Область техники
Настоящее изобретение относится к микробиологической промышленности, в частности к способу получения L-аминокислоты с использованием бактерии семейства Enterobacteriaceae, модифицированной таким образом, что экспрессия гена ybiV указанной бактерии ослаблена.
Описание предшествующего уровня техники
Ген ybiV бактерии Escherichia coli кодирует белок YbiV, белок с гипотетической функцией, аминокислотная последовательность которого имеет гомологию с белками суперсемейства галоациддегалогеназы (HAD). Несмотря на то, что были идентифицированы многие члены этого семейства, только для некоторых из них известны функции. На основании рентгено-структурного анализа, анализа аминокислотной последовательности и биохимического анализа белок YbiV был охарактеризован как HAD-фосфатаза. С помощью рентгено-структурного анализа было показано, что белок YbiV состоит из двух доменов: одного домена с вторичной структурой, типичной для HAD-гидролазы, и другого домена с альфа/бета-структурой. Для выяснения механизма действия были изучены комплексы белка YbiV с фторидом бериллия (BeF3-) и с трифторидом алюминия (AlF3). Данные комплексы имитируют, соответственно, фосфорилированное промежуточное соединение и соединение в переходном состоянии в процессе гидролиза, по аналогии с другими HAD-фосфатазами. С помощью анализа данных структур была обнаружена впадина для связывания субстрата, обладающая гидрофильной природой. На основании анализа структуры белка YbiV и гомологии аминокислотной последовательности было сделано предположение о том, что данный белок может являться сахарной фосфатазой. Это предположение также подтверждается с помощью биохимического анализа на основании измерения высвобождения свободного фосфата с использованием ряда сахаро-подобных субстратов (Roberts, A. et al. YbiV from Escherichia coli K12 is a HAD phosphatase. Proteins, 2005, 58(4):790-801).
Но в настоящее время нет сообщений, описывающих использование инактивации гена ybiV для получения L-аминокислот.
Описание изобретения
Целями настоящего изобретения являются повышение продуктивности штаммов-продуцентов L-аминокислоты и представление способа получения L-аминокислоты с использованием этих штаммов.
Вышеупомянутые цели были достигнуты путем установления того факта, что инактивация гена ybiV может привести к повышению продукции L-аминокислот, таких как L-треонин, L-лизин, L-цистеин, L-лейцин, L-гистидин, L-глутаминовая кислота, L-фенилаланин, L-триптофан, L-пролин и L-аргинин.
Настоящее изобретение представляет бактерию семейства Enterobacteriaceae, обладающую способностью к повышенной продукции аминокислот, таких как L-треонин, L-лизин, L-цистеин, L-лейцин, L-гистидин, L-глутаминовая кислота, L-фенилаланин, L-триптофан, L-пролин и L-аргинин.
Целью настоящего изобретения является представление бактерии-продуцента L-аминокислоты семейства Enterobacteriaceae, модифицированной таким образом, что экспрессия гена ybiV указанной бактерии ослаблена.
Также целью настоящего изобретения является представление описанной выше бактерии, в которой ослабление экспрессии указанного гена ybiV осуществлено путем инактивации указанного гена ybiV.
Также целью настоящего изобретения является представление описанной выше бактерии, при этом указанная бактерия принадлежит к роду Escherichia.
Также целью настоящего изобретения является представление описанной выше бактерии, при этом указанная бактерия принадлежит к роду Pantoea.
Также целью настоящего изобретения является представление описанной выше бактерии, при этом указанная L-аминокислота выбрана из группы, состоящей из ароматической L-аминокислоты и неароматической L-аминокислоты.
Также целью настоящего изобретения является представление описанной выше бактерии, при этом ароматическая L-аминокислота выбрана из группы, состоящей из L-фенилаланина, L-тирозина и L-триптофана.
Также целью настоящего изобретения является представление описанной выше бактерии, при этом неароматическая L-аминокислота выбрана из группы, состоящей из L-треонина, L-лизина, L-цистеина, L-метионина, L-лейцина, L-изолейцина, L-валина, L-гистидина, L-глицина, L-серина, L-аланина, L-аспарагина, L-аспартата, L-глутамина, L-глутаминовой кислоты, L-пролина и L-аргинина.
Также целью настоящего изобретения является представление способа получения L-аминокислоты, который включает в себя:
- выращивание описанной выше бактерии в питательной среде с целью продукции и накопления L-аминокислоты в питательной среде и
- выделение указанной L-аминокислоты из культуральной жидкости.
Также целью настоящего изобретения является представление описанного выше способа, при этом указанная L-аминокислота выбрана из группы, состоящей из ароматической L-аминокислоты и неароматической L-аминокислоты.
Также целью настоящего изобретения является представление описанного выше способа, при этом указанная ароматическая L-аминокислота выбрана из группы, состоящей из L-фенилаланина, L-тирозина и L-триптофана.
Также целью настоящего изобретения является представление описанного выше способа, при этом указанная неароматическая L-аминокислота выбрана из группы, состоящей из L-треонина, L-лизина, L-цистеина, L-метионина, L-лейцина, L-изолейцина, L-валина, L-гистидина, L-глицина, L-серина, L-аланина, L-аспарагина, L-аспартата, L-глутамина, L-глутаминовой кислоты, L-пролина и L-аргинина.
Более детально настоящее изобретение описано ниже.
Подробное описание наилучшего способа осуществления изобретения
1. Бактерия согласно настоящему изобретению
Бактерия согласно настоящему изобретению - это бактерия-продуцент L-аминокислоты семейства Enterobacteriaceae, модифицированная таким образом, что экспрессия гена ybiV в указанной бактерии ослаблена.
Согласно настоящему изобретению "бактерия-продуцент L-аминокислоты" означает бактерию, обладающую способностью к продукции и выделению L-аминокислоты в питательную среду, когда бактерия согласно настоящему изобретению выращивается в указанной питательной среде.
Используемый здесь термин "бактерия-продуцент L-аминокислоты" также означает бактерию, которая способна к продукции L-аминокислоты и вызывает накопление L-аминокислоты в ферментационной среде в больших количествах, по сравнению с природным или родительским штаммом Е. coli, таким как штамм Е. coli К-12, и, предпочтительно означает, что указанный микроорганизм способен накапливать в среде целевую L-аминокислоту в количестве не менее чем 0.5 г/л, более предпочтительно, не менее чем 1.0 г/л. Термин "L-аминокислота" включает в себя L-аланин, L-аргинин, L-аспарагин, L-аспарагиновую кислоту, L-цистеин, L-глутаминовую кислоту, L-глутамин, L-глицин, L-гистидин, L-изолейцин, L-лейцин, L-лизин, L-метионин, L-фенилаланин, L-пролин, L-серин, L-треонин, L-триптофан, L-тирозин и L-валин.
Термин "ароматическая L-аминокислота" включает в себя L-фенилаланин, L-тирозин и L-триптофан. Термин "неароматическая L-аминокислота" включает в себя L-треонин, L-лизин, L-цистеин, L-метионин, L-лейцин, L-изолейцин, L-валин, L-гистидин, L-глицин, L-серин, L-аланин, L-аспарагин, L-аспартат, L-глутамин, L-глутаминовую кислоту, L-пролин и L-аргинин. Наиболее предпочтительны L-треонин, L-лизин, L-цистеин, L-лейцин, L-гистидин, L-глутаминовая кислота, L-фенилаланин, L-триптофан, L-пролин и L-аргинин.
Семейство Enterobacteriaceae включает в себя бактерии, принадлежащие к родам Escherichia, Enterobacter, Erwinia, Klebsiella, Pantoea, Photorhabdus, Providencia, Salmonella, Serratia, Shigella, Morganella, Yersinia и т.д. Более конкретно, могут быть использованы бактерии, классифицируемые как принадлежащие к семейству Enterobacteriaceae в соответствии с таксономией, используемой в базе данных NCBI (National Center for Biotechnology Information)
(http://www.ncbi.nlm.nih.gov/htbinpost/Taxonomy/wgetorg?mode=Tree&id=1236&lvl=3&keep=1&srchmode=1&unlock). Бактерия, принадлежащая к родам Escherichia или Pantoea, предпочтительна.
Термин "бактерия, принадлежащая к роду Escherichia" означает, что бактерия относится к роду Escherichia в соответствии с классификацией, известной специалисту в области микробиологии. В качестве примера микроорганизма, принадлежащего к роду Escherichia, использованного в настоящем изобретении, может быть упомянута бактерия Escherichia coli (E. coli).
Круг бактерий, принадлежащих к роду Escherichia, которые могут быть использованы в настоящем изобретении, не ограничен каким-либо образом, однако, например, бактерии, описанные в книге Neidhardt, F.C. et al. (Escherichia coli and Salmonella typhimurium, American Society for Microbiology, Washington D.C., 1208, Таблица 1), могут быть включены в число бактерий согласно настоящему изобретению.
Термин "бактерия, принадлежащая к роду Pantoea" означает, что бактерия относится к роду Pantoea в соответствии с классификацией, известной специалисту в области микробиологии. Недавно несколько видов Enterobacter agglomerans были классифицированы как Pantoea agglomerans, Pantoea ananatis, Pantoea stewartii или подобные им, на основе анализа нуклеотидной последовательности 16S рРНК и т.д. (Int. J. Syst. BacterioL, 1993, 43:162-173).
Термин "бактерия модифицирована таким образом, что экспрессия гена ybiV ослаблена" означает, что указанная бактерия была модифицирована таким образом, что в результате модификации такая бактерия содержит пониженное количество белка YbiV по сравнению с немодифицированной бактерией или указанная бактерия не способна синтезировать белок YbiV.
Термин "бактерия модифицирована таким образом, что экспрессия гена ybiV ослаблена" также означает, что целевой ген модифицирован таким образом, что кодирует мутантный белок (белки), обладающие пониженной активностью.
Термин "инактивация гена ybiV" означает, что указанный ген модифицирован таким образом, что такой модифицированный ген кодирует полностью неактивный белок. Также возможно, что естественная экспрессия модифицированного участка ДНК невозможна из-за делеции целевого гена или его части, сдвига рамки считывания данного гена, введения missense/nonsense мутации (мутаций) или модификации прилегающих к гену областей, которые включают последовательности, контролирующие экспрессию гена, такие как промотор(ы), энхансер(ы), аттенуатор(ы), сайт(ы) связывания рибосомы, и т.д.
Ген ybiV кодирует белок YbiV, HAD-фосфатазу (синоним - b0822). Ген ybiV (номера нуклеотидов с 859251 по 858436 в нуклеотидной последовательности с инвентарным номером NC_000913.2 в базе данных GenBank; gi: 49175990; SEQ ID NO: 1) расположен на хромосоме штамма Е.coli K-12 между генами ybiU и ybiW. Нуклеотидная последовательность гена ybiV и соответствующая ей аминокислотная последовательность белка YbiV, кодируемого геном ybiV, приведены в Списке последовательностей под номерами 1 (SEQ ID NO: 1) и 2 (SEQ ID NO: 2), соответственно.
Поскольку у представителей различных родов и штаммов семейства Enterobacteriaceae возможны некоторые вариации в нуклеотидных последовательностях, понятие инактивируемого гена ybiV не ограничивается геном, последовательность которого приведена в Списке последовательностей под номером 1 (SEQ ID No: 1), но также может включать и гены, гомологичные последовательности, приведенной в Списке последовательностей под номером 1 (SEQ ID No: 1). Таким образом, вариант белка, кодируемого геном ybiV, может быть представлен белком с гомологией не менее 80%, предпочтительно, не менее 90%, и, наиболее предпочтительно, не менее 95%, по отношению к полной аминокислотной последовательности, приведенной в Списке последовательностей под номером 2 (SEQ ID NO. 2), при условии, что до инактивации сохраняется способность белка YbiV к регуляции транскрипции ряда генов.
Кроме того, ген ybiV может быть представлен вариантом, который гибридизуется в жестких условиях с нуклеотидной последовательностью, приведенной в Списке последовательностей под номером 1 (SEQ ID NO: 1), или с зондом, который может быть синтезирован на основе указанной нуклеотидной последовательности, при условии, что указанный вариант до инактивации кодирует функциональный белок YbiV. "Жесткие условия" включают такие условия, при которых специфические гибриды образуются, а неспецифические гибриды не образуются. Практическим примером жестких условий является однократная отмывка, предпочтительно двух- или трехкратная, при концентрации солей, соответствующей стандартным условиям отмывки при гибридизации по Саузерну, например, 1×SSC, 0.1% SDS, предпочтительно 0.1×SSC, 0.1% SDS, при 60°С. Длина зонда может быть выбрана в зависимости от условий гибридизации, обычно она составляет от 100 п.н. до 1 т.п.н.
Инактивация указанного гена может быть произведена традиционными методами, такими как мутагенез с использованием УФ-излучения или обработка нитрозогуанидином (N-метил-N'-нитро-N-нитрозогуанидин), сайт-направленный мутагенез, инактивация гена с помощью гомологичной рекомбинации, или/и инсерционно-делеционного мутагенеза (Yu, D. et al., Proc. Natl. Acad. Sci. USA, 2000, 97:12: 5978-83 и Datsenko K.A. and Wanner B.L., Proc. Natl. Acad. Sci. USA, 2000, 97:12:6640-45), также называемого "Red-зависимой интеграцией".
Наличие активности белка YbiV может быть определено путем комплементации мутации ybiV, к примеру, с помощью метода, описанного Roberts, A. et al. YbiV from Escherichia coli K12 is a HAD phosphatase. Proteins, 2005, 58(4):790-801. Таким образом, снижение или отсутствие активности белка YbiV в бактерии согласно настоящему изобретению может быть определено путем сравнения указанной бактерии с родительской немодифицированной бактерией. Кроме того, уровень экспрессии гена можно оценить путем измерения количества мРНК, транскрибируемой с целевого гена, с использованием различных известных методик, включая гибридизацию по Нозерну (Northern blotting), количественный метод ОТ-ПЦР (RT-PCR) и подобные им. Количество белка, кодируемого данным геном, может быть измерено с помощью известных методов, включающих метод SDS-PAGE с последующим иммуноблотингом (Western blotting) и подобные им.
Методами получения плазмидной ДНК, разрезания и лидирования ДНК, трансформации, выбора олигонуклеотидов в качестве праймеров и подобными им могут являться обычные методы, хорошо известные специалисту в данной области. Эти методы описаны, например, в книге Sambrook, J., Fritsch, E.F., and Maniatis, Т., "Molecular Cloning: A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press (1989).
Бактерия-продуцент L-аминокислоты
В качестве бактерии согласно настоящему изобретению, модифицированной таким образом, что экспрессия гена ybiV ослаблена, может быть использована бактерия, способная к продукции ароматической или неароматической L-аминокислоты.
Бактерия согласно настоящему изобретению может быть получена путем инактивации гена ybiV в бактерии, уже обладающей способностью к продукции L-аминокислот. С другой стороны, бактерия согласно настоящему изобретению может быть получена путем придания бактерии, в которой ген ybiV уже инактивирован, способности к продукции L-аминокислот.
Бактерия-продуцент L-треонина
Примеры родительского штамма для получения бактерии-продуцента L-треонина согласно настоящему изобретению включают, но не ограничиваются штаммами, принадлежащими к роду Escherichia, такими как штамм Е. coli TDH-6/p VIC40 (ВКПМ В-3996) (патенты США 5175107 и 5705371), штамм Е. coli NRRL-21593 (патент США 5939307), штамм Е. coli FERM BP-3756 (патент США 5474918), штаммы Е. coli FERM ВР-3519 и FERM BP-3520 (патент США 5376538), штамм Е. coli MG442 (Гусятинер и др., Генетика, 14, 947-956 (1978)), штаммы Е.coli VL643 и VL2055 (Европейская патентная заявка ЕР 1149911 А) и подобные им.
Штамм TDH-6 является дефектным по гену thrC, способен ассимилировать сахарозу и содержит ген ilvA с мутацией типа "leaky". Указанный штамм содержит мутацию в гене rhtA, которая обуславливает устойчивость к высоким концентрациям треонина и гомосерина. Штамм В-3996 содержит плазмиду pVIC40, которая была получена путем введения в вектор, производный от вектора RSF1010, оперона thrA*BC, включающего мутантный ген thrA, кодирующий аспартокиназа-гомосериндегидрогеназу I, у которой существенно снижена чувствительность к ингибированию треонином по типу обратной связи. Штамм В-3996 был депонирован 19 ноября 1987 года во Всесоюзном научном центре антибиотиков (РФ, 117105 Москва, Нагатинская ул., 3-А) с инвентарньм номером РИА 1867. Указанный штамм также был депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) (РФ, 117545 Москва, 1-й Дорожный проезд, 1) с инвентарным номером В-3996.
Предпочтительно, чтобы бактерия согласно настоящему изобретению была далее модифицирована таким образом, чтобы иметь повышенную экспрессию одного или нескольких следующих генов:
- мутантного гена thrA, кодирующего аспартокиназа-гомосериндегидрогеназу I, устойчивую к ингибированию треонином по типу обратной связи;
- гена thrB, кодирующего гомосеринкиназу;
- гена thrC, кодирующего треонинсинтазу;
- гена rhtA, предположительно кодирующего трансмембранный белок;
- гена asd, кодирующего аспартат-β-семиальдегиддегидрогеназу, и
- гена aspC, кодирующего аспартатаминотрансферазу (аспартаттрансаминазу).
Нуклеотидная последовательность гена thrA, кодирующего аспартокиназа-гомосериндегидрогеназу I из Escherichia coli, известна (номера нуклеотидов с 337 по 2799 в последовательности с инвентарным номером NC_00913.2 в базе данных GenBank, gi: 49175990). Ген thrA расположен на хромосоме штамма Е. coli K-12 между генами thrL и thrB. Нуклеотидная последовательность гена thrB, кодирующего гомосеринкиназу из Escherichia coli, известна (номера нуклеотидов с 2801 по 3733 в последовательности с инвентарным номером NC_000913.2 в базе данных GenBank, gi: 49175990). Ген thrB расположен на хромосоме штамма Е. coli K-12 между генами thrA и thrC. Нуклеотидная последовательность гена thrC, кодирующего треонинсинтазу из Escherichia coli, известна (номера нуклеотидов с 3734 по 5020 в последовательности с инвентарным номером NC_000913.2 в базе данных GenBank, gi: 49175990). Ген thrC расположен на хромосоме штамма Е. coli K-12 между геном thrB и открытой рамкой считывания уааХ. Все три указанных гена функционируют как один треониновый оперон.
Мутантный ген thrA, кодирующий аспартокиназу-гомосериндегидрогеназу I, устойчивую к ингибированию треонином по типу обратной связи, так же, как и гены thrB и thrC, могут быть получены в виде единого оперона из хорошо известной плазмиды pVIC40, которая представлена в штамме-продуценте Е. coli ВКПМ В-3996. Плазмида pVIC40 подробно описана в патенте США 5705371.
Ген rhtA расположен на 18 минуте хромосомы Е. coli около оперона glnHPQ, который кодирует компоненты транспортной системы глутамина, ген rhtA идентичен ORF1 (ген ybiF, номера нуклеотидов с 764 по 1651 в последовательности с инвентарным номером ААА218541 в базе данных GenBank, gi: 440181), расположен между генами рехВ и ompX. Участок ДНК, экспрессирующийся с образованием белка, кодируемого рамкой считывания ORF1, был назван геном rhtA (rht: resistance to homoserine and threonine). Также было показано, что мутация rhtA23 представляет собой замену А-на-G в положении -1 по отношению к старт кодону ATG (ABSTRACTS of 17th International Congress of Biochemistry and Molecular Biology in conjugation with 1997 Annual Meeting of the American Society for Biochemistry and Molecular Biology, San Francisco, California August 24-29, 1997, abstract No.457, EP 1013765 A).
Нуклеотидная последовательность гена asd из E. coli известна (номера нуклеотидов с 3572511 по 3571408 в последовательности с инвентарным номером NC_000913.1 в базе данных GenBank, gi: 16131307) и может быть получена с помощью ПЦР (полимеразная цепная реакция; ссылка на White, T.J. et al., Trends Genet., 5, 185 (1989)) с использованием праймеров, синтезированных на основе нуклеотидной последовательности указанного гена. Гены asd из других микроорганизмов могут быть получены сходным образом.
Также нуклеотидная последовательность гена aspC из E. coli известна (номера нуклеотидов с 983742 по 984932 в последовательности с инвентарным номером NC_000913.1 в базе данных GenBank, gi: 16128895) и может быть получена с помощью ПЦР. Гены aspC из других микроорганизмов могут быть получены сходным образом.
Бактерия-продуцент L-лизина
Примеры бактерий-продуцентов L-лизина, принадлежащих к роду Escherichia, включают мутанты, обладающие устойчивостью к аналогу L-лизина. Аналог L-лизина ингибирует рост бактерий, принадлежащих к роду Escherichia, но это ингибирование полностью или частично снимается, когда в среде также присутствует L-лизин. Примеры аналога L-лизина включают, но не ограничиваются оксализином, лизингидроксаматом, S-(2-аминоэтил)-L-цистеином (AEC), γ-метиллизином, α-хлорокапролактамом и так далее. Мутанты, обладающие устойчивостью к указанным аналогам лизина, могут быть получены путем обработки бактерий, принадлежащих к роду Escherichia, традиционными мутагенами. Конкретные примеры бактериальных штаммов, используемых для получения L-лизина, включают штамм Escherichia coli AJ11442 (FERM BP-1543, NRRL В-12185; см. патент США 4346170) и штамм Escherichia coli VL611. В этих микроорганизмах аспартокиназа устойчива к ингибированию L-лизином по принципу обратной связи.
Штамм WC196 может быть использован в качестве бактерии-продуцента L-лизина Escherichia coli. Данный бактериальный штамм был получен путем селекции фенотипа устойчивости к АЕС у штамма W3110, производного от штамма Escherichia coli К-12. Полученный штамм был назван Escherichia coli AJ13069 и был депонирован в Национальном Институте Биологических Наук и Человеческих Технологий, Агенство Промышленной Науки и Технологии (National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology), в настоящее время называющийся Национальный Институт Прогрессивной Промышленной Науки и Технологии, Международный Депозитарий Организмов для Целей Патентования, Централ 6,1-1, Хигаши 1-Чоме, Тсукуба-ши, Ибараки-кен, 305-8566, Япония (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary, Central 6,1-1, Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, 305-8566, Japan), 6 декабря 1994 года и получил инвентарный номер FERM Р-14690. Затем было произведено международное депонирование этого штамма согласно условиям Будапештского Договора 29 сентября 1995 года, и штамм получил инвентарный номер FERM BP-5252 (см. патент США 5827698).
Бактерия-продуцент L-цистеина
Примеры родительских штаммов, используемых для получения бактерии-продуцента L-цистеина согласно настоящему изобретению, включают в себя, но не ограничиваются штаммами, принадлежащими к роду Escherichia, такими как штамм Е. coli JM15, трансформированный различными аллелями гена cysE, кодирующими устойчивые к ингибированию по типу обратной связи серинацетилтрансферазы (патент США 6218168, патентная заявка РФ 2003121601); штамм Е. coli W3110, содержащий гены с повышенной экспрессией, кодирующие белок, способный к секреции соединений, токсичных для клетки (патент США 5972663); штаммы Е. coli, содержащие цистеиндесульфогидразу со сниженной активностью (патент Японии JP 11155571А2); штамм Е. coli W3110 с повышенной активностью позитивного транскрипционного регулятора цистеинового регулона, кодируемого геном cysB (международная заявка РСТ WO 0127307 A1) и подобные им.
Бактерия-продуцент L-лейцина
Примеры родительских штаммов, используемых для получения бактерии-продуцента L-лейцина согласно настоящему изобретению, включают в себя, но не ограничиваются штаммами, принадлежащими к роду Escherichia, такими как штаммы Е. coli, устойчивые к аналогам лейцина, включающие, например, β-2-тиенилаланин, 3-гидроксилейцин, 4-азалейцин и 5,5,5-трифлуоролейцин (выложенные патентные заявки Японии 62-34397 и 8-70879), штаммы Е. coli, полученные с помощью генно-инженерных методов, описанных в заявке РСТ 96/06926; Е. coli штамм Н-9068 (JP 08-70879 А), и подобные им.
Бактерия согласно настоящему изобретению может быть улучшена путем усиления экспрессии одного или нескольких генов, вовлеченных в биосинтез L-лейцина. Примеры таких генов включают в себя гены оперона leuABCD, и предпочтительно представлены мутантным геном leuA, кодирующим изопропилмалатсинтазу со снятым ингибированием L-лейцином по типу обратной связи (патент США 6403342). Кроме того, бактерия согласно настоящему изобретению может быть улучшена путем усиления экспрессии одного или нескольких генов, кодирующих белки, которые экспортируют L-аминокислоту из бактериальной клетки. Примеры таких генов включают в себя гены b2682 и b2683 (гены ygaZH) (патентная заявка РФ 2001117632).
Бактерия-продуцент L-гистидина
Примеры родительских штаммов, используемых для получения бактерии-продуцента L-гистидина согласно настоящему изобретению, включают в себя, но не ограничиваются бактериями-продуцентами L-гистидина, принадлежащими к роду Escherichia, такими как штамм Е. coli 24 (ВКПМ В-5945, патент РФ 2003677); штамм Е. coli 80 (ВКПМ В-7270, патент РФ 2119536); штаммы Е. coli NRRL В-12116 - В12121 (патент США 4388405); штаммы Е. coli Н-9342 (FERM ВР-6675) и Н-9343 (FERM ВР-6676) (патент США 6344347); штамм Е. coli H-9341 (FERM BP-6674) (Европейский патент 1085087); штамм Е. coli AI80/pFM201 (патент США 6258554) и подобные им.
Бактерия-продуцент L-глутаминовой кислоты
Примеры родительских штаммов, используемых для получения бактерии-продуцента L-глутаминовой кислоты согласно настоящему изобретению, включают в себя, но не ограничиваются штаммами, принадлежащими к роду Escherichia, такими как штамм Е. coli VL334thrC+ (Европейский патент ЕР 1172433). Штамм Е. coli VL334 (ВКПМ В-1641) является ауксотрофом по L-изолейцину и L-треонину с мутациями в генах thrC и ilvA (патент США 4278765). В этот штамм была перенесена природная аллель гена thrC методом общей трансдукции с использованием бактериофага Р1, выращенного на клетках природного штамма Е. coli K12 (ВКПМ В-7). В результате был получен штамм, ауксотроф по L-изолейцину, VL334thrC+ (ВКПМ В-8961). Этот штамм обладает способностью к продукции L-глутаминовой кислоты.
Примеры родительских штаммов, используемых для получения бактерии-продуцента L-глутаминовой кислоты согласно настоящему изобретению, включают в себя мутантные штаммы, лишенные активности α-кетоглутаратдегидрогеназы или обладающие сниженной активностью α-кетоглутаратдегидрогеназы. Бактерии, принадлежащие к роду Escherichia, лишенные активности α-кетоглутаратдегидрогеназы или обладающие сниженной активностью α-кетоглутаратдегидрогеназы, и способы их получения описаны в патентах США 5378616 и 5573945. Конкретно, примеры таких штаммов включают в себя следующие штаммы:
E. coli W3110sucA::Kmr
Е. coli AJ12624 (FERM ВР-3853)
Е. coli AJ12628 (FERM BP-3854)
Е. coli AJ12949 (FERM BP-4881)
Штамм Е. coli W3110sucA::Kmr был получен в результате разрушения гена α-кетоглутаратдегидрогеназы (далее называемого "ген sucA") в штамме Е. coli W3110. У этого штамма активность α-кетоглутаратдегидрогеназы отсутствует полностью.
Другие примеры бактерии-продуцента L-глутаминовой кислоты включают в себя бактерии, принадлежащие к роду Pantoea, которые лишены активности α-кетоглутаратдегидрогеназы или имеют сниженную активность α-кетоглутаратдегидрогеназы и могут быть получены описанным выше способом. Примерами таких штаммов являются штамм Pantoea ananatis AJ13356 (патент США 6331419), штамм Pantoea ananatis AJ13356, депонированный в Национальном Институте Биологических Наук и Человеческих Технологий, Агенство Промышленной Науки и Технологии, Министерство Международной Торговли и Промышленности (National Institute of Bioscience and Human-Technology, Agency of Industrial Science and Technology, Ministry of International Trade and Industry), в настоящее время называющийся Национальный Институт Прогрессивной Промышленной Науки и Технологии, Международный Депозитарий Организмов для Целей Патентования, Централ 6, 1-1, Хигаши 1-Чоме, Тсукуба-ши, Ибараки-кен, 305-8566, Япония (National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary, Central 6,1-1, Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, 305-8566, Japan), 19 февраля 1998 года и получивший инвентарный номер FERM Р-16645. Затем было произведено международное депонирование этого штамма согласно условиям Будапештского Договора от 11 января 1999 года, и штамм получил инвентарный номер FERM ВР-6615. В штамме Pantoea ananatis AJ13356 отсутствует активность α-KGDH в результате разрушения гена субъединицы αKGDH-E1 (sucA). Вышеупомянутый штамм при выделении был идентифицирован как Enterobacter agglomerans и депонирован как штамм Enterobacter agglomerans AJ13355. Тем не менее, позднее он был классифицирован как Pantoea ananatis на основе нуклеотидной последовательности 16S рРНК и других доказательств (см. раздел Примеры). Несмотря на то, что оба штамма - AJ13355 и полученный из него штамм AJ13356 - были депонированы в указанный выше депозитарий как Enterobacter agglomerans, для целей данного описания они будут упоминаться как Pantoea ananatis.
Бактерия-продуцент L-фенилаланина
Примеры родительских штаммов, используемых для получения бактерии-продуцента L-фенилаланина согласно настоящему изобретению, включают в себя, но не ограничиваются штаммами, принадлежащими к роду Escherichia, такими как штамм E. coli AJ12739 (tyrA::Tn10, tyrR) (ВКМП В-8197); штамм E. coli HW1089 (АТСС-55371), содержащий ген pheA34 (патент США 5354672); мутантный штамм E. coli MWEC101-b (KR8903681); штаммы E. coli NRRL B-12141, NRRL B-12145, NRRL В-12146 и NRRL B-12147 (патент США 4407952) и подобные им. Также в качестве родительских штаммов могут быть использованы бактерии, принадлежащие к роду Escherichia, - продуценты L-фенилаланина, такие как штамм E.coli K-12[W3110(tyrA)/pPHAB] (FERM BP-3566), штамм E. coli K-12[W3110(tyrA)/pPHAD] (FERM BP-12659). штамм E. coli K-12[W3110(tyrA)/pPHATerm] (FERM BP-12662) и штамм E. coli K-12[W3110(tyrA)/pBR-aroG4, рАСМАВ], названный как AJ12604 (FERM BP-3579) (Европейский патент ЕР 488424 В1). Кроме того, также могут быть использованы бактерии-продуценты L-фенилаланина, принадлежащие к роду Escherichia с повышенной активностью белков, кодируемых геном yedA или геном yddG (патентные заявки США 2003/0148473 А1 и 2003/0157667 А1).
Бактерия-продуцент L-триптофана
Примеры родительских штаммов, используемых для получения бактерии-продуцента L-триптофана согласно настоящему изобретению, включают в себя, но не ограничиваются бактериями-продуцентами L-триптофана, принадлежащими к роду Escherichia, такими как штаммы Е. coli JP4735/pMU3028 (DSM10122) и JP6015/pMU91 (DSM10123), лишенные активности триптофанил-тРНК синтетазы, кодируемой мутантным геном trpS (патент США 5756345); штамм Е. coli SV164 (pGH5), содержащий аллель гена serA, кодирующего фермент, не ингибируемый серином по типу обратной связи (патент США 6180373); штаммы Е. coli AGX17 (pGX44) (NRRL В-12263) и AGX6(pGX50)aroP (NRRL В-12264), лишенные активности триптофаназы (патент США 4371614); штамм Е. coli AGX17/pGX50,pACKG4-pps, в котором усилена способность к синтезу фосфоенолпирувата (международная заявка 9708333, патент США 6319696), и подобные им.
Ранее было показано, что природная аллель гена yddG, кодирующего мембранный белок, не участвующий в путях биосинтеза ни одной из L-аминокислот, амплифицированная на многокопийном векторе в микроорганизме, придает этому микроорганизму устойчивость к L-фенилаланину и нескольким аналогам этой аминокислоты. Кроме того, введение в клетки бактерий-продуцентов L-фенилаланина или L-триптофана дополнительных копий гена yddG может положительно влиять на продукцию соответствующих аминокислот (международная заявка РСТ WO 03044192). Таким образом, желательно, чтобы бактерия-продуцент L-триптофана была далее модифицирована таким образом, что в этой бактерии усилена экспрессия открытой рамки считывания yddG.
Бактерия-продуцент L-пролина
Примеры бактерий-продуцентов L-пролина, используемых в качестве родительского штамма согласно настоящему изобретению, включают в себя, но не ограничиваются штаммами, принадлежащими к роду Escherichia, такими как штамм Е. coli 702ilvA (ВКПМ В-8012), дефицитного по гену ilvA и способного к продукции L-пролина (Европейский патент ЕР 1172433). Бактерия согласно настоящему изобретению может быть улучшена путем усиления экспрессии одного или нескольких генов, вовлеченных в биосинтез L-пролина. Предпочтительно, примеры таких генов для бактерий-продуцентов L-пролина, включают ген proB, кодирующий глутаматкиназу с десенсибилизированной регуляцией L-пролином по типу обратной связи (патент Германии 3127361). Кроме того, бактерия согласно настоящему изобретению может быть улучшена путем усиления экспрессии одного или нескольких генов, кодирующих белки, экскретирующие L-аминокислоту из бактериальной клетки. Примерами таких генов являются гены b2682 и b2683 (ygaZH гены) (Европейская патентная заявка ЕР 1239041 А2).
Примеры бактерий, принадлежащих к роду Escherichia и обладающих способностью к продукции L-пролина, включают следующие штаммы Е. coli: NRRL В-12403 и NRRL В-12404 (патент Великобритании GB 2075056), ВКПМ В-8012 (патентная заявка РФ 2000124295), плазмидные мутанты, описанные в патенте Германии DE 3127361, плазмидные мутанты, описанные у Bloom F.R. et al (The 15th Miami winter symposium, 1983, p.34), и подобные им.
Бактерия-продуцент L-аргинина
Примеры родительских штаммов, используемых для получения бактерии-продуцента L-аргинина согласно настоящему изобретению, включают в себя, но не ограничиваются штаммами, принадлежащими к роду Escherichia, такими как штамм Е. coli 237 (ВКПМ В-7925) (патентная заявка США US 2002058315) и его производные, содержащие мутантную N-ацетилглутаматсинтазу (патентная заявка РФ 2001112869), штамм Е. coli 382 (ВКПМ В-7926) (Европейская патентная заявка ЕР 1170358), штамм-продуцент аргинина, в который введен ген argA, кодирующий N-ацетилглутаматсинтетазу (выложенная патентная заявка Японии 57-5693 А), и подобные им.
2. Способ согласно настоящему изобретению
Способом согласно настоящему изобретению является способ получения L-аминокислоты, включающий стадии выращивания бактерии согласно настоящему изобретению в питательной среде с целью продукции и накопления L-аминокислоты в питательной среде, и выделения L-аминокислоты из культуральной жидкости.
Согласно настоящему изобретению выращивание, выделение и очистка L-аминокислоты из культуральной или подобной ей жидкости могут быть осуществлены способом, подобным традиционным способам ферментации, в которых аминокислота продуцируется с использованием бактерии.
Питательная среда, используемая для выращивания, может быть как синтетической, так и натуральной, при условии, что указанная среда содержит источники углерода, азота, минеральные добавки и, если необходимо, соответствующее количество питательных добавок, необходимых для роста микроорганизмов. К источникам углерода относятся различные углеводы, такие как глюкоза и сахароза, а также различные органические кислоты. В зависимости от характера ассимиляции используемого микроорганизма могут использоваться спирты, такие как этанол и глицерин. В качестве источника азота могут использоваться различные неорганические соли аммония, такие как аммиак и сульфат аммония, другие соединения азота, такие как амины, природные источники азота, такие как пептон, гидролизат соевых бобов, ферментолизат микроорганизмов. В качестве минеральных добавок могут использоваться фосфат калия, сульфат магния, хлорид натрия, сульфат железа, сульфат марганца, хлорид кальция и подобные им соединения. В качестве витаминов могут использоваться тиамин, дрожжевой экстракт и подобные им соединения.
Выращивание осуществляется предпочтительно в аэробных условиях, таких как перемешивание культуральной жидкости на качалке, взбалтывание с аэрацией, при температуре в пределах от 20 до 40°С, предпочтительно в пределах от 30 до 38°С. рН среды поддерживают в пределах от 5 до 9, предпочтительно от 6.5 до 7.2. рН среды может регулироваться аммиаком, карбонатом кальция, различными кислотами, основаниями и буферными растворами. Обычно выращивание в течение от 1 до 5 дней приводит к накоплению целевой L-аминокислоты в культуральной жидкости.
После выращивания твердые остатки, такие как клетки, могут быть удалены из культуральной жидкости методом центрифугирования или фильтрацией через мембрану, а затем L-аминокислота может быть выделена и очищена методами ионообменной хроматографии, концентрирования и/или кристаллизации.
Краткое описание чертежей
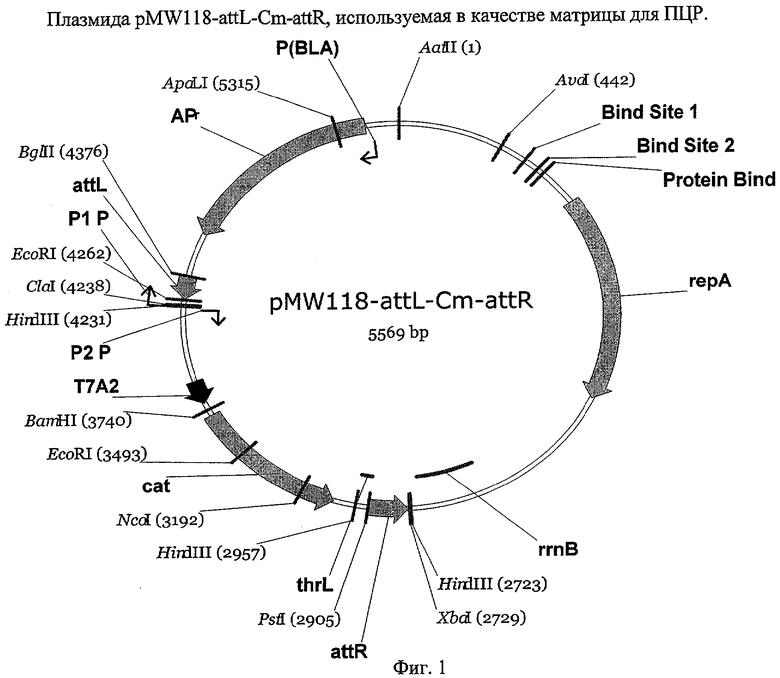
На Фиг.1 изображено конструирование плазмиды pMW118-attL-Cm-attR, используемой в качестве матрицы для ПЦР.
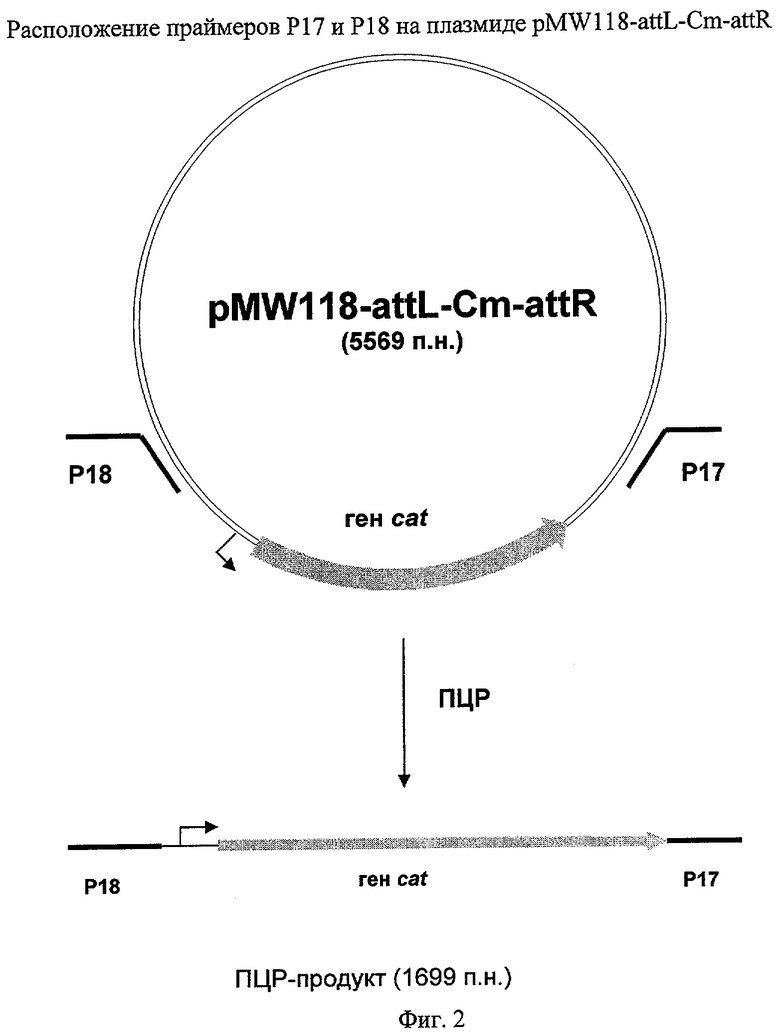
На Фиг.2 изображены относительные положения праймеров Р17 и Р18 на плазмиде pMW118-attL-Cm-attR, используемой для ПЦР-амплификации гена cat.
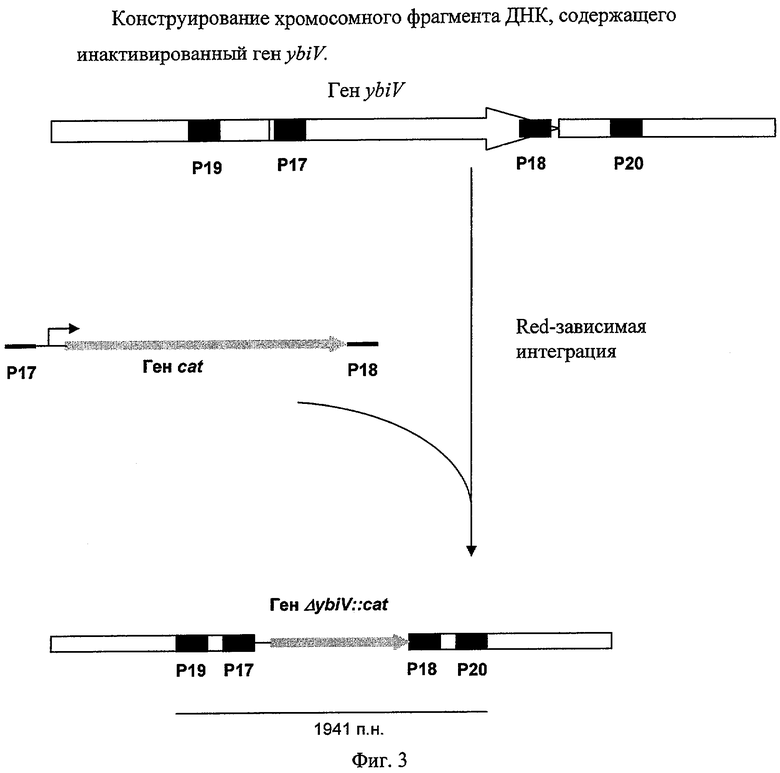
На Фиг.3 изображено конструирование фрагмента хромосомной ДНК, содержащего инактивированный ген ybiV.
Примеры
Настоящее изобретение будет более подробно описано ниже со ссылкой на следующие неограничивающие настоящее изобретение Примеры.
Пример 1. Конструирование ПЦР-матриды и хелперных плазмид
Плазмида pMW118-attL-Cm-attR, используемая в качестве матрицы для ПЦР, и хелперная плазмида pMW-intxis-ts были получены как описано ниже.
(1) pMW118-attL-Cm-attR
Плазмида pMW118-attL-Cm-attR была сконструирована на основе плазмиды pMW118-attL-Tc-attR, полученной путем сшивки следующих четырех фрагментов ДНК:
1) фрагмента BglII-EcoRI (114 п.н.), несущего attL (SEQ ID NO: 3), который был получен путем ПЦР-амплификации соответствующего участка хромосомы штамма Е. coli W3350 (содержащего профаг λ) с использованием олигонуклеотидов Р1 и Р2 (SEQ ID NOS: 4 и 5) в качестве праймеров (данные праймеры содержали дополнительные сайты узнавания для рестриктаз BglII и EcoRI);
2) фрагмента PstI-HindIII (182 п.н.), несущего attR (SEQ ID NO: 6), который был получен путем ПЦР-амплификации соответствующего участка хромосомы штамма Е. coli W3350 (содержащего профаг λ) с использованием олигонуклеотидов Р3 и Р4 (SEQ ID NOS: 7 и 8) в качестве праймеров (данные праймеры содержали дополнительные сайты узнавания для рестриктаз PstI и HindIII);
3) большого фрагмента BglII-HindIII (3916 п.н.) плазмиды pMW118-ter_rrnB. Плазмида pMW118-ter_rrnB была получена путем сшивки следующих трех фрагментов ДНК:
- большого ДНК-фрагмента (2359 п.н.), несущего фрагмент AatII-EcoRI плазмиды pMW118, полученный следующим способом: pMW118 расщепляли с помощью рестриктазы EcoRI, обрабатывали с помощью фрагмента Кленова ДНК-полимеразы I (Klenow fragment of DNA polymerase I), а затем расщепляли с помощью рестриктазы AatII;
- малого фрагмента AatII-BglII (1194 п.н.) плазмиды pUC19, несущего ген устойчивости к ампициллину bla (АрR), который был получен путем ПЦР-амплификации соответствующего участка плазмиды pUC19 с использованием олигонуклеотидов Р5 и Р6 (SEQ ID NOS: 9 и 10) в качестве праймеров (данные праймеры содержали дополнительные сайты узнавания для рестриктаз AatII и BglII);
- малого фрагмента BglII-PstIpol (363 п.н.) терминатора транскрипции ter_rrnB, полученного ПЦР-амплификацией соответствующего участка хромосомы штамма Е. coli MG1655, используя олигонуклеотиды Р7 и Р8 (SEQ ID NOS: 11 и 12) в качестве праймеров (данные праймеры содержали дополнительные сайты узнавания для рестриктаз BglII и PstI);
4) малого фрагмента EcoRI-PstI (1388 п.н.) (SEQ ID NO: 13) плазмиды pML-Tc-tet_thrL, несущей ген устойчивости к тетрациклину и терминатор транскрипции ter_thrL; плазмида pML-Tc-ter_thrL была получена в два этапа:
- плазмида pML-ter_thrL была получена путем расщепления плазмиды pML-MCS (Mashko, S.V. et al., Biotekhnologiya (in Russian), 2001, no.5, 3-20) с помощью рестриктаз Xbal and BamHl и последующей сшивки большого фрагмента (3342 п.н.) и фрагмента XbaI-BamHI (68 п.н.), несущего терминатор ter_thrL и полученного путем ПЦР-амплификации соответствующего участка хромосомы штамма Е. coli MG1655 с использованием олигонуклеотидов Р9 и Р10 (SEQ ID NOS: 14 и 15) в качестве праймеров (данные праймеры содержали дополнительные сайты узнавания для рестриктаз XbaI и BamHI);
- плазмида pML-Tc-ter_thrL была получена путем расщепления плазмиды pML-ter_thrL с помощью рестриктаз KpnI и XbaI и последующей обработки с помощью фрагмента Кленова ДНК-полимеразы I (Klenow fragment of DNA polymerase I) и сшивки с малым фрагментом EcoRI-Van91I (1317 п.н.) плазмиды pBR322, несущей ген устойчивости к тетрациклину (pBR322 расщепляли с помощью рестриктаз EcoRI и Van91I и затем обрабатывали с помощью фрагмента Кленова ДНК-полимеразы I).
Плазмида pMW118-attL-Cm-attR была сконструирована путем сшивки большого фрагмента BamHI-XbaI (4413 п.н.) плазмиды pMW118-attL-Tc-attR и синтетического DNA-фрагмента BglII-XbaI (1162 п.н.), содержащего промотор PA2 (ранний промотор фага Т7), ген устойчивости к хлорамфениколу cat (CmR), терминатор транскрипции ter_thrL и участок attR. Синтетический DNA-фрагмент (SEQ ID NO: 16) был получен следующим образом:
1. Плазмиду pML-MCS расщепляли при помощи рестриктаз KpnI и XbaI и сшивали с малым фрагментом KpnI-XbaI (120 п.н.), содержащим промотор РA2 (ранний промотор фага Т7) и полученным путем ПЦР-амплификации соответствующего участка ДНК фага Т7 с использованием олигонуклеотидов Р11 и P12 (SEQ ID NOS: 17 и 18, соответственно) в качестве праймеров (данные праймеры содержали дополнительные сайты узнавания для рестриктаз KpnI и XbaI). В результате была получена плазмида pML-PA2-MCS.
2. Сайт XbaI был удален из плазмиды pML-РA2-MCS, в результате чего была получена плазмида pML-PA2-MCS(XbaI-).
3. Малый фрагмент BglII-HindIII (928 п.н.) плазмиды pML-PA2-MCS(XbaI-), содержащий промотор РA2 (ранний промотор фага Т7) и ген устойчивости к хлорамфениколу cat (CmR), был сшит с малым фрагментом HindIII-HindIII (234 п.н.) плазмиды pMW118-attL-Tc-attR, содержащим терминатор транскрипции ter_thrL и участок attR.
4. Синтетический DNA-фрагмент (1156 п.н.) был получен путем ПЦР-амплификации лигированной смеси с использованием олигонуклеотидов Р9 и Р4 (SEQ ID NOS: 14 и 8) в качестве праймеров (данные праймеры содержали дополнительные сайты узнавания для рестриктаз HindIII и XbaI).
(2) pMW-intxis-ts
Рекомбинантная плазмида pMW-intxis-ts, содержащая ген-репрессор cI и гены int-xis фага λ под контролем промотора PR, была сконструирована на основе вектора pMWPlaclacI-ts. Вектор pMWPlaclacI-ts был получен путем замещения фрагмента AatII-EcoRV плазмиды pMWPlaclacI (Skorokhodova, A.Yu. et al., Biotekhnologiya (in Russian), 2004, no.5, 3-21) на фрагмент AatII-EcoRV плазмиды pMAN997 (Tanaka, К. et al., J. BacterioL, 2001, 183(22): 6538-6542), несущий локусы par и ori, а также ген repAts репликона pSC101.
Два ДНК-фрагмента были амплифицированы с использованием ДНК фага λ ("Fermentas") в качестве матрицы. Первый ДНК-фрагмент содержал последовательность ДНК с 37168 по 38046 нуклеотид, ген-репрессор cI, промоторы РRM и PR и лидерную последовательность гена его. Данный ДНК-фрагмент был амплифицирован с помощью ПЦР с использованием олигонуклеотидов Р13 и Р14 (SEQ ID NOS: 19 и 20) в качестве праймеров. Второй ДНК-фрагмент, содержащий гены xis-int фага λ и последовательность DNA с 27801 по 29100 нуклеотид, был амплифицирован с помощью ПЦР с использованием олигонуклеотидов Р15 и Р16 (SEQ ID NOS: 21 и 22) в качестве праймеров. Все праймеры содержали соответствующие рестрикционные сайты.
Первый ДНК-фрагмент, полученный с помощью ПЦР-амплификации и несущий ген-репрессор cI, был расщеплен с помощью рестриктазы ClaI, обработан с помощью фрагмента Кленова ДНК-полимеразы I (Klenow fragment of DNA polymerase I) и затем расщеплен с помощью рестриктазы EcoRI. Второй ДНК-фрагмент, полученный с помощью ПЦР-амплификации, был расщеплен с помощью EcoRI и PstI. Плазмида pMWPlaclacI-ts была расщеплена с помощью рестриктазы BglII, обработана с помощью фрагмента Кленова ДНК-полимеразы I (Klenow fragment of DNA polymerase I) и затем расщеплена с помощью рестриктазы PstI. Векторный фрагмент плазмиды pMWPlaclacI-ts был элюирован из агарозного геля и сшит с двумя вышеупомянутыми ДНК-фрагментами, в результате чего была получена рекомбинантная плазмида pMW-intxis-ts.
Пример 2. Конструирование штамма с инактивированным геном ybiV
1. Делеция гена ybiV
Штамм, содержащий делецию гена ybiV, был сконструирован с использованием методики, разработанной Datsenko, K.A. и Wanner, B.L. (Proc. Natl. Acad. Sci. USA, 2000, 97(12), 6640-6645), известной как "Red-зависимая интеграция". Фрагмент ДНК, содержащий маркер CmR, кодируемый геном cat, был получен при помощи ПЦР с использованием праймеров Р17 (SEQ ID NO: 23) и Р18 (SEQ ID NO: 24) и плазмиды pMW118-attL-Cm-attR в качестве матрицы (конструирование плазмиды описано в Примере 1). Праймер Р17 содержит участок ДНК размером в 36 н., комплементарный участку ДНК, расположенному на 5'-конце гена ybiV, а также участок ДНК, комплементарный участку attL. Праймер Р18 содержит участок ДНК размером в 35 н., комплементарный участку ДНК, расположенному на 3'-конце гена ybiV, а также участок ДНК, комплементарный участку attR. Использовали следующий температурный профиль для ПЦР: денатурация при 95°С в течение 3 мин; два первых цикла: 1 мин при 95°С, 30 сек при 50°С, 40 сек при 72°С; последующие 25 циклов: 30 сек при 95°С, 30 сек при 54°С, 40 сек при 72°С; и заключительная полимеризация: 5 мин при 72°С.
Полученный ПЦР-продукт длиной 1699 п.н. (Фиг.2), очищенный в агарозном геле, был использован для электропорации в штамм Е. coli MG1655 (АТСС 700926), содержащий плазмиду pKD46 с термочувствительным репликоном. Плазмида pKD46 (Datsenko, K.A. and Wanner, B.L., Proc. Natl. Acad. Sci. USA, 2000, 97(12):6640-6645) содержит ДНК-фрагмент фага λ длиной 2154 п.н. (позиции с 31088 по 33241 нуклеотидной последовательности с инвентарным номером J02459 в базе данных GenBank), а также содержит гены λ Red-гомологичной системы рекомбинации (гены γ, β, ехо) под контролем промотора РaraB, индуцируемого арабинозой. Плазмида pKD46 необходима для интеграции продукта ПЦР в хромосому штамма MG1655.
Электрокомпетентные клетки были получены следующим образом: ночную культуру штамма Е. coli MG1655 выращивали при 30°С в среде LB с добавкой ампициллина (100 мг/л), разводили в 100 раз, добавив 5 мл среды SOB (Sambrook et al, "Molecular Cloning: A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press, 1989), содержащей ампициллин и L-арабинозу (1 мМ). Полученную культуру растили с перемешиванием при 30°С до достижения OD600≈0.6, после чего делали клетки электрокомпетентными путем концентрирования в 100 раз и трехкратного отмывания ледяной деионизированной Н2О. Электропорацию проводили с использованием 70 мкл клеток и ≈100 нг ПЦР-продукта. После электропорации клетки инкубировали в 1 мл среды SOC (Sambrook et al, "Molecular Cloning: A Laboratory Manual, Second Edition", Cold Spring Harbor Laboratory Press, 1989) при 37°С в течение 2.5 часов, после чего высевали на чашки с L-агаром и выращивали при 37°С для отбора CmR-рекомбинантов. Затем для удаления плазмиды pKD46 проводили 2 пассажа на L-агаре с Cm при 42°С и полученные колонии проверяли на чувствительность к ампициллину.
2. Подтверждение делеции гена ybiV с помощью ПЦР
Мутанты с делетированным геном ybiV, содержащие ген устойчивости Cm, были проверены с помощью ПЦР. Локус-специфичные праймеры Р19 (SEQ ID NO: 25) и Р20 (SEQ ID NO: 26) были использованы для проверки делеции с помощью ПЦР. Использовался следующий температурный профиль для ПЦР-проверки: денатурация при 94°С в течение 3 мин; профиль для 30 циклов: 30 сек при 94°С, 30 сек при 54°С, 1 мин при 72°С; заключительный шаг: 7 мин при 72°С. Длина продукта ПЦР, полученного в результате реакции с использованием в качестве матрицы клеток родительского штамма ybiV+ MG1655, составляет 945 п.н. Длина продукта ПЦР, полученного в результате реакции с использованием в качестве матрицы клеток мутантного штамма MG1655 ΔybiV::cat, составляет 1941 п.н. (Фиг.3).
Пример 3. Продукция L-треонина штаммом Е. coli B-3996-ΔybiV.
Для оценки влияния инактивации гена ybiV на продукцию треонина ДНК-фрагменты хромосомы описанного выше штамма Е. coli MG1655 ΔybiV::cat были перенесены в штамм-продуцент L-треонина Е. coli В-3996 (ВКПМ В-3996) с помощью P1-трансдукции (Miller, J.H. (1972) Experiments in Molecular Genetics, Cold Spring Harbor Lab. Press, Plainview, NY).
Оба штамма Е.Coli, B-3996 и В-3996-ΔybiV, были выращены в течение 18-24 часов при температуре 37°С на чашках с L-агаром, содержащим хлорамфеникол (30 мкг/мл). Для получения посевной культуры указанные штаммы были выращены при 32°С в течение 18 часов на роторной качалке (250 об/мин) в пробирках размером 20×200 мм, содержащих 2 мл L-бульона с 4% сахарозой. Затем в ферментационную среду было внесено по 0.21 мл (10%) посевной культуры. Ферментация была проведена в 2 мл минимальной ферментационной среды в пробирках размером 20×200 мм. Клетки были выращены в течение 65 часов при 32°С с перемешиванием (250 об/мин).
После выращивания количество накопленного в среде L-треонина было определено с помощью бумажной хроматографии, с использованием подвижной фазы следующего состава: бутанол:уксусная кислота:вода=4:1:1 (v/v). Раствор (2%) нингидрина в ацетоне был использован для визуализации. Пятно, содержащее L-треонин, было вырезано; L-треонин был элюирован 0.5% водным раствором CdCl2, после чего количество L-треонина было оценено спектрофотометрическим методом при длине волны 540 нм. Результаты десяти независимых пробирочных ферментаций приведены в Таблице 1. Как следует из Таблицы 1, штамм В-3996-ΔybiV накапливал большее количество L-треонина по сравнению со штаммом В-3996.
Была использована ферментационная среда следующего состава (г/л):
Глюкозу и сульфат магния стерилизовали отдельно. СаСО3 стерилизовали сухим жаром при 180°С в течение 2 часов. рН доводили до 7.0. Антибиотик добавляли в среду после стерилизации.
Пример 4. Продукция L-лизина штаммом Е. coli AJ11442-ΔybiV
Для оценки влияния инактивации гена ybiV продукцию лизина ДНК-фрагменты хромосомы описанного выше штамма Е. coli MG1655 ΔybiV::cat могут быть перенесены в штамм-продуцент L-лизина Е. coli WC196 (pCABD2) с помощью Р1-трансдукции (Miller, J.H. (1972) Experiments in Molecular Genetics, Cold Spring Harbor Lab. Press, Plainview, NY). pCABD2 - это плазмида, содержащая ген dapA, кодирующий мутантную дигидропиколинатсинтазу, устойчивую к ингибированию L-лизином по типу обратной связи, ген lysC, кодирующий мутантную аспартокиназу III, устойчивую к ингибированию L-лизином по типу обратной связи, ген dapB, кодирующий дигидропиколинатредуктазу, и ген ddh, кодирующий диаминопимелатдегидрогеназу (патент США 6040160).
Оба штамма Е. coli, WC196(pCABD2) и WC196(pCABD2)-ΔybiV, могут быть выращены в L-среде, содержащей 50 мг/л хлорамфеникола и 20 мг/л стрептомицина, при 37°С; и 0.3 мл полученных культур может быть внесено в 20 мл ферментационной среды, содержащей необходимые антибиотики, в колбы объемом 500 мл. Культивирование может производиться при 37°С в течение 16 часов с использованием возвратно-поступательной качалки со скоростью перемешивания 115 об/мин. После выращивания количество L-лизина и остаточной глюкозы в среде может быть измерено известным способом (Biotech-analyzer AS210, производитель - Sakura Seiki Co.). Затем для каждого из штаммов может быть рассчитан выход L-лизина в пересчете на потребленную глюкозу.
Может быть использована ферментационная среда следующего состава (г/л):
рН доводят до 7.0 с помощью КОН и среду автоклавируют при 115°С в течение 10 мин. Глюкозу и MgSO4×7H2O стерилизуют отдельно. Также добавляют СаСО3 до концентрации 30 г/л, предварительно простерилизованного сухим жаром при 180°С в течение 2 часов.
Пример 5. Продукция L-цистеина штаммом Е. coli JM15(ydeD)-ΔybiV
Для оценки влияния инактивации гена ybiV на продукцию L-цистеина ДНК-фрагменты хромосомы описанного выше штамма Е. coli MG1655 ΔybiV::cat могут быть перенесены в штамм-продуцент L-цистеина Е. coli JM15(ydeD) с помощью Р1-трансдукции (Miller, J.H. (1972) Experiments in Molecular Genetics, Cold Spring Harbor Lab. Press, Plainview, NY), в результате чего может быть получен штамм JM15(ydeD)-ΔybiV.
Штамм Е.coli JM15(ydeD) является производным штамма Е. coli JM15 (патент США 6218168), который может быть трансформирован ДНК, содержащей ген ydeD, кодирующий мембранный белок, не вовлеченный в пути биосинтеза ни одной из L-аминокислот (патент США 5972663).
Условия ферментации для оценки продукции L-цистеина детально описаны в Примере 6 патента США 6218168.
Пример 6. Продукция L-лейцина штаммом Е. coli 57-ΔybiV
Для оценки влияния инактивации гена ybiV на продукцию L-лейцина ДНК-фрагменты хромосомы описанного выше штамма Е. coli MG1655 ΔybiV::cat могут быть перенесены в штамм-продуцент L-лейцина Е. coli 57 (ВКПМ В-7386, патент США 6124121) с помощью P1-трансдукции (Miller, J.H. (1972) Experiments in Molecular Genetics, Cold Spring Harbor Lab. Press, Plainview, NY), в результате чего может быть получен штамм 57-pMW-ΔybiV.
Оба штамма Е. coli, 57 и 57-ΔybiV, могут быть выращены в течение 18-24 часов при температуре 37°С на чашках с L-агаром, содержащим хлорамфеникол (30 мкг/мл). Для получения посевной культуры указанные штаммы могут быть выращены на роторной качалке (250 об/мин) при 32°С в течение 18 часов в пробирках размером 20×200 мм, содержащих 2 мл L-бульона с 4% сахарозы. Затем в ферментационную среду может быть внесено по 0.21 мл (10%) посевной культуры. Ферментацию можно проводить в 2 мл минимальной ферментационной среды в пробирках размером 20×200 мм. Клетки могут быть выращены в течение 48-72 часов при 32°С с перемешиванием (250 об/мин). Количество L-лейцина может быть измерено с помощью бумажной хроматографии (состав подвижной фазы:бутанол-уксусная кислота-вода=4:1:1).
Может быть использована ферментационная среда (рН 7.2) следующего состава (г/л):
Глюкозу и мел стерилизуют отдельно.
Пример 7. Продукция L-гистидина штаммом Е. coli 80-ΔybiV
Для оценки влияния инактивации гена ybiV на продукцию L-гистидина ДНК-фрагменты хромосомы описанного выше штамма Е. coli MG1655 ΔybiV::cat могут быть перенесены в штамм-продуцент L-гистидина Е. coli 80 с помощью P1-трансдукции (Miller, J.H. (1972) Experiments in Molecular Genetics, Cold Spring Harbor Lab. Press, Plainview, NY). Штамм 80 описан в патенте РФ 2119536 и депонирован во Всероссийской коллекции промышленных микроорганизмов (Россия, 117545 Москва, 1-й Дорожный проезд, 1) с инвентарным номером ВКПМ В-7270.
Оба штамма Е. coli, 80 и 80-ΔybiV, могут быть выращены на L-бульоне, содержащим хлорамфеникол (30 мг/мл), при 29°С в течение 6 часов. Затем по 0.1 мл полученных культур может быть внесено в 2 мл ферментационной среды в пробирки размером 20×200 мм, и культуры могут быть выращены при 29°С в течение 65 часов на роторной качалке (350 об/мин). После выращивания количество накопленного в среде гистидина может быть определено с помощью бумажной хроматографии. Может быть использована подвижная фаза следующего состава: n-бутанол-уксусная кислота-вода=4:1:1 (v/v). Раствор нингидрина (0.5%) в ацетоне может быть использован для визуализации.
Может быть использована ферментационная среда (рН 6.0) следующего состава (г/л):
Глюкозу, пролин, бетаин и СаСО3 стерилизуют отдельно. рН доводят до 6.0 перед стерилизацией.
Пример 8. Продукция L-глутаминовой кислоты штаммом Е. coli VL334thrC+-ΔybiV
Для оценки влияния инактивации гена ybiV на продукцию L-глутаминовой кислоты ДНК-фрагменты хромосомы описанного выше штамма Е. coli MG1655 ΔybiV::cat могут быть перенесены в штамм-продуцент L-глутаминовой кислоты Е. VL334thrC+ (EP 1172433) с помощью P1-трансдукции (Miller, J.H. (1972) Experiments in Molecular Genetics, Cold Spring Harbor Lab. Press, Plainview, NY), в результате чего может быть получен штамм VL334thrC+-ΔybiV.
Оба штамма, VL334thrC+ и VL334thrC+-ΔybiV, могут быть выращены на чашках с L-агаром, содержащим хлорамфеникол (30 мкг/мл), при 37°С в течение 18-24 часов. Далее одна петля клеток может быть перенесена в пробирки, содержащие 2 мл ферментационной среды. Ферментационная среда может содержать глюкозу - 60 г/л, сульфат аммония - 25 г/л, КН2PO4 - 2 г/л, MgSO4 - 1 г/л, тиамин - 0.1 мг/мл, L-изолейцин - 70 мкг/мл и мел - 25 г/л (рН 7.2). Глюкозу и мел стерилизуют отдельно. Выращивание может производиться при 30°С в течение 3 дней с перемешиванием. После выращивания количество полученной L-глутаминовой кислоты может быть определено с помощью бумажной хроматографии (состав подвижной фазы:бутанол-уксусная кислота-вода=4:1:1) с последующим окрашиванием нингидрином (1% раствор в ацетоне) и дальнейшим элюированием полученных соединений в 50% этаноле с 0.5% CdCl2
Пример 9. Продукция L-фенилаланина штаммом Е. coli АJ12739-ΔybiV
Для оценки влияния инактивации гена ybiV на продукцию L-фенилаланина ДНК-фрагменты хромосомы описанного выше штамма Е. coli MG1655 ΔybiV::cat могут быть перенесены в штамм-продуцент L-фенилаланина Е. coli AJ12739 с помощью Р1-трансдукции (Miller, J.H. (1972) Experiments in Molecular Genetics, Cold Spring Harbor Lab. Press, Plainview, NY). Штамм АJ12739 депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) (Россия, 117545 Москва, 1ый Дорожный проезд, 1) 6 ноября 2001 года с инвентарным номером ВКПМ В-8197.
Оба штамма, АJ12739 и AJ12739-ΔybiV, могут быть выращены при 37°С в течение 18 часов в питательном бульоне, содержащем хлорамфеникол (30 мкг/мл); 0.3 мл полученных культур может быть внесено в 3 мл ферментационной среды в пробирки размером 20×200 мм, и культуры могут быть выращены при 37°С в течение 48 часов на роторной качалке. По окончании ферментации количество накопленного в среде фенилаланина может быть определено с помощью тонкослойной хроматографии (TLC). Для этой цели могут быть использованы TLC-пластинки размером 10×15 см, покрытые 0.11 мм - слоем силикагеля Сорбфил без флуоресцентного индикатора (Акционерное Общество Сорбполимер, Краснодар, Россия). Пластинки Сорбфил могут быть экспонированы в подвижной фазе следующего состава: пропан-2-ол:этилацетат:25% водного аммиака:вода=40:40:7:16 (v/v). Раствор (2%) нингидрина в ацетоне может быть использован для визуализации.
Может быть использована ферментационная среда следующего состава (г/л):
Глюкозу и сульфат магния стерилизуют отдельно. СаСО3 стерилизуют сухим жаром при 180°С в течение 2 часов. рН доводят до 7.0.
Пример 10. Продукция L-триптофана штаммом Е. coli SV164 (pGH5)-ΔybiV
Для оценки влияния инактивации гена ybiV на продукцию L-триптофана ДНК-фрагменты хромосомы описанного выше штамма Е. coli MG1655 ΔybiV::cat могут быть перенесены в штамм-продуцент L-триптофана Е. coli SV164 (pGH5) с помощью Р1-трансдукции (Miller, J.H. (1972) Experiments in Molecular Genetics, Cold Spring Harbor Lab. Press, Plainview, NY). Штамм SV164 (pGH5) подробно описан в патенте США 6180373 или Европейском патенте 0662143.
Оба штамма, SV164(pGH5) и SV164(pGH5)-ΔybiV, могут быть выращены с перемешиванием при 37°С в течение 18 часов в 3 мл питательного бульона с добавлением тетрациклина (маркера плазмиды pGH5,20 мг/мл) и хлорамфеникола (30 мкг/мл). По 0.3 мл полученных культур может быть внесено в 3 мл ферментационной среды, содержащей тетрациклин (20 мг/мл), в пробирках размером 20×200 мм; и культуры могут быть выращены при 37°С в течение 48 часов на роторной качалке при 250 об/мин. После выращивания количество накопленного в среде триптофана может быть определено с помощью TLC, как описано в Примере 8. Компоненты ферментационной среды, которая может быть использована, представлены в Таблице 3; группы компонентов А, В, С, D, Е, F и Н стерилизуют отдельно, как и показано в Таблице 3, чтобы избежать нежелательных взаимодействий во время стерилизации.
Пример 11. Продукция L-пролина штаммом Е. coli 702ilvA-ΔybiV
Для оценки влияния инактивации гена ybiV на продукцию L-пролина ДНК-фрагменты из хромосомы описанного выше штамма Е. coli MG1655 ΔybiV::cat могут быть перенесены в штамм-продуцент L-пролина Е. coli 702ilvA с помощью Р1-трансдукции (Miller, J.H. (1972) Experiments in Molecular Genetics, Cold Spring Harbor Lab. Press, Plainview, NY). Штамм 702ilvA депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) (Россия, 117545 Москва, 1ый Дорожный проезд, 1) с инвентарным номером ВКПМ В-8012.
Оба штамма Е.Coli, 702ilvA и 702ilvA-ΔybiV, могут быть выращены в течение 18-24 часов при температуре 37°С на чашках с L-агаром, содержащим хлорамфеникол (30 мкг/мл). Затем ферментация с использованием этих штаммов может производиться в тех же условиях, как описано в Примере 8.
Пример 12. Продукция L-аргинина штаммом Е. coli 382-ΔybiV
Для оценки влияния инактивации гена ybiV продукцию L-аргинина ДНК-фрагменты хромосомы описанного выше штамма Е. coli MG1655 ΔybiV::cat были перенесены в штамм-продуцент L-аргинина Е. coli 382 с помощью P1-трансдукции (Miller, J.H. (1972) Experiments in Molecular Genetics, Cold Spring Harbor Lab. Press, Plainview, NY). Штамм 382 депонирован во Всероссийской коллекции промышленных микроорганизмов (ВКПМ) (Россия, 117545 Москва, 1ый Дорожный проезд, 1) 10 апреля 2000 года с инвентарным номером ВКПМ В-7926.
Оба штамма, 382 и 382-ΔybiV, были выращены с перемешиванием при 37°С в течение 18 часов в 3 мл питательного бульона, содержащего хлорамфеникол (30 мг/мл); по 0.3 мл полученных культур было внесено в 3 мл ферментационной среды в пробирки размером 20×200 мм, и культуры были выращены при 32°С в течение 48 часов на роторной качалке.
После выращивания количество накопленного в среде L-аргинина было определено с помощью бумажной хроматографии, при этом был использован следующий состав подвижной фазы: бутанол:уксусная кислота:вода=4:1:1 (v/v). Раствор нингидрина (2%) в ацетоне был использован для визуализации. Пятно, содержащее L-аргинин, было вырезано; L-аргинин был элюирован 0.5% водным раствором CdCl2, после чего количество L-аргинина было определено спектрофотометрическим методом при длине волны 540 нм. Результаты десяти независимых пробирочных ферментаций приведены в Таблице 2. Как следует из Таблицы 2, штамм 382-ΔybiV накапливал большее количество L-аргинина по сравнению со штаммом 382.
Была использована ферментационная среда следующего состава (г/л):
Глюкозу и сульфат магния стерилизовали раздельно. СаСО3 стерилизовали сухим жаром при 180°С в течение 2 часов. рН доводили до 7.0.
Пример 13. Удаление гена устойчивости Cm (гена cat) из хромосомы штаммов-продуцентов L-аминокислот бактерии Е. coli
Ген устойчивости Cm (ген cat) может быть удален из хромосомы штаммов-продуцентов L-аминокислот путем использования системы int-xis. С этой целью штамм-продуцент L-аминокислоты, в который были перенесены ДНК-фрагменты из вышеописанного штамма Е. coli MG1655 ΔybiV::cat при помощи P1-трансдукции (см. Примеры 3-12), может быть трансформирован плазмидой pMWts-Int/Xis. Селекция трансформированных клонов может быть произведена с использованием среды LB, содержащей ампициллин (100 мкг/мл). Клетки могут быть инкубированы при 30°С в течение ночи. Ген cat может быть удален из трансформированных клонов путем выращивания отдельных колоний при 37°С (при этой температуре репрессор CIts в некоторой мере инактивируется, тогда как гены int/xis активируются) с последующей селекцией вариантов CmSАрR Удаление гена cat из хромосомы штаммов может быть подтверждено при помощи ПЦР. Локус-специфичные праймеры Р21 (SEQ ID NO: 27) и Р22 (SEQ ID NO: 28) могут быть использованы для такого подтверждения путем ПЦР. Условия проведения ПЦР могут быть такими же, как описано выше. Продукт ПЦР, полученный в случае клеток с удаленным геном cat, должен быть длиной в 0,2 т.п.н. Таким образом, может быть получен штамм-продуцент L-аминокислот, в котором инактивирован ген ybiV и из которого удален ген cat.
Хотя указанное изобретение описано в деталях со ссылкой на наилучший способ осуществления изобретения, для специалиста в указанной области техники очевидно, что могут быть совершены различные изменения и произведены эквивалентные замены, и такие изменения и замены не выходят за рамки настоящего изобретения.
Каждому из упомянутых выше документов соответствует ссылка, и все цитируемые документы являются частью описания настоящего изобретения.










Изобретение относится к биотехнологии и представляет собой способ получения L-треонина или L-аргинина, включающий культивирование бактерии, принадлежащей к роду Escherichia, которая модифицирована таким образом, что ген ybiV в указанной бактерии инактивирован, и выделение L-треонина или L-аргинина из культуральной жидкости. Изобретение позволяет получать L-треонин или L-аргинин с высокой степенью эффективности. 2 н. и 1 з.п. ф-лы, 3 ил., 3 табл.
| ROBERTS A et | |||
| al | |||
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| Proteins | |||
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| US 4430430, 07.02.1984 | |||
| Штамм бактерий ЕSснеRIснIа coLI - продуцент L-треонина | 1987 |
|
SU1694643A1 |

