости) с последовательностью, соответствующей последовательности, выбранной из группы, состоящей из SEQ ID NO:1 - SEQ ID NO:57, или последовательности, выбранной из группы, состоящей из SEQ ID NO:58 - SEQ ID NO:114. Кроме того, настоящее изобретение относится к нуклеиновым кислотам, которые приблизительно на 60%, предпочтительно на 65%, более предпочтительно на 70% и даже более предпочтительно на 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% гомологичны последовательностям, соответствующим последовательности, выбранной из группы, состоящей из последовательностей SEQ ID NO:1 - SEQ ID NO:57, или последовательности, выбранной из группы, состоящей из SEQ ID NO:58 - SEQ ID NO:114.
В более предпочтительном варианте настоящее изобретение относится к полинуклеотиду, выделенному из нитчатых грибов, предпочтительно из Aspergillus, а в частности из A.niger.
В одном из вариантов осуществления настоящее изобретение относится к выделенному полинуклеотиду, содержащему последовательность нуклеиновой кислоты, кодирующую полипептид, имеющий аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:115 - SEQ ID NO:171, или его функциональный эквивалент.
В другом предпочтительном варианте осуществления настоящее изобретение относится к выделенному полинуклеотиду, кодирующему, по крайней мере, один функциональный домен полипептида, соответствующего последовательности, выбранной из группы, состоящей из последовательностей SEQ ID NO:115 - SEQ ID NO:171 или их функциональных эквивалентов.
В предпочтительном варианте осуществления настоящее изобретение относится к гену протеазы, имеющему последовательность, выбранную из группы, состоящей из SEQ ID NO:1 - SEQ ID NO:57. В другом своем аспекте настоящее изобретение относится к полинуклеотиду, предпочтительно к кДНК, кодирующей протеазу A.niger, выбранную из группы, состоящей из SEQ ID NO:115 - SEQ ID NO:171, или к вариантам или фрагментам этого полипептида. В предпочтительном варианте осуществления изобретения указанная кДНК имеет последовательность, выбранную из группы, состоящей из SEQ ID NO:58 - SEQ ID NO:114 или их функциональных эквивалентов.
Геномный клон, кодирующий полипептид настоящего изобретения, может быть также получен путем выбора подходящих зондов для специфической амплификации геномной области, соответствующей любой из последовательностей SEQ ID NO:1 - SEQ ID NO:57 или ее фрагментов; гибридизации этого зонда в подходящих условиях с геномной ДНК, полученной из подходящего организма, такого как Aspergillus, например, A.niger, амплификации нужного фрагмента, например, с помощью ПЦР (полимеразная цепная реакция) с последующей очисткой и клонированием амплифицированного фрагмента.
В еще более предпочтительном варианте осуществления настоящее изобретение относится к полинуклеотиду, включающему кодирующую последовательность геномных полинуклеотидов настоящего изобретения, при этом предпочтительной является полинуклеотидная последовательность, выбранная из группы, состоящей из последовательностей SEQ ID NO:58 - SEQ ID NO:114.
В другом предпочтительном варианте осуществления настоящее изобретение относится к кДНК, полученной путем клонирования и экспрессии последовательности, выбранной из группы, состоящей из SEQ ID NO:1 - SEQ ID NO:57, в подходящем организме-хозяине, таком как A.niger.
Полипептид настоящего изобретения может также быть получен путем клонирования и экспрессии последовательности, выбранной из группы, состоящей из SEQ ID NO:1 - SEQ ID NO:57 в подходящем организме-хозяине, таком как A.niger.
Настоящее изобретение также относится к векторам, содержащим полинуклеотидную последовательность настоящего изобретения, и к праймерам, зондам и фрагментам, которые могут быть использованы для амплификации или для детекции ДНК настоящего изобретения.
В другом предпочтительном варианте осуществления настоящее изобретение относится к вектору, в котором полинуклеотидная последовательность настоящего изобретения функционально связана с регуляторными последовательностями, подходящими для экспрессии кодируемой аминокислотной последовательности в подходящей клетке-хозяине, такой как A.niger или A.oryzea. Настоящее изобретение также относится к способам получения полинуклеотидов и векторов настоящего изобретения.
Кроме того, настоящее изобретение относится к рекомбинантно продуцированным клеткам-хозяевам, которые содержат гетерологичные или гомологичные полинуклеотиды настоящего изобретения.
В одном из вариантов своего осуществления настоящее изобретение относится к рекомбинантным клеткам-хозяевам, где значительно снижена экспрессия протеазы настоящего изобретения или снижена ее активность или где указанная протеаза даже является инактивированной. Такие рекомбинанты особенно предпочтительны для экспрессии гомологичных или гетерологичных белков.
В другом варианте осуществления настоящее изобретение относится к рекомбинантным клеткам-хозяевам, в которых уровень экспрессии протеазы настоящего изобретения значительно увеличен или увеличена ее активность. Такие рекомбинанты являются особенно предпочтительными для экспрессии гомологичных или гетерологичных белков, созревание которых значительно затруднено в случае, когда требуемое протеолитическое расщепление становится скорость-ограничивающей стадией.
В другом варианте осуществления настоящее изобретение относится к рекомбинантно продуцированной клетке-хозяину, которая содержит гетерологичную или гомологичную ДНК настоящего изобретения, а предпочтительно ДНК, кодирующую белки, несущие сигнальные последовательности, где указанная клетка способна продуцировать функциональную протеазу настоящего изобретения, а предпочтительно сверхэкспрессировать протеазу настоящего изобретения, где указанная клетка является, например, штаммом Aspergillus, содержащим увеличенное число копий гена или кДНК настоящего изобретения.
В другом своем варианте настоящее изобретение относится к рекомбинантно продуцированной клетке-хозяину, которая содержит гетерологичную или гомологичную ДНК настоящего изобретения, где указанная клетка способна секретировать функциональную протеазу настоящего изобретения, а предпочтительно сверхэкспрессировать и секретировать протеазу настоящего изобретения, и где указанная клетка является, например, штаммом Aspergillus, содержащим увеличенное число копий гена или кДНК настоящего изобретения.
В еще одном аспекте настоящее изобретение относится к очищенному полипептиду. Полипептидами настоящего изобретения являются полипептиды, кодируемые полинуклеотидами настоящего изобретения. Особенно предпочтительным является полипептид, имеющий последовательность, выбранную из группы, состоящей из SEQ ID NO:115 - SEQ ID NO:171, или его функциональные эквиваленты.
Настоящее изобретение также относится к антителам, взаимодействующим с полипептидом настоящего изобретения. Эти антитела могут быть поликлональными, а предпочтительно моноклональными. Такие антитела являются особенно подходящими для очистки полипептидов настоящего изобретения.
В объем настоящего изобретения входят также гибридные белки, содержащие полипептид настоящего изобретения. Настоящее изобретение также относится к способам получения полипептидов настоящего изобретения.
Кроме того, настоящее изобретение относится к способу диагностики аспергиллеза путем детекции присутствия полипептида настоящего изобретения или его функциональных эквивалентов либо путем детекции кДНК настоящего изобретения, или ее фрагментов, или функциональных эквивалентов.
Настоящее изобретение также относится к использованию протеазы настоящего изобретения в описанном здесь промышленном способе.
Подробное описание изобретения
Полинуклеотиды
Настоящее изобретение относится к полинуклеотидам, кодирующим протеазы, имеющие аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:115 - SEQ ID NO:171, или их функциональные эквиваленты. Последовательность этих генов определяют путем секвенирования геномного клона, полученного из Aspergillus niger. Настоящее изобретение относится к полинуклеотидным последовательностям, содержащим ген, кодирующий эти протеазы, а также к их полной кДНК-последовательности и ее кодирующей последовательности. В соответствии с этим настоящее изобретение относится к выделенному полинуклеотиду, включающему нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO:1 - SEQ ID NO:57, или последовательность, выбранную из группы, состоящей из SEQ ID NO:58 - SEQ ID NO:114 или их функциональных эквивалентов.
Более конкретно, настоящее изобретение относится к выделенному полинуклеотиду, гибридизуемому в жестких условиях с полинуклеотидом, выбранным из группы, состоящей из SEQ ID NO:1 - SEQ ID NO:57, или с последовательностью, выбранной из группы, состоящей из SEQ ID NO:58 - SEQ ID NO:114, предпочтительно в условиях высокой жесткости. Предпочтительно такие полинуклеотиды могут быть получены из нитчатых грибов, а в частности из Aspergillus niger. Более конкретно, настоящее изобретение относится к выделенному полинуклеотиду, имеющему нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO:1 - SEQ ID NO:57, или последовательность, выбранную из группы, состоящей из SEQ ID NO:58 - SEQ ID NO:114.
Настоящее изобретение также относится к выделенному полинуклеотиду, кодирующему, по крайней мере, один функциональный домен полипептида, имеющий последовательность, выбранную из группы, состоящей из SEQ ID NO:115 - SEQ ID NO:171 или их функциональных эквивалентов.
Используемые здесь термины “ген” и “рекомбинантный ген” означают молекулы нуклеиновой кислоты, которые могут быть выделены из хромосомной ДНК и которые включают открытую рамку считывания, кодирующую белок, например, протеазу A.niger. Ген может включать кодирующие последовательности, некодирующие последовательности, интроны или регуляторные последовательности. Кроме того, термин “ген” может означать выделенную молекулу нуклеиновой кислоты, определенную в настоящем описании изобретения.
Молекула нуклеиновой кислоты настоящего изобретения, такая как молекула нуклеиновой кислоты, имеющая нуклеотидную последовательность, выбранную из группы, состоящей из SEQ ID NO:1 - SEQ ID NO:57, или последовательность, выбранную из группы, состоящей из SEQ ID NO:58 - SEQ ID NO:114 или ее функциональных эквивалентов, может быть выделена стандартными методами, применяемыми в молекулярной биологии, и с использованием представленной здесь информации, относящейся к этой последовательности. Так, например, с использованием всей или части последовательности нуклеиновой кислоты, выбранной из группы, состоящей из SEQ ID NO:1 - SEQ ID NO:57 или нуклеотидной последовательности, выбранной из группы, состоящей из SEQ ID NO:58 - SEQ ID NO:114, в качестве гибридизационного зонда молекулы нуклеиновой кислоты настоящего изобретения могут быть выделены с использованием стандартной техники гибридизации и клонирования (например, как описано в Sambrook, J., Fritsh, E.F., & Maniatis T., Molecular Cloning: A Laboratory Manual, 2nd, ed., Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989).
Кроме того, молекула нуклеиновой кислоты, включающая всю или часть последовательности, выбранной из группы, состоящей из SEQ ID NO:1 - SEQ ID NO:57, или последовательности, выбранной из группы, состоящей из SEQ ID NO:58 - SEQ ID NO:114, может быть выделена с помощью полимеразной цепной реакции (ПЦР) с использованием синтетических олигонуклеотидных праймеров, сконструированных на основе информации о последовательности, содержащейся в последовательности, выбранной из группы, состоящей из SEQ ID NO:1 - SEQ ID NO:57, или последовательности, выбранной из группы, состоящей из SEQ ID NO:58 - SEQ ID NO:114.
Нуклеиновая кислота настоящего изобретения может быть амплифицирована с использованием кДНК, мРНК или, альтернативно, геномной ДНК в качестве матрицы и с использованием соответствующих олигонуклеотидных праймеров в соответствии со стандартными методами ПЦР-амплификации. Амплифицированная таким образом нуклеиновая кислота может быть клонирована в соответствующий вектор и охарактеризована с помощью анализа ДНК-последовательности.
Кроме того, олигонуклеотиды, соответствующие нуклеотидным последовательностям настоящего изобретения или гибридизуемые с ними, могут быть получены стандартными методами синтеза, например, с использованием автоматического ДНК-синтезатора.
В предпочтительном варианте осуществления изобретения выделенная молекула нуклеиновой кислоты настоящего изобретения включает нуклеотидную последовательность, представленную в последовательности, выбранной из группы, состоящей из SEQ ID NO:58 - SEQ ID NO:114. Последовательность, выбранная из группы, состоящей из SEQ ID NO:58 - SEQ ID NO:114, соответствует кодирующей области кДНК протеазы A.niger. Эта кДНК включает последовательности, кодирующие полипептид протеазы A.niger, имеющей последовательность, выбранную из группы, состоящей из SEQ ID NO:115 - SEQ ID NO:171.
В другом предпочтительном варианте осуществления изобретения выделенная молекула нуклеиновой кислоты настоящего изобретения включает молекулу нуклеиновой кислоты, которая комплементарна нуклеотидной последовательности, представленной в последовательности, выбранной из группы, состоящей из SEQ ID NO:1 - SEQ ID NO:57, или в последовательности, выбранной из группы, состоящей из SEQ ID NO:58 - SEQ ID NO:114, или функциональному эквиваленту этих нуклеотидных последовательностей.
Молекула нуклеиновой кислоты, комплементарная другой нуклеотидной последовательности, представляет собой молекулу, которая по существу комплементарна другой нуклеотидной последовательности так, что может гибридизоваться с этой другой нуклеотидной последовательностью с образованием стабильного дуплекса.
В одном из аспектов настоящее изобретение относится к выделенным молекулам нуклеиновой кислоты, кодирующим полипептид настоящего изобретения или его функциональный эквивалент, такой как биологически активный фрагмент или домен, а также к молекулам нуклеиновой кислоты, подходящим для использования в качестве гибридизационных зондов в целях идентификации молекул нуклеиновой кислоты, кодирующих полипептид настоящего изобретения, и к фрагментам таких молекул нуклеиновой кислоты, подходящих для использования в качестве ПЦР-праймеров в целях амплификации или мутагенеза молекул нуклеиновой кислоты.
Термины “выделенный полинуклеотид” или “выделенная нуклеиновая кислота” означают ДНК или РНК, не являющуюся непосредственно смежной с обеими кодирующими последовательностями, с которыми она является непосредственно смежной (с одной на 5'-конце и с другой на 3'-конце) в природном геноме организма, от которого она происходит. Таким образом, в одном из вариантов осуществления изобретения выделенная нуклеиновая кислота включает несколько или все 5'-некодирующие (например, промоторные) последовательности, которые являются непосредственно смежными с кодирующей последовательностью. Поэтому термин включает, например, рекомбинантную ДНК, которая встроена в вектор, в автономно реплицирующуюся плазмиду или вирус, или в геномную ДНК прокариота или эукариота, или которая существует в виде отдельной молекулы (например, в виде кДНК или фрагмента геномной ДНК, продуцируемых с помощью ПЦР или путем обработки рестриктазами), независимой от других последовательностей. Этот термин также включает рекомбинантную ДНК, которая является частью гибридного гена, кодирующего дополнительный полипептид, в основном, не содержащей клеточного материала, вирусного материала или культуральной среды (при продуцирования методами рекомбинантных ДНК), либо химических предшественников или других химических соединений (при химическом синтезе). Кроме того, термин “выделенный фрагмент нуклеиновой кислоты” означает фрагмент нуклеиновой кислоты, который обычно не встречается в виде фрагмента и не обнаруживается в природе.
Используемые здесь термины “полинуклеотид” или “молекулы нуклеиновой кислоты” означают молекулы ДНК (например, кДНК или геномную ДНК), молекулы РНК (например, мРНК) и аналоги ДНК или РНК, генерированные с использованием нуклеотидных аналогов. Молекула нуклеиновой кислоты может быть одноцепочечной или двухцепочечной, но предпочтительно, она представляет собой двухцепочечную ДНК. Нуклеиновая кислота может быть синтезирована с использованием олигонуклеотидных аналогов или производных (например, инозиновых или фосфортиоатных нуклеотидов). Такие олигонуклеотиды могут быть использованы, например, для получения нуклеиновых кислот, которые обладают измененной способностью к образованию пар оснований или повышенной резистентностью к нуклеазам.
В другом варианте осуществления изобретения настоящее изобретение относится к выделенной молекуле нуклеиновой кислоты, которая является антисмысловой по отношению к молекуле нуклеиновой кислоты протеазы, например, к кодирующей цепи молекулы нуклеиновой кислоты протеазы. В объем настоящего изобретения также входят комплементарные цепи описанных здесь молекул нуклеиновой кислоты.
Ошибки секвенирования
Представленная здесь информация о последовательности не должна рассматриваться в узком смысле как требование включения ошибочно идентифицированных оснований. Описанные здесь конкретные последовательности могут быть с успехом использованы для выделения полного гена из нитчатых грибов, а в частности из A.niger, который, в свою очередь, может быть легко подвергнут последующему анализу последовательностей, идентифицируя тем самым ошибки секвенирования.
Если это не указано особо, то все нуклеотидные последовательности, определенные здесь путем секвенирования молекулы ДНК, были определены с использованием автоматического ДНК-секвенатора, и все аминокислотные последовательности полипептидов, кодируемые определенными здесь молекулами ДНК, были предсказаны посредством трансляции ДНК-последовательности, определенной выше. Поэтому, как известно специалистам, любая определенная здесь нуклеотидная последовательность для любой ДНК-последовательности, определенной таким автоматизированным способом, может содержать определенное число ошибок. Обычно нуклеотидные последовательности, определенные автоматизированным способом, по крайней мере, примерно на 90%, более предпочтительно, по крайней мере, примерно на 95%-99,9% идентичны реальной нуклеотидной последовательности секвенированной молекулы ДНК. Реальная последовательность может быть более точно определена другими способами, включая способы секвенирования ДНК вручную, хорошо известные специалистам. Как также известно специалистам, одна инсерция или делеция в определенной нуклеотидной последовательности по сравнению с фактической последовательностью, будет приводить к сдвигу рамки считывания при трансляции нуклеотидной последовательности, в результате чего предсказанная аминокислотная последовательность, кодируемая определенной нуклеотидной последовательностью, будет полностью отличаться от фактической аминокислотной последовательности, кодируемой секвенированной молекулой ДНК, начиная с точки такой инсерции или делеции.
Каждый специалист может выявить такие неправильно идентифицированные основания и исправить такие ошибки.
Фрагменты нуклеиновых кислот, зонды и праймеры
Молекулы нуклеиновой кислоты настоящего изобретения могут включать только часть или фрагмент последовательности нуклеиновой кислоты, представленной в последовательности, выбранной из группы, состоящей из SEQ ID NO:1-57, или последовательности, выбранной из группы, состоящей из SEQ ID NO:58 - SEQ ID NO:114, например, фрагмент, который может быть использован в качестве зонда или праймера, или фрагмент, кодирующий часть белка протеазы. Нуклеотидная последовательность, определенная в результате клонирования гена протеазы, и кДНК позволяет генерировать зонды и праймеры, сконструированные для идентификации и/или клонирования других членов семейства протеаз, а также гомологов протеаз других видов. Зонд/праймер обычно включает, по существу, очищенный олигонуклеотид, обычно содержащий область нуклеотидной последовательности, которая гибридизуется, предпочтительно, в условиях высокой жесткости, по крайней мере, примерно с 12 или 15, предпочтительно примерно 18 или 20, более предпочтительно примерно с 22 или 25, а наиболее предпочтительно примерно с 30, 35, 40, 45, 50, 55, 60, 65 или 75 нуклеотидами, или с более близко расположенными нуклеотидами нуклеотидной последовательности, представленной в последовательности, выбранной из группы, состоящей из SEQ ID NO:1 - SEQ ID NO:57, или последовательности, выбранной из группы, состоящей из SEQ ID NO:58-114, или их функциональных эквивалентов.
Зонды, основанные на нуклеотидных последовательностях протеазы, могут быть использованы для детекции транскриптов или геномных последовательностей протеаз, кодирующих те же самые или гомологичные белки, происходящие, например, от других организмов. В предпочтительных вариантах осуществления изобретения указанный зонд дополнительно включает присоединенную к нему группу-метку, например, такой группой-меткой может быть радиоизотоп, флуоресцентное соединение, фермент или кофактор фермента. Такие зонды могут быть также использованы как часть диагностического тест-набора для идентификации клеток, экспрессирующих белок протеазы.
Идентификация и гомология
Используемые здесь термины “гомология” или “процент идентичности” являются взаимозаменяемыми. В целях настоящего изобретения для определения процента идентичности двух аминокислотных последовательностей или двух последовательностей нуклеиновой кислоты было проведено оптимальное сравнение последовательностей (например, в последовательности первой аминокислотной последовательности или последовательности нуклеиновой кислоты могут быть введены “бреши” в целях ее оптимального сопоставления со второй аминокислотной последовательностью или последовательностью нуклеиновой кислоты). Затем было проведено сравнение аминокислотных остатков или нуклеотидов в соответствующих положениях аминокислот или в положениях нуклеотидов. Если положение в первой последовательности занято тем же самым аминокислотным остатком или нуклеотидом, как и в соответствующем положении второй последовательности, то эти молекулы являются идентичными в данном положении. Процент идентичности между двумя последовательностями представляет собой функцию числа идентичных положений для двух сравниваемых последовательностей (то есть % идентичности = число идентичных положений/общее число положений (то есть перекрывающихся положений)×100).
Предпочтительно, чтобы эти две последовательности имели одинаковую длину.
Каждому специалисту известно, что для определения гомологии между двумя последовательностями существуют несколько различных компьютерных программ. Так, например, сравнение последовательностей и определение процента идентичности между двумя последовательностями может быть осуществлено с помощью математического алгоритма. В предпочтительном варианте осуществления изобретения процент идентичности между двумя аминокислотными последовательностями определяют с использованием алгоритма Needleman & Wunsch (J. Mol. Biol. (48):444-453 (1970)), который был введен в программу GAP программного обеспечения GCG (доступного в http://www.gcg.com) с использованием либо матрицы Blossom 62, либо матрицы РАМ250 и с использованием веса “бреши” 16, 14, 12, 10, 8, 6 или 4 и веса длины 1, 2, 3, 4, 5 или 6. Для специалиста очевидно, что все эти различные параметры дают слегка отличающиеся результаты, но общий процент идентичности двух последовательностей значительно не меняется при использовании различных алгоритмов.
В другом варианте осуществления изобретения процент идентичности между двумя нуклеотидными последовательностями определяют с использованием программы GAP, программного обеспечения GCG (доступного в http://www.gcg.com) с использованием матрицы NWSgapdna.СМР и с использованием веса “бреши” 40, 50, 60, 70 или 80 и веса длины 1, 2, 3, 4, 5 или 6. В другом варианте осуществления изобретения процент идентичности двух аминокислотных или нуклеотидных последовательностей определяют с использованием алгоритма E. Meyer & W. Miller (CABIOS, 4:11-17 (1989), который был включен в программу ALIGN (версия 2.0) (доступной в http://vega/igh.cnrs.fr/bin/align-guess.cgi), с использованием таблицы веса остатков РАМ120, штрафа на “брешь”-удлинение 12 и штрафа на “брешь-пропуск” 4.
Последовательности нуклеиновой кислоты и белка настоящего изобретения могут быть также использованы в качестве “запрашиваемой последовательности” для осуществления поиска в доступных базах данных, например, в целях идентификации других членов семейств или родственных последовательностей. Такой поиск может быть осуществлен с использованием программ NBLAST и XBLAST (версия 2.0), Altschul et al. (1990) J. Mol. Biol. 215:403-10. Поиск нуклеотидов в BLAST может быть осуществлен с использованием программы NBLAST, метка для отсчета = 100, длина слова = 12 для получения нуклеотидных последовательностей, гомологичных молекулам нуклеиновой кислоты протеазы настоящего изобретения. Поиск белков в BLAST может быть осуществлен с использованием программы ХBLAST, метка для отсчета = 50, длина слова = 3 для получения аминокислотных последовательностей, гомологичных молекулам белка протеазы настоящего изобретения. Для осуществления сопоставления с введением “брешей” может быть использована программа BLAST с введенными “брешами”, как описано Altschul et al., (1997) Nucleic Acids Res. 25(17):3389-3402. При применении программ BLAST и BLAST с введенными брешами могут быть использованы параметры соответствующих программ по умолчанию (например, XBLAST и NBLAST). См. http://www.ncbi.nim.nih.gov.
Гибридизация
Используемый здесь термин “гибридизация” означает условия гибридизации и промывки, при которых нуклеотидные последовательности, являющиеся, по крайней мере, примерно на 50%, по крайней мере, примерно на 60%, по крайней мере, примерно на 70%, более предпочтительно, по крайней мере, на 80%, еще более предпочтительно, по крайней мере, примерно на 85-90%, а наиболее предпочтительно, по крайней мере, на 95% гомологичными друг другу, гибридизуются друг с другом.
Предпочтительным неограничивающим примером таких условий гибридизации являются гибридизация в 6× хлориде натрия/цитрате натрия (SSC) приблизительно при 45°С с последующей одной или более промывками в 1×SSC, 0,1% ДСН при 50°С, предпочтительно при 55°С, более предпочтительно при 60°С, а еще более предпочтительно 65°С.
Условиями высокой жесткости являются, например, гибридизация при 68°С в 5×SSC/5× раствор Денхардта/1,0% ДСН и промывка при 0,2×SSC/0,1% ДСН при комнатной температуре. Альтернативно, промывка может быть осуществлена при 42°С.
Каждому специалисту известно, в каких случаях необходимо применять жесткие условия и условия высокой жесткости гибридизации. Дополнительную информацию о таких условиях можно найти в литературе, например, в руководстве Sambrook et al., 1989, Molecular Cloning, A Laboratory Manual, Cold Spring Harbor Press, N.Y.; и Ausubel et al. (eds.), 1995, Current Protocols in Molecular Biology (John Wiley & Sons, N.Y.).
Само собой разумеется, что полинуклеотид, который гибридизуется только с poly(A)-последовательностью (такой как 3'-концевой poly(A)-хвост мРНК) или с комплементарным фрагментом из остатков Т (или U), не входит в полинуклеотид настоящего изобретения, используемый для специфической гибридизации с частью нуклеиновой кислоты настоящего изобретения, поскольку такой полинуклеотид должен гибридизоваться с любой молекулой нуклеиновой кислоты, содержащей poly(A)-хвост, или с комплементарной последовательностью (например, с любым двухцепочечным ДНК-клоном).
Получение полноразмерной ДНК из других микроорганизмов
С применением обычной методики могут быть скринированы кДНК-библиотеки, сконструированные из других микроорганизмов, например, из нитчатых грибов, а в частности, из грибов вида Aspergillus.
Так, например, штаммы Aspergillus могут быть скринированы на гомологичные полинуклеотиды протеазы с помощью Нозерн-блот-анализа. После детекции транскриптов, гомологичных полинуклеотидам настоящего изобретения, кДНК-библиотеки могут быть сконструированы из РНК, выделенной из соответствующего штамма, стандартными методами, хорошо известными специалистам. Альтернативно, вся геномная ДНК-библиотека может быть скринирована с использованием зонда, гибридизуемого с полинуклеотидом протеазы настоящего изобретения.
Гомологичные генные последовательности могут быть выделены, например, посредством ПЦР с использованием двух олигонуклеотидных праймеров или двух пулов вырожденных олигонуклеотидных праймеров, сконструированных на основе описанных здесь нуклеотидных последовательностей.
Матрицей для реакции может быть кДНК, полученная посредством обратной транскрипции мРНК, выделенной из штаммов, которые, как известно или как предполагается, экспрессируют полинуклеотид настоящего изобретения. ПЦР-продукт может быть субклонирован и секвенирован для гарантии того, что эти амплифицированные последовательности являются последовательностями нуклеиновой кислоты новой протеазы или ее функционального эквивалента.
ПЦР-фрагмент может быть затем использован для выделения полноразмерного кДНК-клона различными известными методами. Так, например, амплифицированный фрагмент может быть выделен и использован для скрининга бактериофага или космидной кДНК-библиотеки. Альтернативно, меченый фрагмент может быть использован для скрининга геномной библиотеки.
Для выделения полноразмерных последовательностей кДНК из других микроорганизмов может быть использована ПЦР-технология. Так, например, РНК может быть выделена в соответствии со стандартными процедурами из подходящего клеточного или тканевого источника. Для запуска синтеза первой цепи реакция обратной транскрипции может быть осуществлена на РНК с использованием олигонуклеотидного праймера, специфичного, главным образом, к 5'-концу амплифицированного фрагмента.
Полученный гибрид РНК/ДНК может быть затем “удлинен” (например, гуанинами) посредством стандартной терминальной трансферазной реакции, и этот гибрид может быть расщеплен РНКазой Н, после чего может быть запущен синтез второй цепи (например, с использованием поли-С-праймера). Таким образом, кДНК-последовательности, расположенные выше амплифицированного фрагмента, могут быть легко выделены. Обзор подходящих стратегий клонирования можно найти, например, у Sambrook et al., см., выше; и Ausubel et al., см. выше.
Векторы
В другом своем аспекте настоящее изобретение относится к векторам, а предпочтительно, к экспрессирующим векторам, содержащим нуклеиновую кислоту, кодирующую белок протеазы или его функциональный эквивалент. Используемый здесь термин “вектор” означает молекулу нуклеиновой кислоты, способную переносить другую нуклеиновую кислоту, к которой он был присоединен. Одним из типов векторов является “плазмида”, представляющая собой кольцевую двухцепочечную ДНК-петлю, в которую могут быть лигированы дополнительные ДНК-сегменты. Другим типом вектора является вирусный вектор, где добавочные ДНК-сегменты могут быть лигированы в вирусный геном. Некоторые векторы способны к автономной репликации в клетке-хозяине, в которую они введены (например, бактериальные векторы, имеющие бактериальный источник репликации, и эписомные векторы млекопитающих). Другие векторы (например, неэписомные векторы млекопитающих) интегрируются в геном клетки-хозяина после введения в эту клетку, и таким образом, реплицируются вместе с геномом хозяина. Кроме того, некоторые векторы способны направлять экспрессию генов, с которыми они функционально связаны. Такие векторы называются здесь “экспрессирующими векторами”. В основном, экспрессирующие векторы часто используются в технике рекомбинантных ДНК в форме плазмид. Термины “плазмида” и “вектор” могут быть взаимозаменяемыми, поскольку плазмида является наиболее распространенной формой вектора. Однако настоящее изобретение относится и к другим формам экспрессирующих векторов, таких как вирусные векторы (например, дефектные по репликации ретровирусы, аденовирусы и аденоассоциированные вирусы), которые обладают эквивалентными функциями.
Рекомбинантные экспрессирующие векторы настоящего изобретения включают нуклеиновую кислоту настоящего изобретения в форме, подходящей для экспрессии этой нуклеиновой кислоты в клетке-хозяине, что означает, что этот рекомбинантный экспрессирующий вектор включает одну или более регуляторных последовательностей, выбранных в соответствии с используемыми для экспрессии клетками-хозяевами и функционально связанных с экспрессируемой последовательностью нуклеиновой кислоты. Используемый по отношению к рекомбинантному экспрессирующему вектору термин “функционально присоединенный” означает, что присутствующая в нем и представляющая интерес нуклеотидная последовательность присоединена к регуляторной последовательности(ям) способом, обеспечивающим экспрессию данной нуклеотидной последовательности (например, в in vitro системе транскрипции/трансляции или в клетке-хозяине, если указанный вектор введен в эту клетку-хозяина). Термин “регуляторная последовательность” означает промоторы, энхансеры и другие регулирующие экспрессию элементы (например, сигнал полиаденилирования). Такие регуляторные элементы описаны, например, Goeddel, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1990). Регуляторными последовательностями являются последовательности, которые направляют конститутивную экспрессию нуклеотидной последовательности в клетки-хозяева многих типов, и последовательности, которые направляют экспрессию нуклеотидной последовательности только в некоторые клетки-хозяева (например, тканеспецифические регуляторные последовательности). При этом следует отметить, что конструирование экспрессирующего вектора может зависеть от таких факторов, как выбор трансформируемой клетки-хозяина, уровень экспрессии нужного белка и т.п. Экспрессирующие векторы настоящего изобретения могут быть введены в клетки-хозяева, в результате чего они будут продуцировать белки или пептиды, кодируемые описанными здесь нуклеиновыми кислотами (например, белки протеаз, мутантные формы белков протеаз, их фрагменты, варианты или функциональные эквиваленты, гибридные белки и т.п.).
Рекомбинантные экспрессирующие векторы настоящего изобретения могут быть сконструированы для экспрессии белков протеаз в прокариотических или в эукариотических клетках. Так, например, белки протеаз могут быть экспрессированы в бактериальных клетках, таких как E.coli, в клетках насекомых (с использованием бакуловирусных экспрессирующих векторов), в дрожжевых клетках или в клетках млекопитающих. Подходящие клетки-хозяева подробно обсуждаются в работе Goeddel, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, CA (1990). Альтернативно, рекомбинантный экспрессирующий вектор может быть транскрибирован и транслирован in vitro с использованием промоторных регуляторных последовательностей Т7 и полимеразы Т7.
Экспрессирующими векторами, используемыми в настоящем изобретении, являются хромосомные, эписомные и вирусные векторы, например, векторы, происходящие от бактериальных плазмид, бактериофага, дрожжевой эписомы, дрожжевых хромосомных элементов; вирусы, такие как бакуловирусы, паповавирусы, вакцинные вирусы, аденовирусы, вирусы оспы домашней птицы, псевдорабивирусы и ретровирусы, а также векторы, полученные из их комбинаций, такие как векторы, происходящие из плазмиды и бактериофаговых генетических элементов, таких как космиды и фагмиды.
ДНК-вставка должна быть функционально присоединена к подходящему промотору, такому как промоторы РL фага лямбда, промоторы lac, trp и tac E.coli, ранние и поздние промоторы SV40 и промоторы ретровирусных LТR и т.п. Специалистам хорошо известны и другие подходящие промоторы. В конкретном варианте осуществления изобретения предпочтительными являются промоторы, способные обеспечивать высокий уровень экспрессии протеаз в нитчатых грибах. Такие промоторы известны специалистам. Экспрессирующие конструкции могут содержать сайты инициации транскрипции, терминации транскрипции, а в транскрибированной области сайт связывания с рибосомой для трансляции. Кодирующая часть зрелых транскриптов, экспрессируемых этими конструкциями, включает сайт инициации трансляции АUG в начале последовательности и кодон терминации трансляции, расположенный в соответствующем положении у конца транслируемого полипептида.
Векторная ДНК может быть введена в прокариотические или эукариотические клетки стандартными методами трансформации или трансфекции. Используемые здесь термины “трансформация” и “трансфекция” означают ряд известных специалистам методов введения чужеродной нуклеиновой кислоты (например, ДНК) в клетку-хозяина, включая ко-преципитацию фосфатом кальция или хлоридом кальция, опосредованную DЕАЕ-декстраном трансфекцию, трансдукцию, инфицирование, липофекцию, опосредованную катионным липидом трансфекцию или электропорацию. Подходящие методы трансформации или трансфекции клеток-хозяев можно найти в руководстве у Sambrook et al. (Molecular Cloning: A Laboratory Manual, 2nd ed., Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989), Davis et al., Basic Methods in Molecular Biology (1986) и в других лабораторных руководствах.
Что касается стабильной трансфекции клеток млекопитающих, то известно, что лишь небольшая фракция клеток может интегрировать чужеродную ДНК в свой геном в зависимости от используемого экспрессирующего вектора и метода трансфекции. Для идентификации и отбора этих интегрантов ген, кодирующий селективный маркер (например, маркер резистентности к антибиотикам), обычно вводят в клетки-хозяева вместе с нужным геном. Предпочтительными селективными маркерами являются маркеры, которые придают резистентность к лекарственным средствам, таким как G418, гигромицин и метотрексат. Нуклеиновая кислота, кодирующая селективный маркер, может быть введена в клетку-хозяин на том же самом векторе, который кодирует белок протеазы, либо она может быть введена на отдельном векторе. Клетки, стабильно трансфицированные введенной нуклеиновой кислотой, могут быть идентифицированы путем отбора по резистентности к лекарственному средству (например, клетки, в которые был введен селективный маркерный ген, выживают, а другие клетки погибают).
Экспрессию белков в прокариотах, в большинстве случаев, осуществляют в клетках E.coli, включающих векторы, содержащие конститутивные или индуцибельные промоторы, регулирующие экспрессию гибридных или негибридных белков. Гибридные векторы присоединяют ряд аминокислот к кодируемому в нем белку, например, к аминоконцу рекомбинантного белка. Такие гибридные векторы обычно служат трем целям, а именно: 1) усиливают экспрессию рекомбинантного белка; 2) увеличивают растворимость рекомбинантного белка и 3) облегчают очистку рекомбинантного белка, действуя как лиганд в аффинной очистке. В большинстве случаев в гибридных экспрессирующих векторах сайт протеолитического расщепления вводят на стыке гибридной молекулы и рекомбинантного белка, что позволяет отделять рекомбинантный белок от гибридной молекулы, а затем проводят очистку этого гибридного белка. Такими ферментами и их последовательностями когнатного распознавания являются фактор Ха, тромбин и энтерокиназа.
Как было указано выше, экспрессирующие векторы, предпочтительно, содержат селективные маркеры. Такими маркерами являются ген дигидрофолатредуктазы или ген резистентности к неомицину для эукариотических клеточных культур и ген резистентности к тетрациклину или ампициллину для культивирования в E.coli и в других бактериях. Репрезентативными примерами подходящего хозяина являются бактериальные клетки, такие как E.coli, Streptomyces и Salmonella typhimurium; клетки грибов, такие как дрожжи; клетки насекомых, такие как Drosophila S2 и Spodoptera Sf9; клетки животных, такие как СНО, СОS и меланома Боуэса; и клетки растений. Подходящие культуральные среды и условия культивирования вышеуказанных клеток-хозяев известны специалистам.
Векторами, предпочтительными для использования в бактериях, являются векторы pQE70, pQE60 и PQE-9, поставляемые от Qiagen; векторы pBS, векторы Phagescript, векторы Bluescript, pNH8A, pNH16A, pNH18A, pNH46A, поставляемые от Stratagene; и ptrc99a, pKK223-3, pKK233-3, pDR540, pRIT5, поставляемые от Pharmacia. Предпочтительными эукариотическими векторами являются векторы PWLNEO, pSV2CAT, pOG44, pZT1 и pSG, поставляемые от Stratagene; и векторы pSVK3, pBРV, pMSG и pSVL, поставляемые от Pharmacia. Другие подходящие векторы хорошо известны специалистам.
Известными бактериальными промоторами, используемыми в настоящем изобретении, являются промоторы lacl и lacZ E.coli промоторы Т3 и Т7, промотор gpt, промоторы лямбда PR, промотор PL и промотор trp, промотор тимидинкиназы HSV, ранний и поздний промоторы SV40, промоторы LTR ретровирусов, таких как вирус саркомы Рауса (“RSV”), и промоторы металлотионеина, такие как промотор мышиного металлотионеина-1.
Транскрипция ДНК, кодирующей полипептиды настоящего изобретения, у высших эукариотов может быть усилена посредством введения в вектор энхансерной последовательности. Энхансеры являются цис-действующими элементами ДНК, которые обычно имеют длину приблизительно 10-300 п.н. и которые усиливают транскрипционную активность промотора в клетке-хозяине данного типа. Примерами энхансеров являются энхансер SV40, расположенный на позднем участке сайта инициации репликации и имеющий длину 100-270 п.н., энхансер раннего промотора цитомегаловируса, энхансер полиомы, расположенный на позднем участке сайта инициации репликации, и аденовирусные энхансеры.
Для секреции транслированного белка в область просвета эндоплазматического ретикулума в периплазматическое пространство или во внеклеточное пространство, в экспрессируемый полипептид может быть введен соответствующий сигнал секреции. Эти сигналы могут быть эндогенными для полипептида, либо они могут быть гетерологичными.
Полипептид может экспрессироваться в модифицированной форме, такой как гибридный белок, и может включать не только сигналы секреции, но также и дополнительные гетерологичные функциональные области. Так, например, область, состоящая из других аминокислот, а в частности, заряженных аминокислот, может быть добавлена к N-концу данного полипептида для повышения его стабильности и устойчивости в клетке-хозяине в процессе очистки или в процессе последующей обработки и хранения. Кроме того, пептидные молекулы могут быть добавлены в полипептид для облегчения очистки.
Полипептиды настоящего изобретения
Настоящее изобретение относится к выделенному полипептиду, имеющему аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:115 - SEQ ID NO:171; аминокислотную последовательность, получаемую путем экспрессии полинуклеотида настоящего изобретения, либо, в предпочтительном варианте изобретения, последовательность, выбранную из группы, состоящей из SEQ ID NO:1 - SEQ ID NO:57 в соответствующем хозяине, а также аминокислотную последовательность, получаемую путем экспрессии полинуклеотидных последовательностей, выбранных из группы, состоящей из SEQ ID NO:58 - SEQ ID NO:114 в соответствующем хозяине. Кроме того, в объем настоящего изобретения входит пептид или полипептид, содержащий функциональный эквивалент вышеуказанных полипептидов. Вышеуказанные полипептиды, в целом, называются “полипептидами настоящего изобретения”.
Термины “пептид” и “олигопептид” рассматриваются как синонимы (в общепринятом смысле), и эти термины являются взаимозаменяемыми в зависимости от контекста, где требуется указать, что цепь состоит, по крайней мере, из двух аминокислот, связанных пептидильными связями. Используемый здесь термин “полипептид” означает цепи, содержащие более чем семь аминокислотных остатков. Все олигопептидные и полипептидные формулы или их последовательности записываются слева направо, в направлении от аминоконца к карбоксиконцу. Однобуквенный код используемых аминокислот хорошо известен специалистам, и его можно найти в руководстве Sambrook et al. (Molecular Cloning: A Laboratory Manual, 2nd ed., Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989).
Термин “выделенный” полипептид или белок означает полипептид или белок, экстрагированный из его природного окружения. Так, например, рекомбинантно продуцируемые полипептиды и белки, экспрессируемые в клетках-хозяевах, считаются выделенными в целях настоящего изобретения, так, как это было определено для нативных или рекомбинантных полипептидов, которые были по существу очищены любыми подходящими методами, такими как, например, метод одностадийной очистки, описанный Smith & Johnson, Gene 67:31-40 (1988).
Протеазы настоящего изобретения могут быть выделены и очищены из рекомбинантных культур хорошо известными методами, включая преципитацию сульфатом аммония или этанолом, экстракцию кислотой, анионо- или катионообменную хроматографию, хроматографию на фосфоцеллюлозе, гидрофобную хроматографию взаимодействия, аффинную хроматографию, хроматографию на гидроксиаппатитах и хроматографию на лектине. Для очистки в аналитических целях наиболее предпочтительно использовать высокоэффективную жидкостную хроматографию (“ВЭЖХ”).
Полипептидами настоящего изобретения являются природные очищенные продукты, продукты химического синтеза и продукты, продуцируемые рекомбинантными методами из прокариотических или эукариотическмх хозяев, включая, например, бактериальные клетки, дрожжевые клетки, клетки высших растений, клетки насекомых и клетки млекопитающих. В зависимости от используемого хозяина в методах рекомбинантного продуцирования полипептиды настоящего изобретения могут быть гликозилированными или негликозилированными. Кроме того, полипептиды настоящего изобретения могут также включать модифицированный инициирующий метиониновый остаток, который, в некоторых случаях, образуется в результате клеточно-опосредованных процессов в хозяине.
Кроме того, белком настоящего изобретения может быть белок-предшественник, такой как зимоген; гибридный белок; и белок, полученный в виде про-последовательности или пре-про-последовательности либо в виде другой незрелой формы любого типа.
Фрагменты белков
Настоящее изобретение также относится к биологически активным фрагментам полипептидов настоящего изобретения.
Биологически активными фрагментами полипептидов настоящего изобретения являются полипептиды, включающие аминокислотные последовательности, которые по существу идентичны аминокислотной последовательности белка протеазы или происходят от этой последовательности (например, аминокислотной последовательности, содержащейся в последовательности, выбранной из группы, состоящей из SEQ ID NO:115 - SEQ ID NO:171), и которые включают меньшее число аминокислот, чем полноразмерный белок, и обладают, по крайней мере, одной биологической активностью, соответствующей активности полноразмерного белка. Обычно биологически активные фрагменты включают домен или мотив, обладающий, по крайней мере, одной активностью белка протеазы. Биологически активный фрагмент белка настоящего изобретения может представлять собой полипептид, который имеет длину, например, в 10, 25, 50, 100 или более аминокислот. Кроме того, другие биологически активные части, в которых делетированы другие области белка, могут быть получены рекомбинантными методами и оценены на одну или более биологических активностей, присущих нативной форме полипептида настоящего изобретения.
Настоящее изобретение также относится к фрагментам нуклеиновой кислоты, которые кодируют вышеописанные биологически активные фрагменты белка протеазы.
Гибридные белки
Белки настоящего изобретения или их функциональные эквиваленты, например, их биологически активные части, могут быть функционально присоединены к непротеазному полипептиду (например, к гетерологичным аминокислотным последовательностям) с получением гибридных белков. Используемый здесь термин “химерный белок” или “гибридный белок” протеазы означает полипептид протеазы, функционально присоединенный к непротеазному полипептиду. Термин “протеазный полипептид” означает полипептид, имеющий аминокислотную последовательность, соответствующую полипептидной последовательности настоящего изобретения, а термин “непротеазный полипептид” означает полипептид, имеющий аминокислотную последовательность, соответствующую белку, который по существу не является гомологичным белку настоящего изобретения, например, белку, который отличается от протеазного белка и происходит от одного и того же или другого организма. В протеазном гибридном белке протеазный полипептид может соответствовать всему белку или части белка настоящего изобретения. В предпочтительном варианте осуществления изобретения гибридный белок протеазы включает, по крайней мере, один биологически активный фрагмент белка настоящего изобретения. В другом предпочтительном варианте осуществления изобретения гибридный белок протеазы включает, по крайней мере, две биологически активных части белка настоящего изобретения. Используемый по отношению к гибридному белку термин “функционально присоединенный” указывает на то, что протеазный полипептид и непротеазный полипептид слиты друг с другом с сохранением рамки считывания. Непротеазный полипептид может быть слит с N-концом или с С-концом протеазного полипептида.
Так, например, в одном из вариантов осуществления изобретения гибридный белок представляет собой гибридный белок “GST-протеаза”, где указанные протеазные последовательности слиты с С-концом последовательностей GST.
Такие гибридные белки могут облегчать очистку рекомбинантной протеазы. В другом варианте осуществления изобретения гибридным белком является протеазный белок, содержащий гетерологичную сигнальную последовательность у его N-конца. В некоторых клетках-хозяевах (например, в клетках млекопитающих и в дрожжевых клетках-хозяевах) экспрессия и/или секреция протеазы могут быть усилены с использованием гетерологичной сигнальной последовательности.
В другом примере секреторная последовательность gp67 белка оболочки бакуловируса может быть использована как гетерологичная сигнальная последовательность (Current Protocols in Molecular Biology, Ausubel et al., eds., John Wiley & Sons, 1992). Другими примерами эукариотических гетерологичных сигнальных последовательностей являются секреторные последовательности мелитина и щелочной фосфатазы плаценты человека (Stratagene; La Jolla, California). Еще в одном примере подходящими прокариотическими гетерологичными сигнальными последовательностями являются секреторный сигнал рhoA (Sambrook et al., см. выше) и секреторный сигнал белка А (Pharmacia Biotech; Piscataway, New Jersey).
Сигнальная последовательность может быть использована для облегчения секреции и выделения белка или полипептида настоящего изобретения. Сигнальные последовательности обычно характеризуются коровыми гидрофобными аминокислотами, которые обычно отщепляются от зрелого белка в процессе секреции в одном или более событиях отщепления. Такие сигнальные пептиды содержат сайты процессинга, которые позволяют отщеплять сигнальную последовательность от зрелых белков, поскольку они проходят по секреторному пути. Сигнальная последовательность направляет секрецию белка, например, из эукариотического хозяина, в который был введен экспрессирующий вектор, а после этого или одновременно с этим сигнальная последовательность отщепляется. Затем белок может быть легко очищен от внеклеточной среды известными методами. Альтернативно, сигнальная последовательность может быть присоединена к нужному белку с использованием последовательности, которая облегчает очистку, например, с использованием домена GST. Так, например, последовательность, кодирующая полипептид, может быть слита с маркерной последовательностью, например, с последовательностью, кодирующей пептид, который облегчает очистку гибридного полипептида. В некоторых предпочтительных вариантах этого аспекта настоящего изобретения маркерная последовательность представляет собой гексагистидиновый пептид, такой как метка, присутствующая в векторе рQE (Qiagen, Inc.), и другие метки, многие из которых являются коммерчески доступными. Как, например, описано в работе Gentz et al., Proc.Natl. Acad. Sci., USA, 86:821-824 (1989), гексагистидин обеспечивает удобную очистку гибридного белка. Метка НА представляет собой другой пептид, используемый для очистки, который соответствует эпитопу, происходящему от белка гемаглютинина вируса гриппа, и который был описан, например, Wilson et al., Cell 37:767 (1984).
Предпочтительно, химерный или гибридный белок протеазы настоящего изобретения получают с использованием стандартной техники рекомбинантных ДНК. Так, например, ДНК-фрагменты, кодирующие различные полипептидные последовательности, лигируют вместе в той же рамке считывания стандартными методами, например, с использованием затупленных или липких концов для лигирования, затем расщепляют рестриктазами с получением соответствующих концов и, если это необходимо, достраивают липкие концы, обрабатывают щелочной фосфатазой во избежание нежелательного присоединения и подвергают ферментативному лигированию. В другом варианте осуществления изобретения гибридный ген может быть синтезирован стандартными методами, включая методы с использованием автоматических ДНК-синтезаторов. Альтернативно, ПЦР-амплификация генных фрагментов может быть осуществлена с использованием “якорных” праймеров, сконструированных так, чтобы они были комплементарны выступающим концам, между двумя смежными генными фрагментами, которые могут быть затем гибридизированы и повторно амплифицированы с получением химерной генной последовательности (см., например, Current Protocols in Molecular Biology, eds. Ausubel et al., John Wiley & Sons, 1992). Кроме того, многие экспрессирующие векторы являются коммерчески доступными и кодируют гибридную часть (например, полипептид GST). Кодирующая протеазу нуклеиновая кислота может быть клонирована в указанный экспрессирующий вектор так, чтобы эта гибридная часть была присоединена к протеазному белку с сохранением рамки считывания.
Функциональные эквиваленты
Термины “функциональные эквиваленты” и “функциональные варианты” являются взаимозаменяемыми. Функциональные эквиваленты ДНК настоящего изобретения представляют собой выделенные ДНК-фрагменты, кодирующие полипептид, который обладает конкретной функцией определенной здесь протеазы A.niger. Функциональным эквивалентом полипептида настоящего изобретения является полипептид, который обладает, по крайней мере, одной функцией определенной здесь протеазы A.niger.
Функциональные эквиваленты белка или полипептида могут содержать лишь консервативные замены одной или более аминокислот в последовательности, выбранной из группы, состоящей из SEQ ID NO:115 - SEQ ID NO:171, или замены, инсерции или делеции заменимых аминокислот. В соответствии с этим заменимый аминокислотный остаток представляет собой остаток, который может быть заменен в последовательности, выбранной из группы, состоящей из SEQ ID NO:115 - SEQ ID NO:171, без значительного изменения биологической функции. Так, например, предполагается, что особенно неподходящими для модификации являются консервативные аминокислотные остатки протеазных белков настоящего изобретения. Кроме того, консервативные аминокислоты в протеазных белках настоящего изобретения и в других протеазах также являются неподходящими для модификации.
Термин “консервативная замена” означает замену, при которой аминокислотный остаток заменен аминокислотным остатком, имеющим аналогичную боковую цепь. Эти семейства аминокислот известны специалистам, и к ним относятся аминокислоты с основными боковыми цепями (например, лизин, аргинин и гистидин), с кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), с незаряженными полярными боковыми цепями (например, глицин, аспарагины, глутамин, серин, треонин, тирозин, цистеин), с неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), с бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и с ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин).
Функциональные эквиваленты нуклеиновых кислот могут, в основном, содержать молчащие мутации или мутации, которые не изменяют биологическую функцию кодируемого полипептида. В соответствии с этим настоящее изобретение относится к молекулам нуклеиновой кислоты, кодирующим протеазные белки, которые содержат аминокислотные остатки, в основном, не влияющие на конкретную биологическую активность. Такие протеазные белки по своей аминокислотной последовательности отличаются от последовательности, выбранной из группы, состоящей из SEQ ID NO:115 - SEQ ID NO:171, однако они сохраняют, по крайней мере, одну биологическую активность. В одном из вариантов осуществления изобретения выделенная молекула нуклеиновой кислоты включает нуклеотидную последовательность, кодирующую белок, где указанный белок содержит аминокислотную последовательность, по существу гомологичную, по крайней мере, примерно на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или более аминокислотной последовательности, представленной в последовательности, выбранной из группы, состоящей из SEQ ID NO:115 - SEQ ID NO:171.
Так, например, руководство по осуществлению фенотипически молчащих аминокислотных замен можно найти в работе Bowie J.U. et al., Science, 247:1306-1310 (1990), где авторы указывают на то, что существует два основных метода исследования толерантности аминокислотной последовательности к заменам. Первый метод основан на процессе эволюции, в котором мутации либо приобретаются, либо подавляются в результате естественного отбора. Второй метод предусматривает генетическое конструирование для введения аминокислотных замен в конкретные положения клонированного гена, а также отбор или скрининг для идентификации последовательностей, которые сохраняют свои функциональные свойства. Как указывают авторы, эти исследования неожиданно выявили, что белки являются толерантными к аминокислотным заменам. Кроме того, авторы указывают, что эти замены вероятно являются допустимыми в определенном положении данного белка. Так, например, наиболее скрытым аминокислотным остаткам требуются неполярные боковые цепи, тогда как некоторые элементы поверхностных боковых цепей являются, в основном, консервативными. Другие такие фенотипически молчащие замены описаны, например, в работе Bowie et al., см. выше, и в указанных там ссылках.
Выделенная молекула нуклеиновой кислоты, кодирующая протеазный белок, гомологичный белку, выбранному из группы, состоящей из SEQ ID NO:115 - SEQ ID NO:171, может быть сконструирована путем введения одной или более нуклеотидных замен, добавлений или делеций в кодирующие нуклеотидные последовательности, выбранные из группы, состоящей из SEQ ID NO:1 - SEQ ID NO:57, или последовательности, выбранные из группы, состоящей из SEQ ID NO:58 - SEQ ID NO:114, так, чтобы одна или более аминокислотных замен, делеций или инсерций были введены в кодируемый белок. Указанные мутации могут быть введены стандартными методами, такими как сайт-направленный мутагенез и ПЦР-опосредованный мутагенез.
Термин “функциональные эквиваленты” также охватывает ортологи протеазного белка A.niger. Ортологи протеазного белка A.niger представляют собой белки, которые могут быть выделены из других штаммов или видов и которые обладают аналогичной или идентичной биологической активностью. Такие ортологи могут быть легко идентифицированы как содержащие аминокислотную последовательность, по существу гомологичную последовательности, выбранной из группы, состоящей из SEQ ID NO:115 - SEQ ID NO:171.
Используемый здесь термин “по существу гомологичный” относится к первой аминокислотной или нуклеотидной последовательности, которая содержит достаточное или минимальное число аминокислот или нуклеотидов, идентичных или эквивалентных (например, с аналогичной боковой цепью) аминокислотам или нуклеотидам второй аминокислотной или нуклеотидной последовательности, так, чтобы указанные первая и вторая аминокислотная или нуклеотидная последовательности имели общий домен. Например, аминокислотные или нуклеотидные последовательности, которые содержат общий домен, идентичный примерно на 60%, предпочтительно на 65%, более предпочтительно на 70%, а еще более предпочтительно на 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99%, и являются, как определено выше, достаточно идентичными.
Кроме того, в объем настоящего изобретения входят нуклеиновые кислоты, кодирующие другие члены протеазного семейства, которые имеют нуклеотидную последовательность, отличающуюся от последовательности, выбранной из группы, состоящей из SEQ ID NO:1 - SEQ ID NO:57, или последовательности, выбранной из группы, состоящей из SEQ ID NO:58 - SEQ ID NO:114. Кроме того, в объем настоящего изобретения входят нуклеиновые кислоты, кодирующие протеазные белки от других видов, которые имеют нуклеотидную последовательность, отличающуюся от последовательности, выбранной из группы, состоящей из SEQ ID NO:1 - SEQ ID NO:57, или последовательности, выбранной из группы, состоящей из SEQ ID NO:58 - SEQ ID NO:114.
Молекулы нуклеиновой кислоты, соответствующие вариантам (например, природным аллельным вариантам) и гомологам ДНК протеазы настоящего изобретения, могут быть выделены исходя из их гомологии по отношению к описанным здесь нуклеиновым кислотам протеазы, с использованием описанной здесь кДНК или их подходящих фрагментов, в качестве гибридизационного зонда в соответствии со стандартными методами гибридизации, предпочтительно, в условиях гибридизации высокой жесткости.
Специалистам известно, что помимо использования природных аллельных вариантов последовательности протеазы, в нуклеотидные последовательности, содержащиеся в последовательности, выбранной из группы, состоящей из SEQ ID NO:1 - SEQ ID NO:57, или последовательности, выбранной из группы, состоящей из SEQ ID NO:58 - SEQ ID NO:114, могут быть внесены модификации, которые приводят к изменениям в аминокислотной последовательности белка протеазы, но не оказывают заметного влияния на функцию протеазного белка.
В другом своем аспекте настоящее изобретение относится к улучшенным протеазным белкам. “Улучшенные” протеазные белки представляют собой белки, которые обладают, по крайней мере, одной улучшенной биологической активностью. Такие белки могут быть получены путем введения неспецифических мутаций по всей кодирующей последовательности протеазы или ее части, например, методом насыщающего мутагенеза, и полученные мутанты могут быть рекомбинантно экспрессированы и скринированы на их биологическую активность. Так, например, в литературе описаны стандартные анализы для измерения ферментативной активности протеаз, и улучшенные таким образом белки могут быть легко отобраны.
В предпочтительном варианте осуществления изобретения протеазный белок имеет аминокислотную последовательность, соответствующую последовательности, выбранной из группы, состоящей из SEQ ID NO:115 - SEQ ID NO:171. В другом варианте осуществления изобретения полипептид протеазы, в основном, гомологичен аминокислотной последовательности, соответствующей последовательности, выбранной из группы, состоящей из SEQ ID NO:115 - SEQ ID NO:171, и сохраняет, по крайней мере, одну биологическую активность полипептида, соответствующего последовательности, выбранной из группы, состоящей из SEQ ID NO:115-171, но при этом он отличается по своей аминокислотной последовательности, что обусловлено природными изменениями или мутагенезом, как описано выше.
В еще одном предпочтительном варианте осуществления изобретения протеазный белок имеет аминокислотную последовательность, кодируемую выделенным фрагментом нуклеиновой кислоты, способным гибридизоваться с нуклеиновой кислотой соответствующей последовательности, выбранной из группы, состоящей из SEQ ID NO:1 - SEQ ID NO:57, или последовательности, выбранной из группы, состоящей из SEQ ID NO:58 - SEQ ID NO:114, предпочтительно, в условиях гибридизации высокой жесткости.
В соответствии с этим протеазный белок представляет собой белок, включающий аминокислотную последовательность, которая, по крайней мере, примерно на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или более гомологична аминокислотной последовательности, представленной в последовательности, выбранной из группы, состоящей из SEQ ID NO:115 - SEQ ID NO:171, и сохраняет, по крайней мере, одну функциональную активность полипептида, соответствующего последовательности, выбранной из группы, состоящей из SEQ ID NO:115 - SEQ ID NO:171.
Функциональные эквиваленты белка настоящего изобретения могут быть также идентифицированы, например, путем скрининга комбинаторных библиотек мутантов, например усеченных мутантов белка настоящего изобретения, на протеазную активность. В одном из вариантов осуществления изобретения генеририруют дискретную библиотеку вариантов путем комбинаторного мутагенеза на уровне нуклеиновой кислоты. Такая дискретная библиотека вариантов может быть продуцирована, например, путем ферментативного лигирования смеси синтетических олигонуклеотидов в генную последовательность, так, чтобы вырожденная серия потенциальных белковых последовательностей экспрессировалась в виде отдельных полипептидов или, альтернативно, в виде набора более крупных гибридных белков (например, для фагового представления). Для продуцирования библиотек потенциальных вариантов полипептидов настоящего изобретения из вырожденной олигонуклеотидной последовательности может быть использован ряд методов. Методы синтеза вырожденных олигонуклеотидов известны специалистам (см. например, Narang (1983) Tetrahedron 39:3; Itakura et al. (1984) Annu. Rev. Biochem. 53:323; Itakura et al. (1984) Science 198:1056; Ike et al. (1983) Nucleic Acid Res. 11:477).
Кроме того, библиотеки фрагментов кодирующей последовательности полипептида настоящего изобретения могут быть использованы для генерирования дискретной популяции полипептидов в целях скрининга для последующего отбора вариантов. Так, например, библиотека фрагментов кодирующей последовательности может быть генерирована путем обработки двухцепочечного ПЦР-фрагмента нужной кодирующей последовательности нуклеазой в условиях, при которых на одну молекулу приходится только один однонитевый разрыв (“ник”); путем денатурации двухцепочечной ДНК; ренатурации ДНК с образованием двухцепочечной ДНК, которая может включать смысловую/антисмысловую пары от различных ник-продуктов; удаления одноцепочечных фрагментов из вновь образованных дуплексов путем обработки нуклеазой S1; и лигирования полученной библиотеки фрагментов в экспрессирующий вектор. Таким методом может быть получена экспрессионная библиотека, которая кодирует N-концевые и внутренние фрагменты нужного белка различных размеров.
Специалистам известны несколько методов скрининга генных продуктов комбинаторных библиотек, полученных посредством точковых мутаций с усечением, и скрининга кДНК-библиотек генных продуктов, имеющих выбранное свойство. Методы, подходящие для высокоэффективных анализов и наиболее широко используемые для скрининга крупных генных библиотек, обычно предусматривают клонирование генной библиотеки в реплицируемые экспрессирующие векторы, трансформацию соответствующих клеток полученной библиотекой векторов и экспрессию комбинированных генов в условиях, при которых детекция нужной активности облегчает выделение вектора, включающего ген, продукты которого были детектированы. Рекурсивный множественный мутагенез (RЕМ), который представляет собой способ увеличения частоты функциональных мутантов в библиотеках, может быть использован в комбинации со скрининг-анализами для идентификации вариантов белка настоящего изобретения (Аrkin & Yourvan (1992) Proc.Natl. Acad. Sci., USA, 89:7811-7815; Delgrave et al. (1993), Protein Engineering 6(3):327-331).
Помимо последовательности протеазного гена, представленной в последовательности, выбранной из группы, состоящей из SEQ ID NO:1-SEQ ID NO:57, для специалиста очевидно, что в данной популяции может присутствовать полиморфизм ДНК-последовательности, который может приводить к изменениям в аминокислотной последовательности протеазного белка. Такой генетический полиморфизм может присутствовать в клетках различных популяций или внутри одной популяции, что обусловлено природной аллельной вариацией. Аллельные варианты могут также включать функциональные эквиваленты.
Фрагменты полинуклеотида настоящего изобретения могут также включать полинуклеотиды, которые не кодируют функциональные полипептиды. Такие полинуклеотиды могут функционировать как зонды или праймеры для ПЦР-реакции. Такие полинуклеотиды могут быть также использованы в том случае, если желательно предотвратить функциональную активность протеазы в конкретном организме (“нокаут”-мутанты).
Нуклеиновые кислоты настоящего изобретения независимо от того, кодируют ли они функциональные или нефункциональные полипептиды, могут быть использованы в качестве гибридизационных зондов или праймеров полимеразной цепной реакции (ПЦР). Использование молекул нуклеиновой кислоты настоящего изобретения, которые не кодируют полипептид, обладающий протеазной активностью, включает, помимо прочего, (1) выделение гена, кодирующего протеазный белок, или его аллельных вариантов из кДНК-библиотеки, например, из других организмов, не являющихся A.niger; (2) in situ гибридизацию (например, FISH) с препаратами метафазной хромосомы в целях обеспечения точной хромосомной локализации гена протеазы, как описано Verma et al., Human Chromosomes: A Manual of Basic Techniques, Pergamon Press, New York (1988); (3) Нозерн-блот-анализ для детекции экспрессии мРНК протеазы в специфических тканях и/или клетках и (4) возможное использование зондов и праймеров в качестве диагностического инструмента для анализа на присутствие нуклеиновой кислоты, гибридизуемой с протеазным зондом в данном биологическом образце (например, в ткани).
В объем настоящего изобретения также входит способ получения функционального эквивалента гена протеазы или кДНК. Такой способ предусматривает получение меченого зонда, который включает выделенную нуклеиновую кислоту, кодирующую всю или часть последовательности, соответствующей последовательности, выбранной из группы, состоящей из SEQ ID NO:115 - SEQ ID NO:171, или ее варианта; скрининг библиотеки фрагментов нуклеиновых кислот с использованием меченого зонда в условиях, позволяющих проводить гибридизацию зонда с фрагментами нуклеиновых кислот в библиотеке с образованием дуплексов нуклеиновых кислот; и продуцирование полноразмерной генной последовательности из фрагментов нуклеиновых кислот в любом меченом дуплексе с получением гена, родственного гену протеазы.
В одном из вариантов изобретения нуклеиновая кислота протеазы настоящего изобретения, по крайней мере, примерно на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более гомологична последовательности нуклеиновой кислоты, представленной в последовательности, выбранной из группы, состоящей из SEQ ID NO:1 - SEQ ID NO:57, или последовательности, выбранной из группы, состоящей из SEQ ID NO:58-SEQ ID NO:114, или комплементарных им последовательностей.
В другом предпочтительном варианте полипептид протеазы, по крайней мере, на 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более гомологичен аминокислотной последовательности, представленной в последовательности, выбранной из группы, состоящей из SEQ ID NO:115 - SEQ ID NO:171.
Клетки-хозяева
В другом варианте своего осуществления настоящее изобретение относится к клеткам, например, к трансформированным клеткам-хозяевам или к рекомбинантным клеткам-хозяевам, которые содержат нуклеиновую кислоту, входящую в объем настоящего изобретения. Термин “трансформированная клетка” или “рекомбинантная клетка” означает клетку (или ее предшественник), в которую, методами рекомбинантных ДНК, введена нуклеиновая кислота настоящего изобретения. Прокариотическими и эукариотическими клетками являются, например, бактерии, грибы, дрожжи и т.п., а особенно предпочтительными клетками являются клетки нитчатых грибов, а в частности, Aspergillus niger.
Клетка-хозяин может быть выбрана так, чтобы в ней осуществлялась модуляция экспрессии встроенных последовательностей либо осуществлялась модификация и процессинг генного продукта в соответствии со специфическим желаемым механизмом. Такие модификации (например, гликозилирование) и процессинг (например, отщепление) белковых продуктов могут облегчать оптимальное функционирование белка.
Различные клетки-хозяева имеют характерные и специфические механизмы посттрансляционного процессинга и модификации белков и генных продуктов. Соответствующие клеточные линии или системы хозяев, известные специалистам в области молекулярной биологии и/или микробиологии, могут быть выбраны так, чтобы они обеспечивали нужную и правильную модификацию и процессинг экспрессируемого чужеродного белка. Для этой цели могут быть использованы эукариотические клетки-хозяева, обладающие клеточным механизмом, обеспечивающим соответствующий процессинг первичного транскрипта, гликозилирование и фосфорилирование генного продукта. Такие клетки-хозяева хорошо известны специалистам.
Клетками-хозяевами также являются, но не ограничиваются ими, клеточные линии млекопитающих, такие как СНО, VЕRO, ВНК, НеLa, СОS, МDCK, 293, 3Т3, WI38 и клеточные линии хороидного сплетения.
Если необходимо, то полипептиды настоящего изобретения могут быть продуцированы с помощью стабильно трансфецированной клеточной линии. Существует ряд векторов, подходящих для стабильной трансфекции клеток млекопитающих и доступных специалистам, и известны методы конструирования таких клеточных линий, описанные, например, Ausubel et al. (см. выше).
Антитела
Кроме того, настоящее изобретение относится к антителам, таким как моноклональные или поликлональные антитела, которые специфически связываются с белками протеазы настоящего изобретения.
Используемые здесь термины “антитело” (Ab) или “моноклональное антитело” (mAb) означают интактные молекулы, а также фрагменты антител (такие как, например, Fab- и F(ab')2-фрагменты), которые способны специфически связываться с протеазным белком. Fab- и F(ab')2-фрагменты не содержат Fc-фрагмент интактного антитела; быстрее выводятся из кровотока и могут иметь меньшую степень неспецифического связывания, чем интактное антитело (Wahl et al., J. Nucl. Med. 24:316-325 (1983)). А поэтому указанные фрагменты являются предпочтительными.
Антитела настоящего изобретения могут быть получены любыми методами. Так, например, клетки, экспрессирующие протеазный белок или его антигенный фрагмент, могут быть введены животному для стимуляции продуцирования сыворотки, содержащей поликлональные антитела. В предпочтительном методе получают и очищают препарат протеазного белка, который, по существу не содержит природных примесей. Затем такой препарат вводят животному для продуцирования поликлональной антисыворотки с более высокой специфической активностью.
В наиболее предпочтительном способе антителами настоящего изобретения являются моноклональные антитела (или их фрагменты, связывающиеся с протеазным белком). Такие моноклональные антитела могут быть получены с использованием гибридомной техники (Kohler et al., Nature 256:495 (1975); Kohler et al., Eur. J. Immunol. 6:511 (1976); Hammerling et al., Monoclonal Antibodies and T-Cell Hybridomas, Elsevier, N.Y., pp. 563-681 (1981)). В основном, такие процедуры предусматривают иммунизацию животного (предпочтительно, мыши) антигеном протеазного белка или клеткой, экспрессирующей протеазный белок. Спленоциты таких мышей выделяют и подвергают слиянию с подходящей миеломной клеточной линией. В соответствии с настоящим изобретением может быть использована любая подходящая миеломная клеточная линия, однако предпочтительно использовать родительскую миеломную клеточную линию (SР2O), имеющуюся в Американской коллекции типовых культур, Роквилл, шт. Мэриленд. После слияния полученные гибридомные клетки выдерживают в селективной среде НАТ, а затем клонируют путем ограниченного разведения, как описано Wands et al. (Gastro-enterology, 80:225-232 (1981)). Гибридомные клетки, полученные посредством такого отбора, затем анализируют для идентификации клонов, которые секретируют антитела, способные связываться с антигеном протеазного белка. Вообще говоря, эти полипептиды могут быть присоединены к белку-носителю, такому как КLН, как описано Ausubel et al. (см. выше), смешаны с адъювантом и инъецированы млекопитающему-хозяину.
В частности, различные животные-хозяева могут быть иммунизованы путем инъекции нужного полипептида. Примерами подходящих животных-хозяев являются кролики, мыши, морские свинки и крысы. Для повышения иммунологического ответа могут быть использованы различные адъюванты в зависимости от вида хозяина, включая, но не ограничиваясь ими, адъювант Фрейнда (полный и неполный), адъювант на основе минеральных гелей, такой как гидроксид алюминия, поверхностно-активные вещества, такие как лизолецитин, полиолы-плюроники, полианионы, пептиды, масляные эмульсии, гемоцианин лимфы улитки, динитрофенол, БЦЖ (бацилла Кальмета-Герена) и Corynebacterium parvum. Поликлональные антитела представляют собой гетерогенные популяции молекул антител, присутствующих в сыворотке иммунизированных животных.
Такими антителами может быть любой иммуноглобулин классов IgG, IgМ, IgЕ, IgА, IgD и любого из их подклассов. Гибридомы, продуцирующие mAb настоящего изобретения, могут быть культивированы in vitro или in vivo.
После продуцирования поликлональные или моноклональные антитела тестируют на специфическое распознавание протеазного полипептида или его функционального эквивалента в иммуноанализе, таком как Вестерн-блот-анализ или анализ на иммунопреципитацию с использованием стандартной техники, например, описанной Ausubel et al. (см. выше). В настоящем изобретении могут быть использованы антитела, которые специфически связываются с протеазными белками или с их функциональными эквивалентами. Так, например, такие антитела могут быть использованы в иммуноанализе для детекции протеазы в патогенных или непатогенных штаммах Aspergillus (например, в экстрактах Aspergillus).
Антитела настоящего изобретения предпочтительно продуцируют с использованием полипептидных фрагментов протеазы, которые, вероятно, также являются антигенными, на что указывает такой критерий, как высокая частота заряженных остатков. Например, такие фрагменты могут быть генерированы стандартными ПЦР-методами, а затем клонированы в экспрессирующий вектор рGEX (Ausubel et al., см. выше). Затем гибридные белки могут быть экспрессированы в E.coli и очищены с использованием аффинной матрицы с глутатион-агарозой, как описано Ausubel et al. (см. выше). Если необходимо, то для каждого белка могут быть генерированы несколько (например, два или три) гибридов, и каждый гибрид может быть инъецирован, по крайней мере, двум кроликам. Антисыворотка может быть продуцирована путем последовательных инъекций, обычно, по крайней мере, трех бустер-инъекций. В основном, антисыворотку оценивают на ее способность к иммунопреципитации рекомбинантного полипептида протеазы или его функциональных эквивалентов, тогда как несвязанные белки могут служить в качестве контроля для оценки специфичности иммунной реакции.
Альтернативно, методы, описанные для продуцирования одноцепочечных антител (в патентах США № 4946778 и 4704692), могут быть адаптированы для продуцирования одноцепочечных антител против протеазного полипептида или его функциональных эквивалентов. Существуют коммерчески доступные наборы для генерирования и скрининга библиотек фагового представления, поставляемые, например, фирмой Pharmacia.
Кроме того, примеры методов и реагентов, особенно подходящих для использования в генерировании и скрининге библиотеки представления антител, можно найти, например, в патенте США № 5223409; в публикации РСТ № WO 92/18619; в публикации РСТ № WO 91/17271; в публикации РСТ № WO 20791; в публикации РСТ № WO 92/20791; в публикации РСТ № WO 92/15679; в публикации РСТ № WO 93/01288; в публикации РСТ № WO 92/01047; в публикации РСТ № WO 92/09690; в публикации РСТ № WO 90/02809; Fuchs et al. (1991) Bio/Technology, 9:1370-1372; Hay et al. (1992) Hum. Antibod. Hybridomas, 3:81-85; Huse et al. (1989) Science 246:1275-1281; Griffiths et al. (1993) EMBO J. 12:725-734.
Поликлональные и моноклональные антитела, которые специфически связываются с полипептидами протеазы или с их функциональными эквивалентами, могут быть использованы, например, для детекции экспрессии гена протеазы или его функциональных эквивалентов, например, в другом штамме Aspergillus.
Так, например, полипептид протеазы может быть легко детектирован в стандартных иммуноанализах клеток или экстрактов Aspergillus. Примерами подходящих анализов являются, но не ограничиваются ими, Вестерн-блоттинг, ELISA, радиоиммуноанализы и т.п.
Термин “специфическое связывание” означает, что антитело распознает конкретный антиген и связывается с ним, например, полипептид протеазы, и, по существу, не распознает другие неродственные молекулы в образце и не связывается с ними.
Антитела могут быть очищены, например, методами аффинной хроматографии, в которых полипептидный антиген иммобилизован на смоле.
Антитело, направленное против полипептида настоящего изобретения (например, моноклональное антитело), может быть использовано для выделения полипептида стандартными методами, такими как аффинная хроматография или иммунопреципитация. Кроме того, такое антитело может быть использовано в целях детекции белка (например, в клеточном лизате или в клеточном супернатанте) для оценки уровня и характера экспрессии этого полипептида. Эти антитела могут быть также использованы в диагностических целях для мониторинга уровня белков в клетках или в тканях, как часть процедуры клинического тестирования, например, для определения эффективности данной схемы лечения или для установления диагноза аспергиллеза.
Детекция может быть облегчена путем связывания антитела с детектируемым веществом. Примерами детектируемых веществ являются различные ферменты, простетические группы, флуоресцентные материалы, люминесцентные материалы, биолюминесцентные материалы и радиоактивные материалы. Примерами подходящих ферментов являются пероксидаза хрена, щелочная фосфатаза, β-галактозидаза или ацетилтрансфераза; примерами подходящих флуоресцентных материалов являются умбеллиферон, флуоресцеин, флуоресцеинизотиоцианат, родамин, дихлортриазиниламинфлуоресцеин, данзилхлорид или фикоэритрин; примером люминесцентного материала является люминол; примерами биолюминесцентных материалов являются люцифераза, люциферин и экворин, а подходящими примерами радиоактивных материалов являются 125I, 131I, 35S или 3Н.
Предпочтительными эпитопами, охватываемыми антигенным пептидом, являются области, локализованные на поверхности белка, например, гидрофильные области. Графики гидрофобности белков настоящего изобретения могут быть использованы для идентификации гидрофобных участков. Антигенный пептид белка настоящего изобретения включает, по крайней мере, 7 (предпочтительно 10, 15, 20 или 30) смежных аминокислотных остатков аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO:115 - SEQ ID NO:171, и содержит эпитоп белка, который стимулирует продуцирование антител против пептида, образующих специфический иммунный комплекс с белком.
Предпочтительными эпитопами, охватываемыми антигенным пептидом, являются области протеазы, которые локализованы на поверхности белка, например гидрофильные области, гидрофобные области, альфа-области, бета-области, спиральные области, области витка и гибкие области.
Иммуноанализы
Качественное или количественное определение полипептида настоящего изобретения в биологическом образце может быть осуществлено любым известным методом. Техника с использованием антител является особенно предпочтительной для оценки конкретных уровней полипептидов в биологическом образце.
При этом специфическое распознавание осуществляется “первым” антителом (поликлональным или моноклональным антителом), а во вторичной системе детекции могут быть использованы антитела, конъюгированные с флуоресцентными веществами, ферментами, или другие конъюгированные “вторые” антитела. В результате получают иммунокомплекс.
В соответствии с этим настоящее изобретение относится к способу диагностики Aspergillus-инфекции у определенного организма, включающему стадии:
- выделения биологического образца из указанного организма, который, как предполагается, инфицирован Aspergillus,
- взаимодействия указанного биологического образца с антителом настоящего изобретения,
- оценки на образование иммунокомплексов.
Для высвобождения белка в целях проведения Вестерн-блот-анализа или дот/блот-анализа ткани могут также быть экстрагированы, например, мочевиной и нейтральным детергентом. Эта техника может быть также применена и к физиологическим жидкостям.
Другими методами на основе антител, используемыми для детекции экспрессии гена протеазы, являются иммуноанализы, такие как твердофазный иммуноферментный анализ (ELISA) и радиоммуноанализ (РИА). Так, например, моноклональные антитела, специфичные к протеазе, могут быть использованы в качестве иммуноабсорбента и в качестве меченного ферментом зонда для детекции и количественной оценки протеазного белка. Количество протеазного белка, присутствующего в данном образце, может быть вычислено по количеству белка, присутствующему в стандартном препарате, с использованием компьютерного алгоритма линейной регрессии. В другом ELISA-анализе могут быть использованы два различных специфических моноклональных антитела для детекции протеазного белка в биологической жидкости. В этом анализе одно из антител используется в качестве иммуноабсорбента, а другое - в качестве меченного ферментом зонда.
Вышеуказанные методы могут быть осуществлены, в основном, в виде “одностадийного” или “двухстадийного” анализа. “Одностадийный анализ” предусматривает контактирование протеазного белка с иммобилизованным антителом без промывки и контактирование этой смеси с меченым антителом. “Двухстадийный анализ” предусматривает промывку, а затем контактирование этой смеси с меченым антителом. Могут быть также использованы и другие подходящие методы. Обычно оказывается предпочтительным иммобилизовать один компонент аналитической системы на носителе, что позволяет другим компонентам этой системы вступать в контакт с указанным компонентом и легко выделяться из образца.
Подходящими ферментными метками являются, например, метки, происходящие от оксидазной группы, которая катализирует продуцирование перекиси водорода посредством взаимодействия с субстратом. Активность оксидазной метки может быть проанализирована путем измерения концентрации перекиси водорода, образованной посредством взаимодействия меченного ферментом антитела с субстратом.
Помимо ферментов, другими подходящими метками являются радиоизотопы, такие как йод (125I, 121I), углерод (14C), сера (35S), тритий (3Н), индий (112In) и технеций (99mТс), а также флуоресцентные метки, такие как флуоресцеин и родамин, и биотин.
Специфическое связывание тестируемого соединения с полипептидом протеазы может быть детектировано, например, in vitro, путем обратимой или необратимой иммобилизации протеазного полипептида на субстрате, например, на поверхности лунки 96-луночного полистиролового микротитрационного планшета. Методы иммобилизации полипептидов и других небольших молекул хорошо известны специалистам. Так, например, микротитрационные планшеты могут быть сенсибилизированы полипептидом протеазы путем добавления в каждую лунку полипептида в растворе (обычно в концентрации от 0,05 до 1 мг/мл в объеме 1-100 мкл) и инкубирования этих планшетов при температуре в пределах от комнатной температуры до 37°С в течение 0,1-36 часов. Полипептиды, не связанные с планшетом, могут быть удалены путем “вытряхивания” избытка раствора из планшетов, с последующей промывкой планшета (однократной или многократной) водой или буфером. Обычно полипептид содержится в воде или в буфере. Затем планшет промывают буфером, не содержащим связанного полипептида. Для блокирования сайтов, связывающихся со свободным белком на планшетах, эти планшеты блокируют белком, который не является родственным связанному полипептиду. Например, подходящим для этой цели является 300 мкл бычьего сывороточного альбумина (ВSA) в концентрации 2 мг/мл в Трис-HCl. Подходящими субстратами являются субстраты, которые содержат определенный сшивающий химический агент (например, пластиковые субстраты, такие как полистироловые, стироловые или, например, полипропиленовые субстраты от Corning Costar Corp. (Cambridge, MA). Если необходимо, то в качестве субстрата может быть использована гранулированная частица, например, гранулированная агароза или гранулированная сефароза.
Связывание тестируемых соединений с полипептидами настоящего изобретения может быть детектировано любым из известных методов. Так, например, в иммуноанализе может быть использовано специфическое антитело. Если необходимо, то это антитело может быть помечено (например, флуоресцентной меткой или радиоизотопом) и детектировано прямым методом (см., например, West & McMahon J. Cell. Biol. 74:264, 1977). Альтернативно, для детекции может быть использовано “второе” антитело (например, меченое антитело, которое связывается с Fc-частью анти-АN97 антитела). В альтернативном методе детекции протеазный полипептид подвергают мечению (например, мечению протеазного полипептида радиоизотопом, флуорофором, хромофором или т.п.) и эту метку детектируют. В еще одном способе полипептид протеазы продуцируют в виде гибридного белка, который может быть оптически детектирован, например, с белком, флуоресцирующим в зеленом диапазоне спектра (который может быть детектирован при УФ-излучении). В альтернативном методе полипептид протеазы может быть ковалентно присоединен к ферменту или слит с ферментом, обладающим детектируемой ферментативной активностью, таким как пероксидаза хрена, щелочная фосфатаза, альфа-галактозидаза или глюкозооксидаза. Гены, кодирующие все указанные ферменты, были клонированы и легко доступны каждому специалисту. Если необходимо, то гибридный белок может включать антиген, и такой антиген может быть детектирован и измерен с использованием поликлонального или моноклонального антитела стандартными методами. Подходящими антигенами являются ферменты (например, пероксидаза хрена, щелочная фосфатаза и α-галактозидаза) и неферментные полипептиды (например, сывороточные белки, такие как ВSA и глобулины, а также молочные белки, такие как казеины).
Эпитопы, антигены и иммуногены
В другом аспекте настоящее изобретение относится к пептиду или полипептиду, содержащему эпитопнесущую часть полипептида настоящего изобретения. Эпитоп этой части полипептида представляет собой иммуногенный или антигенный эпитоп полипептида настоящего изобретения. Термин “иммуногенный эпитоп” определяют как часть белка, которая вырабатывает гуморальный ответ в том случае, если весь белок является иммуногенным. Очевидно, что эти иммуногенные эпитопы ограничены на молекуле несколькими локусами. С другой стороны, область молекулы белка, с которой может связываться антитело, определяют как “антигенный эпитоп”. Число иммуногенных эпитопов белка обычно меньше, чем число антигенных эпитопов. См., например, Geysen H.M. et al., Proc. Natl. Acad. Sci. USA 81:3998-4002 (1984).
Что касается выбора пептидов или полипептидов, несущих антигенный эпитоп (то есть содержащих область молекулы белка, с которой может связываться антитело), то специалистам хорошо известно, что относительно короткие синтетические пептиды, имитирующие часть последовательности белка, обычно способны продуцировать антисыворотку, которая реагирует с частично имитированным белком. См. например, Sutcliffe J.G. et al., Science 219:660-666 (1984). Пептиды, способные продуцировать реагирующую с белком сыворотку и часто представленные в первичной последовательности белка, могут быть охарактеризованы в соответствии с простыми химическими правилами и не ограничиваются ни иммунодоминантными областями интактного белка (то есть иммуногенными эпитопами), ни амино- или карбоксиконцами. Пептиды, которые являются в высокой степени гидрофобными, и пептиды, состоящие из шести или менее остатков, обычно являются неэффективными в индуцировании антител, связывающихся с имитированным белком, тогда как более длинные растворимые пептиды, особенно пептиды, содержащие пролиновые остатки, обычно являются эффективными. Sutcliffe et al., см.выше. Так, например, 18-20 пептидов, сконструированных в соответствии с вышеуказанными руководствами и содержащих 8-39 остатков, охватывающих 75% последовательности полипептидной цепи гемагглютинина НАI вируса гриппа, индуцировали антитела, которые реагировали с белком НАI или с интактным вирусом; а пептиды 12/12, происходящие от полимеразы MuLV, и пептид 18/18, происходящий от гликопротеина рабивируса, индуцировали антитела, которые осаждали соответствующие белки.
Пептиды и полипептиды настоящего изобретения, несущие антигенный эпитоп, могут быть использованы для продуцирования антител, включая моноклональные антитела, которые специфически связываются с полипептидом настоящего изобретения. Таким образом, большая часть гибридом, полученных путем слияния клеток селезенки, взятых у доноров, иммунизованных пептидом, несущим антигенный эпитоп, обычно секретирует антитело, реагирующее с нативным белком. Sutcliffe et al., см.выше, 663. Антитела, продуцируемые антигенным эпитопом, несущим пептиды или полипептиды, могут быть использованы для детекции имитированного белка, а антитела против различных пептидов могут быть использованы для слежения за “судьбой” различных участков белков-предшественников, которые подвергаются посттрансляционному процессингу. Пептиды и антитела против пептидов могут быть использованы в различных качественных или количественных анализах в целях определения имитированного белка, например, в анализах на конкурентное связывание, поскольку, как было показано, даже короткие пептиды (например, состоящие приблизительно из 9 аминокислот) могут связываться и замещать более крупные пептиды в иммунопреципитационных анализах. См. например, Wilson I.A. et al., Cell 37:767-778, at 777 (1984). Антитела против пептидов настоящего изобретения могут быть также использованы для очистки имитированного белка, например, путем адсорбционной хроматографии методами, хорошо известными специалистам.
Антигенные эпитопнесущие пептиды и полипептиды настоящего изобретения, сконструированные в соответствии с вышеуказанными инструкциями, содержат последовательность, состоящую, по крайней мере, из семи, более предпочтительно, по крайней мере, из девяти, а наиболее предпочтительно примерно из 15-30 аминокислот, присутствующих в аминокислотной последовательности полипептида настоящего изобретения. Однако пептиды или полипептиды, включающие более крупную часть аминокислотной последовательности полипептида настоящего изобретения, содержащую примерно от 30 до 50 аминокислот или имеющую любую длину, включая полную аминокислотную последовательность полипептида настоящего изобретения также рассматриваются как эпитопнесущие пептиды или полипептиды настоящего изобретения и могут быть использованы для индуцирования антител, реагирующих с имитированным белком. Предпочтительно, аминокислотную последовательность эпитопнесущего пептида выбирают так, чтобы она обеспечивала хорошую растворимость в водных растворителях (то есть так, чтобы эта последовательность включала относительно гидрофильные остатки и, предпочтительно, в основном, не содержала в высокой степени гидрофобных остатков); а особенно предпочтительными являются последовательности, содержащие пролиновые остатки.
Эпитопнесущие пептиды и полипептиды настоящего изобретения могут быть продуцированы любым стандартным способом получения пептидов или полипептидов, включая рекомбинантные методы с использованием молекул нуклеиновой кислоты настоящего изобретения. Так, например, короткая эпитопнесущая аминокислотная последовательность может быть слита с более крупным действующим как носитель полипептидом в процессе рекомбинантного продуцирования и рекомбинантной очистки, а также в процессе иммунизации для продуцирования антител против пептидов.
Эпитопнесущие пептиды могут быть также синтезированы известными методами химического синтеза. Так, например, Houghten описал простой метод синтеза большого числа пептидов, таких как 10-20 мг из 248 различных пептидов, состоящих из 13 остатков и представляющих собой варианты сегмента полипептида НАI, которые отличаются на одну аминокислоту и которые были получены и охарактеризованы (в анализах по связыванию типа ELISA) за менее чем четыре недели. Houghten R.A. Proc. Natl. Acad. Sci. USA 82:5131-5135 (1985). Этот процесс “одновременного синтеза множества пептидов (SMPS)” был подробно описан в патенте США № 4631211, Houghten et al. (1986). В этой процедуре отдельные смолы для твердофазного синтеза различных пептидов содержатся в отдельных проницаемых для растворителя пакетах, что позволяет оптимально проводить много идентичных повторяющихся стадий в твердофазных методах.
Осуществляемая вручную процедура позволяет одновременно проводить 500-1000 или более синтезов. Houghten et al., см.выше, 5134.
Эпитопнесущие пептиды и полипептиды настоящего изобретения используются для индуцирования антител в соответствии с методами, хорошо известными специалистам. См., например, Sutcliffe et al., см. выше; Wilson et al., см. выше; Chow M. et al., Proc. Natl. Acad. Sci. USA 82:910-914; и Bittle, F.J. et al., J. Gen. Virol. 66:2347-2354 (1985).
В общих чертах, животные могут быть иммунизованы свободным пептидом; однако титр антитела против пептида может быть повышен путем связывания пептида с макромолекулярным носителем, таким как гемоцианин лимфы улитки (КLH) или столбнячный токсоид. Так, например, пептиды, содержащие цистеин, могут быть связаны с носителем посредством линкера, такого как малеимидобензоил-N-гидроксисукцинимидоэфир (МВS), а другие пептиды могут быть связаны с носителем посредством более общего связывающего агента, такого как глутаральдегид.
Животных, таких как кролики, крысы и мыши, иммунизируют либо свободными, либо связанными с носителем пептидами, например, путем внутрибрюшинной и/или внутрикожной инъекции эмульсий, содержащих 10 мкг пептида или белка-носителя и адъюванта Фрейнда. Может оказаться необходимым введение нескольких бустер-инъекций, например, через интервалы времени примерно в две недели, для получения нужного титра антитела против пептида, которое может быть детектировано, например, с помощью ELISA-анализа с использованием свободного пептида, адсорбированного на твердой поверхности. Титр антител против пептида в сыворотке иммунизованного животного может быть увеличен путем отбора антител против пептида, например, посредством адсорбции к пептиду на твердом носителе и элюирования отобранных антител в соответствии с методами, хорошо известными специалистам.
Иммуногенные эпитопнесущие пептиды настоящего изобретения, то есть те части белка, которые вырабатывают гуморальный ответ, если весь белок является иммуногеном, идентифицируют методами, известными специалистам. Так, например, Geysen et al., 1984, см. выше, описал процедуру быстрого конкурентного синтеза на твердых носителях сотен пептидов с чистотой, достаточной для реакции в твердофазном иммуноферментном анализе. Взаимодействие синтезированных пептидов с антителами может быть затем легко детектировано без их отделения от носителя. В этом способе пептид, несущий иммуногенный эпитоп нужного белка, может быть идентифицирован стандартными методами, известными специалистам. Так, например, локализация иммунологически важного эпитопа в оболочечном белке вируса ящура с разрешением в семь аминокислот была определена Geysen и др. путем синтеза перекрывающейся серии из всех 208 возможных гексапептидов, покрывающих всю последовательность белка из 213 аминокислот. Затем была синтезирована полная серия пептидов с заменами, в которой все 20 аминокислот, в свою очередь, были заменены в каждом положении внутри эпитопа, и были определены конкретные аминокислоты, сообщающие специфичность для реакции с антителом. Таким образом, пептидные аналоги эпитопнесущих пептидов настоящего изобретения могут быть легко получены указанным методом. В патенте США № 4708781, Geysen (1987), кроме того, описан способ идентификации пептида, несущего иммуногенный эпитоп нужного белка.
Кроме того, в патенте США № 5194392, Geysen (1990) описан общий метод детекции или определения последовательности мономеров (аминокислот или других соединений), представляющей собой топологический эквивалент эпитопа (то есть “мимотоп”), который является комплементарным конкретному паратопу (ангигенсвязывающему сайту) нужного антитела. Более конкретно, в патенте США № 4433092, Geysen (1989) описан способ детекции или определения последовательности мономеров, которая представляет собой топографический эквивалент лиганда, комплементарный конкретному лигандсвязывающему сайту нужного конкретного рецептора. Аналогичным образом, в патенте США № 5480971, Houghten, R.A. et al. (1996), относящемся к смесям пералкилированных олигопептидов, описаны линейные пералкилированные С1-С7-алкил-олигопептиды; наборы и библиотеки таких пептидов, а также методы использования таких наборов и библиотек олигопептидов для определения последовательности пералкилированного олигопептида, который связывается преимущественно с нужной молекулой-акцептором. Таким образом, непептидные аналоги эпитопнесущих пептидов настоящего изобретения также могут быть рутинно изготовлены этими методами.
Подавление или снижение протеазной активности
Настоящее изобретение также относится к способам продуцирования мутантной клетки, полученной из родительской клетки, которые включают дизрупцию или делецию последовательности нуклеиновой кислоты, кодирующей протеазу или ее регуляторную последовательность, что приводит к образованию мутантной клетки, продуцирующей меньшее количество протеазы, чем родительская клетка.
Конструирование штаммов, которые имеют пониженную протеазную активность, может быть, в основном, осуществлено путем модификации или инактивации последовательности нуклеиновой кислоты, необходимой для экспрессии протеазной активности в клетке. Модифицируемая или инактивируемая последовательность нуклеиновой кислоты может представлять собой, например, последовательность нуклеиновой кислоты, кодирующую протеазу или ее часть, играющую важную роль для обеспечения протеазной активности, либо эта последовательность нуклеиновой кислоты может обладать регуляторной функцией, необходимой для экспрессии протеазы из кодирующей последовательности нуклеиновой кислоты. Примером такой регуляторной или контрольной последовательности может быть промоторная последовательность или ее функциональная часть, то есть часть, которая является достаточной для влияния на экспрессию протеазы. Другими регуляторными последовательностями для возможной модификации являются, но не ограничиваются ими, лидерная последовательность, последовательность полиаденилирования, пропептидная последовательность, сигнальная последовательность и сайт терминации.
Модификация или инактивация последовательности нуклеиновой кислоты может быть осуществлена путем мутагенеза данной клетки и отбора клеток, в которых способность к продуцированию протеазы была снижена или подавлена. Мутагенез, который может быть специфическим или неспецифическим, может быть осуществлен, например, с использованием подходящего физического или химического мутагенизирующего агента, с использованием подходящего олигонуклеотида или путем ПРЦ-генерированного мутагенеза ДНК-последовательности. Кроме того, мутагенез может быть осуществлен с использованием любой комбинации таких мутагенизирующих агентов. Примерами физических или химических мутагенизирующих агентов, подходящих для использования в настоящем изобретении, являются ультрафиолетовое (УФ) излучение, гидроксиламин, N-метил-N'-нитро-N-нитрозогуанидин (MNNG), О-метилгидроксиламин, азотистая кислота, этилметансульфонат (EMS), бисульфит натрия, муравьиная кислота и нуклеотидные аналоги. При использовании таких агентов мутагенез обычно осуществляют посредством инкубирования мутагенизированной клетки в присутствии выбранного мутагенизирующего агента в подходящих условиях и отбора клеток, обладающих пониженной протеазной активностью или не экспрессирующих такую протеазную активность.
Модификация или инактивация продуцирования протеазы настоящего изобретения может быть осуществлена путем введения, замены или удаления одного или более нуклеотидов в последовательности нуклеиновой кислоты, кодирующей протеазу, или в регуляторном элементе, необходимом для ее транскрипции или трансляции. Так, например, нуклеотиды могут быть встроены или удалены так, чтобы это приводило к введению стоп-кодона, удалению старт-кодона или к изменению открытой рамки считывания. Такая модификация или инактивация может быть осуществлена с помощью сайт-направленного мутагенеза или ПЦР-генерированного мутагенеза методами, известными специалистам.
Хотя, в принципе, модификация может быть осуществлена in vivo, то есть непосредственно в клетке, экспрессирующей модифицируемую последовательность нуклеиновой кислоты, предпочтительно, чтобы модификация была осуществлена in vitro, как описано ниже.
Примером подходящего пути инактивации или снижения продуцирования протеазы выбранной клеткой-хозяином является способ, основанный на замене гена или прерывания гена. Так, например, в способе прерывания гена последовательность нуклеиновой кислоты, соответствующую эндогенному гену или фрагменту нужного гена, подвергают мутагенезу in vitro с продуцированием дефектной последовательности нуклеиновой кислоты, которую затем трансформируют в клетку-хозяина с продуцированием дефектного гена. Посредством гомологичной рекомбинации дефектная последовательность нуклеиновой кислоты заменяет эндогенный ген или фрагмент гена. Может оказаться желательным, чтобы дефектный ген или генный фрагмент также кодировал маркер, который может быть использован для отбора трансформантов, в которых ген, кодирующий протеазу, был модифицирован или разрушен.
Альтернативно, модификация или инактивация последовательности нуклеиновой кислоты, кодирующая протеазу настоящего изобретения, может быть осуществлена известными методами с использованием антисмысловой нуклеотидной последовательности, комплементарной последовательности, кодирующей протеазу. Более конкретно, продуцирование протеазы клеткой может быть снижено или подавлено посредством введения нуклеотидной последовательности, комплементарной последовательности нуклеиновой кислоты, кодирующей протеазу, которая может транскрибироваться в клетке и способна гибридизоваться с мРНК протеазы, продуцированной в этой клетке. В условиях, позволяющих комплементарной антисмысловой нуклеотидной последовательности гибридизоваться с мРНК протеазы, количество транслируемой таким образом протеазы снижается или подавляется.
При этом, предпочтительно, чтобы клетка, модифицированная способами настоящего изобретения, имела микробное происхождение, например, происходила от грибкового штамма, подходящего для продуцирования нужных белковых продуктов, являющихся гомологичными либо гетерологичными для данной клетки.
Кроме того, настоящее изобретение относится к мутантной клетке, полученной из родительской клетки и включающей дизрупцию или делецию в последовательности нуклеиновой кислоты, кодирующей протеазу, или в регуляторной последовательности, в результате чего эта мутантная клетка продуцирует меньшее количество протеазы, чем родительская клетка.
Полученные таким образом дефицитные по протеазе мутантные клетки являются особенно подходящими в качестве клеток-хозяев, используемых для экспрессии гомологичных и/или гетерологичных полипептидов. Поэтому настоящее изобретение относится к способам продуцирования гомологичного или гетерологичного полипептида, включающим (а) культивирование мутантной клетки в условиях, подходящих для продуцирования полипептида, и (b) выделение полипептида. В контексте настоящего описания используемый здесь термин “гетерологичные полипептиды” означает полипептиды, которые не являются нативными для клетки-хозяина; нативный белок, в который были внесены модификации для изменения нативной последовательности, или нативный белок, экспрессия которого была количественно изменена путем модификации клетки-хозяина методами рекомбинантных ДНК.
Способы настоящего изобретения для продуцирования, по существу, не содержащего протеазы продукта представляют особый интерес для продуцирования эукариотических полипептидов, в частности, грибковых белков, таких как ферменты. Дефицитные по протеазе клетки могут быть также использованы для экспрессии гетерологичных белков, представляющих интерес для пищевой или фармацевтической промышленности.
Использование протеаз в промышленном производстве
Настоящее изобретение также относится к использованию протеазы настоящего изобретения в некоторых способах, применяемых в промышленности и в фармацевтике. Несмотря на многолетний опыт применения этих способов, протеаза настоящего изобретения имеет ряд значительных преимуществ по сравнению с ферментами, используемыми в настоящее время. В зависимости от конкретного применения такими преимуществами могут быть такие факторы, как более низкая себестоимость продукта, более высокая специфичность к субстрату, более низкая антигенность, меньший уровень нежелательных побочных эффектов, более высокие выходы при продуцировании в подходящем микроорганизме, более подходящие диапазоны рН и температур, лучшие вкусовые качества конечного продукта, а также более высокое качество пищи и ее кошерность.
В крупномасштабном промышленном производстве кормов или пищевых продуктов протеолитические ферменты обычно используются для улучшения таких параметров, как растворимость белка, выходы продуктов экстракции, вязкость или вкус, консистенция, питательная ценность, минимизация антигенности или уровня вредных факторов, цвет или функциональность, а также для улучшения технологических параметров, таких как фильтруемость белкового сырья. Для этих применений белковое сырье может быть получено от животного или растения, и его примерами могут служить растительные белки, такие как белок сои, глютен пшеницы, белок семян рапса, белок гороха, белок люцерны, белок подсолнечника, белок бобовых растений, белок хлопчатника или кунжута, белок кукурузы, белок ячменя, белок сорго, картофельный белок, рисовый белок, белки кофе и белки животного происхождения, такие как молочный белок (например, казеин, белок молочной сыворотки), яичный белок, рыбный белок, мясной белок, включая желатин, коллаген, белок крови (например, гемоглобин), белки волос, перьев и рыбной муки.
Важное свойство протеаз настоящего изобретения заключается в том, что они действуют при оптимальном диапазоне рН и оптимальной температуре, которые идеально подходят для различных целей. Так, например, для минимизации риска возникновения микробных инфекций многие крупномасштабные процессы желательно проводить при относительно высоких температурах обработки, т.е. при 50°С или выше. Некоторые протеазы настоящего изобретения удовлетворяют этим требованиям, но при этом они не обладают чрезмерной термостабильностью, такой, которая противостояла бы всем попыткам инактивировать этот фермент путем дополнительной тепловой обработки. Последний из указанных отличительных признаков позволяет осуществлять продуцирование, которое дает конечные продукты, не содержащие остаточной протеолитической активности. Аналогичным образом, многие корма и пищевые продукты имеют слабокислотные значения рН, а поэтому для их обработки предпочтительно использовать протеазы с оптимальным кислотным рН или почти нейтральным рН. Протеаза настоящего изобретения также удовлетворяет этим требованиям.
Специфичность эндопротеаз обычно определяют по предпочтительному расщеплению связей между карбоксилом аминокислотного остатка в положении Р1 и аминогруппой этого остатка в положении Р1' соответственно. Предпочтительность может определяться преимущественно либо Р1 (например, положительно заряженными остатками в субстратах для трипсина), либо Р1' (например, гидрофобными остатками при расщеплении термолизином), либо Р1 и Р2 (например, специфическими расщеплениями связей между двумя положительно заряженными остатками сериновой эндопротеазой мозгового вещества надпочечника). В некоторых случаях более удаленные остатки могут определять предпочтительность расщепления, например, Р2 является предпочтительным для стрептококковой пептидазы А. Известно, что некоторые остатки негативно влияют на отщепление, и также хорошо известно, что пролиновые связи в положении Р1' резистентны к действию многих протеаз. Большинство эндопротеаз расщепляют преимущественно либо в гидрофобном окружении, либо поблизости от отрицательно заряженных остатков. Так, например, используемые в промышленности эндопротеазы, такие как химотрипсин (полученный из бычьей поджелудочной железы) или субтилизин, нейтральная металлоэндопротеаза или термолизин (полученный из видов Bacillus), имеют тенденцию к расщеплению на участке “позади” гидрофобных аминокислот, таких как -Phe, -Leu и -Tyr. Другими используемыми в промышленности эндопротеазами являются трипсин (полученный из бычьей поджелудочной железы), расщепляющий преимущественно позади -Arg и -Lys, и папаин (сложная смесь различных ферментов, включая протеазы, полученные из плодов папайи), расщепляющий преимущественно позади -Arg. В противоположность этому, пептидные связи, образованные небольшими по размеру остатками, такими как Ala, Gly, Ser, Thr, а также Ile и Pro являются плохими субстратами (Keil B. et al., Protein Seq Data Anal. (1993), 5; 401-407). Такая ситуация создает определенные трудности для фармацевтической промышленности, а именно в производстве пищевых продуктов и напитков, в сельском хозяйстве и в химической промышленности. Протеаза настоящего изобретения обладает необычной предпочтительностью в расщеплении.
Экзопептидазы действуют только вблизи концов полипептидных цепей. Такое действие пептидаз у свободного N-конца приводит к высвобождению одного аминокислотного остатка (так называемых аминопептидаз), или дипептида, или трипептида (так называемых дипептидилпептидаз и трипептидилпептидаз). Такое действие пептидаз у свободного С-конца приводит к высвобождению одного аминокислотного остатка (так называемых карбоксипептидаз) или дипептида (так называемых пептидил-дипептидаз). По своему каталитическому механизму карбоксипептидазы можно разделить на три группы, а именно карбоксипептидазы серинового типа, металлокарбоксипептидазы и карбоксипептидазы цистеинового типа. Другие экзопептидазы являются специфичными для дипептидов (так называемые дипептидазы) или способными расщеплять пептидные связи, не являющиеся связями альфа-карбоксильных или альфа-аминогрупп (так называемые омега-пептидазы). Примерами таких новых омега-пептидаз являются пироглутамилпептидаза и ацитаминоацилпептидаза, идентифицированные в настоящем изобретении (см. таблицу 1, гены 18 и 45 соответственно).
Типичными примерами промышленного применения, которое зависит от использования чистых эндопротеаз и в котором протеаза настоящего изобретения может, как предполагается, обладать превосходными свойствами, являются обработка материалов растительного или животного происхождения. Эти стадии обработки могут быть предназначены для модификации большого массива характеристик сырого материала или (частично) очищенной белковой фракции. Так, например, эти стадии обработки могут быть осуществлены для максимизации растворимости продукта, фильтруемости, способности к выделению, выходов белка после экстракции и его усвояемости или для минимизации токсичности, постороннего привкуса и вязкости. Кроме того, такая обработка может быть направлена на изменение физико-химических свойств сырого материала или очищенного (или частично очищенного) белка. Эти преимущества могут быть использованы не только в тех случаях, когда эндопротеаза настоящего изобретения применяется для облегчения обработки в промышленном производстве, но также в случаях, когда применяется в качестве активного ферментного компонента в корме для животных. В частности, эндопротеаза настоящего изобретения может быть использована в качестве улучшителя хлеба в хлебопекарной промышленности, например, для замедления черствения хлеба или для уменьшения вязкости теста. Либо эндопротеаза может быть использована в пивоваренной и винодельческой промышленности для предупреждения или для минимизации образования нежелательных белковых помутнений. Альтернативно, она может быть использована в пивоваренной промышленности для оптимизации выходов продуктов экстракции белка зерновых, используемых для получения сусла. Кроме того, она может быть также преимущественно использована в молочной промышленности в качестве агента свертывания молока с улучшенными свойствами или для оптимизации текстурирования, пенообразования или свертывающих свойств различных компонентов молока. Другим применением новой протеазы настоящего изобретения в молочной промышленности является ее использование для приготовления модифицированных ферментированных сыров (МФС).
Кроме того, различные белковые субстраты могут быть подвергнуты воздействию эндопротеазы настоящего изобретения обычно в комбинации с другими протеолитическими ферментами с получением гидролизатов, предназначенных для использования в медицинских и в немедицинских целях. Неожиданно было обнаружено, что эндопротеаза настоящего изобретения является эффективной для осуществления полного гидролиза белкового субстрата, такого, что при нем полностью гидролизуются даже части, резистентные к протеазе, и, кроме того, было неожиданно обнаружено, что эта эндопротеаза также обладает активностью, направленной на минимизацию аллергенности конечного гидролизата или на подавление образования горького привкуса.
Более конкретно, эндопротеаза настоящего изобретения отличается своими предпочтениями в расщеплении необычных пептидных связей белков, особенно в аминокислотных остатках небольшого размера, таких как Ala, Gly, Ser и Thr, или в остатках Ile и Pro, в положении Р1 или в Р1' (Keil B. et al., Protein Seq Data Anal. (1993) 5; 401-407). В результате этого полученные фракции белковых исходных материалов, которые оказались резистентными к гидролизу после использования известных эндопротеаз, могут быть растворены и гидролизованы с помощью эндопротеазы настоящего изобретения. Неограничивающими примерами таких резистентных к протеазе фракций являются так называемые экстензины в растительных материалах, коллаген, желатин, а также специфические молочные компоненты в материалах животного происхождения.
Различные пищевые продукты, такие как соевые бобы, содержат ингибиторы трипсина. Эти белки ингибируют трипсиновую активность в желудочно-кишечном тракте, например, у свиней и домашних птиц. Эта ингибирующая трипсин активность приводит к недостаточной усвояемости белка этими животными, а поэтому к увеличению отходов и к повышению материальных затрат. Эта проблема может быть частично решена поджариванием соевых бобов при высоких температурах. В соевых бобах были идентифицированы ингибиторы трипсина двух различных типов, а именно ингибиторы трипсина Боумана-Бирка и ингибиторы трипсина Куница.
Настоящее изобретение также относится к альтернативному пути разрушения трипсин-ингибирующей активности при поджаривании, который обеспечивается цистеиновыми протеазами (ЕС 3.4.22, таблица 1), способными расщеплять пептидные связи лейцин 176 - аспартат 177 возле карбоксильного конца ингибитора трипсина Куница (как описано Wilson (1988), CRC Critical Reviews in Biotechnology 8(3):197-216). Такой путь приводит к инактивации указанного ингибитора трипсина в сое. Было неожиданно обнаружено, что цистеиновые протеазы, секретируемые грибами Aspergillus niger, удовлетворяют этим критериям гораздо лучше, чем аналогичные ферменты, происходящие от других организмов.
Протеазы также широко используются в производстве сыра. При производстве сыра молоко для сыроделия необходимо подвергать свертыванию для того, чтобы отделить сырные вещества от молочной сыворотки, например, казеин. Были описаны некоторые ферменты свертывания молока, называемые также коагулянтами, и такими ферментами являются (коровий) химозин, коровий пепсин, свиной пепсин, а также микробные ферменты, такие как протеаза Rhizomucor miehei, протеаза Rhizomucor pusillus и протеаза Cryptonectria parasitica. Химозин может быть получен из желудка теленка, но он также может продуцироваться микробами, например, Kluyveromyces lactis. Все эти ферменты характеризуются тем, что они обладают специфичностью к пептидной связи между остатком 105 (фенилаланин) и остатком 106 (метионин) или к связи, смежной со связью в ĸ-казеине. Это означает, что при использовании таких ферментов в производстве сыра ĸ-казеин расщепляется на стыке между пара-ĸ-казеином и макропептидной частью, называемой гликомакропептидом (GМР) и несущей отрицательные заряды. Если макропептид диффундирует в молочную сыворотку, то его стабилизирующее действие на растворимость казеиновых мицелл утрачивается, и эти казеиновые мицеллы могут начать агрегироваться после достаточного гидролиза каппа-казеина. Подробное описание ферментативного свертывания молока можно найти, например, у D.G. Dalgleish, Advanced Dairy Chemistry, vol.1 ed. by P.F. Fox, Elsevier, London, 1992.
Используемые в настоящее время коагулянты позволяют получить достаточно высокий выход сыра, однако следует учесть, что это обусловлено огромными объемами производимого сыра, и увеличение выхода на величину порядка одной десятой процента может дать ощутимую экономическую выгоду. Следовательно, в настоящее время имеется крайняя необходимость в получении коагулянтов, дающих увеличение (даже небольшое) выхода.
Коагулянты характеризуются своей высокой специфичностью к субстрату, которая обычно зависит от рН и температуры. В обычных процессах производства сыра рН может изменяться от начального значения 6,3 до более низких значений в пределах 4,5-5,5, причем конечное значение рН зависит от условий, используемых в процессе приготовления сыра. Некоторые коагулянты являются более восприимчивыми к изменениям рН, чем другие. Так, например, протеаза Rhizomucor pusillus является более чувствительной к изменениям рН, чем хитозин. Помимо рН, на специфичность протеазы могут влиять и другие параметры, такие как температура и содержание воды. Хорошо известно, что большинство коагулянтов обнаруживают изменение специфичности к субстрату с изменением рН, что приводит к изменению протеолитической активности на последних стадиях приготовления сыра. Также очень хорошо известно, что коагулянты отличаются по степени протеолиза казеина; и, кроме того, они могут обнаруживать различия в типах пептидов, продуцируемых в процессе протеолиза. Они являются важными факторами в процессе созревания сыра и могут влиять на свойства сыра, такие как вкус, запах и консистенция. В некоторых случаях коагулянты продуцируют нежелательные эффекты, такие как появление горького привкуса, вызываемого пептидами, или постороннего привкуса. Кроме того, изменение протеолитической специфичности может приводить к снижению выхода. Пепсин, хорошо известный компонент, присутствующий во многих препаратах коровьего химозина, является примером протеазы, которая дает более низкие выходы и худшие вкусовые эффекты, чем чистый химозин. Поэтому необходимо получить коагулянты, которые придавали бы сыру новую улучшенную консистенцию и вкус. Такие новые коагулянты будут приводить к ускоренному формированию вкусовых характеристик и профилей консистенции, ассоциированных с созреванием сыра, что может дать ощутимую экономическую выгоду.
Хорошо известно, что свободные аминокислоты имеют очень важное значение для формирования вкуса и запаха. В частности, аминокислоты лейцин, фенилаланин, метионин и валин играют важную роль в образовании компонентов типичного сырного вкуса и запаха. Превращение свободных аминокислот происходит посредством ферментации под действием микроорганизмов, которые добавляют в процессе приготовления сыра к соединениям, формирующим нужный запах и вкус, таким как метандиол, диметилдисульфид, метилпропановая кислота и метилпропаналь. Экзопептидазы играют важную роль в образовании свободных аминокислот. Однако они могут быть эффективными только в том случае, если они действуют совместно с эндопротеазой соответствующей специфичности. В приготовлении сыров могут быть использованы соответствующие комбинации экзо- и эндопептидаз, в результате чего могут быть получены сыры с новыми и улучшенными вкусовыми качествами.
Ферменты настоящего изобретения могут быть использованы для гидролиза белковых материалов животного происхождения, таких как цельное молоко, сепарированное молоко, казеин, белок молочной сыворотки или смеси казеина и белка молочной сыворотки. Такие смеси казеина и белка молочной сыворотки могут быть использованы, например, в отношениях, соответствующих отношениям, обнаруживаемым в человеческом молоке. Кроме того, смесь ферментов настоящего изобретения может быть использована для гидролиза белковых материалов растительного происхождения, таких как, например, глютен пшеницы, солодовый или несолодовый ячмень или другие злаки, используемые для производства пива, соевое молоко, его концентраты или изоляты, белок кукурузы, его концентраты или изоляты и рисовые белки.
Что касается крупномасштабного промышленного производства, некоторые применения основаны на использовании только эндопротеаз, тогда как другие применения основаны, в основном, на комбинациях эндопротеаз и экзопротеаз. Типичными примерами процессов, зависящих от использования чистых эндопротеаз, в которых протеазы настоящего изобретения могут обладать превосходной продуктивностью, являются такие процессы, как обработка белков сои, гороха или зерновых для минимизации вязкости или оптимизации пенообразования или других физико-химических свойств; использование улучшителей хлеба в хлебопекарной промышленности, которое также способствует снижению вязкости теста; использование технологических добавок в пивоварении и винодельческой промышленности, которое способствует предотвращению белковых помутнений или оптимизации выхода после экстракции зерновых; использование кормовых добавок в производстве биокормов, которое способствует усилению внутрикишечной абсорбции или модуляции микробной активности в кишечнике; использование технологических добавок в молочной промышленности, которое способствует оптимизации свертывания, пенообразования или свертывающих свойств различных компонентов молока. Кроме того, для специфических сегментов рынка молочные или соевые белки или коллаген подвергают действию протеаз в целях продуцирования так называемых гидролизатов белка. Хотя гидролизаты этих белков, в основном, применяются для изготовления продуктов детского питания и пищевых продуктов для госпитализированных больных, однако они могут быть также предназначены для индивидуумов, не нуждающихся в медицинской помощи, таких как спортсмены или люди, находящиеся на диете для похудения, и являются объектом быстро растущего спроса на рынке. Во всех этих применениях гидролизаты белков обладают такими весьма привлекательными свойствами, как пониженная аллергенность, улучшенная усвояемость в желудочно-кишечном тракте, низкий уровень химического разложения нужных аминокислот, таких как глутамин и цистеин, и, наконец, отсутствие белкового осадка в кислых напитках во время продолжительного хранения. Все эти преимущества могут быть объединены, если гидролизат представляет собой смесь ди- и трипептидов. Однако все современные коммерчески доступные гидролизаты получают с использованием комбинации нескольких эндопротеаз. Этот последний способ дает неоднородное и неполное разложение белка. Для получения нужной смеси ди- и трипептидов было бы идеальным использовать в процессе гидролиза комбинацию различных ди- и трипептидилпептидаз. К сожалению, известно лишь несколько ферментов пищевого назначения и микроорганизмов, приемлемых для использования в промышленности, не говоря уже о ферментах, фактически доступных для использования в промышленности. В соответствии с настоящим изобретением несколько широко используемых ди- и трипептидилпептидаз может быть получено экономически выгодным способом и в относительно чистом виде. Предпочтительными являются такие ди- и трипептидилпептидазы, которые обладают низкой селективностью по отношению к расщепляемому субстрату, то есть обладают предпочтением лишь к расщеплению минимального числа аминокислотных остатков. Предпочтительными являются комбинации таких ди- и трипептидилпептидаз, которые гидролизуют высокий процент природных пептидных связей. Несмотря на такую высокую активность по отношению к природным пептидным связям, полный гидролиз всех аминокислот зависит от природы ди- и трипептидилпептидаз. Предпочтительными также являются ди- и трипептидилпептидазы, которые обладают оптимальной активностью при рН 4-8 и имеют адекватную термостабильность. Адекватная термостабильность означает, что, по крайней мере, 40%, предпочтительно, по крайней мере, 60%, а более предпочтительно 70-100% начальной гидролитической активности сохраняется после нагревания фермента вместе с субстратом в течение 1 часа при 50°С.
Хотя процесс, направленный на эффективное продуцирование смесей ди- или трипептидов, зависит от доступности ферментов настоящего изобретения, однако начальным ферментом для инкубирования с белковым субстратом обычно является эндопротеаза. Предпочтительной эндопротеазой с широким спектром эндопептидазной активности, подходящей для данного случая, является, например, субтилизин (Delvolase от DSM), нейтральная металлопротеаза (нейтраза от NOVO) или термолизин (термоаза от Daiwa Kasei), применяемый в условиях, близких к нейтральным, и пепсин или аспегиллопепсин (например, Sumizyme AP от Shin Nihon, Japan), применяемый в кислых условиях. Целью этого начального расщепления является повышение растворимости, снижение вязкости и снижение термоосаждающей способности смеси воды и белка. Кроме того, эта предварительная обработка эндонуклеазой имеет важное значение для создания достаточных начальных точек для ди- и трипептидилпептидаз и приводит к ускорению процесса образования ди- или трипептида. Протеаза, используемая для устранения горечи гидролизата, может быть, но необязательно, включена в эту стадию рассматриваемого способа или позже вместе с ди- и трипептидилпептидазами.
Главной целью использования последних гидролизатов является минимизация аллергенности продукта или стимуляция усвояемости в желудочно-кишечном тракте. При продуцировании таких гидролизатов использование дипептидил- и трипептидилпептидаз является особенно важным и представляет собой эффективный способ продуцирования гидролизатов.
Другое применение в производстве пищевых продуктов и кормов, в основном, основано на комбинациях одной или более эндопротеаз с одной или более экзопротеазами. Такие комбинации эндопротеазы с экзопротеазой обычно используются в промышленности для улучшения таких параметров, как вкус и цвет конечного продукта. Основанием для этого является то, что образование вкуса и цвета зависит, главным образом, от присутствия свободных аминокислот. Свободные аминокислоты могут продуцироваться не только протеазами, такими как карбоксипептидазы и аминопептидазы, но также и пептидилдипептидазами. При комбинировании с эндопротеазами или даже с дипептидил- или трипептидилпептидазами карбоксипептидазы, аминопептидазы и пептидилдипептидазы могут продуцировать большее количество свободных аминокислот за меньший период времени. Однако во всех этих процессах неконтролируемое высвобождение аминокислот или даже небелковых компонентов должно быть предотвращено для минимизации нежелательных побочных реакций.
При этом свободные аминокислоты сами по себе могут продуцировать ряд вкусовых ощущений, и эти вкусовые ощущения являются самыми главными (горький, сладкий, кислый и “умами”), однако для различения этих вкусов необходима очень высокая концентрация аминокислот. Несмотря на высокие пороговые значения, свободные аминокислоты способны создавать значительные сенсорные эффекты при более низких концентрациях благодаря ряду усиливающих вкус механизмов. Одним из этих механизмов является комбинирование свободных аминокислот с сахарами в так называемых реакциях Майяра. По сравнению со свободными аминокислотами эти продукты Майяра с очень сложным букетом вкуса и запаха могут вырабатываться с пороговыми значениями, которые на несколько порядков величины ниже, чем те, которые вырабатываются свободными аминокислотами. Продукты Майяра могут образовываться при повышенных температурах, обычно во время кулинарной обработки, выпекания или жарки при приготовлении пищевых продуктов или кормов. Во время такой обработки развивается цвет и большой букет ароматов. В этих реакциях, на первой стадии, аминогруппы реагируют с восстанавливающими соединениями, что в конечном счете приводит к осуществлению целого каскада реакций. Аминосоединениями, преимущественно присутствующими в пищевых продуктах или в кормах, являются свободные аминокислоты, которые высвобождаются из белкового сырья различными протеазами, а необходимыми восстанавливающими соединениями являются, главным образом, восстанавливающие сахара. Отсюда следует, что для минимизации постороннего привкуса, который генерируется в процессе проведения последующих стадий нагревания, как, например, в процессе распылительной сушки или стерилизации, необходимо предотвратить нежелательное высвобождение аминокислот и сахаров во время обработки сырья. Это лишний раз указывает на то, что желательно использовать ферменты настоящего изобретения, которые имеют высокую степень чистоты и низкую себестоимость.
Помимо реакций Майяра, аминокислоты могут также подвергаться важным химическим превращениям при комнатной температуре. Более поздними типами превращения являются процессы, зависящие от ферментов, и процессы, общие для всех ферментированных пищевых продуктов, такие как созревание пива, йогурта и сыра, а также процессы созревания мяса и вина. В этих процессах ферментации свободные аминокислоты высвобождаются из сырьевого материала, используемого протеазами или протеолитическими ферментами, присутствующими в используемом сыром материале или микробной закваске. Затем в фазе созревания свободные аминокислоты под действием микробной метаболической активности превращаются в свои производные, имеющие повышенные сенсорные свойства. Так, например, L-лейцин, L-изолейцин и L-валин при пивном брожении способствуют образованию ценных сивушных спиртов, таких как амиловые спирты и изобутанол. Аналогичным образом было установлено, что сырные летучие вещества, такие как метанэтиол и диметилдисульфид, указывают на присутствие в сыре метионина, а метилпропановая кислота и метилпропаналь - на присутствие валина. И наконец, свободная аминокислота глутамат может давать эффект усиления приятного пряного вкуса в результате ее синергического действия вместе с продуктами расщепления РНК, так называемыми 5'-рибонуклеотидами. Известно, что при комбинировании с соответствующими концентрациями 5'-рибонуклеотидов, таких как 5'-ИМФ и 5'-ГМФ, порог детекции образования вкуса “умами”, генерируемого глутаматом, снижается почти на два порядка величины.
Для получения явно выраженных и точно определенных вкусовых эффектов во всех этих процедурах белковые субстраты должны быть гидролизованы с использованием комбинации эндо- и экзопротеаз, где, по крайней мере, одна эндо- или экзопротеаза, а предпочтительно как эндопротеаза, так и экзопротеаза, являются очищенными и предпочтительно селективными по отношению к конкретной серии аминокислот или, преимущественно, расщепляют предпочтительные аминокислоты. Такие предпочтительные протеазы характеризуются высокой селективностью по отношению к аминокислотным последовательностям, которые могут быть расщеплены ферментами, относящимися к категории Aspergillus, известными как “матуразы” и представляющими особую ценность.
Помимо использования в пищевой промышленности и в производстве кормов, протеазы обычно также применяются в химической и фармацевтической промышленности, а также в производстве диагностических реагентов и средств личной гигиены. В производстве средств личной гигиены протеазы используются для получения пептидов, которые добавляют в различные продукты для улучшения таких свойств, как чувствительность и глянцевитость кожи, или для защиты кожи. Кроме того, наблюдается новая тенденция к непосредственному местному применению протеаз. Аналогичным образом этот фермент может быть использован в кожевенной промышленности, и главной целью такого применения является очистка, удаление шерсти с кожи и умягчение кожи.
В химической и фармацевтической промышленности протеазы рассматриваются как важный инструмент для продуцирования ценных ингредиентов или промежуточных соединений. В этих областях промышленности протеазы используются не только благодаря их гидролитической активности, но также благодаря их способности к синтезу пептидов из природных или неприродных аминокислот. Последний из возможных вариантов применения был явно продемонстрирован возможностью синтезировать аспартам из структурных элементов, состоящих из аминокислот, с использованием эндопротеазы, такой как термолизин.
В отличие от условий, используемых при производстве пищевых продуктов и кормов, для осуществления нужного химического превращения очень важными факторами являются стерео- и региоселективность протеаз, хотя в этом случае могут потребоваться нестандартные реакционные условия. Типичными примерами применения протеаз в такой промышленности является использование эндопротеаз, аминопептидаз, а также карбоксипептидаз при продуцировании различных промежуточных соединений в целях получения лекарственных средств, таких как инсулин, антибиотики, ренин и АСЕ-ингибиторы. Обзор такого применения представлен в Industrial Biotransformations, A. Liese, K. Seelbach, C. Wandrey, Wiley-VCH; ISBH 3-527-30094-5.
Что касается нужной специфичности, стерео- и региоселективности, то важными преимуществами протеазы настоящего изобретения является отсутствие побочных эффектов и резистентности к необычным реакционным условиям, таким как высокие концентрации растворителя, и повышенная продуктивность.
С фармацевтической точки зрения, роль протеаз представлена в широком ряде работ Martindale, “The Extra Pharmacopoeia” (Pharmaceutical Press, London, UK). Кроме того, важная роль высокоспецифических протеаз в регуляции всех видов биологических процессов подтверждается тем фактом, что многие гормоны становятся активными только после процессинга, главным образом, неактивной молекулы-предшественника указанной протеазой с очень высокой специфичностью. Ингибиторы, которые являются активными по отношению к некоторым категориям таких специфических протеаз, были использованы при разработке всех видов новых лекарственных средств. Следовательно, новые и эффективные ингибиторы протеазы могут быть идентифицированы с использованием описанных здесь последовательностей.
Полное описание каждого цитируемого здесь документа вводится в настоящую заявку посредством ссылки.
ПРИМЕРЫ
Пример 1
Анализ на протеолитическую активность и специфичность
Специфичность протезы может быть исследована с использованием различных пептидных субстратов. Синтетические субстраты широко используются для детекции протеолитических ферментов в скрининге, в ферментации, в процессе выделения, для анализа ферментативной активности, для определения концентраций фермента, для исследования специфичности или для исследования взаимодействия с ингибиторами. Пептидные п-нитроанилиды, предпочтительно, используют для анализа протеазной активности, и поскольку такая активность может непрерывно прослеживаться, то это позволяет проводить кинетические измерения. Мониторинг расщепления пептидных п-нитроанилидов может быть осуществлен путем измерения увеличения адсорбции при 410 нм после высвобождения 4-нитроанилида. Паранитроанилидные субстраты обычно используются для сериновой и цистеиновой протеаз. Кроме того, используются тиоэфиры пептидов и производные 7-амино-п-метилкумариновых пептидов. Тиоэфиры пептидов являются очень чувствительными субстратами для сериновой протеазы и металлопротеаз, которые имеют относительно высокую скорость функционального цикла, поскольку тиоэфирная связь расщепляется легче, чем амидная связь. Расщепление тиоэфиров может прослеживаться с использованием тиолового реагента, такого как 4,4-дитиопиридин (324 нм) или 5,5-дитиобис-2-нитробензойной кислоты (405 нм). Такая же повышенная скорость функционального цикла обычно наблюдается при расщеплении сложноэфирных связей по сравнению со скоростью при расщеплении амидных связей. Наиболее хорошо известными субстратами для анализа эстеразной активности протеаз являются производные п-нитрофенола. Высвобождение п-нитрофенола может прослеживаться при различных длинах волн в зависимости от используемого рН; так, например, при почти нейтральном рН используется длина волны 340 нм, а при рН выше 9 мониторинг проводят примерно при 405 нм. Кроме того, мониторинг гидролиза сложных эфиров может также осуществляться путем титрования с использованием оборудования рН-стата. В случае качественного измерения эстеразной активности могут быть использованы рН-чувствительные красители.
В качестве альтернативы, пептиды могут быть присоединены к флуоресцентной уходящей группе. Протеолиз сопровождается увеличением флуоресценции при мониторинге на соответствующих длинах волн. Наиболее часто используются пептидил-2-нафтиламиды и пептидил-4-метил-7-кумариламиды. Высвобождение, например, 7-амино-4-метилкумарина измеряли при длине волны возбуждения 350 нм и при длине волны излучения 460 нм. Использование 7-амино-4-трифторметилкумарина имеет то преимущество, что в нем уходящая группа является хромогенной (поглощение на 380 нм), а также флуорогенной (возбужение на 400 нм, излучение на 505 нм). Если необходимо, чтобы на обеих сторонах расщепляемой связи присутствовала аминокислота, то может оказаться полезным введение группы, которая будет гасить флуоресценцию. Общие свойства таких субстратов заключаются в том, что в них флуоресцентная донорная группа отделяется от акцепторной группы пептидной последовательностью, которая действует как гаситель флуоресценции. Расщепление пептидной связи между гасящей группой и флуорофором может приводить к значительному усилению флуоресценции. Было описано несколько пар “донор-акцептор”, включая о-аминобензойную кислоту (Abz), используемую в качестве донора, и 2,4-динитрофенил (Dnp), используемый в качестве акцептора, или 5-[(2'-аминоэтил)амино]нафталинсульфоновую кислоту (EDANS), используемую в качестве донора, и 4-[[4'-(диметиламино]фенил]азо]бензойную кислоту (DABCYL), используемую в качестве акцептора. Abz/EDDnp представляет собой наиболее подходящую пару “донор-акцептор”, поскольку после полного гидролиза интенсивность флуоресценции увеличивается в 7-100 раз, а спектр поглощения EDDnp не изменяется с изменением рН. Кроме того, пептидная последовательность может содержать до 10 остатков, но при этом теряет свое гасящее действие. По мере увеличения размера соединяющих пептидов положение данной расщепляемой связи может стать менее специфическим. Поэтому помимо установления события протеолиза может оказаться необходимым проведение дополнительного анализа этих продуктов. Это может быть осуществлено с помощью анализа и выделения продуцированных пептидов с помощью ВЭЖХ и определения аминокислотной последовательности этих фрагментов. Кроме того, пептидный состав гидролизата может быть непосредственно проанализирован комбинированным методом с использованием ВЭЖХ/масс-спектроскопии.
Помимо использования пептидов определенной последовательности, для исследования специфичности протеазы могут быть также использованы библиотеки синтетических пептидов. Пептиды были синтезированы с помощью твердофазного синтеза рандомизированным или полурандомизированным способом. Так, например, Meldal et al. (PNAS USA 91, 3314, 1994) сообщают о получении семейства субстратов для протеаз путем присоединения Н-Lys(Abz)-смолы, удлинения смолы пептидами длиной до 6 аминокислот и, наконец, присоединения Tyr(NO2) к этим пептидам. Каждая гранула смолы имеет уникальную последовательность, и после обработки протеазами наиболее чувствительные гранулы становятся флуоресцентными по мере высвобождения Tyr(NО2)-содержащего пептида. Анализ последовательности этих пептидов на их чувствительность будет давать информацию о специфичности протеазы.
Протеазная активность обычно выражается в единицах. Вообще говоря, “международная стандартная единица” (МЕ) определяется как количество фермента, которое в определенных условиях переносит один микромоль субстрата в минуту. В частности, что касается протеаз, то МЕ должна соответствовать гидролизу одного микромоля пептидной связи в минуту. Однако, в случае протеазы, отклонения в определении ее единиц от международного стандарта является скорее правилом, чем исключением. В случае пептидов-моделей, которые специфически расщепляются по одной связи, вычисление МЕ является узко направленной процедурой, а для белковых субстратов, где расщепление данной протеазой может осуществляться в различных положениях и в различной степени, может быть принято другое определение, существенно отличающееся от стандартного. Помимо используемого определения единиц активности, любой эксперимент по гидролизу требует адекватного описания условий, при которых проводят измерения этих единиц. Такие условия включают, например, концентрацию субстрата, отношение “фермент-субстрат”, рН и температуру. Типичные анализы для определения специфической активности протеаз включают использование белкового субстрата, такого как, например, денатурированный гемоглобин, инсулин или казеин. Полипептидный субстрат расщепляется протеазой при фиксированных условиях в течение строго определенного промежутка времени. Нерасщепленные и крупные полипептиды осаждают с использованием ТСА и ТСА-растворимый продукт оценивают путем измерения оптической плотности при 220 или 280 нм либо путем титрования растворимых пептидов фолиновым реагентом, нингидрином, фтор-2,4-динитробензолом/данзилхлоридом, методом с использованием TNBS или флуоресцеина. Вместо мечения продукта после гидролиза могут быть также использованы полипептидные субстраты, которые были уже помечены специфическими красителями или флуорофорами, такими как, например, флуоресцеин. Помимо стандартных методов, аминокислотный анализ может быть осуществлен с использованием стандартных лабораторных анализаторов. Для оценки распределения по размерам пептидов, генерированных протеазой, могут быть осуществлены эксперименты с использованием гель-хроматографии. Помимо этого, для лучшего выделения типов пептидов, генерируемых протеазой, может быть использована ВЭЖХ с применением обращенно-фазовой техники. Ход гидролиза белковых субстратов обычно оценивают по степени гидролиза или DH. В случае использования рН-стата для слежения за ходом гидролиза DН может определяться исходя из поглощения основания в процессе гидролиза (Enzymatic Hydrolysis of Food Protein, J. Adler-Nissen, 1986, Elsevier Applied Science Publishers LTD). DН означает различные полезные функциональные свойства гидролизата, такие как растворимость, эмульгирующая способность, пенообразование и стабильность к пенообразованию, увеличение в объеме при взбивании, органолептические качества. Кроме того, важным параметром пищевых гидролизатов является вкус. Горечь может оказаться главной проблемой при использовании гидролизатов белка. Завершение реакции гидролиза может быть осуществлено путем изменения рН, термоинактивации и применения денатурирующих агентов, таких как ДСН, ацетонитрил и т.п.
Полипептиды, представленные в таблице 1, были экспрессированы и, по крайней мере, частично очищены стандартными методами, известными специалистам. Эти полипептиды были проанализированы, по крайней мере, одним из способов, описанных выше, и было обнаружено, что они обладают активностями, перечисленными в таблице 1.
Пример 2
Прямое определение отношения kcat/Кm для субстратов протеазы
Для мониторинга ферментативной активности в процессе очистки, для определения концентрации субстрата, для определения констант ингибирования или для исследования специфичности к субстрату были использованы синтетические субстраты. Определение отношения kcat/Кm позволяет измерить специфичность к субстрату. Оно также позволяет проводить сравнение специфичностей различных субстратов для одного и того же фермента или сравнение скоростей гидролиза различными ферментами, расщепляющими один и тот же субстрат. Это отношение представляет собой константу скорости, выражаемую в единицах второго порядка, и определяется как 1/(концентрация · время). Субстраты, имеющие отношение kcat/Кm в пределах 10,5-10,6 М-1·сек-1, считаются очень хорошими субстратами, то есть обладающими хорошей аффинностью и быстрым функциональным циклом. Однако некоторые субстраты могут быть очень специфичными, то есть иметь значения kcat/Кm, составляющие порядка 10,4 М-1·сек-1.
Отношение kcat/Кm может быть вычислено после определения отдельных параметров. В этом случае Кm и Vm могут быть определены из различных линейных кривых (построенных, например, методом Ханса или Корниша-Боудена) или методом нелинейной регрессии. Известно, что Vm=kcat·Et (где Et означает конечную концентрацию активного фермента), а следовательно, kcat=Vm/Et. Определение отношения kcat/Кm описанным выше методом может быть затруднено, если происходит ингибирование продукта или субстрата либо если субстрат осаждается при высокой концентрации. Однако можно получить точное значение отношения kcat/Кm в условиях реакции первого порядка, то есть когда концентрация субстрата значительного ниже расчетного Кm. В этих условиях уравнение Михаэлиса-Ментена: v=(Vm·S)/(Кm+S) имеет вид:
v=(Vm·S)/Кm, если S << Кm
или v=(Vm/Кm)·S= kobs·S=-dS/dt,
и после интегрирования получают: lnS=-kobs·t+lnSо, где Sо означает исходную концентрацию субстрата, а S означает концентрацию субстрата в данный момент времени. Скорость пропорциональна концентрации субстрата. Другими словами, гидролиз субстрата протекает в соответствии с реакцией первого порядка, где kobs - константа скорости первого порядка: kobs=Vm/Кm=(kcat·Et)/Кm, если Vm=kcat·Et. Непрерывная регистрация гидролиза субстрата дает возможность для графического определения kobs по кривой зависимости lnS от времени. Отношение kcat/Кm просто определяют исходя из kobs, при условии, что известна концентрация активного фермента:
kcat/Кm=kobs/Et.
Метод анализа: использование начальной концентрации исходного субстрата, составляющей намного ниже расчетного Кm, и низкой концентрации фермента позволяет регистрировать ход гидролиза субстрата. Исходя из этого, для генерирования продукта получают кривую первого порядка. После полного гидролиза субстрата оптическая плотность (или единицы флуоресценции) продукта может быть точно определена исходя из Sо, ибо Pt=So·kobs определяется из кривой зависимости lnS от времени, или альтернативно, она может быть определена с использованием программы подбора кривой по точкам (Enzfitter, SigmaPlot...).
NВ: Следует помнить, что необходимо вычислить концентрацию субстрата для данного момента времени исходя из концентрации продукта (S=Sо-Р), поскольку зависимость Р от времени не дает правильного значения kobs (dP/dt=kobs·S не интегрируется аналогичным образом).
Альтернативно, можно измерить последовательные величины t1/2 (полупериод) исходя из кривой продуцирования продукта в процессе реакции первого порядка:
t1/2=ln2/kobs=0,693/kobs, а kobs=0,693/t1/2
Использование этого метода позволяет контролировать истинное затухание первого порядка (идентичные величины для последовательных значений t1/2).
Пример 3
Инактивация генов протеазы в Aspergillus
Наиболее подходящим способом инактивации генов протеазы в геноме Aspergillus является техника генной замены (так называемая “одностадийная дизрупция гена”). Принципы этой техники описаны Rothstein R.J., Meth. Enzymol. 101, p.202, 1983. В основном, эта техника основана на гомологичной рекомбинации трансформированных ДНК-фрагментов с геномной ДНК клеток грибов. Посредством двойного кроссинговера инактивируемый ген заменяется (частично) ДНК-фрагментом, которым трансформируется клетка. Предпочтительно трансформирующий ДНК-фрагмент содержит селективный маркерный ген Aspergillus niger. Модификацию ДНК и генерирование инактивирующей конструкции осуществляют, в основном, с использованием общих методов молекулярной биологии. Сначала геномную ДНК выделяют из штамма Aspergillus niger, который затем используют для инактивации гена протеазы. Геномная ДНК A.niger может быть выделена любыми описанными методами, например, методом, описанным Graaff et al. (1988) Curr. Genet. 13, 315-321, и методами, хорошо известными специалистам. Эту геномную ДНК используют в качестве матрицы для амплификации фланкирующих областей гена протеазы посредством полимеразной цепной реакции (ПЦР; Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual, 2nd edition, Cold Spring Harbor Laboratory Press, New York). Термин “фланкирующие области” означает некодирующие области, расположенные выше и ниже инактивируемого гена протеазы. Предпочтительно, чтобы каждая фланкирующая область имела длину более 1,0 т.п.н.
Для осуществления ПЦР-амплификации каждой фланкирующей области в качестве праймеров использовали два одноцепочечных ДНК-олигонуклеотида. Для 5'-фланкирующей области использовали один праймер гомологичной ДНК-последовательности, расположенной выше инициирующего кодона кодирующей последовательности гена протеазы. Предпочтительно, чтобы гомологичная область была локализована на участке, расположенном более чем на 1,0 т.п.н. выше сайта инициации трансляции. Второй праймер был гомологичен комплементарной и инвертированной ДНК-последовательности, расположенной непосредственно выше кодирующей последовательности гена протеазы.
Для 3'-фланкирующей области использовали один праймер, который был гомологичен ДНК-последовательности, расположенной непосредственно ниже кодирующей последовательности гена протеазы. Второй праймер был гомологичен комплементарной и инвертированной ДНК-последовательности, локализованной, предпочтительно, на участке, расположенном более чем на 1,0 т.п.н. ниже кодирующей последовательности гена протеазы.
ДНК-последовательность, включенная во все праймеры и гомологичная геному A.niger, должна иметь длину минимум 15 нуклеотидов, а предпочтительно более чем 18 нуклеотидов. В основном, все праймеры должны содержать ДНК-последовательность, кодирующую сайт распознавания для подходящих рестриктирующих ферментов, расположенный выше последовательности, гомологичной геному A.niger. Эти дополнительные сайты распознавания облегчают процесс клонирования.
Праймеры и геномную ДНК A.niger использовали в ПЦР-реакции в условиях, известных специалистам. Температура отжига праймеров может быть вычислена для части ДНК-последовательности, которая гомологична геному A.niger. Оба фрагмента, содержащие 5'-фланкирующую область и 3'-фланкирующую область, клонировали в вектор, который может быть реплицирован в E.coli с использованием общих методов молекулярной биологии. Затем ген, который может быть использован в качестве селективного маркера в Aspergillus niger, клонировали между двумя фланкирующими областями. Как правило, маркерный ген находится под контролем промотора, который регулирует экспрессию в A.niger, а предпочтительно эндогенного промотора A.niger. Ориентация вставки маркерного гена, предпочтительно, совпадает с ориентацией исходного гена протеазы. Конечный инактивирующий фрагмент содержит 5'-фланкирующую область, селективный маркерный ген, предпочтительно, находящийся под контролем эндогенного промотора A.niger, и 3'-фланкирующую область, причем все они имеют одно и то же направление и одну и ту же ориентацию. ДНК конечной конструкции клонируют в вектор, который может реплицироваться в E.coli.
Инактивирующую конструкцию расщепляют подходящими рестриктазами для удаления векторных последовательностей E.coli и инактивирующий фрагмент выделяют стандартными методами (Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual, 2nd edition, Cold Spring Harbor Laboratory Press, New York). И наконец, Aspergillus niger трансформируют инактивирующим фрагментом с использованием метода, описанного в литературе, например, метода, описанного Kusters-van Someren et al. (1991) Curr. Genet. 20, 293-299. Трансформированные клетки отбирают путем высевания смеси для трансформации на чашки с агаром, которые являются селективными в отношении роста штаммов Aspergillus niger, экспрессирующих маркерный ген. После очистки трансформированных штаммов Aspergillus путем высевания реплик ряд репрезентативных штаммов анализируют методом Саузерн-блоттинга с использованием стандартной методики (Sambrook et al. (1989) Molecular Cloning: A Laboratory Manual, 2nd edition, Cold Spring Harbor Laboratory Press, New York). Для этого геномную ДНК мицелия трансформированных штаммов выделяют и расщепляют подходящими рестриктазами. Рестрикционные фрагменты разделяют с использованием электрофореза на агарозном геле, подвергают блоттингу на нитроцеллюлозных мембранах и зондируют меченым фрагментом маркерного гена. Гибридизацию и промывку осуществляют в жестких условиях. Подходящими считаются штаммы, которые содержат меченые рестрикционные фрагменты нужной длины. Этим методом могут быть отобраны штаммы A.niger, содержащие нужный инактивированный ген протеазы.
Пример 4
Выделение протеаз с помощью ионообменной хроматографии
Небольшие количества протеазы, кодируемой описанной здесь нуклеотидной последовательностью, получали путем конструирования экспрессирующей плазмиды, содержащей релевантную ДНК-последовательность; трансформирования штамма A.niger плазмидой; и культивирования штамма A.niger в подходящей среде. После сбора бульона, не содержащего контаминирующих клеток, нужная протеаза может быть очищена.
Для выделения протеазы, кодируемой рассматриваемой нуклеотидной последовательностью, по существу в чистой форме могут быть использованы различные стратегии. Все эти стратегии хорошо описаны в соответствующей научной литературе (см., например, руководство “Protein Purification Handbook, 18-1132-29, Edition AA, Amersham Pharmacia Biotech, Uppsala, Sweden). В этом руководстве также описана процедура, подходящая для очистки протеаз из комплексных смесей. Важно то, что подходящий анализ является доступным и селективным в отношении свойств рассматриваемых ферментов. Для протеаз обычно используется хромогенный, синтетический пептидный субстрат, описанный в примере 1. Такие пептидные субстраты могут быть селективными для эндопротеаз, карбоксипептидаз, аминопептидаз или омегапептидаз. В примере 11 описана селективность для специфической трипептидилпептидазы. Протеазы с нужной специфичностью могут быть отобраны путем выбора соответствующих аминокислотных остатков в релевантном синтетическом пептиде.
Сначала необходимо определить, секретируется ли данная протеаза в среду, так как в зависимости от экспрессионной системы, выбранной для продуцирования протеазы, она может секретироваться из клетки либо содержаться в клетке. Если протеаза секретируется в ферментационную среду, то продуцирующие клетки или фрагменты этих клеток необходимо удалить путем центрифугирования или фильтрации, а полученную прозрачную или осветленную среду можно использовать как исходный материал для последующей очистки. В случаях если рассматриваемая протеаза не секретируется, то продуцирующие клетки необходимо подвергнуть дизрупции для выделения протеазы. В таких случаях собранную клеточную массу тщательно измельчают с использованием абразива, размалывают в шаровой мельнице, подвергают ультразвуковой обработке и обработке на прессе Френча или в гомогенизаторе Мэнтона-Голина, а затем фильтруют или центрифугируют. В случае если данная протеаза является гидрофобной или мембрано-ассоциированной, то перед стадией фильтрования или центрифугирования может оказаться необходимым добавление неионогенного детергента для солюбилизации протеазы.
Для получения неизвестных протеаз, по существу в чистом виде, после стадии осветления может быть осуществлена трехфазная стратегия очистки. Во всех или в некоторых из этих трех фаз может оказаться необходимым добавление детергента.
В первой фазе, или в фазе иммобилизации, нужную протеазу выделяют, частично очищают и концентрируют. В последующей промежуточной фазе очистки большую часть всего объема примесей удаляют, а в конечной фазе очистки удаляют следовые количества оставшихся примесей более крупных близкородственных веществ и фермент растворяют в нужном буфере. Специалист может в зависимости от природы и физических свойств исследуемой протеазы оптимизировать эти три фазы, используя слегка модифицированные варианты различных связывающихся с белком материалов и изменяя до определенной степени условия. Однако во всех случаях необходимо проводить селективный аналитический тест, поскольку это дает возможность осуществлять непрерывный мониторинг возрастающей активности очищенного протеолитического фермента. Аналитические тесты, подходящие для этих целей, предусматривают использование хромогенных пептидных субстратов, упомянутых выше.
В первой иммобилизирующей фазе очистки предпочтительно использовать сильную ионообменную смолу анионного типа для осветления и обессоливания фермент-содержащей среды. Для гарантии связывания нужной протеолитической активности со смолой проводят тест при трех или четырех различных значениях рН среды и смолы в условиях низкой проводимости. В этих тестах смолу всегда уравновешивают буфером с такими же значениями рН и проводимости, как и в ферментсодержащей среде. Затем среду наносят на колонку в условиях рН, которые, как было показано, позволяют осуществлять адекватное связывание протеазы со смолой, то есть так, чтобы ни одна из нужных ферментативных активностей не была обнаружена в проходящей среде. Затем нужную ферментативную активность элюируют из ионообменной смолы с использованием непрерывного солевого градиента, где градиентное элюирование начинают с уравновешивающего смолу буфера и заканчивают тем же самым буфером, в который добавляют 1 моль/литр NаСl. В соответствии с данным анализом элюированные фракции, содержащие нужную активность, собирают, а затем приготавливают для дополнительной стадии очистки. Проведение этой дополнительной стадии очистки зависит от чистоты нужного фермента в собранной фракции: если он почти чистый, то необходимо проводить дополнительную адекватную стадию гель-фильтрации; а если не является достаточно чистым, то необходимо проводить хроматографию на гидрофобной смоле, а затем стадию гель-фильтрации.
Хроматографию на гидрофобной смоле осуществляли сначала путем увеличения содержания соли в собранной фракции, полученной с ионообменной смолы, до 4 моль/литр NаСl, с последующим удалением любого образовавшегося осадка. Если полученная осветленная фракция не содержит нужной активности, то эта активность, очевидно, присутствует в осадке и не может быть выделена в по существу чистой форме. Если в данном анализе полученная прозрачная фракция продолжает обладать нужной активностью, то эту жидкость наносят на фенилсефарозную смолу (Pharmacia), уравновешенную в буфере с высокой концентрацией соли, имеющем идентичные рН и проводимость. Если фракция с нужной ферментативной активностью связывается с фенилсефарозной смолой, то эту фракцию элюируют непрерывным градиентом со снижением содержания соли, а затем промывкой, не содержащей соли, и если необходимо, хаотропным агентом. Как было описано ранее, фракции, полученные градиентным элюированием фракции и проявляющие активность в анализе, собирали и в конце подвергали гель-фильтрации. Если фракция с нужной ферментативной активностью не связывается с фенилсефарозной смолой, то в этом случае присутствует много примесей, а поэтому для получения нужной протеолитической активности, присутствующей в пустом объеме колонки, требуется проведение лишь дополнительных стадий ультрафильтрации в целях получения активности в более концентрированной форме, которую затем наносят на колонку для гель-фильтрации. Колоночная гель-хроматография позволяет не только удалять следовые примеси, но также вводить фермент в буфер, который необходим для последующего использования.
Хотя этот метод является, в основном, подходящим для выделения и очистки протеазы настоящего изобретения, однако в примере 4 описан более специфический способ выделения. В этом примере описано выделение протеазы Aspergillus с использованием иммобилизованного бацитрацина, пептида-антибиотика, который, как известно, селективно взаимодействует с протеазами различных типов.
Пример 5
Выделение протеаз с помощью аффинной хроматографии
Альтернативным методом очистки небольших количеств протеазы является аффинная хроматография. Для получения протеазы в очищенной форме 100 миллилитров культуры выращивали в хорошо проветриваемой шейкерной колбе. После центрифугирования для удаления любого нерастворимого вещества супернатант наносили на 40-миллилитровую колонку с бацитрацином-сефарозой, уравновешенную 0,05 моль/литр ацетата натрия, рН 5,0. Протеазы, связанные с колонкой, элюировали ацетатным буфером, в который было добавлено 1 моль/литр NаСl и 10% (об./об.) изопропанола (J. Appl. Biochem. 1983, pp.420-428). Активные фракции собирали, диализовали против дистиллированной воды и наносили на 20-миллилитровую колонку с бацитрацином-сефарозой, после чего снова уравновешивали ацетатным буфером. Как и ранее, элюирование осуществляли с использованием ацетатного буфера, в который были добавлены NаСl и изопропанол. Активные фракции, то есть фракции, проявляющие нужные активности, собирали, диализовали против 5 миллимоль/литр ацетатного буфера, рН 5,0, а затем концентрировали путем ультрафильтрации на мембране Amicon РМ-10. Для получения по существу чистой протеазы концентрированную жидкость хроматографировали на колонке с супердексом 75, уравновешенной 0,05 моль/литр натрийацетатного буфера, рН 5,0, в который было добавлено 0,5 моль/литр NаСl. Последующие эксперименты, проводимые с использованием очищенного фермента путем электрофореза на ПАГ, могут подтвердить, совпадает ли его молекулярная масса с молекулярной массой, предсказанной исходя из имеющихся данных о последовательности. Конечное подтверждение может быть получено путем проведения анализа неполной, N-концевой аминокислотной последовательности.
Пример 6
Свойства новой цистеиновой протеазы, происходящей от A.niger
В этом примере ген Aspergillus № 28 клонировали и сверхэкспрессировали в A.niger, как описано выше. Полученный фермент очищали в соответствии с процедурами, описанными в примере 4, и использовали для разрушения трипсин-ингибирующей активности соевых бобов в различных условиях. В качестве эталонных материалов использовали папаин и бромелаин. Бромелаин был получен от фирмы Sigma, а папаин был получен от фирмы DSM Food Specialties Business Unit Beverage Ingredients, PO Box 1, 2600 MA Delft, the Netherlands.
Ингибирование трипсина измеряли методом, описанным Kakade M.L., Rackis, J.J., McGhee, J.E. & Puski, G. (1974): J. Cereal Chemistry 51:376-382. Для измерения активности трипсина осуществляли расщепление субстрата N-бензоил-L-аргинин-п-нитроанилина на N-бензоил-L-аргинин и п-нитроанилин. Трипсин закупали у British Drug Houses Ltd и получали из коровьей поджелудочной железы, содержащей более чем 0,54 единиц Anson на грамм продукта. Соевый ингибитор Куница получали от Sigma.
Этот ингибитор трипсина предварительно инкубировали при концентрации 2 мг/мл с вышеупомянутыми ферментами сериновыми протеазами при рН 3 в 50 мМ Nа-ацетатного буфера с последующей оценкой ингибирования трипсина. Ферменты добавляли в отношении ферментный белок:ингибитор трипсина, 1:100 (мас./мас.). В качестве негативного контроля для ферментов служил альбумин. Остаточную трипсиновую активность измеряли после инкубирования в течение 3 часов при 37оС. Результаты представлены в таблице 2.
Действие различных цистеиновых протеаз
на ферментативную инактивацию ингибитора трипсина Куница, выделенного из соевых бобов
Эксперименты повторяли в присутствии пепсина в процессе предварительного инкубирования цистеиновых протеаз с ингибитором трипсина. Пепсин добавляли в конечной концентрации 1,3 мг/мл. Результаты представлены в столбце 3.
Другие серии экспериментов осуществляли для оценки термостабильности. Цистеиновые протеазы инкубировали при 75 и 90°С в течение 5 минут, а затем добавляли ферменты для предварительного инкубирования с ингибиторами трипсина. Результаты представлены столбцами 4 и 5.
Эти результаты ясно продемонстрировали превосходную активность новых цистеиновых протеаз Aspergillus niger по сравнению с уже известными цистеиновыми протеазами, используемыми для инактивации ингибиторов трипсина в корме для животных.
Пример 7
Экзопептидазы, стимулирующие созревание сыра и формирование сырного вкуса
Аминопетидазы, кодируемые генами № 20 и 54 (см. таблицу 1), были сверхэкспрессированы в A.niger методами, описанными выше. Очистку этих ферментов осуществляли в соответствии с процедурами, описанными в примере 4. Активность образцов очищенного фермента определяли при рН 7,2 в водном фосфатном буфере (50 мМ), содержащем паранитроанилидное производное ряда гидрофобных аминокислот (3 мМ) в качестве субстрата. Превращение субстрата аминопептидазой оценивали путем мониторинга изменения оптической плотности при 400 нм в результате превращения субстрата, при этом в качестве контроля использовали раствор, не содержащий фермента. Активность (А) вычисляли как изменение OD в минуту и выражали, например, как единицы Phe-АР, Leu-АР или Val-АР в зависимости от используемого субстрата. Нормальное молоко для производства сыра инокулировали заквасочной культурой Delvo-tecтм DX 31 (DSM Food Specialities Delft, The Netherlands) для получения сыра типа Гауда, а свертывание осуществляли с использованием средних доз коагулянта (50 IMCU на литр сыропригодного молока). Кроме того, к двум экспериментальным сырам добавляли 25 Phe-единиц каждой экзопротеазы, тогда как контроль не содержал ни одной экзопротеазы. Параметры, используемые при получении обоих сыров, соответствовали параметрам, применяемым в производстве полутвердых сыров. При этом в образовании вкуса и аромата между экспериментальными сырами и контрольным сыром наблюдались различия, а именно экспериментальные сыры приобретали большинство из своих органолептических свойств через три (3) недели, тогда как контрольный сыр приобретал аналогичные свойства через шесть (6) недель. Было показано, что у экспериментальных сыров уровень свободных аминокислот через три недели был в два раза выше, а через шесть недель созревания сыра он был снова сравнимым. Анализ аминокислот осуществляли методом Picotag, Waters (Milford MA, USA).
Эти данные позволяют предположить, что продукт был готов для продажи на три недели раньше и при этом по своему качеству он не уступал качеству нормального сыра. Органолептичесие свойства экспериментальных сыров отличались от органолептических свойств контрольного сыра тем, что в противоположность контрольному сыру у экспериментальных сыров в присутствии аминопептидазы пропадал слабо выраженный слегка горьковатый привкус. Было обнаружено, что консистенция этих сыров также была несколько более нежной.
Пример 8
Новая специфичность протеазы, кодируемой геном 55
Как объяснялось выше, некоторые белки могут быть резистентны к ферментативному гидролизу, что обусловлено конкретным аминокислотным составом или наличием специфических третичных структур. В таких случаях количество пептидов, которые присутствуют в этих резистентных к протеазе белках и которые могут быть солюбилизированы, может быть резко увеличено с использованием протеаз, обладающих новыми специфичностями.
Бета-казеин представляет собой белок, имеющий очень ограниченную третичную структуру, но исключительно высокий уровень пролиновых остатков. Многие протеазы не могут расщеплять пролинсодержащие последовательности, а поэтому гидролиз бета-казеина известными протеазами дает гидролизат, относительно обогащенный крупными резистентными к протеазе пептидами. Эти резистентные пептиды могут быть ответственны за ряд нежелательных свойств гидролизата. Так, например, хорошо известно, что эти более крупные пептиды обладают относительно сильным действием с точки зрения аллергенности и имеют горький привкус. Более того, эти пептиды резистентны к последующему разложению на свободные аминокислоты, а поэтому в некоторых процессах присутствие этих крупных резистентных к протеазам пептидов равносильно потере выхода. Следовательно, доступность и использование протеаз, способных расщеплять резистентные к протеазе части белков, будет значительно способствовать техническому прогрессу и получению экономической выгоды.
Бета-казеин представляет собой одну из главных казеиновых фракций коровьего молока. Этот белок был хорошо охарактеризован с точки зрения аминокислотной последовательности и является коммерчески доступным почти в чистой форме. По существу, бета-казеин представляет собой превосходный тест-субстрат, используемый для исследования взаимосвязи между сайтами расщепления ферментом и длиной различных пептидов, образованных в процессе ферментативного гидролиза.
В этом примере продемонстрировано, что, несмотря на широкий спектр расщепления эндопротеазы субтилизина, добавление в высокой степени специфичного фермента, такого как пролилэндопептидаза, кодируемого геном 55 (см. таблицу 1), оказывает огромное воздействие на размер образуемых фрагментов бета-казеина.
Бета-казеин коровьего молока (лиофилизованного, по существу, не содержащего соли сухого молока), содержащего, по крайней мере, 90% бета-казеина, был закуплен у Sigma. Субтилизин, выделенный из B.licheniformis (Delvolase ®, 560000 DU на грамм), был закуплен у DSM Food Specialities (Seclin, France). Пролин-специфическую эндопептидазу, кодируемую геном 55, сверхэкспрессировали в A.niger и очищали с использованием процедур, описанных в примере 4.
Порошок бета-казеина растворяли в концентрации 10 мас.% вместе с 0,1 мас.% порошка Delvolaseтм в 0,1 моль/литр фосфатного буфера рН 7,0. После инкубирования в течение 24 часов при 45°С в водяной бане со встряхиванием реакцию прекращали путем нагревания раствора в течение 15 минут при 90°С. К одной половине раствора (1 мл, содержащего 100 миллиграммов бета-казеина) добавляли 100 микролитров пролин-специфической протеазы и реакцию продолжали еще 24 часа при 45°С. После еще одного теплового шока при 90°С образцы материала бета-казеина, обработанного Delvolaseтм и Delvolaseтм + пролин-специфической протеазой, анализировали с использованием ЖХ/МС-оборудования для исследования точного распределения пептидов по размеру в двух образцах.
ЖХ/МС-анализ
Характеризацию гидролизатов ферментного белка, продуцированного с использованием смеси ферментов настоящего изобретения, осуществляли с помощью ВЭЖХ с применением масс-спектрометра с ионной ловушкой (ThermoquestTM, Breda, The Netherlands), подсоединенного к насосу Р4000 (ThermoquestTM, Breda, The Netherlands). Полученные пептиды разделяли на колонке PEPMAP C18 300A (MIC-15-03-C18-PM, LC Packings, Amsterdam, The Netherlands), элюируя градиентом 0,1% муравьиной кислоты в воде Milli Q (Millipore, Bedford, MA, USA; раствор А) и 0,1% муравьиной кислоты в ацетонитриле (раствор В). Это градиентное элюирование начинали со 100% раствора А, с увеличением до 70% раствора В за 45 минут, и поддерживали при таком соотношении еще в течение 5 минут. Используемый объем инъекции составлял 50 микролитров, скорость потока составляла 50 микролитров в минуту, а температура колонки поддерживалась при 30°С. Концентрация белка инъецированного образца составляла приблизительно 50 микрограммов/миллилитр.
Подробная информация об отдельных пептидах была получена с использованием алгоритма “зависимости от сканирования” MS/MS, который представляет собой характеристический алгоритм для масс-спектрометра с ионной ловушкой. После полного сканирующего анализа проводили сканирующий анализ путем масштабирования для определения заряда наиболее сильного иона в пределах всей сканируемой массы. Последующий MS/MS-анализ более позднего иона давал информацию о неполной пептидной последовательности, которая может быть использована для поиска в базе данных с использованием приложения SEQUEST от Xcalibur Bioworks (ThermoquestTM, Breda, The Netherlands). Используемые банки данных были взяты из банка данных OWL.fasta, доступного в NCBI (Национальный центр биотехнологической информации) и содержащего белки, представляющие интерес для используемого приложения.
С использованием этой техники как способа сканирования в качестве пептидов, подходящих для последующего анализа путем МS-секвенирования, рассматривались только пептиды, имеющие массу в пределах приблизительно 400-2000 Да.
Ангиотензин (М=1295,6) использовали для настройки оптимальной чувствительности в режиме МS и для оптимальной фрагментации в режиме МS/МS, при осуществлении непрерывной инфузии 60 мкг/мл, что приводило, главным образом, к получению двухзарядного и трехзарядного ионов в режиме МS и оптимальной энергии столкновения примерно 35% в режиме МS/МS.
В образце, расщепляемом только Delvolase, LC/МS/МS-анализ позволил идентифицировать 40 пептидов, охватывающих различные части молекулы бета-казеина. Все вместе эти пептиды составляют 79% от общей последовательности бета-казеина. Было установлено, что пептиды длиной в пределах от 2 до 23 аминокислотных остатков имеют отличающееся время удерживания на колонке С18. Было обнаружено, что в целом <15% пептидов имели размер менее 6 аминокислот. В образце, расщепленном DelvolaseТМ и пролин-специфической протеазой, также образовывалось большое количество пептидов, идентифицируемых в бета-казеине. Все вместе эти пептиды охватывали >50% всей последовательности белка бета-казеина. В указанном образце распределение пептидов по размерам было достаточно гомогенным, поскольку эти пептиды имели длину только в пределах от 2 до 6 остатков. Полученные результаты показали, что гидролизат, полученный с использованием пролин-специфической протеазы, содержал большую фракцию пептидов, имеющих 2-6 аминокислот, что указывало на явно положительный эффект совместного инкубирования с эндопротеазой, обладающей необычной специфичностью. Из этих экспериментов также очевидно, что эндопротеаза, кодируемая геном 55, представляет собой эндопротеазу, которая расщепляет пептидную цепь у карбоксиконца пролинового остатка.
Пример 9
Селективное высвобождение специфических аминокислот для стимуляции формирования вкуса
Свободные аминокислоты, такие как лейцин и фенилаланин, не только участвуют в реакциях Майяра, но также являются предшественниками, образующими нужные запахи при ферментации различных пищевых продуктов. Для стимуляции образования таких запахов в процессе ферментации пищевых продуктов или в процессе нагревания, жарки или выпекания пищевых продуктов может оказаться предпочтительным вводить в эти продукты гидролизат белка, который содержит относительно высокие уровни указанных специфических аминокислот в свободной форме. В данном примере авторы описывают получение дрожжевых экстрактов, селективно обогащенных лейцином и фенилаланином. Такое обогащение достигается путем комбинирования эндопротеазы, обладающей предпочтением к расщеплению выбранного ряда аминокислотных остатков, с экзопротеазой, которая обладает предпочтением к высвобождению аналогичного ряда аминокислотных остатков. Такое предпочтение указанной эндопротеазы должно соответствовать предпочтению используемой экзопротеазы. Так, например, авторами было установлено, что аминопептидазы, кодируемые генами 20 и 54 (см. таблицу 1), обладают определенным предпочтением к высвобождению лейциновых и фенилаланиновых остатков, которое совпадает с предпочтением термолизина в его гидролизующем действии. Карбоксипептидазы, кодируемые генами 23 и 24, обладают предпочтением к высвобождению аргининовых и лизиновых остатков, которое совпадает с предпочтением трипсина в его гидролизующем действии. Карбоксипептидаза, кодируемая геном 5, обладает в высокой степени необычным предпочтением к высвобождению глицина и может быть объединена с некоторыми эндопротеазами, присутствующими в папаине. Карбоксипептидаза, кодируемая геном 51, способна удалять остатки глутамата, и эта способность совпадает со способностью глутамат-специфичной протеазы, кодируемой геном 43.
Известно, что эндопротеаза термолизин (имеющаяся в продаже под названием термоаза) С180, поставляемая фирмой Daiwa Kasei KK (Osaka, Japan), расщепляет пептидные связи со стороны аминоконца объемных гидрофобных аминокислот, таких как Leu и Phe. Для высвобождения обработанных таким образом аминокислот из уже образованных пептидов авторами были использованы аминопептидазы, кодируемые генами №№ 20 и 54 (см. таблицу 1). Эти гены были сверхэкспрессированы в A.niger описанными ранее методами, а очистку указанных ферментов осуществляли в соответствии с процедурами, описанными в примере 4.
Очевидно, что для высвобождения лейцина и фенилаланина и, насколько это возможно, без сопутствующего высвобождения нежелательных аминокислот, с использованием этой комбинации ферментов, очевидно, необходимо правильно выбрать условия, используемые в процессе ферментативного гидролиза. Кроме того, присутствующие в дрожжах эндогенные (и вероятно неспецифические) протеазы, должны быть инактивированы. После проведения ряда тест-инкубирований был разработан протокол, в котором использовались эти два новых фермента и который неожиданно обнаруживал селективное и эффективное высвобождение лейцина и фенилаланина из дрожжевых белков.
Для инактивации дрожжевых эндогенных протеаз дрожжевую суспензию выдерживали в течение 5 минут при 95°С. Затем эту суспензию быстро охлаждали до нужной температуры и рН доводили до 7,0 с использованием 4н NаОН. Дрожжи, термолизин и одну из аминопептидаз инкубировали одновременно в условиях, описанных ниже. После теплового шока рН 2000 миллилитров дрожжевой суспензии доводили до 7,0, после чего добавляли 680 миллиграммов термоазы и после перемешивания добавляли очищенную аминопептидазу. Эту смесь инкубировали при перемешивании при 50°С в течение 3 часов и центрифугировали. Для подавления всех ферментативных активностей рН супернатанта доводили до 4 и подвергали дополнительной тепловой обработке в течение 45 минут при 95°С. После дополнительного центрифугирования из супернатанта брали образец для аминокислотного анализа. Осажденное или нерастворенное вещество удаляли центрифугированием в течение 15 минут при 3500 об/мин в мегацентрифуге Hereaus Megafuge 2.0R. Супернатант удаляли и замораживали при -20°С. Сразу после оттаивания образцы супернатанта анализировали на содержание аминокислот методом Picotag, Waters (Milford MA, USA).
В этих аминокислотных анализах значения для Trp и Cys опускали, а значения для Asp и Asn суммировали с получением одной величины. В соответствии с этими данными в полученном гидролизате отношение аланина к лейцину (21,3:11,7) составляло 1:0,5. Коммерчески доступные дрожжевые гидролизаты обычно имели отношение аланина к лейцину 1:0,3.
Во втором эксперименте получали дрожжевой экстракт, который был обогащен свободным глутаматом. Для достижения этого были использованы эндопротеаза (кодируемая геном № 43 согласно таблице 1), обладающая предпочтением к расщеплению у С-конца глутаматных остатков, и карбоксипептидаза (кодируемая геном № 51 согласно таблице 1), способная удалять обработанные ею глутаматные остатки. Эндопротеаза, кодируемая геном 43, и карбоксипептидаза, кодируемая геном 51 (см. таблицу 1), были сверхэкспрессированы в A.niger методами, описанными ранее. Очистку этих ферментов проводили в соответствии с процедурами, описанными в примере 4.
Главная роль свободного глутамата в ряде процессов образования запахов хорошо описана, и известно, что MSG, натриевая соль глутаминовой кислоты, является единственным и самым важным компонентом, способствующим усилению вкуса.
В этом примере рН 200 мл дрожжевой суспензии, подвергнутой тепловому шоку, доводили до 8,0, а затем добавляли очищенный ферментный продукт, кодируемый геном 43, и смесь инкубировали в течение 4 часов при 50°С. Затем рН понижали до 5,0 и суспензию центрифугировали. К 100 миллилитрам супернатанта добавляли очищенный генный продукт гена 51. Инкубирование с этой карбоксипептидазой проводили в течение 30 минут при 50°С и при постоянной коррекции рН. После прекращения инкубирования фермента путем тепловой обработки в течение 5 минут при 95°С полученный материал снова центрифугировали (см. выше) и брали образец для аминокислотного анализа.
В соответствии с данными, полученными для аминокислот, в продуцированном гидролизате отношение аланина к глутамату (30,0:48,7) составляло 1:1,6. Коммерчески доступные дрожжевые гидролизаты обычно имели отношение аланина к глутамату 1:1.
Пример 10
Органолептическая оценка дрожжевых гидролизатов, обогащенных специфическими аминокислотами
Для подтверждения того, что гидролизат белка настоящего изобретения, обогащенный специфическими аминокислотами, может генерировать специфические запахи, был проведен ряд экспериментов с использованием дрожжевых гидролизатов, описанных в предыдущем примере. Для этой цели получали более крупные фрагменты этих гидролизатов и лиофилизовали. Свойства полученных порошков сравнивали со свойствами коммерчески доступного дрожжевого экстракта (Gistex LS, поставляемый фирмой DSM Food Specialties, Delft, The Netherlands) в стандартизированной смеси при нескольких реакционных условиях. Стандартизированная смесь содержала один из гидролизатов, смесь оснований и воду.
Смесь оснований содержала 22 грамма порошка (Maxarome Plus Powder) (специализированного дрожжевого экстракта с высоким содержанием природных нуклеотидов, также поставляемого фирмой DSM Food Specialties), 29,2 грамма глюкозы, 9 граммов жира REFEL-F (гидрогенизированного соевого масла, поставляемого фирмой Barentz, Hoofddorp, The Netherlands) и 0,2 грамма стеароиллактилата кальция (эмульгатора, поставляемого фирмой Abitec, Northampton, UK), тщательно перемешанных в ступке.
Все стандартизированные смеси, содержали 5 граммов порошкообразного дрожжевого гидролизата (то есть материала, обогащенного либо лейцином, либо глутаматом, либо коммерчески доступным дрожжевым экстрактом), 3 грамма смеси оснований и 3 грамма воды. После тщательного перемешивания эти три суспензии подвергали тепловой обработке в различных режимах, то есть либо выдерживали в течение 65 минут при 90-95°С в реакционном сосуде (жидкостная реакция), либо сушили под давлением 20 миллибар при 120°С в вакуумной печи (реакция вакуумной жарки), либо нагревали в открытом реакционном сосуде при 120°С в течение 10 минут после выпаривания всей воды (реакция жарки).
После тепловой обработки все три продукта оценивали на цвет в диапазоне от темно-коричневого до почти черного. В случае реакции вакуумной жарки были использованы только слабо окрашенные верхние слои. Оценку вкуса нагретого продукта проводили путем измельчения почерневших слоев с получением тонкодисперсных порошков и растворения этих порошков до концентрации 2% (мас./мас.) в воде, содержащей 0,6% (мас./мас.) NаСl. Дегустационные наблюдения представлены в таблице 3.
Пример 11
Неаллергенные гидролизаты белка молочной сыворотки, образованные трипептидилпептидазами
Дипептидилпептидазы, кодируемые генами 19 и 55, а также трипептидилпептидазы, кодируемые генами 4, 9, 10, 12, 26, 35, 46 и 50 (см. таблицу 1), были сверхпродуцированы, как описано выше, и они могут быть очищены методами, описанными в примере 4. После очистки оптимальный рН и термостабильность каждого отдельного фермента могут быть установлены любыми доступными методами, известными специалистам. Кроме того, специфичность каждого отдельного фермента может быть определена методами, описанными в примере 1. Селективность трипептидилпептидаз проиллюстрирована в следующем эксперименте.
Фермент, кодируемый геном 12, сверхпродуцировали в клетке-хозяине Aspergillus niger и очищали в соответствии с процедурами, описанными в примере 4. Полученный таким образом фермент инкубировали при рН 5 и при 50°С с различными синтетическими хромогенными субстратами, то есть Ala-Ala-Phe-рNA и Ala-Phe-рNA (оба поставлялись фирмой Bachem, Switserland). Инкубирование с субстратом Ala-Ala-Phe-рNA приводило к значительному увеличению оптической плотности при 410 нм, тогда как инкубирование с Ala-Phe-рNA не давало такого эффекта. Это наблюдение ясно продемонстрировало, что трипептидилпептидазы отщепляют трипептиды и не проявляют аминопептидазной активности, которая может приводить к нежелательному увеличению числа свободных аминокислот.
Кроме того, фермент, кодируемый геном 12, проявляет благоприятные свойства ферментативной стабильности, как было показано в следующем эксперименте. Четыре образца фермента инкубировали при рН 5 в течение одного часа при 0, 40, 50 и 60°С соответственно. Затем каждый образец фермента инкубировали с вышеуказанным субстратом Ala-Ala-Phe-рNA в цитратном буфере при рН 5 и остаточную активность в каждом отдельном образце определяли путем измерения увеличения оптической плотности при 410 нм. При 0°С образец проявлял 100%-ную активность, при 40°С образец проявлял 96%-ную остаточную активность, при 50°С образец проявлял 92%-ную остаточную активность, а при 60°С образец проявлял 88%-ную остаточную активность.
В обычном способе получения гидролизата с высоким содержанием трипептидов белок молочной сыворотки (WРС 75) может быть растворен/суспендирован в концентрации 100 граммов белка/литр в водной среде, имеющей рН 8,5. Первый фермент инкубировали с субтилизином, эндопротеазой широкого спектра (Delvolase®, 560000 DU на грамм, от DSM). После предварительного расщепления молочной сыворотки этим ферментом, взятым в концентрации 0,5% фермента на грамм белка, в течение 2 часов при 60°С смесь подвергали тепловой обработке для инактивации используемой эндопротеазы. Затем температуру доводили до 50°С, добавляли трипептидилпептидазу и всю смесь инкубировали до получения нужного уровня трипептидов. Последующие стадии обработки полученного таким образом гидролизата будут зависеть от цели конкретного применения, но они могут также включать микрофильтрацию или центрифугирование с последующим выпариванием и распылительной сушкой.
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЕ ГЕНЫ, КОДИРУЮЩИЕ НОВЫЕ ПРОТЕОЛИТИЧЕСКИЕ ФЕРМЕНТЫ | 2002 |
|
RU2296160C2 |
| ГЕН НОВОЙ СЕРИНОВОЙ ПРОТЕАЗЫ, РОДСТВЕННОЙ DPPIV | 2001 |
|
RU2305133C2 |
| НОВАЯ ПРОТЕАЗА ГРИБОВ И ЕЕ ПРИМЕНЕНИЕ | 2010 |
|
RU2566549C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ ПРОТЕАЗНОЙ АКТИВНОСТЬЮ, И НУКЛЕИНОВЫЕ КИСЛОТЫ, КОДИРУЮЩИЕ УКАЗАННЫЕ ПОЛИПЕПТИДЫ | 2002 |
|
RU2318018C2 |
| РЕКОМБИНАНТНАЯ ЭКСПРЕССИЯ БЕЛКОВ В ДВУХЦЕПОЧЕЧНОЙ ФОРМЕ С ДИСУЛЬФИДНЫМ МОСТИКОМ | 2006 |
|
RU2412253C2 |
| СПОСОБЫ И МАТЕРИАЛЫ ДЛЯ ЛЕЧЕНИЯ БОЛЕЗНИ ПОМПЕ | 2013 |
|
RU2644258C2 |
| НОВЫЕ ФУКОЗИЛТРАНСФЕРАЗЫ И ИХ ПРИМЕНЕНИЕ | 2011 |
|
RU2642307C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ ЭНДОПЕПТИДАЗНОЙ АКТИВНОСТЬЮ, И КОДИРУЮЩИЕ ИХ ПОЛИНУКЛЕОТИДЫ | 2011 |
|
RU2583293C2 |
| НОВЫЕ ФУКОЗИЛТРАСФЕРАЗЫ И ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2628307C2 |
| ЭКСПРЕССИОННЫЕ ВЕКТОРЫ ДЛЯ УЛУЧШЕННОЙ СЕКРЕЦИИ БЕЛКА | 2012 |
|
RU2661790C2 |













































































































































































































































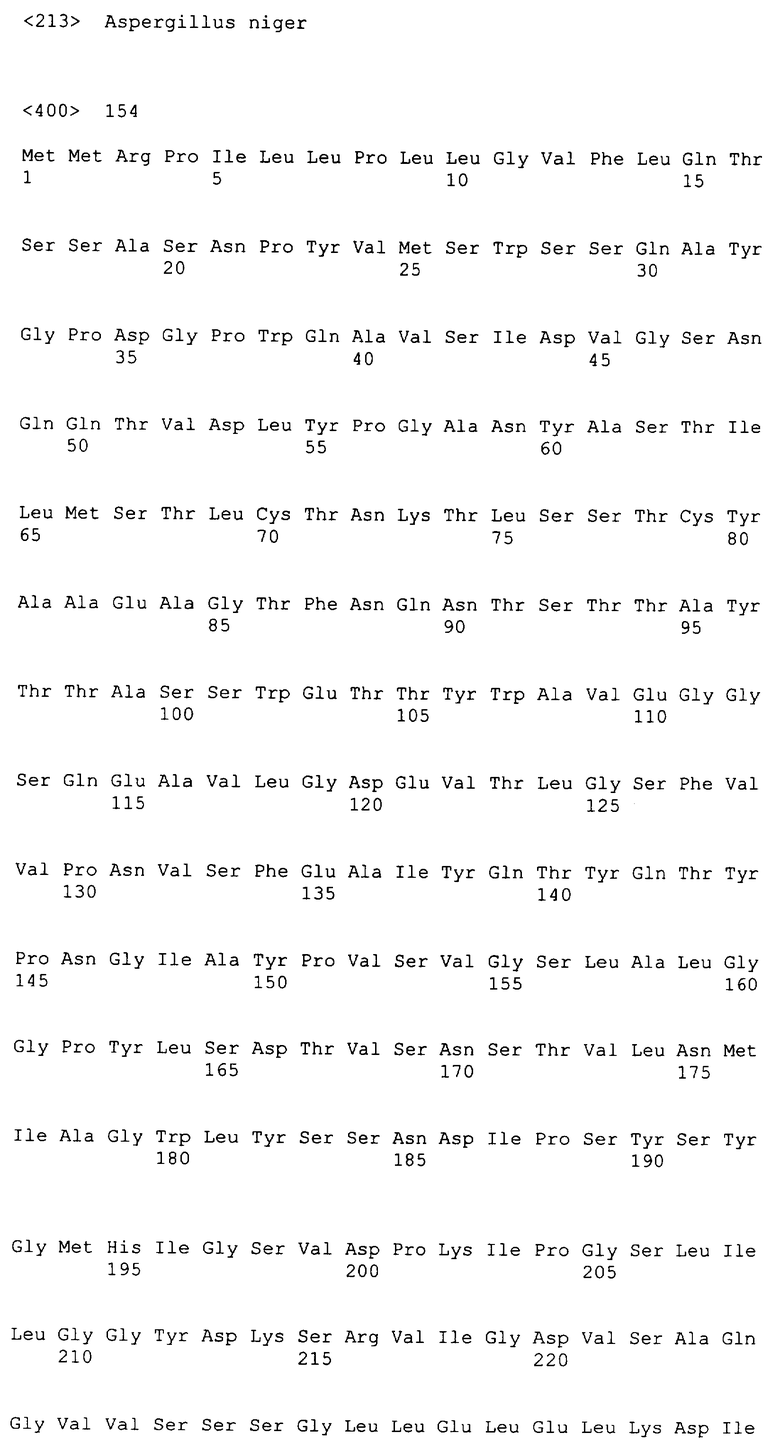








































Настоящее изобретение относится к биотехнологии. Представлен выделенный полинуклеотид, кодирующий аминопептидазу, содержащий нуклеотидную последовательность, приведенную в описании. Также представлен полинуклеотид, гибридизующийся с указанным полинуклеотидом в условиях высокой жесткости. Описаны экспрессионный вектор, содержащий указанный полинуклеотид, и подходящая клетка-хозяин. Представлен полипептид, соответствующий последовательности, приведенной в описании, и являющийся аминопептидазой. Предложен способ получения обозначенного полипептида, предусматривающий стадии трансформации подходящей клетки-хозяина указанным полинуклеотидом или вектором, культивирования клетки в условиях, подходящих для экспрессии указанного полинуклеотида, и, необязательно, очистку полипептида из указанной клетки или культуральной среды. Кроме того, был описан способ диагностики Aspergillus-инфекции в организме, предусматривающий стадии: а) выделения биологического образца из указанного организма, который, как предполагается, инфицирован Aspergillus, b) выделения нуклеиновой кислоты из указанного образца, с) определения, содержит ли указанная выделенная нуклеиновая кислота полинуклеотиды, гибридизующиеся с полинуклеотидом, выделенным из Aspergillus niger. Изобретение позволяет диагностировать Aspergillus-инфекцию в организме. 7 н. и 6 з.п. ф-лы, 3 табл.
1. Выделенный полинуклеотид, кодирующий аминопептидазу, гибридизующийся в условиях высокой жесткости с последовательностью, выбранной из группы, состоящей из SEQ ID NO:57 и 114.
2. Выделенный полинуклеотид по п.1, полученный из нитчатых грибов.
3. Выделенный полинуклеотид по п.2, полученный из A.niger.
4. Выделенный полинуклеотид, кодирующий аминопептидазу, содержащую аминокислотную последовательность, соответствующую последовательности SEQ ID NO:171.
5. Выделенный полинуклеотид, кодирующий аминопептидазу, содержащий нуклеотидную последовательность, соответствующую последовательности, выбранной из группы, состоящей из SEQ ID NO:57 и 114.
6. Экспрессионный вектор, содержащий полинуклеотидную последовательность по любому из пп.1-5.
7. Экспрессионный вектор по п.6, отличающийся тем, что указанная полинуклеотидная последовательность по любому из пп.1-5, функционально связана с регуляторными последовательностями, подходящими для экспрессии указанной полинуклеотидной последовательности в подходящей клетке-хозяине.
8. Экспрессионный вектор по п.7, отличающийся тем, что указанной подходящей клеткой-хозяином являются нитчатые грибы.
9. Выделенный полипептид, представляющий собой аминопептидазу, соответствующий последовательности SEQ ID NO:171, который можно получить путем экспрессии полинуклеотида по пп.1-5 или вектора по пп.6-8 в соответствующей клетке-хозяине.
10. Выделенный полипептид по п.9, полученный из Aspergillus niger.
11. Способ получения полипептида по любому из п.9 или 10, предусматривающий стадии трансформации подходящей клетки-хозяина выделенным полинуклеотидом по любому из пп.1-5 или вектором по любому из пп.6-8, культивирования указанной клетки в условиях, подходящих для экспрессии указанного полинуклеотида, и, необязательно, очистку полипептида из указанной клетки или культуральной среды.
12. Рекомбинантная клетка-хозяин, экспрессирующая полипептид по любому из п.9 или 10, содержащая полинуклеотид по любому из пп.1-5 или вектор по любому из пп.6-8.
13. Способ диагностики Aspergillus-инфекции в организме, предусматривающий стадии:
a. выделения биологического образца из указанного организма, который, как предполагается, инфицирован Aspergillus,
b. выделения нуклеиновой кислоты из указанного образца,
c. определения, содержит ли указанная выделенная нуклеиновая кислота полинуклеотиды, гибридизующиеся с полинуклеотидом по любому из пп.1-5.
| US 5989889, 23.11.1999 | |||
| US 6143546, 07.11.2000 | |||
| WO 1996014404 A1, 17.05.1996 | |||
| WO 1996028542 A1, 19.09.1996 | |||
| Наголовник для закрепления вибропогружателя на трубе с фланцем | 1981 |
|
SU1036843A1 |
| WO 1998054314 A1, 03.12.1998 | |||
| RU 2140986 C1, 10.11.1999. | |||

