Изобретение относится к модифицированным фитазам.
Предпосылки изобретения
Фитат содержится в избытке в растениях как форма запасания фосфата. Животные, имеющие один желудок, не способны высвобождать фосфат из фитата и, таким образом, нуждаются в добавке фосфата к их пище. В настоящее время для высвобождения фосфата из фитата к корму животных прибавляют фермент фитазу. Обычно фитазу добавляют к корму животных во время процесса приготовления этого корма. На некоторых стадиях процесса приготовления корма фитаза подвергается воздействию относительно высокой температуры и относительно высокой влажности. Такие условия оказывают отрицательное влияние на активность лабильных соединений, подобных ферментам.
Фитаза, полученная из Aspergillus niger, обычно используется в кормах благодаря своим благоприятным свойствам. Например, она имеет широкий оптимум рН в кислотной области, широкую специфичность, связанную с относительно высокой специфической активностью и высоким химическим сродством с фитиновой кислотой, так, что даже при низких концентрациях фитиновой кислоты фермент эффективно разлагает фитиновую кислоту. Кроме того, она совершенно гладко удаляет пять из шести фосфатов фитата без существенного накопления промежуточных продуктов, она не требует наличия кофакторов активности или стабильности и не является очень чувствительной к ингибированию ингредиентами корма или ионами металла.
Однако термостабильность фитазы Aspergillus niger является относительно низкой. Таким образом, существует потребность в фитазе, которая имеет те же благоприятные свойства, что и фитаза Aspergillus niger, сочетающиеся с высокой стабильностью и активностью при высоких температурах.
Настоящее изобретение относится к модифицированным фитазам с благоприятными свойствами, например, в отношении устойчивости к высокой температуре и влажности.
Описание чертежей
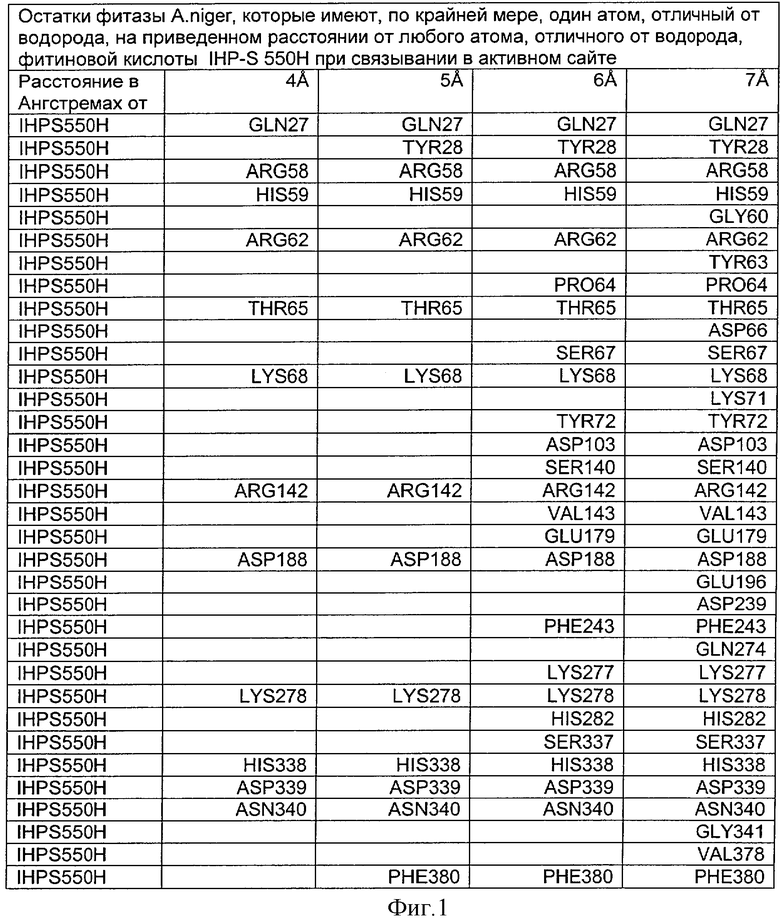
Фиг.1. Остатки сайта активности фитазы Aspergillus niger.
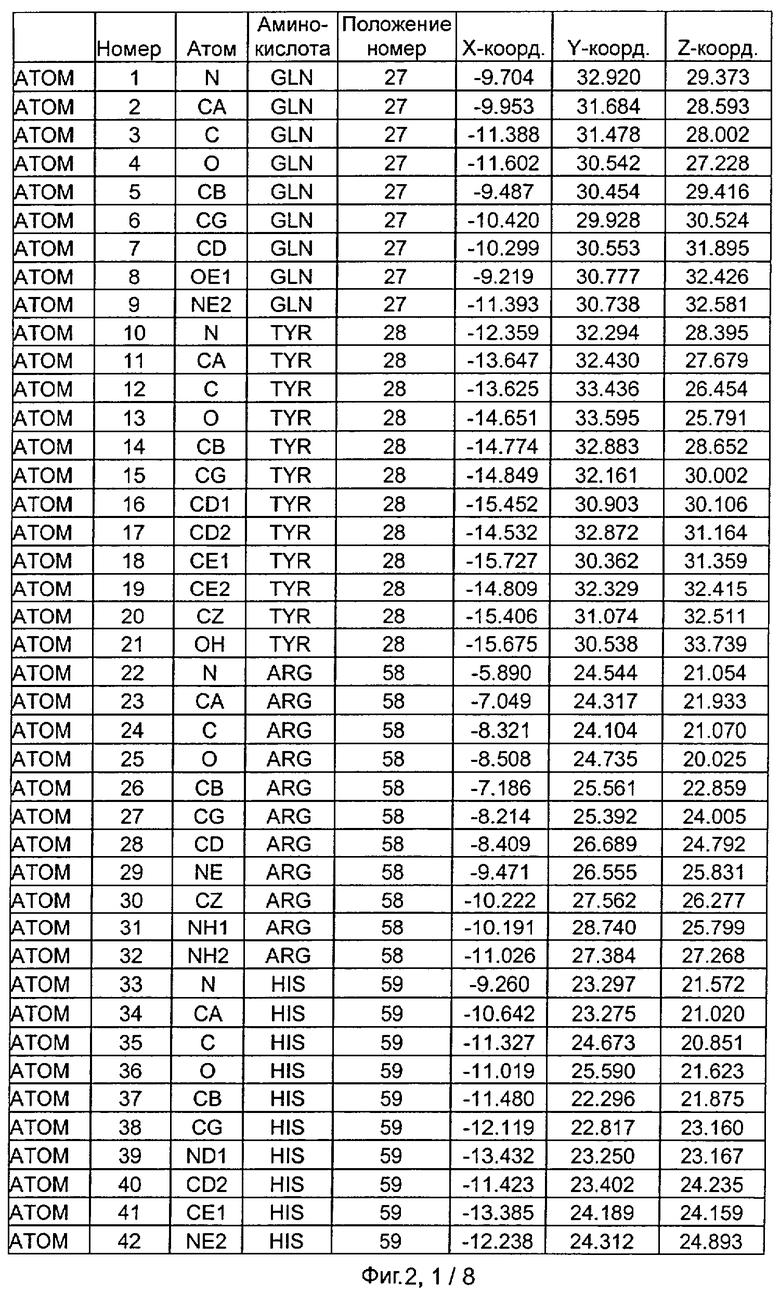
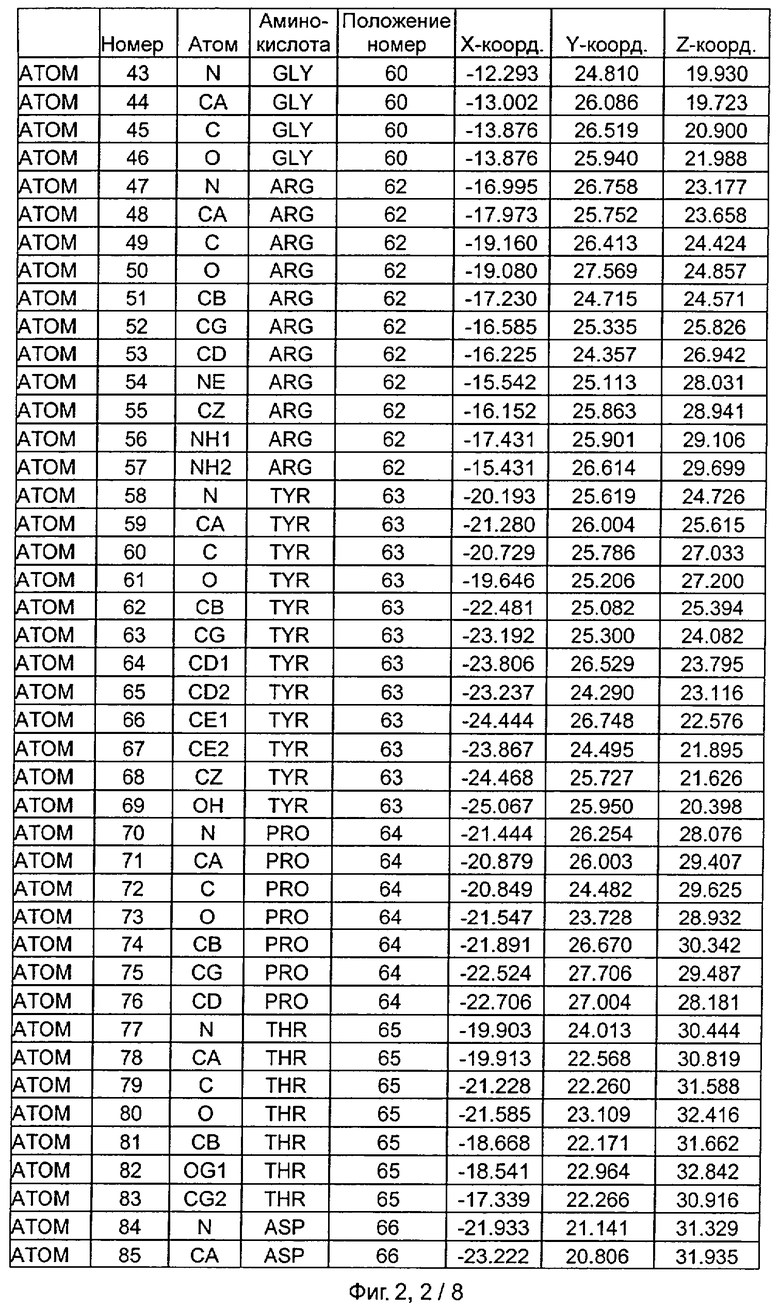
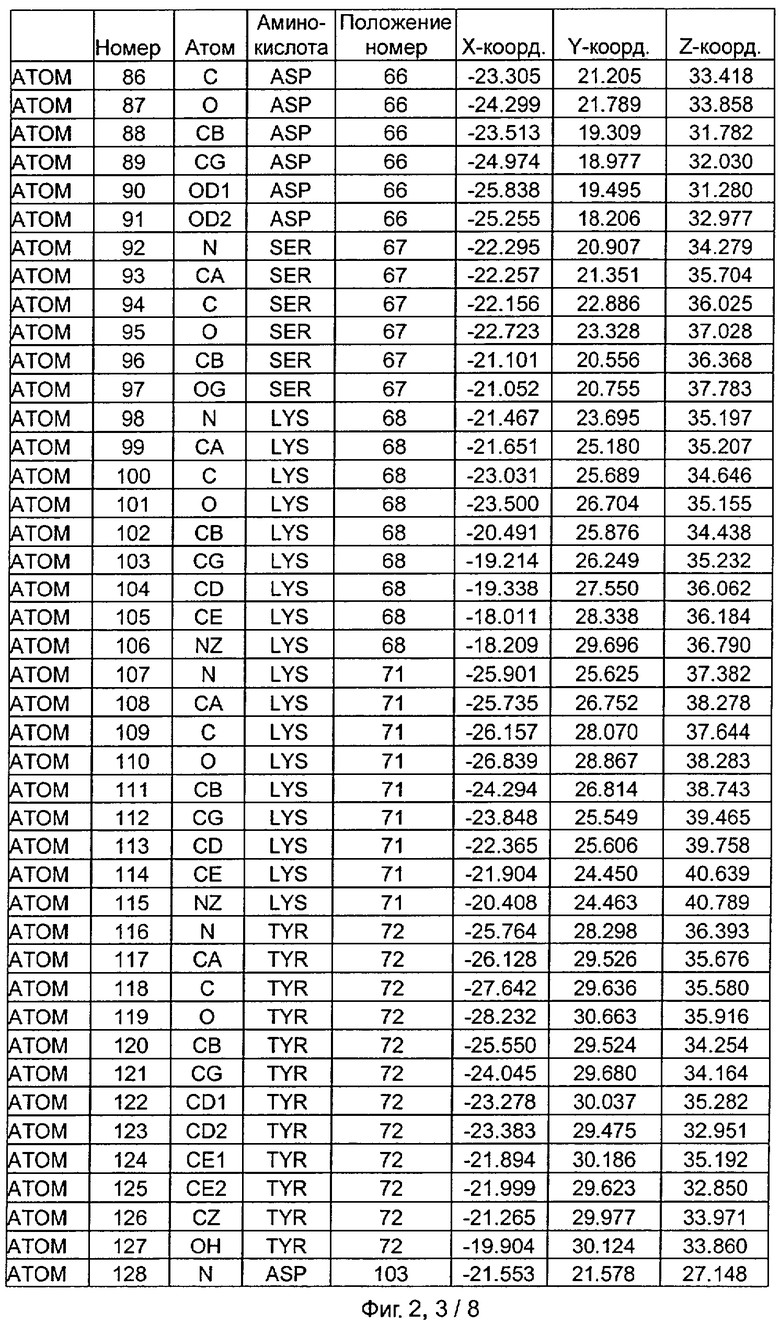
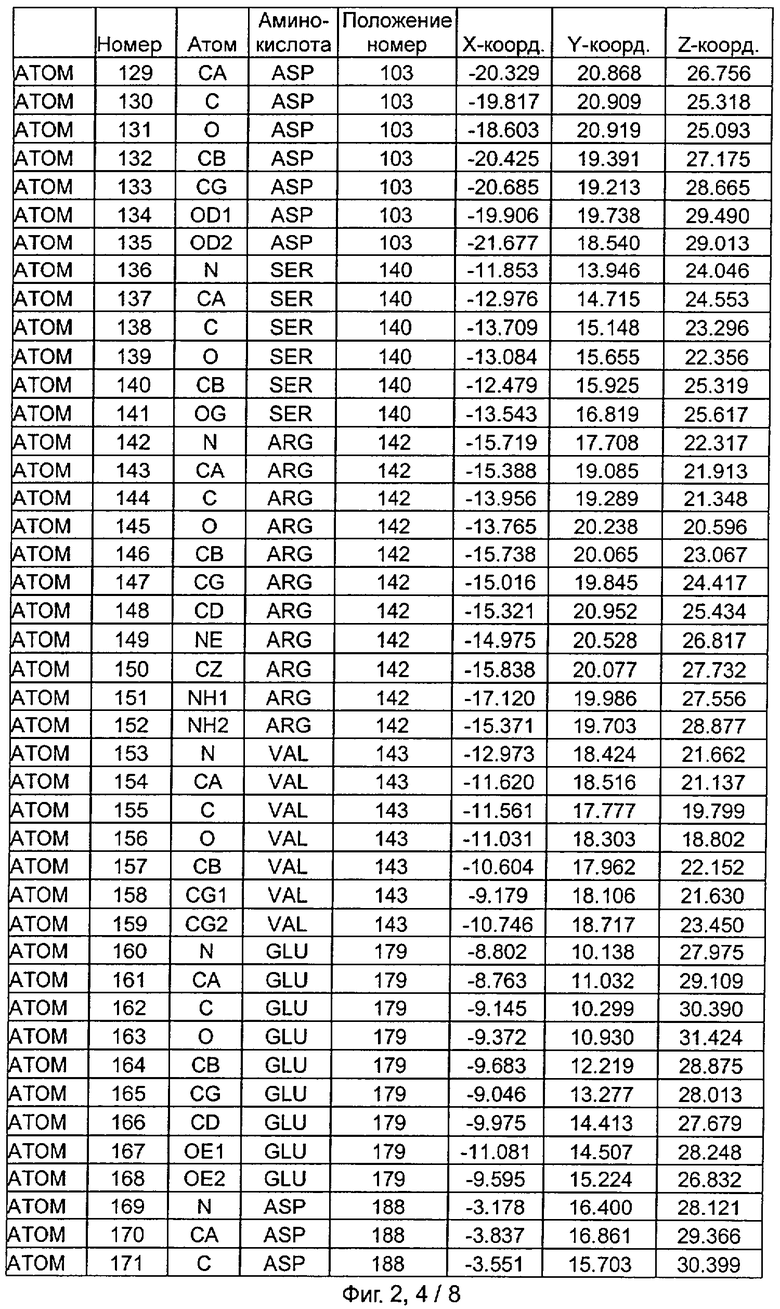
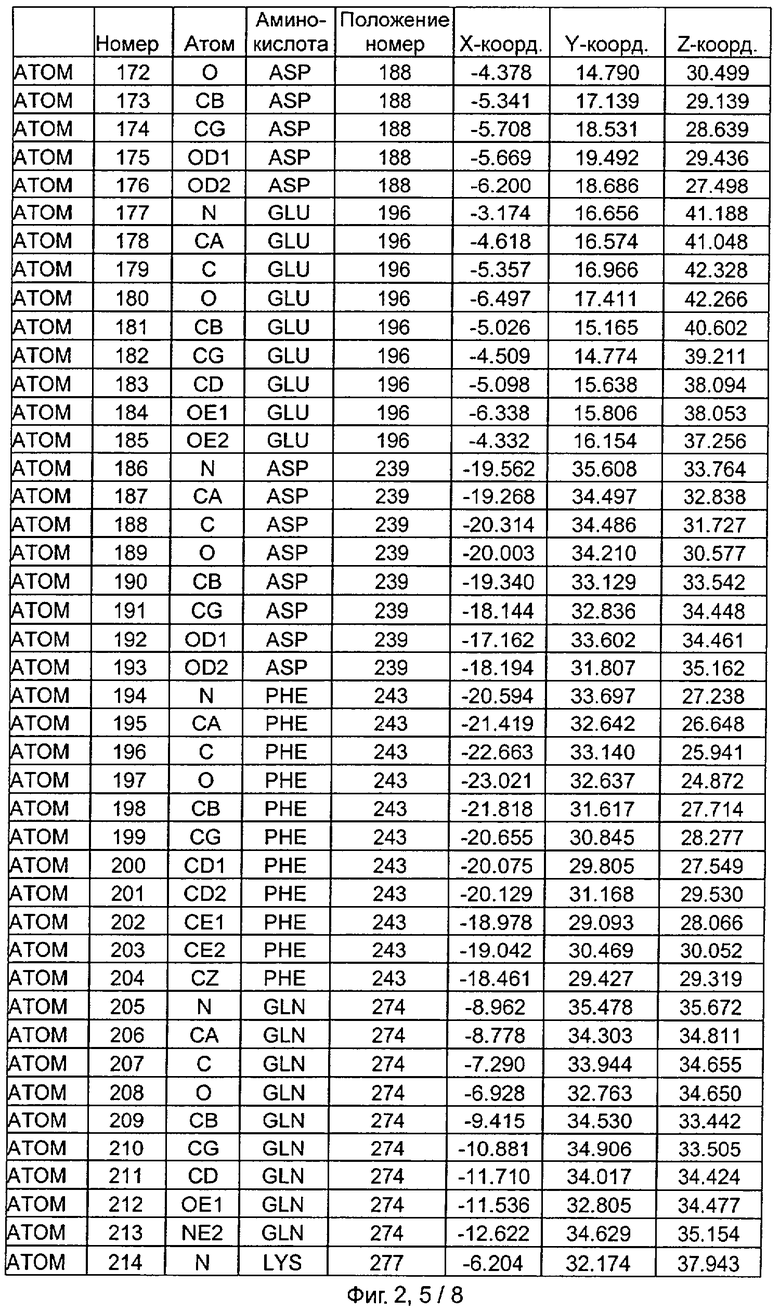
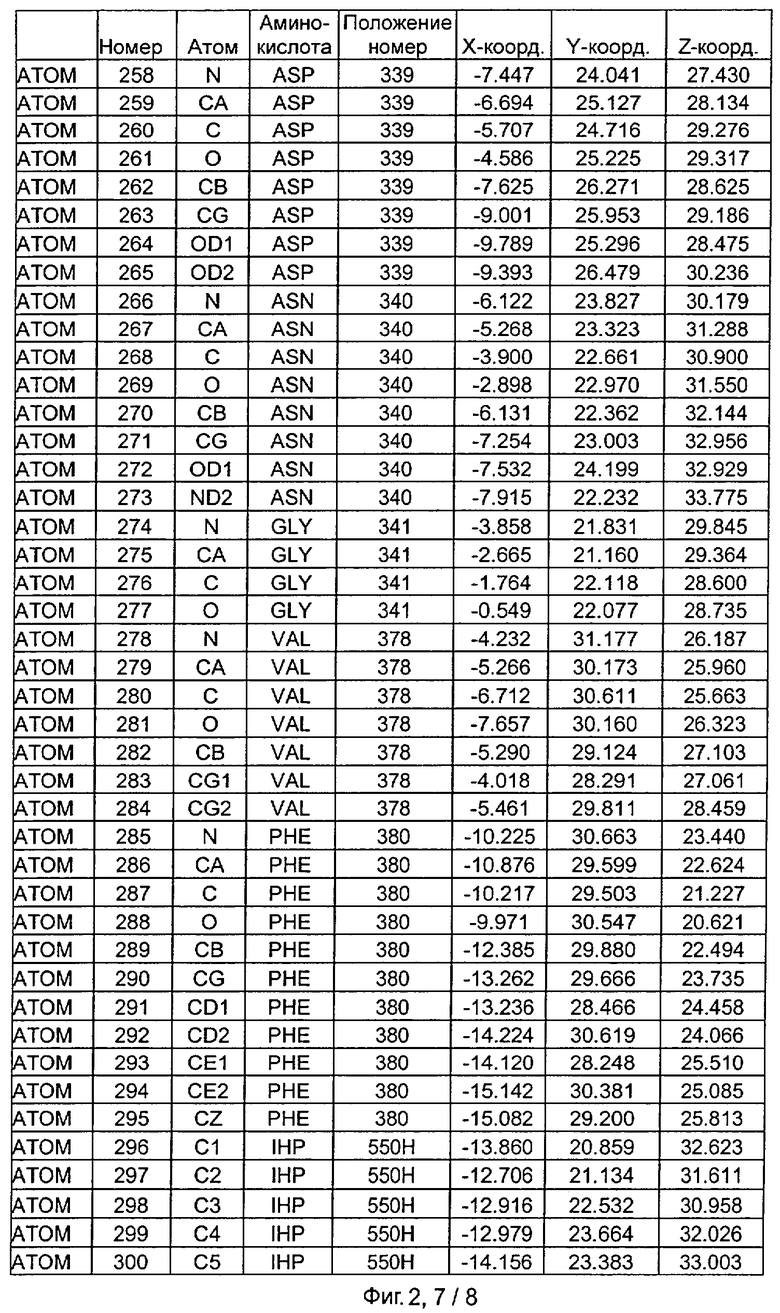
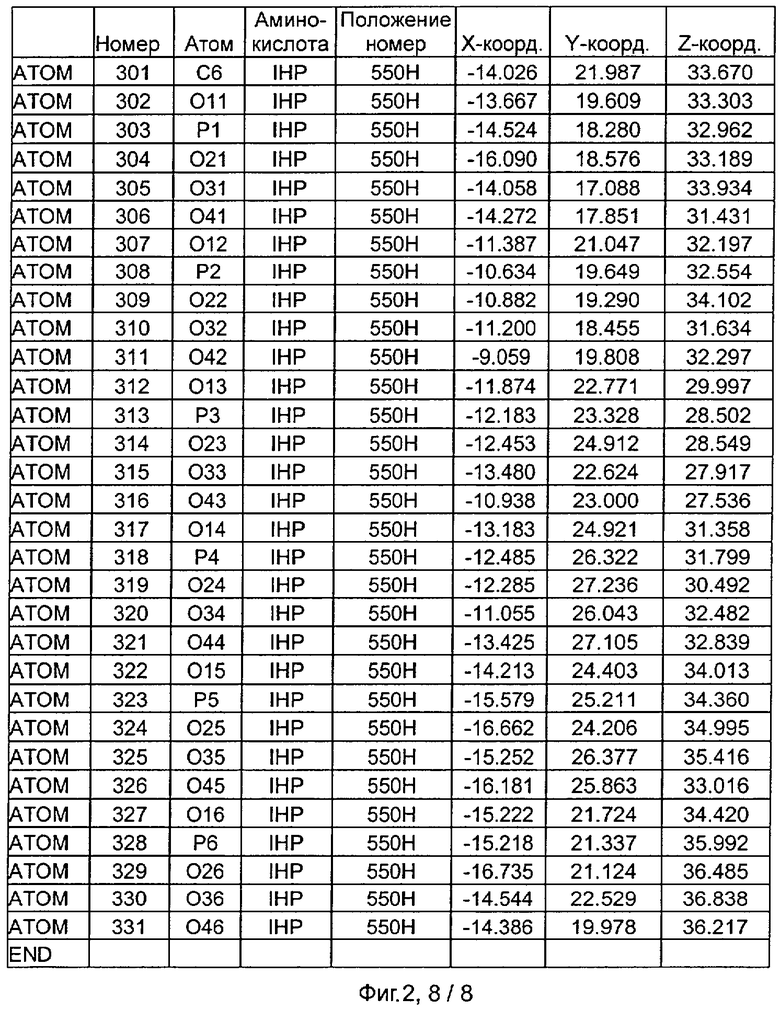
Фиг.2. IHP-S модель координат остатков активного сайта фитазы Aspergillus niger, связанных с фитиновой кислотой (IHP 550H).
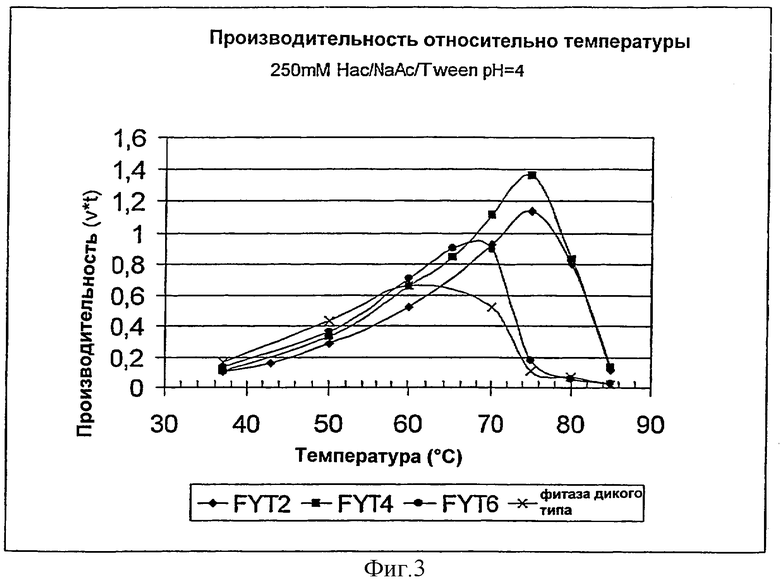
Фиг.3. Эффективность фитаз в зависимости от температуры.
Подробное описание изобретения
В смысле настоящего изобретения фитаза представляет собой фермент, который катализирует гидролиз фитата (гексакисфосфата миоинозитола) до одного или более следующих соединений: миоинозитол пента-, тетра-, три-, ди- и монофосфата и/или миоинозитола. В связи с этим общеизвестно, что некоторые фитазы неспособны к существенному гидролизу монофосфата миоинозитола до миоинозитола. Ферменты фитазы могут быть 3-фитазами или 6-фитазами (ЕС 3.1.3.8 или ЕС 3.1.3.26, соответственно ), что имеет отношение к положению первой сложноэфирной связи, которая подвергается гидролизу.
Первый объект настоящего изобретения касается полипептида, который представляет собой модифицированную фитазу. Полипептид в соответствии с изобретением является модифицированным по сравнению с модельной фитазой таким образом, что полипептид в соответствии с изобретением при сравнении с модельной фитазой содержит модификацию, выбранную из группы, состоящей из замены аминокислоты, которая присутствует в модельной фитазе, на различные другие аминокислоты, делецию аминокислоты, которая присутствует в модельной фитазе, или инсерцию аминокислоты. Совмещение полипептида в соответствии с изобретением с модельной фитазой осуществляют таким образом, чтобы получить максимальное количество гомологичных (идентичных) остатков между полипептидом в соответствии с изобретением и модельной фитазой.
В предпочтительном выполнении изобретения модификация представляет собой замену.
Количество модификаций может быть равно, по меньшей мере, одному, предпочтительно 10, более предпочтительно, по меньшей мере, 20, более предпочтительно, по меньшей мере, 30, более предпочтительно, по меньшей мере, 40, более предпочтительно, по меньшей мере, 50, более предпочтительно, по меньшей мере, 70, особенно предпочтительно, по меньшей мере, 80.
В настоящем изобретении обозначение, подобное, например, "5QS" означает, что аминокислота в положении 5 модельной фитазы в данном случае является замещенной либо Q, либо S. Природа исходного аминокислотного остатка может зависеть от используемой модельной фитазы. Обозначение, подобное "Q5S", означает, что специфический аминокислотный остаток, который присутствует в модельной фитазе, например Q, замещен другой аминокислотой, например S.
Модифицированная фитаза является модифицированной по сравнению с модельной фитазой предпочтительно, по меньшей мере, в одном из следующих положений: 5, 6, 13, 19, 21, 29, 31, 36, 39, 43, 53, 69, 78, 81, 85, 87, 99, 112, 113, 122, 125, 126, 128, 137, 147, 148, 157, 160, 163, 165, 169, 172, 176, 178, 180, 181, 182, 183, 189, 194, 197, 201, 203, 211, 213, 215, 218, 222, 223, 225, 232, 233, 242, 246, 247, 248, 249, 250, 251, 252, 254, 269, 291, 296, 310, 312, 315, 322, 330, 342, 346, 362, 365, 367, 368, 372, 374, 375, 382, 384, 395, 414, 417, 425, 428, 438, 440; или предпочтительно, по меньшей мере, в одном из следующих положений: 13, 19, 21, 29, 31, 36, 39, 43, 53, 69, 78, 81, 85, 87, 99, 112, 113, 122, 125, 126, 128, 137, 147, 148, 157, 160, 163, 165, 169, 172, 176, 178, 180, 181, 182, 183, 189, 194, 197, 201, 203, 211, 213, 215, 218, 222, 223, 225, 232, 233, 242, 246, 247, 248, 249, 250, 251, 252, 254, 269, 291, 296, 310, 312, 315, 322, 330, 342, 346, 362, 365, 367, 368, 372, 374, 375, 382, 384, 395, 414, 417, 425, 428, 438, 440; или предпочтительно, по меньшей мере, в одном из следующих положений: 13, 19, 21, 29, 36, 39, 43, 53, 69, 81, 85, 87, 99, 112, 113, 122, 125, 126, 128, 137, 147, 148, 157, 160, 165, 169, 172, 176, 178, 181, 183, 189, 197, 201, 203, 213, 218, 222, 223, 225, 232, 233, 246, 247, 248, 249, 250, 251, 252, 291, 296, 310, 312, 315, 322, 330, 342, 346, 362, 365, 367, 368, 372, 374, 375, 382, 384, 395, 417, 425, 438, 440. Еще более предпочтительно, когда модифицированная фитаза модифицирована по сравнению с модельной фитазой, по меньшей мере, в одном из следующих положений: 31, 78, 163, 180, 182, 194, 211, 215, 242, 254, 269, 414, 428, 440.
В частности, модифицированная фитаза содержит, по меньшей мере, одну из следующих модификаций по сравнению с модельной фитазой: 5QS, 6SH, 13G, 19Р, 21I, 29S, 31FY, 36D, 39А, 43D, 53V, 69S, 78EA, 81К, 85А, 87К, 99Т, 112Q, 113М, 122R, 125К, 126А, 128А, 137А, 147А, 148Е, 157А, 160А, 163RG, 165N, 169А, 172V, 1761, 178Р, 180AG, 181А, 182STG, 183Y, 189Н, 194VA, 197Е, 201G, 203D, 211TL, 213А, 215SA, 218А, 222А, 223Н, 225Р, 232Е, 233D, 242SP, 246V, 247A, 248R, 249Т, 250S, 251D, 252A, 254КЕ, 269NQ, 291A, 296F, 310Q, 312Н, 315Т, 322N, 330А, 342М, 346F, 362S, 365S, 367Е, 368Е, 372Y, 374А, 375S, 382А, 384А, 395К, 414РА, 417К, 425D, 428RKE, 438N, 440АЕ; или более предпочтительно, по меньшей мере, одну из следующих модификаций по сравнению с модельной фитазой: 5QS, 6SH, 13G, 19Р, 21I, 29S, 31Y, 36D, 39А, 43D, 53V, 69S, 78А, 81К, 85А, 87К, 99Т, 112Q, 113М, 122R, 125К, 126А, 128А, 137А, 147А, 148Е, 157А, 160А, 163G, 165N, 169А, 172V, 1761, 178Р, 180G, 181А, 182G, 183Y, 189Н, 194А, 197Е, 201G, 203D, 211L, 213А, 215А, 218А, 222А, 223Н, 225Р, 232Е, 233D, 242Р, 246V, 247A, 248R, 249Т, 250S, 251D, 252A, 254Е, 269Q, 291A, 296F, 310Q, 312Н, 315Т, 322N, 330А, 342М, 346F, 362S, 365S, 367Е, 368Е, 372Y, 374А, 375S, 382А, 384А, 395К, 414А, 417К, 425D, 428Е, 438N, 440Е.
Наиболее предпочтительно, когда модифицированная фитаза содержит, по меньшей мере, одну из следующих модификаций по сравнению с модельной фитазой: 31Y, 78A, 163G, 180G, 182G, 194А, 211L, 215А, 242Р, 254Е, 269Q, 414А, 428Е, 440Е.
Нумерация положений, которая используется в данном изобретении, приведена в соответствии с нумерацией положений последовательности SEQ ID NO:1.
Модельная фитаза, которая используется в настоящем изобретении, представляет собой фитазу, полученную из нитевидных грибов рода Aspergillus, предпочтительно из видов Aspergillus niger, или вариантную фитазу, которая получена из любой из этих фитаз. Таким образом, известно, что фитазы в пределах индивидуальных штаммов видов Aspergillus niger демонстрируют высокую степень вариации, то есть гомология этих фитаз составляет, по меньшей мере, 90%. Также известно, что виды Aspergillus niger включают виды, которые ранее были известны как Aspergillus ficuum и Aspergillus awamori. Более предпочтительно, когда модельная фитаза представляет собой фитазу, полученную из Aspergillus niger NRRL 3135, как представлено в SEQ ID NO:1.
Особенно предпочтительной модельной фитазой является фитаза, содержащая комбинацию специфических аминокислотных остатков, которые однозначно присутствуют в фитазе Aspergillus niger.
Особенно предпочтительная модельная фитаза содержит те же аминокислотные остатки в активных сайтах, что и аминокислоты, которые присутствуют в фитазе Aspergillus niger в соответствующих положениях. С этой целью изобретение раскрывает способ определения таких остатков в активном сайте фитазы Aspergillus niger, которые находятся на определенном расстоянии от границы фитата.
Аминокислотные остатки, которые образуют активный сайт фитазы Aspergillus niger и которые имеют отношение к каталитическим свойствам деградации фитиновой кислоты с помощью фитазы Aspergillus niger, были идентифицированы при использовании структуры 3D фитазы Aspergillus niger, которая является доступной из базы данных белков (PDB) под номером 1IНР (Kostrewa и др. Nature Structural Biology, 1997, 4, 185). 3D структура Aspergillus niger не содержит субстрата фитиновой кислоты (гексакисфосфат миоинозитола). Однако является доступной 3D структура фитазы Е.coli, связанная с фитиновой кислотой (номер доступа 1DKQ в PDB, Lim и др., Nature Structural Biology, 2000, 7, 108). Несмотря на то, что гомологичность последовательности имеет низкую степень гомологии, обе фитазы продемонстрировали существенное структурное подобие. Наложение атомных координат для фитазы Aspergillus niger (1IHP) и фитазы Е.coli начинали только при использовании альфа-углеродных атомов тех остатков, которые демонстрируют подобную модель сборки в обеих фитазах. Впоследствии увеличивали число включаемых остатков в итеративном процессе до тех пор, пока нельзя было получить дальнейшее улучшение наложения. Качество наложения оценивали при использовании среднеквадратического отклонения атомов, используемых для наложения. При заключительном наложении аминокислотные сегменты 1DKQ: 6-22, 46-66, 83-106, 246-257, 268-278, 296-313, 328-338, 346-351, 375-381, 392-398 были наложены на 1IНР: 48-64, 104-124, 133-156, 270-281, 293-303, 331-348, 379-389, 397-402, 406-412, 422-428.
После наложения субстрат фитиновой кислоты был взят из активного сайта фитазы Е.coli и перенесен в соответствующий сайт фитазы Aspergillus niger. Впоследствии фитазу Aspergillus niger, связанную с фитиновой кислотой, подвергали энергетической минимизации, которая позволяла остаткам субстрата и активного сайта перемещаться при сохранении остатков фиксированной структуры. Энергетическую минимизацию осуществляли в соответствии с программой Insight & Discover (Accelrys, San Diego CA) с усиленным силовым полем CVFF при использовании рабочей станции SGI Octane. Полученную модель для фитазы Aspergillus niger, комплексированную с фитиновой кислотой, обозначали как IHP-S. Было установлено, что расчет растворимой доступной поверхности для тех аминокислот, которые имели, по меньшей мере, один атом на расстоянии 7 Ангстрем от любого атома субстрата фитиновой кислоты, приводит к гладкой непрерывной поверхности, очерчивающей карман, который приспосабливается к фитиновой кислоте почти полностью. Атомные координаты остатков, которые способствуют такому карману активного сайта, приведены на Фиг.1. В дополнение к этому модель IHP-S использовали для идентификации различных зон остатков, находящихся вокруг субстрата. Результаты приведены на Фиг.2.
Таким образом, в контексте данного изобретения аминокислотные остатки активного сайта фитазы Aspergillus niger являются такими аминокислотными остатками, которые находятся на определенном расстоянии от фитиновой кислоты, когда связываются в активном сайте. Предпочтительно, когда указанное расстояние составляет 6 Ангстрем, более предпочтительно 7 Ангстрем.
Таким образом, особенно предпочтительная модель фитазы содержит аминокислоты Q27, Y28, R58, Н59, R62, Р64, Т65, S67, К68, Y72, D103, S140, R142, V143, Е179, D188, F243, KN277, К278, Н282, S337, Н338, D339, N340, F380 (в пределах расстояния 6 Ангстрем), предпочтительно аминокислоты Q27, Y28, R58, Н59, G60, R62, Y63, Р64, Т65, DE66, S67, К68, К71, Y72, D103, S140, R142, V143, Е179, D188, Е196, D239, F243, Q274, KN277, К278, Н282, S337, Н338, D339, N340, G341, V378, F380 (в пределах расстояния 7 Ангстрем).
Особенно предпочтительная модель фитазы дополнительно содержит следующие аминокислоты, которые присутствуют в фитазе Aspergillus niger: A35, А46, N130, S141, G167, Q168, D174, Т191, Е199, Е205, L220, Т235, D244, I268, Н306, G341, К356, А381.
Для неспецифического положения в рамках настоящего изобретения не является существенным, какой аминокислотный остаток может присутствовать. Такое неспецифическое положение представляет собой положение, которое находится вне активного сайта фитазы Aspergillus niger и не является такой аминокислотой, как указано выше, а также не подвергается специфическим модификациям, как указано выше. Совмещение фитаз при использовании общеизвестной программы смещения будет выявлять аминокислоту(ы), которая(ые) типично существуют в определенном положении. В соответствующей неспецифическом положении в полипептиде в соответствии с изобретением может присутствовать любая из таких аминокислот.
Таким образом, предпочтительный полипептид в соответствии с изобретением представляет собой фитазу, которая содержит такие же аминокислотные остатки в активном сайте, что и аминокислоты, присутствующие в фитазе Aspergillus niger в соответствующих положениях, также, как и дополнительно указанные аминокислоты Aspergillus niger (то есть A35, А46, N130, S141, G167, Q168, D174, Т191, Е199, Е205, L220, Т235, D244, I268, Н306, G341, К356, А381), а также содержит модификацию, как указано выше.
Особенно предпочтительный полипептид в соответствии с изобретением представляет собой фитазу, которая дополнительно содержит, по меньшей мере, один из следующих аминокислотных остатков: 31Y, 78А, 163G, 180G, 182G, 194А, 211L, 215А, 242Р, 254Е, 269Q, 414А, 428Е и/или 440Е. Другой особенно предпочтительный полипептид в соответствии с изобретением содержит, по меньшей мере, один из следующих аминокислотных остатков: 180G, 182G, 242Р и/или 440Е; или предпочтительно, по меньшей мере, 180G, 182G и/или 242Р.
В частности, настоящее изобретение раскрывает полипептид, который представляет собой модифицированную фитазу в соответствии с последовательностью SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7.
Полипептид в соответствии с изобретением может содержать все модификации, представленные выше. Кроме того, полипептид в соответствии с изобретением может включать дополнительные модификации, относящиеся к тем положениям в полипептиде, модификация которых не затрагивает сворачивание или активность полипептида. Обычно такие модификации могут быть консервативными заменами, то есть заменами, в которых неполярные, полярные незаряженные, полярные заряженные или ароматические аминокислоты замещаются различными аминокислотами подобной категории.
В одном выполнении полипептид в соответствии с изобретением может включать полипептид, имеющий, по меньшей мере, 91, предпочтительно, по меньшей мере, 92, более предпочтительно, по меньшей мере, 93, более предпочтительно, по меньшей мере, 94, более предпочтительно, по меньшей мере, 95, более предпочтительно, по меньшей мере, 96, более предпочтительно, по меньшей мере, 97, более предпочтительно, по меньшей, 98 или наиболее предпочтительно, по меньшей мере, 99% гомологичности (идентичности) последовательности с SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7.
Полипептид в соответствии с изобретением модифицирован для увеличения термостабильности, и/или для изменения специфической активности, и/или изменения специфичности к определенному субстрату и/или модификации оптимального значения рН фермента, и/или улучшения стабильности при гранулировании, и/или для улучшения биоэффективности, и/или усовершенствования экспрессии, транспорта, созревания и тому подобное, в организме хозяина, который используется для получения модифицированной фитазы, при сравнении ее с модельной фитазой.
В предпочтительном выполнении полипептид в соответствии с изобретением сохраняет некоторые биохимические свойства фитазы Aspergillus niger, в частности фитазы, полученной из Aspergillus niger NRRL 3135. Биохимическое свойство, которое сохраняется, представляет собой значение Km и/или оптимум рН при двух значениях рН приблизительно 5,5 и 2,5, и/или специфическую активность, и/или высокую активность при физиологической температуре.
В предпочтительном выполнении полипептид, полученный в соответствии с изобретением, обладает повышенной термостабильностью. Повышенная термостабильность модифицированной фитазы в соответствии с изобретением по сравнению с модельной фитазой может быть выражена более продолжительным временем жизни при данной повышенной температуре, и/или улучшенными характеристиками повторной сборки/восстановления активности, и/или отсутствием сборки при более высокой температуре.
Неожиданно было обнаружено, что полипептид в соответствии с изобретением сочетает в себе несколько благоприятных свойств фитазы Aspergillus niger c повышенной термостабильностью.
Аминокислоты, которые являются важными, например, для термостабильности или активности полипептида в соответствии с изобретением, и, таким образом, потенциально подвергаются замене, могут быть идентифицированы и модифицированы в соответствии со способами, известными в области техники, такими, как сайт-направленный мутагенез или аланин-сканирующий мутагенез. В более позднем способе мутации вводили в каждый остаток молекулы. Полученные мутантные молекулы исследовали на биологическую активность (например, фитазную активность) для идентификации аминокислотных остатков, которые являются критическими для активности молекулы. Сайты фермент-субстратного взаимодействия также могут быть определены с помощью анализа кристаллической структуры, как определяется с помощью таких методик, как ядерный магнитный резонанс, кристаллография, и фото-аффинное мечение или молекулярное моделирование.
Полипептиды в соответствии с изобретением могут быть получены при использовании способов синтеза, несмотря на то, что обычно их получают рекомбинантным способом путем экспрессии полинуклеотидной последовательности, кодирующей полипептид в приемлемом организме хозяина.
Как ожидается, использование дрожжевых клеток и клеток грибов будет обеспечивать такие пост-трансляционные модификации (например, протеолитический процессинг, миристилирование, гликозилирование, укорачивание, тирозин-, серин- или треонин-фосфорилирование), которые могут быть необходимы для придания оптимальной биологической активности рекомбинантным продуктам экспрессии в соответствии с изобретением.
Полипептиды в соответствии с изобретением могут иметься в такой форме, в которой они находятся за пределами своей природной клеточной окружающей среды. Таким образом, они могут быть существенно изолированы или очищены, как обсуждалось выше, или получены в клетке, в которой они не образуются в природе, например в клетке других видов грибов, животных, дрожжей или бактерий.
Полипептиды в соответствии с изобретением могут подвергаться анализу с помощью любого приемлемого способа, известного специалисту, для оценки усовершенствования по сравнению с модельной фитазой, которая известна в уровне техники.
Во втором аспекте изобретения предлагается (например, изолированный и/или очищенный) полинуклеотид, содержащий полинуклеотидную последовательность, кодирующую полипептид в соответствии с первым аспектом. В частности, настоящее изобретение предлагает полинуклеотид, содержащий полинуклеотидную последовательность, кодирующую аминокислотную последовательность, представленную в SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7, или полинуклеотид, включающий SEQ ID NO:2, SEQ ID NO:4 или SEQ ID NO:6. Полинуклеотиды в соответствии со вторым аспектом дополнительно включают любой вырожденный вариант полинуклеотидной последовательности, кодирующий полипептид в соответствии с первым аспектом. Например, специалист в данной области может при использовании обычных способов осуществить нуклеотидные замены, которые не влияют на белковую последовательность, кодируемую полинуклеотидами в соответствии с изобретением, что отражает предпочтительность кодонов для частного организма-хозяина, в котором экспрессируются полипептиды в соответствии с изобретением.
Полинуклеотидная последовательность в соответствии со вторым аспектом может представлять собой ДНК или РНК и включает геномную ДНК, синтетическую ДНК или кДНК. Предпочтительно, когда полинуклеотид представляет собой ДНК последовательность.
Полинуклеотиды в соответствии с изобретением могут быть синтезированы в соответствии со способами, хорошо известными в уровне техники. Они могут быть получены путем комбинирования олигонуклеотидов, синтезированных в соответствии с нуклеотидной последовательностью полинуклеотида настоящего изобретения. Альтернативно, они могут быть синтезированы путем мутагенеза исходного полинуклеотида в любой желательной точке.
Например, полинуклеотид в соответствии с изобретением конструируется из ряда синтетических олигонукпеотидов длиной 80 нуклеотидов, которые имеют перекрывание размером 20 нуклеотидов. ПЦР обычно проводят в 10 этапов при использовании полимеразы с корректирующей активностью на всех 80-мерных олигонуклеотидах для отжига и удлинения олигонуклеотидов. Дополнительную ПЦР с корректирующей полимеразой проводят с праймерами ПЦР, которые расположены на 5'- и 3'-концах желаемого фрагмента, для синтеза полного желательного фрагмента. Завершенный фрагмент клонируют в приемлемом векторе и секвенируют для подтверждения того факта, является правильной или нет полученная последовательность. Необязательно, ошибки в последовательности корректируют, например, при использовании набора QuickChange от Stratagene в соответствии с инструкциями производителя.
Полинуклеотиды в соответствии с изобретением могут использоваться для получения полинуклеотидов, кодирующих дополнительно модифицированный полипептид, например, путем воздействия на полинуклеотиды в соответствии с изобретением способами мутагенеза. Сайт-направленный метагенез может использоваться для изменения полинуклеотидов в соответствии с изобретением в одном или более положениях. Технология перестановки генов (например, как раскрыто в WO 95/22625, WO 98/27230, WO 98/01581 и/или WO 00/46344) может быть использована для получения вариантов полинуклеотидов со случайной комбинацией любого вариантного положения, которое имеется в любом из членов исходной популяции полинуклеотидов, при этом исходная популяция включает один или более полинуклеотидов в соответствии с изобретением.
Изобретение обеспечивает векторы, содержащие полинуклеотид в соответствии с изобретением, включая клонирующие и экспрессионные векторы.
Вектор, в который встраивают экспрессионную кассету или полинуклеотид в соответствии с изобретением, может быть любым вектором, который может традиционно подвергаться методам рекомбинантной ДНК, и выбор вектора будет часто зависеть от клетки-хозяина, в которую его встраивают. Таким образом, вектор может быть автономно реплицирующимся вектором, то есть вектором, который существует как экстра-хромосомная единица, репликация которой является независимой от хромосомной репликации, например, плазмидой, космидой, вирусным или фаговым вектором, который обычно обеспечивается вместе с источником репликации. Альтернативно вектор может быть таким, который при введении в клетку-хозяина интегрируется с геномом клетки-хозяина и реплицируется вместе с хромосомой(ами), в которую(ые) он интегрирован. Вектор может быть кольцевым, например, плазмидой, или линейным, например, экспрессионной кассетой, полинуклеотидом.
Предпочтительно, когда полинуклеотид в соответствии с изобретением может быть встроен в экспрессионную кассету. В экспрессионной кассете полинуклеотид в соответствии с изобретением оперативно связывают с регуляторной последовательностью, которая способна обеспечить экспрессию полипептида исходя из его кодирующей последовательности, клеткой-хозяином, то есть вектор представляет собой экспрессионный вектор. Термин «оперативно связанный» относится к соседнему участку, в котором описанные компонентны находятся во взаимной связи, позволяющей им функционировать запланированным образом. Регуляторная последовательность, такая как промотор, энхансер или другой экспрессионный регуляторный сигнал, «оперативно связанный» с кодирующей последовательностью, размещается таким образом, что экспрессия полипептида исходя из его кодирующей последовательности, достигается в условиях, совместимых с регуляторными последовательностями.
Экспрессионная кассета для данной клетки-хозяина может включать следующие элементы, оперативно связанные друг с другом в последовательном порядке от 5'-конца до 3'-конца относительно кодирующей цепи последовательности, кодирующей полипептид в соответствии с первым аспектом: промоторную последовательность, способную направлять транскрипцию последовательности ДНК, кодирующей полипептид, в данной клетке-хозяине; необязательно сигнальную последовательность, способную направлять секрецию полипептида из данной клетки-хозяина в культуральную среду; последовательность ДНК, кодирующую зрелую и предпочтительно активную форму полипептида; и предпочтительно также участок терминации транскрипции (терминатор), способный терминировать транскрипцию ниже последовательности ДНК, кодирующей полипептид.
Отдельно от природного промотора гена, кодирующего существующий в природе предшественник полипептида в соответствии с изобретением, могут использоваться другие промоторы для непосредственной экспрессии полипептида в соответствии с изобретением. Промотор может быть выбран в соответствии с его эффективностью относительно направления экспрессии полипептида в соответствии с изобретением в желательном экспрессионном хозяине.
Промоторы/энхансеры и другие экспрессионные регуляторные сигналы могут быть выбраны таким образом, чтобы быть совместимыми с клеткой-хозяином, для которой предназначена экспрессионная кассета или вектор. Предпочтительно, когда промоторная последовательность имеет происхождение от высоко экспрессируемого гена. В контексте настоящего изобретения высоко экспрессируемый ген представляет собой ген, мРНК которого может составлять, по меньшей мере, 0,01% (масс./масс.) от общей мРНК клетки, например, в индуцированных условиях, или альтернативно, ген, продукт которого может составлять, по меньшей мере, 0,2% (масс./масс.) от общего количества белка клетки, или, в случае секретируемого генного продукта, может быть секретирован до уровня, по меньшей мере, 0,05 г/л. Примеры предпочтительных высоко экспрессируемых генов, от которых предпочтительно происходят промоторы, и/или которые включены в предпочтительный предварительно определенный целевой локус для интеграции с экспрессионными кассетами, включают, но не ограничены, гены, кодирующие гликолитические ферменты, такие как триозофосфатизомеразы (TPI), глицеральдегид дегидрогеназы (GAPDH), фосфоглицераткиназы (PGK), пируваткиназы (PYK), алкогольдегидрогеназы (ADH), а также гены, кодирующие амилазы, глюкоамилазы, протеазы, глюканазы, целлобиогидролазы, β-галактозидазы, алкоголь(метанол)оксидазы, факторы элонгации и рибосомальные белки. Специфические примеры приемлемых высоко экспрессируемых генов включают, например, ген LAC4 из Kluyveromyces sp., гены метанолоксидазы (AOX и МОХ) из Hansenula и Pichia, соответственно, гены глюкоамилазы (glaA) из A.niger и A.awamori, ген ТАКА-амилазы A.oryzae, ген gpdA A.nidulans и гены целлобиогидролазы T.reesei. Для эффективной экспрессии в бактериях могут использоваться прокариотические промоторы, в частности, те, которые приемлемы для использования в штаммах E.coli. Примеры сильных бактериальных промоторов представляют собой α-амилазы и промоторы SPo2, а также промоторы из генов экстрацеллюлярной протеазы.
Дрожжевые промоторы включают промоторы GAL4 и ADH S. cerevisiae, промотор nmt 1 S.pombe и промотор adh. Примеры сильных дрожжевых промоторов представляют собой те, которые получены из генов алкогольдегидрогеназы, лактазы, 3-фосфоглицерат киназы и триозофосфатизомеразы.
Примеры сильных конститутивных и/или индуцибельных промоторов, которые являются предпочтительными для использования в грибковых экспрессионных хозяевах, представляют собой те, которые получены из грибковых генов ксиланазы (xlnA), фитазы, АТФ-синтетазы, субъединицы 9 (oliC), триозофосфатизомеразы (tpi), алкогольдегидрогеназы (AdhA), α-амилазы (amy), амилогликозидазы (AG - из гена glaA), ацетамидазы (amdS) и промоторы глицеральдегид-3-фосфат дегидрогеназы (gpd).
Промоторы, приемлемые для растительных клеток, включают промоторы нопалинсинтазы (nos), октопинсинтазы (ocs), маннопинсинтазы (mas), малой субъединицы рибулозы (rubisco ssu), гистона, актина риса, фазеолина, 35S и 19S субъединиц вируса мозаики цветной капусты (CMV), а также промоторы цирковируса. Все эти промоторы являются легко доступными из уровня техники. Если полипептид получают как секретируемый белок, то полинуклеотидная последовательность, кодирующая зрелую форму полипептида в экспрессионной кассете, является оперативно связанной с полинуклеотидной последовательностью, кодирующей сигнальный пептид.
Предпочтительно, когда сигнальная последовательность является природной (гомологичной) полинуклеотидной последовательности, кодирующей полипептид. В предпочтительном воплощении изобретения сигнальная последовательность получена из гена фитазы Aspergillus niger, в частности сигнальная последовательность раскрыта в ЕР 0420358. Альтернативно сигнальная последовательность представляет собой чужеродную (гетерологическую) по отношению к полинуклеотидной последовательности, кодирующей полипептид, в этом случае сигнальная последовательность предпочтительно является эндогенной для клетки-хозяина, в которой экспрессируется последовательность ДНК. Сигнальная последовательность может использоваться в комбинации с промотором, направляющим экспрессию кодирующей последовательности, из которой получена сигнальная последовательность, например промотор амилоглюкозидазы Aspergillus (niger) (которая также называется (глюко)амилазой) в комбинации с сигнальной последовательностью из гена амилоглюкозидазы (AG), как вариант, содержащий 18, так и вариант, содержащий 24 аминокислоты, а также в комбинации с другими промоторами.
В контексте настоящего изобретения могут использоваться гибридные сигнальные последовательности. Примеры приемлемых сигнальных последовательностей для дрожжевых клеток-хозяинов имеют происхождение от генов дрожжевого α-фактора. Приемлемая сигнальная последовательность для бактерий имеет происхождение от гена α-амилазы (Bacillus).
В некоторых случаях во время прохождения полипептида через секреторный путь метаболизма может происходить расщепление сигнального пептида более, чем в одном положении, что влечет за собой получение зрелого полипептида с вариабельным N-концом.
Ниже полинуклеотидной последовательности, кодирующей полипептид, может иметься 3'-нетранслируемый участок, содержащий один или более сайтов терминации транскрипции (например, терминатор). Происхождение терминатора является не таким существенным. Терминатор может быть, например, природным по отношению к полинуклеотидной последовательности, кодирующей полипептид. Однако преимущественно в дрожжевых клетках-хозяинах используется дрожжевой терминатор, а терминатор нитевидных грибов используется в нитевидных грибковых клетках-хозяинах. Более предпочтительно, когда терминатор является эндогенным по отношению к клетке-хозяину (в которой экспрессируется полинуклеотидная последовательность, кодирующая полипептид).
Вектор может содержать один или более селективных маркерных генов для того, чтобы позволить проводить селекцию трансформированных клеток от большинства нетрансформированных клеток.
Предпочтительно, когда селективные маркеры включают, но не ограничены, те, которые восполняют дефект клетки-хозяина или обеспечивают устойчивость к лекарственному средству. Они включают, например, изменчивые маркерные гены, которые могут использоваться для трансформации большинства нитевидных грибов и дрожжей, такие, как гены ацетамидазы, или кДНК (гены amdS, niaD, facA или кДНК из A.nidulans, A.oryzae, или A.niger), или гены, обеспечивающие устойчивость к антибиотикам, подобным G418, гигромицину, блеомицину, канамицину, флеомицину, или те, которые обеспечивают беномильную устойчивость (benA). Альтернативно, могут использоваться специфические селективные маркеры, такие, как ауксотрофные маркеры, которые требуют соответствующих мутантных штаммов-хозяинов: например, URA3 (из S.cerevisiae или аналогичных генов, полученных из других дрожжей), pyrG или pyrA (из A.nidulans или A.niger), argB (из A.nidulans или A.niger) или trpC. В предпочтительном воплощении изобретения селективный маркер делегируют из трансформированной клетки-хозяина после введения экспрессионной конструкции так, чтобы получить трансформированные клетки-хозяина, способные к продуцированию полипептида, который является свободным от генов селективного маркера.
Другие маркеры включают АТФ синтетазу, субединицу 9 (oliC), оротидин-5'-фосфат-декарбоксилазу (pvrA), бактериальный ген устойчивости G418 (он также может использоваться в дрожжах, но не в грибах), ген резистентности к ампициллину (E. coli), ген резистентности к неомицину (Bacillus) и ген uidA E.coli, кодирующий b-глюкуронидазу (GUS). Векторы могут использоваться in vitro, например, для получения РНК, или использоваться для трансфекции или трансформации клетки-хозяина.
Последовательность ДНК, кодирующую полипептид, предпочтительно вводят в приемлемого хозяина как часть экспрессионной кассеты. Для трансформации приемлемого хозяина с помощью экспрессионной кассеты доступны способы трансформации, которые известны специалисту в данной области. Экспрессионная кассета может использоваться для трансформации хозяина как часть вектора, который несет селективный маркер, или экспрессионная кассета может быть совместно трансформирована как отдельная молекула вместе с вектором, несущим селективный маркер. Вектор может включать один или более селективных маркерных гена.
Для большинства нитевидных грибов и дрожжей вектор или экспрессионную конструкцию предпочтительно интегрируют в геном клетки-хозяина для того, чтобы получить стабильные трансформанты. Однако для определенных дрожжей также доступными являются эписомальные векторы, в которые может быть встроена экспрессионная конструкция для стабильного и высокого уровня экспрессии. Примеры таковых включают векторы, имеющие происхождение от 2m и pKD1 плазмид Saccharomyces и Kluyveromyces, соответственно, или векторов, содержащих последовательность АМА (например, АМА1 из Aspergillus). В случае, когда экспрессионные конструкции являются интегрированными в геном клетки-хозяина, конструкции интегрируют либо в случайном локусе генома или в предварительно определенном целевом локусе при использовании гомологической рекомбинации, в этом случае целевой локус предпочтительно включает высоко экспрессируемый ген. Ряд примеров высоко экспрессируемых генов был представлен ранее.
Также обеспечиваются клетки-хозяина, содержащие полинуклеотид или вектор в соответствии с изобретением. Полинуклеотид может быть гетерологическим по отношению к геному клетки-хозяина. В контексте данного изобретения термин «гетерологический» означает, что полинуклеотид не существует в природе в геноме клетки-хозяина или что полипептид в природных условиях не вырабатывается клеткой.
Приемлемые клетки-хозяева предпочтительно представляют собой прокариотические микроорганизмы, такие как бактерии, или более предпочтительно, эукариотические организмы, например, грибы, такие как дрожжи или нитевидные грибы, или растительные клетки.
Бактерии из рода Bacillus являются весьма приемлемыми в качестве гетерологических хозяев ввиду своей способности секретировать белки в культуральную среду. Другие бактерии, которые приемлемы как хозяева, представляют собой таковые из родов Streptomyces и Pseudomonas.
Предпочтительной дрожжевой клеткой-хозяином для экспрессии последовательности ДНК, кодирующей полипептид в соответствии с изобретением, являются роды Saccharomyces, Kluyveromyces, Hansenula, Pichia, Yarrowia и Schizosaccharomyces. Более предпочтительные дрожжевые клетки-хозяины выбраны из группы, состоящей из видов Saccharomyces cerevisiae, Kluyveromyces lactis (который также известен, как Kluyveromyces marxianus var. lactis), Hansenula polymorpha, Pichia pastoris, Yarrowia lipolytica и Schizosaccharomyces pombe.
Наиболее предпочтительные клетки-хозяева нитевидных грибов выбраны из группы, включающей Aspergillus, Trichoderma, Fusarium, Disporotrichum, Penicillium, Acremonium, Neurospora, Thermoascus, Myceliophtora, Sporotrichum, Thielavia и Talaromyces. Наиболее предпочтительная клетка-хозяин нитевидных грибов является таковой, выбранной из видов Aspergillus oryzae, Aspergillus sojae, Aspergillus nidulans или видов группы Aspergillus niger (как определено Raper и Fennell, The Genus Aspergillus, The Williams & Wilkins Company, Baltimore, стр. 293-344, 1965). Таковые включают, но не ограничены, Aspergillus niger, Aspergillus awamori, Aspergillus tubingensis, Aspergillus aculeatus, Aspergillus foetidus, Aspergillus nidulans, Aspergillus japonicus, Aspergillus oryzae and Aspergillus ficuum, а также включают Trichoderma reesei, Fusarium graminearum, Penicillium chrysogenum, Acremonium alabamense, Neurospora crassa, Myceliophtora thermophilum, Sporotrichum cellulophilum, Disporotrichum dimorphosporum и Thielavia terrestris.
Примеры предпочтительных зкспрессионных хозяев в рамках настоящего изобретения представляют собой грибы, такие как виды Aspergillus, Trichoderma; бактерии, такие как виды Bacillus, например Bacillus subtilis. Bacillus licheniformis, Bacillus amyloliquefaciens, Pseudomonas; и грибы, такие как виды Kluyveromyces, например Kluyveromyces lactis и виды Saccharomyces, например Saccharomyces cerevisiae.
Клетки-хозяева в соответствии с изобретением также включают растительные клетки, и изобретение, таким образом, охватывает трансгенные организмы, такие как растения и их части, которые содержат одну или более клеток в соответствии с изобретением. Трансгенное (или генетически модифицированное) растение содержит встроенную (например, стабильно) в геном последовательность, кодирующую один или более полепептидов в соответствии с изобретением. Трансформация растительных клеток может быть осуществлена при использовании известных способов, например, при использовании Ti- или Ri-плазмид из Agrobacterium tumefadens. Плазмида (или вектор) может, таким образом, содержать последовательности, необходимые для инфицирования растения, при этом могут также использоваться производные Ti- и/или Ri-плазмид.
Альтернативно может быть осуществлено непосредственное инфицирование части растения, такой как листья, корни или стебель. В этом способе растение, которое инфицируют, может быть подвергнуто ранению, например, путем срезания растения с помощью бритвы или прокалывания растения с помощью иглы или втирания в растения абразива. Рану потом инокулируют с помощью Agrobacterium.
Растение или часть растения могут потом выращиваться на приемлемой культуральной среде, при этом ему позволяют развиваться до зрелого растения. Регенерация трансформированных клеток в генетически модифицированные растения может быть достигнута при использовании известных способов, например, путем селекции трансформированных побегов с применением антибиотиков и путем субкультивирования побегов на среде, содержащей приемлемые питательные вещества, ростовые гормоны и тому подобное.
Дальнейший аспект изобретения, таким образом, относится к клеткам-хозяевам, трансформированным или трансфицированным с помощью полинуклеотида или вектора в соответствии с изобретением. Предпочтительно, когда полинуклеотид помещают в вектор для репликации полинуклеотида и экспрессии полипептида. Клетки выбирают таким образом, чтобы они были совместимыми с указанным вектором, они могут быть, например, прокариотическими (например, бактериальными), грибковыми, дрожжевыми или растительными клетками.
Гетерологический хозяин может быть также выбран так, что полипептид в соответствии с изобретением получают в форме, которая является существенно свободной от других полипептидов с такой же активностью, что и полипептид в соответствии с изобретением. Этого достигают путем выбора хозяина, который в норме не вырабатывает такие полипептиды с подобной активностью. Если полинуклеотиды в соответствии с изобретением встроены в рекомбинантный вектор, способный к репликации, то вектор может быть использован для репликации полинуклеотида с совместимой клеткой хозяина.
Таким образом, в дальнейшем аспекте изобретение обеспечивает способ получения полинуклеотида в соответствии с изобретением путем встраивания полинуклеотида в соответствии с изобретением в способный к репликации вектор, встраивания вектора в совместимую клетку-хозяина и выращивания клетки-хозяина в условиях, которые вызывают репликацию вектора. Вектор, содержащий полинуклеотид в соответствии с изобретением, может быть восстановлен из клетки-хозяина. Приемлемые клетки-хозяева включают бактерии, такие как Е.coli.
В дальнейшем аспекте изобретение обеспечивает способ получения полипептида в соответствии с изобретением путем культивирования клетки-хозяина (например, трансформированной или трансфицированной с помощью экспрессионного вектора, как описано выше) в условиях, которые обеспечивают экспрессию (с помощью вектора) полипептида в соответствии с изобретением, и необязательное восстановление экспрессируемого полипептида. Предпочтительно, когда полипептид получают как секретируемый белок, в этом случае полинуклеотидная последовательность, кодирующая зрелую форму полипептида в экспрессионной конструкции, является оперативно связанной с полинуклеотидной последовательностью, кодирующей сигнальный пептид.
Рекомбинантные клетки-хозяева в соответствии с изобретением могут культивироваться при использовании способов, известных в данной области техники. Для каждой комбинации промотора и клетки-хозяина условия культивирования являются доступными, при этом такие условия способствуют экспрессии полипептида в соответствии с изобретением. После достижения желаемого уровня плотности клеток или титра полипептида рост культуры останавливают и полипептид выделяют при использовании известных способов.
Ферментационная среда может включать известную культуральную среду, содержащую источник углерода (например, глюкозу, мальтозу, мелассу), источник азота (например, сульфат аммония, нитрат аммония, хлорид аммония, органические источники азота, например, дрожжевой экстракт, солодовый экстракт, пептон) и другие неорганические источники азота (например, фосфат, магний, калий, цинк, железо и тому подобное). Необязательно может включаться индуктор.
Выбор приемлемой культуральной среды может основываться на выборе экспрессионного хозяина и/или основываться на регуляторных требованиях экспрессионной конструкции. Такие среды известны специалисту в данной области. Среда может, если это является желательным, содержать дополнительные компоненты, более благоприятно воздействующие на трансформированных экспрессионных хозяев по сравнению с другими потенциально контаминирующими микроорганизмами.
Ферментацию проводят в течение периода 0,5-30 дней. Это может быть серийное производство, непрерывный процесс или процесс выращивания подпитываемой культуры, преимущественно при температуре в пределах от 0 до 45°С и, например, при значении рН от 2 до 10. Предпочтительными условиями ферментации являются температура в пределах от 20 до 37°С и/или значение рН от 3 до 9. Приемлемые условия обычно выбирают на основании выбора экспрессионного хозяина и белка, который экспрессируется.
После ферментации, если это является необходимым, удаляют клетки из ферментационного бульона путем центрифугирования или фильтрации. После окончания или после удаления клеток полипептид в соответствии с изобретением может быть восстановлен и, если это является желательным, очищен и изолирован с помощью традиционных способов.
Традиционно полипептид в соответствии с изобретением объединяют с приемлемыми (твердым или жидкими) носителями или разбавителями, включая буферы для получения композиции полипептида. Полипептид может быть присоединен или смешан с носителем, например иммобилизован на твердом носителе. Таким образом, настоящее изобретение обеспечивает в дальнейшем своем аспекте композицию, содержащую полипептид в соответствии с изобретением. Такая композиция может быть в форме, приемлемой для упаковки, транспортировки и/или хранения, предпочтительно, когда активность полипептида сохраняется. Композиции могут быть в форме пасты, жидкости, эмульсии, порошка, слоя, гранулята, пилюли или другой экструзивной формы.
Композиция может также включать дополнительные ингредиенты, такие как один или более (дополнительно) ферментов.
Полипептид типично стабильно рецептируют либо в жидкой, либо в сухой форме. Обычно продукт получают как композицию, которая будет необязательно включать, например, стабилизирующий буфер и/или консервант.
Изобретение дополнительно относится к продуктам питания или к композиции животного корма, или к добавке, которые содержат один или более полипептидов в соответствии с изобретением. Полипептид может присутствовать в пище в концентрации, которая отличается от природной концентрации. Предпочтительные количества составляют от 0,1 до 100, например от 0,5 до 50, предпочтительно от 1 до 10 мг на кг пищи.
Изобретение также относится к способу получения композиции животного корма, способу, включающему добавление к одному или более перевариваемому веществу (веществам) или ингредиенту (ингредиентам) полипептида в соответствии с изобретением. Полипептиды могут прибавляться к композиции животного корма отдельно от пищевых веществ или ингредиентов, индивидуально или в комбинации с другими пищевыми добавками. Полипептид может быть составной частью одного из веществ корма или ингредиентов.
Полипептиды в соответствии с изобретением могут также прибавляться к животным кормам для улучшения разложения растительных составляющих, например фитата, приводя к улучшению усвоения растительных питательных веществ животными. Преимущественно полипептиды в соответствии с изобретением могут продолжать разлагать фитат в корме in vivo. Полипептиды в соответствии с изобретением, полученные с помощью грибов, в частности в общем случае имеют более низкое оптимальное значение рН и способны к высвобождению важных питательных веществ в такой кислой окружающей среде, как желудок животного.
Полипептиды в соответствии с изобретением могут также использоваться при получении заменителей молока (или суррогатов) из сои. Такие заменители молока могут потребляться людьми и/или животными.
Композиция может дополнительно включать (в частности, когда рецептируется для использования в животном корме) один или более ионофоров, окисляющих агентов, сурфактантов, аминокислот, защищенных от действия первого отдела желудка жвачных, усилителей действия ферментов, которые могут быть получены природным путем в желудочно-кишечном тракте животных, потребляющих корм. При добавлении к кормам (включая силос) для жвачных животных или животных с одним отделом желудка (например, домашней птицы или свиней) корма могут включать зерновые культуры, такие как ячмень, пшеница, кукуруза, рожь или овес, или побочные продукты производства круп, такие как пшеничные отруби, или другие растительные материалы, такие как соевые бобы или другие бобовые. Фермент(ы) может(могут) в значительной степени улучшать разложение растительного материала, что ведет к лучшей утилизации растительных питательных веществ животными. Как следствие, может улучшается скорость роста и/или переваривание пищи.
Полипептиды в соответствии с изобретением являются особенно пригодными для животных кормов, поскольку они остаются активными в сильно кислых условиях, таких как желудок животных.
Один способ для (экзогенного) добавления полипептида в соответствии с изобретением заключается в прибавлении полипептида как материала трансгенных растений и/или (например, трансгенных) семян. Полипептид может, таким образом, быть синтезирован посредством гетерологической генной экспрессии, например ген, кодирующий желаемый фермент может быть клонирован в растительный экспрессионный вектор под контролем приемлемых сигналов растительной экспрессии, например ткане-специфического промотора, такого как промотор, специфический для семян. Экспрессионный вектор, содержащий ген, кодирующий полипептид, может быть последовательно трансформирован в цельные растения. Полученные таким образом трансгенные растения могут выращиваться, после чего собирают урожай, а те части растений, которые содержат гетерологический (по отношению к растениям) полипептид могут включаться в одну из композиций либо как таковые, либо после дополнительной обработки. Гетерологический полипептид может содержаться в семенах трансгенных растений или он может содержаться в других частях растений, таких как корни, стебли, листья, древесина, цветы, кора и/или плоды. Приемлемые растения включают злаки, такие как овес, ячмень, пшеница, кукуруза и рис.
Добавление полипептида в форме материала трансгенных растений, например в виде трансгенных семян, может требовать обработки растительного материала так, чтобы сделать полипептид доступным, или, по меньшей мере, улучшить его доступность. Такие технологии обработки могут включать различные механические (например, помол и/или шлифовку) способы или термомеханическую обработку, такую как экструзия или увеличение объема.
Настоящее изобретение, таким образом, также относится к способу улучшения роста и/или переваривания пищи у животных с одним отделом желудка или нежвачных животных, способу, включающему скармливание животному полипептида в соответствии с изобретением. Приемлемые животные включают сельскохозяйственные животные с одним отделом желудка и/или нежвачных животных, таких как свиньи (или поросята), домашнюю птицу (такую, как куры, индюки), телят или водных (например, морских) животных (например, рыбу).
Пример 1
Получение штаммов, вырабатывающих фитазу
Фрагменты ДНК, которые имеют последовательность в соответствии с SEQ ID NO:2, SEQ ID NO:4 и SEQ ID NO:6, были получены синтетическим путем. После проверки последовательности ДНК эти синтетически фрагменты генов были слиты с сигнальной последовательностью фитазы A.niger при использовании ПЦР и клонированы под контролем промотора глюкоамилазы. С этой целью ген phyA в том виде, в котором он присутствует в экспрессионном векторе pGBTOPFYTI, как описано в международной патентной заявке WO 98/46772, заменяли генами модифицированной фитазы, описанными выше, получая при этом векторы pTHFYT2, pTHFYT4 и pTHFYT6, соответственно.
Экспрессионные векторы pTHFYT2, pTHFYT4 и pTHFYT6 были встроены в Aspergillus niger CBS 646.97 (описан в WO 98/46772). При использовании ПЦР отбирали трансформанты, содержащие pTHFYT2, pTHFYT4 или pTHFYT6. Для того чтобы определить способны ли эти трансформанты секретировать активную фитазу, трансформанты выращивали на планшетах, содержащих фитат, как описано Chen (1998, Biotechnol. Technique 12, 759-761). В этом анализе ореолы вокруг колоний Aspergillus становились видимыми в том случае, если секретировалась активная фитаза, благодаря разложению фитата. При использовании этого анализа было показано, что все экспрессионные векторы приводили к образованию трансформантов, секретирующих активную фитазу FYT2, FYT4 или FYT6 в среду.
Трансформанты, продемонстрировавшие прозрачные ореолы, выращивали в качающейся колбе. Инокулировали 107 спор отобранных трансформантов и контрольных штаммов в качающиеся колбы, содержащие 20 мл жидкой предварительной среды, содержащей на литр: 30 г мальтоза · Н2O; 5 г дрожжевого экстракта; 10 г гидролизованного казеина; 1 г КН2PO4; 0,5 г MgSO4·7Н2О; 0,03 г ZnCl2; 0,02 г CaCl2; 0,01 г MnSO4·4H2O; 0,3 г FeSO4·7H2O; 3 г Твина 80; 10 мл пенициллина (5000 МЕ/мл) / стрептомицин (5000 ед./мл); рН 5,5. Эти культуры выращивали при температуре 34°С в течение 20-24 часов. 10 мл этой культуры инокулировали в 100 мл ферментационной среды А. niger, содержащей на литр: 70 г мальтодекстринов; 25 г гидролизованного казеина; 12,5 г дрожжевого экстракта; 1 г КН3PO4; 2 г K2SO4; 0,5 г MgSO4·7Н2О; 0,03 г ZnCl2; 0,02 г CaCl2; 0,01 г MnSO4 · 4Н2O; 0,3 г FeSO4·7H2O; 10 мл пенициллина (5000 МЕ/мл)/стрептомицина (5000 ед./мл); значение рН доводили до 5,6 с помощью 4 N H2SO4. Эти культуры выращивали при 34°С в течение 6 дней. Образцы, взятые из ферментационного бульона, центрифугировали (10', 5.000 об./мин в качающейся центрифуге) и собирали супернатанты.
Пример 2
Термостабильные FYT2, FYT4 и FYT6
FYT2, FYT4, FYT6 и фитазу A.ficuum (EP 0420358) получали в качающихся колбах в соответствии с Примером 1. Получение фитазы сопровождали отбором образцов через приемлемые временные интервалы и оценкой супернатантов на фитазную активность в соответствии со способом, как описано van Engelen и др. (Journal of AOAC International 1994, 77:760-764). Активность выражали в FTU, где 1 FTU представляет собой количество носителя, который высвобождает 1 мкмоль неорганического ортофосфата за минуту в контрольных условиях (рН 5,5, температура 37°С, 5 мМ фитата натрия). Термостабильность измеряли в супернатантах так, как описано ниже. Когда это является необходимым, то супернатанты фитазы подвергали дальнейшему концентрированию с помощью ультрафильтрации.
Термостабильность измеряли тремя различными путями. Во-первых, определяли значение Т50 для фитазы. Т50 (в °С) представляет собой температуру, при которой теряется 50% активности после нагревания образцов в течение 20 минут. Экспериментальные условия: стресс-тест осуществляли в 250 мМ НАс/NaAc/Твин20 рН 4,0. Фитазу использовали в дозе приблизительно 0,6 FTU/мл. После нагревания образцы немедленно охлаждали на льду. После этого активность фитазы измеряли в 250 мМ НАс/NaAc/Твин20 рН=4,0. Результаты представлены в Таблице 1. FYT2 и FYT4 были приблизительно на 8-9°С более стабильными.
Во-вторых, активность фитаз определяли во время нагревания. При определении в качестве функции температуры нагревания эксперимент давал оптимальную температуру (Topt) фермента в отношении продуктивности. Инкубацию осуществляли в течение 30 минут. Количество превращенного субстрата зависит от ферментативной активности, а также от инактивации. Таким образом, предпочтительно используется термин продуктивности вместо термина активности. Экспериментальные условия: 250 мМ НАс/МаАс/Твин20 рН 4,0, доза фитазы составляет приблизительно 0,012 FTU/мл. Высвобожденный фосфат может быть измерен в соответствии с методами стандартного анализа, как описано выше. Результаты представлены на Фиг.3. FYT2 также, как и FYT4, являются наиболее эффективными в отношении каталитической продуктивности при температуре 75°С, в то время, как контроль дикого типа полностью терял свою каталитическую активность. Несмотря на то, что активность начинает уменьшаться при температуре выше 75°С, Фиг.3 показывает, что FYT2 также, как и FYT4, является каталитически достаточным при температурах, выше приблизительно 85°С. Поведение FYT6 находится между таковым дикого типа и FYT2 или FYT4.
В-третьих, в дополнение к измерению термостабильности посредством приемлемого анализа на активность термостабильность фитаз была также определена непосредственно путем определения температуры, при которой происходит разворачивание трехмерной структуры фермента фитазы. Эффект нагревания, который сопровождает развертывание, может быть измерен непосредственно путем дифференциальной сканирующей калориметрии (DSC). Экспериментальные условия DSC: 250 мМ HAc/NaAc рН 4,0, приблизительно 5 мг/мл фитазы, скорость нагревания составляет 2,5°С/мин. Результаты представлены в Таблице 1. Можно увидеть, что структура нативного фермента поддерживается при температуре, которая на 9°С выше для модифицированных фитаз FYT2 и FYT4, чем для фитазы дикого типа.
Пример 3
Стабильность при гранулировании FYT2, FYT4 и FYT6
Анализы с двумя гранулирующими материалами и, таким образом, с температурными режимами, осуществляли с культуральными фильтратами FYT2, FYT4 и FYT6.
Все грануляты получали путем перемешивания/замеса культурального фильтрата с необходимым количеством кукурузного крахмала (С-гель от Cerestar) и воды (см. Таблицы 2 и 3 для композиции влажной смеси). После перемешивания и замеса смесь подвергали экструзии с помощью экструдера Nica E-220 и подвергали сферонизации с помощью устройства для сферонизации Fuji Paudal QJ-400G. Полученные частицы высушивали в сушилке с псевдоожиженным слоем Glatt GPCG 1.1. Активность гранул составляла от 2500 до 3000 FTU/г.
250 г. гранул перемешивали в 25 кг пищи с композицией в соответствии с Таблицей 4, а также перемешивали непосредственно перед анализом с 225 кг той же рецептуры. 25 кг корма перемешивали в планетарной мешалке Collete MP90 в течение 10 минут. 25 кг и 225 кг корма перемешивали в мешалке Nautat на 1200 литров. Образцы этой смеси брали для определения стабильности при гранулировании. Эти 250 кг смеси дозировали в мешалку/чаны с помощью дозирующего винта при скорости 600 кг/час, когда его подвергали нагреванию с помощью непосредственного вдувания пара до температуры 80°С. Продолжительность обработки составляла приблизительно 10-15 секунд, после чего горячую смесь выдавливали в пресс для гранулирования. Матрикс, используемый в экспериментах, имел 5/45 мм (ширина/длина; RDS 05) или 3/65 мм (ширина/длина; RDS A1). Температура гранул, покидающих пресс, составляла 82-83°С (RDS 05) или 91-93°С (RDS A1). После прессования гранулы падали на охлаждающий ленточный конвейер, с этого конвейера брали образцы для проведения исследования на стабильность.
Опыты по гранулированию показали подобное увеличение термостабильности FYT2, FYT4 и FYT6 по сравнению с диким типом при температуре 82°С, в то время, как при 92°С FYT2 и FYT4 показывали более высокую стабильность по сравнению с FYT6.
Пример 4
Биохимические характеристики фитаз
Специфическую активность фитаз определяли после очистки фитаз из фильтратов, которые были получены после фильтрации ферментационного бульона. Очистку фитазы осуществляли с помощью ионообменной хроматографии, или аффинной хроматографии, или комбинации двух методов.
Аффинную хроматографию гликозилированных фитаз осуществляли при использовании ConA (Concanavalin А) аффинного матрикса (HiTrap Con A, Amersham Pharmacia Biotech). Фитазу связывали с колонкой в 20 мМ Трис/0,5 М NaCl/1 мМ MnCl2/1 мМ CaCl2 / рН 7,4. После обильного промывания колонки фитазу элюировали с помощью 20 мМ Трис/ 0,5 М NaCl/ 0,5 М метил-α-глюкопиранозида/ рН 7,4.
Регенерацию колонки проводили с помощью 20 мМ Трис рН 8,5. Значение рН буферов устанавливали с помощью 4N HCl.
Ионообменную хроматографию осуществляли при использовании анионного обменника (Resource Q, Amersham Pharmacia Biotech). Обессоливание и смену буфера осуществляли при использовании колонки для гельфильтрации PD-10. Уравновешивание колонки проводили в 50 мМ Трис, рН 7,5. После загрузки образца фитазу элюировали при использовании градиента от 0 до 1 М NaCl в 50 мМ Трис рН 7,5.
Содержание белка очищенной фитазы определяли с помощью измерения Е280, при этом 1 мг/мл фитазы соответствует OD280,1cм=0,938. Для FYT2, FYT4 и FYT6, OD280,1cм при 1 мг/мл соответствует 0,995, 0,995 и 0,963 соответственно. Активность определяли в FTU, как описано van Engelen и др. (Journal of AOAC International 1994, 77:760-764).
Значения Km для фитиновой кислоты определяли путем измерения начальной скорости реакции как функции концентрации субстрата. Смеси для анализа содержали либо 1,0, 0,5, 0,2, 0,1, 0,05, 0,025, 0,015 мМ фитиновой кислоты в 250 мМ NaAc буфера рН 5,5. Ферментативную реакцию останавливали с помощью 15% ТСА (1:1). Высвобожденный неорганический фосфат определяли путем перемешивания смеси, полученной после завершения реакции с 0,6 М H2SO4 - 2% аскорбиновой кислоты - 0,5% молибдата аммония (1:1), инкубирования смеси в течение 20 минут при 50°С и измерения поглощения при 820 нм (Wyss и др. AppI Env Microbiol. 1999, 65: 367-373). Результаты представлены в Таблице 7.
Таблица 7 показывает, что по сравнению с диким типом специфическая активность также, как и высокая аффинность для фитиновой кислоты, не подвергаются изменениям вследствие осуществленных модификаций.
Зависимость активности фитаз от рН определяли путем измерения скорости высвобождения фосфата из фитиновой кислоты при различных значениях рН. В общем использовали стандартный анализ на фитазу, за исключением того, что значение рН может варьировать. Активность при рН 5,6 принимали как 100% активность. Использовали следующие буферы для установления рН в эксперименте: 250 мМ глицина в интервале рН от 2,8 до 3,2; 250 мМ NaAc в интервале рН от 3,6 до 5,6; 250 мМ имидазола в интервале рН от 6 до 7 и 250 мМ Трис в интервале рН от 7,5 до 9. Результаты представлены в Таблице 8.
Зависимость активности модифицированных фитаз от рН очень подобна таковой дикого типа. В частности, сохраняется то свойство, что фитаза дикого типа демонстрирует два оптимума рН. Один оптимум рН находится вблизи значения рН 2,5, а второй оптимум находится вблизи значения рН 5,5. Таблица 8 демонстрирует, что это особое свойство фитазы дикого типа Aspergillus niger не подвергается влиянию модификаций, которые приводят к получению FYT2, FYT4 и FYT6.
Кривые прохождения процесса деградации фитиновой кислоты с помощью фитазы как функцию времени регистрировали в 250 мМ NaAc, pH=5,5 при 37°С. Дозировка фитазы составляла 0,05 FTU/мл при концентрации субстрата 0,2 мМ фитиновой кислоты. Ферментативную реакцию останавливали с помощью ТСА (1:1). Высвобожденный неорганический фосфат определяли путем перемешивания 100 мкл реакционной смеси с 1400 мкл 0,3 М H2SO4 - 1% аскорбиновой кислоты - 0,27% молибдата аммония, последующего инкубирования смеси в течение 20 минут при 50°С и измерения поглощения при 820 нм. Результаты представлены в Таблице 9.
Было показано, что модифицированные фитазы ведут себя подобно фитазе дикого типа при высвобождении фосфата из фитиновой кислоты. Через один час кривые протекания процесса достигали плато при значении приблизительно 80-85% высвобожденного фосфата. Все фитазы достигали такого плато при подобной скорости, что свидетельствует о том, что на эффективность фитазы дикого типа в высвобождении фосфатов из фитиновой кислоты не воздействуют модификации, содержащиеся в модифицированных фитазах.
В заключение результаты показывают, что модификации FYT2, FYT4 и FYT6 не влияют на каталитическое поведение данных модифицированных фитаз по сравнению с таковым для дикого типа. Было продемонстрировано, что аннулирование любой мутации аминокислот из тех, что показаны на Фиг.1 (остатки, находящиеся в пределах участка 7 Ангстрем вблизи субстрата), и дальнейшее сохранение дополнительных аминокислот фитазы Aspergillus niger не поддерживает функциональные свойства фитазы дикого типа Aspergillus niger.
Пример 5
Биоэффективность модифицированных фитаз
Эксперимент с жидкими фитазами
Тест по сравнению биоэффективности вновь полученных термостабильных фитаз проводили при использовании их в виде жидкой композиции, которая применялась к гранулам после гранулирования. Ферменты использовали в таких дозах, чтобы прибавленная фитазная активность составляла 100, 200 или 300 FTU/кг корма. Тест проводили на бройлерах (5-33 дня), которых кормили кормом на основе кукурузы и сои (один корм для первых 14 дней опыта, и другой, который немного отличался, в течение последних 14 дней). Содержание всасываемого фосфора в основной диете составляло 2,2 г/кг корма (дни 5-19) и 1,7 г/кг корма (дни 19-33). Эти значения были ниже оцененных потребностей животных. Животных помещали в шестиэтажные клетки для исследования, при этом каждая клетка содержала 14 птиц. Результаты подсчитывали на основе проанализированного содержания фитазы (FTU/кг), используя способы, описанные Finney (1964: Statistical method in biological assay. Charles Griffin, London). Результаты относительно веса животных представлены в Таблице 10.
Наклоны регрессионных линий для всех продуктов значительно отличались от нуля. Как видно из Таблицы 10, существовало незначительное различие между продуктами, за исключением FYT4. Животные, которые потребляли этот фермент, не чувствовали себя также хорошо, как те, которых кормили другими фитазами, но различие не было статистически значительным.
Опыт с гранулированными фитазами
Эксперимент осуществляли для сравнения термостабильных фитаз в гранулированной композиции, которую применяли к гранулам перед гранулированием. Это означает, что фитазы пропускали через процесс гранулирования. В этом эксперименте гранулирование проводили при действительно высоких температурах: температура гранул составляла приблизительно 92°С. Поскольку цель заключалась в получении добавок 100, 200 или 300 FTU/кг к корму, как предлагается для животных, эксперимент предварительного гранулирования осуществляли для оценки потери активности во время этого процесса, в продукты вводили большую дозу в такой мере, чтобы высвобождались упомянутые активности. Эксперимент осуществляли подобным образом, как и в случае опыта с жидким продуктом, при этом использовали бройлеров (5-33 дня), которых кормили кормом на основе кукурузы и сои (только один корм на протяжении всего периода), которые имели содержание усваиваемого фосфора значительно ниже оцененных требований этих животных (1,9 г/кг корма). Животных помещали в шестиэтажные клетки для обработки, каждая клетка содержала 14 птиц. Результаты относительных наклонов регрессионных кривых для достигнутого веса тела представлены в Таблице 11.
Фитаза FYT2 проявляла наилучшие характеристики в этом опыте, за ней шли FYT6, FYT4 и фитаза дикого типа.
Пример 6
Одиночные мутанты фитазы FYT2 и FYT6
Получали следующие одиночные мутанты фитазы FYT2: Y31F, А78Е, G163R, G180A, G182S, A194V, L211T, A215S, P242S, Е254К, Q269N, А414Р, E428R и Е440А, а также фитазы FYT6: Е440А.
С этой целью ген, кодирующий FYT-2 (SEQ ID NO:2) или FYT6 (SEQ ID NO:6), амплифицировали с помощью ПЦР в общем объеме 50 мкл при использовании 2,5 ед. Pwo полимеразы (Roche diagnostics, GmbH, Mannheim, Germany), 100 нг матричной ДНК, 0,5 мМ дМТР, 1 × Pwo буфера, 10 пмоль DSM-1F и 10 пмоль DSM-1R при следующих условиях 5' 94°С, 30× (30′′ 94°С 1′′ 60°С 2' 72°С), 5' 72°С. Амплифицированный фрагмент клонировали в PCR®-Blunt-TOPO (Invitrogen Life Technologies, Carlsbad, CA, USA) вектор и вводили мутации при использовании набора QuickChange от Stratagene (Stratagene, La Jolla, CA, USA) в соответствии с рекомендациями поставщика.
Последовательность праймеров DSM-1F и DSM-1R была такой, как приведено ниже:
DSM-1 F 5' GGCAGTCCCCGCCTCGAGAAAT 3'
DSM-1 R 5' GTCATCGCGATTAATTAATCTAAGCAAAACACTCCTCCCAGTT 3'
Для мутагенеза использовали следующие праймеры, при этом подвергнутые мутации кодоны выделены жирным шрифтом:

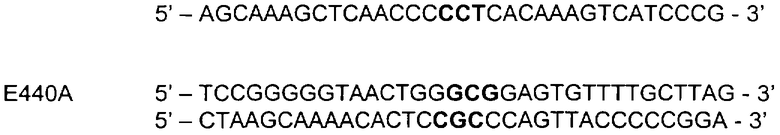
Последовательность полученных фрагментов ДНК фитазы была проверена с помощью анализа последовательности. Последовательности фитазы клонировали в pGBTOPFYT1 и супернатанты культур готовили так, как описано в Примере 1.
Для каждого единичного мутанта FYT2 и FYT6 определяли значение Т50 (см. Пример 2). Результаты показаны в Таблице 12.
Большинство полученных мутантов продемонстрировали значение Т50, которое является сравнимым с таковым для FYT2. Неожиданно было обнаружено, что значение Т50 для FYT6, содержащего комбинацию всех единичных мутаций, было значительно ниже такового для индивидуальных мутантов.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЕН ABFB-1 PENICILLIUM FUNICULOSUM | 2006 |
|
RU2388824C2 |
| ГЕН ABFB-2 PENICILLIUM FUNICULOSUM | 2006 |
|
RU2388820C2 |
| ВАРИАНТНЫЕ ФИТАЗНЫЕ ФЕРМЕНТЫ | 2005 |
|
RU2421520C2 |
| НОВАЯ ФИТАЗА, СПОСОБ ЕЕ ПОЛУЧЕНИЯ И ЕЕ ПРИМЕНЕНИЕ | 2015 |
|
RU2712881C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ ПРОТИВОМИКРОБНЫМ ДЕЙСТВИЕМ, И КОДИРУЮЩИЕ ИХ ПОЛИНУКЛЕОТИДЫ | 2006 |
|
RU2403260C2 |
| КЛОНИРОВАНИЕ, ЭКСПРЕССИЯ И ПРИМЕНЕНИЕ КИСЛЫХ ФОСФОЛИПАЗ | 2010 |
|
RU2567659C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ АНТИМИКРОБНОЙ АКТИВНОСТЬЮ, И ПОЛИНУКЛЕОТИДЫ, КОДИРУЮЩИЕ ИХ | 2006 |
|
RU2415150C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ АНТИМИКРОБНОЙ АКТИВНОВНОСТЬЮ, И ПОЛИНУКЛЕОТИДЫ, КОДИРУЮЩИЕ ИХ | 2006 |
|
RU2512525C2 |
| ПОЛИПЕПТИДЫ С ПРОТИВОМИКРОБНОЙ АКТИВНОСТЬЮ И КОДИРУЮЩИЕ ИХ ПОЛИНУКЛЕОТИДЫ | 2005 |
|
RU2393224C2 |
| ПОЛИПЕПТИДЫ С ЛИЗОЦИМНОЙ АКТИВНОСТЬЮ И ПОЛИНУКЛЕОТИДЫ, КОДИРУЮЩИЕ ИХ | 2012 |
|
RU2619051C2 |

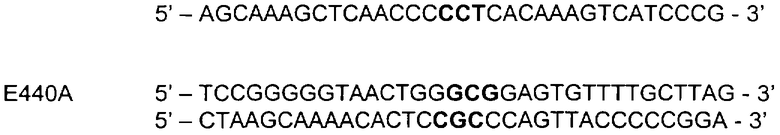













Изобретение относится к биотехнологии. Предложен полипептид, обладающий фитазной активностью. Последовательности приведены в описании. Описан полинуклеотид, кодирующий представленный полипептид. Раскрыто применение указанного полипептида для использования в кормовой композиции или кормовой добавке. Данное изобретение позволяет увеличить устойчивость модифицированной фитазы к температуре и влажности. 3 н. и 3 з.п. ф-лы, 12 табл., 10 ил.
| Способ использования делительного аппарата ровничных (чесальных) машин, предназначенных для мериносовой шерсти, с целью переработки на них грубых шерстей | 1921 |
|
SU18A1 |
| http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=Protein&list_uids=10732783&dopt=GenPept | |||
| Прибор с двумя призмами | 1917 |
|
SU27A1 |
| http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=Protein&list_uids=10732783&dopt=GenPept | |||
| RU 99105347 A, 2002.05.27. | |||

