Ссылка на список последовательностей
Данная заявка содержит список последовательностей в машиночитаемой форме, который включен в данное описание посредством ссылки.
Ссылка на депонирование биологического материала
Данная заявка содержит ссылку на депонирование биологического материала, который включен в данное описание посредством ссылки.
Уровень техники
Область техники, к которой относится изобретения
Настоящее изобретение относится к полипептидам, обладающим эндопептидазной активностью, а также к способам получения и применения таких полипептидов. Изобретение также относится к способам получения гидролизата белка, такого как гидролизат пищевого белка.
Описание уровня техники
Трипсин (EC 3.4.21.4) представляет собой сериновую протеазу, встречающуюся в пищеварительной системе многих позвоночных, где она гидролизует белки. Трипсин расщепляет пептидные цепи в основном на карбоксильном конце аминокислот лизина и аргинина. Трипсин присутствует в большом количестве в поджелудочной железе и может быть очищен довольно легко. Поэтому он широко используется в различных биотехнологических процессах. Трипсин используют в детском питании для его предварительного расщепления. Он способен разрушать белковые молекулы, что помогает ребенку переваривать белок, поскольку детский желудок не развит в достаточной степени для переваривания крупных белковых молекул. Трипсин можно использовать для разрушения белков молока с получением частичного гидролизата молочных белков для детского питания. Например, в WO 93/04593 и US 5039532 описано применение препаратов трипсина поджелудочной железы для получения гипоаллергенных гидролизатов белков молочной сыворотки.
По ряду причин, при производстве пищевых продуктов и, в частности, при производстве детских продуктов или питания для грудных детей использование протеолитических ферментов, полученных из микроорганизмов, таких как бактерии, может обеспечивать преимущества. Например, производство бактериальных ферментов можно легко оптимизировать, чтобы оно было эффективным и легко контролируемым. Вследствие этого, такие ферменты можно получать в больших количествах и с высокой степенью чистоты. Кроме того, использование микробных ферментов поможет преодолеть возрастающие трудности, связанные с системой обеспечения качества, которые возникают в случае экстрагирования ферментов из животных источников.
Одной из целей настоящего изобретения является предложение новых микробных протеаз для потенциального использования, например, в пищевой промышленности.
Сущность изобретения
Авторы настоящего изобретения неожиданно обнаружили, что некоторые бактериальные пептидазы обладают специфичностью расщепления, очень сходной с расщеплением трипсина поджелудочной железы. Кроме того, такие бактериальные эндопептидазы можно использовать для получения гидролизатов пищевых белков, имеющих сходные свойства, например, сходную степень гидролиза и/или сходный пептидный спектр, как у гидролизатов пищевых белков, полученных при помощи трипсина поджелудочной железы.
Кроме того, авторы настоящего изобретения идентифицировали новую бактериальную эндопептидазу, обладающую трипсиноподобной активностью.
Таким образом, настоящее изобретение относится к выделенным полипептидам, обладающим эндопептидазной активностью, выбранным из группы, состоящей из:
(a) полипептида, имеющего по меньшей мере 70% идентичность последовательности со зрелым полипептидом SEQ ID NO:2;
(b) полипептида, кодируемого полинуклеотидом, который гибридизуется в нестрогих условиях с (i) кодирующей зрелый полипептид последовательностью SEQ ID NO:1, (ii) последовательностью геномной ДНК, содержащей кодирующую зрелый полипептид последовательность SEQ ID NO:1, или (iii) полноразмерной комплементарной цепью для (i) или (ii);
(c) полипептида, кодируемого полинуклеотидом, имеющим по меньшей мере 70% идентичность последовательности с кодирующей зрелый полипептид последовательностью SEQ ID NO:1;
(d) варианта, содержащего замену, делецию и/или вставку одной или более (нескольких) аминокислот, зрелого полипептида SEQ ID NO:2; и
(e) фрагмента полипептида по пунктам (a), (b), (c) или (d), обладающего эндопептидазной активностью.
Настоящее изобретение также относится к способам получения данных полипептидов.
Настоящее изобретение также относится к способам применения полипептидов для получения гидролизата белка.
В другом аспекте настоящее изобретение относится к способу получения гидролизата пищевого белка, включающему:
(a) обеспечение раствора, содержащего пищевой белок, подлежащий гидролизу;
(b) добавление к указанному раствору трипсиноподобной эндопептидазы, полученной из бактерии; и
(c) получение гидролизата пищевого белка.
Краткое описание фигур
На фиг.1 представлена карта плазмиды ExpVec8 с геном, кодирующим трипсиноподобную эндопептидазу из Kutzneria albida.
На фиг.2 представлены УФ-хроматограммы бычьего альфа-лактальбумина, гидролизованного трипсиноподобной эндопептидазой из Kutzneria albida (верхняя кривая) или свиным трипсином (нижняя кривая).
Определения
Эндопептидазная активность: Термин «эндопептидазная активность» означает протеолитическую активность, способную приводить к гидролизу любой пептидной связи в пептиде. Однако, поскольку эндопептидазы часто имеют каталитические центры, в которых происходит связывание с несколькими аминокислотами и часто по обе стороны от точки расщепления, эндопептидазы в целом демонстрируют предпочтение в отношении не-концевых пептидных связей, в отличие от экзопептидаз, которые гидролизуют пептидные связи с любого конца пептида. Эндопептидазы, как правило, классифицируются как EC 3.4.21-25. Для целей настоящего изобретения эндопептидазную активность можно определять, используя анализ с Protazyme AK, как описано в примере 2.
Полипептиды по настоящему изобретению могут обладать по меньшей мере 20%, например, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95% или по меньшей мере 100% эндопептидазной активностью зрелого полипептида SEQ ID NO:2.
Трипсиноподобная эндопептидаза: Термин «трипсиноподобная эндопептидаза» или «эндопептидаза, обладающая трипсиноподобной активностью» в данном описании означает эндопептидазу, которая преимущественно расщепляет пептиды или белки в С-концевой части L-изомера аргинина и/или лизина. В предпочтительном варианте осуществления трипсиноподобная эндопептидаза преимущественно расщепляет пептиды или белки в С-концевой части аргинина и лизина. Это означает, что эндопептидаза имеет более высокую специфичность в отношении расщепления после как аргинина, так и лизина, чем она имеет в отношении расщепления после любой другой аминокислоты. В другом предпочтительном варианте осуществления трипсиноподобная эндопептидаза преимущественно расщепляет пептиды или белки в С-концевой части аргинина или лизина. Это означает, что эндопептидаза имеет более высокую специфичность в отношении расщепления после любого из аргинина или лизина, чем она имеет в отношении расщепления после любой другой аминокислоты. В другом предпочтительном варианте осуществления трипсиноподобная эндопептидаза преимущественно расщепляет пептиды или белки в С-концевой части аргинина. Это означает, что эндопептидаза имеет более высокую специфичность в отношении расщепления после аргинина, чем она имеет в отношении расщепления после любой другой аминокислоты. В другом предпочтительном варианте осуществления трипсиноподобная эндопептидаза преимущественно расщепляет пептиды или белки в С-концевой части лизина. Это означает, что эндопептидаза имеет более высокую специфичность в отношении расщепления после лизина, чем она имеет в отношении расщепления после любой другой аминокислоты.
Трипсиновое отношение: «Трипсиновое отношение» определяют как активность фермента при расщеплении после Arg или Lys (в зависимости от того, что больше), деленную на активность фермента при расщеплении после любого из Ala, Asp, Glu, Ile, Leu, Met, Phe или Val (в зависимости от того, что больше). В предпочтительном варианте осуществления трипсиноподобная эндопептидаза по изобретению имеет трипсиновое отношение более 100. То есть, в предпочтительном варианте осуществления трипсиноподобная эндопептидаза по изобретению имеет специфичность в отношении расщепления после Arg или Lys (в зависимости от того, что больше), которая по меньшей мере в 100 раз выше, чем ее специфичность в отношении расщепления после любого из Ala, Asp, Glu, Ile, Leu, Met, Phe или Val (в зависимости от того, что больше). Подобные измерения активности для определения трипсинового отношения следует проводить при pH, при котором активность эндопептидазы составляет по меньшей мере половину активности эндопептидазы при оптимальном для нее pH. Трипсиновое отношение можно определять, как описано в примере 2 настоящего описания.
Выделенный полипептид: Термин «выделенный полипептид» означает полипептид, который модифицирован рукой человека относительно полипептида, который встречается в природе. В одном варианте осуществления полипептид является по меньшей мере на 1% чистым, например, по меньшей мере на 5% чистым, по меньшей мере на 10% чистым, по меньшей мере на 20% чистым, по меньшей мере на 40% чистым, по меньшей мере на 60% чистым, по меньшей мере на 80% чистым или по меньшей мере на 90% чистым, что определяют методом SDS-ПААГ.
По существу чистый полипептид: Термин «по существу чистый полипептид» означает препарат, который содержит не более 10%, не более 8%, не более 6%, не более 5%, не более 4%, не более 3%, не более 2%, не более 1%, или предпочтительно не более 0,5% масс. другого полипептидного материала, с которым он природно или рекомбинантно связан. Предпочтительно, полипептид является по меньшей мере на 92% чистым, например, по меньшей мере на 94% чистым, по меньшей мере на 95% чистым, по меньшей мере на 96% чистым, по меньшей мере на 97% чистым, по меньшей мере на 98% чистым, по меньшей мере на 99%, по меньшей мере на 99,5% чистым или, наиболее предпочтительно, на 100% чистым из расчета на массу общего полипептидного материала, присутствующего в препарате. Полипептиды по настоящему изобретению предпочтительно находятся по существу в чистом виде. Этого можно достичь, например, получая полипептид хорошо известными рекомбинантными способами или классическими методами очистки.
Зрелый полипептид: Термин «зрелый полипептид» означает полипептид в его конечной форме после трансляции и любых посттрансляционных модификаций, таких как N-концевой процессинг, C-концевое усечение, гликозилирование, фосфорилирование и т.д. В одном варианте осуществления зрелый полипептид представляет собой аминокислоты 1-225 SEQ ID NO:2, на основании N-концевого секвенирования и определения молекулярной массы, как описано в примере 2.
Кодирующая зрелый полипептид последовательность: Термин «кодирующая зрелый полипептид последовательность» означает полинуклеотид, который кодирует зрелый полипептид, обладающий эндопептидазной активностью. В одном варианте осуществления кодирующая зрелый полипептид последовательность может представлять собой нуклеотиды 1-675 SEQ ID NO:1, на основании аминокислотной последовательности зрелого полипептида. В другом варианте осуществления кодирующая зрелый полипептид последовательность может представлять собой нуклеотиды 82-756 SEQ ID NO:3.
Идентичность последовательностей: Родство между двумя аминокислотными последовательностями или между двумя нуклеотидными последовательностями описывается параметром «идентичность последовательностей».
Для целей настоящего изобретения степень идентичности последовательностей между двумя аминокислотными последовательностями определяют при помощи алгоритма Нидлмана-Вунша (Needleman-Wunsch) (Needleman and Wunsch, 1970, J. Mol. Biol. 48: 443-453), реализованного в программе Needle пакета программ EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, Trends Genet. 16: 276-277), предпочтительно версии 2.8.0 или более поздней. Дополнительными используемыми параметрами являются: штраф за внесение делеции 10, штраф за продолжение делеции 0,5 и матрица замен EBLOSUM62 (EMBOSS версия BLOSUM62). Результат в программе Needle, помеченный «идентичность наибольшей длины» (полученный при помощи параметра -nobrief) используют в качестве процента идентичности и рассчитывают следующим образом:
(Идентичные остатки×100)/(длина выравнивания-общее число делеций в выравнивании)
Для целей настоящего изобретения степень идентичности последовательностей между двумя дезоксирибонуклеотидными последовательностями определяют при помощи алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, выше), реализованного в программе Needle пакета программ EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, выше), предпочтительно версии 3.0.0 или более поздней. Дополнительными используемыми параметрами являются: штраф за внесение делеции 10, штраф за продолжение делеции 0,5 и матрица замен EDNAFULL (EMBOSS версия NCBI NUC4.4). Результат в программе Needle, помеченный «идентичность наибольшей длины» (полученный при помощи параметра -nobrief) используют в качестве процента идентичности и рассчитывают следующим образом:
(Идентичные дезоксирибонуклеотиды×100)/(длина выравнивания-общее число делеций в выравнивании)
Фрагмент: Термин «фрагмент» означает полипептид, у которого одна или более (несколько) аминокислот делетированы с амино и/или карбоксильного конца зрелого полипептида; при этом фрагмент обладает эндопептидазной активностью. В одном варианте осуществления фрагмент содержит по меньшей мере 100 аминокислотных остатков, по меньшей мере 150 аминокислотных остатков или по меньшей мере 200 аминокислотных остатков.
Подпоследовательность: Термин «подпоследовательность» означает полинуклеотид, у которого один или более (несколько) нуклеотидов делетированы с 5' и/или 3'-конца кодирующей зрелый полипептид последовательности; при этом подпоследовательность кодирует фрагмент, обладающий эндопептидазной активностью. В одном варианте осуществления подпоследовательность содержит по меньшей мере 300 нуклеотидов, например, по меньшей мере 400 нуклеотидов, по меньшей мере 500 нуклеотидов или по меньшей мере 600 нуклеотидов.
Аллельный вариант: Термин «аллельный вариант» означает любую из двух или более альтернативных форм гена, занимающую один и тот же хромосомный локус. Аллельные вариации естественным образом возникают в результате мутации и могут приводить к полиморфизму в популяциях. Генные мутации могут быть «молчащими» (без изменений в кодируемом полипептиде) или могут приводить к кодированию полипептидов, имеющих измененные аминокислотные последовательности. Аллельный вариант полипептида представляет собой полипептид, кодируемый аллельным вариантом гена.
Выделенный полинуклеотид: Термин «выделенный полинуклеотид» означает полинуклеотид, который модифицирован рукой человека относительно полинуклеотида, встречающегося в природе. В одном варианте осуществления выделенный полинуклеотид является по меньшей мере на 1% чистым, например, по меньшей мере на 5% чистым, предпочтительно по меньшей мере на 10% чистым, по меньшей мере на 20% чистым, по меньшей мере на 40% чистым, по меньшей мере на 60% чистым, по меньшей мере на 80% чистым, по меньшей мере на 90% чистым или, более предпочтительно, по меньшей мере на 95% чистым, как определяют электрофорезом в агарозе. Полинуклеотиды могут иметь геномное, кДНК, РНК, полусинтетическое, синтетическое происхождение, либо представлять собой любую их комбинацию.
По существу чистый полинуклеотид: Термин «по существу чистый полинуклеотид» означает препарат полинуклеотида, свободный от других посторонних или нежелательных нуклеотидов и находящийся в форме, подходящей для использования в генно-инженерных системах производства полипептидов. Таким образом, по существу чистый полинуклеотид содержит не более 10%, не более 8%, не более 6%, не более 5%, не более 4%, не более 3%, не более 2%, не более 1% или не более 0,5% масс. другого полинуклеотидного материала, с которым он природно или рекомбинантно связан. По существу чистый полинуклеотид может, однако, содержать 5' и 3'-нетранслируемые области природного происхождения, такие как промоторы и терминаторы. Предпочтительно, полинуклеотид является по меньшей мере на 90% чистым, например, по меньшей мере на 92% чистым, по меньшей мере на 94% чистым, по меньшей мере на 95% чистым, по меньшей мере на 96% чистым, по меньшей мере на 97% чистым, по меньшей мере на 98% чистым, по меньшей мере на 99% чистым или по меньшей мере на 99,5% чистым по массе. Полинуклеотиды по настоящему изобретению предпочтительно находятся по существу в чистом виде.
Кодирующая последовательность: Термин «кодирующая последовательность» означает полинуклеотид, который непосредственно определяет аминокислотную последовательность полипептида. Границы кодирующей последовательности, как правило, определяются открытой рамкой считывания, которая обычно начинается со стартового кодона ATG или альтернативных стартовых кодонов, таких как GTG и TTG, и заканчивается стоп-кодоном, таким как TAA, TAG и TGA. Кодирующая последовательность может представлять собой ДНК, кДНК, синтетический или рекомбинантный полинуклеотид.
кДНК: Термин «кДНК» означает молекулу ДНК, которую можно получать путем обратной транскрипции со зрелой, сплайсированной молекулы мРНК, полученной из эукариотической клетки. У кДНК отсутствуют интронные последовательности, которые могут присутствовать в соответствующей геномной ДНК. Исходный первичный РНК-транскрипт является предшественником мРНК, который процессируется в несколько стадий, включая сплайсинг, перед тем, как стать зрелой сплайсированной мРНК.
Конструкт нуклеиновой кислоты: Термин «конструкт нуклеиновой кислоты» означает молекулу нуклеиновой кислоты, либо одно- либо двухцепочечную, которая выделена из природного гена или модифицирована для содержания сегментов нуклеиновых кислот таким образом, что без этого не могла бы существовать в природе, либо является синтетической. Термин «конструкт нуклеиновой кислоты» является синонимом термину «экспрессионная кассета», когда конструкт нуклеиновой кислоты содержит контрольные последовательности, необходимые для экспрессии кодирующей последовательности по настоящему изобретению.
Контрольные последовательности: Термин «контрольные последовательности» означает все компоненты, необходимые для экспрессии полинуклеотида, кодирующего полипептид по настоящему изобретению. Каждая контрольная последовательность может быть нативной или чужеродной для полинуклеотида, кодирующего полипептид, либо они могут нативными или чужеродными друг для друга. Такие контрольные последовательности включают, но не ограничиваются ими, лидерную последовательность, последовательность полиаденилирования, пропептидную последовательность, промотор, сигнальную пептидную последовательность и последовательность терминации транскрипции. Как минимум, контрольные последовательности включают промотор, а также транскрипционный и трансляционный стоп-сигналы. Контрольные последовательности могут быть снабжены линкерами с целью введения конкретных сайтов рестрикции, облегчающих лигирование контрольных последовательностей с кодирующей областью полинуклеотида, кодирующего полипептид.
Функционально связанные: Термин «функционально связанные» означает конфигурацию, в которой контрольная последовательность помещена в соответствующее положение относительно кодирующей последовательности полинуклеотида таким образом, что контрольная последовательность направляет экспрессию кодирующей последовательности.
Экспрессия: Термин «экспрессия» охватывает любую стадию процесса создания полипептида, включая, но, не ограничиваясь ими, транскрипцию, посттранскрипционные модификации, трансляцию, посттрансляционные модификации, а также секрецию.
Экспрессионный вектор: Термин «экспрессионный вектор» означает линейную или кольцевую молекулу ДНК, содержащую полинуклеотид, который кодирует полипептид и функционально связан с дополнительными нуклеотидами, обеспечивающими его экспрессию.
Клетка-хозяин: Термин «клетка-хозяин» означает любой тип клеток, подверженных трансформации, трансфекции, трансдукции и подобному, при помощи конструкта нуклеиновой кислоты или экспрессионного вектора, содержащего полинуклеотид по настоящему изобретению. Термин «клетка-хозяин» охватывает любое потомство родительской клетки, которое не идентично родительской клетке вследствие мутаций, происходящих в процессе репликации.
Вариант: Термин «вариант» означает полипептид, обладающий эндопептидазной активностью, который содержит изменение, то есть, замену, вставку и/или делецию одного или более (нескольких) аминокислотных остатков в одном или более (нескольких) положениях. Замена означает замену аминокислоты, занимающей некое положение, другой аминокислотой; делеция означает удаление аминокислоты, занимающей некое положение; и вставка означает добавление 1-3 аминокислот рядом с аминокислотой, занимающей некое положение.
Подробное описание изобретения
Полипептиды, обладающие эндопептидазной активностью
Полипептиды
Настоящее изобретение относится к выделенным полипептидам, обладающим эндопептидазной активностью, выбранным из группы, состоящей из:
a) полипептида, имеющего по меньшей мере 70% идентичность последовательности со зрелым полипептидом SEQ ID NO:2;
(b) полипептида, кодируемого полинуклеотидом, который гибридизуется в нестрогих условиях с (i) кодирующей зрелый полипептид последовательностью SEQ ID NO:1, (ii) последовательностью геномной ДНК, содержащей кодирующую зрелый полипептид последовательность SEQ ID NO:1, или (iii) полноразмерной комплементарной цепью для (i) или (ii);
(c) полипептида, кодируемого полинуклеотидом, имеющим по меньшей мере 70% идентичность последовательности с кодирующей зрелый полипептид последовательностью SEQ ID NO:1;
(d) варианта, содержащего замену, делецию и/или вставку одной или более (нескольких) аминокислот, зрелого полипептида SEQ ID NO:2; и
(e) фрагмента полипептида по пунктам (a), (b), (c) или (d), обладающего эндопептидазной активностью.
Настоящее изобретение относится к выделенным полипептидам, имеющим идентичность последовательности со зрелым полипептидом SEQ ID NO:2 по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100%, которые обладают эндопептидазной активностью. В одном варианте осуществления полипептиды отличаются не более чем десятью аминокислотами, например, пятью аминокислотами, четырьмя аминокислотами, тремя аминокислотами, двумя аминокислотами или одной аминокислотой, от зрелого полипептида SEQ ID NO:2.
Полипептид по настоящему изобретению предпочтительно содержит или состоит из аминокислотной последовательности SEQ ID NO:2 или ее аллельного варианта; или представляет собой ее фрагмент, обладающий эндопептидазной активностью. В другом варианте осуществления полипептид содержит или состоит из зрелого полипептида SEQ ID NO:2. В другом предпочтительном варианте осуществления полипептид содержит или состоит из аминокислот 1-225 SEQ ID NO:2.
Настоящее изобретение также относится к выделенным полипептидам, обладающим эндопептидазной активностью, которые кодируются полинуклеотидами, гибридизующимися в очень нестрогих условиях, нестрогих условиях, условиях средней строгости, условиях от средней до высокой степени строгости, строгих условиях или очень строгих условиях с (i) кодирующей зрелый полипептид последовательностью SEQ ID NO:1, (ii) последовательностью геномной ДНК, содержащей кодирующую зрелый полипептид последовательность SEQ ID NO:1, или (iii) полноразмерной комплементарной цепью для (i) или (ii) (J. Sambrook, E.F. Fritsch, and T. Maniatis, 1989, Molecular Cloning, A Laboratory Manual, 2d edition, Cold Spring Harbor, New York).
Полинуклеотид SEQ ID NO:1 или его подпоследовательность, а также аминокислотную последовательность SEQ ID NO:2 или ее фрагмент можно использовать для конструирования нуклеиновокислотных зондов с целью идентификации и клонирования ДНК, кодирующей полипептиды, обладающие эндопептидазной активностью, из штаммов различных родов или видов способами, хорошо известными в данной области. В частности, такие зонды можно использовать для гибридизации с геномной или кДНК представляющего интерес рода или вида, с последующими стандартными процедурами саузерн-блоттинга, с целью идентификации и выделения соответствующего им гена. Такие зонды могут быть значительно короче, чем вся последовательность, однако должны иметь по меньшей мере 14, например, по меньшей мере 25, по меньшей мере 35 или по меньшей мере 70 нуклеотидов в длину. Предпочтительно, нуклеиновокислотный зонд имеет по меньшей мере 100 нуклеотидов в длину, например, по меньшей мере 200 нуклеотидов, по меньшей мере 300 нуклеотидов, по меньшей мере 400 нуклеотидов, по меньшей мере 500 нуклеотидов или по меньшей мере 600 нуклеотидов в длину. Можно использовать как ДНК-, так и РНК-зонды. Как правило, зонды метят для обнаружения соответствующего гена (например, при помощи 32P, 3H, 35S, биотина или авидина). Такие зонды охвачены настоящим изобретением.
Библиотеку геномной ДНК или кДНК, полученную из других таких штаммов, можно подвергать скринингу на ДНК, которая гибридизуется с зондами, описанными выше, и кодирует полипептид, обладающий эндопептидазной активностью. Геномную или другую ДНК из таких других штаммов можно выделять электрофорезом в агарозном или полиакриламидном геле, либо другими методами разделения. ДНК из библиотек или выделенную ДНК можно переносить и иммобилизовать на нитроцеллюлозе или другом подходящем материале-носителе. Для идентификации клона или ДНК, которая гомологична с SEQ ID NO:1 или ее подпоследовательностью, материал-носитель предпочтительно используют в саузерн-блоттинге.
Для целей настоящего изобретения, гибридизация указывает на то, что полинуклеотид гибридизуется с меченым нуклеиновокислотным зондом, соответствующим кодирующей зрелый полипептид последовательности SEQ ID NO:1; последовательности геномной ДНК, содержащей кодирующую зрелый полипептид последовательность SEQ ID NO:1; ее полноразмерной комплементарной цепи; или ее подпоследовательности; в условиях от очень нестрогих до очень строгих. Молекулы, с которыми нуклеиновокислотный зонд гибридизуется в таких условиях, можно обнаруживать, используя, например, рентгеновскую пленку.
В одном варианте осуществления нуклеиновокислотный зонд представляет собой кодирующую зрелый полипептид последовательность SEQ ID NO:1. В другом варианте осуществления нуклеиновокислотный зонд представляет собой нуклеотиды 1-200, нуклеотиды 201-400, нуклеотиды 401-600 или нуклеотиды 401-625 SEQ ID NO:1. В другом варианте осуществления нуклеиновокислотный зонд представляет собой полинуклеотид, который кодирует полипептид SEQ ID NO:2 или его фрагмент. В другом предпочтительном варианте осуществления нуклеиновокислотный зонд представляет собой SEQ ID NO:1. В другом варианте осуществления нуклеиновокислотный зонд представляет собой полинуклеотид, содержащийся в плазмиде ExpVec8, которая содержится в E. coli DSM 23706, при этом полинуклеотид кодирует полипептид, обладающий эндопептидазной активностью. В другом варианте осуществления нуклеиновокислотный зонд представляет собой кодирующую зрелый полипептид область, содержащуюся в плазмиде ExpVec8, которая содержится в E. coli DSM 23706.
Для длинных зондов, имеющих по меньшей мере 100 нуклеотидов в длину, условия от очень нестрогих до очень строгих соответствуют предгибридизации и гибридизации при 42°C в смеси 5X SSPE, 0,3% SDS, 200 микрограмм/мл фрагментированной и денатурированной ДНК спермы лосося и либо 25% формамида для очень нестрогих и нестрогих условий, 35% формамида для условий средней строгости и от средней до высокой степени строгости, либо 50% формамида для строгих и очень строгих условий, после стандартных процедур саузерн-блоттинга в течение 12-24 часов оптимально. В конечном итоге, материал-носитель промывают три раза по 15 минут буфером 2X SSC, 0,2% SDS при 45°C (очень нестрогие условия), при 50°C (нестрогие условия), при 55°C (условия средней строгости), при 60°C (условия от средней до высокой степени строгости), при 65°C (строгие условия) и при 70°C (очень строгие условия).
Для коротких зондов, имеющих примерно от 15 нуклеотидов до примерно 70 нуклеотидов в длину, строгие условия соответствуют предгибридизации и гибридизации при температуре примерно от 5°C до примерно 10°C ниже рассчитанного значения Tm, которое рассчитано согласно Bolton and McCarthy (1962, Proc. Natl. Acad. Sci. USA 48: 1390), в смеси 0,9 M NaCl, 0,09 M Трис-HCl pH 7,6, 6 мМ EDTA, 0,5% NP-40, 1X раствора Денхардта, 1 мМ пирофосфата натрия, 1 мМ одноосновного фосфата натрия, 0,1 мМ АТФ и 0,2 мг дрожжевой РНК на мл после стандартных процедур саузерн-блоттинга в течение 12-24 часов оптимально. В конечном итоге, материал-носитель промывают один раз в 6X SCC плюс 0,1% SDS в течение 15 минут и два раза по 15 минут, используя 6X SSC при температуре от 5°C до 10°C ниже рассчитанного значения Tm.
Настоящее изобретение также относится к выделенным полипептидам, обладающим эндопептидазной активностью, кодируемым полинуклеотидами, имеющими идентичность последовательности с кодирующей зрелый полипептид последовательностью SEQ ID NO:1 по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100%.
Настоящее изобретение также относится к вариантам, содержащим замену, делецию и/или вставку одной или более (или нескольких) аминокислот, зрелого полипептида SEQ ID NO:2, либо его гомологичной последовательности. Предпочтительно, изменения аминокислот носят незначительный характер, то есть консервативные аминокислотные замены или вставки, которые существенно не влияют на сворачивание и/или активность белка; небольшие делеции, как правило, от одной до примерно 30 аминокислот; небольшие удлинения с амино- или карбоксильного конца, такие как амино-концевой остаток метионина; небольшой линкерный пептид длиной примерно до 20-25 остатков; или небольшое удлинение, которое облегчает очистку вследствие изменения суммарного заряда или другой функции, такое как полигистидиновая область, антигенный эпитоп или связывающий домен.
Примеры консервативных замен существуют в группе основных аминокислот (аргинин, лизин и гистидин), кислых аминокислот (глютаминовая кислота и аспарагиновая кислота), полярных аминокислот (глютамин и аспарагин), гидрофобных аминокислот (лейцин, изолейцин и валин), ароматических аминокислот (фенилаланин, триптофан и тирозин) и небольших аминокислот (глицин, аланин, серин, треонин и метионин). Аминокислотные замены, которые, как правило, не изменяют специфическую активность, известны в данной области и описаны, например, H. Neurath and R.L. Hill, 1979, в: The Proteins, Academic Press, New York. Наиболее часто встречающимися заменами являются Ala/Ser, Val/Ile, Asp/Glu, Thr/Ser, Ala/Gly, Ala/Thr, Ser/Asn, Ala/Val, Ser/Gly, Tyr/Phe, Ala/Pro, Lys/Arg, Asp/Asn, Leu/Ile, Leu/Val, Ala/Glu и Asp/Gly.
Альтернативно, существуют аминокислотные замены такой природы, что изменяются физико-химические свойства полипептидов. Например, аминокислотные замены могут улучшать термостабильность полипептида, изменять субстратную специфичность, изменять рН-оптимум и тому подобное.
Незаменимые аминокислоты в родительском полипептиде можно определять методами, известными в данной области, такими как сайт-направленный мутагенез или аланин-сканирующий мутагенез (Cunningham and Wells, 1989, Science 244: 1081-1085). В последнем методе проводят одиночные мутации на аланин каждого остатка в молекуле, и полученные мутантные молекулы тестируют на эндопептидазную активность, предпочтительно, трипсиноподобную эндопептидазную активность, для определения аминокислотных остатков, которые критичны для активности молекулы. См. также, Hilton et al., 1996, J. Biol. Chem. 271: 4699-4708. Активный центр фермента или другие биологические взаимодействия можно также определять путем физического анализа структуры, который осуществляют такими методами, как ядерный магнитный резонанс, кристаллография, дифракция электронов или фотоаффинное мечение, в сочетании с мутацией аминокислот предполагаемого контактного центра. См., например, de Vos et al., 1992, Science 255: 306-312; Smith et al., 1992, J. Mol. Biol. 224: 899-904; Wlodaver et al., 1992, FEBS Lett. 309: 59-64. Конкретные незаменимые аминокислоты также можно определять на основании анализа идентичности с полипептидами, родственными родительскому полипептиду.
Одиночные или множественные аминокислотные замены, делеции и/или вставки можно создавать и тестировать, применяя известные методы мутагенеза, рекомбинации и/или перетасовки, сопровождаемых соответствующей процедурой скрининга, таких как описано в статьях Reidhaar-Olson and Sauer, 1988, Science 241: 53-57; Bowie and Sauer, 1989, Proc. Natl. Acad. Sci. USA 86: 2152-2156; WO 95/17413 или WO 95/22625. Другие методы, которые могут быть использованы, включают ПЦР пониженной точности, фаговый дисплей (например, Lowman et al., 1991, Biochemistry 30: 10832-10837; патент США 5223409; WO 92/06204) и сегмент-направленный мутагенез (Derbyshire et al., 1986, Gene 46: 145; Ner et al., 1988, DNA 7: 127).
Методы мутагенеза/перетасовки можно комбинировать с высокопроизводительными автоматизированными методами скрининга для определения активности клонированных мутагенизированных полипептидов, экспрессируемых клетками-хозяевами (Ness et al., 1999, Nature Biotechnology 17: 893-896). Мутагенизированные молекулы ДНК, которые кодируют активные полипептиды, можно выделять из клеток-хозяев и быстро секвенировать стандартными методами в данной области. Эти методы позволяют быстро определять важность отдельных аминокислотных остатков в полипептиде.
Общее число аминокислотных замен, делеций и/или вставок в зрелом полипептиде SEQ ID NO:2 составляет не более чем 10, например, 1, 2, 3, 4, 5, 6, 7, 8 или 9.
Полипептид может представлять собой гибридный полипептид, в котором часть одного полипептида слита с N-концом или C-концом части другого полипептида.
Полипептид может представлять собой слитый полипептид или расщепляемый слитый полипептид, в котором другой полипептид слит с N-концом или C-концом полипептида по настоящему изобретению. Слитый полипептид получают слиянием полинуклеотида, кодирующего другой полипептид, с полинуклеотидом по настоящему изобретению. Способы получения слитых полипептидов известны в данной области и включают лигирование кодирующих последовательностей, кодирующих полипептиды, таким образом, что они находятся внутри рамки считывания, и экспрессия слитого полипептида находится под контролем одного и того же промотора(ов) и терминатора. Слитые белки можно также конструировать, используя технологию интеинов, в которой слияния создаются посттрансляционно (Cooper et al., 1993, EMBO J. 12: 2575-2583; Dawson et al., 1994, Science 266: 776-779).
Слитый полипептид может дополнительно содержать сайт расщепления между двумя полипептидами. После секреции слитого белка происходит расщепление по сайту, и высвобождаются два полипептида. Примеры сайтов расщепления включают, но не ограничиваются ими, сайты, описанные в статьях Martin et al., 2003, J. Ind. Microbiol. Biotechnol. 3: 568-576; Svetina et al., 2000, J. Biotechnol. 76: 245-251; Rasmussen-Wilson et al., 1997, Appl. Environ. Microbiol. 63: 3488-3493; Ward et al., 1995, Biotechnology 13: 498-503 и Contreras et al., 1991, Biotechnology 378-381; Eaton et al., 1986, Biochemistry 25: 505-512; Collins-Racie et al., 1995, Biotechnology 13: 982-987; Carter et al., 1989, Proteins: Structure, Function, and Genetics 6: 240-248; и Stevens, 2003, Drug Discovery World 4: 35-48.
Предпочтительно, полипептиды по настоящему изобретению являются трипсиноподобными эндопептидазами.
Источники полипептидов, обладающих эндопептидазной активностью
Полипептид, обладающий эндопептидазной активностью, по настоящему изобретению можно получать из микроорганизмов любого рода. Для целей настоящего изобретения используемый в данном описании термин «полученный из» в связи с данным источником должен означать, что полипептид, кодируемый полинуклеотидом, произведен при помощи источника или при помощи штамма, в который полинуклеотид из источника был встроен. В одном варианте осуществления полипептид, полученный из данного источника, секретируется внеклеточно.
Полипептид может быть бактериальным полипептидом. Например, полипептид может быть полипептидом из грамположительных бактерий, таким как полипептид из Actinocynnema, Bacillus, Clostridium, Enterococcus, Geobacillus, Kribbella, Kutzneria, Lactobacillus, Lactococcus, Oceanobacillus, Staphylococcus, Streptococcus или Streptomyces, обладающий эндопептидазной активностью, или полипептидом из грамотрицательных бактерий, таким как полипептид из Campylobacter, E. coli, Flavobacterium, Fusobacterium, Helicobacter, Ilyobacter, Neisseria, Pseudomonas, Salmonella или Ureaplasma.
В одном варианте осуществления полипептид представляет собой полипептид из Bacillus alkalophilus, Bacillus amyloliquefaciens, Bacillus brevis, Bacillus circulans, Bacillus clausii, Bacillus coagulans, Bacillus firmus, Bacillus lautus, Bacillus lentus, Bacillus licheniformis, Bacillus megaterium, Bacillus pumilus, Bacillus stearothermophilus, Bacillus subtilis или Bacillus thuringiensis.
В другом варианте осуществления полипептид представляет собой полипептид из Streptococcus equisimilis, Streptococcus pyogenes, Streptococcus uberis или Streptococcus equi подвид Zooepidemicus.
В другом варианте осуществления полипептид представляет собой полипептид из Streptomyces achromogenes, Streptomyces avermitilis, Streptomyces coelicolor, Streptomyces griseus или Streptomyces lividans.
Полипептид также может быть грибковым полипептидом. Например, полипептид может быть полипептидом из дрожжей, таким как полипептид из Candida, Kluyveromyces, Pichia, Saccharomyces, Schizosaccharomyces или Yarrowia; или полипептидом из нитчатых грибов, таким как полипептид из Acremonium, Agaricus, Alternaria, Aspergillus, Aureobasidium, Botryospaeria, Ceriporiopsis, Chaetomidium, Chrysosporium, Claviceps, Cochliobolus, Coprinopsis, Coptotermes, Corynascus, Cryphonectria, Cryptococcus, Diplodia, Exidia, Filibasidium, Fusarium, Gibberella, Holomastigotoides, Humicola, Irpex, Lentinula, Leptospaeria, Magnaporthe, Melanocarpus, Meripilus, Mucor, Myceliophthora, Neocallimastix, Neurospora, Paecilomyces, Penicillium, Phanerochaete, Piromyces, Poitrasia, Pseudoplectania, Pseudotrichonympha, Rhizomucor, Schizophyllum, Scytalidium, Talaromyces, Thermoascus, Thielavia, Tolypocladium, Trichoderma, Trichophaea, Verticillium, Volvariella или Xylaria.
В другом варианте осуществления полипептид представляет собой полипептид из Saccharomyces carlsbergensis, Saccharomyces cerevisiae, Saccharomyces diastaticus, Saccharomyces douglasii, Saccharomyces kluyveri, Saccharomyces norbensis или Saccharomyces oviformis.
В другом варианте осуществления полипептид представляет собой полипептид из Acremonium cellulolyticus, Aspergillus aculeatus, Aspergillus awamori, Aspergillus foetidus, Aspergillus fumigatus, Aspergillus japonicus, Aspergillus nidulans, Aspergillus niger, Aspergillus oryzae, Chrysosporium inops, Chrysosporium keratinophilum, Chrysosporium lucknowense, Chrysosporium merdarium, Chrysosporium pannicola, Chrysosporium queenslandicum, Chrysosporium tropicum, Chrysosporium zonatum, Fusarium bactridioides, Fusarium cerealis, Fusarium crookwellense, Fusarium culmorum, Fusarium graminearum, Fusarium graminum, Fusarium heterosporum, Fusarium negundi, Fusarium oxysporum, Fusarium reticulatum, Fusarium roseum, Fusarium sambucinum, Fusarium sarcochroum, Fusarium sporotrichioides, Fusarium sulphureum, Fusarium torulosum, Fusarium trichothecioides, Fusarium venenatum, Humicola grisea, Humicola insolens, Humicola lanuginosa, Irpex lacteus, Mucor miehei, Myceliophthora thermophila, Neurospora crassa, Penicillium funiculosum, Penicillium purpurogenum, Phanerochaete chrysosporium, Thielavia achromatica, Thielavia albomyces, Thielavia albopilosa, Thielavia australeinsis, Thielavia fimeti, Thielavia microspora, Thielavia ovispora, Thielavia peruviana, Thielavia setosa, Thielavia spededonium, Thielavia subthermophila, Thielavia terrestris, Trichoderma harzianum, Trichoderma koningii, Trichoderma longibrachiatum, Trichoderma reesei или Trichoderma viride.
В предпочтительном варианте осуществления полипептид представляет собой полипептид из Kutzneria, например, полипептид, полученный из Kutzneria albida, например, полипептид, полученный из Kutzneria albida ATCC 25243 (доступной из коллекции культур ATCC).
Следует иметь в виду, что для приведенных выше видов изобретение охватывает как совершенные, так и несовершенные стадии, а также другие таксономические эквиваленты, например, анаморфы, независимо от названия вида, под которым они известны. Специалисты в данной области легко установят принадлежность соответствующих эквивалентов.
Штаммы этих видов легкодоступны для общественности в ряде коллекций культур, таких как Американская коллекция типовых культур (ATCC), Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (DSM), Centraalbureau Voor Schimmelcultures (CBS) и Коллекция патентованных культур службы сельскохозяйственных исследований Северного регионального исследовательского центра (NRRL).
Полипептид можно идентифицировать и получать из других источников, включая микроорганизмы, выделенные из природных источников (например, почвы, компоста, воды и т.д.), используя приведенные выше зонды. Методы выделения микроорганизмов из естественных мест обитания хорошо известны в данной области. Затем полинуклеотид, кодирующий полипептид, можно получать путем аналогичного скрининга геномной или кДНК библиотеки другого микроорганизма или смешанного образца ДНК. После того, как полинуклеотид, кодирующий полипептид, был обнаружен при помощи зонда(ов), полинуклеотид можно выделять или клонировать с использованием методов, хорошо известных специалисту в данной области (см., например, Sambrook et al., 1989, выше).
Конструкты нуклеиновых кислот
Настоящее изобретение также относится к конструктам нуклеиновых кислот, содержащим полинуклеотид, кодирующий полипептид по настоящему изобретению, функционально связанный с одной или более (несколькими) контрольными последовательностями, которые направляют экспрессию кодирующей последовательности в подходящей клетке-хозяине в условиях, совместимых с контрольными последовательностями.
Полинуклеотид может быть подвергнут манипуляциям различными способами для обеспечения экспрессии полипептида. Манипуляции с полинуклеотидом до его введения в вектор могут быть желательными или необходимыми в зависимости от экспрессионного вектора. Способы изменения полинуклеотидов с использованием методов рекомбинантной ДНК хорошо известны в данной области.
Контрольная последовательность может представлять собой последовательность промотора, полинуклеотида, который распознается клеткой-хозяином для экспрессии полинуклеотида, кодирующего полипептид по настоящему изобретению. Последовательность промотора содержит контрольные последовательности транскрипции, которые опосредуют экспрессию полипептида. Промотор может быть любым полинуклеотидом, проявляющим транскрипционную активность в выбранной клетке-хозяине, включая мутантные, укороченные и гибридные промоторы, и может быть получен из генов, кодирующих внеклеточные или внутриклеточные полипептиды, либо гомологичные, либо гетерологичные для клетки-хозяина.
Примерами подходящих промоторов для направления транскрипции конструктов нуклеиновых кислот по настоящему изобретению в бактериальной клетке-хозяине являются промоторы, полученные из гена альфа-амилазы (amyQ) Bacillus amyloliquefaciens, гена альфа-амилазы (amyL) Bacillus licheniformis, гена пенициллиназы (penP) Bacillus licheniformis, гена мальтогенной амилазы (amyM) Bacillus stearothermophilus, гена левансукразы (sacB) Bacillus subtilis, генов xylA и xylB Bacillus subtilis, lac-опрерона E. coli, гена агаразы (dagA) Streptomyces coelicolor и гена прокариотической бета-лактамазы (Villa-Kamaroff et al., 1978, Proc. Natl. Acad. Sci. USA 75: 3727-3731), а также tac-промотор (DeBoer et al., 1983, Proc. Natl. Acad. Sci. USA 80: 21-25). Дополнительные промоторы описаны в «Useful proteins from recombinant bacteria» в Gilbert et al., 1980, Scientific American, 242: 74-94; и в Sambrook et al., 1989, выше.
Примерами подходящих промоторов для направления транскрипции конструктов нуклеиновых кислот по настоящему изобретению в клетке-хозяине нитчатых грибов являются промоторы, полученные из генов ацетамидазы Aspergillus nidulans, нейтральной альфа-амилазы Aspergillus niger, кислотостабильной альфа-амилазы Aspergillus niger, глюкоамилазы (glaA) Aspergillus niger или Aspergillus awamori, TAKA-амилазы Aspergillus oryzae, щелочной протеазы Aspergillus oryzae, триозофосфатизомеразы Aspergillus oryzae, трипсиноподобной протеазы Fusarium oxysporum (WO 96/00787), амилоглюкозидазы Fusarium venenatum (WO 00/56900), Daria Fusarium venenatum (WO 00/56900), Quinn Fusarium venenatum (WO 00/56900), липазы Rhizomucor miehei, аспарагиновой протеиназы Rhizomucor miehei, бета-глюкозидазы Trichoderma reesei, целлобиогидролазы I Trichoderma reesei, целлобиогидролазы II Trichoderma reesei, эндоглюканазы I Trichoderma reesei, эндоглюканазы II Trichoderma reesei, эндоглюканазы III Trichoderma reesei, эндоглюканазы IV Trichoderma reesei, эндоглюканазы V Trichoderma reesei, ксиланазы I Trichoderma reesei, ксиланазы II Trichoderma reesei, бета-ксилозидазы Trichoderma reesei, а также промотор NA2-tpi (модифицированный промотор, включающий ген, кодирующий нейтральную альфа-амилазу в Aspergilli, в котором нетранслируемая лидерная последовательность была заменена нетранслируемой лидерной последовательностью из гена, кодирующего триозофосфатизомеразу в Aspergilli; не ограничивающие примеры включают модифицированные промоторы, включая ген, кодирующий нейтральную альфа-амилазу в Aspergillus niger, в котором нетранслируемая лидерная последовательность была заменена нетранслируемой лидерной последовательностью из гена, кодирующего триозофосфатизомеразу в Aspergillus nidulans или Aspergillus oryzae); а также полученные из них мутантные, укороченные и гибридные промоторы.
Для дрожжевого хозяина полезные промоторы получают из генов энолазы (ENO-1) Saccharomyces cerevisiae, галактокиназы (GAL1) Saccharomyces cerevisiae, алкогольдегидрогеназы/глицеральдегид-3-фосфатдегидрогеназы (ADH1, ADH2/GAP) Saccharomyces cerevisiae, триозофосфатизомеразы (TPI) Saccharomyces cerevisiae, металлотионеина (CUP1) Saccharomyces cerevisiae и 3-фосфоглицераткиназы Saccharomyces cerevisiae. Другие полезные промоторы для дрожжевых клеток-хозяев описаны Romanos et al., 1992, Yeast 8: 423-488.
Контрольная последовательность может также представлять собой подходящую последовательность терминатора транскрипции, которая распознается клеткой-хозяином для терминации транскрипции. Последовательность терминатора функционально связана с 3'-концом полинуклеотида, кодирующего полипептид. Любой терминатор, который функционален в выбранной клетке-хозяине, можно использовать по настоящему изобретению.
Предпочтительные терминаторы для клеток-хозяев нитчатых грибов получают из генов антранилатсинтазы Aspergillus nidulans, глюкоамилазы Aspergillus niger, альфа-глюкозидазы Aspergillus niger, TAKA-амилазы Aspergillus oryzae и трипсиноподобной протеазы Fusarium oxysporum.
Предпочтительные терминаторы для дрожжевых клеток-хозяев получают из генов энолазы Saccharomyces cerevisiae, цитохрома C (CYC1) Saccharomyces cerevisiae и глицеральдегид-3-фосфатдегидрогеназы Saccharomyces cerevisiae. Другие полезные терминаторы для дрожжевых клеток-хозяев описаны Romanos et al., 1992, выше.
Контрольная последовательность может также представлять собой подходящую лидерную последовательность, когда транскрибируется нетранслируемая область мРНК, которая важна для трансляции в клетке-хозяине. Лидерная последовательность функционально связана с 5'-концом полинуклеотида, кодирующего полипептид. Можно использовать любую лидерную последовательность, которая функциональна в клетке-хозяине.
Предпочтительные лидерные последовательности для клеток-хозяев нитчатых грибов получают из генов TAKA-амилазы Aspergillus oryzae и триозофосфатизомеразы Aspergillus nidulans.
Подходящие лидерные последовательности для дрожжевых клеток-хозяев получают из генов энолазы (ENO-1) Saccharomyces cerevisiae, 3-фосфоглицераткиназы Saccharomyces cerevisiae, альфа-фактора Saccharomyces cerevisiae и алкогольдегидрогеназы/глицеральдегид-3-фосфатдегидрогеназы (ADH2/GAP) Saccharomyces cerevisiae.
Контрольная последовательность может также представлять собой последовательность полиаденилирования, последовательность, функционально связанную с 3'-концом полинуклеотида, которая, будучи транскрибированной, распознается клеткой-хозяином как сигнал для добавления остатков полиаденозина к транскрибированной мРНК. Можно использовать любую последовательность полиаденилирования, которая функциональна в выбранной клетке-хозяине.
Предпочтительные последовательности полиаденилирования для клеток-хозяев нитчатых грибов получают из генов TAKA-амилазы Aspergillus oryzae, глюкоамилазы Aspergillus niger, антранилатсинтазы Aspergillus nidulans, трипсиноподобной протеазы Fusarium oxysporum и альфа-глюкозидазы Aspergillus niger.
Полезные последовательности полиаденилирования для дрожжевых клеток-хозяев описаны Guo and Sherman, 1995, Mol. Cellular Biol. 15: 5983-5990.
Контрольная последовательность может также представлять собой кодирующую сигнальный пептид область, которая кодирует сигнальный пептид, связанный с N-концом полипептида и направляющий полипептид в секреторный путь клетки. 5'-конец кодирующей последовательности полинуклеотида может исходно содержать последовательность, кодирующую сигнальный пептид, природно-связанную в трансляционной рамке считывания с сегментом кодирующей последовательности, который кодирует полипептид. Альтернативно, 5'-конец кодирующей последовательности может содержать последовательность, кодирующую сигнальный пептид, которая является чужеродной для кодирующей последовательности. Чужеродная кодирующая сигнальный пептид последовательность может быть необходима, если кодирующая последовательность природно не содержит последовательность, кодирующую сигнальный пептид. Альтернативно, чужеродной кодирующей сигнальный пептид последовательностью можно просто заменять природную кодирующую сигнальный пептид последовательность для увеличения секреции полипептида. Однако можно использовать любую кодирующую сигнальный пептид последовательность, которая направляет экспрессированный полипептид в секреторный путь выбранной клетки-хозяина.
Эффективными кодирующими сигнальный пептид последовательностями для бактериальных клеток-хозяев являются кодирующие сигнальный пептид последовательности, полученные из генов мальтогенной амилазы Bacillus NCIB 11837, субтилизина Bacillus licheniformis, бета-лактамазы Bacillus licheniformis, альфа-амилазы Bacillus stearothermophilus, нейтральных протеаз (nprT, nprS, nprM) Bacillus stearothermophilus и prsA Bacillus subtilis. Дополнительные сигнальные пептиды описаны Simonen and Palva, 1993, Microbiological Reviews 57: 109-137.
Эффективными кодирующими сигнальный пептид последовательностями для клеток-хозяев нитчатых грибов являются кодирующие сигнальный пептид последовательности, полученные из генов нейтральной амилазы Aspergillus niger, глюкоамилазы Aspergillus niger, TAKA-амилазы Aspergillus oryzae, целлюлазы Humicola insolens, эндоглюканазы V Humicola insolens, липазы Humicola lanuginosa и аспарагиновой протеиназы Rhizomucor miehei.
Полезные сигнальные пептиды для дрожжевых клеток-хозяев получают из генов альфа-фактора Saccharomyces cerevisiae и инвертазы Saccharomyces cerevisiae. Другие полезные кодирующие сигнальный пептид последовательности описаны Romanos et al., 1992, выше.
Контрольная последовательность может также представлять собой последовательность, кодирующую пропептид, которая кодирует пропептид, расположенный на N-конце полипептида. Полученный в результате полипептид известен как профермент или прополипептид (или зимоген в некоторых случаях). Прополипептид, как правило, неактивен и может превращаться в активный полипептид в результате каталитического или аутокаталитического отщепления пропептида от прополипептида. Кодирующую пропептид последовательность можно получать из генов щелочной протеазы (aprE) Bacillus subtilis, нейтральной протеазы (nprT) Bacillus subtilis, лакказы Myceliophthora thermophila (WO 95/33836), аспарагиновой протеиназы Rhizomucor miehei и альфа-фактора Saccharomyces cerevisiae.
Если обе последовательности для сигнального пептида и для пропептида присутствуют на N-конце полипептида, последовательность пропептида расположена рядом с N-концом полипептида, и последовательность сигнального пептида расположена рядом с N-концом последовательности пропептида.
Может также оказаться желательным добавлять регуляторные последовательности, позволяющие регулировать экспрессию полипептида в зависимости от роста клетки-хозяина. Примерами регуляторных систем являются такие, которые вызывают запуск или прекращение экспрессии гена в ответ на химический или физический стимул, включая присутствие регулирующего соединения. Регуляторные системы в прокариотических системах включают системы операторов lac, tac и trp. В дрожжах можно использовать систему ADH2 или систему GAL1. В нитчатых грибах можно использовать промотор глюкоамилазы Aspergillus niger, промотор TAKA альфа-амилазы Aspergillus oryzae и промотор глюкоамилазы Aspergillus oryzae. Другими примерами регуляторных последовательностей являются такие, которые делают возможной амплификацию гена. В эукариотических системах эти регуляторные последовательности включают ген дигидрофолатредуктазы, который амплифицируется в присутствии метотрексата, и гены металлотионеинов, которые амплифицируются в присутствии тяжелых металлов. В этих случаях, полинуклеотид, кодирующий полипептид, будет функционально связан с регуляторной последовательностью.
Экспрессионные векторы
Настоящее изобретение также относится к рекомбинантным экспрессионным векторам, содержащим полинуклеотид, кодирующий полипептид по настоящему изобретению, промотор, а также стоп-сигналы транскрипции и трансляции. Различные нуклеотидные и контролирующие последовательности можно соединять вместе для получения рекомбинантного экспрессионного вектора, который может содержать один или более (несколько) удобных сайтов рестрикции, позволяющих производить вставку или замену полинуклеотида, кодирующего полипептид, в такие сайты. Альтернативно, полинуклеотид может быть экспрессирован путем вставки полинуклеотида или конструкта нуклеиновой кислоты, содержащего последовательность, в соответствующий вектор для экспрессии. При создании экспрессионного вектора кодирующую последовательность располагают в векторе таким образом, что кодирующая последовательность является функционально связанной с соответствующими контрольными последовательностями для экспрессии.
Рекомбинантный экспрессионный вектор может быть любым вектором (например, плазмидой или вирусом), к которому можно легко применять методы рекомбинантной ДНК, и который может обеспечивать экспрессию полинуклеотида. Выбор вектора, как правило, зависит от совместимости вектора с клеткой-хозяином, в которую предстоит вводить вектор. Вектор может быть линейной или замкнутой кольцевой плазмидой.
Вектор может представлять собой автономно реплицирующийся вектор, то есть, вектор, который существует в виде внехромосомной структуры, репликация которой не зависит от хромосомной репликации, например, плазмиду, внехромосомный элемент, минихромосому или искусственную хромосому. Вектор может содержать любые средства для обеспечения саморепликации. Альтернативно, вектор может представлять собой вектор, который, будучи введен в клетку-хозяина, встраивается в геном и реплицируется вместе с хромосомой(ами), в которую он интегрирован. Кроме того, можно использовать один вектор или плазмиду, либо два или более векторов или плазмид, которые вместе содержат общую ДНК, которая должна быть введена в геном клетки-хозяина, либо транспозон.
Предпочтительно, вектор содержит один или более (несколько) селектируемых маркеров, позволяющих легко отбирать трансформированные, трансфицированные, трансдуцированные и подобные клетки. Селектируемый маркер представляет собой ген, продукт которого обеспечивает резистентность к биоцидам или вирусам, устойчивость к тяжелым металлам, прототрофию ауксотрофам и тому подобное.
Примерами бактериальных селектируемых маркеров являются гены dal из Bacillus subtilis или Bacillus licheniformis, либо маркеры, придающие устойчивость к антибиотикам, например, устойчивость к ампициллину, хлорамфениколу, канамицину или тетрациклину. Подходящими маркерами для дрожжевых клеток-хозяев являются ADE2, HIS3, LEU2, LYS2, MET3, TRP1 и URA3. Селектируемые маркеры для использования в клетках-хозяевах нитчатых грибов включают, но не ограничиваются ими, amdS (ацетамидаза), argB (орнитинкарбамоилтрансфераза), bar (фосфинотрицинацетилтрансфераза), hph (гигромицинфосфотрансфераза), niaD (нитратредуктаза), pyrG (оротидин-5'-фосфатдекарбоксилаза), sC (сульфатаденилтрансфераза) и trpC (антранилатсинтаза), а также их эквиваленты. Предпочтительными для использования в клетках Aspergillus являются гены amdS и pyrG из Aspergillus nidulans или Aspergillus oryzae и ген bar из Streptomyces hygroscopicus.
Предпочтительно, вектор содержит элемент(ы), делающий возможной интеграцию вектора в геном клетки-хозяина или автономную репликацию вектора в клетке независимо от генома.
Для интеграции в геном клетки-хозяина вектор может использовать полинуклеотидную последовательность, кодирующую полипептид, или любой другой элемент вектора для интеграции в геном путем гомологичной или негомологичной рекомбинации. Альтернативно, вектор может содержать дополнительные полинуклеотиды для направления интеграции путем гомологичной рекомбинации в геном клетки-хозяина в точном месте(ах) хромосомы(хромосом). Для повышения вероятности интеграции в точном месте, интеграционные элементы должны содержать достаточное количество нуклеиновых кислот, например, от 100 до 10000 пар оснований, от 400 до 10000 пар оснований и от 800 до 10000 пар оснований, которые имеют высокую степень идентичности последовательности с соответствующей целевой последовательностью для повышения вероятности гомологичной рекомбинации. Интеграционные элементы могут представлять собой любую последовательность, которая гомологична целевой последовательности в геноме клетки-хозяина. Кроме того, интеграционные элементы могут представлять собой некодирующие или кодирующие полинуклеотиды. С другой стороны, вектор может быть интегрирован в геном клетки-хозяина путем негомологичной рекомбинации.
Для автономной репликации вектор может дополнительно содержать точку начала репликации, которая позволяет вектору реплицироваться автономно в нужной клетке-хозяине. Точкой начала репликации может быть любой плазмидный репликатор, опосредующий автономную репликацию, который функционирует в клетке. Термин «точка начала репликации» или «плазмидный репликатор» означает полинуклеотид, который позволяет плазмиде или вектору реплицироваться in vivo.
Примерами бактериальных точек начала репликации являются точки начала репликации плазмид pBR322, pUC19, pACYC177 и pACYC184, допускающие репликацию в E. coli, и pUB110, pE194, pTA1060 и pΑΜβ1, допускающие репликацию в Bacillus.
Примерами точек начала репликации для использования в дрожжевых клетках-хозяевах являются 2-микронная точка начала репликации, ARS1, ARS4, сочетание ARS1 и CEN3, а также сочетание ARS4 и CEN6.
Примерами точек начала репликации, применимых в клетках нитчатых грибов, являются AMA1 и ANS1 (Gems et al., 1991, Gene 98: 61-67; Cullen et al., 1987, Nucleic Acids Res. 15: 9163-9175; WO 00/24883). Выделение гена AMA1 и конструирование плазмид или векторов, содержащих этот ген, можно осуществлять в соответствии с методами, раскрытыми в WO 00/24883.
Для увеличения продукции полипептида в клетку-хозяина можно вводить более одной копии полинуклеотида по настоящему изобретению. Увеличения числа копий полинуклеотида можно добиваться путем интегрирования по меньшей мере одной дополнительной копии последовательности в геном клетки-хозяина или путем включения амплифицируемого гена селектируемого маркера с полинуклеотидом, при этом клетки, содержащие амплифицированные копии гена селектируемого маркера и, следовательно, дополнительные копии полинуклеотида, можно отбирать путем культивирования клеток в присутствии соответствующего селекционного средства.
Методы, используемые для лигирования элементов, описанных выше, с целью конструирования рекомбинантных экспрессионных векторов по настоящему изобретению, хорошо известны специалистам в данной области (см., например, Sambrook et al., 1989, выше).
Клетки-хозяева
Настоящее изобретение также относится к рекомбинантным клеткам-хозяевам, содержащим полинуклеотид, кодирующий полипептид по настоящему изобретению, функционально связанный с одной или более (несколькими) контрольными последовательностями, которые направляют продукцию полипептида по настоящему изобретению. Конструкт или вектор, содержащий полинуклеотид, вводят в клетку-хозяина таким образом, что конструкт или вектор сохраняется в виде составной части хромосомы или в виде самореплицирующегося внехромосомного вектора, как описано выше. Термин «клетка-хозяин» охватывает любое потомство родительской клетки, которое не идентично родительской клетке вследствие мутаций, имеющих место в процессе репликации. Выбор клетки-хозяина будет в значительной степени зависеть от гена, кодирующего полипептид, и его источника.
Клетка-хозяин может быть любой клеткой, применимой для рекомбинантной продукции полипептида по настоящему изобретению, например, прокариотической или эукариотической.
Прокариотическая клетка-хозяин может быть любой грамположительной или грамотрицательной бактерией. Грамположительные бактерии включают, но не ограничиваются ими, Bacillus, Clostridium, Enterococcus, Geobacillus, Lactobacillus, Lactococcus, Oceanobacillus, Staphylococcus, Streptococcus и Streptomyces. Грамотрицательные бактерии включают, но не ограничиваются ими, Campylobacter, E. coli, Flavobacterium, Fusobacterium, Helicobacter, Ilyobacter, Neisseria, Pseudomonas, Salmonella и Ureaplasma.
Бактериальная клетка-хозяин может быть любой клеткой Bacillus, включая, но, не ограничиваясь ими, клетки Bacillus alkalophilus, Bacillus amyloliquefaciens, Bacillus brevis, Bacillus circulans, Bacillus clausii, Bacillus coagulans, Bacillus firmus, Bacillus lautus, Bacillus lentus, Bacillus licheniformis, Bacillus megaterium, Bacillus pumilus, Bacillus stearothermophilus, Bacillus subtilis и Bacillus thuringiensis.
Бактериальная клетка-хозяин может также быть любой клеткой Streptococcus, включая, но, не ограничиваясь ими, клетки Streptococcus equisimilis, Streptococcus pyogenes, Streptococcus uberis и Streptococcus equi подвид Zooepidemicus.
Бактериальная клетка-хозяин может также быть любой клеткой Streptomyces, включая, но, не ограничиваясь ими, клетки Streptomyces achromogenes, Streptomyces avermitilis, Streptomyces coelicolor, Streptomyces griseus и Streptomyces lividans.
Введение ДНК в клетку Bacillus можно, например, осуществлять путем трансформации протопластов (см., например, Chang and Cohen, 1979, Mol. Gen. Genet. 168: 111-115), путем использования компетентных клеток (см., например, Young and Spizizen, 1961, J. Bacteriol. 81: 823-829, или Dubnau and Davidoff-Abelson, 1971, J. Mol. Biol. 56: 209-221), путем электропорации (см., например, Shigekawa and Dower, 1988, Biotechniques 6: 742-751) или путем конъюгации (см., например, Koehler and Thorne, 1987, J. Bacteriol. 169: 5271-5278). Введение ДНК в клетку E. coli можно, например, осуществлять путем трансформации протопластов (см., например, Hanahan, 1983, J. Mol. Biol. 166: 557-580) или электропорации (см., например, Dower et al., 1988, Nucleic Acids Res. 16: 6127-6145). Введение ДНК в клетку Streptomyces можно, например, осуществлять путем трансформации протопластов и электропорации (см., например, Gong et al., 2004, Folia Microbiol. (Praha) 49: 399-405), путем конъюгации (см., например, Mazodier et al., 1989, J. Bacteriol. 171: 3583-3585) или путем трансдукции (см., например, Burke et al., 2001, Proc. Natl. Acad. Sci. USA 98: 6289-6294). Введение ДНК в клетку Pseudomonas можно, например, осуществлять путем электропорации (см., например, Choi et al., 2006, J. Microbiol. Methods 64: 391-397) или путем конъюгации (см., например, Pinedo and Smets, 2005, Appl. Environ. Microbiol. 71: 51-57). Введение ДНК в клетку Streptococcus можно, например, осуществлять путем естественной компетенции (см., например, Perry and Kuramitsu, 1981, Infect. Immun. 32: 1295-1297), путем трансформации протопластов (см., например, Catt and Jollick, 1991, Microbios 68: 189-207), путем электропорации (см., например, Buckley et al., 1999, Appl. Environ. Microbiol. 65: 3800-3804) или путем конъюгации (см., например, Clewell, 1981, Microbiol. Rev. 45: 409-436). Однако можно использовать любой метод, известный в данной области, для введения ДНК в клетку-хозяина.
Клетка-хозяин также может быть эукариотической, например, клеткой млекопитающего, насекомого, растения или гриба.
Клетка-хозяин может быть клеткой гриба. Используемый в данном описании термин «грибы» включает тип Ascomycota, Basidiomycota, Chytridiomycota и Zygomycota (как определено Hawksworth et al., в Ainsworth and Bisby's Dictionary of The Fungi, 8th edition, 1995, CAB International, University Press, Cambridge, UK), а также Oomycota (как указано Hawksworth et al., 1995, выше, станица 171) и все митоспоровые грибы (Hawksworth et al., 1995, выше).
Грибная клетка-хозяин может быть дрожжевой клеткой. Используемый в данном описании термин «дрожжи» включает аскоспорогенные дрожжи (Endomycetales), базидиоспорогенные дрожжи и дрожжи, принадлежащие к несовершенным грибам (Blastomycetes). Поскольку классификация дрожжей может измениться в будущем, для целей настоящего изобретения дрожжи должны быть определены, как описано в Biology and Activities of Yeast (Skinner, F.A., Passmore, S.M., and Davenport, R.R., eds, Soc. App. Bacteriol. Symposium Series No. 9, 1980).
Дрожжевая клетка-хозяин может быть клеткой Candida, Hansenula, Kluyveromyces, Pichia, Saccharomyces, Schizosaccharomyces или Yarrowia, такой как клетка Kluyveromyces lactis, Saccharomyces carlsbergensis, Saccharomyces cerevisiae, Saccharomyces diastaticus, Saccharomyces douglasii, Saccharomyces kluyveri, Saccharomyces norbensis, Saccharomyces oviformis или Yarrowia lipolytica.
Грибная клетка-хозяин может быть клеткой нитчатого гриба. «Нитчатые грибы» включают все нитчатые формы подотдела Eumycota и Oomycota (как определено Hawksworth et al., 1995, выше). Нитчатые грибы, как правило, характеризуются мицелиальной стенкой, состоящей из хитина, целлюлозы, глюкана, хитозана, маннана и других сложных полисахаридов. Вегетативный рост осуществляется путем удлинения гифов, и углеродный катаболизм является облигатно аэробным. Напротив, вегетативный рост дрожжей, таких как Saccharomyces cerevisiae, осуществляется путем почкования одноклеточного слоевища, и углеродный катаболизм может быть ферментативным.
Клетка-хозяин нитчатого гриба может представлять собой клетку Acremonium, Aspergillus, Aureobasidium, Bjerkandera, Ceriporiopsis, Chrysosporium, Coprinus, Coriolus, Cryptococcus, Filibasidium, Fusarium, Humicola, Magnaporthe, Mucor, Myceliophthora, Neocallimastix, Neurospora, Paecilomyces, Penicillium, Phanerochaete, Phlebia, Piromyces, Pleurotus, Schizophyllum, Talaromyces, Thermoascus, Thielavia, Tolypocladium, Trametes или Trichoderma.
Например, клетка-хозяин нитчатого гриба может представлять собой клетку Aspergillus awamori, Aspergillus foetidus, Aspergillus fumigatus, Aspergillus japonicus, Aspergillus nidulans, Aspergillus niger, Aspergillus oryzae, Bjerkandera adusta, Ceriporiopsis aneirina, Ceriporiopsis caregiea, Ceriporiopsis gilvescens, Ceriporiopsis pannocinta, Ceriporiopsis rivulosa, Ceriporiopsis subrufa, Ceriporiopsis subvermispora, Chrysosporium inops, Chrysosporium keratinophilum, Chrysosporium lucknowense, Chrysosporium merdarium, Chrysosporium pannicola, Chrysosporium queenslandicum, Chrysosporium tropicum, Chrysosporium zonatum, Coprinus cinereus, Coriolus hirsutus, Fusarium bactridioides, Fusarium cerealis, Fusarium crookwellense, Fusarium culmorum, Fusarium graminearum, Fusarium graminum, Fusarium heterosporum, Fusarium negundi, Fusarium oxysporum, Fusarium reticulatum, Fusarium roseum, Fusarium sambucinum, Fusarium sarcochroum, Fusarium sporotrichioides, Fusarium sulphureum, Fusarium torulosum, Fusarium trichothecioides, Fusarium venenatum, Humicola insolens, Humicola lanuginosa, Mucor miehei, Myceliophthora thermophila, Neurospora crassa, Penicillium purpurogenum, Phanerochaete chrysosporium, Phlebia radiata, Pleurotus eryngii, Thielavia terrestris, Trametes villosa, Trametes versicolor, Trichoderma harzianum, Trichoderma koningii, Trichoderma longibrachiatum, Trichoderma reesei или Trichoderma viride.
Клетки грибов можно трансформировать методом, включающим образование протопластов, трансформацию протопластов и регенерацию клеточной стенки известными способами. Подходящие методы трансформации клеток-хозяев Aspergillus и Trichoderma описаны в EP 238023 и статье Yelton et al., 1984, Proc. Natl. Acad. Sci. USA 81: 1470-1474. Подходящие методы трансформации вида Fusarium описаны Malardier et al., 1989, в Gene 78: 147-156, и WO 96/00787. Дрожжи можно трансформировать методами, описанными Becker and Guarente, в Abelson, J.N. and Simon, M.I., редакторы, Guide to Yeast Genetics and Molecular Biology, Methods in Enzymology, том 194, страницы 182-187, Academic Press, Inc., New York; Ito et al., 1983, J. Bacteriol. 153: 163; и Hinnen et al., 1978, Proc. Natl. Acad. Sci. USA 75: 1920.
Способы получения
Настоящее изобретение также относится к способам получения полипептида по настоящему изобретению, включающим: (a) культивирование клетки, которая в своей форме дикого типа продуцирует полипептид, в условиях, благоприятных для продукции полипептида; и (b) выделение полипептида. В предпочтительном варианте осуществления клетка относится к роду Kutzneria. В более предпочтительном варианте осуществления клетка представляет собой Kutzneria albida. В наиболее предпочтительном варианте осуществления клетка представляет собой Kutzneria albida ATCC 25243.
Настоящее изобретение также относится к способам получения полипептида по настоящему изобретению, включающим: (a) культивирование рекомбинантной клетки-хозяина по настоящему изобретению в условиях, благоприятных для продукции полипептида; и (b) выделение полипептида.
Клетки-хозяева культивируют в питательной среде, подходящей для продукции полипептида, используя методы, хорошо известные в данной области. Например, клетки можно культивировать методом культивирования со встряхиванием колбы, а также методом мелкомасштабной или крупномасштабной ферментации (включая непрерывную, периодическую, периодическую с подпиткой или твердофазную ферментации) в лабораторных или промышленных ферментерах, проводя культивирование в соответствующей среде и в условиях, позволяющих экспрессировать и/или выделять полипептид. Культивирование проводят в подходящей питательной среде, содержащей источники углерода и азота и неорганические соли, с использованием методов, известных в данной области. Подходящие среды доступны от коммерческих поставщиков или могут быть получены в соответствии с опубликованными составами (например, в каталогах Американской коллекции типовых культур). Если полипептид секретируется в питательную среду, полипептид можно выделять непосредственно из среды. Если полипептид не секретируется, его можно выделять из клеточных лизатов.
Полипептид можно обнаруживать методами, известными в данной области, которые являются специфическими для полипептидов. Эти методы обнаружения могут включать использование специфических антител, образование ферментного продукта или исчезновение ферментного субстрата. Например, можно использовать ферментный анализ для определения активности полипептида.
Полипептид можно выделять методами, известными в данной области. Например, полипептид можно выделять из питательной среды общепринятыми методами, включая, но, не ограничиваясь ими, центрифугирование, фильтрацию, экстракцию, сушку распылением, выпаривание или осаждение.
Полипептид можно очищать различными методами, известными в данной области, включая, но, не ограничиваясь ими, хроматографию (например, ионообменную, аффинную, гидрофобную, хроматофокусирование и эксклюзионную), электрофоретические методы (например, препаративное изоэлектрофокусирование), дифференциальную растворимость (например, осаждение сульфатом аммония), SDS-ПААГ или экстракцию (см., например, Protein Purification, J.-C. Janson and Lars Ryden, редакторы, VCH Publishers, New York, 1989) для получения по существу чистых полипептидов.
В альтернативном варианте осуществления полипептид не выделяют, а вместо этого клетку-хозяина по настоящему изобретению, экспрессирующую полипептид, используют в качестве источника полипептида.
Растения
Настоящее изобретение также относится к растениям, например, трансгенному растению, части растения или растительной клетке, содержащим выделенный полинуклеотид по настоящему изобретению таким образом, чтобы экспрессировать и продуцировать полипептид в количествах, которые можно выделять. Полипептид можно выделять из растения или части растения. Альтернативно, растение или части растения, содержащие полипептид, можно использовать как таковые для повышения качества продовольствия или корма, например повышения питательной ценности, вкусовых и реологических свойств или для разрушения анти-питательного фактора.
Трансгенное растение может быть двудольным (двудольное растение) или однодольным (однодольное растение). Примерами однодольных растений являются травы, такие как луговые травы (мятлик, Poa), кормовые травы, такие как Festuca, Lolium, травы умеренной климатической зоны, такие как Agrostis, и зерновые, такие как пшеница, овес, рожь, ячмень, рис, сорго и маис (кукуруза).
Примерами двудольных растений являются табак, бобовые, такие как люпин, картофель, сахарная свекла, горох, бобы и соя, а также крестоцветные растения (семейство Brassicaceae), такие как цветная капуста, рапс и близкородственный модельный организм Arabidopsis thaliana.
Примерами частей растения являются стебель, каллус, листья, корни, плоды, семена и клубни, а также отдельные ткани, содержащие эти части, например, эпидермис, мезофилл, паренхима, сосудистая ткань, меристемы. Конкретные компартменты растительной клетки, такие как хлоропласты, апопласты, митохондрии, вакуоли, пероксисомы и цитоплазма, также считаются частями растения. Кроме того, любая растительная клетка, независимо от тканевого происхождения, считается частью растения. Кроме того, части растений, такие как конкретные ткани и клетки, выделенные для облегчения использования изобретения, также считаются частями растения, например, зародыши, эндоспермы, алейрон и оболочки семян.
Также в объем настоящего изобретения входит потомство таких растений, частей растений и растительных клеток.
Трансгенное растение или растительную клетку, экспрессирующую полипептид, можно конструировать методами, известными в данной области. Кратко, растение или растительную клетку конструируют путем включения одного или более (нескольких) экспрессионных конструктов, кодирующих полипептид, в геном растения-хозяина или геном хлоропластов и репродуцирования полученного модифицированного растения или растительной клетки в трансгенное растение или растительную клетку.
Удобно, если экспрессионный конструкт представляет собой конструкт нуклеиновой кислоты, который содержит полинуклеотид, кодирующий полипептид, функционально связанный с соответствующими регуляторными последовательностями, необходимыми для экспрессии полинуклеотида в выбранном растении или части растения. Кроме того, экспрессионный конструкт может содержать селектируемый маркер, полезный для идентификации клеток-хозяев, в которые был введен экспрессионный конструкт, и последовательности ДНК, необходимые для введения конструкта в представляющее интерес растение (последние зависят от способа введения ДНК, который будет использован).
Выбор регуляторных последовательностей, таких как последовательности промотора и терминатора, и необязательно сигнальных или транзитных последовательностей определяется, например, на основании того, когда, где и как полинуклеотид должен быть экспрессирован. Например, экспрессия гена, кодирующего полипептид, может быть конститутивной или индуцируемой, или может быть зависимой от развития, специфичной для стадии развития или ткани, и генный продукт может быть направлен к определенной ткани или части растения, такой как семена или листья. Регуляторные последовательности, например, описаны Tague et al., 1988, в Plant Physiology 86: 506.
Для конститутивной экспрессии можно использовать промотор 35S-CaMV, убиквитина 1 кукурузы и актина 1 риса (Franck et al., 1980, Cell 21: 285-294; Christensen et al., 1992, Plant Mol. Biol. 18: 675-689; Zhang et al., 1991, Plant Cell 3: 1155-1165). Орган-специфическим промотором может быть, например, промотор из накапливающих запасные вещества запасающих тканей растения, таких как семена, клубни картофеля и фрукты (Edwards and Coruzzi, 1990, Ann. Rev. Genet. 24: 275-303), или из метаболических запасающих тканей, таких как меристемы (Ito et al., 1994, Plant Mol. Biol. 24: 863-878), промотор, специфичный для семян, такой как промотор глютелина, проламина, глобулина или альбумина из риса (Wu et al., 1998, Plant Cell Physiol. 39: 885-889), промотор из легумина B4 из Vicia faba и гена неизвестного белка семян из Vicia faba (Conrad et al., 1998, J. Plant Physiol. 152: 708-711), промотор из белка корпуса масличных семян (Chen et al., 1998, Plant Cell Physiol. 39: 935-941), промотор запасного белка napA из Brassica napus, или любой другой специфичный для семян промотор, известный в данной области, например, описанный в WO 91/14772. Кроме того, промотор может представлять собой специфичный для листьев промотор, такой как промотор rbcs из риса или томата (Kyozuka et al., 1993, Plant Physiol. 102: 991-1000), промотор гена аденинметилтрансферазы вируса хлореллы (Mitra and Higgins, 1994, Plant Mol. Biol. 26: 85-93), промотор гена aldP из риса (Kagaya et al., 1995, Mol. Gen. Genet. 248: 668-674), или индуцируемый ранением промотор, такой как промотор pin2 картофеля (Xu et al., 1993, Plant Mol. Biol. 22: 573-588). Кроме того, промотор может быть индуцируемым абиотическими воздействиями, такими как температура, засуха или изменение солености, либо индуцируемым экзогенно применяемыми веществами, которые активируют промотор, например, этанолом, эстрогенами, растительными гормонами, такими как этилен, абсцизовая кислота и гиббереллиновая кислота, а также тяжелыми металлами.
Можно также использовать элемент энхансер промотора для достижения более высокой экспрессии полипептида в растении. Например, элемент энхансер промотора может представлять собой интрон, который находится между промотором и полинуклеотидом, кодирующим полипептид. Например, Xu et al., 1993, выше, описано применение первого интрона гена актина 1 риса для усиления экспрессии.
Ген селектируемого маркера и любые другие части экспрессионного конструкта можно выбирать из тех, что доступны в данной области.
Конструкт нуклеиновой кислоты встраивают в геном растения общепринятыми методами, известными в данной области, включая опосредованную Agrobacterium трансформацию, опосредованную вирусом трансформацию, микроинъекции, бомбардировку частицами, биолистическую трансформацию и электропорацию (Gasser et al., 1990, Science 244: 1293; Potrykus, 1990, Bio/Technology 8: 535; Shimamoto et al., 1989, Nature 338: 274).
В настоящее время, опосредованный Agrobacterium tumefaciens перенос генов является предпочтительным способом создания трансгенных двудольных (для обзора см. Hooykas and Schilperoort, 1992, Plant Mol. Biol. 19: 15-38) и может также быть использован для трансформации однодольных, хотя для этих растений часто используют другие методы трансформации. В настоящее время предпочтительным способом создания трансгенных однодольных является бомбардировка частицами (микроскопическими золотыми или вольфрамовыми частицами, покрытыми трансформирующей ДНК) зародышевых каллусов или развивающихся зародышей (Christou, 1992, Plant J. 2: 275-281; Shimamoto, 1994, Curr. Opin. Biotechnol. 5: 158-162; Vasil et al., 1992, Bio/Technology 10: 667-674). Альтернативный метод трансформации однодольных основан на трансформации протопластов, описанной Omirulleh et al., 1993, Plant Mol. Biol. 21: 415-428. Дополнительные методы трансформации для использования в соответствии с настоящим изобретением, описаны в патентах США 6395966 и 7151204 (полное содержание обоих из которых включено в данное описание посредством ссылок).
После трансформации трансформанты с включенным экспрессионным конструктом отбирают и регенерируют в целые растения с использованием методов, хорошо известных в данной области. Часто метод трансформации предназначен для избирательного удаления селекционных генов либо в процессе регенерации, либо в последующих поколениях, путем использования, например, котрансформации двумя отдельными T-ДНК конструктами или сайт-специфического вырезания селекционного гена при помощи специфической рекомбиназы.
В дополнение к прямой трансформации генотипа конкретного растения конструктом, полученным в соответствии с настоящим изобретением, трансгенные растения можно получать путем скрещивания растения, содержащего конструкт, со вторым растением, не имеющим конструкта. Например, конструкт, кодирующий полипептид, можно вводить в растение конкретного сорта путем скрещивания, без необходимости в прямой трансформации растения данного сорта. Вследствие этого, настоящее изобретение охватывает не только растение, непосредственно регенерированное из клеток, которые были трансформированы в соответствии с настоящим изобретением, но также и потомство таких растений. При использовании в данном описании термин «потомство» может означать потомство любого поколения родительского растения, полученного в соответствии с настоящим изобретением. Такое потомство может содержать конструкт ДНК, полученный согласно настоящему изобретению, или часть конструкта ДНК, полученного согласно настоящему изобретению. Скрещивание приводит к введению трансгена в линию растений путем перекрестного опыления исходной линии с донорской линией растения. Неограничивающие примеры таких шагов более подробно освещены в патенте США 7151204.
Растения можно получать преобразованием посредством процесса обратного скрещивания. Например, растения включают растения, называемые растениями с генотипом, преобразованным обратным скрещиванием, линейные, инбредные или гибридные.
Можно использовать генетические маркеры для облегчения интрогрессии одного или более трансгенов по изобретению из одного генетического фона в другой. Селекция с помощью маркеров дает преимущества по сравнению с традиционной селекцией в том, что ее можно использовать, чтобы избежать ошибок, вызванных фенотипическими вариациями. Кроме того, генетические маркеры могут предоставлять данные об относительной степени наличия элитой зародышевой плазмы в отдельном потомстве от конкретного скрещивания. Например, когда растение с желательной характеристикой, которое в ином случае имело бы нежелательный с агрономической точки зрения генетический фон, скрещивают с элитным родительским растением, генетические маркеры можно использовать для отбора потомства, которое не только обладает представляющей интерес характеристикой, но также обладает относительно высокой долей нужной зародышевой плазмы. Таким образом, число поколений, необходимых для интрогрессии одной или нескольких характеристик в конкретный генетический фон, сведено к минимуму.
Настоящее изобретение также относится к способам получения полипептида по настоящему изобретению, включающим: (a) культивирование трансгенного растения или растительной клетки, содержащих полинуклеотид, кодирующий полипептид, в условиях, благоприятных для продукции полипептида; и (b) выделение полипептида.
Композиции
Настоящее изобретение также относится к композициям, содержащим полипептид по настоящему изобретению. Предпочтительно, композиции обогащены таким полипептидом. Термин «обогащенный» указывает на то, что эндопептидазная активность, предпочтительно трипсиноподобная эндопептидазная активность, композиции является повышенной, например, с коэффициентом обогащения по меньшей мере 1,1.
Композиция может содержать полипептид по настоящему изобретению в качестве основного ферментативного компонента, например, монокомпонентная композиция. Альтернативно, композиция может содержать несколько ферментативных активностей, таких как аминопептидазная, амилазная, карбогидразная, карбоксипептидазная, каталазная, целлюлазная, хитиназная, кутиназная, циклодекстрингликозилтрансферазная, дезоксирибонуклеазная, эстеразная, альфа-галактозидазная, бета-галактозидазная, глюкоамилазная, альфа-глюкозидазная, бета-глюкозидазная, галопероксидазная, инвертазная, лакказная, липазная, маннозидазная, оксидазная, активность пектинолитического фермента, пептидоглютаминазная, пероксидазная, фитазная, полифенолоксидазная, активность протеолитического фермента, рибонуклеазная, трансглутаминазная или ксиланазная. Дополнительный фермент(ы) может продуцироваться, например, микроорганизмом, принадлежащим к роду Aspergillus, например, Aspergillus aculeatus, Aspergillus awamori, Aspergillus foetidus, Aspergillus fumigatus, Aspergillus japonicus, Aspergillus nidulans, Aspergillus niger или Aspergillus oryzae; Fusarium, например, Fusarium bactridioides, Fusarium cerealis, Fusarium crookwellense, Fusarium culmorum, Fusarium graminearum, Fusarium graminum, Fusarium heterosporum, Fusarium negundi, Fusarium oxysporum, Fusarium reticulatum, Fusarium roseum, Fusarium sambucinum, Fusarium sarcochroum, Fusarium sulphureum, Fusarium toruloseum, Fusarium trichothecioides или Fusarium venenatum; Humicola, например, Humicola insolens или Humicola lanuginosa; или Trichoderma, например, Trichoderma harzianum, Trichoderma koningii, Trichoderma longibrachiatum, Trichoderma reesei или Trichoderma viride.
Композиции могут быть получены способами, известными в данной области, и могут находиться в виде жидкой или сухой композиции. Например, композиция может находиться в виде гранулята или микрогранулята. Полипептид можно стабилизировать способами, известными в данной области.
Применения
Настоящее изобретение также относится к способам применения полипептидов по изобретению или их композиций.
Предпочтительно, изобретение относится к применению полипептидов по изобретению или их композиций для получения гидролизата белка, предпочтительно гидролизата пищевого белка, более предпочтительно гидролизата молочного белка, например, гидролизата белка молочной сыворотки или гидролизата казеина.
Изобретение также относится к способу получения гидролизата белка, включающему:
(a) обеспечение раствора, содержащего пищевой белок, подлежащий гидролизу;
(b) добавление к указанному раствору полипептида по изобретению или его композиции; и
(c) получение гидролизата белка.
В предпочтительном варианте осуществления белок, подлежащий гидролизу, является пищевым белком и, таким образом, полученный гидролизат белка представляет собой гидролизат пищевого белка.
Пищевой белок может быть любым пищевым белком. Раствор, содержащий пищевой белок, может быть раствором, содержащим белковый материал из растения, такого как, например, ячмень, канола, люпин, кукуруза, овес, горох, картофель, рис, соя, пшеница или любая их комбинация. Раствор, содержащий пищевой белок, может быть раствором, содержащим белковый материал от животного, такой как, например, белковый материал яйца, белковый материал мяса или белковый материал молока. Он также может представлять собой сочетание белкового материала(ов), полученного из растения, и белкового материала(ов), полученного от животного.
В предпочтительном варианте осуществления пищевой белок является молочным белком. То есть, исходный материал представляет собой раствор, содержащий молочный белок. Такой раствор может содержать белок молочной сыворотки и казеин в любой пропорции, или он может содержать по существу только белок молочной сыворотки или, по существу, только казеин, или он может быть раствором чистого белка молочной сыворотки или чистого казеина. Он может представлять собой, например, молоко, такое как сырое молоко, или любой раствор, содержащий молочный белок, полученный из молока.
В одном варианте осуществления такой раствор является раствором белка молочной сыворотки, который может быть получен из сыворотки, образованной при изготовлении сыра, особенно сладкой молочной сыворотки, такой, которую получают в результате коагуляции казеина при помощи сычуга. Белок молочной сыворотки можно также получать из концентрата белков молочной сыворотки или из изолята белков молочной сыворотки. В предпочтительном варианте осуществления молочный белок представляет собой концентрат белков молочной сыворотки (WPC).
В другом варианте осуществления такой раствор представляет собой раствор казеина. Источником казеина может быть кислый казеин или сухие остатки обезжиренного молока.
Раствор, содержащий пищевой белок, такой как раствор, содержащий молочный белок, предпочтительно содержит примерно 2-35% масс. белка, более предпочтительно примерно 5-30% масс.
В одном варианте осуществления раствор, содержащий молочный белок, предпочтительно раствор, содержащий белок молочной сыворотки, также содержит лактозу.
Следует понимать, что раствор может находиться в форме, которая с технической точки зрения может быть охарактеризована как дисперсия.
Изобретение также относится к применению полипептидов по изобретению для обработки кожи при влажной обработке, то есть, в процессе вымачивания, сгонки волоса и/или мягчения.
В конкретном варианте осуществления изобретения ферментативную обработку кожи полипептидами по изобретению используют в процессе вымачивания. Процесс вымачивания по настоящему изобретению можно проводить в обычных условиях вымачивания, то есть при pH в диапазоне pH 4-11, предпочтительно в диапазоне pH 6-10, температуре в диапазоне 20-30°C, предпочтительно в диапазоне 24-28°C, и времени реакции в диапазоне 2-24 часа, предпочтительно в диапазоне 4-16 часов, и вместе с известными поверхностно-активными веществами и консервантами, при необходимости.
В более конкретном варианте осуществления изобретения ферментативную обработку кожи полипептидами по настоящему изобретению проводят в процессе сгонки волоса. Процесс сгонки волоса по настоящему изобретению можно проводить в обычных условиях, то есть, при pH в диапазоне pH 5,5-12,5, предпочтительно в диапазоне от 6 до 12, более предпочтительно в диапазоне от 7 до 10, температуре в диапазоне от 5°C до 32°C, предпочтительно в диапазоне от 15°C до 30°C, и времени реакции 1-5 часов, предпочтительно от 1,5 до 4 часов.
В другом конкретном варианте осуществления ферментативную обработку кожи проводят в процессе мягчения. В наиболее предпочтительном варианте осуществления ферментативную обработку проводят в процессе мягчения после фазы обеззоливания. Процесс мягчения по настоящему изобретению можно проводить в обычных условиях, то есть, при pH в диапазоне pH 6-9, предпочтительно в диапазоне pH 6,5-8,5, температуре в диапазоне 20-30°C, предпочтительно в диапазоне 25-28°C, и времени реакции в диапазоне 20-90 минут, предпочтительно в диапазоне 40-80 минут.
Способ получения гидролизата пищевого белка
Методика
В другом аспекте настоящее изобретение относится к способу получения гидролизата пищевого белка, включающему:
(a) обеспечение раствора, содержащего пищевой белок, подлежащий гидролизу;
(b) добавление к указанному раствору трипсиноподобной эндопептидазы, полученной из бактерии; и
(c) получение гидролизата пищевого белка.
Пищевой белковый субстрат
Пищевой белок может быть любым пищевым белком. Раствор, содержащий пищевой белок, может быть раствором, содержащим белковый материал из растения, такого как, например, ячмень, канола, люпин, кукуруза, овес, горох, картофель, рис, соя, пшеница или любая их комбинация. Раствор, содержащий пищевой белок, может быть раствором, содержащим белковый материал от животного, такой как, например, белковый материал яйца, белковый материал мяса или белковый материал молока. Он также может представлять собой сочетание белкового материала(ов), полученного из растения, и белкового материала(ов), полученного от животного.
В предпочтительном варианте осуществления пищевой белок является молочным белком, предпочтительно белком молочной сыворотки. То есть исходный материал представляет собой раствор, содержащий молочный белок. Такой раствор может содержать белок молочной сыворотки и казеин в любой пропорции, или он может содержать по существу только белок молочной сыворотки или по существу только казеин, или он может быть раствором чистого белка молочной сыворотки или чистого казеина. Он может представлять собой, например, сырое молоко или любой раствор, содержащий молочный белок, полученный из сырого молока.
В одном варианте осуществления такой раствор является раствором белка молочной сыворотки, который может быть получен из сыворотки, образованной при изготовлении сыра, особенно сладкой молочной сыворотки, такой, которую получают в результате коагуляции казеина при помощи сычуга. Белок молочной сыворотки можно также получать из концентрата белков молочной сыворотки или из изолята белков молочной сыворотки. В предпочтительном варианте осуществления молочный белок представляет собой концентрат белков молочной сыворотки (WPC).
В другом варианте осуществления такой раствор представляет собой раствор казеина. Источником казеина может быть кислый казеин или сухие остатки обезжиренного молока.
Раствор, содержащий пищевой белок, такой как раствор, содержащий молочный белок, предпочтительно содержит примерно 2-35% масс. белка, более предпочтительно примерно 5-30% масс., и даже более предпочтительно примерно 5-20% масс.
В одном варианте осуществления раствор, содержащий молочный белок, предпочтительно раствор, содержащий белок молочной сыворотки, также содержит лактозу.
Следует понимать, что раствор может находиться в форме, которая с технической точки зрения может быть охарактеризована как дисперсия.
Трипсиноподобная эндопептидаза
Раствор, содержащий пищевой белок, такой как раствор, содержащий молочный белок, обрабатывают трипсиноподобной эндопептидазой, полученной из бактерии.
Для целей настоящего изобретения термин «полученный из», используемый в данном описании в связи с полипептидом, полученным из данного источника (то есть, биологического организма), может означать, что полипептид идентичен или является вариантом полипептида, природно кодируемого геномом данного источника, независимо от того, продуцируется ли полипептид другим источником, например, штаммом, в который полинуклеотид из источника, кодирующий эндопептидазу, был введен.
Трипсиноподобную эндопептидазу можно получать из грамположительного бактериального штамма, такого как штамм Actinosynnema, Bacillus, Clostridium, Enterococcus, Geobacillus, Kribbella, Kutzneria, Lactobacillus, Lactococcus, Oceanobacillus, Staphylococcus, Streptococcus или Streptomyces, или грамотрицательного бактериального штамма, такого как штамм Campylobacter, E. coli, Flavobacterium, Fusobacterium, Helicobacter, Ilyobacter, Neisseria, Pseudomonas, Salmonella или Ureaplasma.
В одном варианте осуществления трипсиноподобную эндопептидазу получают из штамма Bacillus alkalophilus, Bacillus amyloliquefaciens, Bacillus brevis, Bacillus circulans, Bacillus clausii, Bacillus coagulans, Bacillus firmus, Bacillus lautus, Bacillus lentus, Bacillus licheniformis, Bacillus megaterium, Bacillus pumilus, Bacillus stearothermophilus, Bacillus subtilis или Bacillus thuringiensis.
В другом варианте осуществления трипсиноподобную эндопептидазу получают из штамма Streptococcus equisimilis, Streptococcus pyogenes, Streptococcus uberis или Streptococcus equi подвид Zooepidemicus.
В другом варианте осуществления трипсиноподобную эндопептидазу получают из штамма Streptomyces achromogenes, Streptomyces avermitilis, Streptomyces coelicolor, Streptomyces griseus или Streptomyces lividans.
Такая бактериальная трипсиноподобная эндопептидаза может иметь оптимальную протеолитическую активность при pH примерно от 5,0 до примерно 11,0, предпочтительно при pH примерно от 6 до примерно 10, и при температуре примерно от 40°C до примерно 75°C, предпочтительно при температуре примерно от 50°C до примерно 70°C.
В предпочтительном варианте осуществления трипсиноподобная эндопептидаза имеет идентичность последовательности со зрелым полипептидом любой из SEQ ID NO: 2, 5 или 6 по меньшей мере 40%, по меньшей мере 45%, по меньшей мере 50%, по меньшей мере 55%, по меньшей мере 60%, по меньшей мере 65%, по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100%. В другом предпочтительном варианте осуществления трипсиноподобная эндопептидаза отличается не более чем десятью аминокислотами, например, пятью аминокислотами, четырьмя аминокислотами, тремя аминокислотами, двумя аминокислотами или одной аминокислотой от зрелого полипептида любой из SEQ ID NO: 2, 5 или 6.
Зрелый полипептид SEQ ID NO:2 может представлять собой аминокислоты 1-225. Зрелый полипептид SEQ ID NO:5 может представлять собой аминокислоты 38-261. Зрелый полипептид SEQ ID NO:6 может представлять собой аминокислоты 40-266.
В предпочтительном варианте осуществления трипсиноподобную эндопептидазу получают из актинобактерии.
В более предпочтительном варианте осуществления трипсиноподобную эндопептидазу получают из штамма Kutzneria, например, из Kutzneria albida.
В другом более предпочтительном варианте осуществления трипсиноподобная эндопептидаза имеет идентичность последовательности со зрелым полипептидом SEQ ID NO:2 по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100%. В другом предпочтительном варианте осуществления трипсиноподобная эндопептидаза отличается не более чем десятью аминокислотами, например, пятью аминокислотами, четырьмя аминокислотами, тремя аминокислотами, двумя аминокислотами или одной аминокислотой от зрелого полипептида SEQ ID NO:2. Такая трипсиноподобная эндопептидаза может иметь оптимальную протеолитическую активность при pH примерно от 5,0 до примерно 11,0, предпочтительно при pH примерно от 6 до примерно 10, и при температуре примерно от 40°C до примерно 75°C, предпочтительно при температуре примерно от 50°C до примерно 70°C. Она может ингибироваться апротинином.
В другом более предпочтительном варианте осуществления трипсиноподобную эндопептидазу получают из штамма Actinosynnema, например, из Actinosynnema mirum. Она может, например, иметь аминокислотную последовательность зрелого полипептида SEQ ID NO:5 по настоящей изобретению (UNIPROT:C6WDM8).
В другом более предпочтительном варианте осуществления трипсиноподобная эндопептидаза имеет идентичность последовательности со зрелым полипептидом SEQ ID NO:5 по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100%. В другом предпочтительном варианте осуществления трипсиноподобная эндопептидаза отличается не более чем десятью аминокислотами, например, пятью аминокислотами, четырьмя аминокислотами, тремя аминокислотами, двумя аминокислотами или одной аминокислотой от зрелого полипептида SEQ ID NO:5. Такая трипсиноподобная эндопептидаза может иметь оптимальную протеолитическую активность при pH примерно от 5,0 до примерно 11,0, предпочтительно при pH примерно от 8 до примерно 10, и при температуре примерно от 40°C до примерно 75°C, предпочтительно при температуре примерно от 50°C до примерно 70°C. Она может ингибироваться PMSF.
В другом более предпочтительном варианте осуществления трипсиноподобную эндопептидазу получают из штамма Kribbella, например, из Kribbella flavida. Она может, например, иметь аминокислотную последовательность зрелого полипептида SEQ ID NO:6 по настоящей изобретению (UNIPROT:D2PZJ1).
В другом более предпочтительном варианте осуществления трипсиноподобная эндопептидаза имеет идентичность последовательности со зрелым полипептидом SEQ ID NO:6 по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100%. В другом предпочтительном варианте осуществления трипсиноподобная эндопептидаза отличается не более чем десятью аминокислотами, например, пятью аминокислотами, четырьмя аминокислотами, тремя аминокислотами, двумя аминокислотами или одной аминокислотой от зрелого полипептида SEQ ID NO:6. Такая трипсиноподобная эндопептидаза может иметь оптимальную протеолитическую активность при pH примерно от 5,0 до примерно 11,0, предпочтительно при pH примерно от 8 до примерно 11, и при температуре примерно от 40°C до примерно 75°C, предпочтительно при температуре примерно от 40°C до примерно 60°C. Она может ингибироваться EDTA.
Трипсиноподобную эндопептидазу можно получать при помощи штамма, из которого она происходит. Или ее можно получать из рекомбинантной клетки-хозяина, содержащей полинуклеотид, кодирующий эндопептидазу, функционально связанный с одной или более (несколькими) контрольными последовательностями, которые направляют ее продукцию. Конструкт или вектор, содержащий полинуклеотид, вводят в клетку-хозяина так, что конструкт или вектор сохраняется в виде составной части хромосомы или в виде самореплицирующегося внехромосомного вектора.
Клетка-хозяин может быть любой клеткой, применимой для рекомбинантной продукции эндопептидазы, например, прокариотической или эукариотической.
Прокариотическая клетка-хозяин может быть любой грамположительной или грамотрицательной бактерией. Грамположительные бактерии включают, но не ограничиваются ими, Bacillus, Clostridium, Enterococcus, Geobacillus, Lactobacillus, Lactococcus, Oceanobacillus, Staphylococcus, Streptococcus и Streptomyces. Грамотрицательные бактерии включают, но не ограничиваются ими, Campylobacter, E. coli, Flavobacterium, Fusobacterium, Helicobacter, Ilyobacter, Neisseria, Pseudomonas, Salmonella и Ureaplasma.
Бактериальная клетка-хозяин может быть любой клеткой Bacillus, включая, но, не ограничиваясь ими, клетки Bacillus alkalophilus, Bacillus amyloliquefaciens, Bacillus brevis, Bacillus circulans, Bacillus clausii, Bacillus coagulans, Bacillus firmus, Bacillus lautus, Bacillus lentus, Bacillus licheniformis, Bacillus megaterium, Bacillus pumilus, Bacillus stearothermophilus, Bacillus subtilis и Bacillus thuringiensis.
Бактериальная клетка-хозяин может также быть любой клеткой Streptococcus, включая, но, не ограничиваясь ими, клетки Streptococcus equisimilis, Streptococcus pyogenes, Streptococcus uberis и Streptococcus equi подвид Zooepidemicus.
Бактериальная клетка-хозяин может также быть любой клеткой Streptomyces, включая, но, не ограничиваясь ими, клетки Streptomyces achromogenes, Streptomyces avermitilis, Streptomyces coelicolor, Streptomyces griseus и Streptomyces lividans.
Клетка-хозяин также может быть эукариотической, например, клеткой млекопитающего, насекомого, растения или гриба.
Трипсиноподобная эндопептидаза предпочтительно представляет собой выделенную трипсиноподобную эндопептидазу. Концентрация трипсиноподобной эндопептидазы может составлять 100-500000 трипсиновых единиц USP на г пищевого белка, например, 250-250000 или 500-100000.
Одна трипсиновая единица USP представляет собой активность, вызывающую изменение в поглощении при 253 нм, соответствующее 0,003 при pH 7,6 и 25°C при использовании этилового эфира гидрохлорида N-бензоил-L-аргинина (BAEE) в качестве субстрата.
Специфическая активность может варьироваться весьма существенно среди различных трипсиноподобных эндопептидаз, однако специалист в данной области сможет легко определить, в каком количестве трипсиноподобную эндопептидазу следует использовать, например, в зависимости от степени гидролиза.
Отношение трипсиноподобной эндопептидазы к пищевому белку предпочтительно составляет 0,01-5% масс., более предпочтительно 0,01-2%, более предпочтительно 0,05-0,8%, даже более предпочтительно 0,1-0,6% и наиболее предпочтительно примерно 0,2%.
Необязательное включение другой эндопептидазы, являющейся менее специфичной
В предпочтительном варианте осуществления по меньшей мере одну другую эндопептидазу добавляют в раствор до, во время или после стадии (b).
Предпочтительно, по меньшей мере одна другая эндопептидаза представляет собой выделенную эндопептидазу.
Предпочтительно, по меньшей мере одна другая эндопептидаза представляет собой сериновую эндопептидазу.
В предпочтительном варианте осуществления по меньшей мере одна другая эндопептидаза обладает активностью, которая менее специфична, чем активность трипсиноподобной эндопептидазы.
В другом предпочтительном варианте осуществления по меньшей мере одна другая эндопептидаза обладает активностью, которая напоминает активность химотрипсина млекопитающих, например, химотрипсина, экстрагированного из ткани поджелудочной железы свиней.
В другом предпочтительном варианте осуществления по меньшей мере одна другая эндопептидаза обладает более высокой специфичностью в отношении расщепления по карбоксильному концу любого из тирозина, фенилаланина, триптофана, лейцина, метионина или гистидина, чем в отношении расщепления по карбоксильному концу любой другой природной аминокислоты.
В другом предпочтительном варианте осуществления по меньшей мере одна другая эндопептидаза обладает специфичностью в отношении расщепления по карбоксильному концу по меньшей мере одного из тирозина, фенилаланина, триптофана, лейцина, метионина или гистидина, которая по меньшей мере в 3 раза выше, предпочтительно по меньшей мере в 5 раз выше, чем ее специфичность в отношении расщепления по карбоксильному концу любого из аланина, аргинина, аспарагина, аспарагиновой кислоты, цистеина, глютаминовой кислоты, глютамина, глицина, изолейцина, лизина, пролина, серина, треонина и валина.
В другом предпочтительном варианте осуществления по меньшей мере одна другая эндопептидаза обладает более высокой специфичностью в отношении расщепления по карбоксильному концу каждой из по меньшей мере трех аминокислот из группы, состоящей из тирозина, фенилаланина, триптофана, лейцина, метионина и гистидина, чем в отношении расщепления по карбоксильному концу аргинина.
В другом предпочтительном варианте осуществления по меньшей мере одна другая эндопептидаза обладает более высокой специфичностью в отношении расщепления по карбоксильному концу каждой из по меньшей мере трех аминокислот из группы, состоящей из тирозина, фенилаланина, триптофана, лейцина, метионина и гистидина, чем в отношении расщепления по карбоксильному концу лизина.
В другом предпочтительном варианте осуществления по меньшей мере одна другая эндопептидаза обладает более высокой специфичностью в отношении расщепления по карбоксильному концу каждого из тирозина, фенилаланина, триптофана, лейцина, метионина и гистидина, чем в отношении расщепления по карбоксильному концу и аргинина и лизина.
В другом предпочтительном варианте осуществления по меньшей мере одна другая эндопептидаза обладает химотрипсиновым отношением, составляющим по меньшей мере 3, предпочтительно по меньшей мере 5. Химотрипсиновое отношение, равное по меньшей мере 5, означает, что активность фермента при расщеплении после одного из Phe, Leu или Met (в зависимости от того, что больше) по меньшей мере в пять раз выше, чем активность при расщеплении после любого из Ala, Arg, Asp, Glu, Ile, Lys или Val (в зависимости от того, что больше). То есть по меньшей мере одна другая эндопептидаза обладает специфичностью в отношении расщепления после одного из Phe, Leu или Met (в зависимости от того, что больше), которая по меньшей мере в 3 раза выше, предпочтительно по меньшей мере в 5 раз выше, чем ее специфичность в отношении расщепления после любого из Ala, Arg, Asp, Glu, Ile, Lys или Val (в зависимости от того, что больше). Такие измерения активности для определения химотрипсинового отношения следует проводить при pH, при котором активность эндопептидазы составляет по меньшей мере половину от активности эндопептидазы при оптимальном для нее pH. Химотрипсиновое отношение можно определять, как описано в примере 1 патентной заявки № WO 2010/112546.
В другом предпочтительном варианте осуществления по меньшей мере одна другая эндопептидаза представляет собой бактериальную эндопептидазу.
В более предпочтительном варианте осуществления по меньшей мере одна другая эндопептидаза получена из штамма Nocardiopsis, предпочтительно из Nocardiopsis sp. NRRL 18262 (ранее описанного, например, в WO 88/03947). Она может, например, обладать аминокислотной последовательностью зрелого полипептида SEQ ID NO:7 по настоящему изобретению. ДНК и аминокислотная последовательности протеазы, полученной из Nocardiopsis sp. NRRL 18262, ранее опубликованы, например, в патентной заявке Дании № 1996 00013.
В другом более предпочтительном варианте осуществления по меньшей мере одна другая эндопептидаза получена из Metarhizium, предпочтительно Metarhizium anisopliae, и, например, обладает аминокислотной последовательностью зрелого полипептида SEQ ID NO:8 по настоящему изобретению (TREMBL:Q9Y843). В другом более предпочтительном варианте осуществления по меньшей мере одна другая эндопептидаза получена из Brachysporiella, предпочтительно Brachysporiella gayana, и, например, обладает аминокислотной последовательностью зрелого полипептида SEQ ID NO:9 по настоящему изобретению (CGMCC 0865). ДНК и аминокислотные последовательности протеаз, полученных из Metarhizium anisopliae и Brachysporiella gayana, ранее опубликованы, например, в WO 04072279.
В другом предпочтительном варианте осуществления по меньшей мере одна другая эндопептидаза имеет идентичность последовательности со зрелым полипептидом любой из SEQ ID NO: 7, 8 или 9 по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100%. В другом предпочтительном варианте осуществления по меньшей мере одна другая эндопептидаза отличается не более чем десятью аминокислотами, например, пятью аминокислотами, четырьмя аминокислотами, тремя аминокислотами, двумя аминокислотами или одной аминокислотой от зрелого полипептида любой из SEQ ID NO: 7, 8 или 9.
Зрелый полипептид SEQ ID NO:7 может представлять собой аминокислоты 1-188. Зрелый полипептид SEQ ID NO:8 может представлять собой аминокислоты 187-374. Зрелый полипептид SEQ ID NO:9 может представлять собой аминокислоты 190-375.
В более предпочтительном варианте осуществления по меньшей мере одна другая эндопептидаза имеет идентичность последовательности со зрелым полипептидом SEQ ID NO:7 по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100%. В другом предпочтительном варианте осуществления по меньшей мере одна другая эндопептидаза отличается не более чем десятью аминокислотами, например, пятью аминокислотами, четырьмя аминокислотами, тремя аминокислотами, двумя аминокислотами или одной аминокислотой от зрелого полипептида SEQ ID NO:7.
Концентрация по меньшей мере одной другой эндопептидазы предпочтительно составляет 100-100000 химотрипсиновых единиц USP на г пищевого белка, более предпочтительно 500-50000 и наиболее предпочтительно 1000-20000.
Одна химотрипсиновая единица USP представляет собой активность, вызывающую изменение в поглощении при 237 нм, соответствующее 0,0075 при pH 7,0 и 25°C при использовании этилового эфира N-ацетил-L-тирозина (ATEE) в качестве субстрата.
Специфическая активность может варьироваться весьма существенно среди различных эндопептидаз, однако специалист в данной области сможет легко определить, в каком количестве по меньшей мере одну другую эндопептидазу следует использовать, например, в зависимости от степени гидролиза.
Отношение по меньшей мере одной другой эндопептидазы к пищевому белку предпочтительно составляет 0,001-1% масс., более предпочтительно 0,001-0,5%, более предпочтительно 0,005-0,25% и наиболее предпочтительно 0,01-0,05%.
Предпочтительно, по меньшей мере одну другую эндопептидазу добавляют в концентрации, составляющей от 1% до 50% от концентрации добавленной трипсиноподобной эндопептидазы, в расчете на массу эндопептидаз. В одном предпочтительном варианте осуществления по меньшей мере одну другую эндопептидазу добавляют в концентрации, составляющей от 2% до 20% масс./масс. трипсиноподобной эндопептидазы, предпочтительно от 3% до 5%. В другом предпочтительном варианте осуществления по меньшей мере одну другую эндопептидазу добавляют в концентрации, составляющей от 5% до 15% масс./масс. трипсиноподобной эндопептидазы, предпочтительно примерно 10%.
Гидролизат пищевого белка
Необязательной предварительной стадией перед гидролизом является предварительное нагревание раствора, содержащего пищевой белок. В предпочтительном варианте осуществления осуществляют стадию предварительной обработки, которая включает нагревание пищевого белка примерно при 75-95°C в течение примерно 5-30 минут. В другом предпочтительном варианте осуществления осуществляют стадию предварительной обработки, которая включает нагревание пищевого белка при температуре выше 135°C в течение примерно 1-5 секунд. В другом предпочтительном варианте осуществления осуществляют стадию предварительной обработки, которая включает нагревание пищевого белка примерно при 130°C в течение примерно 30-60 секунд.
Специалисту в данной области известно, какие условия предпочтительнее применять для реакции гидролиза. Ее можно, например, проводить при температуре примерно от 40°C до 60°C в течение 1-6 часов при pH в диапазоне от 6,5 до 8,5, предпочтительно от 6,5 до 8.
В предпочтительном варианте осуществления после первой обработки эндопептидазой белковый материал дополнительно подвергают второму протеолитическому гидролизу, с последующей инактивацией эндопептидазы. В более предпочтительном варианте осуществления белковый материал подвергают термообработке в промежутке между первым и вторым протеолитическим гидролизом, как описано в патенте США 5039532, который включен в данное описание посредством ссылки.
Независимо от условий гидролиза, гидролизат предпочтительно подвергают дополнительной стадии инактивации эндопептидазы. Эта инактивация пептидазы в предпочтительном варианте осуществления включает термообработку в течение примерно 0,1-30 минут при температуре примерно 70-110°C, предпочтительно 75-95°C. Альтернативно, эндопептидазу можно инактивировать путем стерилизации при сверхвысокой температуре (например, примерно при 130°C в течение примерно 30-60 секунд).
Полученный гидролизат пищевого белка может быть дополнительно обработан. Его можно осветлять. Его можно хранить в жидком состоянии. Гидролизат также можно подвергать ультрафильтрации, его можно концентрировать, например, выпариванием, и его можно сушить, например, распылительной сушкой или лиофилизацией.
В предпочтительном варианте осуществления полученный гидролизат пищевого белка имеет умеренную степень гидролиза. В другом предпочтительном варианте осуществления полученный гидролизат пищевого белка представляет собой частичный гидролизат. В другом предпочтительном варианте осуществления полученный гидролизат пищевого белка имеет степень гидролиза от 5 до 30%, предпочтительно от 6 до 15%. Особенно предпочтительная степень гидролиза составляет примерно 14%. Другая особенно предпочтительная степень гидролиза составляет примерно 15%.
Степень гидролиза (DH) выражает степень гидролиза белка, полученную данным способом. В контексте изобретения степень гидролиза (DH) определяют следующим образом:
DH=(число расщепленных пептидных связей/общее число пептидных связей)×100%.
Степень гидролиза (DH) полученного гидролизата белка можно измерять спектрофотометрически по методу Church, F.C. et al. (1983) Spectrophotometric Assay Using o-Phthaldialdehyde for Determination of Proteolysis in Milk and Isolated Milk Proteins, J. Dairy Sci. 66: 1219-1227.
Молекулярно-массовое распределение пептидов в полученном гидролизате пищевого белка можно определить, например, методом эксклюзионной хроматографии (SEC). В предпочтительном варианте осуществления гидролизат по изобретению состоит из пептидов, среди которых менее чем 1% по массе имеет молекулярную массу выше, чем 20000 кДа.
Гидролизат пищевого белка, полученный способом по изобретению, предпочтительно лишен поддающегося обнаружению интактного пищевого белка. Отсутствие интактного пищевого белка в гидролизате можно демонстрировать методом электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-ПААГ). Прямое сравнение гидролизата и негидролизованного исходного белкового материала можно проводить в том же геле. В предпочтительном варианте осуществления гидролизат по изобретению состоит из пептидов, среди которых менее чем 1% по массе представляют собой интактный пищевой белок.
Остаточную антигенность гидролизата, полученного способом по изобретению, можно определять при помощи иммуноферментного анализа (ELISA). Негидролизованный пищевой белок иммобилизуют на твердой фазе в концентрации, находящейся в диапазоне линейной зависимости доза-ответ, установленном в данном анализе. Препараты гидролизатов иммобилизуют аналогичным образом. Затем последовательные инкубации с антителом кролика и ферментативным конъюгатом, реагирующим с IgG кролика, выявляют наличие антигенно-распознаваемых белков и пептидов. Результаты, полученные с гидролизатом, сравнивают на основе массы с результатами, полученными с не гидролизованным исходным белковым материалом. Затем рассчитывают уменьшение антигенности гидролизата в процентах.
В предпочтительном варианте осуществления у гидролизата пищевого белка, полученного способом по изобретению, антигенность снижается по меньшей мере примерно на 80%, предпочтительно по меньшей мере примерно на 85%, более предпочтительно по меньшей мере примерно на 90% или по меньшей мере примерно на 95%, наиболее предпочтительно по меньшей мере примерно на 98%, и даже еще более предпочтительно, снижение антигенности составляет по меньшей мере примерно 99%, по сравнению с соответствующим не гидролизованным пищевым белковым материалом, что измеряют методом ELISA.
Настоящее изобретение дополнительно описано с помощью следующих примеров, которые не должны рассматриваться как ограничивающие объем изобретения.
Примеры
Пример 1
Клонирование и экспрессия трипсиноподобной эндопептидазы из Kutznena albida
Ген
Трипсиноподобную эндопептидазу из Kutznena albida экспрессировали из синтетического гена в Bacillus subtilis. Последовательность синтетического гена конструировали на основе SEQ ID NO:2 (зрелая трипсиноподобная эндопептидаза Kutznena albida) и оптимизировали по кодонам для экспрессии в Bacillus subtilis. Экспрессированная последовательность ДНК представляла собой SEQ ID NO:3. Трипсиноподобную эндопептидазу Kutznena albida экспрессировали с сигналом секреции савиназы® (со следующей аминокислотной последовательностью: MKKPLGKIVASTALLISVAFSSSIASA (SEQ ID NO:4)), заменяющим природный сигнал секреции. Нуклеотиды 1-81 в SEQ ID NO:3 представляют собой последовательность ДНК сигнала секреции савиназы, и нуклеотиды 82-756 кодируют зрелый полипептид.
Экспрессионный вектор
ExpVec8 использовали в качестве экспрессионного вектора, и плазмидная карта представлена на фиг.1. Вектор делает возможной интеграцию генного конструкта на хромосому Bacillus subtilis путем гомологичной рекомбинации в локус пектат-лиазы хозяина Bacillus subtilis. Ген экспрессировали из тройной промоторной системы, состоящей из промоторов из гена альфа-амилазы (amyL) Bacillus licheniformis, гена альфа-амилазы (amyQ) Bacillus amyloliquefaciens и промотора cryIIIA Bacillus thuringiensis, включая стабилизирующую последовательность (как описано в WO 99/43835). ExpVec8 содержит сигнальный пептид савиназы, терминатор савиназы, ген устойчивости к хлорамфениколу, чтобы сделать возможным отбор на интегрированные элементы в Bacillus subtilis, ген устойчивости к ампициллину (bla), ген устойчивости к канамицину (neo) и точку начала репликации E. coli.
Экспрессионное клонирование
Стандартный клонирующий вектор, содержащий синтетический ген, трансформировали в E. coli (dam-/dcm- от New England BioLabs, дефицитную по метилированию остатков аденина и цитозина в ДНК). Плазмиду очищали и расщепляли ClaI и MluI для высвобождения вставки с синтетическим геном. ExpVec8 использовали в качестве экспрессионного вектора и расщепляли теми же ферментами рестрикции (ClaI и MluI). Вектор и очищенный фрагмент лигировали и трансформировали в E. coli (Top10, Invitrogen). Экспрессионную плазмиду, содержащую вставку, очищали из одного из трансформантов и повторно трансформировали в Bacillus subtilis. Рекомбинантный клон Bacillus subtilis, содержащий интегрированный экспрессионный конструкт, выращивали в жидкой культуре. Содержащий фермент супернатант собирали, и фермент очищали, как описано в примере 2.
Пример 2
Очистка и характеристика трипсиноподобной эндопептидазы AC3 из Kutzneria albida
Анализ на активность при очистке
Субстрат pNA: Boc-VLGR-pNA (Bachem L-1205).
Температура: комнатная температура (25°C).
Буфер для анализа: 100 мМ янтарная кислота, 100 мМ HEPES, 100 мМ CHES, 100 мМ CABS, 1 мМ CaCl2, 150 мМ KCl, 0,01% Triton X-100, pH 6,0.
По 20 мкл протеазы (разведенной в 0,01% Triton X-100) вносили в лунки планшета для микротитрования. Анализ начинали добавлением 200 мкл субстрата pNA (50 мг Boc-VLGR-pNA, разведенного в 1,0 мл ДМСО и дополнительно разведенного 90x буфером для анализа). Первоначальное увеличение OD405 отслеживали как показания протеазной активности.
Анализы для характеристики активности:
1) анализ pNA:
Субстраты pNA:
Boc-VLGR-pNA (Bachem L-1205)
Suc-AAPF-pNA (Bachem L-1400)
Suc-AAPA-pNA (Bachem L-1775)
Suc-AAPR-pNA (Bachem L-1720)
Suc-AAPD-pNA (Bachem L-1835)
Suc-AAPE-pNA (Bachem L-1710)
Suc-AAPI-pNA (Bachem L-1790)
Suc-AAPL-pNA (Bachem L-1390)
Suc-AAPK-pNA (Bachem L-1725)
Suc-AAPM-pNA (Bachem L-1395)
Suc-AAPV-pNA (Bachem L-1770)
Температура: комнатная температура (25°C)
Буферы для анализа: 100 мМ янтарная кислота, 100 мМ HEPES, 100 мМ CHES, 100 мМ CABS, 1 мМ CaCl2, 150 мМ KCl, 0,01% Triton X-100, доведенный до pH 2,0, 2,5, 3,0, 3,5, 4,0, 5,0, 6,0, 7,0, 8,0, 9,0, 10,0, 11,0 и 12,0 при помощи HCl или NaOH.
20 мкл протеазы (разведенной в 0,01% Triton X-100) смешивали с 100 мкл буфера для анализа. Анализ начинали добавлением 100 мкл субстрата pNA (50 мг разведенного в 1,0 мл ДМСО и дополнительно разведенного 45× в 0,01% Triton X-100). Первоначальное увеличение OD405 отслеживали как показания протеазной активности.
2) Анализ с Protazyme AK:
Субстрат: таблетка Protazyme AK (перекрестно сшитый и окрашенный казеин; от Megazyme).
Температура: контролируемая (температура анализа).
Буферы для анализа: 100 мМ янтарная кислота, 100 мМ HEPES, 100 мМ CHES, 100 мМ CABS, 1 мМ CaCl2, 150 мМ KCl, 0,01% Triton X-100, доведенный до pH 2,0, 2,5, 3,0, 3,5, 4,0, 5,0, 6,0, 7,0, 8,0, 9,0, 10,0 и 11,0 при помощи HCl или NaOH.
Таблетку Protazyme AK суспендировали в 2,0 мл 0,01% Triton X-100 при осторожном перемешивании. 500 мкл этой суспензии и 500 мкл буфера для анализа смешивали в пробирке эппендорф и помещали на лед. Добавляли 20 мкл образца протеазы (разведенного в 0,01% Triton X-100). Анализ начинали перенесением пробирки эппендорф в термомиксер Eppendorf, установленный на температуру анализа. Пробирку инкубировали в течение 15 минут на термомиксере Eppendorf при самой высокой скорости встряхивания (1400 оборотов в минуту). Инкубацию останавливали перенесением пробирки обратно в ледяную баню. Затем пробирку центрифугировали в охлажденной до температуры льда центрифуге в течение нескольких минут, и 200 мкл супернатанта переносили в планшет для микротитрования. Показатели OD650 являлись показателем протеазной активности. Пустой буфер включали в анализ (вместо фермента).
Очистка трипсиноподобной протеазы AC3 из Kutzneria albida
Культуральную жидкость из примера 1 центрифугировали (20000×g, 20 мин) и супернатанты осторожно сливали с осадков. Объединенные супернатанты фильтровали через 0,2-мкм фильтрационную ячейку Nalgene для удаления остатков клеток-хозяев Bacillus. 0,2-мкм фильтрат осаждали сульфатом аммония, добавляя сухой сульфат аммония до конечной концентрации 3,4M (NH4)2SO4. Образовавшийся осадок собирали центрифугированием (20000×g, 20 мин) и осадок растворяли в минимальном объеме деионизированной воды. Раствор переводили в буфер 50 мМ H3BO3, 5 мМ MES, 1 мМ CaCl2, pH 6 на колонке с сефадексом® G25 и наносили на колонку с S-сефарозой® HP, уравновешенную в том же буфере. После интенсивного промывания колонки уравновешивающим буфером протеазу элюировали линейным градиентом NaCl (0-->0,5 M) в том же буфере. Фракции с колонки анализировали на протеазную активность (анализ на активность при очистке). Сухой сульфат аммония добавляли к фракциям, соответствующим пику активности в отношении BOC-VLGR-pNA, с колонки с S-сефарозой HP для получения 1,6M конечной концентрации (NH4)2SO4 в растворе фермента. Раствор фермента осторожно перемешивали магнитной мешалкой в процессе добавления ((NH4)2SO4, и перемешивание продолжали в течение 30 минут после добавления, чтобы привести систему в равновесие. Затем раствор фермента наносили на колонку с фенилсефарозой FF (high sub), уравновешенную в буфере 100 мМ H3BO3, 10 мМ MES, 2 мМ CaCl2, 1,6 M (NH4)2SO4, pH 6. После интенсивного промывания колонки уравновешивающим буфером трипсиновую протеазу AC3 элюировали линейным градиентом (NH4)2SO4 (1,6-->0M) в том же буфере. Фракции с колонки анализировали на протеазную активность (анализ на активность при очистке) и активные фракции дополнительно анализировали методом SDS-ПААГ. Фракции, для которых в окрашенном кумасси SDS-ПААГ геле была видна только одна полоса, объединяли и переводили в буфер 100 мМ H3BO3, 10 мМ MES, 2 мМ CaCl2, pH 6 на колонке с сефадексом G25 как очищенный препарат и использовали для дальнейшей характеристики.
Характеристика: зависимость активности от pH, зависимость стабильности от pH и зависимость активности от температуры
Анализ с pNA использовали для получения профиля зависимости активности от pH в отношении Boc-VLGR-pNA и в отношении Suc-AAPR-pNA, а также для получения профиля зависимости стабильности от pH. Для профиля зависимости стабильности от pH протеазу разводили 10× в буферах для анализа и инкубировали в течение 2 часов при 37°C. После инкубации перед анализом на остаточную активность образцы протеазы доводили до pH 9 путем разведения в буфере для анализа с pH 9. Анализ с Protazyme AK использовали для получения профиля зависимости активности от pH при 37°C и профиля зависимости активности от температуры при pH 7. Результаты приведены ниже в таблицах 1-3. В таблице 1 активности приведены относительно активности при оптимальном pH для фермента для каждого субстрата. В таблице 2 активности представляют собой остаточные активности относительно образца, который хранили в стабильных условиях (5°C, pH 9). В таблице 3 активности приведены относительно активности при оптимальной температуре для фермента при pH 7.
Профили зависимости активности от pH
Профиль зависимости стабильности от pH
Профиль зависимости активности от температуры при pH 7,0
Характеристика: P1-специфичность в отношении субстратов Suc-AAPX-pNA и расчет трипсинового отношения
Анализ с pNA использовали для определения P1-специфичности для трипсиновой протеазы AC3, используя 10 субстратов Suc-AAPX-pNA при двух различных pH: pH 7,0 и pH 9,0. Активности также использовали для расчета трипсинового отношения, как описано в WO 2010/112546. Результаты приведены ниже в таблице 4. В таблице 4 активность для каждого субстрата Suc-AAPX-pNA приведена относительно активности для наилучшего субстрата Suc-AAPX-pNA (Suc-AAPR-pNA).
P1-специфичность в отношении Suc-AAPX-pNA и трипсиновое отношение
Другие характеристики
Трипсиновая протеаза AC3 ингибируется апротинином.
Относительная молекулярная масса, определенная методом SDS-ПААГ, составляла Mr=26 кДа.
N-концевая последовательность была определена как: IVGGTKASTSTY.
Интактная молекулярная масса была определена как ММ=23087,67 Да.
Эти данные указывают на то, что зрелая трипсиновая протеаза AC3 имеет следующую последовательность (аминокислоты 1-225 в SEQ ID NO:2):

Пример 3
Анализ специфичности расщепления трипсиноподобной эндопептидазы AC3 из Kutzneria albida
Введение
Специфичность протеолитического расщепления микробной трипсиноподобной эндопептидазы AC3 сравнивали со свиным трипсином.
Анализы специфичности расщепления проводили путем инкубации описанных эндопептидаз с естественным модельным субстратом бычьим BioPURE-альфа-лактальбумином от Davisco Foods international. Пептидные профили полученных протеолитических пептидов определяли путем разделения методом ОФ-ВЭЖХ с УФ-детекцией сигнала при 214 нм.
Образцы
Протеазы: свиной трипсин (регистрационный номер UniProt: P00761)
AC3 (трипсиноподобная эндопептидаза из Kutzneria albida)
Субстрат: Альфа-лактальбумин от Davisco, BE-2009-00036, 97% белок от Davisco Foods international
Протеолиз
1 г альфа-лактальбумина растворяли в 17,6 мл 5 мМ CaCl2. Образец нагревали до 55°С и рН доводили до 7,5 0,25М NaOH. Фермент добавляли в соотношении 2 мг фермента на грамм альфа-лактальбумина. Титрование pH проводили на Titralab 856 в течение 120 минут. 200 мкл образцов отбирали через 120 минут и титрование останавливали, добавляя 2,2 мкл трифторуксусной кислоты (ТФУ). Образец хранили при -20°C до анализа ОФ-ВЭЖХ.
Анализ ОФ-ВЭЖХ
Протеолитические образцы анализировали на системе ОФ-ВЭЖХ, состоящей из колонки Waters C18 (ACQUITY UPLC® BEH C18, 1,7 мкм, 2,1×100 мм) и системы жидкостной хроматографии Accela от Thermo Scientific. Все образцы разводили 5× в 0,1% ТФУ (10 мкл образца +40 мкл 0,1% ТФУ). Объем 5 мкл вводили инжектором в колонку.
Пептиды разделяли при следующем градиенте:
Все образцы разводили 5×. Объем инъекции: 5 мкл.
Растворитель A: 0,1% ТФУ (CAS номер 76-05-1) в воде UHQ и растворитель B: 0,08% ТФУ в ацетонитриле (CAS номер 75-05-8).
Элюируемые пептиды подвергали мониторингу онлайн при помощи УФ-детектора при 214 нм.
Результаты
Для сравнения, УФ-хроматограмма анализа AC3 (верхняя кривая) приведена на фиг.2 вместе с анализом свиного трипсина (нижняя кривая).
Вывод
ОФ-ВЭЖХ профиль пептидов альфа-лактальбумина, гидролизованного AC3, сходен с профилем пептидов, полученных с помощью свиного трипсина; таким образом, AC3 обладает трипсиноподобной специфичностью.
Пример 4
Гидролиз концентрата белков молочной сыворотки и альфа-лактальбумина трипсиноподобной протеазой AC3 в сравнении со свиным трипсином
Материалы
Концентрат белков молочной сыворотки (WPC), 80% сухой белок, лакпродан 80, Aria Foods Ingredients, DK.
Альфа-лактальбумин (ALA), 97% сухой белок, Davisco Foods International, MN, US.
CaCl2 безводный, Merck артикул 2387.
NaOH, Prolabo 31627.368.
Гидролизный анализ
Растворяли 1 г альфа-лактальбумина в 17,6 мл 5 мМ CaCl2 или 1,2 г лакпродана 80 в 17,4 мл 5 мМ CaCl2 с получением конечной концентрации белка 5%.
Образцы нагревали до 55°С и рН доводили до 7,5. Объем NaOH, использованный для доведения pH, записывали. Добавляли фермент, и титрование pH проводили на Titralab 856 (радиометр) в течение 120 минут. Затраты NaOH подвергали мониторингу и переводили в %DH.
Степень гидролиза
Степень гидролиза суспензии измеряли при помощи pH-стата, как описано в Adler-Nissen, J. 1986, Enzymatic Hydrolysis of Food Proteins, глава 6.
Степень гидролиза (DH), определенную как процент расщепленных пептидных связей, можно отслеживать методом pH-статирования: DH=h/htot×100
h: число расщепленных пептидных связей, пропорциональное количеству основания, потребленного в процессе реакции;
htot: число пептидных связей в белке, рассчитанное на основании аминокислотного состава.
htot для ALA=8,16 и htot для WPC=8,8
h=B×Nb×1/α×1/Mp
B = потребление основания (мл);
Nb = нормальность основания (0,25 Н);
1/α = средняя степень диссоциации α-NH2 групп;
Mp = масса белка (г), (N×фактор Кьельдаля).
Использованные эндопептидазы:
Данные:
Вывод
Как в случае с альфа-лактальбумином белковой фракции молочной сыворотки, так и в случае концентрата белков молочной сыворотки, очевидно, можно добиться степени гидролиза (DH) в том же диапазоне при использовании трипсиноподобной протеазы AC3 микробного происхождения, что и при использовании свиного трипсина, хроматографически очищенного из PTN 6,0 S.
Пример 5
Очистка и характеристика трипсиноподобной эндопептидазы из Actinosynnema mirum
Анализ на активность при очистке
Субстрат pNA: Suc-AAPR-pNA (Bachem L-1720).
Температура: комнатная температура (25°C).
Буфер для анализа: 100 мМ янтарная кислота, 100 мМ HEPES, 100 мМ CHES, 100 мМ CABS, 1 мМ CaCl2, 150 мМ KCl, 0,01% Triton X-100, pH 9,0.
По 20 мкм протеазы (разведенной в 0,01% Triton X-100) вносили в лунки планшета для микротитрования. Анализ начинали добавлением 200 мкл субстрата pNA (50 мг Suc-AAPR-pNA, разведенного в 1,0 мл ДМСО и дополнительно разведенного 90× буфером для анализа). Первоначальное увеличение OD405 отслеживали как показания протеазной активности.
Анализы для характеристики активности:
1) анализ pNA:
Субстраты pNA:
Suc-AAPR-pNA (Bachem L-1720)
Suc-AAPF-pNA (Bachem L-1400)
Suc-AAPA-pNA (Bachem L-1775)
Suc-AAPD-pNA (Bachem L-1835)
Suc-AAPE-pNA (Bachem L-1710)
Suc-AAPI-pNA (Bachem L-1790)
Suc-AAPL-pNA (Bachem L-1390)
Suc-AAPK-pNA (Bachem L-1725)
Suc-AAPM-pNA (Bachem L-1395)
Suc-AAPV-pNA (Bachem L-1770)
Температура: комнатная температура (25°C).
Буферы для анализа: 100 мМ янтарная кислота, 100 мМ HEPES, 100 мМ CHES, 100 мМ CABS, 1 мМ CaCl2, 150 мМ KCl, 0,01% Triton X-100, доведенный до pH 2,0, 3,0, 4,0, 5,0, 6,0, 7,0, 8,0, 9,0, 10,0 и 11,0 при помощи HCl или NaOH.
20 мкл протеазы (разведенной в 0,01% Triton X-100) смешивали с 100 мкл буфера для анализа. Анализ начинали добавлением 100 мкл субстрата pNA (50 мг разведенного в 1,0 мл ДМСО и дополнительно разведенного 45× в 0,01% Triton X-100). Первоначальное увеличение OD405 отслеживали как показания протеазной активности.
2) Анализ с Protazyme AK:
Субстрат: таблетка Protazyme AK (перекрестно сшитый и окрашенный казеин; от Megazyme).
Температура: контролируемая (температура анализа).
Буферы для анализа: 100 мМ янтарная кислота, 100 мМ HEPES, 100 мМ CHES, 100 мМ CABS, 1 мМ CaCl2, 150 мМ KCl, 0,01% Triton X-100, доведенный до pH 2,0, 2,5, 3,0, 3,5, 4,0, 5,0, 6,0, 7,0, 8,0, 9,0, 10,0 и 11,0 при помощи HCl или NaOH.
Таблетку Protazyme AK суспендировали в 2,0 мл 0,01 % Triton X-100 при осторожном перемешивании. 500 мкл этой суспензии и 500 мкл буфера для анализа смешивали в пробирке эппендорф и помещали на лед. Добавляли 20 мкл образца протеазы (разведенного в 0,01% Triton X-100). Анализ начинали перенесением пробирки эппендорф в термомиксер Eppendorf, установленный на температуру анализа. Пробирку инкубировали в течение 15 минут на термомиксере Eppendorf при самой высокой скорости встряхивания (1400 оборотов в минуту). Инкубацию останавливали перенесением пробирки обратно в ледяную баню. Затем пробирку центрифугировали в охлажденной до температуры льда центрифуге в течение нескольких минут и 200 мкл супернатанта переносили в планшет для микротитрования. Показатели OD650 являлись показателем протеазной активности. Пустой буфер включали в анализ (вместо фермента).
Очистка трипсиноподобной протеазы из штамма RBPO0013
RBPO0013 представляет собой штамм Bacillus subtilis, сконструированный для экспрессии в культуральную среду зрелой трипсиноподобной протеазы из Actinosynnema mirum, аминокислотная последовательность которой приведена в виде SEQ ID NO:5 по настоящему изобретению.
Культуральную жидкость от RBPO0013 центрифугировали (20000×g, 20 мин) и супернатанты осторожно сливали с осадков. Объединенные супернатанты фильтровали через 0,2-мкм фильтрационную ячейку Nalgene для удаления остатков клеток-хозяев Bacillus. Сухой сульфат аммония добавляли к 0,2-мкм фильтрату до конечной концентрации 1,8M (NH4)2SO4. Фильтрат осторожно перемешивали магнитной мешалкой в процессе добавления (NH4)2SO4, и перемешивание продолжали в течение 30 минут после добавления, чтобы привести систему в равновесие. Затем раствор наносили на колонку с Phenyl Toyopearl 650S (TosoHaas), уравновешенную в буфере 100 мМ H3BO3, 10 мМ MES, 2 мМ CaCl2, 1,8 M (NH4)2SO4, pH 6. После интенсивного промывания колонки уравновешивающим буфером трипсиноподобную протеазу элюировали линейным градиентом (NH4)2SO4 (1,6-->0M) в том же буфере. Фракции с колонки анализировали на трипсиноподобную протеазную активность (анализ на активность при очистке) и фракции, соответствующие основному пику активности, объединяли. Объединенные фракции с колонки Phenyl Toyopearl переводили в буфер 50 мМ H3BO3, 5 мМ MES, 1 мМ CaCl2, pH 6 на колонке с сефадексом G25 (GE Healthcare) и наносили на колонку с S-сефарозой FF (GE Healthcare), уравновешенную в том же буфере. После интенсивного промывания колонки уравновешивающим буфером протеазу элюировали линейным градиентом NaCl (0-->0,5M) в том же буфере. Фракции с колонки анализировали на трипсиноподобную протеазную активность (анализ на активность при очистке) и активные фракции дополнительно анализировали методом SDS-ПААГ. Фракции, которые выглядели как имеющие по меньшей мере 90% чистоту при окрашивании кумасси SDS-ПААГ геля, объединяли и использовали для дальнейшей характеристики.
Характеристика: зависимость активности от pH, зависимость стабильности от pH и зависимость активности от температуры
Анализ с pNA использовали для получения профиля зависимости активности от pH в отношении Suc-AAPR-pNA, а также для получения профиля зависимости стабильности от pH. Для профиля зависимости стабильности от pH протеазу разводили 10× в буферах для анализа и инкубировали в течение 2 часов при 37°C. После инкубации перед анализом на остаточную активность образцы протеазы доводили до pH 9 путем разведения в буфере для анализа с pH 9. Анализ с Protazyme AK использовали для получения профиля зависимости активности от температуры при pH 7. Результаты приведены ниже в таблицах 5-7. В таблице 5 активности приведены относительно активности при оптимальном pH для фермента. В таблице 6 активности представляют собой остаточные активности относительно образца, который хранили в стабильных условиях (5°C, pH 9). В таблице 7 активности приведены относительно активности при оптимальной температуре для фермента при pH 7.
Профиль зависимости активности от pH
Профиль зависимости стабильности от pH (остаточная активность через 2 часа при 37°C)
Профиль зависимости активности от температуры при pH 7,0
Характеристика: P1-специфичность в отношении субстратов Suc-AAPX-pNA и расчет трипсинового отношения
Анализ с pNA использовали для определения P1-специфичности для трипсиноподобной протеазы из Actinosynnema mirum, используя 10 субстратов Suc-AAPX-pNA при pH 9,0. Активности также использовали для расчета трипсинового отношения, как описано в WO 2010/112546. Результаты приведены ниже в таблице 8. В таблице 8 активность для каждого субстрата Suc-AAPX-pNA приведена относительно активности для наилучшего субстрата Suc-AAPX-pNA (Suc-AAPR-pNA).
P1-специфичность в отношении Suc-AAPX-pNA и трипсиновое отношение при pH 9,0
Другие характеристики
Трипсиноподобная протеаза из Actinosynnema mirum ингибируется PMSF.
Относительная молекулярная масса, определенная методом SDS-ПААГ, составляла Mr=26 кДа.
N-концевая последовательность была определена как: IVGGTRA.
Интактная молекулярная масса была определена как ММ=22460,9 Да.
Эти данные указывают на то, что зрелая трипсиноподобная протеаза из Actinosynnema mirum имеет следующую последовательность (аминокислоты 38-261 в SEQ ID NO:5):

Пример 6
Очистка и характеристика трипсиноподобной эндопептидазы из Kribbella flavida
Анализ на активность при очистке
Субстрат pNA: Suc-AAPR-pNA (Bachem L-1720).
Температура: комнатная температура (25°C).
Буфер для анализа: 100 мМ янтарная кислота, 100 мМ HEPES, 100 мМ CHES, 100 мМ CABS, 1 мМ CaCl2, 150 мМ KCl, 0,01% Triton X-100, pH 9,0.
По 20 мкм протеазы (разведенной в 0,01% Triton X-100) вносили в лунки планшета для микротитрования. Анализ начинали добавлением 200 мкл субстрата pNA (50 мг Suc-AAPR-pNA, разведенного в 1,0 мл ДМСО и дополнительно разведенного 90× буфером для анализа). Первоначальное увеличение OD405 отслеживали как показания протеазной активности.
Анализы для характеристики активности:
1) анализ pNA:
Субстраты pNA:
Suc-AAPR-pNA (Bachem L-1720)
Suc-AAPF-pNA (Bachem L-1400)
Suc-AAPA-pNA (Bachem L-1775)
Suc-AAPD-pNA (Bachem L-1835)
Suc-AAPE-pNA (Bachem L-1710)
Suc-AAPI-pNA (Bachem L-1790)
Suc-AAPL-pNA (Bachem L-1390)
Suc-AAPK-pNA (Bachem L-1725)
Suc-AAPM-pNA (Bachem L-1395)
Suc-AAPV-pNA (Bachem L-1770)
Температура: комнатная температура (25°C).
Буферы для анализа: 100 мМ янтарная кислота, 100 мМ HEPES, 100 мМ CHES, 100 мМ CABS, 1 мМ CaCl2, 150 мМ KCl, 0,01% Triton X-100, доведенный до pH 2,0, 3,0, 4,0, 5,0, 6,0, 7,0, 8,0, 9,0, 10,0 и 11,0 при помощи HCl или NaOH.
20 мкл протеазы (разведенной в 0,01% Triton X-100) смешивали с 100 мкл буфера для анализа. Анализ начинали добавлением 100 мкл субстрата pNA (50 мг разведенного в 1,0 мл ДМСО и дополнительно разведенного 45× в 0,01% Triton X-100). Первоначальное увеличение OD405 отслеживали как показания протеазной активности.
2) Анализ с Protazyme AK:
Субстрат: таблетка Protazyme AK (перекрестно сшитый и окрашенный казеин; от Megazyme).
Температура: контролируемая (температура анализа).
Буферы для анализа: 100 мМ янтарная кислота, 100 мМ HEPES, 100 мМ CHES, 100 мМ CABS, 1 мМ CaCl2, 150 мМ KCl, 0,01% Triton X-100, доведенный до pH 2,0, 2,5, 3,0, 3,5, 4,0, 5,0, 6,0, 7,0, 8,0, 9,0, 10,0 и 11,0 при помощи HCl или NaOH.
Таблетку Protazyme AK суспендировали в 2,0 мл 0,01 % Triton X-100 при осторожном перемешивании. 500 мкл этой суспензии и 500 мкл буфера для анализа смешивали в пробирке эппендорф и помещали на лед. Добавляли 20 мкл образца протеазы (разведенного в 0,01% Triton X-100). Анализ начинали перенесением пробирки эппендорф в термомиксер Eppendorf, установленный на температуру анализа. Пробирку инкубировали в течение 15 минут на термомиксере Eppendorf при самой высокой скорости встряхивания (1400 оборотов в минуту). Инкубацию останавливали перенесением пробирки обратно в ледяную баню. Затем пробирку центрифугировали в охлажденной до температуры льда центрифуге в течение нескольких минут и 200 мкл супернатанта переносили в планшет для микротитрования. Показатели OD650 являлись показателем протеазной активности. Пустой буфер включали в анализ (вместо фермента).
Очистка трипсиноподобной протеазы из штамма THFF0037
THFF0037 представляет собой штамм Bacillus subtilis, сконструированный для экспрессии в культуральную среду зрелой трипсиноподобной протеазы из Kribbella flavida, аминокислотная последовательность которой приведена в виде SEQ ID NO:6 по настоящему изобретению.
Культуральную жидкость от THFF0037 центрифугировали (20000×g, 20 мин) и супернатанты осторожно сливали с осадков. Объединенные супернатанты фильтровали через 0,2-мкм фильтрационную ячейку Nalgene для удаления остатков клеток-хозяев Bacillus. Сухой сульфат аммония добавляли к 0,2-мкм фильтрату до конечной концентрации 1,5M (NH4)2SO4. Фильтрат осторожно перемешивали магнитной мешалкой в процессе добавления (NH4)2SO4, и перемешивание продолжали в течение 30 минут после добавления, чтобы привести систему в равновесие. Затем раствор наносили на колонку с Phenyl Toyopearl 650S (TosoHaas), уравновешенную в буфере 100 мМ H3BO3, 10 мМ MES, 2 мМ CaCl2, 1,5 M (NH4)2SO4, pH 6. После интенсивного промывания колонки уравновешивающим буфером трипсиноподобную протеазу элюировали линейным градиентом (NH4)2SO4 (1,5-->0M) в том же буфере. Фракции с колонки анализировали на трипсиноподобную протеазную активность (анализ на активность при очистке) и фракции, соответствующие основному пику активности, объединяли. Объединенные фракции с колонки Phenyl Toyopearl переводили в буфер 50 мМ H3BO3, 5 мМ MES, 1 мМ CaCl2, pH 6 на колонке с сефадексом G25 (GE Healthcare) и pH доводили до pH 4,5 20% CH3COOH. Раствор с доведенным pH наносили на колонку SOURCE S (GE Healthcare), уравновешенную в буфере 10 мМ CH3COOH/NaOH, 1 мМ CaCl2, pH 4,5. После интенсивного промывания колонки уравновешивающим буфером протеазу элюировали линейным градиентом NaCl (0-->0,5M) в том же буфере. Фракции с колонки анализировали на трипсиноподобную протеазную активность (анализ на активность при очистке) и фракции, соответствующие пику активности, объединяли. Объединенные фракции с колонки SOURCE S наносили на колонку с Superdex 75 (GE Healthcare), уравновешенную в буфере 100 мМ H3BO3, 10 мМ MES, 2 мМ CaCl2, 100 мМ NaCl, pH 6. Колонку элюировали тем же буфером, фракции с колонки анализировали на трипсиноподобную протеазную активность (анализ на активность при очистке) и активные фракции дополнительно анализировали методом SDS-ПААГ. Фракции, которые выглядели как имеющие по меньшей мере 90% чистоту при окрашивании кумасси SDS-ПААГ геля, объединяли и использовали для дальнейшей характеристики.
Характеристика: зависимость активности от pH, зависимость стабильности от pH и зависимость активности от температуры
Анализ с pNA использовали для получения профиля зависимости активности от pH в отношении Suc-AAPR-pNA, а также для получения профиля зависимости стабильности от pH. Для профиля зависимости стабильности от pH протеазу разводили 10× в буферах для анализа и инкубировали в течение 2 часов при 37°C. После инкубации перед анализом на остаточную активность образцы протеазы доводили до pH 9 путем разведения в буфере для анализа с pH 9. Анализ с Protazyme AK использовали для получения профиля зависимости активности от температуры при pH 7. Результаты приведены ниже в таблицах 9-11. В таблице 9 активности приведены относительно активности при оптимальном pH для фермента. В таблице 10 активности представляют собой остаточные активности относительно образца, который хранили в стабильных условиях (5°C, pH 9). В таблице 11 активности приведены относительно активности при оптимальной температуре для фермента при pH 7.
Профиль зависимости активности от pH
Профиль зависимости стабильности от pH (остаточная активность через 2 часа при 37°C)
Профиль зависимости активности от температуры при pH 7,0
Характеристика: P1-специфичность в отношении субстратов Suc-AAPX-pNA и расчет трипсинового отношения
Анализ с pNA использовали для определения P1-специфичности для трипсиноподобной протеазы из Kribbella flavida, используя 10 субстратов Suc-AAPX-pNA при pH 9,0. Активности также использовали для расчета трипсинового отношения, как описано в WO 2010/112546. Результаты приведены ниже в таблице 12. В таблице 12 активность для каждого субстрата Suc-AAPX-pNA приведена относительно активности для наилучшего субстрата Suc-AAPX-pNA (Suc-AAPR-pNA).
P1-специфичность в отношении Suc-AAPX-pNA и трипсиновое отношение при pH 9,0
Другие характеристики
Трипсиноподобная протеаза из Kribbella flavida ингибируется EDTA.
Относительная молекулярная масса, определенная методом SDS-ПААГ, составляла Mr=26 кДа.
N-концевая последовательность была определена как: IVGGSL.
Интактная молекулярная масса была определена как ММ=23169,0 Да.
Эти данные указывают на то, что зрелая трипсиноподобная протеаза из Kribbella flavida имеет следующую последовательность (аминокислоты 40-266 в SEQ ID NO:6):

Рассчитанная на основании этой зрелой последовательности молекулярная масса составила 23168,7 Да.
Пример 7
Гидролиз концентрата белков молочной сыворотки и альфа-лактальбумина трипсиноподобными протеазами из Actinosynnema mirum и Kribbella flavida
Материалы
Концентрат белков молочной сыворотки (WPC), 80% сухой белок, лакпродан 80, Aria Foods Ingredients, DK.
Альфа-лактальбумин (ALA), 97% сухой белок, Davisco Foods International, MN, US.
CaCl2 безводный, Merck артикул 2387.
NaOH, Prolabo 31627.368.
Гидролизный анализ
Растворяли 1 г альфа-лактальбумина в 17,6 мл 5 мМ CaCl2 или 1,2 г лакпродана 80 в 17,4 мл 5 мМ CaCl2 для получения конечной концентрации белка 5%.
Образцы нагревали до 55°С или 50°C и рН доводили до 7,5. Объем NaOH, использованный для доведения pH, записывали. Добавляли фермент, и титрование pH проводили на Titralab 856 (радиометр) в течение 120 минут. Затраты NaOH подвергали мониторингу и переводили в %DH.
Степень гидролиза
Степень гидролиза суспензии измеряли при помощи pH-стата, как описано в Adler-Nissen, J. 1986, Enzymatic Hydrolysis of Food Proteins, глава 6.
Степень гидролиза (DH), определенную как процент расщепленных пептидных связей, можно отслеживать методом pH-статирования: DH=h/htot×100
h: число расщепленных пептидных связей, пропорциональное количеству основания, потребленного в процессе реакции;
htot: число пептидных связей в белке, рассчитанное на основании аминокислотного состава.
htot для ALA=8,16 и htot для WPC=8,8
h=B×Nb×1/α×1/Mp
B = потребление основания (мл);
Nb = нормальность основания (0,25 Н);
1/α = средняя степень диссоциации α-NH2 групп;
Mp = масса белка (г), (N × фактор Кьельдаля).
Использованные эндопептидазы:
Для информации об аминокислотных последовательностях трипсиноподобной протеазы из Fusarium oxysporum и химотрипсиноподобной протеазы из Nocardiopsis sp. NRRL 18262, см. WO 2010/112546.
Данные:
Депонирование биологического материала
Следующий биологический материал был депонирован в соответствии с условиями Будапештского договора компанией Deutsche Sammlung von Mikroorganismen und Zellkulturen GmbH (DSM), Mascheroder Weg 1 B, D-38124 Braunschweig, Germany, и получил следующий регистрационный номер:
Данный штамм были депонирован в условиях, которые гарантируют, что доступ к культуре будет предоставлен на время рассмотрения данной патентной заявки лицу, определенному иностранными патентными законами как лицо, имеющее на это право. Депозит представляет собой по существу чистую культуру депонированного штамма. Депозит доступен в соответствии с требованиями иностранных патентных законов в странах, где зарегистрированы эквиваленты или производные от данного объекта заявки. Тем не менее, следует понимать, что доступность депозита не является лицензией на применение на практике настоящего изобретения в нарушение патентных прав, предоставляемых действиями со стороны правительства.
Изобретение, описанное и заявленное в данном описании, не должно быть ограничено в объеме конкретными вариантами осуществления, раскрытыми в данном описании, поскольку данные варианты осуществления предназначены только для иллюстрации некоторых вариантов осуществления изобретения. Любые эквивалентные варианты осуществления должны входить в объем настоящего изобретения. Действительно, различные модификации изобретения в дополнение к тем, которые приведены и описаны в данном описании, будут очевидны для специалиста в данной области на основании приведенного выше описания. Такие модификации также должны входить в объем прилагаемой формулы изобретения. В случае конфликта настоящее описание изобретения, включая определения, будет иметь преимущественную силу.
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛИПЕПТИДЫ С ПРОТИВОМИКРОБНОЙ АКТИВНОСТЬЮ И КОДИРУЮЩИЕ ИХ ПОЛИНУКЛЕОТИДЫ | 2005 |
|
RU2393224C2 |
| ПРИМЕНЕНИЕ ВАРИАНТОВ ПРОТЕАЗЫ | 2010 |
|
RU2639534C2 |
| СПОСОБЫ УСИЛЕНИЯ ДЕГРАДАЦИИ ИЛИ ПРЕВРАЩЕНИЯ ЦЕЛЛЮЛОЗНОГО МАТЕРИАЛА | 2006 |
|
RU2441912C2 |
| ПРОТИВОМИКРОБНЫЕ ПОЛИПЕПТИДЫ ИЗ PSEUDOPLECTANIA NIGRELLA | 2002 |
|
RU2336278C2 |
| ВАРИАНТЫ СУБТИЛАЗ | 2010 |
|
RU2651525C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ ПРОТЕАЗНОЙ АКТИВНОСТЬЮ, И НУКЛЕИНОВЫЕ КИСЛОТЫ, КОДИРУЮЩИЕ УКАЗАННЫЕ ПОЛИПЕПТИДЫ | 2002 |
|
RU2318018C2 |
| МОЮЩАЯ КОМПОЗИЦИЯ | 2015 |
|
RU2737535C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ АНТИМИКРОБНОЙ АКТИВНОВНОСТЬЮ, И ПОЛИНУКЛЕОТИДЫ, КОДИРУЮЩИЕ ИХ | 2006 |
|
RU2512525C2 |
| СПОСОБЫ ПОВЫШЕНИЯ УСИЛИВАЮЩЕЙ ЦЕЛЛЮЛОЛИТИЧЕСКОЙ АКТИВНОСТИ ПОЛИПЕПТИДА | 2008 |
|
RU2510417C2 |
| ПОЛИПЕПТИДЫ С ЛИЗОЦИМНОЙ АКТИВНОСТЬЮ И ПОЛИНУКЛЕОТИДЫ, КОДИРУЮЩИЕ ИХ | 2012 |
|
RU2619051C2 |
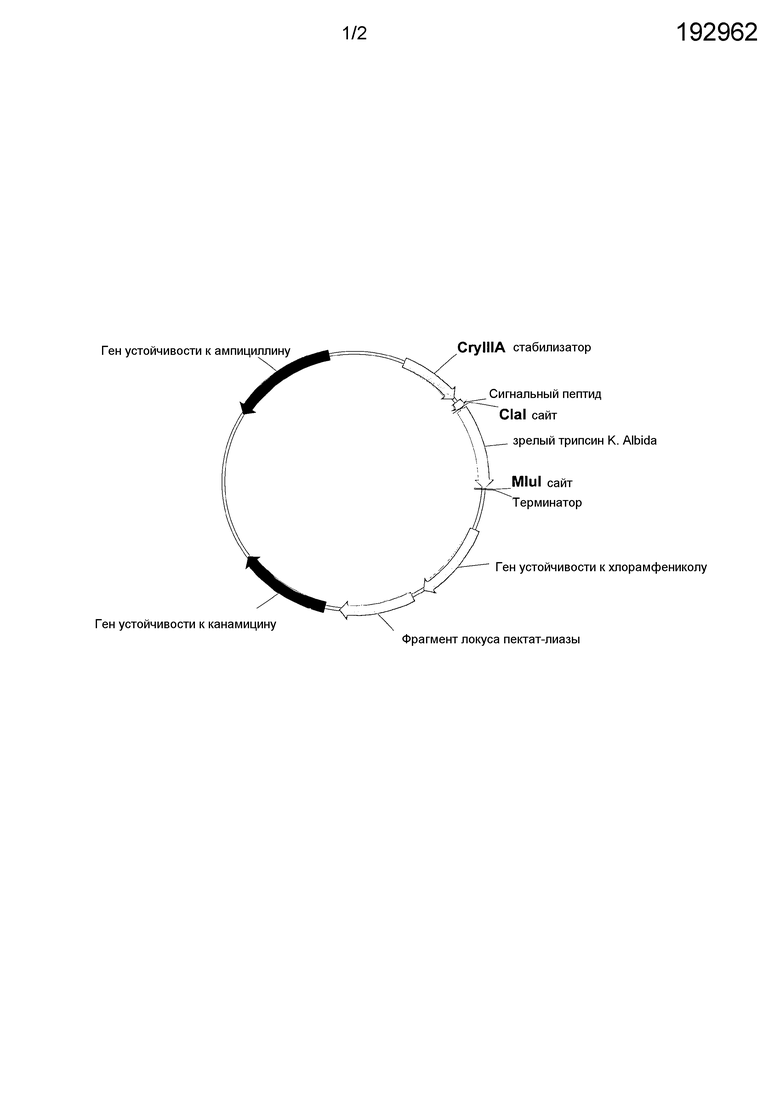
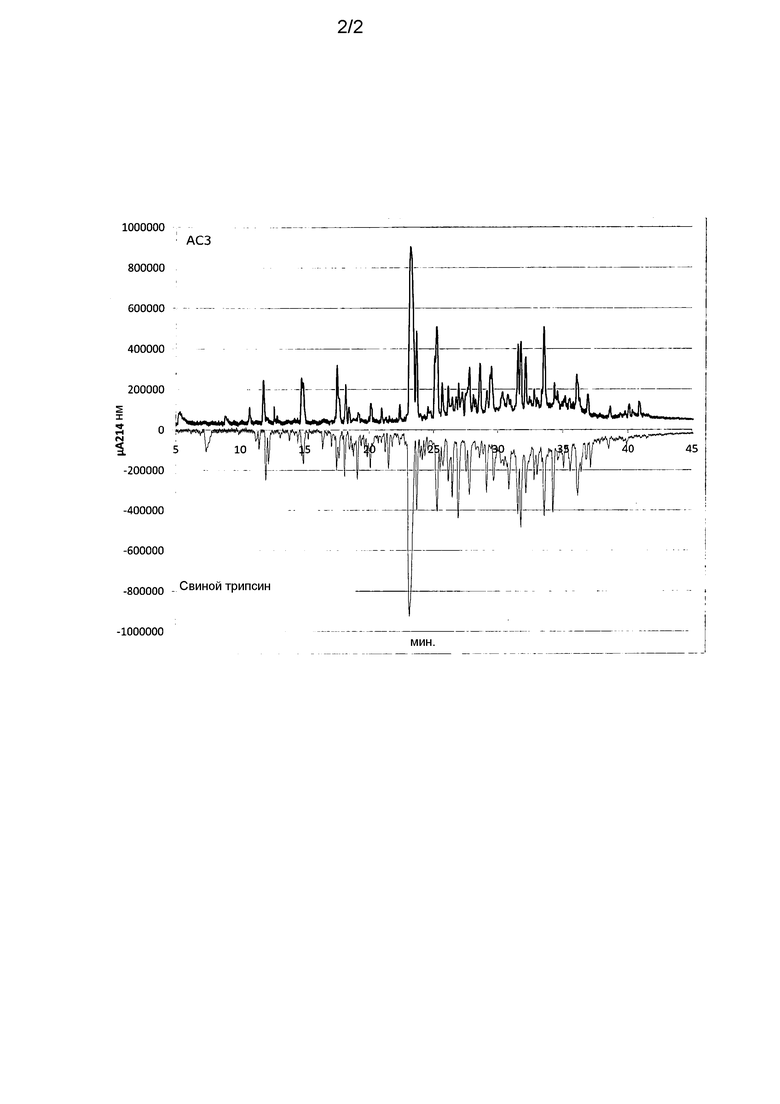









Изобретение относится к биотехнологии и представляет собой выделенный полипептид, обладающий трипсиноподобной эндопептидазной активностью. Изобретение касается также способа получения такого полипептида, включающего культивирование рекомбинантной клетки-хозяина Bacillus subtilis, содержащей полинуклеотид, кодирующий заявленный полипептид, функционально связанный с одной или более контрольными последовательностями, которые направляют продукцию полипептида в условиях, благоприятных для продукции полипептида; и выделения полипептида. Изобретение относится также к применению заявленного полипептида для получения гидролизата белка молочной сыворотки. Изобретение позволяет эффективно использовать трипсиноподобную эндопептидазу для гидролиза белка молочной сыворотки. 4 н. и 1 з.п. ф-лы, 2 ил., 12 табл., 7 пр.
1. Выделенный полипептид, обладающий трипсиноподобной эндопептидазной активностью, выбранный из группы, состоящей из:
(а) полипептида, имеющего по меньшей мере 90% идентичность последовательности со зрелым полипептидом SEQ ID NO: 2;
(b) полипептида, кодируемого полинуклеотидом, имеющим по меньшей мере 90% идентичность последовательности с кодирующей зрелый полипептид последовательностью SEQ ID NO: 1; и
(с) фрагмента полипептида по пунктам (а) или (b), обладающего эндопептидазной активностью.
2. Полипептид по п. 1, имеющий по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичность последовательности со зрелым полипептидом SEQ ID NO: 2.
3. Способ получения полипептида по любому из пп. 1 и 2, включающий:
(a) культивирование рекомбинантной клетки-хозяина Bacillus subtilis, содержащей полинуклеотид, кодирующий полипептид по любому из пп. 1 и 2, функционально связанный с одной или более контрольными последовательностями, которые направляют продукцию полипептида в условиях, благоприятных для продукции полипептида; и
(b) выделение полипептида.
4. Применение полипептида по любому из пп. 1 и 2 для получения гидролизата белка молочной сыворотки.
5. Способ получения гидролизата белка молочной сыворотки, включающий:
(a) получение раствора, содержащего белок молочной сыворотки, подлежащий гидролизу;
(b) добавление к указанному раствору полипептида по любому из пп. 1 и 2 и
(c) получение гидролизата белка молочной сыворотки.
| NAGAMINE-NATSUKA Y | |||
| et al | |||
| Molecular cloning, nucleotide sequence, and expression of the gene encoding a trypsin-like protease from Streptomyces erythraeus, J Biochem | |||
| Топка с качающимися колосниковыми элементами | 1921 |
|
SU1995A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Л.Я.ТЕЛИШЕВСКАЯ Белковые гидролизаты | |||
| Получение, состав, применение, Москва 2000, стр.78. | |||

